SupportedbytheSpecialScientificResearchofForestryPublicWelfareProfessionofChina(201404312) the Major Project of Agricultural (Forestry) Breeding of Zhejiang Province(2016C02056-10)
Abstract AimsLiquidambar formosanais a fast-growing and multipurpose native tree species in China. Our aim is to determine the phenotypic variations, adaption and distribution patterns of seed and fruit characteristics of L. formosana populations.Methods Data for 10 phenotypic traits of seeds and fruits including fruit diameter, peduncle length, thousand kernel weight, seed length, seed width, seed wing length, seed wing width, seed width ratio, seed wing width ratio, ratio of seed length and seed wing length, were sampled from 36 natural L. formosana populations across China. Analysis of variance (ANOVA), correlation analysis, and regression analysis were used to analyze the phenotypic variations among and within populations.Important findings Results showed that the mean phenotypic differentiation coefficient was 57.55%, which indicated that the phenotypic variation among populations was the main source of the phenotypic variation in L. formosana. The mean coefficient of variation was 15.83%, ranging from 10.05% to 24.31%, suggesting that the phenotypic variation of seeds and fruits was very plentiful. The coefficient of variation was different among populations. The mean coefficient of variation in Anfu was the highest (16.73%), while that of Jianyang was the lowest (11.48%). The pattern of geographic variation of most seed and fruit phenotypic traits was random. There was no significant correlation between the adjacent populations in phenotypic traits. The mean temperature in January, altitude and latitude played important roles in affecting the phenotypic traits. The length-width ratio of seeds decreased with the increase of altitude, which can be used to infer the altitude of the distribution of a population. Our results provide useful information for the collection of wild resources and the breeding of this economically important species. Keywords:Liquidambar formosana;seed traits;fruit characteristics;phenotypic variations;nature populations
PDF (1196KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 何庆海, 杨少宗, 李因刚, 沈鑫, 柳新红. 枫香树种群种子与果实表型性状变异分析. 植物生态学报, 2018, 42(7): 752-763 doi:10.17521/cjpe.2017.0229 HE Qing-Hai, YANG Shao-Zong, LI Yin-Gang, SHEN Xin, LIU Xin-Hong. Phenotypic variations in seed and fruit traits of Liquidambar formosana populations. Chinese Journal of Plant Ecology, 2018, 42(7): 752-763 doi:10.17521/cjpe.2017.0229
Table 1 表1 表1枫香树种群取样点的地理位置及主要气候因子 Table 1Geographic locations and main climatic factors for sampling sites of Liquidambar formosana populations
取样地点 Sampling site
取样株数 No. of individuals sampled
经度Longitude (E)
纬度Latitude (N)
海拔Altitude (m)
年平均气 温 AMAT (℃)
1月平均气温 MAT in Jan (℃)
7月平均气温 MAT in July (℃)
无霜期 Frost-free season (d)
年降水量 AP (mm)
海南黎母山 Mt. Limu, Hainan
20
109.16°
18.89°
309
24.7
19.5
28.2
365
1 400.0
海南佳西 Jiaxi, Hainan
12
109.79°
19.27°
209
22.0
16.0
26.0
362
1 951.4
海南霸王岭 Mt. Bawangling, Hainan
9
109.28°
19.14°
250
24.3
18.4
28.8
365
1 395.0
广东紫金 Zijin, Guangdong
6
115.00°
23.59°
388
20.6
11.8
27.2
357
1 761.1
广东英德 Yingde, Guangdong
7
113.40°
24.45°
273
20.7
10.9
28.9
315
1 849.0
广西凭祥 Pingxiang, Guangxi1)
15
106.79°
22.21°
178
21.3
13.2
27.6
347
1 376.0
广西隆林 Longlin, Guangxi1)
13
105.62°
25.03°
1 390
19.1
10.0
25.5
331
1 422.8
广西融水 Rongshui, Guangxi1)
12
109.37°
25.18°
329
19.3
7.5
26.5
320
1 824.8
云南富宁 Funing, Yunnan1)
15
105.73°
24.13°
1 161
19.3
11.0
25.4
327
1 176.9
福建建阳 Jianyang, Fujian
17
117.93°
27.45°
288
17.7
8.9
37.5
255
1 742.0
福建长汀 Changting, Fujian
8
116.44°
25.83°
462
17.3
9.8
27.2
260
1 711.6
福建尤溪 Youxi, Fujian
17
112.89°
25.10°
1 032
17.7
8.0
26.6
264
1 600.0
福建华安 Hua’an, Fujian
8
117.45°
24.96°
140
20.9
18.5
33.9
320
1 598.7
浙江淳安 Chun’an, Zhejiang2)
10
118.77°
29.58°
146
17.0
5.0
28.9
263
1 450.0
浙江泰顺 Taishun, Zhejiang2)
8
119.70°
27.66°
489
16.1
5.7
27.5
242
1 980.6
浙江开化 Kaihua, Zhejiang2)
9
118.41°
29.08°
338
16.3
4.6
27.8
250
1 762.1
浙江丽水 Lishui, Zhejiang2)
15
119.99°
28.54°
259
18.0
7.3
22.9
257
1 399.6
浙江文成 Wencheng, Zhejiang2)
14
120.11°
27.88°
227
18.0
7.3
28.3
286
307.0
浙江桐庐 Tonglu, Zhejiang2)
20
119.67°
29.91°
77
16.5
4.1
28.8
253
1 462.0
浙江天台 Tiantai, Zhejiang2)
18
121.10°
29.11°
464
16.8
5.0
28.3
234
1 332.0
浙江舟山 Zhoushan, Zhejiang2)
18
122.24°
30.27°
0
16.2
5.4
28.4
305
927.3
江苏句容 Jurong, Jiangsu
16
119.31°
32.13°
19
15.1
2.0
27.8
229
1 000.0
江西安福 Anfu, Jiangxi
20
114.70°
27.62°
133
17.9
6.0
28.9
282
1 516.8
江西大余 Dayu, Jiangxi
22
114.52°
25.62°
267
18.4
7.5
27.6
301
1 591.5
湖南桑植 Sangzhi, Hunan
20
110.29°
29.44°
390
16.5
4.8
28.2
255
2 029.1
湖南隆回 Longhui, Hunan
14
111.12°
27.27°
520
17.0
5.1
28.1
281
1 427.5
湖北恩施 Enshi, Hubei
13
110.21°
30.68°
958
16.3
4.9
27.1
292
1 440.0
湖北罗田 Luotian, Hubei
11
115.43°
30.83°
223
16.4
3.6
28.5
228
1 292.6
四川广元 Guangyuan, Sichuan3)
8
105.64°
32.31°
977
16.2
5.2
26.2
270
1 186.0
贵州德江 Dejiang, Guizhou
21
108.17°
28.29°
855
16.0
5.0
26.3
295
1 229.6
贵州罗甸 Luodian, Guizhou1)
21
106.59°
25.32°
916
15.2
10.0
27.0
330
1 200.0
河南西峡 Xixia, Henan3)
6
111.75°
33.65°
940
15.1
1.5
27.1
236
881.0
陕西宁强 Ningqiang, Shaanxi3)
12
106.02°
33.05°
928
12.9
1.4
23.6
247
1 178.0
陕西安康 Ankang, Shaanxi3)
11
107.86°
32.18°
917
15.5
3.1
27.6
253
799.3
陕西镇巴 Zhenba, Shaanxi3)
13
108.00°
32.74°
688
13.7
-1.3
30.6
236
1 310.2
甘肃文县 Wenxian, Gansu3)
11
105.17°
32.77°
1 450
14.4
3.6
24.8
224
574.7
1)表示西南枫香树种群; 2)表示东南枫香树种群; 3)表示西北枫香树种群。 1), southwest L. formosana populations; 2), southeast L. formosana populations; 3), northwest L. formosana populations. AMAT, annual mean air temperature; MAT, mean air temperature; AP, annual precipitation.
Table 2 表2 表2枫香树种子与果实表型性状的方差分量及种群间表型分化系数 Table 2The variance components of the seed and fruit traits of Liquidambar formosana and the phenotype differential coefficients of different groups
Table 5 表5 表5枫香树种子与果实性状及地理因子的相关分析 Table 5Correlation coefficients between the seed and fruit traits of Liquidambar formosana and geographical and environmental variables
相关系数 Correlation coefficients
果柄长 PL (mm)
果径 FD (mm)
千粒重 TKW (g)
种子长 SL (mm)
种子宽 SW (mm)
种翅长 SWL (mm)
种翅宽 SW (mm)
种子长宽比 SWR
种翅长宽比 SWWR
种长翅长比 SLSWLR
果柄长 PL
1.00
0.12
0.44**
0.46**
0.46**
0.22
0.48**
0.01
-0.04
-0.06
果径 FD
1.00
0.15
0.32*
-0.08
0.15
0.01
0.36*
0.19
-0.02
种子千粒重 TKW
1.00
0.37*
0.51**
0.04
0.48**
-0.13
-0.22
0.10
种子长 SL
1.00
0.34*
0.57**
0.49**
0.57**
0.33*
-0.28*
种子宽 SW
1.00
0.02
0.73**
-0.58**
-0.42**
0.09
种翅长S WL
1.00
0.43**
0.49**
0.83**
-0.94**
种翅宽 SWW
1.00
-0.21
-0.15
-0.30*
种子长宽比 SWR
1.00
0.66**
-0.30*
种翅长宽比 SWWR
1.00
-0.84**
种长翅长比 SLSWLR
1.00
经度 Longitude (°N)
-0.20
0.09
-0.12
0.02
-0.23
-0.01
-0.17
0.19
0.08
0.03
纬度 Latitude (°E)
0.30*
0.12
0.29*
0.10
0.24
-0.05
0.36*
-0.15
-0.27
0.13
海拔 Altitude (m)
0.23
-0.09
0.24
-0.13
0.33*
-0.08
0.36*
-0.40**
-0.30*
0.05
年平均气温 AMAT (℃)
-0.26
-0.16
-0.25
-0.04
-0.22
0.11
-0.35*
0.19
0.34*
-0.18
1月平均气温 MAT in Jan. (℃)
-0.37*
-0.08
-0.23
-0.05
-0.16
0.16
-0.27
0.11
0.33*
-0.24
7月平均气温 MAT in July (℃)
-0.18
-0.01
-0.11
0.15
-0.19
0.18
-0.11
0.27
0.27
-0.13
无霜期 frost-free season (d)
-0.30*
-0.20
-0.21
-0.18
-0.13
0.04
-0.24
-0.02
0.20
-0.16
年降水量 AP (mm)
-0.21
0.10
-0.17
0.07
-0.25
0.05
-0.23
0.28*
0.20
-0.05
FD, fruit diameter; PL, peduncle long; TKW, thousand kernel weight; SL, seed length; SW, seed width; SWL, seed wing length; SWW, seed wing width; SWR, seed width ratio; SWWR, seed wing width ratio; SLSWLR, seed length and seed wing length ratio; AMAT, annual mean air temperature; MAT in Jan., mean air temperature in January; MAT in July, mean air temperature in July; AP, annual precipitation. **, p < 0.01; *, p < 0.05.
Table 6 表6 表6地理气候因子对种子与果实性状影响的逐步回归分析 Table 6Best model for the effects of geographic and climatic factors on seed and fruit traits evaluated by stepwise regressions
种子与果实 性状 Seed and fruit trait
回归方程 Regression equations
R2
F
p
容忍度 Tolerance
果径 FD (Y1)
Y1 = -0.101 X1 + 25.022
0.135 3
5.322
0.027
1
种子宽 SW (Y2)
Y2 = 0.00006 X2 + 1.8379
0.111 7
4.274
0.046
1
种翅宽 SWW (Y3)
Y3 = 0.0123 X3 + 1.7559
0.129 5
5.058
0.031
1
种子长宽比 SWR (Y4)
Y4 = -0.0002 X2 + 4.1477
0.155 8
6.275
0.017
1
种翅长宽比 SWWR (Y5)
Y5 = 0.0203 X1 + 1.1182
0.118 6
4.282
0.046
1
X1, 1月平均气温; X2, 海拔; X3, 纬度. X1, mean annual temperature in January; X2, altitude; X3, latitude. FD, fruit diameter; SW, seed width; SWW, seed wing width; SWR, seed width ratio; SWWR, seed wing width ratio.
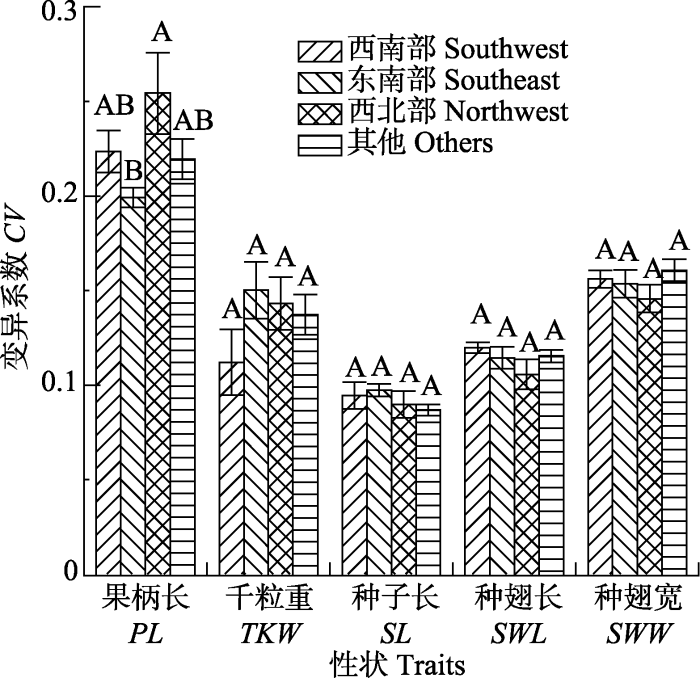
Fig. 1The coefficients of variation (CV) of the seed and fruit traits of different Liquidambar formosana populations in different regions (mean ± SE). PL, peduncle long; TKW, thousand kernel weight; SL, seed length; SWL, seed wing length; SWW, seed wing width. Different letters indicated that the differences were significant at the level of p = 0.05.
Fig. 2The seed length of Liquidambar formosana from different populations (mean ± SE). 1), southwest L. formosana populations; 2), southeast L. formosana populations; 3), northwest L. formosana populations. Different lowercase letters indicate significant differences at the level of p = 0.05.
CuiC, ZhouQY, WangLJ, TanZF, GongJ, XieXR ( 2016). Main composition analysis and comprehensive evaluation of the main agronomic traits of Flax germplasm Journal of Southwest University (Natural Science Edition), 38(12), 10-18. [本文引用: 1]
HuJZ, TianQ, LuXA ( 2007). Relationship between changes of the pigments content and leaf color changing in Phoebe chekiangensis .Journal of Northwest A&F University (Natural Science Edition), 35(10), 219-223. DOI:10.3321/j.issn:1671-9387.2007.10.042URL [本文引用: 1] The changes of the chlorophyll,carotenoid and anthocyanin content of leaves in Liquidambar formosanawere measured continuously by using spectrometer from August to December.The relationship between the leaves color and the content of pigments was also explored.The result showed that the chloro-phyll content of Liquidambar formosanadecreased from August to December in a whole.The changing trends of the chlorophyll a,b and the ratio of chlorophyll a/chlorophyll b were similar to the total content of chlorophyll basically.The anthocyanin and carotenoid content kept relatively stable during different time and the pH value of leaves decreased so that the color of anthocyanin appeared.And the leaves color turned into red even deep red.Therefore,there was a positive correlation between the leaves color and anthocya-nin/chlorophyll,but highly negative correlation with the total chlorophyll contents,and the leaves color had nothing to do with the absolute content of carotenoid and anthocyanin. [ 胡敬志, 田旗, 鲁心安 ( 2007). 枫香叶片色素含量变化及其与叶色变化的关系 西北农林科技大学学报: 自然科学版, 35(10), 219-223.] DOI:10.3321/j.issn:1671-9387.2007.10.042URL [本文引用: 1] The changes of the chlorophyll,carotenoid and anthocyanin content of leaves in Liquidambar formosanawere measured continuously by using spectrometer from August to December.The relationship between the leaves color and the content of pigments was also explored.The result showed that the chloro-phyll content of Liquidambar formosanadecreased from August to December in a whole.The changing trends of the chlorophyll a,b and the ratio of chlorophyll a/chlorophyll b were similar to the total content of chlorophyll basically.The anthocyanin and carotenoid content kept relatively stable during different time and the pH value of leaves decreased so that the color of anthocyanin appeared.And the leaves color turned into red even deep red.Therefore,there was a positive correlation between the leaves color and anthocya-nin/chlorophyll,but highly negative correlation with the total chlorophyll contents,and the leaves color had nothing to do with the absolute content of carotenoid and anthocyanin.
JiMF, ZhangXW, HanJ, DingDL ( 2013). Phenotypic diversity of cone and seed in natural Pinus tabuliformis populations in China Acta Botanica Boreali-Occidentalia Sinica , 33, 1898-1905. [本文引用: 2]
JingGZ, RenHD, YaoXH, WangKL, YangSP ( 2013). Study on phenotypic diversity of seed and fruit in natural populations ofCamellia reticulate .Journal of Nanjing Forestry University (Natural Science Edition), 37(6), 53-58. DOI:10.3969/j.issn.1000-2006.2013.06.011URL [本文引用: 1] 运用变异系数、相对极差、相关分析、巢式方差分析、方差分量比、表型分化系数及层次聚类等方法对腾冲红花油茶12个天然居群种实性状表型多样性进行分析.结果显示:腾冲红花油茶各种实性状在居群间和居群内均存在显著或极显著的差异,且居群内的变异远大于居群间的变异;不同性状变异程度最高的是单果鲜籽质量(42.99%)、单果质量(38.17%)和单籽质量(31.66%),其次是果高(15.69%)和果径(14.01%),果形指数变异程度(13.75%)最小;不同居群变异程度最高的是居群MS1 (30.05%),最低的是居群MS3(15.3%),平均居群变异程度为24.83%;不同性状表型分化程度不同,果实形状分化程度最高,果径分化程度最低;除果高和单果质量外其余各性状间均存在显著或极显著的相关性,果径和单果质量的Person简单相关系数高达0.90;果高与经度和海拔显著正相关,与纬度显著负相关,存在明显的地理变异趋势. [ 靳高中, 任华东, 姚小华, 王开良, 杨水平 ( 2013). 滇西腾冲红花油茶天然居群种实表型性状变异分析 南京林业大学学报(自然科学版), 37(6), 53-58.] DOI:10.3969/j.issn.1000-2006.2013.06.011URL [本文引用: 1] 运用变异系数、相对极差、相关分析、巢式方差分析、方差分量比、表型分化系数及层次聚类等方法对腾冲红花油茶12个天然居群种实性状表型多样性进行分析.结果显示:腾冲红花油茶各种实性状在居群间和居群内均存在显著或极显著的差异,且居群内的变异远大于居群间的变异;不同性状变异程度最高的是单果鲜籽质量(42.99%)、单果质量(38.17%)和单籽质量(31.66%),其次是果高(15.69%)和果径(14.01%),果形指数变异程度(13.75%)最小;不同居群变异程度最高的是居群MS1 (30.05%),最低的是居群MS3(15.3%),平均居群变异程度为24.83%;不同性状表型分化程度不同,果实形状分化程度最高,果径分化程度最低;除果高和单果质量外其余各性状间均存在显著或极显著的相关性,果径和单果质量的Person简单相关系数高达0.90;果高与经度和海拔显著正相关,与纬度显著负相关,存在明显的地理变异趋势.
LengHN, ChenYT, RaoLB, DuanHP, ShiX, HuangXF ( 2009). Response difference of NRA, shoot/root ratio and biomass of Liquidambar formosana provenances to different nitrogen supplies .Forest Research, 22, 190-195. DOI:10.3321/j.issn:1001-1498.2009.02.007URL [本文引用: 1] 为研究不同纬度枫香种源经过无营养胁迫生长后对供氮水平的响应差异,按种源与氮2因素裂区设计砂培试验,以N水平为主处理,枫香种源为副处理,3次重复,随机排列(水平内种源间)。结果表明:低氮胁迫会刺激枫香叶片提前形成花青素变红,且低纬区种源变红早于高纬区种源;另外低纬区种源与高纬区种源硝酸还原酶活性(NRA)存在各自的变化规律。以最高生物量为对照,在0 N水平下福建延平种源生物量降低最少(54.54%),且具有较高的生物量,但其在2N水平生物量迅速降低,耐肥性较差;其它种源生物量变化趋势一致。各种源根冠比从0 N到1/2 N水平均迅速降低:广西南丹福建延平浙江永康安徽泾县,即随着纬度的升高,枫香种源根冠比受N的影响越来越小。统计分析发现,福建延平种源在无N及低N水平时NRA极显著高于其它种源,且具有较高生物量,受环境影响较小(主要是棚内重复间光照的差异),可以认为福建延平是优良的耐低N型种源。 [ 冷华妮, 陈益泰, 饶龙兵, 段红平, 施翔, 黄秀凤 ( 2009). 枫香种源生物量和NRA对供氮水平的响应差异 林业科学研究, 22, 190-195.] DOI:10.3321/j.issn:1001-1498.2009.02.007URL [本文引用: 1] 为研究不同纬度枫香种源经过无营养胁迫生长后对供氮水平的响应差异,按种源与氮2因素裂区设计砂培试验,以N水平为主处理,枫香种源为副处理,3次重复,随机排列(水平内种源间)。结果表明:低氮胁迫会刺激枫香叶片提前形成花青素变红,且低纬区种源变红早于高纬区种源;另外低纬区种源与高纬区种源硝酸还原酶活性(NRA)存在各自的变化规律。以最高生物量为对照,在0 N水平下福建延平种源生物量降低最少(54.54%),且具有较高的生物量,但其在2N水平生物量迅速降低,耐肥性较差;其它种源生物量变化趋势一致。各种源根冠比从0 N到1/2 N水平均迅速降低:广西南丹福建延平浙江永康安徽泾县,即随着纬度的升高,枫香种源根冠比受N的影响越来越小。统计分析发现,福建延平种源在无N及低N水平时NRA极显著高于其它种源,且具有较高生物量,受环境影响较小(主要是棚内重复间光照的差异),可以认为福建延平是优良的耐低N型种源。
LiB, GuWC, LuBM ( 2002). A study on phenotypic diversity of seeds and cones characteristic in Pinus bungeana Biodiversity Science, 10, 181-188. DOI:10.3321/j.issn:1005-0094.2002.02.008URL [本文引用: 1] Totally 13 natural populations of Pinus bungeana were collected. Five characteristics, including seed length (SL), seed width (SW), cone length (FL), cone width (FW) and weight (WT) of 1000 seeds, were measured. The variance analysis of the five characteristics showed that there were significant differences among populations with the value from 2.44 to 14.68, and there were also significant differences among families within a population with the value from 7.48 to 44.53. The variance factors analysis indicated that the variation within a population was the main part of the phenotypic variation in ST =22.8%), which meant that there was a strong adaptability to environment pressure in ) and relative extreme value (i) within each population show similar tendency. Based on Duncan's Multi comparison analysis, all the 13 populations were divided into four groups. The populations in groups A and B have higher phenotypic diversity while the populations in group C and group D have lower phenotypic diversity. Seed characteristics display a significant ecological gradient variation. There is close relationship between phenotypic diversity of seeds and economic characteristics of progeny. [ 李斌, 顾万春, 卢宝明 ( 2002). 白皮松天然群体种实性状表型多样性研究 生物多样性, 10, 181-188.] DOI:10.3321/j.issn:1005-0094.2002.02.008URL [本文引用: 1] Totally 13 natural populations of Pinus bungeana were collected. Five characteristics, including seed length (SL), seed width (SW), cone length (FL), cone width (FW) and weight (WT) of 1000 seeds, were measured. The variance analysis of the five characteristics showed that there were significant differences among populations with the value from 2.44 to 14.68, and there were also significant differences among families within a population with the value from 7.48 to 44.53. The variance factors analysis indicated that the variation within a population was the main part of the phenotypic variation in ST =22.8%), which meant that there was a strong adaptability to environment pressure in ) and relative extreme value (i) within each population show similar tendency. Based on Duncan's Multi comparison analysis, all the 13 populations were divided into four groups. The populations in groups A and B have higher phenotypic diversity while the populations in group C and group D have lower phenotypic diversity. Seed characteristics display a significant ecological gradient variation. There is close relationship between phenotypic diversity of seeds and economic characteristics of progeny.
LiW, LinFR, ZhengYQ, LiB ( 2013). Phenotypic diversity of pods and seeds in natural populations of Gleditsia sinensis in southern China Chinese Journal of Plant Ecology , 37, 61-69. [本文引用: 1]
LiYQ, LiYC, WuZZ ( 2013). Variation in phenotype characters and starch content of Quercus mongolica Fisch seed from different provenances Forest Research , 26, 528-532. [本文引用: 1]
SuSP, LiY, ChongPF, GaoQ ( 2013). Correlati on analysis of phenotypic traits of Reaumuria soongorica seed in different natural populations in the Gansu Corridor Acta Prataculturae Sinica , 22(1), 87-94. DOI:10.11686/cyxb20130111URLMagsci [本文引用: 4] 采用方差分析、多重比较、相关分析、聚类分析等多种分析方法对河西走廊红砂天然分布区10个群体种子千粒重、种子长、种子宽、种子长宽比4个表型性状进行研究,以系统揭示不同群体红砂种子表型性状的变异程度和变异规律。研究结果表明,4性状在群体间差异极显著(<em><strong>P</strong></em><0.01),在群体内没有显著差异;4性状平均变异系数为0.0193,变异幅度0.0018~0.0685,其中千粒重最大,为0.0685,种子长最小,为0.0018,各群体4性状平均变异系数最大的是张掖1,为0.0484,最小的是酒泉2,为0.0066;各性状变异来源以群体间为主,占到总变异的68%以上;各表型性状均呈现出以经度、纬度和海拔为主的梯度变异规律;通过对4个表型性状各测量值的聚类分析,将10个天然群体拟划分为4个类群。对不同群体红砂种子表型性状的研究,初步了解和掌握河西走廊红砂种子的变异趋势和变异规律,可为今后开展种质资源收集与保存、遗传评价及种质创新利用以及人工辅助更新及群体遗传多样性研究提供理论依据。 [ 苏世平, 李毅, 种培芳, 高茜 ( 2013). 河西走廊不同红砂天然群体种子表型性状相关性研究 草业学报, 22(1), 87-94.] DOI:10.11686/cyxb20130111URLMagsci [本文引用: 4] 采用方差分析、多重比较、相关分析、聚类分析等多种分析方法对河西走廊红砂天然分布区10个群体种子千粒重、种子长、种子宽、种子长宽比4个表型性状进行研究,以系统揭示不同群体红砂种子表型性状的变异程度和变异规律。研究结果表明,4性状在群体间差异极显著(<em><strong>P</strong></em><0.01),在群体内没有显著差异;4性状平均变异系数为0.0193,变异幅度0.0018~0.0685,其中千粒重最大,为0.0685,种子长最小,为0.0018,各群体4性状平均变异系数最大的是张掖1,为0.0484,最小的是酒泉2,为0.0066;各性状变异来源以群体间为主,占到总变异的68%以上;各表型性状均呈现出以经度、纬度和海拔为主的梯度变异规律;通过对4个表型性状各测量值的聚类分析,将10个天然群体拟划分为4个类群。对不同群体红砂种子表型性状的研究,初步了解和掌握河西走廊红砂种子的变异趋势和变异规律,可为今后开展种质资源收集与保存、遗传评价及种质创新利用以及人工辅助更新及群体遗传多样性研究提供理论依据。
SunHJ, WangSH, ChenYT ( 2012). Response to water logging stress of different five Liquidambar formosana provenances .Journal of Nanjing Forestry University (Natural Science Edition), 36(3), 43-48. [本文引用: 2]
SunRX, LinFR, HuangP, ZhengYQ ( 2016). Moderate genetic diversity and genetic differentiation in the relict tree Liquidambar formosana Hance revealed by genetic simple sequence repeat markers Front Plant Science, 7, 1411. DOI: 10.3389/fpls.2016.01411. DOI:10.3389/fpls.2016.01411URLPMID:5030344 [本文引用: 1] Chinese sweetgum (Liquidambar formosana) is a relatively fast-growing ecological pioneer species. It is widely used for multiple purposes. To assess the genetic diversity and genetic differentiation of the species, genic SSR markers were mined from transcriptome data for subsequent analysis of the genetic diversity and population structure of natural populations. A total of 10645 potential genic SSR loci were identified in 80482 unigenes. The average frequency was one SSR per 5.12 kb, and the dinucleotide unit was the most abundant motif. A total of 67 alleles were found, with a mean of 6.091 alleles per locus and a mean polymorphism information content of 0.390. Moreover, the species exhibited a relatively moderate level of genetic diversity (He= 0.399), with the highest was found in population XY (He= 0.469). At the regional level, the southwestern region displayed the highest genetic diversity (He= 0.435) and the largest number of private alleles (n = 5), which indicated that the Southwestern region may be the diversity hot spot ofL. formosana.The AMOVA results showed that variation within populations (94.02%) was significantly higher than among populations (5.98%), which was in agreement with the coefficient of genetic differentiation (Fst= 0.076). According to the UPGMA analysis and principal coordinate analysis and confirmed by the assignment test, 25 populations could be divided into three groups, and there were different degrees of introgression among populations. No correlation was found between genetic distance and geographic distance (P > 0.05). These results provided further evidence that geographic isolation was not the primary factor leading to the moderate genetic differentiation ofL. formosana. As most of the genetic diversity ofL. formosanaexists among individuals within a population, individual plant selection would be an effective way to use natural variation in genetic improvement programs. This would be helpful to not only protect the genetic resources but also attain effective management and exploit genetic resources.
SunRX, ZhengYQ, ZhangCH, HeY, ZongYC, YuXD ( 2011). The study on phenotype variation of seed phenoltypes in different groups of Sophora japonica Journal of Hebei Agricultural University, 34(3), 65-70. DOI:10.3969/j.issn.1000-1573.2011.03.014URL [本文引用: 2] 为揭示国槐(Sophora japonica L.)种子变异程度和变异规律,本试验对国槐7个群体130个单株的种子长、种子宽、种子长宽比、表面积、体积和千粒重等6个种子表型性状作了观测分析,采用平均值,变异系数,巢氏方差分析和相关分析等方法分析了表型变异及规律。结果表明:国槐种子各表型性状存在不同程度的变异,变异系数范围为5.24%~14.57%;国槐在群体间和群体内两个层次上差异显著,种子的6个表型性状在群体间和群体内都达到了极显著水平;6个性状的平均表型分化系数为53.11%,表明群体间变异略高于群体内变异;经纬度与种子各个表型性状的相关性较低。利用系统聚类可以将7个国槐群体分为3类。国槐种子随地理变异没有明显的规律,可能是地理环境或人为栽培造成的,也可能是由于采样的不连续性造成的。 [ 孙荣喜, 郑勇奇, 张川红, 何燕, 宗亦臣, 于雪丹 ( 2011). 不同群体国槐种子表型变异研究 河北农业大学学报, 34(3), 65-70.] DOI:10.3969/j.issn.1000-1573.2011.03.014URL [本文引用: 2] 为揭示国槐(Sophora japonica L.)种子变异程度和变异规律,本试验对国槐7个群体130个单株的种子长、种子宽、种子长宽比、表面积、体积和千粒重等6个种子表型性状作了观测分析,采用平均值,变异系数,巢氏方差分析和相关分析等方法分析了表型变异及规律。结果表明:国槐种子各表型性状存在不同程度的变异,变异系数范围为5.24%~14.57%;国槐在群体间和群体内两个层次上差异显著,种子的6个表型性状在群体间和群体内都达到了极显著水平;6个性状的平均表型分化系数为53.11%,表明群体间变异略高于群体内变异;经纬度与种子各个表型性状的相关性较低。利用系统聚类可以将7个国槐群体分为3类。国槐种子随地理变异没有明显的规律,可能是地理环境或人为栽培造成的,也可能是由于采样的不连续性造成的。
WangF ( 2010). Persistent and advanced reddening of sweet gum leaves after major veins severing Journal of Forestry Research, 21, 465-468. DOI:10.1007/s11676-010-0099-7URL [本文引用: 1] The effects of major veins severing on morphological and physiological features of sweetgum ( Liquidambar styraciflua L.) leaves were investigated by observing leaf color change and measuring leaf temperature, green/luminance (G/L) value of half-lobes, leaf stomata conductance, and water content in Yamaguchi University, Japan. The palmately veined leaves of sweetgum ( Liquidambar styraciflua L.) were found more sensitive to the major vein severing than that of other species. Major veins severing resulted in serious water stresses, as indicated by the persistent reddening and/or advanced reddening of local leaf, lower leaf stomatal conductance, and higher leaf temperature, etc. Severed leaf can be clearly divided into non-severed area, transitional area, and stressed area, which the three areas have different colours and temperature. The major vein barrier can also be seen clearly. The persistent reddening and advanced reddening seem consistent with the phenomenon of red crown top of some sweetgum trees and may have similar mechanism.
WangHF, LüGH, ZhouYZ, CaoJ ( 2017). Effects of functional diversity and functional redundancy on the stability of desert plant communities under different water and salt gradients Acta Ecologica Sinica, 37, 7928-7937. DOI:10.5846/stxb201610192139URL [本文引用: 1] 功能多样性和功能冗余是物种多样性的两个组成部分,也是影响群落稳定性的两个重要因素。基于不同水盐梯度下植物功能多样性、功能冗余、物种多样性和群落稳定性及其相关关系的计算结果,分析功能多样性和功能冗余对群落稳定性的影响,结果表明:(1)功能多样性、物种多样性和群落稳定性均表现为高水高盐和中水中盐群落显著高于低水低盐群落(P 〈 0.05);(2)高水高盐群落,功能多样性与物种多样性的相关系数小于功能冗余与物种多样性的相关系数,且功能多样性与稳定性的相关系数也小于功能冗余与稳定性的相关系数,而中水中盐和低水低盐群落的相关系数则呈现相反的规律;(3)中水中盐和低水低盐群落的功能多样性的标准化偏回归系数均大于功能冗余的标准化偏回归系数;(4)典范对应分析中,土壤含水量可以解释总特征根的22.7%,而土壤含盐量仅可以解释总特征根的1.3%;(5)高水高盐群落的稳定比最接近20/80,稳定性最高;低水低盐群落远离20/80,稳定性最低。改进后的Godron稳定性测定方法与物种种群密度变异系数方法得出的结果相同。综上可知,功能多样性和功能冗余两者中与物种多样性关系更为密切者对群落稳定性的影响也越大,且两者均可提高群落稳定性,也就证明冗余假说在温带干旱荒漠区域的隐域性植物群落中是成立的;群落稳定性、功能多样性、功能冗余及物种多样性主要是受土壤含水量的影响,土壤含盐量对其影响较小。 [ 王恒方, 吕光辉, 周耀治, 曹靖 ( 2017). 不同水盐梯度下功能多样性和功能冗余对荒漠植物群落稳定性的影响 生态学报, 37, 7928-7937.] DOI:10.5846/stxb201610192139URL [本文引用: 1] 功能多样性和功能冗余是物种多样性的两个组成部分,也是影响群落稳定性的两个重要因素。基于不同水盐梯度下植物功能多样性、功能冗余、物种多样性和群落稳定性及其相关关系的计算结果,分析功能多样性和功能冗余对群落稳定性的影响,结果表明:(1)功能多样性、物种多样性和群落稳定性均表现为高水高盐和中水中盐群落显著高于低水低盐群落(P 〈 0.05);(2)高水高盐群落,功能多样性与物种多样性的相关系数小于功能冗余与物种多样性的相关系数,且功能多样性与稳定性的相关系数也小于功能冗余与稳定性的相关系数,而中水中盐和低水低盐群落的相关系数则呈现相反的规律;(3)中水中盐和低水低盐群落的功能多样性的标准化偏回归系数均大于功能冗余的标准化偏回归系数;(4)典范对应分析中,土壤含水量可以解释总特征根的22.7%,而土壤含盐量仅可以解释总特征根的1.3%;(5)高水高盐群落的稳定比最接近20/80,稳定性最高;低水低盐群落远离20/80,稳定性最低。改进后的Godron稳定性测定方法与物种种群密度变异系数方法得出的结果相同。综上可知,功能多样性和功能冗余两者中与物种多样性关系更为密切者对群落稳定性的影响也越大,且两者均可提高群落稳定性,也就证明冗余假说在温带干旱荒漠区域的隐域性植物群落中是成立的;群落稳定性、功能多样性、功能冗余及物种多样性主要是受土壤含水量的影响,土壤含盐量对其影响较小。
WangJ, YangZL, YangX, HeZS ( 2012). Effects of orientation and canopy oil deformation of fruits and seeds of Phoebe chekiangensis .Journal of Ecology and Rural Environment, 28, 385-388. [本文引用: 1]
WenCH, ChuFH ( 2017). A R2R3-MYB gene LfMYB113 is responsible for autumn leaf coloration in Formosan sweet gum (Liquidambar formosana Hance) Plant & Cell Physiology, 58, 508-521. DOI:10.1093/pcp/pcw228URLPMID:28115495 [本文引用: 1] Abstract The regulation of autumn leaf coloration in deciduous trees has long been an enigma. Due to the fact that different coloration phenotypes may be considered when planting, more understanding of the regulation mechanism is needed. In this study, a R2R3-MYB transcription factor gene LfMYB113 was identified from a subtropical deciduous tree species Formosan sweet gum (Liquidambar formosana Hance). The expression patterns of LfMYB113 in four selected phenotypes were different and were positively correlated with leaf anthocyanin content. In a 35S::LfMYB113 transgenic Nicotiana tabacum plant, both the early and late genes in the anthocyanin biosynthetic pathway were shown to be up-regulated. It was also shown that LfMYB113 can activate the promoter sequence of LfDFR1 and LfDFR2. Transient overexpression of LfMYB113 in Nicotiana benthamiana showed strong anthocyanin accumulation and pre-senescence; the latter was confirmed by up-regulation of senescence-associated genes. In addition, the activation of proLfSGR::YFP by LfMYB113 in transient experiments indicated that LfMYB113 may have a role in regulation of Chl degradation. To our knowledge, this is the first time a R2R3-MYB transcription factor has been functionally identified as one of the key regulators of autumn leaf coloration and autumn leaf senescence.
WenCH, LinSS, Shih-Shun, ChuFH ( 2015). Transcriptome analysis of a subtropical deciduous tree: Autumn leaf senescence gene expression profile of Formosan gum Plant & Cell Physiology, 56, 163-174. DOI:10.1093/pcp/pcu160URLPMID:25392065 [本文引用: 1] Abstract Autumn leaf senescence is a spectacular natural phenomenon; however, the regulation networks controlling autumnal colors and the leaf senescence program remain largely unelucidated. Whether regulation of leaf senescence is similar in subtropical deciduous plants and temperate deciduous plants is also unknown. In this study, the gene expression of a subtropical deciduous tree, Formosan gum (Liquidambar formosana Hance), was profiled. The transcriptomes of April leaves (green leaves, 'G') and December leaves (red leaves, 'R') were investigated by next-generation gene sequencing. Out of 58,402 de novo assembled contigs, 32,637 were annotated as putative genes. Furthermore, the L. formosana-specific microarray designed based on total contigs was used to extend the observation period throughout the growing seasons of 2011-2013. Network analysis from the gene expression profile focused on the genes up-regulated when autumn leaf senescence occurred. LfWRKY70, LfWRKY75, LfWRKY65, LfNAC1, LfSPL14, LfNAC100 and LfMYB113 were shown to be key regulators of leaf senescnece, and the genes regulated by LfWRKY75, LfNAC1 and LfMYB113 are candidates to link chlorophyll degradation and anthocyanin biosynthesis to senescence. In summary, the gene expression profiles over the entire year of the developing leaf from subtropical deciduous trees were used for in silico analysis and the putative gene regulation in autumn coloration and leaf senescence is discussed in this study. The Author 2014. Published by Oxford University Press on behalf of Japanese Society of Plant Physiologists. All rights reserved. For permissions, please email: journals.permissions@oup.com.
XieQJ, XuXY, ZhengJR, ZhongYT ( 2015). Study on the effect of Liquidambar formosana leaf extract on K562 cells Journal of Chinese Medicinal Materials , 38, 1493-1495. [本文引用: 1]
YangL, ShenHL, LiangLD, CuiXT, WangAZ ( 2009). Phenotypic diversity of the fruit and seed of wild Sorbus pohuashanensis in different growing areas . Journal of Northeast Forestry University, 37(2), 8-10. DOI:10.3969/j.issn.1000-5382.2009.02.004URL [本文引用: 1] 以东北地区6个野生花楸种源的新采收果实和种子为材料,研究了天然花楸果实和种子表型变异情况,结果表明:花楸果实和种子具有极丰富的表型变异,种源内的分化高于种源间;果长、果径、种长及种宽是果实和种子的关键表型性状;种子表型变异在空间分布上呈现以纬度为主的单向变异模式。 [ 杨玲, 沈海龙, 梁立东, 崔晓涛, 王爱芝 ( 2009). 不同产区野生花楸果实和种子的表型多样性 东北林业大学学报, 37(2), 8-10.] DOI:10.3969/j.issn.1000-5382.2009.02.004URL [本文引用: 1] 以东北地区6个野生花楸种源的新采收果实和种子为材料,研究了天然花楸果实和种子表型变异情况,结果表明:花楸果实和种子具有极丰富的表型变异,种源内的分化高于种源间;果长、果径、种长及种宽是果实和种子的关键表型性状;种子表型变异在空间分布上呈现以纬度为主的单向变异模式。
ZhangQ, XueJH, LiuCG ( 2011). Effects of complex pollution of CTAB and Cd 2+ on the growth of Chinese sweetgum seedlings Acta Ecologica Sinica, 31, 5824-5831. DOI:10.1002/etc.434URL [本文引用: 1] Effects of single and complex pollution of CTAB and cadmium on the growth and physiological and biochemical indices of Chinese sweetgum(Liquidambar formosana Hance) seedlings were studied by means of pot planting experiments.The results showed that lower concentrations(0.1—5g/kg) of CTAB could promote the growth of Chinese sweetgum,while CTAB was harmful to Chinese sweetgum seedlings under higher concentrations(0.5—2g/kg).The growth of Chinese sweetgum seedlings was seriously inhibited in the soil with Cd2+ concentration of 100mg/kg.The plant height,dry weight,chlorophyll content and SOD、POD、CAT activities of leaves increased first,then decreased with increasing CTAB concentrations in the same Cd2+ polluted soil.Compared with that during the fast-growing stage,total chlorophyll content dropped significantly at the end of growth stage.Moreover,SOD and POD activities of leaves at the end of growth stage were lower than those during the fast-growing stage due to a long time of stress;MDA accumulation quantity of leaves increased over time.
ZhaoXY, WangJH, ZhangJF, ZhangSG, ZhangJG, MaJW ( 2008). Study on phenotypic traits and germination characters of four taxons of Catalpa ovata genus seed .Journal of Northwest A&F University (Natural Science Edition), 36(12), 149-154. DOI:10.3321/j.issn:1671-9387.2008.12.025URL [本文引用: 1] 【Objective】 The research on seeds of C.fargesii C.ovata C.fargesii Bureau were just the opposite.Seeds soaked for 24 hours were much easier to germinate than those soaked for 12 or 48 hours and Variable temperature condition of 30 ℃ 8 hour light/20 ℃ 16 hour dark was more conducive to germination.Germination potential and germination index both had significantly positive correlation with length,width,and one thousand-seed weight.【Conclusion】 Phenotypic traits and germination features of different species differed significantly. [ 赵曦阳, 王军辉, 张金凤, 张守攻, 张建国, 马建伟 ( 2008). 梓树属4个种种子表型性状和发芽特性的研究 西北农林科技大学学报(自然科学版), 36(12), 149-154.] DOI:10.3321/j.issn:1671-9387.2008.12.025URL [本文引用: 1] 【Objective】 The research on seeds of C.fargesii C.ovata C.fargesii Bureau were just the opposite.Seeds soaked for 24 hours were much easier to germinate than those soaked for 12 or 48 hours and Variable temperature condition of 30 ℃ 8 hour light/20 ℃ 16 hour dark was more conducive to germination.Germination potential and germination index both had significantly positive correlation with length,width,and one thousand-seed weight.【Conclusion】 Phenotypic traits and germination features of different species differed significantly.
Moderate genetic diversity and genetic differentiation in the relict tree Liquidambar formosana Hance revealed by genetic simple sequence repeat markers 1 2016
 ,*浙江省林业科学研究院, 杭州 310023
,*浙江省林业科学研究院, 杭州 310023
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT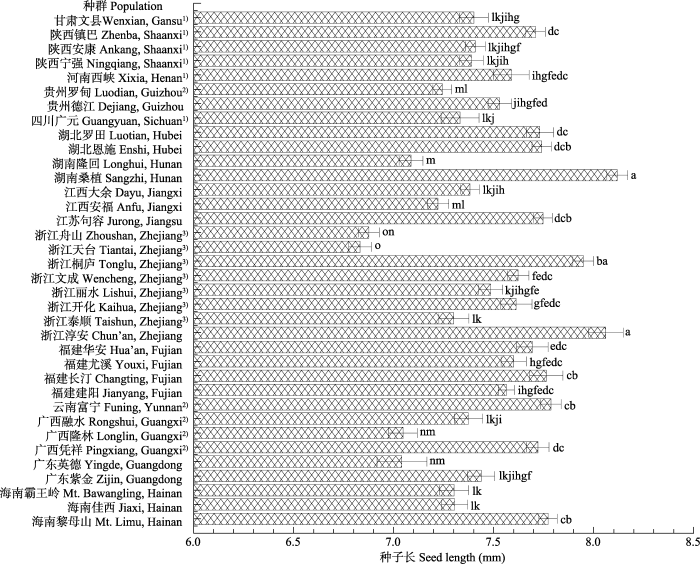 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}