 ,*北京林业大学园林学院, 北京 100083; 城乡生态环境北京实验室, 北京 100083
,*北京林业大学园林学院, 北京 100083; 城乡生态环境北京实验室, 北京 100083Seasonal dynamic of plant phenophases in Beijing—A case study in Beijing Botanical Garden
XING Xiao-Yi, HAO Pei-Yao, LI Guan-Heng, LI Hui, DONG Li,*School of Landscape Architecture, Beijing Forestry University, Beijing 100083, China; and Beijing Laboratory of Urban and Rural Ecological Environment, Beijing 100083, China通讯作者: E-mail:jshe@pku.edu.cn
编委: 陈效逑
责任编辑: 李 敏
收稿日期:2018-05-15修回日期:2018-09-11网络出版日期:2018-09-20
Received:2018-05-15Revised:2018-09-11Online:2018-09-20

摘要
季节性是北京植物景观的典型特征, 而个体物候是植物景观季节动态的基础。该研究基于北京植物园内120种落叶树种的周年物候数据, 对北京植物景观的物候季节动态进行分析。物候观测以中国物候观测法为标准, 采用a、b、c三级物候代码进行物候记录; 数据分析以二十四节气中的候(5日)为基本时间变量, 基于频率分布型法探究北京物候季节划分及其物候构成动态, 基于SPSS 20.0频数分布统计等探究各类物候现象发生期及持续期的时间分布特征等。物候季节划分及物候构成特征结果为: 6-19候为春, 物候期发生频数占全年总量的54.03%, 以发芽、展叶、开花为主要物候特征, 后期有少数树种结果; 20-45候为夏, 物候量占全年的12.95%, 此期全部观测树种完成展叶, 春花树种进入结果期, 并有较少夏花开放及秋色叶出现; 46-59候为秋, 物候量占全年的27.19%, 以秋色叶及落叶物候为主并伴有较少结果和开花物候发生; 60候至次年春季起始前为冬, 其中60-72候物候量仅占全年的0.6%, 全为落叶物候。各类物候期的时间分布特征如下: 展叶物候期分布于3-23候, 华北珍珠梅(Sorbaria kirilowii)、旱柳(Salix matsudana)等展叶最早, 展叶期平均持续3.27候。秋色物候期分布于40-63候, 49-56候为最佳观赏期, 蒙椴(Tilia mongolica)、山杏(Armeniaca sibirica)等最早显秋色; 秋色期平均持续8.2候, 卫矛(Euonymus alatus)、接骨木(Sambucus williamsii)等秋色期较长。叶幕期平均持续44.2候, 糯米条(Abelia chinensis)、旱柳、棣棠(Kerria japonica)等叶幕期最长。花物候期分布于1-53候, 始花期为1-41候, 盛花期平均发生于始花后1.81候, 8-23候为集中观赏期, 蜡梅(Chimonanthus praecox)、迎春(Jasminum nudiflorum)、榆(Ulmus pumila)、毛白杨(Populus tomentosa)等开花最早, 木香薷(Elsholtzia stauntoni)开花最晚; 花期平均持续 6.7候, 华北珍珠梅、木槿(Hibiscus syriacus)、紫薇(Lagerstroemia indica)等夏秋开花树种花期最长。果物候期分布于8-59候, 榆、郁香忍冬(Lonicera fragrantissima)等果实成熟最早; 持果期平均持续29.0候, 果实宿存树种及黑果荚蒾(Viburnum melanocarpum)、‘金叶’风箱果(Physocarpus opulifolius ‘Luteus’)等具有较长的果实观赏期。与20世纪80年代同类研究结果对比分析, 北京2017年的物候季节与30年前相比, 入春提早1候, 夏季延长4候, 入秋延后3候, 秋季缩短2候, 且不同季节持续期长短的差距加大。
关键词:
Abstract
Keywords:
PDF (1338KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
邢小艺, 郝培尧, 李冠衡, 李慧, 董丽. 北京植物物候的季节动态特征——以北京植物园为例. 植物生态学报[J], 2018, 42(9): 906-916 DOI:10.17521/cjpe.2018.0113
XING Xiao-Yi, HAO Pei-Yao, LI Guan-Heng, LI Hui, DONG Li.
北京四季分明, 以暖温带落叶阔叶林为典型地域植被类型, 植物景观具有明显的季节性特征, 即季相。物候学主要是研究自然界植物、动物和环境条件的周期变化之间相互关系的科学(竺可桢和宛敏渭, 1963), 植物物候指植物周期性生命现象的时序规律及其对气候等环境因子的响应(Schwartz, 2013)。季相与物候有密切的相关性, 季相是各物种个体植株的物候特征在群落尺度的组合呈现, 因而物候可作为植物景观季节动态研究的切入点。本研究以北京市植物园内120种常见落叶树种2017年的物候数据为基础, 以中国传统计时单位——“候” (5日)为基本时间变量, 对北京物候季节划分及其物候景观特征、各类物候期发生的时间分布格局进行量化分析, 以期将物候学理论及研究方法应用于植物景观研究, 揭示北京物候季节的整体景观特征及相关规律。
1 研究地和研究方法
1.1 研究地概况
北京地区(39.93° N, 116.33° E)位于华北平原西北边缘, 西部和北部有太行山、燕山余脉环抱。该区属典型暖温带半湿润大陆性季风气候, 四季分明, 夏季炎热多雨, 冬季寒冷干燥; 春、秋短促, 植物景观丰富; 夏、冬漫长, 季相单调乏新。北京植物园位于北京市西北郊卧佛寺附近, 建园于1956年, 园内收集展示各类植物10 000余种, 包括北京城区园林绿地中常见树种, 且植物整体生长发育情况良好, 作为物候观测对象较为稳定可靠。1.2 时间单位
时间是物候特征分析研究中的关键因素。本研究以“日-候-节气-季节”为物候数据采集及数据分析的时间单位体系, 以“日”为物候数据记录的最小时间单位, 以“候” (pentad)(5日)为物候期时间分布特征分析的基本时间单位, 分别以候序(pentad sequence)和候期(pentad period)为描述物候现象发生早晚和持续期长短的基本变量。二十四节气、七十二候是中国传统计时单位, 二十四节气以太阳在黄道上的位置为时间划分依据, 七十二候则利用动植物物候及自然环境的变化为划分标志; 每一节气持续15-16天, 划分为3候, 例如1候(1st pentad)指公历2017年2月3日-7日的时间段, 因是立春节气的第一候, 亦称为“立春一候”, 以此类推。“候”于中国古代被用于物候现象及其发生规律的记录与描述; 且自1996年起, “候” (pentad)即已作为气温及降水统计分析的一个基本单位在气象学研究中得到大量应用, 而考虑到气象条件是影响植物物候的最主要环境因素, 以“候”为单位进行物候特征的分析也是合理的。1.3 物候数据采集方法
1.3.1 物候观测记录方法本研究的物候观测以《中国物候观测方法》(宛敏渭和刘秀珍, 1979)为标准, 采用a、b、c三级物候代码进行物候期记录, 该方法由欧洲通用的二级物候划分方法——BBCH-scale (Meier, 2001)改进而来, 通过增加物候数据分级来提高数据精度。a、b、c分别表示物候期、物候亚期及过渡期, 过渡期(c)根据物候亚期(a, b)的过渡程度划分为1-4 (5即进入下一个物候亚期)。前人研究表明应用BBCH-scale法可在物候数据取样密度较低的情况下获得较为理想的物候数据(Cornelius et al., 2011), a、b、c三级物候代码借鉴了BBCH的典型优点, 可进一步丰富物候数据信息、提高数据精确性。本研究主要观测记录以下15个物候期及物候亚期, 在《中国物候观测方法》观测规范指定的14个物候亚期外, 新增了“结果始期”以用于持果期的分析研究, 结果始期指幼果开始出现的时间。
发芽物候期——芽膨大期(1.1)、芽开放期(1.2), 展叶物候期——展叶始期(1.3)、展叶盛期(1.4), 花物候期——现蕾/花序期(2.3)、始花期(2.4)、盛花期(2.5)、末花期(2.6), 结果物候期——结果始期(3.1)、果实成熟期(3.2)、果实脱落期(3.3), 秋色物候期——叶色始变期(4.1)、秋色叶盛期(4.2), 落叶物候期——落叶始期(5.1)、落叶末期(5.2)。根据不同树种的芽发育特征, 芽开放期包括叶芽开放期、花芽开放期或混合芽开放期, 花芽开放期与叶芽开放期区分标记。
1.3.2 物候观测对象及时间
观测对象为北京植物园内120种北京常见落叶树种(表1), 其对北京地域植物景观的树种构成具有较好代表性。每一树种尽量选择3-5株长势良好、开花结实3年以上的中龄树作为观测对象; 位置以四周空间开阔、南北侧无明显遮挡处为佳, 以尽可能减少南侧遮阴及北侧小气候等因素的影响。
Table 1
表1
表1物候观测树种
Table 1
| 生活型 Life form | 物种 Species | |
|---|---|---|
| 落叶针叶种(2种) Deciduous coniferous species (2 species) | 水杉、华北落叶松 Metasequoia glyptostroboides, Larix principis-rupprechtii | |
| 落叶阔叶种(118种) Deciduous broad-leaved species (118 species) | 乔木(58种) Arbor (58 species) | 旱柳、毛白杨、加杨、刺槐、槐、皂荚、臭椿、元宝槭、梣叶槭*、茶条槭、丝绵木、美国红梣*、流苏树、北京丁香、雪柳、榆、榉树、黑弹树、二球悬铃木*、银杏、栾树、文冠果、柿、毛泡桐、玉兰*、望春玉兰*、二乔玉兰*、梧桐、胡桃、枫杨、七叶树、梓、楸、蒙椴、小花扁担杆、毛梾、灯台树、四照花*、蒙古栎、白桦、黄连木、黄栌、山桃、白花山碧桃*、碧桃*、山杏、榆叶梅、紫叶李*、紫叶稠李*、毛樱桃、‘美人’梅*、‘三轮玉碟’梅*、‘丰后’杏梅*、西府海棠、山楂、蜡梅*、柽柳、沙枣 Salix matsudana, Populus tomentosa, Populus × canadensis, Robinia pseudoacacia, Sophora japonica, Gleditsia sinensis, Ailanthus altissima, Acer truncatum, Acer negundo*, Acer ginnala, Euonymus maackii, Fraxinus pennsylvanica*, Chionanthus retusus, Syringa pekinensis, Fontanesia fortune, Ulmus pumila, Zelkova serrata, Celtis bungeana, Platanus acerifolia*, Ginkgo biloba, Koelreuteria paniculate, Xanthoceras sorbifolium, Diospyros kaki, Paulownia tomentosa, Magnolia denudata*, Magnolia biondii*, Magnolia soulangeana*, Firmiana platanifolia, Juglans regia, Pterocarya stenoptera, Aesculus chinensis, Catalpa ovata, Catalpa bungee, Tilia mongolica, Grewia biloba var. parviflora, Swida walteri, Bothrocaryum controversum, Dendrobenthamia japonica var. chinensis*, Quercus mongolica, Betula platyphylla, Pistacia chinensis, Cotinus coggygria, Amygdalus davidiana, Amygdalus davidiana ‘Albo-plena’*, Amygdalus persica var. persica f. duplex*, Armeniaca sibirica, Amygdalus trilobal, Prunus cerasifera f. atropurpurea*, Prunus virginiana ‘Canada Red’*, Cerasus tomentosa, Armeniaca × blireana ‘Meiren’*, Armeniaca mume var. typica ‘Sanlunyudie’*, Armeniaca mume var. bungo ‘Fenghou’*, Malus × micromalus, Crataegus pinnatifida, Chimonanthus praecox*, Tamarix chinensis, Elaeagnus angustifolia |
| 灌木(54种) Shrub (54 species) | 连翘、迎春*、紫丁香、欧丁香*、花叶丁香*、巧玲花、白鹃梅、郁李、麦李*、皱皮木瓜*、重瓣棣棠*、黄刺玫、金叶风箱果*、三裂绣线菊、麻叶绣线菊、华北珍珠梅、水栒子、平枝栒子*、鸡麻*、野蔷薇、红瑞木、山茱萸*、牡丹*、金银木、郁香忍冬*、新疆忍冬*、天目琼花、欧洲雪球*、香荚蒾*、黑果荚蒾*、红蕾荚蒾*、锦带花、‘红王子’锦带花*、蝟实*、糯米条、六道木、接骨木、黄芦木、卫矛、太平花、大叶醉鱼草*、莸*、牡荆*、锦鸡儿、紫荆*、木香薷、木槿*、紫薇*、蚂蚱腿子、薄皮木、牛奶子、迎红杜鹃、照山白、小叶鼠李 Forsythia suspensa, Jasminum nudiflorum*, Syringa oblata, Syringa vulgaris*, Syringa × persica*, Syringa pubescens, Exochorda racemose, Cerasus japonica, Cerasus glandulosa*, Chaenomeles speciosa*, Kerria japonica f. pleniflora*, Rosa xanthine, Physocarpus opulifolius ‘Luteus’*, Spiraea trilobata, Spiraea cantoniensis, Sorbaria kirilowii, Cotoneaster multiflorus, Cotoneaster horizontalis*, Rhodotypos scandens*, Rosa multiflora, Swida alba, Cornus officinalis*, Paeonia suffruticosa*, Lonicera maackii, Lonicera fragrantissima*, Lonicera tatarica*, Viburnum opulus var. calvescens, Viburnum opulus ‘Roseum’*, Viburnum farreri*, Viburnum melanocarpum*, Viburnum carlesii*, Weigela florida, Weigela florida ‘Red Prince’*, Kolkwitzia amabilis*Abelia chinensis, Abelia biflora, Sambucus williamsii, Berberis amurensis, Euonymus alatus, Philadelphus pekinensis, Buddleja davidii*, Caryopteris divaricata*, Vitex negundo var. cannabifolia*, Caragana sinica, Cercis chinensis*, Elsholtzia stauntoni, Hibiscus syriacus*, Lagerstroemia indica*, Myripnois dioica, Leptodermis oblonga, Elaeagnus umbellate, Rhododendron mucronulatum, Rhododendron micranthum, Rhamnus parvifolia | |
| 藤本(6种) Liana (6 species) | 紫藤、五叶地锦*、美国凌霄*、木香*、盘叶忍冬* Wisteria sinensis, Parthenocissus quinquefolia*, Campsis radicans*, Rosa banksiae*, Lonicera tragophylla* | |
新窗口打开|下载CSV
物候观测持续一年72候, 于2017年立春至大寒(2017-02-03至2018-02-03), 观测时间基本涵盖北京大部分落叶树种的完整生命周期。由1-2名固定观测者于每日14:00开始进行观测(因一天中一般以13:00-14:00为气温最高时段, 植物物候现象常于高温后出现)。观测周期随季节及物候变化速率作相应调整: 春秋两季气温升降幅度大, 物候变化急剧, 以1-2天为观测周期; 夏冬两季物候变化平缓, 以3-5天为观测周期。
1.4 数据分析
1.4.1 物候季节划分物候季节划分采用陈效逑和曹志萍(1999)提出的频率分布型法, 将120种观测树种立春至大寒72候的物候数据组成混合样本, 以候为单位计算各物候期的出现频数及频率, 绘制累积频率曲线, 根据曲线的波动阶段性及累积频率值, 确定各季节的起讫日期。
1.4.2 频数分布统计
采用SPSS 20.0及Excel 2016等统计软件对各物候期发生期及持续期进行频数分布的统计描述等, 以探究各物候现象的时间分布格局及季节性规律。本研究采用物候量描述特定时间段内所有观测树种被观测到的某种、某类或全部物候期的总发生频数。
2 结果
2.1 基于频率分布型法的物候季节划分及其物候构成特征
如图1所示, 根据累积频率曲线的斜率变化及拐点位置, 可将整个年周期(1-72候)分为5个时段, 春夏秋冬4个物候季节。1-5候为早春来临前的暮冬时节(2016年冬季)。6-19候曲线斜率明显增大, 呈急剧上升趋势, 定为春季; 20-45候曲线趋于平缓, 有小幅增势且斜率保持稳定, 定为夏季; 46-59候曲线又呈明显抬升, 定为秋季; 60候之后, 物候期累积频率短期内达到最大值后再无变化, 定为冬季。需注意的是, 考虑到季节完整性, 冬季并非到72候即结束, 而是持续到次年春季起始前, 且次年早春前的暮冬时节会有少量植物呈现萌动迹象并伴随物候现象发生, 但因其为次年新一轮植物生命周期的起始, 且考虑到跨年度的问题, 故不将其纳入本研究物候季节划分的时间范围, 而将2016年冬季的暮冬时节, 即2017年1-5候纳入研究范围。图1
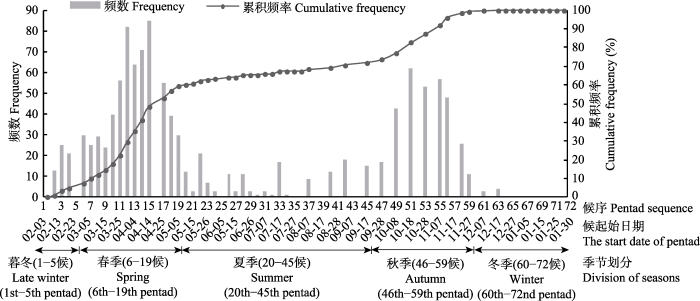 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1物候期累积频率曲线及频数分布直方图。候起始日期MM-DD表示公历日期MM月DD日, 1-65候为2017年, 66-72候为2018年。
Fig. 1The cumulative frequency and frequency distribution of phenophases. For the start date of pentad, MM-DD refers to the date of DD in the month of MM in Gregorian Calendar; 1st-65th pentads are in the year of 2017, while the year of 2018 starts from 66th pentad.
根据季节划分结果, 立春伊始(1候)春季尚未到来, 但少部分树种伴随气温缓慢回升已呈现芽膨大
等萌动迹象, 截至雨水二候(5候), 物候发生量占全年总量的5.23%, 以发芽物候及开花物候为主; 发芽物候占4.03%, 其中芽膨大期占3.34%, 芽开放期占0.69%; 开花物候仅占1.20%, 且多由现蕾/花序期构成(0.94%)。
春季始于雨水三候(6候), 止于立夏一候(19候), 持续14候, 物候量占全年总物候量的54.03%。图2显示, 此期以发芽、展叶、开花为主要物候特征, 后期有少数树种结果。发芽物候期物候量占全年总物候量的8.92%, 其中芽膨大期占1.80%, 芽开放期占7.12%。展叶物候期占15.35%, 其中展叶始期占7.80%, 展叶盛期占7.55%。开花物候期占25.56%, 其中现蕾/花序期占6.26%, 始花期占6.86%, 盛花期占6.60%, 末花期占5.83%。结果物候期占4.20%, 其中结果始期占3.52%, 果实成熟期占0.34%, 果实脱落期占0.34%。
图2
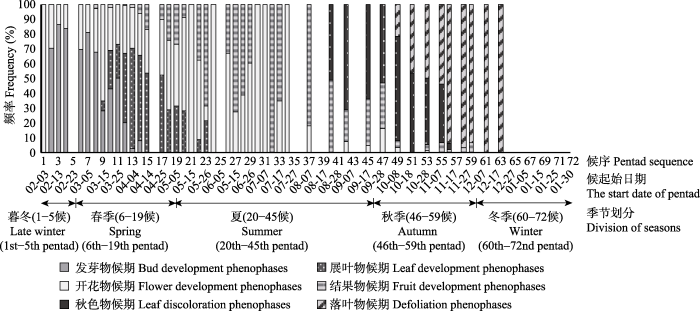 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2各类物候期频率堆积柱形图。候起始日期MM-DD表示公历日期MM月DD日, 1-65候为2017年, 66-72候为2018年。
Fig. 2The frequency and composition of different phenophases. For the start date of pentad, MM-DD refers to the date of DD in the month of MM in Gregorian Calendar; 1st-65th pentads are in the year of 2017, while the year of 2018 starts from 66th pentad.
据此期间累积频率曲线的斜率变化, 又可将春季分为初春、仲春、暮春3个季段。6-9候曲线斜率维持在较低值, 物候发生速率较缓, 为草木萌动之时, 定为初春, 物候现象以发芽、开花为主, 物候量占全年9.26%, 其中发芽物候占4.46%, 开花物候占4.63%, 展叶物候占0.09%, 结果物候占0.09%。10-15候曲线斜率显著增大并维持在较大值, 物候变化急剧, 定为仲春, 至此全部观测树种已萌动发芽, 早发芽树种纷纷进入展叶期、花期甚至果期; 此阶段物候量占全年总物候量的34.13%, 其中开花物候占15.09%, 展叶物候占12.44%, 发芽物候占4.46%, 结果物候占2.14%。16-19候, 曲线趋于平缓, 定为暮春, 物候量占全年的10.63%, 此时, 展叶、开花、结果的物候景观并现, 其中开花物候占5.83%, 展叶物候占2.83%, 结果物候占1.97%。
夏季始于立夏二候(20候), 止于白露三候(45候), 持续期为26候, 夏初至22候, 全部观测树种完成展叶, 春花树种纷纷进入结果期, 并有较少夏花开放, 暮夏(40候)已有少部分树种始显秋色叶。夏季物候量占全年总量的12.95%。开花物候占5.32%, 其中现蕾/花序期占0.43%, 始花期、盛花期及末花期均占1.63%; 结果物候占5.23%, 其中结果始期占1.72%, 果实成熟期占2.14%, 果实脱落期占1.37%; 秋色物候占2.06%, 全由秋色始期构成; 展叶物候仅占0.34%, 且全为展叶盛期。
秋季始于秋分一候(46候), 止于小雪二候(59候), 共持续14候, 物候期以秋色叶及落叶物候为主, 并伴随有部分结果物候和极少开花现象发生。物候量占全年总量的27.19%。落叶物候期占全年物候总量的14.92%, 其中落叶始期及落叶末期分别占7.38%及7.55%; 秋色物候期占9.95%, 其中叶色始变期及秋色盛期分别占5.40%及4.55%; 结果物候期占1.72%, 其中结果始期占0.09%, 果实成熟期占1.03%, 果实脱落期占0.60%; 开花物候期占0.60%, 其中盛花期占0.17%, 末花期占0.43%。
据累积频率曲线的斜率变化, 又可将秋季分为初秋、仲秋、暮秋三个季段。46-47候曲线有轻微抬升, 定作初秋, 物候量占全年总量的1.46%, 由秋色物候期、开花物候期及结果物候期构成, 其中秋色物候占0.60%, 结果物候占0.51%, 开花物候占0.34%。48-56候曲线显著抬升, 物候变化渐急剧, 定作仲秋, 是植物秋色景观最显著的时期, 亦是落叶季的开端; 此阶段物候量占总量的22.56%, 其中落叶物候占11.92%, 秋色物候占9.35%, 结果物候占1.03%, 开花物候占0.26%。57-59候曲线斜率增势减缓, 定作暮秋, 是北京落叶树种的集中落叶期; 物候现象占全年总量的3.17%, 其中落叶物候占3.00%, 结果物候占0.17%。
冬季始于小雪三候(60候), 止于次年春季; 截至72候(大寒三候), 全部观测树种2017年的生命周期及物候现象已经结束, 60-72候的物候量仅占全年总量的0.6%, 且全为落叶物候, 冬初63候是少数晚落叶树种叶幕期的终点, 此后百木尽疏。
2.2 各类物候期时间分布特征分析
对叶、花、果三类物候期发生期及持续期的时间分布特征进行分析。2.2.1 叶物候期时间分布特征
叶物候期包括展叶物候期、秋色物候期及落叶物候期, 展叶物候期包括展叶始期及展叶盛期, 秋色物候期包括秋色始期及秋色盛期, 落叶物候期包括落叶始期及落叶末期(秋色末期)。因叶芽开放作为叶发育的起点, 与展叶是相继发生的, 故将叶芽开放期也纳入展叶物候期分析中。
(1) 展叶物候期
由图3可知, 展叶物候期始于立春三候(3候), 止于小满二候(23候), 持续20候。叶芽开放期发生于仲春及其之前的3-14候。华北珍珠梅(Sorbaria kirilowii)、平枝栒子(Cotoneaster horizontalis)、牡丹(Paeonia suffruticosa)等为最早发芽树种(3-4候), 而槐(Sophora japonica)、毛泡桐(Paulownia tomentosa)、梧桐(Firmiana platanifolia)、紫薇(Lagerstroemia indica)等则最晚发芽展叶。展叶始期全部发生于仲春时段的10-15候, 华北珍珠梅等最早进入展叶期(9候), 此外, 旱柳(Salix matsudana)、平枝栒子、丝绵木(Euonymus maackii)、糯米条(Abelia chinensis)、皱皮木瓜(Chaenomeles speciosa)、蝟实(Kolkwitzia amabilis)等也是早春绿意萌动的标志树种。展叶盛期发生于仲春至初夏的12-23候。
图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3叶物候期累积频率曲线及频数分布直方图。候起始日期MM-DD表示公历日期MM月DD日, 1-65候为2017年, 66-72候为2018年。
Fig. 3The cumulative frequency and frequency distribution of leaf phenophases. For the start date of pentad, MM-DD refers to the date of DD in the Month of MM in Gregorian Calendar; 1st-65th pentads are in the year of 2017, while the year of 2018 starts from 66th pentad.
对展叶持续期(展叶始期至展叶盛期)的数量特征进行分析。展叶持续期最短1候、最长8候, 平均持续3.27候, 众数为2候(35.2%), 数据整体呈正偏态分布, 集中分布于1-5候, 偏度系数0.802, 即展叶持续期偏长的树种整体占比较少。发芽持续期(叶芽开放期至展叶始期)最短1候, 最长9候, 平均持续3.03候, 众数2候(29%), 数据整体呈正偏态分布, 集中分布于1-6候。
(2)秋色及落叶物候期
秋色期始于初秋的处暑一候(40候), 止于初冬的大雪三候(63候), 共持续24候。
秋色始期发生于暮夏至仲秋的40-55候(处暑一候至立冬一候); 秋色呈现最早的树种有蒙椴(Tilia mongolica)、卫矛(Euonymus alatus)(40候), 而紫叶李(Prunus cerasifera f. atropurpurea)、棣棠(Kerria japonica)、木香薷(Elsholtzia stauntoni)等则最晚(55候)进入秋色期。秋色盛期发生于仲秋49-56候, 为秋色景观最佳观赏期, 也常伴随落叶现象发生; 一定的叶量是秋色呈现的基础, 而有46.2%的观测树种因其落叶与叶变色物候近同步发生而未能呈现秋色盛期景象, 即不能算作典型秋色叶树种。落叶期发生于仲秋至暮秋的49-63候, 其中55-59候是落叶急剧发生期, 截至59候, 92.6%的落叶树种已达叶幕期终点。
对秋色持续期(秋色始期至落叶末期)及落叶持续期(落叶始期至落叶末期)的数量特征进行分析, 结果见图4。 秋色持续期为3-21候, 均值8.16候, 中值7候, 众数6候(18.5%), 集中分布于3-13候, 数据整体呈正偏态分布, 偏度系数0.886。文冠果(Xanthoceras sorbifolium)、木香薷、碧桃(Amygdalus persica var. persica f. duplex)、山茱萸(Cornus officinalis)、紫叶李等秋色持续期最短(3候), 胡桃(Juglans regia)秋色持续期最长(21候), 此外, 秋色持续期较长的树种还有卫矛(18候)、接骨木(Sambucus williamsii) (16候)、榉树(Zelkova serrata) (16候)等。
图4
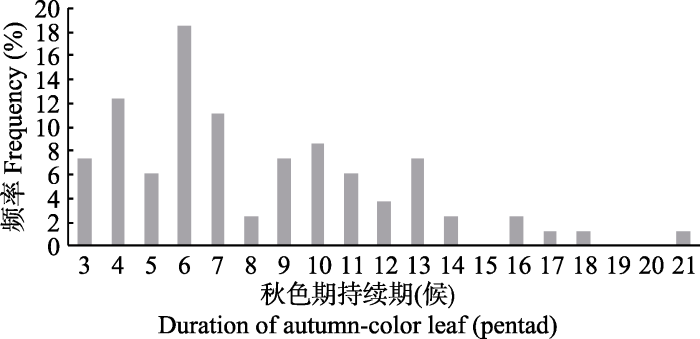 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4秋色期持续期频率分布直方图。
Fig. 4The frequency distribution of duration of autumn-color leaf.
落叶持续期(落叶始期至落叶末期)最短1候, 最长7候, 平均持续3.25候, 中值、众数为3候(48.8%), 水杉(Metasequoia glyptostroboides)、柿(Diospyros kaki)、楸(Catalpa bungei)、灯台树(Bothrocaryum controversum)、山茱萸、牡丹、紫薇等树种的落叶持续期最短, 仅为1候, 而糯米条、木槿(Hibiscus syriacus)的落叶持续期可长达7候。
(3)叶幕期及绿色期
叶幕期从展叶始期至落叶末期, 代表植物叶片具有景观观赏性的时期(包括展叶期、秋色期和落叶期以及夏季漫长的叶色稳定期); 绿色期指从展叶始期至秋季叶色始变期(杨国栋和陈效逑, 2000), 一定程度上反映植物一年中光合作用的有效时间; 两者相比, 叶幕期更能反映叶物候的整体特征, 故对叶幕期数量特征进行重点分析。
观测树种的叶幕期最短持续36候、最长持续53候。统计数据服从近似正态分布, 如图5所示, 叶幕期持续期集中分布于41-49候, 均值为44.17候, 中值43.50候, 众数43候, 且有34.7%的树种叶幕期持续42-43候, 糯米条、沙枣(Elaeagnus angustifolia)、旱柳、棣棠、平枝栒子等是叶幕期最长的树种(持续49-53候), 而小叶鼠李(Rhamnus parvifolia)、紫薇、楸、白桦(Betula platyphylla)则为短叶幕期树种的代表(持续36-40候), 长叶幕期树种对于延长北京植物观赏期, 改善冬季至早春时段绿色匮乏的植物景观现状具有十分重要的应用价值。
图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5叶幕期持续期频率分布直方图。
Fig. 5The frequency distribution of duration of canopy.
2.2.2 花物候期时间分布特征
花物候期始于立春一候(1候), 止于霜降二候(53候), 由暮冬至仲秋, 共持续53候。
现蕾/花序期分布于春夏季节的2-28候, 初春至仲春的3-15候是现蕾急剧发生期。春季(6-19候)现蕾/花序期的累积发生频率达82.02%, 蜡梅(Chimonanthus praecox)、迎春(Jasminum nudiflorum)、毛白杨(Populus tomentosa)、郁香忍冬(Lonicera fragrantissima)、香荚蒾(Viburnum farreri)等早于立春三候即已现蕾/花序, 特别是蜡梅的早花品种于2016年秋季即已完成花芽发育, 是最早进入花发育期的树种。
始花期发生于暮冬至初秋的1-42候, 初春至夏初的6-22候为始花急剧发生期(累积发生量88.60%)。盛花期分布于初春至仲秋的6-49候, 平均发生于始花后1.81候, 与始花期的时间分布特征基本一致, 初春至初夏的8-23候期间有83.68%的树种呈现盛花, 是春花景观的最佳观赏期。末花期起止于9-53候, 集中发生于仲春至夏的10-26候(85.09%)。蜡梅、迎春、榆(Ulmus pumila)、毛白杨、山桃(Amygdalus davidiana)、郁香忍冬、山茱萸、连翘(Forsythia suspensa)、望春玉兰(Magnolia biondii)等先花后叶树种于春分(10候)前即已始花, 是北京花季序幕开启者; 而木香薷于处暑三候(42候)才见始花, 霜降二候(53候)末花, 可视作北京花期的落幕。
对开花持续期(始花期至末花期)的数量分布特征分析, 开花持续期最短2候, 最长28候, 均值6.70候, 众数为5候(21.9%)(图6); 数据整体呈极显著正偏态分布, 偏度系数2.792; 开花持续期时长多为2-8候, 其对应树种多为春花树种, 如毛白杨、加杨(Populus × canadensis)、白桦、榉树、胡桃等花期仅持续2候; 而开花持续期较长的树种则多于夏秋开花, 如华北珍珠梅(28候)、木槿(19候)、紫薇、海州常山(Clerodendrum trichotomum)(16候)、柽柳(Tamarix chinensis)(15候)等, 且大多数夏花植物的开花期都并非一次, 而是多次, 这对于丰富北京夏季植物景观具有重要应用价值。
图6
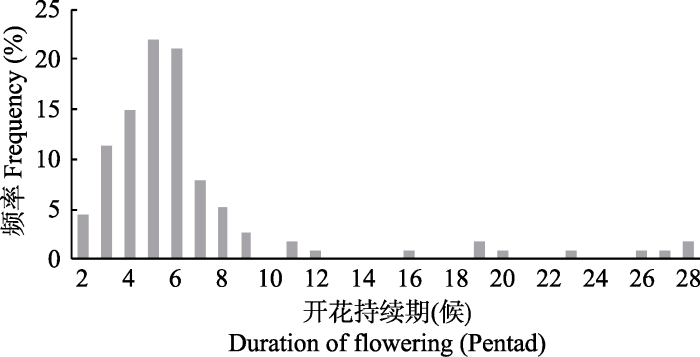 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6开花持续期频率分布直方图。
Fig. 6The frequency distribution of duration of flowering.
2.2.3 果物候期时间分布特征
果物候期始于初春8候(惊蛰二候), 止于暮秋59候(小雪二候), 持续52候。
结果始期分布于初春至初秋的8-45候, 集中于仲春至夏季时段, 多发生于末花期及前后1候, 因而与末花期的季节分布特征基本一致。结果最早树种为榆(8候)、玉兰(Magnolia denudata) (12候)、山桃(13候)、郁香忍冬(13候)等早花树种, 而莸(Caryopteris divaricata)等晚花树种于45候才进入结果期。
果实成熟期发生于暮春至暮秋的15-58候, 集中在夏季至仲秋时段。榆、郁香忍冬、毛樱桃(Cerasus tomentosa)等树种果实成熟最早(早于21候), 而雪柳(Fontanesia fortunei)、白鹃梅(Exochorda racemosa)、莸、毛泡桐等果实则晚于54候方才成熟; 果实成熟期时间分布的季节性特征并不显著, 而相对而言, 入秋后果实成熟发生渐急剧, 40候(处暑一候)呈现小高峰。果实脱落期与果实成熟期的时间分布特征基本一致, 75.3%的树种果实脱落期发生于果实成熟期后1-3候; 所余24.7%的观测树种存在果实宿存现象, 即其在观测期(72候)内虽多少有落果现象发生, 但果实未全部脱落, 故无果实脱落末期的记录, 如木槿、紫薇、毛泡桐、白鹃梅、槐、刺槐(Robinia pseudoacacia)、紫荆(Cercis chinensis)等。
对结果持续期/持果期(结果始期至果实脱落期)的数量特征进行分析, 将果实宿存树种的果实脱落期定为72候以便于种间比较。观测树种的持果期为7-57候, 均值29.02候, 众数为18候, 数据的频率分布较为均匀, 无明显正态性特征(图7)。榆与郁香忍冬持果期最短(7候), 果实宿存树种的持果期可长达50-57候, 果实脱落树种中, 紫藤(Wisteria sinensis)、雪柳、黑果荚蒾(Viburnum melanocarpum)、金叶风箱果(Physocarpus opulifolius ‘Luteus’)等也具有较长的果实观赏期(35-39候)。
图7
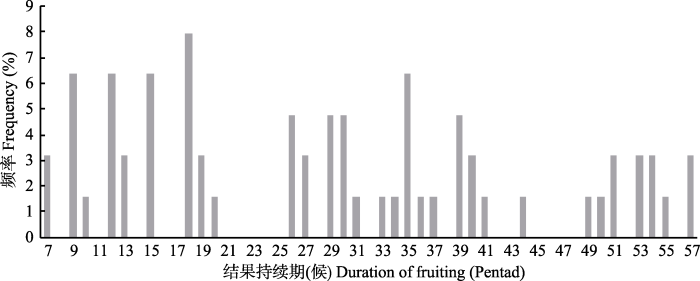 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7结果持续期频率分布直方图。
Fig. 7The frequency distribution of duration of fruiting.
3 讨论
3.1 物候季节划分及特征研究
本研究以陈效逑和曹志萍(1999)提出的频率分布型法为基本方法, 以120种园林树种的15种物候期组成的混合样本为数据基础, 对2017年北京的自然季节进行了划分, 结果如下: 6-19候为春(6-9候为初春, 10-15候为仲春, 16-19候为暮春); 20-45候为夏; 46-59候为秋(46-47候为初秋, 48-56候为仲秋, 57-59候为暮秋, 60候至次年春季开始前为冬。陈效逑和曹志萍(1999)以小西山卧佛寺一带(北京植物园区域) 1979-1987年间26种树种、14种物候期组成的样本为基础数据, 对北京物候季节进行了划分; 此研究以自然年为候序划定标准, 即1月1日-1月5日定为1候, 而本研究以二十四节气为候序划分标准, 即立春一候(2月3日-2月7日)定为1候, 故以本研究方法为标准, 对其研究结果进行校正, 得到20世纪80年代北京地区的物候季节: 7-20候为春(7-9候为初春, 10-17候为仲春, 18-20候为暮春); 21-42候为夏; 43-58候为秋(43-47候为初秋, 48-51候为仲秋, 52-58候为暮秋); 59至次年春季为冬。北京的物候季节与30年前相比, 入春时间提早1候, 夏季延长4候, 入秋时间延后3候, 秋季缩短2候, 不同季节持续期长短的差距加大; 而其他相关研究也表明, 在北半球的中高纬度地区, 近年来受全球变暖影响, 植物春季物候提前, 秋季物候推迟, 生长季变长等, 已成为日益普遍的现象(Schwartz et al., 2006; Shutova et al., 2006; Ge et al., 2011; 仲舒颖等, 2012)。由表2可知, 2017年春、夏季的平均日均温均高于20世纪80年代, 且与30年前相比, 生长季特别是春季物候期的累积发生频率及候均发生频率(物候期候均发生频率=物候期累积频率/持续候期)明显增多, 而秋季物候的累积频率则明显减少, 进一步说明30年来的气候变化可能导致物候发生时间更加集中, 发生期整体提前。此外, 对于同物候期的年际波动, 一般较晚发生的物候期比较早发生的物候期更加稳定(Chen et al., 2017), 这在一定程度上可以解释两研究结果中初春与初秋的起始时间有较大差异, 而仲春与仲秋的起始时间则较为一致的问题。
Table 2
表2
表230年前后物候季节划分及相关特征差异
Table 2
| 季节 Season | 物候季节划分(候) Division of phenological seasons (Pentad) | 持续候期/候 Duration pentads | 物候期累积频率(%) Cumulative frequency of phenophases (%) | 物候期候均发生频率(%) Pentad-average cumulative frequency of phenophases (%) | 平均日均温(℃) Average mean daily temperature (℃) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| 2017 | 1980’s | 2017 | 1980’s | 2017 | 1980’s | 2017 | 1980’s | 2017 | 1980’s | |
| 春季 Spring | 6-19 | 7-20 | 14 | 14 | 54.03 | 44.00 | 3.86 | 3.14 | 13.6 | 11.3 |
| 夏季 Summer | 20-45 | 21-42 | 26 | 22 | 12.95 | 12.00 | 0.50 | 0.55 | 25.2 | 24.1 |
| 秋季 Autumn | 46-59 | 43-58 | 14 | 16 | 27.19 | 38.00 | 1.94 | 2.39 | 9.4 | 12.1 |
新窗口打开|下载CSV
物候数据样本量与观测年份是影响本研究物候季节划分结果可靠性的主要因素。本研究虽然仅基于一年的物候数据划分季节并探讨各季节内植物物候期的变化特征, 无法代表多年平均的物候季节特征, 但实现了通过一年快速、详细地群落植物物候观测, 获取当年植物群落物候季节和景观季相的信息, 具有显著的时效性, 可为下一年物候季节的划分以及植物园季节性旅游活动的设计提供专业化的指导和参考。此外, 北京的物候具有准年周期性, 即同一物候期在相邻年份间的发生时距大约为1年, 其平均年际波动维持在一定幅度之内(5-16天)(杨国栋和陈效逑, 1995; Chen et al., 2017), 这是由于物候是外部环境因素与植物内在调控共同作用的结果, 特别以乡土植物为代表的对生长地具有较强生态适应性的树种在应对年际气温波动的响应方面通常表现出较高的表型可塑性(phenotypic plasticity)(Liang & Schwartz, 2014)。其次, 物候现象的发生具有顺序相关性, 即各物候现象每年都按一定先后顺序出现, 且在一定时段内具有大体同步提前或延后的特点(杨国栋和陈效逑, 1995; Chen et al., 2017)。基于上述规律, 一年的物候数据在物候期时间分布格局、各物候现象发生的相对时序规律等问题的探究中仍具有一定代表性。
3.2 物候的季节动态及时间分布特征研究
季节性是北京植物景观的典型特征, 而个体物候是植物景观季节动态的基础。关于物候在植物景观研究中的应用, 许多****从不同地域特征和应用方向已进行了一定的研究(姚永正等, 1991; 杨国栋和陈效逑, 2000; 许涵和王晓明, 2004; 吴迪和赵兰勇, 2014), 但有关北京地域性物候特征的研究较少。本研究从各季节的物候构成特征和各类物候期的时间分布特征两个角度对北京植物景观的物候动态进行了定量分析。在季节物候特征的研究方面, 杨国栋及陈效逑等人从物候角度对北京植物季相特征进行了定性归纳总结(杨国栋和陈效逑, 1995; 陈效逑和曹志萍, 1999), 而本研究在此基础上基于120种树种的大体量物候数据, 以候(5日)为基本时间单位, 采用累积频率对各季节及季段的物候构成特征进行了定量分析, 进一步揭示了物候期构成随时间变化的动态特征。
在物候期的时间分布特征方面, 前人多基于物候特征对植物进行分组分类, 如赵小雷等(2013)根据萌芽和展叶物候期的时间分布特征对上海崇明岛45种园林绿化树种划分为多类萌芽期至展叶期类型; 杨国栋和陈效逑(2000)根据各类物候期发生早晚及其组合对北京常见木本植物进行物候相分类等。本研究在前人研究基础上, 针对叶物候、花物候和果物候这三大类景观效果突出的物候类型, 对北京城市植被的发芽物候、展叶物候、开花物候、结果物候、秋色物候及落叶物候六大类物候期的发生时序规律、季节分布格局、持续期分布特征及相应物候相景观等进行了综合分析, 较为系统地阐述了北京地区植物景观的物候季节动态, 在将物候学理论及方法体系应用于植物景观研究方面做出了探索。

致谢
感谢北京园林绿化增彩延绿科技创新工程科研项目(CEG-2017-01)资助。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOIURLMagsci [本文引用: 6]

<p>在前人物候季节划分研究的基础上,提出了划分自然景观季节的物候频率分布型法,并利用北京小西山山前一带物候历中的资料,将当地的物候季节划分为12个季段。划分结果表明,每个季节阶段都具有典型的植物物候形态组合与色彩组合,独特的季相特征,并且与气温和降水的季节分配状况相吻合。本方法与以往的物候季节划分方法相比,具有划分指标定量且综合、划分季段详细、季节的内涵丰富、适用区域范围广阔等特点。</p>
DOIURLMagsci [本文引用: 6]
<p>在前人物候季节划分研究的基础上,提出了划分自然景观季节的物候频率分布型法,并利用北京小西山山前一带物候历中的资料,将当地的物候季节划分为12个季段。划分结果表明,每个季节阶段都具有典型的植物物候形态组合与色彩组合,独特的季相特征,并且与气温和降水的季节分配状况相吻合。本方法与以往的物候季节划分方法相比,具有划分指标定量且综合、划分季段详细、季节的内涵丰富、适用区域范围广阔等特点。</p>
DOIURLPMID [本文引用: 3]
Using woody plant phenological data in the Beijing Botanical Garden from 1979 to 2013, we revealed three levels of phenology rhythms and examined their coherence with temperature rhythms. First, the s
DOIURLPMID [本文引用: 1]
The BBCH scale is a two-digit key of growth stages in plants that is based on standardised definitions of plant development stages. The extended BBCH scale, used in this paper, enables the coding of the entire development cycle of all mono- and dicotyledonous plants. Using this key, the frequency distribution of phenological stages was recorded which required a less intense sampling frequency. The onset dates of single events were later estimated from the frequency distribution of BBCH codes. The purpose of this study was to present four different methods from which those onset dates can be estimated. Furthermore, the effects of (1) a less detailed observation key and (2) changes in the sampling frequency on estimates of onset dates were assessed. For all analyses, phenological data from the entire development cycle of four grass species were used. Estimates of onset dates determined by Weighted Plant Development (WPD), Pooled pre-/post-Stage Development (PSD), Cumulative Stage Development (CSD) and Ordinal Logistic Regression (OLR) methods can all be used to determine the phenological progression of plants. Moreover, results show that a less detailed observation key still resulted in similar onset dates, unless more than two consecutive stages were omitted. Further results reveal that the simulation of a less intense sampling frequency had only small impacts on estimates of onset dates. Thus, especially in remote areas where an observation interval of a week is not feasible, estimates derived from the frequency distribution of BBCH codes appear to be appropriate.
DOIURL [本文引用: 1]
AbstractConfirming the results of previous regional studies on changes in first bloom dates (FBD) in China, this study provides evidence that complements conclusions drawn from studies of phenological changes in other dynamic climate systems in the Northern Hemisphere. Furthermore, increased occurrences of yearly second blooms (YSB) further reinforce results derived from other studies indicating a recent trend of generalized climate warming across China. Additionally, ascertaining changes in FBD and YSB against a recent background not only provides a hitherto poorly formulated autumnal equivalent to the well-studied shifts in FBD, but also formulates both spring and autumn flowering changes in recent years. Data in this study are derived from observations made from 1963 to 2006 by the Chinese Phenological Observation Network (CPON) nationwide system of monitoring stations that has made observations of over 173 species from across China since 1963. At each site, the mean value of each species annual deviation and spring mean surface temperatures were calculated. For each species, years and locations were also recorded for species in which second blooms (YSB) occurred. Of the 46 FBD samples, 31 showed advances from the mean, blooming earlier over the course of the study period. Notably, although only 8 of the 46 FBD samples showed significance levels of 0.1 or better, the average FBD did advance by 5.3 days. After the 1980s, the frequency of YSB occurrence remained steady, declining a little from the peak in the 1980s, but still exhibiting occurrences far more than were observed earlier. The data from this study clearly indicate that both the phenological advance of FBD in spring and the increased occurrence of YSB are consistent with climate warming.
DOIURLPMID [本文引用: 1]
Variation in the timing of plant phenology caused by phenotypic plasticity is a sensitive measure of how organisms respond to weather and climate variability. Although continental-scale gradients in climate and consequential patterns in plant phenology are well recognized, the contribution of underlying genotypic difference to the geography of phenology is less well understood. We hypothesize that different temperate plant genotypes require varying amount of heat energy for resuming annual growth and reproduction as a result of adaptation and other ecological and evolutionary processes along climatic gradients. In particular, at least for some species, the growing degree days (GDD) needed to trigger the same spring phenology events (e.g., budburst and flower bloom) may be less for individuals originated from colder climates than those from warmer climates. This variable intrinsic heat energy requirement in plants can be characterized by the term growth efficiency and is quantitatively reflected in the timing of phenophases arlier timing indicates higher efficiency (i.e., less heat energy needed to trigger phenophase transitions) and vice versa compared to a standard reference (i.e., either a uniform climate or a uniform genotype). In this study, we tested our hypothesis by comparing variations of budburst and bloom timing of two widely documented plants from the USA National Phenology Network (i.e., red maple- Acer rubrum and forsythia- Forsythia spp.) with cloned indicator plants (lilac- Syringa x chinensis ed Rothomagensis ) at multiple eastern US sites. Our results indicate that across the accumulated temperature gradient, the two non-clonal plants showed significantly more gradual changes than the cloned plants, manifested by earlier phenology in colder climates and later phenology in warmer climates relative to the baseline clone phenological response. This finding provides initial evidence supporting the growth efficiency hypothesis, and suggests more work is warranted. More studies investigating genotype-determined phenological variations will be useful for better understanding and prediction of the continental-scale patterns of biospheric responses to climate change.
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
Recent warming of Northern Hemisphere (NH) land is well documented and typically greater in winter/spring than other seasons. Physical environment responses to warming have been reported, but not details of large-area temperate growing season impacts, or consequences for ecosystems and agriculture. To date, hemispheric-scale measurements of biospheric changes have been confined to remote sensing. However, these studies did not provide detailed data needed for many investigations. Here, we show that a suite of modeled and derived measures (produced from daily maximum–minimum temperatures) linking plant development (phenology) with its basic climatic drivers provide a reliable and spatially extensive method for monitoring general impacts of global warming on the start of the growing season. Results are consistent with prior smaller area studies, confirming a nearly universal quicker onset of early spring warmth (spring indices (SI) first leaf date, 611.2 days decade 611 ), late spring warmth (SI first bloom date, 611.0 days decade 611 ; last spring day below 5°C, 611.4 days decade 611 ), and last spring freeze date (611.5 days decade 611 ) across most temperate NH land regions over the 1955–2002 period.
DOIURLPMID [本文引用: 1]
The phenophases first greening (bud burst) and yellowing of Nordic mountain birch ( Betula pubescens ssp. tortuosa , also called B . p . ssp. czerepanovii ) were observed at three sites on the Kola Peninsula in northernmost Europe during the period 1964–2003, and at two sites in the trans-boundary Pasvik-Enare region during 1994–2003. The field observations were compared with satellite images based on the GIMMS-NDVI dataset covering 1982–2002 at the start and end of the growing season. A trend for a delay of first greening was observed at only one of the sites (Kandalaksha) over the 4002year period. This fits well with the delayed onset of the growing season for that site based on satellite images. No significant changes in time of greening at the other sites were found with either field observations or satellite analyses throughout the study period. These results differ from the earlier spring generally observed in other parts of Europe in recent decades. In the coldest regions of Europe, e.g. in northern high mountains and the northernmost continental areas, increased precipitation associated with the generally positive North Atlantic Oscillation in the last few decades has often fallen as snow. Increased snow may delay the time of onset of the growing season, although increased temperature generally causes earlier spring phenophases. Autumn yellowing of birch leaves tends towards an earlier date at all sites. Due to both later birch greening and earlier yellowing at the Kandalaksha site, the growing season there has also become significantly shorter during the years observed. The sites showing the most advanced yellowing in the field throughout the study period fit well with areas showing an earlier end of the growing season from satellite images covering 1982–2002. The earlier yellowing is highly correlated with a trend at the sites in autumn for earlier decreasing air temperature over the study period, indicating that this environmental factor is important also for autumn phenophases.
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
针对当前园林景观营造的实际需求,借鉴相关学科的理念与优势,融合景观学与物候学2个交叉学科的基本理论,提出景观物候学概念,进而论述其时代背景、内涵、特点、研究途径及应用意义。景观物候学,既可以充分考虑景观学的观赏属性需求,又可以借用物候学的准确特质,定能极大提高园林植物造景的档次和精准度,推动园林事业更好更快发展。
URL [本文引用: 1]
针对当前园林景观营造的实际需求,借鉴相关学科的理念与优势,融合景观学与物候学2个交叉学科的基本理论,提出景观物候学概念,进而论述其时代背景、内涵、特点、研究途径及应用意义。景观物候学,既可以充分考虑景观学的观赏属性需求,又可以借用物候学的准确特质,定能极大提高园林植物造景的档次和精准度,推动园林事业更好更快发展。
DOIURL [本文引用: 1]
Based on the intrinsic vegetation landscape characteristics of Lianhuashan Park, Shenzhen city, the phenological change of dominant species and their effects on the landscape features were described and analyzed. The vegetation landscape characteristics of dominant communities, and the positive and negative relationships between vegetation landscape and phenological varieties were also discussed, and suggestions on the rebuilding of vegetation landscape were put forward.
DOIURL [本文引用: 1]
Based on the intrinsic vegetation landscape characteristics of Lianhuashan Park, Shenzhen city, the phenological change of dominant species and their effects on the landscape features were described and analyzed. The vegetation landscape characteristics of dominant communities, and the positive and negative relationships between vegetation landscape and phenological varieties were also discussed, and suggestions on the rebuilding of vegetation landscape were put forward.
[本文引用: 4]
[本文引用: 4]
DOIURLMagsci [本文引用: 3]
<p>木本植物物候相的更替鲜明地显示出自然景观外貌及其色彩的季节变化,成为园林设计中不容忽视的重要因素。在植物造景工作中,恰当地运用木本植物的物候相变化及其组合特征进行配植,可以增添空间构图的韵律,显示时间演变的节奏,协调不同时段之间景观季相的匹配关系,从而表现景观的时间与空间之美。为此,本文利用多年的物候观测资料,依据木本植物的展叶、开花、叶变色和落叶等物候相出现日期的早晚,对北京市植物园的70余种木本植物进行了物候相组合分类的研究。这一工作为比较客观地评价一个地区各种园林木本植物季相的观赏价值,认识其存在的主要问题,从物候学角度对造园木本植物材料进行类型划分,提供了一种有效的方法。</p>
DOIURLMagsci [本文引用: 3]
<p>木本植物物候相的更替鲜明地显示出自然景观外貌及其色彩的季节变化,成为园林设计中不容忽视的重要因素。在植物造景工作中,恰当地运用木本植物的物候相变化及其组合特征进行配植,可以增添空间构图的韵律,显示时间演变的节奏,协调不同时段之间景观季相的匹配关系,从而表现景观的时间与空间之美。为此,本文利用多年的物候观测资料,依据木本植物的展叶、开花、叶变色和落叶等物候相出现日期的早晚,对北京市植物园的70余种木本植物进行了物候相组合分类的研究。这一工作为比较客观地评价一个地区各种园林木本植物季相的观赏价值,认识其存在的主要问题,从物候学角度对造园木本植物材料进行类型划分,提供了一种有效的方法。</p>
[本文引用: 1]
[本文引用: 1]
URL
研究园林绿化树种在盐土地上的物候特征,不仅可以为盐土地的绿化确定最佳的造林时间,还可以为植物群落的 景观配置提供科学依据.本研究以上海崇明东滩湿地公园45种园林绿化树种为对象,对其萌芽和展叶物候进行了观测.结果表明:萌芽期从3月中旬持续到4月下 旬,其中22种集中在3月下旬,建议造林在4月底之前完成;展叶期从3月下旬持续到5月下旬,高峰期在4月中旬至5月上旬,可以根据树种展叶时间的早晚进 行景观配置;根据45种树种萌芽和展叶物候特征的差异,划分为4种类型,同一类型内树种具有相似的物候匹配关系,因此,在进行景观配置时,具有一定的相互 替代功能.最后给出了在植树造林和树种配置上的建议.
URL
研究园林绿化树种在盐土地上的物候特征,不仅可以为盐土地的绿化确定最佳的造林时间,还可以为植物群落的 景观配置提供科学依据.本研究以上海崇明东滩湿地公园45种园林绿化树种为对象,对其萌芽和展叶物候进行了观测.结果表明:萌芽期从3月中旬持续到4月下 旬,其中22种集中在3月下旬,建议造林在4月底之前完成;展叶期从3月下旬持续到5月下旬,高峰期在4月中旬至5月上旬,可以根据树种展叶时间的早晚进 行景观配置;根据45种树种萌芽和展叶物候特征的差异,划分为4种类型,同一类型内树种具有相似的物候匹配关系,因此,在进行景观配置时,具有一定的相互 替代功能.最后给出了在植树造林和树种配置上的建议.
DOIURL [本文引用: 1]
AimsMethodsImportant findings In comparison with the original natural calendar, the first dates of phenological spring and summer were advanced by two days and five days, respectively, while the first dates of autumn and winter were delayed by one day and four days, respectively. The lengths of summer and autumn were prolonged by six days and three days, while the lengths of spring and winter were shortened by three days and six days, respectively. The order of spring, autumn, winter phenophases changed with different degrees. The average, earliest, latest date of phenology showed advances mainly in spring and summer and delay in autumn and winter. Temperature change before the first date of spring, summer, winter and the sunshine duration before the first date of autumn are probably the main reason for the changes of phenological season in Beijing Summer Palace. Different responses of different species and phenophase result in change of phenophase order in seasons.
DOIURL [本文引用: 1]
AimsMethodsImportant findings In comparison with the original natural calendar, the first dates of phenological spring and summer were advanced by two days and five days, respectively, while the first dates of autumn and winter were delayed by one day and four days, respectively. The lengths of summer and autumn were prolonged by six days and three days, while the lengths of spring and winter were shortened by three days and six days, respectively. The order of spring, autumn, winter phenophases changed with different degrees. The average, earliest, latest date of phenology showed advances mainly in spring and summer and delay in autumn and winter. Temperature change before the first date of spring, summer, winter and the sunshine duration before the first date of autumn are probably the main reason for the changes of phenological season in Beijing Summer Palace. Different responses of different species and phenophase result in change of phenophase order in seasons.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
植物物候期的频率分布型及其在季节划分中的应用
6
1999
... 物候季节划分采用
... 本研究以
... 提出的频率分布型法为基本方法, 以120种园林树种的15种物候期组成的混合样本为数据基础, 对2017年北京的自然季节进行了划分, 结果如下: 6-19候为春(6-9候为初春, 10-15候为仲春, 16-19候为暮春); 20-45候为夏; 46-59候为秋(46-47候为初秋, 48-56候为仲秋, 57-59候为暮秋, 60候至次年春季开始前为冬.
... Pentad-average cumulative frequency of phenophases = Cumulative frequency of phenophases / Duration pentads. The meteorological data of 2017 in the table come from Chinese Meteorological Data Network (data.cma.cn), and all the data of 1980’s come from
... 物候期候均发生频率=物候期累积频率/持续候期.表中2017年气象数据来自中国气象数据网(data.cma.cn), 1980’s数据来自
... 在季节物候特征的研究方面, 杨国栋及陈效逑等人从物候角度对北京植物季相特征进行了定性归纳总结(
植物物候期的频率分布型及其在季节划分中的应用
6
1999
... 物候季节划分采用
... 本研究以
... 提出的频率分布型法为基本方法, 以120种园林树种的15种物候期组成的混合样本为数据基础, 对2017年北京的自然季节进行了划分, 结果如下: 6-19候为春(6-9候为初春, 10-15候为仲春, 16-19候为暮春); 20-45候为夏; 46-59候为秋(46-47候为初秋, 48-56候为仲秋, 57-59候为暮秋, 60候至次年春季开始前为冬.
... Pentad-average cumulative frequency of phenophases = Cumulative frequency of phenophases / Duration pentads. The meteorological data of 2017 in the table come from Chinese Meteorological Data Network (data.cma.cn), and all the data of 1980’s come from
... 物候期候均发生频率=物候期累积频率/持续候期.表中2017年气象数据来自中国气象数据网(data.cma.cn), 1980’s数据来自
... 在季节物候特征的研究方面, 杨国栋及陈效逑等人从物候角度对北京植物季相特征进行了定性归纳总结(
Temporal coherence of phenological and climatic rhythmicity in Beijing
3
2017
... 北京的物候季节与30年前相比, 入春时间提早1候, 夏季延长4候, 入秋时间延后3候, 秋季缩短2候, 不同季节持续期长短的差距加大; 而其他相关研究也表明, 在北半球的中高纬度地区, 近年来受全球变暖影响, 植物春季物候提前, 秋季物候推迟, 生长季变长等, 已成为日益普遍的现象(
... 物候数据样本量与观测年份是影响本研究物候季节划分结果可靠性的主要因素.本研究虽然仅基于一年的物候数据划分季节并探讨各季节内植物物候期的变化特征, 无法代表多年平均的物候季节特征, 但实现了通过一年快速、详细地群落植物物候观测, 获取当年植物群落物候季节和景观季相的信息, 具有显著的时效性, 可为下一年物候季节的划分以及植物园季节性旅游活动的设计提供专业化的指导和参考.此外, 北京的物候具有准年周期性, 即同一物候期在相邻年份间的发生时距大约为1年, 其平均年际波动维持在一定幅度之内(5-16天)(
... ;
A comparison of methods to estimate seasonal phenological development from BBCH scale recording
1
2011
... 本研究的物候观测以《中国物候观测方法》(
Advances in first bloom dates and increased occurrences of yearly second blooms in eastern China since the 1960s: Further phenological evidence of climate warming
1
2011
... 北京的物候季节与30年前相比, 入春时间提早1候, 夏季延长4候, 入秋时间延后3候, 秋季缩短2候, 不同季节持续期长短的差距加大; 而其他相关研究也表明, 在北半球的中高纬度地区, 近年来受全球变暖影响, 植物春季物候提前, 秋季物候推迟, 生长季变长等, 已成为日益普遍的现象(
Testing a growth efficiency hypothesis with continental-scale phenological variations of common and cloned plants
1
2014
... 物候数据样本量与观测年份是影响本研究物候季节划分结果可靠性的主要因素.本研究虽然仅基于一年的物候数据划分季节并探讨各季节内植物物候期的变化特征, 无法代表多年平均的物候季节特征, 但实现了通过一年快速、详细地群落植物物候观测, 获取当年植物群落物候季节和景观季相的信息, 具有显著的时效性, 可为下一年物候季节的划分以及植物园季节性旅游活动的设计提供专业化的指导和参考.此外, 北京的物候具有准年周期性, 即同一物候期在相邻年份间的发生时距大约为1年, 其平均年际波动维持在一定幅度之内(5-16天)(
Growth Stages of Mono- and Dicotyledonous Plants—BBCH Monograph
1
2001
... 本研究的物候观测以《中国物候观测方法》(
Phenology: An Integrative Environmental Science
1
2013
... 北京四季分明, 以暖温带落叶阔叶林为典型地域植被类型, 植物景观具有明显的季节性特征, 即季相.物候学主要是研究自然界植物、动物和环境条件的周期变化之间相互关系的科学(
Onset of spring startingearlier across the northern hemisphere
1
2006
... 北京的物候季节与30年前相比, 入春时间提早1候, 夏季延长4候, 入秋时间延后3候, 秋季缩短2候, 不同季节持续期长短的差距加大; 而其他相关研究也表明, 在北半球的中高纬度地区, 近年来受全球变暖影响, 植物春季物候提前, 秋季物候推迟, 生长季变长等, 已成为日益普遍的现象(
Growing seasons of Nordic mountain birch in northernmost Europe as indicated by long-term field studies and analyses of satellite images
1
2006
... 北京的物候季节与30年前相比, 入春时间提早1候, 夏季延长4候, 入秋时间延后3候, 秋季缩短2候, 不同季节持续期长短的差距加大; 而其他相关研究也表明, 在北半球的中高纬度地区, 近年来受全球变暖影响, 植物春季物候提前, 秋季物候推迟, 生长季变长等, 已成为日益普遍的现象(
1
1979
... 本研究的物候观测以《中国物候观测方法》(
1
1979
... 本研究的物候观测以《中国物候观测方法》(
试论园林植物景观物候学
1
2004
... 季节性是北京植物景观的典型特征, 而个体物候是植物景观季节动态的基础.关于物候在植物景观研究中的应用, 许多****从不同地域特征和应用方向已进行了一定的研究(
试论园林植物景观物候学
1
2004
... 季节性是北京植物景观的典型特征, 而个体物候是植物景观季节动态的基础.关于物候在植物景观研究中的应用, 许多****从不同地域特征和应用方向已进行了一定的研究(
深圳莲花山公园景观植物物候特点及对公园景观的影响
1
2004
... 季节性是北京植物景观的典型特征, 而个体物候是植物景观季节动态的基础.关于物候在植物景观研究中的应用, 许多****从不同地域特征和应用方向已进行了一定的研究(
深圳莲花山公园景观植物物候特点及对公园景观的影响
1
2004
... 季节性是北京植物景观的典型特征, 而个体物候是植物景观季节动态的基础.关于物候在植物景观研究中的应用, 许多****从不同地域特征和应用方向已进行了一定的研究(
4
1995
... 物候数据样本量与观测年份是影响本研究物候季节划分结果可靠性的主要因素.本研究虽然仅基于一年的物候数据划分季节并探讨各季节内植物物候期的变化特征, 无法代表多年平均的物候季节特征, 但实现了通过一年快速、详细地群落植物物候观测, 获取当年植物群落物候季节和景观季相的信息, 具有显著的时效性, 可为下一年物候季节的划分以及植物园季节性旅游活动的设计提供专业化的指导和参考.此外, 北京的物候具有准年周期性, 即同一物候期在相邻年份间的发生时距大约为1年, 其平均年际波动维持在一定幅度之内(5-16天)(
... ).其次, 物候现象的发生具有顺序相关性, 即各物候现象每年都按一定先后顺序出现, 且在一定时段内具有大体同步提前或延后的特点(
... 在季节物候特征的研究方面, 杨国栋及陈效逑等人从物候角度对北京植物季相特征进行了定性归纳总结(
... 在物候期的时间分布特征方面, 前人多基于物候特征对植物进行分组分类,
4
1995
... 物候数据样本量与观测年份是影响本研究物候季节划分结果可靠性的主要因素.本研究虽然仅基于一年的物候数据划分季节并探讨各季节内植物物候期的变化特征, 无法代表多年平均的物候季节特征, 但实现了通过一年快速、详细地群落植物物候观测, 获取当年植物群落物候季节和景观季相的信息, 具有显著的时效性, 可为下一年物候季节的划分以及植物园季节性旅游活动的设计提供专业化的指导和参考.此外, 北京的物候具有准年周期性, 即同一物候期在相邻年份间的发生时距大约为1年, 其平均年际波动维持在一定幅度之内(5-16天)(
... ).其次, 物候现象的发生具有顺序相关性, 即各物候现象每年都按一定先后顺序出现, 且在一定时段内具有大体同步提前或延后的特点(
... 在季节物候特征的研究方面, 杨国栋及陈效逑等人从物候角度对北京植物季相特征进行了定性归纳总结(
... 在物候期的时间分布特征方面, 前人多基于物候特征对植物进行分组分类,
木本植物物候相组合分类研究——以北京市植物园栽培树种为例
3
2000
... 叶幕期从展叶始期至落叶末期, 代表植物叶片具有景观观赏性的时期(包括展叶期、秋色期和落叶期以及夏季漫长的叶色稳定期); 绿色期指从展叶始期至秋季叶色始变期(
... 季节性是北京植物景观的典型特征, 而个体物候是植物景观季节动态的基础.关于物候在植物景观研究中的应用, 许多****从不同地域特征和应用方向已进行了一定的研究(
... 在物候期的时间分布特征方面, 前人多基于物候特征对植物进行分组分类,
木本植物物候相组合分类研究——以北京市植物园栽培树种为例
3
2000
... 叶幕期从展叶始期至落叶末期, 代表植物叶片具有景观观赏性的时期(包括展叶期、秋色期和落叶期以及夏季漫长的叶色稳定期); 绿色期指从展叶始期至秋季叶色始变期(
... 季节性是北京植物景观的典型特征, 而个体物候是植物景观季节动态的基础.关于物候在植物景观研究中的应用, 许多****从不同地域特征和应用方向已进行了一定的研究(
... 在物候期的时间分布特征方面, 前人多基于物候特征对植物进行分组分类,
1
1991
... 季节性是北京植物景观的典型特征, 而个体物候是植物景观季节动态的基础.关于物候在植物景观研究中的应用, 许多****从不同地域特征和应用方向已进行了一定的研究(
1
1991
... 季节性是北京植物景观的典型特征, 而个体物候是植物景观季节动态的基础.关于物候在植物景观研究中的应用, 许多****从不同地域特征和应用方向已进行了一定的研究(
上海崇明岛园林绿化树种的春季物候特征与物候相分类
2013
上海崇明岛园林绿化树种的春季物候特征与物候相分类
2013
近30年北京自然历的主要物候期、物候季节及归因
1
2012
... 北京的物候季节与30年前相比, 入春时间提早1候, 夏季延长4候, 入秋时间延后3候, 秋季缩短2候, 不同季节持续期长短的差距加大; 而其他相关研究也表明, 在北半球的中高纬度地区, 近年来受全球变暖影响, 植物春季物候提前, 秋季物候推迟, 生长季变长等, 已成为日益普遍的现象(
近30年北京自然历的主要物候期、物候季节及归因
1
2012
... 北京的物候季节与30年前相比, 入春时间提早1候, 夏季延长4候, 入秋时间延后3候, 秋季缩短2候, 不同季节持续期长短的差距加大; 而其他相关研究也表明, 在北半球的中高纬度地区, 近年来受全球变暖影响, 植物春季物候提前, 秋季物候推迟, 生长季变长等, 已成为日益普遍的现象(
1
1963
... 北京四季分明, 以暖温带落叶阔叶林为典型地域植被类型, 植物景观具有明显的季节性特征, 即季相.物候学主要是研究自然界植物、动物和环境条件的周期变化之间相互关系的科学(
1
1963
... 北京四季分明, 以暖温带落叶阔叶林为典型地域植被类型, 植物景观具有明显的季节性特征, 即季相.物候学主要是研究自然界植物、动物和环境条件的周期变化之间相互关系的科学(
1
1963
... 北京四季分明, 以暖温带落叶阔叶林为典型地域植被类型, 植物景观具有明显的季节性特征, 即季相.物候学主要是研究自然界植物、动物和环境条件的周期变化之间相互关系的科学(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}