 ,*, 石明, 杨聪, 代俊西南林业大学林学院, 昆明 650224
,*, 石明, 杨聪, 代俊西南林业大学林学院, 昆明 650224Community characteristics and population structure of Ferrocalamus strictus, a rare species in Mojiang, Yunnan, China
YU Ding-Xiang, DU Fan,*, SHI Ming, YANG Cong, DAI JunCollege of Forestry, Southwest Forestry University, Kunming 650224, China通讯作者: kmdufan@163.com
编委: 李 睿
责任编辑: 李 敏
收稿日期:2018-03-07修回日期:2018-09-10网络出版日期:2018-09-20
Received:2018-03-07Revised:2018-09-10Online:2018-09-20

摘要
铁竹(Ferrocalamus strictus)分布在云南南部局部区域, 其数量少, 分布范围狭窄, 为云南省极小种群物种。对铁竹野生种群结构及其所在群落特征进行研究可以为了解其濒危机制提供科学依据, 也可为其保护提供前期研究基础。该研究通过对墨江铁竹野生资源的调查, 从物种组成、种群密度、种群存活曲线和死亡因素等方面, 分析铁竹的种群结构及其所在群落特征。结果表明: (1)墨江铁竹种群所在群落的外貌、物种组成均具有山地雨林的特征; (2)墨江铁竹种群密度为2.04株·m -2; 铁竹种群的存活曲线介于Deevey-I型和Deevey-II型之间; 铁竹种群的净增殖率(R0)为1.10, 表明铁竹种群处于增长阶段; (3)导致铁竹死亡的原因有人为砍伐、自然枯立、退笋及昆虫啃食, 其中人为砍伐所占比例最大; (4)铁竹为南亚热带森林竹种, 属中型竹类, 节间长度从基部3-4节起突然伸长, 最长超过1 m, 在竹类植物中居于前列, 这与其适应热带山地雨林环境密切相关。
关键词:
Abstract
Keywords:
PDF (1558KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
喻丁香, 杜凡, 石明, 杨聪, 代俊. 云南墨江极小种群物种铁竹的种群结构与群落特征. 植物生态学报[J], 2018, 42(9): 938-945 DOI:10.17521/cjpe.2018.0055
YU Ding-Xiang, DU Fan, SHI Ming, YANG Cong, DAI Jun.
种群是构成群落的基本单位, 了解其结构不仅有助于了解种群数量动态及发展趋势, 也是群落及生态系统研究的基础; 而群落以种群为基础, 对种群的数量、空间结构和分布又产生实质性影响(康华靖等, 2007; 黄祥童等, 2018)。由于环境变化和生境消失, 使许多物种的种群数量低于稳定存活界限的最小生存界线, 濒临灭绝危险, 需要优先保护(孙卫邦和韩春艳, 2015; 黄祥童等, 2018)。极小种群是近年来保护生物学领域提出的新概念, 在“分布地域狭窄”、“受到外界因素胁迫干扰”和“种群及个体数量少”等物种选列指标上有具体量化, 突出了野生植物保护的重点和保护的本质, 从种群的角度提出更注重实际的保护措施(杨文忠等, 2015)。分析极小种群野生植物的种群结构及群落特征, 有助于评估其生存状况和预测未来的发展动态(黄祥童等, 2018), 对揭示其长期生存潜力和种群维持机制有重要意义(Hedrick, 2005; Aguilar et al., 2006)。
铁竹(Ferrocalamus strictus)系禾本科铁竹属(Ferrocalamus)中型散生竹类; 节间超过1 m, 是节间最长的竹类植物; 其笋质硬、味苦, 不堪食用; 秆极坚硬, 是制作弩、竹筷和毛线编织针棒的上乘材料。铁竹属由竹类学家耿伯介和薛纪如于1982年建立(耿伯介和薛纪如, 1982), 建立时仅含模式种铁竹, 零星分布于云南南部金平和绿春, 近年来又发现于纬度更偏北的墨江哈尼族自治县(李恩良等, 2015)。据最新资料, 铁竹属含3种, 其中铁竹和裂箨铁竹(F. rimosivaginus)分布于云南南部(温太辉, 1984), 另一种F. fibrillosus分布于越南(Nguyen et al., 2012)。模式种铁竹分布区狭窄, 个体数量少, 2009年被云南省林业厅和云南省科学技术厅列为云南省极小种群物种。目前有关铁竹的研究除新种的描述外, 未见其他报道。本研究通过对铁竹种群所在群落的物种组成、种群密度、种群存活曲线、死亡因素等进行分析, 以期阐明其濒危原因, 为铁竹的保护提供理论依据。
1 研究区概况
墨江哈尼族自治县地处云南南部哀牢山中段主脉以西, 介于101.13°-102.07° E, 22.85°-23.98° N之间, 地势自西北向东南倾斜, 地貌类型复杂多样, 属南亚热带半湿润山地季风气候。县城(海拔约1 315 m)年平均气温17.8 ℃, 最冷月平均气温11.5 ℃, 极端最低温-4 ℃, 无霜期306天; 全年总日照时间2 161.2 h; 年降水量1 338 mm, 干湿季分明, 5-10月为雨季, 11月至次年4月为干季; 平均年蒸发量1 696.7 mm, 相对湿度80%。研究区位于县城北边的杨仕岭(101.72° E, 23.46° N), 海拔1 500-1 690 m, 该区为墨江县城周边重要的水源林, 2000年被划为省级公益林, 其地带性植被属于季风常绿阔叶林, 以单果柯(Lithocarpus pseudoreinwardtii)、多穗柯(L. polystachyus)、耳叶柯(L. grandifolius)等壳斗科乔木为优势种, 森林植被保存完好。该区有3小片铁竹分布点, 面积分别约6 580 m2、430 m2和260 m2; 海拔1 510-1 530 m, 地形为沟箐, 坡度20°-40°, 地表湿润, 枯枝落叶层厚8-12 cm, 腐质层厚6-15 cm, 土壤为红壤, 微酸性, 厚度45-65 cm (李恩良等, 2015)。2 研究方法
2.1 野外调查
2017年6月和9月进行野外调查, 共计15天。铁竹分布零散, 把3小片分布区视为3个样地, 各设置1个15 m × 15 m的样方用于铁竹种群所在群落的结构调查, 详细记录样方的坡度、坡向、坡位、海拔、受干扰程度; 记录构成群落乔木层、灌木层、草本层、层间植物的名称、盖度、高度、胸径、物候和生活力等。另在铁竹的3个样地内设置16个面积为5 m × 5 m的小样方用于铁竹种群结构调查, 记录铁竹的株数、年龄结构、发笋数、死亡株数、死亡原因等, 并推算样地内铁竹的总株数。
在样地内选取5株标准竹(本研究标准竹是指符合样地内平均胸径的铁竹), 自秆基处截断, 调查内容包括胸径(本研究的胸径是指铁竹高1.3 m处的节节间中部的粗度)、节间长度、竹节数(从铁竹竹兜以上开始计数, 将竹节依次编号为1、2、3、4……称为节位数)等。
2.2 数据处理
2.2.1 铁竹秆龄划分参照前人对竹子年龄结构的划分方法(秦自生等, 1993; 周芳纯, 1998; 辉朝茂和杨宇明, 2002), 结合铁竹的特征, 以竹秆表面颜色、秆箨保留情况、分枝发生与生长、叶片颜色及叶量、梢头状态、附生物等指标, 将铁竹分为1年生(竹笋-幼竹)、2年生(翌年)、3年生、4年生、5年生、> 5年生共计6个龄级。
2.2.2 铁竹种群密度
铁竹种群密度: Nd = S/A (1)
式中, Nd为种群密度, S为样地内铁竹的总株数, A为样地面积。
2.2.3 铁竹种群静态生命表、存活曲线和生殖力表的编制
统计铁竹各秆龄株数, 以其为基础, 编制铁竹种群静态生命表, 实际分析中, 存在死亡率为负数的情况, 故采用匀滑技术对数据进行处理(江洪, 1992); >5年生的铁竹不能区分到具体的秆龄, 故未作分析。根据铁竹种群静态生命表, 以秆龄为横坐标, 以各秆龄内标准化存活数为纵坐标, 绘制铁竹种群的存活曲线。计算公式如下:
nx = (ax/a0) × 1000 (2)
dx = nx - nx + 1 (3)
qx = (dx/nx) × 100% (4)
Lx = (nx + nx+ 1)/2 (5)
Tx= Lx + Lx+ 1+Lx+ 2 … + Lx+I(6)
ex = Tx/nx (7)
式中, x为秆龄; ax为x秆龄内实际存活数; nx为x年龄开始时标准化的存活数(一般校正为1 000); dx为x到x + 1秆龄间隔期内标准化死亡数; qx为x年龄期死亡数dx与该期开始个数nx的比值; Lx为从x到x + 1年龄期间还存活的个体数; Tx为x年龄至超过x年龄的个体总数; ex为进入x年龄个体的生命期望。
依据静态生命表进一步编制生殖力表: 分年龄组计算lx、bx、lxbx、xlxbx的值, 再按以下公式计算种群净增殖率(R0)、种群内禀增殖率(rm)、种群世代平均周期(T)(本研究指铁竹种群发笋平均周期)和种群加倍时间(t) (江洪, 1992)。
${{R}_{0}}=\sum{{{l}_{x}}{{b}_{x}}}$ (8)
rm = lnR0/T (9)
$T={\sum{x{{l}_{x}}{{b}_{x}}}}/{\sum{{{l}_{x}}{{b}_{x}}}}\;$ (10)
t = ln2/rm (11)
式中, lx为在x秆龄时的存活率; bx为在x秆龄所有植株个体的发笋总数除以该秆龄中所有植株个体总数的平均值。
3 结果和分析
3.1 物种组成
墨江铁竹片段化生长在杨仕岭局部生境湿润的沟箐边。群落分为4层, 即乔木层、灌木层、草本层、层间层(表1)。Table 1
表1
表1铁竹种群所在群落的物种组成
Table 1
| 物种 Species | 相对优势度 Relative dominance | 相对多度 Relative abundance | 相对频度 Relative frequency | 重要值 Important value | 物种 Species | 相对优势度 Relative dominance | 相对多度 Relative abundance | 相对频度 Relative frequency | 重要值 Important value |
|---|---|---|---|---|---|---|---|---|---|
| 铁竹 Ferrocalamus strictus | 48.41 | 98.27 | 2.78 | 49.82 | 尖子木 Oxyspora paniculata | 0.30 | 0.01 | 1.85 | 0.72 |
| 绒毛肉实树 Sarcosperma kachinense | 11.53 | 0.09 | 2.78 | 22.19 | 毛狗骨柴 Diplospora fruticosa | 0.30 | 0.01 | 1.85 | 0.72 |
| 西南木荷 Schima wallichii | 6.45 | 0.05 | 2.78 | 3.80 | 云南瓦理棕 Wallichia mooreana | 0.46 | 0.02 | 0.93 | 0.47 |
| 胡椒 Piper nigrum | 6.92 | 0.23 | 2.78 | 3.31 | 露兜树 Pandanus tectorius | 0.46 | 0.01 | 0.93 | 0.47 |
| 黄花胡椒 Piper flaviflorum | 4.38 | 0.15 | 2.78 | 2.44 | 云南斑籽木 Baliospermum calycinum | 0.46 | 0.01 | 0.93 | 0.47 |
| 岭罗麦 Tarennoidea wallichii | 0.92 | 0.02 | 2.78 | 2.33 | 山姜 Alpinia japonica | 0.46 | 0.01 | 0.93 | 0.47 |
| 尼泊尔水东哥 Saurauia napaulensis | 1.84 | 0.01 | 0.93 | 2.23 | 云南叶轮木 Ostodes katharinae | 0.23 | 0.01 | 0.93 | 0.39 |
| 桂木 Artocarpus nitidus subsp. lingnanensis | 1.15 | 0.02 | 2.78 | 2.16 | 三对节 Clerodendrum serratum | 0.23 | 0.01 | 0.93 | 0.39 |
| 纸叶琼楠 Beilschmiedia pergamentacea | 0.69 | 0.02 | 2.78 | 2.00 | 亮叶龙船花 Ixora fulgens | 0.23 | 0.01 | 0.93 | 0.39 |
| 钝叶桂 Cinnamomum bejolghota | 0.69 | 0.02 | 2.78 | 1.72 | 玉兰叶木姜子 Litsea magnoliifolia | 0.23 | 0.01 | 0.93 | 0.39 |
| 罗伞 Brassaiopsis glomerulata | 0.69 | 0.02 | 2.78 | 1.59 | 楤木 Aralia chinensis | 0.23 | 0.01 | 0.93 | 0.39 |
| 云桂暗罗 Polyalthia petelotii | 0.69 | 0.02 | 2.78 | 1.48 | 大果榕 Ficus auriculata | 0.23 | 0.01 | 0.93 | 0.39 |
| 地不容 Stephania epigaea | 0.92 | 0.05 | 2.78 | 1.25 | 散生女贞 Ligustrum confusum | 0.23 | 0.01 | 0.93 | 0.39 |
| 潺槁木姜子 Litsea glutinosa | 0.69 | 0.02 | 2.78 | 1.24 | 西南山茶 Camellia pitardii | 0.23 | 0.01 | 0.93 | 0.39 |
| 四角蒲桃 Syzygium tetragonum | 0.69 | 0.01 | 1.85 | 1.24 | 滇南赤车 Pellionia paucidentata | 0.23 | 0.01 | 0.93 | 0.39 |
| 石韦 Pyrrosia lingua | 0.69 | 0.07 | 2.78 | 1.18 | 大叶仙茅 Curculigo capitulata | 0.23 | 0.01 | 0.93 | 0.39 |
| 石柑子 Pothos chinensis | 0.69 | 0.06 | 2.78 | 1.18 | 金毛狗蕨 Cibotium barometz | 0.23 | 0.01 | 0.93 | 0.39 |
| 蛇根草 Ophiorrhiza mungos | 0.69 | 0.05 | 2.78 | 1.17 | 鸭跖草 Commelina communis | 0.23 | 0.01 | 0.93 | 0.39 |
| 紫金牛 Ardisia japonica | 0.69 | 0.03 | 2.78 | 1.17 | 花蘑芋 Amorphophallus konjac | 0.23 | 0.01 | 0.93 | 0.39 |
| 三叶崖爬藤 Tetrastigma hemsleyanum | 0.53 | 0.04 | 2.78 | 1.12 | 爬树龙 Rhaphidophora decursiva | 0.18 | 0.02 | 0.93 | 0.38 |
| 铁角蕨 Asplenium trichomanes | 0.41 | 0.15 | 2.78 | 1.11 | 巴豆藤 Craspedolobium schochii | 0.12 | 0.01 | 0.93 | 0.35 |
| 匍匐堇菜 Viola pilosa | 0.53 | 0.03 | 2.78 | 1.11 | 密花豆 Spatholobus suberectus | 0.05 | 0.01 | 0.93 | 0.33 |
| 莎草一种 Cyperus sp. | 0.30 | 0.04 | 2.78 | 1.04 | 桂叶素馨 Jasminum laurifolium | 0.02 | 0.03 | 0.93 | 0.33 |
| 肉实树 Sarcosperma laurinum | 0.69 | 0.01 | 0.93 | 1.04 | 带叶卷瓣兰 Bulbophyllum taeniophyllum | 0.02 | 0.03 | 0.93 | 0.33 |
| 瓦韦 Lepisorus thunbergianus | 0.46 | 0.11 | 1.85 | 0.81 | 水晶兰 Monotropa uniflora | 0.02 | 0.01 | 0.93 | 0.32 |
| 乌毛蕨 Blechnum orientale | 0.46 | 0.03 | 1.85 | 0.78 | 毛柱马钱 Strychnos nitida | 0.02 | 0.01 | 0.93 | 0.32 |
| 小绿刺 Capparis urophylla | 0.46 | 0.01 | 1.85 | 0.78 | 买麻藤 Gnetum montanum | 0.02 | 0.01 | 0.93 | 0.32 |
| 九节 Psychotria asiatica | 0.46 | 0.01 | 1.85 | 0.78 | 薯蓣 Dioscorea polystachya | 0.02 | 0.01 | 0.93 | 0.32 |
| 三桠苦 Evodia lepta | 0.30 | 0.06 | 1.85 | 0.74 | 独子藤 Celastrus monospermus | 0.02 | 0.01 | 0.93 | 0.32 |
新窗口打开|下载CSV
乔木层盖度30%-40%, 高15-23 m, 共12种, 如绒毛肉实树(Sarcosperma kachinense)、钝叶桂(Cinnamomum bejolghota)、云桂暗罗(Polyalthia petelotii)等。
灌木层盖度约90%。以铁竹为优势, 盖度50%-80%, 高5-10 m; 其他还有云南瓦理棕(Wallichia mooreana)、尖子木(Oxyspora paniculata)、云南叶轮木(Ostodes katharinae)、云南斑籽木(Baliospermum calycinum)、露兜树(Pandanus tectorius)等散布其中, 高0.5-4.5 m。
由于灌木层盖度高, 草本层发育较弱、疏散, 盖度约5%, 高0.1-1.2 m。但是种类丰富, 共计14种, 如金毛狗蕨(Cibotium barometz)、大叶仙茅(Curculigo capitulate)、山姜(Alpinia japonica)、黄花胡椒(Piper flaviflorum)等。
层间植物丰富, 如巴豆藤(Craspedolobium unijugum)、爬树龙(Rhaphidophora decursiva)、买麻藤(Gnetum montanum)等; 此外还有石柑子(Pothos chinensis)、铁角蕨(Asplenium trichomanes)等附生植物; 林下有腐生植物水晶兰(Monotropa uniflora)出现, 表明铁竹生境高度湿润。
从外貌看, 铁竹种群所在群落的林层较高, 层次结构不明显, 层间植物发达。组成物种则包括典型山地雨林成分, 如绒毛肉实树、云桂暗罗、云南瓦理棕、露兜树、金毛狗蕨、爬树龙等。虽然杨仕岭的地带性植被属于季风常绿阔叶林, 但铁竹生长的小环境为更加湿润的静风沟箐, 有较多山地雨林的物种出现, 因此其群落片段所属的植被类型具有山地雨林的特征。
3.2 铁竹种群数量
3.2.1 铁竹种群密度种群密度是影响种群数量动态的主要因素, 通常被认为是种群自身数量调节的作用系统(秦自生等, 1993)。杨仕岭铁竹的分布面积很小, 3个分布点的总面积约7 270 m2。根据样方调查估算, 研究区铁竹数量约14 870株, 种群密度为2.04株·m-2 (表2)。其中样地1约14 400株, 其面积约6 580 m2, 种群密度为2.20株·m-2; 样地2约350株, 其面积约430 m2, 种群密度为0.81株·m-2; 样地3约120株, 其面积约260 m2, 种群密度为0.46株·m-2。
Table 2
表2
表2铁竹种群密度
Table 2
| 样地 Plot | 总数 Total (ind.) | 面积 Area (m2) | 种群密度 Population density (ind.·m-2) |
|---|---|---|---|
| 1 | 14 400 | 6 580 | 2.20 |
| 2 | 350 | 430 | 0.81 |
| 3 | 120 | 260 | 0.46 |
| Total | 14 870 | 7 270 | 2.04 |
新窗口打开|下载CSV
3.2.2 铁竹种群静态生命表和存活曲线
生命表分析结果(表3)显示, 随着铁竹种群年龄的增加, 存活数逐渐减少, 个体的生命期望值呈递减趋势。由存活曲线(图1)可以看出, 铁竹种群的存活曲线介于Deevey-I型和Deevey-II型之间, 死亡数先减少后增加再减少, 且3-5年生的死亡率分别大于1年生和2年生的死亡率, 以3年生最大。表明铁竹种群从竹笋-幼竹进入到2-3年生的比例相对较大,但是从3年生进入到4-5年生的机会比较少。出现此种现象可能与铁竹的死亡原因有关。
Table 3
表3
表3铁竹种群静态生命表
Table 3
| x | ax | nx | dx | qx | Lx | Tx | ex |
|---|---|---|---|---|---|---|---|
| 1 | 84 | 1 000 | 143 | 14.30 | 928.50 | 3 040.50 | 3.04 |
| 2 | 72 | 857 | 36 | 4.20 | 839.00 | 2 112.00 | 2.46 |
| 3 | 69 | 821 | 214 | 26.07 | 714.00 | 1 273.00 | 1.55 |
| 4 | 51 | 607 | 96 | 15.82 | 559.00 | 559.00 | 0.92 |
| 5 | 43 | 511 |
新窗口打开|下载CSV
图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1铁竹种群的存活曲线。
Fig. 1Survivorship curve of the Ferrocalamus strictus population.
3.2.3 铁竹种群生殖力生殖力表(表4)分析结果显示, 铁竹种群的净增殖率(R0)为1.10, R0 > 1, 表明铁竹种群为增长型(按发笋率计算)。内禀增殖率(rm)为0.04, 瞬时出生率大于死亡率, 说明铁竹种群在理想的状态下, 其种群数量具有增长的潜能。铁竹的平均发笋周期(T)大致为2.71 a, 种群加倍时间(t)理论上是19.70 a, 该值主要是从生存与营养空间无限的指数增长的前提下的一个推断, 这与铁竹种群实际增长情况可能不符, 因此种群约20 a加一倍的推论只是一个理论的参考。
Table 4
表4
表4铁竹种群生殖力表
Table 4
| x | lx | bx | lxbx | xlxbx |
|---|---|---|---|---|
| 1 | 1.00 | 0.00 | 0.00 | 0.00 |
| 2 | 0.86 | 0.41 | 0.35 | 0.71 |
| 3 | 0.82 | 0.88 | 0.72 | 2.17 |
| 4 | 0.61 | 0.04 | 0.02 | 0.10 |
| 5 | 0.51 | 0.00 | 0.00 | 0.00 |
| R0 = 1.10 | ||||
| rm = 0.04 | ||||
| T = 2.71 | ||||
| t = 19.70 |
新窗口打开|下载CSV
3.3 铁竹种群的死亡因素
导致铁竹死亡的原因有人为砍伐、自然枯立、退笋、昆虫啃食(图2)。所占比例最大的是人为砍伐, 占45%, 因铁竹幼笋质硬、味苦, 不能食用, 很少被采挖, 但其秆极坚硬, 常被用作菜园的架子, 依据伐桩痕迹, 遭砍伐的铁竹秆龄多≥3年生。其次是自然枯立, 占32%, 枯立竹的秆龄多>3年生, 可能与种内竞争和自疏有关。退笋和昆虫啃食的比例都比较低, 分别约15%、8%, 早期的退笋可能主要与母竹的营养有关; 直锥大象(Cyrtotrachelus thompsoni)啃食过的铁竹幼笋, 很少死亡, 大部分第二年会从被啃食竹节下方的芽发出新的分枝, 但是影响干材的干型和竹材。图2
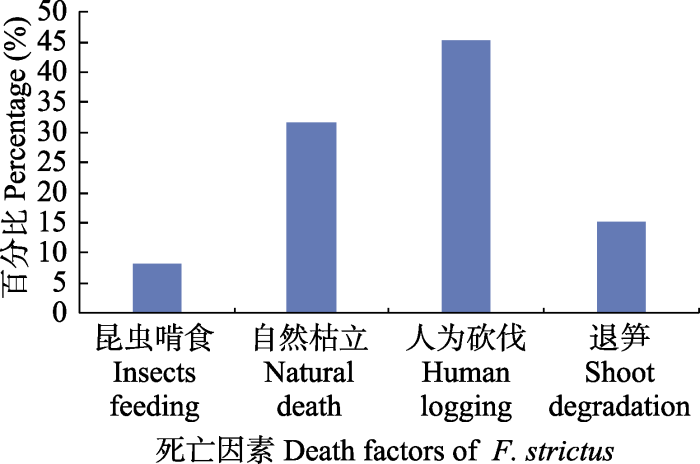 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2铁竹种群的死亡率。
Fig. 2Death rate of Ferrocalamus strictus
3.4 铁竹节间特征
节间长是竹类植物的个体特征之一, 具有明显较长的节是竹类植物快速生长的需要。铁竹是南亚热带的森林竹种, 其节间特征显得特别突出。铁竹节间长度超过1 m。铁竹同一竹秆节间长度变化通常呈单峰曲线规律, 即“短-长-短”变化趋势。铁竹节数11-20节, 峰值出现在第7-9节(图3)。尤为特别的是, 其基部节间伸长变化幅度巨大, 如从1-2节的2.5-16.0 cm跳跃到第3-4节的44-60 cm, 增加幅度为174%-936%, 这种节间突然伸长的特性(图4D), 与耿伯介和薛纪如对铁竹竹秆节间“自短缩节以上的节间即突然正常增长”的描述相一致(耿伯介和薛纪如, 1982)。
图3
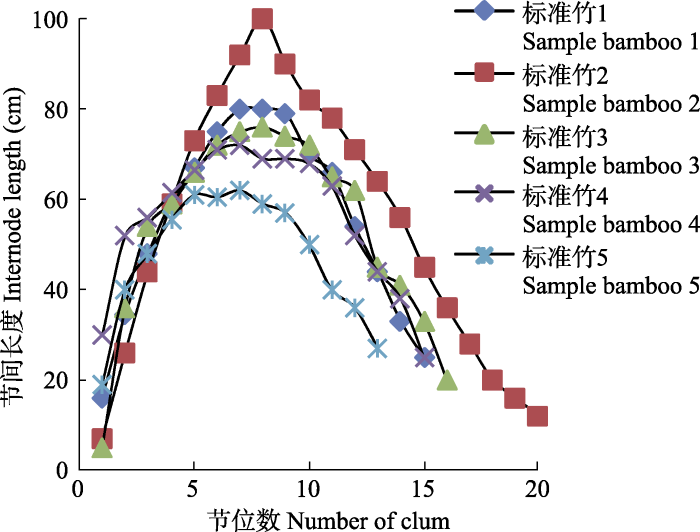 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3铁竹节间生长曲线。
Fig. 3Internode growth curve of Ferrocalamus strictus.
图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4铁竹地下茎(A)、箨叶(B)、秆具类似“多丛性”的现象(C)和铁竹秆基部节间长(D)。
Fig. 4Rhizomes (A), leaf sheaths (B), a similar phenomenon with caespitose of clum (C) and internode length at the base (D) of Ferrocalamus strictus.
4 讨论
4.1 铁竹种群所在群落的特征
墨江杨仕岭地处北回归线, 海拔1 500-1 690 m, 地带性植被为季风常绿阔叶林。铁竹仅分布该区沟箐, 林下生长水晶兰科腐生草本植物水晶兰, 地表腐殖层厚, 比周边山坡季风常绿阔叶林的生境更湿润。在外貌上, 林层较高, 层次结构不明显, 层间植物发达; 在垂直结构上, 铁竹位于灌木层, 且高度明显高于其他物种, 均表明铁竹是南亚热带的一种森林竹种; 在物种组成上, 山榄科绒毛肉实树、番荔枝科云桂暗罗、茜草科岭罗麦(Tarennoidea wallichii)、猕猴桃科尼泊尔水东哥(Saurauia napaulensis)、桃金娘科四角蒲桃(Syzygium tetragonum)、棕榈科云南瓦理棕、露兜树科露兜树、大型蕨类植物金毛狗蕨、大型附生植物爬树龙、大型草本植物大叶仙茅等物种都是云南南部山地雨林较为典型的成分。因此虽然杨仕岭的地带性植被属于季风常绿阔叶林, 但从铁竹种群所在群落片段的外貌和物种组成看, 其群落表现出山地雨林的特征。4.2 铁竹种群现状和发展趋势
铁竹种群的净增殖率(R0)为1.10, 生殖参数值R0 > 1, 表明铁竹种群为增长型。且内禀增殖率(rm)为0.04, rm > 0, 说明未来铁竹种群数量在理想的状态下会不断扩大, 种群加倍时间(t)理论值约20 a。然而铁竹喜湿润, 铁竹种群所在群落具有山地雨林的特征。墨江杨仕岭海拔超过1 500 m, 其地带性植被属于季风常绿阔叶林。该区植被具备山地雨林特征的范围狭小, 仅见于局部湿润沟箐。研究区沿沟箐向上30-40 m的范围, 其植被迅速转变为以单果柯、多穗柯等壳斗科乔木为主的季风常绿阔叶林。与铁竹种群所在群落的生境相比, 季风常绿阔叶林较干燥, 不适宜铁竹生长。因此在墨江杨仕岭, 当铁竹种群扩大到有限程度后, 由于受到较干燥的生境限制, 难以再扩大。所以现在的铁竹种群并非连片分布, 而是很零散地分布在沟箐周围, 且面积狭小; 铁竹作为散生竹类, 也出现种群密度高达2.04株·m-2的现象。同时铁竹种群数量也受到死亡因素的影响。存活曲线一般分为3种类型, 其中Deevey-I型呈凸曲线,早期死亡率很低, 但到达一定生理年龄时则大量死亡; Deevey-II型呈直线, 各年龄内的死亡率相近; Deevey-III型呈凹曲线, 幼龄期死亡率极高, 但成年以后死亡率变低而稳定(Deevey, 1947)。铁竹种群的存活曲线介于Deevey-I型和Deevey-II型之间, 死亡数先减少后增加再减少, 且3-5年生的死亡率分别大于1年生和2年生的死亡率, 以3年生最大。铁竹种群从竹笋-幼竹进入到2-3年生的比例相对较大, 但是从3年生进入到4-5年生的机会比较少。结合死亡原因分析, 铁竹幼笋期死亡原因主要为退笋和昆虫啃食, ≥3年生的铁竹死亡原因多为人为砍伐和自然枯立, 且人为砍伐和自然枯立的比例远大于退笋和昆虫啃食的比例, 因此3-5年生的死亡率分别大于1年生和2年生的死亡率。
因为杨仕岭一直是墨江县城周边重要的水源林, 近年来, 附近居民多在该区修建取用饮用水的管道, 人为活动较多, 加之铁竹秆极坚硬的特性, 常用作菜园的架子, 因此砍伐铁竹的现象比较普遍。建议当地部门做好宣传工作, 帮助当地百姓认识铁竹, 了解铁竹的濒危程度及重要性, 在铁竹生长区域树立保护牌, 协调好铁竹保护工作与居民之间的利益, 加强就地保护。
4.3 铁竹发笋及生长特性
墨江铁竹是该物种分布纬度最北、海拔最高(超过1 500 m)的类群; 其发笋时间为5-9月, 比纬度偏南的金平县(102.52°-103.63° E, 22.43°-23.07° N)铁竹(分布海拔1 200 m以下)的发笋时间晚两个月, 与墨江雨季相一致。文献记载铁竹地下茎为单轴散生、箨片直立(耿伯介和薛纪如, 1982), 观察表明, 墨江铁竹的地下茎有复轴型, 且地面秆类似“多丛性”, 箨片有先直立后反折的现象(图4)。而文献描述铁竹的主要特征之一是地下茎为单轴散生型。因此铁竹地下茎的类型是否真为复轴型还是个别现象, 地面秆类似“多丛性”是否是地下茎类型的表现, 还是因种群密度大, 竹鞭之间相互交错而形成? 墨江铁竹与现在铁竹的分布中心——金平, 无论在地理上还是海拔上都相隔甚远, 是什么原因导致目前的分布特点? 这些生态学问题均值得进一步研究。铁竹节间超过1 m, 且节间长度在秆基部和分枝部位表现出特有的迅速跳跃式生长特性, 这可能与铁竹是南亚热带的森林竹类有关。铁竹高6-10 m, 粗4-5 cm, 节数为11-20节, 平均每节长30-50 cm。乔木状竹类慈竹(Bambusa emeiensis), 高7-9 m, 粗3.8-4.0 cm, 相对应的节数为28-32节, 平均每节长21-28 cm; 高5-6 m, 粗2-3 cm的刚竹(Phyllostachys sulphurea var. viridis), 节数为30-40节, 平均每节长12.5-20.0 cm。可见与其他竹类植物相比, 铁竹的节间数量明显少, 而平均节间长度却明显长。铁竹是南亚热带高大森林下的灌木状竹种, 为争夺阳光, 需要迅速生长, 而节数少, 就必须依靠超长的节间和跳跃式的伸长, 才能快速生长到有利于进行光合作用的空间。这或许是铁竹节位数少而节间长度超长, 且基部节间呈迅速跳跃式生长的原因。

致谢
西南林业大学科技创新基金(C15073)资助。感谢墨江县林业局李恩良工程师、李帅峰副研究员、徐远杰博士等在野外调查和论文写作中给予的帮助。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
China Forestry Publishing House, Beijing.
[本文引用: 1]
[本文引用: 1]
DOIURLPMID [本文引用: 1]

Interpretation of genetic differentiation values is often problematic because of their dependence on the level of genetic variation. For example, the maximum level of GSTis less than the average within population homozygosity so that for highly variable loci, even when no alleles are shared between subpopulations, GSTmay be low. To remedy this difficulty, a standardized measure of genetic differentiation is introduced here, one which has the same range, 0-1, for all levels of genetic variation. With this measure, the magnitude is the proportion of the maximum differentiation possible for the level of subpopulation homozygosity observed. This is particularly important for situations in which the mutation rate is of the same magnitude or higher than the rate of gene flow. The standardized measure allows comparison between loci with different levels of genetic variation, such as allozymes and microsatellite loci, or mtDNA and Y-chromosome genes, and for genetic differentiation for organisms with different effective population sizes.
DOIURL [本文引用: 3]
以国家Ⅱ级保护极小种群野生植物——对开蕨(Phyllitis scolopendrium)为研究对象,分析了在海拔729、1008m群落内,其种群大小、分布频度和密度,个体形态特征指标及其在种群内、种群间差异,苗高分布规律、相对苗高组替代年龄级结构,分布格局和群落各层次的物种多样性,群落的相似性.结果表明:对开蕨在自然分布区内为偶见种,呈斑块状分布.在400m2内,海拔1008m处(01群落)种群密度为31株,样地分布频度为43.75%,最大密度15株/25m2;海拔729m处(02群落)种群密度为91株,样地分布频度为93.75%,最大密度30株/25m2.通过对其自然苗高,叶片数量,有孢子囊叶片数量,最大叶片长、宽值,最小叶片长、宽值,冠径,叶片厚度7个形态指标的分析显示,种群内变异较大,随着植株高度(年龄)的增加,其变异系数均随之减小而趋于稳定;种群间在自然苗高,最大叶片长度和宽度,成熟孢子叶片数量,冠径,叶片厚度上达到极显著差异(P<0.01).其苗高分布呈现双峰型,01种群波谷出现在15.1-20.0cm处;02种群波谷出现在10.1-15.0cm处,两群落均显示有一次较大的更新过程,同时01种群在40.1-45.0cm出现间断.年龄级分析得出,01种群划分为5个龄级,近于正态分布;02种群划分为4个龄级,为倒J型分布;两种群分别处于中龄期和幼龄期,没有出现衰退型年龄结构.格局分析得出,对开蕨种群均为聚集分布.两群落的乔木、灌木、草本层的Shannon-Wiener指数、Pielou均匀度指数、Marglef丰富度指数、生态优势度和种间相遇机率较低(与地带顶级植被相比)而且分布不均;对开蕨在两群落的草本层中重要值较低,仅为伴生种.两群落相似性分析显示,乔木、灌木、草本层的相似度指数分别为66.67%、69.23%和38.46%,草本层差异较大.
DOIURL [本文引用: 3]
以国家Ⅱ级保护极小种群野生植物——对开蕨(Phyllitis scolopendrium)为研究对象,分析了在海拔729、1008m群落内,其种群大小、分布频度和密度,个体形态特征指标及其在种群内、种群间差异,苗高分布规律、相对苗高组替代年龄级结构,分布格局和群落各层次的物种多样性,群落的相似性.结果表明:对开蕨在自然分布区内为偶见种,呈斑块状分布.在400m2内,海拔1008m处(01群落)种群密度为31株,样地分布频度为43.75%,最大密度15株/25m2;海拔729m处(02群落)种群密度为91株,样地分布频度为93.75%,最大密度30株/25m2.通过对其自然苗高,叶片数量,有孢子囊叶片数量,最大叶片长、宽值,最小叶片长、宽值,冠径,叶片厚度7个形态指标的分析显示,种群内变异较大,随着植株高度(年龄)的增加,其变异系数均随之减小而趋于稳定;种群间在自然苗高,最大叶片长度和宽度,成熟孢子叶片数量,冠径,叶片厚度上达到极显著差异(P<0.01).其苗高分布呈现双峰型,01种群波谷出现在15.1-20.0cm处;02种群波谷出现在10.1-15.0cm处,两群落均显示有一次较大的更新过程,同时01种群在40.1-45.0cm出现间断.年龄级分析得出,01种群划分为5个龄级,近于正态分布;02种群划分为4个龄级,为倒J型分布;两种群分别处于中龄期和幼龄期,没有出现衰退型年龄结构.格局分析得出,对开蕨种群均为聚集分布.两群落的乔木、灌木、草本层的Shannon-Wiener指数、Pielou均匀度指数、Marglef丰富度指数、生态优势度和种间相遇机率较低(与地带顶级植被相比)而且分布不均;对开蕨在两群落的草本层中重要值较低,仅为伴生种.两群落相似性分析显示,乔木、灌木、草本层的相似度指数分别为66.67%、69.23%和38.46%,草本层差异较大.
[本文引用: 2]
[本文引用: 2]
URL [本文引用: 3]
正 铁竹属为顶生圆锥花序,小穗多花,具柄,雄蕊及鳞被均为3枚以及叶型和分枝习性均与箬竹属Indocalamus Nakai相近,但本属地下茎为单轴型而非复轴型,外稃先端显著具毛茸,内稃先端不呈裂齿状但具毛茸,花柱单一,特别是果皮肉质而与箬竹属显然有别。本属 在外形上又很象乔草竹属Dendrochloa Parkinson但本属秆较细矮,散生而不是疏稀丛生,花序为单次
URL [本文引用: 3]
正 铁竹属为顶生圆锥花序,小穗多花,具柄,雄蕊及鳞被均为3枚以及叶型和分枝习性均与箬竹属Indocalamus Nakai相近,但本属地下茎为单轴型而非复轴型,外稃先端显著具毛茸,内稃先端不呈裂齿状但具毛茸,花柱单一,特别是果皮肉质而与箬竹属显然有别。本属 在外形上又很象乔草竹属Dendrochloa Parkinson但本属秆较细矮,散生而不是疏稀丛生,花序为单次
DOIURL [本文引用: 1]
在大盘山自然保护区设置了4个有代表性的样地,应用相邻格子法进 行调查获得野外资料.对香果树种群进行统计,绘制径级结构图和高度结构图、编制种群的特定时间生命表;应用理论分布模型和聚集强度指数进行种群分布格局分 析.结果表明:香果树种群结构呈纺锤型,幼苗严重不足,种群有衰退的趋势;其更新是以无性繁殖为主;种群格局整体上呈集群分布,但在不同发育阶段,分布格 局有所差异:幼树阶段一般为集群分布,中树和大树向随机分布发展,这种分布格局变化与其生物学和生态学特性密切相关;其格局规模在介于8S和32S之间. 分析认为,香果树的种群结构和分布格局受到了多种因素的影响,如群落区域小生境、香果树生物学特征以及人为干扰外等,在以上各因素的综合作用下,使得该种 群形成典型的小种群.
DOIURL [本文引用: 1]
在大盘山自然保护区设置了4个有代表性的样地,应用相邻格子法进 行调查获得野外资料.对香果树种群进行统计,绘制径级结构图和高度结构图、编制种群的特定时间生命表;应用理论分布模型和聚集强度指数进行种群分布格局分 析.结果表明:香果树种群结构呈纺锤型,幼苗严重不足,种群有衰退的趋势;其更新是以无性繁殖为主;种群格局整体上呈集群分布,但在不同发育阶段,分布格 局有所差异:幼树阶段一般为集群分布,中树和大树向随机分布发展,这种分布格局变化与其生物学和生态学特性密切相关;其格局规模在介于8S和32S之间. 分析认为,香果树的种群结构和分布格局受到了多种因素的影响,如群落区域小生境、香果树生物学特征以及人为干扰外等,在以上各因素的综合作用下,使得该种 群形成典型的小种群.
DOIURL [本文引用: 2]
铁竹(Ferrocalamus strictus Hsueh et Keng f.)是1982年耿伯介、薛纪如发现的一新属新种,因其个体数量极少,被列为云南省极小种群物种之一。2015年4月,在墨江县北回归线野生植物调查中,发现墨江县有野生铁竹群落分布。墨江县野生铁竹林的发现不仅扩大了铁竹在云南的自然分布区域,还为今后铁竹的培育提供极为难得的优良种源。
DOIURL [本文引用: 2]
铁竹(Ferrocalamus strictus Hsueh et Keng f.)是1982年耿伯介、薛纪如发现的一新属新种,因其个体数量极少,被列为云南省极小种群物种之一。2015年4月,在墨江县北回归线野生植物调查中,发现墨江县有野生铁竹群落分布。墨江县野生铁竹林的发现不仅扩大了铁竹在云南的自然分布区域,还为今后铁竹的培育提供极为难得的优良种源。
[本文引用: 1]
URL [本文引用: 2]
四川西南部和西北部中山、亚高 山森森下,竹源丰富,其中:拐棍竹和冷箭竹是大熊猫的主食竹种,作者1982—1987年进行竹类生态研究;通过固定样方观察和带标志茎秆记录分析,发现 竹杆和笋箨的颜色,以及主枝叶鞘与枝节数的变化和竹子秆龄的增长密切相关,可以作为鉴定竹秆年龄的依据。根据竹子的秆龄结构,应用负指数函数模型 (Lny=Lnyo-bx),可以估算竹子种群的增长量和死亡率,评估竹子的种群动态变化,可为主管部门制定保护大熊猫规划提供竹类资源消长的依据。
URL [本文引用: 2]
四川西南部和西北部中山、亚高 山森森下,竹源丰富,其中:拐棍竹和冷箭竹是大熊猫的主食竹种,作者1982—1987年进行竹类生态研究;通过固定样方观察和带标志茎秆记录分析,发现 竹杆和笋箨的颜色,以及主枝叶鞘与枝节数的变化和竹子秆龄的增长密切相关,可以作为鉴定竹秆年龄的依据。根据竹子的秆龄结构,应用负指数函数模型 (Lny=Lnyo-bx),可以估算竹子种群的增长量和死亡率,评估竹子的种群动态变化,可为主管部门制定保护大熊猫规划提供竹类资源消长的依据。
DOIURL [本文引用: 1]
近年来,中国在生物多样性保护方面取得了一系列成就,但由于人类对自然资源的不断索取和全球变化,一些野生植物仍面临灭绝的风险。目前,仅有1999年国务院批准的《国家重点保护野生植物名录(第一批)》所列种类受到法律的保护。早在20世纪80年代,我国就引入国际自然保护联盟(IUCN)红色名录的评估体系开展了受威胁物种的评估工作,
DOIURL [本文引用: 1]
近年来,中国在生物多样性保护方面取得了一系列成就,但由于人类对自然资源的不断索取和全球变化,一些野生植物仍面临灭绝的风险。目前,仅有1999年国务院批准的《国家重点保护野生植物名录(第一批)》所列种类受到法律的保护。早在20世纪80年代,我国就引入国际自然保护联盟(IUCN)红色名录的评估体系开展了受威胁物种的评估工作,
URL [本文引用: 1]
正 秆高9米,直径7厘米许,节间长达38厘米,绿色无毛,秆环不明显,箨环隆起,基部节有时呈不规则靠近或重叠。箨鞘革质,近矩形、绿色、被脱落性稀疏褐色 刺毛,基部较密,边缘无毛,先端不收缩,两边不均齐;箨舌隆起,高5毫米,先端边缘齿状,具短繸毛;假箨耳椭圆形与箨片、箨鞘相连接,棕褐色、粗糙、皱 摺,边缘有长3—6毫米稀疏繸毛;箨片三角形,直立,基部收缩,先端渐尖,具纵脉、无毛、基部边缘有繸毛。小枝多较簇
URL [本文引用: 1]
正 秆高9米,直径7厘米许,节间长达38厘米,绿色无毛,秆环不明显,箨环隆起,基部节有时呈不规则靠近或重叠。箨鞘革质,近矩形、绿色、被脱落性稀疏褐色 刺毛,基部较密,边缘无毛,先端不收缩,两边不均齐;箨舌隆起,高5毫米,先端边缘齿状,具短繸毛;假箨耳椭圆形与箨片、箨鞘相连接,棕褐色、粗糙、皱 摺,边缘有长3—6毫米稀疏繸毛;箨片三角形,直立,基部收缩,先端渐尖,具纵脉、无毛、基部边缘有繸毛。小枝多较簇
DOIURL [本文引用: 1]
近年来我国提出极小种群野生植物(plant species with extremely small populations,PSESP)的概念并启动实施了全国极小种群野生植物拯救保护工程规划,然而PSESP概念自提出后就不断受到质疑,认为不确切的概念及其引领的保护工程难以取得显著成效。为了更好地理解PSESP概念和相应的保护对策,本文在回顾我国野生植物保护进程的基础上,通过梳理珍稀濒危植物、重点保护植物和极小种群植物等概念,提出从物种选列标准和极小种群临界值等角度理解PSESP概念的途径;并通过比较各个时期采取的保护策略措施,指出基于种群管理的PSESP保护能体现物种以种群形式得以维持和延续的本质,但急需完善传统的资源调查、就地保护、迁地保护方法,探索种群调查分析、近地保护和种群恢复重建等新方法。最后认为PSESP概念的提出及其拯救保护工程的实施在我国野生植物保护领域具有里程碑意义,能促进保护实践和科学研究的相互融合,实现二者共同发展。
DOIURL [本文引用: 1]
近年来我国提出极小种群野生植物(plant species with extremely small populations,PSESP)的概念并启动实施了全国极小种群野生植物拯救保护工程规划,然而PSESP概念自提出后就不断受到质疑,认为不确切的概念及其引领的保护工程难以取得显著成效。为了更好地理解PSESP概念和相应的保护对策,本文在回顾我国野生植物保护进程的基础上,通过梳理珍稀濒危植物、重点保护植物和极小种群植物等概念,提出从物种选列标准和极小种群临界值等角度理解PSESP概念的途径;并通过比较各个时期采取的保护策略措施,指出基于种群管理的PSESP保护能体现物种以种群形式得以维持和延续的本质,但急需完善传统的资源调查、就地保护、迁地保护方法,探索种群调查分析、近地保护和种群恢复重建等新方法。最后认为PSESP概念的提出及其拯救保护工程的实施在我国野生植物保护领域具有里程碑意义,能促进保护实践和科学研究的相互融合,实现二者共同发展。
URL [本文引用: 1]
URL [本文引用: 1]
Plant reproductive susceptibility to habitat fragmentation: Review and synthesis through a metaanalysis
1
2006
... 种群是构成群落的基本单位, 了解其结构不仅有助于了解种群数量动态及发展趋势, 也是群落及生态系统研究的基础; 而群落以种群为基础, 对种群的数量、空间结构和分布又产生实质性影响(
Life tables for natural populations of animals
1
1947
... 同时铁竹种群数量也受到死亡因素的影响.存活曲线一般分为3种类型, 其中Deevey-I型呈凸曲线,早期死亡率很低, 但到达一定生理年龄时则大量死亡; Deevey-II型呈直线, 各年龄内的死亡率相近; Deevey-III型呈凹曲线, 幼龄期死亡率极高, 但成年以后死亡率变低而稳定(
Chinese Bamboo Cultivation and Utilization Manual
1
2002
... 参照前人对竹子年龄结构的划分方法(
Chinese Bamboo Cultivation and Utilization Manual
1
2002
... 参照前人对竹子年龄结构的划分方法(
A standardized genetic differentiation measure
1
2005
... 种群是构成群落的基本单位, 了解其结构不仅有助于了解种群数量动态及发展趋势, 也是群落及生态系统研究的基础; 而群落以种群为基础, 对种群的数量、空间结构和分布又产生实质性影响(
极小种群野生植物对开蕨种群结构特征和群落物种多样性
3
2018
... 种群是构成群落的基本单位, 了解其结构不仅有助于了解种群数量动态及发展趋势, 也是群落及生态系统研究的基础; 而群落以种群为基础, 对种群的数量、空间结构和分布又产生实质性影响(
... ;
... ).分析极小种群野生植物的种群结构及群落特征, 有助于评估其生存状况和预测未来的发展动态(
极小种群野生植物对开蕨种群结构特征和群落物种多样性
3
2018
... 种群是构成群落的基本单位, 了解其结构不仅有助于了解种群数量动态及发展趋势, 也是群落及生态系统研究的基础; 而群落以种群为基础, 对种群的数量、空间结构和分布又产生实质性影响(
... ;
... ).分析极小种群野生植物的种群结构及群落特征, 有助于评估其生存状况和预测未来的发展动态(
2
1992
... 统计铁竹各秆龄株数, 以其为基础, 编制铁竹种群静态生命表, 实际分析中, 存在死亡率为负数的情况, 故采用匀滑技术对数据进行处理(
... 依据静态生命表进一步编制生殖力表: 分年龄组计算lx、bx、lxbx、xlxbx的值, 再按以下公式计算种群净增殖率(R0)、种群内禀增殖率(rm)、种群世代平均周期(T)(本研究指铁竹种群发笋平均周期)和种群加倍时间(t) (
2
1992
... 统计铁竹各秆龄株数, 以其为基础, 编制铁竹种群静态生命表, 实际分析中, 存在死亡率为负数的情况, 故采用匀滑技术对数据进行处理(
... 依据静态生命表进一步编制生殖力表: 分年龄组计算lx、bx、lxbx、xlxbx的值, 再按以下公式计算种群净增殖率(R0)、种群内禀增殖率(rm)、种群世代平均周期(T)(本研究指铁竹种群发笋平均周期)和种群加倍时间(t) (
铁竹属——我国竹类一新属
3
1982
... 铁竹(Ferrocalamus strictus)系禾本科铁竹属(Ferrocalamus)中型散生竹类; 节间超过1 m, 是节间最长的竹类植物; 其笋质硬、味苦, 不堪食用; 秆极坚硬, 是制作弩、竹筷和毛线编织针棒的上乘材料.铁竹属由竹类学家耿伯介和薛纪如于1982年建立(
... 铁竹节间长度超过1 m.铁竹同一竹秆节间长度变化通常呈单峰曲线规律, 即“短-长-短”变化趋势.铁竹节数11-20节, 峰值出现在第7-9节(
... 墨江铁竹是该物种分布纬度最北、海拔最高(超过1 500 m)的类群; 其发笋时间为5-9月, 比纬度偏南的金平县(102.52°-103.63° E, 22.43°-23.07° N)铁竹(分布海拔1 200 m以下)的发笋时间晚两个月, 与墨江雨季相一致.文献记载铁竹地下茎为单轴散生、箨片直立(
铁竹属——我国竹类一新属
3
1982
... 铁竹(Ferrocalamus strictus)系禾本科铁竹属(Ferrocalamus)中型散生竹类; 节间超过1 m, 是节间最长的竹类植物; 其笋质硬、味苦, 不堪食用; 秆极坚硬, 是制作弩、竹筷和毛线编织针棒的上乘材料.铁竹属由竹类学家耿伯介和薛纪如于1982年建立(
... 铁竹节间长度超过1 m.铁竹同一竹秆节间长度变化通常呈单峰曲线规律, 即“短-长-短”变化趋势.铁竹节数11-20节, 峰值出现在第7-9节(
... 墨江铁竹是该物种分布纬度最北、海拔最高(超过1 500 m)的类群; 其发笋时间为5-9月, 比纬度偏南的金平县(102.52°-103.63° E, 22.43°-23.07° N)铁竹(分布海拔1 200 m以下)的发笋时间晚两个月, 与墨江雨季相一致.文献记载铁竹地下茎为单轴散生、箨片直立(
大盘山自然保护区香果树种群结构与分布格局
1
2007
... 种群是构成群落的基本单位, 了解其结构不仅有助于了解种群数量动态及发展趋势, 也是群落及生态系统研究的基础; 而群落以种群为基础, 对种群的数量、空间结构和分布又产生实质性影响(
大盘山自然保护区香果树种群结构与分布格局
1
2007
... 种群是构成群落的基本单位, 了解其结构不仅有助于了解种群数量动态及发展趋势, 也是群落及生态系统研究的基础; 而群落以种群为基础, 对种群的数量、空间结构和分布又产生实质性影响(
墨江县发现野生铁竹群落
2
2015
... 铁竹(Ferrocalamus strictus)系禾本科铁竹属(Ferrocalamus)中型散生竹类; 节间超过1 m, 是节间最长的竹类植物; 其笋质硬、味苦, 不堪食用; 秆极坚硬, 是制作弩、竹筷和毛线编织针棒的上乘材料.铁竹属由竹类学家耿伯介和薛纪如于1982年建立(
... 墨江哈尼族自治县地处云南南部哀牢山中段主脉以西, 介于101.13°-102.07° E, 22.85°-23.98° N之间, 地势自西北向东南倾斜, 地貌类型复杂多样, 属南亚热带半湿润山地季风气候.县城(海拔约1 315 m)年平均气温17.8 ℃, 最冷月平均气温11.5 ℃, 极端最低温-4 ℃, 无霜期306天; 全年总日照时间2 161.2 h; 年降水量1 338 mm, 干湿季分明, 5-10月为雨季, 11月至次年4月为干季; 平均年蒸发量1 696.7 mm, 相对湿度80%.研究区位于县城北边的杨仕岭(101.72° E, 23.46° N), 海拔1 500-1 690 m, 该区为墨江县城周边重要的水源林, 2000年被划为省级公益林, 其地带性植被属于季风常绿阔叶林, 以单果柯(Lithocarpus pseudoreinwardtii)、多穗柯(L. polystachyus)、耳叶柯(L. grandifolius)等壳斗科乔木为优势种, 森林植被保存完好.该区有3小片铁竹分布点, 面积分别约6 580 m2、430 m2和260 m2; 海拔1 510-1 530 m, 地形为沟箐, 坡度20°-40°, 地表湿润, 枯枝落叶层厚8-12 cm, 腐质层厚6-15 cm, 土壤为红壤, 微酸性, 厚度45-65 cm (
墨江县发现野生铁竹群落
2
2015
... 铁竹(Ferrocalamus strictus)系禾本科铁竹属(Ferrocalamus)中型散生竹类; 节间超过1 m, 是节间最长的竹类植物; 其笋质硬、味苦, 不堪食用; 秆极坚硬, 是制作弩、竹筷和毛线编织针棒的上乘材料.铁竹属由竹类学家耿伯介和薛纪如于1982年建立(
... 墨江哈尼族自治县地处云南南部哀牢山中段主脉以西, 介于101.13°-102.07° E, 22.85°-23.98° N之间, 地势自西北向东南倾斜, 地貌类型复杂多样, 属南亚热带半湿润山地季风气候.县城(海拔约1 315 m)年平均气温17.8 ℃, 最冷月平均气温11.5 ℃, 极端最低温-4 ℃, 无霜期306天; 全年总日照时间2 161.2 h; 年降水量1 338 mm, 干湿季分明, 5-10月为雨季, 11月至次年4月为干季; 平均年蒸发量1 696.7 mm, 相对湿度80%.研究区位于县城北边的杨仕岭(101.72° E, 23.46° N), 海拔1 500-1 690 m, 该区为墨江县城周边重要的水源林, 2000年被划为省级公益林, 其地带性植被属于季风常绿阔叶林, 以单果柯(Lithocarpus pseudoreinwardtii)、多穗柯(L. polystachyus)、耳叶柯(L. grandifolius)等壳斗科乔木为优势种, 森林植被保存完好.该区有3小片铁竹分布点, 面积分别约6 580 m2、430 m2和260 m2; 海拔1 510-1 530 m, 地形为沟箐, 坡度20°-40°, 地表湿润, 枯枝落叶层厚8-12 cm, 腐质层厚6-15 cm, 土壤为红壤, 微酸性, 厚度45-65 cm (
Ferrocalamus fibrillosus( Poaceae: Bambusoideae), a new species from Vietnam
1
2012
... 铁竹(Ferrocalamus strictus)系禾本科铁竹属(Ferrocalamus)中型散生竹类; 节间超过1 m, 是节间最长的竹类植物; 其笋质硬、味苦, 不堪食用; 秆极坚硬, 是制作弩、竹筷和毛线编织针棒的上乘材料.铁竹属由竹类学家耿伯介和薛纪如于1982年建立(
大熊猫主食竹种秆龄鉴定及种群动态评估
2
1993
... 参照前人对竹子年龄结构的划分方法(
... 种群密度是影响种群数量动态的主要因素, 通常被认为是种群自身数量调节的作用系统(
大熊猫主食竹种秆龄鉴定及种群动态评估
2
1993
... 参照前人对竹子年龄结构的划分方法(
... 种群密度是影响种群数量动态的主要因素, 通常被认为是种群自身数量调节的作用系统(
论极小种群野生植物的研究及科学保护
1
2015
... 种群是构成群落的基本单位, 了解其结构不仅有助于了解种群数量动态及发展趋势, 也是群落及生态系统研究的基础; 而群落以种群为基础, 对种群的数量、空间结构和分布又产生实质性影响(
论极小种群野生植物的研究及科学保护
1
2015
... 种群是构成群落的基本单位, 了解其结构不仅有助于了解种群数量动态及发展趋势, 也是群落及生态系统研究的基础; 而群落以种群为基础, 对种群的数量、空间结构和分布又产生实质性影响(
我国竹类新分类群 (之一)
1
1984
... 铁竹(Ferrocalamus strictus)系禾本科铁竹属(Ferrocalamus)中型散生竹类; 节间超过1 m, 是节间最长的竹类植物; 其笋质硬、味苦, 不堪食用; 秆极坚硬, 是制作弩、竹筷和毛线编织针棒的上乘材料.铁竹属由竹类学家耿伯介和薛纪如于1982年建立(
我国竹类新分类群 (之一)
1
1984
... 铁竹(Ferrocalamus strictus)系禾本科铁竹属(Ferrocalamus)中型散生竹类; 节间超过1 m, 是节间最长的竹类植物; 其笋质硬、味苦, 不堪食用; 秆极坚硬, 是制作弩、竹筷和毛线编织针棒的上乘材料.铁竹属由竹类学家耿伯介和薛纪如于1982年建立(
极小种群野生植物的概念及其对我国野生植物保护的影响
1
2015
... 种群是构成群落的基本单位, 了解其结构不仅有助于了解种群数量动态及发展趋势, 也是群落及生态系统研究的基础; 而群落以种群为基础, 对种群的数量、空间结构和分布又产生实质性影响(
极小种群野生植物的概念及其对我国野生植物保护的影响
1
2015
... 种群是构成群落的基本单位, 了解其结构不仅有助于了解种群数量动态及发展趋势, 也是群落及生态系统研究的基础; 而群落以种群为基础, 对种群的数量、空间结构和分布又产生实质性影响(
竹植物的秆形结构
1
1998
... 参照前人对竹子年龄结构的划分方法(
竹植物的秆形结构
1
1998
... 参照前人对竹子年龄结构的划分方法(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}