Supported by the Zhejiang Provincial Natural Science Foundation of China.LY14C030003 Supported by the Zhejiang Provincial Natural Science Foundation of China.LY18C020002
Abstract Aims Darwin’s naturalization conundrum describes the paradox that the relationship of exotic species to native residents could either promote or hinder invasion success through opposing mechanisms: niche pre-adaptation or competitive interactions. Previous Darwin’s naturalization studies have showed invasion success could vary at stages, sites, and spatial and phylogenetic scales. Our objective was to assess the effects of exotic-native species relationship on invasion process of exotic plant species in China, where related research is still lacking. Methods Generalized linear mixed models were used to examine relationship between exotic-native species relationship and performance of exotic species at different spatial scale (provincial, municipal and community) and invasion stages (naturalization, dispersal and invasion). At community scale, we measured environmental factors of communities we investigated to control the effect of habitat heterogeneity among them. Important findings At the provincial and municipal scales, exotic species closely related to native flora were more likely to be naturalized and distributed, which is more consistent with the expectation of the pre-adaptation hypothesis. On the community scale, the exotic-native species relationship was not related to establishment and abundance of exotic species in the community. The results suggested that exotic species did not strongly compete with their close native relatives in communities, but were better adapted to areas where their close relatives had lived. Considering their high potential of naturalization and invasion, special attention should be paid to those exotic species that closely related to the native flora in the management of invasive species. Keywords:phylogenetic distance;scale;niche overlap;habitat filtering;stages of invasion
PDF (979KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 郑珊珊, 陈旭波, 许微楠, 骆争荣, 夏更寿. 外来种-本地种亲缘关系对外来植物归化和入侵的影响. 植物生态学报[J], 2018, 42(10): 990-999 DOI:10.17521/cjpe.2018.0101 ZHENG Shan-Shan, CHEN Xu-Bo, XU Wei-Nan, LUO Zheng-Rong, XIA Geng-Shou. Effects of exotic-native species relationship on naturalization and invasion of exotic plant species. Chinese Journal of Plant Ecology[J], 2018, 42(10): 990-999 DOI:10.17521/cjpe.2018.0101
2014-2016年(每年的5-12月), 我们在浙江省丽水市莲都区(1 502.1 km2, 年降水量1 395.5 mm, 年平均气温18.5 ℃)的乡村路边、弃耕地、溪边、林缘等处调查了109个1 m × 1 m的植物群落样方。我们在野外收割每个样方的所有地上部分植物材料, 带回实验室后对他们进行归类、称鲜质量、计数并鉴定。物种定名根据《Flora of China》(Editorial Committee of Flora of China, 2013)。
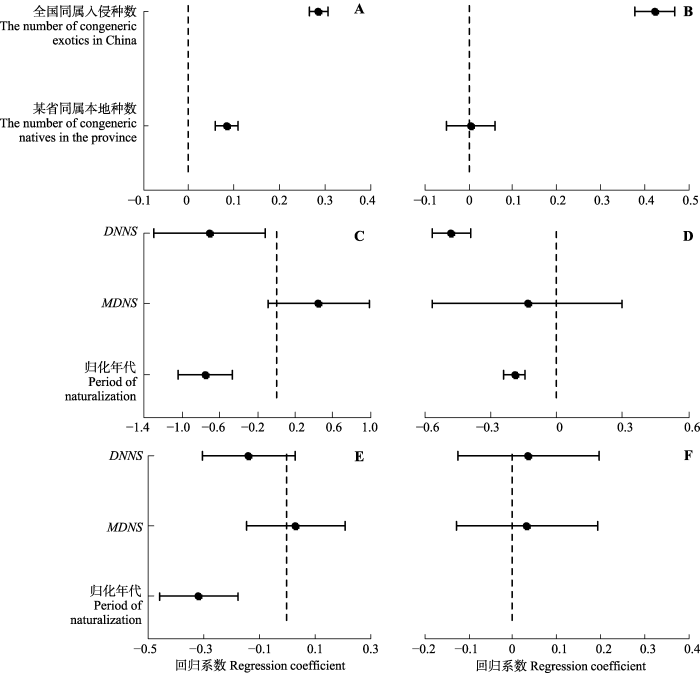
Fig. 1The effects of the number of congeneric natives, phylogenetic distance to the nearest native species (DNNS) and mean phylogenetic distance (MDNS) to all native species in the same community (or flora) on naturalization, dispersal and invasion at different scales. A, Effects on the number of exotics naturalized in a province. B, Effects on the number of exotics naturalized in a province without double-zero records. C, Effects on probability of exotics successfully naturalized in Wenzhou. D, Effects on frequency of exotics naruralized in Wenzhou. E, Effects on exotics successfully invade in communities we examined. F, Effects on invasive species abundance in communities we examined. Solid line segments are 95% credible intervals of parameter distribution. Dash lines are zero lines. If a solid line intersects with a dash line, it means the corresponding coefficient was not significant. Period of naturalization, evaluated at the species level, ranges from 1 to 5, with 5 being more recent naturalization, so a negative coefficient indicates that species introduced more recently perform inferior.
由于达尔文归化假说以近缘种间具有较大的生态位重叠以及随之而来的竞争为理论基础, 生态学家一般认为该假说主要在小尺度上表现(Diez et al., 2008; Thuiller et al., 2010; Li et al., 2015; Ma et al., 2016)。然而, 我们的研究结果表明, 在群落尺度上控制生境差异后, 外来种-本地种亲缘距离与外来种在群落中侵入成功与否及其危害程度无关。这种差异可能与本研究的谱系尺度有关(Castro et al., 2014)。Thuiller等(2010)认为如果谱系尺度适中(如与本地种间的谱系距离接近), 外来种和本地种会因生态位分化而实现共存, 且不受空间尺度影响。我们比较了34种外来植物对各群落的DNNS值和每个本地种与其所在群落内其他本地种之间的最近谱系距离(NTD), 并发现在这一尺度上DNNS值的变异范围略小于NTD值的变异范围(附录II)。这说明本研究调查的群落中, 外来种-本地种谱系尺度接近本地植物间真实的谱系距离。莲都区的外来植物在生态位分化上已经和本地种类似; 外来种和本地种之间保持了合理的生态位分化, 从而保证了这些外来植物和本地植物能够长期共存。
总体上看, 本研究的结果表明, 在同一尺度下外来种-本地种亲缘关系对整个入侵过程的影响没有因入侵阶段不同而不同。在温州, 外来种-本地种亲缘关系较近不仅有利于外来种归化也有利于其扩散; 而在莲都区的群落中, 外来种-本地种亲缘关系对外来种的侵入和危害程度都无影响。这和来自世界其他地方的研究结果不太一致(Diez et al., 2008; Bezeng et al., 2015; Li et al., 2015), 说明中国地区的外来种-本地种亲缘关系对入侵过程的影响可能具有自身特点。
和前人的研究结果类似, 本研究的结果显示无论在区域还是局域尺度上, 无论是归化、扩散还是危害程度, MDNS与外来种入侵过程都没有显著关系(Carboni et al., 2013; Skóra et al., 2015)。理论上来说, 在群落中(尤其是多样性较高的群落)很少会存在极其相似或差异极大的物种, 大部分物种仅存在中等程度的功能差异(Skóra et al., 2015)。Skóra等(2015)指出在不同情况下, 外来种可能仅和少量本地种近似也可能与许多本地种近似, 从而使MDNS的变异非常大; 当多样性增加时, MDNS会变得更加不确定。另一方面, 达尔文提出的归化假说和预适应假说表述的都是外来种归化与近缘物种(同属物种)存在与否的关系(Darwin, 1859), 而MDNS衡量的是外来种与整个本地群落(或区系)所有物种的平均谱系距离。因此, MDNS可能是一个弱指标, 在达尔文归化难题研究中, 应该与其他谱系距离的测度指标结合使用, 以确保准确反映外来种-本地种亲缘关系对外来植物入侵过程的影响。
BakerHG ( 1965). Characteristics and modes of origin of weeds In: Baker HG, Stebbins GL eds. The Genetics of Colonizing Species. Academic Press, New York. 147-172. [本文引用: 1]
BezengSB, DaviesJT, YessoufouK, MaurinO, van der BankM ( 2015). Revisiting Darwin’s naturalization conundrum: Explaining invasion success of non-native trees and shrubs in southern Africa Journal of Ecology, 103, 871-879. DOIURL [本文引用: 4]
CampbellSE, MandrakNE ( 2017). Dissecting spatiotemporal patterns of functional diversity through the lens of Darwin’s naturalization conundrum Ecology and Evolution, 7, 3861-3869. DOIURL [本文引用: 5]
CastroSA, Escobedo1VM, ArandaJ, CarvalloGO ( 2014). Evaluating Darwin’s naturalization hypothesis in experimental plant assemblages: Phylogenetic relationships do not determine colonization success PLOS ONE, 9, e105535. DOI: 10.1371/journal.pone.0105535. DOIURL [本文引用: 1]
CayuelaL, Granzow-de la CerdaI, AlbuquerqueFS, GolicherJD ( 2012). Taxonstand: An R package for species names standardization in vegetation databases Methods in Ecology and Evolution, 3, 1078-1083. DOIURL [本文引用: 1]
ChessonP, HuntlyN ( 1997). The roles of harsh and fluctuating conditions in the dynamics of ecological communites The American Naturalist, 150, 519-553. DOIURL [本文引用: 1]
DaehlerCC ( 2001). Darwin’s naturalization hypothesis revisited The American Naturalist, 158, 324-330. DOIURL [本文引用: 2]
DarwinC ( 1859). On the Origin of Species John Murray, London. [本文引用: 2]
Editorial Committee of Flora of China ( 2013). Flora of China. Science Press, Beijing. [本文引用: 2]
FengJM, DongXD, XuCD ( 2009). Spatial patterns of floristic composition of invasive alien plants in large scale and their climatic interpretation Journal of Wuhan Botanical Research, 27, 159-164. [本文引用: 3]
GaoM, HuRY, ChenXX, LiWC, DingBY ( 2011). Effects of disturbance, topography, and soil conditions on the distribution of invasive plants in Wenzhou Biodiversity Science, 19, 424-431.
GongWN, XieBY, WanFH ( 2011). Basic theory of phenotype plasticity and adaptive evolution in invasive species In: Wan FH, Xie BY, Yang GQ eds. Invasion Biology. Science Press, Beijing. 124-138. [本文引用: 1]
HuRY, DingBY, ChenXX, CaiYB, LeiZP ( 2011). Study on species composition and flora characteristics of alien invasive plants in Wenzhou Journal of Wenzhou University (Natural Sciences), 32(3), 18-25. [本文引用: 1]
HuangQQ ( 2009). Patterns of Plant Invasions in China: Role of Species Traits, Environment and Human Activities. PhD dissertation, Zhejiang University, Hangzhou. 46-54. [本文引用: 5]
JonesEI, NuismerSL, GomulkiewiczR ( 2013). Revisiting Darwin’s conundrum reveals a twist on the relationship between phylogenetic distance and invasibility Proceedings of the National Academy of Sciences of the United States of America, 110, 20627-20632. DOIURL [本文引用: 5]
LiS, CadotteMW, MeinersSJ, HuaZ, ShuH, LiJ, ShuW ( 2015). The effects of phylogenetic relatedness on invasion success and impact: Deconstructing Darwin’s naturalisation conundrum Ecology Letters, 18, 1285-1292. DOIURL [本文引用: 5]
MaC, LiS, PuZ, TanJ, LiuM, ZhouJ, LiH, JiangL ( 2016). Different effects of invader-native phylogenetic relatedness on invasion success and impact: A meta-analysis of Darwin’s naturalization hypothesis Proceedings of the Royal Society B: Biological Sciences, 283, 20160663. DOI: 10.1098/rspb.2016.0663. DOIURL
MaJS ( 2013). The Checklist of the Chinese Invasive Plants. Higher Education Press, Beijing. [本文引用: 4]
MackRN, SimberloffD, LonsdaleWM, EvansH, CloutM, BazzazFA ( 2000). Biotic invasions: Causes, epidemiology, global consequences, and control Ecological Application, 10, 689-710. DOIURL [本文引用: 1]
MarxHE, GiblinDE, DunwiddiePW, TankDC ( 2016). Deconstructing Darwin’s naturalization conundrum in the San Juan Islands using community phylogenetics and functional traits Diversity and Distributions, 22, 318-331. DOIURL
NathanJB, KraftB, AckerlyDD ( 2008). Functional traits and nich-based tree community assembly in an Amazonian forest Science, 322, 580-582. DOIURL [本文引用: 1]
National Agricultural Technology Extension Service Center ( 2006). The Technical Manual of Soil Analysis. China Agriculture Press, Beijing. [本文引用: 1]
PejcharL, MooneyHA ( 2009). Invasive species, ecosystem services and human well-being Trends in Ecology and Evolution, 24, 497-504. DOIURL [本文引用: 1]
QianH, JinY ( 2015). An updated megaphylogeny of plants, a tool for generating plant phylogenies, and an analysis of phylogenetic community structure Journal of Plant Ecology, 9, 233-239. [本文引用: 2]
RejmanekM, RichardsonDM ( 1996). What attributes make some plant species more invasive? Ecology, 77, 1655-1661. DOIURL [本文引用: 1]
RejmanekM, RichardsonDM, PysekP ( 2013). Plant invasions and invasibility of plant communities In: van der Maarel E, Franklin J eds. Vegetation Ecology. 2nd edn. John Wiley & Sons, New York. 387-424. [本文引用: 2]
RevellLJ ( 2012). Phytools: An R package for phylogenetic comparative biology (and other things) Methods in Ecology and Evolution, 3, 217-223. DOIURL [本文引用: 3]
RichardsonDM, PysekP ( 2012). Naturalization of introduced plants: Ecological drivers of biogeographical patterns New Phytologist, 196, 383-396. DOIURL [本文引用: 1]
SheaK, ChessonP ( 2002). Community ecology theory as a framework for biological invasion Trends in Ecology and Evolution, 17, 170-176. DOIURL [本文引用: 3]
SkóraF, AbilhoaV, PadialAA, VituleJRS ( 2015). Darwin’s hypotheses to explain colonization trends: Evidence from a quasi-natural experiment and a new conceptual model Diversity and Distributions, 21, 583-594. DOIURL [本文引用: 2]
StraussSY, WebbCO, SalaminN ( 2006). Exotic taxa less related to native species are more invasive Proceedings of the National Academy of Science of the United States of America, 103, 5841-5845. DOIURL [本文引用: 12]
ThuillerW, GallienL, BoulangeatI, de BelloF, MunkemullerT, RoquetC, LavergneS ( 2010). Resolving Darwin’s naturalization conundrum: A quest for evidence Diversity and Distributions, 16, 461-475. DOIURL [本文引用: 2]
WuSH, SunHT, TengYC, RejmanekM, ChawSM, YangTYA, HsiehCF ( 2010). Patterns of plant invasions in China: Taxonomic, biogeographic, climatic approaches and anthropogenic effects Biological Invasions, 12, 2179-2206. DOI [本文引用: 1]
WuZY, SunH, ZhouZK, LiDZ, PengH ( 2010). Floristics of Seed Plants from China. Science Press, Beijing. [本文引用: 2]
YanXL, LiuQR, ShouHY, ZengXF, ZhangY, ChenL, LiuY, MaHY, QiSY, MaJS ( 2014 a). The categorization and analysis on the geographic distribution patterns of Chinese alien invasive plants Biodiversity Science, 22, 667-676. [本文引用: 2]
Characteristics and modes of origin of weeds 1 1965
... 随着全球经济一体化进程的加快, 生物扩散的自然屏障被打破, 地区间物种交换的比例和空间范围大大增加(Campbell & Mandrak, 2017).部分外来物种到达新的分布区域后在生态、经济和社会等方面给当地造成了严重的后果(Mack et al., 2000; Pejchar & Mooney, 2009).面对生物入侵的严峻挑战, 预测外来种的潜在入侵性和生态影响已经成为生态学家需要解决的重要研究议题(Mack et al., 2000).早期的研究从归纳入侵种的功能特征(特别是资源利用、生殖潜力和扩散能力有关的特征)入手, 总结入侵物种的共性特征(Baker, 1965; Rejmanek & Richardson, 1996).然而, 这些方法仍不能得出一致的结论, 与高入侵性有关的生活史特征在不同类群和生境中存在很大差异(Bezeng et al., 2015; Li et al., 2015).而且一些重要的功能性状很难鉴别和测量, 需要投入大量的人力和物力(Bezeng et al., 2015). ...
Revisiting Darwin’s naturalization conundrum: Explaining invasion success of non-native trees and shrubs in southern Africa 4 2015
... 随着全球经济一体化进程的加快, 生物扩散的自然屏障被打破, 地区间物种交换的比例和空间范围大大增加(Campbell & Mandrak, 2017).部分外来物种到达新的分布区域后在生态、经济和社会等方面给当地造成了严重的后果(Mack et al., 2000; Pejchar & Mooney, 2009).面对生物入侵的严峻挑战, 预测外来种的潜在入侵性和生态影响已经成为生态学家需要解决的重要研究议题(Mack et al., 2000).早期的研究从归纳入侵种的功能特征(特别是资源利用、生殖潜力和扩散能力有关的特征)入手, 总结入侵物种的共性特征(Baker, 1965; Rejmanek & Richardson, 1996).然而, 这些方法仍不能得出一致的结论, 与高入侵性有关的生活史特征在不同类群和生境中存在很大差异(Bezeng et al., 2015; Li et al., 2015).而且一些重要的功能性状很难鉴别和测量, 需要投入大量的人力和物力(Bezeng et al., 2015). ...
... ).而且一些重要的功能性状很难鉴别和测量, 需要投入大量的人力和物力(Bezeng et al., 2015). ...
... 前人针对达尔文归化难题的研究得到的矛盾结果常被归因于这些研究在研究区域、类群以及在概念框架和研究方法(如不同的空间和谱系尺度、入侵阶段、量度的参数和生境条件)上存在巨大差异(Thuiller et al., 2010; Richardson & Pysek, 2012; Li et al., 2015).空间尺度和入侵阶段被认为对入侵过程的生态学机制具有重要的影响(Ma et al., 2016).****们普遍认为达尔文归化假说主要在较小空间尺度和入侵危害(或扩散)阶段起作用, 而预适应假说可能在大尺度和归化阶段起主导作用(Diez et al., 2008; Thuiller et al., 2010; Bezeng et al., 2015; Li et al., 2015), 但同时也存在很多不同意见(Jones et al., 2013; Ma et al., 2016; Campbell & Mandrak, 2017).由于外来种和本地种间稳定共存的潜力并不是随谱系距离线性(而是S形曲线)增加的, 因此研究对象间的谱系距离取值范围(即谱系尺度)对外来种和本地种互作的结果也具有重要影响(Thuiller et al., 2010).另外, 外来种的入侵过程常受多种因素影响, 其中生境条件起着基础性作用.生境条件不仅影响外来种的入侵过程, 而且同样影响本地物种的组成(Chesson & Huntly, 1997; Shea & Chesson, 2002; Richardson & Pysek, 2012), 从而影响外来种-本地种亲缘与外来种归化、扩散和危害的关系.虽然目前有不少****已经在同一研究中考虑了空间尺度和入侵阶段的影响, 但在调查性实证研究中仍很少考虑谱系尺度和生境条件差异. ...
... 总体上看, 本研究的结果表明, 在同一尺度下外来种-本地种亲缘关系对整个入侵过程的影响没有因入侵阶段不同而不同.在温州, 外来种-本地种亲缘关系较近不仅有利于外来种归化也有利于其扩散; 而在莲都区的群落中, 外来种-本地种亲缘关系对外来种的侵入和危害程度都无影响.这和来自世界其他地方的研究结果不太一致(Diez et al., 2008; Bezeng et al., 2015; Li et al., 2015), 说明中国地区的外来种-本地种亲缘关系对入侵过程的影响可能具有自身特点. ...
Dissecting spatiotemporal patterns of functional diversity through the lens of Darwin’s naturalization conundrum 5 2017
... 随着全球经济一体化进程的加快, 生物扩散的自然屏障被打破, 地区间物种交换的比例和空间范围大大增加(Campbell & Mandrak, 2017).部分外来物种到达新的分布区域后在生态、经济和社会等方面给当地造成了严重的后果(Mack et al., 2000; Pejchar & Mooney, 2009).面对生物入侵的严峻挑战, 预测外来种的潜在入侵性和生态影响已经成为生态学家需要解决的重要研究议题(Mack et al., 2000).早期的研究从归纳入侵种的功能特征(特别是资源利用、生殖潜力和扩散能力有关的特征)入手, 总结入侵物种的共性特征(Baker, 1965; Rejmanek & Richardson, 1996).然而, 这些方法仍不能得出一致的结论, 与高入侵性有关的生活史特征在不同类群和生境中存在很大差异(Bezeng et al., 2015; Li et al., 2015).而且一些重要的功能性状很难鉴别和测量, 需要投入大量的人力和物力(Bezeng et al., 2015). ...
... 前人针对达尔文归化难题的研究得到的矛盾结果常被归因于这些研究在研究区域、类群以及在概念框架和研究方法(如不同的空间和谱系尺度、入侵阶段、量度的参数和生境条件)上存在巨大差异(Thuiller et al., 2010; Richardson & Pysek, 2012; Li et al., 2015).空间尺度和入侵阶段被认为对入侵过程的生态学机制具有重要的影响(Ma et al., 2016).****们普遍认为达尔文归化假说主要在较小空间尺度和入侵危害(或扩散)阶段起作用, 而预适应假说可能在大尺度和归化阶段起主导作用(Diez et al., 2008; Thuiller et al., 2010; Bezeng et al., 2015; Li et al., 2015), 但同时也存在很多不同意见(Jones et al., 2013; Ma et al., 2016; Campbell & Mandrak, 2017).由于外来种和本地种间稳定共存的潜力并不是随谱系距离线性(而是S形曲线)增加的, 因此研究对象间的谱系距离取值范围(即谱系尺度)对外来种和本地种互作的结果也具有重要影响(Thuiller et al., 2010).另外, 外来种的入侵过程常受多种因素影响, 其中生境条件起着基础性作用.生境条件不仅影响外来种的入侵过程, 而且同样影响本地物种的组成(Chesson & Huntly, 1997; Shea & Chesson, 2002; Richardson & Pysek, 2012), 从而影响外来种-本地种亲缘与外来种归化、扩散和危害的关系.虽然目前有不少****已经在同一研究中考虑了空间尺度和入侵阶段的影响, 但在调查性实证研究中仍很少考虑谱系尺度和生境条件差异. ...
... 外来种-本地种谱系关系或分类关系对生物入侵和归化的影响是近20年来入侵生态学中争论最多的问题之一(Thuiller et al., 2010; Ma et al., 2016).前人的许多研究表明在较大尺度(如区域)上, 与本地种亲缘关系较近的外来种更易归化或入侵; 而在较小尺度(如局域或者同一生境水平)上, 成功归化或入侵的外来种与本地种的亲缘关系较远(Diez et al., 2008; Carboni et al., 2013; Campbell & Mandrak, 2017).我们多尺度多入侵阶段的研究表明, 在省级和市级(区域)尺度上, 与本地种亲缘关系较近的外来植物更有可能在当地归化和扩散, 符合预适应假说的预期; 而在群落(局域)尺度上, 外来种-本地种亲缘距离与外来种是否在群落中成功定居及其危害程度无关. ...
... 和前人的研究结果类似, 本研究的结果显示无论在区域还是局域尺度上, 无论是归化、扩散还是危害程度, MDNS与外来种入侵过程都没有显著关系(Carboni et al., 2013; Skóra et al., 2015).理论上来说, 在群落中(尤其是多样性较高的群落)很少会存在极其相似或差异极大的物种, 大部分物种仅存在中等程度的功能差异(Skóra et al., 2015).Skóra等(2015)指出在不同情况下, 外来种可能仅和少量本地种近似也可能与许多本地种近似, 从而使MDNS的变异非常大; 当多样性增加时, MDNS会变得更加不确定.另一方面, 达尔文提出的归化假说和预适应假说表述的都是外来种归化与近缘物种(同属物种)存在与否的关系(Darwin, 1859), 而MDNS衡量的是外来种与整个本地群落(或区系)所有物种的平均谱系距离.因此, MDNS可能是一个弱指标, 在达尔文归化难题研究中, 应该与其他谱系距离的测度指标结合使用, 以确保准确反映外来种-本地种亲缘关系对外来植物入侵过程的影响. ...
Evaluating Darwin’s naturalization hypothesis in experimental plant assemblages: Phylogenetic relationships do not determine colonization success 1 2014
... 由于达尔文归化假说以近缘种间具有较大的生态位重叠以及随之而来的竞争为理论基础, 生态学家一般认为该假说主要在小尺度上表现(Diez et al., 2008; Thuiller et al., 2010; Li et al., 2015; Ma et al., 2016).然而, 我们的研究结果表明, 在群落尺度上控制生境差异后, 外来种-本地种亲缘距离与外来种在群落中侵入成功与否及其危害程度无关.这种差异可能与本研究的谱系尺度有关(Castro et al., 2014).Thuiller等(2010)认为如果谱系尺度适中(如与本地种间的谱系距离接近), 外来种和本地种会因生态位分化而实现共存, 且不受空间尺度影响.我们比较了34种外来植物对各群落的DNNS值和每个本地种与其所在群落内其他本地种之间的最近谱系距离(NTD), 并发现在这一尺度上DNNS值的变异范围略小于NTD值的变异范围(附录II).这说明本研究调查的群落中, 外来种-本地种谱系尺度接近本地植物间真实的谱系距离.莲都区的外来植物在生态位分化上已经和本地种类似; 外来种和本地种之间保持了合理的生态位分化, 从而保证了这些外来植物和本地植物能够长期共存. ...
Taxonstand: An R package for species names standardization in vegetation databases 1 2012
... 在构建进化树之前, 我们先将所有种下分类单位合并到种.我们对莲都区群落调查数据进行整理得到一个包含279种维管束植物(蕨类4种, 被子植物275种)的物种名录(其中外来植物34种); 将温州地区种子植物名录和入侵浙江的外来种名录合并得到一个市级尺度的物种名录.由于《Flora of China》认定的植物学名和世界被子植物系统发育树的物种学名存在一定差异, 我们用R语言软件包“Taxonstand” (Cayuela, 2012)对物种名录进行了检查核对.利用Qian和Jin (2015)建立的PhytoPhylo大型系统发育树作为骨架, 我们构建了上述279个物种的进化树和温州地区市级尺度的进化树(2 457种, 含全省的归化种).PhytoPhylo大型进化树是在Zanne等(2014)建立的进化树上提出的.该进化树是使用了7个基因的序列数据和化石数据构建的目前最大且最新的具时间标定的物种水平的种子植物进化树(Zanne et al., 2014). ...
The roles of harsh and fluctuating conditions in the dynamics of ecological communites 1 1997
... 前人针对达尔文归化难题的研究得到的矛盾结果常被归因于这些研究在研究区域、类群以及在概念框架和研究方法(如不同的空间和谱系尺度、入侵阶段、量度的参数和生境条件)上存在巨大差异(Thuiller et al., 2010; Richardson & Pysek, 2012; Li et al., 2015).空间尺度和入侵阶段被认为对入侵过程的生态学机制具有重要的影响(Ma et al., 2016).****们普遍认为达尔文归化假说主要在较小空间尺度和入侵危害(或扩散)阶段起作用, 而预适应假说可能在大尺度和归化阶段起主导作用(Diez et al., 2008; Thuiller et al., 2010; Bezeng et al., 2015; Li et al., 2015), 但同时也存在很多不同意见(Jones et al., 2013; Ma et al., 2016; Campbell & Mandrak, 2017).由于外来种和本地种间稳定共存的潜力并不是随谱系距离线性(而是S形曲线)增加的, 因此研究对象间的谱系距离取值范围(即谱系尺度)对外来种和本地种互作的结果也具有重要影响(Thuiller et al., 2010).另外, 外来种的入侵过程常受多种因素影响, 其中生境条件起着基础性作用.生境条件不仅影响外来种的入侵过程, 而且同样影响本地物种的组成(Chesson & Huntly, 1997; Shea & Chesson, 2002; Richardson & Pysek, 2012), 从而影响外来种-本地种亲缘与外来种归化、扩散和危害的关系.虽然目前有不少****已经在同一研究中考虑了空间尺度和入侵阶段的影响, 但在调查性实证研究中仍很少考虑谱系尺度和生境条件差异. ...
... 早在1859年, 达尔文就注意到了外来物种和本地物种亲缘关系对外来种成功归化的影响(Darwin, 1859).这为入侵种特性的研究提供了简单的方法学框架; 只需对外来种-本地种亲缘和外来种入侵关系进行研究, 就可以方便地预测哪些物种容易在哪些生态系统中入侵(Strauss et al., 2006; Marx et al., 2016), 因此一直受到入侵生态学家的重视.达尔文归化假说认为由于同属的物种间存在强烈竞争, 在含有同属本地种的地方, 被引入的外来植物建立自我维持的野生种群的可能性将减少(Daehler, 2001).尽管如此, 达尔文同时也认识到外来种在含有同属本地种的生境成功归化也是有可能的, 因为它们和本地种具有相似的特征, 使其对当地环境有更好的适应性而有利于成功归化, 即预适应假说(Daehler, 2001; Diez et al., 2008).随着谱系生态学的发展, 这两个假说也逐渐扩展为描述外来种-本地群落谱系距离与外来物种成功归化(或入侵)的关系(Strauss et al., 2006; Thuiller et al., 2010).近20年来, ****们对这两个对立的假说进行了很多实证研究, 但得到了很不一致的结果(Thuiller et al., 2010; Jones et al., 2013; Ma et al., 2016).达尔文归化假说和预适应假说也因此被合称为达尔文归化难题(Diez et al., 2008). ...
... ).尽管如此, 达尔文同时也认识到外来种在含有同属本地种的生境成功归化也是有可能的, 因为它们和本地种具有相似的特征, 使其对当地环境有更好的适应性而有利于成功归化, 即预适应假说(Daehler, 2001; Diez et al., 2008).随着谱系生态学的发展, 这两个假说也逐渐扩展为描述外来种-本地群落谱系距离与外来物种成功归化(或入侵)的关系(Strauss et al., 2006; Thuiller et al., 2010).近20年来, ****们对这两个对立的假说进行了很多实证研究, 但得到了很不一致的结果(Thuiller et al., 2010; Jones et al., 2013; Ma et al., 2016).达尔文归化假说和预适应假说也因此被合称为达尔文归化难题(Diez et al., 2008). ...
On the Origin of Species 2 1859
... 早在1859年, 达尔文就注意到了外来物种和本地物种亲缘关系对外来种成功归化的影响(Darwin, 1859).这为入侵种特性的研究提供了简单的方法学框架; 只需对外来种-本地种亲缘和外来种入侵关系进行研究, 就可以方便地预测哪些物种容易在哪些生态系统中入侵(Strauss et al., 2006; Marx et al., 2016), 因此一直受到入侵生态学家的重视.达尔文归化假说认为由于同属的物种间存在强烈竞争, 在含有同属本地种的地方, 被引入的外来植物建立自我维持的野生种群的可能性将减少(Daehler, 2001).尽管如此, 达尔文同时也认识到外来种在含有同属本地种的生境成功归化也是有可能的, 因为它们和本地种具有相似的特征, 使其对当地环境有更好的适应性而有利于成功归化, 即预适应假说(Daehler, 2001; Diez et al., 2008).随着谱系生态学的发展, 这两个假说也逐渐扩展为描述外来种-本地群落谱系距离与外来物种成功归化(或入侵)的关系(Strauss et al., 2006; Thuiller et al., 2010).近20年来, ****们对这两个对立的假说进行了很多实证研究, 但得到了很不一致的结果(Thuiller et al., 2010; Jones et al., 2013; Ma et al., 2016).达尔文归化假说和预适应假说也因此被合称为达尔文归化难题(Diez et al., 2008). ...
... 和前人的研究结果类似, 本研究的结果显示无论在区域还是局域尺度上, 无论是归化、扩散还是危害程度, MDNS与外来种入侵过程都没有显著关系(Carboni et al., 2013; Skóra et al., 2015).理论上来说, 在群落中(尤其是多样性较高的群落)很少会存在极其相似或差异极大的物种, 大部分物种仅存在中等程度的功能差异(Skóra et al., 2015).Skóra等(2015)指出在不同情况下, 外来种可能仅和少量本地种近似也可能与许多本地种近似, 从而使MDNS的变异非常大; 当多样性增加时, MDNS会变得更加不确定.另一方面, 达尔文提出的归化假说和预适应假说表述的都是外来种归化与近缘物种(同属物种)存在与否的关系(Darwin, 1859), 而MDNS衡量的是外来种与整个本地群落(或区系)所有物种的平均谱系距离.因此, MDNS可能是一个弱指标, 在达尔文归化难题研究中, 应该与其他谱系距离的测度指标结合使用, 以确保准确反映外来种-本地种亲缘关系对外来植物入侵过程的影响. ...
Darwin’s naturalization conundrum: Dissecting taxonomic patterns of species invasions 14 2008
... 早在1859年, 达尔文就注意到了外来物种和本地物种亲缘关系对外来种成功归化的影响(Darwin, 1859).这为入侵种特性的研究提供了简单的方法学框架; 只需对外来种-本地种亲缘和外来种入侵关系进行研究, 就可以方便地预测哪些物种容易在哪些生态系统中入侵(Strauss et al., 2006; Marx et al., 2016), 因此一直受到入侵生态学家的重视.达尔文归化假说认为由于同属的物种间存在强烈竞争, 在含有同属本地种的地方, 被引入的外来植物建立自我维持的野生种群的可能性将减少(Daehler, 2001).尽管如此, 达尔文同时也认识到外来种在含有同属本地种的生境成功归化也是有可能的, 因为它们和本地种具有相似的特征, 使其对当地环境有更好的适应性而有利于成功归化, 即预适应假说(Daehler, 2001; Diez et al., 2008).随着谱系生态学的发展, 这两个假说也逐渐扩展为描述外来种-本地群落谱系距离与外来物种成功归化(或入侵)的关系(Strauss et al., 2006; Thuiller et al., 2010).近20年来, ****们对这两个对立的假说进行了很多实证研究, 但得到了很不一致的结果(Thuiller et al., 2010; Jones et al., 2013; Ma et al., 2016).达尔文归化假说和预适应假说也因此被合称为达尔文归化难题(Diez et al., 2008). ...
... ).达尔文归化假说和预适应假说也因此被合称为达尔文归化难题(Diez et al., 2008). ...
... 前人针对达尔文归化难题的研究得到的矛盾结果常被归因于这些研究在研究区域、类群以及在概念框架和研究方法(如不同的空间和谱系尺度、入侵阶段、量度的参数和生境条件)上存在巨大差异(Thuiller et al., 2010; Richardson & Pysek, 2012; Li et al., 2015).空间尺度和入侵阶段被认为对入侵过程的生态学机制具有重要的影响(Ma et al., 2016).****们普遍认为达尔文归化假说主要在较小空间尺度和入侵危害(或扩散)阶段起作用, 而预适应假说可能在大尺度和归化阶段起主导作用(Diez et al., 2008; Thuiller et al., 2010; Bezeng et al., 2015; Li et al., 2015), 但同时也存在很多不同意见(Jones et al., 2013; Ma et al., 2016; Campbell & Mandrak, 2017).由于外来种和本地种间稳定共存的潜力并不是随谱系距离线性(而是S形曲线)增加的, 因此研究对象间的谱系距离取值范围(即谱系尺度)对外来种和本地种互作的结果也具有重要影响(Thuiller et al., 2010).另外, 外来种的入侵过程常受多种因素影响, 其中生境条件起着基础性作用.生境条件不仅影响外来种的入侵过程, 而且同样影响本地物种的组成(Chesson & Huntly, 1997; Shea & Chesson, 2002; Richardson & Pysek, 2012), 从而影响外来种-本地种亲缘与外来种归化、扩散和危害的关系.虽然目前有不少****已经在同一研究中考虑了空间尺度和入侵阶段的影响, 但在调查性实证研究中仍很少考虑谱系尺度和生境条件差异. ...
... 由于没有构建全国性和各省的种子植物进化树, 在省级尺度分析中, 我们以外来种所在属的本地植物种数作为衡量外来种与本地种亲缘距离 (Diez et al., 2008).在市级尺度和群落尺度的分析中, 我们采用外来种与群落(或区系)中亲缘最近的本地种的谱系距离(DNNS)和外来种与其在群落(或区系)中所有本地种的平均谱系距离(MDNS)作为衡量外来种与本地种亲缘距离的测度(Thuiller et al., 2010).Thuiller等(2010)认为可以直接用DNNS、MDNS等谱系相似性指标或者分类等级来衡量谱系尺度, 即谱系相似性越低或者物种间共同归属的分类等级越高则谱系尺度越大. ...
... 由于达尔文归化假说以近缘种间具有较大的生态位重叠以及随之而来的竞争为理论基础, 生态学家一般认为该假说主要在小尺度上表现(Diez et al., 2008; Thuiller et al., 2010; Li et al., 2015; Ma et al., 2016).然而, 我们的研究结果表明, 在群落尺度上控制生境差异后, 外来种-本地种亲缘距离与外来种在群落中侵入成功与否及其危害程度无关.这种差异可能与本研究的谱系尺度有关(Castro et al., 2014).Thuiller等(2010)认为如果谱系尺度适中(如与本地种间的谱系距离接近), 外来种和本地种会因生态位分化而实现共存, 且不受空间尺度影响.我们比较了34种外来植物对各群落的DNNS值和每个本地种与其所在群落内其他本地种之间的最近谱系距离(NTD), 并发现在这一尺度上DNNS值的变异范围略小于NTD值的变异范围(附录II).这说明本研究调查的群落中, 外来种-本地种谱系尺度接近本地植物间真实的谱系距离.莲都区的外来植物在生态位分化上已经和本地种类似; 外来种和本地种之间保持了合理的生态位分化, 从而保证了这些外来植物和本地植物能够长期共存. ...
... 总体上看, 本研究的结果表明, 在同一尺度下外来种-本地种亲缘关系对整个入侵过程的影响没有因入侵阶段不同而不同.在温州, 外来种-本地种亲缘关系较近不仅有利于外来种归化也有利于其扩散; 而在莲都区的群落中, 外来种-本地种亲缘关系对外来种的侵入和危害程度都无影响.这和来自世界其他地方的研究结果不太一致(Diez et al., 2008; Bezeng et al., 2015; Li et al., 2015), 说明中国地区的外来种-本地种亲缘关系对入侵过程的影响可能具有自身特点. ...
... 针对含有第1-4类外来植物的152个属, 我们统计了每个属的外来种数量, 以及分布在每个省、自治区或直辖市的该属的外来种数和本地种数.本地种数量统计根据《Flora of China》(Editorial Committee of Flora of China, 2013)的记载. ...
... 2014-2016年(每年的5-12月), 我们在浙江省丽水市莲都区(1 502.1 km2, 年降水量1 395.5 mm, 年平均气温18.5 ℃)的乡村路边、弃耕地、溪边、林缘等处调查了109个1 m × 1 m的植物群落样方.我们在野外收割每个样方的所有地上部分植物材料, 带回实验室后对他们进行归类、称鲜质量、计数并鉴定.物种定名根据《Flora of China》(Editorial Committee of Flora of China, 2013). ...
... 早在1859年, 达尔文就注意到了外来物种和本地物种亲缘关系对外来种成功归化的影响(Darwin, 1859).这为入侵种特性的研究提供了简单的方法学框架; 只需对外来种-本地种亲缘和外来种入侵关系进行研究, 就可以方便地预测哪些物种容易在哪些生态系统中入侵(Strauss et al., 2006; Marx et al., 2016), 因此一直受到入侵生态学家的重视.达尔文归化假说认为由于同属的物种间存在强烈竞争, 在含有同属本地种的地方, 被引入的外来植物建立自我维持的野生种群的可能性将减少(Daehler, 2001).尽管如此, 达尔文同时也认识到外来种在含有同属本地种的生境成功归化也是有可能的, 因为它们和本地种具有相似的特征, 使其对当地环境有更好的适应性而有利于成功归化, 即预适应假说(Daehler, 2001; Diez et al., 2008).随着谱系生态学的发展, 这两个假说也逐渐扩展为描述外来种-本地群落谱系距离与外来物种成功归化(或入侵)的关系(Strauss et al., 2006; Thuiller et al., 2010).近20年来, ****们对这两个对立的假说进行了很多实证研究, 但得到了很不一致的结果(Thuiller et al., 2010; Jones et al., 2013; Ma et al., 2016).达尔文归化假说和预适应假说也因此被合称为达尔文归化难题(Diez et al., 2008). ...
... 前人针对达尔文归化难题的研究得到的矛盾结果常被归因于这些研究在研究区域、类群以及在概念框架和研究方法(如不同的空间和谱系尺度、入侵阶段、量度的参数和生境条件)上存在巨大差异(Thuiller et al., 2010; Richardson & Pysek, 2012; Li et al., 2015).空间尺度和入侵阶段被认为对入侵过程的生态学机制具有重要的影响(Ma et al., 2016).****们普遍认为达尔文归化假说主要在较小空间尺度和入侵危害(或扩散)阶段起作用, 而预适应假说可能在大尺度和归化阶段起主导作用(Diez et al., 2008; Thuiller et al., 2010; Bezeng et al., 2015; Li et al., 2015), 但同时也存在很多不同意见(Jones et al., 2013; Ma et al., 2016; Campbell & Mandrak, 2017).由于外来种和本地种间稳定共存的潜力并不是随谱系距离线性(而是S形曲线)增加的, 因此研究对象间的谱系距离取值范围(即谱系尺度)对外来种和本地种互作的结果也具有重要影响(Thuiller et al., 2010).另外, 外来种的入侵过程常受多种因素影响, 其中生境条件起着基础性作用.生境条件不仅影响外来种的入侵过程, 而且同样影响本地物种的组成(Chesson & Huntly, 1997; Shea & Chesson, 2002; Richardson & Pysek, 2012), 从而影响外来种-本地种亲缘与外来种归化、扩散和危害的关系.虽然目前有不少****已经在同一研究中考虑了空间尺度和入侵阶段的影响, 但在调查性实证研究中仍很少考虑谱系尺度和生境条件差异. ...
... 早在1859年, 达尔文就注意到了外来物种和本地物种亲缘关系对外来种成功归化的影响(Darwin, 1859).这为入侵种特性的研究提供了简单的方法学框架; 只需对外来种-本地种亲缘和外来种入侵关系进行研究, 就可以方便地预测哪些物种容易在哪些生态系统中入侵(Strauss et al., 2006; Marx et al., 2016), 因此一直受到入侵生态学家的重视.达尔文归化假说认为由于同属的物种间存在强烈竞争, 在含有同属本地种的地方, 被引入的外来植物建立自我维持的野生种群的可能性将减少(Daehler, 2001).尽管如此, 达尔文同时也认识到外来种在含有同属本地种的生境成功归化也是有可能的, 因为它们和本地种具有相似的特征, 使其对当地环境有更好的适应性而有利于成功归化, 即预适应假说(Daehler, 2001; Diez et al., 2008).随着谱系生态学的发展, 这两个假说也逐渐扩展为描述外来种-本地群落谱系距离与外来物种成功归化(或入侵)的关系(Strauss et al., 2006; Thuiller et al., 2010).近20年来, ****们对这两个对立的假说进行了很多实证研究, 但得到了很不一致的结果(Thuiller et al., 2010; Jones et al., 2013; Ma et al., 2016).达尔文归化假说和预适应假说也因此被合称为达尔文归化难题(Diez et al., 2008). ...
... 前人针对达尔文归化难题的研究得到的矛盾结果常被归因于这些研究在研究区域、类群以及在概念框架和研究方法(如不同的空间和谱系尺度、入侵阶段、量度的参数和生境条件)上存在巨大差异(Thuiller et al., 2010; Richardson & Pysek, 2012; Li et al., 2015).空间尺度和入侵阶段被认为对入侵过程的生态学机制具有重要的影响(Ma et al., 2016).****们普遍认为达尔文归化假说主要在较小空间尺度和入侵危害(或扩散)阶段起作用, 而预适应假说可能在大尺度和归化阶段起主导作用(Diez et al., 2008; Thuiller et al., 2010; Bezeng et al., 2015; Li et al., 2015), 但同时也存在很多不同意见(Jones et al., 2013; Ma et al., 2016; Campbell & Mandrak, 2017).由于外来种和本地种间稳定共存的潜力并不是随谱系距离线性(而是S形曲线)增加的, 因此研究对象间的谱系距离取值范围(即谱系尺度)对外来种和本地种互作的结果也具有重要影响(Thuiller et al., 2010).另外, 外来种的入侵过程常受多种因素影响, 其中生境条件起着基础性作用.生境条件不仅影响外来种的入侵过程, 而且同样影响本地物种的组成(Chesson & Huntly, 1997; Shea & Chesson, 2002; Richardson & Pysek, 2012), 从而影响外来种-本地种亲缘与外来种归化、扩散和危害的关系.虽然目前有不少****已经在同一研究中考虑了空间尺度和入侵阶段的影响, 但在调查性实证研究中仍很少考虑谱系尺度和生境条件差异. ...
Revisiting Darwin’s conundrum reveals a twist on the relationship between phylogenetic distance and invasibility 5 2013
... 随着全球经济一体化进程的加快, 生物扩散的自然屏障被打破, 地区间物种交换的比例和空间范围大大增加(Campbell & Mandrak, 2017).部分外来物种到达新的分布区域后在生态、经济和社会等方面给当地造成了严重的后果(Mack et al., 2000; Pejchar & Mooney, 2009).面对生物入侵的严峻挑战, 预测外来种的潜在入侵性和生态影响已经成为生态学家需要解决的重要研究议题(Mack et al., 2000).早期的研究从归纳入侵种的功能特征(特别是资源利用、生殖潜力和扩散能力有关的特征)入手, 总结入侵物种的共性特征(Baker, 1965; Rejmanek & Richardson, 1996).然而, 这些方法仍不能得出一致的结论, 与高入侵性有关的生活史特征在不同类群和生境中存在很大差异(Bezeng et al., 2015; Li et al., 2015).而且一些重要的功能性状很难鉴别和测量, 需要投入大量的人力和物力(Bezeng et al., 2015). ...
... 前人针对达尔文归化难题的研究得到的矛盾结果常被归因于这些研究在研究区域、类群以及在概念框架和研究方法(如不同的空间和谱系尺度、入侵阶段、量度的参数和生境条件)上存在巨大差异(Thuiller et al., 2010; Richardson & Pysek, 2012; Li et al., 2015).空间尺度和入侵阶段被认为对入侵过程的生态学机制具有重要的影响(Ma et al., 2016).****们普遍认为达尔文归化假说主要在较小空间尺度和入侵危害(或扩散)阶段起作用, 而预适应假说可能在大尺度和归化阶段起主导作用(Diez et al., 2008; Thuiller et al., 2010; Bezeng et al., 2015; Li et al., 2015), 但同时也存在很多不同意见(Jones et al., 2013; Ma et al., 2016; Campbell & Mandrak, 2017).由于外来种和本地种间稳定共存的潜力并不是随谱系距离线性(而是S形曲线)增加的, 因此研究对象间的谱系距离取值范围(即谱系尺度)对外来种和本地种互作的结果也具有重要影响(Thuiller et al., 2010).另外, 外来种的入侵过程常受多种因素影响, 其中生境条件起着基础性作用.生境条件不仅影响外来种的入侵过程, 而且同样影响本地物种的组成(Chesson & Huntly, 1997; Shea & Chesson, 2002; Richardson & Pysek, 2012), 从而影响外来种-本地种亲缘与外来种归化、扩散和危害的关系.虽然目前有不少****已经在同一研究中考虑了空间尺度和入侵阶段的影响, 但在调查性实证研究中仍很少考虑谱系尺度和生境条件差异. ...
... ; Li et al., 2015), 但同时也存在很多不同意见(Jones et al., 2013; Ma et al., 2016; Campbell & Mandrak, 2017).由于外来种和本地种间稳定共存的潜力并不是随谱系距离线性(而是S形曲线)增加的, 因此研究对象间的谱系距离取值范围(即谱系尺度)对外来种和本地种互作的结果也具有重要影响(Thuiller et al., 2010).另外, 外来种的入侵过程常受多种因素影响, 其中生境条件起着基础性作用.生境条件不仅影响外来种的入侵过程, 而且同样影响本地物种的组成(Chesson & Huntly, 1997; Shea & Chesson, 2002; Richardson & Pysek, 2012), 从而影响外来种-本地种亲缘与外来种归化、扩散和危害的关系.虽然目前有不少****已经在同一研究中考虑了空间尺度和入侵阶段的影响, 但在调查性实证研究中仍很少考虑谱系尺度和生境条件差异. ...
... 由于达尔文归化假说以近缘种间具有较大的生态位重叠以及随之而来的竞争为理论基础, 生态学家一般认为该假说主要在小尺度上表现(Diez et al., 2008; Thuiller et al., 2010; Li et al., 2015; Ma et al., 2016).然而, 我们的研究结果表明, 在群落尺度上控制生境差异后, 外来种-本地种亲缘距离与外来种在群落中侵入成功与否及其危害程度无关.这种差异可能与本研究的谱系尺度有关(Castro et al., 2014).Thuiller等(2010)认为如果谱系尺度适中(如与本地种间的谱系距离接近), 外来种和本地种会因生态位分化而实现共存, 且不受空间尺度影响.我们比较了34种外来植物对各群落的DNNS值和每个本地种与其所在群落内其他本地种之间的最近谱系距离(NTD), 并发现在这一尺度上DNNS值的变异范围略小于NTD值的变异范围(附录II).这说明本研究调查的群落中, 外来种-本地种谱系尺度接近本地植物间真实的谱系距离.莲都区的外来植物在生态位分化上已经和本地种类似; 外来种和本地种之间保持了合理的生态位分化, 从而保证了这些外来植物和本地植物能够长期共存. ...
... 总体上看, 本研究的结果表明, 在同一尺度下外来种-本地种亲缘关系对整个入侵过程的影响没有因入侵阶段不同而不同.在温州, 外来种-本地种亲缘关系较近不仅有利于外来种归化也有利于其扩散; 而在莲都区的群落中, 外来种-本地种亲缘关系对外来种的侵入和危害程度都无影响.这和来自世界其他地方的研究结果不太一致(Diez et al., 2008; Bezeng et al., 2015; Li et al., 2015), 说明中国地区的外来种-本地种亲缘关系对入侵过程的影响可能具有自身特点. ...
The effects of phylogenetic relatedness on invasion success and impact: Deconstructing Darwin’s naturalisation conundrum 5 2015
... 早在1859年, 达尔文就注意到了外来物种和本地物种亲缘关系对外来种成功归化的影响(Darwin, 1859).这为入侵种特性的研究提供了简单的方法学框架; 只需对外来种-本地种亲缘和外来种入侵关系进行研究, 就可以方便地预测哪些物种容易在哪些生态系统中入侵(Strauss et al., 2006; Marx et al., 2016), 因此一直受到入侵生态学家的重视.达尔文归化假说认为由于同属的物种间存在强烈竞争, 在含有同属本地种的地方, 被引入的外来植物建立自我维持的野生种群的可能性将减少(Daehler, 2001).尽管如此, 达尔文同时也认识到外来种在含有同属本地种的生境成功归化也是有可能的, 因为它们和本地种具有相似的特征, 使其对当地环境有更好的适应性而有利于成功归化, 即预适应假说(Daehler, 2001; Diez et al., 2008).随着谱系生态学的发展, 这两个假说也逐渐扩展为描述外来种-本地群落谱系距离与外来物种成功归化(或入侵)的关系(Strauss et al., 2006; Thuiller et al., 2010).近20年来, ****们对这两个对立的假说进行了很多实证研究, 但得到了很不一致的结果(Thuiller et al., 2010; Jones et al., 2013; Ma et al., 2016).达尔文归化假说和预适应假说也因此被合称为达尔文归化难题(Diez et al., 2008). ...
... 前人针对达尔文归化难题的研究得到的矛盾结果常被归因于这些研究在研究区域、类群以及在概念框架和研究方法(如不同的空间和谱系尺度、入侵阶段、量度的参数和生境条件)上存在巨大差异(Thuiller et al., 2010; Richardson & Pysek, 2012; Li et al., 2015).空间尺度和入侵阶段被认为对入侵过程的生态学机制具有重要的影响(Ma et al., 2016).****们普遍认为达尔文归化假说主要在较小空间尺度和入侵危害(或扩散)阶段起作用, 而预适应假说可能在大尺度和归化阶段起主导作用(Diez et al., 2008; Thuiller et al., 2010; Bezeng et al., 2015; Li et al., 2015), 但同时也存在很多不同意见(Jones et al., 2013; Ma et al., 2016; Campbell & Mandrak, 2017).由于外来种和本地种间稳定共存的潜力并不是随谱系距离线性(而是S形曲线)增加的, 因此研究对象间的谱系距离取值范围(即谱系尺度)对外来种和本地种互作的结果也具有重要影响(Thuiller et al., 2010).另外, 外来种的入侵过程常受多种因素影响, 其中生境条件起着基础性作用.生境条件不仅影响外来种的入侵过程, 而且同样影响本地物种的组成(Chesson & Huntly, 1997; Shea & Chesson, 2002; Richardson & Pysek, 2012), 从而影响外来种-本地种亲缘与外来种归化、扩散和危害的关系.虽然目前有不少****已经在同一研究中考虑了空间尺度和入侵阶段的影响, 但在调查性实证研究中仍很少考虑谱系尺度和生境条件差异. ...
... 外来种-本地种谱系关系或分类关系对生物入侵和归化的影响是近20年来入侵生态学中争论最多的问题之一(Thuiller et al., 2010; Ma et al., 2016).前人的许多研究表明在较大尺度(如区域)上, 与本地种亲缘关系较近的外来种更易归化或入侵; 而在较小尺度(如局域或者同一生境水平)上, 成功归化或入侵的外来种与本地种的亲缘关系较远(Diez et al., 2008; Carboni et al., 2013; Campbell & Mandrak, 2017).我们多尺度多入侵阶段的研究表明, 在省级和市级(区域)尺度上, 与本地种亲缘关系较近的外来植物更有可能在当地归化和扩散, 符合预适应假说的预期; 而在群落(局域)尺度上, 外来种-本地种亲缘距离与外来种是否在群落中成功定居及其危害程度无关. ...
... 由于达尔文归化假说以近缘种间具有较大的生态位重叠以及随之而来的竞争为理论基础, 生态学家一般认为该假说主要在小尺度上表现(Diez et al., 2008; Thuiller et al., 2010; Li et al., 2015; Ma et al., 2016).然而, 我们的研究结果表明, 在群落尺度上控制生境差异后, 外来种-本地种亲缘距离与外来种在群落中侵入成功与否及其危害程度无关.这种差异可能与本研究的谱系尺度有关(Castro et al., 2014).Thuiller等(2010)认为如果谱系尺度适中(如与本地种间的谱系距离接近), 外来种和本地种会因生态位分化而实现共存, 且不受空间尺度影响.我们比较了34种外来植物对各群落的DNNS值和每个本地种与其所在群落内其他本地种之间的最近谱系距离(NTD), 并发现在这一尺度上DNNS值的变异范围略小于NTD值的变异范围(附录II).这说明本研究调查的群落中, 外来种-本地种谱系尺度接近本地植物间真实的谱系距离.莲都区的外来植物在生态位分化上已经和本地种类似; 外来种和本地种之间保持了合理的生态位分化, 从而保证了这些外来植物和本地植物能够长期共存. ...
Different effects of invader-native phylogenetic relatedness on invasion success and impact: A meta-analysis of Darwin’s naturalization hypothesis 2016
4 2013
... 随着全球经济一体化进程的加快, 生物扩散的自然屏障被打破, 地区间物种交换的比例和空间范围大大增加(Campbell & Mandrak, 2017).部分外来物种到达新的分布区域后在生态、经济和社会等方面给当地造成了严重的后果(Mack et al., 2000; Pejchar & Mooney, 2009).面对生物入侵的严峻挑战, 预测外来种的潜在入侵性和生态影响已经成为生态学家需要解决的重要研究议题(Mack et al., 2000).早期的研究从归纳入侵种的功能特征(特别是资源利用、生殖潜力和扩散能力有关的特征)入手, 总结入侵物种的共性特征(Baker, 1965; Rejmanek & Richardson, 1996).然而, 这些方法仍不能得出一致的结论, 与高入侵性有关的生活史特征在不同类群和生境中存在很大差异(Bezeng et al., 2015; Li et al., 2015).而且一些重要的功能性状很难鉴别和测量, 需要投入大量的人力和物力(Bezeng et al., 2015). ...
... ).面对生物入侵的严峻挑战, 预测外来种的潜在入侵性和生态影响已经成为生态学家需要解决的重要研究议题(Mack et al., 2000).早期的研究从归纳入侵种的功能特征(特别是资源利用、生殖潜力和扩散能力有关的特征)入手, 总结入侵物种的共性特征(Baker, 1965; Rejmanek & Richardson, 1996).然而, 这些方法仍不能得出一致的结论, 与高入侵性有关的生活史特征在不同类群和生境中存在很大差异(Bezeng et al., 2015; Li et al., 2015).而且一些重要的功能性状很难鉴别和测量, 需要投入大量的人力和物力(Bezeng et al., 2015). ...
Biotic invasions: Causes, epidemiology, global consequences, and control 1 2000
... 早在1859年, 达尔文就注意到了外来物种和本地物种亲缘关系对外来种成功归化的影响(Darwin, 1859).这为入侵种特性的研究提供了简单的方法学框架; 只需对外来种-本地种亲缘和外来种入侵关系进行研究, 就可以方便地预测哪些物种容易在哪些生态系统中入侵(Strauss et al., 2006; Marx et al., 2016), 因此一直受到入侵生态学家的重视.达尔文归化假说认为由于同属的物种间存在强烈竞争, 在含有同属本地种的地方, 被引入的外来植物建立自我维持的野生种群的可能性将减少(Daehler, 2001).尽管如此, 达尔文同时也认识到外来种在含有同属本地种的生境成功归化也是有可能的, 因为它们和本地种具有相似的特征, 使其对当地环境有更好的适应性而有利于成功归化, 即预适应假说(Daehler, 2001; Diez et al., 2008).随着谱系生态学的发展, 这两个假说也逐渐扩展为描述外来种-本地群落谱系距离与外来物种成功归化(或入侵)的关系(Strauss et al., 2006; Thuiller et al., 2010).近20年来, ****们对这两个对立的假说进行了很多实证研究, 但得到了很不一致的结果(Thuiller et al., 2010; Jones et al., 2013; Ma et al., 2016).达尔文归化假说和预适应假说也因此被合称为达尔文归化难题(Diez et al., 2008). ...
Deconstructing Darwin’s naturalization conundrum in the San Juan Islands using community phylogenetics and functional traits 2016
Functional traits and nich-based tree community assembly in an Amazonian forest 1 2008
... 由于在大尺度上存在更高的生境异质性, 生境过滤作用也主要发生在较大的空间尺度上, 大多数生态学家认为预适应假说应主要表现在大尺度上(Diez et al., 2008; Nathan et al., 2008; Thuiller et al., 2010).本研究的结果表明在省级和市级尺度上, 与本地种亲缘关系较近有利于外来植物在当地归化和扩散, 有力地支持了上述观点.我国本地植物区系较为古老且与世界其他植物区系存在广泛联系(吴征镒等, 2010), 从而有利于与当地植物亲缘相近的外来种归化到相应地区.例如, 东亚和北美地区存在紧密的生物地理学联系, 原产北美的植物非常适应我国的生存环境, 造成在我国的大量入侵(闫小玲等, 2014a). ...
Phytools: An R package for phylogenetic comparative biology (and other things) 3 2012
... 前人针对达尔文归化难题的研究得到的矛盾结果常被归因于这些研究在研究区域、类群以及在概念框架和研究方法(如不同的空间和谱系尺度、入侵阶段、量度的参数和生境条件)上存在巨大差异(Thuiller et al., 2010; Richardson & Pysek, 2012; Li et al., 2015).空间尺度和入侵阶段被认为对入侵过程的生态学机制具有重要的影响(Ma et al., 2016).****们普遍认为达尔文归化假说主要在较小空间尺度和入侵危害(或扩散)阶段起作用, 而预适应假说可能在大尺度和归化阶段起主导作用(Diez et al., 2008; Thuiller et al., 2010; Bezeng et al., 2015; Li et al., 2015), 但同时也存在很多不同意见(Jones et al., 2013; Ma et al., 2016; Campbell & Mandrak, 2017).由于外来种和本地种间稳定共存的潜力并不是随谱系距离线性(而是S形曲线)增加的, 因此研究对象间的谱系距离取值范围(即谱系尺度)对外来种和本地种互作的结果也具有重要影响(Thuiller et al., 2010).另外, 外来种的入侵过程常受多种因素影响, 其中生境条件起着基础性作用.生境条件不仅影响外来种的入侵过程, 而且同样影响本地物种的组成(Chesson & Huntly, 1997; Shea & Chesson, 2002; Richardson & Pysek, 2012), 从而影响外来种-本地种亲缘与外来种归化、扩散和危害的关系.虽然目前有不少****已经在同一研究中考虑了空间尺度和入侵阶段的影响, 但在调查性实证研究中仍很少考虑谱系尺度和生境条件差异. ...
Naturalization of introduced plants: Ecological drivers of biogeographical patterns 1 2012
... 前人针对达尔文归化难题的研究得到的矛盾结果常被归因于这些研究在研究区域、类群以及在概念框架和研究方法(如不同的空间和谱系尺度、入侵阶段、量度的参数和生境条件)上存在巨大差异(Thuiller et al., 2010; Richardson & Pysek, 2012; Li et al., 2015).空间尺度和入侵阶段被认为对入侵过程的生态学机制具有重要的影响(Ma et al., 2016).****们普遍认为达尔文归化假说主要在较小空间尺度和入侵危害(或扩散)阶段起作用, 而预适应假说可能在大尺度和归化阶段起主导作用(Diez et al., 2008; Thuiller et al., 2010; Bezeng et al., 2015; Li et al., 2015), 但同时也存在很多不同意见(Jones et al., 2013; Ma et al., 2016; Campbell & Mandrak, 2017).由于外来种和本地种间稳定共存的潜力并不是随谱系距离线性(而是S形曲线)增加的, 因此研究对象间的谱系距离取值范围(即谱系尺度)对外来种和本地种互作的结果也具有重要影响(Thuiller et al., 2010).另外, 外来种的入侵过程常受多种因素影响, 其中生境条件起着基础性作用.生境条件不仅影响外来种的入侵过程, 而且同样影响本地物种的组成(Chesson & Huntly, 1997; Shea & Chesson, 2002; Richardson & Pysek, 2012), 从而影响外来种-本地种亲缘与外来种归化、扩散和危害的关系.虽然目前有不少****已经在同一研究中考虑了空间尺度和入侵阶段的影响, 但在调查性实证研究中仍很少考虑谱系尺度和生境条件差异. ...
Community ecology theory as a framework for biological invasion 3 2002
... 和前人的研究结果类似, 本研究的结果显示无论在区域还是局域尺度上, 无论是归化、扩散还是危害程度, MDNS与外来种入侵过程都没有显著关系(Carboni et al., 2013; Skóra et al., 2015).理论上来说, 在群落中(尤其是多样性较高的群落)很少会存在极其相似或差异极大的物种, 大部分物种仅存在中等程度的功能差异(Skóra et al., 2015).Skóra等(2015)指出在不同情况下, 外来种可能仅和少量本地种近似也可能与许多本地种近似, 从而使MDNS的变异非常大; 当多样性增加时, MDNS会变得更加不确定.另一方面, 达尔文提出的归化假说和预适应假说表述的都是外来种归化与近缘物种(同属物种)存在与否的关系(Darwin, 1859), 而MDNS衡量的是外来种与整个本地群落(或区系)所有物种的平均谱系距离.因此, MDNS可能是一个弱指标, 在达尔文归化难题研究中, 应该与其他谱系距离的测度指标结合使用, 以确保准确反映外来种-本地种亲缘关系对外来植物入侵过程的影响. ...
Darwin’s hypotheses to explain colonization trends: Evidence from a quasi-natural experiment and a new conceptual model 2 2015
... 早在1859年, 达尔文就注意到了外来物种和本地物种亲缘关系对外来种成功归化的影响(Darwin, 1859).这为入侵种特性的研究提供了简单的方法学框架; 只需对外来种-本地种亲缘和外来种入侵关系进行研究, 就可以方便地预测哪些物种容易在哪些生态系统中入侵(Strauss et al., 2006; Marx et al., 2016), 因此一直受到入侵生态学家的重视.达尔文归化假说认为由于同属的物种间存在强烈竞争, 在含有同属本地种的地方, 被引入的外来植物建立自我维持的野生种群的可能性将减少(Daehler, 2001).尽管如此, 达尔文同时也认识到外来种在含有同属本地种的生境成功归化也是有可能的, 因为它们和本地种具有相似的特征, 使其对当地环境有更好的适应性而有利于成功归化, 即预适应假说(Daehler, 2001; Diez et al., 2008).随着谱系生态学的发展, 这两个假说也逐渐扩展为描述外来种-本地群落谱系距离与外来物种成功归化(或入侵)的关系(Strauss et al., 2006; Thuiller et al., 2010).近20年来, ****们对这两个对立的假说进行了很多实证研究, 但得到了很不一致的结果(Thuiller et al., 2010; Jones et al., 2013; Ma et al., 2016).达尔文归化假说和预适应假说也因此被合称为达尔文归化难题(Diez et al., 2008). ...
... ).随着谱系生态学的发展, 这两个假说也逐渐扩展为描述外来种-本地群落谱系距离与外来物种成功归化(或入侵)的关系(Strauss et al., 2006; Thuiller et al., 2010).近20年来, ****们对这两个对立的假说进行了很多实证研究, 但得到了很不一致的结果(Thuiller et al., 2010; Jones et al., 2013; Ma et al., 2016).达尔文归化假说和预适应假说也因此被合称为达尔文归化难题(Diez et al., 2008). ...
Exotic taxa less related to native species are more invasive 12 2006
... 早在1859年, 达尔文就注意到了外来物种和本地物种亲缘关系对外来种成功归化的影响(Darwin, 1859).这为入侵种特性的研究提供了简单的方法学框架; 只需对外来种-本地种亲缘和外来种入侵关系进行研究, 就可以方便地预测哪些物种容易在哪些生态系统中入侵(Strauss et al., 2006; Marx et al., 2016), 因此一直受到入侵生态学家的重视.达尔文归化假说认为由于同属的物种间存在强烈竞争, 在含有同属本地种的地方, 被引入的外来植物建立自我维持的野生种群的可能性将减少(Daehler, 2001).尽管如此, 达尔文同时也认识到外来种在含有同属本地种的生境成功归化也是有可能的, 因为它们和本地种具有相似的特征, 使其对当地环境有更好的适应性而有利于成功归化, 即预适应假说(Daehler, 2001; Diez et al., 2008).随着谱系生态学的发展, 这两个假说也逐渐扩展为描述外来种-本地群落谱系距离与外来物种成功归化(或入侵)的关系(Strauss et al., 2006; Thuiller et al., 2010).近20年来, ****们对这两个对立的假说进行了很多实证研究, 但得到了很不一致的结果(Thuiller et al., 2010; Jones et al., 2013; Ma et al., 2016).达尔文归化假说和预适应假说也因此被合称为达尔文归化难题(Diez et al., 2008). ...
... ).近20年来, ****们对这两个对立的假说进行了很多实证研究, 但得到了很不一致的结果(Thuiller et al., 2010; Jones et al., 2013; Ma et al., 2016).达尔文归化假说和预适应假说也因此被合称为达尔文归化难题(Diez et al., 2008). ...
... 前人针对达尔文归化难题的研究得到的矛盾结果常被归因于这些研究在研究区域、类群以及在概念框架和研究方法(如不同的空间和谱系尺度、入侵阶段、量度的参数和生境条件)上存在巨大差异(Thuiller et al., 2010; Richardson & Pysek, 2012; Li et al., 2015).空间尺度和入侵阶段被认为对入侵过程的生态学机制具有重要的影响(Ma et al., 2016).****们普遍认为达尔文归化假说主要在较小空间尺度和入侵危害(或扩散)阶段起作用, 而预适应假说可能在大尺度和归化阶段起主导作用(Diez et al., 2008; Thuiller et al., 2010; Bezeng et al., 2015; Li et al., 2015), 但同时也存在很多不同意见(Jones et al., 2013; Ma et al., 2016; Campbell & Mandrak, 2017).由于外来种和本地种间稳定共存的潜力并不是随谱系距离线性(而是S形曲线)增加的, 因此研究对象间的谱系距离取值范围(即谱系尺度)对外来种和本地种互作的结果也具有重要影响(Thuiller et al., 2010).另外, 外来种的入侵过程常受多种因素影响, 其中生境条件起着基础性作用.生境条件不仅影响外来种的入侵过程, 而且同样影响本地物种的组成(Chesson & Huntly, 1997; Shea & Chesson, 2002; Richardson & Pysek, 2012), 从而影响外来种-本地种亲缘与外来种归化、扩散和危害的关系.虽然目前有不少****已经在同一研究中考虑了空间尺度和入侵阶段的影响, 但在调查性实证研究中仍很少考虑谱系尺度和生境条件差异. ...
... ; Thuiller et al., 2010; Bezeng et al., 2015; Li et al., 2015), 但同时也存在很多不同意见(Jones et al., 2013; Ma et al., 2016; Campbell & Mandrak, 2017).由于外来种和本地种间稳定共存的潜力并不是随谱系距离线性(而是S形曲线)增加的, 因此研究对象间的谱系距离取值范围(即谱系尺度)对外来种和本地种互作的结果也具有重要影响(Thuiller et al., 2010).另外, 外来种的入侵过程常受多种因素影响, 其中生境条件起着基础性作用.生境条件不仅影响外来种的入侵过程, 而且同样影响本地物种的组成(Chesson & Huntly, 1997; Shea & Chesson, 2002; Richardson & Pysek, 2012), 从而影响外来种-本地种亲缘与外来种归化、扩散和危害的关系.虽然目前有不少****已经在同一研究中考虑了空间尺度和入侵阶段的影响, 但在调查性实证研究中仍很少考虑谱系尺度和生境条件差异. ...
... 由于达尔文归化假说以近缘种间具有较大的生态位重叠以及随之而来的竞争为理论基础, 生态学家一般认为该假说主要在小尺度上表现(Diez et al., 2008; Thuiller et al., 2010; Li et al., 2015; Ma et al., 2016).然而, 我们的研究结果表明, 在群落尺度上控制生境差异后, 外来种-本地种亲缘距离与外来种在群落中侵入成功与否及其危害程度无关.这种差异可能与本研究的谱系尺度有关(Castro et al., 2014).Thuiller等(2010)认为如果谱系尺度适中(如与本地种间的谱系距离接近), 外来种和本地种会因生态位分化而实现共存, 且不受空间尺度影响.我们比较了34种外来植物对各群落的DNNS值和每个本地种与其所在群落内其他本地种之间的最近谱系距离(NTD), 并发现在这一尺度上DNNS值的变异范围略小于NTD值的变异范围(附录II).这说明本研究调查的群落中, 外来种-本地种谱系尺度接近本地植物间真实的谱系距离.莲都区的外来植物在生态位分化上已经和本地种类似; 外来种和本地种之间保持了合理的生态位分化, 从而保证了这些外来植物和本地植物能够长期共存. ...
Resolving Darwin’s naturalization conundrum: A quest for evidence 2 2010
... 由于没有构建全国性和各省的种子植物进化树, 在省级尺度分析中, 我们以外来种所在属的本地植物种数作为衡量外来种与本地种亲缘距离 (Diez et al., 2008).在市级尺度和群落尺度的分析中, 我们采用外来种与群落(或区系)中亲缘最近的本地种的谱系距离(DNNS)和外来种与其在群落(或区系)中所有本地种的平均谱系距离(MDNS)作为衡量外来种与本地种亲缘距离的测度(Thuiller et al., 2010).Thuiller等(2010)认为可以直接用DNNS、MDNS等谱系相似性指标或者分类等级来衡量谱系尺度, 即谱系相似性越低或者物种间共同归属的分类等级越高则谱系尺度越大. ...
... 由于达尔文归化假说以近缘种间具有较大的生态位重叠以及随之而来的竞争为理论基础, 生态学家一般认为该假说主要在小尺度上表现(Diez et al., 2008; Thuiller et al., 2010; Li et al., 2015; Ma et al., 2016).然而, 我们的研究结果表明, 在群落尺度上控制生境差异后, 外来种-本地种亲缘距离与外来种在群落中侵入成功与否及其危害程度无关.这种差异可能与本研究的谱系尺度有关(Castro et al., 2014).Thuiller等(2010)认为如果谱系尺度适中(如与本地种间的谱系距离接近), 外来种和本地种会因生态位分化而实现共存, 且不受空间尺度影响.我们比较了34种外来植物对各群落的DNNS值和每个本地种与其所在群落内其他本地种之间的最近谱系距离(NTD), 并发现在这一尺度上DNNS值的变异范围略小于NTD值的变异范围(附录II).这说明本研究调查的群落中, 外来种-本地种谱系尺度接近本地植物间真实的谱系距离.莲都区的外来植物在生态位分化上已经和本地种类似; 外来种和本地种之间保持了合理的生态位分化, 从而保证了这些外来植物和本地植物能够长期共存. ...
Patterns of plant invasions in China: Taxonomic, biogeographic, climatic approaches and anthropogenic effects 1 2010
 ,*, 夏更寿丽水学院生态学院, 浙江丽水 323000
,*, 夏更寿丽水学院生态学院, 浙江丽水 323000
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}