Physiological and ecological responses to drought and heat stresses in Osmanthus fragrans ‘Boyejingui’
WANG Jun-Yu, WANG Xiao-Dong, MA Yuan-Dan, FU Lu-Cheng, ZHOU Huan-Huan, WANG Bin, ZHANG Ru-Min, GAO Yan,*The State Key Laboratory of Subtropical Silviculture, Zhejiang Agriculture and Forestry University, Lin’an, Zhejiang 311300, China
Abstract Aims Drought and heat stresses are two critical environmental factors affecting the growth and development of plants; climate change has exacerbated the occurrence of these conditions in many parts of the world. To elucidate the mechanisms of responses to drought and heat stresses in Osmanthus fragrans, we studied changes in nonstructural carbohydrate (NSC) and volatile organic compounds (VOCs) emissions using 3-year-old seedlings (cultivar ‘Boyejingui’) under conditions of low drought (LD), moderate drought (MD) and severe drought (SD), interactively with heat treatments (40 °C in daytime (12 h) and 30 °C at night (12 h)).Methods The VOCs emissions were measured using the dynamic headspace air-circulation method, and the composition and content of VOCs were analyzed using the thermal desorption system/gas chromatography/mass spectrometer technique (TDS-GC-MS). We determined the content of NSC and activities of lipoxygenase (LOX), 3-hydroxy-3-methylglutaryl-CoA reductase (HMGR) and 1-deoxy-D-xylulose 5-phosphate reductoisomerase (DXR) in seedlings under different drought and heat treatments.Important findings The content of NSC in leaves was not significantly affected by drought stress. The contents of soluble sugar and starch were significantly reduced by heat stress as well as combination of drought and heat stresses. The combination of severe drought and heat treatment significantly reduced the contents of glucose, fructose, sucrose and starch by 47.7%, 46.4%, 34.4% and 38.2% (p < 0.05), respectively. There were differences among the activities of HMGR, DXR and LOX in response to drought and heat stresses. Under drought and combined stresses, the activities of HMGR and DXR increased first and then decreased, and the LOX activity increased. The activities of HMGR, DXR and LOX were significantly higher under heat stress than in controls. The release of terpenes increased first and then decreased with the level of drought treatments. MD and SD stress stimulated the release of terpenes by 37.9% and 32.3% (p < 0.05), respectively, but the combined stresses induced a reduction in the release of terpenes. The release of aldehydes increased significantly in response to drought, heat treatment, and their combination. The results indicated that ‘Boyejingui’ seedlings can enhance resistance to drought stress by adjusting contents of non-structural carbohydrate and releasing terpene compounds. They also adjusted the biosynthesis of terpene compounds and GLVs emission to resist against heat stress. However, the biosynthesis pathway of terpene compounds was blocked in the occurrence of both drought and heat stresses, when the enhanced release of GLVs facilitated the resistance. Under heat stress, moderate and severe drought caused severe damage to cell membranes, thus reducing the self-regulatory capacity. Keywords:Osmanthus fragrans;drought and heat stress;non-structural carbohydrate;volatile organic compounds;secondary metabolic enzymes
PDF (1116KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 汪俊宇, 王小东, 马元丹, 傅卢成, 周欢欢, 王彬, 张汝民, 高岩. ‘波叶金桂’对干旱和高温胁迫的生理生态响应. 植物生态学报[J], 2018, 42(6): 681-691 doi:10.17521/cjpe.2018.0017 WANG Jun-Yu. Physiological and ecological responses to drought and heat stresses in Osmanthus fragrans ‘Boyejingui’. Chinese Journal of Plant Ecology[J], 2018, 42(6): 681-691 doi:10.17521/cjpe.2018.0017
非结构性碳水化合物(NSC)作为植物体内的可溶性碳库, 不仅为植物生长发育提供碳源与能源, 还能调控植物生理活动(Hoch et al., 2003); 同时, 植物体内NSC的积累是一种重要的渗透调节保护机制(Lin et al., 2014; Murcia et al., 2015)。干旱胁迫导致欧洲云杉(Picea abies)树根NSC含量减少, 在地上和地下组织之间的碳分配、使用和运输方面有显著差异(Hartmann et al., 2013), 随干旱胁迫时间的延长, 刺槐(Robinia pseudoacacia)叶片NSC含量降低(Zhang et al., 2015)。Marias等(2017)研究发现热处理能够诱导西黄松(Pinus ponderosa)和花旗松(Pseudotsuga menziesii)淀粉含量减少, 果糖和葡萄糖含量增加, 总NSC和蔗糖含量并没有受到影响。随着高温干旱协同胁迫加剧, 新疆梨(Pyrus sinkiangensis)可溶性糖含量呈上升趋势, 淀粉含量逐渐降低(穆蓁蓁, 2015)。受到环境胁迫时, 植物初级代谢发生紊乱, 可溶性糖的积累受到严重影响。蔗糖裂解为果糖和葡萄糖, 不仅作为代谢物质和能量来源, 还充当信号分子, 通过信号转导途径识别, 协调合成代谢和分解代谢过程, 促进植物生长发育(Li & Sheen, 2016)。
植物VOCs释放受到相关酶的调控。甲羟戊酸和甘油醛磷酸/丙酮酸两条途径是合成萜烯类化合物的主要途径(Dudareva et al., 2013), 3-羟基-3-甲基戊二酸单酰辅酶A还原酶(3-hydroxy-3-methylglutaryl CoA reductase, HMGR)和1-脱氧木酮糖-5-磷酸还原酶(1-deoxy-D-xylulose 5-phosphate reductoisomerase, DXR)分别是两个途径的关键限速酶(Ramak et al., 2013), 直接调控萜烯类化合物的生物合成(Moses et al., 2013)。植物释放绿叶挥发物(GLVs)是由环境胁迫诱发的一种防御反应, 植物通过调控其中关键酶——脂氧合酶(lipoxygenase, LOX)——催化不饱和脂肪酸形成GLVs (Hassan et al., 2015)。因而HMGR、DXR和LOX活性的提高诱导VOCs的形成是植物增强抗胁迫能力的重要前提。
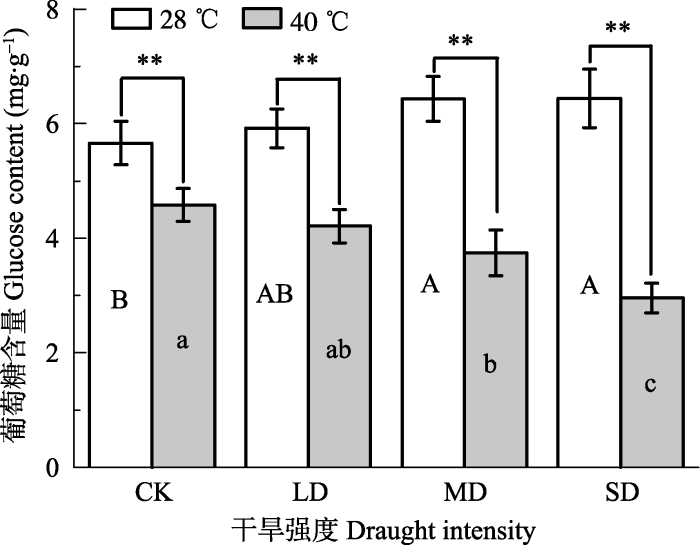
新窗口打开|下载原图ZIP|生成PPT 图1干旱高温处理对波叶‘波叶金桂’叶片葡萄糖含量的影响(平均值±标准误差, n = 5)。不同大写字母表示干旱处理的差异显著, 不同小写字母表示高温处理的差异显著(p < 0.05, 根据LSD测验)。**, p < 0.01。CK, 对照; LD, 轻度干旱; MD, 中度干旱; SD, 重度干旱。
Fig. 1Effects of drought and heat stress on glucose content in leaves of Osmanthus fragrans cv. ‘Boyejingui’ (mean ± SE, n = 5). Different uppercase letters indicate significant differences of drought and different lowercase letters denote statistically significant differences of heat stress treatment (p < 0.05, according to LSD test). p < 0.01. CK, control; LD, light drought; MD, moderate drought; SD, severe drought.
新窗口打开|下载原图ZIP|生成PPT 图2干旱高温处理对‘波叶金桂’叶片果糖含量的影响(平均值±标准误差, n = 5)。不同大写字母表示干旱的差异显著, 不同小写字母表示高温的差异显著(p < 0.05, 根据LSD测验)。**, p < 0.01。CK, 对照; LD, 轻度干旱; MD, 中度干旱; SD, 重度干旱。
Fig. 2Effects of drought and heat stress on fructose content in leaves of Osmanthus fragrans cv. ‘Boyejingui’ (mean ± SE, n = 5). Different uppercase letters indicate significant differences of drought and different lowercase letters denote statistically significant differences of heat stress treatment (p < 0.05, according to LSD test). **, p < 0.01. CK, control; LD, light processing; MD, moderate drought; SD, severe drought.
Fig. 3Effects of drought and heat stress on sucrose content in leaves of Osmanthus fragrans cv. ‘Boyejingui’ (mean ± SE, n = 5). Different uppercase letters indicate significant differences of drought and different lowercase letters denote statistically significant differences of heat stress treatment (p < 0.05, according to LSD test). p < 0.01. CK, control; LD, light drought; MD, moderate drought; SD, severe drought.
新窗口打开|下载原图ZIP|生成PPT 图4干旱高温处理对‘波叶金桂’叶片淀粉含量的影响(平均值±标准误差, n = 5)。不同大写字母表示干旱的差异显著, 不同小写字母表示高温的差异显著(p < 0.05, 根据LSD测验)。**, p < 0.01。CK, 对照; LD, 轻度干旱; MD, 中度干旱; SD, 重度干旱。
Fig. 4Effects of drought and heat stress on starch content in leaves of Osmanthus fragrans cv. ‘Boyejingui’ (mean ± SE, n = 5). Different uppercase letters indicate significant differences of drought and different lowercase letters denote statistically significant differences of heat stress treatment (p < 0.05, according to LSD test). **, p < 0.01. CK, control; LD, light drought; MD, moderate drought; SD, severe drought.
新窗口打开|下载原图ZIP|生成PPT 图5干旱高温处理对‘波叶金桂’叶片3-羟基-3-甲基戊二酸单酰辅酶A还原酶活性的影响(平均值±标准误差, n = 5)。不同大写字母表示干旱的差异显著, 不同小写字母表示高温的差异显著(p < 0.05, 根据LSD测验)。**, p < 0.01。CK, 对照; LD, 轻度干旱; MD, 中度干旱; SD, 重度干旱。
Fig. 5Effects of drought and heat stress on 3-hydroxy-3- methylglutaryl CoA reductase (HMGR) activity in leaves of Osmanthus fragrans cv. ‘Boyejingui’ (mean ± SE, n = 5). Different uppercase letters indicate significant differences of drought and different lowercase letters denote statistically significant differences of heat stress treatment (p < 0.05, according to LSD test). **, p < 0.01. CK, control; LD, light drought; MD, moderate drought; SD, severe drought.
新窗口打开|下载原图ZIP|生成PPT 图6干旱高温处理对‘波叶金桂’ 叶片1-脱氧木酮糖-5-磷酸还原酶活性的影响(平均值±标准误差, n = 5)。不同大写字母表示干旱的差异显著, 不同小写字母表示高温的差异显著(p < 0.05, 根据LSD测验)。**, p < 0.01。CK, 对照; LD, 轻度干旱; MD, 中度干旱; SD, 重度干旱。
Fig. 6Effects of drought and heat stress on 1-deoxy- D-xylulose 5-phosphate reductoisomerase (DXR) activity in leaves of Osmanthus fragrans cv. ‘Boyejingui’ (mean ± SE, n = 5). Different uppercase letters indicate significant differences of drought and different lowercase letters denote statistically significant differences of d heat stress treatment (p < 0.05, according to LSD test). **, p < 0.01;. CK, control; LD, light drought; MD, moderate drought; SD, severe drought.
新窗口打开|下载原图ZIP|生成PPT 图7干旱高温处理对‘波叶金桂’叶片脂氧合酶活性的影响(平均值±标准误差, n = 5)。不同大写字母表示干旱的差异显著, 不同小写字母表示高温的差异显著(p < 0.05, 根据LSD测验)。*, p < 0.05; **, p < 0.01;。CK, 对照; LD, 轻度干旱; MD, 中度干旱; SD, 重度干旱。
Fig. 7Effects of drought and heat stress on lipoxygenase (LOX) activity in leaves of Osmanthus fragrans cv. ‘Boyejingui’ (mean ± SE, n = 5). Different uppercase letters indicate significant differences of different lowercase letters denote statistically significant differences of from drought and heat stress treatment (p < 0.05, according to LSD test). **, p < 0.01. CK, control; LD, light drought; MD, moderate drought; SD, severe drought.
Table 1 表1 表1波叶金桂’叶片挥发性有机化合物的主要成分(峰面积, A × 105·10 g-1)(平均值±标准误差) Table 1The main components of the volatile organic compounds (VOCs) from the leaf of Osmanthus fragrans ‘Boyejingui’ (peak area, A × 105·10 g-1) (mean ± SE)
AgrawalSB, SinghS, AgrawalM ( 2009). Ultraviolet-B induced changes in gene expression and antioxidants in plants Advances in Botanical Research, 52, 47-86. DOI:10.1016/S0065-2296(10)52003-2URL [本文引用: 1]
ArabL, KreuzwieserJ, KruseJ, ZimmerI, AcheP, AlfarrajS, Al-RasheidKAS, SchnitzlerJP, HedrichR, RennenbergH ( 2016). Acclimation to heat and drought—Lessons to learn from the date palm ( Phoenix dactylifera). Environmental & Experimental Botany, 125, 20-30. [本文引用: 2]
BourtsoukidisE, KawaletzH, RadackiD, SchützS, HakolaH, HellénH, NoeS, M?lderI, AmmerC, BonnB ( 2014). Impact of flooding and drought conditions on the emission of volatile organic compounds of Quercus robur and Prunus serotina. Trees, 28, 193-204. [本文引用: 1]
BradfordMM ( 1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding Analytical Biochemistry, 72, 248-254. DOI:10.1016/0003-2697(76)90527-3URL [本文引用: 1]
ChenKC ( 2013). The warming trend and the distribution of the seasons in Hangzhou from 1961 to 2012 Chinese Agricultural Science Bulletin, 29, 345-350. URL [本文引用: 1] In order to clarify the seasonal variation characteristics of Hangzhou under the background of the climate warming and the influence from the temperature variation to the seasonal variation, using the daily temperature data of the Hangzhou station from 1961 to 2012, according to the newest delineation standard of climatic season, the length and beginning date of four seasons were divided. The variation features of the temperature and the length and beginning date of four seasons in recent 52 years and the correlations among them were investigated with the modern statistical diagnostic methods, such as climatic trend rate, correlation analysis, and abrupt detection. The results showed that: summer was the longest season in Hangzhou, while autumn was the shortest season. Meanwhile the summer was becoming longer significantly at a rate of 5.33 d/10 a, and the winter was getting shorter obviously at a rate of -3.62 d/10 a. The lengthening of the summer was influenced by the advancing of the beginning date of the summer and the delaying of the beginning date of the autumn. The shortening of the winter was influenced by the advancing of the beginning date of the spring and delaying of the beginning date of the winter, furthermore the influence of former was more than that of the latter. The summer length becoming longer, the spring and summer beginning date advancing, and the autumn beginning date delaying occurred at the same abrupt change time of the early of the 1990s. The air temperature of Hangzhou had increased clearly with the climatic rate 0.35 /10 a in recent 52 years. The yearly mean air temperature had relationship with the length and beginning date of the four seasons. It mainly showed that when the air temperature increased, the beginning date of the spring and summer advanced and the two seasons lengthened, and the beginning date of the autumn and winter delayed and the two seasons shortened. [ 陈柯辰 ( 2013). 1961-2012年杭州的升温趋势和四季分配之变化 中国农学通报, 29, 345-350.] URL [本文引用: 1] In order to clarify the seasonal variation characteristics of Hangzhou under the background of the climate warming and the influence from the temperature variation to the seasonal variation, using the daily temperature data of the Hangzhou station from 1961 to 2012, according to the newest delineation standard of climatic season, the length and beginning date of four seasons were divided. The variation features of the temperature and the length and beginning date of four seasons in recent 52 years and the correlations among them were investigated with the modern statistical diagnostic methods, such as climatic trend rate, correlation analysis, and abrupt detection. The results showed that: summer was the longest season in Hangzhou, while autumn was the shortest season. Meanwhile the summer was becoming longer significantly at a rate of 5.33 d/10 a, and the winter was getting shorter obviously at a rate of -3.62 d/10 a. The lengthening of the summer was influenced by the advancing of the beginning date of the summer and the delaying of the beginning date of the autumn. The shortening of the winter was influenced by the advancing of the beginning date of the spring and delaying of the beginning date of the winter, furthermore the influence of former was more than that of the latter. The summer length becoming longer, the spring and summer beginning date advancing, and the autumn beginning date delaying occurred at the same abrupt change time of the early of the 1990s. The air temperature of Hangzhou had increased clearly with the climatic rate 0.35 /10 a in recent 52 years. The yearly mean air temperature had relationship with the length and beginning date of the four seasons. It mainly showed that when the air temperature increased, the beginning date of the spring and summer advanced and the two seasons lengthened, and the beginning date of the autumn and winter delayed and the two seasons shortened.
ChengLL, ChenM, LengPS, HuZH ( 2015). Effect of different temperature levels on the emission of terpenoid volatile compounds in Ocimum basilicum ‘Purple Ruffles’. Journal of Beijing University of Agriculture, 30(2), 78-82. DOI:10.13473/cnki.issn.1002-3186.2015.0023URL [本文引用: 1] 以盆栽紫罗勒Ocimum basilicum‘Purple Ruffles’为试验材料,经3个不同温度(20,30,40℃)处理后,使用动态顶空法采集香气,然后采用自动热脱附-气相色谱/质量色谱联用技术(ATD-GC/MS)对萜烯类香气成分和释放量进行鉴定和分析。结果表明,紫罗勒香气中的萜烯类化合物数量随温度升高表现出降低的趋势,并且相对含量也表现出明显变化,说明温度能显著影响紫罗勒萜烯化合物的释放,但各个萜烯类化合物的变化规律不同。在检测到的萜烯化合物中桉叶油醇含量最高,3个温度下的平均含量达到了81%,远远高于其他成分,可初步判断桉叶油醇为紫罗勒挥发物中萜烯化合物的主要成分。 [ 程璐璐, 陈敏, 冷平生, 胡增辉 ( 2015). 不同温度对紫罗勒萜烯类挥发物释放的影响 北京农学院学报, 30(2), 78-82.] DOI:10.13473/cnki.issn.1002-3186.2015.0023URL [本文引用: 1] 以盆栽紫罗勒Ocimum basilicum‘Purple Ruffles’为试验材料,经3个不同温度(20,30,40℃)处理后,使用动态顶空法采集香气,然后采用自动热脱附-气相色谱/质量色谱联用技术(ATD-GC/MS)对萜烯类香气成分和释放量进行鉴定和分析。结果表明,紫罗勒香气中的萜烯类化合物数量随温度升高表现出降低的趋势,并且相对含量也表现出明显变化,说明温度能显著影响紫罗勒萜烯化合物的释放,但各个萜烯类化合物的变化规律不同。在检测到的萜烯化合物中桉叶油醇含量最高,3个温度下的平均含量达到了81%,远远高于其他成分,可初步判断桉叶油醇为紫罗勒挥发物中萜烯化合物的主要成分。
ChoatB, JansenS, BrodribbTJ, CochardH, DelzonS, BhaskarR, BucciSJ, FeildTS, GleasonSM, HackeUG, JacobsenAL, LensF, MaheraliH, Martínez-VilaltaJ, MayrS, MencucciniM, MitchellPJ, NardiniA, PittermannJ, PrattRB, SperryJS, WestobyM, WrightIJ, ZanneAE ( 2012). Global convergence in the vulnerability of forests to drought Nature, 491, 752-755. DOI:10.1038/nature11688URL [本文引用: 1]
CopoloviciL, K?nnasteA, RemmelT, Niinemetsü ( 2014). Volatile organic compound emissions from Alnus glutinosa under interacting drought and herbivory stresses. Environmental & Experimental Botany, 100, 55-63. [本文引用: 1]
DickeM, LoretoF ( 2010). Induced plant volatiles: From genes to climate change Trends in Plant Science, 15, 115-117. DOI:10.1016/j.tplants.2010.01.007URLPMID:20137997 [本文引用: 1] Plants are multifaceted chemical factories that produce a multitude of compounds ranging from ethylene and methanol to complex terpenoids and nitrogen-containing alkaloids. More than 100,000 chemical products are known to be produced by plants and at least 1700 of these are known to be volatile. The study of plant volatiles has long been restricted to floral volatiles, but studies on plant volatiles emitted from the vegetative tissues are rapidly increasing. The amounts of Volatile Organic Compounds that plants disseminate into the atmosphere (VOCs or BVOCs from their ''Biogenic'' origin) are enormous. For instance, terrestrial plants are estimated to release about 500 tera-gram of carbon per year as isoprene, and probably a similar amount of carbon as monoterpenes. Other plant volatiles include e.g. green leaf volatiles, nitrogen-containing compounds and aromatic compounds. The emission of plant VOCs is not restricted to aboveground tissues but also occurs belowground from their roots.
DudarevaN, KlempienA, MuhlemannJK, KaplanI ( 2013). Biosynthesis, function and metabolic engineering of plant volatile organic compounds New Phytologist, 198, 16-32. DOI:10.1111/nph.12145URL [本文引用: 1]
GaoY, JinYJ, LiHD, ChenHJ ( 2005). Volatile organic compounds and their roles in bacteriostasis in five conifer species Journal of Integrative Plant Biology, 47, 499-507. DOI:10.1111/jipb.2005.47.issue-4URL [本文引用: 1]
HartikainenK, NergAM, Kivim?enp??M, Kontunen-SoppelaS, M?enp??M, OksanenE, RousiM, HolopainenT ( 2009). Emissions of volatile organic compounds and leaf structural characteristics of European aspen ( Populus tremula) grown under elevated ozone and temperature. Tree Physiology, 29, 1163-1173. [本文引用: 1]
HartmannH, ZieglerW, TrumboreS ( 2013). Lethal drought leads to reduction in nonstructural carbohydrates in Norway spruce tree roots but not in the canopy Functional Ecology, 27, 413-427. DOI:10.1111/fec.2013.27.issue-2URL [本文引用: 1]
HassanMN, ZainalZ, IsmailI ( 2015). Green leaf volatiles: biosynthesis, Biological functions and their applications in biotechnology Plant Biotechnology Journal, 13, 727-739. DOI:10.1111/pbi.12368URLPMID:25865366 [本文引用: 1] Summary Plants have evolved numerous constitutive and inducible defence mechanisms to cope with biotic and abiotic stresses. These stresses induce the expression of various genes to activate defence-related pathways that result in the release of defence chemicals. One of these defence mechanisms is the oxylipin pathway, which produces jasmonates, divinylethers and green leaf volatiles (GLVs) through the peroxidation of polyunsaturated fatty acids (PUFAs). GLVs have recently emerged as key players in plant defence, plant–plant interactions and plant–insect interactions. Some GLVs inhibit the growth and propagation of plant pathogens, including bacteria, viruses and fungi. In certain cases, GLVs released from plants under herbivore attack can serve as aerial messengers to neighbouring plants and to attract parasitic or parasitoid enemies of the herbivores. The plants that perceive these volatile signals are primed and can then adapt in preparation for the upcoming challenges. Due to their ‘green note’ odour, GLVs impart aromas and flavours to many natural foods, such as vegetables and fruits, and therefore, they can be exploited in industrial biotechnology. The aim of this study was to review the progress and recent developments in research on the oxylipin pathway, with a specific focus on the biosynthesis and biological functions of GLVs and their applications in industrial biotechnology.
HasunumaT, TakenoS, HayashiS, SendaiM, BambaT, YoshimuraS, TomizawaK, FukusakiE, MiyakeC ( 2008). Overexpression of 1-Deoxy-D-xylulose-5-phosphate reductoisomerase gene in chloroplast contributes to increment of isoprenoid production Journal of Bioscience and Bioengineering, 105, 518-526. DOI:10.1263/jbb.105.518URL [本文引用: 1]
HatanakaA ( 1993). The biogeneration of green odour by green leaves Phytochemistry, 34, 1201-1218. DOI:10.1016/0031-9422(91)80003-JURL [本文引用: 2] The ‘green odour’ of green leaves arises from eight volatile compounds, C 6 -aldehydes and C 6 -alcohols which include leaf aldehyde, (2 E )-
HijiokaY, LinE, PereiraJ, CorlettR, CuiX, InsarovG, LascoR, LindgrenE, SurjanA ( 2014). Asia. In: Christopher B, Vicente R eds. Climate Change 2014: Impacts, Adaptation, and Vulnerability, Part B: Regional Aspects, Contribution of Working Group II, Fifth Assessment Report of the Intergovernmental Panel on Climate Change Cambridge University Press, Cambridge,UK . 1327-1370. [本文引用: 1]
HochG, RichterA, K?rnerC ( 2003). Non-structural carbon compounds in temperate forest trees Plant, Cell & Environment, 26, 1067-1081. DOI:10.1046/j.0016-8025.2003.01032.xURL [本文引用: 1] The current carbon supply status of temperate forest trees was assessed by analysing the seasonal variation of non-structural carbohydrate (NSC) concentrations in leaves, branch wood and stem sapwood of 10 tree species (six deciduous broad-leafed, one deciduous conifer and three evergreen conifer trees) in a temperate forest that is approximately 100 years old. In addition, all woody tissue was analysed for lipids (acylglycerols). The major NSC fractions were starch, sucrose, glucose and fructose, with other carbohydrates (e.g. raffinose and stachyose) and sugar alcohols (cyclitols and sorbitol) playing only a minor quantitative role. The radial distribution of NSC within entire stem cores, assessed here for the first time in a direct interspecific comparison, revealed large differences in the size of the active sapwood fraction among the species, reflecting the specific wood anatomy (ring-porous versus diffuse-porous xylem). The mean minimum NSC concentrations in branch wood during the growing season was 55% of maximum, and even high NSC concentrations were maintained during times of extensive fruit production in masting Fagus sylvestris . The NSC in stem sapwood varied very little throughout the season (cross species mean never below 67% of maximum), and the small reductions observed were not significant for any of the investigated species. Although some species contained substantial quantities of lipids in woody tissues ('fat trees'; Tilia , Pinus , Picea , Larix ), the lipid pools did not vary significantly across the growing season in any species. On average, the carbon stores of deciduous trees would permit to replace the whole leave canopy four times. These data imply that there is not a lot of leeway for a further stimulation of growth by ongoing atmospheric CO 2 enrichment. The classical view that deciduous trees rely more on C-reserves than evergreen trees, seems unwarranted or has lost its justification due to the greater than 30% increase in atmospheric CO 2 concentrations over the last 150 years.
Joóé, DewulfJ, AmelynckC, SchoonN, PokorskaO, ?impragaM, SteppeK, AubinetM, van LangenhoveH ( 2011). Constitutive versus heat and biotic stress induced BVOC emissions in Pseudotsuga menziesii. Atmospheric Environment, 45, 3655-3662. [本文引用: 1]
JudW, VanzoE, LiZ, GhirardoA, ZimmerI, SharkeyTD, Hansel1A, SchnitzlerJP ( 2016). Effects of heat and drought stress on post-illumination bursts of volatile organic compounds in isoprene-emitting and non-emitting poplar Plant, Cell & Environment, 39, 1204-1215. [本文引用: 1]
KleinT, HochG, DanY, K?rnerC ( 2014). Drought stress, growth and nonstructural carbohydrate dynamics of pine trees in a semi-arid forest Tree Physiology, 34, 981-992. DOI:10.1093/treephys/tpu071URL [本文引用: 1]
LiL, SheenJ ( 2016). Dynamic and diverse sugar signaling Current Opinion in Plant Biology, 33, 116-125. DOI:10.1016/j.pbi.2016.06.018URLPMID:5050104 [本文引用: 1] Sugars fuel life and exert numerous regulatory actions that are fundamental to all life forms. There are two principal mechanisms underlie sugar ‘perception and signal transduction’ in biological systems. Direct sensing and signaling is triggered via sugar-binding sensors with a broad range of affinity and specificity, whereas sugar-derived bioenergetic molecules and metabolites modulate signaling proteins and indirectly relay sugar signals. This review discusses the emerging sugar signals and potential sugar sensors discovered in plant systems. The findings leading to informative understanding of physiological regulation by sugars are considered and assessed. Comparative transcriptome analyses highlight the primary and dynamic sugar responses and reveal the convergent and specific regulators of key biological processes in the sugar-signaling network.
LiP, SunYF, WangSG, WangXD, CaiX, ZhuWZ, PaoloC, LiMH ( 2008). Altitudinal changes in leaf mass per unit area and tissue non-structural carbohydrates content of Abies fabri on Gongga Mountain of southwest China. Chinese Journal of Applied Ecology, 19, 8-12. URL [本文引用: 1] 以贡嘎山地区磨西山谷东北坡和康定山谷西南坡为样地,研究了高山林线和低海拔冷杉休眠期和生长旺盛期针叶比叶质量和组织非结构性碳水化合物含量.结果表明:生长在温暖湿润的磨西山谷东北坡的冷杉针叶比叶质量和组织非结构性碳水化合物含量均高于炎热干燥的康定山谷西南坡;高山林线冷杉比叶质量大于低海拔冷杉;高山林线冷杉组织非结构性碳水化合物含量总体高于低海拔冷杉,且生长期比休眠期更为显著.研究结果不支持“碳供应不足导致高山林线形成”的假说. [ 李蟠, 孙玉芳, 王三根, 王小丹, 蔡小虎, 朱万泽, CherybiniPaolo, 李迈和 ( 2008). 贡嘎山地区不同海拔冷杉比叶质量和非结构性碳水化合物含量变化 应用生态学报, 19, 8-12.] URL [本文引用: 1] 以贡嘎山地区磨西山谷东北坡和康定山谷西南坡为样地,研究了高山林线和低海拔冷杉休眠期和生长旺盛期针叶比叶质量和组织非结构性碳水化合物含量.结果表明:生长在温暖湿润的磨西山谷东北坡的冷杉针叶比叶质量和组织非结构性碳水化合物含量均高于炎热干燥的康定山谷西南坡;高山林线冷杉比叶质量大于低海拔冷杉;高山林线冷杉组织非结构性碳水化合物含量总体高于低海拔冷杉,且生长期比休眠期更为显著.研究结果不支持“碳供应不足导致高山林线形成”的假说.
LinSK, LinJ, LiuQL, AiYF, KeYQ, ChenC, ZhangZY, HeH ( 2014). Time-course of photosynthesis and non-?structural carbon compounds in the leaves of tea plants ( Camellia sinensis, L.) in response to deficit irrigation. Agricultural Water Management, 144, 98-106. [本文引用: 1]
LiuF, ZuoZJ, XuGP, WuXB, ZhengJ, GaoRF, ZhangRM, GaoY ( 2013). Physiological responses to drought stress and the emission of induced volatile organic compounds in Rosmarinus officinalis. Chinese Journal of Plant Ecology, 37, 454-463. [本文引用: 2]
LoretoF, SchnitzlerJP ( 2010). Abiotic stresses and induced BVOCs Trends in Plant Science, 15, 154-166. DOI:10.1016/j.tplants.2009.12.006URLPMID:20133178 [本文引用: 3] Plants produce a wide spectrum of biogenic volatile organic compounds (BVOCs) in various tissues above and below ground to communicate with other plants and organisms. However, BVOCs also have various functions in biotic and abiotic stresses. For example abiotic stresses enhance BVOCs emission rates and patterns, altering the communication with other organisms and the photochemical cycles. Recent new insights on biosynthesis and eco-physiological control of constitutive or induced BVOCs have led to formulation of hypotheses on their functions which are presented in this review. Specifically, oxidative and thermal stresses are relieved in the presence of volatile terpenes. Terpenes, C6 compounds, and methyl salicylate are thought to promote direct and indirect defence by modulating the signalling that biochemically activate defence pathways.
MaC, FengYL, ZhangJ, WangHZ, YuanJL, LiYJ ( 2017). Effects of exogenous methyl jasmonate on endogenous hormones and yield formation in wheat after anthesis under drought stress Plant Physiology Journal, 53, 1051-1058. [本文引用: 1]
MaguireAJ, KobeRK ( 2015). Drought and shade deplete non-structural carbohydrate reserves in seedlings of five temperate tree species Ecology and Evolution, 5, 5711-5721. DOI:10.1002/ece3.1819URL [本文引用: 1]
MariasDE, MeinzerFC, WoodruffDR, McCullohKA ( 2017). Thermotolerance and heat stress responses of Douglas-fir and ponderosa pine seedling populations from contrasting climates Tree Physiology, 37, 301-315. [本文引用: 1]
McDowellN, PockmanWT, AllenCD, BreshearsDD, CobbN, KolbT, PlautJ, SperryJ, WestA, WilliamsDG, YepezEA ( 2008). Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytologist, 178, 719-739. DOI:10.1111/j.1469-8137.2008.02436.xURL [本文引用: 1]
MorshedlooMR, CrakerLE, SalamiA, NazeriV, SangH, MaggiF ( 2017). Effect of prolonged water stress on essential oil content, compositions and gene expression patterns of mono- and sesquiterpene synthesis in two oregano (Origanum vulgare L.) subspecies. Plant Physiology & Biochemistry, 111, 119-128. [本文引用: 1]
MosesT, PollierJ, TheveleinJM, GoossensA ( 2013). Bioengineering of plant (tri) terpenoids: From metabolic engineering of plants to synthetic biology in vivo and in vitro. New Phytologist, 200, 27-43. [本文引用: 1]
MuZZ ( 2015). Research on the Impacts of High Temperature and Drought on Korla Fragrant Pear Photosynthetic Characteristics. Master degree dissertation, Xinjiang Agricultural University ürümqi. DOI:10.11937/bfyy.201506005URL [本文引用: 3] 本文以库尔勒香梨为材料,在36℃高温条件下,采用高温干旱、灌水、喷水3种处理方式,研究高温干旱对库尔勒香梨光合特性的影响。研究结果如下: 1、高温干旱条件下库尔勒香梨的净光合速率的变化表现为“双峰型”,还有“午休”现象的出现,而且各时间段的净光合速率均小于灌水处理。 2、高温干旱条件下库尔勒香梨的蒸腾速率与气孔导度变化趋势相同,呈凹型,且气孔导度在各时间段均小于灌水处理。 3、高温干旱条件下库尔勒香梨细胞间二氧化碳浓度可能受净光合速率的影响,呈现出中午降低的趋势;灌水处理下库尔勒香梨的细胞间二氧化碳浓度变化幅度较... [ 穆蓁蓁 ( 2015). 高温干旱对库尔勒香梨光合特性的影响研究 新疆农业大学, 乌鲁木齐.] DOI:10.11937/bfyy.201506005URL [本文引用: 3] 本文以库尔勒香梨为材料,在36℃高温条件下,采用高温干旱、灌水、喷水3种处理方式,研究高温干旱对库尔勒香梨光合特性的影响。研究结果如下: 1、高温干旱条件下库尔勒香梨的净光合速率的变化表现为“双峰型”,还有“午休”现象的出现,而且各时间段的净光合速率均小于灌水处理。 2、高温干旱条件下库尔勒香梨的蒸腾速率与气孔导度变化趋势相同,呈凹型,且气孔导度在各时间段均小于灌水处理。 3、高温干旱条件下库尔勒香梨细胞间二氧化碳浓度可能受净光合速率的影响,呈现出中午降低的趋势;灌水处理下库尔勒香梨的细胞间二氧化碳浓度变化幅度较...
MurciaG, PontinM, ReinosoH, BaraldiR, BertazzaG, Gómez-TalquencaS, BottiniR, PiccoliPN ( 2015). ABA and GA3 increase carbon allocation in different organs of grapevine plants by inducing accumulation of non-?structural carbohydrates in leaves, enhancement of phloem area and expression of sugar transporters Physiologia Plantarum, 156, 323-337. [本文引用: 1]
NaharK, HasanuzzamanM, AlamMM, FujitaM ( 2015). Exogenous glutathione confers high temperature stress tolerance in mung bean ( Vigna radiate L.) by modulating antioxidant defense and methylglyoxal detoxification system. Environmental & Experimental Botany, 112, 44-54. [本文引用: 1]
PanQM, HanXG, BaiYF, YangJC ( 2002). Advances in physiology and ecology studies on stored non-structure carbohydrates in plants Chinese Bulletin of Botany, 19, 30-38. DOI:10.3969/j.issn.1674-3466.2002.01.004URL [本文引用: 1] 非结构性碳水化合物是参与植物生命过程的重要物质。蔗糖不仅是植物体内碳水化合物运输的主要形式 ,而且可以在基因表达水平上对细胞内的代谢进行调节。果聚糖是植物营养组织碳水化合物的主要暂贮形式 ;淀粉是植物主要的长期贮存物质之一。植物体内非结构性碳水化合物的代谢在很大程度上影响着植株的生长发育和对环境因子的响应。综述了植物非结构性贮藏碳水化合物的生理生态学研究进展 ,着重介绍了蔗糖、果聚糖和淀粉代谢的生理过程及对环境因子 (温度和水分 )和人为因素的响应机制。 [ 潘庆民, 韩兴国, 白永飞, 杨景成 ( 2002). 植物非结构性贮藏碳水化合物的生理生态学研究进展 植物学报, 19, 30-38.] DOI:10.3969/j.issn.1674-3466.2002.01.004URL [本文引用: 1] 非结构性碳水化合物是参与植物生命过程的重要物质。蔗糖不仅是植物体内碳水化合物运输的主要形式 ,而且可以在基因表达水平上对细胞内的代谢进行调节。果聚糖是植物营养组织碳水化合物的主要暂贮形式 ;淀粉是植物主要的长期贮存物质之一。植物体内非结构性碳水化合物的代谢在很大程度上影响着植株的生长发育和对环境因子的响应。综述了植物非结构性贮藏碳水化合物的生理生态学研究进展 ,着重介绍了蔗糖、果聚糖和淀粉代谢的生理过程及对环境因子 (温度和水分 )和人为因素的响应机制。
RamakP, OsalooSK, EbrahimzadehH, SharifiM, BehmaneshM ( 2013). Inhibition of the mevalonate pathway enhances carvacrol biosynthesis and DXR gene expression in shoot cultures of Satureja khuzistanica Jamzad. Journal of Plant Physiology, 170, 1187-1193. [本文引用: 1]
SantinoA, BonsegnaS, DomenicoSD, PoltronieriP ( 2010). Plant oxylipins and their contribution to plant defense Current Topics in Plant Biology, 11, 103-111. URL [本文引用: 1] Plants have evolved complex signalling pathways to cope with different biotic stresses. Complex interactions among these pathways permit a tight control between development and stress response. Oxylipin metabolism represents one of the main defence mechanisms employed by plants. It begins with the oxygenation of polyunsaturated fatty acids by lipoxygenases (LOX), to form fatty acid hydroperoxid...
SilvaEN, FerreirasilvaSL, FonteneleADV, RibeiroRV, ViégasRA, SilveiraJAG ( 2010). Photosynthetic changes and protective mechanisms against oxidative damage subjected to isolated and combined drought and heat stresses in Jatropha curcas plants. Journal of Plant Physiology, 167, 1157-1164. [本文引用: 2]
StaudtM, MorinX, ChuineI ( 2017). Contrasting direct and indirect effects of warming and drought on isoprenoid emissions from Mediterranean oaks Regional Environmental Change, 17, 2121-2133. DOI:10.1007/s10113-016-1056-6URL [本文引用: 1]
ToroserD, HuberSC ( 1998). 3-Hydroxy-3-methylglutaryl-coenzyme A reductase kinase and sucrose-phosphate synthase kinase activities in cauliflower florets: Ca 2+ dependence and substrate specificities Archives Biochemistry & Biophysics, 355, 291-300. [本文引用: 1]
TrifilòP, CasoloV, RaimondoF, PetrussaE, BoscuttiF, GulloMAL, NardiniA ( 2017). Effects of prolonged drought on stem non-structural carbohydrates content and post-drought hydraulic recovery in Laurus nobilis L.: The possible link between carbon starvation and hydraulic failure. Plant Physiology and Biochemistry, 120, 232-241. [本文引用: 2]
VelikovaV, VarkonyiZ, SzaboM, MaslenkovaL, NoguesI, KovacsL, PeevaV, BushevaM, GarabG, SharkeyTD, LoretoF ( 2011). Increased thermostability of thylakoid membranes in isoprene-emitting leaves probed with three biophysical techniques Plant Physiology, 157, 905-916. DOI:10.1104/pp.111.182519URL [本文引用: 1]
VickersCE, PossellM, CojocariuCI, VelikovaVB, LaothawornkitkulJ, RyanA, MullineauxPM, Nicholas HewittC ( 2009). Isoprene synthesis protects transgenic tobacco plants from oxidative stress Plant, Cell & Environment, 32, 520-531. DOI:10.1111/j.1365-3040.2009.01946.xURLPMID:19183288 [本文引用: 1] Isoprene emission represents a significant loss of carbon to those plant species that synthesize this highly volatile and reactive compound. As a tool for studying the role of isoprene in plant physiology and biochemistry, we developed transgenic tobacco plants capable of emitting isoprene in a similar manner to and at rates comparable to a naturally emitting species. Thermotolerance of photosynthesis against transient high-temperature episodes could only be observed in lines emitting high levels of isoprene; the effect was very mild and could only be identified over repetitive stress events. However, isoprene-emitting plants were highly resistant to ozone-induced oxidative damage compared with their non-emitting azygous controls. In ozone-treated plants, accumulation of toxic reactive oxygen species (ROS) was inhibited, and antioxidant levels were higher. Isoprene-emitting plants showed remarkably decreased foliar damage and higher rates of photosynthesis compared to non-emitting plants immediately following oxidative stress events. An inhibition of hydrogen peroxide accumulation in isoprene-emitting plants may stall the programmed cell death response which would otherwise lead to foliar necrosis. These results demonstrate that endogenously produced isoprene provides protection from oxidative damage.
VitaleM, SalvatoriE, LoretoF, FaresS, ManesF ( 2008). Physiological responses of Quercus ilex leaves to water stress and acute ozone exposure under controlled conditions. Water, Air & Soil Pollution, 189, 113-125. [本文引用: 1]
ZhangT, CaoY, ChenY, LiuG ( 2015). Non-structural carbohydrate dynamics in Robinia pseudoacacia, saplings under three levels of continuous drought stress. Trees, 29, 1837-1849. [本文引用: 1]
ZhaoC, WangHY, LiuMZ ( 2017). Metabolic regulation of soluble sugar, starch and related enzymes in cassava stems under drought stress Plant Physiology Journal, 53, 795-806. [本文引用: 1]
ZhouS, LinFP, WangYK, ShenYB, ZhangRM, GaoRF, GaoY ( 2012). Effects of mechanical damage of leaves on volatile organic compounds and chlorophyll fluorescence parameters in seedlings of Cinnamomum camphora. Chinese Journal of Plant Ecology, 36, 671-680. [本文引用: 1]
Asia. In: Christopher B, Vicente R eds. Climate Change 2014: Impacts, Adaptation, and Vulnerability, Part B: Regional Aspects, Contribution of Working Group II, Fifth Assessment Report of the Intergovernmental Panel on Climate Change 1 2014
Effect of prolonged water stress on essential oil content, compositions and gene expression patterns of mono- and sesquiterpene synthesis in two oregano (Origanum vulgare L.) subspecies. 1 2017
ABA and GA3 increase carbon allocation in different organs of grapevine plants by inducing accumulation of non-?structural carbohydrates in leaves, enhancement of phloem area and expression of sugar transporters 1 2015
... 非结构性碳水化合物(NSC)作为植物体内的可溶性碳库, 不仅为植物生长发育提供碳源与能源, 还能调控植物生理活动(Hoch et al., 2003); 同时, 植物体内NSC的积累是一种重要的渗透调节保护机制(Lin et al., 2014; Murcia et al., 2015).干旱胁迫导致欧洲云杉(Picea abies)树根NSC含量减少, 在地上和地下组织之间的碳分配、使用和运输方面有显著差异(Hartmann et al., 2013), 随干旱胁迫时间的延长, 刺槐(Robinia pseudoacacia)叶片NSC含量降低(Zhang et al., 2015).Marias等(2017)研究发现热处理能够诱导西黄松(Pinus ponderosa)和花旗松(Pseudotsuga menziesii)淀粉含量减少, 果糖和葡萄糖含量增加, 总NSC和蔗糖含量并没有受到影响.随着高温干旱协同胁迫加剧, 新疆梨(Pyrus sinkiangensis)可溶性糖含量呈上升趋势, 淀粉含量逐渐降低(穆蓁蓁, 2015).受到环境胁迫时, 植物初级代谢发生紊乱, 可溶性糖的积累受到严重影响.蔗糖裂解为果糖和葡萄糖, 不仅作为代谢物质和能量来源, 还充当信号分子, 通过信号转导途径识别, 协调合成代谢和分解代谢过程, 促进植物生长发育(Li & Sheen, 2016). ...
Exogenous glutathione confers high temperature stress tolerance in mung bean ( 1 2015
Contrasting direct and indirect effects of warming and drought on isoprenoid emissions from Mediterranean oaks 1 2017
... 植物释放的挥发性有机物(VOCs)能够维持类囊体膜的完整性, 起到抗氧化作用(Velikova et al., 2011; Arab et al., 2016); 在抵御逆境胁迫方面发挥重要的信号作用(Loreto & Schnitzler, 2010; Bourtsoukidis et al., 2014).Arab等(2016)研究海枣(Phoenix dactylifera)发现, 高温诱导使异戊二烯排放大幅增加; 同时, 在干旱胁迫下脂肪酸增加.Staudt等(2017)发现温度升高能够提高Quercus ilex和Quercus pubescens类异戊二烯的排放量; 干旱胁迫提高单萜释放量, 同时减少类异戊二烯的排放.高温胁迫后花旗松倍半萜、水杨酸甲酯、芳樟醇比单萜排放释放量增加更快(Joó et al., 2011).刘芳等(2013)研究发现, 迷迭香(Rosmarinus officinalis) VOCs中萜类和醛类化合物受到干旱胁迫诱导表现出显著增加.VOCs释放有助于诱导植物非生物应激性抵抗, 增强植物抵御非生物胁迫能力(周帅等, 2012). ...
3-Hydroxy-3-methylglutaryl-coenzyme A reductase kinase and sucrose-phosphate synthase kinase activities in cauliflower florets: Ca 2+ dependence and substrate specificities 1 1998
 ,*浙江农林大学亚热带森林培育国家重点实验室, 浙江临安 311300
,*浙江农林大学亚热带森林培育国家重点实验室, 浙江临安 311300
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT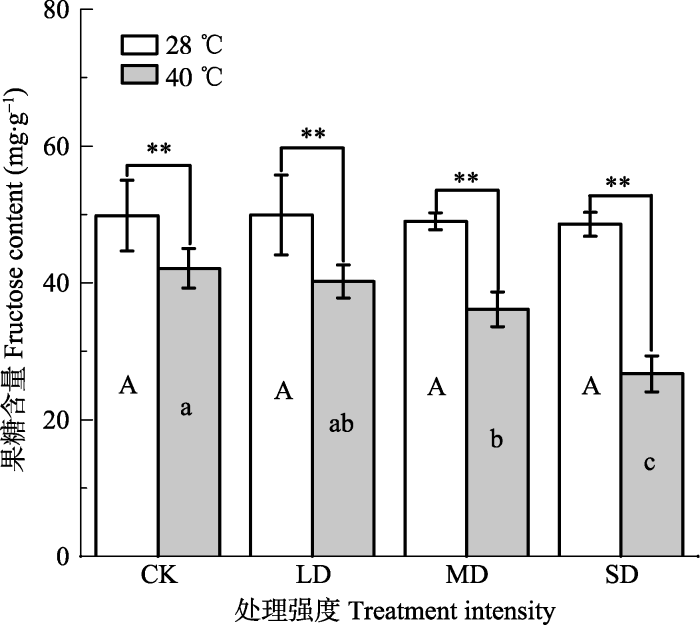 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT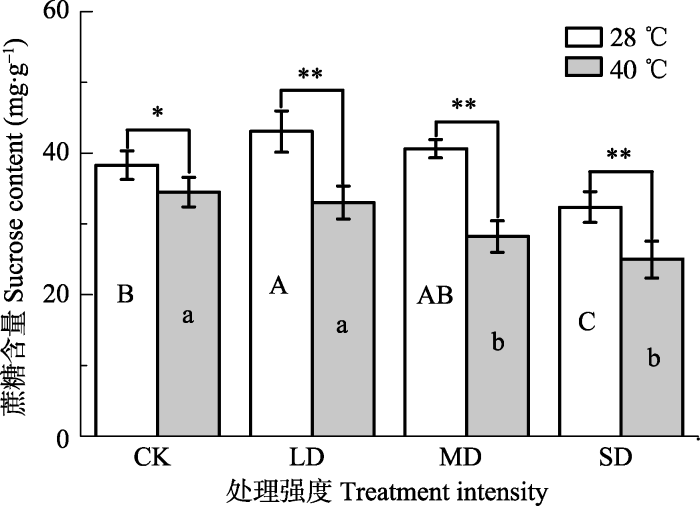 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT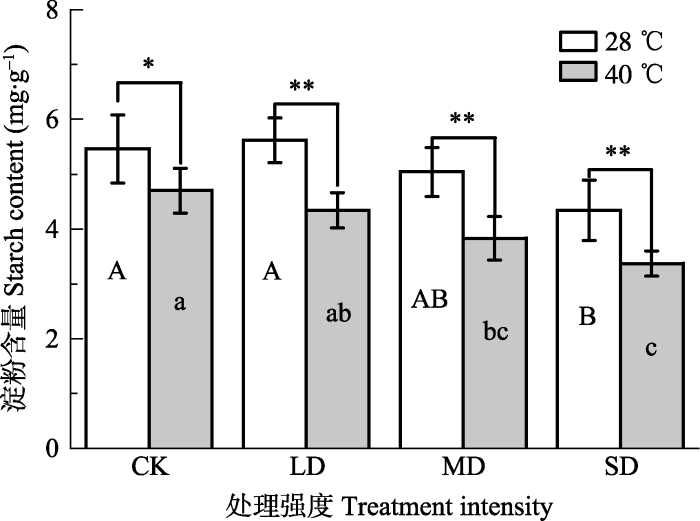 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT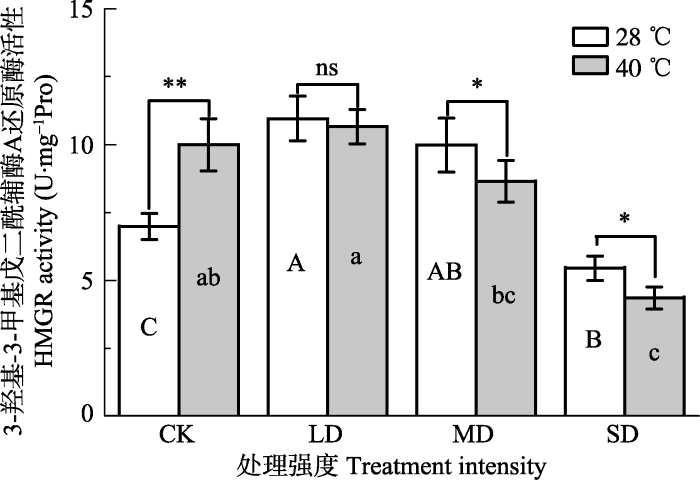 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT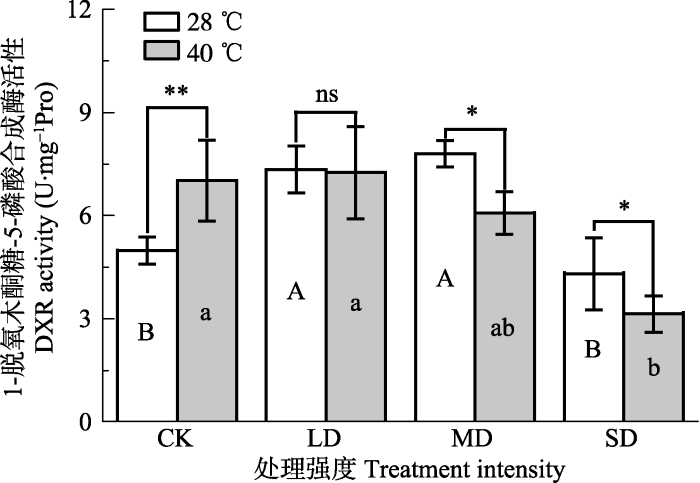 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT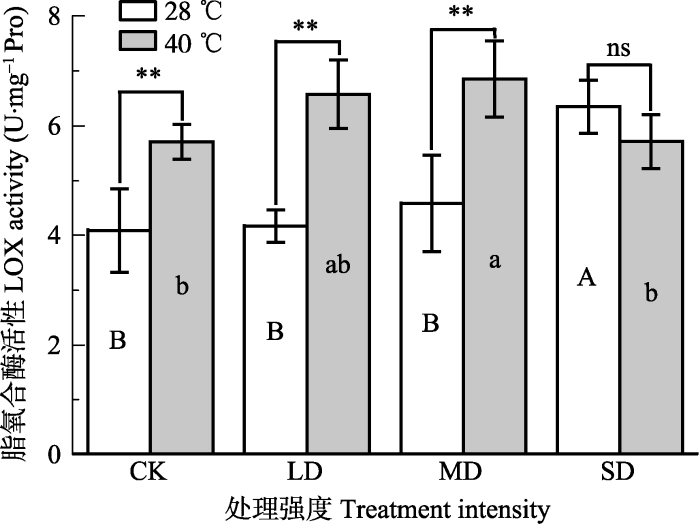 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}