Effect of Pb pollution on the growth, biomass allocation and photosynthesis of Phragmites australis in flood and drought environment
ZHANG Na1,2,3, ZHU Yang-Chun1, LI Zhi-Qiang5, LU Xin1, FAN Ru-Qin1, LIU Li-Zhu1, TONG Fei1, CHEN Jing3, MU Chun-Sheng,4,*, ZHANG Zhen-Hua1,*1Institute of Agricultural Resource and Environment, Jiangsu Academy of Agricultural Sciences, Nanjing 210014, China 2Key Laboratory of Food Quality and Safety of Jiangsu Province, Jiangsu Academy of Agricultural Sciences, Nanjing 210014, China 3Key Laboratory for Palygorskite Science and Applied Technology of Jiangsu Province, Huaiyin Institute of Technology, Huai’an, Jiangsu 223003, China; 4Key Laboratory of Vegetation Ecology, Ministry of Education, Institute of Grassland Science, Northeast Normal University, Changchun 130024, China 5Institution of Climate Change and Public Polices, School of Public Administration, Nanjing University of Information Science and Technology, Nanjing 210044, China
通讯作者: 通讯作者Author for correspondence (E-mail: jshe@pku.edu.cn)
SupportedbytheChinaPostdoctoralScienceFoundation.(2017M621670) the National Basic Research Program of China.( 2017M621670) the Agricultural Science and Technology Independent Innovation Fund of Jiangsu Province.(CX(16)1051)
Abstract Aims Reed (Phragmites australis) is a typical perennial rhizomatic plant with extensive tolerance to environmental stress. In order to better understand the adaptation and tolerance of reeds subjected to heavy metal pollution in different levels of water, we conducted a study on the effects of Pb pollution on growth, biomass and photosynthesis of reeds in flood and drought environment. This research would provide theoretical basis for application of reeds in wetland restoration and remediation.Methods We conducted a pot experiment with destructive sampling after 90 days of growth. The water treatments were main plot, including two water levels. The Pb treatments were secondary plot (nested within water treatments), including five levels (0, 500, 1 500, 3 000, 4 500 mg·kg-1). There were 10 treatments with 12 replicates per treatment.Important findings In the flood environment, Pb pollution significantly inhibited the growth of buds and rhizomes, but had no significant effect on the number of offspring shoots. The offspring shoots had higher growth rate per day, net photosynthetic rate and biomass compared to the parent shoots. In the drought environment, Pb pollution inhibited the growth of roots, buds and rhizomes, and biomass accumulation of parent and offspring shoots as well as photosynthetic parameters. These parameters were lower under the drought condition than in the flood environment. The Pb was mostly concentrated in roots compared to rhizomes and offspring shoots. In both flood and drought environments, the concentration of Pb in parent shoots was about three times of that in offspring shoots. The Pb concentration in offspring shoots under the flood condition was less than that in the drought environment. Overall, these results indicated that the synergistic effect of Pb and drought significantly inhibited the growth, biomass accumulation and photosynthesis of reeds, which might result in reduced offspring productivity and population density and may lead to population decline. However, the flooded reeds could adopt some strategies of Pb allocation to alleviate the negative effect of Pb on the growth, physiology and clonal propagation, benefiting the population reproduction and stabilization. Keywords:Pb pollution;water;growth;biomass allocation;photosynthesis
PDF (1534KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 张娜, 朱阳春, 李志强, 卢信, 范如芹, 刘丽珠, 童非, 陈静, 穆春生, 张振华. 淹水和干旱生境下铅对芦苇生长、生物量分配和光合作用的影响. 植物生态学报, 2018, 42(2): 229-239 doi:10.17521/cjpe.2017.0218 ZHANG Na, ZHU Yang-Chun, LI Zhi-Qiang, LU Xin, FAN Ru-Qin, LIU Li-Zhu, TONG Fei, CHEN Jing, MU Chun-Sheng, ZHANG Zhen-Hua. Effect of Pb pollution on the growth, biomass allocation and photosynthesis of Phragmites australis in flood and drought environment. Chinese Journal of Plant Ecology, 2018, 42(2): 229-239 doi:10.17521/cjpe.2017.0218
Table 1 表1 表1铅和水分影响芦苇母株、子株生长的重复测量和双因素方差分析结果 Table 1Results of Repeated-measures and two-way ANOVA of the effect of Pb and water stress on growth of mother and daughter shoots of Phragmites australis
Table 2 表2 表2不同时间内铅污染对不同生境下芦苇母株生长的影响(平均值±标准误差, n = 40) Table 2The effects of Pb on the growth of parent shoots of Phragmites australis grown in flood and dry environment after 30, 60 and 90 days of treatment (mean ± SE, n = 40)
水分 Water level
处理时间 Treatment time (d)
指标 Parameter
铅处理浓度 Pb concentration of treatments (mg·kg-1)
对照 Control
500
1 500
3 000
4 500
淹水 Flood
30
PSL (cm)
18.13 ± 0.81a
16.64 ± 0.75ab
15.54 ± 0.40b
13.10 ± 0.52c
13.24 ± 0.29c
PDLN (No.)
1.28 ± 0.05b
1.54 ± 0.12ab
1.73 ± 0.11a
1.90 ± 0.06a
1.84 ± 0.13a
PGR (cm·d-1)
0.44 ± 0.03a
0.39 ± 0.02ab
0.35 ± 0.01b
0.27 ± 0.02c
0.28 ± 0.01c
60
PSL (cm)
23.86 ± 0.83a
22.34 ± 1.52ab
22.71 ± 0.72ab
19.22 ± 1.04b
18.17 ± 0.54b
PDLN (No.)
3.57 ± 0.15b
3.65 ± 0.22b
4.38 ± 0.14a
4.50 ± 0.24a
4.56 ± 0.08a
PGR (cm·d-1)
0.19 ± 0.02a
0.19 ± 0.03a
0.24 ± 0.01a
0.21 ± 0.04a
0.16 ± 0.02a
90
PSL (cm)
26.00 ± 0.96a
24.03 ± 1.84a
24.21 ± 0.97a
22.80 ± 0.57a
21.68 ± 1.20a
PDLN (No.)
7.02 ± 0.06a
6.59 ± 0.43a
6.61 ± 0.22a
6.80 ± 0.16a
6.43 ± 0.15a
PGR (cm·d-1)
0.07 ± 0.03a
0.06 ± 0.01a
0.05 ± 0.01a
0.12 ± 0.04a
0.12 ± 0.03a
干旱 Drought
30
PSL (cm)
12.17 ± 0.88a*
11.90 ± 0.73a*
9.30 ± 0.37b*
7.87 ± 0.36b*
7.72 ± 0.42b*
PDLN (No.)
1.64 ± 0.06c*
2.08 ± 0.15b*
2.13 ± 0.05ab*
2.18 ± 0.10ab*
2.42 ± 0.17a*
PGR (cm·d-1)
0.24 ± 0.03a*
0.23 ± 0.02a*
0.15 ± 0.01b*
0.10 ± 0.01b*
0.09 ± 0.01b*
60
PSL (cm)
17.30 ± 0.54a*
15.06 ± 0.89b*
11.15 ± 0.57c*
9.09 ± 0.42d*
7.58 ± 0.61d*
PDLN (No.)
2.60 ± 0.04b
3.19 ± 0.27a*
3.33 ± 0.12a*
3.51 ± 0.17a*
3.65 ± 0.15a*
PGR (cm·d-)
0.17 ± 0.03a
0.11 ± 0.01b*
0.06 ± 0.01bc*
0.04 ± 0.00c*
0.02 ± 0.01c*
90
PSL (cm)
20.29 ± 0.94a*
18.14 ± 0.74a*
13.61 ± 0.94b*
9.43 ± 0.10c*
8.31 ± 0.53c*
PDLN (No.)
7.32 ± 0.16a
5.46 ± 0.15b*
4.82 ± 0.13bc*
4.73 ± 0.34bc*
4.08 ± 0.36c*
PGR (cm·d-1)
0.10 ± 0.02a
0.10 ± 0.03a
0.08 ± 0.03ab
0.02 ± 0.01b
0.03 ± 0.01b*
PDLN, dead leaf number of parent shoot; PGR, growth rate of parent shoots; PSL, stem length of parent shoot. Different lowercase letters indicate significant differences (p ≤ 0.05) between Pb levels within one water treatment level, and * indicates significant difference (p ≤ 0.05) between water treatment and drought treatment within one Pb level. PDLN, 母株叶片死亡数; PGR, 母株生长速率; PSL, 母株茎长。不同的小写字母表示同一水分处理水平上, 不同铅处理间的差异(p ≤ 0.05); *表示同一铅处理水平上, 淹水和干旱处理间差异显著(p ≤ 0.05)。
Table 3 表3 表3不同时间内铅污染对不同生境下芦苇子株生长的影响(平均值±标准误差, n = 40) Table 3The effects of Pb on the growth of offspring shoots of Phragmites australis grown in flood and dry environment after 30, 60 and 90 days of treatment (means ± SE, n = 40)
水分 Water level
处理时间 Treatment time (d)
指标 Parameter
铅处理浓度 Pb concentration of treatments (mg·kg-1)
对照 Control
500
1 500
3 000
4 500
淹水 Flood
30
OSL (cm)
15.08 ± 0.88a
13.20 ± 0.99ab
11.28 ± 0.95b
8.42 ± 0.26c
7.92 ± 0.35c
ODLN (No.)
0.27 ± 0.09a
0.28 ± 0.05a
0.06 ± 0.03b
0.06 ± 0.03b
0.00 ± 0.00b
OGR (cm·d-1)
0.76 ± 0.05a
0.66 ± 0.05ab
0.57 ± 0.05b
0.42 ± 0.01c
0.40 ± 0.02c
60
OSL (cm)
31.06 ± 0.37a
30.08 ± 0.42a
28.53 ± 0.77a
20.12 ± 0.73b
15.24 ± 0.83c
ODLN (No.)
1.49 ± 0.10a
1.50 ± 0.17a
1.74 ± 0.34a
1.82 ± 0.20a
1.96 ± 0.22a
OGR (cm·d-1)
0.53 ± 0.03a
0.57 ± 0.02a
0.58 ± 0.02a
0.39 ± 0.03b
0.24 ± 0.03c
90
OSL (cm)
41.22 ± 1.14a
40.97 ± 1.19a
40.65 ± 1.68a
32.98 ± 1.78b
22.41 ± 0.74c
ODLN (No.)
3.28 ± 0.15a
2.99 ± 0.27ab
2.95 ± 0.22ab
3.10 ± 0.20ab
2.53 ± 0.35b
OGR (cm·d-1)
0.34 ± 0.05ab
0.36 ± 0.03a
0.40 ± 0.04a
0.43 ± 0.04a
0.24 ± 0.03b
干旱Drought
30
OSL (cm)
4.50 ± 0.65a*
3.38 ± 0.63a*
3.38 ± 0.55a*
3.50 ± 0.29a*
1.50 ± 0.05b*
ODLN (No.)
0.00 ± 0.00a*
0.00 ± 0.00a*
0.00 ± 0.00a
0.00 ± 0.00a
0.00 ± 0.00a
OGR (cm·d-1)
0.23 ± 0.03a*
0.17 ± 0.03a*
0.17 ± 0.03a*
0.18 ± 0.01a*
0.07 ± 0.00b*
60
OSL (cm)
19.95 ± 1.08a*
11.23 ± 0.43b*
5.55 ± 0.75c*
4.44 ± 0.87c*
3.65 ± 0.61c*
ODLN (No.)
0.10 ± 0.04a*
0.14 ± 0.05a*
0.15 ± 0.05a*
0.25 ± 0.16a*
0.06 ± 0.06a*
OGR (cm·d-1)
0.52 ± 0.03a
0.26 ± 0.03b*
0.07 ± 0.04cd*
0.03 ± 0.02d*
0.14 ± 0.04c
90
OSL (cm)
28.82 ± 1.16a*
24.56 ± 0.76b*
14.46 ± 1.00c*
8.45 ± 0.25d*
4.13 ± 0.12e*
ODLN (No.)
2.23 ± 0.28a*
1.83 ± 0.11ab*
1.40 ± 0.26ab*
1.86 ± 0.34ab*
1.21 ± 0.44b*
OGR (cm·d-1)
0.30 ± 0.06b
0.44 ± 0.03a
0.30 ± 0.05b
0.13 ± 0.03c*
0.03 ± 0.02c*
ODLN, dead Leaf number of offspring shoots; OGR, growth rate of offspring shoots; OSL, stem length of offspring shoots. Different lowercase letters indicate significant differences (p ≤ 0.05) between Pb levels within one water treatment level, and * indicates significant difference (p ≤ 0.05) between water treatment and drought treatment within one Pb level. ODLN, 子株叶片死亡数; OGR, 子株生长速率; OSL, 子株茎长。不同的小写字母表示同一水分处理水平上, 不同铅处理间的差异 (p ≤ 0.05); *表示同一铅处理水平上, 淹水和干旱处理间差异显著(p ≤ 0.05)。
Table 4 表4 表4铅和水分处理对芦苇根茎生长、芽和子株数的双因素方差分析结果 Table 4Results of two-way ANOVA of the effect of Pb and water stress on rhizome growth, number of buds and offspring shoots of Phragmites australis
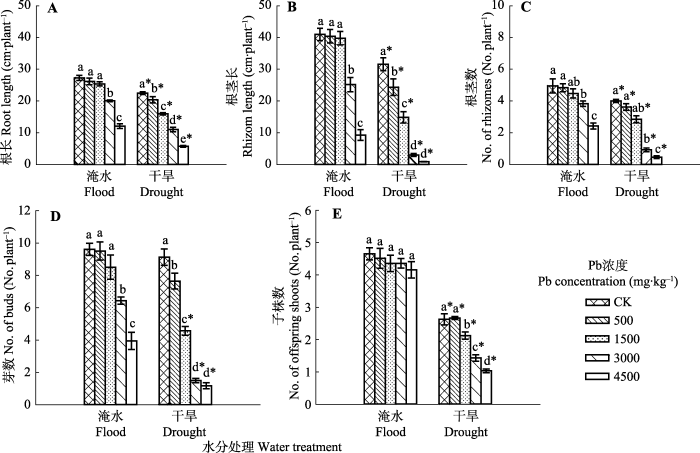
Fig. 1Effects of Pb pollution on growth of below-ground organs and abundance of above-ground offspring shoots of Phragmites australis in flood and dry environment (mean ± SE, n = 40). Different lowercase letters indicate significant differences (p ≤ 0.05) between Pb levels within one water treatment level, and * indicates significant difference (p ≤ 0.05) between water treatment and drought treatment within one Pb level.
Fig. 2The effects of Pb pollution on biomass accumulation of different organs of Phragmites australis in flood and dry environment (mean ± SE, n = 40). Different lowercase letters indicate significant differences (p ≤ 0.05) between Pb levels within one water treatment level, and * indicates significant difference (p ≤ 0.05) between water treatment and drought treatment within one Pb level.
Fig. 3The percentage of different organ biomass in total biomass of Phragmites australis subjected to Pb concentration in flood and dry environment (mean ± SE, n = 40). Different lowercase letters indicate significant differences (p ≤ 0.05) between Pb levels within one water treatment level, and * indicates significant difference (p ≤ 0.05) between water treatment and drought treatment within one Pb level.
Fig. 4The effects of Pb pollution on photosynthesis of parent and offspring shoots of Phragmites australis in flood and dry environment (mean ± SE, n = 16). Different lowercase letters indicate significant differences (p ≤ 0.05) between Pb levels within one water treatment level, and * indicates significant difference (p ≤ 0.05) between water treatment and drought treatment within one Pb level.
Fig. 5The Pb concentrations in different parts of Phragmites australis subjected to Pb in flood and dry environment (mean ± SE, n = 4). Different lowercase letters indicate significant differences (p ≤0.05) between Pb levels within one water treatment level, and * indicates significant difference (p ≤ 0.05) between water treatment and drought treatment within one Pb level.
Table 7 表7 表7铅和水分处理对芦苇各器官中铅含量影响的双因素方差分析结果 Table 7Results of two-way ANOVA of the effect of Pb and water stress on Pb concentrations in organs of Phragmites australis
AhmadMSA, HussainM, IjazS, AlviAK (2008). Photosynthetic performance of two mung bean (Vigna radiata) cultivars under lead and copper stress. International Journal of Agriculture and Biology, 10, 167-172. [本文引用: 1]
BensonEJ (2001). Effects of Fire on Tallgrass Prairie Plant Population Dynamics. Master degree thesis, Kansas State University Manhattan. [本文引用: 1]
BensonEJ, HartnettDC (2006). The role of seed and vegetative reproduction in plant recruitment and demography in tallgrass prairie .Plant Ecology, 187, 163-178. DOI:10.1007/s11258-005-0975-yURL [本文引用: 2]
BrewerJS, BertnessMD (1996). Disturbance and intraspecific variation in the clonal morphology of salt marsh perennials .Oikos, 77, 107-116. DOI:10.2307/3545590URL [本文引用: 1]
CaoM, HuangPW, ZhangN, ChengLY, MuCS (2016). Effects of lead contamination on underground bud and output of aboveground shoots of Phragmites australis (common reed) under different water regimes. Journal of Southwest University for Nationalities (Natural Science Edition), 42(2), 131-138. DOI:10.11920/xnmdzk.2016.02.003URL [本文引用: 1] 为明确铅胁迫对不同水分生境条 件下芦苇的不同克隆繁殖构件数量和繁殖能力的影响,从芽库和子株角度,采用盆栽实验模拟芦苇的水层和干旱生境,设置五个梯度的铅胁迫浓度处理(0、 500、1500、3000和4500mg·kg~(-1))进行了研究.结果表明:分蘖节芽数在水层生境中,处理前期随着铅胁迫浓度的增加而显著减少, 而在处理中期和后期却均显著增加,在干旱生境中处理前、中、后期铅胁迫对分蘖节芽数无显著影响.根茎节芽、根茎顶芽和总芽数无论是在水层生境还是在干旱生 境,均随着铅胁迫浓度的增加而显著减少.分蘖节子株数在水层生境中,处理前期随着铅胁迫浓度的增加而显著减少,而在处理中期和后期却无显著变化,在干旱生 境中随着铅胁迫浓度的增加而显著减少.根茎节子株不存在,根茎顶子株无论是在水层生境还是在干旱生境,均随着铅胁迫浓度的增加而显著减少.总子株数在水层 生境中处理中期显著减少,处理后期无显著变化,在干旱生境中处理中期和后期均显著减少,并且总芽数+总子株数在胁迫前期和中期随铅胁迫浓度的增加显著减 少.说明不同类型芽具有不同的抵御铅胁迫的策略,分蘖节芽采取了耐性策略而不受影响甚至增加,其他类型的芽采取了逃避策略数量减少;水生生境的芦苇具有逐 步适应铅胁迫的能力,并出现了补偿生长效应,而干旱生境进一步加剧了铅胁迫的作用,铅胁迫和干旱具有协同效应. [曹明, 黄蓬万, 张娜, 程露瑶, 穆春生 (2016). 不同水分生境下铅胁迫对芦苇地下芽库及其输出子株能力的影响 西南民族大学学报(自然科学版), 42(2), 131-138.] DOI:10.11920/xnmdzk.2016.02.003URL [本文引用: 1] 为明确铅胁迫对不同水分生境条 件下芦苇的不同克隆繁殖构件数量和繁殖能力的影响,从芽库和子株角度,采用盆栽实验模拟芦苇的水层和干旱生境,设置五个梯度的铅胁迫浓度处理(0、 500、1500、3000和4500mg·kg~(-1))进行了研究.结果表明:分蘖节芽数在水层生境中,处理前期随着铅胁迫浓度的增加而显著减少, 而在处理中期和后期却均显著增加,在干旱生境中处理前、中、后期铅胁迫对分蘖节芽数无显著影响.根茎节芽、根茎顶芽和总芽数无论是在水层生境还是在干旱生 境,均随着铅胁迫浓度的增加而显著减少.分蘖节子株数在水层生境中,处理前期随着铅胁迫浓度的增加而显著减少,而在处理中期和后期却无显著变化,在干旱生 境中随着铅胁迫浓度的增加而显著减少.根茎节子株不存在,根茎顶子株无论是在水层生境还是在干旱生境,均随着铅胁迫浓度的增加而显著减少.总子株数在水层 生境中处理中期显著减少,处理后期无显著变化,在干旱生境中处理中期和后期均显著减少,并且总芽数+总子株数在胁迫前期和中期随铅胁迫浓度的增加显著减 少.说明不同类型芽具有不同的抵御铅胁迫的策略,分蘖节芽采取了耐性策略而不受影响甚至增加,其他类型的芽采取了逃避策略数量减少;水生生境的芦苇具有逐 步适应铅胁迫的能力,并出现了补偿生长效应,而干旱生境进一步加剧了铅胁迫的作用,铅胁迫和干旱具有协同效应.
DalgleishHJ, HartnettDC (2006). Below-ground bud banks increase along a precipitation gradient of the North American Great Plains: A test of the meristem limitation hypothesis .New Phytologist, 171, 81-89. DOI:10.1111/nph.2006.171.issue-1URL [本文引用: 2]
DaviesBE (1990). Lead. In: Alloway BJ ed. Heavy Metals in Soils. John Wiley & Sons,New York. 177-196. [本文引用: 1]
FernandesPM, VegaJA, JiménezE, RigolotE (2008). Fire resistance of European pines .Forest Ecology and Management, 256, 246-255. DOI:10.1016/j.foreco.2008.04.032URL [本文引用: 1] Pine resistance to low- to moderate-intensity fire arises from traits (namely related to tissue insulation from heat) that enable tree survival. Predictive models of the likelihood of tree mortality after fire are quite valuable to assist decision-making after wildfire and to plan prescribed burning. Data and models pertaining to the survival of European pines following fire are reviewed. The type and quality of the current information on fire resistance of the various European species is quite variable. Data from low-intensity fire experiments or regimes is comparatively abundant for Pinus pinaster and Pinus sylvestris, while tree survival after wildfire has been modelled for Pinus pinea and Pinus halepensis. P. pinaster and P. pinea, and Pinus canariensis in special, are better equipped to survive fire, but low-intensity fire is tolerated even by species often referred to as fire-sensitive ( P. halepensis and Pinus radiata). The relative fire resistance of European pine species is assessed on the basis of (i) morphological and experimental data, and (ii) mortality modelling that considers fire behaviour. Limitations of these approaches to rate fire resistance are discussed, and the current knowledge gaps are indicated.
HarperJL (1977). Population Biology of Plants. Academic Press, London. [本文引用: 2]
HartnettDC, SetshogoMP, DalgleishHJ (2006). Bud banks of perennial savanna grasses in Botswana .African Journal of Ecology, 44, 256-263. [本文引用: 1]
HechmiN, AissaNB, AbdenaceurHA, JedidiN (2014). Evaluating the phytoremediation potential of Phragmites australis grown in pentachlorophenol and cadmium co-?contaminated soils. Environmental Science and Pollution Research, 21, 1304-1313. [本文引用: 1]
HenryC, AmorosC (1996). Are the banks a source of recolonization after disturbance: An experiment on aquatic vegetation in a former channel of the Rh?ne River .Hydrobiologia, 330, 151-162 [本文引用: 1]
HuR, SunK, SuX, PanYX, ZhangYF, WangXP (2012). Physiological responses and tolerance mechanisms to Pb in two xerophils: Salsola passerina Bunge and Chenopodium album L Journal of Hazardous Materials, 205-206, 131-138. [本文引用: 3]
IslamE, LiuD, LiTQ, YangXE, JinXF, MahmoodQ, TianS, LiJY (2008). Effect of Pb toxicity on leaf growth, physiology and ultrastructure in the two ecotypes of Elsholtzia argyi. Journal of Hazardous Materials, 154, 914-926. [本文引用: 4]
LiZL, ZhangYT, YuDF, ZhangN, LinJX, ZhangJW, TangJH, WangJF, MuCS (2014). The influence of precipitation regimes and elevated CO2 on photosynthesis and biomass accumulation and partitioning in seedlings of the rhizomatous perennial grass Leymus chinensis. PLOS ONE, 9, e103633. DOI: 10.1371/journal.pone.0103633. DOI:10.1371/journal.pone.0103633 [本文引用: 1]
LiuB, LiuZM, WangLX, WangZN (2014). Responses of rhizomatous grass Phragmites communis to wind erosion: Effects on biomass allocation. Pant and Soil, 380, 389-398. [本文引用: 1]
MonyC, PuijalonS, BornetteG (2011). Resprouting response of aquatic clonal plants to cutting may explain their risistance to spate flooding . Flia Geobotanic, 46, 155-164. [本文引用: 1]
NishihiroJ, ArakiS, FujiwaraN, WashitaniI (2004). Germination characteristics of lakeshore plants under an arti?cially stabilizedwater regime .Aquatic Botany, 79, 333-343. [本文引用: 1]
SharmaP, DubeyRS (2005). Lead toxicity in plants .Brazilian Journal of Plant Physiology, 17(1), 35-52. [本文引用: 1]
WangJF, GaoS, LinJX, MuYG, MuCS (2010). Summer warming effects on biomass production and clonal growth of Leymus chinensis. Crop Pasture Science, 61, 670-676. [本文引用: 3]
WangPF, ZhangSH, WangC, LuJ (2012). Effects of Pb on the oxidative stress and antioxidant response in a Pb bioaccumulator plant Vallisneria natans. Ecotoxicology and Environmental Safety, 78, 28-34. [本文引用: 1]
WeisJS, WeisP (2004). Metal uptake, transport and release by wetland plants: Implications for phytoremediation and restoration .Environment International, 30, 685-700. [本文引用: 1]
WindhamL, WeisJS, WeisP (2001). Lead uptake, distribution, and effects in two dominant salt marsh macrophytes, Spartina alterniflora (cordgrass) and Phragmites australis 42, 811-816. [本文引用: 1]
YeZH, BakerAJM, WongMH, WillisAJ (1997). Zinc, lead and cadmium tolerance, uptake and accumulation by the common reed, phragmites australis(Cav.) Trin. ex Steudel. Annal of Botany, 80, 363-370. [本文引用: 2]
ZhangN, ZhangJW, YangYH, LiXY, LinJX, LiZL, ChengLY, WangJF, MuCS, WangAX (2015). Effects of lead contamination on the clonal propagative ability of Phragmites australis(common reed) grown in wet and dry environments. Plant Biology, 17, 893-903. [本文引用: 3]
ZhuTC (2004). The Bio-ecology of Leymus chinensis.Jilin Science and Technology Press, Changchun. 85-89. [本文引用: 1]
... 铅(Pb)是环境中的有毒重金属污染物之一, 当土壤中Pb含量达100-500 mg·kg-l时, 就会对植物产生毒性(Davies, 1990).例如, Pb可能引起植物体内抗氧化酶活性的改变、膜脂过氧化增强, 干扰矿质元素吸收, 抑制光合作用等(Islam et al., 2008; Hu et al., 2012; Wang et al., 2012).其中光合作用被认为是对Pb最为敏感的一个过程.Pb抑制光合作用的原因可能有: 气孔关闭、叶绿体结构破坏、光合色素合成抑制、阻碍电子运输和抑制卡尔文循环中的酶活性(Sharma & Dubey, 2005; Islam et al., 2008; Hu et al., 2012), 这些改变将最终抑制植物生长和生物量积累.生物量分配是反映植物对环境的敏感性和抵抗能力的重要指标(Liu et al., 2014).对于多年生植物来说, 地上生产力主要来源于母株和子株(Wang et al., 2010; Li et al., 2014).目前, 有关多年生植物生物量在母株和子株的分配还不清楚. ...
Fire resistance of European pines 1 2008
... 地下芽库影响着多年生植物的地上生产力和种群密度(Dalgleish & Hartnett, 2006; Wang et al., 2010).大量的地下芽储存可能促进植物对干扰的抗性和恢复速率, 例如火烧、放牧和洪水等(Hartnett et al., 2006; Fernandes et al., 2008; Mony et al., 2011).然而, 很少有人关注重金属对地下芽库的影响, 特别是对水生植物(Zhang et al., 2015; 曹明等, 2016).尽管地下芽与地上生产力密切相关, 但是附着有芽的根茎或其他多年生器官的延伸或生长以及芽输出成子株的能力也是影响地上生产力和种群密度的重要因子.所以, 在重金属污染环境中, 多年生植物克隆器官的生长需要更多关注. ...
Physiological responses and tolerance mechanisms to Pb in two xerophils: Salsola passerina Bunge and Chenopodium album L 3 2012
... 铅(Pb)是环境中的有毒重金属污染物之一, 当土壤中Pb含量达100-500 mg·kg-l时, 就会对植物产生毒性(Davies, 1990).例如, Pb可能引起植物体内抗氧化酶活性的改变、膜脂过氧化增强, 干扰矿质元素吸收, 抑制光合作用等(Islam et al., 2008; Hu et al., 2012; Wang et al., 2012).其中光合作用被认为是对Pb最为敏感的一个过程.Pb抑制光合作用的原因可能有: 气孔关闭、叶绿体结构破坏、光合色素合成抑制、阻碍电子运输和抑制卡尔文循环中的酶活性(Sharma & Dubey, 2005; Islam et al., 2008; Hu et al., 2012), 这些改变将最终抑制植物生长和生物量积累.生物量分配是反映植物对环境的敏感性和抵抗能力的重要指标(Liu et al., 2014).对于多年生植物来说, 地上生产力主要来源于母株和子株(Wang et al., 2010; Li et al., 2014).目前, 有关多年生植物生物量在母株和子株的分配还不清楚. ...
... ; Hu et al., 2012), 这些改变将最终抑制植物生长和生物量积累.生物量分配是反映植物对环境的敏感性和抵抗能力的重要指标(Liu et al., 2014).对于多年生植物来说, 地上生产力主要来源于母株和子株(Wang et al., 2010; Li et al., 2014).目前, 有关多年生植物生物量在母株和子株的分配还不清楚. ...
... 生长, 可能是因为母株生长初期主要将能量用于地下芽或根茎的生长及子株输出, 依靠资源顶性转移提高种群抵抗力, 而子株输出后具有较强的光合能力(图4E-4H), 因此储存的部分能量也可能通过根茎输送至母株, 发生资源的基性转移供母株生长(祝廷成, 2004).Pb和干旱的协同作用对母株产生了更强的毒性效应, 导致母株生长抑制和死亡, 特别是高浓度Pb处理(表2).母株的生长响应或中毒症状(例如叶片死亡和矮化生长)可能与光合过程变化相关.因为Pb可能代替必需元素如Mg、Fe或Mn, 不仅会抑制叶绿素的合成, 而且可能破坏基粒的形成、电子转移以及抑制光系统II (PSII)和卡尔文循环的酶活性, 抑制植物的光合作用和生长(Islam et al., 2008; Hu et al., 2012). ...
Effect of Pb toxicity on leaf growth, physiology and ultrastructure in the two ecotypes of Elsholtzia argyi. 4 2008
... 铅(Pb)是环境中的有毒重金属污染物之一, 当土壤中Pb含量达100-500 mg·kg-l时, 就会对植物产生毒性(Davies, 1990).例如, Pb可能引起植物体内抗氧化酶活性的改变、膜脂过氧化增强, 干扰矿质元素吸收, 抑制光合作用等(Islam et al., 2008; Hu et al., 2012; Wang et al., 2012).其中光合作用被认为是对Pb最为敏感的一个过程.Pb抑制光合作用的原因可能有: 气孔关闭、叶绿体结构破坏、光合色素合成抑制、阻碍电子运输和抑制卡尔文循环中的酶活性(Sharma & Dubey, 2005; Islam et al., 2008; Hu et al., 2012), 这些改变将最终抑制植物生长和生物量积累.生物量分配是反映植物对环境的敏感性和抵抗能力的重要指标(Liu et al., 2014).对于多年生植物来说, 地上生产力主要来源于母株和子株(Wang et al., 2010; Li et al., 2014).目前, 有关多年生植物生物量在母株和子株的分配还不清楚. ...
... ; Islam et al., 2008; Hu et al., 2012), 这些改变将最终抑制植物生长和生物量积累.生物量分配是反映植物对环境的敏感性和抵抗能力的重要指标(Liu et al., 2014).对于多年生植物来说, 地上生产力主要来源于母株和子株(Wang et al., 2010; Li et al., 2014).目前, 有关多年生植物生物量在母株和子株的分配还不清楚. ...
... 生长, 可能是因为母株生长初期主要将能量用于地下芽或根茎的生长及子株输出, 依靠资源顶性转移提高种群抵抗力, 而子株输出后具有较强的光合能力(图4E-4H), 因此储存的部分能量也可能通过根茎输送至母株, 发生资源的基性转移供母株生长(祝廷成, 2004).Pb和干旱的协同作用对母株产生了更强的毒性效应, 导致母株生长抑制和死亡, 特别是高浓度Pb处理(表2).母株的生长响应或中毒症状(例如叶片死亡和矮化生长)可能与光合过程变化相关.因为Pb可能代替必需元素如Mg、Fe或Mn, 不仅会抑制叶绿素的合成, 而且可能破坏基粒的形成、电子转移以及抑制光系统II (PSII)和卡尔文循环的酶活性, 抑制植物的光合作用和生长(Islam et al., 2008; Hu et al., 2012). ...
The influence of precipitation regimes and elevated CO2 on photosynthesis and biomass accumulation and partitioning in seedlings of the rhizomatous perennial grass 1 2014
... 铅(Pb)是环境中的有毒重金属污染物之一, 当土壤中Pb含量达100-500 mg·kg-l时, 就会对植物产生毒性(Davies, 1990).例如, Pb可能引起植物体内抗氧化酶活性的改变、膜脂过氧化增强, 干扰矿质元素吸收, 抑制光合作用等(Islam et al., 2008; Hu et al., 2012; Wang et al., 2012).其中光合作用被认为是对Pb最为敏感的一个过程.Pb抑制光合作用的原因可能有: 气孔关闭、叶绿体结构破坏、光合色素合成抑制、阻碍电子运输和抑制卡尔文循环中的酶活性(Sharma & Dubey, 2005; Islam et al., 2008; Hu et al., 2012), 这些改变将最终抑制植物生长和生物量积累.生物量分配是反映植物对环境的敏感性和抵抗能力的重要指标(Liu et al., 2014).对于多年生植物来说, 地上生产力主要来源于母株和子株(Wang et al., 2010; Li et al., 2014).目前, 有关多年生植物生物量在母株和子株的分配还不清楚. ...
Responses of rhizomatous grass Phragmites communis to wind erosion: Effects on biomass allocation. 1 2014
... 铅(Pb)是环境中的有毒重金属污染物之一, 当土壤中Pb含量达100-500 mg·kg-l时, 就会对植物产生毒性(Davies, 1990).例如, Pb可能引起植物体内抗氧化酶活性的改变、膜脂过氧化增强, 干扰矿质元素吸收, 抑制光合作用等(Islam et al., 2008; Hu et al., 2012; Wang et al., 2012).其中光合作用被认为是对Pb最为敏感的一个过程.Pb抑制光合作用的原因可能有: 气孔关闭、叶绿体结构破坏、光合色素合成抑制、阻碍电子运输和抑制卡尔文循环中的酶活性(Sharma & Dubey, 2005; Islam et al., 2008; Hu et al., 2012), 这些改变将最终抑制植物生长和生物量积累.生物量分配是反映植物对环境的敏感性和抵抗能力的重要指标(Liu et al., 2014).对于多年生植物来说, 地上生产力主要来源于母株和子株(Wang et al., 2010; Li et al., 2014).目前, 有关多年生植物生物量在母株和子株的分配还不清楚. ...
Resprouting response of aquatic clonal plants to cutting may explain their risistance to spate flooding 1 2011
... 地下芽库影响着多年生植物的地上生产力和种群密度(Dalgleish & Hartnett, 2006; Wang et al., 2010).大量的地下芽储存可能促进植物对干扰的抗性和恢复速率, 例如火烧、放牧和洪水等(Hartnett et al., 2006; Fernandes et al., 2008; Mony et al., 2011).然而, 很少有人关注重金属对地下芽库的影响, 特别是对水生植物(Zhang et al., 2015; 曹明等, 2016).尽管地下芽与地上生产力密切相关, 但是附着有芽的根茎或其他多年生器官的延伸或生长以及芽输出成子株的能力也是影响地上生产力和种群密度的重要因子.所以, 在重金属污染环境中, 多年生植物克隆器官的生长需要更多关注. ...
Germination characteristics of lakeshore plants under an arti?cially stabilizedwater regime 1 2004
... 铅(Pb)是环境中的有毒重金属污染物之一, 当土壤中Pb含量达100-500 mg·kg-l时, 就会对植物产生毒性(Davies, 1990).例如, Pb可能引起植物体内抗氧化酶活性的改变、膜脂过氧化增强, 干扰矿质元素吸收, 抑制光合作用等(Islam et al., 2008; Hu et al., 2012; Wang et al., 2012).其中光合作用被认为是对Pb最为敏感的一个过程.Pb抑制光合作用的原因可能有: 气孔关闭、叶绿体结构破坏、光合色素合成抑制、阻碍电子运输和抑制卡尔文循环中的酶活性(Sharma & Dubey, 2005; Islam et al., 2008; Hu et al., 2012), 这些改变将最终抑制植物生长和生物量积累.生物量分配是反映植物对环境的敏感性和抵抗能力的重要指标(Liu et al., 2014).对于多年生植物来说, 地上生产力主要来源于母株和子株(Wang et al., 2010; Li et al., 2014).目前, 有关多年生植物生物量在母株和子株的分配还不清楚. ...
Summer warming effects on biomass production and clonal growth of 3 2010
... 铅(Pb)是环境中的有毒重金属污染物之一, 当土壤中Pb含量达100-500 mg·kg-l时, 就会对植物产生毒性(Davies, 1990).例如, Pb可能引起植物体内抗氧化酶活性的改变、膜脂过氧化增强, 干扰矿质元素吸收, 抑制光合作用等(Islam et al., 2008; Hu et al., 2012; Wang et al., 2012).其中光合作用被认为是对Pb最为敏感的一个过程.Pb抑制光合作用的原因可能有: 气孔关闭、叶绿体结构破坏、光合色素合成抑制、阻碍电子运输和抑制卡尔文循环中的酶活性(Sharma & Dubey, 2005; Islam et al., 2008; Hu et al., 2012), 这些改变将最终抑制植物生长和生物量积累.生物量分配是反映植物对环境的敏感性和抵抗能力的重要指标(Liu et al., 2014).对于多年生植物来说, 地上生产力主要来源于母株和子株(Wang et al., 2010; Li et al., 2014).目前, 有关多年生植物生物量在母株和子株的分配还不清楚. ...
... 地下芽库影响着多年生植物的地上生产力和种群密度(Dalgleish & Hartnett, 2006; Wang et al., 2010).大量的地下芽储存可能促进植物对干扰的抗性和恢复速率, 例如火烧、放牧和洪水等(Hartnett et al., 2006; Fernandes et al., 2008; Mony et al., 2011).然而, 很少有人关注重金属对地下芽库的影响, 特别是对水生植物(Zhang et al., 2015; 曹明等, 2016).尽管地下芽与地上生产力密切相关, 但是附着有芽的根茎或其他多年生器官的延伸或生长以及芽输出成子株的能力也是影响地上生产力和种群密度的重要因子.所以, 在重金属污染环境中, 多年生植物克隆器官的生长需要更多关注. ...
Effects of Pb on the oxidative stress and antioxidant response in a Pb bioaccumulator plant 1 2012
... 铅(Pb)是环境中的有毒重金属污染物之一, 当土壤中Pb含量达100-500 mg·kg-l时, 就会对植物产生毒性(Davies, 1990).例如, Pb可能引起植物体内抗氧化酶活性的改变、膜脂过氧化增强, 干扰矿质元素吸收, 抑制光合作用等(Islam et al., 2008; Hu et al., 2012; Wang et al., 2012).其中光合作用被认为是对Pb最为敏感的一个过程.Pb抑制光合作用的原因可能有: 气孔关闭、叶绿体结构破坏、光合色素合成抑制、阻碍电子运输和抑制卡尔文循环中的酶活性(Sharma & Dubey, 2005; Islam et al., 2008; Hu et al., 2012), 这些改变将最终抑制植物生长和生物量积累.生物量分配是反映植物对环境的敏感性和抵抗能力的重要指标(Liu et al., 2014).对于多年生植物来说, 地上生产力主要来源于母株和子株(Wang et al., 2010; Li et al., 2014).目前, 有关多年生植物生物量在母株和子株的分配还不清楚. ...
Metal uptake, transport and release by wetland plants: Implications for phytoremediation and restoration 1 2004
 ,4,*, 张振华1,*1
,4,*, 张振华1,*1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT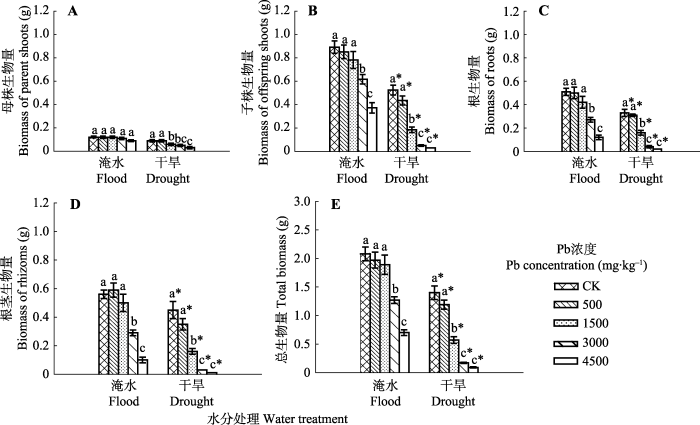 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT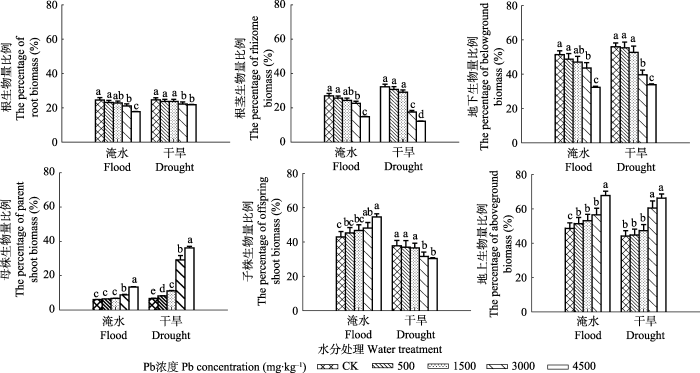 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT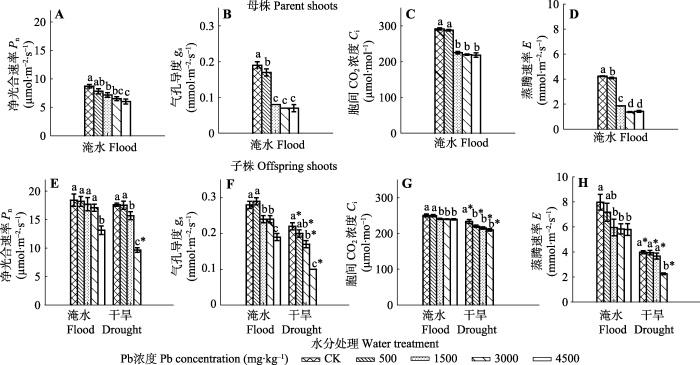 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT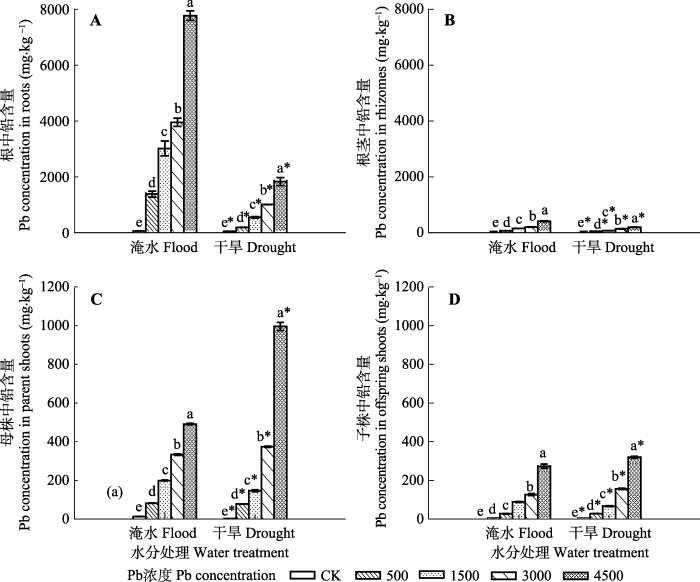 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}