 ,1*, 徐姗1, 熊鑫1,2, 夏艳菊1,2, 刘世忠1, 孟泽1, 褚国伟11
,1*, 徐姗1, 熊鑫1,2, 夏艳菊1,2, 刘世忠1, 孟泽1, 褚国伟11 2
Long-term (1992-2015) dynamics of community composition and structure in a monsoon evergreen broad-leaved forest in Dinghushan Biosphere Reserve
ZOU Shun1,2, ZHOU Guo-Yi1, ZHANG Qian-Mei,1*, XU Shan1, XIONG Xin1,2, XIA Yan-Ju1,2, LIU Shi-Zhong1, MENG Ze1, CHU Guo-Wei11 2
通讯作者: (zqm@scib.ac.cn)
编委: 李意德
责任编辑: 王葳
| 基金资助: |
Online:2018-04-20
| Fund supported: |

摘要
季风常绿阔叶林是中国南亚热带地区的地带性植被, 其群落结构及其动态变化直接影响着该森林能为该地区提供的生态系统服务功能质量。该文利用1992-2015年共24年的长期定点监测数据, 从群落种类组成、生物量、径级、密度等数量特征方面研究了鼎湖山季风常绿阔叶林群落结构的动态变化。结果表明: 1)到2015年该森林群落林分个体密度增加了42.7%, 总生物量减少了5.1%; 24年间基于生物量的β多样性指数群落差异为37.4%, 基于个体数的差异则高达65.6%; 2)灌木和小乔木的个体数剧增, 生物量增大, 中乔木和大乔木的个体数变化虽不显著, 但生物量显著降低; 3)小径级(胸径<15 cm)个体数呈显著增加趋势, 其他径级个体数也有显著变化, 但各径级生物量变化基本不显著; 4)香楠(Aidia canthioides)、鼎湖血桐(Macaranga sampsonii)、柏拉木(Blastus cochinchinensis)等物种个体数急剧增加, 锥(Castanopsis chinensis)、木荷(Schima superba)、黄果厚壳桂(Cryptocarya concinna)等树种生物量大量减少, 白颜树(Gironniera subaequalis)、窄叶半枫荷(Pterospermum lanceifolium)等树种生物量增加, 这些物种是群落结构变化的主要贡献者。鼎湖山季风常绿阔叶林群落结构在1992-2015年发生了巨大变化, 与演替、虫害和气候变化等影响有关。
关键词:
Abstract
Keywords:
PDF (1283KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
邹顺, 周国逸, 张倩媚, 徐姗, 熊鑫, 夏艳菊, 刘世忠, 孟泽, 褚国伟. 1992-2015年鼎湖山季风常绿阔叶林群落结构动态. 植物生态学报[J], 2018, 42(4): 442-452 DOI:10.17521/cjpe.2017.0171
ZOU Shun, ZHOU Guo-Yi, ZHANG Qian-Mei, XU Shan, XIONG Xin, XIA Yan-Ju, LIU Shi-Zhong, MENG Ze, CHU Guo-Wei.
森林具有调节气候、涵养水源、碳固存、生物多样性保育等不可替代的生态系统功能和服务功能(肖强等, 2014; Trumbore et al., 2015), 尤其在全球CO2浓度持续上升、地表平均温度增加、降水格局改变等环境变化背景下, 森林的巨大碳汇功能及其在减缓气候变化中的作用显得愈加重要(Pan et al., 2011)。森林生态系统的这些功能都受森林群落结构的强烈影响, 例如群落径级结构及其多样性与群落生产力呈正相关关系(Zhang & Chen, 2015), 并影响森林生态系统碳固存和物质周转速率(Xiao et al., 2014); 森林群落结构影响森林遭受干扰时的抵抗能力和干扰过后的恢复能力(Jayakaran et al., 2014); 群落结构变化后还常常引起森林产水量和集水区溪水来源组成的改变(Jayakaran et al., 2014; Wehner & Stednick, 2017)。
森林群落结构是指树木个体在环境中的分布及其与周围环境之间相互作用所形成的组分和构造, 包括个体密度、生物量大小、物种-多度分布、径级结构等(宋永昌, 2001)。演替、自然干扰、气候条件变化等都能改变群落结构。演替和自然干扰一直是森林生态学的重要内容, 演替理论认为自然条件下的森林都有向其顶极状态演替的趋势, 并伴随着群落结构的变化, 例如个体密度增大、生物量增加、物种组成改变及物种多样性提高等(Guariguata & Ostertag, 2001); 而极端气候事件、虫害、火灾等自然干扰能够短时间改变群落结构或引起群落结构的根本性变化(Anderegg et al., 2015; Frank et al., 2015); 而近10年来, 全球环境变化, 尤其是气候条件变化对森林群落结构影响的问题成为研究热点, Allen等(2015)认为气候变化引起了包括北方森林、温带森林和热带森林的全球不同类型森林群落结构的变化, 并且气候变化对森林内不同物种和不同径级个体的影响大小不同(Bennett et al., 2015; Anderegg et al., 2016)。由于森林群落结构及其影响因素的复杂性, 群落结构的长期定点监测仍是我们了解森林群落结构变化的主要方法(Allen et al., 2010)。
季风常绿阔叶林是中国南亚热带地区的地带性植被, 该类型森林面积广阔, 物种多样性高(宋永昌等, 2001; 丁圣彦和宋永昌, 2004)。同时, 该地区也是人口稠密地区, 季风常绿阔叶林的功能直接关系该地区的生态安全。在20世纪, 彭少麟等人对该地区季风常绿阔叶林群落的群落结构及其演替趋势做了较多研究(王伯荪和彭少麟, 1985; 彭少麟和方炜, 1995; 彭少麟等, 1998; 彭少麟和李丹, 1999), 但近年来对该森林群落结构动态的长期监测研究较少(张咏梅等, 2003; 叶万辉等, 2008)。有研究表明, 过去几十年该地区升高的年平均温度和愈加极端的降雨格局已经引起土壤湿度的持续下降(Zhou et al., 2011); 该地区同时还是全球三大高氮沉降区域之一和受酸雨严重影响的区域(Fang et al., 2011)。这些因素都可能引起该区域森林群落结构的变化。因此, 本文利用1992-2015年鼎湖山季风常绿阔叶林1 hm2永久样地7次群落调查数据, 分析不同调查时间点群落的个体数和生物量总量及组成, 并结合多样性指数方法, 探究我国南亚热带无人为干扰的近演替顶极森林群落结构随时间的演变, 为在全球变化背景下该地区自然林的评估和管理提供科学依据。
1 材料和方法
1.1 研究区样地概况
鼎湖山国家级自然保护区(23.16°-23.19° N, 112.51°-112.56° E)位于广东省肇庆市鼎湖区, 总占地面积1 155 hm2, 主要地形为丘陵和低山, 海拔100-700 m, 最高峰鸡笼山海拔1 000.3 m (熊鑫等, 2016)。该地区属于南亚热带季风气候, 年降水量为1 714 mm, 年平均湿度为76%, 4-9月为湿季, 10月到次年3月为旱季, 湿季降水量约占80%, 干湿季分明。 年平均气温为22.5 ℃, 最冷月(1月)和最热月(7月)平均气温分别为13.8 ℃和28.8 ℃; 地质基础主要为泥盆纪砂岩、砂页岩、页岩和石英砂岩, 地带性土壤为南亚热带赤红壤(Zhou et al., 2013), 自然酸化严重, 土壤的水浸pH值约为3.7。鼎湖山自然保护区的季风常绿阔叶林(以下简称季风林, MEBF)主要分布于核心区三宝峰, 海拔100-490 m, 具有400多年的保护历史, 属于近演替顶极阶段(熊鑫等, 2016)。该群落终年常绿, 郁闭度约95%。乔木层郁闭度约80%, 可分为两个亚层: 第一亚层为锥(Castanopsis chinensis)、木荷(Schima superba)、黄果厚壳桂(Cryptocarya concinna)、厚壳桂(Cryptocarya chinensis)、白颜树(Gironniera subaequalis)等; 第二亚层为鼎湖血桐(Macaranga sampsonii)、香楠(Aidia canthioides)等。灌木层盖度约50%, 优势种为柏拉木(Blastus cochinchinensis)。草本层盖度约40%, 优势种为华山姜(Alpinia oblongifolia)等。样地位于低山中坡, 坡度25°-35°, 土层厚度为30-90 cm (黄忠良等, 1998)。
1.2 研究方法
1.2.1 样地设置与调查鼎湖山季风林永久样地投影面积为1 hm2。样地采用全站仪将10 000 m2划分为25个20 m × 20 m的样方, 每个20 m × 20 m样方用插值法细分为4个 10 m × 10 m的样方, 每个10 m × 10 m样方细划为4个5 m × 5 m的小样方。在1992-2015年间, 共进行了7次群落调查, 记录样地内所有胸径(DBH)大于等于1.0 cm个体的物种名、胸径、高度、坐标及生长状况等信息(张咏梅等, 2003)。
1.2.2 生物量估算
每个个体生物量由生物量方程(温达志等, 1997) 估算, 并进而计算各物种、生活型和径级的生物量。
1.2.3 生活型分类
基于生活型分类系统把群落物种分为两类: 生活型I为中乔木和大乔木, 在季风林群落内成熟个体高度大于等于8 m, 占据冠层位置, 生活型II为小乔木和灌木, 在季风林群落内成熟个体高度小于8 m (宋永昌, 2001)。
1.2.4 多样性指数计算
α多样性指群落的物种多样性, 是群落内物种丰富度和物种多度格局的综合体现, 常用于度量群落物种多样性的大小。α多样性指数非常多, 但各指数具有较强的相关性, 本文采用常用的Shannon- Wiener指数计算群落的α多样性(陈廷贵和张金屯, 1999)。
长时间的群落结构和多样性的变化可以区分为群落总量(总个体数、总生物量)的变化和群落内物种个体数或生物量分配的变化。为了描述群落结构的变化, 以及区分上述两种原因的贡献, 我们分别计算了以下4个相互关联的指数: 分别描述群落间相似性、差异性的相似指数(SI)和β多样性指数(BI), 描述群落内物种个体数或生物量分配变化的替换指数(RI), 以及描述群落总量变化的总量差异指数(DI) (Fekete et al., 2017)。
$\begin{align}
& R{{I}_{1992j}}={\left( \sum\limits_{i\ =\ 1}^{n}{|{{x}_{1992i}}-{{x}_{ji}}|}-\left| \sum\limits_{i\ =\ 1}^{n}{{{x}_{1992i}}}-\sum\limits_{i\ =\ 1}^{n}{{{x}_{ji}}} \right| \right)}/{{}}\; \\
& \ \ \ \ \ \ \ \ \ \ \ \ \ \ \sum\limits_{i\ =\ 1}^{n}{\max \text{ }\!\!\{\!\!\text{ }{{x}_{1992i}},{{x}_{ji}}\text{ }\!\!\}\!\!\text{ }} \\
\end{align}$ (2)
其中, xji表示第j年(j = 1994, 1999, 2004, 2008, 2010和2015)第i个物种的个体数或生物量。SI1992j、RI1992j和DI1992j分别表示第j年时群落与1992年群落之间的相似指数、替换指数和总量差异指数。而β多样性指数与相似指数, 以及与替换指数和总量差异指数之间有如下关系, 其中BI1992j表示第j年时群落与1992年群落之间的β多样性指数。
$\begin{align}
& B{{I}_{1992j}}=R{{I}_{1992j}}+D{{I}_{1992j}} \\
& \ \ \ \ \ \ \ \ \ \ \ {=\sum\limits_{i\ =\ 1}^{n}{|{{x}_{1992i}}-{{x}_{ji}}|}}/{\sum\limits_{i\ =\ 1}^{n}{\max \{{{x}_{1992i}},{{x}_{ji}}\}}}\; \\
\end{align}$ (4)
1.2.5 重要值与群落主要物种
采用重要值((相对多度+相对显著度+相对频度)/3)表征物种在群落中的优势程度(叶万辉等, 2008)。分别计算各次调查各物种的重要值, 如果某物种7次群落调查中出现过至少1次物种重要值≥1.0, 则我们认为它是群落中的主要物种。鼎湖山季风林有29种主要物种(表1), 其重要值、个体数和生物量之和在群落中的比例分别达88%、87%和96%。
Table 1
表1
表1鼎湖山季风林样地群落主要物种(IV ≥ 1.0)
Table 1
| 序号 Order | 物种 Species | 生活型 Life form | 物种代码 Species code | 序号 Order | 物种 Species | 生活型 Life form | 物种代码 Species code |
|---|---|---|---|---|---|---|---|
| 1 | 锥 Castanopsis chinensis | I | Cch | 16 | 笔罗子 Meliosma rigida | I | Mr |
| 2 | 白颜树 Gironniera subaequalis | I | Gs | 17 | 广东金叶子 Craibiodendron scleranthum | I | Cs |
| 3 | 肖蒲桃 Syzygium acuminatissimum | I | Sa | 18 | 香楠 Aidia canthioides | II | Ac |
| 4 | 木荷 Schima superba | I | Ss | 19 | 鼎湖血桐 Macaranga sampsonii | II | Ms |
| 5 | 窄叶半枫荷 Pterospermum lanceifolium | I | Pl | 20 | 柏拉木 Blastus cochinchinensis | II | Bc |
| 6 | 厚壳桂 Cryptocarya chinensis | I | Crc | 21 | 云南银柴 Aporosa yunnanensis | II | Ay |
| 7 | 橄榄 Canarium album | I | Ca | 22 | 褐叶柄果木 Mischocarpus pentapetalus | II | Mip |
| 8 | 白楸 Mallotus paniculatus | I | Map | 23 | 红枝蒲桃 Syzygium rehderianum | II | Sr |
| 9 | 黄果厚壳桂 Cryptocarya concinna | I | Cco | 24 | 光叶红豆 Ormosia glaberrima | II | Og |
| 10 | 臀果木 Pygeum topengii | I | Pt | 25 | 黄叶树 Xanthophyllum hainanense | II | Xh |
| 11 | 肉实树 Sarcosperma laurinum | I | Sl | 26 | 罗伞树 Ardisia quinquegona | II | Aq |
| 12 | 观光木 Michelia odora | I | Mo | 27 | 黄毛榕 Ficus esquiroliana | II | Fe |
| 13 | 华润楠 Machilus chinensis | I | Mc | 28 | 鼎湖钓樟 Lindera chunii | II | Lc |
| 14 | 鹅掌柴 Schefflera heptaphylla | I | Sh | 29 | 九节 Psychotria asiatica | II | Pa |
| 15 | 黄杞 Engelhardia roxburghiana | I | Er |
新窗口打开|下载CSV
2 结果
2.1 群落个体数和生物量总体变化趋势
1992-2015年样地个体数先略微下降, 在2004年以后急剧上升, 个体数总体呈显著上升趋势(R2 = 0.63, p = 0.02), 2015年样地总个体数为5 682个, 是1992年的1.43倍, 净增个体数为1 701个(图1A)。群落物种-多度格局改变, 优势种个体数急剧增加(图2A), 其在群落中所占比例增大(图2B), 基于个体数的群落Shannon-Wiener指数先上升后下降(表2)。随着时间变化, 各调查年群落与1992年群落间的β多样性指数BI (基于个体数)持续增大(Logistic拟合, R2 = 0.99, p < 0.001), 其中替换指数RI所占比例都大于总量差异指数DI (1994年除外)(图3A)。图1
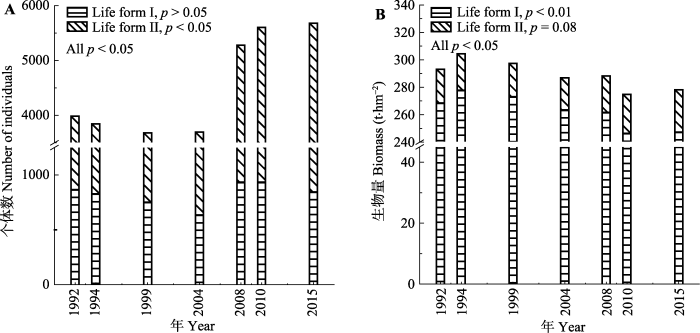 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图11992-2015年鼎湖山季风林1 hm2样地整个群落(All)、大乔木和中乔木(Life form I)、小乔木和灌木(Life form II)个体数(A)和生物量(B)变化(线性拟合)。
Fig. 1Changes in stand density (A) and biomass (B) of community (All), middle and big arbor (Life form I) and small arbor and shrub (Life form II) in the 1 hm2 monsoon evergreen broad-leaved forest sample plot between 1992 and 2015 (linear fitting).
Table 2
表2
表2鼎湖山季风林群落(胸径≥ 1.0 cm)物种数以及分别基于多度和生物量的Shannon-Wiener指数变化
Table 2
| 年 Year | 物种数 Species richness | 基于个体数 Individual based | 基于生物量 Biomass based | ||||
|---|---|---|---|---|---|---|---|
| 群落 Population | 生活型I Life form I | 生活型II Life form II | 群落 Population | 生活型I Life form I | 生活型II Life form II | ||
| 1992 | 86 | 2.90 | 2.39 | 2.30 | 2.16 | 1.86 | 2.17 |
| 1994 | 86 | 2.90 | 2.40 | 2.29 | 2.19 | 1.88 | 2.20 |
| 1999 | 90 | 2.99 | 2.52 | 2.39 | 2.27 | 1.97 | 2.21 |
| 2004 | 94 | 3.07 | 2.63 | 2.53 | 2.16 | 1.82 | 2.29 |
| 2008 | 90 | 2.98 | 2.62 | 2.42 | 2.22 | 1.88 | 2.50 |
| 2010 | 90 | 2.91 | 2.64 | 2.34 | 2.30 | 1.93 | 2.49 |
| 2015 | 80 | 2.71 | 2.62 | 2.15 | 2.38 | 2.01 | 2.41 |
| p | >0.05 | >0.05 | <0.05 | >0.05 | =0.05 | >0.05 | <0.05 |
新窗口打开|下载CSV
1992-2015年, 样地生物量呈显著下降趋势(R2 = 0.64, p < 0.01), 2015年样地总生物量为278.1 t, 比1992年减少了15.0 t, 下降了5.1% (图1B)。群落物种-生物量格局改变, 优势种生物量降低(图2C), 其在群落中所占比例减少(图2D), 导致了基于生物量的群落Shannon-Wiener指数上升(p = 0.05)(表2)。随着时间变化, 各调查年群落与1992年群落间的β多样性指数BI (基于生物量)持续增大(线性拟合, R2 = 0.99, p < 0.001), 其中总量差异指数DI所占比例都大于替换指数RI (1994年除外)(图3B)。
图2
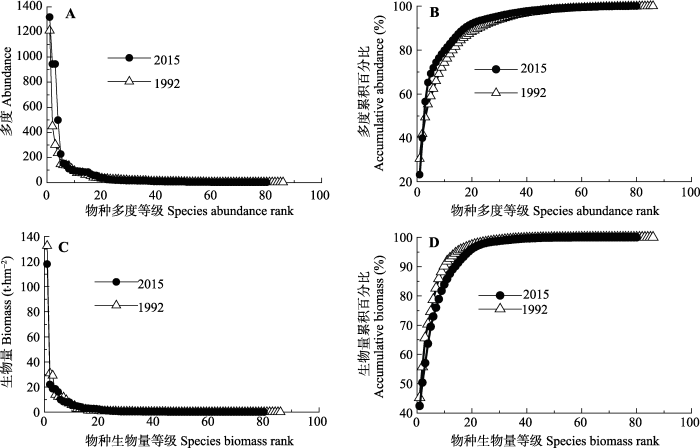 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2鼎湖山季风林1 hm2样地物种-个体数和物种-生物量格局变化。A, 物种-多度分布。B, 物种-多度累积。C, 物种-生物量分布。D, 物种-生物量累积。
Fig. 2Changes in species-abundance and species-biomass relationships in the 1 hm2 monsoon evergreen broad-leaved forest sample plot. A, species-abundance distribution. B, accumulative species-abundance. C, species-biomass distribution. D, species-biomass accumulation.
图3
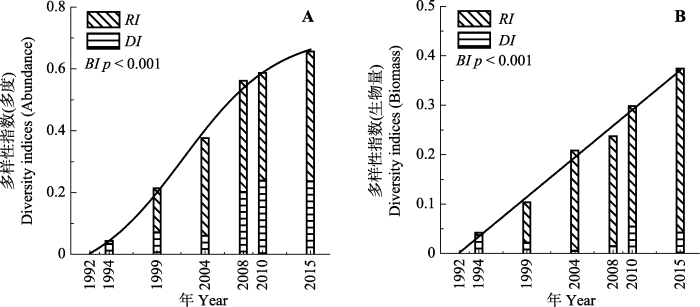 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图31992-2015年群落替换指数(RI)、总量差异指数(DI)和β多样性指数(BI)变化。A, 基于多度(Logistic拟合)。B, 基于生物量(线性拟合)。
Fig. 3Changes in the replacement index (RI), difference index (DI) and β diversity index (BI) with year from 1992 to 2015. A, Abundance-based logistic fitting. B, Biomass-based linear fitting.
2.2 群落生活型组成变化
由图1A所示, 中乔木和大乔木的个体数无显著变化(p > 0.05); 小乔木和灌木的个体数总体呈显著增加趋势(R2 = 0.72, p < 0.01), 2015年群落小乔木和灌木个体数4 836个, 与1992年相比增加1 723个, 占群落个体数净增量的101.3%, 导致小乔木和灌木个体数在群落所占比例增大, 由1992年的74.4%上升到2015年的82.3%。而基于个体数的中乔木和大乔木Shannon-Wiener指数显著上升(p < 0.05), 小乔木和灌木先上升后下降(表2)。
由图1B可见, 中乔木和大乔木总生物量呈显著下降趋势(R2 = 0.73, p < 0.01), 与1992年相比, 2015年群落大乔木和中乔木生物量减少了21.1 t; 小乔木和灌木生物量(R2 = 0.37, p = 0.08)有增加趋势, 但变化在α = 0.05的置信水平下不显著, 与1992年相比, 其生物量增加了6.1 t。基于生物量的中乔木和大乔木Shannon-Wiener指数增大, 但趋势不显著(p > 0.05), 而小乔木和灌木Shannon-Wiener指数呈显著增加趋势(p < 0.05)(表2)。
2.3 群落径级结构变化
由图4可见, 1992-2015年群落DBH < 15 cm (R2 = 0.60, p < 0.05)和DBH 35-50 cm (R2 = 0.59, p < 0.05)径级个体数呈增加趋势, DBH在15-35 cm (R2 = 0.43, p = 0.06)和DBH ≥ 50 cm (R2 = 0.88, p < 0.05)径级个体数呈减小趋势; 群落DBH在15-35 cm径级生物量显著下降(R2 = 0.57, p < 0.05), 而其他径级生物量没有显著变化趋势。图4
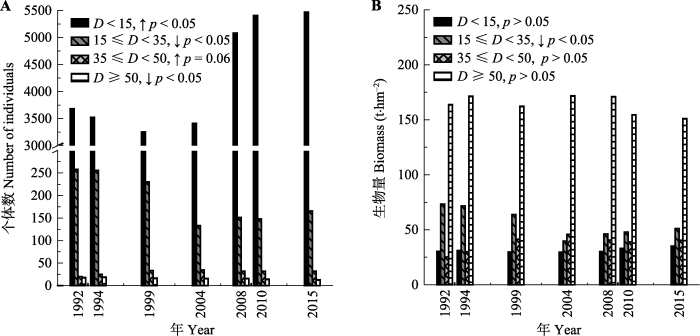 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图41992-2015年鼎湖山季风林1 hm2样地内群落个体数(A)和生物量(B)的径级结构变化(线性拟合)。D, 胸径; ↑, 显著(p < 0.05)上升; ↓, 显著(p < 0.05)下降。
Fig. 4Dynamics of size class(diameters at breast height) distributions of individuals (A) and biomass (B) from 1992 to 2015 (linear fitting) in the 1 hm2 monsoon evergreen broad-leaved forest sample plot. D, diameters at breast height; ↑, significant (p < 0.05) increase; ↓, significant (p < 0.05) decrease.
2.4 主要物种个体数和生物量变化
与1992年比较, 到2015年, 群落中29个主要物种中的17个中乔木和大乔木树种中只有3个个体数增量(增加量或减少量)绝对值超过1992年群落总个体数1.0%, 包括厚壳桂、白楸(Mallotus paniculatus)和黄果厚壳桂, 其中, 厚壳桂和白楸个体数分别增加了51和80个, 种群增大1.6倍和16倍, 黄果厚壳桂个体数减少213个, 种群减小70.8%; 这样的小乔木和灌木树种有7个(共12个), 包括香楠、鼎湖血桐、柏拉木、云南银柴(Aporosa yunnanensis)、褐叶柄果木(Mischocarpus pentapetalus)、黄毛榕(Ficus esquiroliana)和九节(Psychotria asiatica), 其中, 香楠、鼎湖血桐、柏拉木、褐叶柄果木和黄毛榕个体数分别增加了1181、890、497、164和60个, 种群依次增大了8.6倍、16.2倍、1.1倍、2.6倍和7.5倍, 云南银柴和九节个体数分别减少了715个和228个, 种群减小59.0%和97.0% (图5A)。
图5
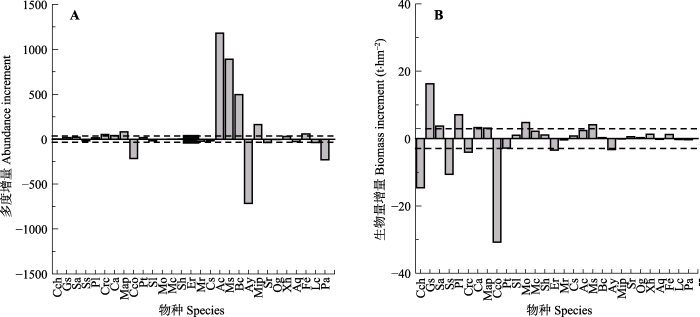 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图51992和2015年鼎湖山季风林1 hm2样地内29个主要物种的多度(A)和生物量(B)变化。A图和B图中的虚线分别为1992年总个体数的1.0% (36.4个)和1992年总生物量的1.0% (2.93 t)。图中各物种代码与物种的对应关系见
Fig. 5The changes in the abundance (A) and biomass (B) of 29 common species in the monsoon evergreen broad-leaved forest (MEBF) sample plot during 1992-2015 in the 1 hm2 MEBF sample plot. Dashed lines in plot A and B represent 1.0% of the total individuals (36.4) and biomass (2.93 t) in 1992, respectively. The one-to-one relationship between codes and species is provided in
与1992年比较, 2015年群落中29个主要物种中有11个中乔木和大乔木树种生物量增量(增加量或减少量)绝对值超过1992年群落总生物量1.0%, 包括锥、白颜树、肖蒲桃、木荷、窄叶半枫荷(Pterospermum lanceifolium)、厚壳桂(Cryptocarya chinensis)、橄榄(Canarium album)、白楸、黄果厚壳桂、观光木(Michelia odora)、黄杞(Engelhardia roxburghiana), 其中, 白颜树、肖蒲桃、窄叶半枫荷、橄榄、白楸和观光木生物量分别增加了16.3 t、3.7 t、7.1 t、3.3 t、3.1 t和4.7 t, 种群生物量增大了283.8%、29.6%、262.0%、67.2%、3581.7%和34.2%, 锥、木荷、厚壳桂、黄果厚壳桂和黄杞生物量分别减少了14.6 t、10.6 t、4.0 t、30.8 t和3.4 t, 种群生物量减小了11.0%、36.4%、33.5%、99.6%和36.6%, 这样的小乔木和灌木树种有2个, 是鼎湖血桐和云南银柴, 鼎湖血桐生物量增加4.1 t, 增大333.7%, 云南银柴生物量减少3.2 t, 减小了28.0% (图5B)。
图6
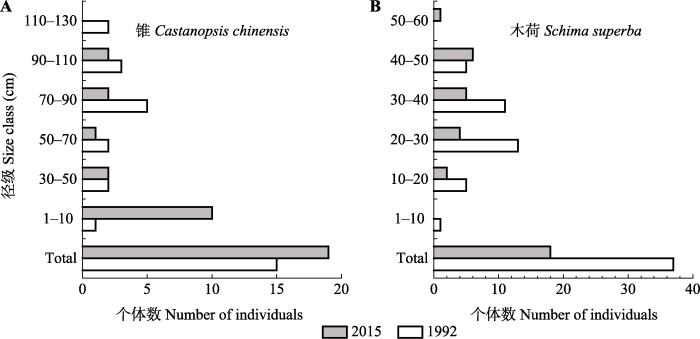 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图61992和2015年鼎湖山季风林1 hm2样地锥(A)和木荷(B)的种群。Total, 总个体数。
Fig. 6Population sizes of Castanopsis chinensis (A) and Schima superba (B) between 1992 and 2015 in the 1 hm2 monsoon evergreen broad-leaved forest sample plot. Total, total number of individuals.
如上所述, 1992-2015年锥和木荷生物量下降(图5B); 锥各大径级个体数下降, 但有更新个体(DBH < 10 cm), 种群总个体数略有增加, 木荷DBH在10-40 cm径级个体数下降, DBH ≥ 40 cm径级个体数略有增加, 但无更新个体, 种群总个体数下降51.4%; 在2015年锥和木荷生物量分别占群落总生物量的42.4%和6.6%。
3 讨论
研究结果表明, 1992-2015年鼎湖山季风林总个体数增加了42.7%, 其直接原因是香楠、鼎湖血桐、柏拉木等少数灌木和小乔木树种种群大小的急剧增加, 这已经使得群落小个体占据更大比例, 基于个体数的群落物种多样性降低, 2015年和1992年间基于个体数的群落差异(β多样性指数)达65.6%, 群落总个体数的变化和物种间多度比例的变化都有很大贡献, 但总的来说, 群落总个体数和β多样性指数在2008年以后的增长速率下降, 群落总个体数和物种-多度分布可能在未来变化较小。而1992-2015年群落总生物量下降了5.1%, 远小于个体数的变化, 虽然锥、木荷、黄果厚壳桂等中乔木和大乔木优势树种生物量大量减少, 但白颜树、窄叶半枫荷等树种生物量的增加具有部分补偿作用, 这引起了群落物种-生物量分布相对更加均匀, 基于生物量的群落物种多样性增大, 2015年和1992年间基于生物量的群落差异(β多样性指数)为37.4%, 其中与总生物量的变化相比, 物种间生物量比例变化更加剧烈, 贡献了群落差异的89.0%, 群落总生物量和β多样性指数在1992-2015年一直呈线性变化趋势, 未来群落生物量总量和组成可能继续变化。
鼎湖山季风林是演替近顶极群落, 该地区森林群落的演替顺序为马尾松林, 马尾松、锥和木荷等树种为主的针阔混交林, 季风林(王伯荪和彭少麟, 1985), 所以季风林中的阳生性树种锥和木荷主要是演替上一阶段针阔混交林时期的遗留(彭少麟和方炜, 1995; 彭少麟等, 1998), 锥和木荷种群小, 大个体逐渐死亡, 少或无更新个体, 但季风林中的锥和木荷多为大个体, 2015年两个树种生物量之和仍占群落总生物量的49.0%, 所以锥和木荷在鼎湖山季风林生态系统功能中仍发挥重要作用, 但其个体不断死亡将给群落造成巨大波动(彭闪江等, 2003; 张池等, 2006), 甚至群落总生物量持续下降。而在气候变化背景下, 群落内大径级个体更容易受到干旱胁迫(Bennett et al., 2015), 因此气候变化可能会加快鼎湖山季风林内锥和木荷种群的衰退, 影响群落的演替进程(王纪军和裴铁璠, 2004)。
随着向气候顶极群落演替, 中生性的乔木树种黄果厚壳桂和厚壳桂将成为季风林中的优势物种(彭少麟和李丹, 1999), 1992年黄果厚壳桂和厚壳桂生物量之和占群落总生物量的14.6%, 到2015年其生物量只占2.9%, 它们的优势度下降和虫害爆发有关。黄忠良(2000)在鼎湖山季风林8年的调查研究, 发现樟翠尺蛾(Thalassodes quadraia)种群爆发对群落内包括黄果厚壳桂和厚壳桂在内的樟科植物造成了极大破坏, 并且分析发现虫害爆发和降水量及降水格局变化有密切关系; 张池等(2006)对鼎湖山季风林1992-2004年树木个体死亡动态进行了研究, 认为樟翠尺蛾的持续爆发导致了该群落90%以上的黄果厚壳桂个体死亡。陈志成和万贤崇(2016)认为气候变化提高了虫害的爆发频度, 虫害对森林的危害加重; Clark等(2016)同样发现由干旱及其伴随的虫害改变了美国森林群落的结构和生物多样性。在华南地区降水变率增大的背景下(Zhou et al., 2011), 虫害发生与气候变化等的关系及其对季风林的危害程度还需要深入研究。
近10年来, 有关全球环境变化对森林群落结构影响的研究和讨论有很多, 变暖和降水减少(或降水变率加大)诱导的干旱对森林造成影响, 中国亚热带地区面临同样的环境问题, 研究认为气温升高和降水变率增大减少了土壤水分(Zhou et al., 2011; 刘效东等, 2014), 并诱导了季风林树木死亡率和更新率的上升(Zhou et al., 2013), 亚热带常绿阔叶林群落个体数及其组成变化(Zhou et al., 2014); Li等(2015)通过测定植物的功能, 发现季风林群落中具有与抗旱和快速生长相关功能性状的物种香楠、鼎湖血桐、柏拉木等多度增加, 同样认为气候变干已经引起该森林群落结构变化。该群落的研究结果与McIntyre等(2015)在美国整个加利福尼亚地区森林长达70年的研究结果一致, 他们也发现群落大径级个体下降, 小径级个体增加, 导致林分密度增加了30%, 基面积下降了19%, 而橡树逐渐取代松树的优势地位, 并认为群落结构改变与该地区气温上升和水分胁迫有关。但也有许多研究认为干旱会同时降低群落个体数和生物量(Allen et al., 2015), 而Zhang等(2015)研究了加拿大西部地区森林长达50年的群落结构动态变化, 认为气候变化并不是主要因素, 气候变化下群落内部的竞争关系变化才是引起群落结构变化的主要原因。
4 结论
通过以上分析, 我们得到以下主要结论: 1)从1992到2015年该森林群落林分密度增加了42.7%, 生物量减少了5.1%, 与1992年相比, 基于β多样性指数的群落生物量差异为37.4%, 主要原因是群落内物种生物量分配的变化, 而群落个体差异则高达65.6%, 群落总个体数变化和群落内物种个体数分配的改变都有很大贡献; 2)小乔木和灌木个体数剧增, 生物量增大, 中乔木和大乔木个体数变化不显著, 但生物量显著下降; 3)小径级(DBH < 15 cm)个体数呈显著增加趋势, 其他径级个体数也有显著变化, 但各径级生物量变化基本不显著; 4)香楠、鼎湖血桐、柏拉木等物种个体数急剧增加, 锥、木荷、黄果厚壳桂等树种生物量大量减少, 白颜树、窄叶半枫荷等树种生物量增加, 这些物种是群落结构变化的主要贡献者。鼎湖山季风林群落结构在1992-2015年发生了巨大变化, 可能与演替进程、虫害和气候变化等因素的影响有关。但我们对季风林的演替进程, 季风林如何响应环境变化, 演替进程与气候变化间的相互作用, 虫害爆发和气候的关系等问题的了解不多, 也缺乏季风林群落结构改变对其生态系统服务功能影响的研究, 未来需要对这些问题进行深入研究, 加强对我国季风林的监测和管理。
扫码加入读者圈
听语音, 看问答
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOIURL [本文引用: 1]

Abstract Here we draw attention to the potential for pelagic bloom-forming cyanobacteria to have substantial effects on nutrient cycling and ecosystem resilience across a wide range of lakes. Specifically, we hypothesize that cyanobacterial blooms can influence lake nutrient cycling, resilience, and regime shifts by tapping into pools of nitrogen (N) and phosphorus (P) not usually accessible to phytoplankton. The ability of many cyanobacterial taxa to fix dissolved N2 gas is a well-known potential source of N, but some taxa can also access pools of P in sediments and bottom waters. Both of these nutrients can be released to the water column via leakage or mortality, thereby increasing nutrient availability for other phytoplankton and microbes. Moreover, cyanobacterial blooms are not restricted to high nutrient (eutrophic) lakes: blooms also occur in lakes with low nutrient concentrations, suggesting that changes in nutrient cycling and ecosystem resilience mediated by cyanobacteria could affect lakes across a gradient of nutrient concentrations. We used a simple model of coupled N and P cycles to explore the effects of cyanobacteria on nutrient dynamics and resilience. Consistent with our hypothesis, parameters reflecting cyanobacterial modification of N and P cycling alter the number, location, and/or stability of model equilibria. In particular, the model demonstrates that blooms of cyanobacteria in low-nutrient conditions can facilitate a shift to the high-nutrient state by reducing the resilience of the low-nutrient state. This suggests that cyanobacterial blooms warrant attention as potential drivers of the transition from a low-nutrient, clear-water regime to a high-nutrient, turbid-water regime, a prediction of particular concern given that such blooms are reported to be increasing in many regions of the world due in part to global climate change.
DOIURL [本文引用: 1]
Les émissions de gaz à effet de serre ont significativement changé le climat mondial et continueront à le faire dans l'avenir. L'augmentation de la fréquence, la durée ou la sévérité des sécheresses et des canicules pourraient fondamentalement changer la composition, la structure et la biogéographie des forêts de nombreuses régions. Il en résulte un accroissement potentiel de la mortalité des arbres, associée aux stress physiologiques et aux interactions avec d'autres processus comme des explosions démographiques d'insectes et les incendies. Malgré ce risque, les prévisions actuelles de mortalité d'arbre sont basées sur des modèles qui manquent fonctionnellement de réalismes. Il n'y a eu à ce jour aucune tentative de suivi global de la mortalité forestière induite par le climat. Nous présentons dans cet article la première évaluation mondiale des mortalités récentes d'arbres attribuées aux stress thermiques et hydriques. / Greenhouse gas emissions have significantly altered global climate, and will continue to do so in the future. Increases in the frequency, duration, and/or severity of drought and heat stress associated with climate change could fundamentally alter the composition, structure, and biogeography of forests in many regions. Of particular concern are potential increases in tree mortality associated with climate-induced physiological stress and interactions with other climate-mediated processes such as insect outbreaks and wildfire. Despite this risk, existing projections of tree mortality are based on models that lack functionally realistic mortality mechanisms, and there has been no attempt to track observations of climate-driven tree mortality globally. Here we present the first global assessment of recent tree mortality attributed to drought and heat stress.
DOIURLPMID [本文引用: 1]
Summary Climate change is expected to drive increased tree mortality through drought, heat stress, and insect attacks, with manifold impacts on forest ecosystems. Yet, climate-induced tree mortality and biotic disturbance agents are largely absent from process-based ecosystem models. Using data sets from the western USA and associated studies, we present a framework for determining the relative contribution of drought stress, insect attack, and their interactions, which is critical for modeling mortality in future climates. We outline a simple approach that identifies the mechanisms associated with two guilds of insects – bark beetles and defoliators – which are responsible for substantial tree mortality. We then discuss cross-biome patterns of insect-driven tree mortality and draw upon available evidence contrasting the prevalence of insect outbreaks in temperate and tropical regions. We conclude with an overview of tools and promising avenues to address major challenges. Ultimately, a multitrophic approach that captures tree physiology, insect populations, and tree–insect interactions will better inform projections of forest ecosystem responses to climate change.
DOIURLPMID [本文引用: 1]
Abstract Drought-induced tree mortality has been observed globally and is expected to increase under climate change scenarios, with large potential consequences for the terrestrial carbon sink. Predicting mortality across species is crucial for assessing the effects of climate extremes on forest community biodiversity, composition, and carbon sequestration. However, the physiological traits associated with elevated risk of mortality in diverse ecosystems remain unknown, although these traits could greatly improve understanding and prediction of tree mortality in forests. We performed a meta-analysis on species' mortality rates across 475 species from 33 studies around the globe to assess which traits determine a species' mortality risk. We found that species-specific mortality anomalies from community mortality rate in a given drought were associated with plant hydraulic traits. Across all species, mortality was best predicted by a low hydraulic safety margin-the difference between typical minimum xylem water potential and that causing xylem dysfunction-and xylem vulnerability to embolism. Angiosperms and gymnosperms experienced roughly equal mortality risks. Our results provide broad support for the hypothesis that hydraulic traits capture key mechanisms determining tree death and highlight that physiological traits can improve vegetation model prediction of tree mortality during climate extremes.
DOIURLPMID [本文引用: 2]
Abstract The frequency of severe droughts is increasing in many regions around the world as a result of climate change(1-3). Droughts alter the structure and function of forests(4,5). Site- and region-specific studies suggest that large trees, which play keystone roles in forests(6) and can be disproportionately important to ecosystem carbon storage(7) and hydrology(8), exhibit greater sensitivity to drought than small trees(4,5,9,10). Here, we synthesize data on tree growth and mortality collected during 40 drought events in forests worldwide to see whether this size-dependent sensitivity to drought holds more widely. We find that droughts consistently had a more detrimental impact on the growth and mortality rates of larger trees. Moreover, drought-related mortality increased with tree size in 65% of the droughts examined, especially when community-wide mortality was high or when bark beetles were present. The more pronounced drought sensitivity of larger trees could be underpinned by greater inherent vulnerability to hydraulic stress(11-14), the higher radiation and evaporative demand experienced by exposed crowns(4,15), and the tendency for bark beetles to preferentially attack larger trees(16). We suggest that future droughts will have a more detrimental impact on the growth and mortality of larger trees, potentially exacerbating feedbacks to climate change.
URL [本文引用: 1]
用关帝山植被数据对15个物种多样性指数进行了比较研究,结果表明:各指数间有着密切的关系,它们都能较好地反映多样性特征,但其中较好的丰富度指数依次为:Patrick指数、Menhinick指数、Margalef指数。较好的均匀度指数依次为:修正的Hil指数、Alatalo指数、Pielou指数(e)、Pielou指数(Jsw)、Pielou指数(Jsi)、McIntosh指数。较好的多样性指数依次为:Shannon-Wiener指数、Simpson指数、Audair和Gof指数、种间相遇机率、McIntosh指数、Hurlbert指数。相关系数分析也很好地说明了这个问题。
URL [本文引用: 1]
用关帝山植被数据对15个物种多样性指数进行了比较研究,结果表明:各指数间有着密切的关系,它们都能较好地反映多样性特征,但其中较好的丰富度指数依次为:Patrick指数、Menhinick指数、Margalef指数。较好的均匀度指数依次为:修正的Hil指数、Alatalo指数、Pielou指数(e)、Pielou指数(Jsw)、Pielou指数(Jsi)、McIntosh指数。较好的多样性指数依次为:Shannon-Wiener指数、Simpson指数、Audair和Gof指数、种间相遇机率、McIntosh指数、Hurlbert指数。相关系数分析也很好地说明了这个问题。
DOIURL
大规模虫害爆发可造成区域森林死亡,近年的气候变化进一步增加了虫害的频度和危害程度。森林和林地植物死亡会导致植被生产力降低,改变生态系统结构和功能,使森林由一个净的碳汇转变为一个碳源。因此,加深虫害对树木危害机制的认识有重要意义。虫害造成的叶损失(虫害叶损失)降低树木光合作用能力,增加非结构性碳(NSC)消耗,使得树木体内碳储备降低,NSC降低到一定程度会导致树木因碳饥饿而死亡。外部环境和树木自身的补偿性机制也会对这个过程产生正或负的影响。在近年气候变化背景下,树木死亡在全球尺度上增多,重新激起了人们对碳饥饿的重视,碳饥饿被视为解释树木死亡的主要生理机制之一。该文介绍了碳饥饿的定义,综述了虫害叶损失减少树木NSC储备与树木生长、死亡的关系,以及树木虫害和叶损失与气候变化之间的关系,并对今后的研究进行了展望。
DOIURL
大规模虫害爆发可造成区域森林死亡,近年的气候变化进一步增加了虫害的频度和危害程度。森林和林地植物死亡会导致植被生产力降低,改变生态系统结构和功能,使森林由一个净的碳汇转变为一个碳源。因此,加深虫害对树木危害机制的认识有重要意义。虫害造成的叶损失(虫害叶损失)降低树木光合作用能力,增加非结构性碳(NSC)消耗,使得树木体内碳储备降低,NSC降低到一定程度会导致树木因碳饥饿而死亡。外部环境和树木自身的补偿性机制也会对这个过程产生正或负的影响。在近年气候变化背景下,树木死亡在全球尺度上增多,重新激起了人们对碳饥饿的重视,碳饥饿被视为解释树木死亡的主要生理机制之一。该文介绍了碳饥饿的定义,综述了虫害叶损失减少树木NSC储备与树木生长、死亡的关系,以及树木虫害和叶损失与气候变化之间的关系,并对今后的研究进行了展望。
DOIURLPMID
Abstract We synthesize insights from current understanding of drought impacts at stand-to-biogeographic scales, including management options, and we identify challenges to be addressed with new research. Large stand-level shifts underway in western forests already are showing the importance of interactions involving drought, insects, and fire. Diebacks, changes in composition and structure, and shifting range limits are widely observed. In the eastern US, the effects of increasing drought are becoming better understood at the level of individual trees, but this knowledge cannot yet be confidently translated to predictions of changing structure and diversity of forest stands. While eastern forests have not experienced the types of changes seen in western forests in recent decades, they too are vulnerable to drought and could experience significant changes with increased severity, frequency, or duration in drought. Throughout the continental United States, the combination of projected large climate-induced shifts in suitable habitat from modeling studies and limited potential for the rapid migration of tree populations suggests that changing tree and forest biogeography could substantially lag habitat shifts already underway. Forest management practices can partially ameliorate drought impacts through reductions in stand density, selection of drought-tolerant species and genotypes, artificial regeneration, and the development of multistructured stands. However, silvicultural treatments also could exacerbate drought impacts unless implemented with careful attention to site and stand characteristics. Gaps in our understanding should motivate new research on the effects of interactions involving climate and other species at the stand scale and how interactions and multiple responses are represented in models. This assessment indicates that, without a stronger empirical basis for drought impacts at the stand scale, more complex models may provide limited guidance.
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
Data have been compiled from published sources on nitrogen (N) fluxes in precipitation, throughfall, and leaching from 69 forest ecosystems at 50 sites throughout China, to examine at a national level: (1) N input in precipitation and throughfall, (2) how precipitation N changes after the interaction with canopy, and (3) whether N leaching increases with increasing N deposition and, if so, to what extent. The deposition of dissolved inorganic N (DIN) in precipitation ranged from 2.6 to 48.2 kg N ha0908081 year0908081, with an average of 16.6 kg N ha0908081 year0908081. Ammonium was the dominant form of N at most sites, accounting for, on average, 63% of total inorganic N deposition. Nitrate accounted for the remaining 37%. On average, DIN fluxes increased through forest canopies, by 40% and 34% in broad-leaved and coniferous forests, respectively. No significant difference in throughfall DIN inputs was found between the two forest types. Overall, 22% of the throughfall DIN input was leached from forest ecosystems in China, which is lower than the 5009000959% observed for European forests. Simple calculations indicate that Chinese forests have great potential to absorb carbon dioxide from the atmosphere, because of the large forest area and high N deposition.
DOIURLPMID [本文引用: 1]
Abstract Forest vegetation and soils have been suggested as potentially important sinks for carbon (C) with appropriate management and thus are implicated as effective tools in stabilizing climate even with increasing anthropogenic release of CO 2 . Drought, however, which is often predicted to increase in models of future climate change, may limit net primary productio (NPP) of dry forest types, with unknown effects on soil C storage. We studied C dynamics of a deciduous temperate forest of Hungary that has been subject to significant decreases in precipitation and increases in temperature in recent decades. We resampled plots that were established in 1972 and repeated the full C inventory by analyzing more than 4 decades of data on the number of living trees, biomass of trees and shrubs, and soil C content. Our analyses show that the decline in number and biomass of oaks started around the end of the 1970s with a 71% reduction in the number of sessile oak stems by 2014. Projected growth in this forest, based on the yield table's data for Hungary, was 4.60002kg C/m 2 . Although new species emerged, this new growth and small increases in oak biomass resulted in only 1.90002kg C/m 2 increase over 410002years. The death of oaks increased inputs of coarse woody debris to the surface of the soil, much of which is still identifiable, and caused an increase of 15.5%, or 2.60002kg C/m 2 , in the top 10002m of soil. Stability of this fresh organic matter input to surface soil is unknown, but is likely to be low based on the results of a colocated woody litter decomposition study. The effects of a warmer and drier climate on the C balance of forests in this region will be felt for decades to come as woody litter inputs decay, and forest growth remains impeded. 0008 2017 John Wiley & Sons Ltd.
DOIURLPMID [本文引用: 1]
Extreme droughts, heat waves, frosts, precipitation, wind storms and other climate extremes may impact the structure, composition and functioning of terrestrial ecosystems, and thus carbon cycling and its feedbacks to the climate system. Yet, the interconnected avenues through which climate extremes drive ecological and physiological processes and alter the carbon balance are poorly understood. Here, we review the literature on carbon cycle relevant responses of ecosystems to extreme climatic events. Given that impacts of climate extremes are considered disturbances, we assume the respective general disturbance-induced mechanisms and processes to also operate in an extreme context. The paucity of well-defined studies currently renders a quantitative meta-analysis impossible, but permits us to develop a deductive framework for identifying the main mechanisms (and coupling thereof) through which climate extremes may act on the carbon cycle. We find that ecosystem responses can exceed the duration of the climate impacts via lagged effects on the carbon cycle. The expected regional impacts of future climate extremes will depend on changes in the probability and severity of their occurrence, on the compound effects and timing of different climate extremes, and on the vulnerability of each land-cover type modulated by management. Although processes and sensitivities differ among biomes, based on expert opinion, we expect forests to exhibit the largest net effect of extremes due to their large carbon pools and fluxes, potentially large indirect and lagged impacts, and long recovery time to regain previous stocks. At the global scale, we presume that droughts have the strongest and most widespread effects on terrestrial carbon cycling. Comparing impacts of climate extremes identified via remote sensing vs. ground-based observational case studies reveals that many regions in the (sub-)tropics are understudied. Hence, regional investigations are needed to allow a global upscaling of the impacts of climate extremes on global carbon limate feedbacks.
DOIURL [本文引用: 1]
URL
URL
DOIURL [本文引用: 1]
鼎湖山自然保护区于1982年分别在针叶林、针阔叶混交林和常绿阔叶林内建立了3个永久样地.样地建立前,针叶林常受到人类的干扰,其它2种一直受到较好的保护.本文植物物种动态的研究和分析基于对3个样地在1982年和1994年的两次调查.结果表明,物种多样性的顺序是阔叶林>混交林>针叶林.就乔木(DBH>2.5cm)而言,3个样地的物种多样性和均匀度指数从1982年至1994年都已增长.除阔叶林外,两次调查的结果差别很大,这是因为在保护之下它们正产生进展演替.至于林下层植物则正好相反,除阔叶林外,针阔叶混交林和常绿阔叶林的多样性及均匀度指数均下降.物种的丰富度和个体数的变化同多样性指数变化一致.物种的组成变化符合演替规律,随着针叶林发展成为混交林,混交林发展成为阔叶林,其林冠变得较为密集,因而有更多的耐阴植物侵入,而一些喜光(不耐阴的)种消失.研究结果表明,人类的干扰将降低植物的物种多样性,而保护将使物种多样性得到恢复和提高.
DOIURL [本文引用: 1]
鼎湖山自然保护区于1982年分别在针叶林、针阔叶混交林和常绿阔叶林内建立了3个永久样地.样地建立前,针叶林常受到人类的干扰,其它2种一直受到较好的保护.本文植物物种动态的研究和分析基于对3个样地在1982年和1994年的两次调查.结果表明,物种多样性的顺序是阔叶林>混交林>针叶林.就乔木(DBH>2.5cm)而言,3个样地的物种多样性和均匀度指数从1982年至1994年都已增长.除阔叶林外,两次调查的结果差别很大,这是因为在保护之下它们正产生进展演替.至于林下层植物则正好相反,除阔叶林外,针阔叶混交林和常绿阔叶林的多样性及均匀度指数均下降.物种的丰富度和个体数的变化同多样性指数变化一致.物种的组成变化符合演替规律,随着针叶林发展成为混交林,混交林发展成为阔叶林,其林冠变得较为密集,因而有更多的耐阴植物侵入,而一些喜光(不耐阴的)种消失.研究结果表明,人类的干扰将降低植物的物种多样性,而保护将使物种多样性得到恢复和提高.
DOIURL [本文引用: 2]
Hurricanes are infrequent but influential disruptors of ecosystem processes in the southeastern Atlantic and Gulf coasts. Every southeastern forested wetland has the potential to be struck by a tropical cyclone. We examined the impact of Hurricane Hugo on two paired coastal watersheds in South Carolina in terms of stream flow and vegetation dynamics, both before and after the hurricanes passage in 1989. The study objectives were to quantify the magnitude and timing of changes including a reversal in relative streamflow-difference between two paired watersheds, and to examine the selective impacts of a hurricane on the vegetative composition of the forest. We related these impacts to their potential contribution to change watershed hydrology through altered evapotranspiration processes. Using over thirty years of monthly rainfall and streamflow data we showed that there was a significant transformation in the hydrologic character of the two watersheds a transformation that occurred soon after the hurricanes passage. We linked the change in the rainfall-runoff relationship to a catastrophic shift in forest vegetation due to selective hurricane damage. While both watersheds were located in the path of the hurricane, extant forest structure varied between the two watersheds as a function of experimental forest management techniques on the treatment watershed. We showed that the primary damage was to older pines, and to some extent larger hardwood trees. We believe that lowered vegetative water use impacted both watersheds with increased outflows on both watersheds due to loss of trees following hurricane impact. However, one watershed was able to recover to pre hurricane levels of canopy transpiration at a quicker rate due to the greater abundance of pine seedlings and saplings in that watershed.
DOIURLPMID
Abstract Significant changes in the composition of tree species have been observed in various forests worldwide. We hypothesised that these changes might result from variable sensitivities of species to global change, and species sensitivities might be quantified, using functional traits. Employing long-term (1978-2010) species abundance data of 48 tree species from a permanent subtropical forest plot, where multiple global change factors have been observed, including soil drying, we examined the relationships between temporal trends in abundance and suits of functional traits. We found that species with high photosynthesis rates, leaf phosphorus and nitrogen concentrations, specific leaf area, hydraulic conductivity, turgor loss point and predawn leaf water potential had increased in abundance, while species with opposite trait patterns had decreased. Our results demonstrate that functional traits underlie tree species abundance dynamics in response to drought stress, thus linking traits to compositional shifts in this subtropical forest under global changes.
[本文引用: 1]
[本文引用: 1]
DOIURLPMID
We document changes in forest structure between historical (1930s) and contemporary (2000s) surveys of California vegetation through comparisons of tree abundance and size across the state and within several ecoregions. Across California, tree density in forested regions increased by 30% between the two time periods, whereas forest biomass in the same regions declined, as indicated by a 19% reduction in basal area. These changes reflect a demographic shift in forest structure: larger trees (>61 cm diameter at breast height) have declined, whereas smaller trees (<30 cm) have increased. Large tree declines were found in all surveyed regions of California, whereas small tree increases were found in every region except the south and central coast. Large tree declines were more severe in areas experiencing greater increases in climatic water deficit since the 1930s, based on a hydrologic model of water balance for historical climates through the 20th century. Forest composition in California in the last century has also shifted toward increased dominance by oaks relative to pines, a pattern consistent with warming and increased water stress, and also with paleohistoric shifts in vegetation in California over the last 150,000 y.
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
Based on a long term survey of plant population dynamics, we investigated 26 gaps to study the factors that affect sapling species composition and biodiversity in gaps at different succession stages in Dinghushan Biosphere Reserve. Gap formation, gap size and age, topography and location, adult tree around gaps, saplings of tree species growing in the gaps were investigated. The formation of gaps resulted from landslide, storm, pest damage, etc. Sapling composition, sapling density and diversity in gaps were different due to different causes of gap formation. Species growing in the gaps were quite different because of their differences in shade tolerance and in succession stages. Differentiation of regeneration niche was caused by variation of gap size, age and topographic factor, which resulted in the appearance of sapling guild in the gaps.
DOIURL [本文引用: 1]
Based on a long term survey of plant population dynamics, we investigated 26 gaps to study the factors that affect sapling species composition and biodiversity in gaps at different succession stages in Dinghushan Biosphere Reserve. Gap formation, gap size and age, topography and location, adult tree around gaps, saplings of tree species growing in the gaps were investigated. The formation of gaps resulted from landslide, storm, pest damage, etc. Sapling composition, sapling density and diversity in gaps were different due to different causes of gap formation. Species growing in the gaps were quite different because of their differences in shade tolerance and in succession stages. Differentiation of regeneration niche was caused by variation of gap size, age and topographic factor, which resulted in the appearance of sapling guild in the gaps.
URL [本文引用: 2]
The processes of forest succession could be expressed by the dynamic characteristics of dominant populations in forest. The dynamic characteristics of two heliophyte species, Castanopsis chinensis and Schima superba in forest succession in Dinghushan were studied. The main results were as follows: 1. The niche breadths of the 2 species increased after their intruding in the needle-leaved forest, and reached the maximum when they became the dominant populations in evergreen broad-leaved forest, and then reduced gradually, but did not disappear at the climax stage. 2. In the process of the succession, changes in distribution patterns of the two populations were from clustering to random. 3. During the succession, the association and niche overlap of these two populations with Pinus massoniana population decreased, while with some mesophyte populations increased at first and reached at the highest in the evergreen broad-leaved forest stage, then reduced at the late stage. The results suggested that the features of the two populations were that of pioneer species; and the cycle and regeneration of mature community could cause the uneven and inlaying in community structure which is the reason for existence of these heliophyte population at the late stage of succession.
URL [本文引用: 2]
The processes of forest succession could be expressed by the dynamic characteristics of dominant populations in forest. The dynamic characteristics of two heliophyte species, Castanopsis chinensis and Schima superba in forest succession in Dinghushan were studied. The main results were as follows: 1. The niche breadths of the 2 species increased after their intruding in the needle-leaved forest, and reached the maximum when they became the dominant populations in evergreen broad-leaved forest, and then reduced gradually, but did not disappear at the climax stage. 2. In the process of the succession, changes in distribution patterns of the two populations were from clustering to random. 3. During the succession, the association and niche overlap of these two populations with Pinus massoniana population decreased, while with some mesophyte populations increased at first and reached at the highest in the evergreen broad-leaved forest stage, then reduced at the late stage. The results suggested that the features of the two populations were that of pioneer species; and the cycle and regeneration of mature community could cause the uneven and inlaying in community structure which is the reason for existence of these heliophyte population at the late stage of succession.
[本文引用: 2]
[本文引用: 2]
DOIURL [本文引用: 2]
通过对黄果厚壳桂的生态学特征研究表明,黄果厚壳桂是南亚热带季风绿阔叶林中地生性顶极群落中的优势种群,在南亚热带低山区域有广泛的分布,在森林群落演替的过程中,黄果厚壳桂是最早进入群落的中性性树种之一,其具有较高的光合速率和摄取营养物质的能力,一旦获得适应的生境,其生物量和第一生产力将在群落占较大比重.黄果厚壳桂种群为优势种组成的生态系统内小气候相对稳定,各生理,生态特性差异较小,黄果厚壳桂是地带性植
DOIURL [本文引用: 2]
通过对黄果厚壳桂的生态学特征研究表明,黄果厚壳桂是南亚热带季风绿阔叶林中地生性顶极群落中的优势种群,在南亚热带低山区域有广泛的分布,在森林群落演替的过程中,黄果厚壳桂是最早进入群落的中性性树种之一,其具有较高的光合速率和摄取营养物质的能力,一旦获得适应的生境,其生物量和第一生产力将在群落占较大比重.黄果厚壳桂种群为优势种组成的生态系统内小气候相对稳定,各生理,生态特性差异较小,黄果厚壳桂是地带性植
DOIURLPMID [本文引用: 1]
Abstract The boreal forest, one of the largest biomes on Earth, provides ecosystem services that benefit society at levels ranging from local to global. Currently, about two-thirds of the area covered by this biome is under some form of management, mostly for wood production. Services such as climate regulation are also provided by both the unmanaged and managed boreal forests. Although most of the boreal forests have retained the resilience to cope with current disturbances, projected environmental changes of unprecedented speed and amplitude pose a substantial threat to their health. Management options to reduce these threats are available and could be implemented, but economic incentives and a greater focus on the boreal biome in international fora are needed to support further adaptation and mitigation actions. Copyright 2015, American Association for the Advancement of Science.
[本文引用: 2]
[本文引用: 2]
URL [本文引用: 1]
森林演替是森林生态动力源驱动下森林再生的生态学过程,自20世纪初建立群落演替理论以来,演替研究成为生态学研究中的热点.客观准确地认识森林演替规律,研究森林演替动力学机理及其模型,是科学管理森林生态系统的需要;对于天然林保护工程与森林植被的恢复重建,具有重要的理论与实际意义.干扰是森林循环的驱动力,导致森林生态系统时空异质性,是更新格局和生态学过程的主要影响因素.它可改变资源的有效性,干扰导致的林隙是森林循环的起点.回顾了目前演替研究的几种方法,即马尔科夫模型、林窗模型(GAP)、陆地生物圈模型(BIOME)和非线性演替模式.介绍了气候变化对森林演替的影响;并在已有成果的基础上,提出了目前研究存在的问题及未来的发展方向.
URL [本文引用: 1]
森林演替是森林生态动力源驱动下森林再生的生态学过程,自20世纪初建立群落演替理论以来,演替研究成为生态学研究中的热点.客观准确地认识森林演替规律,研究森林演替动力学机理及其模型,是科学管理森林生态系统的需要;对于天然林保护工程与森林植被的恢复重建,具有重要的理论与实际意义.干扰是森林循环的驱动力,导致森林生态系统时空异质性,是更新格局和生态学过程的主要影响因素.它可改变资源的有效性,干扰导致的林隙是森林循环的起点.回顾了目前演替研究的几种方法,即马尔科夫模型、林窗模型(GAP)、陆地生物圈模型(BIOME)和非线性演替模式.介绍了气候变化对森林演替的影响;并在已有成果的基础上,提出了目前研究存在的问题及未来的发展方向.
DOIURL [本文引用: 1]
Natural or human-influenced disturbances are important to the health and diversity of forests,which in turn,are important to the water quantity and quality exported from a catchment.However,human-induced disturbances (prescribed fire and harvesting) have been decreasing,and natural disturbances (fires and insects) have been increasing in frequency and severity.One such natural disturbance is the mountain pine beetle (MPB),(Dendroctonus ponderosae) an endemic species.A recent epidemic resulted in the mortality of millions of hectares of lodgepole pine (Pinus contorta) forests in Colorado,USA.Beetle-induced tree mortality brings about changes to the hydrologic cycle,including decreased transpiration and interception with the loss of canopy cover.This study examined the effect of the mountain pine beetle kill on source water contributions to streamflow in snowmeltdominated headwater catchments using stable isotopes (2H and 18O) as tracers.Study catchments with varying level of beetle-killed forest area (6 to 97) were sampled for groundwater,surface water,and precipitation.Streams were sampled to assess whether beetle-killed forests have altered source water contributions to streamflow.Groundwater contributions increased with increasing beetle-killed forest area (p =0.008).Both rain and snow contributions were negatively correlated with beetle-killed forest area (p =0.035 and p =0.011,respectively).As the beetle-killed forest area increases,so does fractional groundwater contribution to streamflow.
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
DOIURLPMID
The finding that mature forest ecosystems increase carbon in woody tissues and mineral soils indicates that the original equilibriums are being pushed to a higher state. The final driving forces will probably be increasing and nitrogen deposition, global warming, and changes to precipitation patterns. However, which part of a mature forest bears the direct impacts of environmental changes and reactivates the balanced ecosystem processes remains unclear. Here, we investigated the living biomass of mature forests in the tropical and subtropical biomes in China and found that active organs and small individuals have accumulated carbon at a rate of 203 kg C ha(-1) yr(-1) in recent decades, whereas the woody tissues did not display carbon accumulation with statistical significance. Our findings indicate that the increased labile plant inputs may have shifted mature forests from their previous equilibrium and caused them to enter a new non-equilibrium state.
[本文引用: 2]
[本文引用: 2]
DOIURL [本文引用: 2]
鼎湖山南亚热带常绿阔叶林是北回归线附近保存较完好的地带性植被,是在南亚热带季风湿润型气候条件下发育的植被类型,具有热带向亚热带的过渡性质,群落结构相对复杂,组成种类相对丰富。参照巴拿马巴洛科罗拉多岛(Barro Colorado Island,BCI)50hm^2热带雨林样地的技术规范,采用中国森林生物多样性监测网络的统一调查研究方法,于2005年在鼎湖山南亚热带常绿阔叶林建立了20hm^2固定监测样地,调查并鉴定了样方内胸径(Diameter at breast height,DBH)大于1cm的木本植物。首次调查结果表明,群落内共有木本植物210种,71617个活的个体,分属于56科119属。从乔木区系的组成及其特点可以看出,其南亚热带的区系成分占绝对优势,并呈现出由亚热带向热带过渡的趋势。群落垂直结构复杂,地上成层现象较明显,乔木可分为3层,其中重要值最大的锥(Casttmopsis chinensis)、木荷(Schima sumrba)和黄杞(Engelhardtia raxburghiana)均是乔木上层的优势种;中层是群落的主要层,由厚壳桂(Cryptocarya chinensis)、黄叶树(Xamhophyllum hainanense)和华润楠(Machilus chinensis)等中生和耐阴树种组成;下层成分较复杂,物种多样性高,不同地段的物种组成差异较大。样地内物种十分丰富,种.面积曲线拟合显示其物种数量接近于BCI。稀有种比例极高,有110种,占总物种数的52.38%,其中有45%的稀有种源于物种本身的特性,有20%源于区系交汇,人为或自然干扰造成的稀有种占30%以上。样地中所有个体的径级分布(以1cm等级排列)明显呈倒“J”形,表示群落稳定与正常生长状态。根据对优势种径级结构的分布分析,将各树种的径级结构归纳为4种类型:1)峰型(中径级个体储备型),此类物种为乔木上层优势种;2)倒“J”型(正常型),此类物种为乔木
DOIURL [本文引用: 2]
鼎湖山南亚热带常绿阔叶林是北回归线附近保存较完好的地带性植被,是在南亚热带季风湿润型气候条件下发育的植被类型,具有热带向亚热带的过渡性质,群落结构相对复杂,组成种类相对丰富。参照巴拿马巴洛科罗拉多岛(Barro Colorado Island,BCI)50hm^2热带雨林样地的技术规范,采用中国森林生物多样性监测网络的统一调查研究方法,于2005年在鼎湖山南亚热带常绿阔叶林建立了20hm^2固定监测样地,调查并鉴定了样方内胸径(Diameter at breast height,DBH)大于1cm的木本植物。首次调查结果表明,群落内共有木本植物210种,71617个活的个体,分属于56科119属。从乔木区系的组成及其特点可以看出,其南亚热带的区系成分占绝对优势,并呈现出由亚热带向热带过渡的趋势。群落垂直结构复杂,地上成层现象较明显,乔木可分为3层,其中重要值最大的锥(Casttmopsis chinensis)、木荷(Schima sumrba)和黄杞(Engelhardtia raxburghiana)均是乔木上层的优势种;中层是群落的主要层,由厚壳桂(Cryptocarya chinensis)、黄叶树(Xamhophyllum hainanense)和华润楠(Machilus chinensis)等中生和耐阴树种组成;下层成分较复杂,物种多样性高,不同地段的物种组成差异较大。样地内物种十分丰富,种.面积曲线拟合显示其物种数量接近于BCI。稀有种比例极高,有110种,占总物种数的52.38%,其中有45%的稀有种源于物种本身的特性,有20%源于区系交汇,人为或自然干扰造成的稀有种占30%以上。样地中所有个体的径级分布(以1cm等级排列)明显呈倒“J”形,表示群落稳定与正常生长状态。根据对优势种径级结构的分布分析,将各树种的径级结构归纳为4种类型:1)峰型(中径级个体储备型),此类物种为乔木上层优势种;2)倒“J”型(正常型),此类物种为乔木
[本文引用: 1]
[本文引用: 1]
DOIURLPMID
Abstract Tree mortality, growth, and recruitment are essential components of forest dynamics and resiliency, for which there is great concern as climate change progresses at high latitudes. Tree mortality has been observed to increase over the past decades in many regions, but the causes of this increase are not well understood, and we know even less about long-term changes in growth and recruitment rates. Using a dataset of long-term (1958-2009) observations on 1,680 permanent sample plots from undisturbed natural forests in western Canada, we found that tree demographic rates have changed markedly over the last five decades. We observed a widespread, significant increase in tree mortality, a significant decrease in tree growth, and a similar but weaker trend of decreasing recruitment. However, these changes varied widely across tree size, forest age, ecozones, and species. We found that competition was the primary factor causing the long-term changes in tree mortality, growth, and recruitment. Regional climate had a weaker yet still significant effect on tree mortality, but little effect on tree growth and recruitment. This finding suggests that internal community-level processes-more so than external climatic factors-are driving forest dynamics.
DOIURL [本文引用: 1]
Summary Despite the mounting evidence for positive diversity–productivity relationships found in controlled experiments, diversity effects on productivity in natural systems remain hotly debated. Understanding the multivariate links between diversity and productivity in natural systems, in particular natural forests that host the majority of terrestrial biodiversity and provide essential services for humanity, remains a critical challenge for ecologists. We analysed data from 448 plots of varying tree species diversity, stand ages and local nutrient availability in Canada's boreal forest (52°30′–55°24′02N latitude and 102°36′–108°02W longitude). We used structural equation models to link multivariate relationships between above-ground biomass, tree species diversity, stand age and soil nutrient availability. Above-ground biomass increased with diversity indirectly via increasing tree size inequality, increased with stand age and was higher on sites of medium soil nutrient regime directly as well as indirectly via increased tree size inequality. Synthesis . Our results demonstrate positive diversity effects on above-ground biomass in natural forests of diverse forest ages and soil resource availability. Furthermore, we show that tree size inequality acts as a mechanism for the positive diversity effects on above-ground biomass and as a mechanism in regulating above-ground biomass and species diversity simultaneously via interactions among individuals in natural forests.
DOIURL [本文引用: 2]
根据广东省中部鼎湖山自然保护区季风常绿阔叶林锥栗_荷木_黄果厚壳桂 (Castanopsischinensis_Schimasu perba_Cryptocaryaconcinna)群落 8年的监测 ,用成对双样本均值分析的方法对群落个体数、生物量等进行了对比研究 ,初步揭示了群落真正时间意义上的变化 :1 )群落个体数呈逐年递减的趋势 ,1 992、1 994、1 999年分别为 3 979、3 82 2、3 531株·hm-2 ,1 994、1 999年比 1 992年分别减少了 3 .9%、1 1 .3 % ,经t_检验 ,差异极显著。类似的 ,群落生物量大体上也呈下降的趋势 ,1 992、1 994和 1 999年分别为 2 93 .1、30 4 .3和 2 4 8.5t·hm-2 ,1 994、1 999年分别为 1 992年的1 0 3 .8%、84.8% ,差异不显著。 2 )不同径级树木个体数和生物量年间存在差异。胸径≤ 30cm的树木个体数年间差异极显著。胸径 90cm以上的树木个体数在 1 992~ 1 999年间减少 50 %。胸径 40cm以下和 90cm以上树木生物量年间差异不显著。 1 994、1 999年中径级 (尤其是 40~ 50cm)树木的生物量有一定的发展 ,分别增长了 1 .36 %、8.7%。 3)黄果厚壳桂、云南银柴物种个体数极显著减少 ,锥栗 1 992~ 1 999年间生物量减少 31 .1 % ,黄果厚壳桂 1 999年生物量显著低于 1 992年。 4)鼎湖山季风常绿阔叶林正朝着中生顶级群落演替。
DOIURL [本文引用: 2]
根据广东省中部鼎湖山自然保护区季风常绿阔叶林锥栗_荷木_黄果厚壳桂 (Castanopsischinensis_Schimasu perba_Cryptocaryaconcinna)群落 8年的监测 ,用成对双样本均值分析的方法对群落个体数、生物量等进行了对比研究 ,初步揭示了群落真正时间意义上的变化 :1 )群落个体数呈逐年递减的趋势 ,1 992、1 994、1 999年分别为 3 979、3 82 2、3 531株·hm-2 ,1 994、1 999年比 1 992年分别减少了 3 .9%、1 1 .3 % ,经t_检验 ,差异极显著。类似的 ,群落生物量大体上也呈下降的趋势 ,1 992、1 994和 1 999年分别为 2 93 .1、30 4 .3和 2 4 8.5t·hm-2 ,1 994、1 999年分别为 1 992年的1 0 3 .8%、84.8% ,差异不显著。 2 )不同径级树木个体数和生物量年间存在差异。胸径≤ 30cm的树木个体数年间差异极显著。胸径 90cm以上的树木个体数在 1 992~ 1 999年间减少 50 %。胸径 40cm以下和 90cm以上树木生物量年间差异不显著。 1 994、1 999年中径级 (尤其是 40~ 50cm)树木的生物量有一定的发展 ,分别增长了 1 .36 %、8.7%。 3)黄果厚壳桂、云南银柴物种个体数极显著减少 ,锥栗 1 992~ 1 999年间生物量减少 31 .1 % ,黄果厚壳桂 1 999年生物量显著低于 1 992年。 4)鼎湖山季风常绿阔叶林正朝着中生顶级群落演替。
DOIURLPMID [本文引用: 1]
<P>There is evidence that climate change induced tree mortalities in boreal and temperate forests and increased forest turnover rates (both mortality and recruitment rates) in Amazon forests. However, no study has examined China's tropical and subtropical evergreen broadleaved forests (TEBF) that cover >26% of China's terrestrial land. The sustainability of this biome is vital to the maintenance of local ecosystem services (e.g., carbon sequestration, biodiversity conservation, climatic regulation), many of which may influence patterns of atmospheric circulation and composition at regional to global scales. Here, we analyze time-series data collected from thirteen permanent plots within China's unmanaged TEBF to study whether and how this biome has changed over recent decades. We find that the numbers of individuals and species for shrub and small tree have increased since 1978, whereas the numbers of individuals and species for tree have decreased over this same time period. The shift in species composition is accompanied by a decrease in the mean diameter at breast height (DBH) for all individuals combined. China's TEBF may thereby be transitioning from cohorts of fewer and larger individuals to ones of more and smaller individuals, which shows a unique change pattern differing from the documented. Regional-scale drying is likely responsible for the biome's reorganization. This biome-wide reconstitution would deeply impact the regimes of carbon sequestration and biodiversity conservation and have implications for the sustainability of economic development in the area.</P>
DOIURLPMID [本文引用: 2]
Recent studies have suggested that tropical forests may not be resilient against climate change in the long term, primarily owing to predicted reductions in rainfall and forest productivity, increased tree mortality, and declining forest biomass carbon sinks. These changes will be caused by drought-induced water stress and ecosystem disturbances. Several recent studies have reported that climate change has increased tree mortality in temperate and boreal forests, or both mortality and recruitment rates in tropical forests. However, no study has yet examined these changes in the subtropical forests that account for the majority of China's forested land. In this study, we describe how the monsoon evergreen broad-leaved forest has responded to global warming and drought stress using 32 years of data from forest observation plots. Due to an imbalance in mortality and recruitment, and changes in diameter growth rates between larger and smaller trees and among different functional groups, the average DBH of trees and forest biomass have decreased. Sap flow measurements also showed that larger trees were more stressed than smaller trees by the warming and drying environment. As a result, the monsoon evergreen broad-leaved forest community is undergoing a transition from a forest dominated by a cohort of fewer and larger individuals to a forest dominated by a cohort of more and smaller individuals, with a different species composition, suggesting that subtropical forests are threatened by their lack of resilience against long-term climate change.
DOIURL [本文引用: 3]
Responses of hydrological processes to climate change are key components in the Intergovernmental Panel for Climate Change (IPCC) assessment. Understanding these responses is critical for developing appropriate mitigation and adaptation strategies for sustainable water resources management and protection of public safety. However, these responses are not well understood and little long-term evidence exists. Herein, we show how climate change, specifically increased air temperature and storm intensity, can affect soil moisture dynamics and hydrological variables based on both long-term observation and model simulations using the Soil and Water Assessment Tool (SWAT) in an intact forested watershed (the Dinghushan Biosphere Reserve) in Southern China. Our results show that, although total annual precipitation changed little from 1950 to 2009, soil moisture decreased significantly. A significant decline was also found in the monthly 7-day low flow from 2000 to 2009. However, the maximum daily streamflow in the wet season and unconfined groundwater tables have significantly increased during the same 10-year period. The significant decreasing trends on soil moisture and low flow variables suggest that the study watershed is moving towards drought-like condition. Our analysis indicates that the intensification of rainfall storms and the increasing number of annual no-rain days were responsible for the increasing chance of both droughts and floods. We conclude that climate change has indeed induced more extreme hydrological events (e.g. droughts and floods) in this watershed and perhaps other areas of Southern China. This study also demonstrated usefulness of our research methodology and its possible applications on quantifying the impacts of climate change on hydrology in any other watersheds where long-term data are available and human disturbance is negligible.
On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene.
1
2015
... 近10年来, 有关全球环境变化对森林群落结构影响的研究和讨论有很多, 变暖和降水减少(或降水变率加大)诱导的干旱对森林造成影响, 中国亚热带地区面临同样的环境问题, 研究认为气温升高和降水变率增大减少了土壤水分(
A global overview of drought and heat- induced tree mortality reveals emerging climate change risks for forests.
1
2010
... 森林群落结构是指树木个体在环境中的分布及其与周围环境之间相互作用所形成的组分和构造, 包括个体密度、生物量大小、物种-多度分布、径级结构等(
Tree mortality from drought, insects, and their interactions in a changing climate.
1
2015
... 森林群落结构是指树木个体在环境中的分布及其与周围环境之间相互作用所形成的组分和构造, 包括个体密度、生物量大小、物种-多度分布、径级结构等(
Meta-analysis reveals that hydraulic traits explain cross-species patterns of drought- induced tree mortality across the globe.
1
2016
... 森林群落结构是指树木个体在环境中的分布及其与周围环境之间相互作用所形成的组分和构造, 包括个体密度、生物量大小、物种-多度分布、径级结构等(
Larger trees suffer most during drought in forests worldwide.
2
2015
... 森林群落结构是指树木个体在环境中的分布及其与周围环境之间相互作用所形成的组分和构造, 包括个体密度、生物量大小、物种-多度分布、径级结构等(
... 鼎湖山季风林是演替近顶极群落, 该地区森林群落的演替顺序为马尾松林, 马尾松、锥和木荷等树种为主的针阔混交林, 季风林(
十五个物种多样性指数的比较研究
1
1999
... α多样性指群落的物种多样性, 是群落内物种丰富度和物种多度格局的综合体现, 常用于度量群落物种多样性的大小.α多样性指数非常多, 但各指数具有较强的相关性, 本文采用常用的Shannon- Wiener指数计算群落的α多样性(
十五个物种多样性指数的比较研究
1
1999
... α多样性指群落的物种多样性, 是群落内物种丰富度和物种多度格局的综合体现, 常用于度量群落物种多样性的大小.α多样性指数非常多, 但各指数具有较强的相关性, 本文采用常用的Shannon- Wiener指数计算群落的α多样性(
虫害叶损失造成的树木非结构性碳减少与树木生长、死亡的关系研究进展
2016
虫害叶损失造成的树木非结构性碳减少与树木生长、死亡的关系研究进展
2016
The impacts of increasing drought on forest dynamics, structure, and biodiversity in the United States.
2016
常绿阔叶林植被动态研究进展
1
2004
... 季风常绿阔叶林是中国南亚热带地区的地带性植被, 该类型森林面积广阔, 物种多样性高(
常绿阔叶林植被动态研究进展
1
2004
... 季风常绿阔叶林是中国南亚热带地区的地带性植被, 该类型森林面积广阔, 物种多样性高(
Atmospheric deposition and leaching of nitrogen in Chinese forest ecosystems.
1
2011
... 季风常绿阔叶林是中国南亚热带地区的地带性植被, 该类型森林面积广阔, 物种多样性高(
Long-term effects of climate change on carbon storage and tree species composition in a dry deciduous forest.
1
2017
... 长时间的群落结构和多样性的变化可以区分为群落总量(总个体数、总生物量)的变化和群落内物种个体数或生物量分配的变化.为了描述群落结构的变化, 以及区分上述两种原因的贡献, 我们分别计算了以下4个相互关联的指数: 分别描述群落间相似性、差异性的相似指数(SI)和β多样性指数(BI), 描述群落内物种个体数或生物量分配变化的替换指数(RI), 以及描述群落总量变化的总量差异指数(DI) (
Effects of climate extremes on the terrestrial carbon cycle: Concepts, processes and potential future impacts.
1
2015
... 森林群落结构是指树木个体在环境中的分布及其与周围环境之间相互作用所形成的组分和构造, 包括个体密度、生物量大小、物种-多度分布、径级结构等(
Neotropical secondary forest succession: Changes in structural and functional characteristics.
1
2001
... 森林群落结构是指树木个体在环境中的分布及其与周围环境之间相互作用所形成的组分和构造, 包括个体密度、生物量大小、物种-多度分布、径级结构等(
樟翠尺蛾种群动态与植物群落结构及气候因子的关系
2000
樟翠尺蛾种群动态与植物群落结构及气候因子的关系
2000
鼎湖山植物物种多样性动态
1
1998
... 鼎湖山自然保护区的季风常绿阔叶林(以下简称季风林, MEBF)主要分布于核心区三宝峰, 海拔100-490 m, 具有400多年的保护历史, 属于近演替顶极阶段(
鼎湖山植物物种多样性动态
1
1998
... 鼎湖山自然保护区的季风常绿阔叶林(以下简称季风林, MEBF)主要分布于核心区三宝峰, 海拔100-490 m, 具有400多年的保护历史, 属于近演替顶极阶段(
Hurricane impacts on a pair of coastal forested watersheds: Implications of selective hurricane damage to forest structure and streamflow dynamics.
2
2014
... 森林具有调节气候、涵养水源、碳固存、生物多样性保育等不可替代的生态系统功能和服务功能(
... ); 群落结构变化后还常常引起森林产水量和集水区溪水来源组成的改变(
Are functional traits a good predictor of global change impacts on tree species abundance dynamics in a subtropical forest?
2015
南亚热带森林演替过程中小气候的改变及对气候变化的响应
1
2014
... 近10年来, 有关全球环境变化对森林群落结构影响的研究和讨论有很多, 变暖和降水减少(或降水变率加大)诱导的干旱对森林造成影响, 中国亚热带地区面临同样的环境问题, 研究认为气温升高和降水变率增大减少了土壤水分(
南亚热带森林演替过程中小气候的改变及对气候变化的响应
1
2014
... 近10年来, 有关全球环境变化对森林群落结构影响的研究和讨论有很多, 变暖和降水减少(或降水变率加大)诱导的干旱对森林造成影响, 中国亚热带地区面临同样的环境问题, 研究认为气温升高和降水变率增大减少了土壤水分(
Twentieth-century shifts in forest structure in California: Denser forests, smaller trees, and increased dominance of oaks.
2015
A large and persistent carbon sink in the world’s forests.
1
2011
... 森林具有调节气候、涵养水源、碳固存、生物多样性保育等不可替代的生态系统功能和服务功能(
鼎湖山林窗形成特征及其对幼树组成和多样性的影响
1
2003
... 鼎湖山季风林是演替近顶极群落, 该地区森林群落的演替顺序为马尾松林, 马尾松、锥和木荷等树种为主的针阔混交林, 季风林(
鼎湖山林窗形成特征及其对幼树组成和多样性的影响
1
2003
... 鼎湖山季风林是演替近顶极群落, 该地区森林群落的演替顺序为马尾松林, 马尾松、锥和木荷等树种为主的针阔混交林, 季风林(
鼎湖山植被演替过程中椎栗和荷木种群的动态
2
1995
... 季风常绿阔叶林是中国南亚热带地区的地带性植被, 该类型森林面积广阔, 物种多样性高(
... 鼎湖山季风林是演替近顶极群落, 该地区森林群落的演替顺序为马尾松林, 马尾松、锥和木荷等树种为主的针阔混交林, 季风林(
鼎湖山植被演替过程中椎栗和荷木种群的动态
2
1995
... 季风常绿阔叶林是中国南亚热带地区的地带性植被, 该类型森林面积广阔, 物种多样性高(
... 鼎湖山季风林是演替近顶极群落, 该地区森林群落的演替顺序为马尾松林, 马尾松、锥和木荷等树种为主的针阔混交林, 季风林(
鼎湖山厚壳桂群落演替过程的组成和结构动态
2
1998
... 季风常绿阔叶林是中国南亚热带地区的地带性植被, 该类型森林面积广阔, 物种多样性高(
... 鼎湖山季风林是演替近顶极群落, 该地区森林群落的演替顺序为马尾松林, 马尾松、锥和木荷等树种为主的针阔混交林, 季风林(
鼎湖山厚壳桂群落演替过程的组成和结构动态
2
1998
... 季风常绿阔叶林是中国南亚热带地区的地带性植被, 该类型森林面积广阔, 物种多样性高(
... 鼎湖山季风林是演替近顶极群落, 该地区森林群落的演替顺序为马尾松林, 马尾松、锥和木荷等树种为主的针阔混交林, 季风林(
黄果厚壳桂种群生态学特征
2
1999
... 季风常绿阔叶林是中国南亚热带地区的地带性植被, 该类型森林面积广阔, 物种多样性高(
... 随着向气候顶极群落演替, 中生性的乔木树种黄果厚壳桂和厚壳桂将成为季风林中的优势物种(
黄果厚壳桂种群生态学特征
2
1999
... 季风常绿阔叶林是中国南亚热带地区的地带性植被, 该类型森林面积广阔, 物种多样性高(
... 随着向气候顶极群落演替, 中生性的乔木树种黄果厚壳桂和厚壳桂将成为季风林中的优势物种(
2001
2001
Forest health and global change.
1
2015
... 森林具有调节气候、涵养水源、碳固存、生物多样性保育等不可替代的生态系统功能和服务功能(
鼎湖山森林群落分析V. 群落演替的线性系统与预测
2
1985
... 季风常绿阔叶林是中国南亚热带地区的地带性植被, 该类型森林面积广阔, 物种多样性高(
... 鼎湖山季风林是演替近顶极群落, 该地区森林群落的演替顺序为马尾松林, 马尾松、锥和木荷等树种为主的针阔混交林, 季风林(
鼎湖山森林群落分析V. 群落演替的线性系统与预测
2
1985
... 季风常绿阔叶林是中国南亚热带地区的地带性植被, 该类型森林面积广阔, 物种多样性高(
... 鼎湖山季风林是演替近顶极群落, 该地区森林群落的演替顺序为马尾松林, 马尾松、锥和木荷等树种为主的针阔混交林, 季风林(
气候变化对森林演替的影响
1
2004
... 鼎湖山季风林是演替近顶极群落, 该地区森林群落的演替顺序为马尾松林, 马尾松、锥和木荷等树种为主的针阔混交林, 季风林(
气候变化对森林演替的影响
1
2004
... 鼎湖山季风林是演替近顶极群落, 该地区森林群落的演替顺序为马尾松林, 马尾松、锥和木荷等树种为主的针阔混交林, 季风林(
Effects of mountain pine beetle- killed forests on source water contributions to streamflow in headwater streams of the Colorado Rocky Mountains.
1
2017
... 森林具有调节气候、涵养水源、碳固存、生物多样性保育等不可替代的生态系统功能和服务功能(
鼎湖山锥栗+黄果厚壳桂+荷木群落生物量及其特征
1
1997
... 每个个体生物量由生物量方程(
鼎湖山锥栗+黄果厚壳桂+荷木群落生物量及其特征
1
1997
... 每个个体生物量由生物量方程(
重庆市森林生态系统服务功能价值评估
2
2014
... 森林具有调节气候、涵养水源、碳固存、生物多样性保育等不可替代的生态系统功能和服务功能(
... ), 并影响森林生态系统碳固存和物质周转速率(
重庆市森林生态系统服务功能价值评估
2
2014
... 森林具有调节气候、涵养水源、碳固存、生物多样性保育等不可替代的生态系统功能和服务功能(
... ), 并影响森林生态系统碳固存和物质周转速率(
Increasing active biomass carbon may lead to a breakdown of mature forest equilibrium.
2014
鼎湖山森林演替序列植物-土壤碳氮同位素特征
2
2016
... 鼎湖山国家级自然保护区(23.16°-23.19° N, 112.51°-112.56° E)位于广东省肇庆市鼎湖区, 总占地面积1 155 hm2, 主要地形为丘陵和低山, 海拔100-700 m, 最高峰鸡笼山海拔1 000.3 m (
... 鼎湖山自然保护区的季风常绿阔叶林(以下简称季风林, MEBF)主要分布于核心区三宝峰, 海拔100-490 m, 具有400多年的保护历史, 属于近演替顶极阶段(
鼎湖山森林演替序列植物-土壤碳氮同位素特征
2
2016
... 鼎湖山国家级自然保护区(23.16°-23.19° N, 112.51°-112.56° E)位于广东省肇庆市鼎湖区, 总占地面积1 155 hm2, 主要地形为丘陵和低山, 海拔100-700 m, 最高峰鸡笼山海拔1 000.3 m (
... 鼎湖山自然保护区的季风常绿阔叶林(以下简称季风林, MEBF)主要分布于核心区三宝峰, 海拔100-490 m, 具有400多年的保护历史, 属于近演替顶极阶段(
鼎湖山南亚热带常绿阔叶林20公顷样地群落特征研究
2
2008
... 季风常绿阔叶林是中国南亚热带地区的地带性植被, 该类型森林面积广阔, 物种多样性高(
... 采用重要值((相对多度+相对显著度+相对频度)/3)表征物种在群落中的优势程度(
鼎湖山南亚热带常绿阔叶林20公顷样地群落特征研究
2
2008
... 季风常绿阔叶林是中国南亚热带地区的地带性植被, 该类型森林面积广阔, 物种多样性高(
... 采用重要值((相对多度+相对显著度+相对频度)/3)表征物种在群落中的优势程度(
鼎湖山季风常绿阔叶林木本植物个体死亡动态
1
2006
... 鼎湖山季风林是演替近顶极群落, 该地区森林群落的演替顺序为马尾松林, 马尾松、锥和木荷等树种为主的针阔混交林, 季风林(
鼎湖山季风常绿阔叶林木本植物个体死亡动态
1
2006
... 鼎湖山季风林是演替近顶极群落, 该地区森林群落的演替顺序为马尾松林, 马尾松、锥和木荷等树种为主的针阔混交林, 季风林(
Half-century evidence from western Canada shows forest dynamics are primarily driven by competition followed by climate.
2015
Individual size inequality links forest diversity and above-ground biomass.
1
2015
... 森林具有调节气候、涵养水源、碳固存、生物多样性保育等不可替代的生态系统功能和服务功能(
南亚热带季风常绿阔叶林锥栗-荷木-黄果厚壳桂群落发展趋势探讨
2
2003
... 季风常绿阔叶林是中国南亚热带地区的地带性植被, 该类型森林面积广阔, 物种多样性高(
... 鼎湖山季风林永久样地投影面积为1 hm2.样地采用全站仪将10 000 m2划分为25个20 m × 20 m的样方, 每个20 m × 20 m样方用插值法细分为4个 10 m × 10 m的样方, 每个10 m × 10 m样方细划为4个5 m × 5 m的小样方.在1992-2015年间, 共进行了7次群落调查, 记录样地内所有胸径(DBH)大于等于1.0 cm个体的物种名、胸径、高度、坐标及生长状况等信息(
南亚热带季风常绿阔叶林锥栗-荷木-黄果厚壳桂群落发展趋势探讨
2
2003
... 季风常绿阔叶林是中国南亚热带地区的地带性植被, 该类型森林面积广阔, 物种多样性高(
... 鼎湖山季风林永久样地投影面积为1 hm2.样地采用全站仪将10 000 m2划分为25个20 m × 20 m的样方, 每个20 m × 20 m样方用插值法细分为4个 10 m × 10 m的样方, 每个10 m × 10 m样方细划为4个5 m × 5 m的小样方.在1992-2015年间, 共进行了7次群落调查, 记录样地内所有胸径(DBH)大于等于1.0 cm个体的物种名、胸径、高度、坐标及生长状况等信息(
Substantial reorganization of China’s tropical and subtropical forests: Based on the permanent plots.
1
2014
... 近10年来, 有关全球环境变化对森林群落结构影响的研究和讨论有很多, 变暖和降水减少(或降水变率加大)诱导的干旱对森林造成影响, 中国亚热带地区面临同样的环境问题, 研究认为气温升高和降水变率增大减少了土壤水分(
A climate change-induced threat to the ecological resilience of a subtropical monsoon evergreen broad-leaved forest in Southern China.
2
2013
... 鼎湖山国家级自然保护区(23.16°-23.19° N, 112.51°-112.56° E)位于广东省肇庆市鼎湖区, 总占地面积1 155 hm2, 主要地形为丘陵和低山, 海拔100-700 m, 最高峰鸡笼山海拔1 000.3 m (
... 近10年来, 有关全球环境变化对森林群落结构影响的研究和讨论有很多, 变暖和降水减少(或降水变率加大)诱导的干旱对森林造成影响, 中国亚热带地区面临同样的环境问题, 研究认为气温升高和降水变率增大减少了土壤水分(
Quantifying the hydrological responses to climate change in an intact forested small watershed in Southern China.
3
2011
... 季风常绿阔叶林是中国南亚热带地区的地带性植被, 该类型森林面积广阔, 物种多样性高(
... 随着向气候顶极群落演替, 中生性的乔木树种黄果厚壳桂和厚壳桂将成为季风林中的优势物种(
... 近10年来, 有关全球环境变化对森林群落结构影响的研究和讨论有很多, 变暖和降水减少(或降水变率加大)诱导的干旱对森林造成影响, 中国亚热带地区面临同样的环境问题, 研究认为气温升高和降水变率增大减少了土壤水分(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}