 ,*海南大学热带农林学院, 环南海陆域生物多样性研究中心, 海口 570228
,*海南大学热带农林学院, 环南海陆域生物多样性研究中心, 海口 570228Diversity and evolution of samara in angiosperm
TAN Ke, DONG Shu-Peng, LU Tao, ZHANG Ya-Jing, XU Shi-Tao, REN Ming-Xun,*Center for Terrestrial Biodiversity of the South China Sea, Institute of Tropical Agriculture and Forestry, Hainan University, Haikou 570228, China通讯作者:
编委: 谭敦炎
责任编辑: 李敏
收稿日期:2018-03-5网络出版日期:2018-08-20
| 基金资助: |
Received:2018-03-5Online:2018-08-20
| Supported by: | SupportedbytheNationalNaturalScienceFoundationofChina( |

摘要
关键词:
Abstract
Keywords:
PDF (2079KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
谭珂, 董书鹏, 卢涛, 张亚婧, 徐诗涛, 任明迅. 被子植物翅果的多样性及演化. 植物生态学报, 2018, 42(8): 806-817 doi:10.17521/cjpe.2018.0053
TAN Ke, DONG Shu-Peng, LU Tao, ZHANG Ya-Jing, XU Shi-Tao, REN Ming-Xun.
翅果(samara)是指果实的果皮或其他部位延伸呈翅状并依靠风力传播的一类果实, 是被子植物果实适应风力传播的一个“关键创新性状”(Green, 1980; 王磊等, 2012), 可能是被子植物早期快速扩散与分化的一个主要原因(Herrera, 1989; Eriksson et al., 2000)。狭义的翅果仅指果皮延伸成翅且不开裂的干果(Harris & Harris, 2001), 而广义的翅果则涵括各类依靠风力传播的带翅的果实, 包括由非果皮部分(如苞片或萼片等)特化成翅状的各类果实(Eriksson & Bremer, 1992; Friis et al., 2011; van der Niet & Johnson, 2012)。
化石证据表明, 早在晚中新世的陉属(Fraxinus)中就出现了翅果(王磊等, 2012), 第三纪出现了槭属(Acer)植物翅果(黄永江等, 2013), 说明翅果在被子植物演化早期就已出现(Eriksson et al., 2000)。翅果在果翅形态、数量与着生位置等方面均表现出极大的多样性(Augspurger, 1986, 1988; Manchester & O’Leary, 2010), 具有不同的演化历史和适应意义。
前人对翅果的风力传播机制、运动方式与演化发育等方面开展了一些实验研究(Norberg, 1973; Augspurger, 1986, 1988; Seter & Rosen, 1992; Manchester & O’Leary, 2010), 但有关被子植物翅果类型的多样性、适应意义及其在系统分类中的价值还缺乏针对性的分析和归纳, 对不同类型翅果在适应风力传播等功能上的差异也缺少实验研究和系统总结。本文总结了广义翅果的主要类型及其系统分布、演化历史与适应意义, 并分析了各类翅果的运动方式与扩散能力以及翅果类型在植物经典分类中的价值, 以期归纳出翅果的多样性及其在被子植物扩散与物种分化历史中的可能作用, 并指出今后进一步研究的方向。
1 翅果的类型
根据果翅形态、着生位置及发育来源的不同, 本文把翅果分为6大类: 单侧翅果、周位翅果、披针翅果、棱翅果、翼状萼翅果、叶状苞翅果(图1)。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1被子植物翅果的6种主要类型。
Fig. 1Six types of samaras of angiosperms.
(Green, 1980; Augspurger, 1986; Peck & Lersten, 1991)。
1.1 单侧翅果(single-winged samara)
单侧翅果是指果实只有一个狭长果翅, 且果翅位于果实的单侧(图1)。单侧翅果是被子植物最简单的翅果类型, 也是目前实验研究最多的翅果类型 单侧翅果通常是不开裂干果, 具1种子, 可简称为“单翅果” (Seter & Rosen, 1992)。单翅果主要出现在木犀科陉属(Fraxinus)、梧桐科银叶树属(Heritiera)、远志科蝉翼藤属(Securidaca)、鼠李科翼核果属(Ventilago)和榆科刺榆属(Hemiptelea)等木本植物中(表1)。Table 1
表1
表1被子植物翅果主要类型及其系统分布
Table 1
| 翅果类型 Samara type | 主要分布类群 Systematic distribution |
|---|---|
| 1 单侧翅果 Single-winged samara (SWS) | |
| 1.1 单翅果 Mono-SWS | 金虎尾科(异翅藤属, Acridocarpus, Banisteriopsis, Barnebya, Bronwenia, Cordobia, Cottsia, Dinemagonum, Diplopterys, Ectopopterys, Janusia, Peixotoa, Sphedamnocarpus, Stigmaphyllon), 木犀科陉属, 远志科蝉翼藤属, 鼠李科翼核果属, 榆科刺榆属, 梧桐科银叶树属 Malpighiaceae (Heteropterys, Acridocarpus, Banisteriopsis, Barnebya, Bronwenia, Cordobia, Cottsia, Dinemagonum, Diplopterys, Ectopopterys, Janusia, Peixotoa, Sphedamnocarpus, Stigmaphyllon), Fraxinus (Oleaceae), Securidaca (Polygalaceae), Ventilago (Rhamnaceae), Hemiptelea (Ulmaceae), Heritiera (Sterculiaceae) |
| 1.2 双聚单翅果 Di-SWS | 槭树科槭树属, 木犀科陉属 Acer (Aceraceae), Fraxinus (Oleaceae) |
| 1.3 三聚单翅果 Tri-SWS | 金虎尾科翅实藤属, 异翅藤属 Malpighiaceae (Ryssopterys, Heteropterys) |
| 2 周位翅果 Perigynous samara | |
| 2.1 圆翅果 Round-winged samara | 金虎尾科(盾翅藤属, Alicia, Amorimia, Calcicola, Caucanthus, Christianella, Diaspis, Excentradenia, Flabellaria, Madagasikaria, Malpighiodes, Mascagnia, Mezia), 胡桃科(青钱柳属, 化香树属), 豆科(紫檀属, 黄檀属), 十字花科(菘蓝属, 屈曲花属, 菥属), 桦木科(桤木属, 桦木属), 榆科(榆属, 青檀属), 蓼科山蓼属, 木樨科雪柳属, 槭树科金钱槭属, 苦木科臭椿属, 杜仲科杜仲属, 芸香科榆橘属, 蓝果树科喜树属, 莲叶桐科青藤属, 鼠李科马甲子属, 马尾树科马尾树属 Malpighiaceae (Aspidopterys, Alicia, Amorimia, Calcicola, Caucanthus, Christianella, Diaspis, Excentradenia, Flabellaria, Madagasikaria, Malpighiodes, Mascagnia, Mezia), Juglandaceae (Cyclocarya, Platycarya), Fabaceae (Pterocarpus, Dalbergia), Cruciferae (Isatis, Iberis, Thlaspi), Betulaceae (Alnus, Betula), Ulmaceae (Ulmus, Pteroceltis), Oxyria (Polygonaceae), Fontanesia (Oleaceae), Dipteronia (Aceraceae), Ailanthus (Simaroubaceae), Eucommia (Eucommiaceae), Ptelea (Rutaceae), Camptotheca (Nyssaceae), Illigera (Hernandiaceae), Paliurus (Rhamnaceae), Rhoiptelea (Rhoipteleaceae) |
| 2.2 蝶翅果 Butterfly-winged samara | 金虎尾科(Adelphia, Aenigmatanthera, Hiraea, Amorimia, Callaeum, Carolus, Christianella, Dinemandra等), 菊科(金鸡菊属, 偶雏菊属, 蟛蜞菊属), 木犀科(雪柳属, 六道木叶属), 使君子科(诃子属, 榆绿木属, 假榆橘属) Malpighiaceae (Adelphia, Christianella, Aenigmatanthera, Hiraea, Amorimia, Callaeum, Carolus, Dinemandra, etc.), Asterceae (Coreopsis, Boltonia, Sphagneticola), Oleaceae (Fontanesia, Abeliophyllum), Combretaeeae (Terminalia, Anogeissus, Pteleopsis) |
| 3 棱翅果 Rib-winged samara | 金虎尾科(Aspicarpa, Calcicola, Digoniopterys), 蓼科(翅果蓼属, 红药子属, 大黄属, 沙拐枣属, 等), 卫矛科(雷公藤属, 卫矛属, 等), 使君子科(风车子属, 诃子属), 无患子科(车桑子属, 黄梨木属), 椴树科(滇桐属, 蚬木属, 柄翅果属, 一担柴属), 鼠李科咀签属, 莲叶桐科青藤属, 茶茱萸科心翼果属, 薯蓣科薯蓣属, 藜科四翅滨藜, 胡颓子科翅果油树 Malpighiaceae (Aspicarpa, Calcicola, Digoniopterys), Polygonaceae (Parapteropyrum, Pteroxygonum, Rheum, Calligonum etc.), Celastraceae (Tripterygium, Euonymus etc.), Sapindaceae (Dodonaea, Boniodendron), Tiliaceae (Craigia, Excentrodendron, Burretiodendron, Colona), Gouania (Rhamnaceae), Illigera (Hernandiaceae), Cardiopteris (Icacinaceae), Dioscorea (Dioscoreaceae), Atriplex canescens (Chenopodiaceae), Elaeagnus mollis (Elaeagnaceae) |
| 4 披针翅果 Lanceolate-winged samara | 金虎尾科(风筝果属, 三星果属, Lophopterys, Dicella, Microsteira, Niedenzuella, Rhynchophora, Tetrapterys), 椴树科六翅木属, 卫矛科斜翼属, 十字花科沙芥属, 使君子科萼翅藤属 Malpighiaceae (Hiptage, Tristellateia, Lophopterys, Dicella, Microsteira, Niedenzuella, Rhynchophora, Tetrapterys), Berrya (Tiliaceae), Plagiopteron (Celastraceae), Pugionium (Crucificeae), Getonia (Combretaeeae) |
| 5 翼状萼翅果 Sepal-winged samara | 龙脑香科, 钩枝藤科钩枝藤属, 旋花科(飞蛾藤属, 三翅藤属, 白花叶属, 地旋花属) Dipterocarpaceae, Ancistrocladus (Ancistrocladaceae), Convolvulaceae (Dinetus, Tridynamia, Poranopsis, Xenostegia) |
| 6 叶状苞翅果 Bract-winged samara | 桦木科(鹅耳枥属, 铁木属, 桦木属, 桤木属, 榛属), 胡桃科(黄杞属, 枫杨属), 败酱科败酱属, 旋花科盾苞藤属, 马鞭草科楔翅藤属, 檀香科米面蓊属 Betulaceae (Carpinus, Ostrya, Betula, Alnus, Corylus), Juglandaceae (Engelhardia, Pterocarya), Patrinia (Valerianaceae), Neuropeltis (Convolvulaceae), Sphenodesme (Verbenaceae), Buckleya (Santalaceae) |
新窗口打开|下载CSV
单翅果有时会两个或三个聚合在一起, 形成双聚单翅果和三聚单翅果等类型(图1)。双聚单翅果为2枚相连的小坚果, 侧面有长翅, 张开形成各种角度, 主要出现在槭树科槭属(Xu et al., 2008)和金虎尾科Ectopopterys属(Anderson et al., 2006)中。三聚单翅果指果实具3种子、每种子具1翅, 出现在Acer trialatum (邓莉兰等, 2003)以及金虎尾科亚洲特有属翅实藤属(Ryssopterys)和美洲特有属Janusia与Stigmaphyllon等属(表1)。
1.2 周位翅果(perigynous samara)
周位翅果的果翅环绕在果实四周, 呈完整的圆形、椭圆或宽条形, 或果翅略有收缩形成有缺口的扇形(图1)。周位翅果是果翅面积最大、最显眼的一类翅果。根据果翅的形状及有无缺口, 周位翅果又可分为圆翅果(round-winged samara)和蝶翅果(butterfly-winged samara)两大类(图1)。圆翅果的翅呈完全闭合圆形或椭圆形包围果实, 果翅在一个平面上(图1), 因此也被称为“平面翅果” (plain winged fruit; Burrows, 1975)。圆翅果的分布较广, 集中出现在胡桃科、榆科和金虎尾科等近20个科中, 特别是杜仲科杜仲属(Eucommia)、胡桃科青钱柳属(Cyclocarya)、豆科紫檀属(Pterocarpus)和黄檀属(Dalbergia)、榆科榆属(Ulmus)和青檀属(Pteroceltis)、苦木科臭椿属(Ailanthus)以及金虎尾科盾翅藤属(Aspidopterys)等是非常典型的圆翅果。
藜科猪毛菜属(Salsola)翅果比较特殊, 果翅由花被及其附属物特化而来(Yamaguchi et al., 1990; Wei et al., 2008)。这类植物的花被边缘膜质, 果时变硬、背面中下部生翅; 果翅5个, 其中3个为肾形、膜质; 另2个较小、倒卵形。这些果翅围绕果实一周并处于一个平面, 可视为特殊的圆翅果类型。
有些周位翅果的果翅存在着对称的收缩或出现缺口, 整体呈蝶翅状(图1), 可称为“蝶翅果”。蝶翅果主要分布在木犀科雪柳属(Fontanesia; Chang et al., 1996)与六道木叶属(Abeliophyllum)、菊科金鸡菊属(Coreopsis)等(Shi et al., 2011)以及金虎尾科美洲和非洲类群中(Anderson et al., 2006)。
1.3 棱翅果(rib-winged samara)
棱翅果的果翅呈棱状或薄片状, 贯通果实的上下两端, 多为3-5个, 均匀分布在果实周围(图1)。该类翅果主要分布在卫矛科雷公藤属(Tripterygium)和卫矛属(Euonymus)、薯蓣科薯蓣属(Dioscorea)和蓼科的大部分物种中(表1)。卫矛属棱翅果的外形、果翅数量与形态变化均较大, 果翅有的宽短、有的狭长、有的末端呈钩状(Ma et al., 2008)。如短翅卫矛(Euonymus rehderianus)果实近扁球状, 果翅5, 宽短; 藏南卫矛(E. austrotibetanus)具4条2-5 mm长的窄翅; 纤齿卫矛(E. giraldii) 4翅, 但翅极短, 顶端圆钝; 角翅卫矛(E. cornutus) 4或5翅, 翅长可达5-10 mm, 向尖端渐窄, 微呈钩状; 征镒卫矛(E. wui) 4翅, 倒卵形, 长14 mm, 顶端具细尖。此外, 该属刺果卫矛(E. acanthocarpus)、星刺卫矛(E. actinocarpus)和棘刺卫矛(E. echinatus)等物种的球状果实密被棕色细刺(Ma et al., 2008), 可能与棱形果翅有着相同的来源。卫矛科雷公藤属棱翅果的果实外形与果翅形态变化较小, 都为3片宽大的膜质翅(Ma et al., 2008)。
蓼科植物的果实虽然较小, 但仍是典型的棱翅果(Li et al., 2003)。如大黄属(Rheum)的瘦果三棱状、棱缘具翅, 翅上各具1条明显纵脉; 翅果蓼(Parapteropyrum tibeticum)的果翅薄膜质, 淡红色, 边缘近全缘; 红药子(Pteroxygonum giraldii)翅果具3锐棱, 沿棱具黄褐色膜质翅, 基部具3个黑色角状附属物(Li et al., 2003)。
椴树科蚬木属(Excentrodendron)和柄翅果属(Burretiodendron)的棱翅果通常具5个薄膜状的果翅(Tang et al., 2007)。胡颓子科的翅果油树(Elaeagnus mollis)的棱翅数量非常多, 常呈8棱脊分布在多毛的圆形或阔椭圆形果实周围(Qin & Gilbert, 2007)。使君子科风车子属(Combretum)和诃子属(Terminalia)的棱翅果既有3棱、也有5棱的果翅(任明迅, 个人观察)。这些现象表明, 棱翅果的棱翅数量与形态在科属水平具有保守性, 有一定的分类学价值, 但也存在发育原因等导致的种内变异(Al-Mayah & Stace, 1989; Manchester & O’ Leary, 2010)。
1.4 披针翅果(lanceolate-winged samara)
披针翅果指果实具不少于3个狭长呈披针状的果翅(图1)。披针翅果主要分布在钩枝藤科钩枝藤属(Ancistrocladus)、使君子科萼翅藤属(Getonia)、金虎尾科亚洲和非洲类群如风筝果属(Hiptage)及三星果属(Tristellateia)等属(表1)。披针翅果的果翅形态与数量变化较大, 有扁长如螺旋桨片的典型披针状果翅, 如金虎尾科风筝果属, 也有较为细长呈针状的果翅, 如金虎尾科三星果属(Anderson et al., 2006)。椴树科六翅木属(Berrya)与黄麻属(Corchorus)的蒴果每个果室具2翅, 果翅横直, 呈车轮的辐状或风车状(图1); 斜翼属(Plagiopteron)蒴果三角状陀螺形, 3个披针状果翅水平或斜立于果实一端形似降落伞状(Tang et al., 2007)。
与其他翅果类型相比, 披针翅果的果翅通常细长、数量较多, 多呈辐射状均匀排列在果实的赤道面(图1), 有时也全部位于果实一侧(Tang et al., 2007)。
1.5 翼状萼翅果(sepal-winged samara)
有些翅果的果翅不是来自于果皮或种皮的延伸, 而是由萼片或苞片发育而来。如热带地区的优势乔木龙脑香科(Dipterocarpaceae)和钩枝藤科(Ancistrocladaceae)一些物种的花萼筒与果实愈合在一起, 萼片增大, 特化成翅状。这类翅果可称为“翼状萼翅果” (图1)。在钩枝藤科中, 花萼的5个裂片都增大成翅状, 呈不等大的倒卵状匙形, 最大者长4.5 cm、宽1.6 cm, 顶端圆, 有较明显的脉纹; 最小的长不到 2 cm、宽不到1 cm (Wang & Gereau, 2007)。旋花科飞蛾藤属(Dinetus)植物蒴果的5个萼片均增长呈螺旋桨状, 外形类似披针翅果; 但由于果翅来源于萼片, 果翅均位于蒴果下方(Fang & Staples, 1995)。三翅藤属(Tridynamia)、白花叶属(Poranopsis)和地旋花属(Xenostegia)等蒴果的5个萼片不等大增长, 通常位于外方的2或3个萼片显著增大成翅状, 每果翅具9条左右、背面凸起的纵向脉; 而盒果藤属(Operculina)和猪菜藤属(Hewittia)蒴果的萼片则宽展成花瓣状(Fang & Staples, 1995)。
龙脑香科有16属、500多种, 广布于亚洲热带地区, 均具翼状萼翅果(Li et al., 2007)。龙脑香科植物的翼状萼翅果变化较大: 龙脑香属(Dipterocarpus)、坡垒属(Hopea)及青梅属(Vatica)的2枚花萼裂片增大为翅状, 但不伸展, 相互靠合在一起, 呈彗星拖尾状位于果实的一侧(图1); 娑罗双属(Shorea)花萼发育成3长2短的翅; 柳安属(Parashorea)的5个果翅等长或其中3枚较大(Li et al., 2007)。
1.6 叶状苞翅果(bract-winged samara)
叶状苞翅果的果翅由花苞片发育而来, 通常是苞片特化和增大呈叶片状位于整个果实的一侧(图1), 主要分布在败酱科败酱属(Patrinia)、胡桃科黄杞属(Engelhardia)和枫杨属(Pterocarya)等, 马鞭草科楔翅藤属(Sphenodesme)花序的总苞也呈翅状。胡桃科黄杞属的叶状苞翅果的果翅膜质弯曲、3深裂, 中裂片显著长于两侧的裂片(Lu et al., 1999); 枫杨属的坚果基部具1宿存的鳞状苞片及2个苞片发育而来的革质翅, 翅向果实两侧斜上方伸展, 呈元宝状(Lu et al., 1999); 枫杨属的翅果在外形上酷似双聚单翅果, 但后者的果翅来源于果皮、由两个单侧翅果靠生在一起形成, 区别于枫杨属的单个果实2枚苞片形成的果翅(Lu et al., 1999)。檀香科米面蓊属(Buckleya)的叶状苞也非常显眼, 往往是果实长度的3-4倍(Xia & Gilbert, 2003)。
败酱科败酱属的果实本身为瘦果, 但苞片膨大呈翅状包围整个瘦果, 使整个果实为功能性“翅果”。其叶状苞翅状, 通常具2-3条主脉, 网脉明显。如西伯利亚败酱(Patrinia sibirica)的叶状苞倒卵形或倒卵状长圆形, 长6-9 mm、宽4.5-6.5 mm, 先端圆钝或三浅裂, 羽状网脉明显, 常具3主脉(Hong et al., 2011)。
桦木科的鹅耳枥(Carpinus turczaninowii)叶状苞翅果较为典型, 苞片半卵形、半长圆形或半宽卵形, 长近2 cm, 外缘具缺齿、内缘全缘或疏生细齿, 基部具卵形、内折裂片(Li & Skvortsov, 1999)。榛属(Corylus)的果苞总状或管状, 有的种类顶端裂片硬化成针刺状; 坚果近球形, 大部分或全部为果苞所包。桤木(Alnus cremastogyne)果苞木质, 鳞片状, 宿存, 由3枚苞片和2枚小苞片愈合而成, 顶端具5枚浅裂片, 膜质翅宽为果实的1/2 (图1)。桦木科叶状苞翅果的果翅形状与大小以及裂片有无和开裂方式是形态分类的一个主要依据(Li & Skvortsov, 1999)。
旋花科盾苞藤属(Neuropeltis)的苞片紧贴萼下, 在果期显著增大呈宽椭圆形或圆形, 长3-5 cm, 除脉络外, 其他部分均被柔毛(Fang & Staples, 1995)。这种叶状苞翅果在外形上类似圆翅果, 但由于果翅来源于苞片, 整个果翅平面位于蒴果下方一侧, 而非圆翅果的果翅处于果实中部环绕一圈。
马鞭草科楔翅藤属是亚洲热带地区特有的攀援木质藤本, 其花序为头状聚伞花序, 具5-6枚匙状总苞片。果实成熟后, 所有苞片宿存、平展, 似风车状(Chen & Gilbert, 1994)。依靠这些翅状的总苞, 楔翅藤属果序作为一个整体进行传播, 可视为一类特殊的翅果。
总的来看, 果翅的数量、形态与着生位置在科属水平上非常稳定, 可以作为植物形态分类的重要依据(Friis et al., 2011)。如槭树科的槭属为双聚单翅果、金钱槭属为圆翅果; 金虎尾科的盾翅藤属为圆翅果、风筝果属具三条披针状扁平翅、三星果属则有着7条披针状窄翅(Anderson et al., 2006)。蓼科和卫矛科的棱翅果也以果翅数量与形态特征作为一个关键的分类学依据(Li et al., 2003; Ma et al., 2008)。虽然环境因素可能影响果实或种子形态和果翅发育(Jurado et al., 1991; Lord et al., 1997), 但在种一级水平翅果的类型也较为稳定, 也可以作为物种水平的分类依据。翅果也有较多的现存化石, 成为研究植物系统与演化的重要材料(Tanai, 1978, 1983; Eriksson et al., 2000; Manchester & O’ Leary, 2010; 王磊等, 2012; 黄永江等, 2013)。
2 翅果的演化
被子植物约有90个科具翅果(Dallwitz et al., 2000), 不同翅果类型在植物系统发育树上反复出现(图2), 表明翅果在被子植物中可能经历过多次独立的演化。翅果多出现在木本和藤本植物中, 而在草本植物中相对较少(图2; 表1)。具有翅果的科或属也普遍比没有翅果的近缘类群具有较高的物种多样性和较广的地理分布; 如龙脑香科较原始的非洲类群果实硕大无翅, 而扩散较远的类群如热带亚洲分布的娑罗双属、青皮属和坡垒属等均为翼状萼翅果, 尤其是物种数量多达200多种的娑罗双属都具典型的翼状萼翅果, 地理分布范围最广(张金泉和王兰洲, 1985; Gunasekara, 2004; Li et al., 2007)。因此, 翅果的出现有可能促进了被子植物的物种形成与分布区扩散。图2
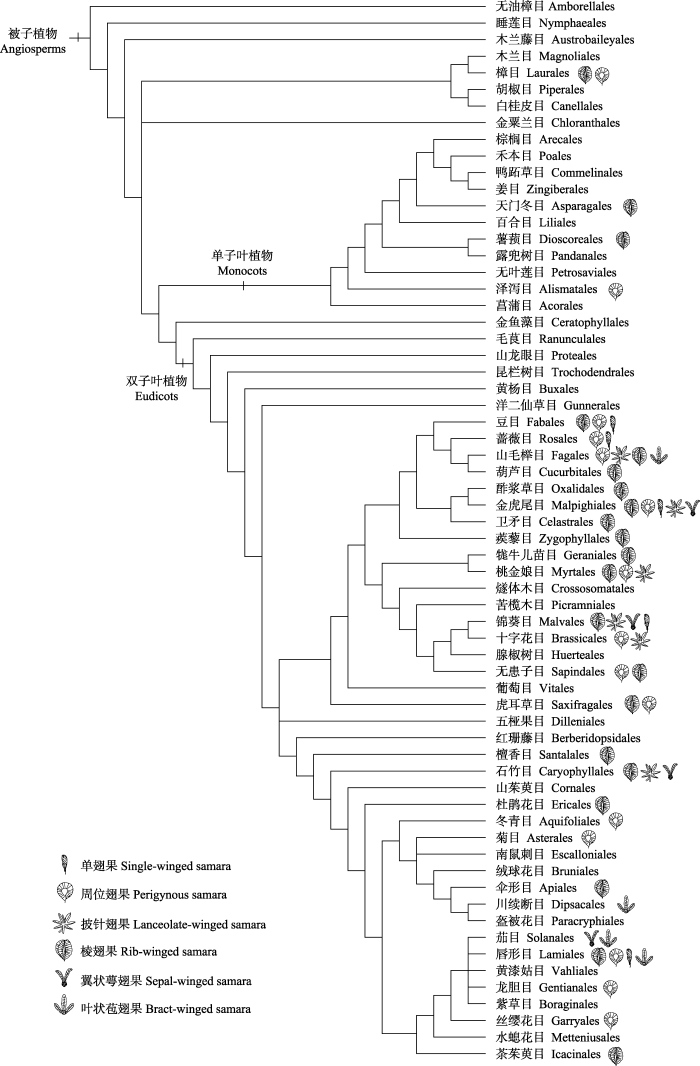 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2被子植物翅果在目一级的系统分布。系统树根据The Angiosperm Phylogeny Group (2016)绘制。
Fig. 2The systematic distribution of samara types across angiosperm orders. The phylogeny of angiosperm orders was drawn according to the Angiosperm Phylogeny Group (2016).
2.1 翅果的起源与演化
从被子植物的系统发育树看, 其基部类群樟目已出现了蝶翅果和棱翅果(图2)。随后, 逐渐出现了单翅果、叶状苞翅果、披针翅果和翼状萼翅果等(Friis et al., 2011)。翅果的整个演化过程呈现果翅数量增加、果翅偏向单侧、果翅负荷降低的趋势(图2), 可能有助于提高扩散距离和适应二次扩散等。单子叶植物的翅果发生频率与翅果类型较少, 仅在天门冬目和薯蓣目有棱翅果, 在泽泻目出现了圆翅果(图2), 这些特征与被子植物基部类群的翅果类型相似。单子叶植物很多果实是浆果或是胚乳丰富的干果, 主要通过吸引动物取食来传播果实与种子, 缺少借助风力传播的翅果。
在双子叶植物中, 基部类群毛茛目、昆栏树目与金鱼藻目并没有出现翅果, 但在物种多样性较高的类群如唇形目、豆目、金虎尾目和锦葵目中, 具有极高的翅果类型多样性(图3)。这些类群都是地理分布范围较广的类群, 与翅果能促进果实传播和物种扩散的特点相一致。在目一级水平上, 单翅果至少独立出现了4次, 披针翅果至少独立出现2次, 叶状苞翅果和翼状萼翅果各自至少独立出现3次; 而棱翅果与周位翅果可能发生过多次、反复的演化(图2)。说明翅果类型可能与物种生活史性状转变、地质变迁和生境类型转换等有关。
图3
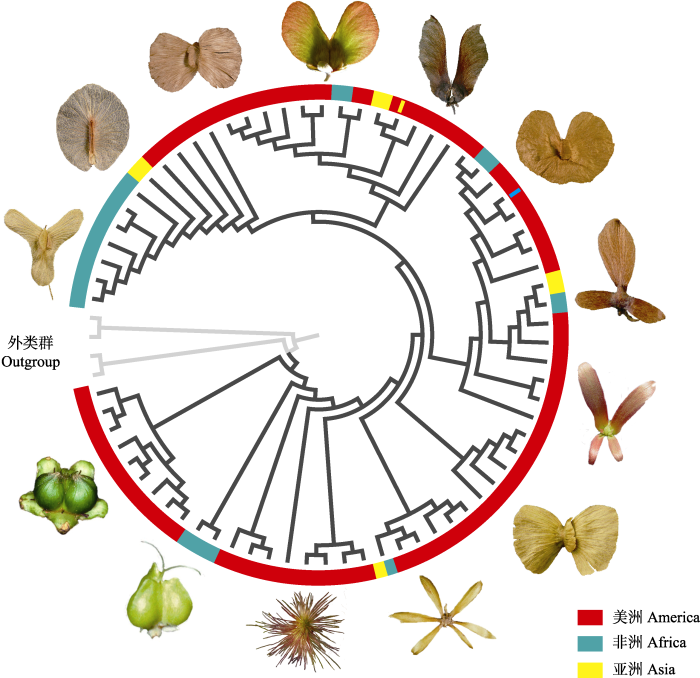 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3金虎尾科翅果类型的多样性与演化。在不同大洲之间的长距离扩散之后都伴随着翅果类型的变化。系统树改自Davis等(2014)。
Fig. 3Samara diversity and evolution in Malpighiaceae. Note there has evolutionary transition of samara types for each long-distance dispersal between continents. The phylogeny is redrawn from Davis et al. (2014).
2.2 “金虎尾路线”的翅果演化
金虎尾科是翅果类型最丰富的一个类群(Anderson et al., 2006; Manchester & O’ Leary, 2010), 除翼状萼翅果和叶状苞翅果外, 其余4大类翅果都在金虎尾科有大量分布(表1; 图3)。此外, 金虎尾科是生物地理学研究模式“金虎尾路线” (Malpighiaceae Route, Davis et al., 2002, 2014)的代表类群, 该科历史上每一次洲际长距离扩散往往都伴随着翅果类型的转变(Davis et al., 2001; 图3), 因此, 该类群可能是揭示翅果适应与演化的一个窗口。“金虎尾路线”认为, 金虎尾科起源于南美洲, 先后9次扩散到非洲和亚洲热带地区, 其中7次沿北大西洋路桥, 2次是近期发生的横渡大西洋扩散(Davis et al., 2002; Davis & Anderson, 2010; 钱贞娜和任明迅, 2016)。南美洲的金虎尾科原始类群均为非翅果, 这些类群的种子很可能通过鸟或水流等方式传播到历史上的中美洲和北美洲一带(Anderson et al., 2006)。随后翅果产生, 使金虎尾科植物可以沿着北大西洋路桥向东扩散至劳亚古陆, 直至现今的非洲和亚洲热带地区(Davis et al., 2001, 2002; 钱贞娜和任明迅, 2016)。在此过程中, 翅果类型多样性逐渐增加(图3; 表1)。
Carolus支系为整个金虎尾家族严格体现金虎尾路线完整扩散过程的支系, 在南美洲、中美洲、非洲以及亚洲都有亲缘关系很近的特有属(钱贞娜和任明迅, 2016)。该支系中, 南美类群为比较原始的核果和浆果(图3), 随后扩散至中美洲的类群演化为周位翅果, 最后扩散至非洲和亚洲。非洲类群(Flabellariopsis)虽不具典型的翅果, 但具有类似翅的果实附属物(Anderson et al., 2006; 图3)。亚洲特有的风筝果属则出现了披针翅果, 扩散能力较强, 使该类群广布于亚洲热带地区并演化出约30个种(Ren, 2015; 钱贞娜和任明迅, 2016)。从整个金虎尾科的翅果多样性及其系统分布(图3)看, 可以猜测金虎尾路线的翅果曾反复出现, 翅果类型发生多次转变。这可能是金虎尾科植物能够实现多次跨洋长距离扩散(Davis et al., 2001, 2002; 钱贞娜和任明迅, 2016)的一个关键因素。
3 翅果的适应意义
3.1 促进果实或种子的传播
Green (1980)研究了7个物种的翅果运动特征, 发现单侧翅果通过自旋转方式延长了在空中的时间, 即降低了翅果降落速度, 使扩散距离更远(图4), 证实了果翅确实借助风力促进了果实的传播。Augspurger (1986)发现, 翅果的降落速率与果实质量/果翅面积的平方根呈极显著相关关系(图4); 他把果实质量与果翅面积的比值称为“果翅负荷” (wing loading), 并作为衡量翅果风力传播能力的一个重要指标。图4
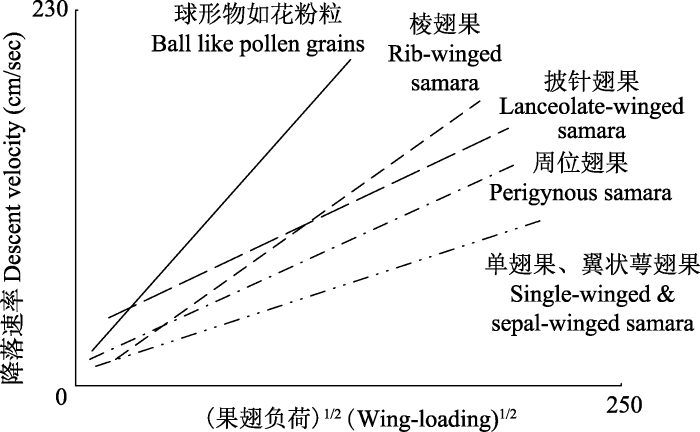 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4不同类型翅果的降落速率与果翅负荷(果实质量/果翅面积)的相关性。改自Augspurger (1986)。
Fig. 4Descent velocity and its relationship with wing loading (fruit mass / wing size) of samaras and pollen grains. Revised from Augspurger (1986).
Augspurger (1986, 1988)把翅果在空中的运动方式分为6类: 自旋式(autogyro)、翻滚自旋式(rolling autogyro)、波浪式(undulator)、直升机式(helicopter)、滚筒式(tumbler)以及不确定式(nonclassified)。自旋式翅果(如单翅果)在空中停留的时间最长, 但传播距离并不是最远的(Augspurger, 1986); 直升机式(如披针翅果、翼状萼翅果和叶状苞翅果等多个果翅位于果实一端)和波浪式(如周位翅果)的空中运动较为稳定, 扩散方向较明确, 是扩散距离较远的一类翅果(Augspurger, 1986; 图4)。翅果降落速率与传播距离不仅与果翅大小、运动方式有关, 还与整个果实的质量分配和水分含量有关(Augspurger, 1988)。
Cremer (1977)提出, 翅果的水平飞行距离可按照模型D = Vw(H/Vf)来进行估算。在该模型中, D是翅果的水平传播距离、Vw是风速、H是翅果降落时的高度、Vf是翅果降落的平均速度。
Vf与果翅负荷密切相关(图4)。由于果翅数量、形态及位置影响到整个翅果的空气动力学特征, 导致果翅对称的翅果(如周位翅果和棱翅果等)与不对称的翅果(单翅果、翼状萼翅果和叶状苞翅果等)有着不同的飞行特性与扩散距离。由于内在的不稳定性, 不对称翅果的传播距离理论上应该不及对称的翅果(Augspurger, 1986, 1988)。然而, Greene和Johnson (1990)发现, 不对称的单翅果(Acer negundo)和两侧对称的双聚单翅果(Fraxinus americana)有着相似的气流状态, 单翅果的自动旋转可以忽略, 两者均可当作重粒子以恒定的速度降落(Seter & Rosen, 1992)。这说明, 需要针对不同类型翅果的空中运动方式开展严谨的实验研究, 才能准确揭示各类翅果在适应风力传播方面可能不同的机制。
Planchuelo等(2016)还发现, 臭椿(Ailanthus altissima)的周位翅果在风力传播之后, 落在水面的臭椿翅果还可依靠宽大的周位翅漂于水面, 随水漂流进行二次扩散。因此, 臭椿的果翅可能同时具有风力传播与水漂传播的适应, 只不过果翅的大小受到了不同的选择: 较宽的果翅风阻较大, 不利于风力作用下的长距离散布, 但却可以增加在水面上的浮力, 促进了水漂传播(Planchuelo et al., 2016)。这可能是臭椿具有果翅大小不同的两类翅果(Delgado et al., 2009)的一个主要原因。水生植物水马齿属(Callitriche)果实的棱翅也有助于果实浮在水面, 促进了水漂传播(Lansdown, 2006)。
有些翅果如棱翅果与披针翅果的果翅均匀分布在果实四周, 可在落至土表之后再次在风吹或重力作用下成为“风滚翅果” (Burrows, 1994), 类似于干旱地区的“风滚植物” (tumble weed)(王静等, 2002; Baskin et al., 2014), 促进了翅果的二次搬运。
可见, 翅果不仅可在重力和风力共同作用下脱离植株实现第一次传播, 还可能在风、水流及动物携带等因素下在地面或水面上发生二次传播, 大大提高了翅果的扩散能力与繁殖成功率。但这种二次扩散的现象在以往的研究中并未得到重视, 其普遍性及在不同类型翅果中的相对重要性尚待进一步的实验研究。
3.2 机械保护作用
由于果翅是果皮的延伸或萼片与苞片的特化, 通常包围着果实, 具有保护果实与种子的作用。特别是周位翅果、棱翅果和披针状翅果等具有环绕翅果四周的果翅, 能够在果实与种子掉落地面时, 避免果实与种子直接接触地表, 可减轻果实在空中运动及落地时的机械损伤和降低地表微生物侵染果实的速率(Willis, 1973)。亚洲热带植物区系的优势类群龙脑香科种子大而重(图1), 翼状萼可减轻果实落地碰撞时所产生的冲击力, 对种子起到一定的保护作用(Willis, 1973; Corlett, 2009)。十字花科的沙芥属、菘蓝属植物和蓼科的沙拐枣属植物生长在沙漠地区, 它们的棱翅果与披针翅果依靠果翅使得种子不直接接触到高温的土表, 有利于其适应干旱高温的沙漠环境(Al-Shehbaz, 1986; Baskin et al., 2014)。
目前, 大多数关于翅果的研究没有考虑果翅可能存在的机械保护作用, 导致还很难界定果翅的物理保护功能在翅果中的普遍性及其重要性。
3.3 调节种子成熟与萌发策略
主要生长于海滨或荒漠盐碱地的藜科猪毛菜属有着由花被及其附属物增厚特化而成的果翅(Yamaguchi et al., 1990), 其果翅及其提取物对种子萌发有抑制作用, 延长了种子休眠时间(Takeno & Yamaguchi, 1991; Wei et al., 2008)。紫翅猪毛菜(Salsola affinis)的果翅虽然不影响水分进入果实和种皮, 但阻碍了胚根突破种皮, 导致翅果比人为去掉果翅的处理萌发率低(Wei et al., 2008)。无翅猪毛菜(Salsola komarovii)在水分较好的环境下产生有翅和无翅的两种果实(Yamaguchi et al., 1990; Wei et al., 2008), 但在水分胁迫的条件下只产生无翅果, 且其翅果比无翅果的休眠期短、种子萌发率更高(Yamaguchi et al., 1990; Takeno & Yamaguchi, 1991), 而无翅果的寿命更长。具圆翅果的胡桃科青钱柳(Cyclocarya paliurus)的种子常需隔年甚至2年后才能萌发(尚旭岚等, 2011)。实验研究发现, 青钱柳种子胚不存在休眠和后熟, 果皮及环绕的圆形果翅造成的机械束缚和含有的香草酸是延滞青钱柳种子萌发的主要原因(尚旭岚等, 2011)。这可能是青钱柳种子适应逆境的机制, 也是青钱柳得以从第四纪冰期存活下来的一个原因。
此外, 黑槭(Acer saccharum ssp. nigrum)果翅在发育早期具有一定的光合作用(Peck & Lersten, 1991), 这可能使得果翅直接向发育中的果实与种子提供了光合物质, 调控了果实与种子的成熟进度, 影响到种子休眠与萌发。槭树科、榆科、胡桃科的大部分翅果都是绿色, 但是, 这些绿色果翅是否具有光合作用的叶绿素、是否影响果实与种子发育, 还尚待研究。
因此, 果翅不仅仅是散布与保护果实与种子的一种适应, 还可能通过改变果皮的理化性质调控种子的成熟、休眠与萌发进程, 避免了种子集中萌发而造成的同胞竞争, 提高了植物适应逆境的能力(Takeno & Yamaguchi, 1991; Wei et al., 2008)。
4 结语
目前, 有关翅果形态与功能的研究还远不如植物繁殖器官“花”那样受人关注, 不同翅果类型可能具有的适应意义还缺乏有力的证据和明确区分, 翅果对被子植物多样性形成与维持的作用也未受到应有的重视。因此, 利用生态适应和演化-发育生物学方法, 研究不同类型翅果在适应风力传播方面的差异, 果翅特别是萼片或苞片来源的果翅的发育机制, 翅果不同类型的演化历史及其对植株物种多样性的影响是值得深入开展的一个课题。值得注意的是, 金虎尾科具有被子植物最为多样的翅果类型, 也是生物地理学研究模式“金虎尾路线” (Davis et al., 2002)的模式类群。该科历史上发生了多达9次的洲际长距离扩散, 大多伴随着翅果类型的转变。以金虎尾科为例, 对被子植物翅果适应与演化开展深入的实验研究与系统发育研究, 有望深入揭示翅果的适应意义及其对被子植物物种形成与扩散的作用。今后的相关研究中, 可以选择翅果类型多样性较高的类群, 对同科同属不同类型翅果的物种进行分子系统发育关系、物种分化速率、果翅发育机制等的比较研究, 揭示翅果类型的演化转变时间、适应机制及其对物种多样性水平的可能影响。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.2307/2399697URL [本文引用: 1]

Revision of the family Combretaceae for accounts in eight tropical American floras has revealed the following twelve novelties: Terminalia chicharronia subsp. neglecta (Bisse) Alwan & Stace, comb. nov.; T. chicharronia subsp. domingensis (Urban) Alwan & Stace, comb. nov.; T. chicharronia subsp. orientensis (Monach.) Alwan & Stace, comb. nov.; T. ramatuella Alwan & Stace, nom. nov.; T. virens (Spruce ex Eichler) Alwan & Stace, comb. nov.; T. crispialata (Ducke) Alwan & Stace, comb. nov.; T. steyermarkii Alwan & Stace, sp. nov.; T. kuhlmannii Alwan & Stace, sp. nov.; T. uleana Engl. ex Alwan & Stace, sp. nov.; T. eichleriana Alwan & Stace, nom. nov.; Bucida molineti (M. G贸mez) Alwan & Stace, comb. nov.; and Buchenavia amazonia Alwan & Stace, sp. nov.
DOI:10.5962/bhl.part.27392URL [本文引用: 1]
JOURNAL OF THEARNOLD ARBORETUMVOLUME 67 JULY 1986 NUMBER 3 THE GENERA OF LEPIDIEAE (CRUCIFERAE; BRASSICACEAE) IN THE SOUTHEASTERN UNITED STATES'2 IHSAN A. AL-SHEHBAZ3 Tribe Lepidieae A. P. de Candolle, Syst. Nat. 2: 151. 1821. Annual, biennial, or perennial herbs [sometimes subshrubs or shrubs], gla-brous or with simple or rarely stellate [or bifurcate or dendritic] trichomes.Inflorescence an ebracteate [or bracteate], corymbose raceme, usually elongatedin fruit; flowers few to many [rarely solitary on long scapes], actinomorphic orslightly [to strongly] zygomorphic, perfect [or very rarely imperfect and plantsmonoecious or dioecious]. Sepals erect to ascending or spreading, not saccate[or strongly saccate] at base. Petals present or absent, equal or unequal. Nectarglands 2, 4, or 6, distinct or connate, subtending [or surrounding] the bases of 'Prepared for the Generic Flora of the Southeastern United States, a long-term project made possibleby grants from the National Science Foundation and currently supported by BSR-8415769 (C. E.Wood, Jr., principal investigator), under which this research was completed, and BSR-8415637 (N. G.Miller, principal investigator). This account, the 11 th in the series, was begun under BSR-8111520(C. E. Wood, Jr., principal investigator). It follows the format established in the first paper (Jour.Arnold Arb. 39: 296-346. 1958) and continued to the present. The area covered by the Generic Floraincludes North and South Carolina, Georgia, Florida, Tennessee, Alabama, Mississippi, Arkansas,and Louisiana. The descriptions are based primarily on the plants of this area, with informationabout extraregional members of a family or genus in brackets [ ]. The references that I have notverified are marked with asterisks. I am most grateful to Carroll Wood for his help with many aspects of this paper, especially hiscritical review of the manuscript. I am variously indebted to Reed C. Rollins, Norton G. Miller,George K. Rogers, R. Dale Thomas, John W. Thieret, and Jimmy R. Massey, as well as to BarbaraNimblett, who typed the manuscript. I also wish to thank Elizabeth B. Schmidt and Stephen A.Spongberg for their editorial advice. The illustrations were made by Karen Stoutsenberger under earlier grants. Carroll Wood and/orKenneth R. Robertson prepared the material and supervised the illustrations. Fresh material wascontributed by Wood, and the fruits and seeds are from herbarium specimens in the Gray Herbariumor the Arnold Arboretum. 2For an account of the family and its tribes, see Al-Shehbaz, The tribes of Cruciferae (Brassicaceae)in the southeastern United States. Jour. Arnold Arb. 65: 343-373. 1984. 'Arnold Arboretum, Harvard University, 22 Divinity Avenue, Cambridge, Massachusetts 02138.锟 President and Fellows of Harvard College, 1986.Journal of the Arnold Arboretum 67: 265-311. July, 1986.
URL [本文引用: 13]
DOI:10.1002/j.1537-2197.1986.tb12048.xURL [本文引用: 8]
Morphological and aerodynamic traits affecting mean potential dispersal distance are quantified for wind-dispersed diaspores of tree species on Barro Colorado Island, Panama. The sample includes 34 species in 16 families and represents six aerodynamic groups. Mass and area (maximum cross section) each vary over six orders of magnitude among the species. In contrast, wing-loading, defined as weight divided by area, varies over only one order of magnitude, as does the rate of descent. While the regression of rate of descent on the square root of wing-loading is significant overall, the slopes vary significantly among five aerodynamic groups. At comparable wing-loading values, diaspores of fluffy kapok fall faster than four other aerodynamic groups and rolling autogyros fall faster than non-rolling autogyros. Assuming the diaspores are released from their typical tree height and experience a mean windspeed of 1.75 m sec-1, the expected mean dispersal distance varies among the 34 species from 22 to 194 m. Rate of descent is weakly correlated with shade tolerance of seedlings for a subset of 18 species; rate of descent is more strongly correlated with the log of dry mass of seed for all 34 species. Given these wide differences in dispersal potential, any generalizations about tropical trees that use wind dispersal are of dubious value.
DOI:10.1111/j.1469-8137.1988.tb04174.xURL [本文引用: 3]
Among 34 wind-dispersed tree species on Barro Colorado Island, Panama, the wet mass of diaspores ranges over six orders of magnitude, The seed mass as a percentage of diaspore mass ( S/D ) varies greatly among species from 14 to 94% with a mean value of 61% The mean percent moisture of diaspores is 10% no consistent differences occur between seed and non-seed components of diaspores in percent moisture. Wing-loading (weight/area) and hence dispersal capacity varies over one order of magnitude among these species and is correlated more highly with S/D than with percent moisture of diaspores. Compared to fruit diaspores, seed diaspores have less mass and a greater S/D , and are slightly more dehydrated. As a result, seed diaspores have lower wing-loading and greater dispersal capacity than fruit diaspores. The morphological/aerodynamic features of a diaspore also have a significant effect on the mass variables. Overall, the type of diaspore (fruit or seed) is of major importance in explaining mass differences of diaspores among these wind-dispersed species; morphological features and dehydration ability contribute secondarily.
DOI:10.1016/j.ppees.2014.02.004URL [本文引用: 1]
The cold deserts of northern Xinjiang Province in northwest China are rich in diaspore-heteromorphic species, and diaspore biology has been compared in more heteromorphic species native to this biogeoclimatic region than for any other region worldwide. Our primary purpose was to compare the dispersal ability and degree of dormancy in heteromorphic diaspores in Xinjiang desert plants via a review of the Chinese and English literature. We located literature on 20 heteromorphic species native to these deserts. Fourteen of the species are chenopods (Amaranthaceae). All 20 species are heterodiasporous (dimorphic, 14; trimorphic, 4; polymorphic, 2) annuals, and none is amphicarphic sensu stricto. Thirteen of the species are heterocarpic, six heterospermic, one amphi-basicarpic and none heteroarthrocarpic. Diaspores of 19 of the species differ in seed dormancy/germination. Thirteen of the 14 species to which both diaspore dispersal ability and degree of dormancy could be assigned had at least one morph with high (or relatively high) dispersal ability and low degree of dormancy and at least one with low dispersal ability and high (or relatively high) degree of dormancy. Conceptual models of the dynamics of seed dormancy of the two morphs for each of three dimorphic species suggest the ecological significance of heterodiaspory in the cold desert annuals. However, the ecological significance of seed/fruit heteromorphism needs to be more thoroughly demonstrated via long-term field studies that compare the life history/demography of plants derived from different morphs.
DOI:10.1080/0028825X.1994.10410366URL [本文引用: 1]
In Ahuriri Summit Bush Scenic Reserve, a 5 ha area of mixed angiosperm-podocarp forest, are 56 wild woody plant species, including five adventives. Lists are given for fruit types, seed numbers per fruit, fruit sizes, degree of fleshiness of the pericarp or accessory tissues of ripe fruit, fruit colours, and dispersal modes for the seeds. Of the 22 tree species, 86% have fruit with fleshy pericarps or accessory tissues; corresponding values for the 20 shrub species are 80%, and for the 14 vine species 50%. Most of these fleshy fruit (87%) are between 2 and 8 mm in diameter. The fruit of only four species exceed 10 mm in diameter. The most frequent fleshy fruit colours are black to dark purplish-black (40%), or red (25%). Some fruit are purple, blue, orange, yellow, or white.
DOI:10.1111/j.1469-8137.1975.tb01404.xURL
Seeds and fruits of widely differing sizes and shapes and which employ different flight styles in wind-borne movement during dispersal are discussed qualitatively in general aerodynamic terms, and grouped according to aerodynamic rather than botanical features. Most of these groups are considered separately, though not in any great detail, and the salient features of the differing motions belonging to each are described. The response of species, in particular to variations in vertical wind velocities, is mentioned as an important factor affecting-deposition and dispersal range, and how because of this response some forced deposition may be possible with the introduction of suitably shaped obstacles to wind flow.
In: Wu ZY, Raven PH, Hong DY eds.
[本文引用: 1]
In: Wu ZY, Raven PH, Hong DY eds.
[本文引用: 2]
DOI:10.1111/j.1744-7429.2009.00503.xURL [本文引用: 1]
Most predictions of vegetation responses to anthropogenic climate change over the next 100 yr are based on plant physiological tolerances and do not account for the ability of plant species to migrate over the distances required in the time available, or the impact of habitat fragmentation on this ability. This review assesses the maximum routine dispersal distances achievable in tropical East Asia and their vulnerability to human impacts. Estimates for various plant090009vector combinations range from < 10 m, for species dispersed by ants or mechanical means, to > 10 km for some species dispersed by wind (tiny seeds), water, fruit pigeons, large fruit bats (tiny seeds), elephants, rhinoceroses, and people. Most plant species probably have maximum dispersal distances in the 1000900091000 m range, but the widespread, canopy-dominant Dipterocarpaceae and Fagaceae are normally dispersed < 100 m. Large fruit bats and fruit pigeons are particularly important for long-distance dispersal in fragmented landscapes and should be protected from hunting. The maximum seed dispersal distances estimated in this study are potentially sufficient for many plant species to track temperature changes in steep topography, but are far too small for a significant role in mitigating climate change impacts in the lowlands, where temperature and rainfall gradients are much more shallow.
URL [本文引用: 5]
DOI:10.3732/ajb.1000146URLPMID:21616850 [本文引用: 1]
Abstract 61 PREMISE OF THE STUDY: The Malpighiaceae include 651300 tropical flowering plant species in which generic definitions and intergeneric relationships have long been problematic. The goals of our study were to resolve relationships among the 11 generic segregates from the New World genus Mascagnia, test the monophyly of the largest remaining Malpighiaceae genera, and clarify the placement of Old World Malpighiaceae. 61 METHODS: We combined DNA sequence data for four genes (plastid ndhF, matK, and rbcL and nuclear PHYC) from 338 ingroup accessions that represented all 77 currently recognized genera with morphological data from 144 ingroup species to produce a complete generic phylogeny of the family. 61 KEY RESULTS AND CONCLUSIONS: The genera are distributed among 14 mostly well-supported clades. The interrelationships of these major subclades have strong support, except for the clade comprising the wing-fruited genera (i.e., the malpighioid+Amorimia, Ectopopterys, hiraeoid, stigmaphylloid, and tetrapteroid clades). These results resolve numerous systematic problems, while others have emerged and constitute opportunities for future study. Malpighiaceae migrated from the New to Old World nine times, with two of those migrants being very recent arrivals from the New World. The seven other Old World clades dispersed much earlier, likely during the Tertiary. Comparison of floral morphology in Old World Malpighiaceae with their closest New World relatives suggests that morphological stasis in the New World likely results from selection by neotropical oil-bee pollinators and that the morphological diversity found in Old World flowers has evolved following their release from selection by those bees.
DOI:10.1073/pnas.102175899URLPMID:11983870 [本文引用: 4]
Explanations for biogeographic disjunctions involving South America and Africa typically invoke vicariance of western Gondwanan biotas or long distance dispersal. These hypotheses are problematical because many groups originated and diversified well after the last known connection between Africa and South America ( 105 million years ago), and it is unlikely that "sweepstakes" dispersal accounts for many of these disjunctions. Phylogenetic analyses of the angiosperm clade Malpighiaceae, combined with fossil evidence and molecular divergence-time estimates, suggest an alternative hypothesis to account for such distributions. We propose that Malpighiaceae originated in northern South America, and that members of several clades repeatedly migrated into North America and subsequently moved via North Atlantic land connections into the Old World during episodes starting in the Eocene, when climates supported tropical forests. This Laurasian migration route may explain many other extant lineages that exhibit western Gondwanan distributions.
DOI:10.1073/pnas.1403157111URLPMID:24706921 [本文引用: 4]
Abstract Many major branches in the Tree of Life are marked by stereotyped body plans that have been maintained over long periods of time. One possible explanation for this stasis is that there are genetic or developmental constraints that restrict the origin of novel body plans. An alternative is that basic body plans are potentially quite labile, but are actively maintained by natural selection. We present evidence that the conserved floral morphology of a species-rich flowering plant clade, Malpighiaceae, has been actively maintained for tens of millions of years via stabilizing selection imposed by their specialist New World oil-bee pollinators. Nine clades that have lost their primary oil-bee pollinators show major evolutionary shifts in specific floral traits associated with oil-bee pollination, demonstrating that developmental constraint is not the primary cause of morphological stasis in Malpighiaceae. Interestingly, Malpighiaceae show a burst in species diversification coinciding with the origin of this plant-pollinator mutualism. One hypothesis to account for radiation despite morphological stasis is that although selection on pollinator efficiency explains the origin of this unique and conserved floral morphology, tight pollinator specificity subsequently permitted greatly enhanced diversification in this system.
DOI:10.2307/3558360URLPMID:21669617 [本文引用: 1]
The Malpighiaceae are a family of ~1250 species of predominantly New World tropical flowering plants. Infrafamilial classification has long been based on fruit characters. Phylogenetic analyses of chloroplast DNA nucleotide sequences were analyzed to help resolve the phylogeny of Malpighiaceae. A total of 79 species, representing 58 of the 65 currently recognized genera, were studied. The 3 region of the gene ndhF was sequenced for 77 species and the noncoding intergenic spacer region trnL-F was sequenced for 65 species; both sequences were obtained for the outgroup, Humiria (Humiriaceae). Phylogenetic relationships inferred from these data sets are largely congruent with one another and with results from combined analyses. The family is divided into two major clades, recognized here as the subfamilies Byrsonimoideae (New World only) and Malpighioideae (New World and Old World). Niedenzu's tribes are all polyphyletic, suggesting extensive convergence on similar fruit types; only de Jussieu's tribe Gaudichaudieae and Anderson's tribes Acmanthereae and Galphimieae are monophyletic. Fleshy fruits evolved three times in the family and bristly fruits at least three times. Among the wing-fruited vines, which constitute more than half the diversity in the family, genera with dorsal-winged samaras are fairly well resolved, while the resolution of taxa with lateral-winged samaras is poor. The trees suggest a shift from radially symmetrical pollen arrangement to globally symmetrical pollen at the base of one of the clades within the Malpighioideae. The Old World taxa fall into at least six and as many as nine clades.
URLPMID:20121015
Abstract We have specifically carried out a greenhouse experiment to assess relationship between samara weight and seed success. Relationship assessed as dispersal potential, germination level, germination rate and early seedling mass for the invasive species Ailanthus altissima. For this purpose, we considered two close stands as seeds source. We found no correlation between samara size and neither germination level, germination rate, nor seedling mass, but a positive correlation with samara projected area. These results suggest that samara weight is not directly related to germination, dispersal and invasion potential neither. Nevertheless, stands differed in the invasion potential of their samaras; one stand presented samaras with higher projected area per weight unit whereas the other one presented samaras that produced heavier seedlings. Whatever the origin, (genetic or environmental) of this differences it should be advantageous for a colonizing invader species such as A. altissima since it could imply a wider range of habitats susceptible to invasion.
DOI:10.3969/j.issn.2095-0845.2003.02.017URL [本文引用: 2]
篇首: 本种与三角槭Acer buergerianum Miq.的区别为:果为具3翅的坚果,稀为双翅果或4翅果,小坚果连翅较短,长1.5~2cm,叶较短,长2~5cm,易于区别.
DOI:10.3969/j.issn.2095-0845.2003.02.017URL [本文引用: 2]
篇首: 本种与三角槭Acer buergerianum Miq.的区别为:果为具3翅的坚果,稀为双翅果或4翅果,小坚果连翅较短,长1.5~2cm,叶较短,长2~5cm,易于区别.
DOI:10.1111/j.1558-5646.1992.tb02000.xURLPMID:28564968 [本文引用: 1]
From southeastern Queensland to southern Victoria over a transect of 11-degrees latitude, the Moreton taxon of the Australian grasshopper Caledia captiva exhibits a cline in chromosome structure that involves change from a metacentric to an acrocentric genome. In this study, we show that embryonic development time covaries with chromosome structure along the transect. Both development time and chromosome short arm length exhibit an overall negative correlation with latitude, but with maxima just south of the northern limit of the taxon's distribution. Selection for such a pattern appears to arise from changes in voltinism along the cline in season length that exists along the transect. Populations with the highest temperature thresholds for avoidance of embryonic diapause also have the slowest development time and probably represent the northern extreme of a primarily univoltine life cycle. North of this region bivoltinism increases in frequency and, as expected from a split of the season length, development time decreases. Maximum chromosome short arm length occurs in the vicinity of the northern univoltine populations, rather than at the limit of distribution where bivoltinism prevails. We conclude that variation in chromosome structure could be contributing to the heritable variation for development time that forms the basis for adaptive change in this trait. These results provide justification for investigating causal relationships between chromosome structure and development time, with an ultimate aim of understanding the adaptive significance of chromosomal variation in C. captiva.
DOI:10.1086/303367URLPMID:10824020 [本文引用: 1]
Fossil data from 25 angiosperm floras from the Early Cretaceous (65124 million years ago) to the Pliocene (652 million years ago) were compiled to estimate sizes of seeds and fruits and the relative proportion of two different seed‐dispersal systems by animals and by wind. The results suggest that, first, seed and fruit sizes were generally small during most of the Cretaceous, in agreement with previous suggestions, but the trend of increasing sizes started before the Cretaceous‐Tertiary boundary; second, there was a decrease in both seed and fruit sizes during late Eocene and Oligocene, reaching a level that has continued to the Late Tertiary; third, the fraction of animal dispersal was, in contrast to previous suggestions, rather high also during the Cretaceous but increased drastically in the Early Tertiary and declined congruently with the declining seed and fruit sizes from the late Eocene; and fourth, the fraction of wind dispersal showed a bimodal pattern, being high in the Late Cretaceous and in the Oligocene‐Miocene but with a drop in between. We find that the observed trends are only weakly related to the availability of animal fruit dispersers. Instead, the trends are congruent with a climate‐driven change in environmental conditions for recruitment, where larger seeds are favored by closed forest vegetation. The prevalence of semiopen, dry, and probably herbivore‐disturbed vegetation during the Cretaceous, the development of closed multistratal forests in the Eocene, and the later development of a more open vegetation and grasslands starting in the Oligocene‐Miocene, are reflected in the distribution of angiosperm seed and fruit sizes and in the dispersal systems.
In: Wu ZY, Raven PH, Hong DY eds.
[本文引用: 6]
[本文引用: 3]
DOI:10.1002/j.1537-2197.1980.tb07754.xURL [本文引用: 1]
Strobe photographs were taken of over 200 spinning samaras from seven species of trees. These were used to measure the rate of descent, angular velocity, orientation, and other parameters of the samaras as they fell. These data were then used to compare the aerodynamic behavior of samaras, helicopters, and theoretical ideal rotors. Plotting morphological data for each samara against its rate of descent showed that this rate was highly correlated with the square root of the samara's wing loading (samara weight divided by wing-surface area). This plot demonstrated the existence of two distinct groups of samaras, distinguished by their morphology, spinning motion, and rate of descent. These results are of greatest use in characterizing local dispersal patterns.
DOI:10.1139/b90-340URL [本文引用: 1]
The application of micrometeorological models of heavy particle dispersal to winged diaspores (samaras) requires that the diaspores maintain stable flight and that descent velocity is constant with respect to vertical air motion. It has been argued that these assumptions may be invalid for autorotating samaras and that bilaterally symmetric samaras will disperse farther than asymmetric samaras ...
[本文引用: 1]
[本文引用: 1]
DOI:10.1086/284921URL [本文引用: 2]
Seed dispersal by animals has sometimes been considered one of the factors helping to explain angiosperm diversification. Three predictions related to this hypothesis are examined here, namely, (1) biotic seed dispersal is a feature unique to angiosperms, or this group exhibits it proportionally more often than gymnosperms; (2) extant groups exhibiting this feature tend to be taxonomically more diverse than sister groups lacking it; and (3) the timing of the appearance in the fossil record differs for biotically and abiotically dispersed angiosperm taxa. The analyses have been confined to endozoochory, the most genuine and widespread of the various biotic seed-dispersal methods, and conducted on virtually the entire set of extant gymnosperm and angiosperm families. None of the tests supports a role for animal dispersal per se in angiosperm diversification.
In: Wu ZY, Raven PH, Hong DY eds.
[本文引用: 1]
DOI:10.7677/ynzwyj201313073URL
选取我国西南地区槭属(槭树科)的8个现生种,通过观察和测量馆藏标本的翅果形态,对比同一个种的翅果在不同地区的形态差异性,并通过主成分分析(PCO)检测翅果在种内的形态变异性,发现槭属的翅果形态具有明显的地区间差异性,这种差异性主要表现在总体形状和大小上,这可能与地区间不同的气候环境特征有关。通过对槭属翅果种内形态变异性和多样性的深入研究,为槭属翅果化石的属下鉴定提供一定依据,指出槭属翅果化石的属下鉴定应该考虑翅果形态种内变异性的特征,具有形态多样性的该属翅果化石可能属于一个种或更少的种。
DOI:10.7677/ynzwyj201313073URL
选取我国西南地区槭属(槭树科)的8个现生种,通过观察和测量馆藏标本的翅果形态,对比同一个种的翅果在不同地区的形态差异性,并通过主成分分析(PCO)检测翅果在种内的形态变异性,发现槭属的翅果形态具有明显的地区间差异性,这种差异性主要表现在总体形状和大小上,这可能与地区间不同的气候环境特征有关。通过对槭属翅果种内形态变异性和多样性的深入研究,为槭属翅果化石的属下鉴定提供一定依据,指出槭属翅果化石的属下鉴定应该考虑翅果形态种内变异性的特征,具有形态多样性的该属翅果化石可能属于一个种或更少的种。
DOI:10.2307/2260669URL [本文引用: 1]
(1) Diaspore weight and dispersal were measured in 229 plant species of the central Australian arid zone. The frequency distribution of log diaspore sizes was unimodal, as has been observed for other habitats. The quantity of reserve (embryo plus endosperm) stored in the seed was related to the weight of the dispersal unit. (2) Reserve weight was greater for woody species than for herbaceous species. Within each growth form, there was no clear relationship between perenniality and reserve weight. (3) Most graminoids were wind dispersed, and elaiosomes for ant dispersal were more common among the woody species. The number of species with unassisted seeds was low in central Australia, when compared with other arid communities.
DOI:10.3417/1055-3177(2006)16[354:TGCCIA]2.0.CO;2URL [本文引用: 1]
Taxonomic review is provided for Callitriche taxa occurring in central and eastern Asia east of the Urals and in southeastern Asia north of Irian Jaya Barat. Three new species: Callitriche fuscicarpa, C. glareosa, and C. raveniana are described from Nepal, Bhutan, and Taiwan, China, respectively. Two new varieties are also described: Callitriche palustris L. var. megalata from Siberia in Russia and Callitriche palustris L. var. strumosa from western Sumatra in Indonesia. Callitriche nana is newly referred as a synonym of C. japonica Engelmann ex Hegelmaier. Callitriche oryzetorum Petrov is transferred and reduced in rank to a variety of C. palustris L. Callitriche wightiana Wallich in Wight is lectotypified.
In: Wu ZY, Raven PH, Hong DY eds.
[本文引用: 3]
In: Wu ZY, Raven PH, Hong DY eds.
[本文引用: 2]
In: Wu ZY, Raven PH, Hong DY eds.
[本文引用: 3]
DOI:10.1046/j.1365-2699.1997.00126.xURL [本文引用: 1]
Many species of tropical moist forests have large seeds compared to those found in temperate floras. This could be attributed to a prevalence of woody growth forms, or adaptation to dispersal by vertebrates, or the dense shade of a closed canopy, rather than to an intrinsic tropical v. temperate difference. We compared tropical v. temperate seed mass data at two geographic scales; firstly within Australia between tropical, subtropical and temperate open woodlands, then between six tropical and temperate datasets from five continents. Within Australia we found that seed mass increased with decreasing latitude in ten out of eleven growth form-dispersal mode combinations: only wind-dispersed graminoids showed no significant effect. While the pattern of generally larger seed in the tropical flora was associated with tropical families tending to have larger seeds than temperate families, we also found that tropical species had larger seeds than temperate cofamilials. As a preliminary test of the generality of these findings we included a further four tropical and temperate data sets in a second analysis. Average tropical seed masses were larger than temperate seed masses in all testable growth form-dispersal mode combinations, and statistically larger in five out of severn cases: unassisted, vertebrate- and wind-despersed woody species, and snassisted and vertebrate-dispersed forbs. No difference was found for wind-dispersed forbs or for graminoids lacking dispersal structures. That an effect was found in these combinations rules out explanations based solely on characteristics of tropical v. temperate dispersers, althogh this may be a contributing factor. Instead we suggest that some aspect of tropical systems, for example higher metabolic costs of seedling production, has selected for a general increase in seed mass among tropical taxa.
In: Wu ZY, Raven PH, Hong DY eds.
[本文引用: 8]
In: Wu ZY, Raven PH, Hong DY eds.
[本文引用: 5]
DOI:10.1007/s12229-010-9041-0URL [本文引用: 2]
Fin-winged fruits have two or more wings aligned with the longitudinal axis like the feathers of an arrow, as exemplified by Combretum , Halesia , and Ptelea . Such fruits vary in dispersal mode from those in which the fruit itself is the ultimate disseminule, to schizocarps dispersing two or more mericarps, to capsules releasing multiple seeds. At least 45 families and more than 140 genera are known to possess fin-winged fruits. We present an inventory of these taxa and describe their morphological characters as an aid for the identification and phylogenetic assessment of fossil and extant genera. Such fruits are most prevalent among Eudicots, but occur occasionally in Magnoliids (Hernandiaceae: Illigera ) and Monocots ( Burmannia , Dioscorea , Herreria ). Although convergent in general form, fin-winged fruits of different genera can be distinguished by details of the wing number, texture, shape and venation, along with characters of persistent floral parts and dehiscence mode. Families having genera with fin-winged fruits and epigynous perianth include Aizoaceae, Apiaceae, Araliaceae, Asteraceae, Begoniaceae, Burmanniaceae, Combretaceae, Cucurbitaceae, Dioscoreaceae, Haloragaceae, Lecythidiaceae, Lophopyxidaceae, Loranthaceae, and Styracaceae. Families with genera having fin-winged fruits and hypogynous perianth include Achariaceae, Brassicaceae, Burseraceae, Celastraceae, Cunoniaceae, Cyrillaceae, Fabaceae, Malvaceae, Melianthaceae, Nyctaginaceae, Pedaliaceae, Polygalaceae, Phyllanthaceae, Polygonaceae, Rhamnaceae, Salicaceae sl, Sapindaceae, Simaroubaceae, Trigoniaceae, and Zygophyllaceae. This survey has facilitated the identification of fossil winged fruits such as Combretaceae and Araliaceae in the late Cretaceous of western North America and provides additional evidence toward the identification of various Cenozoic fossils including Brassicaceae, Fabaceae, Polygonaceae, Rutaceae, and Sapindaceae.
DOI:10.1111/j.1469-185X.1973.tb01569.xURL [本文引用: 1]
1. A samara is a winged fruit or seed that autorotates when falling, thereby reducing the sinking speed of the diaspore and increasing the distance it may be transported by winds. Samaras have evolved independently in a large number of plants.
[本文引用: 5]
[本文引用: 1]
DOI:10.17520/biods.2015195URL [本文引用: 1]
以金虎尾科植物地理分布格局及迁移历史总结出来的“金虎尾路线”,是解释热带植物洲际间断分布与长距离扩散格局的重要模式。金虎尾路线阐明了金虎尾科植物历史时期7次独立的从起源中心(南美洲)向旧世界(非洲和亚洲)的洲际长距离扩散事件。本文总结了金虎尾路线植物起源地与扩散地主要类群的花部特征与传粉系统,以探讨这些类群及类似植物长距离扩散后的花进化与传粉转变等适应规律。金虎尾科的南美洲类群都有分泌油脂的萼片腺体,花的形态结构非常保守,是与美洲当地特有的条蜂科集油蜂长期协同进化的结果。金虎尾科的非洲类群花保守性消失,花白色、。辐射对称且无萼片腺体,繁育系统为雄花一两性花异株(功能性的雌雄异株),传粉者是采集花粉的蜜蜂科昆虫。亚洲的一些属发生了类似非洲类群的泛化适应转变,但风筝果属(Hiptage)~现了镜像花、异型雄蕊和极度反折的花瓣,且传粉者是亚洲特有的大蜜蜂(Apisdorsata),显示出了非常特化的适应性转变。风筝果属所在支系的现存类群涵盖了南美洲、中美洲、非洲和亚洲等地的地方特有属,体现了金虎尾路线整个迁移历史过程,是认识金虎尾路线及其进化适应规律的关键类群,值得在今后的研究中加以重视。
DOI:10.17520/biods.2015195URL [本文引用: 1]
以金虎尾科植物地理分布格局及迁移历史总结出来的“金虎尾路线”,是解释热带植物洲际间断分布与长距离扩散格局的重要模式。金虎尾路线阐明了金虎尾科植物历史时期7次独立的从起源中心(南美洲)向旧世界(非洲和亚洲)的洲际长距离扩散事件。本文总结了金虎尾路线植物起源地与扩散地主要类群的花部特征与传粉系统,以探讨这些类群及类似植物长距离扩散后的花进化与传粉转变等适应规律。金虎尾科的南美洲类群都有分泌油脂的萼片腺体,花的形态结构非常保守,是与美洲当地特有的条蜂科集油蜂长期协同进化的结果。金虎尾科的非洲类群花保守性消失,花白色、。辐射对称且无萼片腺体,繁育系统为雄花一两性花异株(功能性的雌雄异株),传粉者是采集花粉的蜜蜂科昆虫。亚洲的一些属发生了类似非洲类群的泛化适应转变,但风筝果属(Hiptage)~现了镜像花、异型雄蕊和极度反折的花瓣,且传粉者是亚洲特有的大蜜蜂(Apisdorsata),显示出了非常特化的适应性转变。风筝果属所在支系的现存类群涵盖了南美洲、中美洲、非洲和亚洲等地的地方特有属,体现了金虎尾路线整个迁移历史过程,是认识金虎尾路线及其进化适应规律的关键类群,值得在今后的研究中加以重视。
In: Wu ZY, Raven PH, Hong DY eds.
[本文引用: 3]
[本文引用: 2]
DOI:10.1111/j.1469-185X.1992.tb01018.xURL [本文引用: 1]
1. Autorotation of a single-winged samara is a highly nonlinear phenomenon that represents a delicate equilibrium between gravity, inertia and aerodynamic effects. Therefore, in order to analyse this phenomenon, an accurate detailed model is necessary. Such a model has not been presented in the past. Recently the authors derived a detailed model which is briefly described in the paper. 2. The aerodynamic contributions present the most complicated part of the phenomenon. These contributions are treated using the blade-element/momentuin method, with certain improvements and additions. These improvements are necessary due to inherent differences between samara wings and other rotary wing systems (aircraft propellers, helicopter rotors, etc.). 3. The cross-sectional aerodynamics of the samara is characterized by relatively small Reynolds numbers, high angles of attack and rough surfaces. While these characteristics are different from other rotary wings, they are typical of the wing cross-sections of insects and birds. Therefore the lift and drag coefficients, which are necessary for the analysis, are obtained using available data for insect and bird wings. 4. The results of the theoretical model are compared with experimental results of tlvo kinds. The first kind includes results for a samara of an Acer platanoides that were reported in the literature. In addition, a special experimental model of a samiira was built and tested. This model offers a simple way of varying the configuration in order to study (experimentally) the effect of different geometric parameters on the autorotation. 5. In the light of the uncertainty in the aerodynamic coefficients, it can be concluded that there is quite a good agreement between the theoretical and experimental results. Thus, after LTalidation, the theoretical model is used for a parametric study to find the influence of different parameters on the autorotation. The important results of this study are outlined below. 6. The spanwise flolv component and the tangential component of the induced velocity have a very small influence and thus can be neglected. 7. It is important to include in the analysis the effects of the axial induced velocity, the tip effect, and the drag of the root region. 8. Since chordwise variations of the centre of pressure location, as a function of the angle of attack, were seen in the past (based on over simplified analyses) as the mechanism which is responsible for the samara stability, this effect is also added to the model. While the influence of this effect on the pitch angle is large and small on the sinking rate, it results in an increase in the deviation between the theoretical and experimental results. <P>9. Autorotation is sensitive to the cross-sectional aerodynamic coefficients. This sensitivity is critical since the available data on these coefficients is, to say the least, unsatisfactory and require significant
DOI:10.11707/j.1001-7488.20110311URLMagsci [本文引用: 3]
<p>青钱柳种子具有深休眠特性。对青钱柳离体胚及不同处理种子的萌发试验、种壳透水性和透气性测定以及种子各部分内源抑制物质的提取、生物测定和鉴定等的研究表明: 1)青钱柳种胚基本不存在休眠,果皮和种皮存在一定的机械束缚和透水、透气性障碍,果皮中存在活性较强的内源抑制物质是引起青钱柳种子休眠的主要原因。 2)种子各部分甲醇浸提液的生物测定结果说明,青钱柳种子各部分均含有内源抑制物质,抑制活性为果皮>种皮>完整种子>种胚。3)对抑制活性最强的果皮甲醇提取液采用系统溶剂法进行初步分离,生物测定结果表明各分离相中乙醚相的抑制活性最强,GC-MS鉴定结果显示乙醚相中相对含量为35.09%的组分是具有较强抑制作用的香草酸。研究结果首次较系统地阐明青钱柳种子的休眠机制。</p>
DOI:10.11707/j.1001-7488.20110311URLMagsci [本文引用: 3]
<p>青钱柳种子具有深休眠特性。对青钱柳离体胚及不同处理种子的萌发试验、种壳透水性和透气性测定以及种子各部分内源抑制物质的提取、生物测定和鉴定等的研究表明: 1)青钱柳种胚基本不存在休眠,果皮和种皮存在一定的机械束缚和透水、透气性障碍,果皮中存在活性较强的内源抑制物质是引起青钱柳种子休眠的主要原因。 2)种子各部分甲醇浸提液的生物测定结果说明,青钱柳种子各部分均含有内源抑制物质,抑制活性为果皮>种皮>完整种子>种胚。3)对抑制活性最强的果皮甲醇提取液采用系统溶剂法进行初步分离,生物测定结果表明各分离相中乙醚相的抑制活性最强,GC-MS鉴定结果显示乙醚相中相对含量为35.09%的组分是具有较强抑制作用的香草酸。研究结果首次较系统地阐明青钱柳种子的休眠机制。</p>
In: Wu ZY, Raven PH, Hong DY eds.
[本文引用: 1]
DOI:10.1007/BF02489453URL
Seed germination behavior in two types of dimorphic fruits in Salsola komarovii Iljin (Chenopodiaceae) was studied. The germination percentage was much higher in the long-winged type than in the short-winged type when the fruits were tested for germinability. If lignified perianths were removed from the fruits, seeds of both types germinated at higher percentages and the difference in the germination percentage between two types decreased. The extract of the perianths inhibited the seed germination. Abscisic acid was detected in the extract of the perianths by enzyme-linked immunosorbent assay and its level was higher in the short-winged type than in the long-winged type. Germinability in both types was lost one year after harvest when the fruits were stored at room temperature and the germination was tested with fruits. When the seeds were tested, however, the short-winged type germinated even two years after the harvest; the germinability of the short-winged type lasted one year longer than that of the long-winged type. Light irradiation was not required for the seed germination in either type regardless of presence or absence of the perianths. No clear difference between the two types was observed either in optimal temperature for the germination or in strength of the resistance to salinity.
[本文引用: 1]
[本文引用: 1]
In: Wu ZY, Raven PH, Hong DY eds.
[本文引用: 4]
DOI:10.1111/boj.12385URL [本文引用: 3]
An update of the Angiosperm Phylogeny Group (APG) classification of the orders and families of angiosperms is presented. Several new orders are recognized: Boraginales, Dilleniales, Icacinales, Metteniusiales and Vahliales. This brings the total number of orders and families recognized in the APG system to 64 and 416, respectively. We propose two additional informal major clades, superrosids and superasterids, that each comprise the additional orders that are included in the larger clades dominated by the rosids and asterids. Families that made up potentially monofamilial orders, Dasypogonaceae and Sabiaceae, are instead referred to Arecales and Proteales, respectively. Two parasitic families formerly of uncertain positions are now placed: Cynomoriaceae in Saxifragales and Apodanthaceae in Cucurbitales. Although there is evidence that some families recognized in APG III are not monophyletic, we make no changes in Dioscoreales and Santalales relative to APG III and leave some genera in Lamiales unplaced (e.g. Peltanthera ). These changes in familial circumscription and recognition have all resulted from new results published since APG III, except for some changes simply due to nomenclatural issues, which include substituting Asphodelaceae for Xanthorrhoeaceae (Asparagales) and Francoaceae for Melianthaceae (Geraniales); however, in Francoaceae we also include Bersamaceae, Ledocarpaceae, Rhynchothecaceae and Vivianiaceae. Other changes to family limits are not drastic or numerous and are mostly focused on some members of the lamiids, especially the former Icacinaceae that have long been problematic with several genera moved to the formerly monogeneric Metteniusaceae, but minor changes in circumscription include Aristolochiaceae (now including Lactoridaceae and Hydnoraceae; Aristolochiales), Maundiaceae (removed from Juncaginaceae; Alismatales), Restionaceae (now re-including Anarthriaceae and Centrolepidaceae; Poales), Buxaceae (now including Haptanthaceae; Buxales), Peraceae (split from Euphorbiaceae; Malpighiales), recognition of Petenaeaceae (Huerteales), Kewaceae, Limeaceae, Macarthuriaceae and Microteaceae (all Caryophyllales), Petiveriaceae split from Phytolaccaceae (Caryophyllales), changes to the generic composition of Ixonanthaceae and Irvingiaceae (with transfer of Allantospermum from the former to the latter; Malpighiales), transfer of Pakaraimaea (formerly Dipterocarpaceae) to Cistaceae (Malvales), transfer of Borthwickia , Forchhammeria , Stixis and Tirania (formerly all Capparaceae) to Resedaceae (Brassicales), Nyssaceae split from Cornaceae (Cornales), Pteleocarpa moved to Gelsemiaceae (Gentianales), changes to the generic composition of Gesneriaceae ( Sanango moved from Loganiaceae) and Orobanchaceae (now including Lindenbergiaceae and Rehmanniaceae) and recognition of Mazaceae distinct from Phrymaceae (all Lamiales).
DOI:10.1016/j.tree.2012.02.002URLPMID:22445687 [本文引用: 3]
Since Darwin, the diversity of flowers has been attributed to selection by pollinators. Although pollinators commonly act as selective agents on floral traits, determining the extent to which they have influenced angiosperm diversification requires a historical perspective. Here we review recent studies that combine species-level phylogenies with pollinator data and show that pollinator shifts are common, being associated with at least a quarter of documented divergence events. However, shift frequency and directionality vary extensively, owing to variation in intrinsic factors such as floral features and phylogenetic history, as well as extrinsic factors such as interactions with local pollinator assemblages. Despite technical advances, phylogenies remain limited in their power to distinguish among various pollinator-driven evolutionary processes.
DOI:10.3321/j.issn:1673-5021.2002.06.008URL
在冬春季节,对内蒙古锡林河中游草原风滚植物进行了调查,共记录 12种风滚植物,隶属于藜科、伞形科、菊科等7科.其中,一二年生草本植物和多年生草本各6种;旱生、中旱生植物7种,旱中生、中生植物5种;在区系地理 成分中温带亚洲成分(6种)最多.风滚植物是草原枯草期植物的一种存在形式,它的存在为积雪的再分配,局部土壤水分、养分的改善,种子的传播、萌发奠定了 基础,具有十分重要的生态学意义.
DOI:10.3321/j.issn:1673-5021.2002.06.008URL
在冬春季节,对内蒙古锡林河中游草原风滚植物进行了调查,共记录 12种风滚植物,隶属于藜科、伞形科、菊科等7科.其中,一二年生草本植物和多年生草本各6种;旱生、中旱生植物7种,旱中生、中生植物5种;在区系地理 成分中温带亚洲成分(6种)最多.风滚植物是草原枯草期植物的一种存在形式,它的存在为积雪的再分配,局部土壤水分、养分的改善,种子的传播、萌发奠定了 基础,具有十分重要的生态学意义.
URL [本文引用: 5]
梣属植物是北半球古近纪和新近纪化石记录中最具代表性的种属之一。对采自云南临沧晚中新世的一种梣属翅果化石Fraxinus cf.honshuensis进行了鉴定和描述,并通过梣属12个翅果化石种和9个现生种的形态特征的研究,分析了该属翅果形态从始新世直至现代的演化趋势及风力传播作用对翅果形态的影响。结果表明:该属翅果在晚渐新世至中中新世时翅果的风力传播能力最强,传播距离最远,并且与化石记录具有较好的对应性。地史分布表明,该属植物最初可能通过白令陆桥和北大西洋陆桥由高纬度地区向低纬度地区迁徙,最终传播到亚热带和热带区域。
URL [本文引用: 5]
梣属植物是北半球古近纪和新近纪化石记录中最具代表性的种属之一。对采自云南临沧晚中新世的一种梣属翅果化石Fraxinus cf.honshuensis进行了鉴定和描述,并通过梣属12个翅果化石种和9个现生种的形态特征的研究,分析了该属翅果形态从始新世直至现代的演化趋势及风力传播作用对翅果形态的影响。结果表明:该属翅果在晚渐新世至中中新世时翅果的风力传播能力最强,传播距离最远,并且与化石记录具有较好的对应性。地史分布表明,该属植物最初可能通过白令陆桥和北大西洋陆桥由高纬度地区向低纬度地区迁徙,最终传播到亚热带和热带区域。
In: Wu ZY, Raven PH, Hong DY eds.
DOI:10.1007/978-3-662-07255-4_9URL [本文引用: 2]
Large woody lianas; branches sympodial with a series of circinate woody hooks in one plane. Leaves alternate, often crowded in rosettes on lateral flowering shoots, simple, entire; stipules small and caducous; petioles articulated, often leaving a saddle-shaped scar on the twigs. Inflorescences racemes, spikes or dichasially branched panicles. Flowers bisexual; regular, usually less than 2 cm in diameter; calyx tube at first short, at length adnate to the base of the ovary; sepals 5, unequal in size; petals 5, slightly fleshy, distinct or slightly connate at base, contorted or imbricate; stamens 5,10 or 15, erect in bud, perigynous; filaments slightly connate at base; anthers basifixed, tetrasporangiate, introrse, opening by longitudinal slits; carpels 3(4), fused to form a syncarpous, half-inferior, unilocular ovary; stylodia 3(4), articulated at apex of ovary, with capitate, crescent-shaped, or pinnatifid stigmas; ovule solitary, basilateral, hemianatropous, bitegmic. Fruit a nut, surrounded by wing-like, accrescent, unequal calyx lobes; seed subglobose, endosperm ruminate; embryo straight, cotyledons remarkably folded.
DOI:10.1016/j.flora.2007.02.003URL
Salsola affinis is a dominant annual inhabiting saline deserts of Xinjiang, China. Experiments were conducted to determine the effects of temperature, winged perianths and NaCl on seed germination and on germination recovery from the effects of saline conditions after transfer to distilled water. Freshly harvested seeds could germinate equally well in light and darkness at 5–30 °C. Attached winged perianths significantly inhibited germination, removal enhanced germination. However, germination was not inhibited in the presence of detached winged perianths in any of the temperature treatments. We suggest that the winged perianth is a mechanical barrier for radicle emergence, not a barrier for water uptake; hence, it inhibited germination. Germination of seeds from which the perianth had been removed was not affected by NaCl at concentrations below 0.4 mol/l, but it was significantly decreased by NaCl at concentrations of 0.6–2.0 mol/l. No seeds germinated at 4.0 mol/l NaCl. Seeds incubated in NaCl at concentrations of 0.05–4.0 mol/l for 14 days recovered after being transferred to distilled water. However, germination was lower than that in the non-saline control, indicating that a portion of the NaCl-treated seeds may lose their ability to germinate.
[本文引用: 1]
In: Wu ZY, Raven PH, Hong DY eds.
[本文引用: 4]
In: Wu ZY, Raven PH, Hong DY eds.
[本文引用: 1]
DOI:10.1007/BF02489624URL [本文引用: 1]
Two different types of dispersal units (called fruits in this study) were observed in Salsola komarovii Iljin. One is a fruit which has dark brown lignified tepals with long wings and a green seed and falls easily from the mother plant (long-winged type). Another has light brown lignified tepals with short wings and a yellow seed and attaches tightly to the mother plant (short-winged type). This difference of fruit type appeared independently from maturity of fruit. Seeds in the short-winged fruits were in dormancy for a longer period of time than those in the long-winged fruits. The germination rate was significantly higher in the seeds of long-winged fruits. The dormancy in seeds of the long-winged fruits was effectively terminated by reducing the temperature but the effect of chilling was very weak in seeds of the short-winged fruits. It was concluded from these observations that there exists a dimorphism in the fruits of, or a heterocarpy in, S. komarovii . The plants grown under water stress produced mostly short-winged fruits and those grown under well-watered conditions bore fruits of both types. Exogenously applied abscisic acid (ABA) tended to produce the short-winged fruits, suggesting that the heterocarpy was, at least partly, regulated by ABA.
[本文引用: 1]
[本文引用: 1]
New species, names, and combinations in American Combretaceae
1
1989
... 椴树科蚬木属(Excentrodendron)和柄翅果属(Burretiodendron)的棱翅果通常具5个薄膜状的果翅(
The genera of Lepidieae(Cruciferae; Brassicaceae) in the southeastern United States
1
1986
... 十字花科的沙芥属、菘蓝属植物和蓼科的沙拐枣属植物生长在沙漠地区, 它们的棱翅果与披针翅果依靠果翅使得种子不直接接触到高温的土表, 有利于其适应干旱高温的沙漠环境(
13
2006
... 化石证据表明, 早在晚中新世的陉属(Fraxinus)中就出现了翅果(
... 前人对翅果的风力传播机制、运动方式与演化发育等方面开展了一些实验研究(
... (
... 单翅果有时会两个或三个聚合在一起, 形成双聚单翅果和三聚单翅果等类型(
... 有些周位翅果的果翅存在着对称的收缩或出现缺口, 整体呈蝶翅状(
... 披针翅果的果翅形态与数量变化较大, 有扁长如螺旋桨片的典型披针状果翅, 如金虎尾科风筝果属, 也有较为细长呈针状的果翅, 如金虎尾科三星果属(
... 总的来看, 果翅的数量、形态与着生位置在科属水平上非常稳定, 可以作为植物形态分类的重要依据(
... 金虎尾科是翅果类型最丰富的一个类群(
... “金虎尾路线”认为, 金虎尾科起源于南美洲, 先后9次扩散到非洲和亚洲热带地区, 其中7次沿北大西洋路桥, 2次是近期发生的横渡大西洋扩散(
... Carolus支系为整个金虎尾家族严格体现金虎尾路线完整扩散过程的支系, 在南美洲、中美洲、非洲以及亚洲都有亲缘关系很近的特有属(
...
... ); 直升机式(如披针翅果、翼状萼翅果和叶状苞翅果等多个果翅位于果实一端)和波浪式(如周位翅果)的空中运动较为稳定, 扩散方向较明确, 是扩散距离较远的一类翅果(
... Vf与果翅负荷密切相关(
Morphology and dispersal potential of wind-dispersed diaspores of neotropical trees
8
1986
... 化石证据表明, 早在晚中新世的陉属(Fraxinus)中就出现了翅果(
... 前人对翅果的风力传播机制、运动方式与演化发育等方面开展了一些实验研究(
...
... 不同类型翅果的降落速率与果翅负荷(果实质量/果翅面积)的相关性.改自
... Descent velocity and its relationship with wing loading (fruit mass / wing size) of samaras and pollen grains. Revised from
...
... ).翅果降落速率与传播距离不仅与果翅大小、运动方式有关, 还与整个果实的质量分配和水分含量有关(
... Vf与果翅负荷密切相关(
Mass allocation, moisture content, and dispersal capacity of wind-dispersed tropical diaspores
3
1988
...
... 有些翅果如棱翅果与披针翅果的果翅均匀分布在果实四周, 可在落至土表之后再次在风吹或重力作用下成为“风滚翅果” (
... 十字花科的沙芥属、菘蓝属植物和蓼科的沙拐枣属植物生长在沙漠地区, 它们的棱翅果与披针翅果依靠果翅使得种子不直接接触到高温的土表, 有利于其适应干旱高温的沙漠环境(
Diaspore dispersal ability and degree of dormancy in heteromorphic species of cold deserts of northwest China: A review
1
2014
... 有些翅果如棱翅果与披针翅果的果翅均匀分布在果实四周, 可在落至土表之后再次在风吹或重力作用下成为“风滚翅果” (
Fruit types and seed dispersal modes of woody plants in Ahuriri summit Bush, Port Hills, western Banks Peninsula, Canterbury, New Zealand
1
1994
... 圆翅果的翅呈完全闭合圆形或椭圆形包围果实, 果翅在一个平面上(
Wind-borne seed and fruit movement
1975
Oleaceae
1
1996
... 有些周位翅果的果翅存在着对称的收缩或出现缺口, 整体呈蝶翅状(
Verbenaceae
2
1994
... 马鞭草科楔翅藤属是亚洲热带地区特有的攀援木质藤本, 其花序为头状聚伞花序, 具5-6枚匙状总苞片.果实成熟后, 所有苞片宿存、平展, 似风车状(
... 由于果翅是果皮的延伸或萼片与苞片的特化, 通常包围着果实, 具有保护果实与种子的作用.特别是周位翅果、棱翅果和披针状翅果等具有环绕翅果四周的果翅, 能够在果实与种子掉落地面时, 避免果实与种子直接接触地表, 可减轻果实在空中运动及落地时的机械损伤和降低地表微生物侵染果实的速率(
Seed dispersal distances and plant migration potential in tropical East Asia
1
2009
...
Distance of seed dispersal in Eucalyptus estimated from seed weights
1977
5
2000
... 被子植物约有90个科具翅果(
... “金虎尾路线”认为, 金虎尾科起源于南美洲, 先后9次扩散到非洲和亚洲热带地区, 其中7次沿北大西洋路桥, 2次是近期发生的横渡大西洋扩散(
... ,
... Carolus支系为整个金虎尾家族严格体现金虎尾路线完整扩散过程的支系, 在南美洲、中美洲、非洲以及亚洲都有亲缘关系很近的特有属(
... 目前, 有关翅果形态与功能的研究还远不如植物繁殖器官“花”那样受人关注, 不同翅果类型可能具有的适应意义还缺乏有力的证据和明确区分, 翅果对被子植物多样性形成与维持的作用也未受到应有的重视.因此, 利用生态适应和演化-发育生物学方法, 研究不同类型翅果在适应风力传播方面的差异, 果翅特别是萼片或苞片来源的果翅的发育机制, 翅果不同类型的演化历史及其对植株物种多样性的影响是值得深入开展的一个课题.值得注意的是, 金虎尾科具有被子植物最为多样的翅果类型, 也是生物地理学研究模式“金虎尾路线” (
A complete generic phylogeny of Malpighiaceae inferred from nucleotide sequence data and morphology
1
2010
... “金虎尾路线”认为, 金虎尾科起源于南美洲, 先后9次扩散到非洲和亚洲热带地区, 其中7次沿北大西洋路桥, 2次是近期发生的横渡大西洋扩散(
Laurasian migration explains Gondwanan disjunctions: Evidence from Malpighiaceae
4
2002
... 金虎尾科是翅果类型最丰富的一个类群(
... )的代表类群, 该科历史上每一次洲际长距离扩散往往都伴随着翅果类型的转变(
... “金虎尾路线”认为, 金虎尾科起源于南美洲, 先后9次扩散到非洲和亚洲热带地区, 其中7次沿北大西洋路桥, 2次是近期发生的横渡大西洋扩散(
... Carolus支系为整个金虎尾家族严格体现金虎尾路线完整扩散过程的支系, 在南美洲、中美洲、非洲以及亚洲都有亲缘关系很近的特有属(
Long-term morphological stasis maintained by a plant-pollinator mutualism
4
2014
... 在双子叶植物中, 基部类群毛茛目、昆栏树目与金鱼藻目并没有出现翅果, 但在物种多样性较高的类群如唇形目、豆目、金虎尾目和锦葵目中, 具有极高的翅果类型多样性(
... Samara diversity and evolution in Malpighiaceae. Note there has evolutionary transition of samara types for each long-distance dispersal between continents. The phylogeny is redrawn from
... 金虎尾科是翅果类型最丰富的一个类群(
...
Phylogeny of Malpighiaceae: Evidence from chloroplast ndhF and trnL-F nucleotide sequences
1
2001
... 单翅果有时会两个或三个聚合在一起, 形成双聚单翅果和三聚单翅果等类型(
Samara size versus dispersal and seedling establishment in Ailanthus altissima(Miller) Swingle
2009
云南槭属植物一新种
2
2003
... 化石证据表明, 早在晚中新世的陉属(Fraxinus)中就出现了翅果(
... 总的来看, 果翅的数量、形态与着生位置在科属水平上非常稳定, 可以作为植物形态分类的重要依据(
云南槭属植物一新种
2
2003
... 化石证据表明, 早在晚中新世的陉属(Fraxinus)中就出现了翅果(
... 总的来看, 果翅的数量、形态与着生位置在科属水平上非常稳定, 可以作为植物形态分类的重要依据(
Pollination systems, dispersal modes, life forms, and diversification rates in angiosperm families
1
1992
... 翅果(samara)是指果实的果皮或其他部位延伸呈翅状并依靠风力传播的一类果实, 是被子植物果实适应风力传播的一个“关键创新性状”(
Seed size, fruit size, and dispersal systems in angiosperms from the Early Cretaceous to the Late Tertiary
1
2000
... 翅果(samara)是指果实的果皮或其他部位延伸呈翅状并依靠风力传播的一类果实, 是被子植物果实适应风力传播的一个“关键创新性状”(
Convolvulaceae
6
1995
... 翅果(samara)是指果实的果皮或其他部位延伸呈翅状并依靠风力传播的一类果实, 是被子植物果实适应风力传播的一个“关键创新性状”(
... (
... 旋花科飞蛾藤属(Dinetus)植物蒴果的5个萼片均增长呈螺旋桨状, 外形类似披针翅果; 但由于果翅来源于萼片, 果翅均位于蒴果下方(
... )蒴果的萼片则宽展成花瓣状(
... 旋花科盾苞藤属(Neuropeltis)的苞片紧贴萼下, 在果期显著增大呈宽椭圆形或圆形, 长3-5 cm, 除脉络外, 其他部分均被柔毛(
...
3
2011
... 翅果(samara)是指果实的果皮或其他部位延伸呈翅状并依靠风力传播的一类果实, 是被子植物果实适应风力传播的一个“关键创新性状”(
... 总的来看, 果翅的数量、形态与着生位置在科属水平上非常稳定, 可以作为植物形态分类的重要依据(
... 从被子植物的系统发育树看, 其基部类群樟目已出现了蝶翅果和棱翅果(
The terminal velocity and dispersal of spinning samaras
1
1980
... 被子植物约有90个科具翅果(
The dispersal of winged fruits and seeds differing in autorotative behaviour
1
1990
... Vf与果翅负荷密切相关(
1
2004
... 翅果(samara)是指果实的果皮或其他部位延伸呈翅状并依靠风力传播的一类果实, 是被子植物果实适应风力传播的一个“关键创新性状”(
1
2001
... 翅果(samara)是指果实的果皮或其他部位延伸呈翅状并依靠风力传播的一类果实, 是被子植物果实适应风力传播的一个“关键创新性状”(
Seed dispersal by animals: A role in angiosperm diversification?
2
1989
... 化石证据表明, 早在晚中新世的陉属(Fraxinus)中就出现了翅果(
... 总的来看, 果翅的数量、形态与着生位置在科属水平上非常稳定, 可以作为植物形态分类的重要依据(
Valerianaceae
1
2011
... 败酱科败酱属的果实本身为瘦果, 但苞片膨大呈翅状包围整个瘦果, 使整个果实为功能性“翅果”.其叶状苞翅状, 通常具2-3条主脉, 网脉明显.如西伯利亚败酱(Patrinia sibirica)的叶状苞倒卵形或倒卵状长圆形, 长6-9 mm、宽4.5-6.5 mm, 先端圆钝或三浅裂, 羽状网脉明显, 常具3主脉(
槭属翅果种内形态变异性及对其化石鉴定的意义
2013
槭属翅果种内形态变异性及对其化石鉴定的意义
2013
Diaspore weight, dispersal, growth form and perenniality of Central Australian plants
1
1991
... 总的来看, 果翅的数量、形态与着生位置在科属水平上非常稳定, 可以作为植物形态分类的重要依据(
The genus Callitriche(Callitrichaceae) in Asia
1
2006
...
Polygonaceae
3
2003
... 蓼科植物的果实虽然较小, 但仍是典型的棱翅果(
... )翅果具3锐棱, 沿棱具黄褐色膜质翅, 基部具3个黑色角状附属物(
... 总的来看, 果翅的数量、形态与着生位置在科属水平上非常稳定, 可以作为植物形态分类的重要依据(
Betulaceae
2
1999
... 桦木科的鹅耳枥(Carpinus turczaninowii)叶状苞翅果较为典型, 苞片半卵形、半长圆形或半宽卵形, 长近2 cm, 外缘具缺齿、内缘全缘或疏生细齿, 基部具卵形、内折裂片(
... ).桦木科叶状苞翅果的果翅形状与大小以及裂片有无和开裂方式是形态分类的一个主要依据(
Dipterocarpaceae
3
2007
... 龙脑香科有16属、500多种, 广布于亚洲热带地区, 均具翼状萼翅果(
... )的5个果翅等长或其中3枚较大(
... 被子植物约有90个科具翅果(
Larger seeds in tropical floras: Consistent patterns independent of growth form and dispersal mode
1
1997
... 总的来看, 果翅的数量、形态与着生位置在科属水平上非常稳定, 可以作为植物形态分类的重要依据(
Juglandaceae
8
1999
... 化石证据表明, 早在晚中新世的陉属(Fraxinus)中就出现了翅果(
... 前人对翅果的风力传播机制、运动方式与演化发育等方面开展了一些实验研究(
... 椴树科蚬木属(Excentrodendron)和柄翅果属(Burretiodendron)的棱翅果通常具5个薄膜状的果翅(
... 胡桃科黄杞属的叶状苞翅果的果翅膜质弯曲、3深裂, 中裂片显著长于两侧的裂片(
... ); 枫杨属的坚果基部具1宿存的鳞状苞片及2个苞片发育而来的革质翅, 翅向果实两侧斜上方伸展, 呈元宝状(
... ); 枫杨属的翅果在外形上酷似双聚单翅果, 但后者的果翅来源于果皮、由两个单侧翅果靠生在一起形成, 区别于枫杨属的单个果实2枚苞片形成的果翅(
... 总的来看, 果翅的数量、形态与着生位置在科属水平上非常稳定, 可以作为植物形态分类的重要依据(
... 金虎尾科是翅果类型最丰富的一个类群(
Celastraceae
5
2008
... 前人对翅果的风力传播机制、运动方式与演化发育等方面开展了一些实验研究(
... 卫矛属棱翅果的外形、果翅数量与形态变化均较大, 果翅有的宽短、有的狭长、有的末端呈钩状(
... )等物种的球状果实密被棕色细刺(
... ), 可能与棱形果翅有着相同的来源.卫矛科雷公藤属棱翅果的果实外形与果翅形态变化较小, 都为3片宽大的膜质翅(
... 总的来看, 果翅的数量、形态与着生位置在科属水平上非常稳定, 可以作为植物形态分类的重要依据(
Phylogenetic distribution and identification of fin-winged fruits
2
2010
... (
... 此外, 黑槭(Acer saccharum ssp. nigrum)果翅在发育早期具有一定的光合作用(
Autorotation, self-stability, and structure of single-winged fruits and seeds (samaras) with comparative remarks on animal flight
1
1973
...
Samara development of black maple (Acer saccharum ssp. nigrum) with emphasis on the wing
5
1991
... “金虎尾路线”认为, 金虎尾科起源于南美洲, 先后9次扩散到非洲和亚洲热带地区, 其中7次沿北大西洋路桥, 2次是近期发生的横渡大西洋扩散(
... ;
... Carolus支系为整个金虎尾家族严格体现金虎尾路线完整扩散过程的支系, 在南美洲、中美洲、非洲以及亚洲都有亲缘关系很近的特有属(
... ;
... ;
Gone with the wind and the stream: Dispersal in the invasive species Ailanthus altissima
1
2016
...
“金虎尾路线”植物的花进化与传粉转变
1
2016
... Carolus支系为整个金虎尾家族严格体现金虎尾路线完整扩散过程的支系, 在南美洲、中美洲、非洲以及亚洲都有亲缘关系很近的特有属(
“金虎尾路线”植物的花进化与传粉转变
1
2016
... Carolus支系为整个金虎尾家族严格体现金虎尾路线完整扩散过程的支系, 在南美洲、中美洲、非洲以及亚洲都有亲缘关系很近的特有属(
Elaeagnaceae
3
2007
... 前人对翅果的风力传播机制、运动方式与演化发育等方面开展了一些实验研究(
... 单侧翅果是指果实只有一个狭长果翅, 且果翅位于果实的单侧(
... 椴树科蚬木属(Excentrodendron)和柄翅果属(Burretiodendron)的棱翅果通常具5个薄膜状的果翅(
The upper reaches of the largest river in Southern China as an “evolutionary front” of tropical plants: Evidences from Asia-endemic genus Hiptage(Malpighiaceae)
2
2015
... 具圆翅果的胡桃科青钱柳(Cyclocarya paliurus)的种子常需隔年甚至2年后才能萌发(
... ).实验研究发现, 青钱柳种子胚不存在休眠和后熟, 果皮及环绕的圆形果翅造成的机械束缚和含有的香草酸是延滞青钱柳种子萌发的主要原因(
Study of the vertical autorotation of a single winged samara
1
1992
... Vf与果翅负荷密切相关(
青钱柳种子休眠机制
3
2011
... 主要生长于海滨或荒漠盐碱地的藜科猪毛菜属有着由花被及其附属物增厚特化而成的果翅(
... ;
... 因此, 果翅不仅仅是散布与保护果实与种子的一种适应, 还可能通过改变果皮的理化性质调控种子的成熟、休眠与萌发进程, 避免了种子集中萌发而造成的同胞竞争, 提高了植物适应逆境的能力(
青钱柳种子休眠机制
3
2011
... 主要生长于海滨或荒漠盐碱地的藜科猪毛菜属有着由花被及其附属物增厚特化而成的果翅(
... ;
... 因此, 果翅不仅仅是散布与保护果实与种子的一种适应, 还可能通过改变果皮的理化性质调控种子的成熟、休眠与萌发进程, 避免了种子集中萌发而造成的同胞竞争, 提高了植物适应逆境的能力(
Asteraceae
1
2011
... 有些周位翅果的果翅存在着对称的收缩或出现缺口, 整体呈蝶翅状(
Diversity in seed germination behavior in relation to heterocarpy in Salsola komarovii Iljin
1991
Taxonomical investigation of the living species of the genus Acer L., based on vein architecture of leaves
1
1978
... 总的来看, 果翅的数量、形态与着生位置在科属水平上非常稳定, 可以作为植物形态分类的重要依据(
Revisions of tertiary Acer from East Asia
1
1983
... 总的来看, 果翅的数量、形态与着生位置在科属水平上非常稳定, 可以作为植物形态分类的重要依据(
Tiliaceae
4
2007
... 翅果(samara)是指果实的果皮或其他部位延伸呈翅状并依靠风力传播的一类果实, 是被子植物果实适应风力传播的一个“关键创新性状”(
... 椴树科蚬木属(Excentrodendron)和柄翅果属(Burretiodendron)的棱翅果通常具5个薄膜状的果翅(
... 披针翅果的果翅形态与数量变化较大, 有扁长如螺旋桨片的典型披针状果翅, 如金虎尾科风筝果属, 也有较为细长呈针状的果翅, 如金虎尾科三星果属(
... 与其他翅果类型相比, 披针翅果的果翅通常细长、数量较多, 多呈辐射状均匀排列在果实的赤道面(
An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV
3
2016
... 被子植物约有90个科具翅果(
... The systematic distribution of samara types across angiosperm orders. The phylogeny of angiosperm orders was drawn according to the Angiosperm
... 有些翅果如棱翅果与披针翅果的果翅均匀分布在果实四周, 可在落至土表之后再次在风吹或重力作用下成为“风滚翅果” (
Phylogenetic evidence for pollinator-driven diversification of angiosperms
3
2012
... 翅果(samara)是指果实的果皮或其他部位延伸呈翅状并依靠风力传播的一类果实, 是被子植物果实适应风力传播的一个“关键创新性状”(
... 化石证据表明, 早在晚中新世的陉属(Fraxinus)中就出现了翅果(
... 总的来看, 果翅的数量、形态与着生位置在科属水平上非常稳定, 可以作为植物形态分类的重要依据(
锡林河中游草原风滚植物区系分析及生态意义
2002
锡林河中游草原风滚植物区系分析及生态意义
2002
云南临沧晚中新世梣属翅果化石及其古植物地理学意义
5
2012
... 藜科猪毛菜属(Salsola)翅果比较特殊, 果翅由花被及其附属物特化而来(
... 主要生长于海滨或荒漠盐碱地的藜科猪毛菜属有着由花被及其附属物增厚特化而成的果翅(
... )的果翅虽然不影响水分进入果实和种皮, 但阻碍了胚根突破种皮, 导致翅果比人为去掉果翅的处理萌发率低(
... ;
... 因此, 果翅不仅仅是散布与保护果实与种子的一种适应, 还可能通过改变果皮的理化性质调控种子的成熟、休眠与萌发进程, 避免了种子集中萌发而造成的同胞竞争, 提高了植物适应逆境的能力(
云南临沧晚中新世梣属翅果化石及其古植物地理学意义
5
2012
... 藜科猪毛菜属(Salsola)翅果比较特殊, 果翅由花被及其附属物特化而来(
... 主要生长于海滨或荒漠盐碱地的藜科猪毛菜属有着由花被及其附属物增厚特化而成的果翅(
... )的果翅虽然不影响水分进入果实和种皮, 但阻碍了胚根突破种皮, 导致翅果比人为去掉果翅的处理萌发率低(
... ;
... 因此, 果翅不仅仅是散布与保护果实与种子的一种适应, 还可能通过改变果皮的理化性质调控种子的成熟、休眠与萌发进程, 避免了种子集中萌发而造成的同胞竞争, 提高了植物适应逆境的能力(
Ancistrocladaceae
2
2007
... 有些翅果的果翅不是来自于果皮或种皮的延伸, 而是由萼片或苞片发育而来.如热带地区的优势乔木龙脑香科(Dipterocarpaceae)和钩枝藤科(Ancistrocladaceae)一些物种的花萼筒与果实愈合在一起, 萼片增大, 特化成翅状.这类翅果可称为“翼状萼翅果” (
... 由于果翅是果皮的延伸或萼片与苞片的特化, 通常包围着果实, 具有保护果实与种子的作用.特别是周位翅果、棱翅果和披针状翅果等具有环绕翅果四周的果翅, 能够在果实与种子掉落地面时, 避免果实与种子直接接触地表, 可减轻果实在空中运动及落地时的机械损伤和降低地表微生物侵染果实的速率(
Factors influencing seed germination of Salsola affinis(Chenopodiaceae), a dominant annual halophyte inhabiting the deserts of Xinjiang, China
2008
1
1973
... 由于果翅是果皮的延伸或萼片与苞片的特化, 通常包围着果实, 具有保护果实与种子的作用.特别是周位翅果、棱翅果和披针状翅果等具有环绕翅果四周的果翅, 能够在果实与种子掉落地面时, 避免果实与种子直接接触地表, 可减轻果实在空中运动及落地时的机械损伤和降低地表微生物侵染果实的速率(
Santalaceae
4
2003
... 胡桃科黄杞属的叶状苞翅果的果翅膜质弯曲、3深裂, 中裂片显著长于两侧的裂片(
... 主要生长于海滨或荒漠盐碱地的藜科猪毛菜属有着由花被及其附属物增厚特化而成的果翅(
... )在水分较好的环境下产生有翅和无翅的两种果实(
... ), 但在水分胁迫的条件下只产生无翅果, 且其翅果比无翅果的休眠期短、种子萌发率更高(
Aceraceae
1
2008
... 单翅果有时会两个或三个聚合在一起, 形成双聚单翅果和三聚单翅果等类型(
Diversities in morphological characteristics and seed germination behavior in fruits of Salsola komarovii Iljin
1
1990
... 藜科猪毛菜属(Salsola)翅果比较特殊, 果翅由花被及其附属物特化而来(
龙脑香科植物的地理分布
1
1985
... 被子植物约有90个科具翅果(
龙脑香科植物的地理分布
1
1985
... 被子植物约有90个科具翅果(
Copyright © 2021 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}