 ,
, ,1,*
,1,*Effects of desertification on the C:N:P stoichiometry of soil, microbes, and extracellular enzymes in a desert grassland
WU Xiu-Zhi1, YAN Xin1, WANG Bo2, LIU Ren-Tao1, AN Hui,,1,*通讯作者: *安慧 ORCID: 0000-0002-5881-4213anhui08@163.com
编委: 韩文轩
责任编辑: 李敏
收稿日期:2018-05-23网络出版日期:2018-10-20
| 基金资助: |
Received:2018-05-23Online:2018-10-20
| Fund supported: |

摘要
为探讨荒漠草地沙漠化对“土壤-微生物-胞外酶”系统生态化学计量的影响机理, 该研究采用空间序列代替时间演替的方法, 研究了宁夏盐池荒漠草地沙漠化过程中土壤、土壤微生物及土壤胞外酶碳(C)、氮(N)、磷(P)生态化学计量的变异特征。结果表明: (1)随着荒漠草地沙漠化的不断加剧, 土壤C、N、P含量和土壤C:P、N:P均呈降低趋势, 而土壤C:N逐渐增加。(2)荒漠草地沙漠化过程中, 土壤微生物生物量C (MBC):微生物生物量P (MBP)、微生物生物量N (MBN):MBP和土壤β-葡萄糖苷酶(BG):N-乙酰氨基葡萄糖苷酶(NAG)逐渐降低, 而土壤BG:磷酸酶(AP)和NAG:AP基本表现为增加趋势。(3)随着荒漠草地沙漠化程度的加剧, 土壤微生物C利用效率CUEC:N和CUEC:P与土壤微生物N利用效率NUEN:C和土壤微生物P利用效率PUEP:C的变化趋势相反。(4)荒漠草地土壤、土壤微生物生物量和土壤胞外酶C:N化学计量(C:N, MBC:MBN, BG:NAG)与土壤、土壤微生物生物量和土壤胞外酶N:P化学计量(N:P, MBN:MBP, NAG:AP)显著负相关, 而土壤和胞外酶C:N化学计量(C:N, BG:NAG)与土壤和胞外酶C:P化学计量(C:P, BG:AP)显著正相关。土壤N:P与土壤MBN:MBP显著正相关, 而与土壤NAG:AP显著负相关。分析表明, 荒漠草地沙漠化过程中土壤微生物生物量及胞外酶活性随着土壤养分的变化而发生变化; 微生物-胞外酶C:N:P生态化学计量与土壤养分存在协变关系, 为理解荒漠草地土壤-微生物系统C、N、P循环机制提供理论依据。
关键词:
Abstract
Aims In order to discuss the underlying mechanism of desertification effect on the ecological stoichiometry of soil, microbes and extracellular enzymes, we studied the changes of soil, soil microbial and extracellular enzyme C:N:P stoichiometry during the desertification process in the desert grassland in Yanchi County, China.
Methods The “space-for-time” method was used.
Important findings The results demonstrated that: (1) Soil C, N, P contents and soil C:P, N:P significantly decreased, but soil C:N gradually increased with increasing desertification. (2) Soil microbial biomass C (MBC):soil microbial biomass P (MBP), soil microbial biomass N (MBN):MBP and soil β-1,4-glucosidase (BG):β-1,4-N- acetylglucosaminidase (NAG) gradually decreased, soil BG:alkaline phosphatase (AP) and NAG:AP basically showed an increasing trend with increasing desertification. (3) Desertification increased the soil microbial carbon use efficiency (CUEC:N and CUEC:P) gradually, while soil microbial nitrogen use efficiency (NUEN:C) and soil microbial phosphorus use efficiency (PUEP:C) basically decreased. (4) Soil, soil microbial and soil extracellular enzyme C:N stoichiometry (C:N, MBC:MBN, BG:NAG) were significantly negatively correlated with the soil, soil microbial and extracellular enzyme N:P stoichiometry (N:P, MBN:MBP, NAG:AP), the soil and extracellular enzymes C:N (C:N, BG:NAG) were significantly positively correlated with the soil and extracellular enzymes C:P (C:P, BG:AP). Soil N:P was significantly positively correlated with the soil MBN:MBP, but was significantly negatively correlated with the soil NAG:AP. The analysis demonstrated that soil microbial biomass and extracellular enzyme activity changed with soil nutrient during the desertification process in the desert grassland. The covariation relationship between soil nutrient and C:N:P stoichiometry of microbial-extracellular enzyme provides a theoretical basis for understanding the underlying mechanism of C, N, P cycling in the soil-microbial system in desert grasslands.
Keywords:
PDF (1387KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
吴秀芝, 阎欣, 王波, 刘任涛, 安慧. 荒漠草地沙漠化对土壤-微生物-胞外酶化学计量特征的影响. 植物生态学报[J], 2018, 42(10): 1022-1032 DOI:10.17521/cjpe.2018.0121
WU Xiu-Zhi, YAN Xin, WANG Bo, LIU Ren-Tao, AN Hui.
生态化学计量学以环境中相互作用的生物体以及生物体与环境之间的物质平衡来认识生态动力学, 以元素组成来揭示生物化学和生态学之间的联系, 从而分析和预测生态系统的结构和动力学(Sterner & Elser, 2002; Michaels, 2003)。C:N:P化学计量学研究已深入到生态学的各个层次(细胞、个体、种群、群落、生态系统)及区域等不同尺度, 生态化学计量成为联系分子水平化学过程与生态系统水平过程的有效工具(Anderson et al., 2004; 王绍强和于贵瑞, 2008; 贺金生和韩兴国, 2010)。近年来, 由于认识到化学计量学研究可以把生态实体的各个层次在元素水平上统一起来, 因此生态化学计量学已成为研究生态系统的新兴工具。
土壤微生物是土壤生态系统的重要组成部分, 参与土壤有机质分解、腐殖质形成、土壤养分转化和循环等过程。土壤胞外酶是微生物分泌到土壤中并从土壤中获取能量和养分的特殊酶, 对土壤微生物分解有机质以及对微生物吸收利用土壤C、N、P等养分元素发挥调节作用(Burns & Dick, 2002)。β-1,4-葡萄糖苷酶(β-1,4-glucosidase, BG)主要用于获取C; β-1,4-N-乙酰葡糖氨糖苷酶(β-1,4-N-acetylglucosaminidase, NAG)和亮氨酸氨基肽酶(leucine aminopeptidase, LAP)主要用于获取N; 酸性(或碱性)磷酸酶(acid or alkaline phosphatase, AP)主要用于获取P。Sinsabaugh等(2009)提出“生态酶化学计量”概念, 即胞外酶的比例关系(BG:(NAG + LAP):AP)或BG:NAG:AP, 可以反映土壤微生物C、N、P养分资源需求状况(Moorhead et al., 2016; Peng & Wang, 2016)。土壤微生物和胞外酶生态化学计量受土壤C、N、P资源有效性的调控, 可以为探讨土壤养分限制提供有价值的信息(Sinsabaugh et al., 2008; Sinsabaugh & Follstad Shah, 2012)。因此, 研究土壤微生物和胞外酶化学计量有助于深入了解土壤微生物在草地生态系统结构和功能、演替及恢复过程中的作用。
土壤C:N:P化学计量可以反映土壤C、N、P循环, 有助于确定生态过程对全球变化的响应, 因而成为土壤C、N、P平衡特征的重要参数(程滨等, 2010)。土壤C:N:P化学计量特征影响微生物生长、群落结构、微生物生物量C:N:P化学计量及微生物代谢活动(周正虎和王传宽, 2016)。由于土壤微生物数量、群落组成以及代谢活动的复杂性, 土壤微生物生物量C:N:P与土壤C:N:P之间的关系仍不明确。中国森林土壤微生物生物量碳氮格局整合分析表明, 土壤微生物生物量C:N随着土壤C:N的增加呈减小趋势(Zhou & Wang, 2015), 然而亚热带和喀斯特地区土壤微生物生物量C:N随着土壤C:N的增加而显著增加(Li et al., 2012; Hu et al., 2016); 也有研究认为土壤微生物生物量C:N变异性很小(Makino et al., 2003)。亚热带地区土壤微生物生物量C:P随着土壤C:P的增加而显著增加, 而土壤微生物生物量N:P随着土壤N:P的增加呈减小趋势(Li et al., 2012)。因此, 土壤微生物生物量C:N:P与土壤C:N:P是否存在协变关系, 土壤微生物化学计量与土壤化学计量关联性和强度还需要进一步验证。
土壤生态酶的相对活性与资源的有效性相耦合。土壤磷酸酶活性通常与土壤有效P含量成反比(Allison et al., 2007)。当土壤中微生物的生长受到P限制时, 土壤微生物将增加对磷酸酶的投资, 从而降低BG:AP和NAG:AP; 受N限制时, 土壤微生物通常不会消耗自身N素而产生更多的与N获取相关的胞外酶(Sinsabaugh & Follstad Shah, 2012); 当受到严重N限制时, 土壤微生物很可能在N限制解除之前处于休眠状态(Sinsabaugh & Follstad Shah, 2012; Blagodatskaya & Kuzyakov, 2013)。尽管目前关于土壤胞外酶对土壤养分变异的适应机制的认识已经取得很大进展, 然而, 由于土壤养分状况、气候条件、土壤质地等众多因素的影响, 不同区域和不同生态系统类型之间土壤胞外酶对土壤养分变异的适应机制仍然存在很大争议(Waring et al., 2014; Peng & Wang, 2016)。
近年来, 由于人类对草地资源的过度利用和全球气候变化, 造成草地生态系统不同程度的退化与沙漠化。草地沙漠化是草地退化的极端表现形式, 草地沙漠化过程中土壤C和养分衰减(李玉强等, 2005; Zuo et al., 2009)。关于土壤C、N、P的变化是否会影响与其关系密切的土壤微生物生物量C、N、P及微生物生物量C:N:P化学计量的变化没有一致结论。目前研究主要集中在草地沙漠化对土壤C、N、P含量的影响, 而对草地沙漠化对土壤C:N:P化学计量特征、土壤微生物生物量C、N、P含量、微生物生物量C:N:P化学计量、土壤胞外酶活性和胞外酶化学计量影响的研究较少。因此, 本研究以不同沙漠化阶段荒漠草地为研究对象, 分析荒漠草地沙漠化过程中土壤、土壤微生物和土壤胞外酶生态化学计量的变化特征及其内在关联性, 探讨荒漠草地沙漠化对土壤、土壤微生物C:N:P生态化学计量的影响, 揭示土壤微生物对土壤生态化学计量变异的响应机制, 为恢复半干旱地区退化荒漠草原, 遏制草地退化进程提供理论依据和实践指导。
1 材料和方法
1.1 研究区概况
研究区位于宁夏中北部盐池县(107.45° E, 37.82° N, 海拔1 295-1 951 m)。与陕西、甘肃、内蒙古毗邻, 属于鄂尔多斯台地和毛乌素沙地的过渡地带。气候属于中温带干旱气候, 年平均气温8.1 ℃, 1月份最冷(平均气温-8.6 ℃), 7月份最热(平均气温22.4 ℃), 年降水量280 mm, 降雨集中在7-9月, 年际降水变化率大, 年蒸发量为2 710 mm, 无霜期约165天, 年平均风速2.8 m·s-1 (刘任涛等, 2012)。地带性土壤主要有灰钙土, 非地带性土壤主要有风沙土、盐碱土和草甸土, 风沙土分布广泛, 土壤结构松散, 肥力极差。草地植被类型以温带草原和荒漠草原为主, 群落中常见植物种类以旱生和中旱生类型为主。主要优势植物有白草(Pennisetum centrasiaticum)、牛枝子(Lespedeza potaninii)、苦豆子(Sophora alopecuroides)、猪毛蒿(Artemisia scoparia)、沙蓬(Agriophyllum squarrosum)等。
1.2 样地选择
不同沙漠化过程伴随着植物群落类型和特征的演变过程, 沙地活化过程总体上表现为固定沙丘(地)—半固定沙丘(地)—半流动沙丘(地)—流动沙丘(地)的动态演化序列(丁国栋, 2004)。本研究采用空间序列代替时间演替的方法, 根据研究区植被的指示性及盖度分级标准在研究区域选择处于荒漠草原不同沙漠化阶段的荒漠草地、固定沙地、半固定沙地和流动沙地作为试验样地, 以荒漠草地为对照(唐庄生等, 2015)。不同程度的沙漠化草地植被特征表现为: 荒漠草地主要植被有猪毛菜(Salsola collina)、牛枝子、黑沙蒿(Artemisia ordosica)、甘草(Glycyrrhiza uralensis)等, 植被覆盖度70%-80%; 固定沙地主要分布的植被有草木樨状黄芪(Astragalus melilotoides)、针茅(Stipa capillata)和阿尔泰狗娃花(Heteropappus altaicus)等, 植被覆盖度60%-70%; 半固定沙地主要植被有苦豆子、白草等, 植被覆盖度40%-50%; 流动沙地主要植被有苦豆子、老瓜头(Cynanchum komarovii), 植被覆盖度小于10%。1.3 土壤样品采集
2017年7月下旬进行土壤样品采集, 在荒漠草地每个沙漠化阶段选择3个50 m × 50 m的重复取样区, 每个取样区间地形与环境条件基本一致。每个取样区内随机设置5个1 m × 1 m的小样方, 每个小样方内按“Z”形选取5点, 用直径9 cm的土钻采集0-10 cm混合土壤样品, 4种生境共取60个土样, 采集的土壤样品立即置于便携式冷藏箱。土壤样品过 2 mm筛后分成2份, 1份放入4 ℃保存2天, 用于测定土壤微生物生物量C、N、P含量和土壤胞外酶活性; 另1份土壤样品风干研磨后用于测定土壤C、N、P含量。1.4 土壤样品测定
土壤C、N、P含量测定: 土壤有机碳含量采用重铬酸钾氧化法测定; 土壤N含量用凯氏定氮法测定; 土壤P含量采用钼锑抗比色法测定。土壤微生物生物量C、N、P含量测定: 采用氯仿熏蒸-K2SO4浸提法测定土壤微生物生物量C、N、P含量。新鲜土壤经氯仿熏蒸处理, 熏蒸后用K2SO4浸提, 采用全自动有机碳分析仪(Vario TOC, Elementar, Langenselbold, Germany)测定微生物生物量C (MBC)含量, 采用全自动凯氏定氮仪(K9840, 海能仪器股份有限公司, 济南)测定微生物生物量N (MBN)含量, 采用钼锑抗比色法测定微生物生物量P (MBP)含量。
土壤胞外酶活性测定: BG、NAG和AP活性采用96微孔酶标板荧光分析法(Bell et al., 2013)测定。具体如下: 称取2.75 g新鲜土壤样品于三角瓶中, 加入91 mL缓冲液(pH值根据所测土壤样品调节), 200-220 r·min-1摇床振荡30 min。接种250 μL浓度为200 μmol·L-1的荧光标记的C、N、P基质液到深孔板中作为底物板。接种250 μL不同浓度的4-甲基伞形酮(MUB)于深孔板作为标线板。吸取800 μL土壤悬液到96孔板中, 所有样品加完以后用不干胶封口膜将深孔板密封, 将深孔板反复倒置摇晃混匀。所有深孔板放置在25 ℃培养箱中避光培养4 h。培养结束后将深孔板于4 ℃、4 800 r·min-1离心3 min后吸取200 μL上清液。在365 nm波长处激发, 450 nm波长处检测荧光。
1.5 数据计算与处理
(1)土壤微生物生物量碳、氮、磷含量的计算公式:式中, F为熏蒸土壤K2SO4浸提液中有机碳和氮含量, uF为未熏蒸土壤K2SO4浸提液中有机碳和氮含量, ${{K}_{\text{c}}}$为转换系数。MBC、MBN的转换系数均为0.45。
式中, Ec为熏蒸土壤NaHCO3提取液中无机P含量与未熏蒸土壤NaHCO3提取液中无机P含量差值, 0.4为浸提液微生物生物量磷占总微生物生物量磷的比例, R为加入无机磷的回收率。
(2)土壤胞外酶活性计算公式(Bell et al., 2013):
式中EA为酶活性; SEC为土壤样品酶浓度; SFR为土壤样品荧光原始值; T为培养时间; Soil为土壤干质量; 0.8表示土壤悬液0.8 mL; 91表示缓冲液91 mL。
(3) 土壤胞外酶生态化学计量计算公式(Sinsabaugh et al., 2009) :
式中EEA为土壤胞外酶活性; BG为β-葡萄糖苷酶活性; NAG为N-乙酰氨基葡萄糖苷酶活性; AP为磷酸酶活性。
土壤胞外酶(LAP+NAG)或NAG均可反映土壤微生物N养分资源的需求状况, 且BG:(NAG + LAP):AP或BG:NAG:AP均可表示土壤胞外酶生态化学计量关系(Sinsabaugh et al., 2009; Robert et al., 2012; Waring et al., 2014), 因此本研究只测定NAG活性, 土壤胞外酶生态化学计量采用BG:NAG:AP表示。
(4)土壤微生物养分利用效率的计算公式(Sinsabaugh et al., 2016):
其中${{S}_{\text{C:X}}}\text{=(1/}EE{{A}_{\text{C:X}}}\text{)(}{{B}_{\text{C:X}}}\text{/}{{L}_{\text{C:X}}}\text{)}$
其中${{S}_{\text{X:C}}}\text{=(1}-EE{{A}_{\text{X:C}}}\text{)(}{{B}_{\text{X:C}}}\text{/}{{L}_{\text{X:C}}}\text{)}$
式中X为N或P, KX、KC为半饱和常数, KX = 0.5, KC = 0.5; CUEmax是C供给微生物生长能量的上限, CUEmax = 0.6; XUEmax是N、P供给微生物生长能量的上限, XUEmax = 1.0; SC:X为微生物生物量C:N或者C:P、胞外酶活性和土壤C:N或者土壤C:P的比值; BC:X为微生物生物量C:N或微生物生物量C:P; LC:X为土壤C:N或者土壤C:P。
(5)数据处理方法
采用SPSS 17.0统计分析软件对实验数据进行分析。土壤C、N、P含量、土壤C:N:P生态化学计量、土壤微生物生物量C:N:P生态化学计量和土壤胞外酶生态化学计量采用单因素方差分析法进行分析, 所有数据均采用最小显著差异(LSD)法进行多重比较; 用Pearson相关系数对土壤-微生物生物量-胞外酶C:N:P生态化学计量进行相关分析; 用Sigmaplot 12.5软件作图。
2 结果和分析
2.1 不同沙漠化阶段土壤C、N、P含量及其生态化学计量特征
荒漠草地沙漠化显著影响土壤C、N、P含量(图1)。随着荒漠草地沙漠化程度的不断加剧, 土壤C、N、P含量均呈降低趋势。荒漠草地土壤C含量与半固定沙地和流动沙地差异显著(p < 0.05), 固定沙地、半固定沙地和流动沙地土壤C含量分别比荒漠草地降低了15.2%、47.0%和52.1%。荒漠草地土壤N含量显著高于其他沙漠化阶段(p < 0.05), 流动沙地较半固定沙地土壤N含量降低了20.0%。荒漠草地和固定沙地土壤P含量显著高于半固定沙地和流动沙地。荒漠草地土壤P含量分别是固定沙地、半固定沙地和流动沙地的1.04、1.33和1.41倍。图1
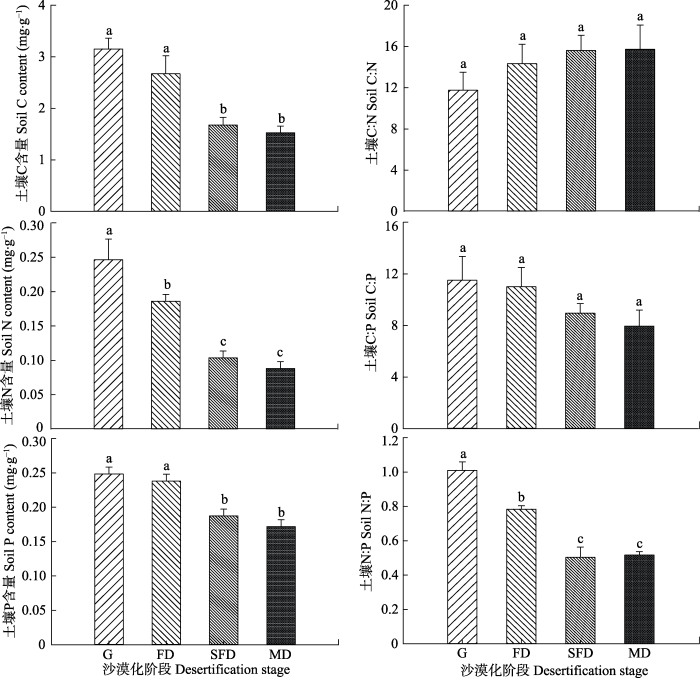 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1土壤C、N、P含量及其生态化学计量特征(平均值±标准偏差)。FD, 固定沙地; G, 荒漠草地; MD, 流动沙地; SFD, 半固定沙地。不同小写字母表示阶段的差异显著(p < 0.05)。
Fig. 1Characteristics of soil C、N、P content and C:N:P ecological stoichiometry (mean ± SD) at different stages of desertification. FD, fixed dunes; G, grasslands; MD, mobile dunes SFD, semi-fixed dunes. Different lowercase letters indicate significance differences among stages (p < 0.05).
荒漠草地沙漠化显著影响土壤N:P, 但对土壤C:N和C:P没有显著影响(图1)。荒漠草地不同沙漠化阶段土壤C:N、C:P差异不显著; 土壤N:P差异显著(p < 0.05)。荒漠草地沙漠化过程中土壤N:P呈不断降低趋势, 固定沙地、半固定沙地和流动沙地土壤N:P分别比荒漠草地降低了22.4%、50.1%和49.0%。
2.2 不同沙漠化阶段土壤微生物生物量C:N:P及土壤胞外酶生态化学计量特征
随着荒漠草地沙漠化程度的不断加剧, 不同沙漠化阶段土壤MBN:MBP差异显著(p < 0.05), 且沙漠化对土壤MBC:MBN和土壤MBC:MBP影响不显著(图2)。荒漠草地沙漠化过程中土壤MBN:MBP呈逐渐降低的趋势。荒漠草地发展至固定沙地, 土壤MBN:MBP降低了37.6%、荒漠草地退化至半固定沙地土壤MBN:MBP降低了46.6%, 荒漠草地退化至流动沙地土壤MBN:MBP降低了82.6%, 降低幅度随沙漠化程度的加剧逐渐增加。图2
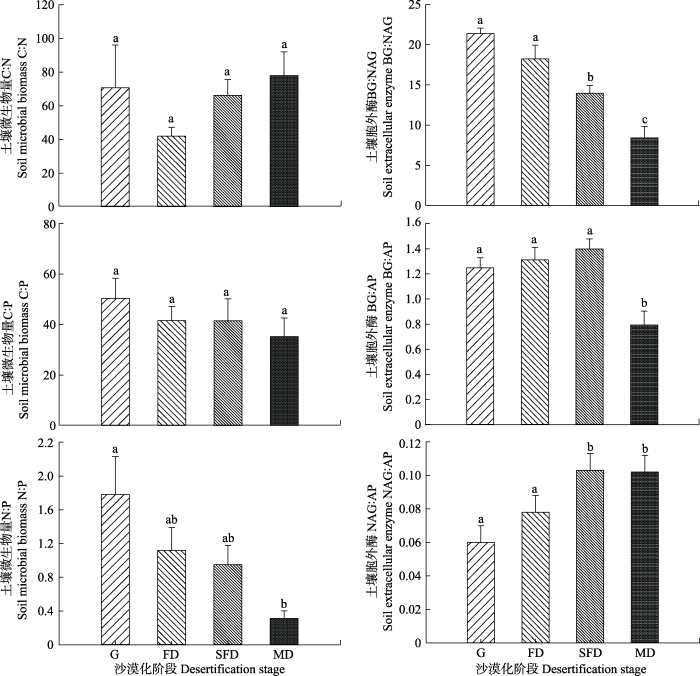 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2土壤微生物生物量及土壤胞外酶C:N:P生态化学计量特征(平均值±标准偏差)。FD, 固定沙地; G, 荒漠草地; MD, 流动沙地; SFD, 半固定沙地。AP, 磷酸酶; BG, β-葡萄糖苷酶; NAG, N-乙酰氨基葡萄糖苷酶。不同小写字母表示阶段的差异显著(p < 0.05)。
Fig. 2C:N:P ecological stoichiometry of soil microbial biomass and soil extracellular enzymes (mean ± SD) at different stages of desertification. FD, fixed dunes; G, grasslands; MD, mobile dunes SFD, semi-fixed dunes. AP, alkaline phosphatase; BG, β-1,4-glucosidase; NAG, β-1,4-N-acetylglucosaminidase. Different lowercase letters indicate significance differences among stages (p < 0.05).
荒漠草地沙漠化对土壤BG:NAG和NAG:AP影响显著(p < 0.05), 而对土壤BG:AP的影响不显著(图2)。随着荒漠草地沙漠化程度的加剧, 土壤BG:NAG呈降低趋势, 而土壤NAG:AP呈增加趋势。与荒漠草地相比, 固定沙地、半固定沙地和流动沙地土壤BG:NAG分别降低14.7%、34.7%和60.6%。荒漠草地到固定沙地、半固定沙地和流动沙地土壤NAG:AP分别增加了30.0%、71.6%和70.0%。土壤BG:AP在荒漠草地各沙漠化阶段整体表现为上升趋势。
2.3 荒漠草地土壤微生物养分利用效率
不同沙漠化阶段土壤微生物C利用效率CUEC:N差异显著(p < 0.05, 表1)。荒漠草地沙漠化过程中土壤微生物C利用效率CUEC:N表现为递增趋势。与荒漠草地相比, 固定沙地、半固定沙地和流动沙地CUEC:N分别增加了4.0%、44.0%和108.0%。荒漠草地沙漠化对土壤微生物N利用效率(NUEN:C)和土壤微生物P利用效率(PUEP:C)影响显著。随着荒漠草地沙漠化的不断加剧, NUEN:C呈先增后减的趋势, 而PUEP:C基本呈降低趋势。Table 1
表1
表1不同沙化阶段土壤微生物养分利用效率特征(平均值±标准偏差)
Table 1
| 沙化阶段 Desertification stage | CUEC:N | CUEC:P | NUEN:C | PUEP:C |
|---|---|---|---|---|
| 荒漠草地 Grasslands | 0.25 ± 0.06a | 0.84 ± 0.02a | 0.37 ± 0.06a | 0.13 ± 0.03a |
| 固定沙地 Fixed dunes | 0.26 ± 0.05a | 0.85 ± 0.03a | 0.40 ± 0.04ab | 0.08 ± 0.03a |
| 半固定沙地 Semi-fixed dunes | 0.36 ± 0.04ab | 0.85 ± 0.02a | 0.29 ± 0.05ab | 0.10 ± 0.02ab |
| 流动沙地 Mobile dunes | 0.52 ± 0.04b | 0.90 ± 0.01a | 0.23 ± 0.02b | 0.01 ± 0.01b |
新窗口打开|下载CSV
2.4 土壤-土壤微生物生物量-胞外酶C:N:P生态化学计量的相关性分析
土壤C:N:P生态化学计量、土壤微生物生物量C:N:P生态化学计量和土壤胞外酶生态化学计量的相关分析表明, 土壤C:N与土壤C:P呈极显著正相关关系, 而与土壤N:P呈极显著负相关关系(p < 0.01, 表2)。土壤N:P与土壤MBC:MBP、MBN:MBP、BG:NAG呈显著正相关, 而与土壤NAG:AP呈显著负相关关系(p < 0.01)。土壤MBN:MBP与土壤MBC:MBN和土壤NAG:AP呈极显著负相关关系, 而与土壤MBC:MBP和土壤BG:NAG呈极显著正相关关系。土壤BG:NAG与土壤BG:AP表现为极显著正相关, 与土壤NAG:AP表现为极显著负相关。荒漠草地土壤C:N, MBC:MBN分别与土壤N:P、MBN:MBP显著负相关, 土壤C:N、BG:NAG分别与土壤C:P、BG:AP显著正相关。土壤微生物生物量MBC:MBP、MBN:MBP随着土壤N:P增加呈增加趋势, 而土壤NAG:AP呈降低趋势。Table 2
表2
表2土壤-微生物-胞外酶生态化学计量的相关性分析
Table 2
| Soil C:N | Soil C:P | Soil N:P | MBC:MBN | MBC:MBP | MBN:MBP | BG:NAG | BG:AP | NAG:AP | ||
|---|---|---|---|---|---|---|---|---|---|---|
| Soil C:N | 1 | 0.760** | -0.396* | 0.149 | -0.118 | -0.143 | -0.249 | -0.194 | 0.131 | |
| Soil C:P | 1 | 0.159 | 0.100 | 0.096 | 0.260 | 0.230 | 0.071 | -0.207 | ||
| Soil N:P | 1 | -0.157 | 0.435* | 0.573** | 0.543** | 0.217 | -0.427** | |||
| MBC:MBN | 1 | -0.038 | -0.604** | -0.150 | -0.185 | 0.039 | ||||
| MBC:MBP | 1 | 0.570** | 0.198 | 0.149 | -0.224 | |||||
| MBN:MBP | 1 | 0.427* | 0.203 | -0.366* | ||||||
| BG:NAG | 1 | 0.542** | -0.706** | |||||||
| BG:AP | 1 | 0.102 | ||||||||
| NAG:AP | 1 | |||||||||
新窗口打开|下载CSV
3 讨论
3.1 荒漠草地沙漠化对土壤C、N、P含量及其生态化学计量的影响
土壤是植物生长发育的重要基质, 其养分含量对于评价土壤质量状况至关重要(徐阳春等, 2002)。土壤C、N、P含量随着荒漠草地沙漠化程度加剧的变化趋势, 与高寒草甸退化过程中土壤C、N、P含量逐渐降低的趋势一致(罗亚勇等, 2012), 表明荒漠草地沙漠化对土壤养分含量的积累有负效应, 原因可能是随着荒漠草地沙漠化的加剧, 中亚白草、赖草、牛枝子等优势植物逐渐减少, 草地生物多样性降低, 植被盖度、生物量、枯落物以及微生物活动均趋于减少(Zhang & McBear, 2016), 影响了土壤有机质的积累。土壤N主要来自土壤有机质; 土壤有机C主要来源于植物凋落物和根系有机碳等(余健等, 2014)。土壤P与土壤C、N不同, 土壤P主要来自岩石风化, 其含量高低更多取决于土壤的质地特征(Chen & Li, 2003), 且随着荒漠草地沙漠化的不断加剧, 土壤选择性风蚀富含养分的土壤细颗粒, 导致了土壤的粗化和沙质化(赵丽莉等, 2013), 使得土壤P含量下降, 此外还可能由于牲畜的采食将P元素移出草地生态系统。土壤C、N、P含量的下降表明荒漠草地沙漠化过程中土壤内的养分积累和能量循环受到限制。土壤C:N:P可以反映土壤内C、N、P养分循环,也可以衡量土壤有机质和土壤养分平衡状况(Tian et al., 2010)。随着荒漠草地沙漠化的不断加剧, 土壤C:N呈逐渐增加的趋势, 土壤C:P和N:P呈降低趋势。其中土壤C:N与高寒草甸随着沙漠化程度的加剧逐渐降低的趋势相反, 表明不同草地生态系统土壤C:N存在较大的异质性(罗亚勇等, 2012)。Chapin等(2011)研究表明, 土壤C:N与土壤有机质分解速率成反比关系, 土壤C:N较高时, N素相对充足, 用来满足微生物的生长代谢, 土壤C:N较低时, 满足微生物生长之余的N素就会释放到土壤中。本研究区荒漠草地阶段土壤C:N最低, 说明荒漠草地阶段土壤有机质分解速率最高, 土壤N素相对充足。土壤C:P和土壤N:P随草地沙漠化不断加剧呈降低的趋势, 荒漠草地沙漠化过程中, 土壤C和N的变化比P的变化更加敏感。另外本研究中土壤C:P的平均值低于全国水平(25.77)(Tian et al., 2010), 参考第二次全国土壤普查分级标准发现, 本研究区土壤C的平均含量(0.225%)处于土壤碳含量极缺乏阶段(程欢等, 2018), 土壤P平均含量(0.021%)也明显低于全国水平(0.056%)(Han et al., 2005), 而土壤C含量相对土壤P含量降低更严重, 进而导致土壤C:P值偏低。荒漠草地土壤N:P高于其他沙漠化阶段, 表明固定沙地、半固定沙地和流动沙地土壤N元素供应相对较弱, 并有可能成为此阶段植被生长的限制元素。
3.2 荒漠草地沙漠化对土壤微生物及胞外酶生态化学计量的影响
土壤微生物作为草地土壤中最活跃的营养库, 其生态化学计量对于探究草地生态系统限制性养分具有重要意义。本研究中土壤MBC:MBN平均值高于中国土壤MBC:MBN平均值7.6, 而土壤MBN:MBP平均值低于中国土壤MBN:MBP的平均值5.6 (Xu et al., 2013), 其主要原因可能是荒漠草地土壤微生物N含量较低; 土壤MBC:MBP与全球土壤MBC:MBP的平均值42.4接近(Xu et al., 2013)。随着荒漠草地沙漠化程度的不断加剧, 土壤MBC: MBP和MBN:MBP均表现出逐渐降低趋势, 这可能与荒漠草地沙漠化过程中生物多样性的降低以及植被变化有关, 而土壤MBN:MBP值的降低也有可能是由于微生物的生长需要更多富含磷的核糖体RNA的投资导致(Elser et al., 2003)。王宝荣等(2018)研究黄土丘陵区关于土壤微生物生物量生态化学计量的影响因素发现, 植被类型的变化会显著影响土壤MBC:MBN和MBC:MBP, 本研究区随着荒漠草地的不断沙化, 优势草本植物相继退化, 沙米、白刺等沙生植物出现, 使得土壤微生物生物量发生变化。此外Cleveland和Liptzin (2007)也认为不同植物种可能导致凋落物质量和数量以及微生物群落组成的改变, 进而会影响土壤微生物生态化学计量。土壤MBC:MBP可作为土壤中微生物吸收固持P潜力的指标, 土壤MBC:MBP一般在7-30之间, 土壤MBC:MBP值低表明微生物在矿化土壤有机质中释放P的潜力较大, 土壤MBC:MBP值高则表明土壤微生物对土壤有效P有同化趋势, 且微生物与植物竞争吸收土壤有效P, 固P能力较强(彭佩钦等, 2006)。本研究中荒漠草地MBC:MBP值在35.25-50.39之间, 表明微生物和荒漠草地植被竞争土壤有效P, 导致荒漠草地植被对土壤P的利用率低。土壤胞外酶活性可以反映微生物生长和代谢过程的能量限制, 并与土壤C、N、P分解密切相关(王冰冰等, 2015)。而不同土壤胞外酶活性的比值又与养分浓度或者土壤C:N:P值密切相关, 可以用来评价土壤微生物C、N、P养分资源的需求状况(Schimel & Weintraub, 2003)。有研究发现土壤P的有效性与土壤磷酸酶活性呈负相关关系, 而土壤N对微生物产生磷酸酶有促进作用(Allison et al., 2007; Olander & Vitousek, 2000)。本研究中, 从荒漠草地退化至固定沙地, 土壤BG:NAG逐渐降低, 而土壤BG:AP、NAG:AP基本表现为上升趋势, 土壤微生物降低了对磷酸酶的投资, 表明本研究区土壤微生物的生长受C、N限制较为突出。另外土壤BG:NAG不断下降也可能与荒漠草地沙漠化过程中土壤养分含量的降低以及植被变化等有关。荒漠草地高的土壤BG:NAG、BG:AP反映出荒漠草地土壤微生物N、P功能的发挥相对较差; 而半固定沙地较高的土壤NAG:AP, 表明半固定沙地土壤微生物N资源功能发挥较好。
3.3 荒漠草地沙漠化对土壤微生物的元素利用效率的影响
消费者驱动的养分循环理论表明土壤C:N:P和土壤微生物C:N:P以及微生物的元素利用效率间的平衡对生态系统C、N、P的循环有直接影响, 微生物消费者驱动的养分循环理论指出微生物会通过保留自身的组成元素来维持代谢需求的限制性元素, 并且能够主动排出多余元素(Sterner & Elser, 2002)。荒漠草地沙漠化过程中, 随着土壤C:N逐渐增加, CUEC:N表现出增加趋势, 而NUEN:C和PUEP:C均有逐渐降低趋势, 这与Mooshammer等(2012)的结论一致, 表明荒漠草地土壤微生物生长主要受C源的限制。而土壤C积累又主要来自土壤有机质的输入(贾国梅等, 2016), 随着荒漠草地沙漠化的不断加剧, 土壤养分含量降低, 使得荒漠草地植被减少, 进一步影响枯落物的覆盖度, 导致土壤有机质的输入受到阻碍, 从而影响到土壤C的积累。流动沙地土壤微生物养分利用效率NUEN:C和PUEP:C达到最低, 表明流动沙地土壤微生物生长受C的限制较为严重。3.4 土壤-微生物-胞外酶C:N:P生态化学计量的相关性分析
不同生态系统土壤N:P与土壤MBN:MBP的相关性存在明显差异, 以往研究显示黄土丘陵地区土壤和微生物N:P显著正相关(吴建平等, 2016); 稻田、高地和林地等生态系统中土壤N:P与土壤MBN:MBP呈负相关关系(Li et al., 2012), 也有研究发现土壤和土壤微生物N:P并无相关关系(Cleveland & Liptzin, 2007; 王宝荣等, 2018)。本研究区土壤N:P与土壤MBN:MBP的正相关关系与黄土丘陵地区结果一致, 反映出荒漠草地土壤微生物生物量N:P不符合“Redfield”比率, 荒漠草地土壤微生物生物量不具有内稳性, 表明“土壤-微生物”系统内物质转换主要受C、N、P养分循环的调控。N、P元素也被认为是陆地生态系统生物生长的主要限制元素, 其比值关系也常用来预测生态系统养分限制状况, 且土壤MBN:MBP比土壤N:P反映养分限制更加敏感。有研究报道全国土壤N:P的平均值为5.2和土壤MBN:MBP的平均值为5.6 (Tian et al., 2010; Xu et al., 2013), 本研究区土壤N:P和MBN:MBP平均值(0.70和1.91)低于全国尺度土壤N:P和MBN:MBP。其主要原因可能是本研究区土壤N素相对贫瘠, 使得土壤微生物所能固持的微生物生物量N含量偏低, 同时表明本研究区生物生长受N素限制较为严重。另外土壤N:P与NAG:AP有极显著的负相关关系, 这与岷江干旱河谷优势灌丛群落土壤生态酶化学计量与土壤生态化学计量的关系基本一致(王冰冰等, 2015), 土壤MBN:MBP与NAG:AP有显著负相关关系, 土壤胞外酶对土壤微生物养分资源的利用可以反映土壤微生物生物量的元素组成, 体现了N、P元素在土壤内的养分循环途径。土壤C:P与土壤MBC:MBP无显著相关关系, 与三峡库区不同植被土壤C:P与土壤MBC:MBP的结果一致(贾国梅等, 2016); 土壤C:N、C:P与土壤MBN:MBP无显著相关性与Cleveland和Liptzin (2007)的研究结果相似, 导致这种现象可能是由于土壤微生物生物量C:N:P受区域气候, 土壤质地等因素的干扰(Zhou & Wang, 2015)。而Mooshammer等(2014)也证实了土壤微生物生物量C:N:P的变异本质更多由微生物群落结构变异来解释。土壤C:N、MBC:MBN、MBC:MBP分别与土壤BG:NAG、BG:AP无相关关系, 表明土壤胞外酶活性和土壤微生物生物量对荒漠草地沙漠化过程中土壤养分变化的响应机理不同。综上所述, 面对土壤C:N:P生态化学计量的变异, 土壤微生物和胞外酶会调整自身生物量C:N:P生态化学计量进行适应。N、P元素在“土壤-微生物-胞外酶”系统的循环机制和土壤微生物C:N:P及胞外酶BG:NAG:AP随土壤养分及其生态化学计量的协变关系能够为理解荒漠草地C、N、P的物质循环机制以及C、N、P在草地生态系统物质循环过程中的耦合关系提供思路。

参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOIURL [本文引用: 2]

During primary succession, the abundance of carbon (C) and nitrogen (N) in soil increases, while phosphorus (P) declines. These changes in nutrient concentrations in organic matter are likely to play an important role in controlling enzyme-mediated nutrient mineralization. We examined how enzyme activity and efficiency changed with successional time in organic and mineral soils taken from the 120 000-year-old Franz Josef soil development sequence, New Zealand, and the relationship between enzyme activity and efficiency and soil nutrient concentrations. We found that the activity of enzymes involved in P mineralization increased with site age across the Franz Josef chronosequence, while the activity of enzymes regulating C and N mineralization declined in organic but not mineral soil. Sulfatase activity peaked at an intermediate-aged site, possibly indicating a transient period of S limitation. The activity of phosphatase enzymes was negatively correlated with the concentration of P in the soil, whereas activity of C-, N- and S-hydrolyzing enzymes was not strongly dependent on nutrient concentrations. When assessed as efficiency (activity per unit microbial biomass), there were strong patterns of increasing efficiency of P-, and decreasing efficiency of C- and N-hydrolyzing enzymes with site age. We suggest that activity patterns for C-, N- and S-hydrolyzing enzymes were obscured by simultaneous and opposing changes in enzyme efficiency and microbial biomass. In mineral soil, efficiency of enzymes was negatively correlated with soil nutrient availability. In contrast, in organic soil, efficiency of C-, N- and S-hydrolyzing enzymes was positively correlated with soil P, while efficiency of P-hydrolyzing enzymes was negatively correlated with soil P. The increase in efficiency of P-hydrolyzing enzymes, and decrease in efficiency of C-, N- and S-hydrolyzing enzymes with site age was accompanied by a shift in microbial community composition towards higher relative abundances of fungi. Changes in enzyme efficiency with site age are likely to be due to both constitutive differences in enzyme production, and down-regulation of enzyme expression.
DOIURL [本文引用: 1]
Ecological stoichiometry is a useful tool for studying how the elemental composition of organisms and their food affects production, nutrient cycling, and food-web dynamics. Two analyses are presented here that show that the use of simple element ratios in stoichiometric calculations may in certain circumstances prove inadequate because of the influence in animal nutrition of biochemical aspects of diet. In the first, a stoichio-metric analysis of herbivores consuming food with varying carbon to nitrogen (C:N) ratios is undertaken, in which the intake of C is segregated into easily assimilated compounds and fiber. Two herbivore strategies emerge from the analysis, both as a means of minimizing limitation by C, not N: fiber eaters that consume high C:N food and have efficient fiber digestion, and selective feeders that consume low C:N food but that do not possess fiber-digesting enzymes. In the second example, the stoichiometric axiom that a single substrate, the one in least supply relative to demand, limits growth is used to identify potentially limiting essential amino acids in the diets of a range of animals. Large consumer-prey imbalances in amino acids were found in several cases, indicating that, at least in theory, growth should be strongly limited by individual amino acids rather than bulk N. In practice such limitation may be offset in consumers by physiological and other factors such as symbiotic relationships. The two analyses emphasize the simplicity of element stoichiometry, highlighting the need to consider biochemical and physiological arguments when undertaking stoichiometric studies of carbon and nutrient transfers in ecosystems.
DOIURLPMID [本文引用: 1]
Microbes in soils and other environments produce extracellular enzymes to depolymerize and hydrolyze organic macromolecules so that they can be assimilated for energy and nutrients. Measuring soil microbial enzyme activity is crucial in understanding soil ecosystem functional dynamics. The general concept of the fluorescence enzyme assay is that synthetic C-, N-, or P-rich substrates bound with a fluorescent dye are added to soil samples. When intact, the labeled substrates do not fluoresce. Enzyme activity is measured as the increase in fluorescence as the fluorescent dyes are cleaved from their substrates, which allows them to fluoresce. Enzyme measurements can be expressed in units of molarity or activity. To perform this assay, soil slurries are prepared by combining soil with a pH buffer. The pH buffer (typically a 50 mM sodium acetate or 50 mM Tris buffer), is chosen for the buffer's particular acid dissociation constant (pKa) to best match the soil sample pH. The soil slurries are inoculated with a nonlimiting amount of fluorescently labeled (i.e. C-, N-, or P-rich) substrate. Using soil slurries in the assay serves to minimize limitations on enzyme and substrate diffusion. Therefore, this assay controls for differences in substrate limitation, diffusion rates, and soil pH conditions; thus detecting potential enzyme activity rates as a function of the difference in enzyme concentrations (per sample). Fluorescence enzyme assays are typically more sensitive than spectrophotometric (i.e. colorimetric) assays, but can suffer from interference caused by impurities and the instability of many fluorescent compounds when exposed to light; so caution is required when handling fluorescent substrates. Likewise, this method only assesses potential enzyme activities under laboratory conditions when substrates are not limiting. Caution should be used when interpreting the data representing cross-site comparisons with differing temperatures or soil types, as in situ soil type and temperature can influence enzyme kinetics.
DOIURL [本文引用: 1]
61We critically evaluated literature on active soil microorganisms and revealed.61Microorganisms in active and potentially active state: tiny versus large pool.61Threshold parameters for microbial activity state in soil.61Relevance of active microbial fraction in calculations of process rates.61Combinations of dynamic approaches enable to evaluate active microbial biomass.
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
生态化学计量学结合生物学、化学和物理学等基本原理,研究能量和碳、氮、磷等化学元素在生态系统中,特别是各种生态系统过程(如竞争、捕食、寄生、共生等)参与者中的变化,以及它们之间的动态平衡,并分析这种平衡对生态系统的影响.目前,C:N:P化学计量学研究已深入到生态学的各个层次(细胞、个体、种群、群落、生态系统)及区域等不同尺度.近年来,由于认识到化学计量学研究可以把生态实体的各个层次在元素水平上统一起来,因此生态化学计量学已成为许多生态系统的新兴研究工具.其中,C:N:P化学计量学是各种生态过程研究中的核心内容.论述了生态化学计量学在物种、群落、生态系统等各层次的应用现状,并指出了C:N:P化学计量学研究的应用前景和发展趋势,以期引起同行的重视并推动该领域的进一步发展.
URL [本文引用: 1]
生态化学计量学结合生物学、化学和物理学等基本原理,研究能量和碳、氮、磷等化学元素在生态系统中,特别是各种生态系统过程(如竞争、捕食、寄生、共生等)参与者中的变化,以及它们之间的动态平衡,并分析这种平衡对生态系统的影响.目前,C:N:P化学计量学研究已深入到生态学的各个层次(细胞、个体、种群、群落、生态系统)及区域等不同尺度.近年来,由于认识到化学计量学研究可以把生态实体的各个层次在元素水平上统一起来,因此生态化学计量学已成为许多生态系统的新兴研究工具.其中,C:N:P化学计量学是各种生态过程研究中的核心内容.论述了生态化学计量学在物种、群落、生态系统等各层次的应用现状,并指出了C:N:P化学计量学研究的应用前景和发展趋势,以期引起同行的重视并推动该领域的进一步发展.
URL [本文引用: 1]
论文通过研究川西亚高山/高山生态系统不同海拔分布典型土壤类型SOC、TN、TP及其生态化学计量学特征,对比《四川土壤》1985年调查成果,评价我国川西亚高山/高山典型土壤恢复状况。测定亚高山草甸土、草甸土、暗棕壤、棕壤、黄棕壤、褐土的腐殖质层、淀积层、母质层土壤SOC、TN、TP含量,计算生态化学计量值。结果表明:土壤SOC含量表现为亚高山草甸土草甸土暗棕壤褐土黄棕壤棕壤,TN含量表现为亚高山草甸土草甸土暗棕壤褐土黄棕壤棕壤,TP含量表现为暗棕壤亚高山草甸土草甸土棕壤褐土黄棕壤;暗棕壤、棕壤基本表现为SOC、TN、TP含量随土层加深递减;依据第二次全国土壤普查分级标准,研究区土壤有机质呈很丰富水平,TN呈丰富水平,TP呈缺乏水平。土壤SOC、TN和TP水平分布从南向北呈先增加后减少。化学计量比特征:土壤碳氮比表现为草甸土褐土黄棕壤亚高山草甸土暗棕壤棕壤,土壤碳磷比表现为草甸土黄棕壤褐土暗棕壤亚高山草甸土棕壤,土壤氮磷比表现为草甸土暗棕壤黄棕壤褐土亚高山草甸土棕壤,TP是主要限制因子。对比1985年调查结果,经过近30 a的恢复,亚高山草甸土、草甸土、黄棕壤、褐土土壤SOC含量呈增加趋势,棕壤SOC含量下降幅度最大,2015年仅为1985年的31.06%;土壤TN变化不大;TP含量呈下降趋势,变化幅度在56.41%~87.85%之间。
URL [本文引用: 1]
论文通过研究川西亚高山/高山生态系统不同海拔分布典型土壤类型SOC、TN、TP及其生态化学计量学特征,对比《四川土壤》1985年调查成果,评价我国川西亚高山/高山典型土壤恢复状况。测定亚高山草甸土、草甸土、暗棕壤、棕壤、黄棕壤、褐土的腐殖质层、淀积层、母质层土壤SOC、TN、TP含量,计算生态化学计量值。结果表明:土壤SOC含量表现为亚高山草甸土草甸土暗棕壤褐土黄棕壤棕壤,TN含量表现为亚高山草甸土草甸土暗棕壤褐土黄棕壤棕壤,TP含量表现为暗棕壤亚高山草甸土草甸土棕壤褐土黄棕壤;暗棕壤、棕壤基本表现为SOC、TN、TP含量随土层加深递减;依据第二次全国土壤普查分级标准,研究区土壤有机质呈很丰富水平,TN呈丰富水平,TP呈缺乏水平。土壤SOC、TN和TP水平分布从南向北呈先增加后减少。化学计量比特征:土壤碳氮比表现为草甸土褐土黄棕壤亚高山草甸土暗棕壤棕壤,土壤碳磷比表现为草甸土黄棕壤褐土暗棕壤亚高山草甸土棕壤,土壤氮磷比表现为草甸土暗棕壤黄棕壤褐土亚高山草甸土棕壤,TP是主要限制因子。对比1985年调查结果,经过近30 a的恢复,亚高山草甸土、草甸土、黄棕壤、褐土土壤SOC含量呈增加趋势,棕壤SOC含量下降幅度最大,2015年仅为1985年的31.06%;土壤TN变化不大;TP含量呈下降趋势,变化幅度在56.41%~87.85%之间。
DOIURL [本文引用: 1]
The effects of human disturbance on soil carbon and nutrients storage are of great concern. We investigated changes in soil carbon and nutrients as a primary forest of mixed broad-leaved Korean pine was changed into seven ecosystems by local human disturbance. We also studied these changes due to brick manufacturing. The primary forest had higher total soil C than potato crop, wetland and plantation types, respectively. The total soil N was higher in grassland than it in the primary forest. The total soil P was lower in plantation type compared with it in the primary forest. The total soil K was higher in potato crop, wetland, secondary forests and plantation than it in the primary forest. More than 40% of soil S was lost as the primary forest was changed into potato crop, wetland and plantation. The ratios of C:N, C:P, C:K and C:S generally were higher in the primary forest than those in the other seven ecosystems. More than 80% of soil C, N and S were lost during brick making. Our study showed that human disturbance could greatly affect soil C and nutrient storage as the primary Korean pine forests were altered.
DOIURL [本文引用: 3]
DOIURL [本文引用: 1]
Based upon analysis to current research status of desertification assessment indicator systems, a new viewpoint that takes vegetation as main factor for desertification assessment is brought forward. Taking Mu Us Sandland as a research example, by analyzing desertification characteristics in different land types, the succession series of the main indicative vegetation community is put forward. Based on the field observation data about regulations of windblown sand movement and sand-driving wind, the protecting effect of vegetation on land surface is studied, and the vegetation cover grading standards for windy erosion desertification assessment are put out.
DOIURL [本文引用: 1]
Based upon analysis to current research status of desertification assessment indicator systems, a new viewpoint that takes vegetation as main factor for desertification assessment is brought forward. Taking Mu Us Sandland as a research example, by analyzing desertification characteristics in different land types, the succession series of the main indicative vegetation community is put forward. Based on the field observation data about regulations of windblown sand movement and sand-driving wind, the protecting effect of vegetation on land surface is studied, and the vegetation cover grading standards for windy erosion desertification assessment are put out.
DOIURL [本文引用: 1]
DOIURLPMID [本文引用: 1]
$\bullet$ Leaf nitrogen and phosphorus stoichiometry of Chinese terrestrial plants was studied based on a national data set including 753 species across the country. $\bullet$ Geometric means were calculated for functional groups based on life form, phylogeny and photosynthetic pathway, as well as for all 753 species. The relationships between leaf N and P stoichiometric traits and latitude (and temperature) were analysed. $\bullet$ The geometric means of leaf N, P, and N : P ratio for the 753 species were 18.6 and $1.21 mg g^{-1}$ and 14.4, respectively. With increasing latitude (decreasing mean annual temperature, MAT), leaf N and P increased, but the N : P ratio did not show significant changes. $\bullet$ Although patterns of leaf N, P and N : P ratios across the functional groups were generally consistent with those reported previously, the overall N : P ratio of China's flora was considerably higher than the global averages, probably caused by a greater shortage of soil P in China than elsewhere. The relationships between leaf N, P and N : P ratio and latitude (and MAT) also suggested the existence of broad biogeographical patterns of these leaf traits in Chinese flora.
DOIURL [本文引用: 1]
篇首: 从1909年丹麦哥本哈根大学Johannes Warming(1841-1924)出版第一本生态学教科书到现在,生态学经历了100年的发展.作为研究生物与生物、生物 与环境相互关系的学科,生态学具有高度的综合性和交叉性,研究的问题也具有复杂性和多尺度的特点.正因为这样的学科特点,传统上生态学家多强调研究对象的 特殊性、研究问题的局域性,缺乏统一的理论框架.
DOIURL [本文引用: 1]
篇首: 从1909年丹麦哥本哈根大学Johannes Warming(1841-1924)出版第一本生态学教科书到现在,生态学经历了100年的发展.作为研究生物与生物、生物 与环境相互关系的学科,生态学具有高度的综合性和交叉性,研究的问题也具有复杂性和多尺度的特点.正因为这样的学科特点,传统上生态学家多强调研究对象的 特殊性、研究问题的局域性,缺乏统一的理论框架.
DOIURL [本文引用: 1]
61Microbial biomass and activity increased with the restoration of soil physicochemical properties.61The increased microbial quotient and decreased metabolic quotient reflect the restoration of degraded habitats.61Microbial C:N ratio tended to increase as an adaptive response to resource stoichiometry.
URL [本文引用: 2]
土壤微生物通过矿化和固持营养而显著影响土壤肥力。以湖北宜昌点军区三种植被类型(柏树、橘树、菜地)的土壤作为研究对象,对土壤微生物量碳氮磷及其生态化学计量进行了研究,探讨了不同植被覆盖土壤微生物量碳氮磷特征及它们之间的联系。结果表明:不同植被覆盖显著改变土壤微生物量碳氮磷含量,土壤微生物量碳(MBC)、氮(MBN)都是柏树地显著大于柑橘地和菜地,而菜地和柑橘地之间无显著性差异。土壤微生物量磷(MBP)的大小顺序为菜地〉柏树地〉橘树地。相关性分析的结果表明土壤有机碳、全氮和微生物量碳氮互相之间具有显著的正相关性(p〈0.01),土壤全磷与土壤微生物量磷之间也具有显著的正相关性(p〈0.01),这意味着土壤微生物量可以作为土壤肥力变化的指标。三种植被类型的土壤MBC/MBN之间都无显著性差异,而土壤MBC/MBP和MBN/MBP的变化顺序是橘树地〉柏树地〉菜地。MBC/MBP和MBN/MBP与MBP之间具有显著的负相关性(p〈0.01),而微生物碳氮与MBC/MBN,MBN/MBP和MBC/MBP之间都无显著的相关性。这意味着土壤MBC/MBP和MBN/MBP的变化主要与微生物量磷具有显著的关系。
URL [本文引用: 2]
土壤微生物通过矿化和固持营养而显著影响土壤肥力。以湖北宜昌点军区三种植被类型(柏树、橘树、菜地)的土壤作为研究对象,对土壤微生物量碳氮磷及其生态化学计量进行了研究,探讨了不同植被覆盖土壤微生物量碳氮磷特征及它们之间的联系。结果表明:不同植被覆盖显著改变土壤微生物量碳氮磷含量,土壤微生物量碳(MBC)、氮(MBN)都是柏树地显著大于柑橘地和菜地,而菜地和柑橘地之间无显著性差异。土壤微生物量磷(MBP)的大小顺序为菜地〉柏树地〉橘树地。相关性分析的结果表明土壤有机碳、全氮和微生物量碳氮互相之间具有显著的正相关性(p〈0.01),土壤全磷与土壤微生物量磷之间也具有显著的正相关性(p〈0.01),这意味着土壤微生物量可以作为土壤肥力变化的指标。三种植被类型的土壤MBC/MBN之间都无显著性差异,而土壤MBC/MBP和MBN/MBP的变化顺序是橘树地〉柏树地〉菜地。MBC/MBP和MBN/MBP与MBP之间具有显著的负相关性(p〈0.01),而微生物碳氮与MBC/MBN,MBN/MBP和MBC/MBP之间都无显著的相关性。这意味着土壤MBC/MBP和MBN/MBP的变化主要与微生物量磷具有显著的关系。
DOIURL [本文引用: 1]
Soil organic carbon and total nitrogen concentrations,soil texture,and carbon and nitrogen densities in different stages of desertification process(Potential,Light,Medium,Severe,and Most severe desertification) in Horqin sandy land were studied.The results showed that along the spectrum from potential desertification to most severe desertification,soil organic carbon and nitrogen concentrations decreased by 91.6% and 85.8%,respectively,to a depth of 100 cm,versus decreases of 95.7% for the fine particle size fractions(<0.1 mm).Carbon density decreased from 4 877.9 to 481.8(C) g/m~2 during deserification process,whereas nitrogen density decreased from 518.0 to 89.4(N) g/m~2.Regression analysis revealed significant positive relationships amongthe soil carbon concentration,soil nitrogen concentration,and soil fine particle size fractions(<0.1 mm) content for all five stages.Soil carbon and nitrogen were lost more rapidly during the initial stages of desertification than the later stages.The soil C/N ratio decreased from 9.385 to 5.541 along the desertification developing.There was a greater proportional decline in soil carbon than in nitrogen.
DOIURL [本文引用: 1]
Soil organic carbon and total nitrogen concentrations,soil texture,and carbon and nitrogen densities in different stages of desertification process(Potential,Light,Medium,Severe,and Most severe desertification) in Horqin sandy land were studied.The results showed that along the spectrum from potential desertification to most severe desertification,soil organic carbon and nitrogen concentrations decreased by 91.6% and 85.8%,respectively,to a depth of 100 cm,versus decreases of 95.7% for the fine particle size fractions(<0.1 mm).Carbon density decreased from 4 877.9 to 481.8(C) g/m~2 during deserification process,whereas nitrogen density decreased from 518.0 to 89.4(N) g/m~2.Regression analysis revealed significant positive relationships amongthe soil carbon concentration,soil nitrogen concentration,and soil fine particle size fractions(<0.1 mm) content for all five stages.Soil carbon and nitrogen were lost more rapidly during the initial stages of desertification than the later stages.The soil C/N ratio decreased from 9.385 to 5.541 along the desertification developing.There was a greater proportional decline in soil carbon than in nitrogen.
DOIURL [本文引用: 3]
[1] Does the soil microbial biomass (SMB) in terrestrial ecosystems present well- constrained atomic carbon:nitrogen:phosphorus (C:N:P) ratios, analogous to the planktonic biomass in marine ecosystems? How do soil microbes respond to changes in the soil environment in terms of their elemental stoichiometry? Following up on the work of Cleveland and Liptzin (2007), we examined the stoichiometry of C, N and P in the soil and SMB and their relationships at both the landscape and land-use levels in subtropical terrestrial ecosystems. 1,069 soil samples were collected at a depth of 0 20 cm from three typical landscapes (a karst mountain, a low hill and a lowland) in southern subtropical China. The landscapes presented various land-use types (e.g., paddy field, upland, woodland, etc.) and intensities of anthropogenic activity. The samples were analyzed to determine soil organic C, total soil N and total soil P contents as well as SMB C, SMB N and SMB P. On average, atomic C:N:P ratios of 80:7.9:1 in the soil and 70.2:6:1 in the SMB were obtained for the region. A clear descending trend of the soil C:N:P ratios (not the SMB C:N:P ratios) was observed across the three landscapes in the order: karst mountain > low hill > lowland. Although significant variations primarily related to human activities were observed in the soil and SMB atomic C:N:P ratios across the landscapes and land-use types, a significant correlation (r = 0.56,p< 0.001) was found between the soil and SMB C:P ratios in the entire data set; however, the correlation for the comparable N:P ratios was not evident. Significant correlations between the soil and SMB C:N, C:P and even N:P ratios (mainly in the woodland) were also observed variably at the finer level of the landscape or land-use. The tendency for a C:N:P stoichiometric relationship to exist between microbes and the soil environment found in this study might suggest possible non-homeostasis of elemental stoichiometry in the SMB of the terrestrial ecosystems in southern subtropical China.
URL [本文引用: 1]
研究荒漠草原区柠条人工林生长过程中土壤-植被系统演变特征,对于揭示柠条人工固沙林对沙化草地生态系统恢复产生的生态效应具有重要的科学意义.以6、15、24和36年生柠条人工林为对象,通过调查每个样地的土壤性质和地表植被特征,分析了荒漠草原区柠条人工林生长过程中土壤-地表植被的变化.结果表明:随着柠条人工林的发育,柠条的冠幅、树高、分枝数和基径极显著增加,土壤粗沙粒和细沙粒含量显著降低,极细沙和粘粉粒含量显著增加,土壤有机碳、全N和全P含量呈现线性增加,而土壤pH值则显著下降;地表植被物种数和密度显著增加,地表植被盖度和高度均表现为24年生>15年生>6年生>36年生林地,相关分析表明,土壤质地、容重、土壤养分与pH值是影响柠条林地地表植被物种数、个体数和盖度分布的主要因素.在荒漠草原区,柠条人工固沙林发育过程有利于土壤条件改善和地表植被恢复,促进沙化草地生态系统的恢复.
URL [本文引用: 1]
研究荒漠草原区柠条人工林生长过程中土壤-植被系统演变特征,对于揭示柠条人工固沙林对沙化草地生态系统恢复产生的生态效应具有重要的科学意义.以6、15、24和36年生柠条人工林为对象,通过调查每个样地的土壤性质和地表植被特征,分析了荒漠草原区柠条人工林生长过程中土壤-地表植被的变化.结果表明:随着柠条人工林的发育,柠条的冠幅、树高、分枝数和基径极显著增加,土壤粗沙粒和细沙粒含量显著降低,极细沙和粘粉粒含量显著增加,土壤有机碳、全N和全P含量呈现线性增加,而土壤pH值则显著下降;地表植被物种数和密度显著增加,地表植被盖度和高度均表现为24年生>15年生>6年生>36年生林地,相关分析表明,土壤质地、容重、土壤养分与pH值是影响柠条林地地表植被物种数、个体数和盖度分布的主要因素.在荒漠草原区,柠条人工固沙林发育过程有利于土壤条件改善和地表植被恢复,促进沙化草地生态系统的恢复.
URL [本文引用: 2]
To clarify the soil stoichiometry characteristics of alpine meadow at its different degradation stages, the native Kobresia meadow, mildly degraded meadow, and seriously degraded sandy meadow were selected to measure their soil organic carbon (SOC), total nitrogen (N), total phosphorous (P), and total potassium (K) contents at different soil depths. With the degradation of alpine meadow, the SOC, N, P, K, C∶N, C∶P, C∶K, N∶P, N∶K, and P∶K in 0-100 cm profiles presently a decreasing trend, and SOC was most sensitive to the degradation, followed by total N, P, and K. The SOC and N in top (0-20 cm) soil could be used as the most sensitive indicators for the degradation degree of alpine meadow. The vertical distribution of SOC, N, P, and K differed with the degradation stage of the alpine meadow. For native Kobresia meadow and mildly degraded meadow, the SOC, N, P, K, C∶N, C∶P, C∶K, N∶P, N∶K, and P∶K in 0-40 cm soil layer decreased sharply with soil depth, but those in 40-100 cm soil layer had a slow decrease and tended to be stable; for seriously degraded sandy meadow, these stoichiometry characteristics remained unchanged with soil depth.
URL [本文引用: 2]
To clarify the soil stoichiometry characteristics of alpine meadow at its different degradation stages, the native Kobresia meadow, mildly degraded meadow, and seriously degraded sandy meadow were selected to measure their soil organic carbon (SOC), total nitrogen (N), total phosphorous (P), and total potassium (K) contents at different soil depths. With the degradation of alpine meadow, the SOC, N, P, K, C∶N, C∶P, C∶K, N∶P, N∶K, and P∶K in 0-100 cm profiles presently a decreasing trend, and SOC was most sensitive to the degradation, followed by total N, P, and K. The SOC and N in top (0-20 cm) soil could be used as the most sensitive indicators for the degradation degree of alpine meadow. The vertical distribution of SOC, N, P, and K differed with the degradation stage of the alpine meadow. For native Kobresia meadow and mildly degraded meadow, the SOC, N, P, K, C∶N, C∶P, C∶K, N∶P, N∶K, and P∶K in 0-40 cm soil layer decreased sharply with soil depth, but those in 40-100 cm soil layer had a slow decrease and tended to be stable; for seriously degraded sandy meadow, these stoichiometry characteristics remained unchanged with soil depth.
DOIURL [本文引用: 1]
1. We examined the relative importance of resource composition (carbon: phosphorus molar ratios which varied between 9 and 933) and growth rate$(0\cdot 5-1\cdot 5\ \text{h}^{-1})$to biomass carbon: nitrogen: phosphorus stoichiometry and nucleic acid content in Escherichia coli grown in chemostats, and in other heterotrophic prokaryotes using published literature. 2. Escherichia coli RNA content and the contribution of RNA-P to total cellular P increased with increasing growth rate at all supply C: P ratios. Growth rate had a much stronger effect on biomass C: P than did supply C: P, and increased RNA content resulted in low biomass C: P and N: P ratios. 3. However, we observed only twofold variations in biomass C: P and N: P ratios in the experiments, despite a difference of two orders of magnitude in C: P and N: P supply. The response of biomass C: P and N: P ratios to alteration of the supply C: P and N: P ratios revealed that E. coli was strongly homeostatic in its elemental composition. 4. This result, and a literature survey, suggest that each heterotrophic bacterial strain regulates its elemental composition homeostatically within a relatively narrow range of characteristic biomass C: P and N: P ratios. 5. Thus shifts in the dominance of different bacterial strains in the environment are probably responsible for the large variation in bacterial biomass C: P, as has been suggested for crustacean zooplankton. These findings indicate that bacteria are more like animals than plants in terms of biomass C: P and N: P homeostasis.
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
Abstract The primary goal of this study was to devise a quantitative method of interpreting the simultaneous microbial C, N, and P acquisition through the activities of four, key extracellular enzymes, 1,4- -glucosidase (BG), leucine aminopeptidase (LAP), 1,4- -N-acetylglucosaminidase (NAG) and acid/alkaline phosphatases (AP). To this end, the proportional activity of C vs. N acquiring enzymes (BG/[BG + NAG + LAP]) was plotted against C vs. P acquiring enzymes (BG/[BG + AP]). We then calculated the length and angle of the vector created by connecting a line between the plot origin and point represented by these proportions; the length quantifies relative C vs. nutrient limitation and the angle quantifies the relative P vs. N limitation. Analyses of large data sets obtained from soil, freshwater periphyton and aquatic sediments revealed that logarithmic, arcsine, arcsine-square-root and logit transformations did little to improve the statistical distribution of data over raw proportions. More importantly, the vector characteristics of enzyme activity loci are much easier to interpret for raw proportions than when data have been previously transformed. Analyses also revealed the importance of using consistent methods, i.e., omitting NAG assays for an acidic soil led to overestimates of P limitations, and the importance of understanding the nature of the system, i.e., soils of the Antarctic Dry Valleys had low BG activities, likely because there is little to no local production of cellulose. Further, analyses of four sites in Luquillo Forest, Puerto Rico, showed no differences among sites in relative C demand despite differences in BG activities, and higher P demand in the cloud than lower montane forest, despite higher AP activity at the latter site. Finally, analyses of three decomposing litter types over time revealed contrasting patterns of change in relative C, N and P demand over time, reflecting both stoichiometric and C quality effects on decomposer communities. Thus enzyme activity vectors reflect the simultaneous, relative resource demands of the microbial community independent of variations in total enzyme activity, such as may result from variations in total microbial biomass.
DOIURLPMID [本文引用: 1]
Resource stoichiometry (C:N:P) is an important determinant of litter decomposition. However, the effect of elemental stoichiometry on the gross rates of microbial N and P cycling processes during litter decomposition is unknown. In a mesocosm experiment, beech (Fagus sylvatica L.) litter with natural differences in elemental stoichiometry (C:N:P) was incubated under constant environmental conditions. After three and six months, we measured various aspects of nitrogen and phosphorus cycling. We found that gross protein depolymerization, N mineralization (ammonification), and nitrification rates were negatively related to litter C:N. Rates of P mineralization were negatively correlated with litter C:P. The negative correlations with litter C:N were stronger for inorganic N cycling processes than for gross protein depolymerization, indicating that the effect of resource stoichiometry on intracellular processes was stronger than on processes catalyzed by extracellular enzymes. Consistent with this, extracellular protein depolymerization was mainly limited by substrate availability and less so by the amount of protease. Strong positive correlations between the interconnected N and P pools and the respective production and consumption processes pointed to feed-forward control of microbial litter N and P cycling. A negative relationship between litter C:N and phosphatase activity (and between litter C:P and protease activity) demonstrated that microbes tended to allocate carbon and nutrients in ample supply into the production of extracellular enzymes to mine for the nutrient that is more limiting. Overall, the study demonstrated a strong effect of litter stoichiometry (C:N:P) on gross processes of microbial N and P cycling in decomposing litter; mineralization of N and P were tightly coupled to assist in maintaining cellular homeostasis of litter microbial communities.
DOIURLPMID [本文引用: 1]
Terrestrial microbial decomposer communities thrive on a wide range of organic matter types that rarely ever meet their elemental demands. In this review we synthesize the current state-of-the-art of microbial adaptations to resource stoichiometry, in order to gain a deeper understanding of the interactions between heterotrophic microbial communities and their chemical environment. The stoichiometric imbalance between microbial communities and their organic substrates generally decreases from wood to leaf litter and further to topsoil and subsoil organic matter. Microbial communities can respond to these imbalances in four ways: first, they adapt their biomass composition toward their resource in a non-homeostatic behavior. Such changes are, however, only moderate, and occur mainly because of changes in microbial community structure and less so due to cellular storage of elements in excess. Second, microbial communities can mobilize resources that meet their elemental demand by producing specific extracellular enzymes, which, in turn, is restricted by the C and N requirement for enzyme production itself. Third, microbes can regulate their element use efficiencies (ratio of element invested in growth over total element uptake), such that they release elements in excess depending on their demand (e.g., respiration and N mineralization). Fourth, diazotrophic bacteria and saprotrophic fungi may trigger the input of external N and P to decomposer communities. Theoretical considerations show that adjustments in element use efficiencies may be the most important mechanism by which microbes regulate their biomass stoichiometry. This review summarizes different views on how microbes cope with imbalanced supply of C, N and P, thereby providing a framework for integrating and linking microbial adaptation to resource imbalances to ecosystem scale fluxes across scales and ecosystems.
DOIURL [本文引用: 1]
Soil microorganisms and plants produce enzymes that mineralize organically bound nutrients. When nutrient availability is low, the biota may be able to increase production of these enzymes to enhance the supply of inorganic nitrogen (N) and phosphorus (P). Regulation of enzyme production may be a point where N and P cycles interact. We measured acid phosphatase and chitinase (N-acetyl -D-glucosaminide) activity in soil across a chronosequence in Hawaii where N and P availability varies substantially among sites and long term fertilizer plots had been maintained for over 4 years. Phosphatase activity was high at all sites. Chitinase activity decreased significantly as age and N availability increased across the chronosequence. Phosphorus addition suppressed phosphatase activity at all sites, while N addition increased phosphatase activity at the young, N-limited site. In contrast, N addition repressed chitinase activity only at the N limited young site, and P additions had no effect on chitinase activity. These results suggest that the regulatory relationship between nutrient supply and nutrient mineralization are asymmetric for N and P, and that the differences could help to explain differences observed in patterns of N and P availability.
DOIURL [本文引用: 1]
以湖南省沅江市典型湖垸为代表,通过密集取样分析,研究了洞庭湖 区不同利用方式条件下农田土壤微生物生物量碳、氮、磷的变化及其和土壤碳、氮、磷的关系,发现水田土壤碳、氮和微生物生物量碳、氮明显高于旱地,水田土壤 中双季稻高于一季稻;土壤磷的含量旱地稍高于水田,但土壤微生物生物量磷水田稍高于旱地.尽管在水田土壤中微生物生物量碳、氮有明显的不同,但水田土壤微 生物生物量磷维持在相对稳定的水平.典型样区土壤微生物生物量碳占有机碳的比例为0.65%~7.24%,平均3.00%;土壤微生物生物量氮占全氮的比 例为0.98%~7.41%,平均3.81%;土壤微生物生物量磷占全磷的比例为0.16%~7.54%,平均2.80%.土壤C/N为 3.87~17.31,平均9.15;BC/BN为4.06~9.29,平均7.26.土壤微生物生物量碳、氮与土壤碳、氮之间存在极其显著的线性相关关 系,但土壤微生物生物量磷占全磷之间相关关系不显著.土壤微生物生物量碳、氮、磷之间的相关关系达到了极显著水平.不同的利用方式和耕作制度导致了土壤 碳、氮和微生物生物量碳、氮的差异,土壤微生物生物量碳、氮能够很好地反映洞庭湖区农田土壤碳、氮水平.
DOIURL [本文引用: 1]
以湖南省沅江市典型湖垸为代表,通过密集取样分析,研究了洞庭湖 区不同利用方式条件下农田土壤微生物生物量碳、氮、磷的变化及其和土壤碳、氮、磷的关系,发现水田土壤碳、氮和微生物生物量碳、氮明显高于旱地,水田土壤 中双季稻高于一季稻;土壤磷的含量旱地稍高于水田,但土壤微生物生物量磷水田稍高于旱地.尽管在水田土壤中微生物生物量碳、氮有明显的不同,但水田土壤微 生物生物量磷维持在相对稳定的水平.典型样区土壤微生物生物量碳占有机碳的比例为0.65%~7.24%,平均3.00%;土壤微生物生物量氮占全氮的比 例为0.98%~7.41%,平均3.81%;土壤微生物生物量磷占全磷的比例为0.16%~7.54%,平均2.80%.土壤C/N为 3.87~17.31,平均9.15;BC/BN为4.06~9.29,平均7.26.土壤微生物生物量碳、氮与土壤碳、氮之间存在极其显著的线性相关关 系,但土壤微生物生物量磷占全磷之间相关关系不显著.土壤微生物生物量碳、氮、磷之间的相关关系达到了极显著水平.不同的利用方式和耕作制度导致了土壤 碳、氮和微生物生物量碳、氮的差异,土壤微生物生物量碳、氮能够很好地反映洞庭湖区农田土壤碳、氮水平.
DOIURL [本文引用: 2]
61Ratios of log-transformed C-, N- and P-acquiring enzymes were 1:1.2:1.4.61Enzyme C:N and C:P ratios declined with soil depth, while enzyme N:P ratio remained stable.61Soil extracellular enzyme activity ratio was not in homeostasis but resource dependent on soil and microbial biomass stoichiometry.
[本文引用: 1]
DOIURL [本文引用: 3]
DOIURL [本文引用: 3]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 3]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 2]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
Changes in enzyme activities and soil microbial community composition along carbon and nutrient gradients at the Franz Josef chronosequence, New Zealand
2
2007
... 土壤生态酶的相对活性与资源的有效性相耦合.土壤磷酸酶活性通常与土壤有效P含量成反比(
... 土壤胞外酶活性可以反映微生物生长和代谢过程的能量限制, 并与土壤C、N、P分解密切相关(
Stoichiometry: Linking elements to biochemical
1
2004
... 生态化学计量学以环境中相互作用的生物体以及生物体与环境之间的物质平衡来认识生态动力学, 以元素组成来揭示生物化学和生态学之间的联系, 从而分析和预测生态系统的结构和动力学(
High-throughput fluorometric measurement of potential soil extracellular enzyme activities
1
2013
... 土壤胞外酶活性测定: BG、NAG和AP活性采用96微孔酶标板荧光分析法(
Active microorganisms in soil: Critical review of estimation criteria and approaches
1
2013
... 土壤生态酶的相对活性与资源的有效性相耦合.土壤磷酸酶活性通常与土壤有效P含量成反比(
1
2002
... 土壤微生物是土壤生态系统的重要组成部分, 参与土壤有机质分解、腐殖质形成、土壤养分转化和循环等过程.土壤胞外酶是微生物分泌到土壤中并从土壤中获取能量和养分的特殊酶, 对土壤微生物分解有机质以及对微生物吸收利用土壤C、N、P等养分元素发挥调节作用(
1
2011
... 土壤C:N:P可以反映土壤内C、N、P养分循环,也可以衡量土壤有机质和土壤养分平衡状况(
生态化学计量学研究进展
1
2010
... 土壤C:N:P化学计量可以反映土壤C、N、P循环, 有助于确定生态过程对全球变化的响应, 因而成为土壤C、N、P平衡特征的重要参数(
生态化学计量学研究进展
1
2010
... 土壤C:N:P化学计量可以反映土壤C、N、P循环, 有助于确定生态过程对全球变化的响应, 因而成为土壤C、N、P平衡特征的重要参数(
川西亚高山/高山典型土壤类型有机碳、氮、磷含量及其生态化学计量特征
1
2018
... 土壤C:N:P可以反映土壤内C、N、P养分循环,也可以衡量土壤有机质和土壤养分平衡状况(
川西亚高山/高山典型土壤类型有机碳、氮、磷含量及其生态化学计量特征
1
2018
... 土壤C:N:P可以反映土壤内C、N、P养分循环,也可以衡量土壤有机质和土壤养分平衡状况(
Change in soil carbon and nutrient storage after human disturbance of a primary Korean pine forest in Northeast China
1
2003
... 土壤是植物生长发育的重要基质, 其养分含量对于评价土壤质量状况至关重要(
C:N:P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomass
3
2007
... 土壤微生物作为草地土壤中最活跃的营养库, 其生态化学计量对于探究草地生态系统限制性养分具有重要意义.本研究中土壤MBC:MBN平均值高于中国土壤MBC:MBN平均值7.6, 而土壤MBN:MBP平均值低于中国土壤MBN:MBP的平均值5.6 (
... 不同生态系统土壤N:P与土壤MBN:MBP的相关性存在明显差异, 以往研究显示黄土丘陵地区土壤和微生物N:P显著正相关(
... 土壤C:P与土壤MBC:MBP无显著相关关系, 与三峡库区不同植被土壤C:P与土壤MBC:MBP的结果一致(
区域荒漠化评价中植被的指示性及盖度分级标准研究——以毛乌素沙区为例
1
2004
... 不同沙漠化过程伴随着植物群落类型和特征的演变过程, 沙地活化过程总体上表现为固定沙丘(地)—半固定沙丘(地)—半流动沙丘(地)—流动沙丘(地)的动态演化序列(
区域荒漠化评价中植被的指示性及盖度分级标准研究——以毛乌素沙区为例
1
2004
... 不同沙漠化过程伴随着植物群落类型和特征的演变过程, 沙地活化过程总体上表现为固定沙丘(地)—半固定沙丘(地)—半流动沙丘(地)—流动沙丘(地)的动态演化序列(
Growth rate-stoichiometry couplings in diverse biota
1
2003
... 土壤微生物作为草地土壤中最活跃的营养库, 其生态化学计量对于探究草地生态系统限制性养分具有重要意义.本研究中土壤MBC:MBN平均值高于中国土壤MBC:MBN平均值7.6, 而土壤MBN:MBP平均值低于中国土壤MBN:MBP的平均值5.6 (
Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China
1
2005
... 土壤C:N:P可以反映土壤内C、N、P养分循环,也可以衡量土壤有机质和土壤养分平衡状况(
生态化学计量学: 探索从个体到生态系统的统一化理论
1
2010
... 生态化学计量学以环境中相互作用的生物体以及生物体与环境之间的物质平衡来认识生态动力学, 以元素组成来揭示生物化学和生态学之间的联系, 从而分析和预测生态系统的结构和动力学(
生态化学计量学: 探索从个体到生态系统的统一化理论
1
2010
... 生态化学计量学以环境中相互作用的生物体以及生物体与环境之间的物质平衡来认识生态动力学, 以元素组成来揭示生物化学和生态学之间的联系, 从而分析和预测生态系统的结构和动力学(
Community size, activity and C:N stoichiometry of soil microorganisms following reforestation in a karst region
1
2016
... 土壤C:N:P化学计量可以反映土壤C、N、P循环, 有助于确定生态过程对全球变化的响应, 因而成为土壤C、N、P平衡特征的重要参数(
三峡库区不同植被土壤微生物生物量碳氮磷生态化学计量特征
2
2016
... 消费者驱动的养分循环理论表明土壤C:N:P和土壤微生物C:N:P以及微生物的元素利用效率间的平衡对生态系统C、N、P的循环有直接影响, 微生物消费者驱动的养分循环理论指出微生物会通过保留自身的组成元素来维持代谢需求的限制性元素, 并且能够主动排出多余元素(
... 土壤C:P与土壤MBC:MBP无显著相关关系, 与三峡库区不同植被土壤C:P与土壤MBC:MBP的结果一致(
三峡库区不同植被土壤微生物生物量碳氮磷生态化学计量特征
2
2016
... 消费者驱动的养分循环理论表明土壤C:N:P和土壤微生物C:N:P以及微生物的元素利用效率间的平衡对生态系统C、N、P的循环有直接影响, 微生物消费者驱动的养分循环理论指出微生物会通过保留自身的组成元素来维持代谢需求的限制性元素, 并且能够主动排出多余元素(
... 土壤C:P与土壤MBC:MBP无显著相关关系, 与三峡库区不同植被土壤C:P与土壤MBC:MBP的结果一致(
科尔沁沙地沙漠化过程中土壤碳氮特征
1
2005
... 近年来, 由于人类对草地资源的过度利用和全球气候变化, 造成草地生态系统不同程度的退化与沙漠化.草地沙漠化是草地退化的极端表现形式, 草地沙漠化过程中土壤C和养分衰减(
科尔沁沙地沙漠化过程中土壤碳氮特征
1
2005
... 近年来, 由于人类对草地资源的过度利用和全球气候变化, 造成草地生态系统不同程度的退化与沙漠化.草地沙漠化是草地退化的极端表现形式, 草地沙漠化过程中土壤C和养分衰减(
Is the C:N:P stoichiometry in soil and soil microbial biomass related to the landscape and land use in southern subtropical China?
3
2012
... 土壤C:N:P化学计量可以反映土壤C、N、P循环, 有助于确定生态过程对全球变化的响应, 因而成为土壤C、N、P平衡特征的重要参数(
... ).亚热带地区土壤微生物生物量C:P随着土壤C:P的增加而显著增加, 而土壤微生物生物量N:P随着土壤N:P的增加呈减小趋势(
... 不同生态系统土壤N:P与土壤MBN:MBP的相关性存在明显差异, 以往研究显示黄土丘陵地区土壤和微生物N:P显著正相关(
荒漠草原区柠条人工固沙林生长过程中地表植被-土壤的变化
1
2012
... 研究区位于宁夏中北部盐池县(107.45° E, 37.82° N, 海拔1 295-1 951 m).与陕西、甘肃、内蒙古毗邻, 属于鄂尔多斯台地和毛乌素沙地的过渡地带.气候属于中温带干旱气候, 年平均气温8.1 ℃, 1月份最冷(平均气温-8.6 ℃), 7月份最热(平均气温22.4 ℃), 年降水量280 mm, 降雨集中在7-9月, 年际降水变化率大, 年蒸发量为2 710 mm, 无霜期约165天, 年平均风速2.8 m·s-1 (
荒漠草原区柠条人工固沙林生长过程中地表植被-土壤的变化
1
2012
... 研究区位于宁夏中北部盐池县(107.45° E, 37.82° N, 海拔1 295-1 951 m).与陕西、甘肃、内蒙古毗邻, 属于鄂尔多斯台地和毛乌素沙地的过渡地带.气候属于中温带干旱气候, 年平均气温8.1 ℃, 1月份最冷(平均气温-8.6 ℃), 7月份最热(平均气温22.4 ℃), 年降水量280 mm, 降雨集中在7-9月, 年际降水变化率大, 年蒸发量为2 710 mm, 无霜期约165天, 年平均风速2.8 m·s-1 (
不同退化阶段高寒草甸土壤化学计量特征
2
2012
... 土壤是植物生长发育的重要基质, 其养分含量对于评价土壤质量状况至关重要(
... 土壤C:N:P可以反映土壤内C、N、P养分循环,也可以衡量土壤有机质和土壤养分平衡状况(
不同退化阶段高寒草甸土壤化学计量特征
2
2012
... 土壤是植物生长发育的重要基质, 其养分含量对于评价土壤质量状况至关重要(
... 土壤C:N:P可以反映土壤内C、N、P养分循环,也可以衡量土壤有机质和土壤养分平衡状况(
Are bacteria more like plants or animals? Growth rate and resource dependence of bacterial C:N:P stoichiometry
1
2003
... 土壤C:N:P化学计量可以反映土壤C、N、P循环, 有助于确定生态过程对全球变化的响应, 因而成为土壤C、N、P平衡特征的重要参数(
The ratios of life
1
2003
... 生态化学计量学以环境中相互作用的生物体以及生物体与环境之间的物质平衡来认识生态动力学, 以元素组成来揭示生物化学和生态学之间的联系, 从而分析和预测生态系统的结构和动力学(
Vector analysis of ecoenzyme activities reveal constraints on coupled C, N and P dynamics
1
2016
... 土壤微生物是土壤生态系统的重要组成部分, 参与土壤有机质分解、腐殖质形成、土壤养分转化和循环等过程.土壤胞外酶是微生物分泌到土壤中并从土壤中获取能量和养分的特殊酶, 对土壤微生物分解有机质以及对微生物吸收利用土壤C、N、P等养分元素发挥调节作用(
Stoichiometric controls of nitrogen and phosphorus cycling in decomposing beech leaf litter
1
2012
... 消费者驱动的养分循环理论表明土壤C:N:P和土壤微生物C:N:P以及微生物的元素利用效率间的平衡对生态系统C、N、P的循环有直接影响, 微生物消费者驱动的养分循环理论指出微生物会通过保留自身的组成元素来维持代谢需求的限制性元素, 并且能够主动排出多余元素(
Stoichiometric imbalances between terrestrial decomposer communities and their resources: Mechanisms and implications of microbial adaptations to their resources
1
2014
... 土壤C:P与土壤MBC:MBP无显著相关关系, 与三峡库区不同植被土壤C:P与土壤MBC:MBP的结果一致(
Regulation of soil phosphatase and chitinase activity by N and P availability
1
2000
... 土壤胞外酶活性可以反映微生物生长和代谢过程的能量限制, 并与土壤C、N、P分解密切相关(
洞庭湖区不同利用方式对土壤微生物生物量碳氮磷的影响
1
2006
... 土壤微生物作为草地土壤中最活跃的营养库, 其生态化学计量对于探究草地生态系统限制性养分具有重要意义.本研究中土壤MBC:MBN平均值高于中国土壤MBC:MBN平均值7.6, 而土壤MBN:MBP平均值低于中国土壤MBN:MBP的平均值5.6 (
洞庭湖区不同利用方式对土壤微生物生物量碳氮磷的影响
1
2006
... 土壤微生物作为草地土壤中最活跃的营养库, 其生态化学计量对于探究草地生态系统限制性养分具有重要意义.本研究中土壤MBC:MBN平均值高于中国土壤MBC:MBN平均值7.6, 而土壤MBN:MBP平均值低于中国土壤MBN:MBP的平均值5.6 (
Stoichiometry of soil extracellular enzyme activity along a climatic transect in temperate grasslands of northern China
2
2016
... 土壤微生物是土壤生态系统的重要组成部分, 参与土壤有机质分解、腐殖质形成、土壤养分转化和循环等过程.土壤胞外酶是微生物分泌到土壤中并从土壤中获取能量和养分的特殊酶, 对土壤微生物分解有机质以及对微生物吸收利用土壤C、N、P等养分元素发挥调节作用(
... 土壤生态酶的相对活性与资源的有效性相耦合.土壤磷酸酶活性通常与土壤有效P含量成反比(
The implications of exoenzyme activity on microbial carbon and nitrogen limitation in soil: A theoretical model
1
2003
... 土壤胞外酶活性可以反映微生物生长和代谢过程的能量限制, 并与土壤C、N、P分解密切相关(
Ecoenzymatic stoichiometry and ecological theory
3
2012
... 土壤微生物是土壤生态系统的重要组成部分, 参与土壤有机质分解、腐殖质形成、土壤养分转化和循环等过程.土壤胞外酶是微生物分泌到土壤中并从土壤中获取能量和养分的特殊酶, 对土壤微生物分解有机质以及对微生物吸收利用土壤C、N、P等养分元素发挥调节作用(
... 土壤生态酶的相对活性与资源的有效性相耦合.土壤磷酸酶活性通常与土壤有效P含量成反比(
... ); 当受到严重N限制时, 土壤微生物很可能在N限制解除之前处于休眠状态(
Ecoenymatic stoichiometry of microbial organic nutrient acquisition in soil and sediment
3
2009
... 土壤微生物是土壤生态系统的重要组成部分, 参与土壤有机质分解、腐殖质形成、土壤养分转化和循环等过程.土壤胞外酶是微生物分泌到土壤中并从土壤中获取能量和养分的特殊酶, 对土壤微生物分解有机质以及对微生物吸收利用土壤C、N、P等养分元素发挥调节作用(
... (3) 土壤胞外酶生态化学计量计算公式(
... 土壤胞外酶(LAP+NAG)或NAG均可反映土壤微生物N养分资源的需求状况, 且BG:(NAG + LAP):AP或BG:NAG:AP均可表示土壤胞外酶生态化学计量关系(
Stoichiometry of soil enzyme activity at global scale
1
2008
... 土壤微生物是土壤生态系统的重要组成部分, 参与土壤有机质分解、腐殖质形成、土壤养分转化和循环等过程.土壤胞外酶是微生物分泌到土壤中并从土壤中获取能量和养分的特殊酶, 对土壤微生物分解有机质以及对微生物吸收利用土壤C、N、P等养分元素发挥调节作用(
Stoichiometry of microbial carbon use efficiency in soils
1
2016
... (4)土壤微生物养分利用效率的计算公式(
2
2002
... 生态化学计量学以环境中相互作用的生物体以及生物体与环境之间的物质平衡来认识生态动力学, 以元素组成来揭示生物化学和生态学之间的联系, 从而分析和预测生态系统的结构和动力学(
... 消费者驱动的养分循环理论表明土壤C:N:P和土壤微生物C:N:P以及微生物的元素利用效率间的平衡对生态系统C、N、P的循环有直接影响, 微生物消费者驱动的养分循环理论指出微生物会通过保留自身的组成元素来维持代谢需求的限制性元素, 并且能够主动排出多余元素(
荒漠草原沙漠化对土壤养分与植被根冠比的影响
1
2015
... 不同沙漠化过程伴随着植物群落类型和特征的演变过程, 沙地活化过程总体上表现为固定沙丘(地)—半固定沙丘(地)—半流动沙丘(地)—流动沙丘(地)的动态演化序列(
荒漠草原沙漠化对土壤养分与植被根冠比的影响
1
2015
... 不同沙漠化过程伴随着植物群落类型和特征的演变过程, 沙地活化过程总体上表现为固定沙丘(地)—半固定沙丘(地)—半流动沙丘(地)—流动沙丘(地)的动态演化序列(
Pattern and variation of C:N:P ratios in China’s soils: A synthesis of observational data
3
2010
... 土壤C:N:P可以反映土壤内C、N、P养分循环,也可以衡量土壤有机质和土壤养分平衡状况(
... 研究表明, 土壤C:N与土壤有机质分解速率成反比关系, 土壤C:N较高时, N素相对充足, 用来满足微生物的生长代谢, 土壤C:N较低时, 满足微生物生长之余的N素就会释放到土壤中.本研究区荒漠草地阶段土壤C:N最低, 说明荒漠草地阶段土壤有机质分解速率最高, 土壤N素相对充足.土壤C:P和土壤N:P随草地沙漠化不断加剧呈降低的趋势, 荒漠草地沙漠化过程中, 土壤C和N的变化比P的变化更加敏感.另外本研究中土壤C:P的平均值低于全国水平(25.77)(
... 不同生态系统土壤N:P与土壤MBN:MBP的相关性存在明显差异, 以往研究显示黄土丘陵地区土壤和微生物N:P显著正相关(
岷江上游干旱河谷优势灌丛群落土壤生态酶化学计量特征
2
2015
... 土壤胞外酶活性可以反映微生物生长和代谢过程的能量限制, 并与土壤C、N、P分解密切相关(
... 不同生态系统土壤N:P与土壤MBN:MBP的相关性存在明显差异, 以往研究显示黄土丘陵地区土壤和微生物N:P显著正相关(
岷江上游干旱河谷优势灌丛群落土壤生态酶化学计量特征
2
2015
... 土壤胞外酶活性可以反映微生物生长和代谢过程的能量限制, 并与土壤C、N、P分解密切相关(
... 不同生态系统土壤N:P与土壤MBN:MBP的相关性存在明显差异, 以往研究显示黄土丘陵地区土壤和微生物N:P显著正相关(
黄土丘陵区植被与地形特征对土壤和土壤微生物生物量生态化学计量特征的影响
2
2018
... 土壤微生物作为草地土壤中最活跃的营养库, 其生态化学计量对于探究草地生态系统限制性养分具有重要意义.本研究中土壤MBC:MBN平均值高于中国土壤MBC:MBN平均值7.6, 而土壤MBN:MBP平均值低于中国土壤MBN:MBP的平均值5.6 (
... 不同生态系统土壤N:P与土壤MBN:MBP的相关性存在明显差异, 以往研究显示黄土丘陵地区土壤和微生物N:P显著正相关(
黄土丘陵区植被与地形特征对土壤和土壤微生物生物量生态化学计量特征的影响
2
2018
... 土壤微生物作为草地土壤中最活跃的营养库, 其生态化学计量对于探究草地生态系统限制性养分具有重要意义.本研究中土壤MBC:MBN平均值高于中国土壤MBC:MBN平均值7.6, 而土壤MBN:MBP平均值低于中国土壤MBN:MBP的平均值5.6 (
... 不同生态系统土壤N:P与土壤MBN:MBP的相关性存在明显差异, 以往研究显示黄土丘陵地区土壤和微生物N:P显著正相关(
生态系统碳氮磷元素的生态化学计量学特征
1
2008
... 生态化学计量学以环境中相互作用的生物体以及生物体与环境之间的物质平衡来认识生态动力学, 以元素组成来揭示生物化学和生态学之间的联系, 从而分析和预测生态系统的结构和动力学(
生态系统碳氮磷元素的生态化学计量学特征
1
2008
... 生态化学计量学以环境中相互作用的生物体以及生物体与环境之间的物质平衡来认识生态动力学, 以元素组成来揭示生物化学和生态学之间的联系, 从而分析和预测生态系统的结构和动力学(
Ecoenzymatic stoichiometry of microbial nutrient acquisition in tropical soils
2
2014
... 土壤生态酶的相对活性与资源的有效性相耦合.土壤磷酸酶活性通常与土壤有效P含量成反比(
... 土壤胞外酶(LAP+NAG)或NAG均可反映土壤微生物N养分资源的需求状况, 且BG:(NAG + LAP):AP或BG:NAG:AP均可表示土壤胞外酶生态化学计量关系(
黄土丘陵区不同植被类型下土壤与微生物C, N, P化学计量特征研究
1
2016
... 不同生态系统土壤N:P与土壤MBN:MBP的相关性存在明显差异, 以往研究显示黄土丘陵地区土壤和微生物N:P显著正相关(
黄土丘陵区不同植被类型下土壤与微生物C, N, P化学计量特征研究
1
2016
... 不同生态系统土壤N:P与土壤MBN:MBP的相关性存在明显差异, 以往研究显示黄土丘陵地区土壤和微生物N:P显著正相关(
A global analysis of soil microbial biomass carbon, nitrogen and phosphorus in terrestrial ecosystems
3
2013
... 土壤微生物作为草地土壤中最活跃的营养库, 其生态化学计量对于探究草地生态系统限制性养分具有重要意义.本研究中土壤MBC:MBN平均值高于中国土壤MBC:MBN平均值7.6, 而土壤MBN:MBP平均值低于中国土壤MBN:MBP的平均值5.6 (
... ), 其主要原因可能是荒漠草地土壤微生物N含量较低; 土壤MBC:MBP与全球土壤MBC:MBP的平均值42.4接近(
... 不同生态系统土壤N:P与土壤MBN:MBP的相关性存在明显差异, 以往研究显示黄土丘陵地区土壤和微生物N:P显著正相关(
长期免耕与施用有机肥对土壤微生物生物量碳、氮、磷的影响
1
2002
... 土壤是植物生长发育的重要基质, 其养分含量对于评价土壤质量状况至关重要(
长期免耕与施用有机肥对土壤微生物生物量碳、氮、磷的影响
1
2002
... 土壤是植物生长发育的重要基质, 其养分含量对于评价土壤质量状况至关重要(
土壤C库构成研究进展
1
2014
... 土壤是植物生长发育的重要基质, 其养分含量对于评价土壤质量状况至关重要(
土壤C库构成研究进展
1
2014
... 土壤是植物生长发育的重要基质, 其养分含量对于评价土壤质量状况至关重要(
Estimation of desertification risk from soil erosion: A case study for Gansu Province, China
1
2016
... 土壤是植物生长发育的重要基质, 其养分含量对于评价土壤质量状况至关重要(
盐池县草地沙漠化过程中土壤微生物的变化
1
2013
... 土壤是植物生长发育的重要基质, 其养分含量对于评价土壤质量状况至关重要(
盐池县草地沙漠化过程中土壤微生物的变化
1
2013
... 土壤是植物生长发育的重要基质, 其养分含量对于评价土壤质量状况至关重要(
Reviews and syntheses: Soil resources and climate jointly drive variations in microbial biomass carbon and nitrogen in China’s forest ecosystems
2
2015
... 土壤C:N:P化学计量可以反映土壤C、N、P循环, 有助于确定生态过程对全球变化的响应, 因而成为土壤C、N、P平衡特征的重要参数(
... 土壤C:P与土壤MBC:MBP无显著相关关系, 与三峡库区不同植被土壤C:P与土壤MBC:MBP的结果一致(
生态系统演替过程中土壤与微生物碳氮磷化学计量关系的变化
1
2016
... 土壤C:N:P化学计量可以反映土壤C、N、P循环, 有助于确定生态过程对全球变化的响应, 因而成为土壤C、N、P平衡特征的重要参数(
生态系统演替过程中土壤与微生物碳氮磷化学计量关系的变化
1
2016
... 土壤C:N:P化学计量可以反映土壤C、N、P循环, 有助于确定生态过程对全球变化的响应, 因而成为土壤C、N、P平衡特征的重要参数(
Vegetation pattern variation, soil degradation and their relationship along a grassland desertification gradient in Horqin Sandy Land, northern China
1
2009
... 近年来, 由于人类对草地资源的过度利用和全球气候变化, 造成草地生态系统不同程度的退化与沙漠化.草地沙漠化是草地退化的极端表现形式, 草地沙漠化过程中土壤C和养分衰减(

{kind=link}
{kind=link}
{kind=link}
{kind=link}