 ,1*, 刘玉洪1,2, 鲁志云1,2, 谢有能31
,1*, 刘玉洪1,2, 鲁志云1,2, 谢有能31 2
3
Species composition and community structure of a 20 hm2 plot of mid-mountain moist evergreen broad-leaved forest on the Mts. Ailaoshan, Yunnan Province, China
WEN Han-Dong1,2, LIN Lu-Xiang1, YANG Jie1, HU Yue-Hua1, CAO Min,1*, LIU Yu-Hong1,2, LU Zhi-Yun1,2, XIE You-Neng31 2
3
通讯作者: (caom@xtbg.ac.cn)
编委: 郝占庆
责任编辑: 王葳
| 基金资助: |
Online:2018-04-20
| Fund supported: |

摘要
中山湿性常绿阔叶林是分布在我国亚热带气候区的一种山地森林植被。为监测此类森林的动态过程, 按照Centre for Tropical Forest Science的建设标准, 于2014年在云南省景东县徐家坝附近的中山湿性常绿阔叶林中建立了一块20 hm 2的森林动态样地, 测量并记录了样地中所有胸径(DBH) ≥ 1 cm的树木直径及其分枝的直径, 并对其挂牌标记、定位和鉴定, 分析了其物种组成和群落结构。结果显示: 样地内共有DBH ≥ 1 cm的木本植物独立个体44 168株, 隶属于36科63属104种。重要值最大的前3个科依次是壳斗科、茶科和樟科。在属的水平上, 热带区系成分占总属数的46.0%, 温带区系成分占38.1%。重要值≥1的物种有22个, 占总个体数的84.6%。重要值最大的3个种依次是蒙自连蕊茶(Camellia forrestii)、云南越桔(Vaccinium duclouxii)和多花山矾(Symplocos ramosissima)。胸高断面积之和排名前4名的物种为硬斗柯(Lithocarpus hancei)、木果柯(L. xylocarpus)、变色锥(Castanopsis wattii)和南洋木荷(Schima noronhae), 它们同时也是该群落林冠层的优势树种。样地内所有个体的径级分布呈倒“J”型, 群落有充足的增补个体, 如蒙自连蕊茶和多花山矾等。稀有种(个体数少于20的物种)共计49种, 占总物种数的47.1%。常绿木本植物在样地内分别占总重要值、总多度和总胸高断面积的94.7%、96.2%和94.0%, 优势明显。
关键词:
Abstract
Keywords:
PDF (1213KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
温韩东, 林露湘, 杨洁, 胡跃华, 曹敏, 刘玉洪, 鲁志云, 谢有能. 云南哀牢山中山湿性常绿阔叶林20 hm2动态样地的物种组成与群落结构 . 植物生态学报[J], 2018, 42(4): 419-429 DOI:10.17521/cjpe.2017.0272
WEN Han-Dong, LIN Lu-Xiang, YANG Jie, HU Yue-Hua, CAO Min, LIU Yu-Hong, LU Zhi-Yun, XIE You-Neng.
常绿阔叶林是分布在我国亚热带气候条件下的一种顶极森林植被, 它是全球亚热带大陆东岸湿润气候和季风气候条件下的产物,此类植被的主要分布区在我国(吴征镒, 1980)。根据气候特征的不同, 我国的常绿阔叶林分布区分为两个亚区域, 即东部常绿阔叶林亚区域和西部常绿阔叶林亚区域(吴征镒, 1980)。分布于云南中部哀牢山自然保护区内的常绿阔叶林属于我国西部常绿阔叶林亚区域。其中徐家坝地区的中山湿性常绿阔叶林以木果柯(Lithocarpus xylocarpus)、硬斗柯(L. hancei)等为优势种, 面积约为2 922 hm2 (游承侠, 1983), 是我国此类森林面积最大、森林保护相对完好的地区。该保护区地处我国青藏高原东南侧以及云南亚热带与热带北缘的过渡区, 热带、亚热带、温带(亚高山)区系成分在这里交错汇集, 并孕育了较多的特有成分, 形成了生物多样性极为丰富和植物区系地理成分极为复杂的格局(邱学忠和谢寿昌, 1998)。鉴于该区所处地域的特殊性, 前人在此开展了部分相关研究工作, 如哀牢山中山湿性常绿阔叶林及其次生林的树种多样性(李宏文, 1983; 钱洪强, 1983; 游承侠, 1983; Young & Wang, 1989; Young et al., 1992; Young & Herwitz, 1995; 何永涛等, 2000; 巩合德等, 2011), 附生植物的多样性和分布格局(徐海清和刘文耀, 2005)。然而, 前期的研究大多是基于较小尺度样地水平上的工作, 难以在大尺度上反映森林树种组成的空间分布规律, 也难以揭示树种分布与关键环境因子之间的关系, 更无法检验控制当地树种分布的生态学机制的理论模型和重要假说。
为此, 中国科学院西双版纳热带植物园于2014年在哀牢山生态站(景东县徐家坝)附近的中山湿性常绿阔叶林中建立了一块面积为20 hm2 (400 m × 500 m)的森林动态样地, 目的在于全面分析样地中的木本植物物种组成、个体多度格局、区系成分、物种的优势度特征和径级结构, 并通过与我国东部常绿阔叶林森林动态样地的比较, 鉴别哀牢山中山湿性常绿阔叶林的群落结构特征, 同时也为今后长期森林动态监测及深入探讨亚热带常绿阔叶林生物多样性的形成与维持机制奠定基础。
1 研究方法
1.1 研究区域概况
哀牢山中山湿性常绿阔叶林动态样地(以下简称哀牢山样地)位于哀牢山国家级自然保护区内, 该保护区主要保护亚热带中山湿性常绿阔叶林生态系统和黑冠长臂猿、绿孔雀、灰叶猴等珍贵野生动物。有资料显示, 哀牢山地区有野生种子植物199科956属2 242种206变种(亚种)(朱华和闫丽春, 2009)。哀牢山样地位于该保护区的徐家坝地区, 海拔 2 400-2 600 m。该区属于西南季风气候, 旱季(11- 4月)、雨季(5-10月)分明。该区10年(2002-2011)的气象观测数据显示年降水量为1 778 mm, 86%以上的降水集中在雨季。年平均气温11.3 ℃, 最热月(7月)气温为15.6 ℃, 最冷月(1月)气温为5.7 ℃ (Wu et al., 2014)。林地土壤为山地黄棕壤, 土壤有机质含量高, 含氮量丰富, 土壤偏酸性(pH < 5); 土壤表面由凋落物层(厚3-7 cm)覆盖, 土壤有机碳、全氮和全磷含量分别为12.91%、0.52%和0.06%, 土壤表面的C/N为14.4, 30-50 cm深度C/N为15.3 (Liu et al., 2002)。哀牢山中山湿性常绿阔叶林乔木树种主要由壳斗科、茶科、樟科和木兰科的树种组成, 其中, 壳斗科的硬斗柯、木果柯、变色锥(Castanopsis wattii), 茶科的南洋木荷(Schima noronhae)、舟柄茶(Hartia sinensis)、蒙自连蕊茶(Camellia forrestii), 樟科的黄心树(Machilus bombycina)、黄丹木姜子(Litsea elongata), 木兰科的红花木莲(Manglietia insignis)、多花含笑(Michelia floribunda)是乔木层的主要优势种。此外, 林中藤本植物和附生植物也相当发达, 层间植物以箭竹(Sinarundinaria nitida)(盖度达75%左右)及木质大藤本最为常见, 附生植物以苔藓和蕨类植物为主, 形成奇特的森林景观(邱学忠和谢寿昌, 1998)。
1.2 样地设置和植物调查
哀牢山样地投影面积为20 hm2, 南北(X轴)长500 m, 东西(Y轴)宽400 m, 样地的具体位置为101.03°-101.03° E, 24.53°-24.54° N, 样地内最低海拔为2 472 m, 最高海拔为2 628 m, 海拔高差为156 m, 平均海拔2 550 m。样地的地形复杂, 有3沟3脊, 坡向总体上呈东西走向(东高西低)(图1)。图1
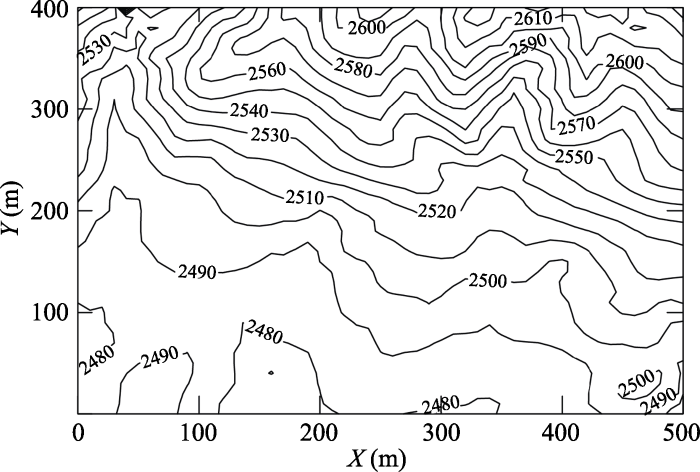 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1哀牢山样地等高线图。
Fig. 1The topographic map of Ailaoshan forest dynamics plot.
样地建设技术按照Center for Tropical Forest Science (CTFS)的建设标准的技术规范进行(Condit, 1998)。采用全站仪(Topcon GTS-336, Topcon Positioning Systems, Tokyo, Japan)将整个样地划分为500个20 m × 20 m的样方, 每个样方又分为16个5 m × 5 m的小样方。测量并记录每个5 m × 5 m样方内的所有胸径(DBH) ≥ 1 cm的木本植物(不包括藤本植物和竹类)的空间位置, 并对其进行物种鉴定、编号挂牌, 于高度1.3 m的胸径处漆上红漆, 用测树钢围尺测量植物的胸径, 并记录植物的编号、树种名称、胸径、空间坐标和生存状态。
1.3 数据处理
物种重要值(IV)的计算公式(叶万辉等, 2008)为:IV = (RD + RF + RP)/3
其中, RD为相对密度, RF为相对频度, RP为相对显著度。计算RF的总样方数为500个(20 m × 20 m), RD = (某种植物的个体数/全部植物的个体数) × 100, RF = (该种的频度/所有种的频度总和) × 100, RP = (样方中该种个体胸高断面积和/样方中所有个体胸高断面积的总和) × 100。
科的重要值(FIV)计算公式(Linares-Palomino & Alvarez, 2005)为:
FIV = RΔ + RD + RA
其中RΔ为相对多样性, 是指一个科的树种数占总树种数的百分比。RD为相对密度, 仅计算独立个体的数量。RA为相对优势度, 计算RA用的胸高断面积包括分枝的断面积。
物种多样性指数采用Shannon-Wiener指数(H)和Pielou均匀度指数(E)(Magurran, 1988)。
$H\text{=}-\sum\limits_{i=1}^{S}{{{p}_{i}}}\mathrm{ln}({{p}_{i}})$
$E={H}/{\mathrm{ln}S}\;$
其中S为总物种数, pi为属于物种i的个体在全部个体中所占的比例。
所有数据的统计分析和作图利用Excel 2016、Sigma Plot 12.5和R3.6.1软件完成。
2 结果
2.1 物种组成
样地内共有DBH ≥ 1 cm的木本植物104种(包括3个未鉴定种), 隶属于36科63属。从科的分布水平来看, 蔷薇科植物最丰富, 共有10属14种; 樟科次之, 有5属10种; 壳斗科有3属8种; 茶科有6属7种。这4个科在科重要值中排名前4位。从属的分布水平来看, 柯属(Lithocarpus)和冬青属(Ilex)的植物最多, 均有6种; 其次为山矾属(Symplocos), 有5种(表1)。另外, 有55.6%的科及71.4%的属仅有一个物种分布。Table 1
表1
表1哀牢山样地重要值前10位的科
Table 1
| 序号 Rank | 科名 Family | 树种数 No. of species | 属数 No. of genera | 个体数 No. of individuals | 胸高断面积 Basal area (cm²) | 相对多样性 Relative diversity | 相对密度 Relative density | 相对优势度 Relative abundance | 科的重要值 Importance value of the family |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 壳斗科 Fagaceae | 8 | 3 | 6 321 | 7 066 975.19 | 7.92 | 14.31 | 57.94 | 80.18 |
| 2 | 茶科 Theaceae | 7 | 6 | 10 466 | 1 508 836.26 | 6.93 | 23.70 | 12.37 | 43.00 |
| 3 | 樟科 Lauraceae | 10 | 5 | 3 833 | 682 608.98 | 9.90 | 8.68 | 5.60 | 24.18 |
| 4 | 蔷薇科 Rosaceae | 14 | 10 | 2 807 | 407 326.40 | 13.86 | 6.36 | 3.34 | 23.56 |
| 5 | 山矾科 Symplocaceae | 5 | 1 | 7 026 | 287 828.73 | 4.95 | 15.91 | 2.36 | 23.22 |
| 6 | 越桔科 Vacciniaceae | 3 | 1 | 4 861 | 528 954.35 | 2.97 | 11.01 | 4.34 | 18.31 |
| 7 | 杜鹃花科 Ericaceae | 6 | 3 | 2 501 | 495 485.15 | 5.94 | 5.66 | 4.06 | 15.67 |
| 8 | 冬青科 Aquifoliaceae | 6 | 1 | 2 059 | 445 327.40 | 5.94 | 4.66 | 3.65 | 14.25 |
| 9 | 木兰科 Magnoliaceae | 3 | 2 | 913 | 217 984.99 | 2.97 | 2.07 | 1.79 | 6.82 |
| 10 | 忍冬科 Caprifoliaceae | 4 | 1 | 84 | 4 286.15 | 3.96 | 0.19 | 0.04 | 4.19 |
| 合计 Total | 66 | 33 | 40 871 | 11 645 613.60 | 65.34 | 92.55 | 95.49 | 253.38 |
新窗口打开|下载CSV
在已知的101个木本植物中, 包括77个(占总数的76.2%)常绿阔叶树种、23个(占总数的22.8%)落叶阔叶树种和1个(占总数的1.0%)常绿针叶树种。其中, 77个常绿阔叶树种分别占总多度、总胸高断面积和总重要值的96.2%、94.0%和94.7%; 23个落叶阔叶树种分别占总多度、总胸高断面积和总重要值的3.8%、6.0%和5.3%。独立个体数大于1 000的全部为常绿阔叶树种(表2)。落叶树种独立个体数最多的是珍珠花(Lyonia ovalifolia), 有446株; 其次为多花山矾(Symplocos ramosissima), 有322株。
2.2 多度格局
整个样地中, DBH ≥ 1 cm的独立木本植物活个体数为44 168株(如果包括分枝则为69 762株), 样地单位面积上的独立个体数量为2 208株·hm-2 (不包括分枝)。从物种多度的累计分布图(图2)可以看出, 个体数最多的前16个物种占了总个体数的75.6%, 个体数最多的前33个物种占了总个体数的95.0%。整个样地中个体数超过1 000株的物种有12个(表2), 占总物种数的11.5%, 占总个体数的67.7%。其中蒙自连蕊茶个体数量最多, 达到6 387株, 占总个体数的14.5%; 随后依次为云南越桔(Vaccinium duclouxii), 有4 843株(11.0%); 多花山矾, 有3 000株(6.8%); 硬斗柯有2 558株(5.1%)。如果按照Hubbell和Foster (1986)的定义, 将每hm2个体数不多于1株的物种定义为稀有种, 1-10的为偶见种, 多于10株的为常见种, 哀牢山样地内有稀有种49个, 占总物种数的47.1%, 稀有种的多度仅占总多度的0.7%, 占总重要值的1.6%; 偶见种有22种, 占总物种数的22.1%, 偶见种的多度仅占总多度的4.4%, 占总重要值的7.8%; 个体数量仅为1株的有13种, 占物种总数的12.5%。Table 2
表2
表2样地内个体数量大于1 000的树种多度及其胸高断面积
Table 2
| 序号 Rank | 种名 Species | 个体数 Abundance | 分枝数 Ramifications | 平均胸径 Mean of DBH (cm) | 胸高断面积 Basal area (m2) |
|---|---|---|---|---|---|
| 1 | 蒙自连蕊茶 Camellia forrestii | 6 387 | 5 847 | 3.42 | 15.95 |
| 2 | 云南越桔 Vaccinium duclouxii | 4 843 | 6 101 | 6.52 | 52.67 |
| 3 | 多花山矾 Symplocos ramosissima | 3 000 | 272 | 4.41 | 15.37 |
| 4 | 硬斗柯 Lithocarpus hancei | 2 558 | 4 195 | 16.81 | 288.69 |
| 5 | 南亚枇杷 Eriobotrya bengalensis | 2 368 | 229 | 9.40 | 27.27 |
| 6 | 变色锥 Castanopsis wattii | 2 008 | 1 140 | 20.28 | 195.83 |
| 7 | 山矾 Symplocos sumuntia | 1 840 | 452 | 4.26 | 9.46 |
| 8 | 丛花山矾 Symplocos poilanei | 1 551 | 657 | 2.68 | 2.08 |
| 9 | 木果柯 Lithocarpus xylocarpus | 1 540 | 777 | 28.99 | 210.85 |
| 10 | 云南柃 Eurya obliquifolia | 1 446 | 737 | 3.53 | 3.33 |
| 11 | 滇润楠 Machilus yunnanensis | 1 236 | 338 | 8.58 | 21.88 |
| 12 | 舟柄茶 Hartia sinensis | 1 118 | 249 | 13.26 | 29.95 |
| 合计 Total | 29 895 | 20 994 | 122.13 | 873.33 |
新窗口打开|下载CSV
图2
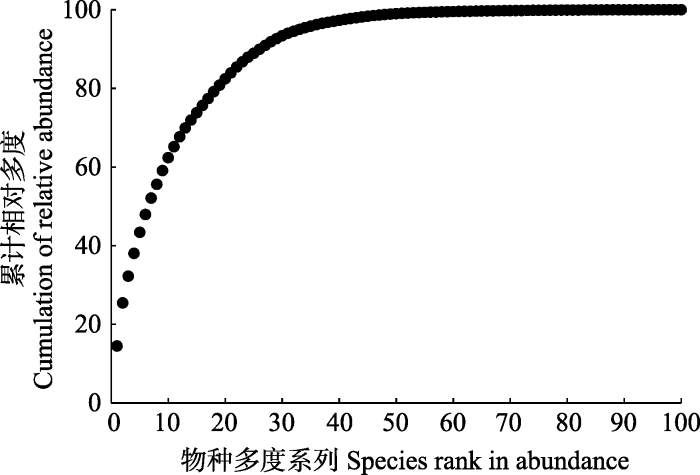 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2哀牢山样地物种多度累计分布图。
Fig. 2Cumulative distribution curve of species abundance in Ailaoshan forest dynamics plot.
样地内104个物种中, 有85种有分枝现象, 占总数的81.7%。分枝数最多的是云南越桔, 其次是蒙自连蕊茶和硬斗柯(表2)。在重要值≥1的物种中, 硬斗柯的分枝率(分枝率=分枝数/独立个体数)最高, 为1.64, 其次是云南越桔和蒙自连蕊茶, 分别为1.26和0.92。因此在重要值≥1的物种中, 这3个种的分枝数和分枝率都处于前3位。77种常绿阔叶树种的平均分枝率为0.51, 而23种落叶阔叶树种的平均分枝率为0.37。
就树种多样性而言, 如果取样地面积为20 hm2, 得到群落的Shannon-Wiener指数为3.32 ± 0.11, Pielou均匀度指数为0.72 ± 0.02。
2.3 属的区系特征
根据吴征镒等(2006)属的分布区类型的划分, 分析了哀牢山样地内63个属的分布区类型(表3)。热带成分的属有29个, 占46.03%; 温带成分的属24个, 占38.10%。热带成分分布最多的是热带亚洲成分, 有12属, 占19.05%; 其次是泛热带和东亚及热带南美间断分布, 各有7属, 各占11.11%。温带成分分布最多的是北温带成分, 有14属, 占22.22%; 其次是东亚及北美间断成分, 有8个属, 占12.70%。由此可以看出, 哀牢山样地内热带成分的属略高于温带成分, 反映了该区系具有从热带向温带的过渡性质。2.4 优势科和优势种分析
对样地内所有科的统计结果显示, 样地内重要值最大的科为壳斗科, 含有3个属8个种, 共有6 321个个体, 其胸高断面积也最大, 达到706.7 m², 占总胸高断面积的57.9%; 茶科是样地内个体数最多的科, 有10 466个个体, 占总个体数的23.7%, 其重要值仅次于壳斗科; 重要值第3的科为樟科; 蔷薇科是样地内物种最丰富的科, 有14个种和10个属, 重要值排名第4。重要值排名前10位的科包含了样地内近92.5%的个体数和63.5%的树种数(表3)。Table 3
表3
表3哀牢山样地种子植物属的分布区类型统计
Table 3
| 序号 Rank | 分布区类型 Areal-type | 属数 No. of genera | 百分比 Percentage |
|---|---|---|---|
| 1 | 世界广布 Widespread | 3 | 4.76 |
| 2 | 泛热带 Pantropic | 7 | 11.11 |
| 3 | 东亚(热带、亚热带) 及热带南美间断 East Asia (Tropical & Subtropical) & Tropical South America disjuncted | 7 | 11.11 |
| 4 | 热带亚洲至热带大洋洲 Tropical Asia to Tropical Australasia Oceania | 2 | 3.17 |
| 5 | 热带亚洲至热带非洲 Tropical Asia to Tropical Africa | 1 | 1.59 |
| 6 | 热带亚洲(热带东南亚至印度-马来, 太平洋诸岛) Tropical Asia (Tropical Southeast Asia to Indo-Malaya & Tropical Southwest Pacific Islands) | 12 | 19.05 |
| 热带成分(2-6)小计 Tropical elements (2-6) sub-total | 29 | 46.03 | |
| 7 | 北温带 North Temperate | 14 | 22.22 |
| 8 | 东亚及北美间断 East Asia & North America disjuncted | 8 | 12.70 |
| 9 | 旧世界温带 Old World Temperate | 2 | 3.17 |
| 温带成分(7-9)小计 Tropical elements(7-9) sub-total | 24 | 38.10 | |
| 10 | 东亚 East Asia | 7 | 11.11 |
| 合计 Total | 63 | 100.00 | |
新窗口打开|下载CSV
样地内重要值≥1的树种有22种, 这些树种的个体数和胸高断面积分别占样地总个体数和总胸高断面积的84.6%和88.4%。重要值排名前3位的物种为蒙自连蕊茶、云南越桔和多花山矾, 均是小乔木层树种, 其胸高断面积仅占样地总胸高断面积的6.9% (表4)。胸高断面积大于1 m2·hm-2的物种有11个, 排名前4位的物种为硬斗柯(14.4 m2·hm-2)、木果柯(10.5 m2·hm-2)、变色锥(9.8 m2·hm-2)和南洋木荷(4.9 m2·hm-2), 胸高断面积和占样地总胸高断面积的65.1%。表明这4个种在该群落中占据了绝对优势, 是该类森林的优势树种。尽管蒙自连蕊茶的重要值在样地内排第一, 但由于蒙自连蕊茶处于群落林冠下层, 多为小乔木, 数量多但平均胸径小(平均胸径为3.42 cm), 远低于处于群落林冠上层的硬斗柯(平均胸径为16.81 cm)、木果柯(平均胸径为28.99 cm)、变色锥(平均胸径为20.28 cm)和南洋木荷(平均胸径为30.72 cm)等树种, 后4种对群落环境的影响也远远大于前者。总体来看, 样地的森林群落类型是以硬斗柯、木果柯、变色锥和南洋木荷等壳斗科与茶科植物为优势树种的亚热带中山湿性常绿阔叶林。
Table 4
表4
表4哀牢山样地重要值前10位的树种
Table 4
| 序号 Rank | 种名 Species | 个体数 No. of individuals | 相对密度 Relative density | 相对频度 Relative frequency | 相对显著度 Relative prominence | 重要值 Importance value |
|---|---|---|---|---|---|---|
| 1 | 蒙自连蕊茶 Camellia forrestii | 6 387 | 14.46 | 5.11 | 0.01 | 6.53 |
| 2 | 云南越桔 Vaccinium duclouxii | 4 843 | 10.96 | 4.27 | 0.04 | 5.09 |
| 3 | 多花山矾 Symplocos ramosissima | 3 000 | 6.79 | 4.14 | 0.01 | 3.65 |
| 4 | 硬斗柯 Lithocarpus hancei | 2 558 | 5.79 | 4.58 | 0.24 | 3.53 |
| 5 | 变色锥 Castanopsis wattii | 2 008 | 4.55 | 4.87 | 0.16 | 3.19 |
| 6 | 南亚枇杷 Eriobotrya bengalensis | 2 368 | 5.36 | 3.98 | 0.02 | 3.12 |
| 7 | 木果柯 Lithocarpus xylocarpus | 1 540 | 3.49 | 4.58 | 0.17 | 2.75 |
| 8 | 山矾 Symplocos sumuntia | 1 840 | 4.17 | 3.62 | 0.01 | 2.60 |
| 9 | 滇润楠 Machilus yunnanensis | 1 236 | 2.80 | 4.48 | 0.02 | 2.43 |
| 10 | 丛花山矾 Symplocos poilanei | 1 551 | 3.51 | 3.06 | 0.00 | 2.19 |
| 合计 Total | 27 331 | 61.88 | 42.69 | 0.68 | 35.08 |
新窗口打开|下载CSV
2.5 种-面积曲线
从种-面积曲线(图3)来看, 在取样面积小于 2.5 hm2的初始阶段, 物种数随着取样面积的增加而迅速上升; 当取样面积超过2.5 hm2时, 物种数随着取样面积增加的速度减缓。当取样面积达到7.5 hm2时, 已涵盖样地总物种数的80%左右; 当取样面积达到15.4 hm2时, 涵盖了样地95%的物种数, 取样面积从15.4 hm2增加到20 hm2时, 仅增加了6个物种(5%)。从个体数-面积曲线来看, 随着取样面积的增加, 个体数随取样面积同步增加, 呈直线上升趋势, 说明个体密度在样地中的分布基本还是均匀的。图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3哀牢山样地种-面积曲线和个体数-面积曲线。CI, 置信区间。
Fig. 3Species-area curve and individual-area curve in Ailaoshan forest dynamics plot. CI, confidence interval.
2.6 径级结构
样地内所有木本植物(DBH ≥ 1 cm)的总胸高断面积为1 220 m2, 平均每hm2胸高断面积为61.0 m2·hm-2, 平均胸径为9.19 cm。胸径最大的个体是变色锥, 为171.0 cm。全部个体的径级分布呈现明显的倒“J”型, 随着DBH等级的增加, 个体数量逐渐减少(图4A)。从径级结构可以看出, DBH < 10 cm的个体数量最多, 有29 602株, 占总个体数量的67.0%。而DBH ≥ 10 cm、≥ 30 cm、≥ 50 cm和≥ 100 cm的个体数分别占总个体数的33.0%、9.1%、2.2%和0.1%。在DBH ≥ 100 cm的39株植物中, 变色锥有15株, 硬斗柯有11株, 木果柯有10株, 南洋木荷、尖叶桂樱(Laurocerasus undulata)和珍珠花各有1株。图4
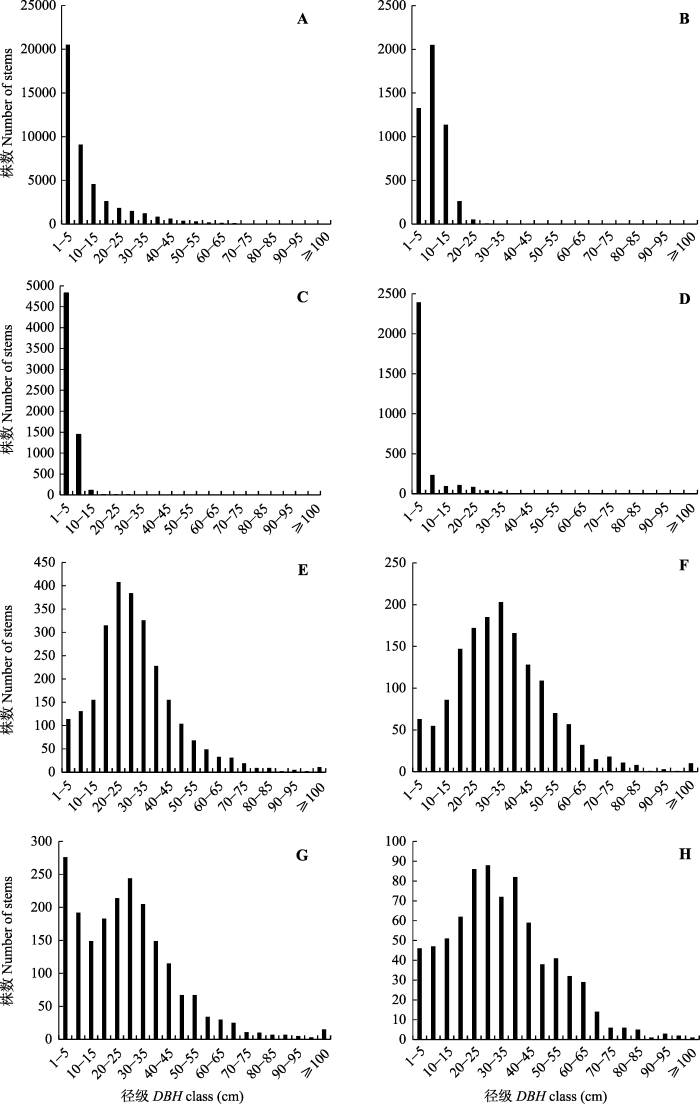 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4哀牢山样地所有个体和主要树种的径级结构图。A, 所有个体。B, 云南越桔。C, 蒙自连蕊茶。D, 多花山矾。E, 硬斗柯。F, 木果柯。G, 变色锥。H, 南洋木荷。
Fig. 4Diameter at breast height (DBH)-class distribution of all individuals and dominant species in Ailaoshan forest dynamics plot. A, All individuals. B, Vaccinium duclouxii. C, Camellia forrestii. D, Symplocos ramosissima. E, Lithocarpus hancei. F, Lithocarpus xylocarpus. G, Castanopsis wattii. H, Schima noronhae.
对样地内主要树种的径级结构的分析结果(图4)表明, 各树种的径级结构可以归纳为4种类型: (1)倒“J”型分布, 此类物种为小乔木层(次林层)的优势种, 径级结构的形状类似于倒置的“J”型, 以1 cm ≤ DBH ≤ 5 cm的个体数最多, 并随着DBH的增加个体数量逐渐下降, 表明物种有足够数量的幼树和幼苗, 能够实现种群的自我更新, 代表种为蒙自连蕊茶和多花山矾(图4C、4D); (2) “泊松”型分布, 如云南越桔(图4B), 它们也多是次林层的优势物种, 因树种的生理限制, 几乎没有大径级; (3)单峰型(正态型)分布, 此类物种为主林层的优势物种, 其径级结构类似于正态分布或偏正态分布, 结构稳定, 个体主要集中在中径级(15 cm ≤ DBH ≤ 40 cm), 小径级与大径级相对较少, 代表种为硬斗柯、木果柯和南洋木荷(图4E、4F、4H); (4)双峰型分布, 此类物种也为主林层的优势物种, 径级结构有两个峰值, 如变色锥(图4G), 在1 cm ≤ DBH ≤ 5 cm与 25 cm ≤ DBH ≤ 30 cm间出现两个峰值, 中径级的个体数较少。
2.7 垂直结构
样地中胸径≥1 cm木本植物的垂直结构可分为3个层次, 即林冠层(高度≥15 m)、亚冠层(5 m ≤ 高度< 15 m)、灌木层(高度< 5 m)。林冠层主要由硬斗柯、木果柯、变色锥和南洋木荷等组成; 亚冠层由蒙自连蕊茶、云南越桔、薄叶马银花(Rhododendron leptothrium)等组成; 灌木层则以多花山矾、云南柃(Eurya obliquifolia)、长柱十大功劳(Mahonia duclouxiana)为优势种。此外, 箭竹(此次未列入清查范围)在局部片区的灌木层也能形成密集的覆盖。总体看来, 各层次的优势种均为常绿植物, 表现出典型的常绿阔叶林特征。3 比较和讨论
哀牢山样地与天童山样地、古田山样地、鼎湖山样地同属于亚热带常绿阔叶林植被型, 但哀牢山样地属于西部常绿阔叶林区域的山地类型, 天童山样地和古田山样地属于东部常绿阔叶林区域中亚热带常绿阔叶林地带, 而鼎湖山样地属于南亚热带常绿阔叶林地带(吴征镒, 1980)。通过对比4块样地的研究结果, 能够更全面地反映亚热带常绿阔叶林的 共性与差异, 同时也能较好地反映各自的群落学 特征。哀牢山样地的物种数低于天童山样地、古田山样地和鼎湖山样地(叶万辉等, 2008; 祝燕等, 2008; 杨庆松等, 2011, 以下讨论中所涉及此3块样地的数据均来自于这3篇文章, 不再标注), 我们分析可能与哀牢山样地海拔高度明显高于这3个样地有关。如果不算分枝数量, 哀牢山样地的总个体数也低于另外3个样地, 也就是说, 哀牢山样地无论是物种数还是总个体数都没有另外3个样地高(表5)。
Table 5
表5
表5哀牢山样地与其他3个东部常绿阔叶林动态样地特征的比较
Table 5
| 样地名称 Name of plot | 经纬度 Longitude, Latitude | 平均海拔 Average altitude (m) | 年平均气温 Mean annual air temp- erature (℃) | 年降水 Annual precipi- tation (mm) | 样地面积 Area of plot (hm2) | 物种数量 No. of species | 个体数量 No. of free standing stems | 稀有种 数量/比例 No. and percentage of rare species | 落叶树种 数量/比例 No. and percentage of deciduous tree species | 平均每公顷的胸高断面积 Average area of stems at breast height per hm2 (m2·hm-2) | 数据来源 Reference |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 哀牢山 Ailaoshan | 101.03°- 101.03° E, 24.53°- 24.54° N | 2 550 | 11.3 | 1 778 | 20 | 104 | 44 168 | 49/47.1% | 23/22.8% | 61.0 | 本研究 This study |
| 天童山 Tiantongshan | 121.86°- 122.03° E, 29.99°- 30.06° N | 447 | 20 | 152 | 94 603 | 55/36.2% | 69/42.8% | Yang et al., 2011 | |||
| 古田山Gutianshan | 118.12°- 118.23° E, 29.28°- 29.35° N | 581 | 15.3 | 1 964 | 24 | 159 | 140 700 | 59/37.1% | 79/49.7% | 36.9 | Zhu et al., 2008 |
| 鼎湖山 Dinghushan | 112.51°- 112.56° E, 23.16°- 23.19° N | 350 | 20.9 | 1 985 | 20 | 210 | 71 617 | 110/52.3% | 30.2 | Ye et al., 2008 |
新窗口打开|下载CSV
从稀有种所占比例来看, 哀牢山样地高于天童山样地和古田山样地, 但是低于鼎湖山样地(表5)。如果只看样地内总个体数为1株的植物, 哀牢山样地有13种, 与天童山样地(13种)相同, 介于古田山样地(11种)和鼎湖山样地(27种)之间。这一方面显示哀牢山样地分布纬度偏南, 致使稀有种比例高于天童山样地和古田山样地, 另一方面, 若与纬度相近的鼎湖山样地相比较, 其较高的海拔导致了稀有种的比例下降, 低于鼎湖山样地。
落叶树种中, 哀牢山样地无论是物种数量还是所占比例都低于天童山和古田山样地(表5), 可能与哀牢山样地的低纬度特性导致其常绿树种数量和比例上升有关。但总体来说, 落叶树种的个体数量在3个样地中所占的比例都相对较低, 哀牢山样地为3.8%、天童山样地为8.8%、古田山样地为14.1%, 说明常绿树种无论在物种或者个体数量(比例)上均占据绝对优势, 这表现出典型亚热带常绿阔叶林的共性特征。
从物种属的分布区类型看, 哀牢山样地有63个属, 热带成分占46.0%, 温带成分占38.1%, 这与古田山样地(热带成分占53.9%, 温带成分占43.1%)和天童山样地(热带成分占52.1%, 温带成分占42.6%)属的区系特征相似, 区别于鼎湖山样地以热带成分(89.9%)为主的特征。说明哀牢山样地与古田山样地和天童山样地一样, 具有热带区系和温带区系的相互交融, 反映了样地内植物区系具有从热带向温带过渡的特征。此外, 哀牢山样地的海拔高度显著高于其他3个样地, 这似乎也导致了其热带成分比例低于其他3个样地, 并且其物种数量也趋于减少。但联系哀牢山样地中的落叶树种比例显著低于天童山样地和古田山样地的事实, 这似乎又表现了该样地强烈的热带性质。从种-面积曲线来看, 哀牢山样地与鼎湖山样地很相似, 取样面积小于2.5 hm2, 物种数随着取样面积的增加而迅速上升; 取样面积达到2.5 hm2之后, 增速减缓, 直至取样面积接近20 hm2时才趋于稳定。综合上述分析结果来看, 哀牢山样地的物种组成、稀有种、落叶成分、属的分布区类型主要受到纬度、海拔高度的影响, 这两者的协同作用导致了现在的分布样式。它同时体现了西部常绿阔叶林与东部常绿阔叶林的共性特征(如常绿树种占绝对优势), 也体现了山地抬升之后产生的隔离效应和温度降低导致的热带性质减弱效应(如物种数量少、在属的水平上热带成分比例偏低)。
径级结构是植物群落稳定性和生长发育状况的重要指标(叶万辉等, 2008)。哀牢山样地内DBH ≥ 1 cm的树木平均每hm2的胸高断面积显著高于古田山样地和鼎湖山样地, 并且哀牢山样地的个体数量还低于后两者的个体数量(表5), 这反映了哀牢山样地森林群落的成熟性和原始性。
哀牢山样地有85种植物有分枝现象, 占总数的81.7%。从个体数量来看, 分枝率为57.9%。在重要值≥1的物种中, 硬斗柯的分枝率最高, 其次是云南越桔和蒙自连蕊茶。这三者的重要值分别占据了所有树种重要值的第4、第2、第1位。天童山样地有112种植物有萌枝现象, 占总数的73.7%。个体分枝率为40.1%。萌枝率最高的分别是长叶石栎、马银花、薄叶山矾, 而这3个树种的重要值分别排在第5、第14、第13位。鼎湖山样地的个体分枝率为12.6%。总体来说, 哀牢山样地的分枝率更高一些。关于分枝率高的原因, 推测主要取决于两个因素: 一是树种自身的生物学特性; 二是群落历史上受干扰的强度、频度和干扰方式。但目前还很难找到直接的科学证据, 有待今后深入探讨。
2008年中国科学院西双版纳热带植物园曾经在哀牢山的相同地点、相同群落类型中建立了一个6 hm2的森林动态样地(巩合德等, 2011)。与这个6 hm2样地的研究结果相比, 现在建立的20 hm2样地在物种组成和物种多度格局等方面都存在较大差异。从物种组成来看, 6 hm2样地DBH ≥ 1 cm的木本植物包括了68种, 20 hm2大样地包括了104种。在6 hm2样地中, 重要值排前两名的树种是林冠层树种硬斗柯和变色锥, 而在20 hm2样地中, 重要值排前两名的树种变成了亚冠层树种(蒙自连蕊茶和云南越桔), 原因是后两者在20 hm2大样地内的植株数量更多, 似乎暗示20 hm2的样地面积能够更加完整地反映哀牢山中山湿性常绿阔叶林的树种多样性和群落结构特征。原6 hm2样地的研究得出了哀牢山常绿阔叶林为多优种林, 优势树种不明显的结论(巩合德等, 2011), 但基于20 hm2样地的研究表明, 此类森林的优势树种是明显的, 其林冠层优势树种即为硬斗柯、木果柯、变色锥和南洋木荷。

扫码加入读者圈
听语音, 看问答
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
URL [本文引用: 1]

This book provides a detailed account of the methods used to establish the Barro Colorado Island plot which has records on 32565000 individual trees and constitutes the largest original forest census in the world. The methodologies used at 11 other large plots that form part of the Center for Tropical Forest Science network are reviewed. Details are given on the design and purpose of a large for...
[本文引用: 3]
[本文引用: 3]
DOIURL [本文引用: 1]
分别测定了滇中地区两类代表性群落即高原水平带上的半温润常绿阔叶林和山地垂直带上的中山温性常绿阔叶林的树种多样性(Shannon-Wiener)指数H,Simpson指数D和α指数),并进行了比较研究。结果表明,中山温性常绿阔叶林的树种多样性(H=2.53±0.14;D=11.76±2.85;α=8.12±1.72)远高于半温润常绿阔叶林(H=1.52±0.35;D=3.88±1.90;α=8.12
DOIURL [本文引用: 1]
分别测定了滇中地区两类代表性群落即高原水平带上的半温润常绿阔叶林和山地垂直带上的中山温性常绿阔叶林的树种多样性(Shannon-Wiener)指数H,Simpson指数D和α指数),并进行了比较研究。结果表明,中山温性常绿阔叶林的树种多样性(H=2.53±0.14;D=11.76±2.85;α=8.12±1.72)远高于半温润常绿阔叶林(H=1.52±0.35;D=3.88±1.90;α=8.12
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
A phytosociological study was conducted in six 1-ha seasonally dry forest samples in selected locations on the Cerros de Amotape Cordillera in north-western Peru. This is the first quantitative study of this vegetation formation in the region, and the findings indicate higher density and richness values than previously thought for the country. The plots varied markedly in species richness from 6 to 25 species, with densities per hectare between 55 and 524 individuals. The most important families were the Bombacaceae, Fabaceae, Bignoniaceae, Combretaceae, Burseraceae and Capparidaceae. At the species level, Caesalpinea glabrata, Tabebuia spp., Terminalia valverdeae, Bursera graveolens, Eriotheca ruizii and Ceiba trichistandra are the most widespread and abundant species. Sorensen similarity indices consistently grouped drier, species poor west facing plots together, while the plots located in more mesic areas either east facing or on the ridges of the Amotape Cordillera were more species rich and similar between them.
DOIURL [本文引用: 1]
Montane evergreen broad-leaved forest was the natural vegetation of subtropical, mountainous central and southern Yunnan, SW China. Pristine natural forest, dominated by Fagaceae ( Lithocarpus xylocarpus, Lithocarpus chintungensis and Castanopsis wattii), is still present at Ailao Mountain National Nature Reserve. Within this forest, older stands are of relatively low density, with scattered large trees, compared with stands of higher density. Biomass and nutrient content are examined in this paper for two stands, one each of relatively high and low density. Tree aerial biomass components were calculated from a stand census applied to previously derived regressions of biomass components and tree size. Tree roots were obtained from cores (20 per stand). Other biomass contributions sampled were: live shrubs (five 4 m 2 samples); herb (ten 1 m 2 samples); dead wood (ten 4 m 2 samples); litterfall (fifteen 1 m 2 samples, monthly); forest floor litter (ten 0.25 m 2 samples, 3-monthly). Three nutrient samples were analysed for each tissue per species for both stands. Total live biomass was 503 and 285 t ha 611 for the high and low density stands, respectively. The biomass ratio of stem-wood to branch-wood and leaf biomass was 10:1 and 43.4:1 in the former and 14:1 and 29.7:1 in the latter, respectively. Standing crops of litter and dead wood were both less in the former (10.3 and 46.2 t ha 611) than the latter (12.9 and 98.5 t ha 611). Litterfall in the high density stand was 5.4 compared with 7.1 t ha 611 in the low density. Total nutrient content in living and dead wood materials was 280 and 195 t ha 611, respectively. The sequence of inorganic nutrient element content decreased in the order, leaves>branches>roots>stems. Elemental nutrient content of the total stand decreased in the order of C>Ca>N>Mg>Al>P>Fe>Mn. The elements C, Ca and N were mainly in stems, while Al and Fe were mainly in roots. Nutrients in dead wood comprised 9.2 and 40.6% of the total nutrients in living trees and nutrients returned to soil through litterfall were estimated at 3120 and 3699 kg ha 611 yr 611 in high and low density stands, respectively.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 3]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 2]
鼎湖山南亚热带常绿阔叶林是北回归线附近保存较完好的地带性植被,是在南亚热带季风湿润型气候条件下发育的植被类型,具有热带向亚热带的过渡性质,群落结构相对复杂,组成种类相对丰富。参照巴拿马巴洛科罗拉多岛(Barro Colorado Island,BCI)50hm^2热带雨林样地的技术规范,采用中国森林生物多样性监测网络的统一调查研究方法,于2005年在鼎湖山南亚热带常绿阔叶林建立了20hm^2固定监测样地,调查并鉴定了样方内胸径(Diameter at breast height,DBH)大于1cm的木本植物。首次调查结果表明,群落内共有木本植物210种,71617个活的个体,分属于56科119属。从乔木区系的组成及其特点可以看出,其南亚热带的区系成分占绝对优势,并呈现出由亚热带向热带过渡的趋势。群落垂直结构复杂,地上成层现象较明显,乔木可分为3层,其中重要值最大的锥(Casttmopsis chinensis)、木荷(Schima sumrba)和黄杞(Engelhardtia raxburghiana)均是乔木上层的优势种;中层是群落的主要层,由厚壳桂(Cryptocarya chinensis)、黄叶树(Xamhophyllum hainanense)和华润楠(Machilus chinensis)等中生和耐阴树种组成;下层成分较复杂,物种多样性高,不同地段的物种组成差异较大。样地内物种十分丰富,种.面积曲线拟合显示其物种数量接近于BCI。稀有种比例极高,有110种,占总物种数的52.38%,其中有45%的稀有种源于物种本身的特性,有20%源于区系交汇,人为或自然干扰造成的稀有种占30%以上。样地中所有个体的径级分布(以1cm等级排列)明显呈倒“J”形,表示群落稳定与正常生长状态。根据对优势种径级结构的分布分析,将各树种的径级结构归纳为4种类型:1)峰型(中径级个体储备型),此类物种为乔木上层优势种;2)倒“J”型(正常型),此类物种为乔木
DOIURL [本文引用: 2]
鼎湖山南亚热带常绿阔叶林是北回归线附近保存较完好的地带性植被,是在南亚热带季风湿润型气候条件下发育的植被类型,具有热带向亚热带的过渡性质,群落结构相对复杂,组成种类相对丰富。参照巴拿马巴洛科罗拉多岛(Barro Colorado Island,BCI)50hm^2热带雨林样地的技术规范,采用中国森林生物多样性监测网络的统一调查研究方法,于2005年在鼎湖山南亚热带常绿阔叶林建立了20hm^2固定监测样地,调查并鉴定了样方内胸径(Diameter at breast height,DBH)大于1cm的木本植物。首次调查结果表明,群落内共有木本植物210种,71617个活的个体,分属于56科119属。从乔木区系的组成及其特点可以看出,其南亚热带的区系成分占绝对优势,并呈现出由亚热带向热带过渡的趋势。群落垂直结构复杂,地上成层现象较明显,乔木可分为3层,其中重要值最大的锥(Casttmopsis chinensis)、木荷(Schima sumrba)和黄杞(Engelhardtia raxburghiana)均是乔木上层的优势种;中层是群落的主要层,由厚壳桂(Cryptocarya chinensis)、黄叶树(Xamhophyllum hainanense)和华润楠(Machilus chinensis)等中生和耐阴树种组成;下层成分较复杂,物种多样性高,不同地段的物种组成差异较大。样地内物种十分丰富,种.面积曲线拟合显示其物种数量接近于BCI。稀有种比例极高,有110种,占总物种数的52.38%,其中有45%的稀有种源于物种本身的特性,有20%源于区系交汇,人为或自然干扰造成的稀有种占30%以上。样地中所有个体的径级分布(以1cm等级排列)明显呈倒“J”形,表示群落稳定与正常生长状态。根据对优势种径级结构的分布分析,将各树种的径级结构归纳为4种类型:1)峰型(中径级个体储备型),此类物种为乔木上层优势种;2)倒“J”型(正常型),此类物种为乔木
[本文引用: 2]
[本文引用: 2]
DOIURL [本文引用: 1]
A cooperative U.S.A.-China ecological study took place at the Xujiaba Nature Sanctuary on the northern crest of the Ailao Shan (mountains) in central-west Yunnan. By making forest access difficult and keeping human immigration low, the mountainous topography of Yunnan has assisted in protecting forest cover. This project studied the vegetational and bird structure and composition of an old growth and a secondary evergreen broad-leaved forest. The study found that the old growth had better developed layers, canopy stems were wider and taller, and there was a greater diversity of bird species. The secondary forest had a higher stem density in the canopy, more even-aged stems in the canopy, more coppiced stems in the canopy, and a greater canopy and overall basal area. Both forest types maintained a high diversity of woody species, but different species were dominant in each forest type and in the canopy and subcanopy of the forests. Natural forests are rare in China and are dwindling quickly. Most natural forest is found in frontier regions and it is here where research and management efforts are needed most urgently. In the nature reserves more ecological research, conservation education, and greater efforts to understand local demands, use, and knowledge about the forests are needed. /// Une étude écologique a été effectuée par les Etats-Unis et la Chine dans le sanctuaire naturel de Xujiaba sur la crête septentrionale des monts Ailao (Ailao Shan), au centre-ouest de la province du Yunnan. La topographie montagneuse du Yunnan a aidé à protéger la couverture forestière en rendant l'accès aux forêts difficile et en limitant ainsi l'immigration humaine. Ce projet a étudié la structure de la végétation et des populations d'oiseaux, ainsi que la composition d'une forêt primaire et d'une forêt secondaire composée d'arbres à larges feuilles persistantes. Cette étude a montré que la forêt primaire avait des couches mieux développées, que les tiges de la couronne étaient plus larges et plus hautes et que les espèces d'oiseaux étaient plus diverses. La forêt secondaire avait une plus haute densité de tiges dans la couronne, plus de tiges d'09ge uniforme dans la couronne, plus de tiges en taillis dans la couronne, ainsi qu'une plus grande surface de couronne et de base. Les deux types de forêt ont maintenu une grande diversité d'espèces ligneuses, mais différentes espèces étaient dominantes dans chaque type de forêt et dans la couronne et la sous-couronne des forêts. Les forêts naturelles sont rares en Chine et disparaissent rapidement. La plupart des forêts naturelles se trouvent dans les régions frontalières et c'est donc là où le besoin de travaux de recherche et de gestion sont les plus urgents. Un accroissement de la recherche écologique et de l'éducation sur la conservation, ainsi que de plus gros efforts visant à comprendre les besoins, l'utilisation et la connaissance des forêts au niveau local sont nécessaires dans les réserves naturelles. /// Im Xujiaba Naturschutzgebiet am n02rdlichen Kamm der Ailao Berge in Zentralwest Yünnan wurde in Zusammenarbeit zwischen den USA und China eine 02kologische Studie durchgeführt. Die bergige Topographie von Yünnan half, den ursprünglichen Waldbestand zu erhalten; der Zugang zu Waldgebieten ist schwierig und menschlicher Einflu08 blieb gering. Dieses Projekt untersuchte die Vegetationsstruktur und die Vogelbest01nde für den Urbewuchs und für gemischt bewachsene Waldgebiete mit altem Baumbestand und mit immergrünen Laubb01umen als Sekund01rbewuchs. Die Studie ergab, da08 die Fl01che des alten Baumbestandes eine besser entwickelte Struktur des Laubwerkes hatte, die St01mme waren dicker und h02her, und es gab eine gr0208ere Vielfalt unter den Vogelarten. Im Sekund01rwald standen die St01mme dichter, der Wuchs war gleichaltriger, es gab mehr Unterholz, Blattd01cher und ihre Grundfl01chen waren allgemein gr02sser. Beide Waldarten hatten viele unterschiedliche Baumsorten, jedoch die Hauptsorte war verschieden für jede Waldart. Prim01rw01lder sind in China selten und verringern sich st01ndig. Die meisten Prim01rw01lder sind in Grenzgebieten und hier sind Forschungs-und Managementbemühungen am n02tigsten. In den Schutzgebieten sind 02kologische Forschung und Schulung auf dem Gebiet des Naturschutzes n02tig, ebenso mu08 es verst01rkte Bemühungen geben, lokale Bedürfnisse, wirtschaftliche Nutzung und die forstwissenschaftliche Erkenntnis zu interpretieren.
DOIURL [本文引用: 1]
Many of the natural forested ecosystems that still remain in mainland China are being cleared with potentially detrimental effects on woody plant species diversity on both local and regional scales. The most extensive stand of subtropical broad-leaved forest remaining in China is located in Yunnan Province. In an effort to document the influence of human-induced disturbance on Yunnan's woody flora, floristic inventories were conducted in a stand of primary forest and in regrowth stands located in its interior and along its outer margin in the Xujiaba Nature Sanctuary in the Ailao Mountain Range. Of particular interest was the location of the disturbance relative to the primary forest source area. A total of 134 woody plant species representing 74 genera and 43 families were recorded. The floristics of the two regrowth stands were significantly different from each other, with < 10% of their respective floras comprised of co-occurring species. The interior regrowth stand had a higher number of co-occurring species with the primary forest; however, > 40% were still non-co-occurring. The principal families represented in the primary forest and the interior regrowth stand were Aquifoliaceae, Berberidaceae, Fagaceae, Lauraceae, Rosaceae, Smilacaceae, Symplocaceae, Theaceae, and Vacciniaceae. The three dominant species with relative importance values ranging from > 5% to 18% in both the primary forest and the interior regrowth stand were Castanopsis wattii, Lithocarpus jingdongensis, and Symplocos sumuntia. The edge regrowth stands had the lowest species diversity and were dominated by the native pine Pinus yunnanensis, with a relative importance of 24%. The principal families represented in the edge regrowth stand were Betulaceae, Ericaceae, Fagaceae, Myricaceae, Pinaceae, and Theaceae. Only the Fagaceae and Theaceae were well-represented in all three stands. The results of the study document the low species diversity in post-cutting regrowth on the margins of the primary forest as compared with post-cutting regrowth in the forest interior.
DOIURL [本文引用: 1]
In southern China, when the original broadleaf evergreen forest is destroyed, pines are generally the first secondary tree species to invade the site. In Yunnan Province (southwestern China), Pinus yunnanensis is the principal secondary tree species. We examined two different secondary pine stands, one on the eastern slope and the other on the western slope of the Ailao Shan (mountains) of Yunnan in the Xujiaba Nature Reserve. Data was collected to determine the structure and composition of secondary forests after the destruction of the primary broadleaf evergreen forest. When compared, the primary broadleaf evergreen forest exhibits a much higher biodiversity and overall density than the secondary pine forests. When the two secondary stands are compared with each other, differences in structure and composition indicate that the Ailao Shans may produce a major climatic border between China's southwestern and southeastern monsoon systems. In addition, bird species were recorded in the stands and observations indicate that the Ailao Shans also mark a significant migratory border. Yunnan is an important biological region in China because it harbors over 50% of the country's plant and animal species. The proper management and protection of Xujiaba and other Nature Reserves in the Province will help maintain Yunnan's biological wealth. The protection of forest cover will also minimize environmental degradation, such as soil erosion, which is being experienced in other developing regions. As China aggressively modernizes, forest management based on ecological data is vital in order to protect remaining forests and help develop a sustainable rural economy.
[本文引用: 1]
[本文引用: 1]
DOIURL
亚热带常绿阔叶林是世界上主要植被类型之一,集中分布于我国,其中以中亚热带的常绿阔叶林最为典型.为了更好地研究常绿阔叶林森林生物多样性维持机理,按照CTFS(Centre for Tropical Forest ScieIlce)样地建设的标准,于2004年11月~2005年9月在浙江开化古田山国家级自然保护区建立了常绿阔叶林24 hm2永久样地.该文对样地内胸径≥1 cm的木本植物进行了统计,初步分析了古田山森林样地(Gutianshan (GTS) forest plot)的群落组成与空间结构.群落的区系类型以热带成分比较多,在属的水平上53个是热带分布,44个是温带分布.共有159种,隶属于49科103属,总计140 700株,以常绿树种为主(91个物种,占总优势度的90.6%,重要值为85.6%,占样地总个体数的85.9%);样地群落有明显的优势物种和大量稀有种(Rare species,每hm2个体数小于1).甜槠(Castanopsis eyrei)、木荷(Schima superba)和马尾松(Pinus massoniana)在群落中占有主要优势;稀有种占总物种数的37.1%(59/159);具有温带落叶阔叶林和热带雨林的相关特征,较好地代表了中亚热带常绿阔叶林群落.垂直结构由林冠层(63个种)、亚乔木层(70个种)和灌木层(26个种)组成.样地所有木本植物物种总径级分布呈倒"J"形,群落更新良好.几个优势种空间分布随着径级增大,聚集程度降低;并且它们的分布表现出与生境紧密关联,不同的物种表现出对生境不同的偏好.该文最后系统地比较了大样地方法与传统取样方法.
DOIURL
亚热带常绿阔叶林是世界上主要植被类型之一,集中分布于我国,其中以中亚热带的常绿阔叶林最为典型.为了更好地研究常绿阔叶林森林生物多样性维持机理,按照CTFS(Centre for Tropical Forest ScieIlce)样地建设的标准,于2004年11月~2005年9月在浙江开化古田山国家级自然保护区建立了常绿阔叶林24 hm2永久样地.该文对样地内胸径≥1 cm的木本植物进行了统计,初步分析了古田山森林样地(Gutianshan (GTS) forest plot)的群落组成与空间结构.群落的区系类型以热带成分比较多,在属的水平上53个是热带分布,44个是温带分布.共有159种,隶属于49科103属,总计140 700株,以常绿树种为主(91个物种,占总优势度的90.6%,重要值为85.6%,占样地总个体数的85.9%);样地群落有明显的优势物种和大量稀有种(Rare species,每hm2个体数小于1).甜槠(Castanopsis eyrei)、木荷(Schima superba)和马尾松(Pinus massoniana)在群落中占有主要优势;稀有种占总物种数的37.1%(59/159);具有温带落叶阔叶林和热带雨林的相关特征,较好地代表了中亚热带常绿阔叶林群落.垂直结构由林冠层(63个种)、亚乔木层(70个种)和灌木层(26个种)组成.样地所有木本植物物种总径级分布呈倒"J"形,群落更新良好.几个优势种空间分布随着径级增大,聚集程度降低;并且它们的分布表现出与生境紧密关联,不同的物种表现出对生境不同的偏好.该文最后系统地比较了大样地方法与传统取样方法.
Tropical Forest Census Plots: Methods and Results from Barro Colorado Island, Panama and a Comparison with Other Plots
1
1998
... 样地建设技术按照Center for Tropical Forest Science (CTFS)的建设标准的技术规范进行(
哀牢山常绿阔叶林树种多样性及空间分布格局
3
2011
... 常绿阔叶林是分布在我国亚热带气候条件下的一种顶极森林植被, 它是全球亚热带大陆东岸湿润气候和季风气候条件下的产物,此类植被的主要分布区在我国(
... 2008年中国科学院西双版纳热带植物园曾经在哀牢山的相同地点、相同群落类型中建立了一个6 hm2的森林动态样地(
... 样地的研究得出了哀牢山常绿阔叶林为多优种林, 优势树种不明显的结论(
哀牢山常绿阔叶林树种多样性及空间分布格局
3
2011
... 常绿阔叶林是分布在我国亚热带气候条件下的一种顶极森林植被, 它是全球亚热带大陆东岸湿润气候和季风气候条件下的产物,此类植被的主要分布区在我国(
... 2008年中国科学院西双版纳热带植物园曾经在哀牢山的相同地点、相同群落类型中建立了一个6 hm2的森林动态样地(
... 样地的研究得出了哀牢山常绿阔叶林为多优种林, 优势树种不明显的结论(
滇中地区常绿阔叶林树种多样性比较研究
1
2000
... 常绿阔叶林是分布在我国亚热带气候条件下的一种顶极森林植被, 它是全球亚热带大陆东岸湿润气候和季风气候条件下的产物,此类植被的主要分布区在我国(
滇中地区常绿阔叶林树种多样性比较研究
1
2000
... 常绿阔叶林是分布在我国亚热带气候条件下的一种顶极森林植被, 它是全球亚热带大陆东岸湿润气候和季风气候条件下的产物,此类植被的主要分布区在我国(
Commonness and rarity in a neotropical forest: Implications for tropical tree conservation In: Soule ME ed. Conservation Biology: Science of Scarcity and Diversity
1986
1
1983
... 常绿阔叶林是分布在我国亚热带气候条件下的一种顶极森林植被, 它是全球亚热带大陆东岸湿润气候和季风气候条件下的产物,此类植被的主要分布区在我国(
1
1983
... 常绿阔叶林是分布在我国亚热带气候条件下的一种顶极森林植被, 它是全球亚热带大陆东岸湿润气候和季风气候条件下的产物,此类植被的主要分布区在我国(
Tree community patterns in seasonally dry tropical forests in the Cerros de Amotape Cordilera, Tumbes, Peru.
1
2005
... 科的重要值(FIV)计算公式(
Biomass and nutrient accumulation in montane evergreen broad-leaved forest (Lithocarpus xylocarpus type) in Ailao Mountains, SW China.
1
2002
... 哀牢山中山湿性常绿阔叶林动态样地(以下简称哀牢山样地)位于哀牢山国家级自然保护区内, 该保护区主要保护亚热带中山湿性常绿阔叶林生态系统和黑冠长臂猿、绿孔雀、灰叶猴等珍贵野生动物.有资料显示, 哀牢山地区有野生种子植物199科956属2 242种206变种(亚种)(
1
1988
... 物种多样性指数采用Shannon-Wiener指数(H)和Pielou均匀度指数(E)(
1
1983
... 常绿阔叶林是分布在我国亚热带气候条件下的一种顶极森林植被, 它是全球亚热带大陆东岸湿润气候和季风气候条件下的产物,此类植被的主要分布区在我国(
1
1983
... 常绿阔叶林是分布在我国亚热带气候条件下的一种顶极森林植被, 它是全球亚热带大陆东岸湿润气候和季风气候条件下的产物,此类植被的主要分布区在我国(
2
1998
... 常绿阔叶林是分布在我国亚热带气候条件下的一种顶极森林植被, 它是全球亚热带大陆东岸湿润气候和季风气候条件下的产物,此类植被的主要分布区在我国(
... 哀牢山中山湿性常绿阔叶林乔木树种主要由壳斗科、茶科、樟科和木兰科的树种组成, 其中, 壳斗科的硬斗柯、木果柯、变色锥(Castanopsis wattii), 茶科的南洋木荷(Schima noronhae)、舟柄茶(Hartia sinensis)、蒙自连蕊茶(Camellia forrestii), 樟科的黄心树(Machilus bombycina)、黄丹木姜子(Litsea elongata), 木兰科的红花木莲(Manglietia insignis)、多花含笑(Michelia floribunda)是乔木层的主要优势种.此外, 林中藤本植物和附生植物也相当发达, 层间植物以箭竹(Sinarundinaria nitida)(盖度达75%左右)及木质大藤本最为常见, 附生植物以苔藓和蕨类植物为主, 形成奇特的森林景观(
2
1998
... 常绿阔叶林是分布在我国亚热带气候条件下的一种顶极森林植被, 它是全球亚热带大陆东岸湿润气候和季风气候条件下的产物,此类植被的主要分布区在我国(
... 哀牢山中山湿性常绿阔叶林乔木树种主要由壳斗科、茶科、樟科和木兰科的树种组成, 其中, 壳斗科的硬斗柯、木果柯、变色锥(Castanopsis wattii), 茶科的南洋木荷(Schima noronhae)、舟柄茶(Hartia sinensis)、蒙自连蕊茶(Camellia forrestii), 樟科的黄心树(Machilus bombycina)、黄丹木姜子(Litsea elongata), 木兰科的红花木莲(Manglietia insignis)、多花含笑(Michelia floribunda)是乔木层的主要优势种.此外, 林中藤本植物和附生植物也相当发达, 层间植物以箭竹(Sinarundinaria nitida)(盖度达75%左右)及木质大藤本最为常见, 附生植物以苔藓和蕨类植物为主, 形成奇特的森林景观(
Influence of interactions between litter decomposition and rhizosphere activity on soil respiration and on the temperature sensitivity in a subtropical montane forest in SW China.
1
2014
... 哀牢山中山湿性常绿阔叶林动态样地(以下简称哀牢山样地)位于哀牢山国家级自然保护区内, 该保护区主要保护亚热带中山湿性常绿阔叶林生态系统和黑冠长臂猿、绿孔雀、灰叶猴等珍贵野生动物.有资料显示, 哀牢山地区有野生种子植物199科956属2 242种206变种(亚种)(
3
1980
... 常绿阔叶林是分布在我国亚热带气候条件下的一种顶极森林植被, 它是全球亚热带大陆东岸湿润气候和季风气候条件下的产物,此类植被的主要分布区在我国(
... ).根据气候特征的不同, 我国的常绿阔叶林分布区分为两个亚区域, 即东部常绿阔叶林亚区域和西部常绿阔叶林亚区域(
... 哀牢山样地与天童山样地、古田山样地、鼎湖山样地同属于亚热带常绿阔叶林植被型, 但哀牢山样地属于西部常绿阔叶林区域的山地类型, 天童山样地和古田山样地属于东部常绿阔叶林区域中亚热带常绿阔叶林地带, 而鼎湖山样地属于南亚热带常绿阔叶林地带(
3
1980
... 常绿阔叶林是分布在我国亚热带气候条件下的一种顶极森林植被, 它是全球亚热带大陆东岸湿润气候和季风气候条件下的产物,此类植被的主要分布区在我国(
... ).根据气候特征的不同, 我国的常绿阔叶林分布区分为两个亚区域, 即东部常绿阔叶林亚区域和西部常绿阔叶林亚区域(
... 哀牢山样地与天童山样地、古田山样地、鼎湖山样地同属于亚热带常绿阔叶林植被型, 但哀牢山样地属于西部常绿阔叶林区域的山地类型, 天童山样地和古田山样地属于东部常绿阔叶林区域中亚热带常绿阔叶林地带, 而鼎湖山样地属于南亚热带常绿阔叶林地带(
1
2006
... 根据
1
2006
... 根据
云南哀牢山山地湿性常绿阔叶林附生植物的多样性和分布
1
2005
... 常绿阔叶林是分布在我国亚热带气候条件下的一种顶极森林植被, 它是全球亚热带大陆东岸湿润气候和季风气候条件下的产物,此类植被的主要分布区在我国(
云南哀牢山山地湿性常绿阔叶林附生植物的多样性和分布
1
2005
... 常绿阔叶林是分布在我国亚热带气候条件下的一种顶极森林植被, 它是全球亚热带大陆东岸湿润气候和季风气候条件下的产物,此类植被的主要分布区在我国(
浙江天童20 ha常绿阔叶林动态监测样地的群落特征
2011
浙江天童20 ha常绿阔叶林动态监测样地的群落特征
2011
鼎湖山南亚热带常绿阔叶林20公顷样地群落特征研究
2
2008
... 物种重要值(IV)的计算公式(
... 径级结构是植物群落稳定性和生长发育状况的重要指标(
鼎湖山南亚热带常绿阔叶林20公顷样地群落特征研究
2
2008
... 物种重要值(IV)的计算公式(
... 径级结构是植物群落稳定性和生长发育状况的重要指标(
2
1983
... 常绿阔叶林是分布在我国亚热带气候条件下的一种顶极森林植被, 它是全球亚热带大陆东岸湿润气候和季风气候条件下的产物,此类植被的主要分布区在我国(
... ;
2
1983
... 常绿阔叶林是分布在我国亚热带气候条件下的一种顶极森林植被, 它是全球亚热带大陆东岸湿润气候和季风气候条件下的产物,此类植被的主要分布区在我国(
... ;
A study of the structure and composition of an old growth and secondary broad-leaved forest in the Ailao Mountains of Yunnan, China.
1
1992
... 常绿阔叶林是分布在我国亚热带气候条件下的一种顶极森林植被, 它是全球亚热带大陆东岸湿润气候和季风气候条件下的产物,此类植被的主要分布区在我国(
Floristic diversity and co-occurrences in a subtropical broad-leaved forest and two contrasting regrowth stands in central-west Yunnan Province, China.
1
1995
... 常绿阔叶林是分布在我国亚热带气候条件下的一种顶极森林植被, 它是全球亚热带大陆东岸湿润气候和季风气候条件下的产物,此类植被的主要分布区在我国(
Comparison of secondary and primary forest in the Ailao Shan region of Yunnan, China.
1
1989
... 常绿阔叶林是分布在我国亚热带气候条件下的一种顶极森林植被, 它是全球亚热带大陆东岸湿润气候和季风气候条件下的产物,此类植被的主要分布区在我国(
1
2009
... 哀牢山中山湿性常绿阔叶林动态样地(以下简称哀牢山样地)位于哀牢山国家级自然保护区内, 该保护区主要保护亚热带中山湿性常绿阔叶林生态系统和黑冠长臂猿、绿孔雀、灰叶猴等珍贵野生动物.有资料显示, 哀牢山地区有野生种子植物199科956属2 242种206变种(亚种)(
1
2009
... 哀牢山中山湿性常绿阔叶林动态样地(以下简称哀牢山样地)位于哀牢山国家级自然保护区内, 该保护区主要保护亚热带中山湿性常绿阔叶林生态系统和黑冠长臂猿、绿孔雀、灰叶猴等珍贵野生动物.有资料显示, 哀牢山地区有野生种子植物199科956属2 242种206变种(亚种)(
古田山中亚热带常绿阔叶林动态监测样地——群落组成与结构
2008
古田山中亚热带常绿阔叶林动态监测样地——群落组成与结构
2008

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}