牟静
1 , 宾振钧
2 , 李秋霞
1 , 卜海燕
1 , 张仁懿
1 , 徐当会
, 1 , * 1 兰州大学生命科学学院, 草地农业生态系统国家重点实验室, 兰州 730000 2 广西壮族自治区亚热带作物研究所, 南宁 530001 Effects of nitrogen and silicon addition on soil nitrogen mineralization in alpine meadows of Qinghai-Xizang Plateau MOU Jing
1 , BIN Zhen-Jun
2 , LI Qiu-Xia
1 , BU Hai-Yan
1 , ZHANG Ren-Yi
1 , XU Dang-Hui
, 1 , * 1 School of Life Science, Lanzhou University, State Key Laboratory of Grassland Agro-ecosystems, Lanzhou 730000, China 2 Guangxi Subtropical Crops Research Institute, Nanning 530001, China 通讯作者: * dhxu@lzu.edu.cn 编委: 程晓莉
责任编辑: 李敏
收稿日期: 2018-09-11
接受日期: 2018-10-31
网络出版日期: 2019-01-20
基金资助: 国家重点研发计划 .2016YFC0501906 国家自然科学基金 .30900171 国家自然科学基金 .31460162 国家自然科学基金 .31600336
Received: 2018-09-11
Accepted: 2018-10-31
Online: 2019-01-20
Fund supported: Supported by the National Key R&D Program .2016YFC0501906 the National Natural Science Foundation .30900171 the National Natural Science Foundation .31460162 the National Natural Science Foundation .31600336
摘要 为了解全球气候变化背景下氮沉降对土壤氮矿化的影响及硅添加对土壤氮矿化的促进作用, 该试验设置不同浓度的氮肥单独添加(0、20、40、60 g·m -2 , 分别为对照CK、N20、N40、N60)以及与硅肥配施(硅酸4 g·m -2 , Si4), 测定不同处理下0-20、20-40、40-60 cm土层土壤硝态氮含量、铵态氮含量、净硝化速率、净氨化速率以及净矿化速率。结果显示: (1)单独添加氮肥, 各土层土壤硝态氮和铵态氮含量均随处理浓度的增加而增加, 0-20 cm土层N20、N40、N60处理下土壤硝态氮和铵态氮分别较CK增加63.48%、126.04%、247.03%和80.66%、152.52%、244.56%; 随着土层深度增加, 土壤硝态氮、铵态氮含量均有下降, 20-40、40-60 cm土层较0-20 cm土层硝态氮含量分别平均减少53.90%、76.05%, 铵态氮含量分别平均减少48.62%、68.23%。(2)土壤净硝化速率、净氨化速率及净矿化速率随着氮肥浓度增加均呈上升趋势。相同氮肥添加浓度下, 土壤净硝化速率、净氨化速率和净矿化速率随着土层深度增加逐渐下降(除CK外)。(3)与单独添加氮肥比较, 氮硅肥配施, 土壤氮含量有显著提高, 在0-20 cm土层硝态氮和铵态氮较CK分别增加98.78%、192.62%、330.16%和99.96%、195.82%、306.32%, 20-40、40-60 cm土层也有类似趋势。同时, 氮硅配施促进了土壤氮矿化行为, 在0-20 cm土层, N60Si4处理下的土壤净硝化速率、净氨化速率较单独施氮时分别增加35.88%、27.41%。以上结果表明, 与单独氮肥添加相比, 氮硅配施不但能提高土壤氮含量, 而且能促进土壤氮的矿化作用, 对大气氮沉降有一定的缓解作用。 关键词: 青藏高原 ;
养分添加 ;
硝态氮 ;
铵态氮 ;
土壤氮矿化 ;
氮沉降 Abstract Aims Methods -2 , namely CK, N20, N40, N60, respectively) and combined with silicon fertilizer (4 g·m -2 silicic acid, namely Si4). Soil nitrate and ammonium nitrogen contents, net nitrification rate, net ammonification rate as well as net mineralization rate in 0-20, 20-40 and 40-60 cm depth profiles were measured.Important findings Keywords: Qinghai-Xizang Plateau ;
nutrients addition ;
nitrate nitrogen ;
ammonium nitrogen ;
soil nitrogen mineralization ;
nitrogen deposition PDF (1229KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 本文引用格式 牟静, 宾振钧, 李秋霞, 卜海燕, 张仁懿, 徐当会. 氮硅添加对青藏高原高寒草甸土壤氮矿化的影响.
植物生态学报 [J], 2019, 43(1): 77-84 doi:10.17521/cjpe.2018.0218
MOU Jing.
Effects of nitrogen and silicon addition on soil nitrogen mineralization in alpine meadows of Qinghai-Xizang Plateau .
Chinese Journal of Plant Ecology [J], 2019, 43(1): 77-84 doi:10.17521/cjpe.2018.0218
青藏高原平均海拔在4 000 m以上, 有“世界屋脊”之称, 分布有世界海拔最高的草地。作为青藏高原自然生态系统的主体(
刘兴元等, 2012 ), 高寒草地面积达1.525 × 10
8 hm
2 , 占青藏高原总面积的59.28% (
张镱锂等, 2013 )。由于海拔高、气候寒冷、干旱等因素, 该地区有机氮的利用率低, 仅占土壤全氮的1% (
白洁冰等, 2011 )。近年来, 随着青藏高原东缘区域性氮沉降逐年增加, 该地区土壤中的可用氮增加(
杨晓霞等, 2014 ), 继而对土壤中氮的活动产生影响。添加营养元素是维持草地生态系统养分平衡, 减缓草地退化的重要措施, 添加氮能有效提高土壤有效氮含量, 促进草地生态系统养分循环(
刘晓东等, 2015 )。硅对植物生长同样至关重要, 不仅可增加植物抗逆、抗倒伏能力, 还可活化土壤有机质, 有效促进植物对氮、磷等的吸收(
张文鹏等, 2016 )。但之前的研究多集中于硅对植物群落多样性、植物抗性等方面的影响, 而对土壤氮矿化的影响研究较少。本研究将土壤矿化作为重点, 探讨氮硅添加对土壤矿化的影响, 以期为维持青藏高原草地生态系统的平衡和制定适当管理措施提供有效的理论依据。
1 研究区域和方法 研究样地设于兰州大学高寒草甸与湿地生态系统定位研究站(阿孜分站), 位于青藏高原东缘甘南藏族自治州玛曲县(33.65° N, 101.88° E)。海拔3 650 m, 年平均气温2.2 ℃, 年降水量672 mm, 降水集中在7-8月, 属于高寒半湿润半干旱气候。年日照时间约2 580 h, 年霜期大于270天。样地内的优势种主要有嵩草(
Kobresia myosuroides )、垂穗披碱草(
Elymus nutans )、草地早熟禾(
Poa pratensis )、长毛风毛菊(
Saussurea hieracioides )、星状雪兔子(
S. stella )、钝苞雪莲(
S. nigrescens )、高原毛茛(
Ranunculus tanguticus )、钝裂银莲花(
Anemone obtusiloba )、草玉梅(
A. rivularis )、中国马先蒿(
Pedicularis chinensis )、短腺小米草(
Euphrasia regelii )、莓叶委陵菜(
Potentilla fragarioides )、多枝黄耆(
Astragalus polycladus )、黄花棘豆(
Oxytropis ochrocephala )等。
1.1 样地设计与处理 样地选自禁牧区, 四周采用围栏围起来以防止牛羊进入破坏。试验采取完全区组设计。在样地内选取48个2 m × 2 m 的样方, 各样方之间留2 m间隔, 每个样方四角用PVC管标记。处理前进行土壤采样, 在每个样方内随机选取3个采样点, 使用5 cm孔径的土钻分别钻取0-20、20-40、40-60 cm深度的土样, 装入自封袋带回实验室, 分析其理化性质, 作为土壤氮矿化的初始值。于2012年5月中旬进行氮、硅添加试验, 均为一次性均匀施肥。根据以往在该地的长期施肥试验资料(
刘慧霞等, 2009 ;
张杰琦等, 2010 )并结合实际情况, 本试验设置4组处理, 分别为对照(CK, 不添加NH
4 NO
3 、H
2 SiO
3 ), 单独氮添加处理(添加NH
4 NO
3 20、40、60 g·m
-2 , 分别记为N20、N40、N60, 折合施氮量分别为7、14、21 g·m
-2 ), 单独硅添加处理(添加H
2 SiO
3 4 g·m
-2 , 即Si4, 折合施硅量1.436 g·m
-2 ), 氮、硅配施处理(在N20、N40、N60基础上, 添加硅酸4 g·m
-2 , 分别记为N20Si4, N40Si4, N60Si4), 共计8个处理, 每处理6个重复。处理前土壤pH值6.33, 有机质含量70.52 g·kg
-1 , 全氮含量3.72 g·kg
-1 , 铵态氮、硝态氮含量分别为10.67 mg·kg
-1 和15.25 mg·kg
-1 ; 全磷含量0.98 g·kg
-1 , 速效磷含量25.27 mg·kg
-1 。
于2012年8月中旬, 在每个样方内随机选取3个采样点, 按0-20、20-40、40-60 cm土层深度, 依次用5 cm孔径的土钻取样, 装入自封袋, 带回实验室进行分析, 作为土壤氮矿化的处理值。
1.2 测定方法与数据处理 按常规分析方法测定土壤理化性质。利用pH仪(PHS-5, 上海艾测电子科技有限公司, 上海)测定土壤pH值; 采用重铬酸钾容量法测定土壤有机质含量; 土样分别采用H
2 SO
4 -K
2 SO
4 :CuSO
4 :Se法和HClO
4 - H
2 SO法催化消煮, 消煮后的溶液经定容、沉淀和稀释后采用SmartChem 200全自动智能化学分析仪(WestCo Scientific Instruments, Brookfield, USA)进行全氮、全磷含量分析测定; 采用钼锑抗比色法测定土壤速效磷含量。土壤硝态氮和铵态氮用2 mol·L
-1 ·KCl溶液浸提后, 用SmartChem 200全自动智能化学分析仪测定其含量。依照下式计算土壤氮素矿化速率:
净硝化速率 = (处理后NO
3 - -N - 初始NO
3 - -N)/培养天数
净氨化速率 = (处理后NH
4 + -N - 初始NH
4 + -N)/培养天数
净矿化速率 = ((处理后NH
4 + -N + 处理后NO
3 - -N) - (初始NH
4 + -N +初始NO
3 - -N))/培养天数
采用Excel 2010、SPSS 22.0等软件对数据进行计算和处理, 采用Origin 8.5作图。不同处理间差异采用单因素方差分析检验; 对土壤深度和施肥处理对土壤净矿化速率的影响进行双因素方差分析, 显著性差异水平为
p < 0.05。
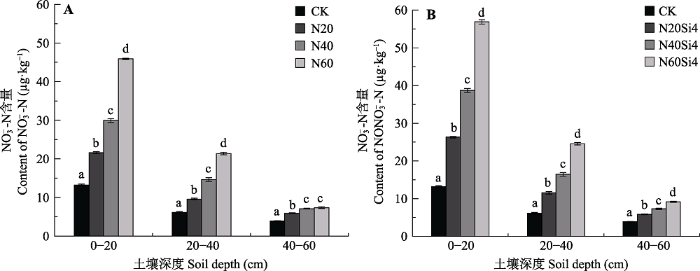
2 结果 2.1 氮硅添加对土壤硝态氮、铵态氮含量的影响 在N20、N40、N60处理下, 0-20 cm土层硝态氮含量较CK分别增加63.48%、126.04%和247.03%; 20-40 cm土层较CK分别增加55.57%、138.20%和247.86%; 40-60 cm土层较CK分别增加52.93%、83.55%和89.55%。随着土壤深度增加, 各处理下土壤硝态氮含量均呈现下降趋势, 20-40、40-60 cm土层较0-20 cm土层硝态氮含量分别平均减少53.90%和76.05% (
图1A )。氮硅配施时, 随着氮肥浓度的增加土壤硝态氮含量显著增加(
p < 0.05), 0-20 cm土层较CK分别增加98.78%、192.62%和330.16%; 20-40 cm土层分别增加88.06%、167.62%和299.70%; 40-60 cm土层分别增加51.43%、89.18%和136.92% (
图1B )。养分处理对土壤硝态氮含量的影响随着土壤深度的增加逐渐减弱。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1青藏高原高寒草甸不同施氮浓度下不同土层硝态氮含量变化(平均值±标准误差, n = 6)。CK、N20、N40、N60, 分别添加NH4 NO3 0、20、40、60 g·m-2 ; Si4, 添加硅酸4 g·m-2 。不同小写字母表示处理间差异显著(p < 0.05)。 Fig. 1Change of NO3 - -N content in alpine meadows of Qinghai-Xizang Plateau under different nitrogen concentrations (mean ± SE, n = 6). CK, N20, N40, N60, added NH4 NO3 0, 20, 40, 60 g·m-2 , respectively; Si4, added silicic acid 4 g·m-2 . Different lowercase letters indicate significant differences among treatments (p < 0.05). 单独添加氮肥时, 0-20 cm土层铵态氮含量较CK分别增加80.66%、152.52%和244.56%。20-40、40-60 cm铵态氮含量较CK分别增加59.94%、56.22%、132.30%和129.81%、217.79%、176.37%。随着土壤深度的增加, 土壤铵态氮含量逐渐减少, 20-40、40-60 cm土层较0-20 cm分别平均减少48.62%、68.23% (
图2A )。当氮硅配施时, 各土层铵态氮含量较硝态氮含量增幅更大, 0-20 cm土层增加99.96%、195.82%和306.32%; 20-40、40-60 cm土层铵态氮含量分别增加82.22%、175.06%、291.49%和82.45%、164.09%、225.67% (
图2B )。随着土壤深度增加, 各养分处理对土壤铵态氮含量的影响也逐渐减弱。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2青藏高原高寒草甸不同施氮浓度下不同土层铵态氮含量变化(平均值±标准误差, n = 6)。CK、N20、N40、N60, 分别添加NH4 NO3 0、20、40、60 g·m-2 ; Si4, 添加硅酸4 g·m-2 。不同小写字母表示处理间差异显著(p < 0.05)。 Fig. 2Change of NH4 + -N content in alpine meadows of Qinghai-Xizang Plateau under different nitrogen concentrations (mean ± SE, n = 6). CK, N20, N40, N60, added NH4 NO3 0, 20, 40, 60 g·m-2 , respectively; Si4, added silicic acid 4 g·m-2 . Different lowercase letters indicate significant differences among treatments (p < 0.05). 2.2 氮硅添加对土壤氮矿化的影响 如
图3A 所示, 施加氮肥前, 各土层净硝化速率均为负值, 其中20-40 cm土壤净硝化速率达最小值-26.65 μg·kg
-1 ·d
-1 。单独施加氮肥时, 各土层净硝化速率随着施氮水平的增加呈上升趋势, 其中0-20 cm土层增幅最大, 20-40 cm次之, 40-60 cm增幅最小(
图3A )。在施硅肥的基础上, 逐浓度增施氮肥, 各土层净硝化速率显著增加, 其最大值462.61 μg·kg
-1 ·d
-1 出现在N60Si4处理下0-20 cm土层, 且各土层增幅与单独施加氮肥时类似, 0-20 cm土层增幅最大, 20-40 cm次之, 40-60 cm 增幅最小(
图3B )。0-20、20-40 cm土层净硝化速率均高于单独添加氮肥时的净硝化速率, 但在40-60 cm 土层, N20Si4处理时则出现了后者高于前者的情况。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3青藏高原高寒草甸不同施氮浓度下土壤净硝化速率(平均值±标准误差, n = 6)。CK、N20、N40、N60, 分别添加NH4 NO3 0、20、40、60 g·m-2 ; Si4, 添加硅酸4 g·m-2 。不同小写字母表示处理间差异显著(p < 0.05)。 Fig. 3Net nitrification rates under different nitrogen concentrations in alpine meadows of Qinghai-Xizang Plateau (mean ± SE, n = 6). CK, N20, N40, N60, added NH4 NO3 0, 20, 40, 60 g·m-2 , respectively; Si4, added silicic acid 4 g·m-2 . Different lowercase letters indicate significant differences among treatments (p < 0.05). 如
图4A 所示, 氮处理前, 各土层净氨化速率均为负值, 其中在0-20 cm达最小值-22.63 μg·kg
-1 ·d
-1 , 且随土壤深度的增加, 净氨化速率逐渐增大。单独添加氮肥时, 各土层的净氨化速率随氮肥浓度的增加而增加, 其中0-20 cm土层增幅较20-40、40-60 cm土层大。以氮硅配施的方法处理, 土壤净氨化速率最大值270.66 μg·kg
-1 ·d
-1 同样出现在0-20 cm土层, 且氮硅配施时各土层净氨化速率均大于单氮施加时的净氨化速率。
图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4青藏高原高寒草甸不同施氮浓度下土壤净氨化速率(平均值±标准误差, n = 6)。CK、N20、N40、N60, 分别添加NH4 NO3 0、20、40、60 g·m-2 ; Si4, 添加硅酸4 g·m-2 。不同小写字母表示处理间差异显著(p < 0.05)。 Fig. 4Net ammonification rates under different nitrogen concentrations in alpine meadows of Qinghai-Xizang Plateau (mean ± SE, n = 6). CK, N20, N40, N60, added NH4 NO3 0, 20, 40, 60 g·m-2 , respectively; Si4, added silicic acid 4 g·m-2 . Different lowercase letters indicate significant differences among treatments (p < 0.05). 土壤净矿化速率在氮处理前均为负值, 且土壤净矿化速率随土层深度的增加而增加, 但始终小于0 (
图5A )。单独施加氮肥时, 0-20 cm土层、N60处理下的净矿化速率达552.89 μg·kg
-1 ·d
-1 , 较N20增加335.69%。氮硅配施, 土壤净矿化速率较单独施加氮肥时显著增加, 在各土层中, N20Si4、N40Si4、N60Si4分别较N20、N40、N60处理下的土壤矿化速率增加55.16%、112.86%、61.19% (
图5B )。
图5 新窗口打开 |
下载原图ZIP |
生成PPT 图5青藏高原高寒草甸不同施氮浓度下土壤净矿化速率(平均值±标准误差, n = 6)。CK、N20、N40、N60, 分别添加NH4 NO3 0、20、40、60 g·m-2 ; Si4, 添加硅酸4 g·m-2 。不同小写字母表示处理间差异显著(p < 0.05)。 Fig. 5Net mineralization rates under different nitrogen concentrations in alpine meadows of Qinghai-Xizang Plateau (mean ± SE, n = 6). CK, N20, N40, N60, added NH4 NO3 0, 20, 40, 60 g·m-2 , respectively; Si4, added silicic acid 4 g·m-2 . Different lowercase letters indicate significant differences among treatments (p < 0.05). 不同施肥处理下各土层间土壤净硝化速率(除CK外)、净氨化速率和净矿化速率间均存在显著性差异(
p < 0.05)(
图3 -5), 故除施肥外, 土壤深度也是影响土壤矿化行为的关键因子, 且两者对土壤净矿化速率存在交互作用(
p < 0.05)(
表1 )。
Table 1 表1 表1 施肥、土壤深度对青藏高原高寒草甸土壤净矿化速率的双因素方差分析结果
Table 1
Two-factor variance analysis between fertilization and soil depths on soil net mineralization rate in alpine meadows of Qinghai-Xizang Plateau 来源 Source 第三类平方和 Type III sum of squares df 均方 Mean Square F 显著性 Sig. 校正模型 Corrected Model 1 436 023.182a 8 179 502.898 5 325.359 0.000 截距 Intercept 1 238 798.353 1 1 238 798.353 36 751.753 0.000 NSi 906 007.721 2 453 003.860 13 439.383 0.000 SD 318 252.753 2 159 126.376 4 720.844 0.000 NSi × SD 211 762.709 4 52 940.677 1 570.605 0.000 误差 Error 606.729 18 33.707 总计 Total 2 675 428.265 27 校正后总数 Corrected total 1 436 629.912 26
a, R 2 = 1.000 (adjusted R 2 = 0.999). NSi, nitrogen and silicon combined; SD, soil depth. a, R 2 = 1.000 (调整的R 2 = 0.999)。NSi, 氮硅配施; SD, 土壤深度。 新窗口打开 |
下载CSV 3 讨论 氮矿化的研究对于揭示生态系统功能, 为维持生态系统稳定提供理论指导有重要意义。氮是植物生长发育必需的营养元素, 是植物吸收量最大的土壤矿质元素(
赵阳等, 2013 ), 也是影响整个陆地系统生产力的重要因子, 通常被认为是植物生长发育重要限制因素(
司晓林等, 2016 )。氮在土壤中绝大部分是以有机形式存在的, 约占土壤全氮含量的92%-98%, 土壤有机氮无法直接被植物吸收利用, 必须借助土壤微生物, 通过矿化作用将其转化为无机氮。土壤中无机氮主要形态有铵态氮和硝态氮, 二者都具水溶性, 更易被植物吸收利用。自18世纪后期, 土壤氮矿化已被视为土壤氮循环的核心, 在控制植物有效氮利用方面起到至关重要的作用(
Aber & Melillo, 2001 )。
本研究结果表明, 施加氮肥可提高土壤有效氮含量, 即硝态氮和铵态氮的含量, 且浅层土壤(0-20 cm)的响应较深层土壤(20-40、40-60 cm)强烈(
图1 ,
图2 ), 这与
郭雅婧等(2015) 的研究结果一致。铵态氮主要以交换态形式存在, 较小的土粒(<0.001 mm)一般呈负电性, 可以吸附交换态的铵态氮, 因此, 土壤中铵态氮的移动范围一般都较小; 相反, 硝态氮呈负电性, 难以被土粒吸附, 在土壤中移动范围较铵态氮大, 故在各土层分布应相对均匀(
奚振邦, 2011 )。但在本研究中, 硝态氮和铵态氮在各土层的分布情况类似, 即在氮处理后, 二者含量均随着土层深度的增加呈现出递减的趋势。可能的解释是硝态氮在土壤中活动性强, 有利于植物深层的根系吸收, 但也易流失。
施肥可促进高寒草甸土壤的矿化作用, 且与土壤深度对土壤的矿化作用存在交互作用(
表1 ), 就本试验而言, 0-20 cm土层矿化行为较20-40、40-60 cm土层强烈, 这与
Aber等(1989) 、
Gundersen等(1998) 对森林生态系统土壤的研究结果一致。但随着研究的不断深入,
Aber和Magill (2004) 发现氮输入后期, 氮输入量与土壤矿化水平呈现一定的负相关关系, 即在长期的氮添加试验中, 森林土壤的矿化速率先随着氮输入量的增加而增大, 继而达到一个峰值, 当氮输入量达到一定水平, 土壤矿化速率便与之呈现一定的负相关关系。而在本试验中未出现此结果, 可能的原因是草地生态系统与森林生态系统土壤对养分添加的响应机制不同, 两种生态系统内部结构差异明显, 植被类型也大不相同, 故而产生不同的响应; 又因本试验为短期试验, 且是一次性均匀施肥, 所以结论可能会与
Aber和Magill (2004) 的研究有所不同。土壤净硝化速率、净氨化速率和净矿化速率在处理前均为负值(
图3 -
图5 ), 其原因可能是土壤中原有的氮含量较少, 微生物仅可用于维持自身生长繁殖, 分解更多有机氮对其来说有较大难度, 故不能为土壤无机氮库做出贡献, 因而土壤净硝化速率、净氨化速率和净矿化速率在处理前均呈现负值。随着无机氮处理浓度增加, 微生物利用量逐渐饱和, 故土壤净硝化速率、净氨化速率和净矿化速率转向正值(
白洁冰等, 2011 )。
硅虽然是地壳中仅次于氧的第二大元素, 但土壤中的大部分硅通常会结合其他物质, 形成氧化性硅酸, 植物无法直接吸收利用, 但硅在植物的生长过程中发挥着不可或缺的作用, 不仅可以活化土壤有机质, 促进植物根对氮磷等矿质元素的吸收, 增加植物抗性(
宾振钧等, 2015 ;
张文鹏等, 2016 ), 提高植物叶片的光合作用和植物群落的物种多样性(
Xu et al ., 2015 ,
2018 ), 还可以缓解环境中污染物, 如可抑制植物光合作用的镉对植物的损害, 且硅对植物的解毒作用因物种、品种及生态型的不同而表现出不同的解毒机制(
刘彩凤等, 2017 )。本研究结果表明, 氮硅配施使土壤中的有效氮含量显著增加, 且显著高于单独施氮时的含量, 土壤矿化活动也较单独施氮时活跃, 即土壤净硝化速率、净氨化速率和净矿化速率均高于单独施氮时。说明施加硅肥能促进土壤矿化行为, 硅元素对高等植物的矿质营养有重要影响, 该结果与
Fu等(2001) 对水稻的研究结果一致。
与单独氮肥添加相比, 氮硅配施不但能提高不同土层的氮含量, 而且能促进土壤氮的矿化作用, 但在本研究中, 只涉及一种浓度的硅, 即Si4与不同浓度氮肥配施, 进而对土壤矿化作用产生影响。若要进一步研究硅肥与氮肥对土壤矿化的交互作用, 需在本试验基础上, 增设多个硅浓度。在后续的研究中可进一步提出在多种氮硅组合中, 是否存在一个最佳氮硅组合, 使得土壤矿化作用达到最优, 继而将其作为衡量草地生态系统的养分循环是否良好及生态系统平衡与否的有效指标。但目前的研究多集中于硅添加对植被地上生物量、生物多样性、群落结构等方面的影响, 对硅肥作用的研究也多集中于其对植物在各种胁迫条件下的抗性、缓解金属污染物对植物损害等方面, 少有研究涉及其对土壤矿化作用的影响, 尤其是和氮肥配施对土壤矿化的促进作用。因此, 养分添加对青藏高原地区土壤氮矿化的影响还有待更广泛、更深入的研究。
致谢 感谢兰州大学生命科学学院理论生态所的所有老师、同学在试验中给予的指导与帮助。
[1] Aber JD Magill AH 2004 ). Chronic nitrogen additions at the Harvard Forest (USA): The first 15 years of a nitrogen saturation experimentForest Ecology and Management, 196, 1 -5 . DOI:
10.1016/j.foreco.2004.03.009 URL [本文引用: 2] [2] Aber JD Melillo JM 2001 ). Terrestrial Ecosystem . 2nd edn. Harcourt Academic Press, San Diego, USA. [本文引用: 1] [3] Aber JD Nadelhoffer KJ Steudler P Melillo JM 1989 ). Nitrogen saturation in northern forest ecosystems hypotheses and implicationsBioscience, 39, 378 -386 . DOI:
10.2307/1311067 URL [本文引用: 1] [4] Bai JB Xu XL Fu G Song MH He YT Jiang J 2011 ). Effects of temperature and nitrogen input on nitrogen mineralization in alpine soils on the Tibetan PlateauJournal of Anhui Agriculture Science, 39, 14698 -14700, 14756 . [本文引用: 2] [ 白洁冰 , 徐兴良 , 付刚 , 宋明华 , 何永涛 , 蒋婧 ( 2011 ). 温度和氮素输入对青藏高原3种高寒草地土壤氮矿化的影响安徽农业科学 , 39, 14698 -14700, 14756 .] [本文引用: 2] [5] Bai JB Xu XL Song MH He YT Jiang J Shi PL 2011 ). Effects of temperature and added nitrogen on carbon mineralization in alpine soils on the Tibetan PlateauEcology and Environmental Sciences, 20, 855 -859 . [ 白洁冰 , 徐兴良 , 宋明华 , 何永涛 , 蒋婧 , 石培礼 ( 2011 ). 温度和氮素输入对青藏高原三种高寒草地土壤碳矿化的影响生态环境学报 , 20, 855 -859 .] [6] Bin ZJ Zhang RY Zhang WP Xu DH 2015 ). Effects of nitrogen, phosphorus and silicon addition on leaf carbon, nitrogen, and phosphorus concentration of Elymus nutans of alpine meadow on Qinghai-Tibetan Plateau, China. Acta Ecologica Sinica, 4699 -4706 . [本文引用: 1] [ 宾振钧 , 张仁懿 , 张文鹏 , 徐当会 ( 2015 ). 氮磷硅添加对青藏高原高寒草甸垂穗披碱草叶片碳氮磷的影响生态学报 , 35, 4699 -4706 .] [本文引用: 1] [7] Fu JR Ho A Kenichi K 2001 ). Nitrogen mineralization in two paddy soils as affected by silica source additionActa Agriculturae Zhejiangensis, 13(3 ), 133 -140 . [本文引用: 1] [8] Gundersen P Emmett BA Kjonaas OJ Koopmans CJ Tietema A 1998 ). Impact of nitrogen deposition on nitrogen cycling in forest: A synthesis of NITREX dataForest Ecology and Management, 101, 37 -55 . DOI:
10.1016/S0378-1127(97)00124-2 URL [本文引用: 1] [9] Guo YJ 2015 ). Effects of nitrogen Addition on Alpine Meadow Vegetation and soil Nitrogen Supply Capacity in the Qinghai-Xizang Plateau. Master degree dissertation, Lanzhou University, Lanzhou. [本文引用: 1] [ 郭雅婧 ( 2015 ). 氮素添加对青藏高原高寒草甸植被和土壤氮素供应能力的影响硕士学位论文, 兰州大学, 兰州 .] [本文引用: 1] [10] Liu CF Shi GR Yu RG Zhang Z 2017 ). Eco-physiological mechanisms of silicon-induced alleviation of cadmium toxicity in plants: A reviewActa Ecologica Sinica, 37, 7799 -7810 . [本文引用: 1] [ 刘彩凤 , 史刚荣 , 余如刚 , 张铮 ( 2017 ). 硅缓解植物镉毒害的生理生态机制生态学报 , 37, 7799 -7810 .] [本文引用: 1] [11] Liu HX Guo ZG Guo XH Zhou XR Hui WS Wang KY 2009 ). Effect of addition of silicon on water use efficiency and yield components of alfalfa under the different soil moistureActa Ecologica Sinica, 29, 3075 -3080 . [本文引用: 1] [ 刘慧霞 , 郭正刚 , 郭兴华 , 周雪荣 , 惠文森 , 王康英 ( 2009 ). 不同土壤水分条件下硅对紫花苜蓿水分利用效率及产量构成要素的影响生态学报 , 29, 3075 -3080 .] [本文引用: 1] [12] Liu XD Yin GL Wu J Chen JG Ma LX Shi SL 2015 ). Effects of nitrogen addition on the physical properties of soil in an alpine meadow on the eastern Qinghai-Tibetan PlateauActa Prataculturae Sinica, 24(10 ), 12 -21 . DOI:
10.11686/cyxb2015154 Magsci [本文引用: 1] 依托设置于青藏高原东部夏河县桑科草原的天然草地刈割型草场培育定位试验,探讨不同氮元素添加量对高寒草甸草地土壤物理性状的影响。本试验共设4个处理,分别为对照(CK,0 kg N/hm<sup>2</sup>)、低氮(LN,50 kg N/hm<sup>2</sup>)、中氮(MN,100 kg N/hm<sup>2</sup>)、高氮(HN,150 kg N/hm<sup>2</sup>)。通过对高寒草甸草地土壤容重、孔隙度、紧实度、土壤团聚体等物理性状进行测定与分析,结果表明,MN与HN处理可显著降低0~10 cm土壤容重与土壤紧实度、增加土壤总孔隙度、非毛管孔隙度与最大持水量。20~30 cm土层中LN、MN、HN处理均可显著增加土壤容重、土壤紧实度,降低土壤总孔隙度、最大持水量。对土壤团聚体的影响,与CK处理相比MN、HN处理可显著提高各土层≥0.25 mm机械稳定性土壤团聚体含量、提升土壤团聚体平均重量直径(MWD);较之CK处理,LN、MN处理可显著提升各土层≥0.25 mm水稳性团聚体含量,同时不同氮元素添加处理均可显著提升水稳性团聚体和MWD,表明氮元素添加对该区土壤侵蚀性有较好的抑制作用,进而减少水土和养分的流失。对地上和地下生物量的影响,MN、HN处理能显著提升草地地上生物量;MN处理显著增加了地下生物量、HN处理显著降低了地下生物量;LN处理对地上与地下生物量的影响不显著。通过试验得出结论MN处理对土壤物理性状的改善效果较好,该处理改善了土壤容重、孔隙度及土壤紧实度,提高了土壤干筛≥0.25 mm机械稳定性团聚体与湿筛≥0.25 mm水稳性团聚体含量及其稳定性(MWD),增加了草地地上生物量和地下生物量。
[ 刘晓东 , 尹国丽 , 武均 , 陈建纲 , 马隆喜 , 师尚礼 ( 2015 ). 青藏高原东部高寒草甸草地土壤物理性状对氮元素添加的响应草业学报 , 24(10 ), 12 -21 .] DOI:
10.11686/cyxb2015154 Magsci [本文引用: 1] 依托设置于青藏高原东部夏河县桑科草原的天然草地刈割型草场培育定位试验,探讨不同氮元素添加量对高寒草甸草地土壤物理性状的影响。本试验共设4个处理,分别为对照(CK,0 kg N/hm<sup>2</sup>)、低氮(LN,50 kg N/hm<sup>2</sup>)、中氮(MN,100 kg N/hm<sup>2</sup>)、高氮(HN,150 kg N/hm<sup>2</sup>)。通过对高寒草甸草地土壤容重、孔隙度、紧实度、土壤团聚体等物理性状进行测定与分析,结果表明,MN与HN处理可显著降低0~10 cm土壤容重与土壤紧实度、增加土壤总孔隙度、非毛管孔隙度与最大持水量。20~30 cm土层中LN、MN、HN处理均可显著增加土壤容重、土壤紧实度,降低土壤总孔隙度、最大持水量。对土壤团聚体的影响,与CK处理相比MN、HN处理可显著提高各土层≥0.25 mm机械稳定性土壤团聚体含量、提升土壤团聚体平均重量直径(MWD);较之CK处理,LN、MN处理可显著提升各土层≥0.25 mm水稳性团聚体含量,同时不同氮元素添加处理均可显著提升水稳性团聚体和MWD,表明氮元素添加对该区土壤侵蚀性有较好的抑制作用,进而减少水土和养分的流失。对地上和地下生物量的影响,MN、HN处理能显著提升草地地上生物量;MN处理显著增加了地下生物量、HN处理显著降低了地下生物量;LN处理对地上与地下生物量的影响不显著。通过试验得出结论MN处理对土壤物理性状的改善效果较好,该处理改善了土壤容重、孔隙度及土壤紧实度,提高了土壤干筛≥0.25 mm机械稳定性团聚体与湿筛≥0.25 mm水稳性团聚体含量及其稳定性(MWD),增加了草地地上生物量和地下生物量。
[13] Liu XY Long RJ Shang ZH 2012 ). Interactive mechanism of service function of alpine rangeland ecosystems in Qinghai?-Tibetan PlateauActa Ecologica Sinica, 32, 7688 -7697 . [本文引用: 1] [ 刘兴元 , 龙瑞军 , 尚占环 ( 2012 ). 青藏高原高寒草地生态系统服务功能的互作机制生态学报 , 32, 7688 -7697 .] [本文引用: 1] [14] Si XL Wang WY Gao XG Xu DH 2016 ). Effects of nitrogen and silicon application on leaf nitrogen content and net photosynthetic rate of Elymus nutans in alpine meadow. Chinese Journal of Plant Ecology, 1238 -1244 . [本文引用: 1] [ 司晓林 , 王文银 , 高小刚 , 徐当会 ( 2016 ). 氮硅添加对高寒草甸垂穗披碱草叶片全氮含量及净光合速率的影响植物生态学报 , 40, 1238 -1244 .] [本文引用: 1] [15] Xi ZB 2011 ). Ammonium nitrogen and nitrate nitrogenPhosphate & Compound Fertilizer, 26(1 ), 62 -65 . [本文引用: 1] [ 奚振邦 ( 2011 ). 硝态氮与铵态氮磷肥与复肥 , 26(1 ), 62 -65 .] [本文引用: 1] [16] Xu DH Fang XW Zhang RY Gao TP Bu HY Du GZ 2015 ). Influences of nitrogen, phosphorus and silicon addition on plant productivity and species richness in an alpine meadow AoB Plants, 10.1093/aobpla/plv125 . [本文引用: 1] [17] Xu DH Gao XG Gao TP Mou J Li JH Bu HY Zhang RY Li QX 2018 ). Interactive effects of nitrogen and silicon addition on growth of five common plant species and structure of plant community in alpine meadowCatena, 169, 80 -89 . DOI:
10.1016/j.catena.2018.05.017 URL [本文引用: 1] [18] Yang XX Ren F Zhou HK He JS 2014 ). Responses of plant community biomass to nitrogen and phosphorus additions in an alpine meadow on the Qinghai-Xizang PlateauChinese Journal of Plant Ecology, 38, 159 -166 . [本文引用: 1] [ 杨晓霞 , 任飞 , 周华坤 , 贺金生 ( 2014 ), 青藏高原高寒草甸植物群落生物量对氮、磷添加的响应植物生态学报 , 38, 159 -166 .] [本文引用: 1] [19] Zhang JQ Li Q Ren ZW Yang X Wang G 2010 ). Effects of nitrogen addition on species richness and relationship between species richness and aboveground productivity of alpine meadow of the Qinghai-Tibetan Platen, ChinaChinese Journal of Plant Ecology, 34, 1125 -1131 . [本文引用: 1] [ 张杰琦 , 李奇 , 任正炜 , 杨雪 , 王刚 . 氮素添加对青藏高原高寒草甸植物群落物种丰富度及其与地上生产力关系的影响植物生态学报 , 34, 1125 -1131 .] [本文引用: 1] [20] Zhang WP Si XL Wang WY Gao TP Xu DH 2016 ). Effects of short-term nitrogen and silicon addition on above-?ground biomass and biodiversity of alpine meadow of the Qinghai-Tibetan Plateau, ChinaPratacultural Science, 33, 38 -45 . [本文引用: 2] [ 张文鹏 , 司晓林 , 王文银 , 高天鹏 , 徐当会 ( 2016 ). 氮硅添加对高寒草甸生物量和多样性的影响 ——以青藏高原为例草业科学 , 33, 38 -45 .] [本文引用: 2] [21] Zhang YL Qi W Zhou CP Ding MJ Liu LS Gao JG Bai WQ Wang ZF Zheng D 2013 ). Spatial and temporal variability in the net primary production (NPP) of alpine grassland on Tibetan Plateau from 1982 to 2009Acta Geographica Sinica, 68, 1197 -1211 . [本文引用: 1] [ 张镱锂 , 祁威 , 周才平 , 丁明军 , 刘林山 , 高俊刚 , 摆万奇 , 王兆锋 , 郑度 ( 2013 ). 青藏高原高寒草地净初级生产力(NPP)时空分异地理学报 , 68, 1197 -1211 .] [本文引用: 1] [22] Zhao Y Zhang C Zhao HF Xu XN 2013 ). Effects of N and P addition on soil nitrogen mineralization in a subtropical evergreen broadleaved forestChinese Journal of Ecology, 32, 1690 -1697 . [本文引用: 1] [ 赵阳 , 张驰 , 赵竑绯 , 徐小牛 ( 2013 ). 氮磷添加对亚热带常绿阔叶林土壤氮素矿化的影响生态学杂志 , 32, 1690 -1697 .] [本文引用: 1] Chronic nitrogen additions at the Harvard Forest (USA): The first 15 years of a nitrogen saturation experiment
2
2004
... 施肥可促进高寒草甸土壤的矿化作用, 且与土壤深度对土壤的矿化作用存在交互作用(
表1 ), 就本试验而言, 0-20 cm土层矿化行为较20-40、40-60 cm土层强烈, 这与
Aber等(1989) 、
Gundersen等(1998) 对森林生态系统土壤的研究结果一致.但随着研究的不断深入,
Aber和Magill (2004) 发现氮输入后期, 氮输入量与土壤矿化水平呈现一定的负相关关系, 即在长期的氮添加试验中, 森林土壤的矿化速率先随着氮输入量的增加而增大, 继而达到一个峰值, 当氮输入量达到一定水平, 土壤矿化速率便与之呈现一定的负相关关系.而在本试验中未出现此结果, 可能的原因是草地生态系统与森林生态系统土壤对养分添加的响应机制不同, 两种生态系统内部结构差异明显, 植被类型也大不相同, 故而产生不同的响应; 又因本试验为短期试验, 且是一次性均匀施肥, 所以结论可能会与
Aber和Magill (2004) 的研究有所不同.土壤净硝化速率、净氨化速率和净矿化速率在处理前均为负值(
图3 -
图5 ), 其原因可能是土壤中原有的氮含量较少, 微生物仅可用于维持自身生长繁殖, 分解更多有机氮对其来说有较大难度, 故不能为土壤无机氮库做出贡献, 因而土壤净硝化速率、净氨化速率和净矿化速率在处理前均呈现负值.随着无机氮处理浓度增加, 微生物利用量逐渐饱和, 故土壤净硝化速率、净氨化速率和净矿化速率转向正值(
白洁冰等, 2011 ). ...
... 发现氮输入后期, 氮输入量与土壤矿化水平呈现一定的负相关关系, 即在长期的氮添加试验中, 森林土壤的矿化速率先随着氮输入量的增加而增大, 继而达到一个峰值, 当氮输入量达到一定水平, 土壤矿化速率便与之呈现一定的负相关关系.而在本试验中未出现此结果, 可能的原因是草地生态系统与森林生态系统土壤对养分添加的响应机制不同, 两种生态系统内部结构差异明显, 植被类型也大不相同, 故而产生不同的响应; 又因本试验为短期试验, 且是一次性均匀施肥, 所以结论可能会与
Aber和Magill (2004) 的研究有所不同.土壤净硝化速率、净氨化速率和净矿化速率在处理前均为负值(
图3 -
图5 ), 其原因可能是土壤中原有的氮含量较少, 微生物仅可用于维持自身生长繁殖, 分解更多有机氮对其来说有较大难度, 故不能为土壤无机氮库做出贡献, 因而土壤净硝化速率、净氨化速率和净矿化速率在处理前均呈现负值.随着无机氮处理浓度增加, 微生物利用量逐渐饱和, 故土壤净硝化速率、净氨化速率和净矿化速率转向正值(
白洁冰等, 2011 ). ...
1
2001
... 氮矿化的研究对于揭示生态系统功能, 为维持生态系统稳定提供理论指导有重要意义.氮是植物生长发育必需的营养元素, 是植物吸收量最大的土壤矿质元素(
赵阳等, 2013 ), 也是影响整个陆地系统生产力的重要因子, 通常被认为是植物生长发育重要限制因素(
司晓林等, 2016 ).氮在土壤中绝大部分是以有机形式存在的, 约占土壤全氮含量的92%-98%, 土壤有机氮无法直接被植物吸收利用, 必须借助土壤微生物, 通过矿化作用将其转化为无机氮.土壤中无机氮主要形态有铵态氮和硝态氮, 二者都具水溶性, 更易被植物吸收利用.自18世纪后期, 土壤氮矿化已被视为土壤氮循环的核心, 在控制植物有效氮利用方面起到至关重要的作用(
Aber & Melillo, 2001 ). ...
Nitrogen saturation in northern forest ecosystems hypotheses and implications
1
1989
... 施肥可促进高寒草甸土壤的矿化作用, 且与土壤深度对土壤的矿化作用存在交互作用(
表1 ), 就本试验而言, 0-20 cm土层矿化行为较20-40、40-60 cm土层强烈, 这与
Aber等(1989) 、
Gundersen等(1998) 对森林生态系统土壤的研究结果一致.但随着研究的不断深入,
Aber和Magill (2004) 发现氮输入后期, 氮输入量与土壤矿化水平呈现一定的负相关关系, 即在长期的氮添加试验中, 森林土壤的矿化速率先随着氮输入量的增加而增大, 继而达到一个峰值, 当氮输入量达到一定水平, 土壤矿化速率便与之呈现一定的负相关关系.而在本试验中未出现此结果, 可能的原因是草地生态系统与森林生态系统土壤对养分添加的响应机制不同, 两种生态系统内部结构差异明显, 植被类型也大不相同, 故而产生不同的响应; 又因本试验为短期试验, 且是一次性均匀施肥, 所以结论可能会与
Aber和Magill (2004) 的研究有所不同.土壤净硝化速率、净氨化速率和净矿化速率在处理前均为负值(
图3 -
图5 ), 其原因可能是土壤中原有的氮含量较少, 微生物仅可用于维持自身生长繁殖, 分解更多有机氮对其来说有较大难度, 故不能为土壤无机氮库做出贡献, 因而土壤净硝化速率、净氨化速率和净矿化速率在处理前均呈现负值.随着无机氮处理浓度增加, 微生物利用量逐渐饱和, 故土壤净硝化速率、净氨化速率和净矿化速率转向正值(
白洁冰等, 2011 ). ...
温度和氮素输入对青藏高原3种高寒草地土壤氮矿化的影响
2
2011
... 青藏高原平均海拔在4 000 m以上, 有“世界屋脊”之称, 分布有世界海拔最高的草地.作为青藏高原自然生态系统的主体(
刘兴元等, 2012 ), 高寒草地面积达1.525 × 10
8 hm
2 , 占青藏高原总面积的59.28% (
张镱锂等, 2013 ).由于海拔高、气候寒冷、干旱等因素, 该地区有机氮的利用率低, 仅占土壤全氮的1% (
白洁冰等, 2011 ).近年来, 随着青藏高原东缘区域性氮沉降逐年增加, 该地区土壤中的可用氮增加(
杨晓霞等, 2014 ), 继而对土壤中氮的活动产生影响.添加营养元素是维持草地生态系统养分平衡, 减缓草地退化的重要措施, 添加氮能有效提高土壤有效氮含量, 促进草地生态系统养分循环(
刘晓东等, 2015 ).硅对植物生长同样至关重要, 不仅可增加植物抗逆、抗倒伏能力, 还可活化土壤有机质, 有效促进植物对氮、磷等的吸收(
张文鹏等, 2016 ).但之前的研究多集中于硅对植物群落多样性、植物抗性等方面的影响, 而对土壤氮矿化的影响研究较少.本研究将土壤矿化作为重点, 探讨氮硅添加对土壤矿化的影响, 以期为维持青藏高原草地生态系统的平衡和制定适当管理措施提供有效的理论依据. ...
... 施肥可促进高寒草甸土壤的矿化作用, 且与土壤深度对土壤的矿化作用存在交互作用(
表1 ), 就本试验而言, 0-20 cm土层矿化行为较20-40、40-60 cm土层强烈, 这与
Aber等(1989) 、
Gundersen等(1998) 对森林生态系统土壤的研究结果一致.但随着研究的不断深入,
Aber和Magill (2004) 发现氮输入后期, 氮输入量与土壤矿化水平呈现一定的负相关关系, 即在长期的氮添加试验中, 森林土壤的矿化速率先随着氮输入量的增加而增大, 继而达到一个峰值, 当氮输入量达到一定水平, 土壤矿化速率便与之呈现一定的负相关关系.而在本试验中未出现此结果, 可能的原因是草地生态系统与森林生态系统土壤对养分添加的响应机制不同, 两种生态系统内部结构差异明显, 植被类型也大不相同, 故而产生不同的响应; 又因本试验为短期试验, 且是一次性均匀施肥, 所以结论可能会与
Aber和Magill (2004) 的研究有所不同.土壤净硝化速率、净氨化速率和净矿化速率在处理前均为负值(
图3 -
图5 ), 其原因可能是土壤中原有的氮含量较少, 微生物仅可用于维持自身生长繁殖, 分解更多有机氮对其来说有较大难度, 故不能为土壤无机氮库做出贡献, 因而土壤净硝化速率、净氨化速率和净矿化速率在处理前均呈现负值.随着无机氮处理浓度增加, 微生物利用量逐渐饱和, 故土壤净硝化速率、净氨化速率和净矿化速率转向正值(
白洁冰等, 2011 ). ...
温度和氮素输入对青藏高原3种高寒草地土壤氮矿化的影响
2
2011
... 青藏高原平均海拔在4 000 m以上, 有“世界屋脊”之称, 分布有世界海拔最高的草地.作为青藏高原自然生态系统的主体(
刘兴元等, 2012 ), 高寒草地面积达1.525 × 10
8 hm
2 , 占青藏高原总面积的59.28% (
张镱锂等, 2013 ).由于海拔高、气候寒冷、干旱等因素, 该地区有机氮的利用率低, 仅占土壤全氮的1% (
白洁冰等, 2011 ).近年来, 随着青藏高原东缘区域性氮沉降逐年增加, 该地区土壤中的可用氮增加(
杨晓霞等, 2014 ), 继而对土壤中氮的活动产生影响.添加营养元素是维持草地生态系统养分平衡, 减缓草地退化的重要措施, 添加氮能有效提高土壤有效氮含量, 促进草地生态系统养分循环(
刘晓东等, 2015 ).硅对植物生长同样至关重要, 不仅可增加植物抗逆、抗倒伏能力, 还可活化土壤有机质, 有效促进植物对氮、磷等的吸收(
张文鹏等, 2016 ).但之前的研究多集中于硅对植物群落多样性、植物抗性等方面的影响, 而对土壤氮矿化的影响研究较少.本研究将土壤矿化作为重点, 探讨氮硅添加对土壤矿化的影响, 以期为维持青藏高原草地生态系统的平衡和制定适当管理措施提供有效的理论依据. ...
... 施肥可促进高寒草甸土壤的矿化作用, 且与土壤深度对土壤的矿化作用存在交互作用(
表1 ), 就本试验而言, 0-20 cm土层矿化行为较20-40、40-60 cm土层强烈, 这与
Aber等(1989) 、
Gundersen等(1998) 对森林生态系统土壤的研究结果一致.但随着研究的不断深入,
Aber和Magill (2004) 发现氮输入后期, 氮输入量与土壤矿化水平呈现一定的负相关关系, 即在长期的氮添加试验中, 森林土壤的矿化速率先随着氮输入量的增加而增大, 继而达到一个峰值, 当氮输入量达到一定水平, 土壤矿化速率便与之呈现一定的负相关关系.而在本试验中未出现此结果, 可能的原因是草地生态系统与森林生态系统土壤对养分添加的响应机制不同, 两种生态系统内部结构差异明显, 植被类型也大不相同, 故而产生不同的响应; 又因本试验为短期试验, 且是一次性均匀施肥, 所以结论可能会与
Aber和Magill (2004) 的研究有所不同.土壤净硝化速率、净氨化速率和净矿化速率在处理前均为负值(
图3 -
图5 ), 其原因可能是土壤中原有的氮含量较少, 微生物仅可用于维持自身生长繁殖, 分解更多有机氮对其来说有较大难度, 故不能为土壤无机氮库做出贡献, 因而土壤净硝化速率、净氨化速率和净矿化速率在处理前均呈现负值.随着无机氮处理浓度增加, 微生物利用量逐渐饱和, 故土壤净硝化速率、净氨化速率和净矿化速率转向正值(
白洁冰等, 2011 ). ...
温度和氮素输入对青藏高原三种高寒草地土壤碳矿化的影响
0
2011
温度和氮素输入对青藏高原三种高寒草地土壤碳矿化的影响
0
2011
氮磷硅添加对青藏高原高寒草甸垂穗披碱草叶片碳氮磷的影响
1
2015
... 硅虽然是地壳中仅次于氧的第二大元素, 但土壤中的大部分硅通常会结合其他物质, 形成氧化性硅酸, 植物无法直接吸收利用, 但硅在植物的生长过程中发挥着不可或缺的作用, 不仅可以活化土壤有机质, 促进植物根对氮磷等矿质元素的吸收, 增加植物抗性(
宾振钧等, 2015 ;
张文鹏等, 2016 ), 提高植物叶片的光合作用和植物群落的物种多样性(
Xu et al ., 2015 ,
2018 ), 还可以缓解环境中污染物, 如可抑制植物光合作用的镉对植物的损害, 且硅对植物的解毒作用因物种、品种及生态型的不同而表现出不同的解毒机制(
刘彩凤等, 2017 ).本研究结果表明, 氮硅配施使土壤中的有效氮含量显著增加, 且显著高于单独施氮时的含量, 土壤矿化活动也较单独施氮时活跃, 即土壤净硝化速率、净氨化速率和净矿化速率均高于单独施氮时.说明施加硅肥能促进土壤矿化行为, 硅元素对高等植物的矿质营养有重要影响, 该结果与
Fu等(2001) 对水稻的研究结果一致. ...
氮磷硅添加对青藏高原高寒草甸垂穗披碱草叶片碳氮磷的影响
1
2015
... 硅虽然是地壳中仅次于氧的第二大元素, 但土壤中的大部分硅通常会结合其他物质, 形成氧化性硅酸, 植物无法直接吸收利用, 但硅在植物的生长过程中发挥着不可或缺的作用, 不仅可以活化土壤有机质, 促进植物根对氮磷等矿质元素的吸收, 增加植物抗性(
宾振钧等, 2015 ;
张文鹏等, 2016 ), 提高植物叶片的光合作用和植物群落的物种多样性(
Xu et al ., 2015 ,
2018 ), 还可以缓解环境中污染物, 如可抑制植物光合作用的镉对植物的损害, 且硅对植物的解毒作用因物种、品种及生态型的不同而表现出不同的解毒机制(
刘彩凤等, 2017 ).本研究结果表明, 氮硅配施使土壤中的有效氮含量显著增加, 且显著高于单独施氮时的含量, 土壤矿化活动也较单独施氮时活跃, 即土壤净硝化速率、净氨化速率和净矿化速率均高于单独施氮时.说明施加硅肥能促进土壤矿化行为, 硅元素对高等植物的矿质营养有重要影响, 该结果与
Fu等(2001) 对水稻的研究结果一致. ...
Nitrogen mineralization in two paddy soils as affected by silica source addition
1
2001
... 硅虽然是地壳中仅次于氧的第二大元素, 但土壤中的大部分硅通常会结合其他物质, 形成氧化性硅酸, 植物无法直接吸收利用, 但硅在植物的生长过程中发挥着不可或缺的作用, 不仅可以活化土壤有机质, 促进植物根对氮磷等矿质元素的吸收, 增加植物抗性(
宾振钧等, 2015 ;
张文鹏等, 2016 ), 提高植物叶片的光合作用和植物群落的物种多样性(
Xu et al ., 2015 ,
2018 ), 还可以缓解环境中污染物, 如可抑制植物光合作用的镉对植物的损害, 且硅对植物的解毒作用因物种、品种及生态型的不同而表现出不同的解毒机制(
刘彩凤等, 2017 ).本研究结果表明, 氮硅配施使土壤中的有效氮含量显著增加, 且显著高于单独施氮时的含量, 土壤矿化活动也较单独施氮时活跃, 即土壤净硝化速率、净氨化速率和净矿化速率均高于单独施氮时.说明施加硅肥能促进土壤矿化行为, 硅元素对高等植物的矿质营养有重要影响, 该结果与
Fu等(2001) 对水稻的研究结果一致. ...
Impact of nitrogen deposition on nitrogen cycling in forest: A synthesis of NITREX data
1
1998
... 施肥可促进高寒草甸土壤的矿化作用, 且与土壤深度对土壤的矿化作用存在交互作用(
表1 ), 就本试验而言, 0-20 cm土层矿化行为较20-40、40-60 cm土层强烈, 这与
Aber等(1989) 、
Gundersen等(1998) 对森林生态系统土壤的研究结果一致.但随着研究的不断深入,
Aber和Magill (2004) 发现氮输入后期, 氮输入量与土壤矿化水平呈现一定的负相关关系, 即在长期的氮添加试验中, 森林土壤的矿化速率先随着氮输入量的增加而增大, 继而达到一个峰值, 当氮输入量达到一定水平, 土壤矿化速率便与之呈现一定的负相关关系.而在本试验中未出现此结果, 可能的原因是草地生态系统与森林生态系统土壤对养分添加的响应机制不同, 两种生态系统内部结构差异明显, 植被类型也大不相同, 故而产生不同的响应; 又因本试验为短期试验, 且是一次性均匀施肥, 所以结论可能会与
Aber和Magill (2004) 的研究有所不同.土壤净硝化速率、净氨化速率和净矿化速率在处理前均为负值(
图3 -
图5 ), 其原因可能是土壤中原有的氮含量较少, 微生物仅可用于维持自身生长繁殖, 分解更多有机氮对其来说有较大难度, 故不能为土壤无机氮库做出贡献, 因而土壤净硝化速率、净氨化速率和净矿化速率在处理前均呈现负值.随着无机氮处理浓度增加, 微生物利用量逐渐饱和, 故土壤净硝化速率、净氨化速率和净矿化速率转向正值(
白洁冰等, 2011 ). ...
氮素添加对青藏高原高寒草甸植被和土壤氮素供应能力的影响
1
2015
... 本研究结果表明, 施加氮肥可提高土壤有效氮含量, 即硝态氮和铵态氮的含量, 且浅层土壤(0-20 cm)的响应较深层土壤(20-40、40-60 cm)强烈(
图1 ,
图2 ), 这与
郭雅婧等(2015) 的研究结果一致.铵态氮主要以交换态形式存在, 较小的土粒(<0.001 mm)一般呈负电性, 可以吸附交换态的铵态氮, 因此, 土壤中铵态氮的移动范围一般都较小; 相反, 硝态氮呈负电性, 难以被土粒吸附, 在土壤中移动范围较铵态氮大, 故在各土层分布应相对均匀(
奚振邦, 2011 ).但在本研究中, 硝态氮和铵态氮在各土层的分布情况类似, 即在氮处理后, 二者含量均随着土层深度的增加呈现出递减的趋势.可能的解释是硝态氮在土壤中活动性强, 有利于植物深层的根系吸收, 但也易流失. ...
氮素添加对青藏高原高寒草甸植被和土壤氮素供应能力的影响
1
2015
... 本研究结果表明, 施加氮肥可提高土壤有效氮含量, 即硝态氮和铵态氮的含量, 且浅层土壤(0-20 cm)的响应较深层土壤(20-40、40-60 cm)强烈(
图1 ,
图2 ), 这与
郭雅婧等(2015) 的研究结果一致.铵态氮主要以交换态形式存在, 较小的土粒(<0.001 mm)一般呈负电性, 可以吸附交换态的铵态氮, 因此, 土壤中铵态氮的移动范围一般都较小; 相反, 硝态氮呈负电性, 难以被土粒吸附, 在土壤中移动范围较铵态氮大, 故在各土层分布应相对均匀(
奚振邦, 2011 ).但在本研究中, 硝态氮和铵态氮在各土层的分布情况类似, 即在氮处理后, 二者含量均随着土层深度的增加呈现出递减的趋势.可能的解释是硝态氮在土壤中活动性强, 有利于植物深层的根系吸收, 但也易流失. ...
硅缓解植物镉毒害的生理生态机制
1
2017
... 硅虽然是地壳中仅次于氧的第二大元素, 但土壤中的大部分硅通常会结合其他物质, 形成氧化性硅酸, 植物无法直接吸收利用, 但硅在植物的生长过程中发挥着不可或缺的作用, 不仅可以活化土壤有机质, 促进植物根对氮磷等矿质元素的吸收, 增加植物抗性(
宾振钧等, 2015 ;
张文鹏等, 2016 ), 提高植物叶片的光合作用和植物群落的物种多样性(
Xu et al ., 2015 ,
2018 ), 还可以缓解环境中污染物, 如可抑制植物光合作用的镉对植物的损害, 且硅对植物的解毒作用因物种、品种及生态型的不同而表现出不同的解毒机制(
刘彩凤等, 2017 ).本研究结果表明, 氮硅配施使土壤中的有效氮含量显著增加, 且显著高于单独施氮时的含量, 土壤矿化活动也较单独施氮时活跃, 即土壤净硝化速率、净氨化速率和净矿化速率均高于单独施氮时.说明施加硅肥能促进土壤矿化行为, 硅元素对高等植物的矿质营养有重要影响, 该结果与
Fu等(2001) 对水稻的研究结果一致. ...
硅缓解植物镉毒害的生理生态机制
1
2017
... 硅虽然是地壳中仅次于氧的第二大元素, 但土壤中的大部分硅通常会结合其他物质, 形成氧化性硅酸, 植物无法直接吸收利用, 但硅在植物的生长过程中发挥着不可或缺的作用, 不仅可以活化土壤有机质, 促进植物根对氮磷等矿质元素的吸收, 增加植物抗性(
宾振钧等, 2015 ;
张文鹏等, 2016 ), 提高植物叶片的光合作用和植物群落的物种多样性(
Xu et al ., 2015 ,
2018 ), 还可以缓解环境中污染物, 如可抑制植物光合作用的镉对植物的损害, 且硅对植物的解毒作用因物种、品种及生态型的不同而表现出不同的解毒机制(
刘彩凤等, 2017 ).本研究结果表明, 氮硅配施使土壤中的有效氮含量显著增加, 且显著高于单独施氮时的含量, 土壤矿化活动也较单独施氮时活跃, 即土壤净硝化速率、净氨化速率和净矿化速率均高于单独施氮时.说明施加硅肥能促进土壤矿化行为, 硅元素对高等植物的矿质营养有重要影响, 该结果与
Fu等(2001) 对水稻的研究结果一致. ...
不同土壤水分条件下硅对紫花苜蓿水分利用效率及产量构成要素的影响
1
2009
... 样地选自禁牧区, 四周采用围栏围起来以防止牛羊进入破坏.试验采取完全区组设计.在样地内选取48个2 m × 2 m 的样方, 各样方之间留2 m间隔, 每个样方四角用PVC管标记.处理前进行土壤采样, 在每个样方内随机选取3个采样点, 使用5 cm孔径的土钻分别钻取0-20、20-40、40-60 cm深度的土样, 装入自封袋带回实验室, 分析其理化性质, 作为土壤氮矿化的初始值.于2012年5月中旬进行氮、硅添加试验, 均为一次性均匀施肥.根据以往在该地的长期施肥试验资料(
刘慧霞等, 2009 ;
张杰琦等, 2010 )并结合实际情况, 本试验设置4组处理, 分别为对照(CK, 不添加NH
4 NO
3 、H
2 SiO
3 ), 单独氮添加处理(添加NH
4 NO
3 20、40、60 g·m
-2 , 分别记为N20、N40、N60, 折合施氮量分别为7、14、21 g·m
-2 ), 单独硅添加处理(添加H
2 SiO
3 4 g·m
-2 , 即Si4, 折合施硅量1.436 g·m
-2 ), 氮、硅配施处理(在N20、N40、N60基础上, 添加硅酸4 g·m
-2 , 分别记为N20Si4, N40Si4, N60Si4), 共计8个处理, 每处理6个重复.处理前土壤pH值6.33, 有机质含量70.52 g·kg
-1 , 全氮含量3.72 g·kg
-1 , 铵态氮、硝态氮含量分别为10.67 mg·kg
-1 和15.25 mg·kg
-1 ; 全磷含量0.98 g·kg
-1 , 速效磷含量25.27 mg·kg
-1 . ...
不同土壤水分条件下硅对紫花苜蓿水分利用效率及产量构成要素的影响
1
2009
... 样地选自禁牧区, 四周采用围栏围起来以防止牛羊进入破坏.试验采取完全区组设计.在样地内选取48个2 m × 2 m 的样方, 各样方之间留2 m间隔, 每个样方四角用PVC管标记.处理前进行土壤采样, 在每个样方内随机选取3个采样点, 使用5 cm孔径的土钻分别钻取0-20、20-40、40-60 cm深度的土样, 装入自封袋带回实验室, 分析其理化性质, 作为土壤氮矿化的初始值.于2012年5月中旬进行氮、硅添加试验, 均为一次性均匀施肥.根据以往在该地的长期施肥试验资料(
刘慧霞等, 2009 ;
张杰琦等, 2010 )并结合实际情况, 本试验设置4组处理, 分别为对照(CK, 不添加NH
4 NO
3 、H
2 SiO
3 ), 单独氮添加处理(添加NH
4 NO
3 20、40、60 g·m
-2 , 分别记为N20、N40、N60, 折合施氮量分别为7、14、21 g·m
-2 ), 单独硅添加处理(添加H
2 SiO
3 4 g·m
-2 , 即Si4, 折合施硅量1.436 g·m
-2 ), 氮、硅配施处理(在N20、N40、N60基础上, 添加硅酸4 g·m
-2 , 分别记为N20Si4, N40Si4, N60Si4), 共计8个处理, 每处理6个重复.处理前土壤pH值6.33, 有机质含量70.52 g·kg
-1 , 全氮含量3.72 g·kg
-1 , 铵态氮、硝态氮含量分别为10.67 mg·kg
-1 和15.25 mg·kg
-1 ; 全磷含量0.98 g·kg
-1 , 速效磷含量25.27 mg·kg
-1 . ...
青藏高原东部高寒草甸草地土壤物理性状对氮元素添加的响应
1
2015
... 青藏高原平均海拔在4 000 m以上, 有“世界屋脊”之称, 分布有世界海拔最高的草地.作为青藏高原自然生态系统的主体(
刘兴元等, 2012 ), 高寒草地面积达1.525 × 10
8 hm
2 , 占青藏高原总面积的59.28% (
张镱锂等, 2013 ).由于海拔高、气候寒冷、干旱等因素, 该地区有机氮的利用率低, 仅占土壤全氮的1% (
白洁冰等, 2011 ).近年来, 随着青藏高原东缘区域性氮沉降逐年增加, 该地区土壤中的可用氮增加(
杨晓霞等, 2014 ), 继而对土壤中氮的活动产生影响.添加营养元素是维持草地生态系统养分平衡, 减缓草地退化的重要措施, 添加氮能有效提高土壤有效氮含量, 促进草地生态系统养分循环(
刘晓东等, 2015 ).硅对植物生长同样至关重要, 不仅可增加植物抗逆、抗倒伏能力, 还可活化土壤有机质, 有效促进植物对氮、磷等的吸收(
张文鹏等, 2016 ).但之前的研究多集中于硅对植物群落多样性、植物抗性等方面的影响, 而对土壤氮矿化的影响研究较少.本研究将土壤矿化作为重点, 探讨氮硅添加对土壤矿化的影响, 以期为维持青藏高原草地生态系统的平衡和制定适当管理措施提供有效的理论依据. ...
青藏高原东部高寒草甸草地土壤物理性状对氮元素添加的响应
1
2015
... 青藏高原平均海拔在4 000 m以上, 有“世界屋脊”之称, 分布有世界海拔最高的草地.作为青藏高原自然生态系统的主体(
刘兴元等, 2012 ), 高寒草地面积达1.525 × 10
8 hm
2 , 占青藏高原总面积的59.28% (
张镱锂等, 2013 ).由于海拔高、气候寒冷、干旱等因素, 该地区有机氮的利用率低, 仅占土壤全氮的1% (
白洁冰等, 2011 ).近年来, 随着青藏高原东缘区域性氮沉降逐年增加, 该地区土壤中的可用氮增加(
杨晓霞等, 2014 ), 继而对土壤中氮的活动产生影响.添加营养元素是维持草地生态系统养分平衡, 减缓草地退化的重要措施, 添加氮能有效提高土壤有效氮含量, 促进草地生态系统养分循环(
刘晓东等, 2015 ).硅对植物生长同样至关重要, 不仅可增加植物抗逆、抗倒伏能力, 还可活化土壤有机质, 有效促进植物对氮、磷等的吸收(
张文鹏等, 2016 ).但之前的研究多集中于硅对植物群落多样性、植物抗性等方面的影响, 而对土壤氮矿化的影响研究较少.本研究将土壤矿化作为重点, 探讨氮硅添加对土壤矿化的影响, 以期为维持青藏高原草地生态系统的平衡和制定适当管理措施提供有效的理论依据. ...
青藏高原高寒草地生态系统服务功能的互作机制
1
2012
... 青藏高原平均海拔在4 000 m以上, 有“世界屋脊”之称, 分布有世界海拔最高的草地.作为青藏高原自然生态系统的主体(
刘兴元等, 2012 ), 高寒草地面积达1.525 × 10
8 hm
2 , 占青藏高原总面积的59.28% (
张镱锂等, 2013 ).由于海拔高、气候寒冷、干旱等因素, 该地区有机氮的利用率低, 仅占土壤全氮的1% (
白洁冰等, 2011 ).近年来, 随着青藏高原东缘区域性氮沉降逐年增加, 该地区土壤中的可用氮增加(
杨晓霞等, 2014 ), 继而对土壤中氮的活动产生影响.添加营养元素是维持草地生态系统养分平衡, 减缓草地退化的重要措施, 添加氮能有效提高土壤有效氮含量, 促进草地生态系统养分循环(
刘晓东等, 2015 ).硅对植物生长同样至关重要, 不仅可增加植物抗逆、抗倒伏能力, 还可活化土壤有机质, 有效促进植物对氮、磷等的吸收(
张文鹏等, 2016 ).但之前的研究多集中于硅对植物群落多样性、植物抗性等方面的影响, 而对土壤氮矿化的影响研究较少.本研究将土壤矿化作为重点, 探讨氮硅添加对土壤矿化的影响, 以期为维持青藏高原草地生态系统的平衡和制定适当管理措施提供有效的理论依据. ...
青藏高原高寒草地生态系统服务功能的互作机制
1
2012
... 青藏高原平均海拔在4 000 m以上, 有“世界屋脊”之称, 分布有世界海拔最高的草地.作为青藏高原自然生态系统的主体(
刘兴元等, 2012 ), 高寒草地面积达1.525 × 10
8 hm
2 , 占青藏高原总面积的59.28% (
张镱锂等, 2013 ).由于海拔高、气候寒冷、干旱等因素, 该地区有机氮的利用率低, 仅占土壤全氮的1% (
白洁冰等, 2011 ).近年来, 随着青藏高原东缘区域性氮沉降逐年增加, 该地区土壤中的可用氮增加(
杨晓霞等, 2014 ), 继而对土壤中氮的活动产生影响.添加营养元素是维持草地生态系统养分平衡, 减缓草地退化的重要措施, 添加氮能有效提高土壤有效氮含量, 促进草地生态系统养分循环(
刘晓东等, 2015 ).硅对植物生长同样至关重要, 不仅可增加植物抗逆、抗倒伏能力, 还可活化土壤有机质, 有效促进植物对氮、磷等的吸收(
张文鹏等, 2016 ).但之前的研究多集中于硅对植物群落多样性、植物抗性等方面的影响, 而对土壤氮矿化的影响研究较少.本研究将土壤矿化作为重点, 探讨氮硅添加对土壤矿化的影响, 以期为维持青藏高原草地生态系统的平衡和制定适当管理措施提供有效的理论依据. ...
氮硅添加对高寒草甸垂穗披碱草叶片全氮含量及净光合速率的影响
1
2016
... 氮矿化的研究对于揭示生态系统功能, 为维持生态系统稳定提供理论指导有重要意义.氮是植物生长发育必需的营养元素, 是植物吸收量最大的土壤矿质元素(
赵阳等, 2013 ), 也是影响整个陆地系统生产力的重要因子, 通常被认为是植物生长发育重要限制因素(
司晓林等, 2016 ).氮在土壤中绝大部分是以有机形式存在的, 约占土壤全氮含量的92%-98%, 土壤有机氮无法直接被植物吸收利用, 必须借助土壤微生物, 通过矿化作用将其转化为无机氮.土壤中无机氮主要形态有铵态氮和硝态氮, 二者都具水溶性, 更易被植物吸收利用.自18世纪后期, 土壤氮矿化已被视为土壤氮循环的核心, 在控制植物有效氮利用方面起到至关重要的作用(
Aber & Melillo, 2001 ). ...
氮硅添加对高寒草甸垂穗披碱草叶片全氮含量及净光合速率的影响
1
2016
... 氮矿化的研究对于揭示生态系统功能, 为维持生态系统稳定提供理论指导有重要意义.氮是植物生长发育必需的营养元素, 是植物吸收量最大的土壤矿质元素(
赵阳等, 2013 ), 也是影响整个陆地系统生产力的重要因子, 通常被认为是植物生长发育重要限制因素(
司晓林等, 2016 ).氮在土壤中绝大部分是以有机形式存在的, 约占土壤全氮含量的92%-98%, 土壤有机氮无法直接被植物吸收利用, 必须借助土壤微生物, 通过矿化作用将其转化为无机氮.土壤中无机氮主要形态有铵态氮和硝态氮, 二者都具水溶性, 更易被植物吸收利用.自18世纪后期, 土壤氮矿化已被视为土壤氮循环的核心, 在控制植物有效氮利用方面起到至关重要的作用(
Aber & Melillo, 2001 ). ...
硝态氮与铵态氮
1
2011
... 本研究结果表明, 施加氮肥可提高土壤有效氮含量, 即硝态氮和铵态氮的含量, 且浅层土壤(0-20 cm)的响应较深层土壤(20-40、40-60 cm)强烈(
图1 ,
图2 ), 这与
郭雅婧等(2015) 的研究结果一致.铵态氮主要以交换态形式存在, 较小的土粒(<0.001 mm)一般呈负电性, 可以吸附交换态的铵态氮, 因此, 土壤中铵态氮的移动范围一般都较小; 相反, 硝态氮呈负电性, 难以被土粒吸附, 在土壤中移动范围较铵态氮大, 故在各土层分布应相对均匀(
奚振邦, 2011 ).但在本研究中, 硝态氮和铵态氮在各土层的分布情况类似, 即在氮处理后, 二者含量均随着土层深度的增加呈现出递减的趋势.可能的解释是硝态氮在土壤中活动性强, 有利于植物深层的根系吸收, 但也易流失. ...
硝态氮与铵态氮
1
2011
... 本研究结果表明, 施加氮肥可提高土壤有效氮含量, 即硝态氮和铵态氮的含量, 且浅层土壤(0-20 cm)的响应较深层土壤(20-40、40-60 cm)强烈(
图1 ,
图2 ), 这与
郭雅婧等(2015) 的研究结果一致.铵态氮主要以交换态形式存在, 较小的土粒(<0.001 mm)一般呈负电性, 可以吸附交换态的铵态氮, 因此, 土壤中铵态氮的移动范围一般都较小; 相反, 硝态氮呈负电性, 难以被土粒吸附, 在土壤中移动范围较铵态氮大, 故在各土层分布应相对均匀(
奚振邦, 2011 ).但在本研究中, 硝态氮和铵态氮在各土层的分布情况类似, 即在氮处理后, 二者含量均随着土层深度的增加呈现出递减的趋势.可能的解释是硝态氮在土壤中活动性强, 有利于植物深层的根系吸收, 但也易流失. ...
Influences of nitrogen, phosphorus and silicon addition on plant productivity and species richness in an alpine meadow
1
2015
... 硅虽然是地壳中仅次于氧的第二大元素, 但土壤中的大部分硅通常会结合其他物质, 形成氧化性硅酸, 植物无法直接吸收利用, 但硅在植物的生长过程中发挥着不可或缺的作用, 不仅可以活化土壤有机质, 促进植物根对氮磷等矿质元素的吸收, 增加植物抗性(
宾振钧等, 2015 ;
张文鹏等, 2016 ), 提高植物叶片的光合作用和植物群落的物种多样性(
Xu et al ., 2015 ,
2018 ), 还可以缓解环境中污染物, 如可抑制植物光合作用的镉对植物的损害, 且硅对植物的解毒作用因物种、品种及生态型的不同而表现出不同的解毒机制(
刘彩凤等, 2017 ).本研究结果表明, 氮硅配施使土壤中的有效氮含量显著增加, 且显著高于单独施氮时的含量, 土壤矿化活动也较单独施氮时活跃, 即土壤净硝化速率、净氨化速率和净矿化速率均高于单独施氮时.说明施加硅肥能促进土壤矿化行为, 硅元素对高等植物的矿质营养有重要影响, 该结果与
Fu等(2001) 对水稻的研究结果一致. ...
Interactive effects of nitrogen and silicon addition on growth of five common plant species and structure of plant community in alpine meadow
1
2018
... 硅虽然是地壳中仅次于氧的第二大元素, 但土壤中的大部分硅通常会结合其他物质, 形成氧化性硅酸, 植物无法直接吸收利用, 但硅在植物的生长过程中发挥着不可或缺的作用, 不仅可以活化土壤有机质, 促进植物根对氮磷等矿质元素的吸收, 增加植物抗性(
宾振钧等, 2015 ;
张文鹏等, 2016 ), 提高植物叶片的光合作用和植物群落的物种多样性(
Xu et al ., 2015 ,
2018 ), 还可以缓解环境中污染物, 如可抑制植物光合作用的镉对植物的损害, 且硅对植物的解毒作用因物种、品种及生态型的不同而表现出不同的解毒机制(
刘彩凤等, 2017 ).本研究结果表明, 氮硅配施使土壤中的有效氮含量显著增加, 且显著高于单独施氮时的含量, 土壤矿化活动也较单独施氮时活跃, 即土壤净硝化速率、净氨化速率和净矿化速率均高于单独施氮时.说明施加硅肥能促进土壤矿化行为, 硅元素对高等植物的矿质营养有重要影响, 该结果与
Fu等(2001) 对水稻的研究结果一致. ...
青藏高原高寒草甸植物群落生物量对氮、磷添加的响应
1
2014
... 青藏高原平均海拔在4 000 m以上, 有“世界屋脊”之称, 分布有世界海拔最高的草地.作为青藏高原自然生态系统的主体(
刘兴元等, 2012 ), 高寒草地面积达1.525 × 10
8 hm
2 , 占青藏高原总面积的59.28% (
张镱锂等, 2013 ).由于海拔高、气候寒冷、干旱等因素, 该地区有机氮的利用率低, 仅占土壤全氮的1% (
白洁冰等, 2011 ).近年来, 随着青藏高原东缘区域性氮沉降逐年增加, 该地区土壤中的可用氮增加(
杨晓霞等, 2014 ), 继而对土壤中氮的活动产生影响.添加营养元素是维持草地生态系统养分平衡, 减缓草地退化的重要措施, 添加氮能有效提高土壤有效氮含量, 促进草地生态系统养分循环(
刘晓东等, 2015 ).硅对植物生长同样至关重要, 不仅可增加植物抗逆、抗倒伏能力, 还可活化土壤有机质, 有效促进植物对氮、磷等的吸收(
张文鹏等, 2016 ).但之前的研究多集中于硅对植物群落多样性、植物抗性等方面的影响, 而对土壤氮矿化的影响研究较少.本研究将土壤矿化作为重点, 探讨氮硅添加对土壤矿化的影响, 以期为维持青藏高原草地生态系统的平衡和制定适当管理措施提供有效的理论依据. ...
青藏高原高寒草甸植物群落生物量对氮、磷添加的响应
1
2014
... 青藏高原平均海拔在4 000 m以上, 有“世界屋脊”之称, 分布有世界海拔最高的草地.作为青藏高原自然生态系统的主体(
刘兴元等, 2012 ), 高寒草地面积达1.525 × 10
8 hm
2 , 占青藏高原总面积的59.28% (
张镱锂等, 2013 ).由于海拔高、气候寒冷、干旱等因素, 该地区有机氮的利用率低, 仅占土壤全氮的1% (
白洁冰等, 2011 ).近年来, 随着青藏高原东缘区域性氮沉降逐年增加, 该地区土壤中的可用氮增加(
杨晓霞等, 2014 ), 继而对土壤中氮的活动产生影响.添加营养元素是维持草地生态系统养分平衡, 减缓草地退化的重要措施, 添加氮能有效提高土壤有效氮含量, 促进草地生态系统养分循环(
刘晓东等, 2015 ).硅对植物生长同样至关重要, 不仅可增加植物抗逆、抗倒伏能力, 还可活化土壤有机质, 有效促进植物对氮、磷等的吸收(
张文鹏等, 2016 ).但之前的研究多集中于硅对植物群落多样性、植物抗性等方面的影响, 而对土壤氮矿化的影响研究较少.本研究将土壤矿化作为重点, 探讨氮硅添加对土壤矿化的影响, 以期为维持青藏高原草地生态系统的平衡和制定适当管理措施提供有效的理论依据. ...
氮素添加对青藏高原高寒草甸植物群落物种丰富度及其与地上生产力关系的影响
1
2010
... 样地选自禁牧区, 四周采用围栏围起来以防止牛羊进入破坏.试验采取完全区组设计.在样地内选取48个2 m × 2 m 的样方, 各样方之间留2 m间隔, 每个样方四角用PVC管标记.处理前进行土壤采样, 在每个样方内随机选取3个采样点, 使用5 cm孔径的土钻分别钻取0-20、20-40、40-60 cm深度的土样, 装入自封袋带回实验室, 分析其理化性质, 作为土壤氮矿化的初始值.于2012年5月中旬进行氮、硅添加试验, 均为一次性均匀施肥.根据以往在该地的长期施肥试验资料(
刘慧霞等, 2009 ;
张杰琦等, 2010 )并结合实际情况, 本试验设置4组处理, 分别为对照(CK, 不添加NH
4 NO
3 、H
2 SiO
3 ), 单独氮添加处理(添加NH
4 NO
3 20、40、60 g·m
-2 , 分别记为N20、N40、N60, 折合施氮量分别为7、14、21 g·m
-2 ), 单独硅添加处理(添加H
2 SiO
3 4 g·m
-2 , 即Si4, 折合施硅量1.436 g·m
-2 ), 氮、硅配施处理(在N20、N40、N60基础上, 添加硅酸4 g·m
-2 , 分别记为N20Si4, N40Si4, N60Si4), 共计8个处理, 每处理6个重复.处理前土壤pH值6.33, 有机质含量70.52 g·kg
-1 , 全氮含量3.72 g·kg
-1 , 铵态氮、硝态氮含量分别为10.67 mg·kg
-1 和15.25 mg·kg
-1 ; 全磷含量0.98 g·kg
-1 , 速效磷含量25.27 mg·kg
-1 . ...
氮素添加对青藏高原高寒草甸植物群落物种丰富度及其与地上生产力关系的影响
1
2010
... 样地选自禁牧区, 四周采用围栏围起来以防止牛羊进入破坏.试验采取完全区组设计.在样地内选取48个2 m × 2 m 的样方, 各样方之间留2 m间隔, 每个样方四角用PVC管标记.处理前进行土壤采样, 在每个样方内随机选取3个采样点, 使用5 cm孔径的土钻分别钻取0-20、20-40、40-60 cm深度的土样, 装入自封袋带回实验室, 分析其理化性质, 作为土壤氮矿化的初始值.于2012年5月中旬进行氮、硅添加试验, 均为一次性均匀施肥.根据以往在该地的长期施肥试验资料(
刘慧霞等, 2009 ;
张杰琦等, 2010 )并结合实际情况, 本试验设置4组处理, 分别为对照(CK, 不添加NH
4 NO
3 、H
2 SiO
3 ), 单独氮添加处理(添加NH
4 NO
3 20、40、60 g·m
-2 , 分别记为N20、N40、N60, 折合施氮量分别为7、14、21 g·m
-2 ), 单独硅添加处理(添加H
2 SiO
3 4 g·m
-2 , 即Si4, 折合施硅量1.436 g·m
-2 ), 氮、硅配施处理(在N20、N40、N60基础上, 添加硅酸4 g·m
-2 , 分别记为N20Si4, N40Si4, N60Si4), 共计8个处理, 每处理6个重复.处理前土壤pH值6.33, 有机质含量70.52 g·kg
-1 , 全氮含量3.72 g·kg
-1 , 铵态氮、硝态氮含量分别为10.67 mg·kg
-1 和15.25 mg·kg
-1 ; 全磷含量0.98 g·kg
-1 , 速效磷含量25.27 mg·kg
-1 . ...
氮硅添加对高寒草甸生物量和多样性的影响 ——以青藏高原为例
2
2016
... 青藏高原平均海拔在4 000 m以上, 有“世界屋脊”之称, 分布有世界海拔最高的草地.作为青藏高原自然生态系统的主体(
刘兴元等, 2012 ), 高寒草地面积达1.525 × 10
8 hm
2 , 占青藏高原总面积的59.28% (
张镱锂等, 2013 ).由于海拔高、气候寒冷、干旱等因素, 该地区有机氮的利用率低, 仅占土壤全氮的1% (
白洁冰等, 2011 ).近年来, 随着青藏高原东缘区域性氮沉降逐年增加, 该地区土壤中的可用氮增加(
杨晓霞等, 2014 ), 继而对土壤中氮的活动产生影响.添加营养元素是维持草地生态系统养分平衡, 减缓草地退化的重要措施, 添加氮能有效提高土壤有效氮含量, 促进草地生态系统养分循环(
刘晓东等, 2015 ).硅对植物生长同样至关重要, 不仅可增加植物抗逆、抗倒伏能力, 还可活化土壤有机质, 有效促进植物对氮、磷等的吸收(
张文鹏等, 2016 ).但之前的研究多集中于硅对植物群落多样性、植物抗性等方面的影响, 而对土壤氮矿化的影响研究较少.本研究将土壤矿化作为重点, 探讨氮硅添加对土壤矿化的影响, 以期为维持青藏高原草地生态系统的平衡和制定适当管理措施提供有效的理论依据. ...
... 硅虽然是地壳中仅次于氧的第二大元素, 但土壤中的大部分硅通常会结合其他物质, 形成氧化性硅酸, 植物无法直接吸收利用, 但硅在植物的生长过程中发挥着不可或缺的作用, 不仅可以活化土壤有机质, 促进植物根对氮磷等矿质元素的吸收, 增加植物抗性(
宾振钧等, 2015 ;
张文鹏等, 2016 ), 提高植物叶片的光合作用和植物群落的物种多样性(
Xu et al ., 2015 ,
2018 ), 还可以缓解环境中污染物, 如可抑制植物光合作用的镉对植物的损害, 且硅对植物的解毒作用因物种、品种及生态型的不同而表现出不同的解毒机制(
刘彩凤等, 2017 ).本研究结果表明, 氮硅配施使土壤中的有效氮含量显著增加, 且显著高于单独施氮时的含量, 土壤矿化活动也较单独施氮时活跃, 即土壤净硝化速率、净氨化速率和净矿化速率均高于单独施氮时.说明施加硅肥能促进土壤矿化行为, 硅元素对高等植物的矿质营养有重要影响, 该结果与
Fu等(2001) 对水稻的研究结果一致. ...
氮硅添加对高寒草甸生物量和多样性的影响 ——以青藏高原为例
2
2016
... 青藏高原平均海拔在4 000 m以上, 有“世界屋脊”之称, 分布有世界海拔最高的草地.作为青藏高原自然生态系统的主体(
刘兴元等, 2012 ), 高寒草地面积达1.525 × 10
8 hm
2 , 占青藏高原总面积的59.28% (
张镱锂等, 2013 ).由于海拔高、气候寒冷、干旱等因素, 该地区有机氮的利用率低, 仅占土壤全氮的1% (
白洁冰等, 2011 ).近年来, 随着青藏高原东缘区域性氮沉降逐年增加, 该地区土壤中的可用氮增加(
杨晓霞等, 2014 ), 继而对土壤中氮的活动产生影响.添加营养元素是维持草地生态系统养分平衡, 减缓草地退化的重要措施, 添加氮能有效提高土壤有效氮含量, 促进草地生态系统养分循环(
刘晓东等, 2015 ).硅对植物生长同样至关重要, 不仅可增加植物抗逆、抗倒伏能力, 还可活化土壤有机质, 有效促进植物对氮、磷等的吸收(
张文鹏等, 2016 ).但之前的研究多集中于硅对植物群落多样性、植物抗性等方面的影响, 而对土壤氮矿化的影响研究较少.本研究将土壤矿化作为重点, 探讨氮硅添加对土壤矿化的影响, 以期为维持青藏高原草地生态系统的平衡和制定适当管理措施提供有效的理论依据. ...
... 硅虽然是地壳中仅次于氧的第二大元素, 但土壤中的大部分硅通常会结合其他物质, 形成氧化性硅酸, 植物无法直接吸收利用, 但硅在植物的生长过程中发挥着不可或缺的作用, 不仅可以活化土壤有机质, 促进植物根对氮磷等矿质元素的吸收, 增加植物抗性(
宾振钧等, 2015 ;
张文鹏等, 2016 ), 提高植物叶片的光合作用和植物群落的物种多样性(
Xu et al ., 2015 ,
2018 ), 还可以缓解环境中污染物, 如可抑制植物光合作用的镉对植物的损害, 且硅对植物的解毒作用因物种、品种及生态型的不同而表现出不同的解毒机制(
刘彩凤等, 2017 ).本研究结果表明, 氮硅配施使土壤中的有效氮含量显著增加, 且显著高于单独施氮时的含量, 土壤矿化活动也较单独施氮时活跃, 即土壤净硝化速率、净氨化速率和净矿化速率均高于单独施氮时.说明施加硅肥能促进土壤矿化行为, 硅元素对高等植物的矿质营养有重要影响, 该结果与
Fu等(2001) 对水稻的研究结果一致. ...
青藏高原高寒草地净初级生产力(NPP)时空分异
1
2013
... 青藏高原平均海拔在4 000 m以上, 有“世界屋脊”之称, 分布有世界海拔最高的草地.作为青藏高原自然生态系统的主体(
刘兴元等, 2012 ), 高寒草地面积达1.525 × 10
8 hm
2 , 占青藏高原总面积的59.28% (
张镱锂等, 2013 ).由于海拔高、气候寒冷、干旱等因素, 该地区有机氮的利用率低, 仅占土壤全氮的1% (
白洁冰等, 2011 ).近年来, 随着青藏高原东缘区域性氮沉降逐年增加, 该地区土壤中的可用氮增加(
杨晓霞等, 2014 ), 继而对土壤中氮的活动产生影响.添加营养元素是维持草地生态系统养分平衡, 减缓草地退化的重要措施, 添加氮能有效提高土壤有效氮含量, 促进草地生态系统养分循环(
刘晓东等, 2015 ).硅对植物生长同样至关重要, 不仅可增加植物抗逆、抗倒伏能力, 还可活化土壤有机质, 有效促进植物对氮、磷等的吸收(
张文鹏等, 2016 ).但之前的研究多集中于硅对植物群落多样性、植物抗性等方面的影响, 而对土壤氮矿化的影响研究较少.本研究将土壤矿化作为重点, 探讨氮硅添加对土壤矿化的影响, 以期为维持青藏高原草地生态系统的平衡和制定适当管理措施提供有效的理论依据. ...
青藏高原高寒草地净初级生产力(NPP)时空分异
1
2013
... 青藏高原平均海拔在4 000 m以上, 有“世界屋脊”之称, 分布有世界海拔最高的草地.作为青藏高原自然生态系统的主体(
刘兴元等, 2012 ), 高寒草地面积达1.525 × 10
8 hm
2 , 占青藏高原总面积的59.28% (
张镱锂等, 2013 ).由于海拔高、气候寒冷、干旱等因素, 该地区有机氮的利用率低, 仅占土壤全氮的1% (
白洁冰等, 2011 ).近年来, 随着青藏高原东缘区域性氮沉降逐年增加, 该地区土壤中的可用氮增加(
杨晓霞等, 2014 ), 继而对土壤中氮的活动产生影响.添加营养元素是维持草地生态系统养分平衡, 减缓草地退化的重要措施, 添加氮能有效提高土壤有效氮含量, 促进草地生态系统养分循环(
刘晓东等, 2015 ).硅对植物生长同样至关重要, 不仅可增加植物抗逆、抗倒伏能力, 还可活化土壤有机质, 有效促进植物对氮、磷等的吸收(
张文鹏等, 2016 ).但之前的研究多集中于硅对植物群落多样性、植物抗性等方面的影响, 而对土壤氮矿化的影响研究较少.本研究将土壤矿化作为重点, 探讨氮硅添加对土壤矿化的影响, 以期为维持青藏高原草地生态系统的平衡和制定适当管理措施提供有效的理论依据. ...
氮磷添加对亚热带常绿阔叶林土壤氮素矿化的影响
1
2013
... 氮矿化的研究对于揭示生态系统功能, 为维持生态系统稳定提供理论指导有重要意义.氮是植物生长发育必需的营养元素, 是植物吸收量最大的土壤矿质元素(
赵阳等, 2013 ), 也是影响整个陆地系统生产力的重要因子, 通常被认为是植物生长发育重要限制因素(
司晓林等, 2016 ).氮在土壤中绝大部分是以有机形式存在的, 约占土壤全氮含量的92%-98%, 土壤有机氮无法直接被植物吸收利用, 必须借助土壤微生物, 通过矿化作用将其转化为无机氮.土壤中无机氮主要形态有铵态氮和硝态氮, 二者都具水溶性, 更易被植物吸收利用.自18世纪后期, 土壤氮矿化已被视为土壤氮循环的核心, 在控制植物有效氮利用方面起到至关重要的作用(
Aber & Melillo, 2001 ). ...
氮磷添加对亚热带常绿阔叶林土壤氮素矿化的影响
1
2013
... 氮矿化的研究对于揭示生态系统功能, 为维持生态系统稳定提供理论指导有重要意义.氮是植物生长发育必需的营养元素, 是植物吸收量最大的土壤矿质元素(
赵阳等, 2013 ), 也是影响整个陆地系统生产力的重要因子, 通常被认为是植物生长发育重要限制因素(
司晓林等, 2016 ).氮在土壤中绝大部分是以有机形式存在的, 约占土壤全氮含量的92%-98%, 土壤有机氮无法直接被植物吸收利用, 必须借助土壤微生物, 通过矿化作用将其转化为无机氮.土壤中无机氮主要形态有铵态氮和硝态氮, 二者都具水溶性, 更易被植物吸收利用.自18世纪后期, 土壤氮矿化已被视为土壤氮循环的核心, 在控制植物有效氮利用方面起到至关重要的作用(
Aber & Melillo, 2001 ). ...
 ,1,*
,1,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT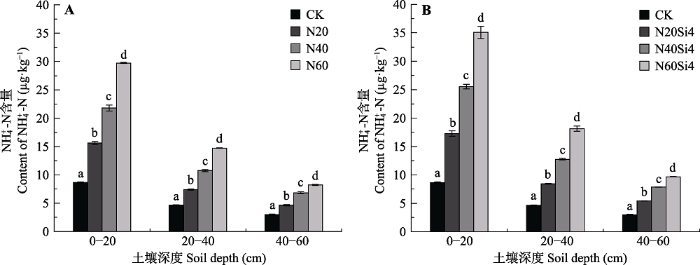 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT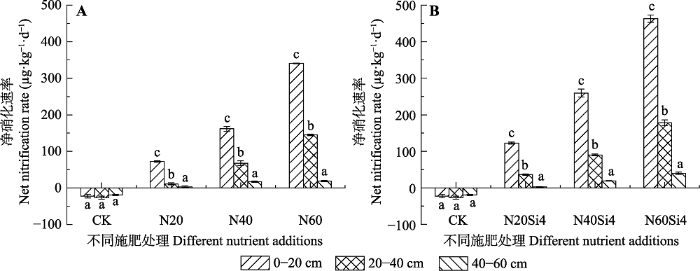 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT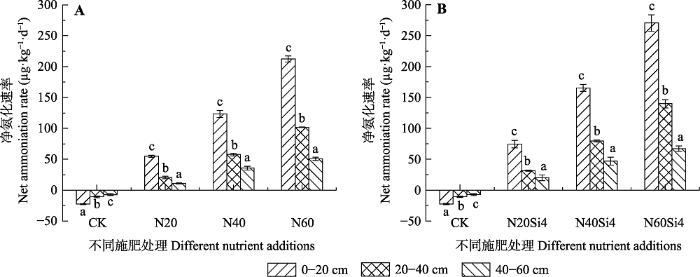 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT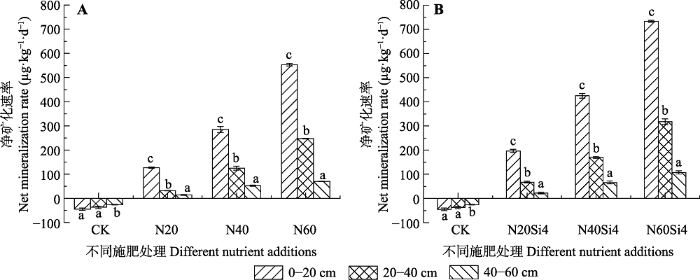 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}