Response of soil enzyme activities to litter input changes in two secondary Castanopsis carlessii forests in subtropical China
WEI Cui-Cui, LIU Xiao-Fei, LIN Cheng-Fang,*, LI Xian-Feng*, LI Yan*, ZHENG Yu-Xiong*School of Geographical Sciences, Fujian Normal University, Fuzhou, 350007, China; State Key Laboratory of Humid Subtropical Mountain Ecology, Fuzhou,350007, China; and Sanming Research Station of Forest Ecosystem and Global Change, Sanming, Fujian 365000, China
Supported by the National Natural Science Foundation of China.31770663 Supported by the National Natural Science Foundation of China.31500407 the Joint Fund for Promotion of Cross-strait Cooperation in Science and Technology .U1505233
Abstract Aims Enzymes play an important role in the decomposition of soil organic matter. Changes in net primary productivity in response to climate change are likely to affect litter inputs to forest soil. However, the effects of altered litter input on soil enzyme activities remain poorly understood in subtropical forests. Thus, this study is designed to find out if litter manipulation has an effect on enzymes in different subtropical Castanopsis carlessii forest.Methods Three treatments including double litter (DL), no litter (NL) and control (CT) were installed in a secondary C. carlesii forest and a human-assisted naturally regenerated C. carlesii forest, to investigate the responses of 6 soil extracellular enzyme activities.Important findings The activities of Cellobiohydrolase (CBH), β-1,4-N-acetylglucosaminidase (NAG), Polyphenol oxidase (PhOx) and Peroxidase (PerOx) in the secondary C. carlesii forest were higher than those in the human-assisted naturally regenerated C. carlesii forest, while acid phosphatase (AP) and β-1,4-glucosidase (βG) activities had no significant difference between the two forests. Compared to control, both NL and DL treatments decreased the activities of AP, βG and NAG, but had no effect on the activities of CBH and PerOx, and DL treatment decreased only the activity of PhOx in two forests. Except for NAG activity, the activities of AP, βG and PerOx decreased more in the human-assisted naturally regenerated C. carlesii forest than in the secondary C. carlesii forest after litter manipulaition. Both Pearson correlation analysis and redundancy analysis showed that soil enzyme activities were significantly correlated with soil moisture content, carbon (C), nitrogen (N), microbial biomass carbon (MBC) and nitrogen (MBN) contents. Therefore, changes in litter input (both increase and decrease) could decrease some major soil enzyme activities such as AP, βG and NAG in both secondary and human-assisted naturally regenerated C. carlesii forests by decreasing soil moisture content, C and N, MBC and MBN contents. Based on the responses of soil enzyme activity, we conclude that the C and N cycling in secondary C. carlesii forest could be faster compared to that in the human-assisted forest of the same species in the subtropical forest ecosystems. Keywords:double litter;no litter;soil enzyme activities;secondary Castanopsis carlesii forest;human-assisted naturally regenerated Castanopsis carlesii forest
PDF (1249KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 魏翠翠, 刘小飞, 林成芳, 李先锋, 李艳, 郑裕雄. 凋落物输入改变对亚热带两种米槠次生林土壤酶活性的影响. 植物生态学报[J], 2018, 42(6): 692-702 doi:10.17521/cjpe.2017.0247 WEI Cui-Cui. Response of soil enzyme activities to litter input changes in two secondary Castanopsis carlessii forests in subtropical China. Chinese Journal of Plant Ecology[J], 2018, 42(6): 692-702 doi:10.17521/cjpe.2017.0247
Table 3 表3 表3两种米槠林添加和去除凋落物处理土壤理化性质和微生物生物量碳、微生物生物量氮含量的变化(平均值±标准偏差, n = 3) Table 3Soil physical, chemical properties and microbial biomass carbon and nitrogen contents in the litter removal, control, and litter addition treatments plots in the two Castanopsis carlesii forests (mean ± SD, n = 3)
同一行小写字母表示同一林分下不同处理间的差异性(p < 0.05), 大写字母表示同一处理不同林分间的差异性(p < 0.05)。 The lowercase letters mean significant differences among treatments in the same forests (p < 0.05), the capital letters mean significant differences between the two forests under the same treatment (p < 0.05).
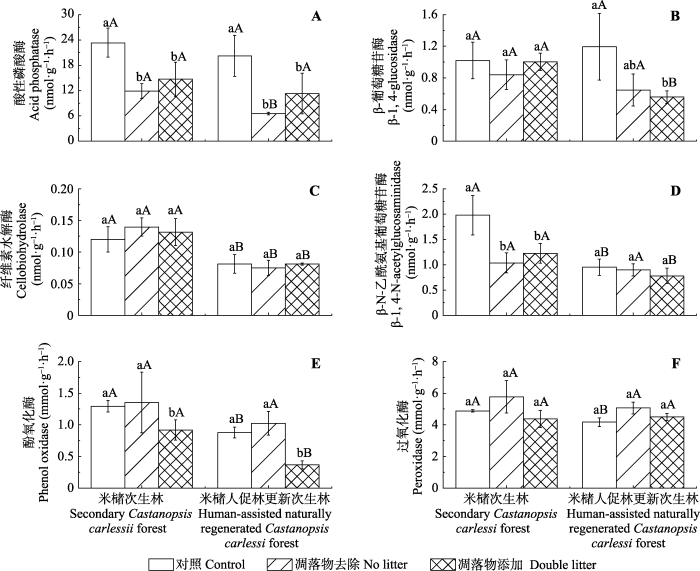
新窗口打开|下载原图ZIP|生成PPT 图1次生林和人促林中凋落物添加和去除后土壤酶活性的变化(平均值±标准偏差, n = 3)。小写字母表示同一林分下不同处理间的差异性(p < 0.05), 大写字母表示不同林分同一处理间的差异性(p < 0.05)。
Fig. 1Soil enzyme activities under litter removal, control, and litter addition treatments in the two Castanopsis carlesii forests (mean ± SD, n = 3). The lowercase letters mean significant differences among treatments in the same forests (p < 0.05), the capital letters mean significant differences between the two forests under the same treatment (p < 0.05).
Fig. 2Redundancy analysis on the relationship of soil enzyme activity and soil physical, chemical properties, microbial biomass carbon, microbial biomass nitrogen.
3 讨论
3.1 凋落物去除和添加对土壤酶活性的影响
本研究中, NL处理降低了土壤AP、βG以及NAG活性, 这与已有凋落物去除降低酶活性的研究结果(Fekete et al., 2011; 郑卫国等, 2011; Weintraub et al., 2013; Veres et al., 2015; 杨洋等, 2016)一致。土壤AP、βG和NAG主要是由微生物产生(杨万勤和杨开运, 2004), 通过这些酶的作用将复杂的有机质水解转化为微生物和植物可利用的P、C、N等小分子物质(Kotroczó et al., 2014)。Xiong等(2008)研究发现随着土壤有机质和全氮含量的下降, 土壤微生物量和微生物活性均显著降低。郑卫国等(2011)研究发现去除凋落物显著降低了土壤微生物数量, 同时降低了土壤微生物的生理活性。本研究中, NL处理显著降低了土壤MBC和MBN的含量, 且相关分析和冗余分析均发现MBC、MBN含量与土壤AP、βG以及NAG活性显著正相关。这表明NL处理可能通过影响微生物生物量, 进而影响土壤AP、βG以及NAG活性。土壤有机质不仅是酶促反应的重要底物, 同时也是微生物活动的主要C源, 去除凋落物减少了土壤有机质的来源, 使微生物可利用的C源减少, 生长受抑制(表现为MBC和MBN的减少), 进而降低了土壤酶活性。微生物活动的变化可以直接引起酶活性的变化, 非生物因素主要通过改变微生物生存环境间接影响土壤酶活性(Kivlin & Treseder, 2014)。本研究中, NL处理降低了土壤含水量, 且土壤水分含量与土壤AP、βG活性显著正相关, 这与Kotroczó等(2014)的研究结果一致, 表明去除凋落物降低土壤酶活性可能与土壤含水量的变化有关。土壤水分含量是限制微生物生物量和活性的重要因素(Rutigliano et al., 2009; Brockett et al., 2012), 杨洋等(2016)认为土壤含水量的降低会通过影响底物和酶的扩散速率影响微生物的变化, 进而降低土壤βG和NAG活性。本研究中去除凋落物使得土壤表层覆盖物减少, 增加了土壤水分的蒸发量, 降低了土壤含水量, 导致微生物生物量发生变化, 从而间接地影响了土壤酶活性。
AllisonSD, VitousekPM ( 2005). Responses of extracellular enzymes to simple and complex nutrient inputs Soil Biology & Biochemistry, 37, 937-944. DOI:10.1016/j.soilbio.2004.09.014URL [本文引用: 2] Soil microbes produce extracellular enzymes that mineralize organic matter and release carbon and nutrients in forms that can be assimilated. Economic theories of microbial metabolism predict that enzyme production should increase when simple nutrients are scarce and complex nutrients are abundant; however, resource limitation could also constrain enzyme production. We tested these hypotheses by monitoring enzyme activities and nutrient pools in soil incubations with added simple and complex nutrient compounds. Over 28 days of incubation, we found that an enzyme's activity increased when its target nutrient was present in complex but not simple form, and carbon and nitrogen were available. β-Glucosidase and acid phosphatase activities also increased in treatments where only carbon and nitrogen were added. Glycine aminopeptidase and acid phosphatase activities declined in response to ammonium and phosphate additions, respectively. In some cases, mineralization responses paralleled changes in enzyme activity—for example, β-glucosidase activity increased and respiration was 5-fold greater in soil incubations with added cellulose, ammonium, and phosphate. However, a doubling of acid phosphatase activity in response to collagen addition was not associated with any changes in phosphorus mineralization. Our results indicate that microbes produce enzymes according to ‘economic rules’, but a substantial pool of mineral stabilized or constitutive enzymes mediates this response. Enzyme allocation patterns reflect microbial nutrient demands and may allow microbes to acquire limiting nutrients from complex substrates available in the soil.
BaldrianP ( 2009). Microbial enzyme-catalyzed processes in soils and their analysis Plant Soil and Environment, 55, 370-378. DOI:10.1007/s11104-009-0098-7URL [本文引用: 1] Currently, measuring enzyme activities in soils or other lignocellulose-based materials is technically feasible; this measurement is particularly suitable for evaluating soil processes of biopolymer (cellulose, hemicelluloses, lignin, chitin and others) degradation by microbes and for assessing cycling and mobilization of principal nutrients including nitrogen, phosphorus and sulfur. With some ...
BlazierMA, HennesseyTC, DengS ( 2005). Effects of fertilization and vegetation control on microbial biomass carbon and dehydrogenase activity in a juvenile loblolly pine plantation Forest Science, 51, 449-459. [本文引用: 1]
BrockettBF, PrescottCE, GraystonSJ ( 2012). Soil moisture is the major factor influencing microbial community structure and enzyme activities across seven biogeoclimatic zones in western Canada Soil Biology & Biochemistry, 44, 9-20. [本文引用: 1]
CaoFQ, LiuZH, LiuM, CuiJF ( 2010). Research progress on the forest litterfall and its decomposition process Guangxi Agricultural Sciences, 41, 693-697. [本文引用: 1]
CaoH, SunH, YangH, SunB, ZhaoQG ( 2003). A review soil enzyme activity and its indication for soil quality Chinese Journal of Applied and Environmental Biology, 9, 105-109. [本文引用: 1]
CernusakLA, WinterK, DallingJW, HoltumJAM, JaramilloC, K?rnerC, LeakeyADW, NorbyRJ, PoulterB, TurnerBL, WrightS ( 2013). Tropical forest responses to increasing atmospheric CO2: Current knowledge and opportunities for future research Functional Plant Biology, 40, 531-551. DOI:10.1071/FP12309URL [本文引用: 1]
ChenSS, ChenSR, MaXQ ( 2001). Effects of different regeneration pattern of secondary broadleaved forest on stand composition and soil fertility Scientia Silvae Sinicae, 37(6), 113-117. DOI:10.11707/j.1001-7488.20010621Magsci [本文引用: 1]
DingSY, SheJY, YangQP, WangSL ( 2015). Effects of thinning and pruning on soil microbial biomass carbon and soilenzyme activities in Chinese fir plantation Journal of Central South University of Forestry & Technology, 35(6), 75-79. DOI:10.14067/j.cnki.1673-923x.2015.06.014URL [本文引用: 2] 土壤微生物量碳和土壤酶活性都是反映土壤养分和土壤生态环境质量的重要指标之一,但间伐、修枝和去除林下植被等经营措施对其影响如何还不太清楚。以湖南省会同县杉木人工林研为究对象,探讨了不同森林经营措施对土壤微生物量碳和酶活性的影响。结果表明,与对照样地相比,间伐、修枝和去除林下植被均显著减小了土壤微生物量碳含量和酶活性,分别为对照样地的79.96%、76.69%和70.70%;土壤微生物量碳和酶活性之间存在极显著的正相关关系。 [ 丁思一, 佘济云, 杨庆朋, 汪思龙 ( 2015). 间伐和修枝对杉木人工林土壤微生物量碳和酶活性的影响 中南林业科技大学学报, 35(6), 75-79.] DOI:10.14067/j.cnki.1673-923x.2015.06.014URL [本文引用: 2] 土壤微生物量碳和土壤酶活性都是反映土壤养分和土壤生态环境质量的重要指标之一,但间伐、修枝和去除林下植被等经营措施对其影响如何还不太清楚。以湖南省会同县杉木人工林研为究对象,探讨了不同森林经营措施对土壤微生物量碳和酶活性的影响。结果表明,与对照样地相比,间伐、修枝和去除林下植被均显著减小了土壤微生物量碳含量和酶活性,分别为对照样地的79.96%、76.69%和70.70%;土壤微生物量碳和酶活性之间存在极显著的正相关关系。
DoughtyCE, MetcalfeDB, GirardinCA, GirardinF, AmezquitaFF, CabreraDG, HuascoWH, Silva-EspejoJE, Araujo-MurakamiA, CostaMC, RochaW, FeldpauschTR, MendozaALM, CostaACL, MeirP, PhillipsOL, MalhiY ( 2015). Drought impact on forest carbon dynamics and fluxes in Amazonia Nature, 519, 78-82. DOI:10.1038/nature14213URLPMID:25739631 [本文引用: 1] Abstract In 2005 and 2010 the Amazon basin experienced two strong droughts, driven by shifts in the tropical hydrological regime possibly associated with global climate change, as predicted by some global models. Tree mortality increased after the 2005 drought, and regional atmospheric inversion modelling showed basin-wide decreases in CO2 uptake in 2010 compared with 2011 (ref. 5). But the response of tropical forest carbon cycling to these droughts is not fully understood and there has been no detailed multi-site investigation in situ. Here we use several years of data from a network of thirteen 1-ha forest plots spread throughout South America, where each component of net primary production (NPP), autotrophic respiration and heterotrophic respiration is measured separately, to develop a better mechanistic understanding of the impact of the 2010 drought on the Amazon forest. We find that total NPP remained constant throughout the drought. However, towards the end of the drought, autotrophic respiration, especially in roots and stems, declined significantly compared with measurements in 2009 made in the absence of drought, with extended decreases in autotrophic respiration in the three driest plots. In the year after the drought, total NPP remained constant but the allocation of carbon shifted towards canopy NPP and away from fine-root NPP. Both leaf-level and plot-level measurements indicate that severe drought suppresses photosynthesis. Scaling these measurements to the entire Amazon basin with rainfall data, we estimate that drought suppressed Amazon-wide photosynthesis in 2010 by 0.38 petagrams of carbon (0.23-0.53 petagrams of carbon). Overall, we find that during this drought, instead of reducing total NPP, trees prioritized growth by reducing autotrophic respiration that was unrelated to growth. This suggests that trees decrease investment in tissue maintenance and defence, in line with eco-evolutionary theories that trees are competitively disadvantaged in the absence of growth. We propose that weakened maintenance and defence investment may, in turn, cause the increase in post-drought tree mortality observed at our plots.
FeketeI, VargaC, KotroczóZ, TóthJA, VárbiróG ( 2011). The relation between various detritus inputs and soil enzyme activities in a Central European deciduous forest Geoderma, 167- 168, 15-21. [本文引用: 1]
GrayDB, MaryKT, JulieEJ ( 2002). Interactions between crop residue and soil organic matter quality and the functional diversity of soil microbial communities Soil Biology & Biochemistry, 34, 1073-1082. [本文引用: 1]
HernandezDL, HobbieSE ( 2010). The effects of substrate composition, quantity, and diversity on microbial activity Plant and Soil, 335, 397-411. DOI:10.1007/s11104-010-0428-9URL [本文引用: 3]
HuangQL, ZhengQR, RongJT, ZhuoMX, GuanXH ( 2012). Selective logging techniques of natural broad-leaved timber forest in the mid-subtropical zone of Fujian, China Journal of Mountain Science, 30(2), 180-185. [本文引用: 1]
KivlinSN, TresederKK ( 2014). Soil extracellular enzyme activities correspond with abiotic factors more than fungal community composition Biogeochemistry, 117, 23-37. DOI:10.1007/s10533-013-9852-2URL [本文引用: 4]
KotroczóZ, VeresZ, FeketeI, KrakompergerK, Tóth JA LajthaK, TóthmérészB ( 2014). Soil enzyme activity in response to long term organic matter manipulation Soil Biology & Biochemistry, 70, 237-243. DOI:10.1016/j.soilbio.2013.12.028URL [本文引用: 4] 61Litter additions did not affect enzyme activities.61Detritus removal significantly reduced enzyme activities.61Root removal significantly reduced enzyme activities.61Enzyme activity are driven primarily by labile carbon.61Aboveground litter is not a significant source of labile carbon to microbes.
LiXJ, LiuXF, XiongDC, LinWS, LinTW, ShiYW ( 2016). Impact of litterfall addition and exclusion on soil respiration in Cunninghamia lanceolata plantation and secondary Castanopsis carlesii forest in mid-subtropical China. Chinese Journal of Plant Ecology, 40, 447-457. DOI:10.17521/cjpe.2015.0404URL [本文引用: 1] 在未来大气CO2浓度升高的背景下,植被净初级生产力的增加将促使森林土壤碳输入增多。凋落物是土壤碳库的重要来源,对土壤呼吸会产生重要影响。为了模拟植物净初级生产力提高、凋落物产量增加情景下凋落物对土壤呼吸和土壤碳库的影响,2013年1月到2014年12月,在福建省三明市陈大镇国有林场,在杉木(Cunninghamia lanceolata)人工林和米槠(Castanopsis carlesii)次生林,通过设置去除凋落物、添加凋落物和对照(保留凋落物,不做任何处理)处理,研究了土壤呼吸和土壤碳库的动态变化。研究发现:土壤含水量在10%–25%范围内,土壤呼吸温度敏感性指数(Q10)随着土壤含水量的增加呈递增趋势,当含水量<10%时,由于干旱胁迫打破了土壤呼吸与温度之间的耦合,改变了Q10值,使得Q10值小于1。土壤呼吸与凋落物输入量呈显著的线性正相关关系,杉木人工林对照和添加凋落物处理及米槠次生林对照处理,土壤呼吸与2个月前的凋落物输入量相关性最好。而米槠次生林添加凋落物处理,土壤呼吸与当月的凋落物输入量相关性最好,不同林分凋落物呼吸对土壤呼吸的贡献率不同,米槠次生林凋落物层呼吸年通量明显大于杉木人工林,分别占各林分土壤总呼吸的34.4%和15.1%,添加凋落物后,杉木人工林和米槠次生林的土壤呼吸速率增加,但添加凋落物处理的土壤呼吸年通量与对照的差值小于年凋落物输入量。因此,在未来全球CO2升高背景下,植被碳储量的增加、凋落物增加并没有引起土壤呼吸成倍增加,更有利于中亚热带地区土壤碳吸存。 [ 李晓杰, 刘小飞, 熊德成, 林伟盛, 林廷武, 施友文 ( 2016). 中亚热带杉木人工林和米槠次生林凋落物添加与去除对土壤呼吸的影响 植物生态学报, 40, 447-457.] DOI:10.17521/cjpe.2015.0404URL [本文引用: 1] 在未来大气CO2浓度升高的背景下,植被净初级生产力的增加将促使森林土壤碳输入增多。凋落物是土壤碳库的重要来源,对土壤呼吸会产生重要影响。为了模拟植物净初级生产力提高、凋落物产量增加情景下凋落物对土壤呼吸和土壤碳库的影响,2013年1月到2014年12月,在福建省三明市陈大镇国有林场,在杉木(Cunninghamia lanceolata)人工林和米槠(Castanopsis carlesii)次生林,通过设置去除凋落物、添加凋落物和对照(保留凋落物,不做任何处理)处理,研究了土壤呼吸和土壤碳库的动态变化。研究发现:土壤含水量在10%–25%范围内,土壤呼吸温度敏感性指数(Q10)随着土壤含水量的增加呈递增趋势,当含水量<10%时,由于干旱胁迫打破了土壤呼吸与温度之间的耦合,改变了Q10值,使得Q10值小于1。土壤呼吸与凋落物输入量呈显著的线性正相关关系,杉木人工林对照和添加凋落物处理及米槠次生林对照处理,土壤呼吸与2个月前的凋落物输入量相关性最好。而米槠次生林添加凋落物处理,土壤呼吸与当月的凋落物输入量相关性最好,不同林分凋落物呼吸对土壤呼吸的贡献率不同,米槠次生林凋落物层呼吸年通量明显大于杉木人工林,分别占各林分土壤总呼吸的34.4%和15.1%,添加凋落物后,杉木人工林和米槠次生林的土壤呼吸速率增加,但添加凋落物处理的土壤呼吸年通量与对照的差值小于年凋落物输入量。因此,在未来全球CO2升高背景下,植被碳储量的增加、凋落物增加并没有引起土壤呼吸成倍增加,更有利于中亚热带地区土壤碳吸存。
LiuXF, LinTC, YangZ, VadeboncoeurMA, LinCF, XiongDC, LinWS, ChenGS, XieJS, LiYQ, YangYS ( 2017). Increased litter in subtropical forests boosts soil respiration in natural forests but not plantations of Castanopsis carlesii. Plant and Soil, 418, 141-148. [本文引用: 1]
LiuX, WangN, ZhaoB, ZhangQ, ZhaoXH ( 2014). Effects of carbon input changes on soil enzyme activities in a Pinus tabulaeformis forest at the Taiyue Mountain. Chinese Journal of Applied and Environmental Biology, 20, 655-661. [本文引用: 2]
OlanderLP, VitousekPM ( 2000). Regulation of soil phosphatase and chitinase activity by N and P availability Biogeochemistry, 49, 175-191. DOI:10.1023/A:1006316117817URL [本文引用: 1] Soil microorganisms and plants produce enzymes that mineralize organically bound nutrients. When nutrient availability is low, the biota may be able to increase production of these enzymes to enhance the supply of inorganic nitrogen (N) and phosphorus (P). Regulation of enzyme production may be a point where N and P cycles interact. We measured acid phosphatase and chitinase (N-acetyl -D-glucosaminide) activity in soil across a chronosequence in Hawaii where N and P availability varies substantially among sites and long term fertilizer plots had been maintained for over 4 years. Phosphatase activity was high at all sites. Chitinase activity decreased significantly as age and N availability increased across the chronosequence. Phosphorus addition suppressed phosphatase activity at all sites, while N addition increased phosphatase activity at the young, N-limited site. In contrast, N addition repressed chitinase activity only at the N limited young site, and P additions had no effect on chitinase activity. These results suggest that the regulatory relationship between nutrient supply and nutrient mineralization are asymmetric for N and P, and that the differences could help to explain differences observed in patterns of N and P availability.
RutiglianoFA, CastaldiS, D’AscoliR, PapaS, CarforaA, MarzaioliR ( 2009). Soil activities related to nitrogen cycle under three plant cover types in Mediterranean environment . Applied Soil Ecology, 43, 40-46. DOI:10.1016/j.apsoil.2009.05.010URL [本文引用: 1]
Saiya-CorkKR, SinsabaughRL, ZakDR ( 2002). The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil. Soil Biology & Biochemistry, 34, 1309-1315. [本文引用: 1]
SayerEJ, PowersJS, TannerEVJ ( 2007). Increased litterfall in tropical forests boosts the transfer of soil CO2 to the atmosphere PLOS ONE, 2, e1299. DOI: 10. 1371/journal. pone.?0001299. DOI:10.1371/journal.pone.0001299URLPMID:18074023 [本文引用: 1] Abstract Aboveground litter production in forests is likely to increase as a consequence of elevated atmospheric carbon dioxide (CO(2)) concentrations, rising temperatures, and shifting rainfall patterns. As litterfall represents a major flux of carbon from vegetation to soil, changes in litter inputs are likely to have wide-reaching consequences for soil carbon dynamics. Such disturbances to the carbon balance may be particularly important in the tropics because tropical forests store almost 30% of the global soil carbon, making them a critical component of the global carbon cycle; nevertheless, the effects of increasing aboveground litter production on belowground carbon dynamics are poorly understood. We used long-term, large-scale monthly litter removal and addition treatments in a lowland tropical forest to assess the consequences of increased litterfall on belowground CO(2) production. Over the second to the fifth year of treatments, litter addition increased soil respiration more than litter removal decreased it; soil respiration was on average 20% lower in the litter removal and 43% higher in the litter addition treatment compared to the controls but litter addition did not change microbial biomass. We predicted a 9% increase in soil respiration in the litter addition plots, based on the 20% decrease in the litter removal plots and an 11% reduction due to lower fine root biomass in the litter addition plots. The 43% measured increase in soil respiration was therefore 34% higher than predicted and it is possible that this 'extra' CO(2) was a result of priming effects, i.e. stimulation of the decomposition of older soil organic matter by the addition of fresh organic matter. Our results show that increases in aboveground litter production as a result of global change have the potential to cause considerable losses of soil carbon to the atmosphere in tropical forests.
ShuYY, HuangJS, ZhaoGJ, BaoWK, PangXY ( 2016). Effects of a forestation with different tree species on soil enzyme activities and nutrient content in eastern Qinghai-?Tibetan Plateau, China Acta Ecologica Sinica, 36, 394-402. DOI:10.5846/stxb201409221877URL [本文引用: 1] 为评价不同树种人工林对土壤酶及养分的影响,选择立地条件和营林方式相同的4种人工林(连香树[CJ]、油松[PT]、落叶松[LK]和华山松[PA])为研究对象,以落叶灌丛(QC)为对照,比较不同树种人工林地土壤酶活性和土壤养分的变化。结果显示:(1)造林降低了土壤酸性磷酸酶、脱氢酶、β-葡萄糖苷酶和过氧化氢酶活性,但人工造林后土壤脲酶活性增加;(2)造林也明显影响了土壤养分,与对照林地相比,除CJ人工林土壤中磷(P)略高外,造林地土壤有机碳(TOC)、氮(N)、水可提取有机碳(WEOC)和氮(WEON)、铵态氮(NH+4-N)和硝态氮(NO-3-N)均降低;(3)不同的人工林树种之间土壤养分及酶活性也存在一定的差异性,CJ和LK人工林土壤C、N、P及相关酶活性明显不同于PT和PA人工林;(4)土壤酶与养分变化有一定的相关性,除转化酶和多酚氧化酶反应较迟钝外,其它酶对环境反应较敏感。综合分析表明,在川西地区选择高密度单一树种造林并没有改善土壤养分和酶活性,在该地区选择落叶或阔叶树种造林可使土壤肥力恢复。 [ 舒媛媛, 黄俊胜, 赵高卷, 包维楷, 李根前, 庞学勇 ( 2016). 青藏高原东缘不同树种人工林对土壤酶活性及养分的影响 生态学报, 36, 394-402. DOI:10.5846/stxb201409221877URL [本文引用: 1] 为评价不同树种人工林对土壤酶及养分的影响,选择立地条件和营林方式相同的4种人工林(连香树[CJ]、油松[PT]、落叶松[LK]和华山松[PA])为研究对象,以落叶灌丛(QC)为对照,比较不同树种人工林地土壤酶活性和土壤养分的变化。结果显示:(1)造林降低了土壤酸性磷酸酶、脱氢酶、β-葡萄糖苷酶和过氧化氢酶活性,但人工造林后土壤脲酶活性增加;(2)造林也明显影响了土壤养分,与对照林地相比,除CJ人工林土壤中磷(P)略高外,造林地土壤有机碳(TOC)、氮(N)、水可提取有机碳(WEOC)和氮(WEON)、铵态氮(NH+4-N)和硝态氮(NO-3-N)均降低;(3)不同的人工林树种之间土壤养分及酶活性也存在一定的差异性,CJ和LK人工林土壤C、N、P及相关酶活性明显不同于PT和PA人工林;(4)土壤酶与养分变化有一定的相关性,除转化酶和多酚氧化酶反应较迟钝外,其它酶对环境反应较敏感。综合分析表明,在川西地区选择高密度单一树种造林并没有改善土壤养分和酶活性,在该地区选择落叶或阔叶树种造林可使土壤肥力恢复。
SinsabaughRL, LauberCL, WeintraubMN, AhmedB, AllisonSD, CrenshawC, ContostaAS, CusackD, FreyS, GalloME, GartnerTB, HobbieSE, HollandK, KeelerBL, PowersJS, StursovaM, Takacs-VesbachC, WaldropMP, WallensteinMD, ZakDR, ZeglinLH ( 2008). Stoichiometry of soil enzyme activity at global scale . Ecology Letters, 11, 1252-1264. DOI:10.1111/j.1461-0248.2008.01245.xURLPMID:18823393 [本文引用: 1] Extracellular enzymes are the proximate agents of organic matter decomposition and measures of these activities can be used as indicators of microbial nutrient demand. We conducted a global-scale meta-analysis of the seven-most widely measured soil enzyme activities, using data from 40 ecosystems. The activities of β-1,4-glucosidase, cellobiohydrolase, β-1,4- N -acetylglucosaminidase and phosphatase g 611 soil increased with organic matter concentration; leucine aminopeptidase, phenol oxidase and peroxidase activities showed no relationship. All activities were significantly related to soil pH. Specific activities, i.e. activity g 611 soil organic matter, also varied in relation to soil pH for all enzymes. Relationships with mean annual temperature (MAT) and precipitation (MAP) were generally weak. For hydrolases, ratios of specific C, N and P acquisition activities converged on 1 : 1 : 1 but across ecosystems, the ratio of C : P acquisition was inversely related to MAP and MAT while the ratio of C : N acquisition increased with MAP. Oxidative activities were more variable than hydrolytic activities and increased with soil pH. Our analyses indicate that the enzymatic potential for hydrolyzing the labile components of soil organic matter is tied to substrate availability, soil pH and the stoichiometry of microbial nutrient demand. The enzymatic potential for oxidizing the recalcitrant fractions of soil organic material, which is a proximate control on soil organic matter accumulation, is most strongly related to soil pH. These trends provide insight into the biogeochemical processes that create global patterns in ecological stoichiometry and organic matter storage.
SongXY, NiuSK, PengB, KeYQ ( 2006). A study on species diversity of different regeneration patterns in Castanopsis carlesii forest in Sanming, Fujian Province. Subtropical Plant Science, 35(2), 31-34. [本文引用: 2]
ThomasH, BenjaminS, IcolinP, KristinaM, PaulM, AlmutA, MartintS ( 2008). CO2 fertilization in temperate FACE experiments not representative of boreal and tropical forests Global Change Biology, 14, 1531-1542. DOI:10.1111/j.1365-2486.2008.01598.xURL [本文引用: 1]
VanceED, BrooksPC, JenkinsonDS ( 1987). An extraction method for measuring soil microbial biomass Soil Biology & Biochemistry, 19, 703-707. DOI:10.1016/0038-0717(87)90052-6URL [本文引用: 1] The effects of fumigation on organic C extractable by 0.5 M K 2SO 4 were examined in a contrasting range of soils. E C (the difference between organic C extracted by 0.5 M K 2SO 4 from fumigated and non-fumigated soil) was about 70% of F C (the flush of CO 2-C caused by fumigation during a 10 day incubation), meaned for ten soils. There was a close relationship between microbial biomass C, measured by fumigation-incubation (from the relationship Biomass C = F C/0.45) and E C given by the equation: Biomass C = (2.64 0.060) E C that accounted for 99.2% of the variance in the data. This relationship held over a wide range of soil pH (3.9 8.0). ATP and microbial biomass N concentrations were measured in four of the soils. The (ATP) (E C) ratios were very similar in the four soils, suggesting that both ATP and the organic C rendered decomposable by CHCl 3 came from the soil microbial biomass. The C:N ratio of the biomass in a strongly acid (pH 4.2) soil was greater (9.4) than in the three less-acid soils (mean C:N ratio 5.1). We propose that the organic C rendered extractable to 0.5 m K 2SO 4 after a 24 h CHCl 3-fumigation ( E C) comes from the cells of the microbial biomass and can be used to estimate soil microbial biomass C in both neutral and acid soils.
VeresZ, KotroczóZ, FeketeI, TóthJA, LajthaK, TownsendK, TóthmérészB ( 2015). Soil extracellular enzyme activities are sensitive indicators of detrital inputs and carbon availability Applied Soil Ecology, 92, 18-23. DOI:10.1016/j.apsoil.2015.03.006URL [本文引用: 4]
WangZH, DuanCQ, HouYP, YangJS ( 2006). The relationship of plant species diversity to ecosystem function in relation to soil conservation in semi-humid evergreen forests, Yunnan Province, China Journal of Plant Ecology (Chinese Version), 30, 392-403. [本文引用: 1]
WeintraubSR, WiederWR, ClevelandCC, TownsendAR ( 2013). Organic matter inputs shift soil enzyme activity and allocation patterns in a wet tropical forest Biogeochemistry, 114, 313-326. DOI:10.1007/s10533-012-9812-2URL [本文引用: 4]
XiongYM, XiaHP, LiZA, CaiXA, FuSL ( 2008). Impacts of litter and understory removal on soil properties in a subtropical Acacia mangium plantation in China. Plant and Soil, 304, 179-188. [本文引用: 2]
YangJT, SuZX, HuJY, WuQG, HeJ ( 2010). Path analysis for soil organic matters and enzyme activities of Davidia involucrate Virgin forest. Chinese Journal of Applied and Environmental Biology, 16, 164-167. [本文引用: 1]
YangWQ, ZhongZC, TaoJP, HeWM ( 2001). Study on relationship between soil enzyme activities and plant species diversity in forest ecosystem of Mt. Jinyun Scientia Silvae Sinicae, 37(4), 124-128. DOI:10.11707/j.1001-7488.20010420Magsci [本文引用: 1]
YangY, WangJF, ZhangXY, LiDD, WangHM, ChenFS, SunXM, WenXF ( 2016). Mechanism of litter and understory vegetation effects on soil carbon and nitrogen hydrolase activities in Chinese fir forests Acta Ecologica Sinica, 36, 8102-8110. [本文引用: 2]
YinP, HuX, WuY ( 2015). Responses of soil enzyme activity on snow cover and litter input in alpine area Journal of Jinan University, 36, 115-118. [本文引用: 1]
YuanXC, LinWS, PuXT, YangZR, ZhengW, ChenYM ( 2016). Effects of forest regeneration patterns on the quantity and chemical structure of soil solution dissolved organic matter in a subtropical forest Chinese Journal of Applied Ecology, 27, 1845-1852. [本文引用: 1]
ZhaoJ ( 2016). Effects of Nitrogen Addition and Litter on Soil Microorganism and Enzyme Activities. Master degree dissertation, Beijing Forestry University Beijing. URL [本文引用: 1] 随着全球气候变化日益加剧,一些人为或者自然的扰动会导致森林地表凋落物和土壤性状发生变化,进而影响到由微生物介导的土壤有机碳周转和养分循环。然而,由于缺少全面、系统性的实验探索,目前在全球变化研究中对凋落物变化和外源氮输入双重影响下的森林土壤微生物过程和功能的变异性仍认识不足。本研究于2014年4月至2015年8月期间,基于山西省太岳山灵空山林场的凋落物处理和氮素添加双因子交互作用实验样地,通过样品采集和土壤性状化验分析,探究了森林土壤微生物和相关土壤酶对不同凋落物输入和氮添加的响应形式、过程和机理。实验样地于2010年9月建立,位于一个典型油松-辽东栎混交林中;样地布设采用随机区组设计,实验处理包含4种凋落物情形(剔除凋落物、混合凋落物加倍、叶凋落物加倍和枝果凋落物加倍)和3个梯度氮添加处理(不施氮、5g·N·m-2·a-1施氮率和10g·N·m-2·a-1施氮率)。主要研究结果概要总结如下:(1)与剔除凋落物、枝果凋落物加倍处理相比,叶凋落物加倍与混合凋落物加倍处理极显著提高了土壤有机碳、全氮、C:N、铵态氮、总无机氮含量,微生物生物量及其活性,以及p-葡萄糖苷酶、N-乙酰-β-葡萄糖苷酶、酚氧化酶、过氧化物酶活性,同时显著提高细菌总量及革兰氏阴性细菌的相对含量。(2) 5gN·m-2·a-1施氮率处理极显著提高了有机碳含量、革兰氏阴性细菌的相对含量以及脲酶活性,而土壤硝态氮、总无机氮含量则随着施氮率升高而上升。不同梯度氮添加处理对微生物量及活性影响并不显著。(3)不同梯度氮添加及凋落物处理对微生物及酶活性交互影响并不显著。仅在2014年10月,N-乙酰-β-葡萄糖苷酶活性在5gN·m2·a-1施氮率处理配合叶凋落物加倍、混合凋落物加倍处理时显著高于其他处理。综上,混合凋落物加倍、叶凋落物加倍处理显著促进了土壤微生物群落的生长、增加了酶活性,相比之下不同氮添加处理以及不同氮添加与凋落物处理之间的交互作用并不显著。 [ 赵静 ( 2016). 氮添加与凋落物对土壤微生物和酶活性的影响 硕士学位论文, 北京林业大学. 北京.] URL [本文引用: 1] 随着全球气候变化日益加剧,一些人为或者自然的扰动会导致森林地表凋落物和土壤性状发生变化,进而影响到由微生物介导的土壤有机碳周转和养分循环。然而,由于缺少全面、系统性的实验探索,目前在全球变化研究中对凋落物变化和外源氮输入双重影响下的森林土壤微生物过程和功能的变异性仍认识不足。本研究于2014年4月至2015年8月期间,基于山西省太岳山灵空山林场的凋落物处理和氮素添加双因子交互作用实验样地,通过样品采集和土壤性状化验分析,探究了森林土壤微生物和相关土壤酶对不同凋落物输入和氮添加的响应形式、过程和机理。实验样地于2010年9月建立,位于一个典型油松-辽东栎混交林中;样地布设采用随机区组设计,实验处理包含4种凋落物情形(剔除凋落物、混合凋落物加倍、叶凋落物加倍和枝果凋落物加倍)和3个梯度氮添加处理(不施氮、5g·N·m-2·a-1施氮率和10g·N·m-2·a-1施氮率)。主要研究结果概要总结如下:(1)与剔除凋落物、枝果凋落物加倍处理相比,叶凋落物加倍与混合凋落物加倍处理极显著提高了土壤有机碳、全氮、C:N、铵态氮、总无机氮含量,微生物生物量及其活性,以及p-葡萄糖苷酶、N-乙酰-β-葡萄糖苷酶、酚氧化酶、过氧化物酶活性,同时显著提高细菌总量及革兰氏阴性细菌的相对含量。(2) 5gN·m-2·a-1施氮率处理极显著提高了有机碳含量、革兰氏阴性细菌的相对含量以及脲酶活性,而土壤硝态氮、总无机氮含量则随着施氮率升高而上升。不同梯度氮添加处理对微生物量及活性影响并不显著。(3)不同梯度氮添加及凋落物处理对微生物及酶活性交互影响并不显著。仅在2014年10月,N-乙酰-β-葡萄糖苷酶活性在5gN·m2·a-1施氮率处理配合叶凋落物加倍、混合凋落物加倍处理时显著高于其他处理。综上,混合凋落物加倍、叶凋落物加倍处理显著促进了土壤微生物群落的生长、增加了酶活性,相比之下不同氮添加处理以及不同氮添加与凋落物处理之间的交互作用并不显著。
ZhengWG, XueL, XuPB, LiangLL, FengHF ( 2011). Soil response to litter in a Pinus caribaea woodland. Journal of South China Agricultural University, 32(1), 21-25. [本文引用: 2]
ZhuTB, ZhugeYP, LiuSJ, LouYH ( 2008). Effects of different fertilizer treatments and water regimes on soil enzyme activities Shandong Agricultural Sciences,( 3), 74-78. [本文引用: 1]
Soil moisture is the major factor influencing microbial community structure and enzyme activities across seven biogeoclimatic zones in western Canada 1 2012
... 本研究中, NL处理降低了土壤AP、βG以及NAG活性, 这与已有凋落物去除降低酶活性的研究结果(Fekete et al., 2011; 郑卫国等, 2011; Weintraub et al., 2013; Veres et al., 2015; 杨洋等, 2016)一致.土壤AP、βG和NAG主要是由微生物产生(杨万勤和杨开运, 2004), 通过这些酶的作用将复杂的有机质水解转化为微生物和植物可利用的P、C、N等小分子物质(Kotroczó et al., 2014).Xiong等(2008)研究发现随着土壤有机质和全氮含量的下降, 土壤微生物量和微生物活性均显著降低.郑卫国等(2011)研究发现去除凋落物显著降低了土壤微生物数量, 同时降低了土壤微生物的生理活性.本研究中, NL处理显著降低了土壤MBC和MBN的含量, 且相关分析和冗余分析均发现MBC、MBN含量与土壤AP、βG以及NAG活性显著正相关.这表明NL处理可能通过影响微生物生物量, 进而影响土壤AP、βG以及NAG活性.土壤有机质不仅是酶促反应的重要底物, 同时也是微生物活动的主要C源, 去除凋落物减少了土壤有机质的来源, 使微生物可利用的C源减少, 生长受抑制(表现为MBC和MBN的减少), 进而降低了土壤酶活性.微生物活动的变化可以直接引起酶活性的变化, 非生物因素主要通过改变微生物生存环境间接影响土壤酶活性(Kivlin & Treseder, 2014).本研究中, NL处理降低了土壤含水量, 且土壤水分含量与土壤AP、βG活性显著正相关, 这与Kotroczó等(2014)的研究结果一致, 表明去除凋落物降低土壤酶活性可能与土壤含水量的变化有关.土壤水分含量是限制微生物生物量和活性的重要因素(Rutigliano et al., 2009; Brockett et al., 2012), 杨洋等(2016)认为土壤含水量的降低会通过影响底物和酶的扩散速率影响微生物的变化, 进而降低土壤βG和NAG活性.本研究中去除凋落物使得土壤表层覆盖物减少, 增加了土壤水分的蒸发量, 降低了土壤含水量, 导致微生物生物量发生变化, 从而间接地影响了土壤酶活性. ...
森林凋落物及其分解过程的研究进展 1 2010
... 森林凋落物是土壤有机质的重要来源, 是森林生态系统能量流动和物质循环的重要载体(曹富强等, 2010), 可为植物和微生物生长提供所需养分.研究表明大气CO2浓度增加和温度升高将导致植物净初级生产力增加, 提高凋落物生产量(Thomas et al., 2008; Cernusak et al., 2013), 而极端干旱事件可能会降低植物的生产力(Doughty et al., 2015).植物生产力发生变化将引起输入土壤中的凋落物数量和质量改变, 从而使土壤微生物生物量、群落组成乃至代谢方式发生变化(Gray et al., 2002), 影响土壤酶活性.但现有研究表明凋落物增加和减少对土壤酶活性的影响并不一致(Weintraub et al., 2013; Kotroczó et al., 2014; 刘星等, 2014; Veres et al., 2015).在温带森林生态系统, 凋落物添加后土壤酶活性未发生显著变化, 而凋落物去除后土壤酶活性显著降低(Kotroczó et al., 2014; 刘星等, 2014; Veres et al., 2015).尹鹏等(2015)在高原土壤生态系统凋落物添加试验中发现土壤蔗糖酶显著降低, 而Weintraub等(2013)在热带雨林添加凋落物增加了土壤酶活性, 去除凋落物降低了土壤酶活性.有研究指出, 土壤酶活性与土壤温度, 土壤含水量, 土壤C、N等含量有关, 且相关程度因生态系统而异(Kivlin & Treseder, 2014).碳输入改变引起土壤物理、化学、生物性质发生变化, 进而导致酶活性发生变化(Weintraub et al., 2013).亚热带作为温带与热带的过渡带, 其温度、降水量等与温带、热带有较大差异, 森林地表凋落物输入改变(增加或减少)对土壤酶活性的影响还是未知. ...
森林凋落物及其分解过程的研究进展 1 2010
... 森林凋落物是土壤有机质的重要来源, 是森林生态系统能量流动和物质循环的重要载体(曹富强等, 2010), 可为植物和微生物生长提供所需养分.研究表明大气CO2浓度增加和温度升高将导致植物净初级生产力增加, 提高凋落物生产量(Thomas et al., 2008; Cernusak et al., 2013), 而极端干旱事件可能会降低植物的生产力(Doughty et al., 2015).植物生产力发生变化将引起输入土壤中的凋落物数量和质量改变, 从而使土壤微生物生物量、群落组成乃至代谢方式发生变化(Gray et al., 2002), 影响土壤酶活性.但现有研究表明凋落物增加和减少对土壤酶活性的影响并不一致(Weintraub et al., 2013; Kotroczó et al., 2014; 刘星等, 2014; Veres et al., 2015).在温带森林生态系统, 凋落物添加后土壤酶活性未发生显著变化, 而凋落物去除后土壤酶活性显著降低(Kotroczó et al., 2014; 刘星等, 2014; Veres et al., 2015).尹鹏等(2015)在高原土壤生态系统凋落物添加试验中发现土壤蔗糖酶显著降低, 而Weintraub等(2013)在热带雨林添加凋落物增加了土壤酶活性, 去除凋落物降低了土壤酶活性.有研究指出, 土壤酶活性与土壤温度, 土壤含水量, 土壤C、N等含量有关, 且相关程度因生态系统而异(Kivlin & Treseder, 2014).碳输入改变引起土壤物理、化学、生物性质发生变化, 进而导致酶活性发生变化(Weintraub et al., 2013).亚热带作为温带与热带的过渡带, 其温度、降水量等与温带、热带有较大差异, 森林地表凋落物输入改变(增加或减少)对土壤酶活性的影响还是未知. ...
 ,*, 李先锋*, 李艳*, 郑裕雄*福建师范大学地理科学学院, 福州 350007; 湿润亚热带山地生态国家重点实验室培育基地, 福州 350007; 福建三明森林生态系统与全球变化研究站, 福建三明 365000
,*, 李先锋*, 李艳*, 郑裕雄*福建师范大学地理科学学院, 福州 350007; 湿润亚热带山地生态国家重点实验室培育基地, 福州 350007; 福建三明森林生态系统与全球变化研究站, 福建三明 365000
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT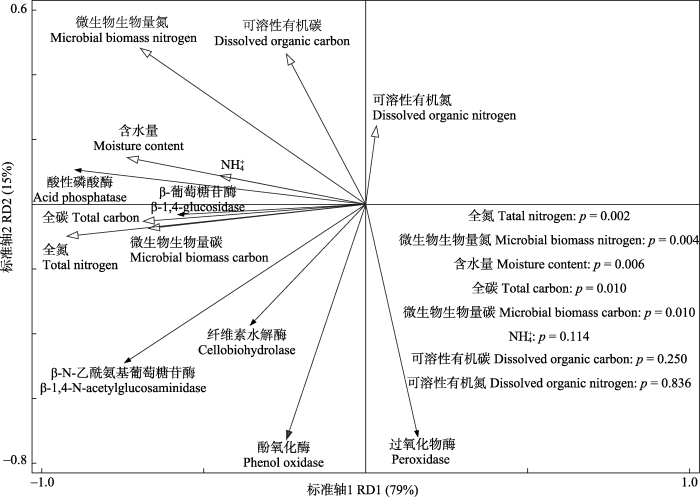 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}