 ,2,3,**
,2,3,**Response of plant biomass to nitrogen addition and precipitation increasing under different climate conditions and time scales in grassland
DIAO Li-Wei1,2,*, LI Ping2,3,*, LIU Wei-Xing2, XU Shan4, QIAO Chun-Lian5, ZENG Hui1,6, LIU Ling-Li,2,3,**通讯作者:
第一联系人:
编委: 吕晓涛
责任编辑: 李敏
收稿日期:2018-03-7网络出版日期:2018-08-20
| 基金资助: |
Received:2018-03-7Online:2018-08-20
| Fund supported: |

摘要
关键词:
Abstract
Methods To investigate the impact of nitrogen addition and precipitation increasing on grassland biomass over climate conditions and temproal scales, a meta-analysis was conducted based on 46 papers that were published during 1990-2017 involving 1 350 observations.
Important findings Results showed that: (1) N addtion, precipitation increasing and the combinations of these two treatments significantly increased the aboveground biomass (37%, 41%, 104%), total biomass (32%, 23%, 60%) and the ratio of aboveground biomass to belowground biomass (29%, 25%, 46%) in grassland ecosystem. Belowground biomass showed no response to single N addtion, but could be significantly enhanced together with increaseing precipitation; (2) The response of grassland biomass under these N addtion and the increasing of precipitation showed obvious spatial pattern under different climate conditions. The N addition tended to increase more aboveground biomass, total biomass and the ratio of aboveground biomass to belowground biomass under high sites with high mean annual air temperature (MAT) and mean annual precipitation (MAP) while precipitation increasing tended to simulate more belowground biomass and total biomass under low MAT and MAP sites; (3) In addition, the response of grassland biomass under these two global change index showed obvious temporal pattern. With the increase of duration of N addition, the belowgound biomass tended to decrease, while the aboveground biomass, total biomass and the ratio of aboveground biomass to belowground biomass tended to increase under N addition. With the increase of duration of precipitation manipulation, the total biomass showed no response to precipitation increasing, while aboveground biomass, belowground biomass and the ratio of aboveground biomass to belowground biomass tended to be enhanced. The results indicated that aboveground biomass was more likely to be enhanced than belowground biomass under N addition or precipitation increasing in the long term.
Keywords:
PDF (5914KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
刁励玮, 李平, 刘卫星, 徐姗, 乔春连, 曾辉, 刘玲莉. 草地生态系统生物量在不同气候及多时间尺度上对氮添加和增雨处理的响应. 植物生态学报[J], 2018, 42(8): 818-830 doi:10.17521/cjpe.2018.0056
DIAO Li-Wei.
草地生态系统作为陆地生态系统重要的组成部分, 其生产力约占陆地总初级生产力的1/3 (Hoekstra et al., 2005), 具有重要的生态系统服务功能(Wrage et al., 2011)。氮和水分是草地生态系统生产力的两个重要限制性因素(Hooper & Johnson, 1999; Yahdjian et al., 2011)。氮进入生态系统后对光合作用和植物生长的影响存在差异, 进而导致植物生产力的不同响应(Ren et al., 2017)。同时, 干旱和湿润的草地生态系统生产力对降水变异所产生的响应也存在差异(Wu et al., 2011)。生物量是构成生态系统生产力的重要组分, 在许多研究中生物量常被近似等同于生态系统生产力(Waring et al., 1998; DeLucia et al., 2007; Goulden et al., 2011)。因此, 在前所未有的全球变化的大背景下, 理解草地生态系统生物量对氮沉降和降水的时空响应显得十分重要。
研究发现, 在低氮条件下, 植物为了获取氮, 会增加向地下分配的光合产物(Dingkuhn et al., 2007; Grechi et al., 2007); 随着外源氮的添加, 草地生态系统地上生物量显著增加(Lee et al., 2010), 分配到地下的生物量减少, 草地群落生产力提高(Bai et al., 2001)。然而, 不同植物种生物量的变化幅度存在差异(Pan et al., 2011), 随着处理年限的增加, 群落物种组成发生变化, 适应环境的植物物种生物量显著提高, 成为优势种(Bai et al., 2010)。在环境较为干旱时, 养分的迁移和传输均受到限制(Rouphael et al., 2012; He & Dijkstra, 2014), 添加的氮不易被植物快速吸收。随着降水量的增加(Hayes & Seastedt, 1987), 养分的可利用性增强(Tilman & Wedin, 1991), 增加的生物量更多地被分配到地上, 导致地上生物量与地下生物量的比值增加(Li et al., 2011)。目前, 地下生物量对增雨的响应尚没有完全统一的结论, 呈增加(Sala et al., 1988; Lauenroth & Sala, 1992; Wang et al., 2012; Kang et al., 2013)、减少(Knapp et al., 2001; Li et al., 2011)或不变的趋势(Fay et al., 2000; Weltzin et al., 2003; Zhou et al., 2009)。
一般而言, 添加氮和增雨交互作用能显著提高群落地上生物量和总生物量(Gao et al., 2011), 然而也可能导致植物群落组成单一化, 进而降低植物多样性(Harpole et al., 2007; 陆婷婷等, 2014)。同时有研究表明, 不同植被类型的生物量对于氮水交互作用的响应存在差异, 呈增加、减少或先增加后减少的趋势(李文娇等, 2015; Ren et al., 2017), 但尚未得到统一的结论。因此, 整合分析草地生态系统生物量如何响应氮添加和增雨及其交互作用非常有必要。
模型预测的结果表明, 未来草地生态系统仍将受到氮沉降加剧(Liu et al., 2013)和降水增加(Cholaw et al., 2003)的影响。据估计, 全球氮沉降水平在未来25年内还会加倍(Neff et al., 2002)。全球大气环流改变引起的降水格局发生显著变化(Sugiyama et al., 2010), 未来高纬度地区的降水量将增加, 大部分地区的单次强降水事件也将增加(Easterling et al., 2000; IPCC, 2012)。目前, 大多数关于草地生态系统生物量的野外控制实验的研究存在很大的局限性: 仅研究单一站点生物量变化及其影响因素; 仅研究氮或降水单一因素对生物量变化的影响; 仅研究生物量在短时间尺度上的变化及其影响因素(Vitousek & Howarth, 1991; Bai et al., 2008; LeBauer & Treseder, 2008; Fay et al., 2015)。
综合以上因素, 在全球尺度上探讨草地生态系统生物量在空间和时间尺度上对氮添加和增雨处理的响应, 有助于更好地应对未来氮沉降加剧和极端降水频发的全球变化, 为草地管理提供科学指导。针对上述现有问题, 本研究从46篇已发表的文献中, 收集全球41个野外实验站点时间跨度约为30年的1 350条独立数据, 进行整合响应比与回归分析, 用以研究草地生态系统生物量对氮增加和增雨两个独立因素及其交互作用的影响, 并进一步探讨生物量随氮添加梯度、增雨处理梯度的变化, 以及在空间格局和时间尺度上的响应。
1 材料和方法
1.1 数据来源
本文数据均提取自Web of ScienceTM和Google Scholar数据库中近30年间(即1990-2017年)有关氮添加和增雨处理对草地生态系统生物量影响的相关文献。通过对相关研究的数据及其来源的空间、时间信息进行收集, 建立数据库, 从而开展本文草地生态系统生物量在空间、时间尺度上对氮添加、增雨处理响应的整合研究。本研究共整合文献46篇, 独立数据1 350条, 整个数据库实验站点经度跨度247.16° (123.61° W-123.51° E), 纬度跨度82.18° (25.75° S-56.43° N), 集中分布在北半球中低纬度地区。实验年限涵盖1-10年, 年平均气温涵盖0.3-24.5 ℃ , 年降水量涵盖159-1 400 mm (图1)。图1
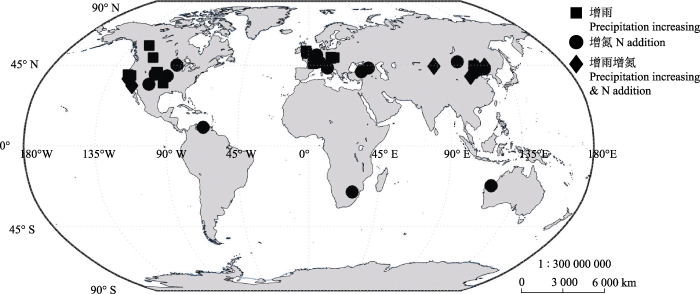 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1整合分析站点分布图。
Fig. 1The map of distribution of all the field experiments in this meta-analysis.
实验数据的筛选标准为: (1)实验设置位点需是野外原位草地生态系统, 非室内控制实验; (2)野外控制实验结果以图或表的形式说明实验组(如: 氮添加)和对照组的处理结果$\bar{X}_{T}$和$\bar{X}_{C}$; 重复次数NT和NC; 标准误SET和SEC; 标准偏差SDT和SDC; (3)单一实验处理至少包含两个处理梯度; (4)增雨处理仅包含雨量的变化, 不含降雨格局的变化; (5)在不同实验处理(如氮添加、增雨处理和同时增氮增雨处理)中, 测定相应变化指标, 主要包括草地生态系统地上、地下、总生物量及地上、地下生物量比。同时, 本研究收集包括野外实验站点经纬度、年平均气温(MAT)、年降水量(MAP)等辅助信息。对于缺失MAT、MAP信息的文献, 本研究通过R软件程序包, 利用Climatic Research Unit Climatology version 2.0 dataset (CRUCL 2.0, New et al., 2002)根据站点的经纬度信息提取相应的MAT和MAP数据(附录I)。
其次, 为保证数据的相对独立性, 发表在不同文献中的同一站点的同一实验数据被剔除, 不同期刊中同一控制实验的不同实验处理, 如氮添加浓度、氮添加形式, 或同一实验处理的不同实验年份视为一条独立数据。数据的提取方法包括从表格中的数据直接摘录和从图中间接提取, 使用Engauge Digitizer (Free Software Foundation, Boston, USA)软件数字化图以提取量化的数据, 并尽量按照统一标准减少人工处理误差, 生物量单位统一为g·m-2·a-1。
为探讨氮添加、增雨处理下, 草地生态系统生物量在不同气候条件和不同时间尺度上的响应, 我们将收集的数据分为两类: 第一类, 不同站点同一实验处理的年均结果, 用来探究不同气候条件上的响应; 第二类, 不同站点同一实验研究的逐年结果, 用来研究时间格局上的响应(表1)。
Table 1
表1
表1数据库结构及参数
Table 1
| 氮添加 Nitrogen addition | ||||
|---|---|---|---|---|
| 实验数量 Study numbers | 地上生物量 AGB | 地下生物量 BGB | 总生物量 TB | 地上地下生物量比 AGB/BGB |
| 年均 Annual mean | 109 | 56 | 59 | 42 |
| 年际 Inter-annual | 169 | 79 | 74 | 37 |
| 增雨处理 Precipitation increasing | ||||
| 实验数量 Study numbers | 地上生物量 AGB | 地下生物量 BGB | 总生物量 TB | 地上地下生物量比 AGB/BGB |
| 年均 Annual mean | 77 | 82 | 71 | 51 |
| 年际 Inter-annual | 62 | 90 | 98 | 32 |
| 增氮增雨处理 Nitrogen addition and precipitation increasing | ||||
| 实验数量 Study numbers | 地上生物量 AGB | 地下生物量 BGB | 总生物量 TB | 地上地下生物量比 AGB/BGB |
| 年均 Annual mean | 19 | 21 | 17 | 14 |
| 年际 Inter-annual | 21 | 32 | 27 | 11 |
新窗口打开|下载CSV
1.2 分析方法
本研究通过数据整合(meta-analysis, Hedges et al., 1999)的方法对表1所列的数据进行分析。利用响应比(response ratio, RR)以及对数响应比(log response ratio, LRR)量化各个参数(草地生态系统地上、地下、总生物量及地上地下生物量比)对氮添加、增雨处理以及同时增氮增雨处理的效应, 即:变异系数v的计算公式为:
其中, $\bar{X}_{C}$、SC和NC分别为对照处理下相应指标的平均值、标准偏差和样本量, $\bar{X}_{T} S_{T}$和${{N}_{\text{T}}}$分别为实验处理水平下相应指标的平均值、标准偏差和样本量。
指标的累积效应$(\bar{E})$通过求权重的方式进行计算, 即:
其中, n为所有实验的总数, ${{W}_{i}}$为第i个实验的权重, 计算方法为其变异系数的倒数, 即:
每个指标RR的计算是通过对$\bar{E}$进行反对数转换以降低偏倚, 使样本近似成正态分布(Hedegs et al., 1999; Koricheva et al., 2013; Lu et al., 2013)。为了在数据样本相对较小时更加精确地计算整合分析数值, 我们采用9 999次重复迭代的方法计算95%的置信区间来作为RR的异质性(Adams et al., 1997)。如果95%置信区间(95%CI)与1没有重叠表明变量有显著的正效应或负效应, 反之则认为变量间不存在显著效应。RR取对数后LRR的95%置信区间则与0比较是否存在显著差异。
将LRR进行标准化以排除处理浓度的影响, 即将单个实验处理水平以数据库平均值进行标准化, 以氮添加对地上生物量影响的标准化为例: 数据库平均施氮浓度为8.4 g·m-2·a-1, 即:
其中, ${{\bar{X}}_{\text{NT}}}$是标准化后的施氮响应值, 效应值${{\bar{X}}_{\text{T}}}$、${{\bar{X}}_{\text{C}}}$分别为原始实验处理和对照数值, 标准化LRR(normalized LRR) = log$\frac{{{{\bar{X}}}_{\text{NT}}}}{{{{\bar{X}}}_{\text{C}}}}$ (Liu et al., 2016)。其他指标标准化过程同理。
以上所有整合分析及整合回归计算由R软件里的metafor软件包实现(Viechtbauer, 2010)。同时, 使用填补法检验偏爱性对研究结果的影响, 结果表明整合分析仅在氮添加处理下低估了总生物量增加的程度, 其余研究结果均不受影响(附录II-V)。
2 结果
2.1 草地生态系统生物量对氮添加和增雨处理及增氮增雨处理的响应
氮添加显著增加草地生态系统的地上生物量和总生物量(37%, 32%, 图2A、2C), 增加生物量的地上部分分配比例(29%, 图2D)。增雨处理显著增加地上、地下和总生物量(41%, 10%, 23%, 图2A-2C), 同时显著增加地上部分的分配比例(25%, 图2D)。地下生物量对单独氮添加处理的响应不显著(图2B), 而同时进行增雨处理时, 地下生物量则显著增加(42%, 图2B), 且其促进作用显著高于增雨处理(10%, 图2B)。图2
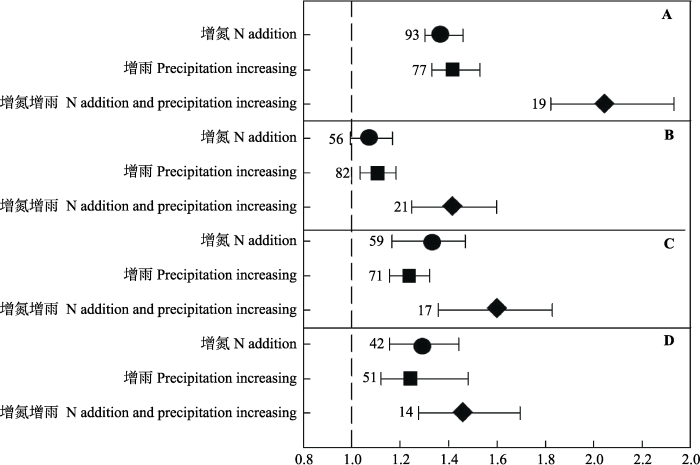 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2氮添加处理(圆圈)、增雨处理(正方形)及同时增氮增雨处理(菱形)对地上生物量(AGB)(A)、地下生物量(BGB)(B)、总生物量(TB)(C)及地上地下生物量比(AGB/BGB)(D)的影响(平均值± 95%CI)。图中数字代表独立研究数量。
Fig. 2Effects of N addition (in circle), precipitation increasing (in square) and N addition plus precipitation increasing (in rhombus) on aboveground biomass (AGB)(A), belowground biomass (BGB)(B), total biomass (TB)(C) and the ratio of aboveground biomass to belowground biomass (AGB/BGB)(D) (mean ± 95%CI). Numbers in the parentheses represent study number.
增氮增雨处理对地上、地下、总生物量(104%, 42%, 60%, 图2A-2C)均有显著的促进作用, 其中对地上生物量的促进作用显著高于地下生物量(104%, 42%, 图2A、2B), 从而显著增加地上地下生物量的比例(46%, 图2D)。
2.2 草地生态系统生物量对氮添加和增雨处理沿处理梯度的变化
氮添加和增雨处理对地上生物量的促进作用沿着氮添加处理水平(p < 0.01, 图3A)、增雨处理梯度(p < 0.001, 图3E)增加而增加, 但地下生物量、总生物量及地上地下生物量比的响应与处理水平并无显著相关性(图3B-3D, 3F-3H)。图3
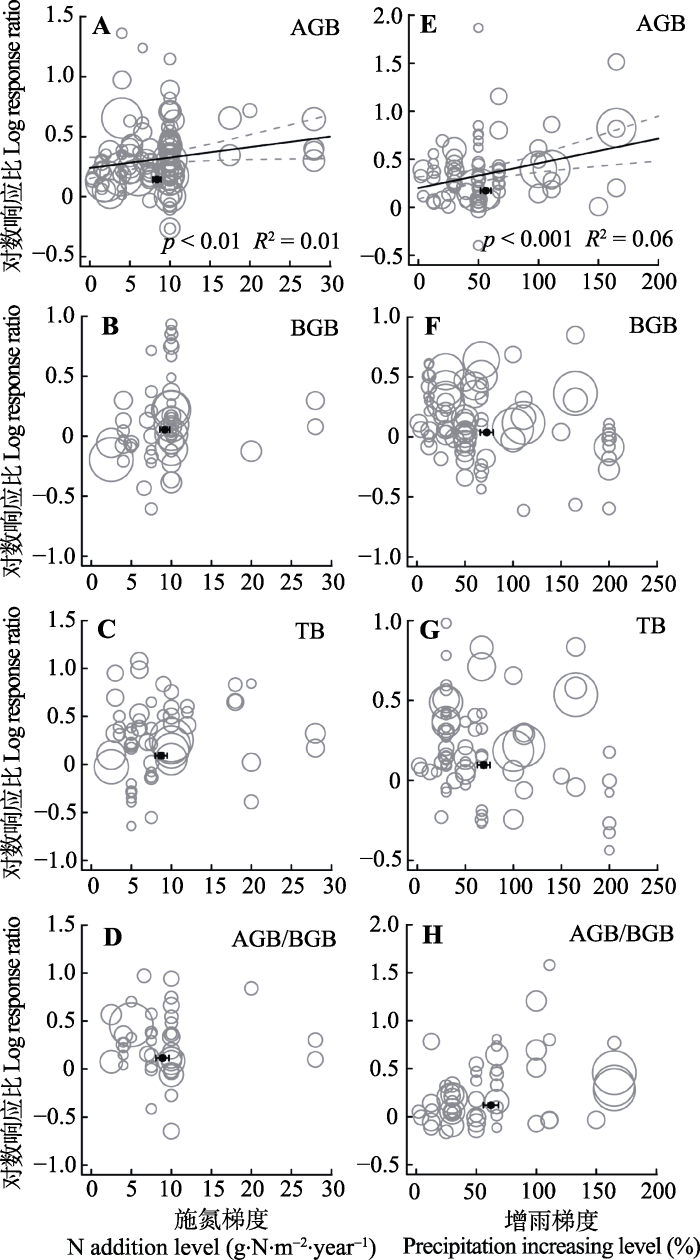 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3整合回归结果显示地上生物量(AGB)(A, E)、地下生物量(BGB)(B, F)、总生物量(TB)(C, G)和地上地下生物量比(AGB/BGB)(D, H)沿处理梯度的变化。A-D代表氮添加, E-H代表增雨处理, 圆圈的大小代表响应比在随机效应meta回归中的权重。圆圈越大, 权重越大, 贡献度越高。黑色圆点纵坐标为响应比平均值±标准误差, 横坐标为处理梯度平均值±标准误差。
Fig. 3Bubble plots of the meta-regression results between the response of aboveground biomass (AGB)(A, E), belowground biomass (BGB)(B, F), total biomass(TB)(C, G) and the ratio of aboveground biomass to belowground biomass (AGB/BGB)(D, H) to the treatment level of N addition and precipitation increasing. A-D represent the N addition treatment; E-H represent the precipitation increasing treatment. The size of the bubble is the relative weight of the effect size (response ratio, RR) in the random-effects meta- regression. Larger bubbles indicate study outcomes that contribute a great overall weight in meta-regression. The y-direction error bars of the black dots represent the standard error of the means of response ratio; the x-direction error bars represent the standard error of treatment level under N addition and precipitation increasing.
2.3 草地生态系统生物量对氮添加和增雨处理的响应随不同气候条件的变化
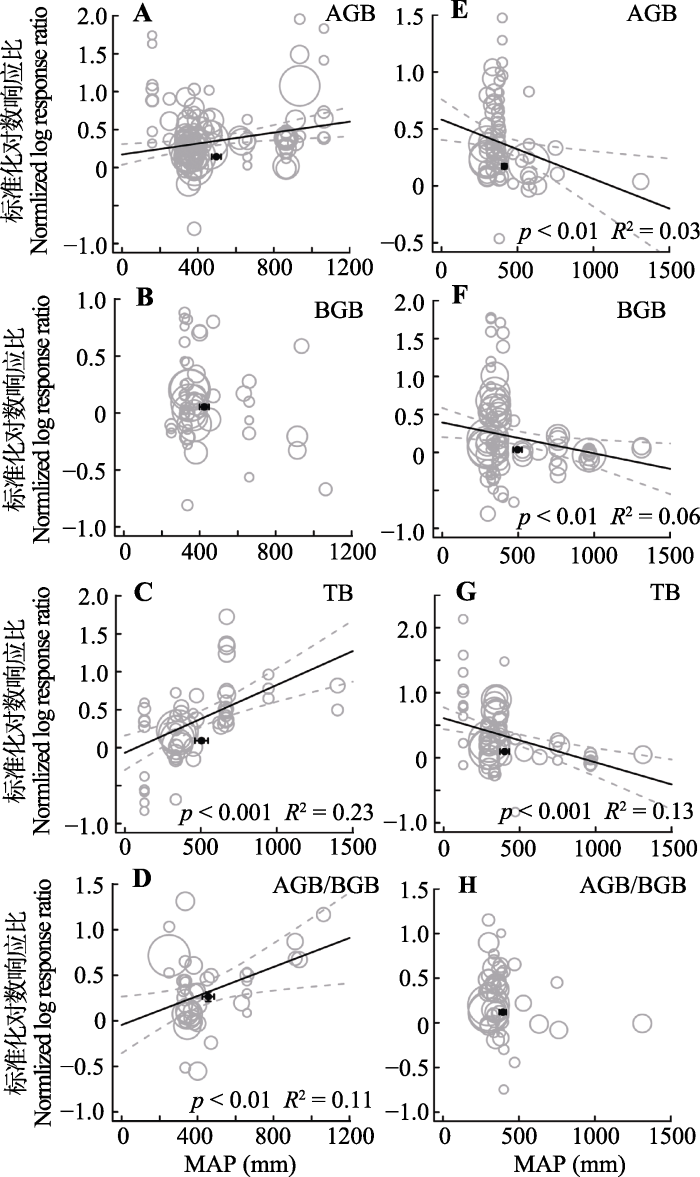
随着年均温升高, 氮添加对年均温较高的草地生态系统的地上生物量、总生物量及生物量向地上分配的促进作用显著增强(p < 0.001; p < 0.001; p < 0.01, 图4A、4C、4D)。随着年降水量增加, 氮添加对降水量较高的草地生态系统的地上生物量、总生物量及生物量向地上分配的促进作用增强(p < 0.001; p < 0.001; p < 0.01, 图5A、5C、5D)。图4
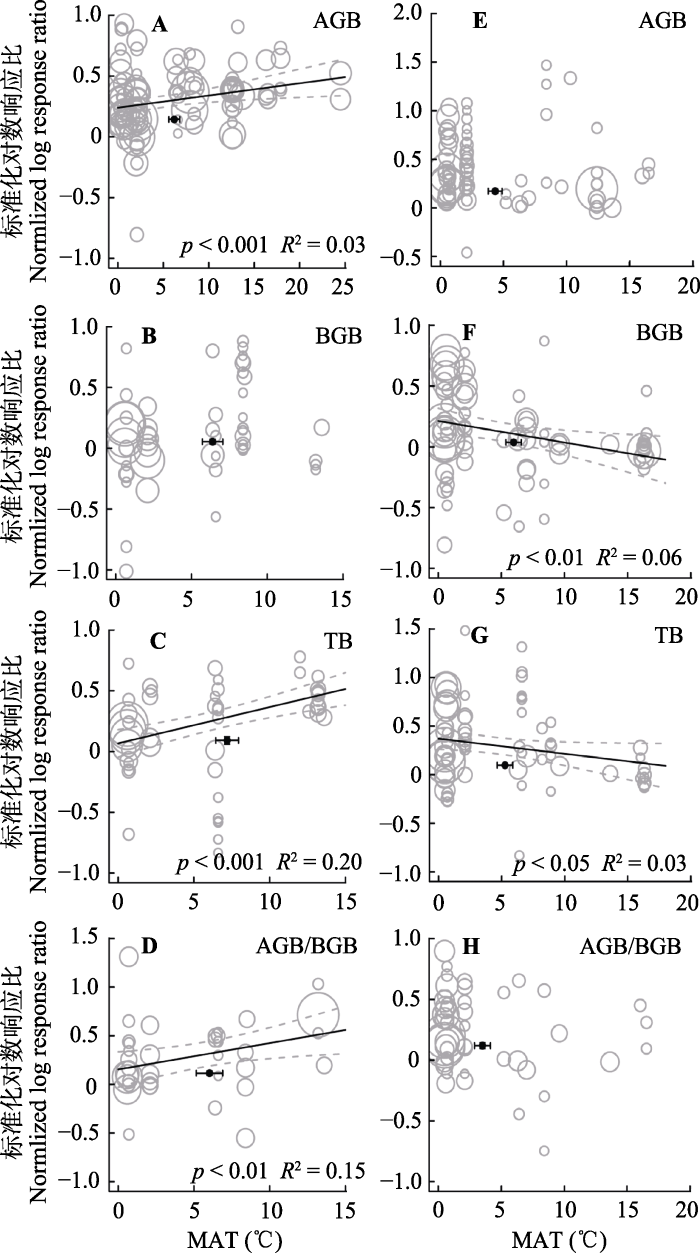 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4整合回归结果显示随着实验站点年平均气温(MAT)升高, 地上生物量(AGB)(A, E)、地下生物量(BGB)(B, F)、总生物量(TB)(C, G)和地上地下生物量比(AGB/BGB)(D, H)在不同处理下的变化。A-D代表氮添加, E-H代表增雨处理, 圆圈的大小代表响应比在随机效应meta回归中的权重。圆圈越大, 权重越大, 贡献度越高。黑色圆点纵坐标为响应比平均值±标准误差, 横坐标为处理梯度平均值±标准误差。
Fig. 4Bubble plots of the meta-regression results between the response of aboveground biomass (AGB)(A, E), belowground biomass (BGB)(B, F), total biomass (TB)(C, G) and the ratio of aboveground biomass to belowground biomass (AGB/BGB)(D, H) to the mean annual temperature (MAT) in the study sites. A-D represent the N addition treatment; E-H represent the precipitation increasing treatment. The size of the bubble is the relative weight of the effect size (response ratio, RR) in the random-effects meta-regression. Larger bubbles indicate study outcomes that contribute a great overall weight in meta- regression. The y-direction error bars of the black dots represent the standard error of the means of response ratio; the x-direction error bars represent the standard error of the means of mean annual temperature (MAT).
图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5整合回归结果显示随着实验站点年降水量(MAP)增加, 地上生物量(AGB)(A, E)、地下生物量(BGB)(B, F)、总生物量(TB)(C, G)和地上地下生物量比(AGB/BGB)(D, H)在不同处理下的变化。A-D代表氮添加, E-H代表增雨处理, 圆圈的大小代表响应比在随机效应meta回归中的权重。圆圈越大, 权重越大, 贡献度越高。黑色圆点纵坐标为响应比平均值±标准误差, 横坐标为处理梯度平均值±标准误差。
Fig. 5Bubble plots of the meta-regression results between the response of aboveground biomass (AGB)(A, E), belowground biomass (BGB)(B, F), total biomass(TB)(C, G) and the ratio of aboveground biomass to belowground biomass (AGB/BGB)(D, H) to the mean annual precipitation (MAP) in the study sites. A-D represent the N addition treatment; E-H represent the precipitation increasing treatment. The size of the bubble is the relative weight of the effect size (response ratio, RR) in the random-effects meta-regression. Larger bubbles indicate study outcomes that contribute a great overall weight in meta-regression. The y-direction error bars of the black dots represent the standard error of the means of response ratio; the x-direction error bars represent the standard error of the means of mean annual precipitation (MAP).
在年平均气温较高的草地生态系统中, 增雨处理对地下生物量、总生物量的促进作用显著低于年均温较低地区的草地生态系统(p < 0. 01; p < 0.05, 图4F、4G)。在年降水量较低的草地生态系统中, 增雨处理促进草地生态系统的地上、地下和总生物量。然而, 随着年降水量的增加, 增雨处理对生物量的促进作用逐渐减弱, 而抑制作用则逐渐增强, 且随年降水量越高抑制作用越强(p < 0.01; p < 0.01; p < 0.001, 图5E-5G)。
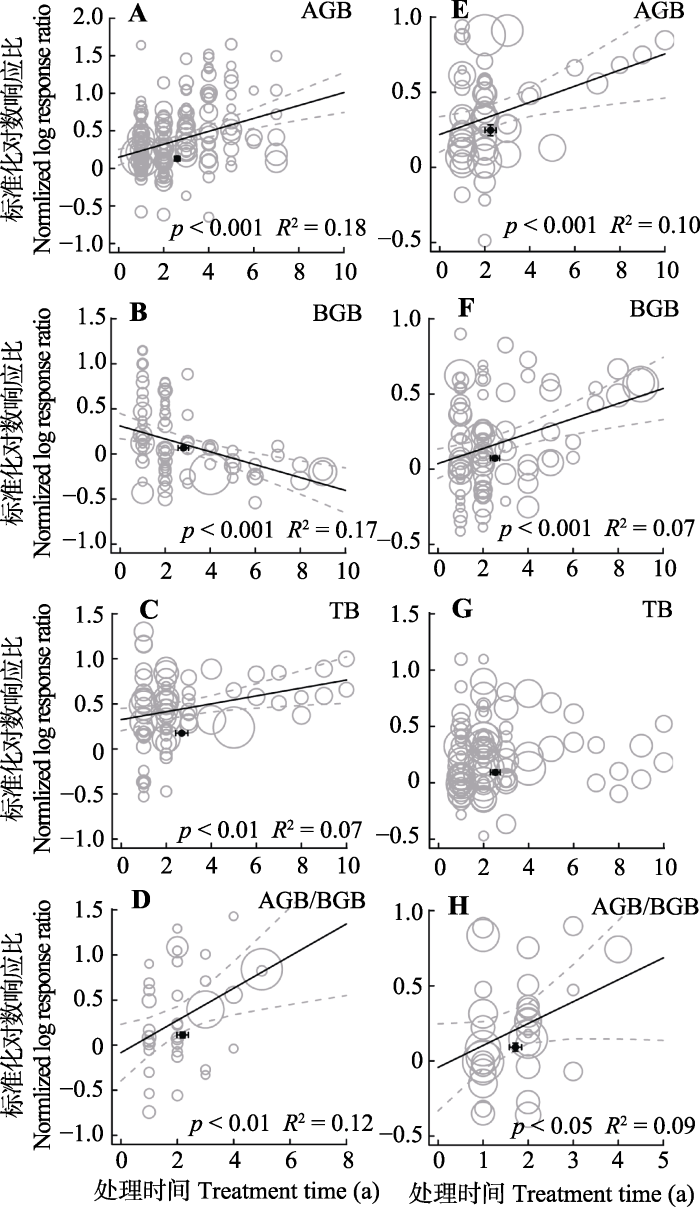
2.4 草地生态系统生物量对氮添加和增雨处理的响应随时间尺度的变化
随着处理年限的增加, 氮添加和增雨对地上生物量以及地上地下生物量分配比例的促进作用显著增强(p < 0.001; p < 0.001; p < 0.001; p < 0.05, 图6A、6E、6D、6H)。总生物量的增加仅在氮添加处理下有显著的增强趋势(p < 0.001, 图6C), 在增雨处理下没有明显的变化。早期施氮可以促进地下生物量, 但随施氮年限的增加, 地下生物量显著下降(p < 0.001, 图6B)。增雨处理下, 地下生物量随着处理年限的增加显著增加(p < 0.001, 图6H)。图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6整合回归结果显示随处理年限增加, 地上生物量(AGB)(A, E)、地下生物量(BGB)(B, F)、总生物量(TB)(C, G)和地上地下生物量比(AGB/BGB)(D, H)在不同处理下的变化。A-D代表氮添加, E-H代表增雨处理, 圆圈的大小代表响应比在随机效应meta回归中的权重。圆圈越大, 权重越大, 贡献度高。黑色圆点纵坐标为响应比平均值±标准误差, 横坐标为处理梯度平均值±标准误差。
Fig. 6Bubble plots of the meta-regression results between the response of aboveground biomass (AGB)(A, E), belowground biomass (BGB)(B, F), total biomass(TB)(C, G)and the ratio of aboveground biomass to belowground biomass (AGB/BGB)(D, H) to the study year. A-D represent the N addition treatment; E-H represent the precipitation increase treatment. The size of the bubble is the relative weight of the effect size (response ratio, RR) in the random-effects meta- regression. Larger bubbles indicate study outcomes that contribute a great overall weight in meta-regression. The y-direction error bars of the black dots represent the standard error of the means of response ratio; the x-direction error bars represent the standard error of the means of treatment time (a).
3 讨论
3.1 氮添加和增雨处理及两者交互作用对草地生态系统生物量的影响
氮添加对草地生态系统生产力的促进作用存在阈值(Fang et al., 2012)。植物生长发育受到氮限制时, 一定量的氮添加能够促进植物叶片的抗衰老过程(韩炳宏等, 2016), 提高光合速率、增加植被高度和叶面积指数(韩会阁等, 2015); 而当氮冗余时, 可能引起土壤酸化、NH4+和Al3+浓度增加, 对植物的生长产生毒害作用(Bobbink et al., 1998; Stevens et al., 2010), 抑制植物的生长发育, 从而降低植物生产力(Mack et al., 2004)。本研究结果中氮添加浓度达到30 g·m-2·a-1、增雨处理水平达到200%时, 地上生物量的促进作用仍有显著的增加趋势(图3A、3E), 说明草地生态系统的植物群落组成可能发生改变, 向着适应高氮高降雨的环境方向发展。因此, 生物量在高氮高降雨下仍有响应, 尚未达到阈值。随氮添加浓度提高, 草地地下、总生物量及地上地下分配比例没有明显的增加趋势(图3B-3D)。一般而言, 氮添加会增加地上生物量, 导致群落透光率降低, 植物对养分的竞争转变为对光的竞争, 加剧物种间的竞争排斥效应, 容易造成低矮植被的丢失(Tilman, 1985; Gough et al., 2000)。植被的改变会进一步影响地上和地下生物量的分配, 从而表现为其生产力增加趋势不明显。此外, 沿增雨处理梯度的提高, 地下、总生物量及地上地下分配比例也没有明显的增加趋势(图3F-3H)。尽管生长季后期降水量减少、土壤含水量下降, 多年生植物通过利用土壤深层水累积地上、地下生物量, 减少次年春季一年生植物可利用的水分(Corbin & D’Antonio, 2004)。但5 mm的有效降水量就能满足一年生植物基本完成生活史, 且一年生小禾草类植物能够根据降水量变化及时调整物候长短以适应生长(闫建成等, 2013)。因此, 有研究表明增雨能通过增强一年生植物的光竞争能力, 减少多年生植物的生物量(毛伟等, 2014)。此外, 降雨量增加能够提高土壤有效水分, 促进光合作用(Thomey et al., 2011), 但也会影响土壤中的其他因素, 如促进土壤氮矿化, 从而提高土壤无机氮含量(Burke et al., 1997)。土壤无机氮含量的增加会减少植被对地下生物量的分配(陈骥等, 2013), 同时, 水分增加还会限制根系呼吸, 从而抑制根系的生长, 导致地下生物量降低(Dukes et al., 2005)。
氮添加的同时进行增雨处理显著增加地下生物量, 而且效果显著高于单独增雨(42%, 10%, 图 2B), 说明水分和土壤氮含量对草地生态系统生产力的影响存在交互作用(Bell et al., 2008; Brueck et al., 2010; Li et al., 2011)。在环境较为干旱时, 养分的迁移和传输均受到限制, 随着降雨量的增加, 土壤中养分限制得以解除, 从而提高养分的可利用性, 促进植物对养分的吸收利用, 最终提高生态系统生产力(Li et al., 2011), 因此氮水的交互作用往往呈协同效应(Niu et al., 2009)。
3.2 草地生态系统生物量对氮添加和增雨处理的响应在不同气候变化尺度下的格局
较大区域尺度上, 温度、降水等环境因子是影响植物生产力的关键气候因素(Fang et al., 2001; Swemmer & Knapp, 2008; Yang et al., 2008)。在氮添加处理下, 温暖性气候区的草地生态系统的地上生物量、总生物量及生物量向地上分配的促进作用较高(图4A、4C、4D), 这可能是由于从温暖性气候区到寒冷性气候区, 年均温逐渐降低, 植被生长季明显缩短(陈智等, 2014), 导致生产力受到的促进作用降低。除了温度的调控作用之外, 不同气候区的陆地生态系统也同时受到水分状况的影响, 无论是热带、亚热带还是温带区域内, 在其相对湿润的气候区具有更高的生产力(陈智等, 2014)。本研究的整合结果也支持该观点, 即在海洋性气候区的草地生态系统中, 施氮处理对地上生物量、总生物量及生物量向地上分配的促进作用显著高于温带大陆性气候区的草地生态系统(图5A、5C、5D)。在增雨处理下, 寒冷性气候区的草地生态系统的地下生物量、总生物量受到的促进作用显著高于温暖性气候区的草地生态系统(图4F、4G)。一般而言, 低温条件下, 草地生态系统土壤碳矿化速率较低(黄文华等, 2014)、微生物活性较弱, 会受到更多的氮限制(Fay et al., 2015)。增雨使土壤中可利用性氮增加(Burke et al., 1997), 从而缓解草地生态系统由低温引发的氮限制, 且研究发现寒冷性气候区的植物生物量比温暖性气候区的植物生物量对氮添加的响应更为敏感(Xia & Wan, 2008)。因此, 相比于荒漠草地、典型草地和温带湿润草地, 冻原和高山草地等受温度限制的草地生态系统增雨后生物量增加更为明显。未来在高纬度、赤道太平洋附近及中纬度的湿润气候区中年降水量呈增加趋势, 而大部分中纬度和亚热带干旱气候区的年降水量将进一步减少(Stocker et al., 2013), 这种区域性的降水量改变将使优势物种的分布区域向高海拔偏移(Kelly & Goulden, 2008), 可能进一步促进寒冷性气候区草地生态系统生产力。
3.3 草地生态系统生物量对氮添加和增雨处理的响应在时间尺度上的格局
长期氮添加处理对地下生物量的促进作用在处理后期转为抑制作用(图6B), 这表明草地生态系统可能发生物种组成的改变(Clark & Tilman, 2008), 植被由深根系向浅根系物种发生转变。长期增雨处理下地上、地下生产量的促进作用都有增加的趋势(图6E、6F), 可能是在水分条件充足时, 植物个体生长得到促进, 地上生物量持续累积, 根系生物量随着个体植株的增大也得到促进(Li et al., 2011)。一般认为, 降水变化对植物地上初级生产力的影响具有累积效应, 群落地上初级生产力与年降水量在空间尺度上呈线性相关关系, 在时间尺度上呈二次曲线关系(王玉辉和周广胜, 2004)。然而, 总生物量的促进作用(图6G)在长期增雨处理下没有显著变化, 说明长期的水分添加可能引起淋溶(Zhu et al., 2016), 使得土壤中氮流失。因此, 生产力长期持续的增加需要外源营养元素添加(Ren et al., 2017)。长期氮、水分的添加有利于改善草地生态系统的养分条件, 促进植物迅速生长, 加快养分周转, 进而有利于竞争得到更多的养分。 但是在养分贫瘠时, 植物生长较慢, 滞留吸收养分的能力较强, 反而可以防止养分损失或被其他竞争性植物吸收利用(Yuan et al., 2006)。长期氮添加、增雨及同时增氮增雨三种处理下, 地上地下生物量比都显著增加, 生物量更多地向地上分配(29%、25%、46%, 图2D), 且随着处理年限的增加, 比值也呈现逐渐增加的趋势(图6D、6H)。在短期时间尺度上, 地上生物量的持续增加有利于在农牧业活动中收获更多作物; 在长时间尺度上, 若缺乏相应合理的草地生态系统管理措施, 则可能因过度放牧、割草等活动打破其系统平衡, 对草地生态系统生物量造成负面影响。

附录
Supplement
附录I 本文46篇文献中的实验站点及环境信息Supplement I All the forty-six field experiments in this meta-analysis and the related environmental factor
http://www.plant-ecology.com/fileup/PDF/cjpe.2018.0056-S1.pdf
附录II 修建填补法检验meta分析发表偏倚
Supplement II Results of publication bias tests using Egger’s regression and the comparison of adjusted effect sizes using the trim and fill method and no-adjusted effect sizes
http://www.plant-ecology.com/fileup/PDF/cjpe.2018.0056-S2.pdf
附录III 氮添加处理下地上、地下、总生物量及地上地下生物量比的漏斗图
Supplement III Funnel plots of AGB (A), BGB (B), TB (C) and AGB/BGB (D) under N addition treatment
http://www.plant-ecology.com/fileup/PDF/cjpe.2018.0056-S3.pdf
附录IV 增雨处理下地上、地下、总生物量及地上地下生物量比的漏斗图
Supplement IV Funnel plots of AGB (A), BGB (B), TB (C) and AGB/BGB (D) under precipitation increasing treatment
http://www.plant-ecology.com/fileup/PDF/cjpe.2018.0056-S4.pdf
附录V 增氮增雨处理下地上、地下、总生物量及地上地下生物量比的漏斗图
Supplement V Funnel plots of AGB (A), BGB (B), TB (C) and AGB/BGB (D) under N addition and precipitation increasing treatment
http://www.plant-ecology.com/fileup/PDF/cjpe.2018.0056-S5.pdf
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1890/0012-9658(1997)078[1277:RTFMAO]2.0.CO;2URL [本文引用: 1]
[本文引用: 1]
DOI:10.1111/j.1365-2486.2009.01950.xURL [本文引用: 1]

Nitrogen (N) deposition is widely considered an environmental problem that leads to biodiversity loss and reduced ecosystem resilience; but, N fertilization has also been used as a management tool for enhancing primary production and ground cover, thereby promoting the restoration of degraded lands. However, empirical evaluation of these contrasting impacts is lacking. We tested the dual effects of N enrichment on biodiversity and ecosystem functioning at different organizational levels (i.e., plant species, functional groups, and community) by adding N at 0, 1.75, 5.25, 10.5, 17.5, and 28.0 g N m612 yr611 for four years in two contrasting field sites in Inner Mongolia: an undisturbed mature grassland and a nearby degraded grassland of the same type. N addition had both quantitatively and qualitatively different effects on the two communities. In the mature community, N addition led to a large reduction in species richness, accompanied by increased dominance of early successional annuals and loss of perennial grasses and forbs at all N input rates. In the degraded community, however, N addition increased the productivity and dominance of perennial rhizomatous grasses, with only a slight reduction in species richness and no significant change in annual abundance. The mature grassland was much more sensitive to N-induced changes in community structure, likely as a result of higher soil moisture accentuating limitation by N alone. Our findings suggest that the critical threshold for N-induced species loss to mature Eurasian grasslands is below 1.75 g N m612 yr611, and that changes in aboveground biomass, species richness, and plant functional group composition to both mature and degraded ecosystems saturate at N addition rates of approximately 10.5 g N m612 yr611. This work highlights the tradeoffs that exist in assessing the total impact of N deposition on ecosystem function.
DOI:10.1890/07-0992.1URLPMID:18724724 [本文引用: 1]
Understanding how the aboveground net primary production (ANPP) of arid and semiarid ecosystems of the world responds to variations in precipitation is crucial for assessing the impacts of climate change on terrestrial ecosystems. Rain-use efficiency (RUE) is an important measure for acquiring this understanding. However, little is known about the response pattern of RUE for the largest contiguous natural grassland region of the world, the Eurasian Steppe. Here we investigated the spatial and temporal patterns of ANPP and RUE and their key driving factors based on a long-term data set from 21 natural arid and semiarid ecosystem sites across the Inner Mongolia steppe region in northern China. Our results showed that, with increasing mean annual precipitation (MAP), (1) ANPP increased while the interannual variability of ANPP declined, (2) plant species richness increased and the relative abundance of key functional groups shifted predictably, and (3) RUE increased in space across different ecosystems but decreased with increasing annual precipitation within a given ecosystem. These results clearly indicate that the patterns of both ANPP and RUE are scale dependent, and the seemingly conflicting patterns of RUE in space vs. time suggest distinctive underlying mechanisms, involving interactions among precipitation, soil N, and biotic factors. Also, while our results supported the existence of a common maximum RUE, they also indicated that its value could be substantially increased by altering resource availability, such as adding nitrogen. Our findings have important implications for understanding and predicting ecological impacts of global climate change and for management practices in arid and semiarid ecosystems in the Inner Mongolia steppe region and beyond.
DOI:10.1007/s00248-007-9333-zURLPMID:18246293 [本文引用: 1]
Global climate change models indicate that storm magnitudes will increase in many areas throughout southwest North America, which could result in up to a 25% increase in seasonal precipitation in the Big Bend region of the Chihuahuan Desert over the next 5002years. Seasonal precipitation is a key limiting factor regulating primary productivity, soil microbial activity, and ecosystem dynamics in arid and semiarid regions. As decomposers, soil microbial communities mediate critical ecosystem processes that ultimately affect the success of all trophic levels, and the activity of these microbial communities is primarily regulated by moisture availability. This research is focused on elucidating soil microbial responses to seasonal and yearly changes in soil moisture, temperature, and selected soil nutrient and edaphic properties in a Sotol Grassland in the Chihuahuan Desert at Big Bend National Park. Soil samples were collected over a 3-year period in March and September (2004–2006) at 0–1502cm soil depth from 12 3 × 302m community plots. Bacterial and fungal carbon usage (quantified using Biolog 96-well micro-plates) was related to soil moisture patterns (ranging between 3.0 and 14%). In addition to soil moisture, the seasonal and yearly variability of soil bacterial activity was most closely associated with levels of soil organic matter, extractable NH 4 –N, and soil pH. Variability in fungal activity was related to soil temperatures ranging between 13 and 2602°C. These findings indicate that changes in soil moisture, coupled with soil temperatures and resource availability, drive the functioning of soil-microbial dynamics in these desert grasslands. Temporal patterns in microbial activity may reflect the differences in the ability of bacteria and fungi to respond to seasonal patterns of moisture and temperature. Bacteria were more able to respond to moisture pulses regardless of temperature, while fungi only responded to moisture pulses during cooler seasons with the exception of substantial increased magnitudes in precipitation occurring during warmer months. Changes in the timing and magnitude of precipitation will alter the proportional contribution of bacteria and fungi to decomposition and nitrogen mineralization in this desert grassland.
DOI:10.1046/j.1365-2745.1998.8650717.xURL [本文引用: 1]
The effects of increased atmospheric nitrogen inputs, from both NO y and NH x, on diversity in various semi-natural and natural ecosystems are reviewed. The severity of these impacts depends on abiotic conditions (e.g. buffering capacity, soil nutrient status and soil factors that influence the nitrification potential and nitrogen immobilization rate) in the particular system. The sensitivity of fresh water ecosystems, wetlands and bogs, species-rich grasslands, heathlands and field layer of forests, all of which have conservational value, are discussed in detail. The most important effects of nitrogen deposition are: (i) accumulation of nitrogenous compounds resulting in enhanced availability of nitrate or ammonium; (ii) soil-mediated effects of acidification; and (iii) increased susceptibility to secondary stress factors. Long-term nitrogen enrichment has gradually increased the availability of nitrogen in several vegetation types, leading to competitive exclusion of characteristic species by more nitrophilic plants, especially under oligo- to mesotrophic soil conditions. Soil acidification (with losses of buffering capacity and increased concentrations of toxic metals) is especially important after nitrification of ammonium in weakly buffered environments: acid-resistant plant species then become dominant at the expense of the often rare plants typical of intermediate pH. The related change in the balance between ammonium and nitrate may also affect the performance of several species. The susceptibility of plant species to secondary stress factors (pathogens; frost and drought) may be affected by air-borne nitrogen but data are only available for a few communities (e.g. dry heathlands). Most global biodiversity is contained within natural and semi-natural vegetation. It is thus crucial to control emissions of nitrogenous compounds to the atmosphere, in order to reduce or prevent effects on diversity in these systems. Most research has focused on acidification in forestry stands and lakes and on the effects on trees. We highlight serious gaps in knowledge of other ecosystems.
DOI:10.1007/s11104-009-0128-5URL [本文引用: 1]
Productivity of semiarid grasslands is primarily limited by seasonal rainfall amount and becomes increasingly limited by nutrient availability under wet conditions. Interactive effects of water and N availability on grassland productivity and parameters related to water use were studied on a grassland site in Inner Mongolia, China, in a 2-factorial experiment with two levels of water (rainfed: 15802mm; irrigated: 839 (N0) and 972 (N1) mm) and N supply (0 or 18002kg02N ha 611 ). RUE was calculated from ANPP and cumulative water supply. Bare soil evaporation (E) was calculated from climatic data and leaf area dynamics, and percolation (D) and transpiration (T) were estimated with HYDRUS-1D. Water-use efficiency (WUE, ANPP / (T + D)) and transpiration efficiency (TE, ANPP / T) were calculated. Resource availability had pronounced effects on the water-use efficiency of semiarid grassland. RUE, WUE, and TE all decreased under irrigated compared to rainfed conditions and were significantly increased with N fertilizer application at both levels of water supply. While the irrigation effect on parameters of water-use efficiency were accordingly reflected in stable carbon isotope signatures, N supply resulted in significantly less negative δ 13 C-values under rainfed but not irrigated conditions. It is concluded, that spatial or temporal gradients in resource availability have pronounced effects on the water-use efficiency of semiarid grassland. The decrease of water use-efficiency under high water supply was related to differences in TE and not to a relative increase of unproductive water loss. Carbon isotope discrimination was highly correlated with WUE and TE, but can be a poor predictor of RUE.
DOI:10.2307/2266128URL [本文引用: 2]
Spatial variability that occurs at large scales has long been used by ecologists as a tool to examine the controls over ecosystem structure and function. Correlations of control variables such as climatic factors and response variables such as vegetation and soil carbon storage across broad regions have played a crucial role in predicting the response of ecosystems to global climate change. Despite the importance of these large-scale space-for-time substitutions, there are substantial limitations. One of these limitations is that many of the possible control factors covary with one another, and only some of the important control factors actually exist in large-scale databases. Thus, the true proximal controls may be difficult to identify. A second limitation is that models of spatial variability may not be appropriately applied to temporal variability. In this paper, we utilize a new approach to determine the extent to which N availability may constrain aboveground primary productivity in the Central Grassland region of the U.S. The strong relationship between average annual primary production and average annual precipitation found in spatial patterns in ecosystems globally has often been interpreted as evidence of a fundamental water limitation. However, temporal variation in annual aboveground net primary production (ANPP) indicates that other factors constrain production. We generated a spatial and temporal database for annual aboveground net primary production and annual net N mineralization by linking a database of input variables (precipitation, temperature, and soils) with predictive models. We generated independent data sets of aboveground net primary production and net N mineralization by using regression models to predict aboveground net primary production, and the Century model to simulate net N mineralization. Our analyses indicate that net primary production and net N mineralization both increase with mean annual precipitation; thus, it is not possible to separate the extent to which ANPP is controlled by water or N availability. Nitrogen use efficiency (NUE) increased with increasing precipitation across the region. Aboveground net primary production decreased with increasing temperature across the region, while N mineralization increased slightly, leading to decreasing (NUE) with increasing temperature. At high precipitation levels, aboveground net primary production increased and N mineralization decreased slightly with increasing soil fineness. Nitrogen use efficiency generally increased with increasing pools of soil organic matter, likely because in grasslands, the proportion of recalcitrant organic matter increases with the total organic matter pools. A comparison of interannual variation in net N mineralization with average spatial variation indicated a high degree of inertia in the response of N availability to precipitation levels. Our simulation results as well as field results of Lauenroth and Sala (1992) raise important questions about the applicability of space-for-time substitutions when dealing with ecosystem function. The structure of the systems appears to provide important constraints on the temporal variability that are not evident in an analysis of spatial variability.
DOI:10.3969/j.issn.1009-5500.2013.06.020URL [本文引用: 1]
碳、氮循环是陆地生态系统化学循环和能量流动的两大重要过程,二者紧密相连;土壤呼吸是陆地碳循环的重要过程,也是陆地生态系统与大气之间进行交换的主要途径。由于施肥等人为因素导致了陆地生态系统氮素的增加。不同陆地生态系统对这一过程做出了不同的响应。综述了不同生态系统土壤呼吸对模拟氮沉降的响应方式和机理,分析了氮素添加对土壤呼吸影响的不确定性,并在此基础上对未来研究方向进行了探讨和展望。
DOI:10.3969/j.issn.1009-5500.2013.06.020URL [本文引用: 1]
碳、氮循环是陆地生态系统化学循环和能量流动的两大重要过程,二者紧密相连;土壤呼吸是陆地碳循环的重要过程,也是陆地生态系统与大气之间进行交换的主要途径。由于施肥等人为因素导致了陆地生态系统氮素的增加。不同陆地生态系统对这一过程做出了不同的响应。综述了不同生态系统土壤呼吸对模拟氮沉降的响应方式和机理,分析了氮素添加对土壤呼吸影响的不确定性,并在此基础上对未来研究方向进行了探讨和展望。
DOI:10.3969/j.issn.1001-7410.2014.04.03URL [本文引用: 2]
Terrestrial vegetation fixes CO from atmosphere via photosynthesis. Understanding the spatial pattern and regional characteristics of the terrestrial ecosystem carbon uptake is of great significance for accurately predicting and assessing the global carbon budget and developing efficient regional ecosystem management policy. In this study, we analyzed the spatial pattern and regional characteristics of terrestrial ecosystem carbon fluxes (Gross Primary Productivity(GPP), Ecosystem Respiration (RE), and Net Ecosystem Productivity (NEP)) based on long-term observational data from ChinaFLUX and literature on studies in the Northern Hemisphere. A total of 732 site-years out of 233 flux sites were included in the analysis. These sites were distributed across the Asian, European and North American regions, spanning from 2.97 N to 74.47 N in latitude, and from 148.88 W to 161.34 E in longitude. These sites covered six climate zones: tropical, subtropical, temperate, boreal, arctic and subarctic, and alpine, and were grouped into four ecosystem types:forests (107 sites), grasslands (65 sites), croplands (33 sites) and wetlands (28 sites). The results showed that GPP and RE linearly decreased with an increase in latitude, with one degree increase in latitude, GPP and RE declined by 22.9g C/m/a, while there was no apparent latitudinal trend in NEP. GPP and NEP were higher in cropland and forest ecosystems than in grassland and wetland ecosystems. RE was highest in forest ecosystems (1185 641g C/m/a on average)but did not differ among other ecosystems. There were no differences among the Asian, European and North-American regions in GPP, RE and NEP of forest, cropland, and wetland. Only GPP and RE in the European grasslands was averaged at 1472 473g C/m/a and 1236 452g C/m/a respectively, which were notably higher than those in Asia and North-America. GPP, RE and NEP tended to decrease from warm environments to cold environments, and exhibited higher values in humid areas under the regulation of water condition. Our findings demonstrate that there are spatial variations but no regional differences in terrestrial ecosystem carbon fluxes in the Northern Hemisphere the differences are significant among biomes and ecosystems types. It shows that environmental conditions (temperature and water) and human activities are important factors commonly regulating the terrestrial ecosystem carbon fluxes in the Northern Hemisphere.
DOI:10.3969/j.issn.1001-7410.2014.04.03URL [本文引用: 2]
Terrestrial vegetation fixes CO from atmosphere via photosynthesis. Understanding the spatial pattern and regional characteristics of the terrestrial ecosystem carbon uptake is of great significance for accurately predicting and assessing the global carbon budget and developing efficient regional ecosystem management policy. In this study, we analyzed the spatial pattern and regional characteristics of terrestrial ecosystem carbon fluxes (Gross Primary Productivity(GPP), Ecosystem Respiration (RE), and Net Ecosystem Productivity (NEP)) based on long-term observational data from ChinaFLUX and literature on studies in the Northern Hemisphere. A total of 732 site-years out of 233 flux sites were included in the analysis. These sites were distributed across the Asian, European and North American regions, spanning from 2.97 N to 74.47 N in latitude, and from 148.88 W to 161.34 E in longitude. These sites covered six climate zones: tropical, subtropical, temperate, boreal, arctic and subarctic, and alpine, and were grouped into four ecosystem types:forests (107 sites), grasslands (65 sites), croplands (33 sites) and wetlands (28 sites). The results showed that GPP and RE linearly decreased with an increase in latitude, with one degree increase in latitude, GPP and RE declined by 22.9g C/m/a, while there was no apparent latitudinal trend in NEP. GPP and NEP were higher in cropland and forest ecosystems than in grassland and wetland ecosystems. RE was highest in forest ecosystems (1185 641g C/m/a on average)but did not differ among other ecosystems. There were no differences among the Asian, European and North-American regions in GPP, RE and NEP of forest, cropland, and wetland. Only GPP and RE in the European grasslands was averaged at 1472 473g C/m/a and 1236 452g C/m/a respectively, which were notably higher than those in Asia and North-America. GPP, RE and NEP tended to decrease from warm environments to cold environments, and exhibited higher values in humid areas under the regulation of water condition. Our findings demonstrate that there are spatial variations but no regional differences in terrestrial ecosystem carbon fluxes in the Northern Hemisphere the differences are significant among biomes and ecosystems types. It shows that environmental conditions (temperature and water) and human activities are important factors commonly regulating the terrestrial ecosystem carbon fluxes in the Northern Hemisphere.
DOI:10.1007/BF02915400URL [本文引用: 1]
This paper applies the newest emission scenarios of the sulfur and greenhouse gases, namely IPCC SRES A2 and B2 scenarios, to investigate the change of the North China climate with an atmosphere-ocean coupled general circulation model. In the last three decades of the 21st century, the global warming enlarges the land-sea thermal contrast, and hence, causes the East Asian summer (winter) monsoon circulation to be strengthened (weakened). The rainfall seasonality strengthens and the summer precipitation increases significantly in North China. It is suggested that the East Asian rainy area would expand northward to North China in the last three decades of the 21st century. In addition, the North China precipitation would increase significantly in September. In July, August, and September, the interannual variability of the precipitation enlarges evidently over North China, implying a risk of flooding in the future.
DOI:10.1038/nature06503URLPMID:18256670 [本文引用: 1]
Abstract Rates of atmospheric deposition of biologically active nitrogen (N) are two to seven times the pre-industrial rates in many developed nations because of combustion of fossil fuels and agricultural fertilization. They are expected to increase similarly over the next 50 years in industrializing nations of Asia and South America. Although the environmental impacts of high rates of nitrogen addition have been well studied, this is not so for the lower, chronic rates that characterize much of the globe. Here we present results of the first multi-decadal experiment to examine the impacts of chronic, experimental nitrogen addition as low as 10 kg N ha(-1) yr(-1) above ambient atmospheric nitrogen deposition (6 kg N ha(-1) yr(-1) at our site). This total input rate is comparable to terrestrial nitrogen deposition in many industrialized nations. We found that this chronic low-level nitrogen addition rate reduced plant species numbers by 17% relative to controls receiving ambient N deposition. Moreover, species numbers were reduced more per unit of added nitrogen at lower addition rates, suggesting that chronic but low-level nitrogen deposition may have a greater impact on diversity than previously thought. A second experiment showed that a decade after cessation of nitrogen addition, relative plant species number, although not species abundances, had recovered, demonstrating that some effects of nitrogen addition are reversible.
DOI:10.1890/02-0744URL [本文引用: 1]
DOI:10.1111/j.1365-2486.2007.01365.xURL [本文引用: 1]
Carbon-use efficiency (CUE), the ratio of net primary production (NPP) to gross primary production (GPP), describes the capacity of forests to transfer carbon (C) from the atmosphere to terrestrial biomass. It is widely assumed in many landscape-scale carbon-cycling models that CUE for forests is a constant value of 0.5. To achieve a constant CUE, tree respiration must be a constant fraction of canopy photosynthesis. We conducted a literature survey to test the hypothesis that CUE is constant and universal among forest ecosystems. Of the 60 data points obtained from 26 papers published since 1975, more than half reported values of GPP that were not estimated independently from NPP; values of CUE calculated from independent estimates of GPP were greater than those calculated from estimates of GPP derived from NPP. The slope of the relationship between NPP and GPP for all forests was 0.53, but values of CUE varied from 0.23 to 0.83 for different forest types. CUE decreased with increasing age, and a substantial portion of the variation among forest types was caused by differences in stand age. When corrected for age the mean value of CUE was greatest for temperate deciduous forests and lowest for boreal forests. CUE also increased as the ratio of leaf mass-to-total mass increased. Contrary to the assumption of constancy, substantial variation in CUE has been reported in the literature. It may be inappropriate to assume that respiration is a constant fraction of GPP as adhering to this assumption may contribute to incorrect estimates of C cycles. A 20% error in current estimates of CUE used in landscape models (i.e. ranging from 0.4 to 0.6) could misrepresent an amount of C equal to total anthropogenic emissions of CO 2 when scaled to the terrestrial biosphere.
URL [本文引用: 1]
ABSTRACT There is a new interest in plant morphogenesis and architecture because molecular genetics is providing new information on their genetic and physiological control. From a crop modeller's point of view, this requires particular attention paid to the regulation of sinks associated with organ development, as well as their interactions with assimilate sources. Existing agronomic and architectural crop models are not capable of simulating such interactions. A conceptual framework is presented for the analysis and simulation of crop growth driven by either assimilate source or sink dynamics, building on the assumption that meristems are the main sites in the plant architecture where sinks are initiated and adjusted to resources. Among the numerous sink ource feedbacks to be considered are sensing of the plant's resource and stress status by meristems (enabling adjustment of morphogenesis), as well as transitory reserves, organ senescence and end-product inhibition of photosynthesis (necessary for the plant to cope with acute imbalances). These feedbacks are to a large extent related to sugar metabolism and can be explained with recent molecular findings on the prominent place in plant development of sugar sensing and the regulation of sucrose cleavage at sink sites. A model integrating these phenomena in a simplified manner, called EcoMeristem, was developed and is being applied in phenotyping for functional-genomics studies on rice. Theoretical evidence and model sensitivity analyses suggested that sink regulation during vegetative growth has a strong effect on plant vigour and growth rate, even at given levels of leaf photosynthetic capacity. However, the usefulness of complex, whole-plant models such as EcoMeristem for heuristic phenotyping approaches remains to be demonstrated. Specific problems are related to the stability of process-based crop parameters across environments, as well as the measurement of such crop parameters that are inaccessible to direct observation. But it is argued that integrated, structural-functional models may be the only means to quantify complex traits, such as those governing adaptive morphology (phenotypic plasticity). Furthermore, such models may be well suited to develop improved plant type concepts in silico.
DOI:10.1371/journal.pbio.0030319URLPMID:1182693 [本文引用: 1]
In this century, increasing concentrations of carbon dioxide (CO2) and other greenhouse gases in the Earth's atmosphere are expected to cause warmer surface temperatures and changes in precipitation patterns. At the same time, reactive nitrogen is entering natural systems at unprecedented rates. These global environmental changes have consequences for the functioning of natural ecosystems, and responses of these systems may feed back to affect climate and atmospheric composition. Here, we report plant growth responses of an ecosystem exposed to factorial combinations of four expected global environmental changes. We exposed California grassland to elevated CO2, temperature, precipitation, and nitrogen deposition for five years. Root and shoot production did not respond to elevated CO2 or modest warming. Supplemental precipitation led to increases in shoot production and offsetting decreases in root production. Supplemental nitrate deposition increased total production by an average of 26%, primarily by stimulating shoot growth. Interactions among the main treatments were rare. Together, these results suggest that production in this grassland will respond minimally to changes in CO2 and winter precipitation, and to small amounts of warming. Increased nitrate deposition would have stronger effects on the grassland. Aside from this nitrate response, expectations that a changing atmosphere and climate would promote carbon storage by increasing plant growth appear unlikely to be realized in this system.
DOI:10.1126/science.289.5487.2068URL [本文引用: 1]
.
DOI:10.1016/S0378-5173(00)00669-4URLPMID:11250095 [本文引用: 1]
The skin permeation of estradiol from various proniosome gel formulations across excised rat skin was investigated in vitro. The encapsulation efficiency and size of niosomal vesicles formed from proniosomes upon hydration were also characterized. The encapsulation (%) of proniosomes with Span surfactants showed a very high value of 100%. Proniosomes with Span 40 and Span 60 increased the permeation of estradiol across skin. Both penetration enhancer effect of non-ionic surfactant and vesicle-skin interaction may contribute to the mechanisms for proniosomes to enhance estradiol permeation. Niosome suspension (diluted proniosomal formulations) and proniosome gel showed different behavior in modulating transdermal delivery of estradiol across skin. Presence or absence of cholesterol in the lipid bilayers of vesicles did not reveal difference in encapsulation and permeation of the associated estradiol. The types and contents of non-ionic surfactant in proniosomes are important factors affecting the efficiency of transdermal estradiol delivery.
DOI:10.1371/journal.pone.0047369URLPMID:3470592 [本文引用: 1]
Although community structure and species richness are known to respond to nitrogen fertilization dramatically, little is known about the mechanisms underlying specific species replacement and richness loss. In an experiment in semiarid temperate steppe of China, manipulative N addition with five treatments was conducted to evaluate the effect of N addition on the community structure and species richness. Species richness and biomass of community in each plot were investigated in a randomly selected quadrat. Root element, available and total phosphorus (AP, TP) in rhizospheric soil, and soil moisture, pH, AP, TP and inorganic N in the soil were measured. The relationship between species richness and the measured factors was analyzed using bivariate correlations and stepwise multiple linear regressions. The two dominant species, a shrubArtemisia frigidaand a grassStipa krylovii, responded differently to N addition such that the former was gradually replaced by the latter.S. kryloviiandA.frigidahad highly-branched fibrous and un-branched tap root systems, respectively.S. kryloviihad higher height thanA.frigidain both control and N added plots. These differences may contribute to the observed species replacement. In addition, the analysis on root element and AP contents in rhizospheric soil suggests that different calcium acquisition strategies, and phosphorus and sodium responses of the two species may account for the replacement. Species richness was significantly reduced along the five N addition levels. Our results revealed a significant relationship between species richness and soil pH, litter amount, soil moisture, AP concentration and inorganic N concentration. Our results indicate that litter accumulation and soil acidification accounted for 52.3% and 43.3% of the variation in species richness, respectively. These findings would advance our knowledge on the changes in species richness in semiarid temperate steppe of northern China under N deposition scenario.
DOI:10.1007/s100210000028URL [本文引用: 1]
Global climate change is predicted to alter growing season rainfall patterns, potentially reducing total amounts of growing season precipitation and redistributing rainfall into fewer but larger individual events. Such changes may affect numerous soil, plant, and ecosystem properties in grasslands and ultimately impact their productivity and biological diversity. Rainout shelters are useful tools for experimental manipulations of rainfall patterns, and permanent fixed-location shelters were established in 1997 to conduct the Rainfall Manipulation Plot study in a mesic tallgrass prairie ecosystem in northeastern Kansas. Twelve 9 14-m fixed-location rainfall manipulation shelters were constructed to impose factorial combinations of 30% reduced rainfall quantity and 50% greater interrainfall dry periods on 6 6-m plots, to examine how altered rainfall regimes may affect plant species composition, nutrient cycling, and above- and belowground plant growth dynamics. The shelters provided complete control of growing season rainfall patterns, whereas effects on photosynthetic photon flux density, nighttime net radiation, and soil temperature generally were comparable to other similar shelter designs. Soil and plant responses to the first growing season of rainfall manipulations (1998) suggested that the interval between rainfall events may be a primary driver in grassland ecosystem responses to altered rainfall patterns. Aboveground net primary productivity, soil CO2flux, and flowering duration were reduced by the increased inter-rainfall intervals and were mostly unaffected by reduced rainfall quantity. The timing of rainfall events and resulting temporal patterns of soil moisture relative to critical times for microbial activity, biomass accumulation, plant life histories, and other ecological properties may regulate longer-term responses to altered rainfall patterns.
DOI:10.1038/nplants.2015.80URLPMID:27250253 [本文引用: 2]
Terrestrial ecosystem productivity is widely accepted to be nutrient limited(1). Although nitrogen (N) is deemed a key determinant of aboveground net primary production (ANPP) 2,3, the prevalence of co-limitation by N and phosphorus (P) is increasingly recognized(4-8). However, the extent to which terrestrial productivity is co-limited by nutrients other than N and P has remained unclear. Here, we report results from a standardized factorial nutrient addition experiment, in which we added N, P and potassium (K) combined with a selection of micronutrients (K+mu), alone or in concert, to 42 grassland sites spanning five continents, and monitored ANPP. Nutrient availability limited productivity at 31 of the 42 grassland sites. And pairwise combinations of N, P, and K+mu co-limited ANPP at 29 of the sites. Nitrogen limitation peaked in cool, high latitude sites. Our findings highlight the importance of less studied nutrients, such as K and micronutrients, for grassland productivity, and point to significant variations in the type and degree of nutrient limitation. We suggest that multiple-nutrient constraints must be considered when assessing the ecosystem-scale consequences of nutrient enrichment.
DOI:10.1007/s00442-010-1890-zURLPMID:21191799 [本文引用: 1]
Productivity of semiarid grasslands is affected by soil water and nutrient availability, with water controlling net primary production under dry conditions and soil nutrients constraining biomass production under wet conditions. In order to investigate limitations on plants by the response of root-shoot biomass allocation to water and nitrogen (N) availability, a field experiment, on restoration plots with rainfed, unfertilized control plots, fertilized plots receiving N (25 kg urea-N ha ) and water (irrigation simulating a wet season), was conducted at two sites with different grazing histories: moderate (MG) and heavy (HG) grazing. Irrigation and N addition had no effect on belowground biomass. Irrigation increased aboveground (ANPP) and belowground net primary production (BNPP) and rain-use efficiency based on ANPP (RUE ANPP ), whereas N addition on rainfed plots had no effect on any of the measured parameters. N fertilizer application on irrigated plots increased ANPP and RUE ANPP and reduced the root fraction (RF: root dry matter/total dry matter), resulting in smaller N effects on total net primary production (NPP) and rain-use efficiency based on NPP. This suggests that BNPP should be included in evaluating ecosystem responses to resource availability from the wholeplant perspective. N effects on all measured parameters were similar on both sites. However, site HG responded to irrigation with higher ANPP and a lower RF when compared to site MG, indicating that species composition had a pronounced effect on carbon allocation pattern due to below-and aboveground niche complementarity.
DOI:10.1046/j.1365-2745.2000.00426.xURL [本文引用: 1]
1 Species diversity in the Arctic varies dramatically across abiotic gradients, including topography, moisture, pH and nutrient availability. We hypothesized that vascular plant species density, richness and diversity in Alaskan tundra would be correlated with: (i) site age, (ii) soil pH, (iii) above-ground productivity and biomass, and (iv) spatial heterogeneity. We sampled moist tussock, snowbed and watertrack communities in three sites that differed in substrate age (11 000-4.5 million years since deglaciation) for a variety of environmental and vegetation data over one growing season. 2 Productivity, biomass and heterogeneity were not consistently correlated with species density. However, variation in canopy height was correlated with species density and richness in a unimodal fashion, suggesting that heterogeneity of the light regime may be important for maintaining higher species numbers. 3 The 11 000-year-old site supported more vascular plant species than the two older sites, primarily due to greater numbers of forb species on the youngest site. 4 Soil pH was significantly positively correlated with species richness (R2= 0.82) and species density (R2= 0.61). In general the species found on acidic substrates (pH < 5.5) also occurred on non-acidic substrates (pH > 5.5). 5 This pattern of higher richness with higher pH occurs across other tundra types throughout the Arctic, suggesting that soil pH is an important filter of the regional species pool within northern regions, although other factors may become more important at local scales.
DOI:10.1111/j.1365-2486.2010.02274.xURL [本文引用: 1]
We combined year-round eddy covariance with biometry and biomass harvests along a chronosequence of boreal forest stands that were 1, 6, 15, 23, 40, 74, and 154 years old to understand how ecosystem production and carbon stocks change during recovery from stand-replacing crown fire. Live biomass (Clive) was low in the 1- and 6-year-old stands, and increased following a logistic pattern to high levels in the 74- and 154-year-old stands. Carbon stocks in the forest floor (Cforest floor) and coarse woody debris (CCWD) were comparatively high in the 1-year-old stand, reduced in the 6- through 40-year-old stands, and highest in the 74- and 154-year-old stands. Total net primary production (TNPP) was reduced in the 1- and 6-year-old stands, highest in the 23- through 74-year-old stands and somewhat reduced in the 154-year-old stand. The NPP decline at the 154-year-old stand was related to increased autotrophic respiration rather than decreased gross primary production (GPP). Net ecosystem production (NEP), calculated by integrated eddy covariance, indicated the 1- and 6-year-old stands were losing carbon, the 15-year-old stand was gaining a small amount of carbon, the 23- and 74-year-old stands were gaining considerable carbon, and the 40- and 154-year-old stands were gaining modest amounts of carbon. The recovery from fire was rapid; a linear fit through the NEP observations at the 6- and 15-year-old stands indicated the transition from carbon source to sink occurred within 11 12 years. The NEP decline at the 154-year-old stand appears related to increased losses from Clive by tree mortality and possibly from Cforest floor by decomposition. Our findings support the idea that NPP, carbon production efficiency (NPP/GPP), NEP, and carbon storage efficiency (NEP/TNPP) all decrease in old boreal stands.
DOI:10.1016/j.envexpbot.2005.11.002URL [本文引用: 1]
Theoretical plant growth models postulate that the relative rates of shoot and root growth are largely modulated by signals related to carbon and nitrogen status of the plant. To test this experimentally, 6-week-old vegetative cuttings of grapevine ( Vitis vinifera L. cv Merlot) were grown aeroponically in different controlled conditions of irradiance (13.8, 8.4 and 5.3 mol PAR m 612 day 611) and/or nitrogen nutrition (0.15, 1.20 and 7.11 mM N). Total non-structural carbohydrates (TNC) and amino acids (FAA) in leaves and roots were analysed 0, 6 and 28 days after treatment initiation. Both whole-plant biomass accumulation as well as C and N contents were highly responsive to light and N availability. At day 28, plant dry weight was significantly reduced in shaded vines (6135% of that of the control plants) and stimulated under the high irradiance environment (+30%). Deprivation of N enhanced root growth (+51%) at the expense of above-ground growth, whereas leaf dry weight was significantly greater in the high-N treatment than in the control. Vines grown under low-N and high irradiance conditions had the highest root-to-shoot ratios and those grown under low light and high N the lowest. Finally, redistribution of biomass among vegetative vine parts was significantly related to different indicators of the vine C:N status measured either at the whole-plant (N concentration) or at the organ level (TNC:FAA ratio), suggesting that root-to-shoot biomass partitioning was controlled by some aspect of plant C:N balance. Such relationships will be useful to improve allocation rules in a process-based growth model of grapevine.
DOI:10.11829/j.issn.1001-0629.2015-0485URL [本文引用: 1]
为研究氮沉降对黄土高原典型草原植物代谢功能的影响,本研究通过模拟大气氮沉降的方法对黄土高原天然草地进行了氮添加试验,分析了天然草地植物长芒草(Stipa bungeana)在不同施氮水平下叶片氮含量、光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、水分利用效率(WUE)以及叶绿素含量的响应特征,探讨了长芒草光合生理特性和叶片衰老的影响对氮添加的响应.结果显示,氮素添加可显著增加长芒草地上生物量(P<0.05),明显增加其所占总生物量的比例,显著增加叶片氮含量、叶绿素含量和净光合速率,而对气孔导度和蒸腾速率没有显著影响(P>0.05),显著提高了植物水分利用效率.另外,氮素添加延缓了长芒草叶绿素的降解速率.以上结果表明,一定范围内的大气氮沉降量有利于长芒草光合能力的提高,可以延缓长芒草叶片的衰老.
DOI:10.11829/j.issn.1001-0629.2015-0485URL [本文引用: 1]
为研究氮沉降对黄土高原典型草原植物代谢功能的影响,本研究通过模拟大气氮沉降的方法对黄土高原天然草地进行了氮添加试验,分析了天然草地植物长芒草(Stipa bungeana)在不同施氮水平下叶片氮含量、光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、水分利用效率(WUE)以及叶绿素含量的响应特征,探讨了长芒草光合生理特性和叶片衰老的影响对氮添加的响应.结果显示,氮素添加可显著增加长芒草地上生物量(P<0.05),明显增加其所占总生物量的比例,显著增加叶片氮含量、叶绿素含量和净光合速率,而对气孔导度和蒸腾速率没有显著影响(P>0.05),显著提高了植物水分利用效率.另外,氮素添加延缓了长芒草叶绿素的降解速率.以上结果表明,一定范围内的大气氮沉降量有利于长芒草光合能力的提高,可以延缓长芒草叶片的衰老.
DOI:10.11733/j.issn.1007-0435.2015.01.011URL [本文引用: 1]
Our objective was to examine the effect of nitrogen(N) enrichment on the specific leaf area (SLA) and important value (IV) of several species in the typical steppe grasslands of the Loess Plateau. A long-term N addition experiment was conducted to investigate the effects of increasing N inputs on the SLA and IV of four dominant species (Stipa bungeana, Leymus secalinus, Heteropappus altaicus and Lespedeza davurica) in the Loess Plateau natural grassland. The results showed that N addition enhanced the SLAs of all species and IVs of Stipa bungeana and Leymus secalinus, but decreased the IVs of Heteropappus altaicus and Lespedeza davurica. IV was not correlated with SLA, however, there was a strong negative correlation between IV and the coefficient of variation. These results indicated that the SLAs of plants in arid ecosystems were conservative and low plasticity. N addition was more favorable to gramineous than to forbs.
DOI:10.11733/j.issn.1007-0435.2015.01.011URL [本文引用: 1]
Our objective was to examine the effect of nitrogen(N) enrichment on the specific leaf area (SLA) and important value (IV) of several species in the typical steppe grasslands of the Loess Plateau. A long-term N addition experiment was conducted to investigate the effects of increasing N inputs on the SLA and IV of four dominant species (Stipa bungeana, Leymus secalinus, Heteropappus altaicus and Lespedeza davurica) in the Loess Plateau natural grassland. The results showed that N addition enhanced the SLAs of all species and IVs of Stipa bungeana and Leymus secalinus, but decreased the IVs of Heteropappus altaicus and Lespedeza davurica. IV was not correlated with SLA, however, there was a strong negative correlation between IV and the coefficient of variation. These results indicated that the SLAs of plants in arid ecosystems were conservative and low plasticity. N addition was more favorable to gramineous than to forbs.
DOI:10.1111/j.1365-2486.2007.01447.xURL [本文引用: 1]
The world's ecosystems are experiencing simultaneous changes in the supply of multiple limiting resources. Two of these, water and nitrogen (N) can strongly limit grassland production and can affect community composition and biogeochemical cycles in different ways. Grassland ecosystems in California may be particularly vulnerable to current and predicted changes in precipitation and N deposition, and ecosystem responses to potential interactive effects of water and N are not well understood. Here, we show strong colimitation of plant production resulting from factorial addition of water and N. In addition, water and N addition in combination led to increased dominance of the two most abundant grass species, while N addition regardless of water availability led to decreased species diversity. Late season carbon (C) flux response to water addition depended on N. Only plots that received additional water, but not N, still showed net ecosystem C uptake at the end of the experiment. Our results suggest that grassland ecosystem response to N deposition will be strongly dependent on future precipitation patterns.
DOI:10.1139/b87-105URL [本文引用: 1]
Abstract Root dynamics were studied using root windows at Konza Prairie, a tallgrass prairie in north central Kansas, during dry (1984) and wet (1985) years. Amounts, production, and disappearance of root length decreased during drought but increased when rains resumed; however, standing crop remained low. The 1985 root lengths increased throughout the growing season, while production and disappearance remained constant. Yearly summaries of amounts, productivity, and decomposition by 10-cm increments in soil depth show that the effect of drought on these variables decreased with increasing soil depth. Turnover rates of root length averaged 564 in the dry year versus 389% in the wet year, with the largest difference noted in the 0- to 10-cm depth (800 in 1984 versus 540% in 1985). Production and decay patterns observed using root windows were also noted in root biomass data (obtained from soil cores). The average total root biomass turnover rate was 31%. Failure to sort below-ground materials into tissue types (rhizomes, roots) and live versus dead status results in reduced estimates of biomass turnover rates. The greatest possible separation of plant components presents the most comprehensive picture of (belowground) growth dynamics.
DOI:10.1111/nph.12952URL [本文引用: 1]
DOI:10.1890/0012-9658(1999)080[1150:TMAORR]2.0.CO;2URL [本文引用: 2]
DOI:10.1111/j.1461-0248.2004.00686.xURL [本文引用: 1]
Abstract Human impacts on the natural environment have reached such proportions that in addition to an ‘extinction crisis’, we now also face a broader ‘biome crisis’. Here we identify the world's terrestrial biomes and, at a finer spatial scale, ecoregions in which biodiversity and ecological function are at greatest risk because of extensive habitat conversion and limited habitat protection. Habitat conversion exceeds habitat protection by a ratio of 8:1 in temperate grasslands and Mediterranean biomes, and 10:1 in more than 140 ecoregions. These regions include some of the most biologically distinctive, species rich ecosystems on Earth, as well as the last home of many threatened and endangered species. Confronting the biome crisis requires a concerted and comprehensive response aimed at protecting not only species, but the variety of landscapes, ecological interactions, and evolutionary pressures that sustain biodiversity, generate ecosystem services, and evolve new species in the future.
[本文引用: 1]
DOI:10.11829\j.issn.1001-0629.2013-0640URL [本文引用: 1]
Global warming has brought great impacts on the ecological system including extending plants growth period, increasing biomass, increasing photosynthetic rate of the plants with lower optimum temperature without changing stomatal conductance, decreasing the content of sodium, potassium and phosphorus in the leaves. With the temperature increasing, the biomass variations of different types of grassland were inconsistent. Kobresia humilis in alpine meadow would be substitute by other grass and Eymus chinensis would be substitute by Stipa grandis. The insects and rodents were mainly studied with warming which showed that insects increased generations and had a tendency to spread to the high altitude and high latitudes and the rodent increased population density without changing of sex ratio and population age structure. Warming enhanced soil respiration, the activities of soil enzyme and the content of soil bacteria. The present paper summarized the response of each component in grassland ecological system which need further study the influence the grassland ecosystem and biosphere.
DOI:10.11829\j.issn.1001-0629.2013-0640URL [本文引用: 1]
Global warming has brought great impacts on the ecological system including extending plants growth period, increasing biomass, increasing photosynthetic rate of the plants with lower optimum temperature without changing stomatal conductance, decreasing the content of sodium, potassium and phosphorus in the leaves. With the temperature increasing, the biomass variations of different types of grassland were inconsistent. Kobresia humilis in alpine meadow would be substitute by other grass and Eymus chinensis would be substitute by Stipa grandis. The insects and rodents were mainly studied with warming which showed that insects increased generations and had a tendency to spread to the high altitude and high latitudes and the rodent increased population density without changing of sex ratio and population age structure. Warming enhanced soil respiration, the activities of soil enzyme and the content of soil bacteria. The present paper summarized the response of each component in grassland ecological system which need further study the influence the grassland ecosystem and biosphere.
[本文引用: 2]
DOI:10.1371/journal.pone.0069561URLPMID:23936045 [本文引用: 1]
Understanding and predicting ecosystem functioning such as biomass accumulation requires an accurate assessment of large-scale patterns of biomass distribution and partitioning in relation to climatic and soil environments. We sampled above- and belowground biomass from 26 sites spanning 1500 km in Inner Mongolian grasslands, compared the difference in aboveground, belowground biomass and below-aboveground biomass ratio (AGB, BGB, and B/A, respectively) among meadow steppe, typical steppe, and desert steppe types. The relationships between AGB, BGB, B/A and climatic and soil environments were then examined. We found that AGB and BGB differed significantly among three types of grasslands while B/A did not differ. Structural equation model analyses indicated that mean annual precipitation was the strongest positive driver for AGB and BGB. AGB was also positively associated with soil organic carbon, whereas B/A was positively associated with total soil nitrogen. These results indicated that precipitation positively influence plant production in Inner Mongolian grasslands. Contrary to the prediction from the optimal partitioning hypothesis, biomass allocation to belowground increased with soil total nitrogen, suggesting that more productive sites may increase belowground allocation as an adaptive strategy to potentially high fire frequencies.
DOI:10.1073/pnas.0802891105URL
DOI:10.1007/s100210000057URL [本文引用: 1]
The frequency and extent of water limitation to aboveground net primary production (ANPP) in a mesic grassland in NE Kansas (Konza Prairie, USA) was assessed with an 8-year irrigation experiment. Since 1991, transects spanning upland and lowland sites in annually burned, ungrazed tallgrass prairie were provided with supplemental water to satisfy evapotranspirational demands. This protocol minimized water limitations during the growing season, as well as interannual variability in water stress. Irrigation of this mesic grassland increased ANPP in 6 of 8 years by an average of 26% when compared to control transects. Although interannual variation in ANPP was greater in uplands than lowlands at nominal levels of precipitation, reducing interannual variability in water availability via irrigation eliminated topographic differences; the irrigation protocol also reduced interannual variability in ANPP by as much as 40%. The addition of supplemental water enabled us to extend the relationship between annual precipitation and ANPP in grasslands to precipitation levels (average, 1153 mm; maximum, 1346 mm) similar to those experienced by more mesic grasslands that today exist only as remnants several hundred kilometers east of Kansas. This relationship was linear$(r^{2}=0.81)$, with maximum ANPP$(738\ {\rm g}/{\rm m}^{2})$similar to values reported for sites in Illinois and Wisconsin. After 8 years of irrigation, production of the C3forb component was twice that in control sites. These results indicate that water limitations in grasslands at the western edge of the presettlement extent of tall-grass prairie affect ANPP in most years and that this high frequency of water limitation may lead to greater dominance of the C4grasses than is seen in more eastern grassland sites.
[本文引用: 2]
DOI:10.2307/1941874URLPMID:27759270 [本文引用: 1]
We evaluated the relationship between annual forage production and annual and seasonal precipitation and temperature at a shortgrass steppe site in north-central Colorado using a long-term data set (52 yr). We also constructed a relationship between forage production and aboveground net primary production (ANPP). Precipitation fluctuated randomly, but temperature had clear warming and cooling trends including a 17-yr warming trend from 1974 to 1990. Forage production was significantly related to both annual and seasonal precipitation but not temperature. Precipitation events between 15 and 30 mm accounted for most of the variability in production because they accounted for most of the variability in precipitation and because they wetted the soil layers that have the largest effect on production. Forage production amplified variability in annual precipitation. Production showed time lags of several years in responding to increases in precipitation. Change in vegetation structure has a characteristic response time, which contrains production responses in wet years. Constraint caused by vegetation structure is the reason why regional ANPP-precipitation models have a steeper slope than long-term models and point out a weakness of exchanging space for time in predicting production patterns.
DOI:10.1890/06-2057.1URL [本文引用: 1]
DOI:10.1098/rstb.2010.0028URLPMID:2880131 [本文引用: 5]
Grassland ecosystems cover vast areas of the Earth's surface and provide many ecosystem services including carbon (C) storage, biodiversity preservation and the production of livestock forage. Predicting the future delivery of these services is difficult, because widespread changes in atmospheric CO60 concentration, climate and nitrogen (N) inputs are expected. We compiled published data from global change driver manipulation experiments and combined these with climate data to assess grassland biomass responses to CO60 and N enrichment across a range of climates. CO60 and N enrichment generally increased aboveground biomass (AGB) but effects of CO60 enrichment were weaker than those of N. The response to N was also dependent on the amount of N added and rainfall, with a greater response in high precipitation regions. No relationship between response to CO60 and climate was detected within our dataset, thus suggesting that other site characteristics, e.g. soils and plant community composition, are more important regulators of grassland responses to CO60. A statistical model of AGB response to N was used in conjunction with projected N deposition data to estimate changes to future biomass stocks. This highlighted several potential hotspots (e.g. in some regions of China and India) of grassland AGB gain. Possible benefits for C sequestration and forage production in these regions may be offset by declines in plant biodiversity caused by these biomass gains, thus necessitating careful management if ecosystem service delivery is to be maximized. An approach such as ours, in which meta-analysis is combined with global scale model outputs to make large-scale predictions, may complement the results of dynamic global vegetation models, thus allowing us to form better predictions of biosphere responses to environmental change.
DOI:10.1007/s11104-010-0612-yURL [本文引用: 1]
While water availability determines grassland productivity in semiarid regions, nutrient availability is the main limiting factor under wet conditions. An experiment was conducted in 2008 at two sites in Inner Mongolia with histories of heavy grazing (HG) and moderate grazing (MG) to study the interactive effects of water and nitrogen on above- and belowground net primary productivity (ANPP and BNPP), biomass partitioning, and plant species composition. The study comprises two water treatments (no irrigation and irrigated when soil water content was below 70% of the field capacity), and two nitrogen (N) levels (0 and 10002kg02N02ha 611 ). Mean values of ANPP at the peak biomass time reached 1,02865±6595 SD g02m 612 at the HG site and 56865±6532 SD g02m 612 at the MG site in irrigated and fertilized treatment. Nitrogen use efficiency (NUE) was significantly higher at irrigated plots compared to rain-fed plots at both HG and MG sites. Water use efficiency (WUEt) based on total water input and ANPP decreased with irrigation at the HG site. Meanwhile, N application significantly increased WUEt, WUEp (based on precipitation), and WUEi (based on irrigation water) at both sites. BNPP was significantly higher at irrigated plots compared to rain-fed plots at both HG and MG sites, and it tended to decrease with N addition. However, the fraction of belowground to total biomass ( f BNPP = BNPP/(ANPP+BNPP) decreased with the addition of supplemental resources and exhibited a negative correlation with ANPP. Species diversity remained lower at the HG site compared to the MG site; it decreased with the addition of supplemental resources at the latter site. The annual Salsola collina contributed the most to the total biomass under irrigation. Based on global climate models, more frequent extreme climates are predicted in the future, which can result in changes in resource availabilities. Therefore, our research results have important implications for predicting the production and other properties of grassland ecosystems.
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/gcb.13156URLPMID:26554753 [本文引用: 1]
Abstract Soil respiration ( R s) is the second-largest terrestrial carbon (C) flux. Although R s has been extensively studied across a broad range of biomes, there is surprisingly little consensus on how the spatiotemporal patterns of R s will be altered in a warming climate with changing precipitation regimes. Here, we present a global synthesis R s data from studies that have manipulated precipitation in the field by collating studies from 113 increased precipitation treatments, 91 decreased precipitation treatments, and 14 prolonged drought treatments. Our meta-analysis indicated that when the increased precipitation treatments were normalized to 28% above the ambient level, the soil moisture, R s, and the temperature sensitivity ( Q 10) values increased by an average of 17%, 16%, and 6%, respectively, and the soil temperature decreased by 611.3%. The greatest increases in R s and Q 10 were observed in arid areas, and the stimulation rates decreased with increases in climate humidity. When the decreased precipitation treatments were normalized to 28% below the ambient level, the soil moisture and R s values decreased by an average of 6114% and 6117%, respectively, and the soil temperature and Q 10 values were not altered. The reductions in soil moisture tended to be greater in more humid areas. Prolonged drought without alterations in the amount of precipitation reduced the soil moisture and R s by 6112% and 616%, respectively, but did not alter Q 10. Overall, our synthesis suggests that soil moisture and R s tend to be more sensitive to increased precipitation in more arid areas and more responsive to decreased precipitation in more humid areas. The responses of R s and Q 10 were predominantly driven by precipitation-induced changes in the soil moisture, whereas changes in the soil temperature had limited impacts. Finally, our synthesis of prolonged drought experiments also emphasizes the importance of the timing and frequency of precipitation events on ecosystem C cycles. Given these findings, we urge future studies to focus on manipulating the frequency, intensity, and seasonality of precipitation with an aim to improving our ability to predict and model feedback between R s and climate change.
DOI:10.1038/nature11917URLPMID:23426264 [本文引用: 1]
China is experiencing intense air pollution caused in large part by anthropogenic emissions of reactive nitrogen(1,2). These emissions result in the deposition of atmospheric nitrogen (N) in terrestrial and aquatic ecosystems, with implications for human and ecosystem health, greenhouse gas balances and biological diversity(1,3-5). However, information on the magnitude and environmental impact of N deposition in China is limited. Here we use nationwide data sets on bulk N deposition, plant foliar N and crop N uptake (from long-term unfertilized soils) to evaluate N deposition dynamics and their effect on ecosystems across China between 1980 and 2010. We find that the average annual bulk deposition of N increased by approximately 8 kilograms of nitrogen per hectare (P < 0.001) between the 1980s (13.2 kilograms of nitrogen per hectare) and the 2000s (21.1 kilograms of nitrogen Per hectare). Nitrogen deposition rates in the industrialized and agriculturally intensified regions of China are as high as the peak levels of deposition in northwestern Europe in the 1980s(6), before the introduction of mitigation measures(7,8). Nitrogen from ammonium (NH4+) is the dominant form of N in bulk deposition, but the rate of increase is largest for deposition of N from nitrate (NO3-), in agreement with decreased ratios of NH3 to NOx emissions since 1980. We also find that the impact of N deposition on Chinese ecosystems includes significantly increased plant foliar N concentrations in natural and semi-natural (that is, non-agricultural) ecosystems and increased crop N uptake from long-term-unfertilized crop-lands. China and other economies are facing a continuing challenge to reduce emissions of reactive nitrogen, N deposition and their negative effects on human health and the environment.
DOI:10.1890/12-0279.1URLPMID:23687898 [本文引用: 1]
Global warming potentially alters the terrestrial carbon (C) cycle, likely feeding back to further climate warming. However, how the ecosystem C cycle responds and feeds back to warming remains unclear. Here we used a meta-analysis approach to quantify the response ratios of 18 variables of the ecosystem C cycle to experimental warming and evaluated ecosystem C-cycle feedback to climate warming. Our results showed that warming stimulated gross ecosystem photosynthesis (GEP) by 15.7%, net primary production (NPP) by 4.4%, and plant C pools from above- and belowground parts by 6.8% and 7.0%, respectively. Experimental warming accelerated litter mass loss by 6.8%, soil respiration by 9.0%, and dissolved organic C leaching by 12.1%. In addition, the responses of some of those variables to experimental warming differed among the ecosystem types. Our results demonstrated that the stimulation of plant-derived C influx basically offset the increase in warming-induced efflux and resulted in insignificant changes in litter and soil C content, indicating that climate warming may not trigger strong positive C-climate feedback from terrestrial ecosystems. Moreover, the increase in plant C storage together with the slight but not statistically significant decrease of net ecosystem exchange (NEE) across ecosystems suggests that terrestrial ecosystems might be a weak C sink rather than a C source under global climate warming. Our results are also potentially useful for parameterizing and benchmarking land surface models in terms of C cycle responses to climate warming.
[本文引用: 1]
[本文引用: 1]
DOI:10.1038/nature02887URLPMID:15386009 [本文引用: 1]
Global warming is predicted to be most pronounced at high latitudes, and observational evidence over the past 25 years suggests that this warming is already under way. One-third of the global soil carbon pool is stored in northern latitudes, so there is considerable interest in understanding how the carbon balance of northern ecosystems will respond to climate warming. Observations of controls over plant productivity in tundra and boreal ecosystems have been used to build a conceptual model of response to warming, where warmer soils and increased decomposition of plant litter increase nutrient availability, which, in turn, stimulates plant production and increases ecosystem carbon storage. Here we present the results of a long-term fertilization experiment in Alaskan tundra, in which increased nutrient availability caused a net ecosystem loss of almost 2,000 grams of carbon per square meter over 20 years. We found that annual aboveground plant production doubled during the experiment. Losses of carbon and nitrogen from deep soil layers, however, were substantial and more than offset the increased carbon and nitrogen storage in plant biomass and litter. Our study suggests that projected release of soil nutrients associated with high-latitude warming may further amplify carbon release from soils, causing a net loss of ecosystem carbon and a positive feedback to climate warming.
DOI:10.3724/SP.J.1258.2014.00011URL
以氮素和水分(冬季增雪和夏季增雨)为控制因子,开展相关田间控制实验,分析不同功能群(以生活史为划分依据)尺度和群落尺度植物生物量分配格局对氮素和水分的响应,得出以下结论:1)一年生植物的繁殖生物量比重明显高于多年生植物,而多年生植物种的叶/地上生物量比值显著高于一年生植物;2)一年生植物对氮素和水分添加的响应剧烈,氮添加耦合夏季增雨、氮添加耦合冬季增雪显著增加了一年生植物的繁殖生物量比重和叶生物量比重。多年生植物对氮素和水分添加的响应不敏感,表现为多年生植物的各器官生物量分配格局对氮素添加和水分添加的响应不明显。3)氮素添加和水分处理改变了群落尺度生物量分配格局:氮素添加耦合冬季增雪处理降低了群落植物的繁殖生物量比重和茎生物量比重,提高了群落植物的叶生物量比重。4)冬季增雪和夏季增雨与氮素添加的交互作用对群落生物量分配格局的改变不同。夏季增雨耦合氮素添加处理下群落的茎生物量比重显著提高,群落茎生物量分配的改变引起群落的垂直结构发生改变。冬季增雪氮素处理下群落的叶生物量比重增加,但茎生物量比重增加不明显。冬季增雪也改变了群落的结构和功能。
DOI:10.3724/SP.J.1258.2014.00011URL
以氮素和水分(冬季增雪和夏季增雨)为控制因子,开展相关田间控制实验,分析不同功能群(以生活史为划分依据)尺度和群落尺度植物生物量分配格局对氮素和水分的响应,得出以下结论:1)一年生植物的繁殖生物量比重明显高于多年生植物,而多年生植物种的叶/地上生物量比值显著高于一年生植物;2)一年生植物对氮素和水分添加的响应剧烈,氮添加耦合夏季增雨、氮添加耦合冬季增雪显著增加了一年生植物的繁殖生物量比重和叶生物量比重。多年生植物对氮素和水分添加的响应不敏感,表现为多年生植物的各器官生物量分配格局对氮素添加和水分添加的响应不明显。3)氮素添加和水分处理改变了群落尺度生物量分配格局:氮素添加耦合冬季增雪处理降低了群落植物的繁殖生物量比重和茎生物量比重,提高了群落植物的叶生物量比重。4)冬季增雪和夏季增雨与氮素添加的交互作用对群落生物量分配格局的改变不同。夏季增雨耦合氮素添加处理下群落的茎生物量比重显著提高,群落茎生物量分配的改变引起群落的垂直结构发生改变。冬季增雪氮素处理下群落的叶生物量比重增加,但茎生物量比重增加不明显。冬季增雪也改变了群落的结构和功能。
DOI:10.1038/nature01136URLPMID:12410307 [本文引用: 1]
Abstract Soils contain the largest near-surface reservoir of terrestrial carbon and so knowledge of the factors controlling soil carbon storage and turnover is essential for understanding the changing global carbon cycle. The influence of climate on decomposition of soil carbon has been well documented, but there remains considerable uncertainty in the potential response of soil carbon dynamics to the rapid global increase in reactive nitrogen (coming largely from agricultural fertilizers and fossil fuel combustion). Here, using 14C, 13C and compound-specific analyses of soil carbon from long-term nitrogen fertilization plots, we show that nitrogen additions significantly accelerate decomposition of light soil carbon fractions (with decadal turnover times) while further stabilizing soil carbon compounds in heavier, mineral-associated fractions (with multidecadal to century lifetimes). Despite these changes in the dynamics of different soil pools, we observed no significant changes in bulk soil carbon, highlighting a limitation inherent to the still widely used single-pool approach to investigating soil carbon responses to changing environmental conditions. It remains to be seen if the effects observed here-caused by relatively high, short-term fertilizer additions-are similar to those arising from lower, long-term additions of nitrogen to natural ecosystems from atmospheric deposition, but our results suggest nonetheless that current models of terrestrial carbon cycling do not contain the mechanisms needed to capture the complex relationship between nitrogen availability and soil carbon storage.
DOI:10.3354/cr021001URL [本文引用: 2]
DOI:10.1007/s10021-009-9265-1URL [本文引用: 1]
Changes in precipitation and nitrogen (N) deposition can influence ecosystem carbon (C) cycling and budget in terrestrial biomes, with consequent feedbacks to climate change. However, little is known about the main and interactive effects of water and N additions on net ecosystem C exchange (NEE). In a temperate steppe of northern China, a field-manipulated experiment was conducted to evaluate the responses of NEE and its components to improve N and water availability from 2005 to 2008. The results showed that both water and N additions stimulated gross ecosystem productivity (GEP), ecosystem respiration (ER), and NEE. Water addition increased GEP by 17%, ER by 24%, and NEE by 11% during the experimental period, whereas N addition increased GEP by 17%, ER by 16%, and NEE by 19%. The main effects of both water and N additions changed with time, with the strongest water stimulation in the dry year and a diminishing N stimulation over time. When water and N were added in combination, there were nonadditive effects of water and N on ecosystem C fluxes, which could be explained by the changes in species composition and the shifts of limiting resources from belowground (water or N) to aboveground (light). The positive water and N additions effects indicate that increasing precipitation and N deposition in the future will favor C sequestration in the temperate steppe. The non-additive effects of water and N on ecosystem C fluxes suggest that multifactor experiments are better able to capture complex interactive processes, thus improving model simulations and projections.
DOI:10.1371/journal.pone.0020078URL [本文引用: 2]
DOI:10.1002/ecm.1262URL [本文引用: 1]
Abstract Given that plant growth is often water-limited in grasslands, it has been proposed that projected increases in precipitation could increase plant productivity and carbon sequestration. However, the existing evidence for this hypothesis comes primarily from observational studies along natural precipitation gradients or from short-term manipulative experiments. It remains unclear whether long-term increased precipitation persistently stimulates grassland productivity. In the world's largest remaining temperate grassland, we found that experimentally increased precipitation enhanced net primary production, soil available nitrogen and foliar nitrogen concentrations during the first six years, but it ceased to do so in the following four years, unless nitrogen was simultaneously added with water. The 15N enrichment of plant and soil nitrogen pools in later years indicates increased nitrogen losses, which exacerbated nitrogen limitation and ended the stimulation of productivity by increased precipitation. Changes in species abundance might have contributed little to the changes in water treatment effects. Our study demonstrates that the long-term response of grassland ecosystems to increased precipitation will be mediated by nitrogen availability. Our results also point to a shift from co-limitation by water and nitrogen early to perhaps limitation by nitrogen only later in this temperate grassland, highlighting significant variations in the type of resource limitation induced by climate change. This article is protected by copyright. All rights reserved.
In: Aroca R ed.
DOI:10.1007/978-3-642-32653-0_7URL [本文引用: 2]
Scarcity of water is a severe environmental limit to plant productivity. Drought-induced loss in crop yield probably exceeds losses from all other causes, since both the severity and duration of the stress are critical. Nutritional imbalance under drought conditions depresses plant growth and therefore productivity by affecting nutrient uptake, transport, and distribution. Despite contradictory reports on the effects of nutrient supply on plant growth under drought conditions, it is generally accepted that an increased nutrient supply will not improve plant growth when the nutrient is already present in sufficient amounts in the soil and the drought is severe. A better understanding of the role of mineral nutrients in plant resistance to drought will contribute to improve fertilizer management in arid and semiarid areas and in regions suffering from temporary drought. Considering that vegetables are concentrated in semiarid zones where water stress is frequent, it is important to ascertain how this type of stress affects the nutrient uptake and assimilation of these crops. This chapter starts with an overview of the recent literature on plant nutrition of vegetables under drought conditions, stressing mainly the effects of drought on nutrient availability, uptake, transport, and accumulation in plants, and also the interactions between nutrient supply and drought response; it then proceeds to identify the means to increase nutrient availability under drought conditions through breeding, grafting, and fertilization.
DOI:10.2307/1943158URL [本文引用: 1]
Aboveground net primary production of grasslands is strongly influenced by the amount and distribution of annual precipitation. Analysis of data collected at 9500 sites throughout the central United States confirmed the overwhelming importance of water availability as a control of production. The regional spatial pattern of production reflected the east-west gradient in annual precipitation. Lowest values of aboveground net primary production were observed in the west and highest values in the east. This spatial pattern was shifted eastward during unfavorable years and westward during favorable years. Variability in production among years was maximum in northern New Mexico and southwestern Kansas and decreased towards the north and south. The regional pattern of production was largely accounted for by annual precipitation. Production at the site level was explained by annual precipitation, soil water-holding capacity, and an interaction term. Our results support the inverse texture hypothesis. When precipitation is <370 mm/yr, sandy soils with low water-holding capacity are more productive than loamy soils with high water-holding capacity, while the opposite pattern occurs when precipitation is >370 mm/yr.
DOI:10.1111/j.1365-2435.2009.01663.xURL [本文引用: 1]
1. Atmospheric deposition of nitrogen (N) is a global problem resulting in negative consequences for biodiversity due to direct toxicity, increases in invasive species, increased susceptibility to environmental stresses and soil-mediated effects of acidification and eutrophication.2. Reductions in plant species richness related to N deposition have been observed in a number of habitats including calcifuge (acid) grasslands but the mechanisms of this decline have not been fully investigated. We test the hypotheses that along a large-scale gradient of N deposition there is (i) an increase in species tolerant of low pH conditions as a result of soil acidification and (ii) an increase in competitive and nitrophilic species as a result of soil eutrophication. As competitive species can occur in low pH habitats, both of these hypotheses could be true.3. Using plant characteristics, we examined changes in vegetation species composition along the gradient of N deposition in the UK. Mean C090009S090009R signatures were used to identify the competitive response of plant communities together with Ellenberg N (nitrogen) scores to identify increases in nitrophilic species. Ellenberg R (reaction, pH) scores were used to identify change in response to soil pH together with an index of soil acidity preference developed using regional survey data.4. Mean C090009S090009R signatures showed no significant correlation with N deposition, nor did mean Ellenberg N scores. Ellenberg R scores and the index of soil acidity preference showed significant relationships with N deposition indicating an increased dominance of acid-tolerant species.5. The results suggest that soil acidification as opposed to eutrophication and consequent competition between species is contributing to shifts in species composition and diversity linked to N deposition in calcifuge grasslands. Soil acidification may be leading to reduced nutrient availability preventing the effects of N addition from being apparent.
[本文引用: 1]
DOI:10.1073/pnas.0903186107URL [本文引用: 1]
Precipitation extreme changes are often assumed to scale with, or are constrained by, the change in atmospheric moisture content. Studies have generally confirmed the scaling based on moisture content for the midlatitudes but identified deviations for the tropics. In fact half of the twelve selected Intergovernmental Panel on Climate Change (IPCC) models exhibit increases faster than the climatological-mean precipitable water change for high percentiles of tropical daily precipitation, albeit with significant intermodel scatter. Decomposition of the precipitation extreme changes reveals that the variations among models can be attributed primarily to the differences in the upward velocity. Both the amplitude and vertical profile of vertical motion are found to affect precipitation extremes. A recently proposed scaling that incorporates these dynamical effects can capture the basic features of precipitation changes in both the tropics and midlatitudes. In particular, the increases in tropical precipitation extremes significantly exceed the precipitable water change in Model for Interdisciplinary Research on Climate (MIROC), a coupled general circulation model with the highest resolution among IPCC climate models whose precipitation characteristics have been shown to reasonably match those of observations. The expected intensification of tropical disturbances points to the possibility of precipitation extreme increases beyond the moisture content increase as is found in MIROC and some of IPCC models.
DOI:10.1890/07-1434.1URLPMID:18959323 [本文引用: 1]
The aboveground net primary productivity (ANPP) of grass communities in grasslands and savannas is primarily determined by precipitation quantity. Recent research, motivated by predictions of changes in the distribution of rainfall events by global climate change models, indicates that ANPP may be affected by rainfall distribution as much as by annual totals. Grazing and community composition are also known to affect grassland ANPP. The manner in which interactions between rainfall distribution, grazing, and community composition affect the relationship between precipitation and ANPP represents a critical knowledge gap. The effects of community composition and grazing on aboveground growth responses to intraseasonal variation in water availability were investigated at seven grassland sites with a nonselective clipping experiment. The aboveground growth of the dominant C species at each site was measured at regular intervals for 2 3 growing seasons in the presence or absence of regular defoliation. In the absence of defoliation, there was a general lack of synchrony of intraseasonal growth among co-occurring species. Variation in growth rates was high and was only partially explained by variation in rainfall. Regular defoliation increased growth synchrony at all sites, but changes in growth responses to rainfall varied between sites. These results suggest that community composition will be important in determining ANPP recipitation relationships under conditions of altered rainfall distribution. However this effect appears to be a result of species responding differently to soil water or other resources rather than to rainfall per se. Grazing may override the effects of community composition by reducing differences in growth patterns between species and has the potential to weaken precipitation controls on ANPP.
DOI:10.1111/j.1365-2486.2010.02363.xURL [本文引用: 1]
Abstract Precipitation regimes are predicted to become more variable with more extreme rainfall events punctuated by longer intervening dry periods. Water-limited ecosystems are likely to be highly responsive to altered precipitation regimes. The bucket model predicts that increased precipitation variability will reduce soil moisture stress and increase primary productivity and soil respiration in aridland ecosystems. To test this hypothesis, we experimentally altered the size and frequency of precipitation events during the summer monsoon (July through September) in 2007 and 2008 in a northern Chihuahuan Desert grassland in central New Mexico, USA. Treatments included (1) ambient rain, (2) ambient rain plus one 20 mm rain event each month, and (3) ambient rain plus four 5 mm rain events each month. Throughout two monsoon seasons, we measured soil temperature, soil moisture content ( ), soil respiration ( R s), along with leaf-level photosynthesis ( A net), predawn leaf water potential ( pd), and seasonal aboveground net primary productivity (ANPP) of the dominant C4 grass, Bouteloua eriopoda . Treatment plots receiving a single large rainfall event each month maintained significantly higher seasonal soil which corresponded with a significant increase in R s and ANPP of B. eriopoda when compared with plots receiving multiple small events. Because the strength of these patterns differed between years, we propose a modification of the bucket model in which both the mean and variance of soil water change as a consequence of interannual variability from 1 year to the next. Our results demonstrate that aridland ecosystems are highly sensitive to increased precipitation variability, and that more extreme precipitation events will likely have a positive impact on some aridland ecosystem processes important for the carbon cycle.
DOI:10.1086/284382URL [本文引用: 1]
The resource-ratio hypothesis assumes that each plant species is a superior competitor for a particular proportion of the limiting resources and predicts that community composition should change whenever the relative availability of two or more limiting resources changes. It is suggested that (1) the major limiting resources for mesic terrestrial habitats are often a soil resource, especially nitrogen, and light; (2) these resources are naturally inversely related, the habitats with poor soils having high-light availability and the habitats with rich soils having low-light availability; (3) the life history of a plant should depend on the point along the soil-resource: light gradient at which the plant is a superior competitor; and (4) primary succession and secondary succession on poor soils result from a temporal gradient in the relative availabilities of a limiting soil resource and light. If, as hypothesized, plants specialized on low-nutrient habitats are relatively short in height, short-lived, and fast growing, and if they reproduce early in life compared to plants dominant when light at the soil surface is limiting, these life history differences could explain the similarity of secondary succession on rich soils to primary succession and secondary succession on poor soils.
DOI:10.2307/2937208URL [本文引用: 1]
Five grass species (Agrostis scabra, Agropyron repens, Poa pratensis, Schizachyrium scoparium and Andropogon gerardi) were grown in monoculture for 3 yr on an experimental nitrogen gradient. The species differed significantly in the levels to which they reduced soil solution (0.01 mol/L KCI extractable) nitrate and ammonium concentrations and light penetration to the soil surface. Soil nitrate concentration was an inverse function of root mass, which explained 73% of the observed variance in nitrate. Other species differences explained an additional 9.2%, and total soil N an additional 5% of this variance. Extractable soil ammonium also depended on these variables, but total soil N explained the most variance. Light penetration to the soil surface in these monocultures was a negative exponential function of aboveground biomass (R^2 - 0.79). Schizachyrium and Andropogon, the species that reduce soil solution N to the lowest levels on infertile soils, had lower vegetative growth rates, higher root allocation, lower reproductive allocation, and lower tissue N than the other species. Many of these traits are associated with plants on infertile habitats, suggesting a direct link between ecophysiology, resource reduction, and distributional patterns. Because all species survived on even our most nitrogen-poor soil (subsurface sand), differential nutrient reduction, not tolerance, may be the main mechanism favoring these traits in infertile habitats. On infertile soils, the three earlier successional species (Agrostis, Agropyron, and Poa) allocated more to reproduction (rhizome or seed) than the later successional species, but did not reduce soil solution nitrate and ammonium to as low levels. This suggests that our early successional species may be superior colonists but inferior nitrogen competitors compared to the prairie bunchgrasses. Our results can be used to make explicit predictions as to the outcome of nitrogen competition among all possible combinations of these five species.
[本文引用: 1]
DOI:10.1111/gcb.12812URLPMID:25418963 [本文引用: 1]
Abstract Elevated ultraviolet (UV) radiation has been demonstrated to stimulate litter decomposition. Despite years of research, it is still not fully understood whether the acceleration in litter degradation is primarily attributed to abiotic photodegradation or the combined effects of abiotic photodegradation and microbial decomposition. In this study, we used meta-analysis to synthesize photodegradation studies and compared the effects of UV radiation on litter decomposition between abiotic and biotic conditions. We also conducted a microcosm experiment to assess the effects of UV radiation on litter biodegradability and microbial activity. Overall, our meta-analysis found that under abiotic photodegradation, UV radiation reduced the remaining litter mass by 1.44% (95% CI: 0.85% to 2.08%), did not affect the remaining lignin and increased the dissolved organic carbon (DOC) concentration by 14.01% (1.49–23.67%). Under combined abiotic photodegradation and microbial decomposition, UV radiation reduced the remaining litter mass and lignin by 1.60% (0.04–3.58%) and 16.07% (9.27–24.23%), respectively, but did not alter DOC concentration. UV radiation had no significant impact on soil microbial biomass carbon (MBC), but it reduced microbial respiration by 44.91% (2.26–78.62%) and altered the composition of the microbial community. In addition, UV radiation reduced nitrogen (N) immobilization by 19.44% (4.77–37.92%). Our microcosm experiment further indicated that DOC concentration and the amount of respired C in UV-treated litter increased with UV exposure time, suggesting that longer UV exposure resulted in greater biodegradability. Overall, our study suggested that UV exposure could increase litter biodegradability by increasing the microbial accessibility of lignin, as well as the labile carbon supply to microbes. However, the remaining litter mass was not different between the abiotic and biotic conditions, most likely because the positive effect of UV radiation on litter biodegradability was offset by its negative effect on microbial activity. Our results also suggested that UV radiation could alter the N cycle during decomposition, primarily by inhibiting N immobilization.
DOI:10.1016/S2095-3119(12)60466-3URL [本文引用: 1]
Grassland plays an important role in the global carbon cycle and climate regulation. However, there are still large uncertainties in grassland carbon pool and also its role in global carbon cycle due to the lack of measured grassland biomass at regional scale or global scale with a unified survey method, particular for below-ground biomass. The present study, based on a total of 44 grassland sampling plots with 220 quadrats across Ningxia, investigated the characteristics of above-ground biomass (AGB), below-ground biomass (BGB), litter biomass (LB), total biomass (TB) and root:shoot ratios (R:S) for six predominantly grassland types, and their relationships with climatic factors. AGB, BGB, LB and TB varied markedly across different grassland types, the median value ranging from 28.2-692.6 g m-2 forAGB, 130.4-2036.6 g m-2 for BGB, 9.2-82.3 g m-2 for LB, and 168.0-2 681.3 g m-2 for TB. R:S showed less variation with median values from 3.2 to 5.3 (excluding marshy meadow). The different grassland types showed similar patterns of biomass allocation, with more than 70% BGB for all types. There is evidence of strong positive effects associated with mean annual precipitation (MAP) and negative effects associated with mean annual temperature (MAT) on AGB, BGB, and LB, although both factors have the opposite effect on R:S.
[本文引用: 1]
[本文引用: 1]
DOI:10.1093/treephys/18.2.129URLPMID:12651397 [本文引用: 1]
Abstract Considerable progress has been made in our ability to model and measure annual gross primary production (GPP) by terrestrial vegetation. But challenges remain in estimating maintenance respiration (R(m)) and net primary production (NPP). To search for possible common relationships, we assembled annual carbon budgets from six evergreen and one deciduous forest in Oregon, USA, three pine plantations in New South Wales, Australia, a deciduous forest in Massachusetts, USA, and a Nothofagus forest on the South Island of New Zealand. At all 12 sites, a standard procedure was followed to estimate annual NPP of foliage, branches, stems, and roots, the carbon expended in synthesis of these organs (R(g)), their R(m), and that of previously produced foliage and sapwood in boles, branches, and large roots. In the survey, total NPP ranged from 120 to 1660 g C m(-2) year(-1), whereas the calculated fraction allocated to roots varied from 0.22 to 0.63. Comparative analysis indicated that the total NPP/GPP ratio was conservative (0.47 +/- 0.04 SD). This finding supports the possibility of greatly simplifying forest growth models. The constancy of the NPP/GPP ratio also provides an incentive to renew efforts to understand the environmental factors affecting partitioning of NPP above and belowground.
DOI:10.1641/0006-3568(2003)053[0941:ATROTE]2.0.CO;2URL [本文引用: 1]
Changes in Earth's surface temperatures caused by anthropogenic emissions of greenhouse gases are expected to affect global and regional precipitation regimes. Interactions between changing precipitation regimes and other aspects of global change are likely to affect natural and managed terrestrial ecosystems as well as human society. Although much recent research has focused on assessing the responses of terrestrial ecosystems to rising carbon dioxide or temperature, relatively little research has focused on understanding how ecosystems respond to changes in precipitation regimes. Here we review predicted changes in global and regional precipitation regimes, outline the consequences of precipitation change for natural ecosystems and human activities, and discuss approaches to improving understanding of ecosystem responses to changing precipitation. Further, we introduce the Precipitation and Ecosystem Change Research Network (PrecipNet), a new interdisciplinary research network assembled to encourage and foster communication and collaboration across research groups with common interests in the impacts of global change on precipitation regimes, ecosystem structure and function, and the human enterprise.
DOI:10.1007/s10531-011-0145-6URL [本文引用: 1]
AbstractPlant diversity has been reported to increase productivity. Farming practices aiming at conserving or increasing plant diversity are, however, usually less profitable than conventional ones. In this review, we aim to find reasons for this discrepancy, discuss ecosystem services of grassland phytodiversity that are useful for farmers, and ways of livestock management most beneficial for diversity. Under agricultural conditions, a clear effect of species richness on a site primary or secondary production has not yet been demonstrated. Reasons could be that species numbers in permanent grassland are above the threshold of five species found effective in experimental plots or that the conditions are more in equilibrium with management than in weeded experimental plots. Other diversity effects on production stability, nutrient and water retention or product quality might convince farmers to increase diversity. However, these should be tested in agricultural situations, as most research has again been carried out in experimental plots. To enhance phytodiversity, grazing has been found superior over mowing, as selective grazing, treading and excreta deposition increase the heterogeneity of a sward and thus the niches available. Especially rotational grazing with intermediate intensity may be advantageous for phytodiversity. However, complex interactions between environmental conditions, sward composition, management and livestock behaviour make it difficult to forecast grazing effects. Thus, ecological and agricultural researchers should cooperate more, e.g. either in interdisciplinary projects or by hiring researchers from the respective other profession and thus diversifying research groups, in order to integrate agricultural management into biodiversity research and biodiversity measurements into agricultural research to advance our understanding of how to make conservation and enhancement of grassland phytodiversity both feasible and sustainable.
DOI:10.1111/j.1365-2486.2010.02302.xURL [本文引用: 1]
Global mean temperature is predicted to increase by 2 7 C and precipitation to change across the globe by the end of this century. To quantify climate effects on ecosystem processes, a number of climate change experiments have been established around the world in various ecosystems. Despite these efforts, general responses of terrestrial ecosystems to changes in temperature and precipitation, and especially to their combined effects, remain unclear. We used meta-analysis to synthesize ecosystem-level responses to warming, altered precipitation, and their combination. We focused on plant growth and ecosystem carbon (C) balance, including biomass, net primary production (NPP), respiration, net ecosystem exchange (NEE), and ecosystem photosynthesis, synthesizing results from 85 studies. We found that experimental warming and increased precipitation generally stimulated plant growth and ecosystem C fluxes, whereas decreased precipitation had the opposite effects. For example, warming significantly stimulated total NPP, increased ecosystem photosynthesis, and ecosystem respiration. Experimentally reduced precipitation suppressed aboveground NPP (ANPP) and NEE, whereas supplemental precipitation enhanced ANPP and NEE. Plant productivity and ecosystem C fluxes generally showed higher sensitivities to increased precipitation than to decreased precipitation. Interactive effects of warming and altered precipitation tended to be smaller than expected from additive, single-factor effects, though low statistical power limits the strength of these conclusions. New experiments with combined temperature and precipitation manipulations are needed to conclusively determine the importance of temperature recipitation interactions on the C balance of terrestrial ecosystems under future climate conditions.
DOI:10.1111/j.1469-8137.2008.02488.xURLPMID:19086179 [本文引用: 1]
Summary Top of page Summary Introduction Materials and Methods Results Discussion Acknowledgements References Supporting Information 6168 Better understanding of the responses of terrestrial plant species under global nitrogen (N) enrichment is critical for projection of changes in structure, functioning, and service of terrestrial ecosystems. 6168 Here, a meta-analysis of data from 304 studies was carried out to reveal the general response patterns of terrestrial plant species to the addition of N. 6168 Across 456 terrestrial plant species included in the analysis, biomass and N concentration were increased by 53.6 and 28.5%, respectively, under N enrichment. However, the N responses were dependent upon plant functional types, with significantly greater biomass increases in herbaceous than in woody species. Stimulation of plant biomass by the addition of N was enhanced when other resources were improved. In addition, the N responses of terrestrial plants decreased with increasing latitude and increased with annual precipitation. 6168 Dependence of the N responses of terrestrial plants on biological realms, functional types, tissues, other resources, and climatic factors revealed in this study can help to explain changes in species composition, diversity, community structure and ecosystem functioning under global N enrichment. These findings are critical in improving model simulation and projection of terrestrial carbon sequestration and its feedbacks to global climate change, especially when progressive N limitation is taken into consideration.
DOI:10.1016/j.jaridenv.2011.03.003URL [本文引用: 1]
Evidence supporting water limitation in arid-semiarid ecosystems includes strong correlations between aboveground net primary production (ANPP) and annual precipitation as well as results from experimental water additions. Similarly, there is evidence of N limitation on ANPP in low precipitation ecosystems, but is this a widespread phenomenon? Are all arid-semiarid ecosystems equally limited by nitrogen? Is the response of N fertilization modulated by water availability? We conducted a meta-analysis of ANPP responses to N fertilization across arid to subhumid ecosystems to quantify N limitation, using the effect-size index R which is the ratio of ANPP in fertilized to control plots. Nitrogen addition increased ANPP across all studies by an average of 50%, and nitrogen effects increased significantly ( P02=020.03) along the 50–65002mm02yr 611 precipitation gradient. The response ratio decreased with mean annual temperature in arid and semiarid ecosystems but was insensitive in subhumid systems. Sown pastures showed significant ( P02=020.007) higher responses than natural ecosystems. Neither plant-life form nor chemical form of the applied fertilizer showed significant effects on the primary production response to N addition. Our results showed that nitrogen limitation is a widespread phenomenon in low-precipitation ecosystems and that its importance increases with annual precipitation from arid to subhumid regions. Both water and N availability limit primary production, probably at different times during the year; with frequency of N limitation increasing and frequency of water limitation decreasing as annual precipitation increases. Expected increase N deposition, which could be significant even in arid ecosystems, would increase aboveground net primary production in water-limited ecosystems that account for 40% of the terrestrial surface.
DOI:10.11686/cyxb20130109URLMagsci [本文引用: 1]
草原与荒漠生态系统中一年生植物是高度顺应气候波动、特别是降水波动的特殊类群。本试验以内蒙古草原与荒漠区4种一年生植物为研究对象,通过室内模拟5,20,70和150mm4个有效降水梯度,分析了4种一年生植物物候及其形态等性状对降水变化的响应。结果表明,草原与荒漠一年生植物类群生长期通常在40~90(<100)d;随降水梯度的增加,一年生植物的生长期、植物高度、叶长、生物量等均增加,5mm 有效降水能基本完成生活史,70mm 有效降水资源分配达到最佳化,结实量最大;一年生猪毛菜类在不同降水条件下的物候期长短的变化比一年生小禾草类小,倾向于以调整生殖生长期的长短的方式,减少结实量为代价加速种子成熟的方式响应降水量不足;一年生小禾草类能根据降水量变化及时调整各物候期长短以适应生长和繁殖;相同降水条件下小禾草类植物的生长期相对较长;在适应方式上,猪毛菜类植物表现出更强的避旱能力,而小禾草类植物则表现出相对更强的耐旱能力。
DOI:10.11686/cyxb20130109URLMagsci [本文引用: 1]
草原与荒漠生态系统中一年生植物是高度顺应气候波动、特别是降水波动的特殊类群。本试验以内蒙古草原与荒漠区4种一年生植物为研究对象,通过室内模拟5,20,70和150mm4个有效降水梯度,分析了4种一年生植物物候及其形态等性状对降水变化的响应。结果表明,草原与荒漠一年生植物类群生长期通常在40~90(<100)d;随降水梯度的增加,一年生植物的生长期、植物高度、叶长、生物量等均增加,5mm 有效降水能基本完成生活史,70mm 有效降水资源分配达到最佳化,结实量最大;一年生猪毛菜类在不同降水条件下的物候期长短的变化比一年生小禾草类小,倾向于以调整生殖生长期的长短的方式,减少结实量为代价加速种子成熟的方式响应降水量不足;一年生小禾草类能根据降水量变化及时调整各物候期长短以适应生长和繁殖;相同降水条件下小禾草类植物的生长期相对较长;在适应方式上,猪毛菜类植物表现出更强的避旱能力,而小禾草类植物则表现出相对更强的耐旱能力。
DOI:10.1029/2008GL035408URL [本文引用: 1]
Aboveground net primary production (ANPP) is strongly correlated with annual precipitation (AP) in grassland ecosystems. However, the relationship between the interannual variation in ANPP and the variability in precipitation remains controversial. In this study, we used long-term data of biomass and precipitation from 118 sites across global grasslands to examine the relationship between variability in ANPP and AP. Our results showed that ANPP increased with precipitation, but leveled off in humid regions, and that increased variation in precipitation led to an increase in the variability in ANPP. The relative ANPP maxima significantly increased with relative AP maxima and the relative ANPP minima also positively correlated with relative AP minima. These suggest that the fluctuations in precipitation can alter the growth of grasslands, which should be incorporated into the prediction and modeling of climate changes.
DOI:10.1007/s00442-006-0409-0URLPMID:16708228 [本文引用: 1]
The concept of nutrient use efficiency is central to understanding ecosystem functioning because it is the step in which plants can influence the return of nutrients to the soil pool and the quality of the litter. Theory suggests that nutrient efficiency increases unimodally with declining soil resources, but this has not been tested empirically for N and water in grassland ecosystems, where plant growth in these ecosystems is generally thought to be limited by soil N and moisture. In this paper, we tested the N uptake and the N use efficiency (NUE) of two Stipa species (S. grandis and S. krylovii) from 20 sites in the Inner Mongolia grassland by measuring the N content of net primary productivity (NPP). NUE is defined as the total net primary production per unit N absorbed. We further distinguished NUE from N response efficiency (NRE; production per unit N available). We found that NPP increased with soil N and water availability. Efficiency of whole-plant N use, up-take, and response increased monotonically with decreasing soil N and water, being higher on infertile (dry) habitats than on fertile (wet) habitats. We further considered NUE as the product of the N productivity (NP the rate of biomass increase per unit N in the plant) and the mean residence time (MRT; the ratio between the average N pool and the annual N uptake or loss). The NP and NUE of S. grandis growing usually in dry and N-poor habitats exceeded those of S. krylovii abundant in wet and N-rich habitats. NUE differed among sites, and was often affected by the evolutionary trade-off between NP and MRT, where plants and communities had adapted in a way to maximize either NP or MRT, but not both concurrently. Soil N availability and moisture influenced the community-level N uptake efficiency and ultimately the NRE, though the response to N was dependent on the plant community examined. These results show that soil N and water had exerted a great impact on the N efficiency in Stipa species. The intraspecific differences in N efficiency within both Stipa species along soil resource availability gradient may explain the differences in plant productivity on various soils, which will be conducive to our general understanding of the N cycling and vegetation dynamics in northern Chinese grasslands.
DOI:10.1007/s10021-009-9296-7URL
Knowledge of how ecosystem carbon (C) processes respond to variations in precipitation is crucial for assessing impacts of climate change on terrestrial ecosystems. In this study, we examined variations of shoot and root biomass, standing and surface litter, soil respiration, and soil C content along a natural precipitation gradient from 430 to 1200 mm in the southern Great Plains, USA. Our results show that shoot biomass and soil respiration increased linearly with mean annual precipitation (MAP), whereas root biomass and soil C content remained relatively constant along the precipitation gradient. Consequently, the root/shoot ratio linearly decreased with MAP. However, patterns of standing, surface, and total litter mass followed quadratic relationships with MAP along the gradient, likely resulting from counterbalance between litter production and decomposition. Those linear/quadratic equations describing variations of eco-system C processes with precipitation could be useful for model development, parameterization, and validation at landscape and regional scales to improve predictions of C dynamics in grasslands in response to climate change. Our results indicated that precipitation is an important driver in shaping ecosystem functioning as reflected in vegetation production, litter mass, and soil respiration in grassland ecosystems.
DOI:10.1073/pnas.1606734113URLPMID:27601643 [本文引用: 1]
Global changes in climate, atmospheric composition, and pollutants are altering ecosystems and the goods and services they provide. Among approaches for predicting ecosystem responses, long-term observations and manipulative experiments can be powerful approaches for resolving single-factor and interactive effects of global changes on key metrics such as net primary production (NPP). Here we combine both approaches, developing multidimensional response surfaces for NPP based on the longest-running, best-replicated, most-multifactor global-change experiment at the ecosystem scale 17-y study of California grassland exposed to full-factorial warming, added precipitation, elevated CO2, and nitrogen deposition. Single-factor and interactive effects were not time-dependent, enabling us to analyze each year as a separate realization of the experiment and extract NPP as a continuous function of global-change factors. We found a ridge-shaped response surface in which NPP is humped (unimodal) in response to temperature and precipitation when CO2 and nitrogen are ambient, with peak NPP rising under elevated CO2 or nitrogen but also shifting to lower temperatures. Our results suggest that future climate change will push this ecosystem away from conditions that maximize NPP, but with large year-to-year variability.
Resampling tests for meta-analysis of ecological data
1
1997
... 每个指标RR的计算是通过对$\bar{E}$进行反对数转换以降低偏倚, 使样本近似成正态分布(
The influence of plant diversity and functional composition on ecosystem stability of four Stipa communities in the Inner Mongolia Plateau
1
2001
... 研究发现, 在低氮条件下, 植物为了获取氮, 会增加向地下分配的光合产物(
Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: Evidence from inner Mongolia grasslands
1
2010
... 研究发现, 在低氮条件下, 植物为了获取氮, 会增加向地下分配的光合产物(
Primary production and rain use efficiency across a precipitation gradient on the Mongolia Plateau
1
2008
... 模型预测的结果表明, 未来草地生态系统仍将受到氮沉降加剧(
Soil microbial responses to temporal variations of moisture and temperature in a Chihuahuan Desert grassland
1
2008
... 氮添加的同时进行增雨处理显著增加地下生物量, 而且效果显著高于单独增雨(42%, 10%,
The effects of air-borne nitrogen pollutants on species diversity in natural and semi-natural European vegetation
1
1998
... 氮添加对草地生态系统生产力的促进作用存在阈值(
Effects of N and water supply on water use-efficiency of a semiarid grassland in Inner Mongolia
1
2010
... 氮添加的同时进行增雨处理显著增加地下生物量, 而且效果显著高于单独增雨(42%, 10%,
Regional and temporal variation in net primary production and nitrogen mineralization in grasslands
2
1997
... 随氮添加浓度提高, 草地地下、总生物量及地上地下分配比例没有明显的增加趋势(
... 在增雨处理下, 寒冷性气候区的草地生态系统的地下生物量、总生物量受到的促进作用显著高于温暖性气候区的草地生态系统(
氮添加对土壤呼吸影响的研究进展
1
2013
... 随氮添加浓度提高, 草地地下、总生物量及地上地下分配比例没有明显的增加趋势(
氮添加对土壤呼吸影响的研究进展
1
2013
... 随氮添加浓度提高, 草地地下、总生物量及地上地下分配比例没有明显的增加趋势(
北半球陆地生态系统碳交换通量的空间格局及其区域特征
2
2014
... 较大区域尺度上, 温度、降水等环境因子是影响植物生产力的关键气候因素(
... ), 导致生产力受到的促进作用降低.除了温度的调控作用之外, 不同气候区的陆地生态系统也同时受到水分状况的影响, 无论是热带、亚热带还是温带区域内, 在其相对湿润的气候区具有更高的生产力(
北半球陆地生态系统碳交换通量的空间格局及其区域特征
2
2014
... 较大区域尺度上, 温度、降水等环境因子是影响植物生产力的关键气候因素(
... ), 导致生产力受到的促进作用降低.除了温度的调控作用之外, 不同气候区的陆地生态系统也同时受到水分状况的影响, 无论是热带、亚热带还是温带区域内, 在其相对湿润的气候区具有更高的生产力(
The change of North China climate in transient simulations using the IPCC SRES A2 and B2 scenarios with a coupled atmosphere-ocean general circulation model
1
2003
... 模型预测的结果表明, 未来草地生态系统仍将受到氮沉降加剧(
Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands
1
2008
... 长期氮添加处理对地下生物量的促进作用在处理后期转为抑制作用(
Competition between native perennial and exotic annual grasses: Implications for an historical invasion
1
2004
... 随氮添加浓度提高, 草地地下、总生物量及地上地下分配比例没有明显的增加趋势(
Forest carbon use efficiency: Is respiration a constant fraction of gross primary production?
1
2007
... 草地生态系统作为陆地生态系统重要的组成部分, 其生产力约占陆地总初级生产力的1/3 (
Is plant growth driven by sink regulation? Implications for crop models, phenotyping approaches and ideotypes
1
2007
... 研究发现, 在低氮条件下, 植物为了获取氮, 会增加向地下分配的光合产物(
Responses of grassland production to single and multiple global environmental changes
1
2005
... 随氮添加浓度提高, 草地地下、总生物量及地上地下分配比例没有明显的增加趋势(
Climate extremes: Observations, modeling, and impacts
1
2000
... 模型预测的结果表明, 未来草地生态系统仍将受到氮沉降加剧(
In vitro skin permeation of estradiol from various proniosome formulations
1
2001
... 较大区域尺度上, 温度、降水等环境因子是影响植物生产力的关键气候因素(
Long-term nitrogen addition leads to loss of species richness due to litter accumulation and soil acidification in a temperate steppe
1
2012
... 氮添加对草地生态系统生产力的促进作用存在阈值(
Altering rainfall timing and quantity in a mesic grassland ecosystem: Design and performance of rainfall manipulation shelters
1
2000
... 研究发现, 在低氮条件下, 植物为了获取氮, 会增加向地下分配的光合产物(
Grassland productivity limited by multiple nutrients
2
2015
... 模型预测的结果表明, 未来草地生态系统仍将受到氮沉降加剧(
... 在增雨处理下, 寒冷性气候区的草地生态系统的地下生物量、总生物量受到的促进作用显著高于温暖性气候区的草地生态系统(
Resource manipulation effects on net primary production, biomass allocation and rain-use efficiency of two semiarid grassland sites in Inner Mongolia, China
1
2011
... 一般而言, 添加氮和增雨交互作用能显著提高群落地上生物量和总生物量(
Vascular plant species richness in Alaskan arctic tundra: The importance of soil pH
1
2000
... 随氮添加浓度提高, 草地地下、总生物量及地上地下分配比例没有明显的增加趋势(
Patterns of NPP, GPP, respiration, and NEP during boreal forest succession
1
2011
... 草地生态系统作为陆地生态系统重要的组成部分, 其生产力约占陆地总初级生产力的1/3 (
Effect of light and nitrogen supply on internal C:N balance and control of root-to-shoot biomass allocation in grapevine
1
2007
... 研究发现, 在低氮条件下, 植物为了获取氮, 会增加向地下分配的光合产物(
氮添加对黄土高原典型草原长芒草光合特性的影响
1
2016
... 氮添加对草地生态系统生产力的促进作用存在阈值(
氮添加对黄土高原典型草原长芒草光合特性的影响
1
2016
... 氮添加对草地生态系统生产力的促进作用存在阈值(
氮添加对黄土高原典型草原植物比叶面积及其重要值的影响
1
2015
... 氮添加对草地生态系统生产力的促进作用存在阈值(
氮添加对黄土高原典型草原植物比叶面积及其重要值的影响
1
2015
... 氮添加对草地生态系统生产力的促进作用存在阈值(
Ecosystem responses to water and nitrogen amendment in a California grassland
1
2007
... 一般而言, 添加氮和增雨交互作用能显著提高群落地上生物量和总生物量(
Root dynamics of tallgrass prairie in wet and dry years
1
1987
... 研究发现, 在低氮条件下, 植物为了获取氮, 会增加向地下分配的光合产物(
Drought effect on plant nitrogen and phosphorus: A meta-analysis
1
2014
... 研究发现, 在低氮条件下, 植物为了获取氮, 会增加向地下分配的光合产物(
The meta-analysis of response ratios in experimental ecology
2
1999
... 本研究通过数据整合(meta-analysis,
... 每个指标RR的计算是通过对$\bar{E}$进行反对数转换以降低偏倚, 使样本近似成正态分布(
Confronting a biome crisis: Global disparities of habitat loss and protection
1
2005
... 草地生态系统作为陆地生态系统重要的组成部分, 其生产力约占陆地总初级生产力的1/3 (
Nitrogen limitation in dryland ecosystems: Responses to geographical and temporal variation in precipitation
1
1999
... 草地生态系统作为陆地生态系统重要的组成部分, 其生产力约占陆地总初级生产力的1/3 (
草地生态系统对模拟大气增温的响应
1
2014
... 在增雨处理下, 寒冷性气候区的草地生态系统的地下生物量、总生物量受到的促进作用显著高于温暖性气候区的草地生态系统(
草地生态系统对模拟大气增温的响应
1
2014
... 在增雨处理下, 寒冷性气候区的草地生态系统的地下生物量、总生物量受到的促进作用显著高于温暖性气候区的草地生态系统(
Managing the Risks of Extreme Events and Disasters to Advance Climate Change Adaptation
2
2012
... 研究发现, 在低氮条件下, 植物为了获取氮, 会增加向地下分配的光合产物(
... 模型预测的结果表明, 未来草地生态系统仍将受到氮沉降加剧(
Biomass and its allocation in relation to temperature, precipitation, and soil nutrients in Inner Mongolia grasslands, China
1
2013
... 在增雨处理下, 寒冷性气候区的草地生态系统的地下生物量、总生物量受到的促进作用显著高于温暖性气候区的草地生态系统(
Rapid shifts in plant distribution with recent climate change
2008
Frequency and extent of water limitation to primary production in a mesic temperate grassland
1
2001
... 研究发现, 在低氮条件下, 植物为了获取氮, 会增加向地下分配的光合产物(
2
2013
... 研究发现, 在低氮条件下, 植物为了获取氮, 会增加向地下分配的光合产物(
... 每个指标RR的计算是通过对$\bar{E}$进行反对数转换以降低偏倚, 使样本近似成正态分布(
Long-term forage production of North American shortgrass steppe
1
1992
... 模型预测的结果表明, 未来草地生态系统仍将受到氮沉降加剧(
Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed
1
2008
... 研究发现, 在低氮条件下, 植物为了获取氮, 会增加向地下分配的光合产物(
A global comparison of grassland biomass responses to CO2 and nitrogen enrichment
5
2010
... 研究发现, 在低氮条件下, 植物为了获取氮, 会增加向地下分配的光合产物(
... ;
... 氮添加的同时进行增雨处理显著增加地下生物量, 而且效果显著高于单独增雨(42%, 10%,
... ).在环境较为干旱时, 养分的迁移和传输均受到限制, 随着降雨量的增加, 土壤中养分限制得以解除, 从而提高养分的可利用性, 促进植物对养分的吸收利用, 最终提高生态系统生产力(
... 长期氮添加处理对地下生物量的促进作用在处理后期转为抑制作用(
Above and belowground net primary productivity of grassland influenced by supplemental water and nitrogen in Inner Mongolia
1
2011
... 一般而言, 添加氮和增雨交互作用能显著提高群落地上生物量和总生物量(
氮和水分添加对贝加尔针茅草原植物多样性及生物量的影响
1
2015
... 其中, ${{\bar{X}}_{\text{NT}}}$是标准化后的施氮响应值, 效应值${{\bar{X}}_{\text{T}}}$、${{\bar{X}}_{\text{C}}}$分别为原始实验处理和对照数值, 标准化LRR(normalized LRR) = log$\frac{{{{\bar{X}}}_{\text{NT}}}}{{{{\bar{X}}}_{\text{C}}}}$ (
氮和水分添加对贝加尔针茅草原植物多样性及生物量的影响
1
2015
... 其中, ${{\bar{X}}_{\text{NT}}}$是标准化后的施氮响应值, 效应值${{\bar{X}}_{\text{T}}}$、${{\bar{X}}_{\text{C}}}$分别为原始实验处理和对照数值, 标准化LRR(normalized LRR) = log$\frac{{{{\bar{X}}}_{\text{NT}}}}{{{{\bar{X}}}_{\text{C}}}}$ (
A cross-biome synthesis of soil respiration and its determinants under simulated precipitation changes
1
2016
... 模型预测的结果表明, 未来草地生态系统仍将受到氮沉降加剧(
Enhanced nitrogen deposition over China
1
2013
... 每个指标RR的计算是通过对$\bar{E}$进行反对数转换以降低偏倚, 使样本近似成正态分布(
Response of ecosystem carbon cycle to experimental warming: A meta-analysis
1
2013
... 一般而言, 添加氮和增雨交互作用能显著提高群落地上生物量和总生物量(
施肥, 灌溉及火烧对荒漠草原土壤养分和植物群落特征的影响
1
2014
... 氮添加对草地生态系统生产力的促进作用存在阈值(
施肥, 灌溉及火烧对荒漠草原土壤养分和植物群落特征的影响
1
2014
... 氮添加对草地生态系统生产力的促进作用存在阈值(
Ecosystem carbon storage in arctic tundra reduced by long-term nutrient fertilization
1
2004
... 随氮添加浓度提高, 草地地下、总生物量及地上地下分配比例没有明显的增加趋势(
沙质草地不同生活史植物的生物量分配对氮和水分添加的响应
2014
沙质草地不同生活史植物的生物量分配对氮和水分添加的响应
2014
Variable effects of nitrogen additions on the stability and turnover of soil carbon
1
2002
... 模型预测的结果表明, 未来草地生态系统仍将受到氮沉降加剧(
A high-resolution data set of surface climate over global land areas
2
2002
... 实验数据的筛选标准为: (1)实验设置位点需是野外原位草地生态系统, 非室内控制实验; (2)野外控制实验结果以图或表的形式说明实验组(如: 氮添加)和对照组的处理结果$\bar{X}_{T}$和$\bar{X}_{C}$; 重复次数NT和NC; 标准误SET和SEC; 标准偏差SDT和SDC; (3)单一实验处理至少包含两个处理梯度; (4)增雨处理仅包含雨量的变化, 不含降雨格局的变化; (5)在不同实验处理(如氮添加、增雨处理和同时增氮增雨处理)中, 测定相应变化指标, 主要包括草地生态系统地上、地下、总生物量及地上、地下生物量比.同时, 本研究收集包括野外实验站点经纬度、年平均气温(MAT)、年降水量(MAP)等辅助信息.对于缺失MAT、MAP信息的文献, 本研究通过R软件程序包, 利用Climatic Research Unit Climatology version 2.0 dataset (CRUCL 2.0,
... 氮添加的同时进行增雨处理显著增加地下生物量, 而且效果显著高于单独增雨(42%, 10%,
Non-additive effects of water and nitrogen addition on ecosystem carbon exchange in a temperate steppe
1
2009
... 研究发现, 在低氮条件下, 植物为了获取氮, 会增加向地下分配的光合产物(
Hierarchical plant responses and diversity loss after nitrogen addition: Testing three functionally-based hypotheses in the Inner Mongolia grassland
2
2011
... 草地生态系统作为陆地生态系统重要的组成部分, 其生产力约占陆地总初级生产力的1/3 (
... 一般而言, 添加氮和增雨交互作用能显著提高群落地上生物量和总生物量(
Exacerbated nitrogen limitation ends transient stimulation of grassland productivity by increased precipitation
1
2017
... 长期氮添加处理对地下生物量的促进作用在处理后期转为抑制作用(
Effects of drought on nutrient uptake and assimilation in vegetable crops
2
2012
... 研究发现, 在低氮条件下, 植物为了获取氮, 会增加向地下分配的光合产物(
... ).目前, 地下生物量对增雨的响应尚没有完全统一的结论, 呈增加(
Primary production of the central grassland region of the United States
1
1988
... 氮添加对草地生态系统生产力的促进作用存在阈值(
Contribution of acidification and eutrophication to declines in species richness of calcifuge grasslands along a gradient of atmospheric nitrogen deposition
1
2010
... 在增雨处理下, 寒冷性气候区的草地生态系统的地下生物量、总生物量受到的促进作用显著高于温暖性气候区的草地生态系统(
Climate Change 2013: The Physical Science Basis
1
2013
... 模型预测的结果表明, 未来草地生态系统仍将受到氮沉降加剧(
Precipitation extreme changes exceeding moisture content increases in MIROC and IPCC climate models
1
2010
... 较大区域尺度上, 温度、降水等环境因子是影响植物生产力的关键气候因素(
Defoliation synchronizes aboveground growth of co-occurring C4 grass species
1
2008
... 随氮添加浓度提高, 草地地下、总生物量及地上地下分配比例没有明显的增加趋势(
Effect of precipitation variability on net primary production and soil respiration in a Chihuahuan Desert grassland
1
2011
... 随氮添加浓度提高, 草地地下、总生物量及地上地下分配比例没有明显的增加趋势(
The resource-ratio hypothesis of plant succession
1
1985
... 研究发现, 在低氮条件下, 植物为了获取氮, 会增加向地下分配的光合产物(
Plant traits and resource reduction for five grasses growing on a nitrogen gradient
1
1991
... 以上所有整合分析及整合回归计算由R软件里的metafor软件包实现(
Conducting meta-analyses in R with the metafor package
2010
Nitrogen limitation on land and in the sea: How can it occur?
1
1991
... 模型预测的结果表明, 未来草地生态系统仍将受到氮沉降加剧(
The interaction between abiotic photodegradation and microbial decomposition under ultraviolet radiation
1
2015
... 研究发现, 在低氮条件下, 植物为了获取氮, 会增加向地下分配的光合产物(
Biomass components and environmental controls in Ningxia grasslands
1
2012
... 长期氮添加处理对地下生物量的促进作用在处理后期转为抑制作用(
内蒙古羊草草原植物群落地上初级生产力时间动态对降水变化的响应
1
2004
... 草地生态系统作为陆地生态系统重要的组成部分, 其生产力约占陆地总初级生产力的1/3 (
内蒙古羊草草原植物群落地上初级生产力时间动态对降水变化的响应
1
2004
... 草地生态系统作为陆地生态系统重要的组成部分, 其生产力约占陆地总初级生产力的1/3 (
Net primary production of forests: A constant fraction of gross primary production?
1
1998
... 研究发现, 在低氮条件下, 植物为了获取氮, 会增加向地下分配的光合产物(
Assessing the response of terrestrial ecosystems to potential changes in precipitation
1
2003
... 草地生态系统作为陆地生态系统重要的组成部分, 其生产力约占陆地总初级生产力的1/3 (
Phytodiversity of temperate permanent grasslands: Ecosystem services for agriculture and livestock management for diversity conservation
1
2011
... 草地生态系统作为陆地生态系统重要的组成部分, 其生产力约占陆地总初级生产力的1/3 (
Responses of terrestrial ecosystems to temperature and precipitation change: A meta-analysis of experimental manipulation
1
2011
... 在增雨处理下, 寒冷性气候区的草地生态系统的地下生物量、总生物量受到的促进作用显著高于温暖性气候区的草地生态系统(
Global response patterns of terrestrial plant species to nitrogen addition
1
2008
... 草地生态系统作为陆地生态系统重要的组成部分, 其生产力约占陆地总初级生产力的1/3 (
Nitrogen limitation in arid-subhumid ecosystems: A meta-analysis of fertilization studies
1
2011
... 随氮添加浓度提高, 草地地下、总生物量及地上地下分配比例没有明显的增加趋势(
草原与荒漠一年生植物性状对降水变化的响应
1
2013
... 较大区域尺度上, 温度、降水等环境因子是影响植物生产力的关键气候因素(
草原与荒漠一年生植物性状对降水变化的响应
1
2013
... 较大区域尺度上, 温度、降水等环境因子是影响植物生产力的关键气候因素(
Relationship between variability in aboveground net primary production and precipitation in global grasslands
1
2008
... 长期氮添加处理对地下生物量的促进作用在处理后期转为抑制作用(
Nitrogen response efficiency increased monotonically with decreasing soil resource availability: A case study from a semiarid grassland in northern China
1
2006
... 研究发现, 在低氮条件下, 植物为了获取氮, 会增加向地下分配的光合产物(
Biomass, litter, and soil respiration along a precipitation gradient in southern Great Plains, USA
2009
Nonlinear, interacting responses to climate limit grassland production under global change
1
2016
... 长期氮添加处理对地下生物量的促进作用在处理后期转为抑制作用(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}