 ,1,2, 王成栋1,2, 张震3, 任侠4, 刘美珍
,1,2, 王成栋1,2, 张震3, 任侠4, 刘美珍 ,1,2,*, 杨帆1,2
,1,2,*, 杨帆1,2Spatial distributions of biomass and carbon density in natural grasslands of Hebei, China
CEN Yu,1,2, WANG Cheng-Dong1,2, ZHANG Zhen3, REN Xia4, LIU Mei-Zhen,1,2,*, YANG Fan1,2通讯作者:
编委: 白永飞
责任编辑: 李敏
| 基金资助: |
Online:2018-03-20
| Supported by: | SupportedbytheStrategicPriorityResearchProgramoftheChineseAcademyofSciences.( |

摘要
关键词:
Abstract
Keywords:
PDF (2318KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
岑宇, 王成栋, 张震, 任侠, 刘美珍, 杨帆. 河北省天然草地生物量和碳密度空间分布格局. 植物生态学报, 2018, 42(3): 265-276 doi:10.17521/cjpe. 2015.0300
CEN Yu, WANG Cheng-Dong, ZHANG Zhen, REN Xia, LIU Mei-Zhen, YANG Fan.
草地资源是陆生生态系统的重要组成部分, 在全球陆地碳循环中起着重要作用。世界上的草地约占全球陆地面积的20%, 主要分布在各大陆内部气候干燥、降水较少的地区(Scurlock & Hall, 1998)。在中国, 草地是我国陆地上面积最大的生态系统, 主要分布在干旱、半干旱的温带地区, 是欧亚中高纬度草地生态系统的重要组成部分(陈佐忠和汪诗平, 2000), 我国的草地面积约有4 × 108 hm2 (中华人民共和国农业部兽医司和全国畜牧兽医总站, 1996), 可利用面积达3.1 × 108 hm2。草地生态系统对畜牧业的发展、生物多样性的保护、水土保持以及生态平衡的维持有重要的作用和价值(谢高地等, 2001)。
河北省的草地资源处于温带欧亚草原与森林过渡地带, 天然草地植被类型多样, 植物资源丰富, 草地畜牧业生产在当地国民经济中占有重要的地位(河北省畜牧水产局, 1990; 河北省畜牧局, 1997)。在2000年之前, 我国对天然草地利用方式多采取粗放的自然放牧方式, 由于没有监管, 大多数草地呈现中度或重度利用、轻建设、无管理的状态, 使得大部分草地出现不同程度的退化(李博, 1997; 韩永伟和高吉喜, 2005; 闫玉春和唐海萍, 2008)。河北草地也面临过度开垦、超载放牧、草地资源破坏严重等生态问题, 再加上频繁的干旱、风蚀、水蚀等自然因素的影响, 导致草地发生了不同程度的退化、沙化及盐碱化, 降低了河北省草地资源的生产力, 严重影响了河北省草地畜牧业的健康发展和生态环境保护(Ellis, 1992; 赵雪和张强, 1997; 张培栋和介小兵, 2007; 宋理明等, 2009; 徐敏云等, 2009, 2010)。
近年来, 相关的****已经对我国草地生物量开展了大量的研究, 对于大尺度范围内草地生物量的研究, ****多采用遥感的方法进行。如方精云等(1996)在实测与遥感相结合的基础上, 对我国北方温带荒漠草原、温带典型草原和温带草甸草原的地上生物量进行了估算; Ni (2004a)基于草地资源清查资料和遥感数据对我国北方以上3种草地类型的地上生物量进行了估算; Piao等(2007)也通过遥感方法估算了我国草地生物量的变化。对于我国草地碳密度与碳储量的分布与变化, 不同的****也开展了相关研究, 如Ni (2001, 2002)利用草地资源调查数据研究了中国草地的碳密度分布与碳储量; Fan等(2008)研究了我国草地地上和地下生物量的碳蓄积; Ma等(2010)研究了1982-2006年我国北方草地碳蓄积量的变化; Fang等(2010)对中国草地生物量平均碳密度进行了估算。不同的****虽然对我国的草地生物量与碳密度进行了大量的研究, 但是由于采用的研究方法、草地类型、数据来源等的不同导致估算的结果差异较大, 尤其是缺乏基于统一的草地生态系统调查规范所获得的大量实测数据对草地生物量与碳密度的估算。本研究采用统一的样地调查法全面分析了河北省境内不同草地类型生物量空间分布特征和碳密度变化规律, 结合海拔梯度和气候因子, 阐明影响草地生物量和碳密度变化的地理和气候因素, 揭示河北省草地植物碳密度变化规律, 为我国草地生态系统碳汇/碳源的深入研究以及系统分析草地植被在气候变化中的生态意义提供基础科学数据, 同时对于合理优化河北草地利用方式、管理利用强度以及加强生态环境保护、提高草地固碳潜力、促进草地畜牧业的可持续发展具有十分重要的意义。
1 材料和方法
1.1 试验地概况
河北省是我国华北地区的腹心地带, 地处113.45°-119.75° E, 36.85°-42.62° N, 全省年平均气温在4-13 ℃之间, 1月最低温-14- -2 ℃, 7月最高温20-27 ℃, 年降水量300-800 mm, 燕山南麓和太行山东降雨较多, 张北高原降水一般不足400 mm, 为季风性暖温带气候, 四季分明。河北省总面积187600 km2, 其中平原约占1/3, 山区和高原约占2/3。天然草地总面积4.74 × 106 hm2, 占全国草地面积的1.2%, 占全省土地面积的35.7%左右。河北省的草地大部分分布在北部与西北山地, 其中80.3%的草地分布在北部的承德、张家口和保定三市, 是坝上地区、北部山地以及太行山区发展畜牧业的基地(繆应庭, 1981)。植物群落的主要优势种有羊草(Leymus chinensis)、冰草(Agropyron cristatum)、龙牙草(Agrimonia pilosa)、荆条(Vitex negundo var. heterophylla)、胡枝子(Lespedeza bicolor)、河蒴荛花(Wikstroemia chamaedaphne)、委陵菜(Potentilla chinensis)、冷蒿(Artemisia frigida)与大针茅(Stipa grandis)等。1.2 取样方法
1.2.1 样地选择在20世纪90年代全国草地类型图和2000年土地利用图基础上, 以中国南、北草地资源调查办公室的草地分类法为依据, 结合1979-1984年河北省草地资源调查结果, 以草地类型为基本单元, 选择具有代表性的区域作为样地, 于2011-2013年进行草地调查。本实验共涉及6种草地类型, 78个取样地(图1)。6种草地类型分别为温性草原、温性草甸、温性山地草甸、低地盐化草甸、暖性草丛和暖性灌草丛。
图1
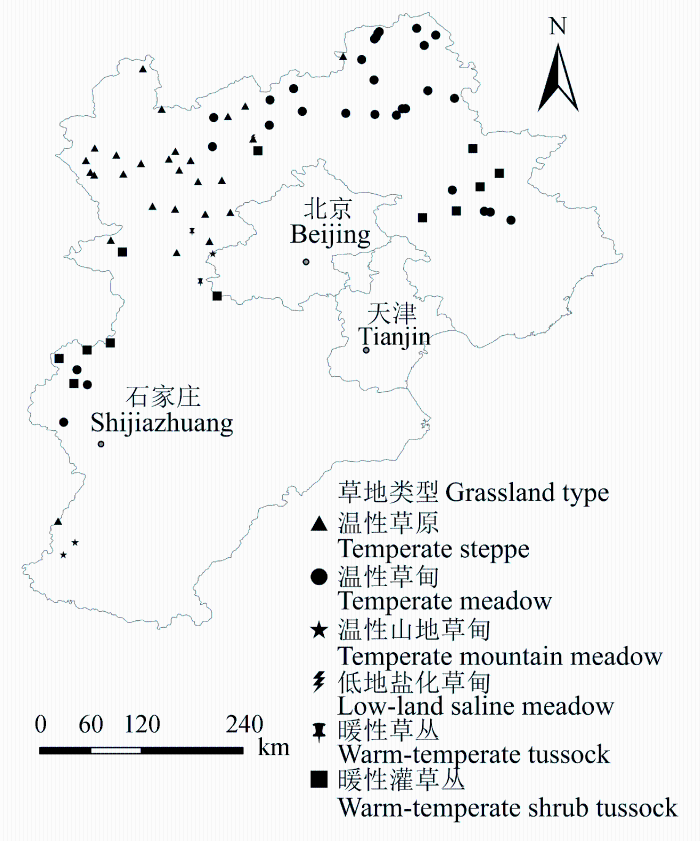 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1实验中6种草地类型的取样地在河北省的分布情况。
Fig. 1The distribution of sampling sites in the experiment in Hebei Province.
1.2.2 样方分布及取样原则
在设置的78个样地中, 每个样地选择一个100 m × 100 m的区域进行取样调查, 沿其对角线设置样线, 在样线上设置10个1 m × 1 m的草本样方, 包括5个草本分种样方和5个草本不分种样方, 草本分种样方主要是确定不同草地类型的优势种, 草本不分种样方用于测定不同草地类型的地上活体、根系和凋落物生物量以及相应的碳含量。
1.2.3 生物量的测定
在不分种样方内, 地上活体生物量采用齐地面刈割法, 将样方内植物地面以上的所有绿色部分用剪刀齐地面剪下, 在65 ℃烘箱中烘干至恒质量, 称量, 得到地上活体生物量。凋落物生物量指的是凋落物现存量的生物量, 凋落物现存量是地面以上还没有发生明显分解的多年累积的植物死亡残体, 调查中用小耙子进行收集采样, 带回实验室烘干至恒质量, 称量, 用干质量除以取样地表面积计算得到单位面积生物量(g·m-2)。
根系生物量的测定采用土钻法分层取样, 根钻直径为7 cm, 测定样方与地上部分测定的样方对应, 在5个样方中, 根系取样分为0-5、5-10、10-20、20-30、30-50、50-70和70-100 cm, 共7层, 每层取5钻土壤样品, 将相同层次分别混合为一个样品, 从各土层中钻取的土样经清水冲洗后拣出所有根系, 置于65 ℃烘箱中烘至恒质量, 称量。然后根据土钻面积和干质量换算为单位土地面积地下生物量(g·m-2)。
1.2.4 碳含量测定及碳密度计算
将烘干的植物样品用球磨仪(MM400, Retsch, Haan, Germany)粉碎, 用元素分析仪(VARIO EL III, Elementar, Langencelbold, Germany)测定植物样品中的总碳含量。然后根据公式计算植物地上活体、根系和凋落物生物量碳密度。植物体地上活体、根系和凋落物各部分碳密度测定估算公式为:
Cp = αM
式中, Cp为植物体碳密度(g C·m-2); α为生物量中总碳含量(g·kg-1); M为地上活体部分(凋落物或根系)生物量(g·m-2)。本研究中碳含量和生物量均采用实测值。
1.3 数据处理
实验所测数据采用Excel处理后, 用SPSS 14.0软件分析, 用混合线性模型分析不同草地类型生物量和碳密度的差异, 以及不同环境因素对生物量空间分布的影响, 用最小差异法(LSD)进行多重比较, 显著性水平α = 0.05。用Excel绘制图表。气象资料的获取: 河北省1960-2010年各县市月平均气象数据(积温、降水量)由国家气象局提供, 用于与草地生物量的回归关系分析。
2 结果
2.1 不同草地类型植物群落主要优势种
本研究区域的草地类型包括了温性草原、温性草甸、温性山地草甸、低地盐化草甸、暖性草丛和暖性灌草丛6种主要草地类型(表1)。温性草原主要分布在河北北部的张家口和承德市, 植被群落主要优势种为羊草、冰草、百里香(Thymus mongolicus)、大针茅等。温性草甸也主要分布在河北北部的张家口和承德市, 植被群落主要优势种为委陵菜、披针薹草(Carex lancifolia)、地榆(Sanguisorba officinalis)等。温性山地草甸主要分布在张家口和承德市的山地, 植被群落主要优势种为龙牙草、篷子菜(Galium verum)、直穗披碱草(Elymus gmelinii)等。低地盐化草甸主要分布在张家口北部和秦皇岛市的沿海岸地带, 由于海岸带盐化草甸面积较小, 本次调查中未涉及海岸带盐化草地, 植被群落主要优势种为叉分蓼(Polygonum divaricatum)、白花马蔺(Iris lactea)、芨芨草(Achnatherum splendens)等。暖性草丛主要分布在河北保定市、承德市和秦皇岛市, 植被群落主要优势种为阿拉伯黄背草(Themeda triandra)、矮蒿(Artemisia lancea)等。暖性灌草丛主要分布在保定市和邢台市, 植被群落主要优势种为胡枝子、绣线菊(Spiraea salicifolia)、金色狗尾草(Setaria glauca)、荆条等。Table 1
表1
表1河北省不同草地类型的样地数和植物群落主要优势种
Table 1
| 草地类型 Grassland type | 样本数 Sample size | 优势种 Dominant species |
|---|---|---|
| 温性草原 Temperate steppe | 30 | 冰草、羊草、百里香、大针茅 Agropyron cristatum, Leymus chinensis, Thymus mongolicus, Stipa grandis |
| 温性草甸 Temperate meadow | 13 | 委陵菜、披针薹草、地榆 Potentilla chinensis, Carex lancifolia, Sanguisorba officinalis |
| 温性山地草甸 Temperate mountain meadow | 9 | 龙牙草、篷子菜、直穗披碱草 Agrimonia pilosa, Galium verum, Elymus gmelinii |
| 低地盐化草甸 Low-land saline meadow | 3 | 叉分蓼、白花马蔺、芨芨草 Polygonum divaricatum, Iris lacteal, Achnatherum splendens |
| 暖性草丛 Warm-temperate tussock | 4 | 阿拉伯黄背草、矮蒿 Themeda triandra, Artemisia lancea |
| 暖性灌草丛 Warm-temperate shrub tussock | 19 | 胡枝子、荆条、绣线菊、金色狗尾草 Lespedeza bicolor, Vitex negundo var. heterophylla, Spiraea salicifolia, Setaria glauca |
新窗口打开|下载CSV
2.2 不同草地类型生物量
2.2.1 地上活体生物量由于不同草地类型的分布地区以及土壤类型不同, 河北省不同草地类型的地上活体生物量差异显著(p < 0.05)。低地盐化草甸的平均地上活体生物量最大, 为285.0 g·m-2, 其次是暖性灌草丛和温性山地草甸, 分别为235.1和203.1 g·m-2, 温性草原的地上活体生物量最低, 为110.6 g·m-2, 是低地盐化草甸的39% (图2)。但单个样地草地地上活体生物量最大值是温性山地草甸, 为366.8 g·m-2。
图2
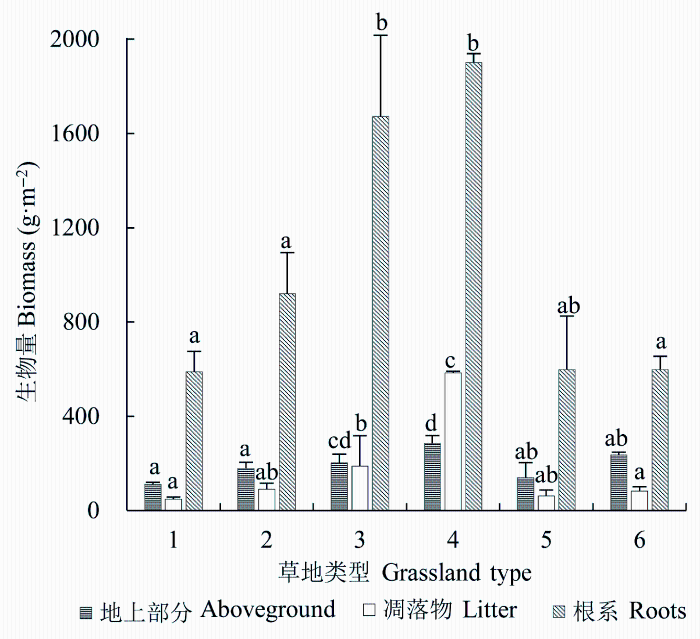 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2不同草地类型地上活体生物量、凋落物生物量与根系生物量(平均值±标准偏差)。1, 温性草原; 2, 温性草甸; 3, 温性山地草甸; 4, 低地盐化草甸; 5, 暖性草丛; 6, 暖性灌草丛。不同的小写字母表示不同草地类型间的生物量差异显著(p < 0.05)。
Fig. 2The aboveground biomass, litter mass and root biomass in different types of grasslands (mean ± SD). 1, temperate steppe; 2, temperate meadow; 3, temperate mountain meadow; 4, low-land saline meadow; 5, warm-temperate tussock; 6, warm- temperate shrub tussock. Different lower-case letters indicate significant differences in biomass among different types of grasslands (p < 0.05).
2.2.2 凋落物生物量
凋落物生物量也是低地盐化草甸最大, 其次是温性山地草甸和温性草甸, 分别为584.0、187.9和91.0 g·m-2。温性草原的凋落物生物量最低, 为48.3 g·m-2, 低地盐化草甸的凋落物生物量约为温性草原的12倍多(图2)。
2.2.3 根系生物量
不同草地类型的根系生物量均大于地上生物量, 与地上生物量相似, 根系生物量最大值同样为低地盐化草甸, 达1901.3 g·m-2, 其次是温性山地草甸和温性草甸, 分别为1671.8和920.1 g·m-2 (图2)。但单个样地的根系生物量最大值出现在温性山地草甸, 为2712.7 g·m-2。
2.3 不同草地类型生物量分配比例
河北省不同草地类型根系生物量与地上生物量各部分的分配比例不同, 其中地上生物量包括地上活体生物量与凋落物生物量两部分。不同草地类型的根系生物量均大于地上活体生物量与凋落物生物量之和, 根系生物量是地上生物量的1.9-4.3倍, 平均根冠比为3.1。6种不同草地类型中根系生物量在总生物量中所占比例差异显著, 温性山地草甸的根系生物量分配比例最大, 达81.0%, 暖性灌草丛的分配比例最小, 为65.3%。温性山地草甸的地上活体生物量的分配比例是最小的, 为9.9%, 暖性灌草丛的分配比例最高, 达到了25.7%, 这与暖性灌草丛中具有生物量较大的灌木植物有关。对凋落物生物量而言, 分配比例最高的是低地盐化草甸, 达21.1% (图3)。图3
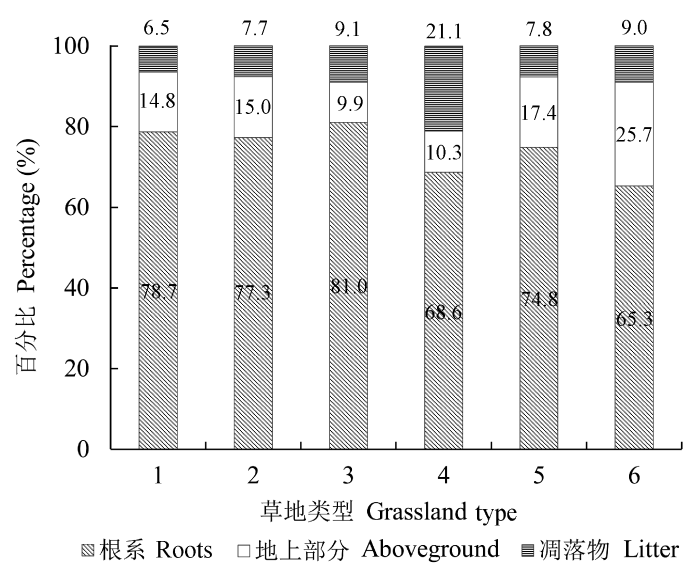 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3不同草地类型地上活体生物量、凋落物生物量与根系生物量各部分分配比例。1, 温性草原; 2, 温性草甸; 3, 温性山地草甸; 4, 低地盐化草甸; 5, 暖性草丛; 6, 暖性灌草丛。
Fig. 3Proportions of aboveground biomass, litter mass and root biomass in different types of grasslands. 1, temperate steppe; 2, temperate meadow; 3, temperate mountain meadow; 4, low- land saline meadow; 5, warm-temperate tussock; 6, warm- temperate shrub tussock.
2.4 不同草地类型植被碳密度
2.4.1 地上活体碳密度河北省不同草地类型地上活体碳密度差异显著(p < 0.05) (图4), 最大值出现在低地盐化草甸, 温性草原地上活体植被碳密度的值最小。不同草地类型地上活体植被碳密度大小顺序为低地盐化草甸(132.7 g C·m-2) >暖性灌草丛(102.5 g C·m-2) >温性山地草甸(87.9 g C·m-2) >温性草甸(84.9 g C·m-2) >暖性草丛(60.0 g C·m-2) >温性草原(47.4 g C·m-2)。低地盐化草甸和暖性灌草丛的地上活体碳密度分别为温性草原的2.8倍和2.2倍, 可见草地植物群落的优势种不同, 导致草地生态系统植物地上活体碳密度值差异显著。
图4
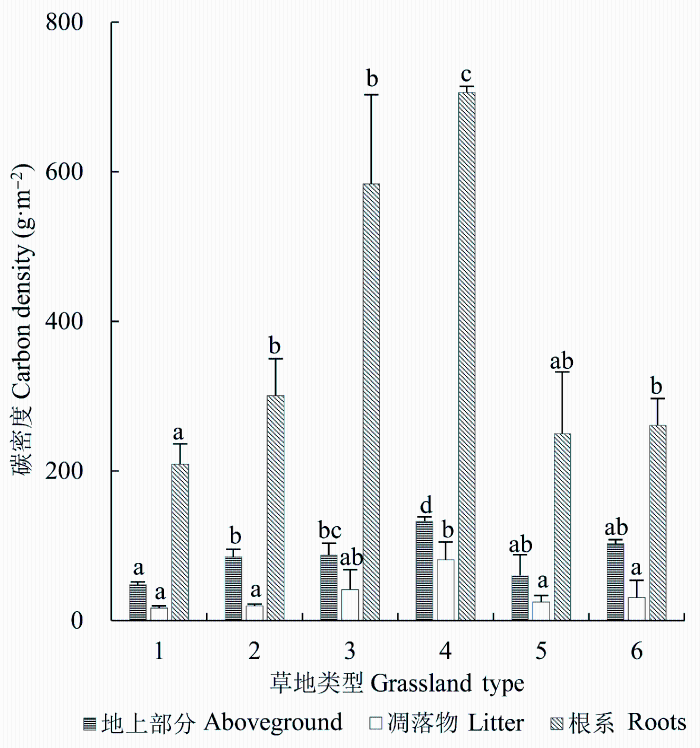 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4不同草地类型地上活体碳密度、凋落物碳密度与根系碳密度(平均值±标准差)。1, 温性草原; 2, 温性草甸; 3, 温性山地草甸; 4, 低地盐化草甸; 5, 暖性草丛; 6, 暖性灌草丛。不同的小写字母表示不同草地类型的生物量之间差异显著(p < 0.05)。
Fig. 4The aboveground carbon density, litter carbon density and root carbon density in different types of grasslands (mean ± SD). 1, temperate steppe; 2, temperate meadow; 3, temperate mountain meadow; 4, low-land saline meadow; 5, warm- temperate tussock; 6, warm-temperate shrub tussock. Different lower-case letters indicate significant differences in carbon density among different types of grasslands (p < 0.05).
2.4.2 凋落物碳密度
低地盐化草甸和温性山地草甸的凋落物生物量最大, 凋落物的碳密度也最大, 两者差异不显著(p > 0.05)(图4), 但低地盐化草甸与其他5种草地类型的凋落物碳密度差异显著(p < 0.05), 其他5种草地类型的凋落物碳密度的差异性均不显著。6种草地类型凋落物平均碳密度大小顺序为低地盐化草甸(81.2 g C·m-2) >温性山地草甸 (41.4 g C·m-2) >暖性灌草丛(30.7 g C·m-2) >暖性草丛(24.8 g C·m-2) >温性草甸(19.4 g C·m-2) >温性草原(16.6 g C·m-2)。
2.4.3 根系碳密度
不同草地类型根系碳密度差异显著(p < 0.05) (图4), 最大值出现在低地盐化草甸和温性山地草甸, 分别为705.9和583.6 g C·m-2, 其次是温性草甸与暖性灌草丛, 分别为300.9和261.1 g C·m-2, 根系碳密度值较小的是暖性草丛和温性草原, 分别是249.7和209.1 g C·m-2, 二者差异不显著。
2.5 不同草地类型植被碳密度分配比例
与草地生物量的分布格局相似, 植物体各部分碳密度在总碳密度中的分配比例不同。根系平均碳密度值远大于地上活体碳密度与凋落物碳密度, 分别是地上活体碳密度与凋落物碳密度的2.5-6.7倍和8.5-15.5倍, 是地上部分总碳密度的2.0-4.5倍。根系碳密度在植物体中分配比例最高的是温性山地草甸, 达81.9%; 温性山地草甸的地上活体碳密度的分配比例是6种草地类型中最低的, 为12.3%。根系碳密度中分配比例最低的是暖性灌草丛, 为66.2%, 暖性灌草丛的地上活体碳密度的分配比例是6种草地类型中最高的, 为26.0%。凋落物碳密度中, 分配比例最高的是低地盐化草甸, 为8.9%; 分配比例最低的是温性草甸, 为4.8% (图5)。图5
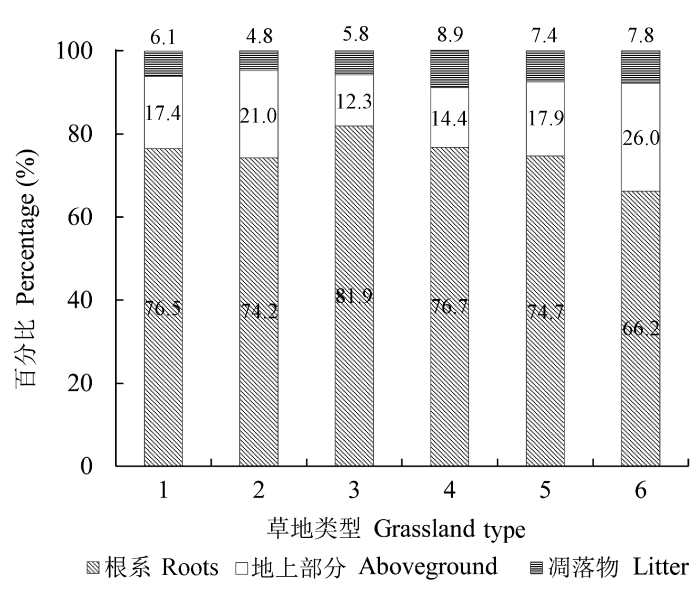 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5不同草地类型地上活体、凋落物与根系各部分碳密度分配比例。1, 温性草原; 2, 温性草甸; 3, 温性山地草甸; 4, 低地盐化草甸; 5, 暖性草丛; 6, 暖性灌草丛。
Fig. 5Proportions of aboveground carbon density, litter carbon density and root carbon density in different types of grasslands. 1, temperate steppe; 2, temperate meadow; 3, temperate mountain meadow; 4, low-land saline meadow; 5, warm- temperate tussock; 6, warm-temperate shrub tussock.
2.6 生物量与环境因子的关系
2.6.1 生物量与海拔的关系河北省草地生态系统植物总生物量和各部分生物量与海拔之间的回归关系分析结果表明, 根系生物量和植物总生物量与海拔之间的关系均达到了极显著水平(p < 0.01)(图6), 且随海拔的变化趋势相似, 生物量随着海拔的升高先减少, 当海拔上升到一定高度时, 生物量减少到最低水平, 之后随着海拔的升高草地生物量又呈现出增加的变化趋势, 根系生物量和总生物量的最低点都出现在海拔750-800 m之间, 分别约为350和600 g·m-2。地上活体生物量和凋落物生物量与海拔之间的关系达到了显著水平(p < 0.05), 随海拔升高先减少后增加左右, (图6), 地上活体生物量的最低值出现在海拔800 m约为120 g·m-2; 凋落物生物量的最低值出现在海拔1000 m左右, 约为50 g·m-2。
图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6草地生态系统地上生物量、凋落物生物量、根系生物量及总生物量与海拔之间的关系。
Fig. 6Relationships of aboveground biomass, litter mass, root biomass, and total biomass with elevation in the grassland ecosystem.
2.6.2 生物量与气候因子的关系
2.6.2.1 生物量与降水量的关系 地上活体生物量、根系生物量与总生物量都是随着年降水量的增加呈现出先增加后降低的变化趋势, 但回归关系均不显著(p > 0.05); 凋落物生物量随着年降水量的增加而增加, 但是回归关系也是不显著(p > 0.05) (图7)。
图7
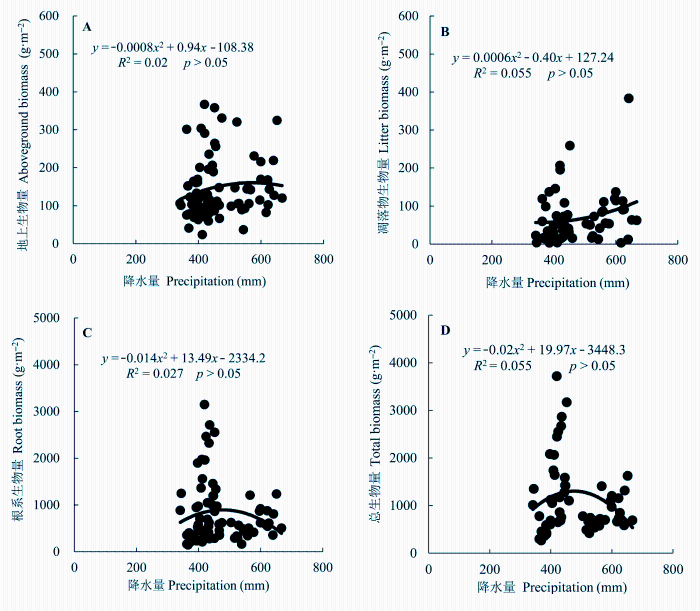 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7草地生态系统地上生物量、凋落物生物量、根系生物量及总生物量与降水量之间的关系。降水量是取样地区的平均年降水量。
Fig. 7Relationships of aboveground biomass, litter mass, root biomass, and total biomass with precipitation in the grassland ecosystem. Here the precipitation is the mean of the annual precipitation of all sampling sites.
2.6.2.2 生物量与积温的关系 在分析草地生物量与温度之间的关系中, 温度采用的是取样地区近50年以来日平均温度10 ℃以上的积温, 回归分析结果表明, 地上活体生物量、凋落物生物量、根系生物量和总生物量都是随着平均积温的增加呈现出先减少后增加的变化趋势, 但地上活体生物量和凋落物生物量与积温之间的回归关系不显著(p > 0.05) (图8); 根系生物量和总生物量与积温之间的关系达到了极显著水平(p < 0.01)。
图8
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8草地生态系统地上生物量、凋落物生物量、根系生物量及总生物量与积温之间的关系。积温采用的是日平均温度大于10 ℃以上的积温。
Fig. 8Relationships of aboveground biomass, litter mass, root biomass, and total biomass with accumulative temperature in the grassland ecosystem. The accumulative temperature is the mean of the accumulative temperatures > 10 °C of all sampling sites.
3 讨论
3.1 生物量与碳密度的空间分布
对于大尺度范围内草地生物量的估算, ****多采用遥感的方法进行。基于我国第一次草地资源清查资料和遥感数据估算我国北方温带荒漠草原、温带典型草原和温带草甸草原的地上生物量分别为45.6、89.0和146.5 g·m-2 (Ni, 2004a); 方精云等(1996)基于实测和遥感相结合的方法估算我国北方以上3种主要草地类型的地上生物量分别为34.2、122.4和135.1 g·m-2。导致两者估算结果不同的原因主要有两个方面, 一方面是由于不同的研究者采用的数据来源不同, 导致结果差异较大; 另一方面, 不同的****对世界范围内的植被分类不同, 也可能引起较大的差异, 所以在比较不同草地类型植被生物量变化时需要采用统一的研究方法。本研究采用统一的草地生态系统调查规范, 获得了第一手的实测草地生物量与碳密度的数据, 所以对整个河北省草地生态系统植被总生物量和碳密度的估算准确性很高。本研究得出河北省不同草地类型的地上生物量差异显著, 其中温带典型草原和温带草甸草原的地上生物量分别为110.6和178.9 g·m-2, 高于遥感估算的结果。河北省6种不同草地类型植被总生物量介于747.6-2770.0 g·m-2之间, 其平均值为1414.6 g·m-2, 低于20世纪80年代第一次草地普查数据全国相同草地类型的平均值1477.0 g·m-2 (辛晓平等, 2009)。但是, 经过30年的草地利用, 不同草地类型植被生物量变化趋势不同, 在20世纪80年代的草地调查中, 全国温性草原、温性草甸草原和暖性草丛的平均生物量分别为872.0、1579.0和1643.0 g·m-2, 均高于本研究的调查结果; 而温性山地草甸、低地盐化草甸的生物量分别为1617.0和1674.0 g·m-2, 远低于本研究的2062.8和2770.2 g·m-2。不同草地类型的生物量变化趋势不同的主要原因有以下几个: 一, 经过30多年的草地利用, 河北省不同草地类型的物种组成发生了变化, 部分草地已经发生沙化、盐碱化, 导致草地生物量发生了变化; 二, 近30年间, 全球气候变化、降水分布等自然环境的改变, 形成了不同草地类型的小生物气候环境, 导致植物物种产生了与环境变化相适应的分布, 最终导致不同草地类型的生物量产生了不同的变化; 三, 人类对不同草地类型的利用强度和干扰程度不同, 以及不同草地类型对于人类活动承受能力的差异最终导致了生物量的差异。在草地生态系统中, 地下生物量显著高于地上生物量, 本研究结果中温性典型草原和温性草甸草原的地下生物量分别为588.7和920.1 g·m-2, 远低于Ni (2004b)通过遥感和根冠比估算的结果1777.7和2415.7 g·m-2。Piao等(2007)根据文献得到的草原植物根冠比为5.3-10.1, 平均值为7.7; Fan等(2008)报道中国北方草地的根冠比范围在2.5-52.3, 平均值为24.6; Yang等(2010)对中国北方草地进行的大量调查分析表明, 不同样地之间的根冠比变化很大, 为0.4-14.3, 中值为5.7; 而本研究结果表明河北省不同草地植被类型根冠比为1.9-4.3, 平均值为3.1, 低于以往的研究结果。出现这种较大变化的主要原因有两个: 一, 不同****采用的数据来源不同, 常用的数据来源基于实测数据加上遥感估算, 而实地调查的结果经过尺度扩展放大到整个区域后, 必然造成误差增大, 导致遥感估测的数值通常偏高; 二, 本研究实地调查中地下生物量只考虑了活根的生物量, 而且地上生物量包括了半分解层和凋落物的生物量, 所以导致根冠比偏小。比较而言, 本研究通过实地调查采用的是收获法, 获得大量的第一手实测数据, 可信度更高。
河北省不同草地植被生物量碳密度差异显著, 为273.0-919.8 g C·m-2, 平均值506.6 g C·m-2, 高于中国草地生物量平均碳密度320.5 g C·m-2 (Fang et al., 2010)。其中低地盐化草甸的碳密度最高, 为919.8 g C·m-2, 其次为温性山地草甸、温性草甸和暖性草草丛, 本研究中不同草地类型植被碳密度平均值的大小与全国和全球草地碳密度的分布规律基本一致(Ni, 2001; Lu & Sun, 2004)。青藏高原的研究表明具有灌木的草地植物地上部分碳密度最高(常天军等, 2007), 而本研究中暖性灌草丛是指矮小灌木郁闭度小于40%的草地植被群落, 因而灌木数量在草地中很少, 对暖性灌草丛地上生物量和碳密度影响较小。综合分析, 本区域影响不同草地类型植被生物量和碳密度差异的主要因素如下: 一是受不同植被类型中优势植物种类的控制, 低地盐化草甸植物种类丰富, 而且生物量较大的薹草类和马蔺等占很大比例; 二是低地盐化草甸中的委陵菜、薹草类植物对于草地利用中践踏的承受能力更强, 加上围封禁牧更加有利于草甸草原植物群落中地榆、薹草类植物的生物量大幅度增加, 从而导致生物量碳密度也增加; 三是由于不同草地类型土壤质地的差异导致植物生物量差异显著, 草甸类草原土壤有机质含量高, 促进植物的生长, 直接影响生物量和碳密度的增加。
3.2 影响草地生物量的环境因素
河北省不同草地类型的植物群落物种组成不同, 这不仅是植物种对环境条件长期适应的结果, 更是反映了草地的生物气候类型(杜国祯等, 2003)。草地生物量表示草地生态系统的植被生产力, 所以研究草地生物量对于了解草地生态系统的结构与功能有重要的意义。诸多研究表明草地生物量主要受降水量与温度的影响(Xiao et al., 1995; 马文红等, 2008; Fang et al., 2010), 同时也受到土壤、海拔、热量等气象因子的影响(白永飞等, 2002; 韩彬等, 2006)。本研究区域海拔梯度较大, 变化范围为250-1700 m, 不同草地类型地上活体生物量、凋落物生物量、根系生物量以及总生物量与海拔之间的关系均达到显著水平(p < 0.05), 且变化趋势基本一致, 都是随着海拔的升高先降低, 当海拔上升到一定高度时, 生物量降低到最低水平, 之后随着海拔的升高草地生物量又呈现出增加的变化趋势。这种变化趋势主要是由于海拔的变化引起了温度、蒸发量及土壤等环境因子的改变, 造成了不同草地类型生物量的差异, 进而造成生物量碳密度的差异。本研究中草地生物量随着平均积温(> 10 ℃)的增加呈现出先减少后增加的变化趋势, 根系生物量和总生物量与积温之间的关系达到了极显著水平(p < 0.01), 尽管地上活体生物量和凋落物生物量与积温之间的回归关系不显著(p > 0.05)。已有研究发现气候是影响草地生物量的重要因素, 尤其是地上生物量的空间变化主要受降水量控制, 在对内蒙古草原的研究中发现, 降水量是影响生物量分布差异的最主要的环境因素, 地上、地下生物量随着年降水量的增加显著增加(马文红等, 2008); 黄土高原的研究也表明草地生物量随着降水量增加而增加(邓蕾和上官周平, 2012)。但是本研究中河北省不同草地类型的地上活体生物量、根系生物量与总生物量随着年降水量的增加而增加的变化趋势不显著(p > 0.05)。综合分析, 导致降水量对生物量影响不显著的原因可能有两点: 第一, 河北省属于草地与森林的过渡地带, 从北部高原到南部山地的海拔变化较大, 降水量变化幅度较小, 为340-667 mm。降水量较大的区域, 主要分布有暖性草丛和暖性灌草丛, 而草地生物量较大的低地盐化草甸、温性草甸草原和温性山地草甸大多分布在降水量为400-600 mm的地区。第二, 本研究中降水量在500-667 mm的区域多为石质山区, 土层很薄, 地表30 cm以下基本为石砾, 与土层较厚的草甸草原相比, 植物根系生物量较小。可见, 在山地草地中, 积温和海拔对植物生物量积累具有显著影响。分析环境因子与草地生物量的关系, 应该综合考虑气候、土壤、地理等因素。致谢 感谢河北农业大学徐敏云、白志英在野外工作中给予的帮助。

参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
(
[本文引用: 1]
(
[本文引用: 1]
(
[本文引用: 1]
(
[本文引用: 1]
DOI:10.1088/1009-1963/11/5/313URL [本文引用: 1]

用样带法调查了草原群落植物功能群组成沿水热梯度的变化。结果表明,随着海拔高度的降低,降水量的减少,热量和干燥度的增加,群落植物生活型功能群组成中,多年生杂类草的相对多度逐渐减少,而多年生丛生禾草的相对多度逐渐增加,多年生丛生禾草对多年生杂类草具有强烈的生态替代作用。在植物生态类群功能群组成中,中旱生植物的作用逐渐减弱,而旱生植物的作用显著增强。草地的利用方式对群落组成具有重要的影响,永久样地和割草场中不耐牧种的相对生物量显著地高于放牧场,而放牧场中耐牧种的相对生物量则显著地高于永久样地和割草场。受草地利用强度的影响,土壤有机碳及全氮含量对上述植物功能群相对多度的影响呈非线性。在样带梯度上,大气降水递减和利用强度增加所引起的群落结构与组成的简单化,可能是导致草原群落生产力和稳定性降低的主要因素。
DOI:10.1088/1009-1963/11/5/313URL [本文引用: 1]
用样带法调查了草原群落植物功能群组成沿水热梯度的变化。结果表明,随着海拔高度的降低,降水量的减少,热量和干燥度的增加,群落植物生活型功能群组成中,多年生杂类草的相对多度逐渐减少,而多年生丛生禾草的相对多度逐渐增加,多年生丛生禾草对多年生杂类草具有强烈的生态替代作用。在植物生态类群功能群组成中,中旱生植物的作用逐渐减弱,而旱生植物的作用显著增强。草地的利用方式对群落组成具有重要的影响,永久样地和割草场中不耐牧种的相对生物量显著地高于放牧场,而放牧场中耐牧种的相对生物量则显著地高于永久样地和割草场。受草地利用强度的影响,土壤有机碳及全氮含量对上述植物功能群相对多度的影响呈非线性。在样带梯度上,大气降水递减和利用强度增加所引起的群落结构与组成的简单化,可能是导致草原群落生产力和稳定性降低的主要因素。
DOI:10.3969/j.issn.1008-8873.2007.05.009URL [本文引用: 1]
采用实地调查与查阅文献相结合的方法估算藏北高寒草地植被碳密度和碳贮量。结果表明:1)藏北高寒草地总面积约39.059×10^6hm^2,植被地上平均碳密度12.158±4.7g·m^-2,植被地下平均碳密度84.458±20.38g·m^-2,植被地上部碳贮量5.171±0.95Tg,植被地下部碳贮量25.223±2.96Tg,植被总碳贮量为30.394±3.91Tg;21不同草地组间植被碳密度和碳贮量差异显著。其中不同草地组间植被碳密度以丛生禾草组碳密度值最低,地上和地下碳密度分别为6.13±1.51g·m^-2和26.04±5.8g·m^-2,具灌木的半灌木组碳密度最高,地上和地下碳密度分别为31±3.4g·m^2和244.59±6.9g·m^-2;而不同草地组间,草地植被碳贮量以小莎草组最大,植被地上和地下碳贮量分别为2.24±0.32Tg和9.52±0.89Tg,半灌木组碳贮量最小,其地上和地下碳贮量分别为0.0124±0.002Tg和0.0981±0.002Tg。3)藏北高寒草地分布各县(区)碳密度和碳贮量的分布也存在显著差异。从碳密度来看,革吉县、札达县、噶尔县和措勤县碳密度较高,植被平均碳密度分别相当于藏北平均植被碳密度的1.76,1.47,1.11和1.06倍,从碳贮量来看,碳贮量集中分布于双湖特别区、札达、尼玛、日土、革吉和改则6县(区),六县(区)草地植被碳贮量为25.2±2.31Tg,占藏北总植被碳量的82.89%。
DOI:10.3969/j.issn.1008-8873.2007.05.009URL [本文引用: 1]
采用实地调查与查阅文献相结合的方法估算藏北高寒草地植被碳密度和碳贮量。结果表明:1)藏北高寒草地总面积约39.059×10^6hm^2,植被地上平均碳密度12.158±4.7g·m^-2,植被地下平均碳密度84.458±20.38g·m^-2,植被地上部碳贮量5.171±0.95Tg,植被地下部碳贮量25.223±2.96Tg,植被总碳贮量为30.394±3.91Tg;21不同草地组间植被碳密度和碳贮量差异显著。其中不同草地组间植被碳密度以丛生禾草组碳密度值最低,地上和地下碳密度分别为6.13±1.51g·m^-2和26.04±5.8g·m^-2,具灌木的半灌木组碳密度最高,地上和地下碳密度分别为31±3.4g·m^2和244.59±6.9g·m^-2;而不同草地组间,草地植被碳贮量以小莎草组最大,植被地上和地下碳贮量分别为2.24±0.32Tg和9.52±0.89Tg,半灌木组碳贮量最小,其地上和地下碳贮量分别为0.0124±0.002Tg和0.0981±0.002Tg。3)藏北高寒草地分布各县(区)碳密度和碳贮量的分布也存在显著差异。从碳密度来看,革吉县、札达县、噶尔县和措勤县碳密度较高,植被平均碳密度分别相当于藏北平均植被碳密度的1.76,1.47,1.11和1.06倍,从碳贮量来看,碳贮量集中分布于双湖特别区、札达、尼玛、日土、革吉和改则6县(区),六县(区)草地植被碳贮量为25.2±2.31Tg,占藏北总植被碳量的82.89%。
[本文引用: 1]
[本文引用: 1]
DOI:10.11733/j.issn.1007-0435.2012.05.006URL [本文引用: 1]
草地生态系统是全球陆地生态系统的重要组成部分,在全球碳循环中扮演着重要角色。以陕西省天然草地为研究对象,基于57个调查样地的地上、地下和凋落物生物量资料,分析不同植被类型草地生物量的空间分布特征及其影响因素,并揭示其之间的相互关系,为我国草地生态系统碳汇研究提供基础数据。结果表明:7种草地类型(暖性草丛类、暖性灌草丛类、温性山地草甸类、温性草甸草原、温性草原、温性荒漠草原类和低地盐化草甸类)的各部分生物量存在显著差异(P0.05),其地上生物量分别为:238.3,293.6,157.0,350.5,156.9,99.9和144.6g·m-2,地下生物量分别为:670.2,560.5,726.3,1072.5,719.5,1156.7和775.0g.m-2,凋落物生物量分别为:332.2,294.0,328.5,271.4,107.4,97.2和155.7g·m-2;但是其群落总生物量差异不显著,分别为:1353.9,1240.8,1405.3,1501.0,983.8,1148.1和1075.3g·m-2。地下生物量均大于地上生物量和凋落物生物量,其分配比例介于38.8%~77.4%之间;地上生物量和凋落物生物量随海拔高度增加而极显著减少(P0.01);地上生物量随生长期降雨量增加而极显著增加(P0.01),随年均温增加而减少(P0.05);地下生物量对降雨和温度的响应均不显著。因此,降水是陕西省天然草地生态系统地上生物量最主要的限制因子,且主要受生长期水分供给的制约,温度尚不是主要限制因子。
DOI:10.11733/j.issn.1007-0435.2012.05.006URL [本文引用: 1]
草地生态系统是全球陆地生态系统的重要组成部分,在全球碳循环中扮演着重要角色。以陕西省天然草地为研究对象,基于57个调查样地的地上、地下和凋落物生物量资料,分析不同植被类型草地生物量的空间分布特征及其影响因素,并揭示其之间的相互关系,为我国草地生态系统碳汇研究提供基础数据。结果表明:7种草地类型(暖性草丛类、暖性灌草丛类、温性山地草甸类、温性草甸草原、温性草原、温性荒漠草原类和低地盐化草甸类)的各部分生物量存在显著差异(P0.05),其地上生物量分别为:238.3,293.6,157.0,350.5,156.9,99.9和144.6g·m-2,地下生物量分别为:670.2,560.5,726.3,1072.5,719.5,1156.7和775.0g.m-2,凋落物生物量分别为:332.2,294.0,328.5,271.4,107.4,97.2和155.7g·m-2;但是其群落总生物量差异不显著,分别为:1353.9,1240.8,1405.3,1501.0,983.8,1148.1和1075.3g·m-2。地下生物量均大于地上生物量和凋落物生物量,其分配比例介于38.8%~77.4%之间;地上生物量和凋落物生物量随海拔高度增加而极显著减少(P0.01);地上生物量随生长期降雨量增加而极显著增加(P0.01),随年均温增加而减少(P0.05);地下生物量对降雨和温度的响应均不显著。因此,降水是陕西省天然草地生态系统地上生物量最主要的限制因子,且主要受生长期水分供给的制约,温度尚不是主要限制因子。
[本文引用: 1]
[本文引用: 1]
DOI:10.17521/cjpe.2003.0019URL [本文引用: 1]
以物种组成较为复杂的青藏高原东部地区典型高寒草甸植物群落为背景,从不同的时间和空间尺度水平研究了植物群落中物种丰富度与生产力(地上部生物量)关系的基本模式.结果表明:1)总体而言,高寒草甸植物群落中物种丰富度与生产力的关系呈对数线性增加关系,这实际上是空间和时间尺度放大时在时间和空间尺度的相互作用下高寒草甸植物群落中物种丰富度与生产力关系的一种总体模式;2)随着研究的时间和空间尺度的改变,物种丰富度与生产力的关系会发生一些相应的变化,但空间尺度比时间尺度对物种丰富度与生产力的关系造成的影响更为显著,这可能与高寒草甸植物群落的生长季短有关.上述结果揭示:时间和空间尺度的不同可能是造成许多试验研究得不到一个统一的物种丰富度-生产力关系模式的主要原因.
DOI:10.17521/cjpe.2003.0019URL [本文引用: 1]
以物种组成较为复杂的青藏高原东部地区典型高寒草甸植物群落为背景,从不同的时间和空间尺度水平研究了植物群落中物种丰富度与生产力(地上部生物量)关系的基本模式.结果表明:1)总体而言,高寒草甸植物群落中物种丰富度与生产力的关系呈对数线性增加关系,这实际上是空间和时间尺度放大时在时间和空间尺度的相互作用下高寒草甸植物群落中物种丰富度与生产力关系的一种总体模式;2)随着研究的时间和空间尺度的改变,物种丰富度与生产力的关系会发生一些相应的变化,但空间尺度比时间尺度对物种丰富度与生产力的关系造成的影响更为显著,这可能与高寒草甸植物群落的生长季短有关.上述结果揭示:时间和空间尺度的不同可能是造成许多试验研究得不到一个统一的物种丰富度-生产力关系模式的主要原因.
[本文引用: 1]
DOI:10.1007/s10584-007-9316-6URL [本文引用: 2]
Above- and below-ground biomass values for 17 types of grassland communities in China as classified by the Chinese Grasslands Resources Survey were obtained from systematic replicated sampling at 78 sites and from published records from 146 sites. Most of the systematic samples were along a 5,000-km-long transect from Hailar, Inner Mongolia (49°15′N, 119°15′E), to Pulan, Tibet (30°15′N, 81°10′E). Above-ground biomass was separated into stem, leaf, flower and fruit, standing dead matter, and litter. Below-ground biomass was measured in 10-cm soil layers to a depth of 3002cm for herbs and to 5002cm for woody plants. Grassland type mean total biomass carbon densities ranged from 2.40002kg m 612 for swamp to 0.14902kg m 612 for alpine desert grasslands. Ratios of below- to above-ground carbon density varied widely from 0.99 for tropical tussock grassland to 52.28 for alpine meadow. Most below-ground biomass was in the 0–1002cm soil depth layer and there were large differences between grassland types in the proportions of living and dead matter and stem and leaf. Differences between grassland types in the amount and allocation of biomass showed patterns related to environments, especially aridity gradients. Comparisons of our estimates with other studies indicated that above-ground biomass, particularly forage-yield biomass, is a poor predictor of total vegetation carbon density. Our estimate for total carbon storage in the biomass of the grasslands of China was 3.3202Pg C, with 56.4% contained in the grasslands of the Tibet-Qinghai plateau and 17.9% in the northern temperate grasslands. The need for further standardized and systematic measurements of vegetation biomass to validate global carbon cycles is emphasised.
[本文引用: 2]
[本文引用: 2]
DOI:10.1007/s11427-010-4029-xURLPMID:20697865 [本文引用: 3]
The knowledge of carbon(C) stock and its dynamics is crucial for understanding the role of grassland ecosystems in China's terrestrial C cycle.To date,a comprehensive assessment on C balance in China's grasslands is still lacking.By reviewing published literature,this study aims to evaluate ecosystem C stocks(both vegetation biomass and soil organic C) and their changes in China's grasslands.Our results are summarized as follows:(1) biomass C density(C stock per area) of China's grasslands differed greatly among previous studies,ranging from 215.8 to 348.1 g C m-2 with an average of 300.2 g C m-2.Likewise,soil C density also varied greatly between 8.5 and 15.1 kg C m-2.In total,ecosystem C stock in China's grasslands was estimated at 29.1 Pg C.(2) Both the magnitude and direction of ecosystem C changes in China's grasslands differed greatly among previous studies.According to recent reports,neither biomass nor soil C stock in China's grasslands showed a significant change during the past 20 years,indicating that grassland ecosystems are C neutral.(3) Spatial patterns and temporal dynamics of grassland biomass were closely correlated with precipitation,while changes in soil C stocks exhibited close associations with soil moisture and soil texture.Human activities,such as livestock grazing and fencing could also affect ecosystem C dynamics in China's grasslands.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLMagsci [本文引用: 1]
草地退化是在过牧、开垦等人为活动及不利自然因素影响下草地生态系统逆行演替的一种过程,是土地荒漠化的主要形式之一。本文在讨论了草地退化指标及其等级划分的基础上,编制了中国北方退化草地分布图。据此图计算,北方草地退化面积达137.77万km#+2,占该草地总面积的50.24%,近15年来以每年1.9%的速度在扩大退化。为此提出了防治对策及草地畜牧业可持续发展战略。
URLMagsci [本文引用: 1]
草地退化是在过牧、开垦等人为活动及不利自然因素影响下草地生态系统逆行演替的一种过程,是土地荒漠化的主要形式之一。本文在讨论了草地退化指标及其等级划分的基础上,编制了中国北方退化草地分布图。据此图计算,北方草地退化面积达137.77万km#+2,占该草地总面积的50.24%,近15年来以每年1.9%的速度在扩大退化。为此提出了防治对策及草地畜牧业可持续发展战略。
[本文引用: 1]
[本文引用: 1]
DOI:10.17521/cjpe.2004.0093URL [本文引用: 1]
Terrestrial ecosystems are a large carbon density and play an important role in the global carbon budget and mitigating global warming by carbon sequestration. To encourage additional carbon sequestration and storage in global vegetation and soils, we need to be clear about the distribution patterns of carbon density and the factors that influence these patterns. Therefore, the characteristics of the distribution patterns of carbon density in terrestrial ecosystems of the world and China are reviewed in this paper. At the global scale, the distribution of carbon in vegetation corresponds with the spatial pattern of biomass and generally decreases from low-latitude to high-latitude with the exception of boreal forests. In contrast, soil organic pools of carbon increase along the same gradient. The largest stores of carbon in biomass are in tropical and boreal forests, and the largest stores of soil carbon are in the high latitude ecosystems (boreal forest and tundra). Total carbon density of both vegetation and soils are highest in tropical and boreal forests. In the tropics more carbon is stored in vegetation than in soils, while in the boreal region far more carbon is stored in the soils. At the regional scale, these patterns might vary due to differences in climate, topography and human influence among regions. Several major factors, including climatic conditions, soil nutrients, biodiversity, climate and atmospheric CO 2 changes, land use and cover changes, all contribute to the storage and maintenance of carbon. For example, carbon density will be high in regions where temperature and precipitation are favorable for abundant plant growth. The enhanced carbon sequestered in response to elevated levels of CO 2 or nitrogen, or their combination, is less in species-poor than in species-rich regions. In a word, they can raise the carbon density in terrestrial ecosystems by directly or indirectly accelerating net primary production, or constraining respiration and decomposition. However, in spite of its great significance for explaining the present distribution patterns of carbon pools and estimating future changes, the mechanisms by which these processes occur are not fully understood. It is critical that we strengthen our research effort in this area of study. Although a great deal of effort has been put into the carbon density researches, there still remains much uncertainty regarding data collection, mechanistic explanations, and model construction in the relevant studies. In the future, we should establish a standardized and unified system to estimate carbon density, produce more accurate and useful models, and perform integrated studies at multi-scales and multi-resolutions levels.
DOI:10.1007/s11427-010-4020-6URLPMID:20697873 [本文引用: 1]
Abstract Grassland covers approximately one-third of the area of China and plays an important role in the global terrestrial carbon (C) cycle. However, little is known about biomass C stocks and dynamics in these grasslands. During 2001-2005, we conducted five consecutive field sampling campaigns to investigate above-and below-ground biomass for northern China's grasslands. Using measurements obtained from 341 sampling sites, together with a NDVI (normalized difference vegetation index) time series dataset over 1982-2006, we examined changes in biomass C stock during the past 25 years. Our results showed that biomass C stock in northern China's grasslands was estimated at 557.5 Tg C (1 Tg=10(12) g), with a mean density of 39.5 g C m(-2) for above-ground biomass and 244.6 g C m(-2) for below-ground biomass. An increasing rate of 0.2 Tg C yr(-1) has been observed over the past 25 years, but grassland biomass has not experienced a significant change since the late 1980s. Seasonal rainfall (January-July) was the dominant factor driving temporal dynamics in biomass C stock; however, the responses of grassland biomass to climate variables differed among various grassland types. Biomass in arid grasslands (i.e., desert steppe and typical steppe) was significantly associated with precipitation, while biomass in humid grasslands (i.e., alpine meadow) was positively correlated with mean January-July temperatures. These results suggest that different grassland ecosystems in China may show diverse responses to future climate changes.
DOI:10.3724/SP.J.1005.2008.01083URL [本文引用: 2]
利用实际观测的113个地面数据,估算了内蒙古温带草地地上、地下生物量的大小,揭示了其空间分布和地下生物量的垂直分布规律,并探讨了不同环境因素对地上、地下生物量的调控作用.主要结果如下:(1)3种草地类型(荒漠草原、典型草原和草甸草原)的生物量存在显著差异;其地上生物量分别为56.6,133.4和196.7g/m2,地下生物量分别为301.0,688.9和1385.2g/m2;(2)地上生物量和地下生物量均呈现自西南向东北增加的空间分布特征;3种草地具有相似的地下生物量垂直分布特征;总体上,温带草地表层(0~10cm)地下生物量约占总地下生物量的一半;(3)降水是导致内蒙古温带草地生物量空间变异的主要因子.地下生物量的垂直分布与降水关系密切,而受土壤质地和草地类型的影响较弱.
DOI:10.3724/SP.J.1005.2008.01083URL [本文引用: 2]
利用实际观测的113个地面数据,估算了内蒙古温带草地地上、地下生物量的大小,揭示了其空间分布和地下生物量的垂直分布规律,并探讨了不同环境因素对地上、地下生物量的调控作用.主要结果如下:(1)3种草地类型(荒漠草原、典型草原和草甸草原)的生物量存在显著差异;其地上生物量分别为56.6,133.4和196.7g/m2,地下生物量分别为301.0,688.9和1385.2g/m2;(2)地上生物量和地下生物量均呈现自西南向东北增加的空间分布特征;3种草地具有相似的地下生物量垂直分布特征;总体上,温带草地表层(0~10cm)地下生物量约占总地下生物量的一半;(3)降水是导致内蒙古温带草地生物量空间变异的主要因子.地下生物量的垂直分布与降水关系密切,而受土壤质地和草地类型的影响较弱.
DOI:10.1023/A:1010728609701URL [本文引用: 2]
The carbon storage of terrestrial ecosystems in China was estimated using acommon carbon density method for vegetation and soils relating to thevegetation types. Usingmedian density estimates, carbon storage of 35.23 Gt (1 Gt = 10 15 g) in biomass and119.76 Gt in soils with total of 154.99 Gt were calculated based on thebaseline distribution of37 vegetation types. Total carbon storage of the median estimates at differentspatial resolutionswas 153.43, 158.08 and 158.54 Gt, respectively, for the fine (10′),median (20′) and coarse (30′)latitude × longitude grids. There were differences of 611.56, +3.09and +3.55 Gt carbon storagebetween baseline vegetation and those at different spatial resolutions. Changein mappingresolution would change area estimates and hence carbon storage estimates. Thefiner the spatialresolution in mapping vegetation, the closer the carbon storage to thebaseline estimation. Carbonstorage in vegetation and soils for baseline vegetation is quite similar tothat of biomes predictedby BIOME3 for the present climate and CO 2 concentration of 340ppmv. Climate changealone as well as climate change with elevated CO 2 concentrationwill produce an increasein carbon stored by vegetation and soils, especially a larger increase in thesoils. Total mediancarbon storage of terrestrial ecosystems in China will increase by 5.09 Gt and15.91 Gt for theclimate scenario at CO 2 concentration of 340 ppmv and 500 ppmv,respectively. This ismainly due to changes in vegetation areas and the effects of changes inclimate and CO 2 concentration.
DOI:10.1006/jare.2001.0902URL [本文引用: 1]
Carbon storage in grasslands of China was estimated by the carbon density method and based on a nationwide grassland resource survey finished by 1991. The grasslands in China were classified into 18 types, which are distributed mostly in the temperate region and on the Tibetan Plateau, and scattered in the warm-temperate and tropical regions. Based on the median estimate, vegetation, soil and total carbon storage of grasslands in China were 3·06, 41·03 and 44·09 Pg C, respectively. Vegetation had low carbon storage and most carbon was stored in soils. Of the four types of regions that have grasslands, alpine region (54·5%) and temperate region (31·6%) hold more than 85% of the total grassland carbon (in both vegetation and soils) in China. Considering specific types within these two regions, three grassland types, alpine meadow (25·6%), alpine steppe (14·5%) and temperate steppe (11%) constituted more than half of all carbon stored in China's grasslands. In general and regardless of regional vegetation types, steppes (38·6%) and meadows (38·2%) made up more than 2/3 of total grassland carbon. The carbon storage in alpine grasslands may have a significant and long-lived effect on global C cycles. This study estimated more carbon storage in vegetation and less in soils than previous studies. The differences of grassland carbon between this study and two previous studies were due probably to four reasons, i.e. different estimation methods, different classification systems of grasslands, different areas of grasslands, and different carbon densities. China's grasslands cover only 6–8% of total world grassland area and have 9–16% of total carbon in the world grasslands. They make a big contribution to the world carbon storage and may have significant effects on carbon cycles, both in global and in arid lands.
DOI:10.1007/s10584-004-0070-8URL [本文引用: 2]
Forage yield-based carbon storage in 18 grasslands of China was estimated according to the detailed investigation of grassland area and forage yield (standing crop), which were derived from a 10-year national grassland survey. The total forage yield carbon in Chinese grasslands is 134.09 Tg C for ca. 299 10 6 ha of grassland area and 1232 kg/ha of mean forage yield. The carbon storage is different depending on grassland types and climatic regions. Meadow, steppe and tussock occupy 93.3% (125.14 Tg C), and desert and swamp only accounts for 6.7% (8.95 Tg C) of total forage yield carbon. Forage yield carbon is stored largely in temperate (38.4%, 51.54 Tg C) and alpine regions (30.4%, 40.78 Tg C), and to less extent in tropical regions (22.1%, 29.66 Tg C). These three regions take 91% of the forage yield carbon in grasslands of China. The warm-temperate region accounts for only 9% (12.1 Tg C) of forage yields carbon. The forage yield-based carbon in grasslands of China is more accurate than the site biomass-based carbon estimate and the carbon density-based estimate. Although, forage yield carbon storage is small compared with the total carbon storage in China, carbon budgets of grasslands are often a dominant component in many regions and provide an important management opportunity to enhance terrestrial carbon sinks in vast areas of China.
DOI:10.1023/B:VEGE.0000049097.85960.10URL [本文引用: 1]
Data on field biomass measurements in temperate grasslands of northern China (141 samples from 74 sites) were obtained from 23 Chinese journals, reports and books. Net primary productivity (NPP) of grasslands was estimated using three algorithms (peak live biomass, peak standing crop and maximum minus minimum live biomass), respectively, based on availability of biomass data in sites. 135 samples which have aboveground biomass (AGB) measurements, have peak AGB ranges from 20 to 2021 g m –2 (mean = 325.3) and the aboveground NPP (ANPP) ranges from 15 to 1647.1 g m –2 per year (mean = 295.7). 72 samples which have belowground biomass (BGB) measurements, have peak BGB ranges from 226.5 to 12827.5 g m –2 (mean = 3116) and the belowground NPP (BNPP) ranges from 15.8 to 12827.5 g m –2 per year (mean = 2425.6). In total 66 samples have the total NPP (TNPP), ranging from 55.3 to 13347.8 g m –2 per year (mean = 2980.3). Mean peak biomass and NPP varied from different geographical sampling locations, but they had a general rough regularity in ten grasslands. Meadow, mountain and alpine grasslands had high biomass and NPP (sometimes including saline grassland). Forested steppe, saline grassland and desert had median values. Meadowed and typical steppes had low biomass and NPP (sometimes including desert). The lowest biomass and NPP occurred in deserted steppe and stepped desert. Grassland ANPP has significant positive relationships with annual and summer precipitation as well as summer temperature (all p<0.01). However, grassland BNPP and TNPP have more significant negative relationships with summer temperature (p<0.01) than with annual temperature (p<0.05). The analysis of climate – productivity correlations implied that aboveground productivity is more controlled by rainfall, whereas belowground and total productivity is more influenced by temperature in the temperate grasslands of northern China. The present study might underestimate grassland NPP in northern China due to limitation of biomass measurements. Data on relative long-term aboveground and belowground biomass dynamics, as well as data of standing dead matter, litterfall, decomposition and turnover, are required if grassland NPP is to be more accurately estimated and the role of temperate grasslands in the regional to global carbon cycles is to be fully appreciated.
DOI:10.1029/2005GB002634URL [本文引用: 2]
[1] Terrestrial ecosystems in the northern latitudes are significant carbon sinks for atmospheric CO2; however, few studies come from grassland ecosystems. Using national grassland resource inventory data, NDVI (normalized difference vegetation index) time series data set, and a satellite-based statistical model, this study identifies changes in the size and distribution of aboveground biomass carbon (C) stocks for China's grasslands between 1982 and 1999. Biomass C stocks averaged 145.4 Tg C for the study period for a total area of 334.1 0103 104 km2, and have increased by 17.7 Tg C (1 Tg = 1012 g) from 136.3 Tg C in the early 1980s (average of 19820900091984) to 154.0 Tg C in the late 1990s (average of 19970900091999), with an annual increase of 0.7%. This suggests that the aboveground biomass of China's grasslands may have functioned as the C sinks in the past 2 decades. Assuming a constant ratio of aboveground to belowground biomass for each grassland type, we also estimated belowground biomass C and its change over time for each grassland type, generating an average estimate of 1051.1 Tg C for the total (aboveground and belowground) biomass C and an annual increase of 126.67 Tg C for China's grasslands over the 18 years. However, the accuracy of these estimates has limitations due primarily to uncertainties in estimates of belowground C, biomass inventories, and satellite time series data sets.
DOI:10.1046/j.1365-2486.1998.00151.xURL [本文引用: 1]
The challenge to identify the biospheric sinks for about half the total carbon emissions from fossil fuels must include a consideration of below-ground ecosystem processes as well as those more easily measured above-ground. Recent studies suggest that tropical grasslands and savannas may contribute more to the 'missing sink' than was previously appreciated, perhaps as much as 0.5 Pg (= 0.5 Gt) carbon per annum. The rapid increase in availability of productivity data facilitated by the Internet will be important for future scaling-up of global change responses, to establish independent lines of evidence about the location and size of carbon sinks.
URLMagsci [本文引用: 1]
<p>基于海北州天然草地退化沙化的事实,简要分析了退化沙化的表现和危害,重点阐述了区域天然草地退化沙化的多层次原因,提出了相应的应对策略和建议。分析表明:海北州天然草地退化沙化的直接原因为超载过牧和生物链遭到破坏,间接原因为牧业人口数量的增加和畜牧技术的应用,内在原因为高寒气候特点和复杂地形造成区域草地生态系统的脆弱性。提出3项对策建议:推行计划生育,增加教育投资,控制并逐步减少牧业人口;大力推行现代高效畜牧业发展模式,发展栽培草地,实施“以草定畜”;树立“不再破坏就是保护”的意识,开展天然草地的有限治理。</p>
URLMagsci [本文引用: 1]
<p>基于海北州天然草地退化沙化的事实,简要分析了退化沙化的表现和危害,重点阐述了区域天然草地退化沙化的多层次原因,提出了相应的应对策略和建议。分析表明:海北州天然草地退化沙化的直接原因为超载过牧和生物链遭到破坏,间接原因为牧业人口数量的增加和畜牧技术的应用,内在原因为高寒气候特点和复杂地形造成区域草地生态系统的脆弱性。提出3项对策建议:推行计划生育,增加教育投资,控制并逐步减少牧业人口;大力推行现代高效畜牧业发展模式,发展栽培草地,实施“以草定畜”;树立“不再破坏就是保护”的意识,开展天然草地的有限治理。</p>
DOI:10.2307/2845965URL [本文引用: 1]
We investigated the effects of global climate change and doubled atmospheric CO2concentration to plant primary production and soil organic matter of typical steppe (Leymus chinense steppe and Stipa grandis steppe) and meadow steppe (Filifolium sibiricum steppe, S. baicalensis steppe and L. chinense steppe) at individual sites in Inner Mongolia, using the CENTURY ecosystem model. In the simulation of climate change, loss of soil organic C ranges from 783 gC. m-2 in meadow steppe to 1485 gC.m-2 in typical steppe, and annual above-ground net primary production (ANPP) decreases by 17.6 gC.m-2 in meadow steppe to 29.5 gC.m-2 in typical steppe under CCC (Canadian Climate Center). While under GFDL (Geophysical Fluid Dynamics Laboratory), loss of soil organic C. varies from 584 gC.m-2 in typical steppe to 1164 gC. m-2 in meadow steppe, and ANPP decreases in the range of 18.3 gC.m-2 in typical steppe to 32.1 gC.m-2 in meadow steppe. In the simulations of climate change plus elevated CO2(from 350 p.p.m. to 700 p.p.m.), ANPP decreases by 5.4 gC.m-2 in meadow steppe to 11.3 gC.m-2 in typical steppe under CCC+CO2, while ANPP varies from an increase of 1.8 gC.m-2 in S. grandis steppe to a decrease of 20.6 gC.m-2 in meadow steppe under GFDL+CO2. Losses of soil organic C are slightly lower (in the range of 42 gC.m-2 to 248 gC.m-2) than losses of soil organic C. under climate change only. These five steppe ecosystems are very sensitive to climate change, dependent upon projected change in temperature and precipitation by GCMs of CCC and GFDL.
DOI:10.3321/j.issn:1000-3037.2001.01.009URL [本文引用: 1]
文中参照 Constaza等提出的方法,在对草地生态系统服务价格根据其生物量订正的基础上,逐项估计了各类草地生态系统的各项生态系统服务价值,得出全国草地生态系统每年的服务价值为 1 497 9× 108US$。其中扰乱管理价值占 16 07%,水管理和水供应价值占 14 44%,侵蚀控制和沉积保存价值占 9 08%,废物处理价值占 42 03%,授粉价值占 7 83%,生物控制价值占 7 2%,食物和原材料生产价值占 22 26%,娱乐和文化价值占 5 78%,其它如气体管理价值、土壤形成价值和栖息地价值低于 5%。受各类草地生物群落分布广度和单位面积生态服务功能强弱的综合影响,各类草地的生态服务价值贡献率有很大差异,其中,沼泽类对草地生态系统总服务价值的贡献率为 52 34%。温性草原类、热性草丛类、热性灌草丛类、低地草甸类和高寒草甸类对总服务价值的贡献分别在 5 03%~ 7 74%之间,其余类型的草地生态服务价值贡献率在 5%以下。
DOI:10.3321/j.issn:1000-3037.2001.01.009URL [本文引用: 1]
文中参照 Constaza等提出的方法,在对草地生态系统服务价格根据其生物量订正的基础上,逐项估计了各类草地生态系统的各项生态系统服务价值,得出全国草地生态系统每年的服务价值为 1 497 9× 108US$。其中扰乱管理价值占 16 07%,水管理和水供应价值占 14 44%,侵蚀控制和沉积保存价值占 9 08%,废物处理价值占 42 03%,授粉价值占 7 83%,生物控制价值占 7 2%,食物和原材料生产价值占 22 26%,娱乐和文化价值占 5 78%,其它如气体管理价值、土壤形成价值和栖息地价值低于 5%。受各类草地生物群落分布广度和单位面积生态服务功能强弱的综合影响,各类草地的生态服务价值贡献率有很大差异,其中,沼泽类对草地生态系统总服务价值的贡献率为 52 34%。温性草原类、热性草丛类、热性灌草丛类、低地草甸类和高寒草甸类对总服务价值的贡献分别在 5 03%~ 7 74%之间,其余类型的草地生态服务价值贡献率在 5%以下。
[本文引用: 1]
[本文引用: 1]
DOI:10.11686/cyxb20100128Magsci [本文引用: 1]
<p><font face="Verdana">根据河北省2005-2007年草地地面调查,基于110个样地、990个样方的调查数据,对河北省草地产草量进行了监测估算,结果表明,1)2005-2007 年河北省鲜草年均产量为62254697.81t,平均单产为13134.57kg/hm<sup>2</sup>;干草年均产量为19438911.94t,平均单产为4101.24kg/hm<sup>2</sup>。2)2005-2007 年各类草地干草平均单产:暖性灌草丛(5133.35kg/hm<sup>2</sup>)>暖性草丛(4182.22kg/hm<sup>2</sup>)>沼泽草地(3376.13kg/hm<sup>2</sup>)>山地草甸(坝上高原区,3268.35kg/hm<sup>2</sup>)>温性草甸草原(2665.94kg/hm<sup>2</sup>)>山地草甸(山地丘陵区,2111.55kg/hm<sup>2</sup>)>低地草甸(1872.44kg/hm<sup>2</sup>)>温性草原(1103.29kg/hm<sup>2</sup>)。暖性灌草丛草地、暖性草丛草地、山地草甸草地干草产量分别占全省草地产量的56.04%,20.85%和18.13%,是河北省草地产量主体。3)2005-2007河北各地市草地干草单产:沧州7134.40kg/hm<sup>2</sup>>保定6981.15kg/hm<sup>2</sup>>承德4850.26kg/hm<sup>2</sup>>石家庄4677.78kg/hm<sup>2</sup>>邯郸2802.83kg/hm<sup>2</sup>>张家口2519.58kg/hm<sup>2</sup>> 邢台2436.47kg/hm<sup>2</sup>> 秦皇岛2227.55kg/hm<sup>2</sup>> 唐山1984.14kg/hm<sup>2</sup>。承德、张家口、保定3地市草地干草产量占全省草地干草产量的84.44%,其他6地市草地干草产量占全省干草产量的15.56%。4)2005-2007年河北省草地干草年均单产与1979-1984年草地年均干草单产相比,干草单产下降36.49%,明显减产,草地干草产量平均每年下降1.46%。山地草甸、温性草原、沼泽草地、低地草甸、温性草甸草原等草地类的草地干草单产降幅属于严重减产,暖性灌丛草、暖性草丛草地类的草地干草单产降幅为明显减产。减产的草地面积占全省草地面积的91.96%,其中,严重减产的草地面积占全省草地面积的31.92%,另有8.04%的草地产量基本持平。</font></p>
DOI:10.11686/cyxb20100128Magsci [本文引用: 1]
<p><font face="Verdana">根据河北省2005-2007年草地地面调查,基于110个样地、990个样方的调查数据,对河北省草地产草量进行了监测估算,结果表明,1)2005-2007 年河北省鲜草年均产量为62254697.81t,平均单产为13134.57kg/hm<sup>2</sup>;干草年均产量为19438911.94t,平均单产为4101.24kg/hm<sup>2</sup>。2)2005-2007 年各类草地干草平均单产:暖性灌草丛(5133.35kg/hm<sup>2</sup>)>暖性草丛(4182.22kg/hm<sup>2</sup>)>沼泽草地(3376.13kg/hm<sup>2</sup>)>山地草甸(坝上高原区,3268.35kg/hm<sup>2</sup>)>温性草甸草原(2665.94kg/hm<sup>2</sup>)>山地草甸(山地丘陵区,2111.55kg/hm<sup>2</sup>)>低地草甸(1872.44kg/hm<sup>2</sup>)>温性草原(1103.29kg/hm<sup>2</sup>)。暖性灌草丛草地、暖性草丛草地、山地草甸草地干草产量分别占全省草地产量的56.04%,20.85%和18.13%,是河北省草地产量主体。3)2005-2007河北各地市草地干草单产:沧州7134.40kg/hm<sup>2</sup>>保定6981.15kg/hm<sup>2</sup>>承德4850.26kg/hm<sup>2</sup>>石家庄4677.78kg/hm<sup>2</sup>>邯郸2802.83kg/hm<sup>2</sup>>张家口2519.58kg/hm<sup>2</sup>> 邢台2436.47kg/hm<sup>2</sup>> 秦皇岛2227.55kg/hm<sup>2</sup>> 唐山1984.14kg/hm<sup>2</sup>。承德、张家口、保定3地市草地干草产量占全省草地干草产量的84.44%,其他6地市草地干草产量占全省干草产量的15.56%。4)2005-2007年河北省草地干草年均单产与1979-1984年草地年均干草单产相比,干草单产下降36.49%,明显减产,草地干草产量平均每年下降1.46%。山地草甸、温性草原、沼泽草地、低地草甸、温性草甸草原等草地类的草地干草单产降幅属于严重减产,暖性灌丛草、暖性草丛草地类的草地干草单产降幅为明显减产。减产的草地面积占全省草地面积的91.96%,其中,严重减产的草地面积占全省草地面积的31.92%,另有8.04%的草地产量基本持平。</font></p>
DOI:10.11686/cyxb20090601Magsci [本文引用: 1]
<FONT face=Verdana>根据河北省2005-2007年草地地面调查,结合1979-1984年河北省草地调查资料,确定了河北省草地类型、面积、分布及物种组成特征,并对河北省草地盖度、高度和产草量年际变化动态进行了监测研究。结果表明,1)草地类型、面积及分布:河北省有温性草甸草原、温性草原、暖性灌草丛、暖性草丛、山地草甸草原、低地草甸、沼泽草地等7个草地类型;草地分布分为坝上高原区、山地丘陵区草地及滨海平原区;2)草地产量(干草):暖性灌草丛<BR>(5133.35 kg/hm<SUP>2</SUP>)>暖性草丛(4182.22 kg/hm<SUP>2</SUP>)>山地草甸(坝上高原区)(3268.35 kg/hm<SUP>2</SUP>)> 温性草甸草原(2665.94 kg/hm<SUP>2</SUP>)> 山地草甸(山地丘陵区)(2111.55 kg/hm<SUP>2</SUP>)> 低地草甸(1872.44 kg/hm<SUP>2</SUP>)> 温性草原(1103.29 kg/hm<SUP>2</SUP>),暖性灌草丛产量最高,温性草原产量最低。3)草地植被盖度:山地草甸(坝上高原区)(93.97%)>山地草甸(山地丘陵区)(82.60%)> 低地草甸(82.26%)> 暖性灌草丛(79.70%)> 暖性草丛(76.91%)>温性草甸草原(71.59%)> 温性草原(57.63%),山地草甸的植被盖度最大,温性草原的植被盖度最小。4)草地植被高度:暖性草丛(49.25cm)>暖性灌草丛(34.18cm)>山地草甸(坝上高原区)(28.88cm)>温性草甸(26.22cm)>山地草甸(山地丘陵区)(21.98cm)>温性草原(19.77cm)>低地草甸(18.77cm),暖性草丛草地植被高度最高,低地草甸草地植被高度最低。5)草地植被动态:2006 年草地植被基本以正向波动为主,2005 和2007年基本以负向波动为主,其中2007年植被负向波动率大于2005年植被负向波动率。按草地类型,暖性草丛类草地年际波动幅度最大,温性草原草地次之,温性草甸草地的年际波动幅度最小,其他类草地年际波动幅度介于暖性草丛类草地、温性草原类草地与低地草甸类草地之间。<BR></FONT>
DOI:10.11686/cyxb20090601Magsci [本文引用: 1]
<FONT face=Verdana>根据河北省2005-2007年草地地面调查,结合1979-1984年河北省草地调查资料,确定了河北省草地类型、面积、分布及物种组成特征,并对河北省草地盖度、高度和产草量年际变化动态进行了监测研究。结果表明,1)草地类型、面积及分布:河北省有温性草甸草原、温性草原、暖性灌草丛、暖性草丛、山地草甸草原、低地草甸、沼泽草地等7个草地类型;草地分布分为坝上高原区、山地丘陵区草地及滨海平原区;2)草地产量(干草):暖性灌草丛<BR>(5133.35 kg/hm<SUP>2</SUP>)>暖性草丛(4182.22 kg/hm<SUP>2</SUP>)>山地草甸(坝上高原区)(3268.35 kg/hm<SUP>2</SUP>)> 温性草甸草原(2665.94 kg/hm<SUP>2</SUP>)> 山地草甸(山地丘陵区)(2111.55 kg/hm<SUP>2</SUP>)> 低地草甸(1872.44 kg/hm<SUP>2</SUP>)> 温性草原(1103.29 kg/hm<SUP>2</SUP>),暖性灌草丛产量最高,温性草原产量最低。3)草地植被盖度:山地草甸(坝上高原区)(93.97%)>山地草甸(山地丘陵区)(82.60%)> 低地草甸(82.26%)> 暖性灌草丛(79.70%)> 暖性草丛(76.91%)>温性草甸草原(71.59%)> 温性草原(57.63%),山地草甸的植被盖度最大,温性草原的植被盖度最小。4)草地植被高度:暖性草丛(49.25cm)>暖性灌草丛(34.18cm)>山地草甸(坝上高原区)(28.88cm)>温性草甸(26.22cm)>山地草甸(山地丘陵区)(21.98cm)>温性草原(19.77cm)>低地草甸(18.77cm),暖性草丛草地植被高度最高,低地草甸草地植被高度最低。5)草地植被动态:2006 年草地植被基本以正向波动为主,2005 和2007年基本以负向波动为主,其中2007年植被负向波动率大于2005年植被负向波动率。按草地类型,暖性草丛类草地年际波动幅度最大,温性草原草地次之,温性草甸草地的年际波动幅度最小,其他类草地年际波动幅度介于暖性草丛类草地、温性草原类草地与低地草甸类草地之间。<BR></FONT>
DOI:10.3321/j.issn:1004-5759.2008.01.015URL [本文引用: 1]
草地退化是草地生态学和草地经营学中倍受关注的问题。在已有研究成果和笔者的研究体会的基础 上总结了草地退化中的几组重要概念,包括生态退化与草场退化、植被退化与土壤退化、结构退化与功能退化、绝对退化与相对退化、参照系统与退化程度以及草地 退化小尺度定位研究与大尺度空间格局研究。对其进行辨析,同时指出这些概念的划分在草地退化理论与实践研究中的重要意义。
DOI:10.3321/j.issn:1004-5759.2008.01.015URL [本文引用: 1]
草地退化是草地生态学和草地经营学中倍受关注的问题。在已有研究成果和笔者的研究体会的基础 上总结了草地退化中的几组重要概念,包括生态退化与草场退化、植被退化与土壤退化、结构退化与功能退化、绝对退化与相对退化、参照系统与退化程度以及草地 退化小尺度定位研究与大尺度空间格局研究。对其进行辨析,同时指出这些概念的划分在草地退化理论与实践研究中的重要意义。
DOI:10.1111/j.1466-8238.2009.00502.xURL [本文引用: 1]
ABSTRACT Aim68 To investigate large-scale patterns of above-ground and below-ground biomass partitioning in grassland ecosystems and to test the isometric theory at the community level. Location68 Northern China, in diverse grassland types spanning temperate grasslands in arid and semi-arid regions to alpine grasslands on the Tibetan Plateau. Methods68 We investigated above-ground and below-ground biomass in China's grasslands by conducting five consecutive sampling campaigns across the northern part of the country during 2001–05. We then documented the root:shoot ratio (R/S) and its relationship with climatic factors for China's grasslands. We further explored relationships between above-ground and below-ground biomass across different grassland types. Results68 Our results indicated that the overall R/S of China's grasslands was larger than the global average (6.3 vs. 3.7). The R/S for China's grasslands did not show any significant trend with either mean annual temperature or mean annual precipitation. Above-ground biomass was nearly proportional to below-ground biomass with a scaling exponent (the slope of log–log linear relationship between above-ground and below-ground biomass) of 1.02 across various grassland types. The slope did not differ significantly between temperate and alpine grasslands or between steppe and meadow. Main conclusions68 Our findings support the isometric theory of above-ground and below-ground biomass partitioning, and suggest that above-ground biomass scales isometrically with below-ground biomass at the community level.
DOI:10.3969/j.issn.1001-0629.2007.09.001URL [本文引用: 1]
对黄河上游甘肃段草地退化的现状和机理进行了系统分析,结果表明,区域环境脆弱、全球气候变化、过度放牧、鼠害严重、草地管理制度不完善等因素是导致黄河上游甘肃段草地退化的主要原因,在这些因素的相互作用下,该区域草地退化的趋势可能进一步加剧.
DOI:10.3969/j.issn.1001-0629.2007.09.001URL [本文引用: 1]
对黄河上游甘肃段草地退化的现状和机理进行了系统分析,结果表明,区域环境脆弱、全球气候变化、过度放牧、鼠害严重、草地管理制度不完善等因素是导致黄河上游甘肃段草地退化的主要原因,在这些因素的相互作用下,该区域草地退化的趋势可能进一步加剧.
[本文引用: 1]
[本文引用: 1]
河北省畜牧水产局
1
1990
... id="C7">河北省的草地资源处于温带欧亚草原与森林过渡地带, 天然草地植被类型多样, 植物资源丰富, 草地畜牧业生产在当地国民经济中占有重要的地位(
河北省畜牧水产局
1
1990
... id="C7">河北省的草地资源处于温带欧亚草原与森林过渡地带, 天然草地植被类型多样, 植物资源丰富, 草地畜牧业生产在当地国民经济中占有重要的地位(
河北省畜牧局
1
1997
... id="C7">河北省的草地资源处于温带欧亚草原与森林过渡地带, 天然草地植被类型多样, 植物资源丰富, 草地畜牧业生产在当地国民经济中占有重要的地位(
河北省畜牧局
1
1997
... id="C7">河北省的草地资源处于温带欧亚草原与森林过渡地带, 天然草地植被类型多样, 植物资源丰富, 草地畜牧业生产在当地国民经济中占有重要的地位(
内蒙古锡林河流域草原植物群落功能群植物组成沿水热梯度变化的样带研究
1
2002
... id="C48">河北省不同草地类型的植物群落物种组成不同, 这不仅是植物种对环境条件长期适应的结果, 更是反映了草地的生物气候类型(
内蒙古锡林河流域草原植物群落功能群植物组成沿水热梯度变化的样带研究
1
2002
... id="C48">河北省不同草地类型的植物群落物种组成不同, 这不仅是植物种对环境条件长期适应的结果, 更是反映了草地的生物气候类型(
藏北高寒草地植被的碳密度与碳贮量
1
2007
... id="C47">河北省不同草地植被生物量碳密度差异显著, 为273.0-919.8 g C·m-2, 平均值506.6 g C·m-2, 高于中国草地生物量平均碳密度320.5 g C·m-2 (
藏北高寒草地植被的碳密度与碳贮量
1
2007
... id="C47">河北省不同草地植被生物量碳密度差异显著, 为273.0-919.8 g C·m-2, 平均值506.6 g C·m-2, 高于中国草地生物量平均碳密度320.5 g C·m-2 (
1
2000
... id="C6">草地资源是陆生生态系统的重要组成部分, 在全球陆地碳循环中起着重要作用.世界上的草地约占全球陆地面积的20%, 主要分布在各大陆内部气候干燥、降水较少的地区(
1
2000
... id="C6">草地资源是陆生生态系统的重要组成部分, 在全球陆地碳循环中起着重要作用.世界上的草地约占全球陆地面积的20%, 主要分布在各大陆内部气候干燥、降水较少的地区(
陕西省天然草地生物量空间分布格局及其影响因素
1
2012
... id="C48">河北省不同草地类型的植物群落物种组成不同, 这不仅是植物种对环境条件长期适应的结果, 更是反映了草地的生物气候类型(
陕西省天然草地生物量空间分布格局及其影响因素
1
2012
... id="C48">河北省不同草地类型的植物群落物种组成不同, 这不仅是植物种对环境条件长期适应的结果, 更是反映了草地的生物气候类型(
1
1996
... id="C6">草地资源是陆生生态系统的重要组成部分, 在全球陆地碳循环中起着重要作用.世界上的草地约占全球陆地面积的20%, 主要分布在各大陆内部气候干燥、降水较少的地区(
1
1996
... id="C6">草地资源是陆生生态系统的重要组成部分, 在全球陆地碳循环中起着重要作用.世界上的草地约占全球陆地面积的20%, 主要分布在各大陆内部气候干燥、降水较少的地区(
高寒草甸植物群落中物种丰富度与生产力的关系研究
1
2003
... id="C48">河北省不同草地类型的植物群落物种组成不同, 这不仅是植物种对环境条件长期适应的结果, 更是反映了草地的生物气候类型(
高寒草甸植物群落中物种丰富度与生产力的关系研究
1
2003
... id="C48">河北省不同草地类型的植物群落物种组成不同, 这不仅是植物种对环境条件长期适应的结果, 更是反映了草地的生物气候类型(
1
1992
... id="C7">河北省的草地资源处于温带欧亚草原与森林过渡地带, 天然草地植被类型多样, 植物资源丰富, 草地畜牧业生产在当地国民经济中占有重要的地位(
Carbon storage in the grasslands of China based on field measurements of above- and below- ground biomass
2
2008
... id="C8">近年来, 相关的****已经对我国草地生物量开展了大量的研究, 对于大尺度范围内草地生物量的研究, ****多采用遥感的方法进行.如
... id="C46">在草地生态系统中, 地下生物量显著高于地上生物量, 本研究结果中温性典型草原和温性草甸草原的地下生物量分别为588.7和920.1 g·m-2,
中国陆地生态系统的碳库. 见:王庚辰, 温玉璞
2
1996
... id="C8">近年来, 相关的****已经对我国草地生物量开展了大量的研究, 对于大尺度范围内草地生物量的研究, ****多采用遥感的方法进行.如
... id="C45">对于大尺度范围内草地生物量的估算, ****多采用遥感的方法进行.基于我国第一次草地资源清查资料和遥感数据估算我国北方温带荒漠草原、温带典型草原和温带草甸草原的地上生物量分别为45.6、89.0和146.5 g·m-2 (
中国陆地生态系统的碳库. 见:王庚辰, 温玉璞
2
1996
... id="C8">近年来, 相关的****已经对我国草地生物量开展了大量的研究, 对于大尺度范围内草地生物量的研究, ****多采用遥感的方法进行.如
... id="C45">对于大尺度范围内草地生物量的估算, ****多采用遥感的方法进行.基于我国第一次草地资源清查资料和遥感数据估算我国北方温带荒漠草原、温带典型草原和温带草甸草原的地上生物量分别为45.6、89.0和146.5 g·m-2 (
Ecosystem carbon stocks and their changes in China’s grasslands
3
2010
... id="C8">近年来, 相关的****已经对我国草地生物量开展了大量的研究, 对于大尺度范围内草地生物量的研究, ****多采用遥感的方法进行.如
... id="C47">河北省不同草地植被生物量碳密度差异显著, 为273.0-919.8 g C·m-2, 平均值506.6 g C·m-2, 高于中国草地生物量平均碳密度320.5 g C·m-2 (
... id="C48">河北省不同草地类型的植物群落物种组成不同, 这不仅是植物种对环境条件长期适应的结果, 更是反映了草地的生物气候类型(
内蒙古草地样带植物群落生物量的梯度研究
1
2006
... id="C48">河北省不同草地类型的植物群落物种组成不同, 这不仅是植物种对环境条件长期适应的结果, 更是反映了草地的生物气候类型(
内蒙古草地样带植物群落生物量的梯度研究
1
2006
... id="C48">河北省不同草地类型的植物群落物种组成不同, 这不仅是植物种对环境条件长期适应的结果, 更是反映了草地的生物气候类型(
中国草地主要生态环境问题分析与防治对策
1
2005
... id="C7">河北省的草地资源处于温带欧亚草原与森林过渡地带, 天然草地植被类型多样, 植物资源丰富, 草地畜牧业生产在当地国民经济中占有重要的地位(
中国草地主要生态环境问题分析与防治对策
1
2005
... id="C7">河北省的草地资源处于温带欧亚草原与森林过渡地带, 天然草地植被类型多样, 植物资源丰富, 草地畜牧业生产在当地国民经济中占有重要的地位(
中国北方草地退化及其防治对策
1
1997
... id="C7">河北省的草地资源处于温带欧亚草原与森林过渡地带, 天然草地植被类型多样, 植物资源丰富, 草地畜牧业生产在当地国民经济中占有重要的地位(
中国北方草地退化及其防治对策
1
1997
... id="C7">河北省的草地资源处于温带欧亚草原与森林过渡地带, 天然草地植被类型多样, 植物资源丰富, 草地畜牧业生产在当地国民经济中占有重要的地位(
河北草地资源与利用
1
1981
... id="C9">河北省是我国华北地区的腹心地带, 地处113.45°-119.75° E, 36.85°-42.62° N, 全省年平均气温在4-13 ℃之间, 1月最低温-14- -2 ℃, 7月最高温20-27 ℃, 年降水量300-800 mm, 燕山南麓和太行山东降雨较多, 张北高原降水一般不足400 mm, 为季风性暖温带气候, 四季分明.河北省总面积187600 km2, 其中平原约占1/3, 山区和高原约占2/3.天然草地总面积4.74 × 106 hm2, 占全国草地面积的1.2%, 占全省土地面积的35.7%左右.河北省的草地大部分分布在北部与西北山地, 其中80.3%的草地分布在北部的承德、张家口和保定三市, 是坝上地区、北部山地以及太行山区发展畜牧业的基地(
河北草地资源与利用
1
1981
... id="C9">河北省是我国华北地区的腹心地带, 地处113.45°-119.75° E, 36.85°-42.62° N, 全省年平均气温在4-13 ℃之间, 1月最低温-14- -2 ℃, 7月最高温20-27 ℃, 年降水量300-800 mm, 燕山南麓和太行山东降雨较多, 张北高原降水一般不足400 mm, 为季风性暖温带气候, 四季分明.河北省总面积187600 km2, 其中平原约占1/3, 山区和高原约占2/3.天然草地总面积4.74 × 106 hm2, 占全国草地面积的1.2%, 占全省土地面积的35.7%左右.河北省的草地大部分分布在北部与西北山地, 其中80.3%的草地分布在北部的承德、张家口和保定三市, 是坝上地区、北部山地以及太行山区发展畜牧业的基地(
A review on the distribution patterns of carbon density in terrestrial ecosystems
1
2004
... id="C47">河北省不同草地植被生物量碳密度差异显著, 为273.0-919.8 g C·m-2, 平均值506.6 g C·m-2, 高于中国草地生物量平均碳密度320.5 g C·m-2 (
Biomass carbon stocks and their changes in northern China’s grasslands during 1982-2006
1
2010
... id="C8">近年来, 相关的****已经对我国草地生物量开展了大量的研究, 对于大尺度范围内草地生物量的研究, ****多采用遥感的方法进行.如
内蒙古温带草地生物量及其与环境因子的关系
2
2008
... id="C48">河北省不同草地类型的植物群落物种组成不同, 这不仅是植物种对环境条件长期适应的结果, 更是反映了草地的生物气候类型(
... 0.05).已有研究发现气候是影响草地生物量的重要因素, 尤其是地上生物量的空间变化主要受降水量控制, 在对内蒙古草原的研究中发现, 降水量是影响生物量分布差异的最主要的环境因素, 地上、地下生物量随着年降水量的增加显著增加(
内蒙古温带草地生物量及其与环境因子的关系
2
2008
... id="C48">河北省不同草地类型的植物群落物种组成不同, 这不仅是植物种对环境条件长期适应的结果, 更是反映了草地的生物气候类型(
... 0.05).已有研究发现气候是影响草地生物量的重要因素, 尤其是地上生物量的空间变化主要受降水量控制, 在对内蒙古草原的研究中发现, 降水量是影响生物量分布差异的最主要的环境因素, 地上、地下生物量随着年降水量的增加显著增加(
Carbon storage in terrestrial ecosystems of China: Estimates at different spatial resolutions and their responses to climate change
2
2001
... id="C8">近年来, 相关的****已经对我国草地生物量开展了大量的研究, 对于大尺度范围内草地生物量的研究, ****多采用遥感的方法进行.如
... id="C47">河北省不同草地植被生物量碳密度差异显著, 为273.0-919.8 g C·m-2, 平均值506.6 g C·m-2, 高于中国草地生物量平均碳密度320.5 g C·m-2 (
Carbon storage in grasslands of China
1
2002
... id="C8">近年来, 相关的****已经对我国草地生物量开展了大量的研究, 对于大尺度范围内草地生物量的研究, ****多采用遥感的方法进行.如
Forage yield-based carbon storage in grasslands of China
2
2004a
... id="C8">近年来, 相关的****已经对我国草地生物量开展了大量的研究, 对于大尺度范围内草地生物量的研究, ****多采用遥感的方法进行.如
... id="C45">对于大尺度范围内草地生物量的估算, ****多采用遥感的方法进行.基于我国第一次草地资源清查资料和遥感数据估算我国北方温带荒漠草原、温带典型草原和温带草甸草原的地上生物量分别为45.6、89.0和146.5 g·m-2 (
Estimating net primary productivity of grasslands from field biomass measurements in temperate northern China
1
2004b
... id="C46">在草地生态系统中, 地下生物量显著高于地上生物量, 本研究结果中温性典型草原和温性草甸草原的地下生物量分别为588.7和920.1 g·m-2,
Changes in biomass carbon stocks in China’s grasslands between 1982 and 1999. Global Biogeochemical Cycles, 21,
2
2007
... id="C8">近年来, 相关的****已经对我国草地生物量开展了大量的研究, 对于大尺度范围内草地生物量的研究, ****多采用遥感的方法进行.如
... id="C46">在草地生态系统中, 地下生物量显著高于地上生物量, 本研究结果中温性典型草原和温性草甸草原的地下生物量分别为588.7和920.1 g·m-2,
The global carbon sink: A grassland perspective
1
1998
... id="C6">草地资源是陆生生态系统的重要组成部分, 在全球陆地碳循环中起着重要作用.世界上的草地约占全球陆地面积的20%, 主要分布在各大陆内部气候干燥、降水较少的地区(
海北州天然草地退化沙化成因与对策
1
2009
... id="C7">河北省的草地资源处于温带欧亚草原与森林过渡地带, 天然草地植被类型多样, 植物资源丰富, 草地畜牧业生产在当地国民经济中占有重要的地位(
海北州天然草地退化沙化成因与对策
1
2009
... id="C7">河北省的草地资源处于温带欧亚草原与森林过渡地带, 天然草地植被类型多样, 植物资源丰富, 草地畜牧业生产在当地国民经济中占有重要的地位(
Sensitivity of Inner Mongolia grasslands to climate change
1
1995
... id="C48">河北省不同草地类型的植物群落物种组成不同, 这不仅是植物种对环境条件长期适应的结果, 更是反映了草地的生物气候类型(
中国自然草地生态系统服务价值
1
2001
... id="C6">草地资源是陆生生态系统的重要组成部分, 在全球陆地碳循环中起着重要作用.世界上的草地约占全球陆地面积的20%, 主要分布在各大陆内部气候干燥、降水较少的地区(
中国自然草地生态系统服务价值
1
2001
... id="C6">草地资源是陆生生态系统的重要组成部分, 在全球陆地碳循环中起着重要作用.世界上的草地约占全球陆地面积的20%, 主要分布在各大陆内部气候干燥、降水较少的地区(
1982-2003年中国草地生物量时空格局变化研究
1
2009
... id="C45">对于大尺度范围内草地生物量的估算, ****多采用遥感的方法进行.基于我国第一次草地资源清查资料和遥感数据估算我国北方温带荒漠草原、温带典型草原和温带草甸草原的地上生物量分别为45.6、89.0和146.5 g·m-2 (
1982-2003年中国草地生物量时空格局变化研究
1
2009
... id="C45">对于大尺度范围内草地生物量的估算, ****多采用遥感的方法进行.基于我国第一次草地资源清查资料和遥感数据估算我国北方温带荒漠草原、温带典型草原和温带草甸草原的地上生物量分别为45.6、89.0和146.5 g·m-2 (
河北省草地产量动态监测
1
2010
... id="C7">河北省的草地资源处于温带欧亚草原与森林过渡地带, 天然草地植被类型多样, 植物资源丰富, 草地畜牧业生产在当地国民经济中占有重要的地位(
河北省草地产量动态监测
1
2010
... id="C7">河北省的草地资源处于温带欧亚草原与森林过渡地带, 天然草地植被类型多样, 植物资源丰富, 草地畜牧业生产在当地国民经济中占有重要的地位(
河北省草地资源分布及植被特征动态
1
2009
... id="C7">河北省的草地资源处于温带欧亚草原与森林过渡地带, 天然草地植被类型多样, 植物资源丰富, 草地畜牧业生产在当地国民经济中占有重要的地位(
河北省草地资源分布及植被特征动态
1
2009
... id="C7">河北省的草地资源处于温带欧亚草原与森林过渡地带, 天然草地植被类型多样, 植物资源丰富, 草地畜牧业生产在当地国民经济中占有重要的地位(
草地退化相关概念辨析
1
2008
... id="C7">河北省的草地资源处于温带欧亚草原与森林过渡地带, 天然草地植被类型多样, 植物资源丰富, 草地畜牧业生产在当地国民经济中占有重要的地位(
草地退化相关概念辨析
1
2008
... id="C7">河北省的草地资源处于温带欧亚草原与森林过渡地带, 天然草地植被类型多样, 植物资源丰富, 草地畜牧业生产在当地国民经济中占有重要的地位(
Large-scale pattern of biomass partitioning across China’s grasslands
1
2010
... id="C46">在草地生态系统中, 地下生物量显著高于地上生物量, 本研究结果中温性典型草原和温性草甸草原的地下生物量分别为588.7和920.1 g·m-2,
黄河上游甘肃段草地退化的现状及机理研究
1
2007
... id="C7">河北省的草地资源处于温带欧亚草原与森林过渡地带, 天然草地植被类型多样, 植物资源丰富, 草地畜牧业生产在当地国民经济中占有重要的地位(
黄河上游甘肃段草地退化的现状及机理研究
1
2007
... id="C7">河北省的草地资源处于温带欧亚草原与森林过渡地带, 天然草地植被类型多样, 植物资源丰富, 草地畜牧业生产在当地国民经济中占有重要的地位(
1
1997
... id="C7">河北省的草地资源处于温带欧亚草原与森林过渡地带, 天然草地植被类型多样, 植物资源丰富, 草地畜牧业生产在当地国民经济中占有重要的地位(
1
1997
... id="C7">河北省的草地资源处于温带欧亚草原与森林过渡地带, 天然草地植被类型多样, 植物资源丰富, 草地畜牧业生产在当地国民经济中占有重要的地位(
Copyright © 2021 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}