Spatio-temporal dynamics of two alpine treeline ecotones and ecological characteristics of their dominate species at the eastern margin of Qinghai-Xizang Plateau
ZHOU Tian-Yang1,2,3, NARAYAN Prasad Gaire4,8, LIAO Li-Bin1,2, ZHENG Li-Li2,5, WANG Jin-Niu,1,3,6,*, SUN Jian5, WEI Yan-Qiang7, XIE Yu1, WU Yan,1,3,*1Chengdu Institute of Biology, Chinese Academy of Sciences, Chengdu 610041, China 2University of Chinese Academy of Sciences, Beijing 101408, China 3Key Laboratory of Mountain Ecological Restoration and Bioresource Utilization, Chinese Academy of Sciences, Chengdu 610041, China 4Central Department of Environmental Science, Tribhuvan University, Kathmandu, Nepal 5Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101, China 6International Center for Integrated Mountain Development, Kathmandu, Nepal 7Northwest Institute of Eco-Environment and Resources, Chinese Academy of Sciences, Lanzhou 730000, China 8Xishuangbanna Tropical Botanical garden, Chinese Academy of Sciences, Xishuangbanna, Yunnan 666303, China
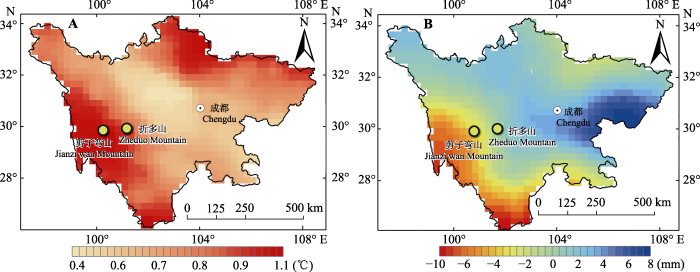
Abstract Aims Temperature limit is the main cause of alpine treeline formation. Therefore, it is important to understand the response mechanisms of alpine treeline as well as their tree species under the global climate change. The present study focused on the spatio-temporal dynamics of treeline and ecological characteristics of the tree species in two treeline ecotones. Methods Two vertical belt-transect plots were established in each treeline ecotone of the Zheduo Mountain and Jianziwan Mountain of the eastern Qinghai-Xizang Plateau. Top and bottom of each transects were lain between species line and forest line, respectively. Detailed information of each tree species treeline, including species name, latitude, longitude, height, age, base diameter, and coordinates, was recorded accordingly. Important findings The temperatures of the two research areas have increased during the past 58 years. The precipitation has decreased slightly in both the Zheduo Mountain and Jianziwan Mountain. The age structure of Abies fabri from the Zheduo Mountain and A. squamata from the Jianziwan Mountain showed a reversed “J” shape curve and a bimodal shape, respectively. Within the two transects, due to the limitation of seed diffusion, the dominate species showed aggregated distributions at the small scale. At the large scale, A. fabri was aggregated at the Zheduo Mountain, while A. squanmata of the Jianziwan Mountain was randomly distributed due to the impact of surrounding environmental factors. Both tree height and base diameter decreased with the increase of elevation. The fir trees (Abies spp.) at the upper part of the treeline ecotone presented an allometric growth, whose height growth rate was higher than that of base growth, while the relationships between height growth and base growth were isometric at almost mid and lower part of the treeline ecotone. Compared with 10 years ago, there was no significant change at the position of treeline and tree species line of the Zheduo Mountain and the Jianziwan Mountain, neither of the tree density in the Jianziwan Mountain. However, the number of trees in the Zheduo Mountain increased by about 25%. Compared with 20 years ago, tree species lines of the Zheduo Mountain and Jianziwan Mountain were shifted upwards by 50 and 30 m, respectively. Besides, their treeline positions were increased by 75 and 40 m, respectively. Furthermore, the number of trees also increased significantly by 220% and 100%, respectively. Therefore, the treeline and its constructive species are mainly affected by temperature at the large spatio-temporal scale, while influenced by temperature and ambient environment at the small spatio-temporal scale. Keywords:treeline;spatio-temporal dynamics;age structure;allometric;altitude gradient
PDF (3410KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 周天阳, NARAYAN Prasad Gaire, 廖礼彬, 郑莉莉, 王金牛, 孙建, 魏彦强, 谢雨, 吴彦. 青藏高原东缘两处高山树线交错带时空动态及其建群种的生态学特征. 植物生态学报[J], 2018, 42(11): 1082-1093 doi:10.17521/cjpe.2018.0082 ZHOU Tian-Yang, NARAYAN Prasad Gaire, LIAO Li-Bin, ZHENG Li-Li, WANG Jin-Niu, SUN Jian, WEI Yan-Qiang, XIE Yu, WU Yan. Spatio-temporal dynamics of two alpine treeline ecotones and ecological characteristics of their dominate species at the eastern margin of Qinghai-Xizang Plateau. Chinese Journal of Plant Ecology[J], 2018, 42(11): 1082-1093 doi:10.17521/cjpe.2018.0082
Fig. 4Point pattern analyses for the Zheduo Mountain (A) and Jianziwan Mountain (B). The two green lines are the confidence intervals for the fitting, and the solid line is from the simulation.
利用树高与树龄间的关系对样地树线分布进行模拟分析(10年前及20年前), 具体树高-树龄模型见表3。建立的两个样地的树高-树龄模型均较好(p < 0.001, R2分别为0.68和0.71), 能较准确地模拟前10至20年的情况。模拟结果见图5, 由模拟分布图可以看出, 目前折多山样地的树线海拔约为4 275 m, 树种线海拔约为4 380 m, 样地最下端为郁闭森林; 与10年前相比, 目前该样地的树线及树种线位置无明显变化, 而树木数量提高了约25%; 20年前树线及树种线的海拔分别约为4 200 m和4 330 m, 与目前相比, 二者分别提高了75 m和50 m, 而树木个体数(54株)仅为目前的1/3。剪子弯山样地目前的树种线位于海拔4 520 m处, 树线位于海拔约4 440 m处; 10年前该样地的树木个体数、树线及树种线位置与目前基本保持一致; 20年前树木个体数(47株)仅为目前的一半, 树种线位于4 490 m处, 而树线位于样地下方具体海拔无法确定, 相比当前样地内树种线上升了30 m, 树线位置至少爬升了40 m。
Table 3 表3 表3折多山与剪子弯山树高(H)与树龄(a)的回归模型 Table 3The regression model of tree height (H) and tree age (a)
Fig. 5The spatio-temporal pattern of treeline dynamics in the Zheduo Mountain (A) and Jianziwan Mountain (B). The solid line is the position of treeline and the dash line is tree species line.
LuckmanB, KavanaghT ( 2000). Impact of climate fluctuations on mountain environments in the Canadian Rockies AMBIO, 29, 371-380. DOI:10.1579/0044-7447-29.7.371URL This paper presents examples of environmental changes in the Canadian Rockies in the context of a 1.5 C increase in mean annual temperatures over the last 100 years. During this period increases in winter temperatures have been more than twice as large as those during spring and summer. Glacier cover has decreased by at least 25% during the present century and glacier fronts have receded to positions last occupied ca. 3000 years ago. These two lines of evidence suggest that the climate of the late 20th century is exceptional in the context of the last 1000 to 3000 years. Detailed studies in three closely located upper treeline sites document variable responses of vegetation to climate change that reflect species differences as well as local differences in microclimate and site conditions. Treeline has advanced upslope in response to climate warming, but site and species differences control the rate and nature of the advance. Human impacts on the environment compound the changes due to climate warming. Historic photographs indicate significant changes in the type and density of forest cover due to the absence of significant forest fires within these National Parks during the last 70-80 years. The visual impact of these changes, which partially reflects a policy of fire suppression, is far greater than the impact of changes associated with more direct tourist-related impacts. It is therefore important that monitoring programs examine vegetation changes over the entire landscape rather than focussing exclusively on supposedly climate-sensitive sites.
MacDonaldGM, SzeiczJM, ClaricoatesJ, DaleKA ( 1998). Response of the central Canadian treeline to recent climatic changes Annals of the Association of American Geographers, 88, 183-208. DOI:10.1111/1467-8306.00090URL [本文引用: 2]
MayrS, Charra-VaskouK ( 2007). Winter at the alpine timberline causes complex within-tree patterns of water potential and embolism in Picea abies Physiologia Plantarum, 131, 131-139. DOI:10.1111/ppl.2007.131.issue-1URL [本文引用: 1]
NiklasKJ ( 2005). Modelling below- and above-ground biomass for non-woody and woody plants Annals of Botany, 95, 315-321. DOI:10.1093/aob/mci028URLPMID:15546927 [本文引用: 1] BACKGROUND AND AIMS: Intraspecific relationships between below- and above-ground biomass (MB and MA, respectively) have been studied extensively to evaluate environmental effects on growth and development at the level of the individual plant. However, no current theoretical model for this relationship exists for broad interspecific trends. The aims of this paper are to provide a model and to test its predictions using a recently assembled, large database (1406 data entries for 257 species). METHODS: An allometric model was derived to predict the relationship between MB and MA for non-woody and woody plants based on previously developed scaling relationships for leaf, stem and root standing biomass and annual growth rates. The predictions of this model were tested by comparing the numerical values of predicted scaling exponents (the slopes of log-log regression curves) with those observed for the database. KEY RESULTS AND CONCLUSIONS: For non-woody plants and the juveniles of woody species, the model predicts an isometric scaling relationship (i.e. MB proportional, variant MA). For woody plants, a complex scaling function is predicted. But, for a particular set of biologically reasonable conditions, the model predicts MB proportional, variant MA across woody plants. These predictions accord reasonably well with observed statistical trends when non-woody and woody plants are studied separately (n=1061 and 345 data entries, respectively). Although the reliability of regression formulas to estimate MB based on MA measurements increased with increasing plant size, estimates of MB can be as much as two orders of magnitude off, even when using regression formulas with r2 >>0.90 and F >>53,000.
PelissierR, GoreaudF ( 2015). Ads package for R: A fast unbiased implementation of the k-function family for studying spatial point patterns in irregular-shaped sampling windows Journal of Statistical Software, 63, 1-18. [本文引用: 1]
RanF, LiangYM, YangY, YangY, WangGX ( 2014). Spatial-temporal dynamics of an Abies fabri population near the alpine treeline in the Yajiageng area of Gongga Mountain, China Acta Ecologica Sinica, 34, 6872-6878. DOI:10.5846/stxb201311252809URL [本文引用: 1] 通过对贡嘎山雅家埂峨眉冷杉种群林线附近6个3000 m2样地(阴阳坡各3个)中峨眉冷杉(Abies fabri Craib)种群的定位调查,分析了过去100a间该区峨眉冷杉种群的时间-空间动态。结果表明:1)雅家埂林线附近峨眉冷杉种群密度在过去100 a(主要是近50 a)有显著的升高,但树线的海拔位置并无明显的爬升;2)阴阳坡林线格局存在显著的坡向分异:阴坡林线和树线的海拔高度显著高于阳坡(分别比阳坡高152.5 m和135.8 m),阳坡林线附近峨眉冷杉早期的生长速率在大于阴坡,但后期的生长速率却低于阴坡;3)热量(温度)控制假说不能完全解释雅家埂目前的树线格局,除气候因素之外,其它因素也限制了雅家梗地区树线位置的变化。 [ 冉飞, 梁一鸣, 杨燕, 杨阳, 王根绪 ( 2014). 贡嘎山雅家埂峨眉冷杉林线种群的时空动态 生态学报, 34, 6872-6878.] DOI:10.5846/stxb201311252809URL [本文引用: 1] 通过对贡嘎山雅家埂峨眉冷杉种群林线附近6个3000 m2样地(阴阳坡各3个)中峨眉冷杉(Abies fabri Craib)种群的定位调查,分析了过去100a间该区峨眉冷杉种群的时间-空间动态。结果表明:1)雅家埂林线附近峨眉冷杉种群密度在过去100 a(主要是近50 a)有显著的升高,但树线的海拔位置并无明显的爬升;2)阴阳坡林线格局存在显著的坡向分异:阴坡林线和树线的海拔高度显著高于阳坡(分别比阳坡高152.5 m和135.8 m),阳坡林线附近峨眉冷杉早期的生长速率在大于阴坡,但后期的生长速率却低于阴坡;3)热量(温度)控制假说不能完全解释雅家埂目前的树线格局,除气候因素之外,其它因素也限制了雅家梗地区树线位置的变化。
RipleyBD ( 1977). Modeling spatial patterns Journal of the Royal Statistical Society Series B—Methodological, 39, 172-212. DOI:10.1111/rssb.1977.39.issue-2URL [本文引用: 1]
ShiCM, Masson-DelmotteV, DauxV, LiZS, CarreM, MooreJC ( 2015). Unprecedented recent warming rate and temperature variability over the east Tibetan Plateau inferred from alpine treeline dendrochronology Climate Dynamics, 45, 1367-1380. DOI:10.1007/s00382-014-2386-zURL [本文引用: 1] Despite instrumental records showing recent large temperature rises on the Tibetan Plateau (TP), only a few tree-ring temperature reconstructions do capture this warming trend. Here, we sampled 260 trees from seven Alpine treeline locations across the southeast TP. Standardized tree-ring width chronologies of Abies squamata and Sabina squamat were produced following Regional Curve Standardization detrending. The leading principal component of these records is well correlated with the regional summer (JJA) minimum temperature (MinT) (R 2 02=020.47, P 02<020.001, 1953–2009). Hence we produce a regional summer MinT reconstruction spanning the last 21202years. This reconstruction reveals a long-term persistent warming trend, starting in the 1820s, at a rate of 0.4502±020.0902°C/century (1820–2009). This trend is also detected since the 1820s in the Asian summer MinT reconstruction produced by the PAGES 2K project, with a very close warming rate (0.4302±020.0802°C/century, 1820–1989). Our record also displays an enhanced multi-decadal variability since the mid-twentieth century. The 1990s–2000s are the warmest of our whole record, due to the superposition of the gradual warming trend and decadal variability during this interval. The strongest decadal cooling occurs during the 1950s and the largest warming trend during the 1970s. The magnitude of warming from 1973 to 2003 was larger than the total warming trend from 1820s to 2009. Extreme events are also more frequent since 1950. The pattern of multi-decadal variability has similarities with the Atlantic multi-decadal oscillation, suggesting common causality. CMIP5 historical simulations fail to capture both the magnitude and timing of this multi-decadal variability. The ensemble CMIP5 average produces a steady warming trend starting in the 1970s, which only accounts for about 6002% of the observed warming trend during this period. We conclude that TP summer temperature could reflect a climate response to increased greenhouse gas concentrations, however modulated by multi-decadal variations common with the Atlantic sector.
ShiJY, HanHR, ChengXQ, DongLL, TianHX, CaiMK, KangFF ( 2017). Age structure and dynamics of Pinus tabuliformis population in the Liaoheyuan Nature Reserve of Hebei Province Chinese Journal of Ecology, 36, 1808-1814. DOI:10.13292/j.1000-4890.201707.019URL [本文引用: 1] 本文基于种群生命表和生存分析理论,应用“空间代替时间”的方法和匀滑技术,编制了辽河源自然保护区油松(Pinus tabuliformis)种群的静态生命表,并绘制了其死亡率曲线、消失率曲线、存活曲线以及生存函数曲线.结果表明:油松种群呈波动式的发展,经历3次死亡高峰;种群年龄结构呈倒“J”型,幼苗数量较多,但从幼苗发展到幼树(第1龄级→第2龄级)阶段需通过环境筛的高强度选择,经历第1次死亡高峰;第2次和第3次死亡高峰分别发生在油松由次林层向主林层过渡时期(第7龄级→第8龄级)和生长后期(第11龄级→第12龄级),分别是竞争导致的自疏作用和步入生理死亡年龄的结果;种群的存活曲线趋近于Deevy-Ⅱ型,为稳定型种群;4个生存函数表明,种群动态具有前期减少,中期稳定,后期衰退的特点. [ 矢佳昱, 韩海荣, 程小琴, 董玲玲, 田慧霞, 蔡锰柯, 康峰峰 ( 2017). 河北辽河源自然保护区油松种群年龄结构和种群动态 生态学杂志, 36, 1808-1814.] DOI:10.13292/j.1000-4890.201707.019URL [本文引用: 1] 本文基于种群生命表和生存分析理论,应用“空间代替时间”的方法和匀滑技术,编制了辽河源自然保护区油松(Pinus tabuliformis)种群的静态生命表,并绘制了其死亡率曲线、消失率曲线、存活曲线以及生存函数曲线.结果表明:油松种群呈波动式的发展,经历3次死亡高峰;种群年龄结构呈倒“J”型,幼苗数量较多,但从幼苗发展到幼树(第1龄级→第2龄级)阶段需通过环境筛的高强度选择,经历第1次死亡高峰;第2次和第3次死亡高峰分别发生在油松由次林层向主林层过渡时期(第7龄级→第8龄级)和生长后期(第11龄级→第12龄级),分别是竞争导致的自疏作用和步入生理死亡年龄的结果;种群的存活曲线趋近于Deevy-Ⅱ型,为稳定型种群;4个生存函数表明,种群动态具有前期减少,中期稳定,后期衰退的特点.
StewartGH, RoseAB ( 1990). The significance of life-history strategies in the developmental history of mixed beech (nothofagus) forests, New-Zealand Vegetatio, 87, 101-114. DOI:10.1007/bf00042947URL [本文引用: 1] Size and age structure analysis, dated past disturbances, treefall replacement patterns, and spatial pattern analysis were used to reconstruct the developmental history of two old-growth Nothofagus fusca/N. menziesii stands, South Island, New Zealand. Diameter and height class distributions suggested that N. menziesii was replacing N. fusca, howeer, stand history reconstruction analysis showed that both species had regenerated intermittently after small-scale disturbances. Although large-scale disturbances such as blowdowns may occasionally generate even-aged stands, gap-phase regeneration maintains the forests in compositional equilibrium. In the absence of other competing tree species and understorey plants the two species appear to coexist by way of different life history strategies, where one species (N. menziesii) has low juvenile mortality and the other (N. fusca) has faster height growth rates and greater longevity and adult survivorship.
van BogaertR, HanecaK, HoogestegerJ, JonassonC, de DapperM, CallaghanTV ( 2011). A century of tree line changes in sub-Arctic Sweden shows local and regional variability and only a minor influence of 20th Century climate warming Journal of Biogeography, 38, 907-921. DOI:10.1111/j.1365-2699.2010.02453.xURL [本文引用: 1] Aim Models project that climate warming will cause the tree line to move to higher elevations in alpine areas and more northerly latitudes in Arctic environments. We aimed to document changes or stability of the tree line in a sub-Arctic model area at different temporal and spatial scales, and particularly to clarify the ambiguity that currently exists about tree line dynamics and their causes.Location The study was conducted in the Tornetrsk area in northern Sweden where climate warmed by 2.5 C between 1913 and 2006. Mountain birch (Betula pubescens ssp. czerepanovii) sets the alpine tree line.Methods We used repeat photography, dendrochronological analysis, field observations along elevational transects and historical documents to study tree line dynamics.Results Since 1912, only four out of eight tree line sites had advanced: on average the tree line had shifted 24 m upslope (+0.2 m year 1 assuming linear shifts). Maximum tree line advance was +145 m (+1.5 m year 1 in elevation and +2.7 m year 1 in actual distance), whereas maximum retreat was 120 m downslope. Counter-intuitively, tree line advance was most pronounced during the cooler late 1960s and 1970s. Tree establishment and tree line advance were significantly correlated with periods of low reindeer (Rangifer tarandus) population numbers. A decreased anthropozoogenic impact since the early 20th century was found to be the main factor shaping the current tree line ecotone and its dynamics. In addition, episodic disturbances by moth outbreaks and geomorphological processes resulted in descent and long-term stability of the tree line position, respectively.Main conclusions In contrast to what is generally stated in the literature, this study shows that in a period of climate warming, disturbance may not only determine when tree line advance will occur but if tree line advance will occur at all. In the case of non-climatic climax tree lines, such as those in our study area, both climate-driven model projections of future tree line positions and the use of the tree line position for bioclimatic monitoring should be used with caution.
WangT, ZhangQB, MaKP ( 2006). Treeline dynamics in relation to climatic variability in the central Tianshan Mountains, northwestern China Global Ecology and Biogeography, 15, 406-415. DOI:10.1111/j.1466-822X.2006.00233.xURL Aim Climate variability may be an important mediating agent of ecosystem dynamics in cold, arid regions such as the central Tianshan Mountains, north-western China. Tree-ring chronologies and the age structure of a Schrenk spruce ( Picea schrenkiana ) forest were developed to examine treeline dynamics in recent decades in relation to climatic variability. Of particular interest was whether tree-ring growth and population recruitment patterns responded similarly to climate warming. Location The study was conducted in eight stands that ranged from 2500 m to 2750 m a.s.l. near the treeline in the Tianchi Nature Reserve (43°45'6143°59' N, 88°00'6188°20' E) in the central Xinjiang Uygur Autonomous Region, northwestern China. Methods Tree-ring cores were collected and used to develop tree-ring chronologies. The age of sampled trees was determined from basal cores sampled as close as possible to the ground. Population age structure and recruitment information were obtained using an age–d.b.h. (diameter at breast height) regression from the sampled cores and the d.b.h. measured on all trees in the plots. Ring-width chronologies and tree age structure were both used to investigate the relationship between treeline dynamics and climate change. Results Comparisons with the climatic records showed that both the radial growth of trees and tree recruitment were influenced positively by temperature and precipitation in the cold high treeline zone, but the patterns of their responses differed. The annual variation in tree rings could be explained largely by the average monthly minimum temperatures during February and August of the current year and by the monthly precipitation of the previous August and January, which had a significant and positive effect on tree radial growth. P. schrenkiana recruitment was influenced mainly by consecutive years of high minimum summer temperatures and high precipitation during spring. Over the last several decades, the treeline did not show an obvious upward shift and new recruitment was rare. Some trees had established at the treeline at least 200 years ago. Recruitment increased until the early 20th century (1910s) but then decreased with poor recruitment over the past several decades (1950–2000). Main conclusions There were strong associations between climatic change and ring-width patterns, and with recruitments in Schrenk spruce. Average minimum temperatures in February and August, and total precipitation in the previous August and January, had a positive effect on tree-ring width, and several consecutive years of high minimum summer temperature and spring precipitation was a main factor favouring the establishment of P. schrenkiana following germination within the treeline ecotone. Both dendroclimatology and recruitment analysis were useful and compatible to understand and reconstruct treeline dynamics in the central Tianshan Mountains.
WangYF, Julio CamareroJ, LuoTX, LiangEY ( 2012). Spatial patterns of smith fir alpine treelines on the south-eastern Tibetan Plateau support that contingent local conditions drive recent treeline patterns Plant Ecology & Diversity, 5, 311-321. DOI:10.1080/17550874.2012.704647URL [本文引用: 2] Background: Recent work has shown little change in the position of the Smith fir treeline on the south-eastern Tibetan Plateau in response to global warming. However, the relationship between tree distribution patterns within the treeline ecotone and low responsiveness is unknown, and additional constraints than climate might be major drivers of these patterns (e.g. microsite availability for regeneration).
WangYF, LiangEY ( 2012). A review on progresses in treeline dynamics and climate change Journal of Earth Environment, 3, 855-861. URL [本文引用: 2] 作为直立树木分布的高海拔上限和高纬度北界,树线对气候变化具有潜在的敏感性,是全球变化研究的热点内容之一。树线变化不仅包含树线位置(本文中指树线上边界位置)的进退,还涉及到林分结构、空间格局和树木的生长变化等方面。尽管****们利用不同的假说来解释树线和林线形成机理,但普遍认为温度是影响树线变化的最重要的环境因子。因此,全球变暖背景下,树线位置和种群密度变化趋势的研究备受关注。然而,树线位置的变化对气候变化会有几十年至几百年的的滞后性,在全球变暖的背景下树线位置有可能仅发生微小的变化。相对于树线位置,森林种群密度的变化是更敏感的生态指标。国际上多处树线研究(包括藏东南一带)都揭示了近100~200年来树线过渡带内种群密度的显著增加。但是,国际上已经报道的大多数树线样点不同程度地受到人为活动的干扰,难以区分气候变化和人为干扰对树线波动的影响。值得提出的是,青藏高原具有全球最高海拔的天然树线,且受人类活动干扰少,是研究气候变化对树线动态与格局影响的理想地点之一。 [ 王亚锋, 梁尔源 ( 2012). 树线波动与气候变化研究进展 地球环境学报, 3, 855-861.] URL [本文引用: 2] 作为直立树木分布的高海拔上限和高纬度北界,树线对气候变化具有潜在的敏感性,是全球变化研究的热点内容之一。树线变化不仅包含树线位置(本文中指树线上边界位置)的进退,还涉及到林分结构、空间格局和树木的生长变化等方面。尽管****们利用不同的假说来解释树线和林线形成机理,但普遍认为温度是影响树线变化的最重要的环境因子。因此,全球变暖背景下,树线位置和种群密度变化趋势的研究备受关注。然而,树线位置的变化对气候变化会有几十年至几百年的的滞后性,在全球变暖的背景下树线位置有可能仅发生微小的变化。相对于树线位置,森林种群密度的变化是更敏感的生态指标。国际上多处树线研究(包括藏东南一带)都揭示了近100~200年来树线过渡带内种群密度的显著增加。但是,国际上已经报道的大多数树线样点不同程度地受到人为活动的干扰,难以区分气候变化和人为干扰对树线波动的影响。值得提出的是,青藏高原具有全球最高海拔的天然树线,且受人类活动干扰少,是研究气候变化对树线动态与格局影响的理想地点之一。
WangYF, LiangEY, LuXM, ZhuHF, PiaoSL, ZhuLP ( 2017). Are treelines advancing in response to climate warming on the Tibetan Plateau? Chinese Journal of Nature, 39, 179-183. DOI:10.3969/j.issn.0253-9608.2017.03.004URL [本文引用: 1] 树线上树木生长主要受生长季低温限制,因此气候变暖将导致高山树线向更高海拔迁移。然而,全球树线调查数据显示,近百年来约50%调查样点树线上升明显,其余则保持静止状态。以上事实揭示,除了气候因素,非气候因素(如竞争等种间关系)也在一定程度上调控着树线的位置变化。青藏高原拥有全球海拔最高的天然树线,是探讨气候和非气候因素对树线变化影响的理想区域。研究发现:树线之上低矮稀疏的灌丛或草丛成为幼苗荫发或生长的“避难所”,从而有利于树线大幅度爬升:而高大茂密的灌丛则会形成一道宽厚的”封锁墙”,从而抑制树线的上升。因此,变暖背景下树线位置倾向于向更高海拔爬升,但爬升速率受到树线之上种间关系的调控。变暖并不一定会造成树线位置的显著上升。 [ 王亚锋, 梁尔源, 芦晓明, 朱海峰, 朴世龙, 朱立平 ( 2017). 气候变暖会使青藏高原树线一直上升吗? 自然杂志, 39, 179-183.] DOI:10.3969/j.issn.0253-9608.2017.03.004URL [本文引用: 1] 树线上树木生长主要受生长季低温限制,因此气候变暖将导致高山树线向更高海拔迁移。然而,全球树线调查数据显示,近百年来约50%调查样点树线上升明显,其余则保持静止状态。以上事实揭示,除了气候因素,非气候因素(如竞争等种间关系)也在一定程度上调控着树线的位置变化。青藏高原拥有全球海拔最高的天然树线,是探讨气候和非气候因素对树线变化影响的理想区域。研究发现:树线之上低矮稀疏的灌丛或草丛成为幼苗荫发或生长的“避难所”,从而有利于树线大幅度爬升:而高大茂密的灌丛则会形成一道宽厚的”封锁墙”,从而抑制树线的上升。因此,变暖背景下树线位置倾向于向更高海拔爬升,但爬升速率受到树线之上种间关系的调控。变暖并不一定会造成树线位置的显著上升。
WartonDI, DuursmaRA, FalsterDS, TaskinenS ( 2012). Smatr 3—An R package for estimation and inference about allometric lines Methods in Ecology and Evolution, 3, 257-259. DOI:10.1111/j.2041-210X.2011.00153.xURL [本文引用: 1]
WartonDI, WrightIJ, FalsterDS, WestobyM ( 2006). Bivariate line-fitting methods for allometry Biological Reviews, 81, 259-291. DOI:10.1017/S1464793106007007URLPMID:16573844 [本文引用: 1] Fitting a line to a bivariate dataset can be a deceptively complex problem, and there has been much debate on this issue in the literature. In this review, we describe for the practitioner the essential features of line-fitting methods for estimating the relationship between two variables: what methods are commonly used, which method should be used when, and how to make inferences from these lines to answer common research questions. A particularly important point for line-fitting in allometry is that usually, two sources of error are present (which we call measurement and equation error), and these have quite different implications for choice of line-fitting method. As a consequence, the approach in this review and the methods presented have subtle but important differences from previous reviews in the biology literature. Linear regression, major axis and standardised major axis are alternative methods that can be appropriate when there is no measurement error. When there is measurement error, this often needs to be estimated and used to adjust the variance terms in formulae for line-fitting. We also review line-fitting methods for phylogenetic analyses. Methods of inference are described for the line-fitting techniques discussed in this paper. The types of inference considered here are testing if the slope or elevation equals a given value, constructing confidence intervals for the slope or elevation, comparing several slopes or elevations, and testing for shift along the axis amongst several groups. In some cases several methods have been proposed in the literature. These are discussed and compared. In other cases there is little or no previous guidance available in the literature. Simulations were conducted to check whether the methods of inference proposed have the intended coverage probability or Type I error. We identified the methods of inference that perform well and recommend the techniques that should be adopted in future work.
WilmkingM, JudayGP, BarberVA, ZaldHSJ ( 2004). Recent climate warming forces contrasting growth responses of white spruce at treeline in Alaska through temperature thresholds Global Change Biology, 10, 1724-1736. DOI:10.1111/j.1365-2486.2004.00826.xURL [本文引用: 1] Northern and high-latitude alpine treelines are generally thought to be limited by available warmth. Most studies of tree-growth limate interaction at treeline as well as climate reconstructions using dendrochronology report positive growth response of treeline trees to warmer temperatures. However, population-wide responses of treeline trees to climate remain largely unexamined. We systematically sampled 1558 white spruce at 13 treeline sites in the Brooks Range and Alaska Range. Our findings of both positive and negative growth responses to climate warming at treeline challenge the widespread assumption that arctic treeline trees grow better with warming climate. High mean temperatures in July decreased the growth of 40% of white spruce at treeline areas in Alaska, whereas warm springs enhance growth of additional 36% of trees and 24% show no significant correlation with climate. Even though these opposing growth responses are present in all sampled sites, their relative proportion varies between sites and there is no overall clear relationship between growth response and landscape position within a site. Growth increases and decreases appear in our sample above specific temperature index values (temperature thresholds), which occurred more frequently in the late 20th century. Contrary to previous findings, temperature explained more variability in radial growth after 1950. Without accounting for these opposite responses and temperature thresholds, climate reconstructions based on ring width will miscalibrate past climate, and biogeochemical and dynamic vegetation models will overestimate carbon uptake and treeline advance under future warming scenarios.
WongMH, DuanCQ, LongYC, LuoYM, XieGQ ( 2010). How will the distribution and size of subalpine Abies georgei forest respond to climate change? A study in northwest Yunnan, China Physical Geography, 31, 319-335. DOI:10.2747/0272-3646.31.4.319URL http://bellwether.metapress.com/openurl.asp?genre=article&id=doi:10.2747/0272-3646.31.4.319
XieCQ, TianMX, ZhaoZR, ZhengWL, WangGY ( 2015). Spatial point pattern analysis of Abies georgei var. smithii in forest of Sygera Mountains in southeast Tibet, China Chinese Journal of Applied Ecology, 26, 1617-1624. URL [本文引用: 3] 基于4 hm2色季拉山长期监测样地调查资料,采用成对相关函数点格局方法,分析西藏色季拉山冷杉林群落优势种急尖长苞冷杉种群活立木不同生长阶段和不同大小死亡木的空间格局及它们之间的空间关联性.结果表明:急尖长苞冷杉种群活立木径级分布连续,呈倒"J"型,为增长型种群.小树在小尺度(0~7 m)上呈显著聚集分布,随着龄级和研究尺度的增加,冷杉种群聚集强度逐渐减弱,最终趋向于均匀或随机分布格局.小树与大树、中树与大树在小尺度(0~35和0~30 m)上呈负关联,在大尺度(45~100和80~100 m)上呈正关联,且个体间龄级相差越大,其关联性越强.冷杉死亡所释放的空间不足以影响大树对小树的距离制约,大径级死亡木与小树在小尺度(0~34和5~27 m)上具有负关联性,在大尺度(49~100和73~100 m)上具有正关联性.自疏作用和Janzen-Connell假说中的距离制约效应可能是色季拉山急尖长苞冷杉种群空间格局形成的重要原因. [ 解传奇, 田民霞, 赵忠瑞, 郑维列, 王国严 ( 2015). 西藏色季拉山急尖长苞冷杉种群点格局分析 应用生态学报, 26, 1617-1624.] URL [本文引用: 3] 基于4 hm2色季拉山长期监测样地调查资料,采用成对相关函数点格局方法,分析西藏色季拉山冷杉林群落优势种急尖长苞冷杉种群活立木不同生长阶段和不同大小死亡木的空间格局及它们之间的空间关联性.结果表明:急尖长苞冷杉种群活立木径级分布连续,呈倒"J"型,为增长型种群.小树在小尺度(0~7 m)上呈显著聚集分布,随着龄级和研究尺度的增加,冷杉种群聚集强度逐渐减弱,最终趋向于均匀或随机分布格局.小树与大树、中树与大树在小尺度(0~35和0~30 m)上呈负关联,在大尺度(45~100和80~100 m)上呈正关联,且个体间龄级相差越大,其关联性越强.冷杉死亡所释放的空间不足以影响大树对小树的距离制约,大径级死亡木与小树在小尺度(0~34和5~27 m)上具有负关联性,在大尺度(49~100和73~100 m)上具有正关联性.自疏作用和Janzen-Connell假说中的距离制约效应可能是色季拉山急尖长苞冷杉种群空间格局形成的重要原因.
XieZQ, ChenWL, LuP, HuD ( 1999). The demography and age structure of the endangered plant population of Cathaya argyrophylla Acta Ecologica Sinica, 19, 523-528. DOI:10.3321/j.issn:1000-0933.1999.06.002URL [本文引用: 1] 通过对银杉主要群落种群的数量统计和年龄组成的分析,编绘了银杉种的特定时间生命表,存活曲线和年龄结构图,结果表明,银杉种群年龄超过200a后,出现植株个体死亡高峰,种群的平均生命期望陡降,预示着种群的生理衰退。银杉种群的存活曲线介于DeeveyI和II型曲线之间整个银杉种群在40龄级以上的年龄结构基本表现为增长型,但不同的银杉群落种群的年龄结构差异很大,大部分种群都存在的幼龄个体缺乏的现象,其中花坪 [ 谢宗强, 陈伟烈, 路鹏, 胡东 ( 1999). 濒危植物银杉的种群统计与年龄结构 生态学报, 19, 523-528.] DOI:10.3321/j.issn:1000-0933.1999.06.002URL [本文引用: 1] 通过对银杉主要群落种群的数量统计和年龄组成的分析,编绘了银杉种的特定时间生命表,存活曲线和年龄结构图,结果表明,银杉种群年龄超过200a后,出现植株个体死亡高峰,种群的平均生命期望陡降,预示着种群的生理衰退。银杉种群的存活曲线介于DeeveyI和II型曲线之间整个银杉种群在40龄级以上的年龄结构基本表现为增长型,但不同的银杉群落种群的年龄结构差异很大,大部分种群都存在的幼龄个体缺乏的现象,其中花坪
YangH, LiYL, ShenL, KangXG, YueG, WangY ( 2014). Spatial distribution patterns of seedling and sapling in a spruce-fir forest in the Changbai Mountains area in northeastern China Acta Ecologica Sinica, 34, 7311-7319. DOI:10.5846/stxb201303150432URL [本文引用: 1] 长白山云冷杉针阔混交林是我国东北主要的森林类型之一,其乔木树种幼苗幼树的结构和动态决定着未来林分的 结构和生长动态.在长白山地区设置一块具有代表性的云冷杉针阔混交林幼苗幼树更新样地,统计分析幼苗幼树更新特征,绘制地径结构图、树高结构图及其空间分 布图.运用点格局分析中的单变量O-ring统计方法,分析更新树种的空间分布格局;用双变量O-ring统计方法,分析更新树种种间的空间关联性.研究 结果表明:(1)更新树种组成有冷杉(Abies nphrolepis)、色木槭(Acer mono)、紫椴(Tilia amurensis)、红皮云杉(Picea koraiensis)、红松(Pinus koraiensis)、蒙古栎(Quercus mongolica)、春榆(Ulmus japonica)7种,其中以冷杉、色木槭为主,更新幼苗幼树的地径近似呈倒J型分布,树高结构近似呈双峰分布;(2)所有更新树种、冷杉、色木槭在小 尺度1-10 m的范围内呈聚集分布,随着尺度增加,聚集程度减弱,逐渐趋于均匀分布和随机分布,紫椴、云杉和红松在空间所有尺度上以随机分布为主;(3)更新树种之间 的空间关联性在小尺度范围上正关联性比较多,较大尺度范围上负关联性比较多,随着尺度增加,空间关联性减弱. [ 杨华, 李艳丽, 沈林, 亢新刚, 岳刚, 王妍 ( 2014). 长白山云冷杉林幼苗幼树空间分布格局及其更新特征 生态学报, 34, 7311-7319.] DOI:10.5846/stxb201303150432URL [本文引用: 1] 长白山云冷杉针阔混交林是我国东北主要的森林类型之一,其乔木树种幼苗幼树的结构和动态决定着未来林分的 结构和生长动态.在长白山地区设置一块具有代表性的云冷杉针阔混交林幼苗幼树更新样地,统计分析幼苗幼树更新特征,绘制地径结构图、树高结构图及其空间分 布图.运用点格局分析中的单变量O-ring统计方法,分析更新树种的空间分布格局;用双变量O-ring统计方法,分析更新树种种间的空间关联性.研究 结果表明:(1)更新树种组成有冷杉(Abies nphrolepis)、色木槭(Acer mono)、紫椴(Tilia amurensis)、红皮云杉(Picea koraiensis)、红松(Pinus koraiensis)、蒙古栎(Quercus mongolica)、春榆(Ulmus japonica)7种,其中以冷杉、色木槭为主,更新幼苗幼树的地径近似呈倒J型分布,树高结构近似呈双峰分布;(2)所有更新树种、冷杉、色木槭在小 尺度1-10 m的范围内呈聚集分布,随着尺度增加,聚集程度减弱,逐渐趋于均匀分布和随机分布,紫椴、云杉和红松在空间所有尺度上以随机分布为主;(3)更新树种之间 的空间关联性在小尺度范围上正关联性比较多,较大尺度范围上负关联性比较多,随着尺度增加,空间关联性减弱.
YangXD, YanER, ZhangZH, SunBW, HuangHX, ArshadA, MaWJ, ShiQR ( 2013). Tree architecture of overlapping species among successional stages in evergreen broad-leaved forests in Tiantong region, Zhejiang Province, China Chinese Journal of Plant Ecology, 37, 611-619. DOI:10.3724/SP.J.1258.2013.00063URL [本文引用: 4] AimsMethodsImportant findings [ 杨晓东, 阎恩荣, 张志浩, 孙宝伟, 黄海侠, Arshad A, 马文济, 史青茹 ( 2013). 浙江天童常绿阔叶林演替阶段共有种的树木构型 植物生态学报, 37, 611-619.] DOI:10.3724/SP.J.1258.2013.00063URL [本文引用: 4] AimsMethodsImportant findings
BaiDZ ( 2012). The Impact Factors of Growth and Regeneration of Picea crassifolia Growing at Timberline in The Qilian Mountains PhD dissertation, Chinese Academy of Forestry, Beijing. URL [本文引用: 2] 林线作为郁闭森林和高山草甸之间的生态过渡带,对自然环境因子变化和人为干扰异常敏感。目前,虽然热量(温度)控制、环境胁迫、生长受限、繁殖更新障碍、碳平衡失调等假说能解释一定地域的高山林线现象,但并不能作为普遍适应的林线理论。本论文通过对祁连山阴坡建群种青海云杉林线自然更新和人工更新研究,探索影响祁连山林线更新的主要因素,以及人工更新对林线位移的影响和气候变暖条件下林线的变化趋势。 本论文主要研究了祁连山地区近50年来的气候变化、西水林区不同海拔的青海云杉幼苗更新、树年轮和枝条生长,以及在林线海拔以上青海云杉播种试验和幼苗移栽,分析了青海云杉林线树木生长、更新的影响因素,主要研究结果如下: 1.祁连山西水林区周边气象台站气象资料分析显示,52年来该地区年均温升高显著;降水略有增加,全年降水主要分布在5、6、7、8、9月份,占到全年降雨量的82%。实验区不同海拔5或6月份-10月份大气温度和土壤温度都在0℃以上,大气和土壤温度都随着海拔升高而降低,每升高100m温度降低0.44℃左右。林线海拔3300m最热月平均温度13.60℃高于全球林线海拔最热月平均温度10℃,林线根际温度6.22℃与全球林线根际温度6.7±0.80℃相同。 2.土壤有机质、全氮、速效磷和速效钾含量随着土层加深而减小;pH值随着土层加深而增大;全磷含量在各土层中变异性较小。有机质和全氮含量随着海拔升高而降低,全钾含量和pH值随着海拔的升高而减小;而速效磷和速效钾含量随着海拔的升高没有明显的规律。 3.随着海拔升高,青海云杉的生长表现为从水分制约向热量制约转变的趋势。低海拔处的青海云杉年生长量较高海拔处大。水分是下限林线青海云杉的制约因子,而随着海拔的升高热量条件逐渐成为主要的制约因子。生长季的秋季结束时间对青海云杉的生长制约较大。 4.海拔2900m和3000m的青海云杉种群林木生长呈现近倒“J”型结构,种群林木生长处于稳定状态;海拔3100m的种群发展呈现线性结构,种群开始出现衰退的现象;海拔3200m和3300m种群林木生长较年轻旺盛,无衰老的苗木出现,3200m处为波浪形变化,说明此海拔区段青海云杉种群发展波动较大。 5.随着海拔梯度的增加,青海云杉林更新幼苗数量呈现逐渐降低的趋势,海拔3200m以下的更新幼苗以中低高度级(25cm、25~55cm、105cm)为主,且海拔越低幼苗数量越多,85~105cm高度级的幼苗缺少,而海拔3300m各高度级幼苗数量相差不大且更新情况较差,甚至某些高度的幼苗缺失。 6.随着海拔的升高,更新幼苗的分布格局由随机分布向集群分布变化。海拔2900m,<25cm的幼苗仅为弱聚集性,表现为随机分布;85cm~105cm的幼苗为均匀分布;在海拔3000m上,85cm~105cm和>105cm两个高度级的幼苗表现为随机分布;3100m,3200m和3300m三个高海拔内,幼苗均呈现集群分布。不同海拔高度25cm的幼苗的生长主要与海拔梯度、土壤含水量、有机质、总地表覆盖度、活地被物盖度存在显著正相关,而与全钾、大气温度显著负相关;影响25cm~55cm和55~85cm的幼苗与土壤pH值、大气温度、全钾呈正相关,与有机质、活地被物盖度、总地表覆盖度呈负相关;影响85cm~105cm和﹥105cm的幼苗与大气温度、乔木层郁闭度、全钾呈正相关,与土壤pH值、土壤含水量、海拔梯度呈负相关。 7.从林线3300m到海拔3500m,播种试验发现,青海云杉种子均可发芽,并随海拔升高,发芽率降低,但无法越冬,而5年左右生青海云杉幼苗移栽,从3300到3700m均能成活,表明在祁连山高海拔地区种子发芽和幼苗成活是限制林线更新的一个重要环节。随着海拔的升高,人工更新幼苗生长越来越慢,长势变差,死亡率升高,海拔对幼苗生长影响明显。高海拔低温可能是限制种子发芽和幼苗成活的关键因素。 8.随海拔升高青海云杉叶片组织中NSC含量持续增加状态,随海拔升高温度降低青海云杉的的生长并未受到碳限制。 在气候变暖条件下,随着高海拔地区温度升高,祁连山青海云杉生长加快,低温对青海云杉种子发芽和幼苗成活的限制影响减小,有利于林线向高海拔扩展。 [ 白登忠 ( 2012). 祁连山青海云杉林线树木生长、更新的影响因素研究 博士学位论文, 中国林业科学研究院, 北京.] URL [本文引用: 2] 林线作为郁闭森林和高山草甸之间的生态过渡带,对自然环境因子变化和人为干扰异常敏感。目前,虽然热量(温度)控制、环境胁迫、生长受限、繁殖更新障碍、碳平衡失调等假说能解释一定地域的高山林线现象,但并不能作为普遍适应的林线理论。本论文通过对祁连山阴坡建群种青海云杉林线自然更新和人工更新研究,探索影响祁连山林线更新的主要因素,以及人工更新对林线位移的影响和气候变暖条件下林线的变化趋势。 本论文主要研究了祁连山地区近50年来的气候变化、西水林区不同海拔的青海云杉幼苗更新、树年轮和枝条生长,以及在林线海拔以上青海云杉播种试验和幼苗移栽,分析了青海云杉林线树木生长、更新的影响因素,主要研究结果如下: 1.祁连山西水林区周边气象台站气象资料分析显示,52年来该地区年均温升高显著;降水略有增加,全年降水主要分布在5、6、7、8、9月份,占到全年降雨量的82%。实验区不同海拔5或6月份-10月份大气温度和土壤温度都在0℃以上,大气和土壤温度都随着海拔升高而降低,每升高100m温度降低0.44℃左右。林线海拔3300m最热月平均温度13.60℃高于全球林线海拔最热月平均温度10℃,林线根际温度6.22℃与全球林线根际温度6.7±0.80℃相同。 2.土壤有机质、全氮、速效磷和速效钾含量随着土层加深而减小;pH值随着土层加深而增大;全磷含量在各土层中变异性较小。有机质和全氮含量随着海拔升高而降低,全钾含量和pH值随着海拔的升高而减小;而速效磷和速效钾含量随着海拔的升高没有明显的规律。 3.随着海拔升高,青海云杉的生长表现为从水分制约向热量制约转变的趋势。低海拔处的青海云杉年生长量较高海拔处大。水分是下限林线青海云杉的制约因子,而随着海拔的升高热量条件逐渐成为主要的制约因子。生长季的秋季结束时间对青海云杉的生长制约较大。 4.海拔2900m和3000m的青海云杉种群林木生长呈现近倒“J”型结构,种群林木生长处于稳定状态;海拔3100m的种群发展呈现线性结构,种群开始出现衰退的现象;海拔3200m和3300m种群林木生长较年轻旺盛,无衰老的苗木出现,3200m处为波浪形变化,说明此海拔区段青海云杉种群发展波动较大。 5.随着海拔梯度的增加,青海云杉林更新幼苗数量呈现逐渐降低的趋势,海拔3200m以下的更新幼苗以中低高度级(25cm、25~55cm、105cm)为主,且海拔越低幼苗数量越多,85~105cm高度级的幼苗缺少,而海拔3300m各高度级幼苗数量相差不大且更新情况较差,甚至某些高度的幼苗缺失。 6.随着海拔的升高,更新幼苗的分布格局由随机分布向集群分布变化。海拔2900m,<25cm的幼苗仅为弱聚集性,表现为随机分布;85cm~105cm的幼苗为均匀分布;在海拔3000m上,85cm~105cm和>105cm两个高度级的幼苗表现为随机分布;3100m,3200m和3300m三个高海拔内,幼苗均呈现集群分布。不同海拔高度25cm的幼苗的生长主要与海拔梯度、土壤含水量、有机质、总地表覆盖度、活地被物盖度存在显著正相关,而与全钾、大气温度显著负相关;影响25cm~55cm和55~85cm的幼苗与土壤pH值、大气温度、全钾呈正相关,与有机质、活地被物盖度、总地表覆盖度呈负相关;影响85cm~105cm和﹥105cm的幼苗与大气温度、乔木层郁闭度、全钾呈正相关,与土壤pH值、土壤含水量、海拔梯度呈负相关。 7.从林线3300m到海拔3500m,播种试验发现,青海云杉种子均可发芽,并随海拔升高,发芽率降低,但无法越冬,而5年左右生青海云杉幼苗移栽,从3300到3700m均能成活,表明在祁连山高海拔地区种子发芽和幼苗成活是限制林线更新的一个重要环节。随着海拔的升高,人工更新幼苗生长越来越慢,长势变差,死亡率升高,海拔对幼苗生长影响明显。高海拔低温可能是限制种子发芽和幼苗成活的关键因素。 8.随海拔升高青海云杉叶片组织中NSC含量持续增加状态,随海拔升高温度降低青海云杉的的生长并未受到碳限制。 在气候变暖条件下,随着高海拔地区温度升高,祁连山青海云杉生长加快,低温对青海云杉种子发芽和幼苗成活的限制影响减小,有利于林线向高海拔扩展。
ZhangJT ( 1998). Analysis of spatial point pattern for plant species Acta Phytocologica Sinica, 22, 57-62. URL [本文引用: 1] 植物种群在群落中的分布格局与空间尺度有着密切关系,传统的样方取样及其格局分析方法,只能人析一种尺度下的格局,本文引入一种新的格局分析方法-点格局分析,其是以种群空间分布的坐标点图为基础,通过本文对美国密西根州克林顿县栎林3个优势格局分析,它有3个明显的优点;1)能够分析各种尺度下的种群格局和种间关系,结果清楚,直观;2)所描述的结果更符合实际,尤其是对群落结构的描述;3)它有利于定点观察,研究时间 [ 张金屯 ( 1998). 植物种群空间分布的点格局分析 植物生态学报, 22, 57-62.] URL [本文引用: 1] 植物种群在群落中的分布格局与空间尺度有着密切关系,传统的样方取样及其格局分析方法,只能人析一种尺度下的格局,本文引入一种新的格局分析方法-点格局分析,其是以种群空间分布的坐标点图为基础,通过本文对美国密西根州克林顿县栎林3个优势格局分析,它有3个明显的优点;1)能够分析各种尺度下的种群格局和种间关系,结果清楚,直观;2)所描述的结果更符合实际,尤其是对群落结构的描述;3)它有利于定点观察,研究时间
BatlloriE, Blanco-MorenoJM, NinotJM, GutierrezE, CarrilloE ( 2009). Vegetation patterns at the alpine treeline ecotone: The influence of tree cover on abrupt change in species composition of alpine communities Journal of Vegetation Science, 20, 814-825. DOI:10.1111/j.1654-1103.2009.01085.xURL [本文引用: 1] Aims: The upper elevation limit of forest vegetation in mountain ranges (the alpine treeline ecotone) is expected to be highly sensitive to global change. Treeline shifts and/or ecotone afforestation could cause fragmentation and loss of alpine habitat, and are expected to trigger considerable alterations in alpine vegetation. We performed an analysis of vegetation structure at the treeline ecotone to evaluate whether distribution of the tree population determines the spatial pattern of vegetation (species composition and diversity) across the transition from subalpine forest to alpine vegetation. Location: Iberian eastern range of the Pyrenees. Methods: We studied 12 alpine Pinus uncinata treeline ecotones. Rectangular plots ranging from 940 to 1900 m were placed along the forest-alpine vegetation transition, from closed forest to the treeless alpine area. To determine community structure and species distribution in the treeline ecotone, species variation along the forest-alpine vegetation transition was sampled using releves of 0.5 m set every 2 m along the length of each plot. Fuzzy C-means clustering was performed to assess the transitional status of the releves in terms of species composition. The relation of P. uncinata canopy cover to spatial pattern of vegetation was evaluated using continuous wavelet transform analysis. Results: Vegetation analyses revealed a large degree of uniformity of the subalpine forest between all treeline ecotone areas studied. In contrast, the vegetation mosaic found upslope displayed great variation between sites and was characterized by abrupt changes in plant community across the treeline ecotone. Plant richness and diversity significantly increased across the ecotone, but tree cover and diversity boundaries were not spatially coincident. Conclusions: Our results revealed that no intermediate communities, in terms of species composition, are present in the treeline ecotone. Ecotone vegetation reflected both bedrock type and fine-scale heterogeneity at ground level, thereby reinforcing the importance of microenvironmental conditions for alpine community composition. Tree cover did not appear to be the principal driver of alpine community changes across the treeline ecotone. Micro-environmental heterogeneity, together with effects of past climatic and land-use changes on ecotone vegetation, may weaken the expected correlation between species distribution and vegetation structure.
CamareroJJ, GutierrezE ( 2004). Pace and pattern of recent treeline dynamics, response of ecotones to climatic variability in the Spanish Pyrenees Climatic Change, 63, 181-200. DOI:10.1023/B:CLIM.0000018507.71343.46URL [本文引用: 1] Treeline ecotones are regarded as sensitive monitors of the recent climatic warming. However, it has been suggested that their sensitivity depends more on changes in tree density than on treeline position. We study these processes and the effect of climate, mainly air temperature, on tree recruitment and recent treeline dynamics. We selected three relatively undisturbed sites in the Spanish Pyrenees, dominated by Pinus uncinata, and analyzed their recent dynamics at local spatial (0.300–0.5 ha) and short temporal scales (10000–300 years). We wanted to establish whether higher temperature was the only climatic factor causing an upward shift of the studied alpine treelines. The data we report show that treelines were ascending until a period of high interannual variability in mean temperature started (195000–95). During the late twentieth century, treeline fluctuation was less sensitive to climate than was the change in tree density within the ecotone. Tree recruitment and treeline position responded to contrasting climatic signals; tree recruitment was favored by high March temperatures whereas treeline position ascended in response to warm springs. We found a negative relationship between mean treeline-advance rate and March temperature variability. According to our findings, if the interannual variability of March temperature increases, the probability of successful treeline ascent will decrease.
ZhangPJ, QingH, ZhangL, XuYD, MuL, YeRH, QiuX, ChangH, ShenHH, YangJ ( 2017). Population structure and spatial pattern of Caragana tibetica communities in Nei Mongol shrub-encroached grassland Chinese Journal of Plant Ecology, 41, 165-174. DOI:10.17521/cjpe.2015.0448URL 灌丛化草原的形成机制和群落结构与功能是植被生态学研究的一个重要科学问题。植物种群空间格局分析是研究植物种群相互作用及种群与环境关系的重要手段。灌丛化草原是内蒙古草原的主要类型。该文选择分布在鄂尔多斯市的毛刺锦鸡儿(Caragana tibetica)灌丛化草原为研究对象,布设一个64 m×64 m的样方,采用“三线定位法”确定样方中灌木的空间位置,用点格局的方法分析灌木空间分布格局与种内关联。结果显示:(1)毛刺锦鸡儿种群为增长型种群,成熟毛刺锦鸡儿个体在半径〈3.7 m的范围内为均匀分布,范围增大则呈随机分布;毛刺锦鸡儿幼年个体在0.3–20.4 m之间为聚集分布,其他范围表现为随机分布;死亡个体在0.7–1.4 m范围为聚集分布,其他范围则随机分布。(2)毛刺锦鸡儿存活个体对死亡个体在0–9.9 m范围内为不明显的正相互作用,其他范围无相关关系;成熟个体对幼年个体在〈2.2 m范围表现为负相互作用,在〉2.2 m范围表现为不相关。研究显示,毛刺锦鸡儿灌丛化草原不同发育阶段的毛刺锦鸡儿种群空间分布格局是不同的,并随空间尺度的变化而变化,如果在干旱区采用毛刺锦鸡儿对退化草原进行恢复重建,灌木间的株行距以4 m为宜。 [ 张璞进, 清华, 张雷, 徐延达, 木兰, 晔薷罕, 邱晓, 常虹, 沈海花, 杨劼 ( 2017). 内蒙古灌丛化草原毛刺锦鸡儿种群结构和空间分布格局 植物生态学报, 41, 165-174.] DOI:10.17521/cjpe.2015.0448URL 灌丛化草原的形成机制和群落结构与功能是植被生态学研究的一个重要科学问题。植物种群空间格局分析是研究植物种群相互作用及种群与环境关系的重要手段。灌丛化草原是内蒙古草原的主要类型。该文选择分布在鄂尔多斯市的毛刺锦鸡儿(Caragana tibetica)灌丛化草原为研究对象,布设一个64 m×64 m的样方,采用“三线定位法”确定样方中灌木的空间位置,用点格局的方法分析灌木空间分布格局与种内关联。结果显示:(1)毛刺锦鸡儿种群为增长型种群,成熟毛刺锦鸡儿个体在半径〈3.7 m的范围内为均匀分布,范围增大则呈随机分布;毛刺锦鸡儿幼年个体在0.3–20.4 m之间为聚集分布,其他范围表现为随机分布;死亡个体在0.7–1.4 m范围为聚集分布,其他范围则随机分布。(2)毛刺锦鸡儿存活个体对死亡个体在0–9.9 m范围内为不明显的正相互作用,其他范围无相关关系;成熟个体对幼年个体在〈2.2 m范围表现为负相互作用,在〉2.2 m范围表现为不相关。研究显示,毛刺锦鸡儿灌丛化草原不同发育阶段的毛刺锦鸡儿种群空间分布格局是不同的,并随空间尺度的变化而变化,如果在干旱区采用毛刺锦鸡儿对退化草原进行恢复重建,灌木间的株行距以4 m为宜。
ChengW, LuoP, WuN ( 2005). Ecological characteristics of minjiang fir (Abies faxoniana Rehd. et Wild) population near timberline on upper Min River Chinese Journal of Applied and Envirnmental Biology, 11, 300-303. DOI:10.3321/j.issn:1006-687X.2005.03.010URL [本文引用: 3] 在松潘县弓杠岭采用样方调查的方法针对岷江冷杉(Abies faxoniana)种群结构、幼苗更新及个体生长及与海拔变化的关系进行了研究.样地分别位于林线下方、林线及树线区,个体年龄采用WinDENRO系 统或侧生轮枝计数的方法确定.结果发现:(1)岷江冷杉种群更新与样地海拔高度、干扰强度有关.(2)岷江冷杉的个体生长过程可分为生长期、成熟期、过熟 期3个阶段,但随着海拔的升高,这3个时期生长速度的差异变得不明显.(3)3个样地内岷江冷杉种群随海拔升高,其平均高度呈现由高到低,胸径由大到小, 个体数量由多到少的变化趋势.(4)岷江冷杉种群大中型立木中出现由于病腐导致空心树的比例随海拔升高有增加趋势.图4表1参28 [ 程伟, 罗鹏, 吴宁 ( 2005). 岷江上游林线附近岷江冷杉种群(Abies faxoniana Rehd. et Wild)的生态学特点 应用与环境生物学报, 11, 300-303.] DOI:10.3321/j.issn:1006-687X.2005.03.010URL [本文引用: 3] 在松潘县弓杠岭采用样方调查的方法针对岷江冷杉(Abies faxoniana)种群结构、幼苗更新及个体生长及与海拔变化的关系进行了研究.样地分别位于林线下方、林线及树线区,个体年龄采用WinDENRO系 统或侧生轮枝计数的方法确定.结果发现:(1)岷江冷杉种群更新与样地海拔高度、干扰强度有关.(2)岷江冷杉的个体生长过程可分为生长期、成熟期、过熟 期3个阶段,但随着海拔的升高,这3个时期生长速度的差异变得不明显.(3)3个样地内岷江冷杉种群随海拔升高,其平均高度呈现由高到低,胸径由大到小, 个体数量由多到少的变化趋势.(4)岷江冷杉种群大中型立木中出现由于病腐导致空心树的比例随海拔升高有增加趋势.图4表1参28
ConditR, AshtonPS, BakerP, BunyavejchewinS, GunatillekeS, GunatillekeN, HubbellSP, FosterRB, ItohA, LafrankieJV, LeeHS, LososE, ManokaranN, SukumarR, YamakuraT ( 2000). Spatial patterns in the distribution of tropical tree species Science, 288, 1414-1418. DOI:10.1126/science.288.5470.1414URLPMID:10827950 [本文引用: 1] Fully mapped tree census plots of large area, 25 to 52 hectares, have now been completed at six different sites in tropical forests, including dry deciduous to wet evergreen forest on two continents. One of the main goals of these plots has been to evaluate spatial patterns in tropical tree populations. Here the degree of aggregation in the distribution of 1768 tree species is examined based on the average density of conspecific trees in circular neighborhoods around each tree. When all individuals larger than 1 centimeter in stem diameter were included, nearly every species was more aggregated than a random distribution. Considering only larger trees ( 10 centimeters in diameter), the pattern persisted, with most species being more aggregated than random. Rare species were more aggregated than common species. All six forests were very similar in all the particulars of these results.
DaiJH, CuiHT ( 1999). A reviewon the studies of alpine timberline Scientia Geographica Sinica, 19(3), 52-58. DOI:10.3969/j.issn.1000-0690.1999.03.010URL [本文引用: 1] 高山林线作为山地郁闭林和高山草甸的分界在山地生态系统中占有重要地位。以往对山地生态的研究远少于对其它类型陆地生态系统的研究,高山林线方面的研究就更为缺乏。讨论了高山林线的概念和内容,对国内外高山林线研究及特点作了简单回顾,指出高山林线的研究意义及在我国开展此项研究的初步方案。 [ 戴君虎, 崔海亭 ( 1999). 国内外高山林线研究综述 地理科学, 19(3), 52-58.] DOI:10.3969/j.issn.1000-0690.1999.03.010URL [本文引用: 1] 高山林线作为山地郁闭林和高山草甸的分界在山地生态系统中占有重要地位。以往对山地生态的研究远少于对其它类型陆地生态系统的研究,高山林线方面的研究就更为缺乏。讨论了高山林线的概念和内容,对国内外高山林线研究及特点作了简单回顾,指出高山林线的研究意义及在我国开展此项研究的初步方案。
FalsterDS, WestobyM ( 2003). Plant height and evolutionary games Trends in Ecology & Evolution, 18, 337-343. DOI:10.1016/S0169-5347(03)00061-2URL [本文引用: 1] In plants, investment in height improves access to light, but incurs costs in construction and maintenance of the stem. Because the benefits of plant height depend on which other height strategies are present, competition for light can usefully be framed as a game-theoretic problem. The vertical structure of the world's vegetation, which is inefficient for plant growth, can then be understood as the outcome of evolutionary and ecological arms races. In addition, game-theoretic models predict taller vegetation on sites of higher leaf area index, and allocation to reproduction only after an initial period of height growth. However, of 14 game-theoretic models for height reviewed here, only one predicts coexistence of a mix of height strategies, a conspicuous feature of most vegetation. We suggest that game-theoretic models could help account for observed mixtures of height strategies if they incorporated processes for coexistence along spectra of light income and time since disturbance.
GaireNP, KoiralaM, BhujuDR, BorgaonkarHP ( 2014). Treeline dynamics with climate change at the Central Nepal Himalaya Climate of the Past, 10, 1277-1290. DOI:10.5194/cp-10-1277-2014URL [本文引用: 1]
GaoJ, WangJN, XuB, XieY, HeJD, WuY ( 2016). Plant leaf traits, height and biomass partitioning in typical ephemerals under different levels of snow cover thickness in an alpine meadow Chinese Journal of Plant Ecology, 40, 775-787. DOI:10.17521/cjpe.2015.0288URL [本文引用: 2] 在高寒生命带,雪被作为重要的综合环境因子,影响着植物的生理生态特征、种群动态及群落演替等过程,进而作用于生态系统的功能与服务。通过在青藏高原东缘高寒草甸设置厚雪、中雪和浅雪3个地段,选取早春开花的常见种紫罗兰报春(Primula purdomii)、甘肃马先蒿(Pedicularis kansuensis)、高原毛茛(Ranunculus tanguticus),研究了三种植物株高、叶片性状和生物量分配随雪被厚度的变化规律,以及三者之间的关系。结果表明:甘肃马先蒿和高原毛茛在生境状况较好的地段比叶面积相对较高,紫罗兰报春由于具有根状茎和肉质根,在厚雪地段比叶面积相对较小;关于单个物种的地上-地下生物量的关系,紫罗兰报春在厚雪和浅雪地段为完全一致的异速生长关系,而甘肃马先蒿和高原毛茛在个别地段并无显著相关关系。总体而言,三种植物整体样本的地上-地下生物量在不同雪被地段均为异速生长关系,不支持等速生长假说,并且地上生物量能够很好地解释地下生物量的变异;功能性状和生物量指标间的相关性,在紫罗兰报春和高原毛茛表现较好,而在甘肃马先蒿表现较弱。植物对环境变化的适应具有一定的规律,同时也表现出物种特异性。今后的研究中,应注重构建植物适应环境变化的功能性状谱,以更好地理解全球变化背景下植物功能性状的响应及其适应策略。 [ 高景, 王金牛, 徐波, 谢雨, 贺俊东, 吴彦 ( 2016). 不同雪被厚度下典型高山草地早春植物叶片性状、株高及生物量分配的研究 植物生态学报, 40, 775-787.] DOI:10.17521/cjpe.2015.0288URL [本文引用: 2] 在高寒生命带,雪被作为重要的综合环境因子,影响着植物的生理生态特征、种群动态及群落演替等过程,进而作用于生态系统的功能与服务。通过在青藏高原东缘高寒草甸设置厚雪、中雪和浅雪3个地段,选取早春开花的常见种紫罗兰报春(Primula purdomii)、甘肃马先蒿(Pedicularis kansuensis)、高原毛茛(Ranunculus tanguticus),研究了三种植物株高、叶片性状和生物量分配随雪被厚度的变化规律,以及三者之间的关系。结果表明:甘肃马先蒿和高原毛茛在生境状况较好的地段比叶面积相对较高,紫罗兰报春由于具有根状茎和肉质根,在厚雪地段比叶面积相对较小;关于单个物种的地上-地下生物量的关系,紫罗兰报春在厚雪和浅雪地段为完全一致的异速生长关系,而甘肃马先蒿和高原毛茛在个别地段并无显著相关关系。总体而言,三种植物整体样本的地上-地下生物量在不同雪被地段均为异速生长关系,不支持等速生长假说,并且地上生物量能够很好地解释地下生物量的变异;功能性状和生物量指标间的相关性,在紫罗兰报春和高原毛茛表现较好,而在甘肃马先蒿表现较弱。植物对环境变化的适应具有一定的规律,同时也表现出物种特异性。今后的研究中,应注重构建植物适应环境变化的功能性状谱,以更好地理解全球变化背景下植物功能性状的响应及其适应策略。
HarschMA, HulmePE, McgloneMS, DuncanRP ( 2009). Are treelines advancing? A global meta-analysis of treeline response to climate warming Ecology Letters, 12, 1040-1049. DOI:10.1111/ele.2009.12.issue-10URL [本文引用: 2]
IPCC (Intergovernmental Panel on Climate Change) (2014). Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, UK. [本文引用: 2]
JinHJ, MaQL, ZhangDK, LiuYJ, YuanHB ( 2012). Analysis on typical shrub plant community characteristics and quantitative characteristics in Ulanbuh desert Acta Botanica Boreall-Occidentalia Sinica, 32, 579-588. DOI:10.3969/j.issn.1000-4025.2012.03.022URL [本文引用: 1] The shrub community is the most important ecosystem components in Ulanbuh desert.The community structure and quantitative characteristics of shrub community was researched,which can provide the theory basis and the practice foundation for protecting biodiversity and sustainable utilization of plant resources in the area.According to the quadrat investigation material,the community structure and quantitative characteristics of typical shrub community was analyzed by the species composition,life form spectrum,coverage characteristics,species diversity and similarity of community.The results showed that:(1)The main community type was the community of diversity index varied between 0.98~3.42.The diversity index varied between 0.415 5~0.879 5,the species composition of each shrub community was relatively simple.(5)The similitude coefficient of each community varied between 0.068 2~0.604 7,so each community similarity did not reached the level of very similar.There was mild similar among the three community,which were N.tangutorum,H.ammodendron and A.mongolicus,but the similitude coefficient between the other communities were less than 0.25.It showed there were great difference between community habitats. [ 靳虎甲, 马全林, 张德魁, 刘有军, 袁洪波 ( 2012). 乌兰布和沙漠典型灌木群落结构及数量特征 西北植物学报, 32, 579-588.] DOI:10.3969/j.issn.1000-4025.2012.03.022URL [本文引用: 1] The shrub community is the most important ecosystem components in Ulanbuh desert.The community structure and quantitative characteristics of shrub community was researched,which can provide the theory basis and the practice foundation for protecting biodiversity and sustainable utilization of plant resources in the area.According to the quadrat investigation material,the community structure and quantitative characteristics of typical shrub community was analyzed by the species composition,life form spectrum,coverage characteristics,species diversity and similarity of community.The results showed that:(1)The main community type was the community of diversity index varied between 0.98~3.42.The diversity index varied between 0.415 5~0.879 5,the species composition of each shrub community was relatively simple.(5)The similitude coefficient of each community varied between 0.068 2~0.604 7,so each community similarity did not reached the level of very similar.There was mild similar among the three community,which were N.tangutorum,H.ammodendron and A.mongolicus,but the similitude coefficient between the other communities were less than 0.25.It showed there were great difference between community habitats.
KimJW, LeeJS ( 2015). Dynamics of alpine treelines: Positive feedbacks and global, regional and local controls Journal of Ecology and Environment, 38, 1-14. DOI:10.5141/ecoenv.2015.001URL
KlasnerFL, FagreDB ( 2002). A half century of change in alpine treeline patterns at Glacier National Park, Montana, USA Arctic Antarctic and Alpine Research, 34, 49-56. DOI:10.1080/15230430.2002.12003468URL [本文引用: 2]
KnowlesP, GrantMC ( 1983). Age and size structure analyses of engelmann spruce, ponderosa pine, lodgepole pine, and limber pine in Colorado Ecology, 64, 1-9. DOI:10.2307/1937322URL [本文引用: 3] Ages and diameters were measured in mature stands for each of 507 ponderosa pine, 541 lodgepole pine, 141 limber pine, and 217 Engelmann spruce trees in the Colorado Front Range of the Rocky Mountains, USA. Cumulative age distributions were statistically different for each species. The spruce and ponderosa pine age distributions each exhibited a strong infection point at @?210 and 125 yr, respectively, while neither lodgepole nor limber pine showed such a sharp inflection point. We suggest that the presence of this inflection point may be indicative of "climax" type in mature stands and interpret the age distributions of these species as reflective of their climax, colonizing and fugitive ecological patterns, respectively. Diameter distribution curves exhibited patterns markedly different from the age patterns. Ponderosa pine and Englemann spruce showed almost identical distribution despite widely disparate age structure. We interpret this result to simply that to the complex of forces which influence size distributions in forest trees is very similar in ponderosa pine and spruce although they form very different ecological communities. Lodgepole and limber pine diameter distributions were quite different. Our conjectures that the presence of a sharp inflection point in cumulative age distributions indicates a "climax" stand and that "climax" species will often have coincident diameter distributions, are offered as empirically testable hypotheses.
KörnerC ( 1998). A re-assessment of high elevation treeline positions and their explanation Oecologia, 115, 445-459. DOI:10.1007/s004420050540URLPMID:28308263 [本文引用: 1] In this review I first compile data for the worldwide position of climate-driven alpine treelines. Causes for treeline formation are then discussed with a global perspective. Available evidence suggests a combination of a general thermal boundary for tree growth, with regionally variable "modulatory" forces, including the presence of certain taxa. Much of the explanatory evidence found in the literature relates to these modulatory aspects at regional scales, whereas no good explanations emerged for the more fundamental global pattern related to temperature per se, on which this review is focused. I hypothesize that the life form "tree" is limited at treeline altitudes by the potential investment, rather than production, of assimilates (growth as such, rather than photosynthesis or the carbon balance, being limited). In shoots coupled to a cold atmosphere, meristem activity is suggested to be limited for much of the time, especially at night. By reducing soil heat flux during the growing season the forest canopy negatively affects root zone temperature. The lower threshold temperature for tissue growth and development appears to be higher than 3 C and lower than 10 C, possibly in the 5.5-7.5 C range, most commonly associated with seasonal means of air temperature at treeline positions. The physiological and developmental mechanisms responsible have yet to be analyzed. Root zone temperature, though largely unknown, is likely to be most critical.
KörnerC, PaulsenJ ( 2004). A world-wide study of high altitude treeline temperatures Journal of Biogeography, 31, 713-732. DOI:10.1111/j.1365-2699.2003.01043.xURL [本文引用: 1] Aim At a coarse scale, the treelines of the world's mountains seem to follow a common isotherm, but the evidence for this has been indirect so far. Here we aim at underpinning this with facts.
KullmanL ( 1993). Tree timit dynamics of Betula pubescens ssp. tortuosa in relation to climate variability: Evidence from Central Sweden Journal of Vegetation Science, 4, 765. [本文引用: 2]
KullmanL ( 2001). 20th century climate warming and tree-limit rise in the southern Scandes of Sweden AMBIO, 30, 72-80. DOI:10.1579/0044-7447-30.2.72URLPMID:11374309 Climate warming by ca. 0.8 C between the late-19th and late-20th century, although with some fluctuations, has forced multispecies elevational tree-limit advance by > 100 m for the principal tree species in the Swedish part of the Scandinavian mountain range. Predominantly, these processes imply growth in height of old established individuals and less frequently upslope migration of new individuals. After a slight retardation during some cooler decades after 1940, a new active phase of tree-limit advance has occurred with a series of exceptionally mild winters and some warm summers during the 1990s. The magnitude of total 20th century tree-limit rise varies with topoclimate and is mainly confined to wind-sheltered and snow-rich segments of the landscape. Thickening of birch tree stands in the "advance belt" has profoundly altered the general character of the subalpine/low alpine landscape and provides a positive feedback loop for further progressive change and resilience to short-term cooling episodes. All upslope tree-limit shifts and associated landscape transformations during the 20th century have occurred without appreciable time lags, which constitutes knowledge fundamental to the generation of realistic models concerning vegetation responses to potential future warming. The new and elevated pine tree-limit may be the highest during the past 400014C years. Thus, it is tentatively inferred that the 20th century climate is unusually warm in a late-Holocene perspective.
LanGY, HuYH, CaoM, ZhuH, WangH, ZhouSS, DengXB, CuiJY, HuangJG, LiuLY, XuHL, SongJP, HeYC ( 2008). Establishment of Xishuangbanna tropical forest dynamics plot: Species compositions and spatial distribution patterns Chinese Journal of Plant Ecology (Chinese Version), 32, 287-298. DOI:10.3773/j.issn.1005-264x.2008.02.006URL [本文引用: 3] 中国科学院机构知识库(CAS IR GRID)以发展机构知识能力和知识管理能力为目标,快速实现对本机构知识资产的收集、长期保存、合理传播利用,积极建设对知识内容进行捕获、转化、传播、利用和审计的能力,逐步建设包括知识内容分析、关系分析和能力审计在内的知识服务能力,开展综合知识管理。 [ 兰国玉, 胡跃华, 曹敏, 朱华, 王洪, 周仕顺, 邓晓保, 崔景云, 黄建国, 刘林云, 许海龙, 宋军平, 何有才 ( 2008). 西双版纳热带森林动态监测样地——树种组成与空间分布格局 植物生态学报, 32, 287-298.] DOI:10.3773/j.issn.1005-264x.2008.02.006URL [本文引用: 3] 中国科学院机构知识库(CAS IR GRID)以发展机构知识能力和知识管理能力为目标,快速实现对本机构知识资产的收集、长期保存、合理传播利用,积极建设对知识内容进行捕获、转化、传播、利用和审计的能力,逐步建设包括知识内容分析、关系分析和能力审计在内的知识服务能力,开展综合知识管理。
LiGC, SongHD, LiQ, BuSH ( 2017). Spatial point pattern analysis of main trees and flowering Fargesia qinlingensis in Abies fargesii forests in Mt. Taibai of the Qinling Mountains, China Chinese Journal of Applied Ecology, 28, 3487-3493. DOI:10.13287/j.1001-9332.201711.003URL [本文引用: 1] 采用单变量和双变量O-ring函数对太白山大熊猫栖息地巴山冷杉林主要树种的空间分布格局、种间关联性及其与林下开花箭竹的空间关联性进行了多尺度分析.结果表明:巴山冷杉林中,巴山冷杉数量最多,但种群结构衰退,白桦种群相对年轻,种群结构稳定,红桦种群也呈衰退趋势;3个主要树种在小尺度上呈聚集分布,随尺度增加,逐渐表现为随机分布.3个树种的空间关联性主要表现在小尺度范围内,随尺度增加,空间分布格局逐渐表现为不关联;巴山冷杉和白桦与开花秦岭箭竹在大、中尺度内呈现正相关,而红桦与开花秦岭箭竹在大、中尺度上表现出负相关.大熊猫栖息地内乔木和林下秦岭箭竹共同推动森林的动态演替和发展,进而影响秦岭大熊猫栖息地的环境变化. [ 李国春, 宋华东, 李琦, 卜书海 ( 2017). 太白山巴山冷杉林主要树种与开花秦岭箭竹的空间点格局分析 应用生态学报, 28, 3487-3493.] DOI:10.13287/j.1001-9332.201711.003URL [本文引用: 1] 采用单变量和双变量O-ring函数对太白山大熊猫栖息地巴山冷杉林主要树种的空间分布格局、种间关联性及其与林下开花箭竹的空间关联性进行了多尺度分析.结果表明:巴山冷杉林中,巴山冷杉数量最多,但种群结构衰退,白桦种群相对年轻,种群结构稳定,红桦种群也呈衰退趋势;3个主要树种在小尺度上呈聚集分布,随尺度增加,逐渐表现为随机分布.3个树种的空间关联性主要表现在小尺度范围内,随尺度增加,空间分布格局逐渐表现为不关联;巴山冷杉和白桦与开花秦岭箭竹在大、中尺度内呈现正相关,而红桦与开花秦岭箭竹在大、中尺度上表现出负相关.大熊猫栖息地内乔木和林下秦岭箭竹共同推动森林的动态演替和发展,进而影响秦岭大熊猫栖息地的环境变化.
LiLP, MohammaA, WangXP ( 2011). Study on relationship between height and DBH of mountain coniferous forests in Xinjiang Arid Zone Research, 28(1), 47-53. Magsci [本文引用: 1] 树木的生长包括了径向生长和垂直生长两个方面,这两方面具有密切的关系,这一关系既受到树木本身的控制,同时也受森林立地条件的影响。通过分析新疆阿尔泰山、天山南北坡以及昆仑山区11个地点120个样方的胸径-树高关系。结果表明:昆仑山的尖削度(taper)显著大于其余3地,其潜在最大树高(potential height, <em>H</em><sup>*</sup>)显著小于阿尔泰山和天山北坡。随着海拔的升高,针叶林尖削度显著上升,潜在最大树高显著下降;随年降水的增加,尖削度显著减小,而潜在最大树高显著增加。一般线性模型(general linear model, GLM)分析表明,尖削度和潜在最大树高受年降水、坡向和林型等因素影响较大,而受年平均气温的影响较小,说明新疆山区针叶林胸径-树高关系主要受水分的影响,即水分是生产力的限制因素。胸径-树高与气候的关系具有区域分异,在北疆,尖削度受坡向、坡位和年平均气温的影响较大,而受年降水的影响较小;在南疆,尖削度主要受年降水和坡向的影响,其次是年平均气温。 [ 李利平, 安尼瓦尔·买买提, 王襄平 ( 2011). 新疆山地针叶林乔木胸径-树高关系分析 干旱区研究, 28(1), 47-53.] Magsci [本文引用: 1] 树木的生长包括了径向生长和垂直生长两个方面,这两方面具有密切的关系,这一关系既受到树木本身的控制,同时也受森林立地条件的影响。通过分析新疆阿尔泰山、天山南北坡以及昆仑山区11个地点120个样方的胸径-树高关系。结果表明:昆仑山的尖削度(taper)显著大于其余3地,其潜在最大树高(potential height, <em>H</em><sup>*</sup>)显著小于阿尔泰山和天山北坡。随着海拔的升高,针叶林尖削度显著上升,潜在最大树高显著下降;随年降水的增加,尖削度显著减小,而潜在最大树高显著增加。一般线性模型(general linear model, GLM)分析表明,尖削度和潜在最大树高受年降水、坡向和林型等因素影响较大,而受年平均气温的影响较小,说明新疆山区针叶林胸径-树高关系主要受水分的影响,即水分是生产力的限制因素。胸径-树高与气候的关系具有区域分异,在北疆,尖削度受坡向、坡位和年平均气温的影响较大,而受年降水的影响较小;在南疆,尖削度主要受年降水和坡向的影响,其次是年平均气温。
LiangEY, LeuschnerC, DulamsurenC, WagnerB, HauckM ( 2016). Global warming-related tree growth decline and mortality on the north-eastern Tibetan Plateau Climatic Change, 13, 163-176. DOI:10.1007/s10584-015-1531-yURL [本文引用: 3] Semi-arid forests at the limit of their existence close to the Gobi Desert in Inner Asia might be vulnerable to warming-induced drought stress. Yet, not much is known about the impact of...
LiangEY, WangYF, EcksteinD, LuoTX ( 2011). Little change in the fir tree-line position on the southeastern Tibetan Plateau after 200 years of warming New Phytologist, 190, 760-769. DOI:10.1111/nph.2011.190.issue-3URL [本文引用: 2]
LiuXD, ChenBD ( 2000). Climatic warming in the Tibetan Plateau during recent decades International Journal of Climatology, 20, 1729-1742. DOI:10.1002/1097-0088(20001130)20:14<1729::AID-JOC556>3.0.CO;2-YURL [本文引用: 1] Abstract Adequate knowledge of climatic change over the Tibetan Plateau (TP) with an average elevation of more than 4000 m above sea level (a.s.l.) has been insufficient for a long time owing to the lack of sufficient observational data. In the present study, monthly surface air temperature data were collected from almost every meteorological station on the TP since their establishment. There are 97 stations located above 2000 m a.s.l. on the TP; the longest records at five stations began before the 1930s, but most records date from the mid-1950s. Analyses of the temperature series show that the main portion of the TP has experienced statistically significant warming since the mid-1950s, especially in winter, but the recent warming in the central and eastern TP did not reach the level of the 1940s warm period until the late 1990s. Compared with the Northern Hemisphere and the global average, the warming of the TP occurred early. The linear rates of temperature increase over the TP during the period 1955–1996 are about 0.16°C/decade for the annual mean and 0.32°C/decade for the winter mean, which exceed those for the Northern Hemisphere and the same latitudinal zone in the same period. Furthermore, there is also a tendency for the warming trend to increase with the elevation in the TP and its surrounding areas. This suggests that the TP is one of the most sensitive areas to respond to global climate change. Copyright 08 2000 Royal Meteorological Society
LloydAH, FastieCL ( 2002). Spatial and temporal variability in the growth and climate response of treeline trees in Alaska Climate Change, 52, 481-509. DOI:10.1023/A:1014278819094URL [本文引用: 1] In this study, we investigated the response of trees growing at the cold margins of the boreal forest to climate variation in the 20th century. Working at eight sites at and near alpine and arctic treeline in three regions in Alaska, we compared tree growth (from measured tree ring-widths) to historical climate data to document how growth has responded to climate variation in the 20th century. We found that there was substantial regional variability in response to climate variation. Contrary to our expectations, we found that after 1950 warmer temperatures were associated with decreased tree growth in all but the wettest region, the Alaska Range. Although tree growth increased from 1900 1950 at almost all sites, significant declines in tree growth were common after 1950 in all but the Alaska Range sites. We also found that there was substantial variability in response to climate variation according to distance to treeline. Inverse growth responses to temperature were more common at sites below the forest margin than at sites at the forest margin. Together, these results suggest that inverse responses to temperature are widespread, affecting even the coldest parts of the boreal forest. Even in such close proximity to treeline, warm temperatures after 1950 have been associated with reduced tree growth. Growth declines were most common in the warmer and drier sites, and thus support the hypothesis that drought-stress may accompany increased warming in the boreal forest.
LloydAH, FastieCL ( 2003). Recent changes in treeline forest distribution and structure in interior Alaska Ecoscience, 10, 176-185. DOI:10.1080/11956860.2003.11682765URL [本文引用: 3] Although the forest-tundra boundary is likely to be sensitive to future climate warming, the degree to which treeline response may lag climate change and the extent to which sensitivity to climate may vary among sites remain largely unknown. We used tree-ring analysis to reconstruct white spruce (Picea glauca) density from 1800 to present at and beyond the current forest limit at seven altitudinal treeline sites in two regions of interior Alaska. Treeline advance was ubiquitous: cone-bearing spruce are present beyond the current forest limit at all but one site, and tree density has increased at and beyond the forest limit in recent decades at all sites. Increases in stand density were positively correlated with summer temperature at most, but not all, sites. The timing of inferred advances in treeline differed significantly between regions, beginning in the mid- to late 1800s in the White Mountains and in the mid-1900s in the Alaska Range. These differences in the timing of treeline advance may be caused by differences in the rate of forest response to climate or by differences in regional climate history, which remains poorly known. Despite the variation in timing of an advance of treeline, the similarities among sites in the pattern (if not the timing) of change at treeline suggest that recent shifts in the location of the forest-tundra border are a widespread response to recent warming in Alaska. Impact of climate fluctuations on mountain environments in the Canadian Rockies 2000
Response of the central Canadian treeline to recent climatic changes 2 1998
Ads package for R: A fast unbiased implementation of the k-function family for studying spatial point patterns in irregular-shaped sampling windows 1 2015
A century of tree line changes in sub-Arctic Sweden shows local and regional variability and only a minor influence of 20th Century climate warming 1 2011
Treeline dynamics in relation to climatic variability in the central Tianshan Mountains, northwestern China 2006
Spatial patterns of smith fir alpine treelines on the south-eastern Tibetan Plateau support that contingent local conditions drive recent treeline patterns 2 2012
 ,1,3,6,*, 孙建5, 魏彦强7, 谢雨1, 吴彦
,1,3,6,*, 孙建5, 魏彦强7, 谢雨1, 吴彦
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT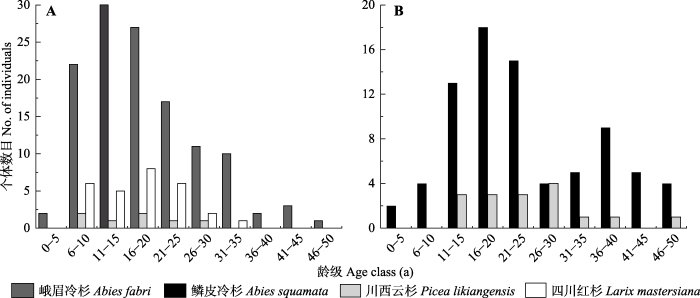 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT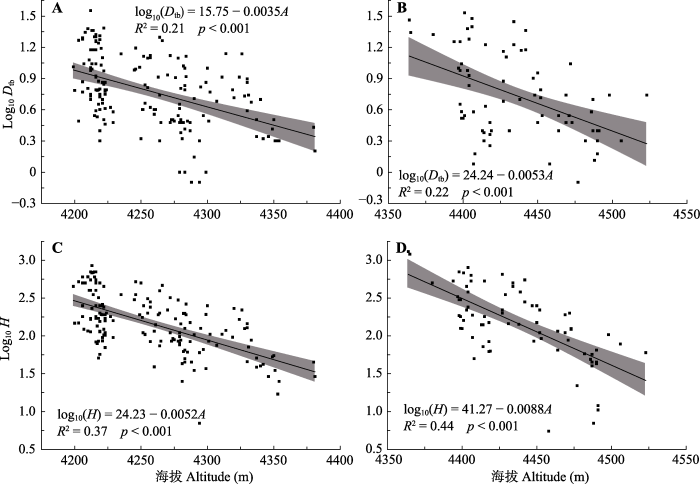 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT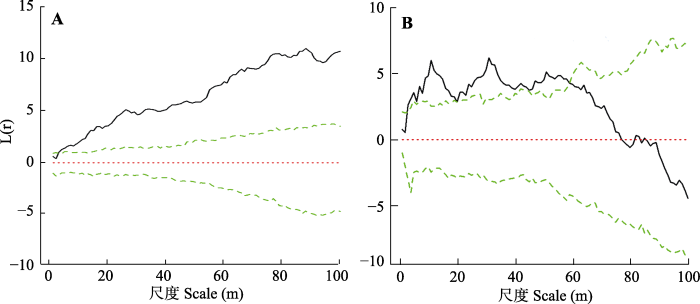 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT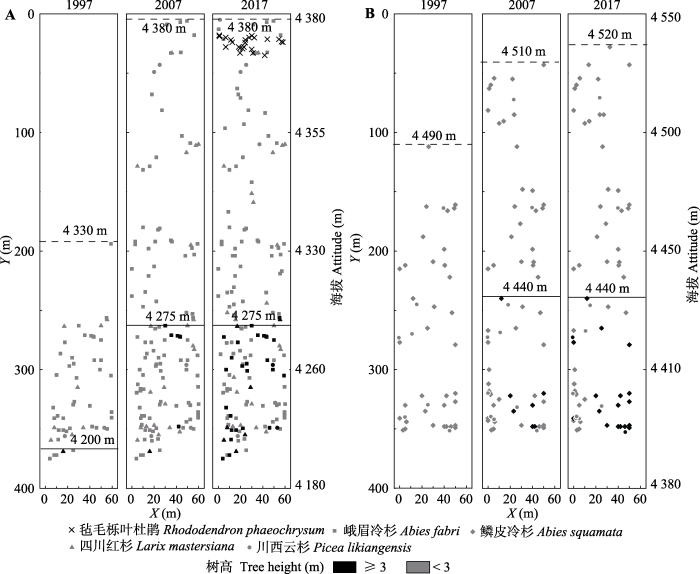 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}