Effects of vegetation restoration on soil organic carbon concentration and density in the mid-subtropical region of China
GU Xiang1, ZHANG Shi-Ji1, LIU Zhao-Dan1, LI Lei-Da1, CHEN Jin-Lei1, WANG Liu-Fang1, FANG Xi,,1,2,3,*1 School of Life Science and Technology, Central South University of Forestry and Technology, Changsha 410004, China 2 National Engineering Laboratory of South China Forestry Ecology Applicable Technologies, Changsha 410004, China 3 Huitong National Field Station for Scientific Observation and Research of Chinese Fir Plantation Ecosystem of Hunan Province, Huitong, Hunan 438107, China
Supported by the National Forestry Public Welfare Industry Research Project.201504411 the National Natural Science Foundation of China.30771720 the National Natural Science Foundation of China.31170426
Abstract Aims Vegetation restoration plays an important role in the accumulation and storage of soil organic carbon (SOC). Our objectives were to investigate the effects of vegetation restoration on SOC and to explain the underlying mechanisms of carbon sequestration during vegetation restoration in the mid-subtropical China. Methods According to the disturbance intensity and the degree of restoration, we used the space-for-time substitution method by selecting four different types of vegetation communities, composed of Loropetalum chinense-Vaccinium bracteatum-Rhododendron simsii scrub-grass-land (LVR), Loropetalum chinense-Cunninghamia lanceolata-Quercus fabri shrubbery (LCQ), Pinus massoniana-Lithocarpus glaber-Loropetalum chinense coniferous-broad leaved mixed forest (PLL), and Lithocarpus glaber-Cleyera japonica-Cyclobalanopsis glauca evergreen broad-leaved forest (LAG) to represent the successional sequence in the secondary forests in Changsha County, Hunan Province, China. Permanent plots were established in each vegetation communities. Soil samples (0-40 cm) were collected and divided into four layers (0-10, 10-20, 20-30 and 30-40 cm). Soil organic carbon concentration (CSOC) and soil organic carbon density (DSOC) were measured. The main influencing factors on CSOC and DSOC were analyzed with Principal Component Analysis and Stepwise Regressions Analysis. Important findings 1) Along vegetation restoration, CSOC and DSOC increased dramatically. The CSOC was the highest in LAG, which was 12.5, 9.3 and 4.7 g·kg -1 higher than in LVR, LCQ and PLL in 0-40 cm soil depth, increasing by 248.5%, 113.1% and 58.5%, respectively. The increments of DSOC in LAG at 0-40 cm soil depth were 67.1, 46.1 and 32.5 t C·hm -2, and increased by 182.0%, 79.7% and 45.6% compared to DSOC in LVR, LCQ and PLL, respectively. 2) Correlation analysis showed that CSOC and DSOC were strongly and positively correlated with species diversity index, community total biomass, aboveground biomass, root biomass, existing biomass in litter layer, nitrogen (N), phosphorus (P) concentration in litter layer, soil total P, soil available P, soil C/N ratio (except CSOC), soil C/P ratio, soil N/P ratio and percentage of soil clay (< 0.002 mm), but significantly and negatively correlated with C/N in litter layer (except DSOC), C/P in litter layer, soil pH and soil bulk density, suggesting that the differences in CSOC and DSOC under different vegetation stages were related to both vegetation and soil properties. 3) The results of principal component analysis and stepwise regression analysis revealed that soil C/P, pH, concentration of soil clay (except CSOC) and C/P in litter layer were the dominant factors affecting CSOC and DSOC during vegetation restoration. Among them, soil C/P ratio ranked first. These results indicated that the differences in soil C/P ratio, pH, soil clay concentration and C/P in litter layer were responsible for the changes in SOC during vegetation restoration. Keywords:central hilly area of Hunan Province;vegetation restoration;scrub-grass-land;shrubbery;coniferous- broad leaved mixed forest;evergreen broad-leaved forest;vegetation factor;soil factor
PDF (1108KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 辜翔, 张仕吉, 刘兆丹, 李雷达, 陈金磊, 王留芳, 方晰. 中亚热带植被恢复对土壤有机碳含量、碳密度的影响. 植物生态学报[J], 2018, 42(5): 595-608 DOI:10.17521/cjpe.2018.0021 GU Xiang, ZHANG Shi-Ji, LIU Zhao-Dan, LI Lei-Da, CHEN Jin-Lei, WANG Liu-Fang, FANG Xi. Effects of vegetation restoration on soil organic carbon concentration and density in the mid-subtropical region of China. Chinese Journal of Plant Ecology[J], 2018, 42(5): 595-608 DOI:10.17521/cjpe.2018.0021
Table 1 表1 表1不同植被恢复阶段样地的基本特征 Table 1Stand characteristics at different stages during vegetation restoration
恢复阶段 Restoration stage
优势植物 Dominant plants
组成比例 Composition proportion (%)
木本植物密度 Density of woody plants (trees·hm-2)
多样性 指数 Diversity index
重要值Important value (%)
平均胸径 Average DBH (cm)
平均树高 Average tree height (m)
海拔 Elevation (m)
坡向 Slope aspect
坡度 Slope
檵木-南烛- 杜鹃灌草丛 LVR
檵木 Loropetalum chinense
34.48
18125
0.87
27.46
-
0.85 (0.3-1.8)
120-131
东南 Southeast
18°
南烛 Vaccinium bracteatum
21.55
18.96
杜鹃 Rhododendron simsii
12.07
14.00
白栎 Quercus fabri
7.76
10.66
板栗 Castanea mollissima
5.17
2.72
其他(8种) Others (8 species)
18.97
26.20
檵木-杉木- 白栎灌木林 LCQ
檵木 Loropetalum chinense
17.47
7633
1.06
22.02
2.74 (1.0-9.8)
3.37 (1.5-6.5)
120-135
西北 Northwest
22°
杉木 Cunninghamia lanceolata
14.85
13.16
白栎 Quercus fabri
12.66
11.10
南烛 Vaccinium bracteatum.
12.66
9.26
木姜子 Litsea spp.
11.35
10.13
其他(16种) Others (16 species)
31.01
34.43
马尾松-柯- 檵木针阔 混交林 PLL
马尾松 Pinus massoniana
39.69
17629
1.91
45.34
5.70 (1.0-28.0)
6.54 (1.5-20.0)
135-160
西南 Southwest
20°
柯 Lithocarpus glaber
25.52
13.87
檵木 Loropetalum chinense
11.06
7.43
红淡比 Cleyera japonica
3.59
4.11
连蕊茶 Camellia cuspidata
3.11
4.02
其他(22种) Others (22 species)
17.03
25.23
柯-红淡比- 青冈常绿 阔叶林 LAG
柯 Lithocarpus glaber
38.78
19970
2.29
25.75
5.63 (1.0-40.0)
5.75 (1.5-20.0)
200-260
东南 Southeast
22°
红淡比 Cleyera japonica
18.70
11.05
青冈 Cyclobalanopsis glauca
5.82
8.90
杉木 Cunninghamia lanceolata
5.36
6.14
格药柃 Eurya muricata Dunn
5.06
5.73
其他(31种) Others (31 species)
26.28
42.44
Values in parenthesis are the range of tree diameter at breast height (DBH) or height of each forest. LAG, Lithocarpus glaber-Cleyera japonica-Cyclobalanopsis glauca evergreen broad-leaved forest; LCQ, Loropetalum chinense-Cunninghamia lanceolata-Quercus fabri shrubbery; LVR, Loropetalum chinense-Vaccinium bracteatum- Rhododendron simsii scrub-grass-land; PLL, Pinus massoniana-Lithocarpus glaber-Loropetalum chinense coniferous-broad leaved mixed forest. 括号内的数据为每个群落中树木胸径或树高的变化范围。
Table 2 表2 表2不同植被恢复阶段的群落生物量和凋落物层养分特征(平均值±标准偏差) Table 2Plant community biomass and nutrient characteristics of litter layer at different stages during vegetation restoration (mean ± SD)
恢复阶段 Restoration stage
群落总生物量 Community total biomass (kg·hm-2)
地上部分 生物量 Aboveground biomass (kg·hm-2)
根系生物量 Root biomass (kg·hm-2)
凋落物层 现存量 Existing biomass in litter layer (kg·hm-2)
凋落物层 C含量 C concentration in litter layer (g·kg-1)
凋落物层 N含量 N concentration in litter layer (g·kg-1)
凋落物层 P含量 P concentration in litter layer (g·kg-1)
凋落物层 C/N比 C/N ratio in litter layer
凋落物层 C/P比 C/P ratio in litter layer
凋落物层N/P比 N/P ratio in litter layer
LVQ
5 185.4 ± 2 702.5a
2 244.5 ± 1 153.9a
2 029.6 ± 1 515.7a
911.4 ± 653. 9a
315.8 ± 25.3ac
9.84 ± 0.3a
0.30 ± 0.02a
35.8 ± 2.6a
1532.2 ± 95.2a
41.5 ± 2.9a
LCQ
2 0591.5 ± 9 728.9b
10 346.7 ± 7 143.2b
5 443.0 ± 3 467.5a
4 801.8 ± 1 030.2b
277.2 ± 33.3a
11.2 ± 0.9a
0.34 ± 0.03ab
26.0 ± 0.2b
914.2 ± 69.4b
35.7 ± 3.6a
PLL
129 112.7 ± 19 713.5c
107 692.6 ± 16 361.6c
16 332.9 ± 3 047.6b
5 087.2 ± 1 246.1b
424.1 ± 8.6b
11.5 ± 0.5a
0.33 ± 0.05ab
38.0 ± 1.6a
1389.7 ± 227.9ab
36.6 ± 4.4a
LAG
148 975.4 ± 43 906.4c
120 695.2 ± 13 647.7c
24 400.4 ± 5 124.8c
3 879.8 ± 1 171.5b
332.3 ± 53.9c
14.0 ± 0.2b
0.38 ± 0.04b
23.9 ± 3.5b
953.1 ± 44.6b
40.3 ± 4.0a
See Table 1for LVQ, LCQ, PLL, LAG. Different letters indicate significant differences among different vegetation restoration stages (p < 0.05). LVQ、LCQ、PLL、LAG同表1。不同字母表示不同恢复阶段之间差异显著(p < 0.05)。
<0.002 mm 黏粒百分含量 Soil clay (<0.002 mm) percentage (%)
pH
TP (g·kg-1)
AP (mg·kg-1)
C/N
C/P
N/P
LVR
0-10
1.4 ± 0.2Aa
43.6 ± 4.8Aa
44.7 ± 6.4Aa
11.7 ± 4.6Aa
4.5 ± 0.2Aa
0.17 ± 0.16Aa
2.1 ± 0.7Aa
22.4 ± 12.5Aa
109.1 ± 44.7Aa
5.3 ± 2.4Aa
10-20
1.5 ± 0.1Aab
39.5 ± 4.1Aa
57.0 ± 5.2Ab
3.5 ± 1.1Ab
4.7 ± 0.2Ab
0.12 ± 0.03Aab
1.4 ± 0.6Ab
20.0 ± 9.3Aab
59.5 ± 51.4Ab
3.3 ± 3.0Ab
20-30
1.5 ± 0.1Aab
42.0 ± 5.6Aa
56.5 ± 6.2Ab
2.7 ± 0.9Ab
4.8 ± 0.2Ab
0.11 ± 0.04Ab
1.3 ± 0.5Ab
17.7 ± 9.5Aab
42.9 ± 40.9Ab
2.5 ± 2.2Ab
30-40
1.5 ± 0.1Ab
43.1 ± 5.6Aa
54.4 ± 6.1ACb
2.5 ± 0.6Ab
5.0 ± 0.2Ac
0.11 ± 0.04Ab
1.2 ± 0.5Ab
13.7 ± 7.7ABb
30.5 ± 37.4Ab
2.5 ± 2.5Ab
LCQ
0-10
1.4 ± 0.1Ab
67.9 ± 2.1Ba
21.6 ± 2.5Ba
10.6 ± 2.4Aa
4.8 ± 0.2Ba
0.12 ± 0.03Aa
2.6 ± 1.1ABa
17.7 ± 6.8Aa
164.4 ± 45.5Ba
10.0 ± 3.1Ba
10-20
1.6 ± 0.1Ab
60.1 ± 11.1Bb
27.2 ± 9.1Bb
12.9 ± 5.0Ba
4.9 ± 0.2ABab
0.10 ± 0.03Ab
1.9 ± 0.6Bab
16.3 ± 4.6ABab
90.2 ± 29.9Bb
6.0 ± 2.8Bb
20-30
1.6 ± 0.1Ab
63.3 ± 3.9Bab
26.0 ± 4.9Bab
10.7 ± 3.5Ba
5.0 ± 0.2ABb
0.09 ± 0.02Ab
1.9 ± 0.7Bb
17.0 ± 7.5ABab
72.4 ± 47.2Bb
4.8 ± 2.84Bbc
30-40
1.7 ± 0.0Bb
63.4 ± 3.7Bab
25.8 ± 5.0Bab
10.8 ± 3.4Ba
5.1 ± 0.2Ab
0.10 ± 0.03Ab
1.6 ± 0.7ABb
12.5 ± 3.2Ab
40.7 ± 17.7ABc
3.5 ± 1.6ABc
PLL
0-10
1.2 ± 0.3Aa
49.0 ± 18.5Aa
35.2 ± 14.0Ca
15.8 ± 6.3Ba
4.4 ± 0.2Ba
0.15 ± 0.05Aa
2.5 ± 0.9ABa
21.5 ± 3.4Aa
217.6 ± 54.6Ca
10.2 ± 2.7Ba
10-20
1.4 ± 0.2Aa
45.2 ± 19.3Aa
43.3 ± 18.0Ca
11.6 ± 8.3Ba
4.5 ± 0.3Bab
0.13 ± 0.06Aa
2.0 ± 0.4BCab
20.1 ±6.5Aab
103.6 ± 33.3Bb
5.5 ± 2.1Bb
20-30
1.5 ± 0.2Aa
43.0 ± 18.2Aa
45.6 ± 19.4Ca
11.5 ± 6.8Ba
4.6 ± 0.3Bab
0.11 ± 0.06Aa
1.7 ± 0.8ABb
16.6 ± 4.9ABb
72.7 ± 26.5Bc
4.5 ± 1.7Bbc
30-40
1.5 ± 0.1Aa
41.2 ± 18.7Aa
48.8 ± 19.7Aa
10.1 ± 7.9Ba
4.7 ± 0.3Ab
0.13 ± 0.08Aa
2.0 ± 1.5Bab
17.1 ± 5.6Bb
46.8 ± 22.4ABc
3.1 ± 2.1Ac
LAG
0-10
1.3 ± 0.1Aa
26.1 ± 7.4Ca
56.5 ± 8.4Da
17.4 ± 4.2Ba
4.4 ± 0.3Ca
0.25 ± 0.05Ba
2.8 ± 0.9Ba
16.5 ± 2.9Aa
151.4 ± 55.8Ba
8.6 ± 3.7Ba
10-20
1.4 ± 0.0Ab
20.1 ± 3.6Cb
61.2 ± 4.8Dab
18.8 ± 4.8Ca
4.6 ± 0.2Ca
0.20 ± 0.04Bb
2.5 ± 0.9Cab
13.2 ± 1.2Bb
84.7 ± 27.3ABb
6.5 ± 2.1Bb
20-30
1.5 ± 0.0Ab
21.8 ± 5.2Cab
62.9 ± 5.0Ab
15.3 ± 4.3Cab
4.6 ± 0.2Ca
0.19 ± 0.04Bb
2.0 ± 1.0Bb
11.9 ± 1.6Bb
67.4 ± 25.0ABb
5.6 ± 1.8Bb
30-40
1.5 ± 0.0Ab
23.9 ± 6.3Cab
62.5 ± 7.3Cb
13.6 ± 4.5Bb
4.6 ± 0.3Ca
0.20 ± 0.05Bb
1.9 ± 0.9ABb
12.0 ± 1.7Ab
63.3 ± 31.3Bb
5.2 ± 2.4Bb
AP, soil available phosphorus concentration; C/N, ratio of carbon to nitrogen in soil; C/P, ratio of carbon to phosphorus in soil; N/P, ratio of nitrogen to phosphorus in soil; pH, soil pH value; TP, soil total phosphorus concentration. See Table 1 for restoration stage. Different capital letters represent significant differences between different vegetation restoration stages in the same soil layer (p < 0.05), and different lowercase letters indicate significant differences between different soil layers at the same vegetation restoration stage (p < 0.05). AP, 土壤速效磷含量; C/N, 土壤碳氮比; C/P, 土壤碳磷比; N/P, 土壤氮磷比; pH, 土壤pH值; TP, 土壤全磷含量。恢复阶段同表1。不同大写字母表示同一土层不同恢复阶段之间差异显著(p < 0.05), 不同小写字母表示同一恢复阶段不同土层之间差异显著(p < 0.05)。
Fig. 1Soil organic carbon concentration at different stages during vegetation restoration (mean ± SD). LVR, Loropetalum chinense-Vaccinium bracteatum-Rhododendron simsii scrub- grass-land; LCQ, Loropetalum chinense-Cunninghamia lanceolata-Quercus fabri shrubbery; PLL, Pinus massoniana- Lithocarpus glaber-Loropetalum chinense coniferous-broad leaved mixed forest; LAG, Lithocarpus glaber-Cleyera japonica- Cyclobalanopsis glauca evergreen broad-leaved forest. Different capital letters represent significant differences among different vegetation restoration stages in the same soil layer (p < 0.05), and different lower letters indicate significant differences among different soil layers at the same vegetation restoration stage (p < 0.05).
AertsR ( 1997). Climate, leaf litter chemistry and leaf litter decomposition in terrestrial ecosystems: A triangular relationship Oikos, 79, 439-449. DOIURL [本文引用: 1]
BergerTW, NeubauerC, GlatzelG ( 2002). Factors controlling soil carbon and nitrogen stores in pure stands of Norway spruce (Picea abies) and mixed species stands in Austria Forest Ecology and Management, 159, 3-14. DOIURL [本文引用: 1] Soil data of 18 pairs of secondary pure spruce stands ( Picea abies) and adjacent mixed stands of spruce and broadleaf trees (at least 40% broadleaf trees, mainly beech, Fagus sylvatica) on comparable sites were used to study effects of species composition and other driving forces on carbon and nitrogen stores. It was hypothesized that carbon input by litter and roots to different soil horizons has a marked effect on carbon and nitrogen stores and, therefore, silvicultural methods, e.g., admixture of beech versus spruce, can be used to manage nitrogen retention and release. The study sites of different stand age (pole to mature stage) were chosen on two different bedrock materials (Flysch and Molasse). In general, soils on Flysch are less acidic, better supplied with nutrients, and less sandy than soils developed on Molasse. Species composition did not affect total stores (forest floor and 0 50 cm mineral soil) of carbon or nitrogen on Molasse, however, paired samples tests indicated that mean total stores were significantly higher both for carbon and nitrogen for pure spruce stands than for mixed species stands on Flysch. Carbon stores of the whole soil profile were best predicted by admixture of spruce, stores of phosphorus, sodium and calcium as well as stand age on Flysch; on Molasse, the only given predictor was aluminum storage, indicating that acidic soil conditions favor accumulation of organic carbon. On Flysch, total nitrogen stores were best explained by stores of sulfur and of the sum of base cations.
BiJD, LiYL, NingZY, ZhaoXY ( 2016). Carbon mineralization and decomposition of litters from dominant plants in the Horqin Sandy Land: Effect of litter quality Journal of Desert Research, 36, 85-92. [本文引用: 1]
CaoJ, YanWD, XiangWH, ChenXY, LeiPF ( 2015). Stoichiometry characterization of soil C, N and P of Chinese fir plantations at three different ages in Huitong, Hunan Province, China Scientia Silvae Sinicae, 51(7), 1-8. DOIURLMagsci [本文引用: 1] <p><strong>[目的]</strong> 研究不同林龄(7,17和25年生)杉木人工林土壤有机碳、全氮和全磷含量及其生态化学计量特征,探讨土壤层次分布规律及随年龄的变化规律,以期为杉木人工林可持续经营提供理论依据。<strong>[方法]</strong> 以湖南会同杉木林生态系统国家野外科学观测研究站基地内3块不同林龄的杉木人工林为研究对象,在每个林分内设置3块20 m×20 m样地,按照随机、等量和多点混合的原则,每块样地内采用"品"字形布点,按0~20, 20~40 和40~60 cm土层取样,测定土壤有机碳、全氮和全磷含量,并计算土壤碳、氮、磷之间的计量比。<strong>[结果]</strong> 7,17和25年生杉木人工林土壤有机碳含量为12.14~19.52 g·kg<sup>-1</sup>,全氮含量为1.43~1.89 g·kg<sup>-1</sup>,全磷含量为0.32~0.40 g·kg<sup>-1</sup>; 3个林分土壤有机碳含量和全氮含量随着土壤深度(0~60 cm)增加而逐渐降低, 全磷含量在土壤垂直结构上的差异不明显; 3个林龄杉木人工林的土壤C:N随林龄增加而下降,分别为11.73,8.70和8.52,对照样地炼山造林前土壤C:N为9.52,7年生幼龄林显著高于对照样地,对照样地显著高于17年生中龄林和25年生近熟林(<em>P</em><0.05); 土壤C:P分别为57.42,36.27和40.54,对照样地为55.70,低于我国平均值61,7年生幼龄林、对照样地显著高于17年生中龄林和25年生近熟林(<em>P</em><0.05); 土壤N:P分别为4.87,4.18,4.76,差异不显著(<em>P</em>>0.05),低于我国土壤平均值5.2,而炼山造林前为5.85,显著高于3个林龄杉木人工林土壤N:P(<em>P</em><0.05); 3个林龄杉木人工林土壤C:N:P分别为56:5:1,36:4:1和41:5:1,炼山造林前为55:6:1,均低于我国平均值(60:5:1)。<strong>[结论]</strong> 通过对不同林龄杉木人工林土壤碳、氮、磷含量的研究发现,随着炼山后造林时间的推移,土壤有机碳、全氮、全磷含量逐渐得到恢复。本研究杉木人工林土壤中的氮含量相对较高,而磷含量较为缺乏,为了达到土壤中磷的相对平衡,在土壤C:P较低的情况下土壤微生物体有机磷会出现净矿化现象。3个林龄杉木人工林土壤C:N,N:P和C:P在不同土壤剖面深度上变化不大,相对稳定。该研究区土壤C:N和C:P主要受土壤有机碳的影响,因此在杉木人工林经营管理过程中如何调控土壤中机碳含量显得十分关键。</p> [ 曹娟, 闫文德, 项文化, 谌小勇, 雷丕锋 ( 2015). 湖南会同3个林龄杉木人工林土壤碳、氮、磷化学计量特征 林业科学, 51(7), 1-8.] DOIURLMagsci [本文引用: 1] <p><strong>[目的]</strong> 研究不同林龄(7,17和25年生)杉木人工林土壤有机碳、全氮和全磷含量及其生态化学计量特征,探讨土壤层次分布规律及随年龄的变化规律,以期为杉木人工林可持续经营提供理论依据。<strong>[方法]</strong> 以湖南会同杉木林生态系统国家野外科学观测研究站基地内3块不同林龄的杉木人工林为研究对象,在每个林分内设置3块20 m×20 m样地,按照随机、等量和多点混合的原则,每块样地内采用"品"字形布点,按0~20, 20~40 和40~60 cm土层取样,测定土壤有机碳、全氮和全磷含量,并计算土壤碳、氮、磷之间的计量比。<strong>[结果]</strong> 7,17和25年生杉木人工林土壤有机碳含量为12.14~19.52 g·kg<sup>-1</sup>,全氮含量为1.43~1.89 g·kg<sup>-1</sup>,全磷含量为0.32~0.40 g·kg<sup>-1</sup>; 3个林分土壤有机碳含量和全氮含量随着土壤深度(0~60 cm)增加而逐渐降低, 全磷含量在土壤垂直结构上的差异不明显; 3个林龄杉木人工林的土壤C:N随林龄增加而下降,分别为11.73,8.70和8.52,对照样地炼山造林前土壤C:N为9.52,7年生幼龄林显著高于对照样地,对照样地显著高于17年生中龄林和25年生近熟林(<em>P</em><0.05); 土壤C:P分别为57.42,36.27和40.54,对照样地为55.70,低于我国平均值61,7年生幼龄林、对照样地显著高于17年生中龄林和25年生近熟林(<em>P</em><0.05); 土壤N:P分别为4.87,4.18,4.76,差异不显著(<em>P</em>>0.05),低于我国土壤平均值5.2,而炼山造林前为5.85,显著高于3个林龄杉木人工林土壤N:P(<em>P</em><0.05); 3个林龄杉木人工林土壤C:N:P分别为56:5:1,36:4:1和41:5:1,炼山造林前为55:6:1,均低于我国平均值(60:5:1)。<strong>[结论]</strong> 通过对不同林龄杉木人工林土壤碳、氮、磷含量的研究发现,随着炼山后造林时间的推移,土壤有机碳、全氮、全磷含量逐渐得到恢复。本研究杉木人工林土壤中的氮含量相对较高,而磷含量较为缺乏,为了达到土壤中磷的相对平衡,在土壤C:P较低的情况下土壤微生物体有机磷会出现净矿化现象。3个林龄杉木人工林土壤C:N,N:P和C:P在不同土壤剖面深度上变化不大,相对稳定。该研究区土壤C:N和C:P主要受土壤有机碳的影响,因此在杉木人工林经营管理过程中如何调控土壤中机碳含量显得十分关键。</p>
CastroH, FortunelC, FreitasH ( 2010). Effects of land abandonment on plant litter decomposition in a Montado system: Relation to litter chemistry and community functional parameters Plant and Soil, 333, 181-190. DOIURL [本文引用: 1] Changes in land use and subsequent shifts in vegetation can influence decomposition through changes in litter quality (chemistry and structure) and alterations of soil temperature and moisture. Our aim was to study the effects of land abandonment on litter decomposition in a Mediterranean area of Montado, South Portugal. We tested the hypothesis that decomposition tends to slow down with abandonment, as woody species, richer in lignified structures, replace herbaceous species. We assessed the decomposition of community litter in situ using litterbag technique. To test the influence of local conditions, we simultaneously incubated a standard litter in situ. Our results showed that the shift from herbaceous to shrub-dominated communities lead to decreased decomposition rates. Changes in litter decomposition were primarily driven by changes in litter quality, even though the uneven pattern of litter mass loss over the experiment might reveal an effect from possible differences in microclimate. Shrub litter had higher nutrient content than herbaceous litter, which seemed to favour higher initial decomposition rates, but lower decomposition rate in the longer term. Shrubs also contribute to woody litter, richer in lignin, and secondary compounds that retard decomposition, and may play a role in increasing pools of slowly decomposing organic matter.
ChenJ, YangN ( 2016). Effect of re-vegetation on active organic carbon pool of purple soils on sloping-land in Hengyang, Hunan Province, China Journal of Tropical and Subtropical Botany, 24, 568-576. DOIURL [本文引用: 2] 为了解植被恢复对土壤活性有机碳库的影响,采用空间代替时间序列方法,对衡阳紫色土丘陵坡地植被恢复对土壤活性有机碳含量和分布进行研究.结果表明,土壤有机碳(SOC)、微生物量碳(MBC)、水溶性有机碳(DOC)、轻组有机碳(LFOC)和易氧化碳(ROC)含量均以乔灌阶段最高(P<0.05),随土层加深显著减小(P<0.05).随恢复进程,MBC/SOC、DOC/SOC和ROC/SOC显著增加(P<0.05);0~20 cm土层,LFOC/SOC随恢复显著增加(P<0.05),而20~40 cm土层,LFOC/SOC的差异不明显(P>0.05).随土层加深,LFOC/SOC显著减小(P<0.05),DOC/SOC、MBC/SOC和ROC/SOC逐渐增加(P>0.05).SOC、MBC、DOC、LFOC和ROC间存在极显著正相关关系(P<0.01).各类活性有机碳库与土壤含水量(SwC)、全氮(TN)、碱解氮(AN)和速效磷(AP)呈显著或极显著正相关关系(P<0.05或P<0.01),与土壤容重(BD)呈极显著负相关关系.因此,植被恢复在一定程度上可提高衡阳紫色土丘陵坡地土壤活性有机碳的形成和积累,增加土壤碳储量. [ 陈璟, 杨宁 ( 2016). 衡阳紫色土丘陵坡地植被恢复对土壤活性有机碳库的影响 热带亚热带植物学报, 24, 568-576.] DOIURL [本文引用: 2] 为了解植被恢复对土壤活性有机碳库的影响,采用空间代替时间序列方法,对衡阳紫色土丘陵坡地植被恢复对土壤活性有机碳含量和分布进行研究.结果表明,土壤有机碳(SOC)、微生物量碳(MBC)、水溶性有机碳(DOC)、轻组有机碳(LFOC)和易氧化碳(ROC)含量均以乔灌阶段最高(P<0.05),随土层加深显著减小(P<0.05).随恢复进程,MBC/SOC、DOC/SOC和ROC/SOC显著增加(P<0.05);0~20 cm土层,LFOC/SOC随恢复显著增加(P<0.05),而20~40 cm土层,LFOC/SOC的差异不明显(P>0.05).随土层加深,LFOC/SOC显著减小(P<0.05),DOC/SOC、MBC/SOC和ROC/SOC逐渐增加(P>0.05).SOC、MBC、DOC、LFOC和ROC间存在极显著正相关关系(P<0.01).各类活性有机碳库与土壤含水量(SwC)、全氮(TN)、碱解氮(AN)和速效磷(AP)呈显著或极显著正相关关系(P<0.05或P<0.01),与土壤容重(BD)呈极显著负相关关系.因此,植被恢复在一定程度上可提高衡阳紫色土丘陵坡地土壤活性有机碳的形成和积累,增加土壤碳储量.
ChenLZ, XieBY, XiaoWF, HuangZL ( 2007). Organic carbon storage in soil under the major forest vegetation types in the Three Gorges Reservior area Resources and Environment in the Yangtze Basin, 16, 640-643. [本文引用: 1]
ChenS, HuangY, ZouJ, ShiY ( 2013). Mean residence time of global topsoil organic carbon depends on temperature, precipitation and soil nitrogen Global and Planetary Change, 100, 99-108. DOIURL [本文引用: 1] Mean residence time (MRT) of topsoil organic carbon is one critical parameter for predicting future land carbon sink dynamics. Large uncertainties remain about controls on the variability in global MRT of soil organic carbon. We estimated global MRT of topsoil (0 20 cm) organic carbon in terrestrial ecosystems and found that mean annual air temperature, annual precipitation, and topsoil nitrogen storage were responsible for the variability in MRT. An empirical climate and soil nitrogen-based (Clim&SN) model could be used to explain the temporal and spatial variability in MRT across various ecosystems. Estimated MRT was lowest in the low-latitude zones, and increased toward high-latitude zones. Global MRT of topsoil organic carbon showed a significant declining tendency between 1960 and 2008, particularly in the high-latitude zone of the northern hemisphere. The largest absolute and relative changes (0.2% per yr) in MRT of topsoil organic carbon from 1960 to 2008 occurred in high-latitude regions, consistent with large carbon stocks in, and greater degree of climate change being experienced by, these areas. Overall, global MRT anomalies (differences between MRT in each year and averaged value of MRT from 1960 to 2008) of terrestrial topsoil organic carbon were decreasing from 1960 to 2008. Global MRT anomalies decreased significantly ( P < 0.001) with the increase of global temperature anomalies, indicating that global warming resulted in faster turnover rates of topsoil organic carbon.
DaiQH, LiuGB, XueS, YuN, ZhangC, LanX ( 2008). Effect of different vegetation restoration on soil carbon and carbon management index in eroded Hilly Loess Plateau Research of Soil and Water Conservation, 15(3), 61-64. URLMagsci [本文引用: 1] 为了解侵蚀环境下植被恢复对土壤活性有机碳及碳库管理指数的影响,以典型侵蚀环境黄土丘陵区纸坊沟流域生态恢复30 a植被长期定位试验点为研究对象,选取坡耕地为参照,分析植被恢复过程中土壤碳库各组分和碳库管理指数的演变特征.结果表明,侵蚀环境下植被恢复后土壤碳库各组分含量都得到显著改善,有机碳、活性有机碳和非活性有机碳含量增幅分别为109%~238%,194%~212%和65%~149%.增加速度非活性有机碳>有机碳>活性有机碳.碳库指数和碳库管理指数较坡耕地明显增加,增幅分别为15%~659%和6.5%~414%,说明土壤经营和管理水平较坡耕地得到了显著改善,土壤系统向着良性方向转变.相关性分析表明有机碳、活性有机碳、非活性有机碳、碳库指数、碳库管理指数与土壤主要肥力因子相关性及其密切,可以作为反映生态恢复过程土壤质量演变的指标.不同植被恢复措施可以显著改善土壤碳库含量,增加土壤碳库管理水平,但改善作用不同,总体来说混交林的效果最好,其次为纯林,最后为荒草地,因此在该地区要通过人工促进生态恢复,应以营造混交林为主,纯林为辅的恢复模式. [ 戴全厚, 刘国彬, 薛萐, 于娜, 张超, 兰雪 ( 2008). 不同植被恢复模式对黄土丘陵区土壤碳库及其管理指数的影响 水土保持研究, 15(3), 61-64.] URLMagsci [本文引用: 1] 为了解侵蚀环境下植被恢复对土壤活性有机碳及碳库管理指数的影响,以典型侵蚀环境黄土丘陵区纸坊沟流域生态恢复30 a植被长期定位试验点为研究对象,选取坡耕地为参照,分析植被恢复过程中土壤碳库各组分和碳库管理指数的演变特征.结果表明,侵蚀环境下植被恢复后土壤碳库各组分含量都得到显著改善,有机碳、活性有机碳和非活性有机碳含量增幅分别为109%~238%,194%~212%和65%~149%.增加速度非活性有机碳>有机碳>活性有机碳.碳库指数和碳库管理指数较坡耕地明显增加,增幅分别为15%~659%和6.5%~414%,说明土壤经营和管理水平较坡耕地得到了显著改善,土壤系统向着良性方向转变.相关性分析表明有机碳、活性有机碳、非活性有机碳、碳库指数、碳库管理指数与土壤主要肥力因子相关性及其密切,可以作为反映生态恢复过程土壤质量演变的指标.不同植被恢复措施可以显著改善土壤碳库含量,增加土壤碳库管理水平,但改善作用不同,总体来说混交林的效果最好,其次为纯林,最后为荒草地,因此在该地区要通过人工促进生态恢复,应以营造混交林为主,纯林为辅的恢复模式.
DengL, WangKB, ChenML, ShangguanZP, SweeneyS ( 2013). Soil organic carbon storage capacity positively related to forest succession on the Loess Plateau, China Catena, 110, 1-7. DOIURL [本文引用: 3] 61Revegetation is recognized as one of the primary contributors to carbon sink.61We examine the effects of long-term revegetation on C stock in soil.61C sequestration mainly occur in lower soil after abandoned farmland.
DingFJ, PanZS, ZhouFJ, WuP ( 2012). Organic carbon contents and vertical distribution characteristics of the soil in the three forest types of the karst regions in central Guizhou Province Journal of Soil and Water Conservation, 26(1), 161-164. URL [本文引用: 1] 在全球气候变化背景下,森林土壤有机碳库已成为全球碳循环研究的 重点之一。研究以黔中喀斯特地区阔叶混交林、针阔混交林和灌木林为对象,分析其土壤及枯落物有机碳含量。结果表明:0-80cm土壤剖面的有机碳含量和碳 密度均值表现为阔叶混交林(19.10g/kg,18.62kg/m2)〉灌木林(9.21g/kg,9.67kg/m2)〉针阔混交林(8.38g /kg,8.60kg/m2),且差异显著(P〈0.05),说明同一地区不同林型土壤有机碳的积累和存储不同;3种林型0-20cm土层有机碳含量和碳 密度均最大,且与各土层之间差异均显著(P〈0.05),并随土层深度的增加而减少,说明森林土壤有机碳含量和密度分布有很强的表聚性,因此,应避免人为 活动干扰,增加植被覆盖和减少水土流失;3种林型的土壤有机碳含量与土壤容重呈负相关关系,拟合度(R2)均较大,说明土壤有机碳含量与土壤容重拟合方程 均较好;3种林型枯落物现存量和有机碳含量差异均显著(P〈0.05),说明同一地区,不同林型的枯落物的积累和分解能力不同。 [ 丁访军, 潘忠松, 周凤娇, 吴鹏 ( 2012). 黔中喀斯特地区3种林型土壤有机碳含量及垂直分布特征 水土保持学报, 26(1), 161-164.] URL [本文引用: 1] 在全球气候变化背景下,森林土壤有机碳库已成为全球碳循环研究的 重点之一。研究以黔中喀斯特地区阔叶混交林、针阔混交林和灌木林为对象,分析其土壤及枯落物有机碳含量。结果表明:0-80cm土壤剖面的有机碳含量和碳 密度均值表现为阔叶混交林(19.10g/kg,18.62kg/m2)〉灌木林(9.21g/kg,9.67kg/m2)〉针阔混交林(8.38g /kg,8.60kg/m2),且差异显著(P〈0.05),说明同一地区不同林型土壤有机碳的积累和存储不同;3种林型0-20cm土层有机碳含量和碳 密度均最大,且与各土层之间差异均显著(P〈0.05),并随土层深度的增加而减少,说明森林土壤有机碳含量和密度分布有很强的表聚性,因此,应避免人为 活动干扰,增加植被覆盖和减少水土流失;3种林型的土壤有机碳含量与土壤容重呈负相关关系,拟合度(R2)均较大,说明土壤有机碳含量与土壤容重拟合方程 均较好;3种林型枯落物现存量和有机碳含量差异均显著(P〈0.05),说明同一地区,不同林型的枯落物的积累和分解能力不同。
DuH, ZengFP, SongTQ, WenYG, LiCG, PengWX, ZhangH, ZengZX ( 2016). Spatial pattern of soil organic carbon of the main forest soils and its influencing factors in Guangxi, China Chinese Journal of Plant Ecology, 40, 282-291. DOIURL [本文引用: 1] 为阐明广西森林土壤有机碳密度分布格局及其主要影响因素,基于森林资源清查资料和345个调查样地的土壤数据,估算了广西主要森林土壤有机碳储量,采用地统计学方法描绘了土壤有机碳密度的空间分布,并利用主成分分析和逐步回归分析方法分析了影响土壤有机碳密度的主要因子.结果表明:广西主要森林土壤有机碳储量(0-100 cm)达到1 686.88 Tg,土壤有机碳密度为124.70 Mg·hm2,低于全国森林土壤平均水平.广西主要森林土壤有机碳密度最佳拟合模型为指数模型,呈中等强度空间相关,Kriging插值显示土壤碳密度高值区在东北区域,低值区在西北区域,表现为喀斯特区域低、非喀斯特区域高的特点.广西主要森林土壤碳密度在不同植被类型和土壤类型下表现出一定差异,其中竹林>落叶阔叶林>暖性针叶林>常绿落叶阔叶林>常绿阔叶林,黄壤>红壤>赤红壤>石灰土.主成分分析和逐步回归分析结果发现土层深度、经纬度、海拔是影响广西森林土壤有机碳的主要因子,其中以土层深度影响最大,主要受岩溶地貌的影响. [ 杜虎, 曾馥平, 宋同清, 温远光, 李春干, 彭晚霞, 张浩, 曾昭霞 ( 2016). 广西主要森林土壤有机碳空间分布及其影响因素 植物生态学报, 40, 282-291.] DOIURL [本文引用: 1] 为阐明广西森林土壤有机碳密度分布格局及其主要影响因素,基于森林资源清查资料和345个调查样地的土壤数据,估算了广西主要森林土壤有机碳储量,采用地统计学方法描绘了土壤有机碳密度的空间分布,并利用主成分分析和逐步回归分析方法分析了影响土壤有机碳密度的主要因子.结果表明:广西主要森林土壤有机碳储量(0-100 cm)达到1 686.88 Tg,土壤有机碳密度为124.70 Mg·hm2,低于全国森林土壤平均水平.广西主要森林土壤有机碳密度最佳拟合模型为指数模型,呈中等强度空间相关,Kriging插值显示土壤碳密度高值区在东北区域,低值区在西北区域,表现为喀斯特区域低、非喀斯特区域高的特点.广西主要森林土壤碳密度在不同植被类型和土壤类型下表现出一定差异,其中竹林>落叶阔叶林>暖性针叶林>常绿落叶阔叶林>常绿阔叶林,黄壤>红壤>赤红壤>石灰土.主成分分析和逐步回归分析结果发现土层深度、经纬度、海拔是影响广西森林土壤有机碳的主要因子,其中以土层深度影响最大,主要受岩溶地貌的影响.
FangJ, ChenA, PengC, ZhaoS, CiL ( 2001). Changes in forest biomass carbon storage in China between 1949 and 1998 Science, 292, 2320-2322. DOIURLPMID [本文引用: 1] Abstract The location and mechanisms responsible for the carbon sink in northern mid-latitude lands are uncertain. Here, we used an improved estimation method of forest biomass and a 50-year national forest resource inventory in China to estimate changes in the storage of living biomass between 1949 and 1998. Our results suggest that Chinese forests released about 0.68 petagram of carbon between 1949 and 1980, for an annual emission rate of 0.022 petagram of carbon. Carbon storage increased significantly after the late 1970s from 4.38 to 4.75 petagram of carbon by 1998, for a mean accumulation rate of 0.021 petagram of carbon per year, mainly due to forest expansion and regrowth. Since the mid-1970s, planted forests (afforestation and reforestation) have sequestered 0.45 petagram of carbon, and their average carbon density increased from 15.3 to 31.1 megagrams per hectare, while natural forests have lost an additional 0.14 petagram of carbon, suggesting that carbon sequestration through forest management practices addressed in the Kyoto Protocol could help offset industrial carbon dioxide emissions.
FiskM, SantangeloS, MinickK ( 2015). Carbon mineralization is promoted by phosphorus and reduced by nitrogen addition in the organic horizon of northern hardwood forests Soil Biology & Biochemistry, 81, 212-218. DOIURL [本文引用: 2] Limitations to the respiratory activity of heterotrophic soil microorganisms exert important controls of CO 2 efflux from soils. In the northeastern US, ecosystem nutrient status varies across the landscape and changes with forest succession following disturbance, likely impacting soil microbial processes regulating the transformation and emission of carbon (C). We tested whether nitrogen (N) or phosphorus (P) limit the mineralization of soil organic C (SOC) or that of added C sources in the Oe horizon of successional and mature northern hardwood forests in three locations in central New Hampshire, USA. Added N reduced mineralization of C from SOC and from added leaf litter and cellulose. Added P did not affect mineralization from SOC; however, it did enhance mineralization of litter- and cellulose- C in organic horizons from all forest locations. Added N increased microbial biomass N and K 2 SO 4 -extractable DON pools, but added P had no effect. Microbial biomass C increased with litter addition but did not respond to either nutrient. The direction of responses to added nutrients was consistent among sites and between forest ages. We conclude that in these organic horizons limitation by N promotes mineralization of C from SOC, whereas limitation by P constrains mineralization of C from new organic inputs. We also suggest that N suppresses respiration in these organic horizons either by relieving the N limitation of microbial biomass synthesis, or by slowing turnover of C through the microbial pool; concurrent measures of microbial growth and turnover are needed to resolve this question.
FuDL, LiuMY, LiuL, ZhangK, ZuoJX ( 2014). Organic carbon density and storage in different soils on the Loess Plateau Arid Zone Research, 1, 44-50. URL [本文引用: 1] 土壤有机质的理化特性是黄土高原地区水土保持及生态修复的重要物质基础,充分了解黄土高原区不同土壤类型的有机碳密度与储量,对生态建设具有重要的实际意义。利用第二次全国土壤普查数据,对黄土高原不同土壤类型0~20 cm表层土体有机碳密度及储量进行估算,并分析两者的空间特征。结果表明:黄土高原区土壤有机碳密度加权平均值为2.00 kg·m^-2,棕壤碳密度值最高,为15.56 kg·m^-2,风沙土最低,仅为0.24 kg· m^-2,空间上呈中间低四周高的分布格局。黄土高原地区总碳储量为1 239.85 Tg(1 Tg=10^12g),灰褐土及黄绵土碳储量较高,两者占总体的46.86%,灰漠土、冻漠土、碱土较低,总量仅占0.17%,空间上呈由西北向东南递增的分布规律。黄绵土、风沙土在黄土高原区分布较广,但两者碳密度较低。因此,在今后的生态修复措施中,提高两者有机碳含量十分关键。 [ 付东磊, 刘梦云, 刘林, 张琨, 左进香 ( 2014). 黄土高原不同土壤类型有机碳密度与储量特征 干旱区研究, 31, 44-50.] URL [本文引用: 1] 土壤有机质的理化特性是黄土高原地区水土保持及生态修复的重要物质基础,充分了解黄土高原区不同土壤类型的有机碳密度与储量,对生态建设具有重要的实际意义。利用第二次全国土壤普查数据,对黄土高原不同土壤类型0~20 cm表层土体有机碳密度及储量进行估算,并分析两者的空间特征。结果表明:黄土高原区土壤有机碳密度加权平均值为2.00 kg·m^-2,棕壤碳密度值最高,为15.56 kg·m^-2,风沙土最低,仅为0.24 kg· m^-2,空间上呈中间低四周高的分布格局。黄土高原地区总碳储量为1 239.85 Tg(1 Tg=10^12g),灰褐土及黄绵土碳储量较高,两者占总体的46.86%,灰漠土、冻漠土、碱土较低,总量仅占0.17%,空间上呈由西北向东南递增的分布规律。黄绵土、风沙土在黄土高原区分布较广,但两者碳密度较低。因此,在今后的生态修复措施中,提高两者有机碳含量十分关键。
GongW, HuXT, WangJY, GongYB, RanH ( 2008). Soil carbon pool and fertility under natural evergreen broad-leaved forest and its artificial regeneration forests in Southern Sichuan Province Acta Ecologica Sinica, 28, 2536-2545. DOIURL [本文引用: 5] 对川南天然常绿阔叶林及其人工更新成檫木林、柳杉林和水杉林后土壤不同形态碳素含量、碳库管理指数、养分含量和酶活性进行研究,并探讨了土壤不同形态碳素及碳库管理指数与土壤肥力之间的关系。结果表明:各季节土壤有机碳、水溶性有机碳、微生物量碳、活性有机碳、稳定态碳、碱解氮、有效磷和速效钾含量及蔗糖酶、磷酸酶和过氧化氢酶活性均为天然常绿阔叶林〉檫木林〉水杉林〉柳杉林,土壤碳库管理指数也为天然常绿阔叶林〉檫木林〉水杉林〉柳杉林,且土壤不同形态碳素含量和碳库管理指数与土壤养分含量及酶活性之间存在显著的相关关系。这说明了天然常绿阔叶林人工更新后土壤不同形态碳素含量、碳库管理指数和土壤肥力下降,且各人工林下降程度小同,而且土壤不同形态碳素含量和碳库管理指数的变化能够较好地表征土壤肥力的变化。研究结果为保护天然常绿阔叶林、选择适宜的更新树种和天然常绿阔叶林人工更新后林地土壤的科学管理提供依据,也为退耕还林中树种的选择提供参考。 [ 龚伟, 胡庭兴, 王景燕, 宫渊波, 冉华 ( 2008). 川南天然常绿阔叶林人工更新后土壤碳库与肥力的变化 生态学报, 28, 2536-2545.] DOIURL [本文引用: 5] 对川南天然常绿阔叶林及其人工更新成檫木林、柳杉林和水杉林后土壤不同形态碳素含量、碳库管理指数、养分含量和酶活性进行研究,并探讨了土壤不同形态碳素及碳库管理指数与土壤肥力之间的关系。结果表明:各季节土壤有机碳、水溶性有机碳、微生物量碳、活性有机碳、稳定态碳、碱解氮、有效磷和速效钾含量及蔗糖酶、磷酸酶和过氧化氢酶活性均为天然常绿阔叶林〉檫木林〉水杉林〉柳杉林,土壤碳库管理指数也为天然常绿阔叶林〉檫木林〉水杉林〉柳杉林,且土壤不同形态碳素含量和碳库管理指数与土壤养分含量及酶活性之间存在显著的相关关系。这说明了天然常绿阔叶林人工更新后土壤不同形态碳素含量、碳库管理指数和土壤肥力下降,且各人工林下降程度小同,而且土壤不同形态碳素含量和碳库管理指数的变化能够较好地表征土壤肥力的变化。研究结果为保护天然常绿阔叶林、选择适宜的更新树种和天然常绿阔叶林人工更新后林地土壤的科学管理提供依据,也为退耕还林中树种的选择提供参考。
GrünebergE, SchöningI, HessenmöllerD, SchulzeED, WeisserWW ( 2013). Organic layer and clay content control soil organic carbon stocks in density fractions of differently managed German beech forests Forest Ecology and Management, 303, 1-10. DOIURL [本文引用: 3] Forest management and associated litter inputs and decomposition rates are thought to affect the carbon storage in mineral soils. Here, we studied the effects of forest management on soil organic carbon (OC) stocks in density fractions of Ah-horizons in soils that developed on loess. We used 82 beech (Fagus sylvatica L.) dominated forest plots in Thuringia, Germany that differed in their management (unmanaged forest, forests under age-class management and forests under selection cutting forest). After density fractionation of the mineral soil with a 1.6 g cm(-3) polytungstate solution we determined OC concentrations and stocks as well as CN-ratios in the free (f-LF) light fraction, the occluded (o-LF) light fraction and in the mineral associated organic matter (MOM) fraction. In our study, Ah-horizons of beech forests stored on average 2.6 +/- 0.2 kg m(-2) (38.7 +/- 1.3 kg m(-3)) OC. The results showed that 37% of the bulk soil OC was stored in the light fractions. We could show that DC stocks in the light fraction were significantly affected by the amount of C stored in organic layers (p = 0.011). The OC stocks in the organic layers, in turn, were higher in unmanaged forests and in forests under selection cutting. This suggests a sensitivity of unprotected OC in the f-LF of beech forests against forest management. In contrast to the f-LF, the OC stocks in the MOM fraction are mainly controlled by pedogenic properties such as clay and iron oxide content. Even after several decades of forest management and with large sample size, an effect of forest management on the stable MOM fraction could not be detected. (c) 2013 Elsevier B.V. All rights reserved.
GuX, FangX, XiangWH, LiSL, SunWJ ( 2013). Soil organic carbon and mineralizable organic carbon in four subtropical forests in hilly region of central Hunan Province, China Chinese Journal of Ecology, 32, 2687-2694. URL [本文引用: 4] 采用短期室内土壤培养法,比较研究了湘中丘陵区4种不同森林类型(杉木人工林、马尾松+石栎针阔混交林、 南酸枣落叶阔叶林、石栎+青冈常绿阔叶林)土壤有机碳、可矿化有机碳的含量.结果表明,4种森林土壤有机碳含量、可矿化有机碳含量以及可矿化有机碳的分配 比例均随土壤深度的增加而下降.同一土层有机碳含量从高至低的排序均表现为:石栎+青冈常绿阔叶林>马尾松+石栎针阔混交林>南酸枣落叶阔叶林>杉木人工 林,0一30 cm土层,南酸枣落叶阔叶林、马尾松+石栎针阔混交林、石栎+青冈常绿阔叶林土壤有机碳含量比杉木人工林分别高出20.8%、29.7%和31.9%.可 矿化有机碳含量及其分配比例从高至低的排序均表现为:石栎+青冈常绿阔叶林>南酸枣落叶阔叶林>马尾松+石栎针阔混交林>杉木人工林,0~30 cm土层,石栎+青冈常绿阔叶林、南酸枣落叶阔叶林、马尾松+石栎针阔混交林可矿化有机碳含量比杉木人工林分别高出51.6%、45.2%和41.0%, 可矿化有机碳的分配比例比杉木人工林分别高出35.8%、29.3%和20.0%,4种森林土壤可矿化有机碳与总有机碳、土壤微生物生物量碳、全N、 C/N之间均呈极显著相关性. [ 辜翔, 方晰, 项文化, 李胜蓝, 孙伟军 ( 2013). 湘中丘陵区4种森林类型土壤有机碳和可矿化有机碳的比较 生态学杂志, 32, 2687-2649.] URL [本文引用: 4] 采用短期室内土壤培养法,比较研究了湘中丘陵区4种不同森林类型(杉木人工林、马尾松+石栎针阔混交林、 南酸枣落叶阔叶林、石栎+青冈常绿阔叶林)土壤有机碳、可矿化有机碳的含量.结果表明,4种森林土壤有机碳含量、可矿化有机碳含量以及可矿化有机碳的分配 比例均随土壤深度的增加而下降.同一土层有机碳含量从高至低的排序均表现为:石栎+青冈常绿阔叶林>马尾松+石栎针阔混交林>南酸枣落叶阔叶林>杉木人工 林,0一30 cm土层,南酸枣落叶阔叶林、马尾松+石栎针阔混交林、石栎+青冈常绿阔叶林土壤有机碳含量比杉木人工林分别高出20.8%、29.7%和31.9%.可 矿化有机碳含量及其分配比例从高至低的排序均表现为:石栎+青冈常绿阔叶林>南酸枣落叶阔叶林>马尾松+石栎针阔混交林>杉木人工林,0~30 cm土层,石栎+青冈常绿阔叶林、南酸枣落叶阔叶林、马尾松+石栎针阔混交林可矿化有机碳含量比杉木人工林分别高出51.6%、45.2%和41.0%, 可矿化有机碳的分配比例比杉木人工林分别高出35.8%、29.3%和20.0%,4种森林土壤可矿化有机碳与总有机碳、土壤微生物生物量碳、全N、 C/N之间均呈极显著相关性.
GuoSL, MaYH, CheSG, SunWY ( 2009). Effects of artificial and natural vegetations on litter production and soil organic carbon change in loess hilly areas Scientia Silvae Sinicae, 45(10), 14-48. DOIMagsci [本文引用: 1] <p><font face="Verdana">在黄土丘陵区选择从耕地、草地、灌木林到乔木林样地,不同样地内设立1 m×1 m(乔木10 m×10 m)的样方,分析样方内凋落物积累量、碳氮含量、土壤有机碳(SOC)和可溶性碳(DOC)含量变化。结果表明:天然草地、灌木林、乔木林凋落物积累量依次为5.3,12.1和32.4 t·hm<sup>-2</sup>; 但人工灌木林和乔木林的凋落物积累量分别为6.7和11.4 t·hm<sup>-2</sup>,分别是天然灌木林和乔木林的1/2和1/3。随着植被的恢复,天然植被凋落物的C/N高于人工植被(刺槐林除外)。与耕地SOC(4.67 g·kg<sup>-1</sup>)相比,天然灌木林地SOC提高5.9倍,人工灌木林地提高1.8倍; 天然乔木林地提高8.0倍,而人工乔木林地仅提高4.0倍。凋落物积累量与0~20 cm土层土壤有机碳存在显著线性相关关系(<em>R</em><sup>2</sup>>0.83),但20 cm以下线性相关关系不显著。凋落物积累量与0~10 cm土壤可溶性碳含量存在显著线性相关关系(<em>R</em><sup>2</sup>>0.893),与10~60 cm土层线性相关关系不显著,与80~100 cm土壤可溶性碳存在显著线性负相关关系。</font></p> [ 郭胜利, 马玉红, 车升国, 孙文义 ( 2009). 黄土区人工与天然植被对凋落物量和土壤有机碳变化的影响 林业科学, 45(10), 14-18.] DOIMagsci [本文引用: 1] <p><font face="Verdana">在黄土丘陵区选择从耕地、草地、灌木林到乔木林样地,不同样地内设立1 m×1 m(乔木10 m×10 m)的样方,分析样方内凋落物积累量、碳氮含量、土壤有机碳(SOC)和可溶性碳(DOC)含量变化。结果表明:天然草地、灌木林、乔木林凋落物积累量依次为5.3,12.1和32.4 t·hm<sup>-2</sup>; 但人工灌木林和乔木林的凋落物积累量分别为6.7和11.4 t·hm<sup>-2</sup>,分别是天然灌木林和乔木林的1/2和1/3。随着植被的恢复,天然植被凋落物的C/N高于人工植被(刺槐林除外)。与耕地SOC(4.67 g·kg<sup>-1</sup>)相比,天然灌木林地SOC提高5.9倍,人工灌木林地提高1.8倍; 天然乔木林地提高8.0倍,而人工乔木林地仅提高4.0倍。凋落物积累量与0~20 cm土层土壤有机碳存在显著线性相关关系(<em>R</em><sup>2</sup>>0.83),但20 cm以下线性相关关系不显著。凋落物积累量与0~10 cm土壤可溶性碳含量存在显著线性相关关系(<em>R</em><sup>2</sup>>0.893),与10~60 cm土层线性相关关系不显著,与80~100 cm土壤可溶性碳存在显著线性负相关关系。</font></p>
Harrison-KirkT, BeareMH, MeenkenED, CondronLM ( 2013). Soil organic matter and texture affect responses to dry/wet cycles: Effects on carbon dioxide and nitrous oxide emissions Soil Biology & Biochemistry, 57, 43-55. DOIURL [本文引用: 1] Soil organic matter (SOM) content and texture are important factors affecting carbon (C) and nitrogen (N) mineralisation under constant soil moisture but their effects on organic matter mineralisation and associated biogenic gas (carbon dioxide (CO 2 ) and nitrous oxide (N 2 O)) production during dry/wet cycles is poorly understood. A laboratory incubation study was conducted to quantify CO 2 and N 2 O production during sequential dry/wet cycles and under constant soil moisture conditions along a gradient of SOM contents in two soil types representing different texture classes (silt loam vs. clay loam). Three soil moisture treatments were established: wet (WW; field capacity), moderately dry (MD; 120% of soil moisture content (SMC) at wilting point (WP)) and very dry (VD; 80% of SMC at WP). To each of the two ‘dry’ treatments two different dry/wet treatments were applied where the soils were either maintained continuously dry (MD & VD) or subjected to three sequential 20-day long dry/wet cycles (MDW & VDW) during the treatment phase of the experiment. At field capacity soil moisture content, the rate of C mineralisation increased with increases in SOC content and the increase per unit of C was twice as high in silt loam (0.3002mg02CO 2 -C02g 611 02SOC02d 611 ) as in clay loam (0.1302mg02CO 2 -C02g 611 02SOC02d 611 ) soils. N 2 O-N emissions also increased with increasing in SOC content. However, in contrast to C mineralisation, the effect was four-fold greater for clay loam (1.3802μg02N 2 O-N02g 611 02SOC02d 611 ) than silt loam (0.3202μg02N 2 O-N02g 611 02SOC02d 611 ) soils. Following rewetting, the VDW and MDW soils produced a short-term C mineralisation flush that was, on average, 30% and 15% greater, respectively, than in WW soils. However, the flush of C mineralisation was not sufficient to compensate for the reduction in mineralisation during the drying phase of each cycle, resulting in a lower total C mineralisation from MDW and VDW soils, on average, compared with WW soils over the three sequential dry/wet cycles. The C mineralisation flush also remained a relatively constant proportion of the total C mineralised from both silt loam (23%) and clay loam soils (22%), irrespective of their SOC content. In contrast, the short-term flush of N 2 O that followed rewetting of dry soil accounted for 62% and 68% of the total N 2 O emissions from silt loam and clay loam soils, respectively. On average, the total N 2 O emissions from dry/wet treatments imposed on silt loam and clay loam soils were 33% and 270% greater, respectively, than from the WW treatments, though the effect varied greatly and depended on SOC content. Overall, N 2 O emissions were highest where we had a combination of fine texture, an adequate supply of available C (i.e. high SOM content), and a water-filled pore space (WFPS)>0.60cmcm 613 at field capacity. Prediction of C mineralisation over dry/wet cycles using mineralisation data from soils at constant moisture content is possible, but knowledge of the stress history for the soil would be required to improve accuracy. The prediction of N 2 O-N emissions during dry/wet cycles using emission data from soils at constant moisture was very inaccurate, due to the inherent spatial variability of N 2 O emissions.
HerbertDA, WilliamsM, RastetterEB ( 2003). Amodel analysis of N and P limitation on carbon accumulation in Amazonian secondary forest after alternate land-use abandonment Biogeochemistry, 65, 121-150. DOIURL [本文引用: 2] Productivity and carbon (C) storage in many mature tropical forests are considered phosphorus (P) limited because of advanced soil weathering. However, disturbance can shift limitation away from P and toward nitrogen (N) because of disproportionately large N losses associated with its mobility relative to P in ecosystems. This shift was illustrated by model analyses in which large disturbances including timber extraction and slash-burn were simulated in a P-limited tropical forest. Re-accumulation of ecosystem C during secondary forest growth was initially N-limited, but long term limitation reverted to P. Mechanisms controlling shifts between N and P limitation included: (1) N volatility during slash combustion produced ash that increased soil solution P more than N, (2) a wide N:P ratio in residual fuel and belowground necromass relative to soil organic matter (SOM) N:P produced a simultaneous P sink and N source during decomposition, (3) a supplemental (to aerosol deposition) external N source via biological N fixation. Redistribution of N and P from low C:nutrient SOM to high C:nutrient vegetation was the most important factor contributing to the resilience of ecosystem C accumulation during secondary growth. Resilience was diminished when multiple harvest and re-growth cycles depleted SOM. Phosphorus losses in particular resulted in long-term reductions of C storage capacity because of slow re-supply rates via deposition and the absence of other external sources. Sensitivity analyses limiting the depth of microbially active SOM in soil profiles further illustrated the importance of elements stored in SOM to ecosystem resilience, pointing to a need for better knowledge on the functioning of deeply buried SOM.
HuangZS, YuLF, FuYH ( 2012). Characteristics of soil mineralizable carbon pool in natural restoration process of karst forest vegetation Chinese Journal of Applied Ecology, 23, 2165-2170. URL [本文引用: 2] 2011年9月,采用空间代替时间方法,研究了茂兰自然保护区喀斯特森林自然恢复过程中土壤可矿化碳库的特征.结果表明:研究期间,喀斯特森林自然恢复过程中不同深度土壤的总有机碳含量、可矿化碳含量和矿化速率随土层加深而减少,随恢复的进程而增加;累积矿化排放量及其速率随恢复的进程增加,其速率随培养时间延长而减小;矿化率随恢复的进程增加,而随土层加深的变化不明显;qCO2值随恢复的进程和土层加深而递减;土壤可矿化碳与凋落物现存量及其分解质量损失率分别呈负相关(r=-0.796)和正相关(r=0.924);土壤生境由早期干扰强烈转向中后期日趋稳定,土壤的固碳能力由早期差、潜力大转向中后期强、潜力小. [ 黄宗胜, 喻理飞, 符裕红 ( 2012). 喀斯特森林植被自然恢复过程中土壤可矿化碳库特征 应用生态学报, 23, 2165-2170.] URL [本文引用: 2] 2011年9月,采用空间代替时间方法,研究了茂兰自然保护区喀斯特森林自然恢复过程中土壤可矿化碳库的特征.结果表明:研究期间,喀斯特森林自然恢复过程中不同深度土壤的总有机碳含量、可矿化碳含量和矿化速率随土层加深而减少,随恢复的进程而增加;累积矿化排放量及其速率随恢复的进程增加,其速率随培养时间延长而减小;矿化率随恢复的进程增加,而随土层加深的变化不明显;qCO2值随恢复的进程和土层加深而递减;土壤可矿化碳与凋落物现存量及其分解质量损失率分别呈负相关(r=-0.796)和正相关(r=0.924);土壤生境由早期干扰强烈转向中后期日趋稳定,土壤的固碳能力由早期差、潜力大转向中后期强、潜力小.
JhaP, DeA, LakariaBL, BiswasAK, SinghM, ReddyKS, RaoAS ( 2012). Soil carbon pools, mineralization and fluxes associated with land use change in vertisols of Central India Natinal Academy Science Letters, 35, 475-483. DOIURL [本文引用: 1] Land use affects soil carbon dynamics which influence the global carbon cycle. Understanding the response of soil organic carbon (SOC) dynamics to land-use changes require size and mean residence time (MRT) of measurable SOC pools. We measured soil carbon pool sizes and decay constants of active and slow carbon pools for different land use systems (agriculture, forest and horticulture) of Central India. Long-term incubation study was carried out for quantification of active (C a ) and slow (C s ) carbon pools and MRT. Chemical fractionation technique (acid hydrolysis) was used for quantification of resistant carbon pool (C r ). The non-hydrolysable carbon (C r ) expressed as a percentage of total C varied between 65 and 6802% in the surface soil of different land use system whereas it was 7802% in sub-surface soil. The incubation study revealed that the Ca pool in different land use system ranged between 3.3 and 1002% of the total C and MRT varied from 25 to 6702days with an average value of 4602days. Our results demonstrate that the active SOC is a sensitive indicator for changes in SOC following land use change. MRT of a slow carbon pool of forest land use was 38.502years. The MRT of slow pool C in the upper soil layer (0–1502cm) did not differ between horticulture and agriculture land use systems and was approximately 4.502years. In agriculture land use, MRT of a slow pool of C under was only 2.302years in sub surface plot. However, MRT of slow carbon pool (C s ) under the Farm yard manure (organic plot) increased by fourfold (17.502years) in comparison to inorganic treatment (recommended dose of fertilizer-RDF) plot. The results clearly depict opportunity of soil carbon buildup in agriculture with good management practices (application of organic manure). The present research work will also help in improving the SOC dynamics predictive capacity of different global bio-geochemical carbon models.
JiaGM, CaoJ, WangC, WangG ( 2005). Microbial biomass and nutrients in soils at the different stages of secondary forest succession in Ziwulin, northwest China Forest Ecology and Management, 217, 117-125. DOIURL [本文引用: 4]
KunkelML, FloresAN, SmithTJ, McnamaraJP, BennerSG ( 2011). A simplified approach for estimating soil carbon and nitrogen stocks in semi-arid complex terrain Geoderma, 165, 1-11. DOIURL [本文引用: 1] 78 The distribution of soil carbon is strongly correlated with elevation and aspect. 78 NDVI and insolation predict soil carbon distribution in semi-arid montane environment. 78 Slope, aspect and elevation reflect the role of insolation in ecosystem productivity. 78 Accounting for past land disturbance improves predictions of soil carbon distribution.
LalR ( 1999). Soil management and restoration for C sequestration to mitigate the accelerated greenhouse effect Progress in Environmental Science, 1, 307-326. DOIURL [本文引用: 1] The accelerated greenhouse effect is caused by increases in the tropospheric concentrations of radiatively active gases (CO2, CH4, N2O, etc.) resulting from such anthropogenic activities as fossil fuel combustion, deforestation and agricultural practices. Soil can be a source (CO2, CH4 and N2O) or sink (CO2 and CH4) of gases depending on land use and management. Soil degradation and decline in ...
LiSJ, QiuLP, ZhangXC ( 2010). Mineralization of soil organic carbon and its relations with soil physical and chemical properties on the Loess Plateau Acta Ecologica Sinica, 30, 1217-1226. URL [本文引用: 1] 土壤有机碳矿化与陆地生态系统碳循环和全球气候变化密切相关。采集98个共7类黄土高原土样,通过密闭培养法对有机碳矿化动态进行分析,探讨了土壤理化性质对有机碳矿化的影响。结果表明,黄土高原地区主要土壤有机碳在培养初期日均矿化量较高,之后逐渐降低。土壤类型对黄土高原土壤有机碳矿化影响较大,不同土壤有机碳的日均矿化量、累积矿化量和矿化率具有显著或极显著的差异。褐土有机碳总矿化量最高,CO2-C达到0.252g.kg-1,风沙土最低,CO2-C仅为0.095g.kg-1。下层土壤的有机碳矿化量较上层的有所下降,但土层深度对有机碳矿化大体上无明显影响。一级动力学方程能很好模拟黄土高原土壤有机碳矿化特征。供试土壤有机碳矿化潜力(Cp)和矿化速率常数(k)值均较低,分别为0.329-0.116g.kg-1和4.55-8.57×10-5d-1。不同土壤的Cp值变异较大,而k和Cp/SOC值无明显的变异。土壤Cp与土壤总有机碳、全氮、全磷、速效钾、粘粒和粉粒含量均呈极显著的正相关,而与k、pH值和砂粒含量呈显著的负相关。研究结果对黄土高原土壤有机碳循环和土壤碳库研究具有重要的科学价值。 [ 李顺姬, 邱莉萍, 张兴昌 ( 2010). 黄土高原土壤有机碳矿化及其与土壤理化性质的关系 生态学报, 30, 1217-1226.] URL [本文引用: 1] 土壤有机碳矿化与陆地生态系统碳循环和全球气候变化密切相关。采集98个共7类黄土高原土样,通过密闭培养法对有机碳矿化动态进行分析,探讨了土壤理化性质对有机碳矿化的影响。结果表明,黄土高原地区主要土壤有机碳在培养初期日均矿化量较高,之后逐渐降低。土壤类型对黄土高原土壤有机碳矿化影响较大,不同土壤有机碳的日均矿化量、累积矿化量和矿化率具有显著或极显著的差异。褐土有机碳总矿化量最高,CO2-C达到0.252g.kg-1,风沙土最低,CO2-C仅为0.095g.kg-1。下层土壤的有机碳矿化量较上层的有所下降,但土层深度对有机碳矿化大体上无明显影响。一级动力学方程能很好模拟黄土高原土壤有机碳矿化特征。供试土壤有机碳矿化潜力(Cp)和矿化速率常数(k)值均较低,分别为0.329-0.116g.kg-1和4.55-8.57×10-5d-1。不同土壤的Cp值变异较大,而k和Cp/SOC值无明显的变异。土壤Cp与土壤总有机碳、全氮、全磷、速效钾、粘粒和粉粒含量均呈极显著的正相关,而与k、pH值和砂粒含量呈显著的负相关。研究结果对黄土高原土壤有机碳循环和土壤碳库研究具有重要的科学价值。
LiangQP, YuXX, PangZ, WangC, LüXZ ( 2010). Study on soil organic carbon density of different forest types Ecology and Environmental Sciences, 19, 889-893. [本文引用: 1]
LiuC, XiangWH, LeiPF, DengXW, TianDL, FangX, PengCH ( 2014). Standing fine root mass and production in four Chinese subtropical forests along a succession and species diversity gradient Plant and Soil, 376, 445-459. DOIURL [本文引用: 1] Background and aims: The influences of succession and species diversity on fine root production are not well known in forests. This study aimed to investigate: (i) whether fine root biomass and production increased with successional stage and increasing tree species diversity; (ii) how forest type affected seasonal variation and regrowth of fine roots. Methods: Sequential coring and ingrowth core methods were used to measure fine root production in four Chinese subtropical forests differing in successional stages and species diversity. Results: Fine root biomass increased from 262 g00·m to 626 g00·m with increasing successional stage and species diversity. A similar trend was also found for fine root production, which increased from 86 to 114 g00·m yr for Cunninghamia lanceolata plantation to 211-240 g00·m yr for Choerospondias axillaries forest when estimated with sequential coring data. Fine root production calculated using the ingrowth core data ranged from 186 g00·m yr for C. lanceolata plantation to 513 g00·m yr for Lithocarpus glaber - Cyclobalanopsis glauca forest. Conclusions: Fine root biomass and production increased along a successional gradient and increasing tree species diversity in subtropical forests. Fine roots in forests with higher species diversity exhibited higher seasonal variation and regrowth rate.
LiuJS, YangJS, YuJB, WangJD ( 2003). Study on vertical distribution of soil organic carbon in wetlands Sanjiang Plain Journal of Soil and Water Conservation, 17(3), 5-8. [本文引用: 2]
LiuSR, WangH, LuanJW ( 2011). A review of research progress and future prospective of forest soil carbon stock and soil carbon process in China Acta Ecologica Sinica, 31, 5437-5448. [本文引用: 2]
LiuW, ChenS, QinX, BaumannF, ScholtenT, ZhouZ, SunW, ZhangT, RenJ, QinD ( 2012). Storage, patterns, and control of soil organic carbon and nitrogen in the northeastern margin of the Qinghai-Tibetan Plateau Environmental Research Letters, 7, 35401-35412. DOIURL [本文引用: 1] This study tested the hypothesis that soil organic carbon (SOC) and total nitrogen (TN) spatial distributions show clear relationships with soil properties and vegetation composition as well as climatic conditions. Further, this study aimed to find the corresponding controlling parameters of SOC and TN storage in high-altitude ecosystems. The study was based on soil, vegetation and climate data from 42 soil pits taken from 14 plots. The plots were investigated during the summers of 2009 and 2010 at the northeastern margin of the Qinghai-Tibetan Plateau. Relationships of SOC density with soil moisture, soil texture, biomass and climatic variables were analyzed. Further, storage and vertical patterns of SOC and TN of seven representative vegetation types were estimated. The results show that significant relationships of SOC density with belowground biomass (BGB) and soil moisture (SM) can be observed. BGB and SM may be the dominant factors influencing SOC density in the topsoil of the study area. The average densities of SOC and TN at a depth of 1 m were about 7.72 kg C m(2) and 0.93 kg N m 2. Both SOC and TN densities were concentrated in the topsoil (0-20 cm) and fell exponentially as soil depth increased. Additionally, the four typical vegetation types located in the northwest of the study area were selected to examine the relationship between SOC and environmental factors (temperature and precipitation). The results indicate that SOC density has a negative relationship with temperature and a positive relationship with precipitation diminishing with soil depth. It was concluded that SOC was concentrated in the topsoil, and that SOC density correlates well with BGB. SOC was predominantly influenced by SM, and to a much lower extent by temperature and precipitation. This study provided a new insight in understanding the control of SOC and TN density in the northeastern margin of the Qinghai-Tibetan Plateau.
LiuWD, SuJR, LiSF, ZhangZJ, LiZW ( 2010). Stoichiometry study of C, N and P in plant and soil at different successional stages of monsoon evergreen broad-leaved forest in Pu’er, Yunnan Province Acta Ecologica Sinica, 30, 6581-6590. URL [本文引用: 1] 通过测定中国西南季风常绿阔叶林不同演替阶段(演替15a,演替30a及原始林)群落中植物叶片与土壤中C、N、P含量,探索了季风常绿阔叶林不同演替阶段群落C、N、P化学计量特征及其与物种丰富度及多度的相关性。结果表明:土壤及植物中全N、全P含量及土壤中C含量均为演替30a群落中最低,而植物中C含量在不同演替阶段群落间无显著性差异。在不同演替阶段共有种中,40%的物种N含量原始林中最高,40%的物种P含量演替15a群落中最高,而80%的物种C含量无显著性差异。土壤中C:N比在不同演替阶段间无显著变化,而N:P及C:P比则随演替呈减小趋势。植物中C:N及C:P比均为演替30a群落最高,而N:P比则随演替呈增加趋势。不同演替阶段共有种的C:N比中,40%的物种原始林中最低,40%的物种无差异,而C:P与N:P比中则均有60%物种无显著性差异,但70%物种在演替15a群落中N:P小于14,演替30a群落中50%物种N:P在14—16,原始林中则有80%物种N:P大于16。群落物种丰富度及个体多度均与C:N、N:P、C:P无显著的相关性,但植物中的N、P与土壤的N、P分别具有显著的线性正相关,说明土壤中N、P供应量影响植物体中的N、P含量。 [ 刘万德, 苏建荣, 李帅锋, 张志钧, 李忠文 ( 2010). 云南普洱季风常绿阔叶林演替系列植物和土壤 C、N、P 化学计量特征 生态学报, 30, 6581-6590.] URL [本文引用: 1] 通过测定中国西南季风常绿阔叶林不同演替阶段(演替15a,演替30a及原始林)群落中植物叶片与土壤中C、N、P含量,探索了季风常绿阔叶林不同演替阶段群落C、N、P化学计量特征及其与物种丰富度及多度的相关性。结果表明:土壤及植物中全N、全P含量及土壤中C含量均为演替30a群落中最低,而植物中C含量在不同演替阶段群落间无显著性差异。在不同演替阶段共有种中,40%的物种N含量原始林中最高,40%的物种P含量演替15a群落中最高,而80%的物种C含量无显著性差异。土壤中C:N比在不同演替阶段间无显著变化,而N:P及C:P比则随演替呈减小趋势。植物中C:N及C:P比均为演替30a群落最高,而N:P比则随演替呈增加趋势。不同演替阶段共有种的C:N比中,40%的物种原始林中最低,40%的物种无差异,而C:P与N:P比中则均有60%物种无显著性差异,但70%物种在演替15a群落中N:P小于14,演替30a群落中50%物种N:P在14—16,原始林中则有80%物种N:P大于16。群落物种丰富度及个体多度均与C:N、N:P、C:P无显著的相关性,但植物中的N、P与土壤的N、P分别具有显著的线性正相关,说明土壤中N、P供应量影响植物体中的N、P含量。
LiuZH, ChenLX, WangLL ( 2009). Soil active carbon in soils under different stages of Korean pine and broadleaf forest Chinese Journal of Soil Science, 40, 1088-1103. URL [本文引用: 2] 采用对比分析方法,研究了红松阔叶混交林不同演替阶段(形成阶段、发展阶段、稳定阶段和顶级群落)、不同土层各形态土壤活性有机碳变化。结果表明:土壤溶解性有机碳(DOC)、微生物量碳(MBC)和易氧化态碳(EOC)含量均随着演替的进展呈上升趋势。土壤溶解性有机碳、微生物量碳、易氧化态碳含量均随着土层深度加深而递减。其中,不同演替阶段易氧化态碳占总有机碳比率随剖面加深有规律地下降,而土壤水溶性有机碳占总有机碳比率随剖面从上到下均表明出上升趋势。A层土壤水溶性碳和微生物量碳占总有机碳比率最大,C层易氧化态碳占总有机碳比率明显高于A层和B层。土壤有机碳总量与各活性有机碳之间以及各类活性有机碳碳与土壤养分之间的相关性均达到显著水平或极显著水平。 [ 刘振花, 陈立新, 王琳琳 ( 2009). 红松阔叶混交林不同演替阶段土壤活性有机碳的研究 土壤通报, 40, 1088-1103.] URL [本文引用: 2] 采用对比分析方法,研究了红松阔叶混交林不同演替阶段(形成阶段、发展阶段、稳定阶段和顶级群落)、不同土层各形态土壤活性有机碳变化。结果表明:土壤溶解性有机碳(DOC)、微生物量碳(MBC)和易氧化态碳(EOC)含量均随着演替的进展呈上升趋势。土壤溶解性有机碳、微生物量碳、易氧化态碳含量均随着土层深度加深而递减。其中,不同演替阶段易氧化态碳占总有机碳比率随剖面加深有规律地下降,而土壤水溶性有机碳占总有机碳比率随剖面从上到下均表明出上升趋势。A层土壤水溶性碳和微生物量碳占总有机碳比率最大,C层易氧化态碳占总有机碳比率明显高于A层和B层。土壤有机碳总量与各活性有机碳之间以及各类活性有机碳碳与土壤养分之间的相关性均达到显著水平或极显著水平。
McKaneRB, RastetterEB, MelilloJM, ShaverGR, HopkinsonCS, FernandesDN ( 1995). Effects of global change on carbon storage in tropical forests of South America Global Biogeochemical Cycle, 9, 329-350. DOIURL [本文引用: 1] We used a process-based model of ecosystem biogeochemistry (MBL-GEM) to evaluate the effects of global change on carbon (C) storage in mature tropical forest ecosystems in the Amazon Basin of Brazil. We first derived a single parameterization of the model that was consistent with all the C stock and turnover data from three intensively studied sites within the Amazon Basin that differed in temperature, rainfall, and cloudiness. The range in temperature, soil moisture, and photosynthetically active radiation (PAR) among these sites is about as large as the anticipated changes in these variables in the tropics under CO 2 -induced climate change. We then tested the parameterized model by predicting C stocks along a 2400-km transect in the Amazon Basin. Comparison of predicted and measured vegetation and soil C stocks along this transect suggests that the model provides a reasonable approximation of how climatic and hydrologic factors regulate present-day C stocks within the Amazon Basin. Finally, we used the model to predict and analyze changes in ecosystem C stocks under projected changes in atmospheric CO 2 and climate. The central hypothesis of this exercise is that changes in ecosystem C storage in response to climate and CO 2 will interact strongly with changes in other element cycles, particularly the nitrogen (N) and phosphorus (P) cycles. We conclude that C storage will increase in Amazonian forests as a result of (1) redistribution of nutrients from soil (with low C:nutrient ratios) to vegetation (with high C:nutrient ratios), (2) increases in the C:nutrient ratio of vegetation and soil, and (3) increased sequestration of external nutrient inputs by the ecosystem. Our analyses suggest that C:nutrient interactions will constrain increases in C storage to a maximum of 63 Mg/ha during the next 200 years, or about 16% above present-day stocks. However, it is impossible to predict how much smaller the actual increase in C storage will be until more is known about the controls on soil P availability. On the basis of these analyses, we identify several topics for further research in the moist tropics that must be addressed to resolve these uncertainties.
MotavalliPP, PalmCA, PartonCA, ElliottET, FreySD ( 1995). Soil pH and organic C dynamics in tropical forest soils: Evidence from laboratory and simulation studies Soil Biology & Biochemistry, 27, 1589-1599. DOIURL [本文引用: 1] Acidic soil pH may affect decomposition of added organic materials in humid tropical forest soils. Our objective was to determine the effects of soil pH on decomposition of added organic materials to tropical forest soils of different soil texture and clay mineralogy. Release of 14 CO 2 and microbial biomass 14 C were measured during a 270-d incubation at 25 C after either [ 14 C]glucose or 14 C-labeled blue grama grass ( Bouteloua gracilis ) material had been added to 13 tropical forest smectitic, kaolinitic, oxidic or allophanic mineralogies. Initial soil pH ranged from 3.9 to 6.7. An additional investigation examined 14 CO 2 release from kaolinitic or oxidic forest soils to which either Ca(OH) 2 or CaSO 4 had been previously applied to obtain 5 soil pH values. Initial soil pH and cumulative 14 CO 2 release in glucose-amended soils were positively related only after 1 and 4 d. In contrast, plant-residue-amended soils had positive relationships between initial soil pH and cumulative 14 CO 2 release after 7 d and continued with that relationship up to 270 d. Microbial biomass 14 C was reduced at lower pH values in both glucose-and plant-residue-amended soils after 270 d. Water-extractable 14 C was also higher at pH > 5.5 in plant-residue-amended soils after 58 d. Differences in soil texture and clay mineralogy had no apparent effect on the relationship between soil pH and decomposition. Simulated results of the experiment using the CENTURY Soil Organic Matter Model diverged from observed results for soils with pH < 6.5. Further research is required to determine the effects of acidic soil pH on decomposition rates of stable C pools and to develop functions for simulation models to account for the short- and long-term effects of soil acidity on decomposition.
PostWM, KwonKC ( 2000). Soil carbon sequestration and land-use change: Processes and Potential Global Change Biology, 6, 317-327. DOIURL [本文引用: 1] Summary When agricultural land is no longer used for cultivation and allowed to revert to natural vegetation or replanted to perennial vegetation, soil organic carbon can accumulate. This accumulation process essentially reverses some of the effects responsible for soil organic carbon losses from when the land was converted from perennial vegetation. We discuss the essential elements of what is known about soil organic matter dynamics that may result in enhanced soil carbon sequestration with changes in land-use and soil management. We review literature that reports changes in soil organic carbon after changes in land-use that favour carbon accumulation. This data summary provides a guide to approximate rates of SOC sequestration that are possible with management, and indicates the relative importance of some factors that influence the rates of organic carbon sequestration in soil. There is a large variation in the length of time for and the rate at which carbon may accumulate in soil, related to the productivity of the recovering vegetation, physical and biological conditions in the soil, and the past history of soil organic carbon inputs and physical disturbance. Maximum rates of C accumulation during the early aggrading stage of perennial vegetation growth, while substantial, are usually much less than 100 g C m 612 y 611. Average rates of accumulation are similar for forest or grassland establishment: 33.8 g C m 612 y 611 and 33.2 g C m 612 y 611 , respectively. These observed rates of soil organic C accumulation, when combined with the small amount of land area involved, are insufficient to account for a significant fraction of the missing C in the global carbon cycle as accumulating in the soils of formerly agricultural land.
RameshT, ManjaiahKM, MohopatraKP, RajasekarK, NgachanSV ( 2015). Assessment of soil organic carbon stocks and fractions under different agroforestry systems in subtropical hill agroecosystems of north-east India Agroforestry Systems, 89, 677-690. DOIURL [本文引用: 2] Soil total organic carbon (TOC) is not sensitive enough to measure the short and medium term changes due to land use change practices, and thus meaningful fractions of soil organic carbon (SOC) should be measured. The objective of this study was to evaluate four agroforestry systems (AFSs) in order to test the hypothesis that agroforestry systems promote an increase in the soil organic carbon stocks and fractions, thus improving soil quality. For this, four 2602years old agroforestry systems consisting of multipurpose tree species (MPTs) ( Michelia oblonga Wall, Parkia roxburghii G. Don, Alnus nepalensis D. Don, and Pinus kesiya Royle ex- Gordon) maintained at ICAR Research Complex, Umiam, Meghalaya, India were tested and compared with control plot for SOC stocks, microbial biomass carbon (MBC), readily oxidizable carbon (ROC), non-labile carbon (C NL ), particulate organic carbon (POC), and carbon management index (CMI). The soil samples were collected at five depths: 0–15, 15–30, 30–45, 45–60, and 60–7502cm. The TOC, POC, ROC, C NL and MBC in soils of agroforestry land use, on average, increased significantly by 26.3, 60.1, 27.1, 46.4 and 3402%, respectively in relative to control plot. However, the AFS consisting A. nepalensis recorded the highest mean SOC stocks (60.202Mg02ha 611 ), TOC (3.6002g 10002g 611 ), POC (5.4202g02kg 611 ), ROC (10.1602g02kg 611 ) and MBC (54802mg02kg 611 ) than other AFSs. Furthermore, the CMI was highest under A. nepalensis (129.6) followed by M. oblonga (119.4), whereas P. kesiya showed the lowest value (91.1). The SOC stocks and fractions were, on average, 35.3 and 7002%, respectively higher in upper soil layer (0–1502cm) compared to subsurface soil layers (15–7502cm) in all the AFSs. The strong and positive correlation ( P 02<020.05) between labile soil carbon fractions with TOC (r02=020.680** for POC; r02=020.683** for ROC and r02=020.758** for MBC) indicates that the changes in TOC content of soils is mainly influenced by the labile C pools and are the sensitive indicators of soil quality improvements. The AFSs, A. nepalensis in particular, can, therefore, be recommended as an alternative soil management strategy for food production, and for the maintenance of soil quality and agricultural sustainability through increased SOC sequestration in the highly fragile agro-ecosystems of northeast India.
ReichsteinM, BahnM, CiaisP, FrankD, Mahecha1MD, SeneviratneSI, ZscheischlerJ, BeerC, BuchmannN, FrankDC, PapaleD, RammigA, SmithP, ThonickeK, van der VeldeM, ViccaS, WalzA, WattenbachM ( 2013). Climate extremes and the carbon cycle Nature, 500, 287-295. DOIURLPMID [本文引用: 1] Abstract The terrestrial biosphere is a key component of the global carbon cycle and its carbon balance is strongly influenced by climate. Continuing environmental changes are thought to increase global terrestrial carbon uptake. But evidence is mounting that climate extremes such as droughts or storms can lead to a decrease in regional ecosystem carbon stocks and therefore have the potential to negate an expected increase in terrestrial carbon uptake. Here we explore the mechanisms and impacts of climate extremes on the terrestrial carbon cycle, and propose a pathway to improve our understanding of present and future impacts of climate extremes on the terrestrial carbon budget.
SáJCDM, LalR ( 2009). Stratification ratio of soil organic matter pools as an indicator of carbon sequestration in a tillage chronosequence on a Brazilian Oxisol Soil and Tillage Research, 103, 46-56. DOIURL [本文引用: 2] Long-term no-tillage (NT) leads to profile stratification of soil organic matter (SOM) pools, and the soil organic carbon (SOC) stratification ratio (SR) is an indicator of soil quality. The objective of this report is to assess the feasibility of using SOC-SR as an index for estimating SOC sequestration in NT soils. The effect of a plow tillage (PT) and NT chronosequence on the SR of SOM pools was assessed in an Oxisol in Southern Brazil (50°23′W and 24°36′S). The chronosequence consisted of six sites: (i) native field (NF); (ii) PT of the native field (PNF-1) involving conversion of natural vegetation to cropland; (iii) NT for 10 years (NT-10); (iv) NT for 20 years (NT-20); (v) NT for 22 years (NT-22); (vi) conventional tillage for 22 years (CT-22). Soil samples were collected from four depths (0–5 cm; 5–10 cm; 10–20 cm; 20–40 cm layer) and soil parameters comprised by SOM pools [i.e., C, N, S, particulate organic C (POC), particulate N (PN), stable C (SC) and stable N (SN), microbial biomass C (MBC) and microbial biomass N (MBN), basal respiration (BR), dissolved organic C (DOC), total polysaccharides (TP) and labile polysaccharides (LP)] were measured. In undisturbed NF soil, the SR of all parameters increased with increase in soil depth. In contrast, the SR decreased in PT, and the SOM was uniformly distributed in the soil profile. All NT treatments restored the SR, and were characterized with higher values of all measured parameters compared to NF. The SR for SOC ranged from 1.12 to 1.51 for CT-22 compared with 1.64–2.61 SR for NT surface and sub-soil layers, respectively. The SR for POC and PN were higher than those for stable C and N. However, SR for the biological pools (e.g., MBC, MBN and BR) were the highest and strongly correlated with the rate of SOC sequestration. An increase in SR of SOC was also positively correlated with the rate and amount of SOC sequestered. Regression analyses indicated a strong correlation between SR of SOC and all parameters monitored in this study. The data showed that the SR of SOC is an efficient indicator of C sequestration in long-term NT management.
SunBW, YangXD, ZhangZH, MaWJ, ArshadA, HuangHX, YanER ( 2013). Relationships between soil carbon pool and vegetation carbon return through succession of evergreen broad-leaved forests in Tiantong region, Zhejiang Province, Eastern China Chinese Journal of Plant Ecology, 37, 803-810. [本文引用: 1]
SunWJ, FangX, XiangWH, ZhangSJ, LiSL ( 2013). Active pools of soil organic carbon in subtropical forests at different successional stages in Central Hunan, China Acta Ecologica Sinica, 33, 7765-7773. DOIURL [本文引用: 2] 为了解天然次生林保护对土壤活 性有机碳库的影响,采用空间替代时间研究方法,对湘中丘陵区不同演替阶段4种林分类型(杉木人工林、马尾松+石栎针阔混交林、南酸枣落叶阔叶林、青冈+石 栎常绿阔叶林)土壤活性有机碳及其与土壤养分相关性进行研究。结果表明:1)各土层总有机碳(TOC)、微生物生物量碳(MBC)、水溶性有机碳 (DOC)、易氧化有机碳(EOC)含量均表现为:青冈+石栎常绿阔叶林南酸枣落叶阔叶林马尾松+石栎针阔混交林杉木人工林,在0—30cm土层,马尾 松+石栎针阔混交林、南酸枣落叶阔叶林、青冈+石栎常绿阔叶林TOC含量比杉木人工林分别高出13.40%、19.40%和29.91%,MBC含量分别 高出15.62%、32.89%和53.33%,DOC含量分别高出8.52%、8.75%和13.76%,EOC含量分别高出32.79%、 38.48%和78.30%;2)天然次生林各土层MBC占TOC的比率以南酸枣落叶阔叶林最高,青冈+石栎常绿阔叶林为其次,马尾松+石栎混交林最低, 均高于同一土层杉木人工林(除马尾松+石栎混交林15—30cm土层外),天然次生林各土层DOC占TOC的比率随着演替进展而下降,均低于同一土层杉木 人工林(除马尾松+石栎混交林0—15cm土层外),天然次生林各土层EOC占TOC的比率随着演替进展而增加,且均高于同一土层杉木人工林;3)土壤 MBC、DOC、EOC含量与TOC含量的相关性均达到极显著水平,且天然次生林土壤MBC、DOC、EOC含量与TOC含量的相关系数随着演替进展而增 高,均高于杉木人工林;4)4种林分土壤TOC、MBC、DOC、EOC含量与土壤全N、碱解N、全P、有效P、全K、速效K含量之间的相关性均达到显著 或极显著水平。 [ 孙伟军, 方晰, 项文化, 张仕吉, 李胜蓝 ( 2013). 湘中丘陵区不同演替阶段森林土壤活性有机碳库特征 生态学报, 33, 7765-7773.] DOIURL [本文引用: 2] 为了解天然次生林保护对土壤活 性有机碳库的影响,采用空间替代时间研究方法,对湘中丘陵区不同演替阶段4种林分类型(杉木人工林、马尾松+石栎针阔混交林、南酸枣落叶阔叶林、青冈+石 栎常绿阔叶林)土壤活性有机碳及其与土壤养分相关性进行研究。结果表明:1)各土层总有机碳(TOC)、微生物生物量碳(MBC)、水溶性有机碳 (DOC)、易氧化有机碳(EOC)含量均表现为:青冈+石栎常绿阔叶林南酸枣落叶阔叶林马尾松+石栎针阔混交林杉木人工林,在0—30cm土层,马尾 松+石栎针阔混交林、南酸枣落叶阔叶林、青冈+石栎常绿阔叶林TOC含量比杉木人工林分别高出13.40%、19.40%和29.91%,MBC含量分别 高出15.62%、32.89%和53.33%,DOC含量分别高出8.52%、8.75%和13.76%,EOC含量分别高出32.79%、 38.48%和78.30%;2)天然次生林各土层MBC占TOC的比率以南酸枣落叶阔叶林最高,青冈+石栎常绿阔叶林为其次,马尾松+石栎混交林最低, 均高于同一土层杉木人工林(除马尾松+石栎混交林15—30cm土层外),天然次生林各土层DOC占TOC的比率随着演替进展而下降,均低于同一土层杉木 人工林(除马尾松+石栎混交林0—15cm土层外),天然次生林各土层EOC占TOC的比率随着演替进展而增加,且均高于同一土层杉木人工林;3)土壤 MBC、DOC、EOC含量与TOC含量的相关性均达到极显著水平,且天然次生林土壤MBC、DOC、EOC含量与TOC含量的相关系数随着演替进展而增 高,均高于杉木人工林;4)4种林分土壤TOC、MBC、DOC、EOC含量与土壤全N、碱解N、全P、有效P、全K、速效K含量之间的相关性均达到显著 或极显著水平。
TangPH, DangKL, WangLH, MaJ ( 2016). Factors affecting soil organic carbon density in Betula albo-sinensis forests on the southern slope of the Qinling Mountains Acta Ecologica Sinica, 36, 1030-1039. DOIURL [本文引用: 3] 以秦岭南坡红桦林为研究对象,利用标准地调查法获得林分、地形、土壤相关数据,分析红桦林土壤有机碳密度(SOCD)分异特征及其与林分因子和地形因子间的关系.结果表明:秦岭南坡红桦林土壤有机碳密度总体均值为(69.02±12.90) t/hm2,原始红桦林土壤有机碳密度均值为(76.21± 10.83) t/hm2,次生红桦林为(65.24± 12.32) t/hm2,原始红桦林土壤有机碳密度比次生红桦林高16.81%,t-检验结果显示两者存在显著差异;在不同林区间,红桦林土壤有机碳密度亦存在显著差异(P<0.05).从地形因子看,红桦林土壤有机碳密度在不同坡位和坡向间未表现出显著差异,而海拔和坡度对红桦林土壤有机碳密度有较为显著的影响.土壤有机碳密度与海拔、林龄、乔木生物量和草本生物量呈显著正相关,与坡度和林分密度呈显著负相关;主成分分析表明:特征值大于1的四个主成分对土壤有机碳密度的方差累积贡献率为85.62%,海拔、坡度、林分密度和郁闭度是影响秦岭南坡红桦林土壤有机碳密度的主要因子;通过逐步回归分析得到利用海拔、坡度、林龄、林分密度、乔木生物量和草本生物量估算红桦林土壤有机碳密度的模型:SOCD=0.015E-0.332G-0.026FD+0.304SA+0.105BA +21.673BH+36.358. [ 唐朋辉, 党坤良, 王连贺, 马俊 ( 2016). 秦岭南坡红桦林土壤有机碳密度影响因素 生态学报, 36, 1030-1039.] DOIURL [本文引用: 3] 以秦岭南坡红桦林为研究对象,利用标准地调查法获得林分、地形、土壤相关数据,分析红桦林土壤有机碳密度(SOCD)分异特征及其与林分因子和地形因子间的关系.结果表明:秦岭南坡红桦林土壤有机碳密度总体均值为(69.02±12.90) t/hm2,原始红桦林土壤有机碳密度均值为(76.21± 10.83) t/hm2,次生红桦林为(65.24± 12.32) t/hm2,原始红桦林土壤有机碳密度比次生红桦林高16.81%,t-检验结果显示两者存在显著差异;在不同林区间,红桦林土壤有机碳密度亦存在显著差异(P<0.05).从地形因子看,红桦林土壤有机碳密度在不同坡位和坡向间未表现出显著差异,而海拔和坡度对红桦林土壤有机碳密度有较为显著的影响.土壤有机碳密度与海拔、林龄、乔木生物量和草本生物量呈显著正相关,与坡度和林分密度呈显著负相关;主成分分析表明:特征值大于1的四个主成分对土壤有机碳密度的方差累积贡献率为85.62%,海拔、坡度、林分密度和郁闭度是影响秦岭南坡红桦林土壤有机碳密度的主要因子;通过逐步回归分析得到利用海拔、坡度、林龄、林分密度、乔木生物量和草本生物量估算红桦林土壤有机碳密度的模型:SOCD=0.015E-0.332G-0.026FD+0.304SA+0.105BA +21.673BH+36.358.
WangH, HallCAS ( 2004). Modeling the effects of Hurricane Hugo on spatial and temporal variation in primary productivity and soil carbon and nitrogen in the Luquillo Experimental Forest, Puerto Rico Plant and Soil, 263, 69-84. DOIURL [本文引用: 1] Hurricanes account for much of the spatial and temporal variation in forest productivity and soil organic matter pools in many forest ecosystems. In this study, we used an ecosystem level model, TOPOECO, to simulate the effects of Hurricane Hugo (18 September 1989) on spatial and temporal patterns of gross primary productivity (GPP), net primary productivity (NPP), soil organic carbon (SOC) and nitrogen over the entire Luquillo Experimental Forest (LEF), Puerto Rico, a tropical rainforest. Our simulation results indicated that simulated annual GPP increased by an average of 30% five years after Hugo in the Tabonuco forest at low elevations where there was a fast recovery of the canopy, whereas simulated GPP decreased by an average of 20% in the Palm and Dwarf forests at high elevations as a result of the slow recovery of the canopy. Simulated annual NPP in the Palm and Dwarf forests also did not recover to pre-Hugo levels within 5 years. Simulated storages of SOC, CO 2 emission from decomposition of SOC and total soil nitrogen increased slightly but N mineralization rate increased significantly in all four vegetation types due to the massive input of plant materials from Hugo at low elevations and the slow decomposition at high elevations.
WangSF, WangXK, OuyangZY ( 2012). Factors controlling soil organic carbon and total nitrogen densities in the upstream watershed of Miyun Reservoir, north China Environmental Science, 33, 946-951. [本文引用: 1]
WangYF, FuBJ, LüYH, ChenLD ( 2011). Effects of vegetation restoration on soil organic carbon sequestration at multiple scales in semi-arid Loess Plateau, China Catena, 85, 58-66. DOIURL [本文引用: 1] Soil organic carbon (SOC) sequestration by vegetation restoration is the theme of much current research. Since 1999, the program of “Grain for Green”has been implemented in the semi-arid Loess Plateau, China. Its scope represents the largest vegetation restoration activity in China. However, it is still unclear for the SOC sequestration effects of vegetation cover change or natural succession promoted by the revegetation efforts at different scales under the semi-arid conditions. In this study, the changes in SOC stocks due to the vegetation restoration in the middle of Loess Plateau were estimated at patch, hill slope transect and small watershed scale from 1998 to 2006. Soil samples were taken from field for the determination of cesium-137 ( 137 Cs) and SOC contents. Vegetation cover change from 1998 to 2006 at the small watershed scale was assessed using Geographic Information System. The results showed that cropland transforming to grassland or shrubland significantly increased SOC at patch scale. Immature woodland, however, has no significant effect. When vegetation cover has no transformation for mature woodland (2502years old), SOC has no significant increase implying that SOC has come to a stable level. At hill slope scale, three typical vegetation cover patterns showed different SOC sequestration effects of 8.6%, 24.6%, and 21.4% from 1998 to 2006, and these SOC increases mainly resulted from revegetation. At the small watershed scale, SOC stocks increased by 19% in the surface soil layer at 0–2002cm soil depth from 1998 to 2006, which was equivalent to an average SOC sequestration rate of 19.9202t02C02y 61021 km 61022 . Meanwhile, SOC contents showed a significant positive correlation (P02<020.001) with the 137 Cs inventory at every soil depth interval. This implied significant negative impacts of soil erosion on SOC sequestration. The results have demonstrated general positive effects of vegetation restoration on SOC sequestration at multiple scales. However, soil erosion under rugged topography modified the spatial distribution of the SOC sequestration effects. Therefore, vegetation restoration was proved to be a significant carbon sink, whereas, erosion could be a carbon source in high erosion sensitive regions. This research can contribute to the performance assessment of ecological rehabilitation projects such as “Grain to Green” and the scientific understanding of the impacts of vegetation restoration and soil erosion on soil carbon dynamics in semi-arid environments.
WeintraubMN, SchimelJP ( 2003). Interactions between carbon and nitrogen mineralization and soil organic matter chemistry in arctic tundra soils Ecosystems, 6, 129-143. DOIURL [本文引用: 1] We used long-term laboratory incubations and chemical fractionation to characterize the mineralization dynamics of organic soils from tussock, shrub, and wet meadow tundra communities, to determine the relationship between soil organic matter (SOM) decomposition and chemistry, and to quantify the relative proportions of carbon (C) and nitrogen (N) in tundra SOM that are biologically available for decomposition. In all soils but shrub, we found little decline in respiration rates over 1 year, although soils respired approximately a tenth to a third of total soil C. The lack of decline in respiration rates despite large C losses indicates that the quantity of organic matter available was not controlling respiration and thus suggests that something else was limiting microbial activity. To determine the nature of the respired C, we analyzed soil chemistry before and after the incubation using a peat fractionation scheme. Despite the large losses of soil C, SOM chemistry was relatively unchanged after the incubation. The decomposition dynamics we observed suggest that tundra SOM, which is largely plant detritus, fits within existing concepts of the litter decay continuum. The lack of changes in organic matter chemistry indicates that this material had already decomposed to the point where the breakdown of labile constituents was tied to lignin decomposition. N mineralization was correlated with C mineralization in our study, but shrub soil mineralized more and tussock soil less N than would have been predicted by this correlation. Our results suggest that a large proportion of tundra SOM is potentially mineralizable, despite the fact that decomposition was dependent on lignin breakdown, and that the historical accumulation of organic matter in tundra soils is the result of field conditions unfavorable to decomposition and not the result of fundamental chemical limitations to decomposition. Our study also suggests that the anticipated increases in shrub dominance may substantially alter the dynamics of SOM decomposition in the tundra.
WynnJG, BirdMI, VellenL, Grand-ClementE, CarterJ, BerrySL ( 2006). Continental-scale measurement of the soil organic carbon pool with climatic, edaphic, and biotic controls Global Biogeochemical Cycles, 20, 1-12. DOIURL [本文引用: 1] [1] We present data on soil organic carbon (SOC) inventory for 7050 soil cores collected from a wide range of environmental conditions throughout Australia. The data set is stratified over the spatial distribution of trees and grass to account for variability of SOC inventory with vegetation distribution. We model controls on SOC inventory using an index of water availability and mean annual temperature to represent the climatic control on the rate of C input into the SOC pool and decomposition of SOC, in addition to the fraction of soil particles <63 m in diameter as a measure of textural control on SOC stabilization. SOC inventories in the top 30 cm of soil increase from 35 mg/cm2 in the driest regions to a modeled plateau with respect to a threshold of water availability at 335 mg/cm2, excluding variables controlling SOC decomposition. Above this threshold, decomposition factors begin to control SOC inventory, which we attribute to energetic control on microbial decomposition rates, and relatively weak stabilization of SOC in association with fine particles. When combined, these relationships provide an overall prediction of SOC inventory that accounts for 89 90% of the variance observed in the measured data set. Deviations from this relationship are most likely due to additional factors that also control decomposition rate such as hydrochemical and soil drainage conditions not accounted for by soil texture. Outliers within this data set are explained with respect to these conditions.
XiangWH, ZhouJ, OuyangS, ZhangSL, LeiPF, LiJX, DengXW, FangX, ForresterDI ( 2016). Species-specific and general allometric equations for estimating tree biomass components of subtropical forests in southern China European Journal of Forest Research, 135, 1-17. DOIURL [本文引用: 1] An increasing number of studies provide evidence that mixed-species stands can overyield monocultures. But it is still hardly understood, how the overyielding at the stand level emerges from the tree, canopy, and size structure. Analyses of 42 triplets with 126 mixed and mono-specific plots in middle-aged, two-species stands of Scots pine (Pinus sylvestris L.), Norway spruce (Picea abies [L.] Karst.), Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco), and European beech (Fagus sylvativa L.) in Central Europe revealed that mixed-species compared with mono-specific stands can have (1) higher tree numbers, higher right skewness and kurtosis of the size distribution, higher inequality of tree sizes, and thereby higher stocking densities and sum of crown projection areas, (2) growth ize relationships with stronger size asymmetry of growth and higher inequality of size growth, and (3) higher stand productivity coupled with higher maximum stand density, canopy space filling, and size asymmetry. These differences depend on the species assemblage. They suggest a deeper entrance of light into the canopy as well as a higher light interception and light-use efficiency as main causes of the overyielding and overdensity. We discuss implications for research and silviculture and draw conclusions for designing and managing resource-efficient production systems.
XueS, LiuGB, PanYP, DaiQH, ZhangC, YuN ( 2009). Evolution of soil labile organic matter and carbon management index in the artificial Robinia of loess hilly area Scientia Agricultura Sinica, 42, 1458-1464. DOIURL [本文引用: 1] 【目的】土壤碳库管理指数是表征土壤碳变化的重要量化指标,研究黄土丘陵区人工刺槐林土壤活性有机碳与碳库管理指数的变化过程对认识该地区生态恢复过程中土壤质量的演变及其效果评价具有重要意义。【方法】采用时空互代法,以典型侵蚀环境纸坊沟流域生态恢复过程中不同年限的人工刺槐林为研究对象,选取坡耕地和天然侧柏林为参照,分析了植被恢复过程中土壤有机碳(TOC)、活性有机碳(LOC)、非活性有机碳(NLOC)及碳库管理指数(CPMI)的演变特征,并运用相关和回归分析方法对生态恢复过程中碳库各组分和恢复年限进行拟合。【结果】营造刺槐林可以显著增加土壤碳库各组分含量,并随恢复年限呈显著线性关系,50a时TOC、LOC、NLOC和碳库指数(CPI)分别较坡耕地增加271%、174%、467%和271%,其中NLOC增加速率略高于LOC,表明植被恢复增加的土壤碳素绝大多数以非活性形态贮存起来,而为了满足生物生长所必须的活性物质来源,土壤碳库必须维持一定的活度状态来满足碳素的动态转化平衡,碳库管理指数在营造刺槐林初期显著降低,随后先增加后降低,与刺槐林生长特性密切相关;但与天然林相比差距仍然较大,恢复50a时TOC、LOC和NLOC仅为侧柏林的49%、34%和61%。【结论】侵蚀环境下的坡耕地由于人为干扰,土壤碳库含量偏低,并处于高速低效率物质转化过程中,人工刺槐林促进生态恢复可以依靠生物的自肥作用增加土壤碳库各组分含量,但要恢复到破坏前该地区顶级群落时的水平,还需要一个漫长的阶段,这个阶段可能需要上百年的时间。 [ 薛萐, 刘国彬, 潘彦平, 戴全厚, 张超, 余娜 ( 2009). 黄土丘陵区人工刺槐林土壤活性有机碳与碳库管理指数演变 中国农业科学, 42, 1458-1464.] DOIURL [本文引用: 1] 【目的】土壤碳库管理指数是表征土壤碳变化的重要量化指标,研究黄土丘陵区人工刺槐林土壤活性有机碳与碳库管理指数的变化过程对认识该地区生态恢复过程中土壤质量的演变及其效果评价具有重要意义。【方法】采用时空互代法,以典型侵蚀环境纸坊沟流域生态恢复过程中不同年限的人工刺槐林为研究对象,选取坡耕地和天然侧柏林为参照,分析了植被恢复过程中土壤有机碳(TOC)、活性有机碳(LOC)、非活性有机碳(NLOC)及碳库管理指数(CPMI)的演变特征,并运用相关和回归分析方法对生态恢复过程中碳库各组分和恢复年限进行拟合。【结果】营造刺槐林可以显著增加土壤碳库各组分含量,并随恢复年限呈显著线性关系,50a时TOC、LOC、NLOC和碳库指数(CPI)分别较坡耕地增加271%、174%、467%和271%,其中NLOC增加速率略高于LOC,表明植被恢复增加的土壤碳素绝大多数以非活性形态贮存起来,而为了满足生物生长所必须的活性物质来源,土壤碳库必须维持一定的活度状态来满足碳素的动态转化平衡,碳库管理指数在营造刺槐林初期显著降低,随后先增加后降低,与刺槐林生长特性密切相关;但与天然林相比差距仍然较大,恢复50a时TOC、LOC和NLOC仅为侧柏林的49%、34%和61%。【结论】侵蚀环境下的坡耕地由于人为干扰,土壤碳库含量偏低,并处于高速低效率物质转化过程中,人工刺槐林促进生态恢复可以依靠生物的自肥作用增加土壤碳库各组分含量,但要恢复到破坏前该地区顶级群落时的水平,还需要一个漫长的阶段,这个阶段可能需要上百年的时间。
YouLH, YeGF, ChenZH, LuoMJ, GaoW, YouSS ( 2014). Litter decomposition and initial nutrient content of major dominant tree species on coastal sandy areas Journal of Fujian Agriculture and Forestry University (Natural Science Edition), 43, 585-591. DOIURL [本文引用: 1] 研究东南沿海4个不同海岸带森林群落9个优势种[潺槁木姜子 (tsea glutinosa)、湿地松(Pinus elliottii)、木麻黄(Casuarina equisetifolia)、黄连木(Pistacia chinensis)、鳝藤(Anodendron affine)、雀梅藤(Sageretia thea)、榕树(Ficus microcarpa)、簕竹(Bambusa blumeana)、朴树(Celtis sinensis]凋落物初始化学组成及其与分解速率的关系,为海岸带防护林树种选择和合理搭配提供参考.采用网袋法进行分解试验.结果表明:①木麻黄和 湿地松凋落物分解最慢,年失重率分别为48.80%和46.00%;潺槁木姜子、黄连木、榕树、簕竹、朴树次之,分别为64.40%、76.80%、 69.30%、74.60、71.40%;鳝藤和雀梅藤分解最快,分别为84.00%和81.40%.②Olson指数衰减模型中,木麻黄和湿地松凋落物 分解50%与95%所需时间最长,分别为1.0354、4.4750和1.1249、4.8617年,鳝藤和雀梅藤最短,分别为0.3782、 1.6347和0.4121、1.7810年.③凋落物分解速率与凋落物初始N、P含量和C/N、C/P比显著相关(P<0.05),与C含量和N/P比 相关性不显著(P>0.05). [ 尤龙辉, 叶功富, 陈增鸿, 罗美娟, 高伟, 游水生 ( 2014). 滨海沙地主要优势树种的凋落物分解及其与初始养分含量的关系 福建农林大学学报(自然科学版), 43, 585-591.] DOIURL [本文引用: 1] 研究东南沿海4个不同海岸带森林群落9个优势种[潺槁木姜子 (tsea glutinosa)、湿地松(Pinus elliottii)、木麻黄(Casuarina equisetifolia)、黄连木(Pistacia chinensis)、鳝藤(Anodendron affine)、雀梅藤(Sageretia thea)、榕树(Ficus microcarpa)、簕竹(Bambusa blumeana)、朴树(Celtis sinensis]凋落物初始化学组成及其与分解速率的关系,为海岸带防护林树种选择和合理搭配提供参考.采用网袋法进行分解试验.结果表明:①木麻黄和 湿地松凋落物分解最慢,年失重率分别为48.80%和46.00%;潺槁木姜子、黄连木、榕树、簕竹、朴树次之,分别为64.40%、76.80%、 69.30%、74.60、71.40%;鳝藤和雀梅藤分解最快,分别为84.00%和81.40%.②Olson指数衰减模型中,木麻黄和湿地松凋落物 分解50%与95%所需时间最长,分别为1.0354、4.4750和1.1249、4.8617年,鳝藤和雀梅藤最短,分别为0.3782、 1.6347和0.4121、1.7810年.③凋落物分解速率与凋落物初始N、P含量和C/N、C/P比显著相关(P<0.05),与C含量和N/P比 相关性不显著(P>0.05).
ZakDR, HolmesWE, BurtonAJ, PregitzerKS, TalhelmAF ( 2008). Simulated atmospheric NO -3 deposition increases soil organic matter by slowing decomposition Ecological Applications, 18, 2016-2027. DOIURLPMID [本文引用: 1] Abstract Presently, there is uncertainty regarding the degree to which anthropogenic N deposition will foster C storage in the N-limited forests of the Northern Hemisphere, ecosystems which are globally important sinks for anthropogenic CO2. We constructed organic matter and N budgets for replicate northern hardwood stands (n = 4) that have received ambient (0.7-1.2 g N x m(-2) x yr(-1) and experimental NO3- deposition (ambient plus 3 g NO3(-)-N x m(-2) x yr(-1)) for a decade; we also traced the flow of a 15NO3- pulse over a six-year period. Experimental NO3- deposition had no effect on organic matter or N stored in the standing forest overstory, but it did significantly increase the N concentration (+19%) and N content (+24%) of canopy leaves. In contrast, a decade of experimental NO3- deposition significantly increased amounts of organic matter (+12%) and N (+9%) in forest floor and mineral soil, despite no increase in detritus production. A greater forest floor (Oe/a) mass under experimental NO3- deposition resulted from slower decomposition, which is consistent with previously reported declines in lignolytic activity by microbial communities exposed to experimental NO3- deposition. Tracing 15NO3- revealed that N accumulated in soil organic matter by first flowing through soil microorganisms and plants, and that the shedding of 15N-labeled leaf litter enriched soil organic matter over a six-year duration. Our results demonstrate that atmospheric NO3- deposition exerts a direct and negative effect on microbial activity in this forest ecosystem, slowing the decomposition of aboveground litter and leading to the accumulation of forest floor and soil organic matter. To the best of our knowledge, this mechanism is not represented in the majority of simulation models predicting the influence of anthropogenic N deposition on ecosystem C storage in northern forests.
ZhangX, HanSJ, WangSQ, GuY, YueLY, FengY, GengSC, ChenZJ ( 2016). Change of soil organic carbon fractions at different successional stages of Betula platyphylla forest in Changbai Mountains Chinese Journal of Ecology, 35, 282-289. DOIURL [本文引用: 2] 为了解长白山天然针阔混交林群落恢复演替土壤碳储量的变化,采用空间代替时间的方法,选取白桦幼龄林、白桦中龄林、白桦成熟林、阔叶红松成熟林和阔叶红松过熟林5个不同演替序列,研究其土壤总有机碳(SOC)、易氧化有机碳(ROC)、微生物生物量碳(MBC)及颗粒有机碳(POC)含量.结果表明:随着白桦林从早期到晚期的演替,SOC、MBC、ROC、POC以及土壤全氮、全磷和碳氮比(C/N)均呈现先逐渐增加后保持稳定的规律.随着土层深度的增加,SOC、MBC、ROC和POC含量均显著降低(P<0.05),5个演替序列内ROC/SOC和POC/SOC的变化范围分别为12.91% ~47.95%和14.21%~69.46%.相关分析表明:MBC、ROC和POC含量与土壤总有机碳(SOC)含量呈极显著正相关(P<0.01),SOC、MBC、ROC和POC含量与全氮、全磷及碳氮比呈极显著正相关(P<0.01).研究结果为了解白桦林在演替过程中土壤有机碳的稳定性变化和固碳潜力提供数据支持. [ 张雪, 韩士杰, 王树起, 谷越, 岳琳艳, 冯月, 耿世聪, 陈志杰 ( 2016). 长白山白桦林不同演替阶段土壤有机碳组分的变化 生态学杂志, 35, 282-289.] DOIURL [本文引用: 2] 为了解长白山天然针阔混交林群落恢复演替土壤碳储量的变化,采用空间代替时间的方法,选取白桦幼龄林、白桦中龄林、白桦成熟林、阔叶红松成熟林和阔叶红松过熟林5个不同演替序列,研究其土壤总有机碳(SOC)、易氧化有机碳(ROC)、微生物生物量碳(MBC)及颗粒有机碳(POC)含量.结果表明:随着白桦林从早期到晚期的演替,SOC、MBC、ROC、POC以及土壤全氮、全磷和碳氮比(C/N)均呈现先逐渐增加后保持稳定的规律.随着土层深度的增加,SOC、MBC、ROC和POC含量均显著降低(P<0.05),5个演替序列内ROC/SOC和POC/SOC的变化范围分别为12.91% ~47.95%和14.21%~69.46%.相关分析表明:MBC、ROC和POC含量与土壤总有机碳(SOC)含量呈极显著正相关(P<0.01),SOC、MBC、ROC和POC含量与全氮、全磷及碳氮比呈极显著正相关(P<0.01).研究结果为了解白桦林在演替过程中土壤有机碳的稳定性变化和固碳潜力提供数据支持.
ZhaoYG, LiuXF, WangZL, ZhaoSW ( 2015). Soil organic carbon fractions and sequestration across a 150-yr secondary forest chronosequence on the Loess Plateau, China Catena, 133, 303-308. DOIURL [本文引用: 4] Restoration of natural vegetation is recommended as an effective approach to restore soil function and rebuild degraded ecosystems. Information is lacking about the long-term results of secondary forest succession on the Loess Plateau with respect to soil organic carbon (SOC) fractions and sequestration in the root-zone soil profile. We investigated the differences in SOC fractions down to 10002cm depths along a 150-yr chronosequence, including cropland (control) and five successional stages (pioneer weeds, herbage, shrub, early forest, and climax forest). Total, labile, and non-labile SOC concentrations increased rapidly at early successional stages (before shrub, <0250-yr) and then gradually leveled off. Total SOC stock was highest at the climax forest stage (64.302Mg02ha 61021 ) and lowest in cropland (39.902Mg02ha 61021 ). Nearly half (~0244.8%) of total SOC stock was stored in surface soils (0–2002cm) and the majority (76.4%) existed in the non-labile fraction. The ratio of labile to non-labile fraction decreased with depth but remained stable across successional stages. The mean SOC sequestration potential and rate relative to cropland were 20.502Mg02ha 61021 and 0.7302Mg02ha 61021 02yr 61021 , respectively. Although the SOC sequestration potential decreased with depth (64.7% for 0–2002cm vs. 35.3% for 20–10002cm), it decreased by 3.102Mg02ha 61021 in the surface and increased by 3.402Mg02ha 61021 in the subsurface from herbage to climax forest stage. This study indicated that long-term secondary forest succession played a positive role in SOC sequestration on the Loess Plateau, especially in the subsurface soil layers.
Effects of global change on carbon storage in tropical forests of South America 1 1995
... 尽管植被恢复过程中CSOC与植被因子、土壤因子密切相关, 但不同因子对CSOC的影响不尽相同.研究表明, SOC的储存在一定程度上是由关键养分N、P的有效性所控制, 高的土壤C/P表征着土壤P有效性较低, 反映土壤P的缺乏, 意味着土壤有机质分解较慢(曹娟等, 2015), P添加可促进土壤SOC分解(Fisk et al., 2015), 森林土壤C/P增大的过程中土壤C储存量会有所增加(刘万德等, 2010).本研究中, 土壤C/P对林地CSOC的影响最显著, CSOC随着土壤C/P的增加而增加(表7).尽管随着植被恢复, 土壤P含量增加, 但土壤C/P总体上也呈增加趋势(表3), 意味着土壤P稀缺程度增大, 微生物活性受到抑制, 有机质分解速率缓慢, 有利于SOC的积累(McKane et al., 1995; Herbert et al., 2003).研究表明, CSOC与土壤pH值呈显著负相关关系(刘景双等, 2003; Jia et al., 2005), 较低的土壤pH值, 通常会抑制微生物的活性和数量, 从而减慢SOC的分解速率, 有利于SOC的积累(Chen et al., 2013); 凋落物C/P是影响凋落物分解速率的重要指标(Aerts, 1997), 较低的凋落物C/P, 能为微生物群落提供充足的养分和能源, 从而提高微生物活性, 促进凋落物的分解(尤龙辉等; 2016), 加快SOC的周转, 不利于SOC的储存(毕京东等, 2014).本研究中, 土壤pH值、凋落物层C/P也显著影响林地CSOC的变化(表7), CSOC随着土壤pH值、凋落物层C/P增大而下降.表明土壤C/P、土壤pH值和凋落物层C/P随着植被恢复的变化对CSOC产生显著影响, SOC积累和存储与限制植物生长的P的供应有关(Herbert et al., 2003). ...
Soil pH and organic C dynamics in tropical forest soils: Evidence from laboratory and simulation studies 1 1995
... 森林SOC主要分布在1 m深度的土壤层中, 对环境变化和人类活动干扰较敏感, 受到诸多因素(如地上植物群落组成结构(龚伟等, 2008; 辜翔等, 2013)、生物量(朱丽琴等, 2017)等生物因素, 地形(唐朋辉等, 2016)、气候(Zak et al., 2008)、土壤理化性质(Grüneberg et al., 2013)及水热条件(Harrison- Kirk et al., 2013)等非生物因素)综合作用的影响.其中, 植被变化是影响SOC库存和积累的重要因素(Wang et al., 2011; Ramesh et al., 2015; Zhao et al., 2015).随着植被恢复, 植物种类组成结构的变化可显著改变凋落物的质量和数量(Ramesh et al., 2015)、根构型及其分泌物(Liu et al., 2014), 显著提高SOC的含量、密度、储量及其稳定性(Deng et al., 2013; Zhao et al., 2015), 全球土壤退化损失的有机碳的60%-75%可重新固定(Lal, 1999); 天然植被恢复对SOC含量(CSOC)的影响比人工植被恢复更显著(郭胜利等, 2009); 原始红桦(Betula albosinensis)林SOC密度(DSOC)显著高于次生红桦林(唐朋辉等, 2016).土壤类型是影响黄土高原地区DSOC和SOC储量的主要因素(付东磊等, 2014); 低温和潮湿环境更有利于SOC的积累(Wynn et al., 2006); 土壤氮(N)是SOC保持和周转的主要影响因素(Weintraub & Schimel, 2003), N添加抑制森林SOC分解, 而磷(P)添加促进土壤SOC分解(Fisk et al., 2015); 土壤颗粒组成在SOC固定、转化过程中起着重要作用(Grüneberg et al., 2013).近20多年来, 在生态系统尺度上, 森林SOC的研究报道已有很多, 但由于森林生态系统各因子之间相互作用的复杂性以及生物、非生物环境的异质性, 使得森林SOC的研究结果仍存在许多不确定性, 特别是有关SOC库动态变化的影响因子方面的研究.研究表明, 森林CSOC与土壤pH值极显著负相关, 土壤酸性越强, 微生物活性越弱, 致使SOC周转速率下降, 越有利于SOC的积累(刘景双等, 2003; Jia et al., 2005).而美国湿润热带SOC与土壤pH值之间具有弱的负相关关系(Motavalli et al., 1995); 陈璟和杨宁(2016)认为, 森林演替过程中, CSOC受土壤pH值变化的影响很小, SOC库不会因此发生较大改变.CSOC与凋落物层现存量的相关性随着森林演替而变小(孙宝伟等, 2013); 而黄宗胜等(2012)发现, 地表凋落物层现存量随着森林演替减少, 显著减慢了SOC分解释放CO2的速率, 有利于SOC的积累.此外, 对于植被恢复或森林演替过程中CSOC与土壤P含量(刘振花等, 2009; 孙伟军等, 2013)、C/N (Jia et al., 2005; 张雪等, 2016)、水分(Jia et al., 2005; Deng et al., 2013)等因子关系的研究, 不同研究的结果也存在较大差异.研究结果的不一致性并不仅仅是某一个因子造成的, 其他SOC周转的控制因素(如植被类型及其种群组成结构、土壤微生物数量及种群组成、土壤质地及其水热状况等)也是造成这种差异的重要原因.因此研究SOC与植被因子、土壤因子之间的关系, 有助于深入了解植被因子、土壤因子对SOC周转的影响方式, 对于发展更加完善的SOC周转模型也是十分必要的.现有森林SOC动态的研究, 主要集中在不同植被类型之间或不同土地利用类型之间SOC库、活性有机碳库的差异(龚伟等, 2008; 刘振花等, 2009; 辜翔等, 2013; 孙伟军等, 2013; 陈璟和杨宁, 2016; 唐朋辉等, 2016; 朱丽琴等, 2017), 在影响机制方面的研究很少涉及森林CSOC、DSOC动态与植被因子、土壤因子间的相互关系(王淑芳等, 2012). ...
Soil carbon sequestration and land-use change: Processes and Potential 1 2000
... 森林土壤有机碳(SOC)库是森林生态系统碳库的重要组成部分, 其储量占森林生态系统碳库的2/3以上, 其微小变化将会影响陆地生态系统碳收支平衡和碳循环过程, 导致全球气候变化.此外, 森林SOC库与土壤物理、化学、生物学性质密切相关, 其质量和数量不仅能够反映森林土壤生产力, 还反映地上植物群落的空间分布和时间演替过程(龚伟等, 2008; 辜翔等, 2013).森林SOC库在调节全球气候变化、维持全球碳平衡以及森林立地生产力等方面起着重要的作用(Post & Kwon, 2000; Wang & Hall, 2004; Reichstein et al., 2013).因此, 森林SOC库动态变化及其影响因素的研究仍然是全球森林生态系统碳循环研究的核心内容(Jha et al., 2012). ...
Assessment of soil organic carbon stocks and fractions under different agroforestry systems in subtropical hill agroecosystems of north-east India 2 2015
... 森林SOC主要分布在1 m深度的土壤层中, 对环境变化和人类活动干扰较敏感, 受到诸多因素(如地上植物群落组成结构(龚伟等, 2008; 辜翔等, 2013)、生物量(朱丽琴等, 2017)等生物因素, 地形(唐朋辉等, 2016)、气候(Zak et al., 2008)、土壤理化性质(Grüneberg et al., 2013)及水热条件(Harrison- Kirk et al., 2013)等非生物因素)综合作用的影响.其中, 植被变化是影响SOC库存和积累的重要因素(Wang et al., 2011; Ramesh et al., 2015; Zhao et al., 2015).随着植被恢复, 植物种类组成结构的变化可显著改变凋落物的质量和数量(Ramesh et al., 2015)、根构型及其分泌物(Liu et al., 2014), 显著提高SOC的含量、密度、储量及其稳定性(Deng et al., 2013; Zhao et al., 2015), 全球土壤退化损失的有机碳的60%-75%可重新固定(Lal, 1999); 天然植被恢复对SOC含量(CSOC)的影响比人工植被恢复更显著(郭胜利等, 2009); 原始红桦(Betula albosinensis)林SOC密度(DSOC)显著高于次生红桦林(唐朋辉等, 2016).土壤类型是影响黄土高原地区DSOC和SOC储量的主要因素(付东磊等, 2014); 低温和潮湿环境更有利于SOC的积累(Wynn et al., 2006); 土壤氮(N)是SOC保持和周转的主要影响因素(Weintraub & Schimel, 2003), N添加抑制森林SOC分解, 而磷(P)添加促进土壤SOC分解(Fisk et al., 2015); 土壤颗粒组成在SOC固定、转化过程中起着重要作用(Grüneberg et al., 2013).近20多年来, 在生态系统尺度上, 森林SOC的研究报道已有很多, 但由于森林生态系统各因子之间相互作用的复杂性以及生物、非生物环境的异质性, 使得森林SOC的研究结果仍存在许多不确定性, 特别是有关SOC库动态变化的影响因子方面的研究.研究表明, 森林CSOC与土壤pH值极显著负相关, 土壤酸性越强, 微生物活性越弱, 致使SOC周转速率下降, 越有利于SOC的积累(刘景双等, 2003; Jia et al., 2005).而美国湿润热带SOC与土壤pH值之间具有弱的负相关关系(Motavalli et al., 1995); 陈璟和杨宁(2016)认为, 森林演替过程中, CSOC受土壤pH值变化的影响很小, SOC库不会因此发生较大改变.CSOC与凋落物层现存量的相关性随着森林演替而变小(孙宝伟等, 2013); 而黄宗胜等(2012)发现, 地表凋落物层现存量随着森林演替减少, 显著减慢了SOC分解释放CO2的速率, 有利于SOC的积累.此外, 对于植被恢复或森林演替过程中CSOC与土壤P含量(刘振花等, 2009; 孙伟军等, 2013)、C/N (Jia et al., 2005; 张雪等, 2016)、水分(Jia et al., 2005; Deng et al., 2013)等因子关系的研究, 不同研究的结果也存在较大差异.研究结果的不一致性并不仅仅是某一个因子造成的, 其他SOC周转的控制因素(如植被类型及其种群组成结构、土壤微生物数量及种群组成、土壤质地及其水热状况等)也是造成这种差异的重要原因.因此研究SOC与植被因子、土壤因子之间的关系, 有助于深入了解植被因子、土壤因子对SOC周转的影响方式, 对于发展更加完善的SOC周转模型也是十分必要的.现有森林SOC动态的研究, 主要集中在不同植被类型之间或不同土地利用类型之间SOC库、活性有机碳库的差异(龚伟等, 2008; 刘振花等, 2009; 辜翔等, 2013; 孙伟军等, 2013; 陈璟和杨宁, 2016; 唐朋辉等, 2016; 朱丽琴等, 2017), 在影响机制方面的研究很少涉及森林CSOC、DSOC动态与植被因子、土壤因子间的相互关系(王淑芳等, 2012). ...
... ).随着植被恢复, 植物种类组成结构的变化可显著改变凋落物的质量和数量(Ramesh et al., 2015)、根构型及其分泌物(Liu et al., 2014), 显著提高SOC的含量、密度、储量及其稳定性(Deng et al., 2013; Zhao et al., 2015), 全球土壤退化损失的有机碳的60%-75%可重新固定(Lal, 1999); 天然植被恢复对SOC含量(CSOC)的影响比人工植被恢复更显著(郭胜利等, 2009); 原始红桦(Betula albosinensis)林SOC密度(DSOC)显著高于次生红桦林(唐朋辉等, 2016).土壤类型是影响黄土高原地区DSOC和SOC储量的主要因素(付东磊等, 2014); 低温和潮湿环境更有利于SOC的积累(Wynn et al., 2006); 土壤氮(N)是SOC保持和周转的主要影响因素(Weintraub & Schimel, 2003), N添加抑制森林SOC分解, 而磷(P)添加促进土壤SOC分解(Fisk et al., 2015); 土壤颗粒组成在SOC固定、转化过程中起着重要作用(Grüneberg et al., 2013).近20多年来, 在生态系统尺度上, 森林SOC的研究报道已有很多, 但由于森林生态系统各因子之间相互作用的复杂性以及生物、非生物环境的异质性, 使得森林SOC的研究结果仍存在许多不确定性, 特别是有关SOC库动态变化的影响因子方面的研究.研究表明, 森林CSOC与土壤pH值极显著负相关, 土壤酸性越强, 微生物活性越弱, 致使SOC周转速率下降, 越有利于SOC的积累(刘景双等, 2003; Jia et al., 2005).而美国湿润热带SOC与土壤pH值之间具有弱的负相关关系(Motavalli et al., 1995); 陈璟和杨宁(2016)认为, 森林演替过程中, CSOC受土壤pH值变化的影响很小, SOC库不会因此发生较大改变.CSOC与凋落物层现存量的相关性随着森林演替而变小(孙宝伟等, 2013); 而黄宗胜等(2012)发现, 地表凋落物层现存量随着森林演替减少, 显著减慢了SOC分解释放CO2的速率, 有利于SOC的积累.此外, 对于植被恢复或森林演替过程中CSOC与土壤P含量(刘振花等, 2009; 孙伟军等, 2013)、C/N (Jia et al., 2005; 张雪等, 2016)、水分(Jia et al., 2005; Deng et al., 2013)等因子关系的研究, 不同研究的结果也存在较大差异.研究结果的不一致性并不仅仅是某一个因子造成的, 其他SOC周转的控制因素(如植被类型及其种群组成结构、土壤微生物数量及种群组成、土壤质地及其水热状况等)也是造成这种差异的重要原因.因此研究SOC与植被因子、土壤因子之间的关系, 有助于深入了解植被因子、土壤因子对SOC周转的影响方式, 对于发展更加完善的SOC周转模型也是十分必要的.现有森林SOC动态的研究, 主要集中在不同植被类型之间或不同土地利用类型之间SOC库、活性有机碳库的差异(龚伟等, 2008; 刘振花等, 2009; 辜翔等, 2013; 孙伟军等, 2013; 陈璟和杨宁, 2016; 唐朋辉等, 2016; 朱丽琴等, 2017), 在影响机制方面的研究很少涉及森林CSOC、DSOC动态与植被因子、土壤因子间的相互关系(王淑芳等, 2012). ...
Climate extremes and the carbon cycle 1 2013
... 森林土壤有机碳(SOC)库是森林生态系统碳库的重要组成部分, 其储量占森林生态系统碳库的2/3以上, 其微小变化将会影响陆地生态系统碳收支平衡和碳循环过程, 导致全球气候变化.此外, 森林SOC库与土壤物理、化学、生物学性质密切相关, 其质量和数量不仅能够反映森林土壤生产力, 还反映地上植物群落的空间分布和时间演替过程(龚伟等, 2008; 辜翔等, 2013).森林SOC库在调节全球气候变化、维持全球碳平衡以及森林立地生产力等方面起着重要的作用(Post & Kwon, 2000; Wang & Hall, 2004; Reichstein et al., 2013).因此, 森林SOC库动态变化及其影响因素的研究仍然是全球森林生态系统碳循环研究的核心内容(Jha et al., 2012). ...
Stratification ratio of soil organic matter pools as an indicator of carbon sequestration in a tillage chronosequence on a Brazilian Oxisol 2 2009
Modeling the effects of Hurricane Hugo on spatial and temporal variation in primary productivity and soil carbon and nitrogen in the Luquillo Experimental Forest, Puerto Rico 1 2004
... 森林土壤有机碳(SOC)库是森林生态系统碳库的重要组成部分, 其储量占森林生态系统碳库的2/3以上, 其微小变化将会影响陆地生态系统碳收支平衡和碳循环过程, 导致全球气候变化.此外, 森林SOC库与土壤物理、化学、生物学性质密切相关, 其质量和数量不仅能够反映森林土壤生产力, 还反映地上植物群落的空间分布和时间演替过程(龚伟等, 2008; 辜翔等, 2013).森林SOC库在调节全球气候变化、维持全球碳平衡以及森林立地生产力等方面起着重要的作用(Post & Kwon, 2000; Wang & Hall, 2004; Reichstein et al., 2013).因此, 森林SOC库动态变化及其影响因素的研究仍然是全球森林生态系统碳循环研究的核心内容(Jha et al., 2012). ...
密云水库上游流域土壤有机碳和全氮密度影响因素研究 1 2012
... 森林SOC主要分布在1 m深度的土壤层中, 对环境变化和人类活动干扰较敏感, 受到诸多因素(如地上植物群落组成结构(龚伟等, 2008; 辜翔等, 2013)、生物量(朱丽琴等, 2017)等生物因素, 地形(唐朋辉等, 2016)、气候(Zak et al., 2008)、土壤理化性质(Grüneberg et al., 2013)及水热条件(Harrison- Kirk et al., 2013)等非生物因素)综合作用的影响.其中, 植被变化是影响SOC库存和积累的重要因素(Wang et al., 2011; Ramesh et al., 2015; Zhao et al., 2015).随着植被恢复, 植物种类组成结构的变化可显著改变凋落物的质量和数量(Ramesh et al., 2015)、根构型及其分泌物(Liu et al., 2014), 显著提高SOC的含量、密度、储量及其稳定性(Deng et al., 2013; Zhao et al., 2015), 全球土壤退化损失的有机碳的60%-75%可重新固定(Lal, 1999); 天然植被恢复对SOC含量(CSOC)的影响比人工植被恢复更显著(郭胜利等, 2009); 原始红桦(Betula albosinensis)林SOC密度(DSOC)显著高于次生红桦林(唐朋辉等, 2016).土壤类型是影响黄土高原地区DSOC和SOC储量的主要因素(付东磊等, 2014); 低温和潮湿环境更有利于SOC的积累(Wynn et al., 2006); 土壤氮(N)是SOC保持和周转的主要影响因素(Weintraub & Schimel, 2003), N添加抑制森林SOC分解, 而磷(P)添加促进土壤SOC分解(Fisk et al., 2015); 土壤颗粒组成在SOC固定、转化过程中起着重要作用(Grüneberg et al., 2013).近20多年来, 在生态系统尺度上, 森林SOC的研究报道已有很多, 但由于森林生态系统各因子之间相互作用的复杂性以及生物、非生物环境的异质性, 使得森林SOC的研究结果仍存在许多不确定性, 特别是有关SOC库动态变化的影响因子方面的研究.研究表明, 森林CSOC与土壤pH值极显著负相关, 土壤酸性越强, 微生物活性越弱, 致使SOC周转速率下降, 越有利于SOC的积累(刘景双等, 2003; Jia et al., 2005).而美国湿润热带SOC与土壤pH值之间具有弱的负相关关系(Motavalli et al., 1995); 陈璟和杨宁(2016)认为, 森林演替过程中, CSOC受土壤pH值变化的影响很小, SOC库不会因此发生较大改变.CSOC与凋落物层现存量的相关性随着森林演替而变小(孙宝伟等, 2013); 而黄宗胜等(2012)发现, 地表凋落物层现存量随着森林演替减少, 显著减慢了SOC分解释放CO2的速率, 有利于SOC的积累.此外, 对于植被恢复或森林演替过程中CSOC与土壤P含量(刘振花等, 2009; 孙伟军等, 2013)、C/N (Jia et al., 2005; 张雪等, 2016)、水分(Jia et al., 2005; Deng et al., 2013)等因子关系的研究, 不同研究的结果也存在较大差异.研究结果的不一致性并不仅仅是某一个因子造成的, 其他SOC周转的控制因素(如植被类型及其种群组成结构、土壤微生物数量及种群组成、土壤质地及其水热状况等)也是造成这种差异的重要原因.因此研究SOC与植被因子、土壤因子之间的关系, 有助于深入了解植被因子、土壤因子对SOC周转的影响方式, 对于发展更加完善的SOC周转模型也是十分必要的.现有森林SOC动态的研究, 主要集中在不同植被类型之间或不同土地利用类型之间SOC库、活性有机碳库的差异(龚伟等, 2008; 刘振花等, 2009; 辜翔等, 2013; 孙伟军等, 2013; 陈璟和杨宁, 2016; 唐朋辉等, 2016; 朱丽琴等, 2017), 在影响机制方面的研究很少涉及森林CSOC、DSOC动态与植被因子、土壤因子间的相互关系(王淑芳等, 2012). ...
密云水库上游流域土壤有机碳和全氮密度影响因素研究 1 2012
... 森林SOC主要分布在1 m深度的土壤层中, 对环境变化和人类活动干扰较敏感, 受到诸多因素(如地上植物群落组成结构(龚伟等, 2008; 辜翔等, 2013)、生物量(朱丽琴等, 2017)等生物因素, 地形(唐朋辉等, 2016)、气候(Zak et al., 2008)、土壤理化性质(Grüneberg et al., 2013)及水热条件(Harrison- Kirk et al., 2013)等非生物因素)综合作用的影响.其中, 植被变化是影响SOC库存和积累的重要因素(Wang et al., 2011; Ramesh et al., 2015; Zhao et al., 2015).随着植被恢复, 植物种类组成结构的变化可显著改变凋落物的质量和数量(Ramesh et al., 2015)、根构型及其分泌物(Liu et al., 2014), 显著提高SOC的含量、密度、储量及其稳定性(Deng et al., 2013; Zhao et al., 2015), 全球土壤退化损失的有机碳的60%-75%可重新固定(Lal, 1999); 天然植被恢复对SOC含量(CSOC)的影响比人工植被恢复更显著(郭胜利等, 2009); 原始红桦(Betula albosinensis)林SOC密度(DSOC)显著高于次生红桦林(唐朋辉等, 2016).土壤类型是影响黄土高原地区DSOC和SOC储量的主要因素(付东磊等, 2014); 低温和潮湿环境更有利于SOC的积累(Wynn et al., 2006); 土壤氮(N)是SOC保持和周转的主要影响因素(Weintraub & Schimel, 2003), N添加抑制森林SOC分解, 而磷(P)添加促进土壤SOC分解(Fisk et al., 2015); 土壤颗粒组成在SOC固定、转化过程中起着重要作用(Grüneberg et al., 2013).近20多年来, 在生态系统尺度上, 森林SOC的研究报道已有很多, 但由于森林生态系统各因子之间相互作用的复杂性以及生物、非生物环境的异质性, 使得森林SOC的研究结果仍存在许多不确定性, 特别是有关SOC库动态变化的影响因子方面的研究.研究表明, 森林CSOC与土壤pH值极显著负相关, 土壤酸性越强, 微生物活性越弱, 致使SOC周转速率下降, 越有利于SOC的积累(刘景双等, 2003; Jia et al., 2005).而美国湿润热带SOC与土壤pH值之间具有弱的负相关关系(Motavalli et al., 1995); 陈璟和杨宁(2016)认为, 森林演替过程中, CSOC受土壤pH值变化的影响很小, SOC库不会因此发生较大改变.CSOC与凋落物层现存量的相关性随着森林演替而变小(孙宝伟等, 2013); 而黄宗胜等(2012)发现, 地表凋落物层现存量随着森林演替减少, 显著减慢了SOC分解释放CO2的速率, 有利于SOC的积累.此外, 对于植被恢复或森林演替过程中CSOC与土壤P含量(刘振花等, 2009; 孙伟军等, 2013)、C/N (Jia et al., 2005; 张雪等, 2016)、水分(Jia et al., 2005; Deng et al., 2013)等因子关系的研究, 不同研究的结果也存在较大差异.研究结果的不一致性并不仅仅是某一个因子造成的, 其他SOC周转的控制因素(如植被类型及其种群组成结构、土壤微生物数量及种群组成、土壤质地及其水热状况等)也是造成这种差异的重要原因.因此研究SOC与植被因子、土壤因子之间的关系, 有助于深入了解植被因子、土壤因子对SOC周转的影响方式, 对于发展更加完善的SOC周转模型也是十分必要的.现有森林SOC动态的研究, 主要集中在不同植被类型之间或不同土地利用类型之间SOC库、活性有机碳库的差异(龚伟等, 2008; 刘振花等, 2009; 辜翔等, 2013; 孙伟军等, 2013; 陈璟和杨宁, 2016; 唐朋辉等, 2016; 朱丽琴等, 2017), 在影响机制方面的研究很少涉及森林CSOC、DSOC动态与植被因子、土壤因子间的相互关系(王淑芳等, 2012). ...
Effects of vegetation restoration on soil organic carbon sequestration at multiple scales in semi-arid Loess Plateau, China 1 2011
... 森林SOC主要分布在1 m深度的土壤层中, 对环境变化和人类活动干扰较敏感, 受到诸多因素(如地上植物群落组成结构(龚伟等, 2008; 辜翔等, 2013)、生物量(朱丽琴等, 2017)等生物因素, 地形(唐朋辉等, 2016)、气候(Zak et al., 2008)、土壤理化性质(Grüneberg et al., 2013)及水热条件(Harrison- Kirk et al., 2013)等非生物因素)综合作用的影响.其中, 植被变化是影响SOC库存和积累的重要因素(Wang et al., 2011; Ramesh et al., 2015; Zhao et al., 2015).随着植被恢复, 植物种类组成结构的变化可显著改变凋落物的质量和数量(Ramesh et al., 2015)、根构型及其分泌物(Liu et al., 2014), 显著提高SOC的含量、密度、储量及其稳定性(Deng et al., 2013; Zhao et al., 2015), 全球土壤退化损失的有机碳的60%-75%可重新固定(Lal, 1999); 天然植被恢复对SOC含量(CSOC)的影响比人工植被恢复更显著(郭胜利等, 2009); 原始红桦(Betula albosinensis)林SOC密度(DSOC)显著高于次生红桦林(唐朋辉等, 2016).土壤类型是影响黄土高原地区DSOC和SOC储量的主要因素(付东磊等, 2014); 低温和潮湿环境更有利于SOC的积累(Wynn et al., 2006); 土壤氮(N)是SOC保持和周转的主要影响因素(Weintraub & Schimel, 2003), N添加抑制森林SOC分解, 而磷(P)添加促进土壤SOC分解(Fisk et al., 2015); 土壤颗粒组成在SOC固定、转化过程中起着重要作用(Grüneberg et al., 2013).近20多年来, 在生态系统尺度上, 森林SOC的研究报道已有很多, 但由于森林生态系统各因子之间相互作用的复杂性以及生物、非生物环境的异质性, 使得森林SOC的研究结果仍存在许多不确定性, 特别是有关SOC库动态变化的影响因子方面的研究.研究表明, 森林CSOC与土壤pH值极显著负相关, 土壤酸性越强, 微生物活性越弱, 致使SOC周转速率下降, 越有利于SOC的积累(刘景双等, 2003; Jia et al., 2005).而美国湿润热带SOC与土壤pH值之间具有弱的负相关关系(Motavalli et al., 1995); 陈璟和杨宁(2016)认为, 森林演替过程中, CSOC受土壤pH值变化的影响很小, SOC库不会因此发生较大改变.CSOC与凋落物层现存量的相关性随着森林演替而变小(孙宝伟等, 2013); 而黄宗胜等(2012)发现, 地表凋落物层现存量随着森林演替减少, 显著减慢了SOC分解释放CO2的速率, 有利于SOC的积累.此外, 对于植被恢复或森林演替过程中CSOC与土壤P含量(刘振花等, 2009; 孙伟军等, 2013)、C/N (Jia et al., 2005; 张雪等, 2016)、水分(Jia et al., 2005; Deng et al., 2013)等因子关系的研究, 不同研究的结果也存在较大差异.研究结果的不一致性并不仅仅是某一个因子造成的, 其他SOC周转的控制因素(如植被类型及其种群组成结构、土壤微生物数量及种群组成、土壤质地及其水热状况等)也是造成这种差异的重要原因.因此研究SOC与植被因子、土壤因子之间的关系, 有助于深入了解植被因子、土壤因子对SOC周转的影响方式, 对于发展更加完善的SOC周转模型也是十分必要的.现有森林SOC动态的研究, 主要集中在不同植被类型之间或不同土地利用类型之间SOC库、活性有机碳库的差异(龚伟等, 2008; 刘振花等, 2009; 辜翔等, 2013; 孙伟军等, 2013; 陈璟和杨宁, 2016; 唐朋辉等, 2016; 朱丽琴等, 2017), 在影响机制方面的研究很少涉及森林CSOC、DSOC动态与植被因子、土壤因子间的相互关系(王淑芳等, 2012). ...
Interactions between carbon and nitrogen mineralization and soil organic matter chemistry in arctic tundra soils 1 2003
... 森林SOC主要分布在1 m深度的土壤层中, 对环境变化和人类活动干扰较敏感, 受到诸多因素(如地上植物群落组成结构(龚伟等, 2008; 辜翔等, 2013)、生物量(朱丽琴等, 2017)等生物因素, 地形(唐朋辉等, 2016)、气候(Zak et al., 2008)、土壤理化性质(Grüneberg et al., 2013)及水热条件(Harrison- Kirk et al., 2013)等非生物因素)综合作用的影响.其中, 植被变化是影响SOC库存和积累的重要因素(Wang et al., 2011; Ramesh et al., 2015; Zhao et al., 2015).随着植被恢复, 植物种类组成结构的变化可显著改变凋落物的质量和数量(Ramesh et al., 2015)、根构型及其分泌物(Liu et al., 2014), 显著提高SOC的含量、密度、储量及其稳定性(Deng et al., 2013; Zhao et al., 2015), 全球土壤退化损失的有机碳的60%-75%可重新固定(Lal, 1999); 天然植被恢复对SOC含量(CSOC)的影响比人工植被恢复更显著(郭胜利等, 2009); 原始红桦(Betula albosinensis)林SOC密度(DSOC)显著高于次生红桦林(唐朋辉等, 2016).土壤类型是影响黄土高原地区DSOC和SOC储量的主要因素(付东磊等, 2014); 低温和潮湿环境更有利于SOC的积累(Wynn et al., 2006); 土壤氮(N)是SOC保持和周转的主要影响因素(Weintraub & Schimel, 2003), N添加抑制森林SOC分解, 而磷(P)添加促进土壤SOC分解(Fisk et al., 2015); 土壤颗粒组成在SOC固定、转化过程中起着重要作用(Grüneberg et al., 2013).近20多年来, 在生态系统尺度上, 森林SOC的研究报道已有很多, 但由于森林生态系统各因子之间相互作用的复杂性以及生物、非生物环境的异质性, 使得森林SOC的研究结果仍存在许多不确定性, 特别是有关SOC库动态变化的影响因子方面的研究.研究表明, 森林CSOC与土壤pH值极显著负相关, 土壤酸性越强, 微生物活性越弱, 致使SOC周转速率下降, 越有利于SOC的积累(刘景双等, 2003; Jia et al., 2005).而美国湿润热带SOC与土壤pH值之间具有弱的负相关关系(Motavalli et al., 1995); 陈璟和杨宁(2016)认为, 森林演替过程中, CSOC受土壤pH值变化的影响很小, SOC库不会因此发生较大改变.CSOC与凋落物层现存量的相关性随着森林演替而变小(孙宝伟等, 2013); 而黄宗胜等(2012)发现, 地表凋落物层现存量随着森林演替减少, 显著减慢了SOC分解释放CO2的速率, 有利于SOC的积累.此外, 对于植被恢复或森林演替过程中CSOC与土壤P含量(刘振花等, 2009; 孙伟军等, 2013)、C/N (Jia et al., 2005; 张雪等, 2016)、水分(Jia et al., 2005; Deng et al., 2013)等因子关系的研究, 不同研究的结果也存在较大差异.研究结果的不一致性并不仅仅是某一个因子造成的, 其他SOC周转的控制因素(如植被类型及其种群组成结构、土壤微生物数量及种群组成、土壤质地及其水热状况等)也是造成这种差异的重要原因.因此研究SOC与植被因子、土壤因子之间的关系, 有助于深入了解植被因子、土壤因子对SOC周转的影响方式, 对于发展更加完善的SOC周转模型也是十分必要的.现有森林SOC动态的研究, 主要集中在不同植被类型之间或不同土地利用类型之间SOC库、活性有机碳库的差异(龚伟等, 2008; 刘振花等, 2009; 辜翔等, 2013; 孙伟军等, 2013; 陈璟和杨宁, 2016; 唐朋辉等, 2016; 朱丽琴等, 2017), 在影响机制方面的研究很少涉及森林CSOC、DSOC动态与植被因子、土壤因子间的相互关系(王淑芳等, 2012). ...
Continental-scale measurement of the soil organic carbon pool with climatic, edaphic, and biotic controls 1 2006
... 森林SOC主要分布在1 m深度的土壤层中, 对环境变化和人类活动干扰较敏感, 受到诸多因素(如地上植物群落组成结构(龚伟等, 2008; 辜翔等, 2013)、生物量(朱丽琴等, 2017)等生物因素, 地形(唐朋辉等, 2016)、气候(Zak et al., 2008)、土壤理化性质(Grüneberg et al., 2013)及水热条件(Harrison- Kirk et al., 2013)等非生物因素)综合作用的影响.其中, 植被变化是影响SOC库存和积累的重要因素(Wang et al., 2011; Ramesh et al., 2015; Zhao et al., 2015).随着植被恢复, 植物种类组成结构的变化可显著改变凋落物的质量和数量(Ramesh et al., 2015)、根构型及其分泌物(Liu et al., 2014), 显著提高SOC的含量、密度、储量及其稳定性(Deng et al., 2013; Zhao et al., 2015), 全球土壤退化损失的有机碳的60%-75%可重新固定(Lal, 1999); 天然植被恢复对SOC含量(CSOC)的影响比人工植被恢复更显著(郭胜利等, 2009); 原始红桦(Betula albosinensis)林SOC密度(DSOC)显著高于次生红桦林(唐朋辉等, 2016).土壤类型是影响黄土高原地区DSOC和SOC储量的主要因素(付东磊等, 2014); 低温和潮湿环境更有利于SOC的积累(Wynn et al., 2006); 土壤氮(N)是SOC保持和周转的主要影响因素(Weintraub & Schimel, 2003), N添加抑制森林SOC分解, 而磷(P)添加促进土壤SOC分解(Fisk et al., 2015); 土壤颗粒组成在SOC固定、转化过程中起着重要作用(Grüneberg et al., 2013).近20多年来, 在生态系统尺度上, 森林SOC的研究报道已有很多, 但由于森林生态系统各因子之间相互作用的复杂性以及生物、非生物环境的异质性, 使得森林SOC的研究结果仍存在许多不确定性, 特别是有关SOC库动态变化的影响因子方面的研究.研究表明, 森林CSOC与土壤pH值极显著负相关, 土壤酸性越强, 微生物活性越弱, 致使SOC周转速率下降, 越有利于SOC的积累(刘景双等, 2003; Jia et al., 2005).而美国湿润热带SOC与土壤pH值之间具有弱的负相关关系(Motavalli et al., 1995); 陈璟和杨宁(2016)认为, 森林演替过程中, CSOC受土壤pH值变化的影响很小, SOC库不会因此发生较大改变.CSOC与凋落物层现存量的相关性随着森林演替而变小(孙宝伟等, 2013); 而黄宗胜等(2012)发现, 地表凋落物层现存量随着森林演替减少, 显著减慢了SOC分解释放CO2的速率, 有利于SOC的积累.此外, 对于植被恢复或森林演替过程中CSOC与土壤P含量(刘振花等, 2009; 孙伟军等, 2013)、C/N (Jia et al., 2005; 张雪等, 2016)、水分(Jia et al., 2005; Deng et al., 2013)等因子关系的研究, 不同研究的结果也存在较大差异.研究结果的不一致性并不仅仅是某一个因子造成的, 其他SOC周转的控制因素(如植被类型及其种群组成结构、土壤微生物数量及种群组成、土壤质地及其水热状况等)也是造成这种差异的重要原因.因此研究SOC与植被因子、土壤因子之间的关系, 有助于深入了解植被因子、土壤因子对SOC周转的影响方式, 对于发展更加完善的SOC周转模型也是十分必要的.现有森林SOC动态的研究, 主要集中在不同植被类型之间或不同土地利用类型之间SOC库、活性有机碳库的差异(龚伟等, 2008; 刘振花等, 2009; 辜翔等, 2013; 孙伟军等, 2013; 陈璟和杨宁, 2016; 唐朋辉等, 2016; 朱丽琴等, 2017), 在影响机制方面的研究很少涉及森林CSOC、DSOC动态与植被因子、土壤因子间的相互关系(王淑芳等, 2012). ...
Species-specific and general allometric equations for estimating tree biomass components of subtropical forests in southern China 1 2016
 ,
, ,1,2,3,*
,1,2,3,*
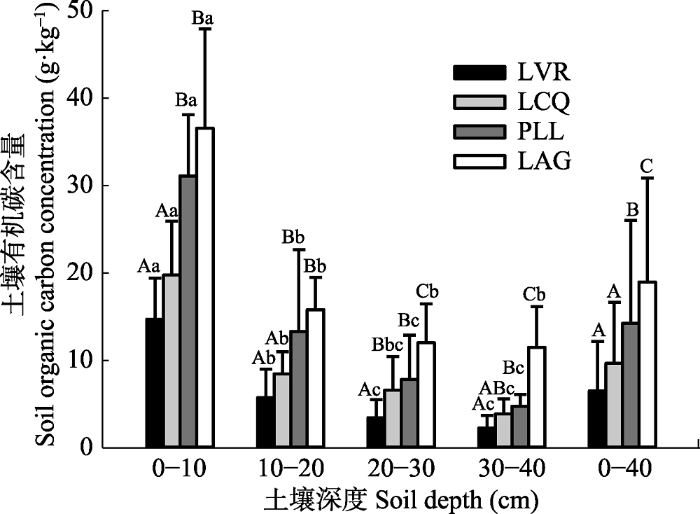 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}