 ,1,3,*, 韩淑梅1,3, 钟云芳,1,*
,1,3,*, 韩淑梅1,3, 钟云芳,1,*Modeling the potential suitable habitat of Impatiens hainanensis, a limestone-endemic plant
NING Yao1, LEI Jin-Rui2, SONG Xi-Qiang,1,3,*, HAN Shu-Mei1,3, ZHONG Yun-Fang,1,*通讯作者: Song XQ:songstrong@hainu.edu.cn;Zhong YF:zhongzyf@163.com;
编委: 倪 健
责任编辑: 李 敏
收稿日期:2018-03-28修回日期:2018-09-11网络出版日期:2018-09-20
| 基金资助: |
Received:2018-03-28Revised:2018-09-11Online:2018-09-20
| Fund supported: |

摘要
模拟物种的潜在分布区是保护管理受威胁物种的重要手段。该研究对海南岛石灰岩特有种、濒危植物——海南凤仙花(Impatiens hainanensis)的潜在适宜生境分布进行预测, 旨在为海南凤仙花的有效保护及重引入工作提供基础的科学依据。研究基于海南凤仙花8个种群分布点和12个环境变量, 利用最大熵模型(MaxEnt)和GIS技术构建海南凤仙花适宜生境预测模型, 模拟了当前时期海南凤仙花在海南岛的潜在分布区; 同时基于5个实际分布数据和5个不存在数据, 采用受试者工作特征曲线下的面积(AUC)、Kappa系数、真实技巧统计值(TSS)及总体精度4个评估指标综合评价模型的预测精度。研究结果表明: 4个评估指标值均在0.9以上, 说明MaxEnt模型能够很好地预测海南凤仙花潜在适宜生境的分布。限制其分布的主要环境因子为坡度、最干季降水量、降水量季节性变异系数。当前, 海南凤仙花的最适宜生境占海南岛总面积的1.8%, 主要分布于白沙西部与南部、昌江中部和南部、东方东部、乐东东北部。海南凤仙花潜在适宜生境分布狭窄, 且破碎化严重, 迫切需要保护。因此建议: 收集海南凤仙花各种群种子, 建立种质资源库; 将东方天安乡、江边乡及乐东东北部(佳西保护区)等可能存在最适宜生境的地区, 作为今后野外深入调查的首选区域和重引入的重点区域。
关键词:
Abstract
Keywords:
PDF (5126KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
宁瑶, 雷金睿, 宋希强, 韩淑梅, 钟云芳. 石灰岩特有植物海南凤仙花潜在适宜生境分布模拟. 植物生态学报[J], 2018, 42(9): 946-954 DOI:10.17521/cjpe.2018.0066
NING Yao, LEI Jin-Rui, SONG Xi-Qiang, HAN Shu-Mei, ZHONG Yun-Fang.
使用最大熵模型(MaxEnt)等物种分布模型预测物种的潜在分布区已成为生物多样性及生物保护领域的研究热点。预测物种的潜在适宜生境分布区对制定有效的生物多样性保护策略(Hernandez et al., 2006), 濒危物种再引入(Rushton et al., 2004; Aguilar et al., 2015), 森林资源规划(甄江红, 2008), 发现物种未知种群(Engler et al., 2004; Fois et al., 2015), 评估入侵物种传播范围及其威胁(Beaumont et al., 2005)以及气候变化对植被分布的影响(吴建国等, 2010)等具有重要意义, 同时也是有效保护受威胁物种的重要手段(Majid et al., 2015)。MaxEnt模型因具有仅利用物种出现点、易于掌握与使用且在多数情况下模拟精度通常优于其他模型的优点, 使其成为现代物种生境适宜性评价方面应用最广泛的一个工具(Hernandez et al., 2006; Merow et al., 2013)。目前, 该模型已被成功应用于珍稀濒危物种如Canacomyrica monticola (Kumar & Stohlgren, 2009), Vanda bicolor (Deb et al., 2017), 青钱柳(Cyclocarya paliurus) (刘清亮等, 2017), 蒙古扁桃(Amygdalus mongolica) (马松梅等, 2014)等的生境评价与分布预测中。Hernandez等(2006)及Deb等(2017)的研究都表明当物种出现点很少(<10), 即使低至4或5个, MaxEnt模型仍能以高精度产生有效的预测结果。尽管目前物种分布模型已广泛用于预测珍稀濒危动植物潜在适宜分布区及其适宜生境分布的变化, 然而研究仅集中于广布种或分布点较多的物种, 对于具有明显狭域性和地域性的特有种, 尤其是单一生境的特有种如石灰岩特有植物却鲜有研究。因此, 开展狭域特有种的适宜生境预测及环境因子对其分布的影响的研究, 对于了解物种适应单一生境的机制具有重要意义, 同时对物种分布模型的发展具有推动作用。
海南凤仙花(Impatiens hainanensis)是海南岛特有种, 属海南岛石灰岩特有植物(Francisco-Ortega et al., 2010), 2006年被列为海南省重点保护植物, 其种群自然分布于海拔190-1 300 m的热带山地雨林, 多见于裸露的石灰岩石缝中, 且不同种群的大小及植株生长状况差异明显(钟云芳, 2014)。经调查, 海南凤仙花主要分布于昌江县及东方市(秦新生等, 2005)。近几十年来, 由于人类活动(如开山采矿、采药)的干扰及外界自然环境尤其是气候变化的影响, 海南凤仙花种群数量和分布面积日益减少, 加上岛屿本地特有种具有地理隔离、分布狭窄的特点, 及石灰岩自然生境易干旱且先天脆弱, 海南凤仙花的生存发展面临着严峻的威胁(冷欣等, 2005; 吴静等, 2010; 钟云芳, 2014)。因此, 海南凤仙花及其生境保护迫在眉睫, 要对其进行有效的保护必须明确其地理分布范围。但目前关于该物种的具体适宜分布范围, 及限制其地理分布的主要环境因子尚不明确。因此, 本研究以海南岛为研究区, 在进行充分调查野外实际生境的基础上, 基于海南凤仙花野外实际分布点和环境数据, 使用MaxEnt, 结合GIS技术, 模拟该物种当前的潜在适宜分布区, 并分析限制其分布的环境因子, 旨在得到它的潜在适生区, 为海南凤仙花的保护管理和重引入工作提供基础的科学依据, 同时也为海南岛其他珍稀濒危植物, 尤其是生境单一物种的生境分布模拟提供参考。
1 材料和方法
1.1 研究区概况与物种分布数据
海南岛位于中国大陆的南端, 属热带北缘, 是中国最大的热带岛屿。岛内呈现中部高四周低的地势。这种特殊的地貌使得整个岛屿内不同地区的光、温、水、热等气候要素的分配差异显著。受热带季风气候的影响, 海南岛具有明显的干季和湿季之分, 5-11月为湿季, 12月至翌年4月为干季; 岛内阳光充足、日照时间长, 年平均气温在22-35 ℃。海南凤仙花为海南岛特有植物, 现今主要分布在昌江县及东方的石灰岩山地, 多见于山顶裸露的石灰岩缝隙中。经2016年6月至2018年5月期间的野外实地调查, 结合前人研究成果, 得到海南凤仙花种群分布点共13个, 各种群分布信息详见图1及附录I。在ArcGIS 10.2中随机选择其中的8个点用于模型建立, 另外的5个点用作模型检验, 此过程重复10次, 得到10套用于模型运行的分布数据。
1.2 环境变量和数据处理
本研究选取的环境变量包括地形和气候两部分。地形数据包括海拔、坡度、坡向3个变量, 收集中国科学院地理空间数据云平台(http://www.gscloud.cn) 90 m分辨率数字高程模型(DEM), 在ArcGIS10.2中运用空间分析模块下的相关工具, 提取得到海拔、坡度、坡向3个栅格图层。当前时期的气候数据从世界气候数据库(http://www.worldclim.org/)下载, 采用2.0版的数据。该数据库通过收集全球各个气象观测站1970-2000年的气候数据, 运用ANUSPLIN程序中的薄板平滑样条函数插值法, 以海拔、经度、纬度为自变量, 从而进行全球范围的插值, 得到30?分辨率(约1 km)的全球连续气候数据( Hijmans et al., 2005)。该数据包括19个生物气候变量, 具有明显的生物学意义, 通常被运用于物种分布建模和相关的生态建模中。本研究为进一步提高该气候数据分辨率, 利用海南岛16个气象观测站1981-2010年共30年的累年日值数据集(包括气温和降水等), 通过计算得到19个气候变量。运用这部分数据减去对应站点上的WorldClim数据值, 得到19个气候变量的残差。再以海南岛的经度、纬度及海拔(90 m) 3个栅格图层作为协变量, 对残差进行协同克里格插值, 生成经过地形校正的90 m分辨率的残差空间分布图层19个, 再将残差图层与研究区的WorldClim数据一起叠加, 得到研究区内的19个气候变量栅格图层。
为保证模型的准确预测, 运用ArcGIS 10.2的波段集统计工具检验22个环境变量间的多重共线性, 对于相关性较高(|r| > 0.8)的变量, 仅保留其中一个, 最终确定选用昼夜温差月平均值、等温性、气温季节性变化标准偏差、气温年较差、最冷季平均气温、年降水量、降水量季节性变异系数、最湿季降水量、最干季降水量、海拔、坡度、坡向12个环境变量(表1)用于MaxEnt模型的预测。所有的环境变量统一坐标WGS-84, 统一边界, 统一分辨率为100 m。
图1
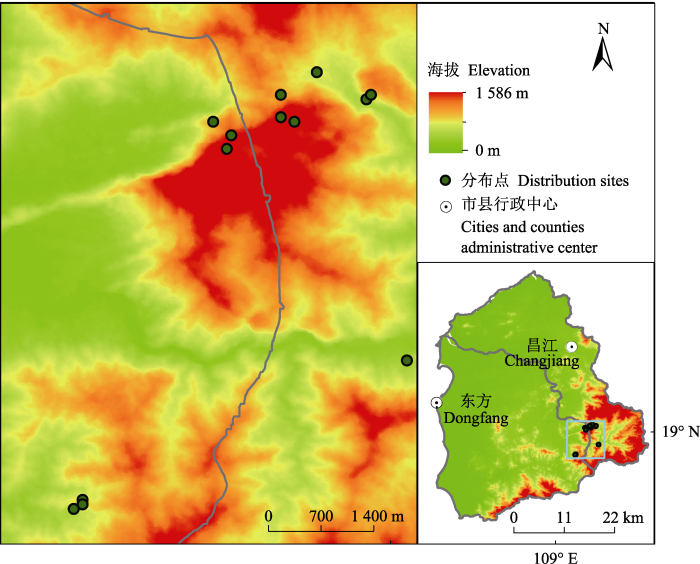 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1海南凤仙花实际地理分布图。
Fig. 1Actual geographic distribution maps of Impatiens hainanensis.
Supplement I
附录I
附录I野外调查的海南凤仙花13个种群的基本情况
Supplement I
| 地点 Sites | 种群编码 Populations code | 经度 Longitude | 纬度 Latitude | 海拔 Altitude (m) |
|---|---|---|---|---|
| 俄贤岭 Exian Moutain | Pop1 | 109.094° E | 19.013° N | 945 |
| Pop 2 | 109.098° E | 19.010° N | 890 | |
| Pop 3 | 109.097° E | 19.007° N | 1 000 | |
| Pop 4 | 109.109° E | 19.104° N | 490 | |
| Pop 5 | 109.112° E | 19.013° N | 1 050 | |
| 王下乡 Wangxia Village | Pop6 | 109.117° E | 19.024° N | 450 |
| Pop7 | 109.109° E | 19.019° N | 590 | |
| Pop8 | 109.128° E | 19.018° N | 350 | |
| Pop9 | 109.129° E | 19.019° N | 460 | |
| 皇帝洞 Huangdidong | Pop10 | 109.137° E | 18.960° N | 250 |
| 猕猴岭保护区 Mihouling Reserve | Pop11 | 109.065° E | 18.929° N | 480 |
| Pop12 | 109.065° E | 18.928° N | 400 | |
| Pop13 | 109.063° E | 18.927° N | 340 |
新窗口打开|下载CSV
Table 1
表1
表1生境评价数据信息表
Table 1
| 变量及数据 Variable and data | 代码 Code | 来源 Source | 原始坐标系 Original coordinate system |
|---|---|---|---|
| 昼夜温差月平均值 Mean diurnal range (Mean of monthly (max temperature - min temperature)) | bio2 | 世界气候数据库 Worldclim | WGS84 |
| 等温性 Isothermality | bio3 | ||
| 气温季节性变化标准偏差 Standard deviation of air temperature seasonality | bio4 | ||
| 气温年较差 Air temperature annual range | bio7 | ||
| 最冷季平均气温 Mean air temperature of coldest quarter | bio11 | ||
| 年降水量 Annual precipitation | bio12 | ||
| 降水量季节性变异系数 Coefficient of variation of precipitation seasonality | bio15 | ||
| 最湿季降水量 Precipitation of wettest quarter | bio16 | ||
| 最干季降水量 Precipitation of driest quarter | bio17 | ||
| 海拔 Elevation | ele | 地理空间数据云 Geospatial data cloud (Computer network information center of Chinese academy of sciences) | WGS84 |
| 坡度 Slope | slo | ||
| 坡向 Aspect | asp | ||
| 海南凤仙花分布点 Distribution points of Impatiens hainanensis | - | 实地调查 Field survey | WGS84 |
| 海南岛土地覆被类型图 Land cover type map of Hainan Island | - | 海南省林业厅 Forestry department of Hainan | WGS84 |
新窗口打开|下载CSV
1.3 模型运行
本研究运用MaxEnt 3.3.3k软件进行预测。将随机选取的8个海南凤仙花分布点(10套)和环境数据导入MaxEnt模型, 勾选创建反应曲线和刀切法检验, 其他参数选项默认设定。根据模型自带的Jackknife刀切法检验得到的增益结果和模型贡献率表综合地确定影响海南凤仙花的主导环境因子。通常研究会选取对模型累积贡献率在85%-95%的环境因子, 作为对物种适宜分布区影响比较重大的因子(石慰, 2013; 应凌霄等, 2016)。模型输出结果为ASCII格式, 可解释为0-1范围的生境适宜值, 将每套数据运行得到的平均的ASCII文件导入ArcGIS 10.2中转为栅格数据, 利用栅格计算器, 求得10套ASCII文件的平均值, 再将该平均值图层与海南岛土地覆被类型图进行相乘, 剔除耕地、草地、水体、城乡工矿地、居民用地等不适生生境(不适生的土地覆被类型栅格单元值为0, 适生的土地覆被类型栅格单元值为1)。再运用重分类功能中的自然间断点分级法(Jenks)进行重分类, 将海南凤仙花的生境适宜性划分为3个等级: 最适宜、次适宜和不适宜。此外, 据秦新生等(2014)的实地考察, 海南岛有石灰岩分布的市县共有6个, 即儋州、白沙、昌江、东方、三亚、保亭, 但并不排除这6个市县以外的地区没有石灰岩的分布, 因为有可能存在未调查区域, 或者其他地区在史前有可能分布着石灰岩, 因此, 本研究中预测得到的海南凤仙花适宜生境分布在这6个市县外的区域不作剔除, 最终得到海南凤仙花生境适宜性分布。
1.4 模型精度评估
为有效地评价基于小样本建模的模型预测精度, 本研究选取受试者工作特征曲线下面积(AUC)、Kappa系数、真实技巧统计值(TSS)及总体精度4个指标综合评估模型预测的准确性。其中Kappa系数、TSS和总体精度受到阈值的影响, 需要物种不存在数据来协助计算, 因此, 本研究运用褚建民等(2017)提到的具体方法, 随机生成与未参与建模的5个实际分布验证点相对应的5个“真实”不存在数据, 此过程重复10次, 之后通过计算混淆矩阵, 根据相关公式得到10套各评价指标的评估值。具体操作方法在此不做叙述, 可参考褚建民等(2017)文献。4种评价指标的优劣划分见表2。Table 2
表2
表24种评估指标(受试者工作特征曲线下面积、卡帕系数、真实技巧统计值、总体精度)的评价标准
Table 2
| 评估指标 Evaluate index | 失败 Fail | 很差 Very poor | 一般 Common | 较准确 Good | 优秀 Excellent | 本实验结果值(平均值±标准偏差) Result value of this experiment (mean ± SD) |
|---|---|---|---|---|---|---|
| AUC | 0.50-0.60 | 0.60-0.70 | 0.70-0.80 | 0.80-0.90 | 0.90-1.00 | 0.98 ± 0.02 |
| Kappa | 0-0.40 | 0.40-0.55 | 0.55-0.70 | 0.70-0.85 | 0.85-1.00 | 0.96 ± 0.08 |
| TSS | 0-0.40 | 0.40-0.55 | 0.55-0.70 | 0.70-0.85 | 0.85-1.00 | 0.96 ± 0.08 |
| Overall accuracy | 0-0.40 | 0-0.55 | 0.55-0.70 | 0.70-0.85 | 0.85-1.00 | 0.98 ± 0.04 |
新窗口打开|下载CSV
2 结果
2.1 影响海南凤仙花分布的主导环境因子
本研究运用AUC、Kappa系数、TSS和总体精度对模型预测精度的评估结果如表2所示。4个评估指标值均大于0.9, 表明模型的预测水平达到优秀, 模型能够作为海南凤仙花潜在适宜分布区的预测。根据Jackknife刀切法检验(图2), 较为重要的环境因子主要是坡度、最干季降水量、降水量季节性变异系数3个因子, 它们对模型的预测结果的增益较大, 也就是说这些环境因子对海南凤仙花的分布概率有着很大的影响。在研究选取的12个环境因子中, 贡献率排在前三的依次是坡度(58.9%)、最干季降水量(bio17, 27.5%)、降水量季节性变异系数(bio15, 12.1%), 累积贡献率达到98.5%, 这3个环境因子对模型预测性能起重要影响, 与刀切法检验结果一致。而其他环境因子的贡献率较小或者为0, 表明它们对模型的影响很小。综合来看, 影响海南凤仙花分布的主导环境因子为坡度、最干季降水量、降水量季节性变异系数这3个环境因素。此外本研究根据单因子反应曲线分析得到: 在坡度为50°以下时, 海南凤仙花分布概率随坡度的增加而增加, 且呈现先缓增后急增再缓增的趋势, 在坡度为16.5°-33.0°时增加最快, 当坡度为50°左右时趋于水平; 分布概率随最干季降水量的增加而减小, 在最干季降水量为23 mm左右时, 分布概率最大; 分布概率随着降水量季节性变异的增大而增加, 两者间存在正相关关系。
图2
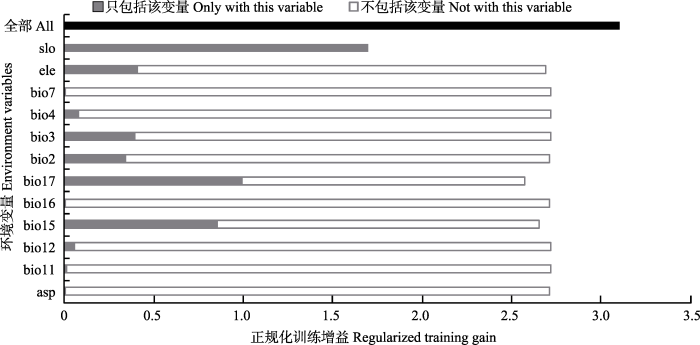 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2海南凤仙花环境因子刀切法检验结果。代码请参见
Fig. 2The jackknife test result of environmental factor for Impatiens hainanensis. See
2.2 海南凤仙花潜在适宜分布区
据图3可知, 海南岛内海南凤仙花的潜在适宜分布区当前主要集中分布于海南岛的西南部和南部地区。总的潜在适宜生境面积为2 871.6 km2, 占海南岛总面积的8.2%。整体的适宜生境分布狭窄, 且呈现严重的破碎化。其中最适宜生境面积为 639.0 km2, 占海南岛总面积的1.8%, 主要分布在白沙西部与南部、昌江中部和南部、东方东部、乐东北部, 包括白沙西北与昌江交界的付鹅岭、邦溪镇、金波乡、青松乡、南开乡及元门乡, 昌江县的七叉镇、王下乡、叉河镇及鸡心村, 东方的东河镇、天安乡及江边乡, 乐东东北部与东方市及昌江县交界的地区(佳西保护区), 此外, 乐东与五指山市交界处及乐东与三亚交界处也存在很小的最适宜区。次适宜生境面积为2 232.6 km2, 占海南岛总面积的6.4%, 分布在白沙县的邦溪镇、荣邦乡、金波乡、青松乡及南开乡, 昌江的保平乡、叉河镇、七叉镇及王下乡, 东方市的东河镇、天安乡、江边乡, 乐东的北部, 及其东部与五指山、三亚交界处, 此外, 五指山市西部地区、三亚北部及保亭西北部和南部有零星的区域可能适宜海南凤仙花生存。图3
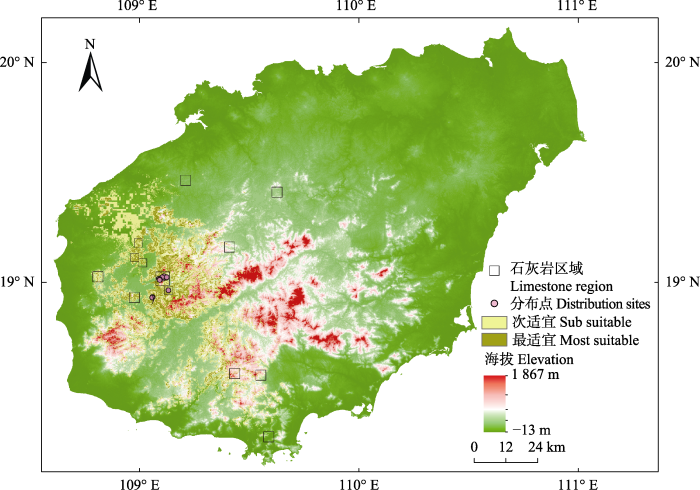 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3海南凤仙花在海南岛的潜在适宜生境分布。
Fig. 3The potential distribution of suitable habitat for Impatiens hainanensis in Hainan Island.
3 讨论
MaxEnt模型运用较少的分布点(8个)也很好地预测了海南凤仙花在当前时期的潜在适宜生境分布区。其最适宜生境主要分布在白沙西部与南部、昌江中部和南部、东方东部、乐东北部等地, 与现今已发现的石灰岩地区有着较好的一致性(图3)。海南凤仙花的潜在适宜生境呈现严重的破碎化, 分布范围极为狭窄, 且在多个地方仅零星分布。造成这一现象的原因可能有以下几点: 1)人为原因: 进入工业时代后人类活动对环境的干扰特别大, 开山采矿、采药等严重破坏了植物生长的适宜环境。2)自然原因: 海南凤仙花所处的石灰岩山地生态环境脆弱, 由于其特殊的碳酸盐岩物质组成, 地表渗水严重、保水性差、温度高, 土壤浅薄, 植物群落结构单一, 破坏后自我恢复能力极差等特点, 使海南凤仙花的生长更新存在一定困难(秦新生等, 2012)。同时外界环境气候变化造成的水分、营养胁迫等, 引起海南凤仙花的可塑性向消极反应, 导致其不适应或不利于适应环境, 从而只支持其在某些地方零星分布(Bennington & Mcgraw, 1996; 胡启鹏等, 2008)。3)自身原因: 因海南凤仙花本身属海岛石灰岩特有种, 形成了独特的传粉通道, 存在严重的传粉者限制, 使其自然结实率较低, 影响其繁殖与扩散(钟云芳, 2014)。3.1 模型环境因子
MaxEnt分析结果表明影响海南凤仙花分布的主导因子主要是坡度、最干季降水量、降水量季节性变异系数。其中坡度的贡献率高达58.9%, 降水量季节性变异系数及最干季降水量这2个与水分相关的因子累计贡献率达到39.6%, 说明海南凤仙花分布受限的最主要因素是地形因子(坡度)和水分因子。有研究发现, 在喀斯特山地森林即石灰岩山地, 坡度是所有地形因素中影响草本植物分布的最主要因子(袁铁象等, 2014)。坡度不仅影响山地地表水源及土壤养分的流向, 同时影响土壤厚度, 进而影响植物的分布格局。本研究中, 坡度是所有环境因子中影响海南凤仙花分布的最重要因素, 在50°的坡度范围内, 海南凤仙花都可以生长。据野外观察, 在低海拔的海南凤仙花种群生长于坡度较缓的环境, 而在中高海拔尤其是高海拔, 种群分布的坡度较陡, 说明海南凤仙花已适应石灰岩复杂多变的地形, 且在较陡的生境也可以生长。降水量季节性变异越大, 海南凤仙花的分布概率越大, 说明在不同季节间降水量差异越大的地区, 越可能适合海南凤仙花生存。由于特殊的地质二元结构和水分的快速漏失, 石灰岩地区植物的水分来源主要依赖于近期的降水(聂云鹏等, 2011; Deng et al., 2012), 而在热带季风气候的影响下, 海南凤仙花生境地区水分资源的季节性变化十分明显。海南凤仙花的繁殖特性与生境地这种水分的时空变异紧密相关, 海南凤仙花的花期集中在雨季, 可充分利用季节性的降水资源, 从而保障繁殖成功, 并演化出了相应的生理生态机制, 如其肉质且基部膨大的根茎可以贮藏大量的水分(钟云芳, 2014)。Lasky等(2016)的研究表明, 为了响应石灰岩这种干湿季变化, 植物通常是选择在雨季集中开花结实, 而本研究野外实地观察发现, 海南凤仙花的开花高峰期处于6-10月, 且10月结实的植株非常多, 本研究结果与其观点一致。本研究模型预测选取的环境因子因数据来源获取的原因, 只考虑了温度、水分等气候因子及地形因子, 未考虑土壤、物种适应性、生物竞争等其他因素, 存在一定的不足之处。土壤直接为植物提供生长发育所需的水分及营养, 对物种的分布结构有着明显的限制作用(高福元, 2012)。目前将土壤因素作为预测因子模拟物种分布的研究还较少。这可能是由于要获取一个研究区内连续分布的土壤属性较为困难, 尤其是对于大尺度的研究; 此外, 在研究过去和未来时期的物种分布时, 也没有能够相匹配的土壤属性, 因此限制了研究的进行。物种对环境的适应性并不是恒定的, 随着环境的改变, 物种表现出可塑性, 通过自然选择及遗传变异将这种可塑性转化为环境适应性(胡启鹏等, 2008)。而当前开展的研究都是假定物种与环境的关系是均衡的, 物种在新环境下仍保持原有的环境关系, 这将导致模型预测的偏差及不稳定性, 因此, 寻找合适的方法将物种的适应机制结合物种分布模型进行预测可以有效地提高结果的准确性(许仲林等, 2015)。
3.2 保护建议
经本课题组实地调查, 发现开山采矿及村民上山采药等人为活动, 不仅直接造成生态系统的失调, 也使海南凤仙花的适宜生境受到了严重的破坏, 尤其是皇帝洞的种群, 植株数量明显减少, 对海南凤仙花及其生境的保护已迫在眉睫。本研究结果表明霸王岭保护区、俄贤岭、皇帝洞及猕猴岭等地区的生境适宜性较高, 同时这几个地区也是海南凤仙花种群分布的核心区, 建议加强这些地方的保护力度, 尤其是昌江王下乡的皇帝洞至今未处于保护区内, 可建立自然保护区, 采取就地保护和迁地保护相结合的措施, 保护处于隔离破碎化状态的种群。东方天安乡、江边乡及乐东东北部(佳西保护区)等可能存在最适宜生境的地区, 作为今后野外深入调查的首选区域和重引入的重点区域。此外建议收集各种群海南凤仙花种子建立种质资源库同时加强其遗传多样性研究。由于受地理隔离的影响, 海南凤仙花不同种群间的基因交流极少, 可以考虑以人工授粉的方式实现不同种群间的基因交流, 促进其发展与进化。最后, 在今后开展的保护工作中, 应着重研究海南凤仙花的生理生态特征, 探讨其适应石灰岩的内在机制, 同时开展石灰岩地区的专项研究, 为更好地保护海南凤仙花及石灰岩特有植物提供科学 指导。
致谢
感谢海南霸王岭国家级自然保护区护林员赵莫强同志, 海南大学热带农林学院戚山江、刘宇、胡翔宇、黄蔚霞同学在野外过程中的协助。感谢中国热带农业科学院环境与植物保护研究所卢辉博士的指导与支持。感谢海南大学科研启动基金(kygd1625和kyqd1642)资助。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
DOIURL [本文引用: 1]

Bioclimatic models are widely used tools for assessing potential responses of species to climate change. One commonly used model is BIOCLIM, which summarises up to 35 climatic parameters throughout a species’ known range, and assesses the climatic suitability of habitat under current and future climate scenarios. A criticism of BIOCLIM is that the use of all 35 parameters may lead to over-fitting of the model, which in turn may result in misrepresentations of species’ potential ranges and to the loss of biological reality. In this study, we investigated how different methods of combining climatic parameters in BIOCLIM influenced predictions of the current distributions of 25 Australian butterflies species. Distributions were modeled using three previously used methods of selecting climatic parameters: (i) the full set of 35 parameters, (ii) a customised selection of the most relevant parameters for individual species based on analysing histograms produced by BIOCLIM, which show the values for each parameter at all of the focal species known locations, and (iii) a subset of 8 parameters that may generally influence the distributions of butterflies. We also modeled distributions based on random selections of parameters. Further, we assessed the extent to which parameter choice influenced predictions of the magnitude and direction of range changes under two climate change scenarios for 2020. We found that the size of predicted distributions was negatively correlated with the number of parameters incorporated in the model, with progressive addition of parameters resulting in progressively narrower potential distributions. There was also redundancy amongst some parameters; distributions produced using all 35 parameters were on average half the size of distributions produced using only 6 parameters. The selection of parameters via histogram analysis was influenced, to an extent, by the number of location records for the focal species. Further, species inhabiting different biogeographical zones may have different sets of climatic parameters limiting their distributions; hence, the appropriateness of applying the same subset of parameters to all species may be reduced under these situations. Under future climates, most species were predicted to suffer range reductions regardless of the scenario used and the method of parameter selection. Although the size of predicted distributions varied considerably depending on the method of selecting parameters, there were no significant differences in the proportional change in range size between the three methods: under the worst-case scenario, species’ distributions decrease by an average of 12.6, 11.4, and 15.7%, using all parameters, the ‘customised set’, and the ‘general set’ of parameters, respectively. However, depending on which method of selecting parameters was used, the direction of change was reversed for two species under the worst-case climate change scenario, and for six species under the best-case scenario (out of a total of 25 species). These results suggest that when averaged over multiple species, the proportional loss or gain of climatically suitable habitat is relatively insensitive to the number of parameters used to predict distributions with BIOCLIM. However, when measuring the response of specific species or the actual size of distributions, the number of parameters is likely to be critical.
[本文引用: 1]
DOIURL [本文引用: 2]
长柄扁桃(Amygdalus pedunculata)在我国分布于内蒙古和陕西,是一种濒危灌木,其资源现状只有零星的文献记录,这限制了对其资源数量和保护现状的评估。为此,本文通过野外调查来确立其自然分布区范围。我们选择了8个环境因子,运用基于规则集的遗传算法(genetic algorithm for rule-set prediction,GARP)模型和最大熵(maximum entropy,Max Ent)模型进行模拟,预测了长柄扁桃在中国的潜在适宜分布区;同时基于4个模型精度评估指标(Kappa、真实技巧统计法、总精度和受试者工作特征曲线下的面积)对模型进行验证,采用刀切法评估了预测变量的重要性。结果表明,两种模型均能准确预测长柄扁桃的地理分布规律,但Max Ent模型的4个预测精度指标都大于GARP模型。根据模型结果可判断长柄扁桃的适宜分布区以内蒙古中部地区为主,东起大兴安岭南部,向西可至贺兰山、乌兰布和沙漠以东,涵盖了毛乌素沙地、库布齐沙漠和浑善达克沙地,以及乌拉山、大青山等土石山区。其低适宜分布区可辐射至辽宁、河北、山西、陕西等省部分地区,另外在宁夏和甘肃中部地区也有零星的低适宜分布区。变量重要性分析结果表明,与降水相关的变量是决定长柄扁桃地理分布的重要环境因素。
DOIURL [本文引用: 2]
长柄扁桃(Amygdalus pedunculata)在我国分布于内蒙古和陕西,是一种濒危灌木,其资源现状只有零星的文献记录,这限制了对其资源数量和保护现状的评估。为此,本文通过野外调查来确立其自然分布区范围。我们选择了8个环境因子,运用基于规则集的遗传算法(genetic algorithm for rule-set prediction,GARP)模型和最大熵(maximum entropy,Max Ent)模型进行模拟,预测了长柄扁桃在中国的潜在适宜分布区;同时基于4个模型精度评估指标(Kappa、真实技巧统计法、总精度和受试者工作特征曲线下的面积)对模型进行验证,采用刀切法评估了预测变量的重要性。结果表明,两种模型均能准确预测长柄扁桃的地理分布规律,但Max Ent模型的4个预测精度指标都大于GARP模型。根据模型结果可判断长柄扁桃的适宜分布区以内蒙古中部地区为主,东起大兴安岭南部,向西可至贺兰山、乌兰布和沙漠以东,涵盖了毛乌素沙地、库布齐沙漠和浑善达克沙地,以及乌拉山、大青山等土石山区。其低适宜分布区可辐射至辽宁、河北、山西、陕西等省部分地区,另外在宁夏和甘肃中部地区也有零星的低适宜分布区。变量重要性分析结果表明,与降水相关的变量是决定长柄扁桃地理分布的重要环境因素。
URL [本文引用: 1]
Advancement in field ofGIS and Information Technology has taken conservation works and strategies astep further as most conservation works are now dependent on thesetechnologies. The present study explores the prediction ability of MAXENT usinga very low sample size by applying jackknife analysis over a well definedsmaller region and using only climate data. Vanda bicolor is a horticultureimportant orchid grown in certain patches of North Eastern region of India andthe species considered to be “Vulnerable”. Present study reports adistribution prediction model using different geo-climatic parameters for asmall area. Model validation by ground truthing gives a significant successful result which clearly defines the ability of MAXENTprediction model to give high success rate (71%) with low training samples. Useof the low sample size over a larger area results in unstable models howeverapplication of these samples in smaller radius around the occurrence pointscould provide good working models.
DOIURL [本文引用: 1]
It is not clear how the water sources are used by trees in typical karst areas of Guangxi, China. Trees growing on rigid carbonate rock can primarily use spring water, precipitation-recharged soil water or a mixture of the two. In this study, the Nongla karst dynamic monitoring station in the southwest of Guangxi, China, was observed. To determine the water sources accessed by trees growing on rigid carbonate rock, seasonal change of isotope ratios (delta O-18 and delta D) in twig sap, soil water, rainfall and spring water were measured. Stem water samples plotted to the right of the meteoric water line indicates utilization of water sources subject to evaporative isotopic enrichment. Trees growing on rigid carbonate rock in the southwest of China used rainfall directly in the rainy season. Adult trees tended to use deeper water stored in the epikarst zone, while the young ones used soil water when precipitation decreased. In the dry season, all trees sampled depend mostly on water stored in the epikarst zone. Spring water was the major water source for all of the tree species investigated in the dry season. In conclusion, these data indicate that trees growing on rigid carbonate rock in southwest China depend mostly on water stored in the epikarst zone. Therefore, water storage in the epikarst zone is the essential water source for trees growing on carbonate rock in southwest China.
DOIURL [本文引用: 1]
1. Few examples of habitat-modelling studies of rare and endangered species exist in the literature, although from a conservation perspective predicting their distribution would prove particularly useful. Paucity of data and lack of valid absences are the probable reasons for this shortcoming. Analytic solutions to accommodate the lack of absence include the ecological niche factor analysis (ENFA) and the use of generalized linear models (GLM) with simulated pseudo-absences. 2. In this study we tested a new approach to generating pseudo-absences, based on a preliminary ENFA habitat suitability (HS) map, for the endangered species Eryngium alpinum. This method of generating pseudo-absences was compared with two others: (i) use of a GLM with pseudo-absences generated totally at random, and (ii) use of an ENFA only. 3. The influence of two different spatial resolutions (i.e. grain) was also assessed for tackling the dilemma of quality (grain) vs. quantity (number of occurrences). Each combination of the three above-mentioned methods with the two grains generated a distinct HS map. 4. Four evaluation measures were used for comparing these HS maps: total deviance explained, best kappa, Gini coefficient and minimal predicted area (MPA). The last is a new evaluation criterion proposed in this study. 5. Results showed that (i) GLM models using ENFA-weighted pseudo-absence provide better results, except for the MPA value, and that (ii) quality (spatial resolution and locational accuracy) of the data appears to be more important than quantity (number of occurrences). Furthermore, the proposed MPA value is suggested as a useful measure of model evaluation when used to complement classical statistical measures. 6. Synthesis and applications. We suggest that the use of ENFA-weighted pseudo-absence is a possible way to enhance the quality of GLM-based potential distribution maps and that data quality (i.e. spatial resolution) prevails over quantity (i.e. number of data). Increased accuracy of potential distribution maps could help to define better suitable areas for species protection and reintroduction.
DOIURL [本文引用: 1]
Species Distribution Models (SDMs) could be an important tool to limit search efforts by selecting the areas where field surveys are to be carried out; due to the constant decrease of financial funds, this challenging purpose is particularly necessary. In particular, these methods are useful when applied to endangered and/or rare species with a poor known distribution area, especially due to difficulties in plant detection and in reaching the study areas. We hereby describe the development of maximum-entropy (Maxent) models for the endangered yellow gentian Gentiana lutea L. in Sardinia with the aims of (i) guiding survey efforts; (ii) estimating SDMs utility by post-test species current/extinct localities through the Observed Positive Predictive Power (OPPP) values; and (iii) evaluating the influence of sample data addition. Besides the Area Under Curve (AUC) values, we used the OPPP (observed/modelled positive localities ratio) to compare results from eight, 24 and 58 presence-only data points. Even with the initial small and biased sample data, we found that surveys could be effectively guided using such methods, whereby the focus of our research was on 48% of our initial 721km2 study area. The high OPPPs values additionally proved the reliability of our results in discovering 16 new localities of G. lutea. Nevertheless, the predictive models should be considered as a complementary tool rather than a replacement for expert knowledge.
DOIURL [本文引用: 1]
Global conservation of plant biodiversity on tropical islands is a major priority, as approximately one third of all endangered plant species are insular endemics. Checklists can be an important first step in determining conservation priorities on islands. Hainan, the largest island in the Indo-Burma Biodiversity Hotspot, and therefore an international focus for conservation, has the most extensive and best preserved tropical forests in China. In this study we enumerate the endemic seed plants of Hainan Island. The checklist was prepared by consulting: (1) several bibliographic/taxonomic data base resources, (2) relevant taxonomic treatments and floras, and (3) plant taxonomists who are actively working with Chinese plants. The checklist also contains information concerning conservation status, the occurrence of Hainan endemics in four protected areas on the island, and available molecular phylogenies. An additional checklist of the species that were until recently thought to be endemic to Hainan, but are no longer considered to be, is also presented. In a separate paper in this issue of Botanical Review the patterns of endemism on Hainan Island are discussed.
URL [本文引用: 1]
在全球变化影响下植物分布格局呈现多元化趋势,繁殖力、适应力强的入侵种容易形成组成简单的单一优势种群,其中在干旱半干旱区,单一优势种群群落已成为植被退化的主要形式之一,这种单一优势种群斑块可能会改变小尺度范围内土壤水分分布格局,从而优化植物种群对环境响应的分布区扩散机制。因此,单一优势种群格局与土壤水分关系的研究,对于揭示植物应对土壤水分变化的可塑性机制,深入理解植物与环境之间的协同关系具有重要意义。祁连山北坡高寒山区特殊的地理环境及气候条件,加之气候变迁和家畜过度放牧干扰下,甘肃臭草凭借自身生物学特征、可塑性以...
URL [本文引用: 1]
在全球变化影响下植物分布格局呈现多元化趋势,繁殖力、适应力强的入侵种容易形成组成简单的单一优势种群,其中在干旱半干旱区,单一优势种群群落已成为植被退化的主要形式之一,这种单一优势种群斑块可能会改变小尺度范围内土壤水分分布格局,从而优化植物种群对环境响应的分布区扩散机制。因此,单一优势种群格局与土壤水分关系的研究,对于揭示植物应对土壤水分变化的可塑性机制,深入理解植物与环境之间的协同关系具有重要意义。祁连山北坡高寒山区特殊的地理环境及气候条件,加之气候变迁和家畜过度放牧干扰下,甘肃臭草凭借自身生物学特征、可塑性以...
DOIURL [本文引用: 2]
Species distribution models should provide conservation practioners with estimates of the spatial distributions of species requiring attention. These species are often rare and have limited known occurrences, posing challenges for creating accurate species distribution models. We tested four modeling methods (Bioclim, Domain, GARP, and Maxent) across 18 species with different levels of ecological specialization using six different sample size treatments and three different evaluation measures. Our assessment revealed that Maxent was the most capable of the four modeling methods in producing useful results with sample sizes as small as 5, 10 and 25 occurrences. The other methods compensated reasonably well (Domain and GARP) to poorly (Bioclim) when presented with datasets of small sample sizes. We show that multiple evaluation measures are necessary to determine accuracy of models produced with presence-only data. Further, we found that accuracy of models is greater for species with small geographic ranges and limited environmental tolerance, ecological characteristics of many rare species. Our results indicate that reasonable models can be made for some rare species, a result that should encourage conservationists to add distribution modeling to their toolbox.
DOIURL [本文引用: 1]
DOIURL [本文引用: 2]
表型可塑性是植物与环境相互作用而表现出的一种适应能力,表现为积极的响应和消极的响应。可塑性不仅可以作为性状遗传,也可接受进化,但适应进化的前提条件是遗传变异和自然选择,同时受可塑性代价和限制条件所局限,这些问题可通过先进的生物手段逐渐找到答案,并得到解决。其次,植物可塑性响应还受到非生物因子的影响,如光照(耐荫性)、营养、水分以及温度等,特别在全球大气变化的影响下,植物的可塑性响应能否反映这种变化值得去关注。同时,国内的表型可塑性研究还需要进一步深入,特别是在大区域尺度上,更多的评价工作需要完善。
DOIURL [本文引用: 2]
表型可塑性是植物与环境相互作用而表现出的一种适应能力,表现为积极的响应和消极的响应。可塑性不仅可以作为性状遗传,也可接受进化,但适应进化的前提条件是遗传变异和自然选择,同时受可塑性代价和限制条件所局限,这些问题可通过先进的生物手段逐渐找到答案,并得到解决。其次,植物可塑性响应还受到非生物因子的影响,如光照(耐荫性)、营养、水分以及温度等,特别在全球大气变化的影响下,植物的可塑性响应能否反映这种变化值得去关注。同时,国内的表型可塑性研究还需要进一步深入,特别是在大区域尺度上,更多的评价工作需要完善。
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
全缘冬青(Ilex integra)为岛屿特有植物,主要分布于浙江舟山群岛,被列为浙江省珍稀濒危植物.作者运用ISSR分子标记对舟山群岛的6个全缘冬青种群(5个自然 种群和1个栽培种群)共57个个体进行了遗传多样性分析.采用9条随机引物扩增出78条清晰谱带,其中45条为多态性条带,多态位点百分比(PPL)为 57.7%.经POPGENE软件分析,全缘冬青种群平均多态位点百分比(PPL)为42.1%,Nei's基因多样度(HE)为0.153,较其他海岛 植物的遗传多样性偏低.5个自然种群遗传分化系数Gst=0.316,AMOVA测度Fst=0.295,Shannon多样性分化系数S=0.304, 遗传分化程度较高.全缘冬青种群间地理距离与遗传距离具有正相关性(r=0.649,P<0.05),岛屿间地理隔离对种群间遗传分化产生了较为显著的影 响.UPGMA聚类分析显示,朱家尖岛与普陀岛种群亲缘关系较近,而舟山岛栽培种群是由桃花岛自然种群移植而来.针对全缘冬青种群遗传多样性较低及遗传分 化程度较高的现状,应加强现有自然种群的就地保护,促进种群自然更新;建立种质资源库,收集不同岛屿的种源进行混合繁殖,促进基因交流.
DOIURL [本文引用: 1]
全缘冬青(Ilex integra)为岛屿特有植物,主要分布于浙江舟山群岛,被列为浙江省珍稀濒危植物.作者运用ISSR分子标记对舟山群岛的6个全缘冬青种群(5个自然 种群和1个栽培种群)共57个个体进行了遗传多样性分析.采用9条随机引物扩增出78条清晰谱带,其中45条为多态性条带,多态位点百分比(PPL)为 57.7%.经POPGENE软件分析,全缘冬青种群平均多态位点百分比(PPL)为42.1%,Nei's基因多样度(HE)为0.153,较其他海岛 植物的遗传多样性偏低.5个自然种群遗传分化系数Gst=0.316,AMOVA测度Fst=0.295,Shannon多样性分化系数S=0.304, 遗传分化程度较高.全缘冬青种群间地理距离与遗传距离具有正相关性(r=0.649,P<0.05),岛屿间地理隔离对种群间遗传分化产生了较为显著的影 响.UPGMA聚类分析显示,朱家尖岛与普陀岛种群亲缘关系较近,而舟山岛栽培种群是由桃花岛自然种群移植而来.针对全缘冬青种群遗传多样性较低及遗传分 化程度较高的现状,应加强现有自然种群的就地保护,促进种群自然更新;建立种质资源库,收集不同岛屿的种源进行混合繁殖,促进基因交流.
DOIURL [本文引用: 1]
【目的】青钱柳是集药用、材用和观赏价值于一身的多用途树种。通过应用贡献率、置换重要值比较以及Jackknife检验等综合分析方法,评价制约青钱柳现代地理分布的主要因子,可为青钱柳资源的科学保护和合理利用提供理论依据。【方法】基于183个青钱柳地理分布记录和8个气候环境因子数据,采用MaxEnt模型软件对青钱柳潜在适宜分布区进行预测,并在此基础上划分适宜栽培区。【结果】采用MaxEnt模型预测青钱柳潜在适宜分布区准确度极高,受试者工作特征曲线下的面积(AUC值)达0.964±0.006。目前,青钱柳高度适宜分布区为浙江南部、福建西北部、安徽南部、湖北东部、江西东部和西部、湖南东部和西部、贵州东部、重庆东部、陕西南部和四川东北部;随着未来气温的升高,青钱柳有向高纬度和高海拔地区迁移的趋势。【结论】影响青钱柳分布的主要气候因子是温度,包括年均温、季节温度变异系数和平均日温差,其中年均温为最主要因子。
DOIURL [本文引用: 1]
【目的】青钱柳是集药用、材用和观赏价值于一身的多用途树种。通过应用贡献率、置换重要值比较以及Jackknife检验等综合分析方法,评价制约青钱柳现代地理分布的主要因子,可为青钱柳资源的科学保护和合理利用提供理论依据。【方法】基于183个青钱柳地理分布记录和8个气候环境因子数据,采用MaxEnt模型软件对青钱柳潜在适宜分布区进行预测,并在此基础上划分适宜栽培区。【结果】采用MaxEnt模型预测青钱柳潜在适宜分布区准确度极高,受试者工作特征曲线下的面积(AUC值)达0.964±0.006。目前,青钱柳高度适宜分布区为浙江南部、福建西北部、安徽南部、湖北东部、江西东部和西部、湖南东部和西部、贵州东部、重庆东部、陕西南部和四川东北部;随着未来气温的升高,青钱柳有向高纬度和高海拔地区迁移的趋势。【结论】影响青钱柳分布的主要气候因子是温度,包括年均温、季节温度变异系数和平均日温差,其中年均温为最主要因子。
DOIURL [本文引用: 1]
为模拟、预测气候变化对孑遗、濒危植物蒙古扁桃(Amygdalus mongolica)潜在分布的影响,利用最大熵(MAXENT)模型模拟、预测、对比、分析、揭示蒙古扁桃在最大冰期(CCSM及MIROC模型)、历史气候(1961-1990年)及未来气候(2020年、2050年-12080年,政府间气候变化专门委员会排放情景特别报告的A2A情景)条件下的适宜分布范围和空间格局的变化。结果表明:(1)蒙古扁桃在历史气候条件下的潜在分布区集中在蒙古的南戈壁省及东戈壁省,我国内蒙古巴彦淖尔市、阿拉善左旗、鄂尔多斯市、锡林郭勒盟西部,河西走廊中部及东部,宁夏北部及陕西北部,以及河北北部的部分地区;(2)与历史气候条件下的潜在分布相比,蒙古扁桃在最大冰期CCSM气候情景下的分布经历了明显的、大范围的向南迁移和范围缩小;(3)未来A2A气候情景下,其潜在分布范围表现出在2020年明显扩大,在2050年减小,到2080年又略有增大的趋势。分布格局表现出不断向我国河北及内蒙古东部,蒙古东部、北部及西部大幅度扩散、迁移的趋势。
DOIURL [本文引用: 1]
为模拟、预测气候变化对孑遗、濒危植物蒙古扁桃(Amygdalus mongolica)潜在分布的影响,利用最大熵(MAXENT)模型模拟、预测、对比、分析、揭示蒙古扁桃在最大冰期(CCSM及MIROC模型)、历史气候(1961-1990年)及未来气候(2020年、2050年-12080年,政府间气候变化专门委员会排放情景特别报告的A2A情景)条件下的适宜分布范围和空间格局的变化。结果表明:(1)蒙古扁桃在历史气候条件下的潜在分布区集中在蒙古的南戈壁省及东戈壁省,我国内蒙古巴彦淖尔市、阿拉善左旗、鄂尔多斯市、锡林郭勒盟西部,河西走廊中部及东部,宁夏北部及陕西北部,以及河北北部的部分地区;(2)与历史气候条件下的潜在分布相比,蒙古扁桃在最大冰期CCSM气候情景下的分布经历了明显的、大范围的向南迁移和范围缩小;(3)未来A2A气候情景下,其潜在分布范围表现出在2020年明显扩大,在2050年减小,到2080年又略有增大的趋势。分布格局表现出不断向我国河北及内蒙古东部,蒙古东部、北部及西部大幅度扩散、迁移的趋势。
[本文引用: 1]
DOIURL [本文引用: 1]
The MaxEnt software package is one of the most popular tools for species distribution and environmental niche modeling, with over 1000 published applications since 2006. Its popularity is likely for two reasons: 1) MaxEnt typically outperforms other methods based on predictive accuracy and 2) the software is particularly easy to use. MaxEnt users must make a number of decisions about how they should select their input data and choose from a wide variety of settings in the software package to build models from these data. The underlying basis for making these decisions is unclear in many studies, and default settings are apparently chosen, even though alternative settings are often more appropriate. In this paper, we provide a detailed explanation of how MaxEnt works and a prospectus on modeling options to enable users to make informed decisions when preparing data, choosing settings and interpreting output. We explain how the choice of background samples reflects prior assumptions, how nonlinear functions of environmental variables (features) are created and selected, how to account for environmentally biased sampling, the interpretation of the various types of model output and the challenges for model evaluation. We demonstrate MaxEnt090005s calculations using both simplified simulated data and occurrence data from South Africa on species of the flowering plant family Proteaceae. Throughout, we show how MaxEnt090005s outputs vary in response to different settings to highlight the need for making biologically motivated modeling decisions.
DOIURL [本文引用: 1]
以桂西北石灰岩地区连片出露石丛生境次生林群落为研究对象,运用稳定性氢氧同位素技术结合 IsoSource模型,分析了5种典型植物f半落叶乔木粉苹婆(Sterculiaeuosma)、落叶乔木菜豆树 (Radermacherasinica)、常绿灌木鹅掌柴(Scheffleraoctophylla)以及落叶灌木红背山麻杆(Alchornea trewioides)和紫弹树(Ceresbiondii))水分来源的季节性差异。结果表明:雨季,除鹅掌柴同时利用部分前期雨水外,其余4种植物均 主要利用最近15天内的雨水(利用比例的平均值之和均超过80%)。旱季,5种植物均主要利用最近一次雨水与前期雨水(一个月前)的混合(利用比例的平均 值之和均超过80%),其中乔木和常绿灌木对前期雨水的利用比例更高(利用比例的平均值均超过50%)。植被良好的石灰岩连片出露石丛生境中,植物主要以 不同时期的雨水为主要水源,这可能与石灰岩发育的裂隙及其储水能力有关。储存在裂隙中的雨水通过植物蒸腾的方式返回大气,这一良好的水文循环过程应得到充 分的保护。
DOIURL [本文引用: 1]
以桂西北石灰岩地区连片出露石丛生境次生林群落为研究对象,运用稳定性氢氧同位素技术结合 IsoSource模型,分析了5种典型植物f半落叶乔木粉苹婆(Sterculiaeuosma)、落叶乔木菜豆树 (Radermacherasinica)、常绿灌木鹅掌柴(Scheffleraoctophylla)以及落叶灌木红背山麻杆(Alchornea trewioides)和紫弹树(Ceresbiondii))水分来源的季节性差异。结果表明:雨季,除鹅掌柴同时利用部分前期雨水外,其余4种植物均 主要利用最近15天内的雨水(利用比例的平均值之和均超过80%)。旱季,5种植物均主要利用最近一次雨水与前期雨水(一个月前)的混合(利用比例的平均 值之和均超过80%),其中乔木和常绿灌木对前期雨水的利用比例更高(利用比例的平均值均超过50%)。植被良好的石灰岩连片出露石丛生境中,植物主要以 不同时期的雨水为主要水源,这可能与石灰岩发育的裂隙及其储水能力有关。储存在裂隙中的雨水通过植物蒸腾的方式返回大气,这一良好的水文循环过程应得到充 分的保护。
DOIURL [本文引用: 1]
Floristic and ecological characteristics of the community with .The dominant families were Orchidaceae(including 16 species), Polypodiaceae(5 species), and Euphorbiaceae(4 species). The distribution types of the genera in the community were mainly Pantropic(including 18 species) and Tropic Asia (India-Malasia) (15 species). The shrub layer(including 29 species) and herb layer(45 species) were the main component of the community. There were 9 species in the liana layer. The dominant species of shrub layer, herb layer, and liana layer were Euphorbia hainanensis, Eria rosea, and Dischidia chinensis, respectively. The life form in the community was dominated by nanophanerophytes(including 57 species).The order of the frequency distribution class was A>B>D>C>E. The A-level frequency was dominant(including 41 species). The community characteristics between I. hainanensis and Eu. hainanensis were compared. Some preliminary suggestions were also put forward on how to effectively protect I. hainanensis.
DOIURL [本文引用: 1]
Floristic and ecological characteristics of the community with .The dominant families were Orchidaceae(including 16 species), Polypodiaceae(5 species), and Euphorbiaceae(4 species). The distribution types of the genera in the community were mainly Pantropic(including 18 species) and Tropic Asia (India-Malasia) (15 species). The shrub layer(including 29 species) and herb layer(45 species) were the main component of the community. There were 9 species in the liana layer. The dominant species of shrub layer, herb layer, and liana layer were Euphorbia hainanensis, Eria rosea, and Dischidia chinensis, respectively. The life form in the community was dominated by nanophanerophytes(including 57 species).The order of the frequency distribution class was A>B>D>C>E. The A-level frequency was dominant(including 41 species). The community characteristics between I. hainanensis and Eu. hainanensis were compared. Some preliminary suggestions were also put forward on how to effectively protect I. hainanensis.
DOIURL [本文引用: 1]
通过对海南岛石灰岩地区植物区系和植被的调查研究表明,海南岛石灰岩地区共有野生分布的国家及省级珍稀濒危保护植物26种,隶属19科,24属,其中蕨类植物4种,裸子植物3种,被子植物19种。并建议将84种海南岛石灰岩地区的珍稀濒危植物列入国家或省级保护植物名录。本文在分析海南岛石灰岩地区珍稀濒危植物种类组成、分布、现状评估、濒危原因的基础上,提出了海南岛石灰岩地区珍稀濒危植物保护对策。
DOIURL [本文引用: 1]
通过对海南岛石灰岩地区植物区系和植被的调查研究表明,海南岛石灰岩地区共有野生分布的国家及省级珍稀濒危保护植物26种,隶属19科,24属,其中蕨类植物4种,裸子植物3种,被子植物19种。并建议将84种海南岛石灰岩地区的珍稀濒危植物列入国家或省级保护植物名录。本文在分析海南岛石灰岩地区珍稀濒危植物种类组成、分布、现状评估、濒危原因的基础上,提出了海南岛石灰岩地区珍稀濒危植物保护对策。
DOIURL [本文引用: 1]
【Objective】The purpose of this study was to reveal the floristic composition and characteristics of seed plants in limestone areas in Hainan.【Method】Extensive field work, specimens collection and identification were conducted to collect data for the analysis.【Result and conclusion】A total of 1 176 species, 58 varieties and 1 forma belonging to 645 genera and 142 families were recorded. The tropical and subtropical distribution elements, especially tropical Asia ones, were dominant in the limestone flora in Hainan. The tropical and subtropical distribution families accounted for 73.24% of the total and the genera accounted for 87.75%. The proportions of temperate distribution families and genera accounted for 11.97% and 7.29%, respectively. The limestone areas contributed about 27.26% to the total number of species with less than 2% land area of the whole island. However, many genera include only one or very few species. There are 113 single and few species families, accounting for 79.58% of the total families. There are 2 genera endemic to China.
DOIURL [本文引用: 1]
【Objective】The purpose of this study was to reveal the floristic composition and characteristics of seed plants in limestone areas in Hainan.【Method】Extensive field work, specimens collection and identification were conducted to collect data for the analysis.【Result and conclusion】A total of 1 176 species, 58 varieties and 1 forma belonging to 645 genera and 142 families were recorded. The tropical and subtropical distribution elements, especially tropical Asia ones, were dominant in the limestone flora in Hainan. The tropical and subtropical distribution families accounted for 73.24% of the total and the genera accounted for 87.75%. The proportions of temperate distribution families and genera accounted for 11.97% and 7.29%, respectively. The limestone areas contributed about 27.26% to the total number of species with less than 2% land area of the whole island. However, many genera include only one or very few species. There are 113 single and few species families, accounting for 79.58% of the total families. There are 2 genera endemic to China.
DOIURL [本文引用: 1]
1. The management of both desirable and undesirable species requires an understanding of the factors determining their distribution. Quantitative distribution models offer simple methods for formulating the species-habitat link and the means not only for predicting where species should occur, but also for understanding the factors involved. Generalized linear modelling, in particular, links the incidence of species to habitat variables, and has increasingly formed the backbone of the modelling approaches used. New 'data technologies', such as remote sensing and geographical information systems, have further broadened these modelling applications to almost any ecological system and any species for which there are distribution data. 2. Many previous approaches have aimed to identify the most parsimonious model with the best suite of predictors, selected on the basis of null hypothesis testing. However, information-theoretic approaches based on Akaike's information criterion allow the selection of a best approximating model or a subset of models from a set of candidates. Information-theoretic approaches require a deeper understanding of the biology of the system modelled and may well become an improved paradigm for species distribution modelling. 3. Synthesis and applications. This special profile of six papers demonstrates the development in methodology used in species distribution modelling. The papers show how information-theoretic approaches can be coupled with emerging data technologies to address issues of conservation significance. With conservation biology and applied ecology at the forefront of many of the basic science developments so far, we expect these methods to pervade other areas of ecological research more fully in future.
URL [本文引用: 1]
基于上世纪80-90年代和少量本世纪初野外调查获得的我国兴安落叶松地理分布数据、以及通过区域气候模式(RegCM3)模拟的气候数据拟合出的12个主要气候因子,利用最大熵(MAXENT)模型和ARCGIS软件的空间分析技术,模拟预估并分析了本世纪内A1B气候变化情景下兴安落叶松的气候适生性及其潜在地理分布的变化趋势。这对编制近期环境保育、区域生态规划和管理工作提供了科学依据与建议,并对制定中、长期生态规划给出了有益参考。 在获得未来气候变化情境下兴安落叶松潜在地理分布的预估结果后,采用土地利用数据过滤了兴安落叶松不可能适生的土地类型,并根据IPCC关于评估“可能性”表述的划分标准划分出四个适生等级,进一步完善和精确了兴安落叶松的潜在适生区。研究的主要结果显示: (1)应用MAXENT模型模拟预测出的“当前”适生区分布与实地调查获得的分布基本一致,模型可用于研究兴安落叶松未来潜在分布与气候变化的关系。 (2)MAXENT模型比较准确地捕捉到了影响兴安落叶松潜在分布的主导生物-气候因子,按照其对模拟的贡献率从大到小排序依次为:春季最高温度、全年平均温度、春季降水量和五月份降水量,其总累计贡献率超过96%。 (3)气温的持续攀升和降水量的缓慢增加,将导致我国兴安落叶松原有的适生区大幅度减少,潜在适生区边界逐渐北移和稍微的东移,北界逐渐与境外的分布区分离;并围绕大兴安岭地区潜在适生中心区域收缩,形成孤立的小林区;而小兴安岭地区和长白山地区的潜在适生区也将逐渐缩小并在世纪末完全消失。
URL [本文引用: 1]
基于上世纪80-90年代和少量本世纪初野外调查获得的我国兴安落叶松地理分布数据、以及通过区域气候模式(RegCM3)模拟的气候数据拟合出的12个主要气候因子,利用最大熵(MAXENT)模型和ARCGIS软件的空间分析技术,模拟预估并分析了本世纪内A1B气候变化情景下兴安落叶松的气候适生性及其潜在地理分布的变化趋势。这对编制近期环境保育、区域生态规划和管理工作提供了科学依据与建议,并对制定中、长期生态规划给出了有益参考。 在获得未来气候变化情境下兴安落叶松潜在地理分布的预估结果后,采用土地利用数据过滤了兴安落叶松不可能适生的土地类型,并根据IPCC关于评估“可能性”表述的划分标准划分出四个适生等级,进一步完善和精确了兴安落叶松的潜在适生区。研究的主要结果显示: (1)应用MAXENT模型模拟预测出的“当前”适生区分布与实地调查获得的分布基本一致,模型可用于研究兴安落叶松未来潜在分布与气候变化的关系。 (2)MAXENT模型比较准确地捕捉到了影响兴安落叶松潜在分布的主导生物-气候因子,按照其对模拟的贡献率从大到小排序依次为:春季最高温度、全年平均温度、春季降水量和五月份降水量,其总累计贡献率超过96%。 (3)气温的持续攀升和降水量的缓慢增加,将导致我国兴安落叶松原有的适生区大幅度减少,潜在适生区边界逐渐北移和稍微的东移,北界逐渐与境外的分布区分离;并围绕大兴安岭地区潜在适生中心区域收缩,形成孤立的小林区;而小兴安岭地区和长白山地区的潜在适生区也将逐渐缩小并在世纪末完全消失。
DOIURL [本文引用: 1]
石灰岩裸露地区普遍具有土体浅薄,保水性差,基质、土壤和水等环境富钙的生态特征,这些特征促进了石灰岩地区植物区系朝着特有化发展.进一步深化石灰岩地 区土壤形成与保护理论研究,大力开展特有植物种的生态及经济性能综合评价,加强造林先锋树种的选育,为人工促进石灰岩地区森林生态系统恢复提供物质保障是 近期研究的发展趋势.
DOIURL [本文引用: 1]
石灰岩裸露地区普遍具有土体浅薄,保水性差,基质、土壤和水等环境富钙的生态特征,这些特征促进了石灰岩地区植物区系朝着特有化发展.进一步深化石灰岩地 区土壤形成与保护理论研究,大力开展特有植物种的生态及经济性能综合评价,加强造林先锋树种的选育,为人工促进石灰岩地区森林生态系统恢复提供物质保障是 近期研究的发展趋势.
DOIURL [本文引用: 1]
分析气候变化对植物分布的影响,对保护生物多样性具有重要意义。利用CART(分类和回归树)模型及A2和B2情景,分析了气候变化对短叶假木贼(Anabasis brevifolia)、裸果木(Gymnocarpos przewalskii)、梭梭(Haloxylon ammoden-dron)、膜果麻黄(Ephedra przewalskii)、驼绒藜(Ceratoides latens)和喀什膜果麻黄(Ephedra przewalskii var.kaschgarica)分布范围及空间格局的影响。结果表明:气候变化下,这些植物目前适宜分布范围减小;从新适宜及总适宜分布范围而言,短叶假木贼和梭梭从1991-2020年到2051-2080年时段增加,之后减小,其它植物从1991-2020年到2081-2100年时段减小;喀什膜果麻黄和驼绒藜适宜分布范围减小并破碎化,其它植物向目前适宜分布的西部、西北部(或青海西南部)、昆仑山、阿尔金山和祁连山区扩展;除驼绒藜和喀什膜果麻黄与年均气温变化具显著相关性外,其它植物分布范围与年均气温和降水量变化的相关性较弱(P〉0.05),除驼绒藜、喀什膜果麻黄和裸果木目前分布范围与年均气温和降水量变化的回归关系较强外,其它植物分布范围与年均气温和年降水量变化多元线性回归关系较弱。上述研究结果表明,气候变化下,这些植物空间分布格局改变,目前分布范围减少,新适宜及总适宜分布范围近期增加,随着气候变化程度的增强,又逐渐减小。
DOIURL [本文引用: 1]
分析气候变化对植物分布的影响,对保护生物多样性具有重要意义。利用CART(分类和回归树)模型及A2和B2情景,分析了气候变化对短叶假木贼(Anabasis brevifolia)、裸果木(Gymnocarpos przewalskii)、梭梭(Haloxylon ammoden-dron)、膜果麻黄(Ephedra przewalskii)、驼绒藜(Ceratoides latens)和喀什膜果麻黄(Ephedra przewalskii var.kaschgarica)分布范围及空间格局的影响。结果表明:气候变化下,这些植物目前适宜分布范围减小;从新适宜及总适宜分布范围而言,短叶假木贼和梭梭从1991-2020年到2051-2080年时段增加,之后减小,其它植物从1991-2020年到2081-2100年时段减小;喀什膜果麻黄和驼绒藜适宜分布范围减小并破碎化,其它植物向目前适宜分布的西部、西北部(或青海西南部)、昆仑山、阿尔金山和祁连山区扩展;除驼绒藜和喀什膜果麻黄与年均气温变化具显著相关性外,其它植物分布范围与年均气温和降水量变化的相关性较弱(P〉0.05),除驼绒藜、喀什膜果麻黄和裸果木目前分布范围与年均气温和降水量变化的回归关系较强外,其它植物分布范围与年均气温和年降水量变化多元线性回归关系较弱。上述研究结果表明,气候变化下,这些植物空间分布格局改变,目前分布范围减少,新适宜及总适宜分布范围近期增加,随着气候变化程度的增强,又逐渐减小。
DOIURL [本文引用: 1]
物种分布模型已被广泛地应用于以保护区规划、气候变化对物种分布的影响等为目的的研究。回顾了已经得到广泛应用的多种物种分布模型,总结了评价模型性能的方法。基于物种分布模型的发展和应用以及性能评价中尚存在的问题,本文认为:在物种分布模型中集成样本选择模块能够避免模型预测过程中的过度拟合及欠拟合,增加变量选择模块可评估和降低变量之间自相关性的影响,增加生物因子以及将物种对环境的适应性机制(及扩散行为特征)和潜在分布模型进行结合,是提高模型预测性能的可行方案;在模型性能的评价方面,采用赤池信息量可对模型的预测性能进行客观评价。相关建议可为物种分布建模提供参考。;Species distribution models (SDMs) have been used in various applications, such as conservation planning, determining the impact of climate change on species distribution, and others. SDMs allow construction of the correlation relationship between occurrence of a target species and environmental conditions (including bioclimatic and anthropogenic conditions). The correlation relationship can then be applied to the entire environmental space to predict the potential distribution of a target species. In the present study, we first review widely-used SDMs and summarize their evaluation approaches. Generally, SDMs can be classified into two categories according to the data required for construction of the correlation relationship, i.e., SDMs that predict the potential distribution of species based on presence-only records (PO models), and SDMs that use presence-absence records (PA models). If reliable absence records are available, PA models are suggested. Additional classification of SDMs is based on output format, namely, SDMs that give prediction results in the format of continuous probabilities (the higher the probability, the more suitable for distribution), and those with results in the format of binary values (1 for suitable and 0 for unsuitable). According to the various SDM output formats, SDM performance can be evaluated by threshold-independent (for models with continuous probabilities) and threshold-dependent (for models with binary prediction) strategies. Threshold-independent strategies can be realized by calculating values of maximum overall accuracy, maximum kappa, maximum vertical distance, area under the receiver operating characteristic (ROC) Curve (AUC), Gini index, point biserial correlation coefficient, mean square error, root mean square error, coefficient of determination, mean absolute prediction error, and others. Threshold-dependent strategies can be realized by calculating values of sensitivity, specificity, positive predictive value, negative predictive value, positive likelihood ratio, negative likelihood ratio, true skill statistic, odds ratio, Yule's Y, Yule's Q, Phi coefficient, Kappa, normalized mutual information, extreme dependency score, and others. Based on existing problems related to the development, application and evaluation of SDMs, the present study suggests the following. (1) Because biased presence samples can influence the result of prediction, integrating a sample selection module in the SDM could improve the reliability of model prediction. (2) Bioclimatic variables (such as WorldClim) that are calculated from a Digital Elevation Model (DEM) may co-linearly correlate with each other, and such collinearity may result in overfitting when modeling the potential distribution of a species. As a result, selecting variables based on calculation of the Variance Inflation Factor (VIF) is a suitable means to avoid overfitting. (3) In addition to abiotic factors, biotic factors are also important determinants for species distribution. Thus, the use of biotic variables could improve the model results, although biotic factors are not easy to delineate within a geographic space. (4) The spatial and temporal extrapolation of SDMs, which deal with problems of species potential distribution at different geographic ranges and time points (past and/or future), respectively, are actually based on the assumption of an equilibrium relationship between the target species and environmental conditions. However, this assumption is challenged, because species have the abilities of adaption and dispersal. (5) The Partial AUC (PAUC) is suitable for evaluation of single model performance, and the Akaike Information Criterion (AIC) could provide an objective evaluation of the performance of several SDMs.
DOIURL [本文引用: 1]
物种分布模型已被广泛地应用于以保护区规划、气候变化对物种分布的影响等为目的的研究。回顾了已经得到广泛应用的多种物种分布模型,总结了评价模型性能的方法。基于物种分布模型的发展和应用以及性能评价中尚存在的问题,本文认为:在物种分布模型中集成样本选择模块能够避免模型预测过程中的过度拟合及欠拟合,增加变量选择模块可评估和降低变量之间自相关性的影响,增加生物因子以及将物种对环境的适应性机制(及扩散行为特征)和潜在分布模型进行结合,是提高模型预测性能的可行方案;在模型性能的评价方面,采用赤池信息量可对模型的预测性能进行客观评价。相关建议可为物种分布建模提供参考。;Species distribution models (SDMs) have been used in various applications, such as conservation planning, determining the impact of climate change on species distribution, and others. SDMs allow construction of the correlation relationship between occurrence of a target species and environmental conditions (including bioclimatic and anthropogenic conditions). The correlation relationship can then be applied to the entire environmental space to predict the potential distribution of a target species. In the present study, we first review widely-used SDMs and summarize their evaluation approaches. Generally, SDMs can be classified into two categories according to the data required for construction of the correlation relationship, i.e., SDMs that predict the potential distribution of species based on presence-only records (PO models), and SDMs that use presence-absence records (PA models). If reliable absence records are available, PA models are suggested. Additional classification of SDMs is based on output format, namely, SDMs that give prediction results in the format of continuous probabilities (the higher the probability, the more suitable for distribution), and those with results in the format of binary values (1 for suitable and 0 for unsuitable). According to the various SDM output formats, SDM performance can be evaluated by threshold-independent (for models with continuous probabilities) and threshold-dependent (for models with binary prediction) strategies. Threshold-independent strategies can be realized by calculating values of maximum overall accuracy, maximum kappa, maximum vertical distance, area under the receiver operating characteristic (ROC) Curve (AUC), Gini index, point biserial correlation coefficient, mean square error, root mean square error, coefficient of determination, mean absolute prediction error, and others. Threshold-dependent strategies can be realized by calculating values of sensitivity, specificity, positive predictive value, negative predictive value, positive likelihood ratio, negative likelihood ratio, true skill statistic, odds ratio, Yule's Y, Yule's Q, Phi coefficient, Kappa, normalized mutual information, extreme dependency score, and others. Based on existing problems related to the development, application and evaluation of SDMs, the present study suggests the following. (1) Because biased presence samples can influence the result of prediction, integrating a sample selection module in the SDM could improve the reliability of model prediction. (2) Bioclimatic variables (such as WorldClim) that are calculated from a Digital Elevation Model (DEM) may co-linearly correlate with each other, and such collinearity may result in overfitting when modeling the potential distribution of a species. As a result, selecting variables based on calculation of the Variance Inflation Factor (VIF) is a suitable means to avoid overfitting. (3) In addition to abiotic factors, biotic factors are also important determinants for species distribution. Thus, the use of biotic variables could improve the model results, although biotic factors are not easy to delineate within a geographic space. (4) The spatial and temporal extrapolation of SDMs, which deal with problems of species potential distribution at different geographic ranges and time points (past and/or future), respectively, are actually based on the assumption of an equilibrium relationship between the target species and environmental conditions. However, this assumption is challenged, because species have the abilities of adaption and dispersal. (5) The Partial AUC (PAUC) is suitable for evaluation of single model performance, and the Akaike Information Criterion (AIC) could provide an objective evaluation of the performance of several SDMs.
.
DOIURL [本文引用: 1]
清香木(Pistacia weinmannifolia)是中国西南干旱河谷植被的特征种。本文利用野外调查的165个清香木分布点信息以及22个环境变量数据,基于最大熵(Maxent)算法构建清香木分布的适宜生境预测模型,并据此模拟清香木在我国西南地区的适宜分布区,以及历史和未来不同气候情景下的分布格局变化。结果表明:清香木生境预测的Maxent模型准确性非常高(AUC=0.974),温度季节性变化、极端低温和降水量是限制其分布的主要气候因子。清香木当前的潜在分布区集中在我国西南干旱河谷区,其适宜生境的气候特征是降水少、温度季节性变化小且无极端低温。对清香木在末次间冰期和末次冰盛期分布的模拟结果表明,其分布区范围均以诸大江河的河谷为中心,随气候变化在我国西南地区主要呈现先向东扩张,然后向西退缩的趋势,并印证了"冰期走出横断山(glacial out-of-Hengduan Mts.)"的观点。在未来(2061–2080年)3种典型浓度路径(representative concentration pathway,RCP)的气候情景下,清香木在我国西南地区的分布都向东扩张,主要分布在云贵高原与四川盆地结合地带的河谷,以及云贵高原与广西西部交界地带的河谷中,这也反映了这些地区河谷地段干旱化的可能,而当前的潜在分布区趋于消失;清香木的潜在适宜分布面积在中低浓度路径情景下均将减少约33%,而在高浓度路径情景下有所增加。
DOIURL [本文引用: 1]
清香木(Pistacia weinmannifolia)是中国西南干旱河谷植被的特征种。本文利用野外调查的165个清香木分布点信息以及22个环境变量数据,基于最大熵(Maxent)算法构建清香木分布的适宜生境预测模型,并据此模拟清香木在我国西南地区的适宜分布区,以及历史和未来不同气候情景下的分布格局变化。结果表明:清香木生境预测的Maxent模型准确性非常高(AUC=0.974),温度季节性变化、极端低温和降水量是限制其分布的主要气候因子。清香木当前的潜在分布区集中在我国西南干旱河谷区,其适宜生境的气候特征是降水少、温度季节性变化小且无极端低温。对清香木在末次间冰期和末次冰盛期分布的模拟结果表明,其分布区范围均以诸大江河的河谷为中心,随气候变化在我国西南地区主要呈现先向东扩张,然后向西退缩的趋势,并印证了"冰期走出横断山(glacial out-of-Hengduan Mts.)"的观点。在未来(2061–2080年)3种典型浓度路径(representative concentration pathway,RCP)的气候情景下,清香木在我国西南地区的分布都向东扩张,主要分布在云贵高原与四川盆地结合地带的河谷,以及云贵高原与广西西部交界地带的河谷中,这也反映了这些地区河谷地段干旱化的可能,而当前的潜在分布区趋于消失;清香木的潜在适宜分布面积在中低浓度路径情景下均将减少约33%,而在高浓度路径情景下有所增加。
URL [本文引用: 1]
Covariance analysis, curvefitting, and canonical correspondence analysis (CCA) were used to explore the effects of topographic factors on the plant diversity and distribution patterns of ground flora with different growth forms in the karst mountains of Southwest Guangxi, China. A total of 152 ground plants were recorded. Among them, 37 species were ferns, 44 species herbs, 9 species lianas, and 62 species shrubs. Covariance analysis revealed that altitude significantly correlated with the individual number and richness of ground plants, and slope aspect had a significant effect on richness. Statistical analyses showed a highly significant nonlinear correlation between the individual number or richness of ground plants and altitude. Results of CCA revealed that slope aspect had a significant effect on the distribution pattern of ferns, and slope had a significant effect on the distribution patterns of herbs, lianas and shrubs. Ferns were more sensitive than herbs, lianas and shrubs to changes in heat and soil water caused by aspect. The effect of slope was stronger than that of elevation on soil water and nutrients, and it was the most important topographic factor that affected the distribution patterns of herbs, lianas and shrubs in this region.
URL [本文引用: 1]
Covariance analysis, curvefitting, and canonical correspondence analysis (CCA) were used to explore the effects of topographic factors on the plant diversity and distribution patterns of ground flora with different growth forms in the karst mountains of Southwest Guangxi, China. A total of 152 ground plants were recorded. Among them, 37 species were ferns, 44 species herbs, 9 species lianas, and 62 species shrubs. Covariance analysis revealed that altitude significantly correlated with the individual number and richness of ground plants, and slope aspect had a significant effect on richness. Statistical analyses showed a highly significant nonlinear correlation between the individual number or richness of ground plants and altitude. Results of CCA revealed that slope aspect had a significant effect on the distribution pattern of ferns, and slope had a significant effect on the distribution patterns of herbs, lianas and shrubs. Ferns were more sensitive than herbs, lianas and shrubs to changes in heat and soil water caused by aspect. The effect of slope was stronger than that of elevation on soil water and nutrients, and it was the most important topographic factor that affected the distribution patterns of herbs, lianas and shrubs in this region.
DOIURL [本文引用: 1]
四合木为鄂尔多斯高原所特有的蒺藜科单种属植物,因起源古老、抗逆性强,并为生物多样性起源和环境演变研究的理想对象,四合木具有重要的学术研究价值且倍受关注。但近年来四合木分布区生态环境与景观格局发生了显著变化,原有植被自然景观破碎化明显,生境岛屿化现象普遍,致使其种群数量锐减,处于濒危状态。为此,本文即以四合木为研究对象,以其分布范围为研究区域,以四合木分布区内不同的种群斑块为生境单元,借助1977、1990、2000与2005年4个时期陆地卫星遥感影像,运用3S技术,在分析四合木种群面积动态变化的基础上,探讨其生境景观格局时空演变特征及其影响因子与作用份额;通过不同生境四合木种群数量与结构特征的比较研究,分析四合木对生境适应性的空间差异及生境与景观格局变化对其种群动态的影响;在深入分析四合木生境需求的基础上,确定影响其生存的主要生态因子及其代用指标,并用评价模型进行生境适宜性评价。主要结论如下: 1.四合木主要分布在黄河与桌子山之间的狭长地带上,分布区总面积2824.31km2,由8个种群斑块组成。在研究时段内其种群面积不断减少,28a年间减少了19.36%,年均减少13.40km2;但在不同时空尺度上的减少速率并不相同。其中,以1990~2000年间减少得最快,1977~1990年间减少得最慢,且其年减少率有逐年增大趋势;在空间分布上四合木面积以桃司兔种群减少得最多,低山种群减少得最少;千里山种群减少速率最大,乌家庙种群减少速率最小。 2.四合木是研究区的基质景观,但1977年以来其生境景观格局发生了较大变化。在整个景观中,斑块数量、斑块密度、景观形状指数及多样性与均匀性指数不断增加,而斑块平均面积与优势度指数逐年减小,表明研究区的景观组分趋于复杂化、破碎化与多元化。各生境类型区中,以石嘴山、桃司兔、千里山与乌达种群的景观格局较为破碎,景观结构趋于复杂;乌家庙、棋盘井与海南种群的景观格局较为完整,景观中有连通度极高的优势斑块类型存在。在各种景观中,四合木是研究区最大的景观组分,但研究时段内其主导地位不断下降,空间分布趋于分散、破碎;而耕地与林地、城镇与居民点、工矿、沙化土地及其它植被面积大幅增长,有连片延续发展且在景观中的作用与影响程度逐渐加大趋势。 3.研究区历年气候波动和人类活动变化的代用指标分析表明,20世纪70年代以来,四合木分布区旱化趋势加剧,水分亏缺严重,加之人口数量剧增及城市化与工业化的巨大压力,使四合木的生存环境面临更为严峻的考验。各因素对四合木生境影响的累加效应分析表明,自然因素与四合木种群面积的关联程度大多以4a左右的平均尺度最高,人为因素的关联程度大多以6a左右的尺度最高,具有在时间尺度及累加作用上大于气候条件变化的特点。从主成分因子的信息贡献来看,人为因素对四合木生境影响作用的贡献率为55.66%,自然因素的贡献率为44.34%,可见人为因素的作用是导致四合木生境劣化的主要原因。 4.不同生境的四合木种群数量与结构特征有所不同,表明四合木对不同生境的适应性有一定差异。其中四合木密度与多度以海南、低山种群较高,棋盘井与乌达种群较低;盖度、高度、冠幅以低山、乌家庙种群较大,乌达、千里山种群较小;重要值和综合优势比以低山、乌家庙、棋盘井种群较高,乌达与千里山种群较低;不同生境中四合木种群的径级、龄级、冠幅、高度结构亦有差别,体现出低山、乌家庙、棋盘井种群中四合木的适应性较高,而乌达与千里山种群中其适应性相对较低的特点。 5.四合木生境适宜性的定量评价与分析表明,研究区内的气候与地形等自然因素不是四合木生长及濒危的限制性因素;而人类活动的干扰使其较不适宜或不适宜的生境面积增加了1.42%,且有33.71km2的可生存生境退化为较不适宜生境,说明人为活动可能是导致四合木局部濒危的关键因素。在各生境类型区内,以乌家庙、低山、棋盘井种群的生境质量较好,而乌达与千里山种群的生境质量较差。
DOIURL [本文引用: 1]
四合木为鄂尔多斯高原所特有的蒺藜科单种属植物,因起源古老、抗逆性强,并为生物多样性起源和环境演变研究的理想对象,四合木具有重要的学术研究价值且倍受关注。但近年来四合木分布区生态环境与景观格局发生了显著变化,原有植被自然景观破碎化明显,生境岛屿化现象普遍,致使其种群数量锐减,处于濒危状态。为此,本文即以四合木为研究对象,以其分布范围为研究区域,以四合木分布区内不同的种群斑块为生境单元,借助1977、1990、2000与2005年4个时期陆地卫星遥感影像,运用3S技术,在分析四合木种群面积动态变化的基础上,探讨其生境景观格局时空演变特征及其影响因子与作用份额;通过不同生境四合木种群数量与结构特征的比较研究,分析四合木对生境适应性的空间差异及生境与景观格局变化对其种群动态的影响;在深入分析四合木生境需求的基础上,确定影响其生存的主要生态因子及其代用指标,并用评价模型进行生境适宜性评价。主要结论如下: 1.四合木主要分布在黄河与桌子山之间的狭长地带上,分布区总面积2824.31km2,由8个种群斑块组成。在研究时段内其种群面积不断减少,28a年间减少了19.36%,年均减少13.40km2;但在不同时空尺度上的减少速率并不相同。其中,以1990~2000年间减少得最快,1977~1990年间减少得最慢,且其年减少率有逐年增大趋势;在空间分布上四合木面积以桃司兔种群减少得最多,低山种群减少得最少;千里山种群减少速率最大,乌家庙种群减少速率最小。 2.四合木是研究区的基质景观,但1977年以来其生境景观格局发生了较大变化。在整个景观中,斑块数量、斑块密度、景观形状指数及多样性与均匀性指数不断增加,而斑块平均面积与优势度指数逐年减小,表明研究区的景观组分趋于复杂化、破碎化与多元化。各生境类型区中,以石嘴山、桃司兔、千里山与乌达种群的景观格局较为破碎,景观结构趋于复杂;乌家庙、棋盘井与海南种群的景观格局较为完整,景观中有连通度极高的优势斑块类型存在。在各种景观中,四合木是研究区最大的景观组分,但研究时段内其主导地位不断下降,空间分布趋于分散、破碎;而耕地与林地、城镇与居民点、工矿、沙化土地及其它植被面积大幅增长,有连片延续发展且在景观中的作用与影响程度逐渐加大趋势。 3.研究区历年气候波动和人类活动变化的代用指标分析表明,20世纪70年代以来,四合木分布区旱化趋势加剧,水分亏缺严重,加之人口数量剧增及城市化与工业化的巨大压力,使四合木的生存环境面临更为严峻的考验。各因素对四合木生境影响的累加效应分析表明,自然因素与四合木种群面积的关联程度大多以4a左右的平均尺度最高,人为因素的关联程度大多以6a左右的尺度最高,具有在时间尺度及累加作用上大于气候条件变化的特点。从主成分因子的信息贡献来看,人为因素对四合木生境影响作用的贡献率为55.66%,自然因素的贡献率为44.34%,可见人为因素的作用是导致四合木生境劣化的主要原因。 4.不同生境的四合木种群数量与结构特征有所不同,表明四合木对不同生境的适应性有一定差异。其中四合木密度与多度以海南、低山种群较高,棋盘井与乌达种群较低;盖度、高度、冠幅以低山、乌家庙种群较大,乌达、千里山种群较小;重要值和综合优势比以低山、乌家庙、棋盘井种群较高,乌达与千里山种群较低;不同生境中四合木种群的径级、龄级、冠幅、高度结构亦有差别,体现出低山、乌家庙、棋盘井种群中四合木的适应性较高,而乌达与千里山种群中其适应性相对较低的特点。 5.四合木生境适宜性的定量评价与分析表明,研究区内的气候与地形等自然因素不是四合木生长及濒危的限制性因素;而人类活动的干扰使其较不适宜或不适宜的生境面积增加了1.42%,且有33.71km2的可生存生境退化为较不适宜生境,说明人为活动可能是导致四合木局部濒危的关键因素。在各生境类型区内,以乌家庙、低山、棋盘井种群的生境质量较好,而乌达与千里山种群的生境质量较差。
.
URL [本文引用: 4]
凤仙花属Impatiens L隶属于凤仙花科Balsaminaceae,是现存被子植物中的特大属之一,亦是一个高度分化和多样化显著的大属。凤仙花属植物对生长环境要求较为严格,特有现象和地域性极为明显。绝大多数种类为我国或某个省区狭域分布的特有种,在我国华南和西南石灰岩地区这种特有现象更为显著。然而,对这些种类与石灰岩基质的关系一直缺乏深入的研究,尤其缺少种群结构、异质性生境、遗传结构、细胞学、生理生态以及种质资源保存等方面的综合研究工作,因而这些特有植物的特有机理及其适应对策目前仍没有得到很好的诠释。 海南凤仙花Impatiens hainanensis Y. L. Chen仅分布于海南石灰岩地区,包括昌江、东方、白沙等地,多见于山顶裸露石灰岩缝隙,是该地区的石灰岩专性特有植物。由于当地少数民族有刀耕火种、采集药材的传统,海南凤仙花的生境遭到了很大的破坏,按照IUCN的濒危物种评估标准,现已处于濒危状态,被列为海南省级重点保护野生植物。 本研究讨论了海南凤仙花保护生态学内容,涉及其特有濒危过程的群落生态学、繁殖生态学和保护遗传学等内容,以期了解石灰岩特有植物适应石灰岩特殊生境的内在机制,分析其特有濒危的主要原因;并试图通过建立基于组织培养的扩繁体系,为有效保护其种质资源、制订科学而合理的保护策略提供科学的决策依据。主要研究成果与创新点如下: 1.海南凤仙花群落生态学研究 应用TWINSPAN和DCA,研究海南凤仙花所在群落植被的分布格局,并探讨了海南凤仙花种群大小及其不同群丛下物种的多样性。研究表明:(1)DCA排序结果与TWINSPAN分类结果一致,将16个样方划分为4个群丛类型;(2)DCA排序能较好地反映各群丛类型与环境因子的生态关系,DCA第一、第二排序轴分布反映了海拔和郁闭度的变化;(3)不同群丛的物种多样性指数和丰富度指数存在差异,且不同层之间也存在差异。海南凤仙花所在群落不同群丛的形成与环境因子关系密切,其中海拔是影响其分布格局和物种多样性的环境因子;(4)不同群落结构下的海南凤仙花的种群大小及幼苗更新能力不同,在今后的保护中应采取不同的保护措施。 2.海南凤仙花繁殖生态学研究 对3个不同海拔梯度的种群进行了开花生物学特性和开花物候、花器官结构、花粉活力和柱头活性、传粉者种类和访花行为及繁育系统的比较研究。结果发现,从花芽萌生至凋谢大约25.2d,花朵开放平均寿命4.10d,雄性期和雌性期分别约为3.95d和0.95d;种群开花峰期在8月初,高海拔种群花期高峰相对滞后。低、中海拔种群开花第2天花粉活力最高,高海拔种群以第1天花粉活力最高,但各开花天次都较中低海拔种群低;柱头活性随开花时间的推移而增强,高海拔种群开花各天次均较低、中海拔种群低。主要传粉昆虫为黄黑无垫蜂Amegilla leptocoma和绿条无垫蜂Amegilla zonata,中低海拔种群以黄黑无垫蜂为主,高海拔种群以绿条无垫蜂为主。无自动自花授粉和无融合生殖现象,人工授粉能明显增加座果率(75%-90%),自然座果率高海拔种群相对较低(40%-60%),说明存在较强的传粉者限制。因此,对于海南凤仙花的保护需要同时关注生境与有效传粉者的保护,促进有效传粉昆虫在不同海拔种群之间的往来,保证种群间的花粉流与种子流,维持海南凤仙花的种群遗传多样性与有效种群大小。 3.海南凤仙花种群遗传学研究 运用ISSR分子标记技术,分析了海南岛石灰岩地区的海南凤仙花自然种群的遗传结构,比较了三个海拔梯度对种群遗传结构的影响。研究结果显示:13个ISSR引物的扩增结果显示其多态性位点百分率为97.7%;AMOVA分析表明海南凤仙花自然种群的变异主要来自种群内(92%),而种群间变异较少(8%);中海拔种群的遗传多样性最为丰富;而高海拔种群的遗传多样性则较低;种群的遗传距离和地理距离表现出显著的高度专一性。分析表明海拔是影响种群基因流的重要因素,高海拔种群可以向中低海拔种群进行传播花粉和种子等基因流动,而中低海拔种群向高海拔种群进行类似的基因流动则较为困难,海南凤仙花数量较少的主要原因可能来自于其生境的限制。因此在以后的保护工作中要特别注重其生境的保护,避免人为因素的破坏。 4.海南凤仙花组织培养研究 以海南凤仙花种子、嫩叶和侧芽为外植体进行离体培养,获得以下优化培养基配方: (1)播种培养基:Hyponex花宝1号3.0g·L-1+蛋白胨1.0g·L-1+椰子汁CM100ml·L-1;(2)叶片诱导愈伤组织培养基:MS+6-BA3.0mg·L-1+NAA0.1mg·L-1;(3)愈伤组织分化培养基:MS+6-BA2.0mg·L-1+NAA0.5mg·L-1;(4)腋芽诱导培养基:MS+6-BA1.0mg·L-1+NAA0.2mg·L-1;(5)丛生芽增殖培养基:MS+6-BA2.0mg·L-1+NAA0.5mg·L-1;(6)生根培养基:MS+NAA0.Smg-l。
URL [本文引用: 4]
凤仙花属Impatiens L隶属于凤仙花科Balsaminaceae,是现存被子植物中的特大属之一,亦是一个高度分化和多样化显著的大属。凤仙花属植物对生长环境要求较为严格,特有现象和地域性极为明显。绝大多数种类为我国或某个省区狭域分布的特有种,在我国华南和西南石灰岩地区这种特有现象更为显著。然而,对这些种类与石灰岩基质的关系一直缺乏深入的研究,尤其缺少种群结构、异质性生境、遗传结构、细胞学、生理生态以及种质资源保存等方面的综合研究工作,因而这些特有植物的特有机理及其适应对策目前仍没有得到很好的诠释。 海南凤仙花Impatiens hainanensis Y. L. Chen仅分布于海南石灰岩地区,包括昌江、东方、白沙等地,多见于山顶裸露石灰岩缝隙,是该地区的石灰岩专性特有植物。由于当地少数民族有刀耕火种、采集药材的传统,海南凤仙花的生境遭到了很大的破坏,按照IUCN的濒危物种评估标准,现已处于濒危状态,被列为海南省级重点保护野生植物。 本研究讨论了海南凤仙花保护生态学内容,涉及其特有濒危过程的群落生态学、繁殖生态学和保护遗传学等内容,以期了解石灰岩特有植物适应石灰岩特殊生境的内在机制,分析其特有濒危的主要原因;并试图通过建立基于组织培养的扩繁体系,为有效保护其种质资源、制订科学而合理的保护策略提供科学的决策依据。主要研究成果与创新点如下: 1.海南凤仙花群落生态学研究 应用TWINSPAN和DCA,研究海南凤仙花所在群落植被的分布格局,并探讨了海南凤仙花种群大小及其不同群丛下物种的多样性。研究表明:(1)DCA排序结果与TWINSPAN分类结果一致,将16个样方划分为4个群丛类型;(2)DCA排序能较好地反映各群丛类型与环境因子的生态关系,DCA第一、第二排序轴分布反映了海拔和郁闭度的变化;(3)不同群丛的物种多样性指数和丰富度指数存在差异,且不同层之间也存在差异。海南凤仙花所在群落不同群丛的形成与环境因子关系密切,其中海拔是影响其分布格局和物种多样性的环境因子;(4)不同群落结构下的海南凤仙花的种群大小及幼苗更新能力不同,在今后的保护中应采取不同的保护措施。 2.海南凤仙花繁殖生态学研究 对3个不同海拔梯度的种群进行了开花生物学特性和开花物候、花器官结构、花粉活力和柱头活性、传粉者种类和访花行为及繁育系统的比较研究。结果发现,从花芽萌生至凋谢大约25.2d,花朵开放平均寿命4.10d,雄性期和雌性期分别约为3.95d和0.95d;种群开花峰期在8月初,高海拔种群花期高峰相对滞后。低、中海拔种群开花第2天花粉活力最高,高海拔种群以第1天花粉活力最高,但各开花天次都较中低海拔种群低;柱头活性随开花时间的推移而增强,高海拔种群开花各天次均较低、中海拔种群低。主要传粉昆虫为黄黑无垫蜂Amegilla leptocoma和绿条无垫蜂Amegilla zonata,中低海拔种群以黄黑无垫蜂为主,高海拔种群以绿条无垫蜂为主。无自动自花授粉和无融合生殖现象,人工授粉能明显增加座果率(75%-90%),自然座果率高海拔种群相对较低(40%-60%),说明存在较强的传粉者限制。因此,对于海南凤仙花的保护需要同时关注生境与有效传粉者的保护,促进有效传粉昆虫在不同海拔种群之间的往来,保证种群间的花粉流与种子流,维持海南凤仙花的种群遗传多样性与有效种群大小。 3.海南凤仙花种群遗传学研究 运用ISSR分子标记技术,分析了海南岛石灰岩地区的海南凤仙花自然种群的遗传结构,比较了三个海拔梯度对种群遗传结构的影响。研究结果显示:13个ISSR引物的扩增结果显示其多态性位点百分率为97.7%;AMOVA分析表明海南凤仙花自然种群的变异主要来自种群内(92%),而种群间变异较少(8%);中海拔种群的遗传多样性最为丰富;而高海拔种群的遗传多样性则较低;种群的遗传距离和地理距离表现出显著的高度专一性。分析表明海拔是影响种群基因流的重要因素,高海拔种群可以向中低海拔种群进行传播花粉和种子等基因流动,而中低海拔种群向高海拔种群进行类似的基因流动则较为困难,海南凤仙花数量较少的主要原因可能来自于其生境的限制。因此在以后的保护工作中要特别注重其生境的保护,避免人为因素的破坏。 4.海南凤仙花组织培养研究 以海南凤仙花种子、嫩叶和侧芽为外植体进行离体培养,获得以下优化培养基配方: (1)播种培养基:Hyponex花宝1号3.0g·L-1+蛋白胨1.0g·L-1+椰子汁CM100ml·L-1;(2)叶片诱导愈伤组织培养基:MS+6-BA3.0mg·L-1+NAA0.1mg·L-1;(3)愈伤组织分化培养基:MS+6-BA2.0mg·L-1+NAA0.5mg·L-1;(4)腋芽诱导培养基:MS+6-BA1.0mg·L-1+NAA0.2mg·L-1;(5)丛生芽增殖培养基:MS+6-BA2.0mg·L-1+NAA0.5mg·L-1;(6)生根培养基:MS+NAA0.Smg-l。
Modeling the potential distribution of Picea chihuahuana Martínez, an endangered species at the Sierra Madre Occidental, Mexico
1
2015
... 使用最大熵模型(MaxEnt)等物种分布模型预测物种的潜在分布区已成为生物多样性及生物保护领域的研究热点.预测物种的潜在适宜生境分布区对制定有效的生物多样性保护策略(
Predicting species distributions: Use of climatic parameters in BIOCLIM and its impact on predictions of species’ current and future distributions
1
2005
... 使用最大熵模型(MaxEnt)等物种分布模型预测物种的潜在分布区已成为生物多样性及生物保护领域的研究热点.预测物种的潜在适宜生境分布区对制定有效的生物多样性保护策略(
Environment-dependence of quantitative genetic parameters in Impatiens pallida
1
1996
... MaxEnt模型运用较少的分布点(8个)也很好地预测了海南凤仙花在当前时期的潜在适宜生境分布区.其最适宜生境主要分布在白沙西部与南部、昌江中部和南部、东方东部、乐东北部等地, 与现今已发现的石灰岩地区有着较好的一致性(
濒危物种长柄扁桃的潜在分布与保护策略
2
2017
... 为有效地评价基于小样本建模的模型预测精度, 本研究选取受试者工作特征曲线下面积(AUC)、Kappa系数、真实技巧统计值(TSS)及总体精度4个指标综合评估模型预测的准确性.其中Kappa系数、TSS和总体精度受到阈值的影响, 需要物种不存在数据来协助计算, 因此, 本研究运用
... 提到的具体方法, 随机生成与未参与建模的5个实际分布验证点相对应的5个“真实”不存在数据, 此过程重复10次, 之后通过计算混淆矩阵, 根据相关公式得到10套各评价指标的评估值.具体操作方法在此不做叙述, 可参考
濒危物种长柄扁桃的潜在分布与保护策略
2
2017
... 为有效地评价基于小样本建模的模型预测精度, 本研究选取受试者工作特征曲线下面积(AUC)、Kappa系数、真实技巧统计值(TSS)及总体精度4个指标综合评估模型预测的准确性.其中Kappa系数、TSS和总体精度受到阈值的影响, 需要物种不存在数据来协助计算, 因此, 本研究运用
... 提到的具体方法, 随机生成与未参与建模的5个实际分布验证点相对应的5个“真实”不存在数据, 此过程重复10次, 之后通过计算混淆矩阵, 根据相关公式得到10套各评价指标的评估值.具体操作方法在此不做叙述, 可参考
Distribution prediction model of a rare orchid species ( Vanda bicolor Griff.) using small sample size
1
2017
... 使用最大熵模型(MaxEnt)等物种分布模型预测物种的潜在分布区已成为生物多样性及生物保护领域的研究热点.预测物种的潜在适宜生境分布区对制定有效的生物多样性保护策略(
Water source partitioning among trees growing on carbonate rock in a subtropical region of Guangxi, China
1
2012
... MaxEnt分析结果表明影响海南凤仙花分布的主导因子主要是坡度、最干季降水量、降水量季节性变异系数.其中坡度的贡献率高达58.9%, 降水量季节性变异系数及最干季降水量这2个与水分相关的因子累计贡献率达到39.6%, 说明海南凤仙花分布受限的最主要因素是地形因子(坡度)和水分因子.有研究发现, 在喀斯特山地森林即石灰岩山地, 坡度是所有地形因素中影响草本植物分布的最主要因子(
An improved approach for predicting the distribution of rare and endangered species from occurrence and pseudo-absence data
1
2004
... 使用最大熵模型(MaxEnt)等物种分布模型预测物种的潜在分布区已成为生物多样性及生物保护领域的研究热点.预测物种的潜在适宜生境分布区对制定有效的生物多样性保护策略(
A practical method to speed up the discovery of unknown populations using species distribution models
1
2015
... 使用最大熵模型(MaxEnt)等物种分布模型预测物种的潜在分布区已成为生物多样性及生物保护领域的研究热点.预测物种的潜在适宜生境分布区对制定有效的生物多样性保护策略(
Endemic seed plant species from Hainan Island: A checklist
1
2010
... 海南凤仙花(Impatiens hainanensis)是海南岛特有种, 属海南岛石灰岩特有植物(
高寒退化草地甘肃臭草种群空间分布格局对土壤水分的响应
1
2012
... 本研究模型预测选取的环境因子因数据来源获取的原因, 只考虑了温度、水分等气候因子及地形因子, 未考虑土壤、物种适应性、生物竞争等其他因素, 存在一定的不足之处.土壤直接为植物提供生长发育所需的水分及营养, 对物种的分布结构有着明显的限制作用(
高寒退化草地甘肃臭草种群空间分布格局对土壤水分的响应
1
2012
... 本研究模型预测选取的环境因子因数据来源获取的原因, 只考虑了温度、水分等气候因子及地形因子, 未考虑土壤、物种适应性、生物竞争等其他因素, 存在一定的不足之处.土壤直接为植物提供生长发育所需的水分及营养, 对物种的分布结构有着明显的限制作用(
The effect of sample size and species characteristics on performance of different species distribution modeling methods
2
2006
... 使用最大熵模型(MaxEnt)等物种分布模型预测物种的潜在分布区已成为生物多样性及生物保护领域的研究热点.预测物种的潜在适宜生境分布区对制定有效的生物多样性保护策略(
... ).MaxEnt模型因具有仅利用物种出现点、易于掌握与使用且在多数情况下模拟精度通常优于其他模型的优点, 使其成为现代物种生境适宜性评价方面应用最广泛的一个工具(
Very high resolution interpolated climate surfaces for global land areas
1
2005
... 当前时期的气候数据从世界气候数据库(http://www.worldclim.org/)下载, 采用2.0版的数据.该数据库通过收集全球各个气象观测站1970-2000年的气候数据, 运用ANUSPLIN程序中的薄板平滑样条函数插值法, 以海拔、经度、纬度为自变量, 从而进行全球范围的插值, 得到30?分辨率(约1 km)的全球连续气候数据(
植物表型可塑性对非生物环境因子的响应研究进展
2
2008
... MaxEnt模型运用较少的分布点(8个)也很好地预测了海南凤仙花在当前时期的潜在适宜生境分布区.其最适宜生境主要分布在白沙西部与南部、昌江中部和南部、东方东部、乐东北部等地, 与现今已发现的石灰岩地区有着较好的一致性(
... 本研究模型预测选取的环境因子因数据来源获取的原因, 只考虑了温度、水分等气候因子及地形因子, 未考虑土壤、物种适应性、生物竞争等其他因素, 存在一定的不足之处.土壤直接为植物提供生长发育所需的水分及营养, 对物种的分布结构有着明显的限制作用(
植物表型可塑性对非生物环境因子的响应研究进展
2
2008
... MaxEnt模型运用较少的分布点(8个)也很好地预测了海南凤仙花在当前时期的潜在适宜生境分布区.其最适宜生境主要分布在白沙西部与南部、昌江中部和南部、东方东部、乐东北部等地, 与现今已发现的石灰岩地区有着较好的一致性(
... 本研究模型预测选取的环境因子因数据来源获取的原因, 只考虑了温度、水分等气候因子及地形因子, 未考虑土壤、物种适应性、生物竞争等其他因素, 存在一定的不足之处.土壤直接为植物提供生长发育所需的水分及营养, 对物种的分布结构有着明显的限制作用(
Maxent modeling for predicting suitable habitat for threatened and endangered tree Canacomyrica monticola in New Caledonia
1
2009
... 使用最大熵模型(MaxEnt)等物种分布模型预测物种的潜在分布区已成为生物多样性及生物保护领域的研究热点.预测物种的潜在适宜生境分布区对制定有效的生物多样性保护策略(
Synchrony, co-mpensatory dynamics, and the functional trait basis ofphonological diversity in a tropical dry forest tree co-mmunity: Effects of rainfall seasonality
1
2016
... MaxEnt分析结果表明影响海南凤仙花分布的主导因子主要是坡度、最干季降水量、降水量季节性变异系数.其中坡度的贡献率高达58.9%, 降水量季节性变异系数及最干季降水量这2个与水分相关的因子累计贡献率达到39.6%, 说明海南凤仙花分布受限的最主要因素是地形因子(坡度)和水分因子.有研究发现, 在喀斯特山地森林即石灰岩山地, 坡度是所有地形因素中影响草本植物分布的最主要因子(
岛屿特有种全缘冬青遗传多样性的ISSR分析
1
2005
... 海南凤仙花(Impatiens hainanensis)是海南岛特有种, 属海南岛石灰岩特有植物(
岛屿特有种全缘冬青遗传多样性的ISSR分析
1
2005
... 海南凤仙花(Impatiens hainanensis)是海南岛特有种, 属海南岛石灰岩特有植物(
基于MaxEnt模型的青钱柳潜在适宜栽培区预测
1
2017
... 使用最大熵模型(MaxEnt)等物种分布模型预测物种的潜在分布区已成为生物多样性及生物保护领域的研究热点.预测物种的潜在适宜生境分布区对制定有效的生物多样性保护策略(
基于MaxEnt模型的青钱柳潜在适宜栽培区预测
1
2017
... 使用最大熵模型(MaxEnt)等物种分布模型预测物种的潜在分布区已成为生物多样性及生物保护领域的研究热点.预测物种的潜在适宜生境分布区对制定有效的生物多样性保护策略(
气候变化对蒙古扁桃适宜分布范围和空间格局的影响
1
2014
... 使用最大熵模型(MaxEnt)等物种分布模型预测物种的潜在分布区已成为生物多样性及生物保护领域的研究热点.预测物种的潜在适宜生境分布区对制定有效的生物多样性保护策略(
气候变化对蒙古扁桃适宜分布范围和空间格局的影响
1
2014
... 使用最大熵模型(MaxEnt)等物种分布模型预测物种的潜在分布区已成为生物多样性及生物保护领域的研究热点.预测物种的潜在适宜生境分布区对制定有效的生物多样性保护策略(
Potential distribution of endemic Scutellaria chamaedrifolia geographic information system and statistical model approach
1
2015
... 使用最大熵模型(MaxEnt)等物种分布模型预测物种的潜在分布区已成为生物多样性及生物保护领域的研究热点.预测物种的潜在适宜生境分布区对制定有效的生物多样性保护策略(
A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter
1
2013
... 使用最大熵模型(MaxEnt)等物种分布模型预测物种的潜在分布区已成为生物多样性及生物保护领域的研究热点.预测物种的潜在适宜生境分布区对制定有效的生物多样性保护策略(
石灰岩地区连片出露石丛生境植物水分来源的季节性差异
1
2011
... MaxEnt分析结果表明影响海南凤仙花分布的主导因子主要是坡度、最干季降水量、降水量季节性变异系数.其中坡度的贡献率高达58.9%, 降水量季节性变异系数及最干季降水量这2个与水分相关的因子累计贡献率达到39.6%, 说明海南凤仙花分布受限的最主要因素是地形因子(坡度)和水分因子.有研究发现, 在喀斯特山地森林即石灰岩山地, 坡度是所有地形因素中影响草本植物分布的最主要因子(
石灰岩地区连片出露石丛生境植物水分来源的季节性差异
1
2011
... MaxEnt分析结果表明影响海南凤仙花分布的主导因子主要是坡度、最干季降水量、降水量季节性变异系数.其中坡度的贡献率高达58.9%, 降水量季节性变异系数及最干季降水量这2个与水分相关的因子累计贡献率达到39.6%, 说明海南凤仙花分布受限的最主要因素是地形因子(坡度)和水分因子.有研究发现, 在喀斯特山地森林即石灰岩山地, 坡度是所有地形因素中影响草本植物分布的最主要因子(
海南俄贤岭石灰岩山地海南凤仙花所在群落植物区系及生态特征
1
2012
... MaxEnt模型运用较少的分布点(8个)也很好地预测了海南凤仙花在当前时期的潜在适宜生境分布区.其最适宜生境主要分布在白沙西部与南部、昌江中部和南部、东方东部、乐东北部等地, 与现今已发现的石灰岩地区有着较好的一致性(
海南俄贤岭石灰岩山地海南凤仙花所在群落植物区系及生态特征
1
2012
... MaxEnt模型运用较少的分布点(8个)也很好地预测了海南凤仙花在当前时期的潜在适宜生境分布区.其最适宜生境主要分布在白沙西部与南部、昌江中部和南部、东方东部、乐东北部等地, 与现今已发现的石灰岩地区有着较好的一致性(
海南岛石灰岩地区珍稀濒危植物及其保护
1
2005
... 海南凤仙花(Impatiens hainanensis)是海南岛特有种, 属海南岛石灰岩特有植物(
海南岛石灰岩地区珍稀濒危植物及其保护
1
2005
... 海南凤仙花(Impatiens hainanensis)是海南岛特有种, 属海南岛石灰岩特有植物(
海南石灰岩地区的种子植物区系
1
2014
... 模型输出结果为ASCII格式, 可解释为0-1范围的生境适宜值, 将每套数据运行得到的平均的ASCII文件导入ArcGIS 10.2中转为栅格数据, 利用栅格计算器, 求得10套ASCII文件的平均值, 再将该平均值图层与海南岛土地覆被类型图进行相乘, 剔除耕地、草地、水体、城乡工矿地、居民用地等不适生生境(不适生的土地覆被类型栅格单元值为0, 适生的土地覆被类型栅格单元值为1).再运用重分类功能中的自然间断点分级法(Jenks)进行重分类, 将海南凤仙花的生境适宜性划分为3个等级: 最适宜、次适宜和不适宜.此外,
海南石灰岩地区的种子植物区系
1
2014
... 模型输出结果为ASCII格式, 可解释为0-1范围的生境适宜值, 将每套数据运行得到的平均的ASCII文件导入ArcGIS 10.2中转为栅格数据, 利用栅格计算器, 求得10套ASCII文件的平均值, 再将该平均值图层与海南岛土地覆被类型图进行相乘, 剔除耕地、草地、水体、城乡工矿地、居民用地等不适生生境(不适生的土地覆被类型栅格单元值为0, 适生的土地覆被类型栅格单元值为1).再运用重分类功能中的自然间断点分级法(Jenks)进行重分类, 将海南凤仙花的生境适宜性划分为3个等级: 最适宜、次适宜和不适宜.此外,
New paradigms for modelling species distributions?
1
2004
... 使用最大熵模型(MaxEnt)等物种分布模型预测物种的潜在分布区已成为生物多样性及生物保护领域的研究热点.预测物种的潜在适宜生境分布区对制定有效的生物多样性保护策略(
气候变化对中国东北兴安落叶松分布的影响
1
2013
... 本研究运用MaxEnt 3.3.3k软件进行预测.将随机选取的8个海南凤仙花分布点(10套)和环境数据导入MaxEnt模型, 勾选创建反应曲线和刀切法检验, 其他参数选项默认设定.根据模型自带的Jackknife刀切法检验得到的增益结果和模型贡献率表综合地确定影响海南凤仙花的主导环境因子.通常研究会选取对模型累积贡献率在85%-95%的环境因子, 作为对物种适宜分布区影响比较重大的因子(
气候变化对中国东北兴安落叶松分布的影响
1
2013
... 本研究运用MaxEnt 3.3.3k软件进行预测.将随机选取的8个海南凤仙花分布点(10套)和环境数据导入MaxEnt模型, 勾选创建反应曲线和刀切法检验, 其他参数选项默认设定.根据模型自带的Jackknife刀切法检验得到的增益结果和模型贡献率表综合地确定影响海南凤仙花的主导环境因子.通常研究会选取对模型累积贡献率在85%-95%的环境因子, 作为对物种适宜分布区影响比较重大的因子(
我国石灰岩地区特有植物研究进展
1
2010
... 海南凤仙花(Impatiens hainanensis)是海南岛特有种, 属海南岛石灰岩特有植物(
我国石灰岩地区特有植物研究进展
1
2010
... 海南凤仙花(Impatiens hainanensis)是海南岛特有种, 属海南岛石灰岩特有植物(
气候变化对6种荒漠植物分布的潜在影响
1
2010
... 使用最大熵模型(MaxEnt)等物种分布模型预测物种的潜在分布区已成为生物多样性及生物保护领域的研究热点.预测物种的潜在适宜生境分布区对制定有效的生物多样性保护策略(
气候变化对6种荒漠植物分布的潜在影响
1
2010
... 使用最大熵模型(MaxEnt)等物种分布模型预测物种的潜在分布区已成为生物多样性及生物保护领域的研究热点.预测物种的潜在适宜生境分布区对制定有效的生物多样性保护策略(
物种分布模型的发展及评价方法
1
2015
... 本研究模型预测选取的环境因子因数据来源获取的原因, 只考虑了温度、水分等气候因子及地形因子, 未考虑土壤、物种适应性、生物竞争等其他因素, 存在一定的不足之处.土壤直接为植物提供生长发育所需的水分及营养, 对物种的分布结构有着明显的限制作用(
物种分布模型的发展及评价方法
1
2015
... 本研究模型预测选取的环境因子因数据来源获取的原因, 只考虑了温度、水分等气候因子及地形因子, 未考虑土壤、物种适应性、生物竞争等其他因素, 存在一定的不足之处.土壤直接为植物提供生长发育所需的水分及营养, 对物种的分布结构有着明显的限制作用(
气候变化情景下基于最大熵模型的中国西南地区清香木潜在分布格局模拟
1
2016
... 本研究运用MaxEnt 3.3.3k软件进行预测.将随机选取的8个海南凤仙花分布点(10套)和环境数据导入MaxEnt模型, 勾选创建反应曲线和刀切法检验, 其他参数选项默认设定.根据模型自带的Jackknife刀切法检验得到的增益结果和模型贡献率表综合地确定影响海南凤仙花的主导环境因子.通常研究会选取对模型累积贡献率在85%-95%的环境因子, 作为对物种适宜分布区影响比较重大的因子(
气候变化情景下基于最大熵模型的中国西南地区清香木潜在分布格局模拟
1
2016
... 本研究运用MaxEnt 3.3.3k软件进行预测.将随机选取的8个海南凤仙花分布点(10套)和环境数据导入MaxEnt模型, 勾选创建反应曲线和刀切法检验, 其他参数选项默认设定.根据模型自带的Jackknife刀切法检验得到的增益结果和模型贡献率表综合地确定影响海南凤仙花的主导环境因子.通常研究会选取对模型累积贡献率在85%-95%的环境因子, 作为对物种适宜分布区影响比较重大的因子(
地形对桂西南喀斯特山地森林地表植物多样性及分布格局的影响
1
2014
... MaxEnt分析结果表明影响海南凤仙花分布的主导因子主要是坡度、最干季降水量、降水量季节性变异系数.其中坡度的贡献率高达58.9%, 降水量季节性变异系数及最干季降水量这2个与水分相关的因子累计贡献率达到39.6%, 说明海南凤仙花分布受限的最主要因素是地形因子(坡度)和水分因子.有研究发现, 在喀斯特山地森林即石灰岩山地, 坡度是所有地形因素中影响草本植物分布的最主要因子(
地形对桂西南喀斯特山地森林地表植物多样性及分布格局的影响
1
2014
... MaxEnt分析结果表明影响海南凤仙花分布的主导因子主要是坡度、最干季降水量、降水量季节性变异系数.其中坡度的贡献率高达58.9%, 降水量季节性变异系数及最干季降水量这2个与水分相关的因子累计贡献率达到39.6%, 说明海南凤仙花分布受限的最主要因素是地形因子(坡度)和水分因子.有研究发现, 在喀斯特山地森林即石灰岩山地, 坡度是所有地形因素中影响草本植物分布的最主要因子(
濒危植物四合木生境的景观动态与适宜性评价研究
1
2008
... 使用最大熵模型(MaxEnt)等物种分布模型预测物种的潜在分布区已成为生物多样性及生物保护领域的研究热点.预测物种的潜在适宜生境分布区对制定有效的生物多样性保护策略(
濒危植物四合木生境的景观动态与适宜性评价研究
1
2008
... 使用最大熵模型(MaxEnt)等物种分布模型预测物种的潜在分布区已成为生物多样性及生物保护领域的研究热点.预测物种的潜在适宜生境分布区对制定有效的生物多样性保护策略(
海南凤仙花保育生态学研究
4
2014
... 海南凤仙花(Impatiens hainanensis)是海南岛特有种, 属海南岛石灰岩特有植物(
... ;
... MaxEnt模型运用较少的分布点(8个)也很好地预测了海南凤仙花在当前时期的潜在适宜生境分布区.其最适宜生境主要分布在白沙西部与南部、昌江中部和南部、东方东部、乐东北部等地, 与现今已发现的石灰岩地区有着较好的一致性(
... MaxEnt分析结果表明影响海南凤仙花分布的主导因子主要是坡度、最干季降水量、降水量季节性变异系数.其中坡度的贡献率高达58.9%, 降水量季节性变异系数及最干季降水量这2个与水分相关的因子累计贡献率达到39.6%, 说明海南凤仙花分布受限的最主要因素是地形因子(坡度)和水分因子.有研究发现, 在喀斯特山地森林即石灰岩山地, 坡度是所有地形因素中影响草本植物分布的最主要因子(
海南凤仙花保育生态学研究
4
2014
... 海南凤仙花(Impatiens hainanensis)是海南岛特有种, 属海南岛石灰岩特有植物(
... ;
... MaxEnt模型运用较少的分布点(8个)也很好地预测了海南凤仙花在当前时期的潜在适宜生境分布区.其最适宜生境主要分布在白沙西部与南部、昌江中部和南部、东方东部、乐东北部等地, 与现今已发现的石灰岩地区有着较好的一致性(
... MaxEnt分析结果表明影响海南凤仙花分布的主导因子主要是坡度、最干季降水量、降水量季节性变异系数.其中坡度的贡献率高达58.9%, 降水量季节性变异系数及最干季降水量这2个与水分相关的因子累计贡献率达到39.6%, 说明海南凤仙花分布受限的最主要因素是地形因子(坡度)和水分因子.有研究发现, 在喀斯特山地森林即石灰岩山地, 坡度是所有地形因素中影响草本植物分布的最主要因子(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}