汤丹丹
1 , 2 , 吴毅
1 , 2 , 刘文耀
, 1 , * , 胡涛
1 , 2 , 黄俊彪
1 , 2 , 张婷婷
1 , 2 1
中国科学院西双版纳热带植物园热带森林生态学重点实验室, 云南勐腊 666303 2
中国科学院大学, 北京 100049 Ecological stoichiometry of two common hemiparasite plants and their relationship with host trees in Ailao Mountain, Yunnan, China Dan-Dan TANG
1 , 2 , Yi WU
1 , 2 , Wen-Yao LIU
, 1 , * , Tao HU
1 , 2 , Jun-Biao HUANG
1 , 2 , Ting-Ting ZHANG
1 , 2 1
CAS Key Laboratory of Tropical Forest Ecology, Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, Mengla, Yunnan 666303, China 2
University of Chinese Academy of Sciences, Beijing 100049, China 通讯作者: *
liuwy@xtbg.ac.cn 编委: 吕晓涛
责任编辑: 李敏
收稿日期: 2018-12-7
修回日期: 2019-03-1
网络出版日期: 2019-03-20
基金资助: 国家自然科学基金 (31770496 )国家自然科学基金 (41471050 )中国科学院生物多样性保护策略项目 (ZSSD-016 )中国科学院"135项目" (2017XTBG-T01 )
Received: 2018-12-7
Revised: 2019-03-1
Online: 2019-03-20
Fund supported: Supported by the National Natural Science Foundation of China (31770496 )Supported by the National Natural Science Foundation of China (41471050 )Biodiversity Conservation Strategy Program of Chinese Academy of Sciences (ZSSD-016 )The "135 Program" of Chinese Academy of Sciences (2017XTBG-T01 )
摘要 为深入探究半寄生植物与其寄主间的养分关系, 在云南哀牢山徐家坝地区选取两种常见半寄生植物椆树桑寄生(
Loranthus delavayi )和柳叶钝果寄生(
Taxillus delavayi ), 研究其寄主枝条-吸器-寄生枝条-寄生叶片间的碳(C)、氮(N)、磷(P)生态化学计量特征关系。结果表明: 1)两种半寄生植物的寄主枝条-吸器-寄生枝条-寄生叶片这一连续体的C、N、P生态化学计量特征变化趋势并不完全相同, 具有物种自身的特性。2)同一半寄生植物的寄主枝条具有相似的C、N、P生态化学计量特征, 寄主物种对半寄生植物的生态化学计量特征没有显著影响。3)寄主枝条的C、N、P生态化学计量特征间具有紧密的相关关系, 吸器弱于寄主枝条, 寄生枝条弱于吸器, 寄生叶片的N、P含量相关关系不显著。4)寄主枝条与寄生叶片间的C含量存在极显著负相关关系。5)吸器与寄主枝条间的C、N、P生态化学计量特征存在紧密的相关关系, 但在生态化学计量特征的数值上吸器与寄生枝条更为接近。吸器作为连接寄主与寄生植物的关键部位, 其与寄主枝条极为显著的相关关系体现了寄主枝条养分对寄生植物的重要性, 而吸器在元素含量、计量比的数值以及相互关系上与寄生枝条更为相似, 则体现了吸器作为寄生植物器官具有与寄生枝条相似的生理功能。通过对寄主枝条-吸器-寄生枝条-寄生叶片C、N、P生态化学计量特征的分析, 为深入研究半寄生植物的养分利用策略与生态适应性提供了重要的基础资料。
关键词: 半寄生植物 ;
生态化学计量学 ;
枝条 ;
吸器 ;
叶片 Abstract Aims The objectives of this study were to characterize the carbon (C) : nitrogen (N) : phosphorus (P) stoichiometry of the “host branches-haustorias-parasitic branches-parasitic leaves” continuum and to better understand nutrient relationship between hemiparasite plants and their hosts.
Methods The study site is located in the Xujiaba area of Ailao Mountain, Yunnan Province. Two common hemiparasite plants
Loranthus delavayi and
Taxillus delavayi were selected, and the C, N and P concentrations of host branches, haustorias, parasitic branches and parasitic leaves were measured.
Important findings The results showed that, the tendency of C, N, P stoichiometry characteristics of host branches-haustorias-parasitic branches-parasitic leaves were species specific, and were not identical between the two hemiparasites. The host branches of the same parasitic plant have similar C, N, and P stoichiometry characteristics, and the host species have no significant effect on the stoichiometry of hemiparasites. There was a close coupling relationship between the C, N, P stoichiometry characteristics in the host branches, and the haustorias was weaker than the host branch, the parasitic branch was weaker than the haustorias, and there was no significant correlation between the N and P concentrations in the parasitic leaf. There was a significant negative correlation between the host branches and the parasitic leaves of C concentration. The C, N, P stoichiometry characteristics of the haustorias were more similar to the parasitic branches, and it had a very significant positive correlation with the host branches. As a key part of the host and parasitic plants, the haustorias had a significant correlation with the host branches, which reflected the importance of the host branch nutrients to the parasitic plants. The element stoichiometry and their relationship of the haustorias were more similar to those of the parasitic branches, which embodied that haustorias as a parasitic plant organ had physiological functions similar to those of the parasitic branches. These results provided important data for in-depth study of nutrient utilization strategies and ecological adaptability of hemiparasitic plants.
Keywords: hemiparasite ;
ecological stoichiometry ;
branch ;
haustoria ;
leaf PDF (1252KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 汤丹丹, 吴毅, 刘文耀, 胡涛, 黄俊彪, 张婷婷. 云南哀牢山两种常见半寄生植物的生态化学计量特征及其与寄主的关系. 植物生态学报, 2019, 43(3): 245-257. DOI:
10.17521/cjpe.2018.0311 TANG Dan-Dan, WU Yi, LIU Wen-Yao, HU Tao, HUANG Jun-Biao, ZHANG Ting-Ting.
Ecological stoichiometry of two common hemiparasite plants and their relationship with host trees in Ailao Mountain, Yunnan, China .
Chinese Journal of Plant Ecology , 2019, 43(3): 245-257. DOI:
10.17521/cjpe.2018.0311 生态化学计量学是主要研究生态系统能量平衡和多重化学元素平衡的科学(
Sterner & Elser, 2002 ;
曾德慧和陈广生, 2005 ), 其核心问题是揭示生物体元素组成差异对其生态功能的影响(
Elser et al ., 2000 ;
蒋利玲等, 2014 )。生态化学计量学从元素比率的角度将不同尺度、不同生物群系以及不同领域的生物学研究联系起来(
Sterner & Elser, 2002 ;
Michaels, 2003 ), 成为生态学研究的主要方法和研究热点(
曾冬萍等, 2013 ;
刘文亭等, 2015 )。这一领域研究最多的化学元素是碳(C)、氮(N)、磷(P), 作为3种最重要的生命元素, C、N、P是所有生命过程的基础, 在植物的生化功能方面紧密耦合(
李威, 2016 )。基于不同生命形式, 系统发育以及光合作用碳同化途径的植物叶片生态化学计量特征及其变异模式的研究已取得重大进展(
Han et al ., 2005 ,
2011 ), 但这些研究的主要对象为非寄生植物。有关研究表明, 非寄生植物生态化学计量特征与土壤养分状况相关(
Reich & Oleksyn, 2004 ;
赵一娉等, 2017 ), 但寄生植物没有根系直接接触土壤, 其生命活动所需要的养分和水分主要来自于寄主植物。目前, 有关寄生植物生态化学计量特征的研究鲜有报道。
半寄生植物是寄生植物的主要类群, 其叶片具有叶绿体, 在生命周期的至少一个阶段可进行光合作用, 仅从寄主体内获取部分水和溶解的矿物质(
Nickrent et al ., 1998 )。在自然界中, 半寄生植物对植物群落结构和多样性具有显著影响(
Phoenix & Press, 2010 ;
Press & Phoenix, 2010 ;
Demey et al ., 2014 ), 而其营养丰富、可快速分解和释放养分的凋落物可能会提高森林土壤养分可利用率(
Demey et al ., 2015 ), 影响生态系统水平凋落物分解和养分循环(
Quested et al ., 2010 )。虽然半寄生植物的丰度和生物量相对较小, 但它们对生物地球化学循环、植物种群结构和多样性的影响是很明确的(
Nave et al ., 2017 ), 因而被描述为影响群落动态的生态系统工程师, 以及影响养分循环的关键物种(
Watson, 2010 ;
Hatcher et al ., 2012 )。
半寄生植物与寄主的复杂关系包括寄生和竞争(
Lep? & Tě?itel, 2015 )。作为混合营养策略的代表, 半寄生植物既能从自身的光合作用中获取碳同化物, 也能通过吸器从寄主体内获取碳水化合物(
Nave et al ., 2017 ;
Selosse et al ., 2017 )。半寄生植物自身的光合作用在同化寄生矿物资源, 并将矿物资源转换化成自身生长和适合度中起着至关重要的作用, 而将木质部所获得的资源转化为生长和适应的关键过程, 可能是其进化稳定性的基础(
Tě?itel et al ., 2015 )。尽管半寄生植物与寄主关系的研究开展较早, 不少****对半寄生植物与寄主之间的水分利用、光合作用以及矿物质营养等方面的关系进行了较为深入的研究(
Tennakoon et al ., 2011 ;
Chen et al ., 2013 ;
Suriyagoda et al ., 2017 ;
Gebauer et al ., 2018 ), 但营养物质从寄主到半寄生植物的转移机制仍未完全研究清楚(
Gebauer et al ., 2012 ,
2018 )。有关研究表明, 半寄生植物
Rhinanthus alectorolophus 中5%-40%的C来自寄主, 而寄主体内的N亦可被半寄生植物获取(
Tě?itel et al ., 2015 ;
Světlíková et al ., 2016 )。寄生植物的吸器同时连接寄主枝条与寄生枝条, 研究寄主枝条-吸器-寄生枝条-寄生叶片这一连续体的生态化学计量特征变化将有助于进一步理解寄生植物与寄主间的养分关系。
哀牢山国家级自然保护区分布着我国目前保存最完整、面积最大的亚热带山地湿性常绿阔叶林, 林内附生植物丰富, 也有相当数量的寄生植物(
游承侠, 1983 ;
Young & Herwitz, 1995 ;
朱华和闫丽春, 2009 ), 不少****对该地区森林群落类型及结构、树种、附生和寄生植物多样性、生物量等方面进行了研究(
邱学忠和谢寿昌, 1998 ), 但对该区域不同类群植物包括寄生植物的生态化学计量特征及养分利用的了解甚少。本研究以哀牢山山地湿性常绿阔叶林中常见的半寄生植物为对象, 对寄生植物的寄主枝条-吸器-寄生枝条-寄生叶片生态化学计量特征及其动态进行研究, 以期进一步理解半寄生植物的养分利用策略, 为山地森林资源保护与管理提供科学依据。
1 材料和方法 1.1 研究区概况 哀牢山国家级自然保护区位于云南省中部哀牢山山脉中北段上部, 地处云贵高原、横断山和青藏高原南缘三大地理区域的结合部, 是生物多样性较丰富及植物区系地理成分荟萃之地, 该保护区内分布着我国目前保存最完整、面积最大的亚热带山地湿性常绿阔叶林(
朱华和闫丽春, 2009 )。本研究区位于哀牢山北段的徐家坝地区(23.35°-24.44° N, 100.54°-101.30° E), 该地区海拔2 000-2 750 m, 面积约5 100 hm
2 , 年降水量1 947 mm, 年蒸发量1 192 mm, 相对湿度85%, 年平均气温11.3 ℃ (
李苏等, 2007 )。该地区常绿阔叶林的土壤全C、全N与全P含量分别为98.65、6.53和1.17 mg·g
-1 , C:N、C:P、N:P为14.96、91.22和5.95 (
鲁志云等, 2017 )。徐家坝地区分布有椆树桑寄生(
Loranthus delavayi )、柳叶钝果寄生(
Taxillus delavayi )、卵叶梨果寄生(
Scurrula chingii )、五蕊寄生(
Dendrophthoe pentandra )、卵叶槲寄生(
Viscum album var.
meridianum )等寄生植物(
朱华和闫丽春, 2009 )。
1.2 样地设置与采样方法 2017年11月对哀牢山徐家坝地区的寄生植物进行初步调查, 在爱山亭、茶园、生态站、小坝子以及生态站通往小坝子、老周山、簸箕坝等的路旁, 因一定程度的干扰或处于林缘地带, 森林林冠较为开阔、阳光较充足的地方发现较多的寄生植物。通过采集标本进行鉴定, 这些寄生植物主要为椆树桑寄生与柳叶钝果寄生, 故选取这两种常见的半寄生植物作为研究对象。基于这两种半寄生植物的分布特点以及调查地实际情况, 本研究采用样带调查法, 在上述区域设置面积为10 m × 10 m-20 m × 4 000 m的样带进行采样(
王煊妮和张玲, 2017 )。
采样时间为2018年3月下旬至4月上旬。采样时一致选取寄主胸径以上、林冠外层朝南方向生长健康的寄生植株, 于吸器往寄主根部方向约10 cm处用高枝剪将寄主枝条和整株寄生植物剪下, 再于吸器往寄生叶片方向约10 cm处用枝剪将寄生枝条剪断, 并收集健康、完整的寄生植物叶片。最后用枝剪、刀斧、锯子等工具将寄主枝条、寄生枝条与吸器分开。本次采样共收集椆树桑寄生样品(寄主枝条-吸器-寄生枝条-寄生叶片为一个样品) 27个, 柳叶钝果寄生样品31个。研究区中椆树桑寄生对寄主有较强的选择性, 本次采样仅发现景东石栎(
Lithocarpus chintungensis )、顺宁厚叶柯(
L. pachyphyllus var.
fruticosus )和硬叶柯(
L. crassifolius ) 3种寄主, 均为壳斗科植物, 其中硬叶柯上采集的样品数为2个, 顺宁厚叶柯为7个, 其余样品均采集于景东石栎上。柳叶钝果寄生的寄主有山矾(
Symplocos sumuntia )、直角荚蒾(
Viburnum foetidum var.
rectangulatum )、地檀香(
Gaultheria forrestii )、花椒(
Zanthoxylum bungeanum )、桃(
Amygdalus persica ), 但这几个寄主上采集的样品数均小于4个; 寄主云南越桔(
Vaccinium duclouxii )、紫药女贞(
Ligustrum delavayanum )上采集的样品数各为4个, 露珠杜鹃(
Rhododendron irroratum )为5个, 其余样品均采集于厚皮香(
Ternstroemia gymnanthera )上。由于寄主树的优势度不同, 为将每种寄主树的样品重复数保持基本一致, 并将尽可能多的寄主包含到研究中以分析寄主物种不同对寄生植物生态化学计量的影响, 本研究将样品数大于4的寄主作为优势寄主, 每种寄主选取样本4-5个重复用于元素测量与数据分析。
1.3 样品处理 将采集到的植物叶片用自来水洗净灰尘等杂质后, 再用超纯水冲洗。由于半寄生植物的大部分矿物质是通过吸器从寄主木质部获取(
Glatzel, 1983 ;
Bell & Adams, 2011 ), 故将寄主枝条的树皮去除后作为半寄生植物的养分来源, 以便更准确地分析寄主枝条-吸器-寄生枝条-寄生叶片的生态化学计量特征变化。将寄主枝条、吸器、寄生枝条与叶片分别装入信封, 于65 ℃条件下烘干至恒质量。烘干后先用刀斧将吸器及寄主枝条、寄生枝条粗加工成小块, 再通过微型植物粉碎机(FZ102型, 天津市泰斯特仪器有限公司, 天津)将样品磨成粉末, 植物叶片则利用高速万能粉碎机(FW80型, 天津市泰斯特仪器有限公司, 天津)磨成粉末, 用于全碳、全氮和全磷含量的测定。C、N含量用碳氮分析仪(Vario MAX CN, Elementar Analysensysteme GmbH, Hanau, Germany)测定, P含量采用电感耦合等离子体原子发射光谱仪(iCAP6300, Thermo Fisher Scientific, Waltham, USA)测定。
1.4 数据处理 采用SPSS 21.0对数据进行整理与分析, 利用SigmaPlot 12.5制图。对寄主枝条、吸器、寄生枝条和寄生叶片间的全碳、全氮和全磷含量(质量含量, 下同)及其化学计量比(质量比, 下同)的差异显著性进行单因素方差分析, 对柳叶钝果寄生4种寄主枝条间全碳、全氮和全磷含量及其化学计量比的差异显著性进行单因素方差分析, 椆树桑寄生2种寄主枝条间的差异则采用独立样本
t 检验进行分析。对椆树桑寄生同一器官C、N、P生态化学计量特征在2种寄主情况下的差异进行独立样本
t 检验。对椆树桑寄生同一器官C、N、P生态化学计量特征在4种寄主情况下的差异进行单因素方差分析。对同一寄生植物不同器官C、N、P生态化学计量特征在不同寄主之间的差异进行双因素方差分析。对寄主枝条、吸器、寄生枝条和寄生叶片4种不同器官的同一元素含量、化学计量比的关系以及4种器官各自的元素含量、化学计量比的相关关系进行Pearson相关性分析。N、P间的计量关系利用lg
Y =
α (lg
X ) +
β 进行拟合, 其中
X 和
Y 代表叶P和N浓度,
α 和
β 分别是回归线斜率和截距。为方便表达, 全碳、全氮、全磷简称为C、N、P, 寄主枝条、寄生枝条、寄生叶片简称为寄主枝、寄生枝与寄生叶。
2 结果和分析 2.1 寄主枝-吸器-寄生枝-寄生叶的C、N、P含量及其化学计量比 寄主枝-吸器-寄生枝-寄生叶这一连续体的C、N、P生态化学计量特征的变化趋势在两种半寄生植物中并不完全相同, 但在同一寄生植物中是一致的(
图1 )。两种半寄生植物N含量从寄主枝到寄生叶总体上都呈现上升趋势(
图1 B), C:N则总体上呈现下降趋势(
图1 D)。C含量在寄主枝、吸器、寄生枝与寄生叶中总体呈现先增加后下降的趋势, 但柳叶钝果寄生叶片C含量的下降趋势强于椆树桑寄生(
图1 A)。椆树桑寄生的P含量呈现先下降再上升的趋势, 柳叶钝果寄生则保持上升趋势(
图1 C)。椆树桑寄生C:P在寄主枝-吸器-寄生枝中变化不大, 在寄生叶中降低, 柳叶钝果寄生则总体上保持下降趋势(
图1 E)。两种半寄生植物的N:P变化规律皆不明显(
图1 F)。椆树桑寄生的两种寄主枝之间, 仅C:N差异显著(
p < 0.05)。柳叶钝果寄生的寄主露珠杜鹃、云南越桔以及紫药女贞两两间也仅紫药女贞的C含量显著低于另外两个寄主(
p < 0.05)。寄主厚皮香的生态化学计量特征与另外3种寄主的差异相对较大, 其P含量显著高于其他3个寄主(
p < 0.05), 而N:P显著低于其他3个寄主(
p < 0.05)。此外, 厚皮香的C含量高于紫药女贞(
p < 0.05), N含量低于、C:N高于露珠杜鹃(
p < 0.05), C:P则显著低于露珠杜鹃与云南越桔(
p < 0.05)。柳叶钝果寄生同一器官的C、N、P生态化学计量特征在4种不同寄主情况下均没有显著差异(
p > 0.05); 椆树桑寄生同一器官的C、N、P生态化学计量特征在2种不同寄主情况下仅枝条的C:P、N:P存在显著差异(
p < 0.05), 生长于景东石栎上的寄生枝条C:P、N:P高于生长在顺宁厚叶柯上的。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1C、N、P生态化学计量特征在哀牢山两种常见半寄生植物寄主枝-吸器-寄生枝-寄生叶中的变化(平均值±标准误差)。a, 寄主枝; b, 吸器; c, 寄生枝; d, 寄生叶。 Fig. 1Changes of C, N, P concentrations and stoichiometry in the host branches, haustorias, parasitic branches, parasitic leaves in two common hemiparasites in Ailao Mountain (mean ± SE ). a, host branch; b, haustorias; c, parasitic branches; d, parasitic leaves. 对同一寄生植物不同器官C、N、P生态化学计量特征在不同寄主之间的差异进行双因素方差分析的结果(
表1 )表明: 在柳叶钝果寄生中, 其寄主物种对寄生植物C、N、P生态化学计量特征没有显著影响(
p > 0.05), 而不同器官的影响是极显著的(
p < 0.001); 在椆树桑寄生中, 其寄主物种仅对C含量具有显著影响(
p < 0.05), 而器官仅对C含量与N:P的影响不显著(
p > 0.05)。值得注意的是, 椆树桑寄生中C含量与N:P的
R 2 较小。两种寄生植物的器官与寄主物种的交互作用对寄生植物C、N、P生态化学计量特征没有显著影响(
p > 0.05)。
Table 1 表1 表1 哀牢山同一寄生植物不同器官C、N、P生态化学计量特征在不同寄主之间的差异
Table 1
Differences in the stoichiometric characteristics of C, N, and P in different organs of the same parasitic plant between different hosts in Ailao Mountain 变量 椆树桑寄生 Loranthus delavayi 柳叶钝果寄生 Taxillus delavayi 物种 Species 器官 Organ 物种×器官 Species × Organ R 2 物种 Species 器官 Organ 物种×器官 Species × Organ R 2 C * ns ns 0.336 ns *** ns 0.863 N ns *** ns 0.657 ns *** ns 0.445 P ns *** ns 0.526 ns *** ns 0.747 C:N ns *** ns 0.712 ns *** ns 0.586 C:P ns *** ns 0.663 ns *** ns 0.826 N:P ns ns ns 0.211 ns *** ns 0.730
ns, no significant. ***, p < 0.001; **, p < 0.01; *, p < 0.05; ns, 不显著。 新窗口打开 |
下载CSV 吸器的C、N、P生态化学计量特征与寄主枝的差异较大, 而与寄生枝的差异较小(
表2 ): 吸器与寄主枝间, 椆树桑寄生仅P含量和C:P差异不显著(
p > 0.05), 柳叶钝果寄生也仅N:P差异不显著(
p > 0.05)。吸器与寄生枝间, 椆树桑寄生仅C含量差异显著(
p < 0.05), 柳叶钝果寄生仅N:P存在显著差异(
p < 0.05)。寄生叶的N、P含量较高, 除椆树桑寄生的寄主枝与寄生叶间P含量差异不显著(
p > 0.05)外, 两种寄生植物叶片中的N、P含量皆显著高于其寄主枝、吸器与寄生枝(
p < 0.05)。椆树桑寄生叶片的C含量与N:P高于寄主枝, 柳叶钝果寄生叶片的C含量与N:P却低于寄主枝。两种半寄生植物寄主枝、吸器、寄生枝与寄生叶中C含量都较稳定, 变异系数均小于3%, N含量的变异系数均小于P含量, C:N的变异系数也都小于C:P。两种半寄生植物的N:P的变异系数同为吸器<寄生枝<寄主枝<寄生叶。
Table 2 Table 2 C, N and P stoichiometry characteristics of host branches, haustorias, parasitic branches, parasitic leaves in two common hemiparasites in Ailao Mountain
平均值 Mean (mg·g-1 ) 变异系数 Coefficient of variation (%) C N P C:N C:P N:P C N P C:N C:P N:P 椆树桑寄生Loranthus 寄主枝 Host branch 449.90c 4.01c 1.55ab 113.55a 345.09ab 3.01b 0.78 11.95 35.38 11.39 55.68 50.37 吸器 Haustoria 474.80b 9.15b 1.18b 52.23b 410.69a 7.89a 2.04 8.22 13.11 9.31 14.80 14.89 寄生枝 Parasitic branch 486.10a 8.90b 1.03b 56.34b 503.09a 9.08a 2.14 20.55 25.82 17.28 26.25 25.43 寄生叶 Parasitic leaf 478.00ab 15.65a 2.90a 31.85c 220.41b 7.20ab 2.76 24.57 56.37 19.60 55.53 55.33 柳叶钝果寄生Taxillus 寄主枝 Host branch 460.28c 2.64c 0.32c 184.03a 1 618.57a 9.37ab 1.60 24.17 39.11 24.54 29.85 35.49 吸器 Haustoria 477.17a 7.67b 0.73b 63.47b 679.07b 10.74a 2.17 15.76 18.88 13.52 19.79 16.11 寄生枝 Parasitic branch 483.50a 7.65b 0.94b 64.18b 564.00b 8.77b 1.39 12.87 33.30 12.19 28.24 25.40 寄生叶 Parasitic leaf 431.61c 10.10a 3.16a 45.82c 158.13c 3.76c 2.98 27.51 37.24 26.87 44.33 58.43 总体 寄主枝 Host branch 456.57b 3.13c 0.76c 158.86a 1 163.70a 7.10cbd 1.75 28.19 90.78 63.46 58.70 31.62 吸器 Haustoria 476.32a 8.20b 0.89b 59.46b 583.22b 9.72a 2.10 15.59 29.32 15.45 29.58 21.25 寄生枝 Parasitic branch 484.43a 8.10b 0.97b 61.38b 542.25b 8.88ab 1.68 17.91 30.38 15.00 27.77 24.99 寄生叶 Parasitic leaf 448.18bc 12.08a 3.06a 40.83c 180.37c 4.99d 5.79 34.24 43.48 30.48 52.63 66.97
同一列同一物种不同小写字母表示差异显著(p < 0.05)。p < 0.05). 新窗口打开 |
下载CSV 2.2 寄主枝、吸器、寄生枝、寄生叶内C、N、P生态化学计量特征的相关关系 寄主枝内元素含量及计量比间的相关关系最为紧密, 吸器内的相关性弱于寄主枝, 其C含量与N、P含量间的相关关系均不显著(
p > 0.05); 寄生枝又弱于吸器, 其C含量与N:P的关系也不显著(
p > 0.05); 寄生叶的特征与前三种器官不甚相同, 仅C和N含量、C含量和N:P显著相关(
p < 0.01)(
表3 )。值得注意的是, N和P含量的相关性在寄主枝与吸器中均为极显著正相关(
p < 0.01), 寄生枝中为显著相关(
p < 0.05), 而寄生叶中为不显著的负相关关系(
p > 0.05)。N、P计量关系的回归斜率在寄主枝、吸器、寄生枝中为0.218、0.370、0.223。寄生叶中N、P计量关系不显著(
p = 0.204) (
图2 )。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2哀牢山两种常见半寄生植物寄主枝、吸器、寄生枝、寄生叶的N-P计量关系。 Fig. 2N-P scaling of host branches, haustorias, parasitic branches and parasitic leaves in two common hemiparasites in Ailao Mountain. Table 3 表3 表3 哀牢山两种常见半寄生植物寄主枝、吸器、寄生枝、寄生叶内C、N、P生态化学计量特征的相关关系
Table 3
Correlation relationship of C, N, P stoichiometry of the host branches, haustorias, parasitic branches and parasitic leaves in two common hemiparasites in Ailao Mountain 组成 Composition 寄主枝 Host branch 吸器 Haustoria 寄生枝 Parasitic branch 寄生叶 Parasitic leaf C和N C and N -0.496** 0.080 0.221 0.595** C和P C and P -0.588** -0.243 -0.126 -0.259 C和N:P C and N:P 0.477* 0.462* 0.242 0.621** N和P N and P 0.716** 0.662** 0.419* -0.257 N和C:P N and C:P -0.619** -0.666** -0.388* 0.285 P和C:N P and C:N -0.599** -0.722** -0.402* 0.196
**, p < 0.01; *, p < 0.05。 新窗口打开 |
下载CSV 2.3 寄主枝、吸器、寄生枝、寄生叶间C、N、P生态化学计量特征的相关关系 由
表4 可知, 寄主枝与吸器间的元素含量及化学计量比具有紧密的相关关系, 两者间仅C含量相关关系未达到显著水平(
p > 0.05), N、P含量以及C:N、C:P、N:P皆为极显著正相关(
p < 0.01); 寄主枝与寄生枝间仅N含量极显著正相关(
p < 0.01), C:N显著正相关(
p < 0.05); 与寄生叶间的N含量显著正相关(
p < 0.05)、C含量极显著负相关(
p < 0.01)。吸器与寄生枝间的C含量、C:P显著正相关(
p < 0.05), P含量极显著正相关(
p < 0.01); 与寄生叶则C、N、P含量及C:N、C:P、N:P皆未达到显著水平(
p > 0.05)。寄生枝与寄生叶间仅N含量和C:N极显著正相关(
p < 0.01)。
Table 4 表4 表4 哀牢山两种常见半寄生植物寄主枝、吸器、寄生枝、寄生叶间C、N、P生态化学计量特征的相关关系
Table 4
Correlation between host branches, haustorias, parasitic branches, and parasitic leaves of C, N, and P stoichiometry in two common hemiparasites in Ailao Mountain 组成 Composition C N P C:N C:P N:P 寄主枝与吸器 Host branch and haustoria 0.163 0.508** 0.772** 0.589** 0. 784** 0.731** 寄主枝与寄生枝 Host branch and parasitic branch 0.142 0.502** 0.154 0.470* 0.178 -0.075 寄主枝与寄生叶 Parasitic branch and parasitic leaf -0.636** 0.409* 0.160 0.289 0.123 -0.134 吸器与寄生枝 Haustoria and parasitic branch 0. 460* 0.216 0.489** 0.184 0.452* 0.289 吸器与寄生叶 Haustoria and parasitic leaf -0.160 0.296 -0.048 0.186 -0.157 -0.133 寄生枝与寄生叶 Parasitic branch and parasitic leaf 0.033 0.674** 0.168 0.673** 0.134 0.371
**, p < 0.01; *, p < 0.05。 新窗口打开 |
下载CSV 3 讨论 3.1 寄主枝-吸器-寄生枝-寄生叶的C、N、P含量及其化学计量比 寄主枝-吸器-寄生枝-寄生叶这一连续体中的C、N、P生态化学计量特征的变化趋势, 在同一寄生植物中一致, 在不同种寄生植物中并不完全相同, 差异主要体现在寄主枝和寄生叶中。椆树桑寄生叶片C含量和N:P高于寄主枝, 柳叶钝果寄生则相反,这一现象可能与两种寄生植物自身生长发育阶段的不同有关。C元素是植物各种生理生化过程的底物和能量来源, 其同化与N、P元素的吸收途径不同, 随生长阶段的变化, C、N、P元素的分配比例也有较大的差别(
贡璐等, 2017 )。本研究采样时, 哀牢山地区椆树桑寄生处于营养生长阶段, 而柳叶钝果寄生已经进入开花期。两种寄生植物各自的寄主枝具有更相似的C、N、P生态化学计量特征, 这有可能是寄生植物对养分浓度有一定的需求和偏好, 从而养分含量相似的物种易被同一种寄生植物侵染。该区椆树桑寄生的寄主皆为壳斗科植物, 亲缘关系上的相近可能是其寄主枝C、N、P生态化学计量特征相似的原因之一。柳叶钝果寄生的寄主厚皮香、云南越桔以及紫药女贞为不同科属植物, 其生态化学计量特征相似的原因需要进一步研究。
大多数寄生植物具有广寄性, 而不同的寄生植物其寄主种类有很大的差异(
Norton & Lange, 1999 ;
胡飞和孔垂华, 2004 )。本次调查记录中椆树桑寄生的寄主均为壳斗科植物, 柳叶钝果寄生的寄主较为广泛, 但未发现与椆树桑寄生具有相同寄主。不同的寄主物种对同一寄生植物的影响可能不同, 有研究发现同一桑寄生植物的挥发性成分组成及含量、总黄酮含量、毒性等会因其寄主物种不同而不同(
李永华等, 2009 ,
2012 ;
周汉华等, 2013 ), 但对寄生植物生态化学计量特征与寄主物种的关系进行探讨的研究较少。在本研究中, 寄生于不同寄主的同一寄生植物的同一器官C、N、P生态化学计量特征基本没有显著的不同, 而对同一寄生植物的不同寄主情况下不同器官之间的生态化学计量特征进行双因素方差分析, 也发现寄主物种的影响不显著。这可能与同一寄生植物的不同寄主枝条本身生态化学计量特征差异不显著有关。
寄生植物不同的营养素富集程度也不同, 被认为是吸器选择性吸收离子的结果, 若摄取是通过蒸腾而被动吸收, 那么营养元素的比值应保持相同(
Gebauer et al ., 2018 )。本研究中, 椆树桑寄生吸器的C:N、C:P及N:P均与其寄主枝条存在显著差异, 柳叶钝果寄生吸器的C:N和C:P也与其寄主枝条存在显著差异, 这在一定程度上说明C、N和P元素并非简单的通过蒸腾作用而被转运至寄生植物体内, 但也不能排除离子通过被动运输进入吸器后, 由于吸器具有合成物质的功能而导致元素比值发生改变(
黄新亚等, 2011 )。植物不同器官C、N、P含量的差异不仅受植物基本生理过程需求的影响, 也受相应器官组织结构和功能分化的影响(
Minden & Kleyer, 2015 )。叶片是植物的同化器官与重要的养分储存器官, 因而寄生植物叶片的N和P含量高于寄主枝、吸器与寄生枝(
Niklas & Cobb, 2005 ;
贺合亮等, 2017 )。植物叶片中C、N、P含量的变异系数普遍存在C < N < P的规律(
Koerselman & Meuleman, 1996 ;
Güsewell & Koerselman, 2002 ;
姜沛沛等, 2017 )。本研究中两种半寄生植物的寄主枝、吸器、寄生枝与寄生叶的C、N、P含量皆符合这个规律, 这是因为C作为构成植物骨架的基本结构物质, 且为多种植物生理活动提供能源, 而N含量可被植物控制在较窄的范围内, 故C在植物体内含量高且变异小, N含量在各器官中的变异性小于P含量(
Vitousek, 1998 ;
贺合亮等, 2017 )。本研究中两种寄生植物C:N的变异性小于C:P,是因为植物光合作用中C的同化与营养元素的吸收并非同一途径, 而C含量高且变异小, C:N和C:P的变异主要受到N和P含量变化的影响(
Hedin, 2004 ;
曾昭霞等, 2015 )。
3.2 寄主枝、吸器、寄生枝、寄生叶内C、N、P生态化学计量特征的相关性 植物体中C与N、P含量一般呈负相关关系是高等陆生植物C、N、P计量的普遍规律(
Sterner & Elser, 2002 ;
贺合亮等, 2017 )。在本研究中, 仅寄主枝符合此规律, 其C含量与N、P含量及C:N、C:P、N:P均存在极显著的相关关系。寄生植物吸器的C含量仅与N:P具有显著的相关关系, 枝条的C含量则与N、P含量及C:N、C:P、N:P均不存在极显著的相关关系。寄生植物叶片的C含量与N含量为正相关关系, 与P含量的相关性未达到显著, 也不同于此规律, 但该结果与荒漠植物叶片C、N、P生态化学计量特征具有一致性(
张珂等, 2014 ;
罗艳等, 2017 )。陆生植物的C主要来自大气中的CO
2 (
廖宗文等, 2014 ), 半寄生植物的C既可以来自自身光合作用, 也可以从寄主处获取(
Tě?itel et al ., 2015 )。规律不同说明半寄生植物在固C过程中对养分(N、P等)的利用策略有别于一般陆生植物。
N和P在植物中为协同元素, 植物叶片N、P含量的正相关关系亦为高等陆生植物养分计量的普遍规律(
Sterner & Elser, 2002 ;
Wright et al ., 2005 )。植物N、P间的耦合关系很早就有文献记录(
Garten, 1976 ), 随后也得到许多研究的验证(
Li et al ., 2014 ;
Hidaka & Kitayama, 2015 ;
崔高阳等, 2015 )。对中国753种陆地植物叶片的研究(
Han et al ., 2005 )以及中国东部南北样带654种植物叶片的研究(
任书杰等, 2007 )表明, 不同生活型或不同系统发育植物的叶片N和P含量大部分也呈极显著相关关系。水生植物N、P含量间同样具有强线性关系, 这可能是大多数植物N、P含量的基本特征(
Duarte, 1992 )。
Hidaka和Kitayama (2015) 认为植物叶片N、P含量间的正相关关系可能部分受到土壤可利用性氮含量的影响, 而植物营养策略(N利用策略受到P利用策略的影响)以及植物细胞中的N和P生态化学计量特征(如蛋白质合成中N和P的生化关系)则可提供另一种解释, 但仍需要进一步分析以阐明相关性的生理背景。本研究中, N和P由寄主枝、吸器中的极显著正相关关系, 寄生枝中显著正相关关系, 到寄生叶中变为了相关关系不显著。
Garten (1976) 的研究中大型水生植物的N和P含量相关性也不显著, 这可能是大型水生植物的N含量范围较小, 或者是当环境P缺乏时对N的奢侈消费造成的。
Han等(2005) 和
任书杰等(2007) 的研究中, 裸子植物和蕨类植物的N、P含量相关性也不显著。本研究中寄生叶N、P间的不协同性可能与寄生植物不直接从土壤吸收养分、混合营养策略以及寄生植物叶片重吸收P但不重吸收N有关(
Suriyagoda et al ., 2017 )。受到N、P不耦合的影响, 寄生叶的N含量和C:P, P含量和C:N也不存在显著的相关关系。
N、P在植物叶片中的关系可以通过幂函数
N =
βPα 进行量化(
Wright et al ., 2004 ;
Reich et al ., 2010 ;
Yan et al ., 2018 )。
Wright等(2004) 基于全球范围内广泛的叶片N和P浓度数据, 认为N对P的幂指数约为2/3,
Niklas (2006) 的文章中该指数为0.73, 接近3/4而不是2/3,
Reich等(2010) 收集了更为全面的叶片N、P浓度数据, 提出了叶片N、P浓度计量的2/3幂法则(
N =
αP 2/3 )。目前已有研究表明, 植物叶片N、P计量关系并非恒定, 而是受到植物功能群、纬度区、生态区、施肥类型与水平等多种因素影响(
Tian et al ., 2018 ;
Yan et al ., 2018 )。在本研究中, 寄主枝、吸器与寄生枝的N、P计量关系的回归斜率均小于3/4与2/3, 尽管叶片N、P的计量关系有一些趋势但没有统计学意义。以往的研究主要基于植物叶片的数据, 结果是否适用于不同器官有待验证。
寄生植物是一类重要而独特的植物类群。本研究中两种半寄生植物叶片的平均N含量为12.08 mg·g
-1 , 明显低于全球陆生植物的N含量20.09 mg·g
-1 (
Reich & Oleksyn, 2004 )、中国陆生植物的N含量18.6 mg·g
-1 (
Han et al ., 2005 ); 而P含量3.06 mg·g
-1 则明显地高于全球和中国陆生植物的P含量, 分别为1.77 mg·g
-1 (
Reich & Oleksyn, 2004 )、1.21 mg·g
-1 (
Han et al ., 2005 )。寄生叶的N含量较低可能与其光合作用较弱有关, 而较高的P含量可能受到研究区土壤P含量的间接影响, 哀牢山地区森林土壤的全P和有效P含量分别为1.17 g·kg
-1 和3.40 mg·kg
-1 (
鲁志云等, 2017 ), 中国土壤全P和有效P含量分别为0.60 g·kg
-1 和2.39 mg·kg
-1 (
汪涛等, 2008 ), 哀牢山地区森林土壤全P含量显著高于全国水平(
鲁志云等, 2017 )。此外, 哀牢山地区山地森林表层土壤(0-20 cm)的C:N、C:P、N:P值分别为14.9、84.9、5.7, 高于中国陆地(0-10 cm)土壤的C:N、C:P、N:P水平(分别为12.3、52.7、3.9)(
Tian et al ., 2010 ), 更接近于全球陆地(0-10 cm)和全球森林土壤水平(分别为12.3、72.0、5.9和12.4、81.9、6.6)(
Cleveland & Liptzin, 2007 )。已有的研究认为我国陆地植物生长普遍受P限制(
Han et al ., 2005 ;
任书杰等, 2007 ), 但哀牢山地区较高的土壤P含量暗示该地区可能并不缺P。关于哀牢山地区森林寄生植物、寄主的生态化学计量特征与土壤化学元素计量的关系将是以后研究的重要内容。
3.3 寄主枝条、吸器、寄生枝条、寄生叶片间C、N、P生态化学计量特征的相关性 寄生植物与其寄主之间的养分相关性蕴含了重要信息, 这些信息尚未被充分理解(
Gebauer et al ., 2018 )。寄主植物中的元素浓度通常是用来预测半寄生植物体内元素浓度的最佳指标(
Glatzel & Geils, 2009 ;
Gebauer et al ., 2012 )。本研究中寄主枝与吸器之间的N、P含量以及C:N、C:P和N:P均呈极显著正相关关系, 说明寄生植物吸器与寄主枝存在非常密切的关系, 但寄主枝与寄生枝间仅N含量和C:N存在显著相关关系, 寄主枝与寄生叶间仅C、N含量显著相关。这说明寄主枝的C、N、P浓度并非同时适用于预测寄生植物不同器官的C、N、P浓度, 如寄主枝的C浓度可能仅适用于预测寄生叶的C浓度。此外, 寄主枝、吸器、寄生枝与寄生叶两两器官间, 仅寄主枝C含量与寄生叶C含量为极显著负相关关系, 说明寄主枝C含量对寄生叶的形成以及生理活动的正常进行可能有着不可或缺的作用。寄主枝与吸器、吸器与寄生枝间P含量极显著正相关, 说明寄主枝、吸器和寄生枝对P的吸收具有一致性(
何苗等, 2015 )。
大多数有关寄生植物的研究主要关注于植物的光合器官, 而对寄生植物不同器官与寄主植物间的关系研究较少。有关研究表明, 植物叶片N、P浓度与枝条N、P浓度具有相关性(
Kerkhoff et al ., 2006 ), 根和叶间的N浓度也具有显著的相关性(
Craine et al ., 2005 )。本研究寄生枝与寄生叶之间的P含量没有呈现出显著的相关性, 但N含量存在极显著正相关关系, 该结果与
杨梅等(2015) 、
胡耀升等(2014) 的研究果相一致。叶和茎间N含量的相关关系反映了叶、茎在生物、生化方面的功能需求, 这是由植株冠层结构所决定的(
Westoby et al ., 2002 ;
胡耀升等, 2014 )。
4 结论 在云南哀牢山徐家坝地区山地森林中, 椆树桑寄生(
Loranthus delavayi )和柳叶钝果寄生(
Taxillus delavayi )两种常见半寄生植物寄主枝-吸器-寄生枝-寄生叶这一连续体的C、N、P生态化学计量特征的变化趋势有所不同, 同一物种半寄生植物的寄主枝条具有相似的C、N、P生态化学计量特征。寄主对半寄生植物的生态化学计量特征没有显著的影响, 半寄生植物的吸器、枝条与叶片C、N、P含量的变异系数符合C < N < P的一般规律, 而C、N、P的生态化学计量特征关系又不同于一般的陆生植物, 尤其是叶片的N、P含量间并不存在显著的相关关系。寄生植物吸器的C、N、P生态化学计量特征的数值与寄生枝条相似, 吸器的C、N、P生态化学计量特征与寄主枝条存在紧密的相关关系; 寄主枝条与寄生叶片之间的C含量存在极显著的负相关关系。本研究表明半寄生植物既具有普通陆生植物生态化学计量的一般特性, 又具有自身独特的特征。
致谢 感谢云南哀牢山森林生态系统国家野外科学观测研究站全体工作人员在本研究中提供的帮助, 感谢李苏副研究员对本研究数据处理部分的指导。
[1] Bell TL Adams MA 2011 ). Attack on all fronts: Functional relationships between aerial and root parasitic plants and their woody hosts and consequences for ecosystemsTree Physiology 3 -15 . [本文引用: 1] [2] Chen L Huang L Li X You S Yang S Zhang Y Wang W 2013 ). Water and nutrient relationships between a mistletoe and its mangrove host under saline conditionsFunctional Plant Biology 475 -483 . [本文引用: 1] [3] Cleveland CC Liptzin D 2007 ). C:N:P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomassBiogeochemistry 235 -252 . [本文引用: 1] [4] Craine JM Lee WG Bond WJ Williams RJ Johnson LC 2005 ). Environmental constraints on a global relationship among leaf and root traits of grassesEcology 12 -19 . [本文引用: 1] [5] Cui GY Cao Y Chen YM 2015 ). Characteristics of nitrogen and phosphorus stoichiometry across components of forest ecosystem in Shaanxi ProvinceChinese Journal of Plant Ecology 1146 -1155 . [本文引用: 1] [ 崔高阳 , 曹扬 , 陈云明 ( 2015 ). 陕西省森林各生态系统组分氮磷化学计量特征植物生态学报 , 39, 1146 -1155 .] [本文引用: 1] [6] Demey A de Frenne P Baeten L Verstraeten G Hermy M Boeckx P Verheyen K 2015 ). The effects of hemiparasitic plant removal on community structure and seedling establishment in semi-natural grasslandsJournal of Vegetation Science 409 -420 . [本文引用: 1] [7] Demey A Rütting T Huygens D Staelens J Hermy M Verheyen K Boeckx P 2014 ). Hemiparasitic litter additions alter gross nitrogen turnover in temperate semi-natural grassland soilsSoil Biology & Biochemistry 419 -428 . [本文引用: 1] [8] Duarte CM 1992 ). Nutrient concentration of aquatic plants: Patterns across speciesLimnology and Oceanography 882 -889 . [本文引用: 1] [9] Elser JJ Fagan WF Denno RF Dobberfuhl DR Folarin A Huberty A Interlandi S Kilham SS Mccauley E Schulz KL 2000 ). Nutritional constraints in terrestrial and freshwater food websNature 578 -580 . [本文引用: 1] [10] Garten CT 1976 ). Correlations between concentrations of elements in plantsNature 686 -688 . [本文引用: 2] [11] Gebauer R Vola?ík D Urban J 2012 ). Quercus pubescens and its hemiparasite Loranthus europaeus: Nutrient dynamics of leaves and twigsActa Physiologiae Plantarum 1801 -1809 . [本文引用: 2] [12] Gebauer R Vola?ík D Urban J 2018 ). Seasonal variations of sulphur, phosphorus and magnesium in the leaves and current-year twigs of hemiparasitic mistletoe Loranthus europaeus Jacq. and its host Quercus pubescens WilldJournal of Forest Science 66 -73 . [本文引用: 4] [13] Glatzel G 1983 ). Mineral nutrition and water relations of hemiparasitic mistletoes: A question of partitioning. Experiments with Loranthus europaeus on Quercus petraea and Quercus roburOecologia 193 -201 . [本文引用: 1] [14] Glatzel G Geils BW 2009 ). Mistletoe ecophysiology: Host- parasite interactionsBotany 10 -15 . [本文引用: 1] [15] Gong L Li HL Liu YT An SQ 2017 ). Effects of nitrogen and phosphorus fertilizers on carbon, nitrogen, and phosphorus stoichiometry of oasis cotton in the upper reaches of Tarim River, Xinjiang, ChinaActa Ecologica Sinica 7689 -7697 . [本文引用: 1] [ 贡璐 , 李红林 , 刘雨桐 , 安申群 ( 2017 ). N、P施肥对塔里木河上游绿洲棉花C、N、P生态化学计量特征的影响生态学报 , 37, 7689 -7697 .] [本文引用: 1] [16] Güsewell S Koerselman W 2002 ). Variation in nitrogen and phosphorus concentrations of wetland plantsPerspectives in Plant Ecology Evolution and Systematics 37 -61 . [本文引用: 1] [17] Han WX Fang JY Guo DL Zhang Y 2005 ). Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in ChinaNew Phytologist 377 -385 . [本文引用: 6] [18] Han WX Fang JY Reich PB Woodward FI Wang ZH 2011 ). Biogeography and variability of eleven mineral elements in plant leaves across gradients of climate, soil and plant functional type in ChinaEcology Letters 788 -796 . [本文引用: 1] [19] Hatcher M Dick JTA Dunn AM 2012 ). Diverse effects of parasites in ecosystems: Linking interdependent processesFrontiers in Ecology and the Environment 186 -194 . [本文引用: 1] [20] He HL Yang XC Wang D Sun YY Yin CY Li T Li YX Zhou GY Zhang L Liu Q 2017 ). Ecological stoichiometric characteristics of soil carbon, nitrogen and phosphorus of Sibiraea angustata shrub in eastern Qinghai-?Tibetan PlateauChinese Journal of Applied and Environmental Biology 1128 -1135 . [本文引用: 3] [ 贺合亮 , 阳小成 , 王东 , 孙誉育 , 尹春英 , 李婷 , 黎云祥 , 周国英 , 张林 , 刘庆 ( 2017 ). 青藏高原东部窄叶鲜卑花灌丛土壤C、N、P生态化学计量学特征应用与环境生物学报 , 21, 1128 -1135 .] [本文引用: 3] [21] He M Chen H Pan CD Hu Y Xiao ZZ Wang B 2015 ). Seasonal variation and correlation of mineral element concentrations between Juglans regia ‘Xinwen 185’ leaves and fruitsXinjiang Agricultural Sciences 1399 -1406 . [本文引用: 1] [ 何苗 , 陈虹 , 潘存德 , 胡渊 , 肖真真 , 王蓓 ( 2015 ). 新温185号核桃叶片和果实矿质元素浓度时节变化及其相关性新疆农业科学 , 52, 1399 -1406 .] [本文引用: 1] [22] Hedin LO 2004 ). Global organization of terrestrial plant-nutrient interactionsProceedings of the National Academy of Sciences of the United States of America 10849 -10850 . [本文引用: 1] [23] Hidaka A Kitayama K 2015 ). Physiological linkage in co-variation of foliar nitrogen and phosphorus in tropical tree species along a gradient of soil phosphorus availabilityJournal of Tropical Ecology 221 -229 . [本文引用: 2] [24] Hu F Kong CH 2004 ). Selectivity and influence of parasite plants on their hostsChinese Journal of Applied Ecology 905 -908 . [本文引用: 1] [ 胡飞 , 孔垂华 ( 2004 ). 寄生植物对寄主的选择和影响应用生态学报 , 15, 905 -908 .] [本文引用: 1] [25] Hu YS Yao XY Liu YH 2014 ). N and P stoichiometric traits of plant and soil in different forest succession stages in Changbai MountainsChinese Journal of Applied Ecology 632 -638 . [本文引用: 2] [ 胡耀升 , 么旭阳 , 刘艳红 ( 2014 ). 长白山森林不同演替阶段植物与土壤氮磷的化学计量特征应用生态学报 , 25, 632 -638 .] [本文引用: 2] [26] Huang XY Guan KY Li AR 2011 ). Biological traits and their ecological significances of parasitic plants: A reviewChinese Journal of Ecology 1838 -1844 . [本文引用: 1] [ 黄新亚 , 管开云 , 李爱荣 ( 2011 ). 寄生植物的生物学特性及生态学效应生态学杂志 , 30, 1838 -1844 .] [本文引用: 1] [27] Jiang LL He S Wu LF Yan YF Weng SF Liu J Wang WQ Zeng CS 2014 ). Characteristics of stoichiometric homeostasis of three plant species in wetlands in Minjiang estuaryWetland Science 293 -298 . [本文引用: 1] [ 蒋利玲 , 何诗 , 吴丽凤 , 颜远烽 , 翁少峰 , 刘静 , 王维奇 , 曾从盛 ( 2014 ). 闽江河口湿地3种植物化学计量内稳性特征湿地科学 , 12, 293 -298 .] [本文引用: 1] [28] Jiang PP Cao Y Chen YM Zhao YP 2017 ). N and P stoichiometric characteristics of leaves, litter, and soil for three dominant tree species in the Shaanxi ProvinceActa Ecologica Sinica 443 -454 . [本文引用: 1] [ 姜沛沛 , 曹扬 , 陈云明 , 赵一娉 ( 2017 ). 陕西省3种主要树种叶片、凋落物和土壤N、P化学计量特征生态学报 , 37, 443 -454 .] [本文引用: 1] [29] Kerkhoff AJ Fagan WF Else JJ Enquist BJ 2006 ). Phylogenetic and growth form variation in the scaling of nitrogen and phosphorus in the seed plantsThe American Naturalist 103 -122 . [本文引用: 1] [30] Koerselman W Meuleman AFM 1996 ). The vegetation N:P ratio: A new tool to detect the nature of nutrient limitationJournal of Applied Ecology 1441 -1450 . [本文引用: 1] [31] Lep? J Tě?itel J 2015 ). Root hemiparasites in productive communities should attack competitive host, and harm them to make regeneration gapsJournal of Vegetation Science 407 -408 . [本文引用: 1] [32] Li S Liu WY Wang LS Yang GP Li DW 2007 ). Species diversity and distribution of epiphytic lichens in the primary and secondary forests in Ailao Mountain, YunnanBiodiversity Science 445 -455 . [本文引用: 1] [ 李苏 , 刘文耀 , 王立松 , 杨国平 , 李达文 ( 2007 ). 云南哀牢山原生林及次生林群落附生地衣物种多样性与分布生物多样性 , 15, 445 -455 .] [本文引用: 1] [33] Li W 2016 ). Global pattern of plant ecological stoichiometrics characteristicsJournal of Nanchang Institute of Technology 6 ), 6 -10 . [本文引用: 1] [ 李威 ( 2016 ). 植物生态化学计量学的全球格局综述南昌工程学院学报 , 35(6 ), 6 -10 .] [本文引用: 1] [34] Li XW Sun K Li FY 2014 ). Variation in leaf nitrogen and phosphorus stoichiometry in the nitrogen-fixing Chinese sea-buckthorn (Hippophae rhamnoides L. subsp. sinensis Rousi) across northern ChinaEcological Research 723 -731 . [本文引用: 1] [35] Li YH Chen SL Lu D Zhu KX Zhao MH Pei HH Ruan JL 2009 ). Study on the contents of total flavonoids of herbataxilli from different host-plantsLishizhen Medicine and Materia Medicare Research 3009 -3010 . [本文引用: 1] [ 李永华 , 陈士林 , 卢栋 , 朱开昕 , 赵明惠 , 裴河欢 , 阮金兰 ( 2009 ). 不同寄主植物桑寄生总黄酮含量研究时珍国医国药 , 20, 3009 -3010 .] [本文引用: 1] [36] Li YH Su BW Zhang XJ Zhu KX Pei HH Zhao MH Lu D 2012 ). Influence of host plant on the volatile components of taxilli herbaLishizhen Medicine and Materia Medicare Research 574 -578 . [本文引用: 1] [ 李永华 , 苏本伟 , 张协君 , 朱开昕 , 裴河欢 , 赵明惠 , 卢栋 ( 2012 ). 寄主植物对桑寄生药材挥发性成分的影响研究时珍国医国药 , 23, 574 -578 .] [本文引用: 1] [37] Liao ZW Mao XY Liu KX 2014 ). Effect of organic Carbon fertlizer on nutrient balance—Analysis of carbon, a short board, in plant nutritionActa Pedologica Sinica 656 -659 . [本文引用: 1] [ 廖宗文 , 毛小云 , 刘可星 ( 2014 ). 有机碳肥对养分平衡的作用初探——试析植物营养中的碳短板土壤学报 , 51, 656 -659 .] [本文引用: 1] [38] Liu WT Wei ZJ Lü SJ Sun SX 2015 ). Research advancees in stoichiometry of grassland in ChinaActa Agrestia Sinica 914 -926 . [本文引用: 1] [ 刘文亭 , 卫智军 , 吕世杰 , 孙世贤 ( 2015 ). 中国草原生态化学计量学研究进展草地学报 , 23, 914 -926 .] [本文引用: 1] [39] Lu ZY Song L Wang X Li YW Zhang YP Sha LQ 2017 ). Ecological stoichiometry characteristics of the litterfall-humus-soil continuum systems under different successional stages of the subtropical forest in SW ChinaMountain Reseach 274 -282 . [本文引用: 3] [ 鲁志云 , 宋亮 , 王训 , 李玉武 , 张一平 , 沙丽清 ( 2017 ). 哀牢山森林凋落物与腐殖质及土壤的生态化学计量特征山地学报 , 35, 274 -282 .] [本文引用: 3] [40] Luo Y Gong L Zhu ML An SQ 2017 ). Stoichometry charateristics of leaves and soil of four shrubs in the upper reacher of the Tarim River DesertActa Ecologica Sinica 8326 -8335 . [本文引用: 1] [ 罗艳 , 贡璐 , 朱美玲 , 安申群 ( 2017 ). 塔里木河上游荒漠区4种灌木植物叶片与土壤生态化学计量特征生态学报 , 37, 8326 -8335 .] [本文引用: 1] [41] Michaels AF 2003 ) . The ratios of lifeScience 906 -907 . [本文引用: 1] [42] Minden V Kleyer M 2015 ). Internal and external regulation of plant organ stoichiometryPlant Biology 897 -907 . [本文引用: 1] [43] Nave LE Heckman KA Mu?oz AB Swanston CW 2017 ). Radiocarbon suggests the hemiparasitic annual Melampyrum lineare Desr. may acquire carbon from stressed hostsRadiocarbon 1 -13 . [本文引用: 2] [44] Nickrent DL Duff RJ Colwell AE Wolfe AD Young ND Steiner KE dePamphilis CW 1998 ). Molecular phylogenetic and evolutionary studies of parasitic plants. Molecular Systematics of Plants IISpringer,New York. 211 -241 . [本文引用: 1] [45] Niklas KJ 2006 ). Plant allometry, leaf nitrogen and phosphorus stoichiometry, and interspecific trends in annual growth ratesAnnals of Botany 155 -163 . [本文引用: 1] [46] Niklas KJ Cobb ED 2005 ). N, P, and C stoichiometry of Eranthis hyemalis (Ranunculaceae) and the allometry of plant growthAmerican Journal of Botany 1256 -1263 . [本文引用: 1] [47] Norton DA Lange PJD 1999 ). Host specificity in Parasitic mistletoes (Loranthaceae) in New ZealandFunctional Ecology 552 -559 . [本文引用: 1] [48] Phoenix GK Press MC 2010 ). Linking physiological traits to impacts on community structure and function: The role of root hemiparasitic Orobanchaceae (ex-Scrophulariaceae)Journal of Ecology 67 -78 . [本文引用: 1] [49] Press MC Phoenix GK 2010 ). Impacts of parasitic plants on natural communitiesNew Phytologist 737 -751 . [本文引用: 1] [50] Qiu XZ Xie SC 1998 ). Studies on the Forest Ecosystem in Ailao Mountains [本文引用: 1] [ 邱学忠 , 谢寿昌 ( 1998 ). 云南哀牢山森林生态系统研究 . 云南科技出版社, 昆明.] [本文引用: 1] [51] Quested HM Callaghan TV Cornelissen JHC Press MC 2010 ). The impact of hemiparasitic plant litter on decomposition: Direct, seasonal and litter mixing effectsJournal of Ecology 87 -98 . [本文引用: 1] [52] Reich PB Oleksyn J 2004 ). Global patterns of plant leaf N and P in relation to temperature and latitudeProceedings of National Academy of Sciences of the United States of America 11001 -11006 . [本文引用: 3] [53] Reich PB Oleksyn J Wright IJ Niklas KJ Hedin L Elser JJ 2010 ). Evidence of a General 2/3-power law of scaling leaf nitrogen to phosphorus among major plant groups and biomesProceedings of the Royal Society B 877 -883 . [本文引用: 2] [54] Ren SJ Yu GR Tao B Wang SQ 2007 ). Leaf nitrogen and phosphorus stoichiometry across 643 terrestrial plant species in NSTECEnvironmental Science 2665 -2673 . [本文引用: 3] [ 任书杰 , 于贵瑞 , 陶波 , 王绍强 ( 2007 ). 中国东部南北样带654种植物叶片氮和磷的化学计量学特征研究环境科学 , 28, 2665 -2673 .] [本文引用: 3] [55] Selosse MA Charpin M Not F 2017 ). Mixotrophy everywhere on land and in water: The grand écart hypothesisEcology Letters 246 . [本文引用: 1] [56] Sterner RW Elser JJ 2002 ). Ecological Stoichiometry:The Biology of Elements From Molecules to the BiospherePrinceton University Press 225 -226 . [本文引用: 4] [57] Suriyagoda LDB Rajapaksha R Pushpakumara G Lambers H 2017 ). Nutrient resorption from senescing leaves of epiphytes, hemiparasites and their hosts in tropical forests of Sri LankaJournal of Plant Ecology 815 -826 . [本文引用: 2] [58] Světlíková P Bla?ek P Mühlsteinová R Tě?itel J 2016 ). Tracing nitrogen flow in a root-hemiparasitic association by foliar stable-isotope labellingPlant Ecology and Evolution 39 -44 . [本文引用: 1] [59] Tennakoon KU Chak WH Bolin JF 2011 ). Nutritional and isotopic relationships of selected Bornean tropical mistletoe-?host associations in Brunei DarussalamFunctional Plant Biology 505 -513 . [本文引用: 1] [60] Tě?itel J Tě?itelová T Fisher JP Lep? J Cameron DD 2015 ). Integrating ecology and physiology of root-hemiparasitic interaction: Interactive effects of abiotic resources shape the interplay between parasitism and autotrophyNew Phytologist 350 -360 . [本文引用: 3] [61] Tian D Yan ZB Niklas KJ Han WX Kattge J Reich PB Luo YK Chen YH Tang ZY Hu HF Wright IJ Schmid B Fang JY 2018 ). Global leaf nitrogen and phosphorus stoichiometry and their scaling exponentNational Science Review 728 -739 . [本文引用: 1] [62] Tian HQ Chen GS Zhang C Melillo JM Hall CAS 2010 ). Pattern and variation of C:N:P ratios in China’s soils: A synthesis of observational dataBiogeochemistry 327 -346 . [本文引用: 1] [63] Vitousek PM 1998 ). Foliar and litter nutrients, nutrient resorption, and decomposition in Hawaiian Metrosideros polymorphaEcosystems 401 -407 . [本文引用: 1] [64] Wang T Yang YH Ma WH 2008 ). Storage, patterns and environmental controls of soil phosphorus in ChinaActa Scientiarum Naturalium Universitatis Pekinensis 945 -952 . [本文引用: 1] [ 汪涛 , 杨元合 , 马文红 ( 2008 ). 中国土壤磷库的大小、分布及其影响因素北京大学学报(自然科学版) , 44, 945 -952 .] [本文引用: 1] [65] Wang XN Zhang L 2017 ). Species diversity and distribution of mistletoes and hosts in four different habitats in Xishuangbanna, Southwest ChinaChinese Journal of Plant Ecology 701 -711 . [本文引用: 1] [ 王煊妮 , 张玲 ( 2017 ). 西双版纳4种生境下的桑寄生与寄主植物多样性及分布特点植物生态学报 , 39, 701 -711 .] [本文引用: 1] [66] Watson DM 2010 ). Parasitic plants as facilitators: More dryad than dracula?Journal of Ecology 1151 -1159 . [本文引用: 1] [67] Westoby M Falster DS Moles AT Vesk PA Wright IJ 2002 ). Plant ecological strategies: Some leading dimensions of variation between speciesAnnual Review of Ecology and Systematics 125 -159 . [本文引用: 1] [68] Wright IJ Reich PB Cornelissen JHC Falster DS Garnier E Hikosaka K Lamont BB Lee W Oleksyn J Osada N Poorter H Villar R Warton DI Westoby M 2005 ). Assessing the generality of global leaf trait relationshipsNew Phytologist 485 -496 . [本文引用: 1] [69] Wright IJ Reich PB Westoby M Ackerly DD Baruch Z Bongers F Cavender-Bares J Chapin FS Cornelissen JHC Diemer M Flexas J Garnier E Groom PK Gulias J Hikosaka K Lamont BB Lee T Lee W Lusk C Midgley JJ Navas M-L Niinemets ü Oleksyn J Osada N Poorter H Poot P Prior L Pyankov VI Roumet C Thomas SC Tjoelker MG Veneklaas EJ Villar R 2004 ). The world- wide leaf economics spectrumNature 821 -827 . [本文引用: 2] [70] Yan ZB Li XP Tian D Han WX Hou XH Shen HS Guo YL Fang JY 2018 ). Nutrient addition affects scaling relationship of leaf nitrogen to phosphorus in Arabidopsis thalianaFunctional Ecology 2689 -2698 . [本文引用: 2] [71] Yang M Wang CQ Yuan DG Li QQ Zeng J Luo Q Lan XM Tang J 2015 ). C, N, P stoichiometry traits of different flue-cured tobacco organs at different growth stages. Chinese Journal ofEco-Agriculture 686 -693 . [本文引用: 1] [ 杨梅 , 王昌全 , 袁大刚 , 李启权 , 曾建 , 罗茜 , 兰兴梅 , 唐杰 ( 2015 ). 不同生长期烤烟各器官C、N、P生态化学计量学特征中国生态农业学报 , 23, 686 -693 .] [本文引用: 1] [72] You CX 1983 ). Classification of Vegetation in Xujiaba Region in Ailao MtsResearch of Forest Ecosystems on Ailao Mountains Yunnan [本文引用: 1] [ 游承侠 ( 1983 ). 哀牢山徐家坝地区的植被分类云南哀牢山森林生态系统研究 . 云南科技出版社, 昆明.] [本文引用: 1] [73] Young SS Herwitz SR 1995 ). Floristic diversity and co-occurrences in a subtropical broad-leaved forest and two contrasting regrowth stands in central-west Yunnan Province, ChinaVegetatio 1 -13 . [本文引用: 1] [74] Zeng DH Chen GS 2005 ). Ecological stoichiometry: A science to explore the complexity of living systemsActa Phytoecologica Sinica 1007 -1019 . [本文引用: 1] [ 曾德慧 , 陈广生 ( 2005 ). 生态化学计量学: 复杂生命系统奥秘的探索植物生态学报 , 29, 1007 -1019 .] [本文引用: 1] [75] Zeng DP Jiang LL Zeng CS Wang WQ Wang C 2013 ). Reviews on the ecological stoichiometry characteristics and its applicationsActa Ecologica Sinica 5484 -5492 . [本文引用: 1] [ 曾冬萍 , 蒋利玲 , 曾从盛 , 王维奇 , 王纯 ( 2013 ). 生态化学计量学特征及其应用研究进展生态学报 , 33, 5484 -5492 .] [本文引用: 1] [76] Zeng ZX Wang KL Liu XL Zeng FP Song TQ Peng WX Zhang H Du H 2015 ). Stoichiometric characteristics of plants, litter and soils in karst plant communities of Northwest GuangxiChinese Journal of Plant Ecology 682 -693 . [本文引用: 1] [ 曾昭霞 , 王克林 , 刘孝利 , 曾馥平 , 宋同清 , 彭晚霞 , 张浩 , 杜虎 ( 2015 ). 桂西北喀斯特森林植物-凋落物-土壤生态化学计量特征植物生态学报 , 39, 682 -693 .] [本文引用: 1] [77] Zhang K He MZ Li XR Tan HJ Gao YH Li G Han GJ Wu YY 2014 ). Foliar carbon, nitrogen and phosphorus stoichiometry of typical desert plants across the Alashan DesertActa Ecologica Sinica 6538 -6547 . [本文引用: 1] [ 张珂 , 何明珠 , 李新荣 , 谭会娟 , 高艳红 , 李刚 , 韩国君 , 吴杨杨 ( 2014 ). 阿拉善荒漠典型植物叶片碳、氮、磷化学计量特征生态学报 , 34, 6538 -6547 .] [本文引用: 1] [78] Zhao YP Cao Y Chen YM Peng SZ 2007 ). Ecological stoichiometry in a forest in the hilly-gully of Loess PlateauActa Ecologica Sinica 5451 -5460 . [本文引用: 1] [ 赵一娉 , 曹扬 , 陈云明 , 彭守璋 ( 2017 ). 黄土丘陵沟壑区森林生态系统生态化学计量特征生态学报 , 37, 5451 -5460 .] [本文引用: 1] [79] Zhou HH Liu XL Qian HB Wang SQ Zhou C 2013 ). Comparative study on toxicities of Taxillus sutchuenensis from different host plantsChinese Journal of Experimental Traditional Medical Formulae 274 -277 . [本文引用: 1] [ 周汉华 , 刘晓龙 , 钱海兵 , 王世清 , 周超 ( 2013 ). 不同寄主上的桑寄生药材毒性的比较研究中国实验方剂学杂志 , 19, 274 -277 .] [本文引用: 1] [80] Zhu H Yan LC 2009 ). List of Seed Plants in the Ailao Mts.of Yunnan Province, China [本文引用: 3] [ 朱华 , 闫丽春 ( 2009 ). 云南哀牢山种子植物 . 云南科技出版社, 昆明.] [本文引用: 3] Attack on all fronts: Functional relationships between aerial and root parasitic plants and their woody hosts and consequences for ecosystems
1
2011
... 将采集到的植物叶片用自来水洗净灰尘等杂质后, 再用超纯水冲洗.由于半寄生植物的大部分矿物质是通过吸器从寄主木质部获取(
Glatzel, 1983 ;
Bell & Adams, 2011 ), 故将寄主枝条的树皮去除后作为半寄生植物的养分来源, 以便更准确地分析寄主枝条-吸器-寄生枝条-寄生叶片的生态化学计量特征变化.将寄主枝条、吸器、寄生枝条与叶片分别装入信封, 于65 ℃条件下烘干至恒质量.烘干后先用刀斧将吸器及寄主枝条、寄生枝条粗加工成小块, 再通过微型植物粉碎机(FZ102型, 天津市泰斯特仪器有限公司, 天津)将样品磨成粉末, 植物叶片则利用高速万能粉碎机(FW80型, 天津市泰斯特仪器有限公司, 天津)磨成粉末, 用于全碳、全氮和全磷含量的测定.C、N含量用碳氮分析仪(Vario MAX CN, Elementar Analysensysteme GmbH, Hanau, Germany)测定, P含量采用电感耦合等离子体原子发射光谱仪(iCAP6300, Thermo Fisher Scientific, Waltham, USA)测定. ...
Water and nutrient relationships between a mistletoe and its mangrove host under saline conditions
1
2013
... 半寄生植物与寄主的复杂关系包括寄生和竞争(
Lep? & Tě?itel, 2015 ).作为混合营养策略的代表, 半寄生植物既能从自身的光合作用中获取碳同化物, 也能通过吸器从寄主体内获取碳水化合物(
Nave et al ., 2017 ;
Selosse et al ., 2017 ).半寄生植物自身的光合作用在同化寄生矿物资源, 并将矿物资源转换化成自身生长和适合度中起着至关重要的作用, 而将木质部所获得的资源转化为生长和适应的关键过程, 可能是其进化稳定性的基础(
Tě?itel et al ., 2015 ).尽管半寄生植物与寄主关系的研究开展较早, 不少****对半寄生植物与寄主之间的水分利用、光合作用以及矿物质营养等方面的关系进行了较为深入的研究(
Tennakoon et al ., 2011 ;
Chen et al ., 2013 ;
Suriyagoda et al ., 2017 ;
Gebauer et al ., 2018 ), 但营养物质从寄主到半寄生植物的转移机制仍未完全研究清楚(
Gebauer et al ., 2012 ,
2018 ).有关研究表明, 半寄生植物
Rhinanthus alectorolophus 中5%-40%的C来自寄主, 而寄主体内的N亦可被半寄生植物获取(
Tě?itel et al ., 2015 ;
Světlíková et al ., 2016 ).寄生植物的吸器同时连接寄主枝条与寄生枝条, 研究寄主枝条-吸器-寄生枝条-寄生叶片这一连续体的生态化学计量特征变化将有助于进一步理解寄生植物与寄主间的养分关系. ...
C:N:P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomass
1
2007
... 寄生植物是一类重要而独特的植物类群.本研究中两种半寄生植物叶片的平均N含量为12.08 mg·g
-1 , 明显低于全球陆生植物的N含量20.09 mg·g
-1 (
Reich & Oleksyn, 2004 )、中国陆生植物的N含量18.6 mg·g
-1 (
Han et al ., 2005 ); 而P含量3.06 mg·g
-1 则明显地高于全球和中国陆生植物的P含量, 分别为1.77 mg·g
-1 (
Reich & Oleksyn, 2004 )、1.21 mg·g
-1 (
Han et al ., 2005 ).寄生叶的N含量较低可能与其光合作用较弱有关, 而较高的P含量可能受到研究区土壤P含量的间接影响, 哀牢山地区森林土壤的全P和有效P含量分别为1.17 g·kg
-1 和3.40 mg·kg
-1 (
鲁志云等, 2017 ), 中国土壤全P和有效P含量分别为0.60 g·kg
-1 和2.39 mg·kg
-1 (
汪涛等, 2008 ), 哀牢山地区森林土壤全P含量显著高于全国水平(
鲁志云等, 2017 ).此外, 哀牢山地区山地森林表层土壤(0-20 cm)的C:N、C:P、N:P值分别为14.9、84.9、5.7, 高于中国陆地(0-10 cm)土壤的C:N、C:P、N:P水平(分别为12.3、52.7、3.9)(
Tian et al ., 2010 ), 更接近于全球陆地(0-10 cm)和全球森林土壤水平(分别为12.3、72.0、5.9和12.4、81.9、6.6)(
Cleveland & Liptzin, 2007 ).已有的研究认为我国陆地植物生长普遍受P限制(
Han et al ., 2005 ;
任书杰等, 2007 ), 但哀牢山地区较高的土壤P含量暗示该地区可能并不缺P.关于哀牢山地区森林寄生植物、寄主的生态化学计量特征与土壤化学元素计量的关系将是以后研究的重要内容. ...
Environmental constraints on a global relationship among leaf and root traits of grasses
1
2005
... 大多数有关寄生植物的研究主要关注于植物的光合器官, 而对寄生植物不同器官与寄主植物间的关系研究较少.有关研究表明, 植物叶片N、P浓度与枝条N、P浓度具有相关性(
Kerkhoff et al ., 2006 ), 根和叶间的N浓度也具有显著的相关性(
Craine et al ., 2005 ).本研究寄生枝与寄生叶之间的P含量没有呈现出显著的相关性, 但N含量存在极显著正相关关系, 该结果与
杨梅等(2015) 、
胡耀升等(2014) 的研究果相一致.叶和茎间N含量的相关关系反映了叶、茎在生物、生化方面的功能需求, 这是由植株冠层结构所决定的(
Westoby et al ., 2002 ;
胡耀升等, 2014 ). ...
陕西省森林各生态系统组分氮磷化学计量特征
1
2015
... N和P在植物中为协同元素, 植物叶片N、P含量的正相关关系亦为高等陆生植物养分计量的普遍规律(
Sterner & Elser, 2002 ;
Wright et al ., 2005 ).植物N、P间的耦合关系很早就有文献记录(
Garten, 1976 ), 随后也得到许多研究的验证(
Li et al ., 2014 ;
Hidaka & Kitayama, 2015 ;
崔高阳等, 2015 ).对中国753种陆地植物叶片的研究(
Han et al ., 2005 )以及中国东部南北样带654种植物叶片的研究(
任书杰等, 2007 )表明, 不同生活型或不同系统发育植物的叶片N和P含量大部分也呈极显著相关关系.水生植物N、P含量间同样具有强线性关系, 这可能是大多数植物N、P含量的基本特征(
Duarte, 1992 ).
Hidaka和Kitayama (2015) 认为植物叶片N、P含量间的正相关关系可能部分受到土壤可利用性氮含量的影响, 而植物营养策略(N利用策略受到P利用策略的影响)以及植物细胞中的N和P生态化学计量特征(如蛋白质合成中N和P的生化关系)则可提供另一种解释, 但仍需要进一步分析以阐明相关性的生理背景.本研究中, N和P由寄主枝、吸器中的极显著正相关关系, 寄生枝中显著正相关关系, 到寄生叶中变为了相关关系不显著.
Garten (1976) 的研究中大型水生植物的N和P含量相关性也不显著, 这可能是大型水生植物的N含量范围较小, 或者是当环境P缺乏时对N的奢侈消费造成的.
Han等(2005) 和
任书杰等(2007) 的研究中, 裸子植物和蕨类植物的N、P含量相关性也不显著.本研究中寄生叶N、P间的不协同性可能与寄生植物不直接从土壤吸收养分、混合营养策略以及寄生植物叶片重吸收P但不重吸收N有关(
Suriyagoda et al ., 2017 ).受到N、P不耦合的影响, 寄生叶的N含量和C:P, P含量和C:N也不存在显著的相关关系. ...
陕西省森林各生态系统组分氮磷化学计量特征
1
2015
... N和P在植物中为协同元素, 植物叶片N、P含量的正相关关系亦为高等陆生植物养分计量的普遍规律(
Sterner & Elser, 2002 ;
Wright et al ., 2005 ).植物N、P间的耦合关系很早就有文献记录(
Garten, 1976 ), 随后也得到许多研究的验证(
Li et al ., 2014 ;
Hidaka & Kitayama, 2015 ;
崔高阳等, 2015 ).对中国753种陆地植物叶片的研究(
Han et al ., 2005 )以及中国东部南北样带654种植物叶片的研究(
任书杰等, 2007 )表明, 不同生活型或不同系统发育植物的叶片N和P含量大部分也呈极显著相关关系.水生植物N、P含量间同样具有强线性关系, 这可能是大多数植物N、P含量的基本特征(
Duarte, 1992 ).
Hidaka和Kitayama (2015) 认为植物叶片N、P含量间的正相关关系可能部分受到土壤可利用性氮含量的影响, 而植物营养策略(N利用策略受到P利用策略的影响)以及植物细胞中的N和P生态化学计量特征(如蛋白质合成中N和P的生化关系)则可提供另一种解释, 但仍需要进一步分析以阐明相关性的生理背景.本研究中, N和P由寄主枝、吸器中的极显著正相关关系, 寄生枝中显著正相关关系, 到寄生叶中变为了相关关系不显著.
Garten (1976) 的研究中大型水生植物的N和P含量相关性也不显著, 这可能是大型水生植物的N含量范围较小, 或者是当环境P缺乏时对N的奢侈消费造成的.
Han等(2005) 和
任书杰等(2007) 的研究中, 裸子植物和蕨类植物的N、P含量相关性也不显著.本研究中寄生叶N、P间的不协同性可能与寄生植物不直接从土壤吸收养分、混合营养策略以及寄生植物叶片重吸收P但不重吸收N有关(
Suriyagoda et al ., 2017 ).受到N、P不耦合的影响, 寄生叶的N含量和C:P, P含量和C:N也不存在显著的相关关系. ...
The effects of hemiparasitic plant removal on community structure and seedling establishment in semi-natural grasslands
1
2015
... 半寄生植物是寄生植物的主要类群, 其叶片具有叶绿体, 在生命周期的至少一个阶段可进行光合作用, 仅从寄主体内获取部分水和溶解的矿物质(
Nickrent et al ., 1998 ).在自然界中, 半寄生植物对植物群落结构和多样性具有显著影响(
Phoenix & Press, 2010 ;
Press & Phoenix, 2010 ;
Demey et al ., 2014 ), 而其营养丰富、可快速分解和释放养分的凋落物可能会提高森林土壤养分可利用率(
Demey et al ., 2015 ), 影响生态系统水平凋落物分解和养分循环(
Quested et al ., 2010 ).虽然半寄生植物的丰度和生物量相对较小, 但它们对生物地球化学循环、植物种群结构和多样性的影响是很明确的(
Nave et al ., 2017 ), 因而被描述为影响群落动态的生态系统工程师, 以及影响养分循环的关键物种(
Watson, 2010 ;
Hatcher et al ., 2012 ). ...
Hemiparasitic litter additions alter gross nitrogen turnover in temperate semi-natural grassland soils
1
2014
... 半寄生植物是寄生植物的主要类群, 其叶片具有叶绿体, 在生命周期的至少一个阶段可进行光合作用, 仅从寄主体内获取部分水和溶解的矿物质(
Nickrent et al ., 1998 ).在自然界中, 半寄生植物对植物群落结构和多样性具有显著影响(
Phoenix & Press, 2010 ;
Press & Phoenix, 2010 ;
Demey et al ., 2014 ), 而其营养丰富、可快速分解和释放养分的凋落物可能会提高森林土壤养分可利用率(
Demey et al ., 2015 ), 影响生态系统水平凋落物分解和养分循环(
Quested et al ., 2010 ).虽然半寄生植物的丰度和生物量相对较小, 但它们对生物地球化学循环、植物种群结构和多样性的影响是很明确的(
Nave et al ., 2017 ), 因而被描述为影响群落动态的生态系统工程师, 以及影响养分循环的关键物种(
Watson, 2010 ;
Hatcher et al ., 2012 ). ...
Nutrient concentration of aquatic plants: Patterns across species
1
1992
... N和P在植物中为协同元素, 植物叶片N、P含量的正相关关系亦为高等陆生植物养分计量的普遍规律(
Sterner & Elser, 2002 ;
Wright et al ., 2005 ).植物N、P间的耦合关系很早就有文献记录(
Garten, 1976 ), 随后也得到许多研究的验证(
Li et al ., 2014 ;
Hidaka & Kitayama, 2015 ;
崔高阳等, 2015 ).对中国753种陆地植物叶片的研究(
Han et al ., 2005 )以及中国东部南北样带654种植物叶片的研究(
任书杰等, 2007 )表明, 不同生活型或不同系统发育植物的叶片N和P含量大部分也呈极显著相关关系.水生植物N、P含量间同样具有强线性关系, 这可能是大多数植物N、P含量的基本特征(
Duarte, 1992 ).
Hidaka和Kitayama (2015) 认为植物叶片N、P含量间的正相关关系可能部分受到土壤可利用性氮含量的影响, 而植物营养策略(N利用策略受到P利用策略的影响)以及植物细胞中的N和P生态化学计量特征(如蛋白质合成中N和P的生化关系)则可提供另一种解释, 但仍需要进一步分析以阐明相关性的生理背景.本研究中, N和P由寄主枝、吸器中的极显著正相关关系, 寄生枝中显著正相关关系, 到寄生叶中变为了相关关系不显著.
Garten (1976) 的研究中大型水生植物的N和P含量相关性也不显著, 这可能是大型水生植物的N含量范围较小, 或者是当环境P缺乏时对N的奢侈消费造成的.
Han等(2005) 和
任书杰等(2007) 的研究中, 裸子植物和蕨类植物的N、P含量相关性也不显著.本研究中寄生叶N、P间的不协同性可能与寄生植物不直接从土壤吸收养分、混合营养策略以及寄生植物叶片重吸收P但不重吸收N有关(
Suriyagoda et al ., 2017 ).受到N、P不耦合的影响, 寄生叶的N含量和C:P, P含量和C:N也不存在显著的相关关系. ...
Nutritional constraints in terrestrial and freshwater food webs
1
2000
... 生态化学计量学是主要研究生态系统能量平衡和多重化学元素平衡的科学(
Sterner & Elser, 2002 ;
曾德慧和陈广生, 2005 ), 其核心问题是揭示生物体元素组成差异对其生态功能的影响(
Elser et al ., 2000 ;
蒋利玲等, 2014 ).生态化学计量学从元素比率的角度将不同尺度、不同生物群系以及不同领域的生物学研究联系起来(
Sterner & Elser, 2002 ;
Michaels, 2003 ), 成为生态学研究的主要方法和研究热点(
曾冬萍等, 2013 ;
刘文亭等, 2015 ).这一领域研究最多的化学元素是碳(C)、氮(N)、磷(P), 作为3种最重要的生命元素, C、N、P是所有生命过程的基础, 在植物的生化功能方面紧密耦合(
李威, 2016 ).基于不同生命形式, 系统发育以及光合作用碳同化途径的植物叶片生态化学计量特征及其变异模式的研究已取得重大进展(
Han et al ., 2005 ,
2011 ), 但这些研究的主要对象为非寄生植物.有关研究表明, 非寄生植物生态化学计量特征与土壤养分状况相关(
Reich & Oleksyn, 2004 ;
赵一娉等, 2017 ), 但寄生植物没有根系直接接触土壤, 其生命活动所需要的养分和水分主要来自于寄主植物.目前, 有关寄生植物生态化学计量特征的研究鲜有报道. ...
Correlations between concentrations of elements in plants
2
1976
... N和P在植物中为协同元素, 植物叶片N、P含量的正相关关系亦为高等陆生植物养分计量的普遍规律(
Sterner & Elser, 2002 ;
Wright et al ., 2005 ).植物N、P间的耦合关系很早就有文献记录(
Garten, 1976 ), 随后也得到许多研究的验证(
Li et al ., 2014 ;
Hidaka & Kitayama, 2015 ;
崔高阳等, 2015 ).对中国753种陆地植物叶片的研究(
Han et al ., 2005 )以及中国东部南北样带654种植物叶片的研究(
任书杰等, 2007 )表明, 不同生活型或不同系统发育植物的叶片N和P含量大部分也呈极显著相关关系.水生植物N、P含量间同样具有强线性关系, 这可能是大多数植物N、P含量的基本特征(
Duarte, 1992 ).
Hidaka和Kitayama (2015) 认为植物叶片N、P含量间的正相关关系可能部分受到土壤可利用性氮含量的影响, 而植物营养策略(N利用策略受到P利用策略的影响)以及植物细胞中的N和P生态化学计量特征(如蛋白质合成中N和P的生化关系)则可提供另一种解释, 但仍需要进一步分析以阐明相关性的生理背景.本研究中, N和P由寄主枝、吸器中的极显著正相关关系, 寄生枝中显著正相关关系, 到寄生叶中变为了相关关系不显著.
Garten (1976) 的研究中大型水生植物的N和P含量相关性也不显著, 这可能是大型水生植物的N含量范围较小, 或者是当环境P缺乏时对N的奢侈消费造成的.
Han等(2005) 和
任书杰等(2007) 的研究中, 裸子植物和蕨类植物的N、P含量相关性也不显著.本研究中寄生叶N、P间的不协同性可能与寄生植物不直接从土壤吸收养分、混合营养策略以及寄生植物叶片重吸收P但不重吸收N有关(
Suriyagoda et al ., 2017 ).受到N、P不耦合的影响, 寄生叶的N含量和C:P, P含量和C:N也不存在显著的相关关系. ...
... 认为植物叶片N、P含量间的正相关关系可能部分受到土壤可利用性氮含量的影响, 而植物营养策略(N利用策略受到P利用策略的影响)以及植物细胞中的N和P生态化学计量特征(如蛋白质合成中N和P的生化关系)则可提供另一种解释, 但仍需要进一步分析以阐明相关性的生理背景.本研究中, N和P由寄主枝、吸器中的极显著正相关关系, 寄生枝中显著正相关关系, 到寄生叶中变为了相关关系不显著.
Garten (1976) 的研究中大型水生植物的N和P含量相关性也不显著, 这可能是大型水生植物的N含量范围较小, 或者是当环境P缺乏时对N的奢侈消费造成的.
Han等(2005) 和
任书杰等(2007) 的研究中, 裸子植物和蕨类植物的N、P含量相关性也不显著.本研究中寄生叶N、P间的不协同性可能与寄生植物不直接从土壤吸收养分、混合营养策略以及寄生植物叶片重吸收P但不重吸收N有关(
Suriyagoda et al ., 2017 ).受到N、P不耦合的影响, 寄生叶的N含量和C:P, P含量和C:N也不存在显著的相关关系. ...
Quercus pubescens and its hemiparasite Loranthus europaeus: Nutrient dynamics of leaves and twigs
2
2012
... 半寄生植物与寄主的复杂关系包括寄生和竞争(
Lep? & Tě?itel, 2015 ).作为混合营养策略的代表, 半寄生植物既能从自身的光合作用中获取碳同化物, 也能通过吸器从寄主体内获取碳水化合物(
Nave et al ., 2017 ;
Selosse et al ., 2017 ).半寄生植物自身的光合作用在同化寄生矿物资源, 并将矿物资源转换化成自身生长和适合度中起着至关重要的作用, 而将木质部所获得的资源转化为生长和适应的关键过程, 可能是其进化稳定性的基础(
Tě?itel et al ., 2015 ).尽管半寄生植物与寄主关系的研究开展较早, 不少****对半寄生植物与寄主之间的水分利用、光合作用以及矿物质营养等方面的关系进行了较为深入的研究(
Tennakoon et al ., 2011 ;
Chen et al ., 2013 ;
Suriyagoda et al ., 2017 ;
Gebauer et al ., 2018 ), 但营养物质从寄主到半寄生植物的转移机制仍未完全研究清楚(
Gebauer et al ., 2012 ,
2018 ).有关研究表明, 半寄生植物
Rhinanthus alectorolophus 中5%-40%的C来自寄主, 而寄主体内的N亦可被半寄生植物获取(
Tě?itel et al ., 2015 ;
Světlíková et al ., 2016 ).寄生植物的吸器同时连接寄主枝条与寄生枝条, 研究寄主枝条-吸器-寄生枝条-寄生叶片这一连续体的生态化学计量特征变化将有助于进一步理解寄生植物与寄主间的养分关系. ...
... 寄生植物与其寄主之间的养分相关性蕴含了重要信息, 这些信息尚未被充分理解(
Gebauer et al ., 2018 ).寄主植物中的元素浓度通常是用来预测半寄生植物体内元素浓度的最佳指标(
Glatzel & Geils, 2009 ;
Gebauer et al ., 2012 ).本研究中寄主枝与吸器之间的N、P含量以及C:N、C:P和N:P均呈极显著正相关关系, 说明寄生植物吸器与寄主枝存在非常密切的关系, 但寄主枝与寄生枝间仅N含量和C:N存在显著相关关系, 寄主枝与寄生叶间仅C、N含量显著相关.这说明寄主枝的C、N、P浓度并非同时适用于预测寄生植物不同器官的C、N、P浓度, 如寄主枝的C浓度可能仅适用于预测寄生叶的C浓度.此外, 寄主枝、吸器、寄生枝与寄生叶两两器官间, 仅寄主枝C含量与寄生叶C含量为极显著负相关关系, 说明寄主枝C含量对寄生叶的形成以及生理活动的正常进行可能有着不可或缺的作用.寄主枝与吸器、吸器与寄生枝间P含量极显著正相关, 说明寄主枝、吸器和寄生枝对P的吸收具有一致性(
何苗等, 2015 ). ...
Seasonal variations of sulphur, phosphorus and magnesium in the leaves and current-year twigs of hemiparasitic mistletoe Loranthus europaeus Jacq. and its host Quercus pubescens Willd
4
2018
... 半寄生植物与寄主的复杂关系包括寄生和竞争(
Lep? & Tě?itel, 2015 ).作为混合营养策略的代表, 半寄生植物既能从自身的光合作用中获取碳同化物, 也能通过吸器从寄主体内获取碳水化合物(
Nave et al ., 2017 ;
Selosse et al ., 2017 ).半寄生植物自身的光合作用在同化寄生矿物资源, 并将矿物资源转换化成自身生长和适合度中起着至关重要的作用, 而将木质部所获得的资源转化为生长和适应的关键过程, 可能是其进化稳定性的基础(
Tě?itel et al ., 2015 ).尽管半寄生植物与寄主关系的研究开展较早, 不少****对半寄生植物与寄主之间的水分利用、光合作用以及矿物质营养等方面的关系进行了较为深入的研究(
Tennakoon et al ., 2011 ;
Chen et al ., 2013 ;
Suriyagoda et al ., 2017 ;
Gebauer et al ., 2018 ), 但营养物质从寄主到半寄生植物的转移机制仍未完全研究清楚(
Gebauer et al ., 2012 ,
2018 ).有关研究表明, 半寄生植物
Rhinanthus alectorolophus 中5%-40%的C来自寄主, 而寄主体内的N亦可被半寄生植物获取(
Tě?itel et al ., 2015 ;
Světlíková et al ., 2016 ).寄生植物的吸器同时连接寄主枝条与寄生枝条, 研究寄主枝条-吸器-寄生枝条-寄生叶片这一连续体的生态化学计量特征变化将有助于进一步理解寄生植物与寄主间的养分关系. ...
... ,
2018 ).有关研究表明, 半寄生植物
Rhinanthus alectorolophus 中5%-40%的C来自寄主, 而寄主体内的N亦可被半寄生植物获取(
Tě?itel et al ., 2015 ;
Světlíková et al ., 2016 ).寄生植物的吸器同时连接寄主枝条与寄生枝条, 研究寄主枝条-吸器-寄生枝条-寄生叶片这一连续体的生态化学计量特征变化将有助于进一步理解寄生植物与寄主间的养分关系. ...
... 寄生植物不同的营养素富集程度也不同, 被认为是吸器选择性吸收离子的结果, 若摄取是通过蒸腾而被动吸收, 那么营养元素的比值应保持相同(
Gebauer et al ., 2018 ).本研究中, 椆树桑寄生吸器的C:N、C:P及N:P均与其寄主枝条存在显著差异, 柳叶钝果寄生吸器的C:N和C:P也与其寄主枝条存在显著差异, 这在一定程度上说明C、N和P元素并非简单的通过蒸腾作用而被转运至寄生植物体内, 但也不能排除离子通过被动运输进入吸器后, 由于吸器具有合成物质的功能而导致元素比值发生改变(
黄新亚等, 2011 ).植物不同器官C、N、P含量的差异不仅受植物基本生理过程需求的影响, 也受相应器官组织结构和功能分化的影响(
Minden & Kleyer, 2015 ).叶片是植物的同化器官与重要的养分储存器官, 因而寄生植物叶片的N和P含量高于寄主枝、吸器与寄生枝(
Niklas & Cobb, 2005 ;
贺合亮等, 2017 ).植物叶片中C、N、P含量的变异系数普遍存在C < N < P的规律(
Koerselman & Meuleman, 1996 ;
Güsewell & Koerselman, 2002 ;
姜沛沛等, 2017 ).本研究中两种半寄生植物的寄主枝、吸器、寄生枝与寄生叶的C、N、P含量皆符合这个规律, 这是因为C作为构成植物骨架的基本结构物质, 且为多种植物生理活动提供能源, 而N含量可被植物控制在较窄的范围内, 故C在植物体内含量高且变异小, N含量在各器官中的变异性小于P含量(
Vitousek, 1998 ;
贺合亮等, 2017 ).本研究中两种寄生植物C:N的变异性小于C:P,是因为植物光合作用中C的同化与营养元素的吸收并非同一途径, 而C含量高且变异小, C:N和C:P的变异主要受到N和P含量变化的影响(
Hedin, 2004 ;
曾昭霞等, 2015 ). ...
... 寄生植物与其寄主之间的养分相关性蕴含了重要信息, 这些信息尚未被充分理解(
Gebauer et al ., 2018 ).寄主植物中的元素浓度通常是用来预测半寄生植物体内元素浓度的最佳指标(
Glatzel & Geils, 2009 ;
Gebauer et al ., 2012 ).本研究中寄主枝与吸器之间的N、P含量以及C:N、C:P和N:P均呈极显著正相关关系, 说明寄生植物吸器与寄主枝存在非常密切的关系, 但寄主枝与寄生枝间仅N含量和C:N存在显著相关关系, 寄主枝与寄生叶间仅C、N含量显著相关.这说明寄主枝的C、N、P浓度并非同时适用于预测寄生植物不同器官的C、N、P浓度, 如寄主枝的C浓度可能仅适用于预测寄生叶的C浓度.此外, 寄主枝、吸器、寄生枝与寄生叶两两器官间, 仅寄主枝C含量与寄生叶C含量为极显著负相关关系, 说明寄主枝C含量对寄生叶的形成以及生理活动的正常进行可能有着不可或缺的作用.寄主枝与吸器、吸器与寄生枝间P含量极显著正相关, 说明寄主枝、吸器和寄生枝对P的吸收具有一致性(
何苗等, 2015 ). ...
Mineral nutrition and water relations of hemiparasitic mistletoes: A question of partitioning. Experiments with Loranthus europaeus on Quercus petraea and Quercus robur
1
1983
... 将采集到的植物叶片用自来水洗净灰尘等杂质后, 再用超纯水冲洗.由于半寄生植物的大部分矿物质是通过吸器从寄主木质部获取(
Glatzel, 1983 ;
Bell & Adams, 2011 ), 故将寄主枝条的树皮去除后作为半寄生植物的养分来源, 以便更准确地分析寄主枝条-吸器-寄生枝条-寄生叶片的生态化学计量特征变化.将寄主枝条、吸器、寄生枝条与叶片分别装入信封, 于65 ℃条件下烘干至恒质量.烘干后先用刀斧将吸器及寄主枝条、寄生枝条粗加工成小块, 再通过微型植物粉碎机(FZ102型, 天津市泰斯特仪器有限公司, 天津)将样品磨成粉末, 植物叶片则利用高速万能粉碎机(FW80型, 天津市泰斯特仪器有限公司, 天津)磨成粉末, 用于全碳、全氮和全磷含量的测定.C、N含量用碳氮分析仪(Vario MAX CN, Elementar Analysensysteme GmbH, Hanau, Germany)测定, P含量采用电感耦合等离子体原子发射光谱仪(iCAP6300, Thermo Fisher Scientific, Waltham, USA)测定. ...
Mistletoe ecophysiology: Host- parasite interactions
1
2009
... 寄生植物与其寄主之间的养分相关性蕴含了重要信息, 这些信息尚未被充分理解(
Gebauer et al ., 2018 ).寄主植物中的元素浓度通常是用来预测半寄生植物体内元素浓度的最佳指标(
Glatzel & Geils, 2009 ;
Gebauer et al ., 2012 ).本研究中寄主枝与吸器之间的N、P含量以及C:N、C:P和N:P均呈极显著正相关关系, 说明寄生植物吸器与寄主枝存在非常密切的关系, 但寄主枝与寄生枝间仅N含量和C:N存在显著相关关系, 寄主枝与寄生叶间仅C、N含量显著相关.这说明寄主枝的C、N、P浓度并非同时适用于预测寄生植物不同器官的C、N、P浓度, 如寄主枝的C浓度可能仅适用于预测寄生叶的C浓度.此外, 寄主枝、吸器、寄生枝与寄生叶两两器官间, 仅寄主枝C含量与寄生叶C含量为极显著负相关关系, 说明寄主枝C含量对寄生叶的形成以及生理活动的正常进行可能有着不可或缺的作用.寄主枝与吸器、吸器与寄生枝间P含量极显著正相关, 说明寄主枝、吸器和寄生枝对P的吸收具有一致性(
何苗等, 2015 ). ...
N、P施肥对塔里木河上游绿洲棉花C、N、P生态化学计量特征的影响
1
2017
... 寄主枝-吸器-寄生枝-寄生叶这一连续体中的C、N、P生态化学计量特征的变化趋势, 在同一寄生植物中一致, 在不同种寄生植物中并不完全相同, 差异主要体现在寄主枝和寄生叶中.椆树桑寄生叶片C含量和N:P高于寄主枝, 柳叶钝果寄生则相反,这一现象可能与两种寄生植物自身生长发育阶段的不同有关.C元素是植物各种生理生化过程的底物和能量来源, 其同化与N、P元素的吸收途径不同, 随生长阶段的变化, C、N、P元素的分配比例也有较大的差别(
贡璐等, 2017 ).本研究采样时, 哀牢山地区椆树桑寄生处于营养生长阶段, 而柳叶钝果寄生已经进入开花期.两种寄生植物各自的寄主枝具有更相似的C、N、P生态化学计量特征, 这有可能是寄生植物对养分浓度有一定的需求和偏好, 从而养分含量相似的物种易被同一种寄生植物侵染.该区椆树桑寄生的寄主皆为壳斗科植物, 亲缘关系上的相近可能是其寄主枝C、N、P生态化学计量特征相似的原因之一.柳叶钝果寄生的寄主厚皮香、云南越桔以及紫药女贞为不同科属植物, 其生态化学计量特征相似的原因需要进一步研究. ...
N、P施肥对塔里木河上游绿洲棉花C、N、P生态化学计量特征的影响
1
2017
... 寄主枝-吸器-寄生枝-寄生叶这一连续体中的C、N、P生态化学计量特征的变化趋势, 在同一寄生植物中一致, 在不同种寄生植物中并不完全相同, 差异主要体现在寄主枝和寄生叶中.椆树桑寄生叶片C含量和N:P高于寄主枝, 柳叶钝果寄生则相反,这一现象可能与两种寄生植物自身生长发育阶段的不同有关.C元素是植物各种生理生化过程的底物和能量来源, 其同化与N、P元素的吸收途径不同, 随生长阶段的变化, C、N、P元素的分配比例也有较大的差别(
贡璐等, 2017 ).本研究采样时, 哀牢山地区椆树桑寄生处于营养生长阶段, 而柳叶钝果寄生已经进入开花期.两种寄生植物各自的寄主枝具有更相似的C、N、P生态化学计量特征, 这有可能是寄生植物对养分浓度有一定的需求和偏好, 从而养分含量相似的物种易被同一种寄生植物侵染.该区椆树桑寄生的寄主皆为壳斗科植物, 亲缘关系上的相近可能是其寄主枝C、N、P生态化学计量特征相似的原因之一.柳叶钝果寄生的寄主厚皮香、云南越桔以及紫药女贞为不同科属植物, 其生态化学计量特征相似的原因需要进一步研究. ...
Variation in nitrogen and phosphorus concentrations of wetland plants
1
2002
... 寄生植物不同的营养素富集程度也不同, 被认为是吸器选择性吸收离子的结果, 若摄取是通过蒸腾而被动吸收, 那么营养元素的比值应保持相同(
Gebauer et al ., 2018 ).本研究中, 椆树桑寄生吸器的C:N、C:P及N:P均与其寄主枝条存在显著差异, 柳叶钝果寄生吸器的C:N和C:P也与其寄主枝条存在显著差异, 这在一定程度上说明C、N和P元素并非简单的通过蒸腾作用而被转运至寄生植物体内, 但也不能排除离子通过被动运输进入吸器后, 由于吸器具有合成物质的功能而导致元素比值发生改变(
黄新亚等, 2011 ).植物不同器官C、N、P含量的差异不仅受植物基本生理过程需求的影响, 也受相应器官组织结构和功能分化的影响(
Minden & Kleyer, 2015 ).叶片是植物的同化器官与重要的养分储存器官, 因而寄生植物叶片的N和P含量高于寄主枝、吸器与寄生枝(
Niklas & Cobb, 2005 ;
贺合亮等, 2017 ).植物叶片中C、N、P含量的变异系数普遍存在C < N < P的规律(
Koerselman & Meuleman, 1996 ;
Güsewell & Koerselman, 2002 ;
姜沛沛等, 2017 ).本研究中两种半寄生植物的寄主枝、吸器、寄生枝与寄生叶的C、N、P含量皆符合这个规律, 这是因为C作为构成植物骨架的基本结构物质, 且为多种植物生理活动提供能源, 而N含量可被植物控制在较窄的范围内, 故C在植物体内含量高且变异小, N含量在各器官中的变异性小于P含量(
Vitousek, 1998 ;
贺合亮等, 2017 ).本研究中两种寄生植物C:N的变异性小于C:P,是因为植物光合作用中C的同化与营养元素的吸收并非同一途径, 而C含量高且变异小, C:N和C:P的变异主要受到N和P含量变化的影响(
Hedin, 2004 ;
曾昭霞等, 2015 ). ...
Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China
6
2005
... 生态化学计量学是主要研究生态系统能量平衡和多重化学元素平衡的科学(
Sterner & Elser, 2002 ;
曾德慧和陈广生, 2005 ), 其核心问题是揭示生物体元素组成差异对其生态功能的影响(
Elser et al ., 2000 ;
蒋利玲等, 2014 ).生态化学计量学从元素比率的角度将不同尺度、不同生物群系以及不同领域的生物学研究联系起来(
Sterner & Elser, 2002 ;
Michaels, 2003 ), 成为生态学研究的主要方法和研究热点(
曾冬萍等, 2013 ;
刘文亭等, 2015 ).这一领域研究最多的化学元素是碳(C)、氮(N)、磷(P), 作为3种最重要的生命元素, C、N、P是所有生命过程的基础, 在植物的生化功能方面紧密耦合(
李威, 2016 ).基于不同生命形式, 系统发育以及光合作用碳同化途径的植物叶片生态化学计量特征及其变异模式的研究已取得重大进展(
Han et al ., 2005 ,
2011 ), 但这些研究的主要对象为非寄生植物.有关研究表明, 非寄生植物生态化学计量特征与土壤养分状况相关(
Reich & Oleksyn, 2004 ;
赵一娉等, 2017 ), 但寄生植物没有根系直接接触土壤, 其生命活动所需要的养分和水分主要来自于寄主植物.目前, 有关寄生植物生态化学计量特征的研究鲜有报道. ...
... N和P在植物中为协同元素, 植物叶片N、P含量的正相关关系亦为高等陆生植物养分计量的普遍规律(
Sterner & Elser, 2002 ;
Wright et al ., 2005 ).植物N、P间的耦合关系很早就有文献记录(
Garten, 1976 ), 随后也得到许多研究的验证(
Li et al ., 2014 ;
Hidaka & Kitayama, 2015 ;
崔高阳等, 2015 ).对中国753种陆地植物叶片的研究(
Han et al ., 2005 )以及中国东部南北样带654种植物叶片的研究(
任书杰等, 2007 )表明, 不同生活型或不同系统发育植物的叶片N和P含量大部分也呈极显著相关关系.水生植物N、P含量间同样具有强线性关系, 这可能是大多数植物N、P含量的基本特征(
Duarte, 1992 ).
Hidaka和Kitayama (2015) 认为植物叶片N、P含量间的正相关关系可能部分受到土壤可利用性氮含量的影响, 而植物营养策略(N利用策略受到P利用策略的影响)以及植物细胞中的N和P生态化学计量特征(如蛋白质合成中N和P的生化关系)则可提供另一种解释, 但仍需要进一步分析以阐明相关性的生理背景.本研究中, N和P由寄主枝、吸器中的极显著正相关关系, 寄生枝中显著正相关关系, 到寄生叶中变为了相关关系不显著.
Garten (1976) 的研究中大型水生植物的N和P含量相关性也不显著, 这可能是大型水生植物的N含量范围较小, 或者是当环境P缺乏时对N的奢侈消费造成的.
Han等(2005) 和
任书杰等(2007) 的研究中, 裸子植物和蕨类植物的N、P含量相关性也不显著.本研究中寄生叶N、P间的不协同性可能与寄生植物不直接从土壤吸收养分、混合营养策略以及寄生植物叶片重吸收P但不重吸收N有关(
Suriyagoda et al ., 2017 ).受到N、P不耦合的影响, 寄生叶的N含量和C:P, P含量和C:N也不存在显著的相关关系. ...
... 的研究中大型水生植物的N和P含量相关性也不显著, 这可能是大型水生植物的N含量范围较小, 或者是当环境P缺乏时对N的奢侈消费造成的.
Han等(2005) 和
任书杰等(2007) 的研究中, 裸子植物和蕨类植物的N、P含量相关性也不显著.本研究中寄生叶N、P间的不协同性可能与寄生植物不直接从土壤吸收养分、混合营养策略以及寄生植物叶片重吸收P但不重吸收N有关(
Suriyagoda et al ., 2017 ).受到N、P不耦合的影响, 寄生叶的N含量和C:P, P含量和C:N也不存在显著的相关关系. ...
... 寄生植物是一类重要而独特的植物类群.本研究中两种半寄生植物叶片的平均N含量为12.08 mg·g
-1 , 明显低于全球陆生植物的N含量20.09 mg·g
-1 (
Reich & Oleksyn, 2004 )、中国陆生植物的N含量18.6 mg·g
-1 (
Han et al ., 2005 ); 而P含量3.06 mg·g
-1 则明显地高于全球和中国陆生植物的P含量, 分别为1.77 mg·g
-1 (
Reich & Oleksyn, 2004 )、1.21 mg·g
-1 (
Han et al ., 2005 ).寄生叶的N含量较低可能与其光合作用较弱有关, 而较高的P含量可能受到研究区土壤P含量的间接影响, 哀牢山地区森林土壤的全P和有效P含量分别为1.17 g·kg
-1 和3.40 mg·kg
-1 (
鲁志云等, 2017 ), 中国土壤全P和有效P含量分别为0.60 g·kg
-1 和2.39 mg·kg
-1 (
汪涛等, 2008 ), 哀牢山地区森林土壤全P含量显著高于全国水平(
鲁志云等, 2017 ).此外, 哀牢山地区山地森林表层土壤(0-20 cm)的C:N、C:P、N:P值分别为14.9、84.9、5.7, 高于中国陆地(0-10 cm)土壤的C:N、C:P、N:P水平(分别为12.3、52.7、3.9)(
Tian et al ., 2010 ), 更接近于全球陆地(0-10 cm)和全球森林土壤水平(分别为12.3、72.0、5.9和12.4、81.9、6.6)(
Cleveland & Liptzin, 2007 ).已有的研究认为我国陆地植物生长普遍受P限制(
Han et al ., 2005 ;
任书杰等, 2007 ), 但哀牢山地区较高的土壤P含量暗示该地区可能并不缺P.关于哀牢山地区森林寄生植物、寄主的生态化学计量特征与土壤化学元素计量的关系将是以后研究的重要内容. ...
... (
Han et al ., 2005 ).寄生叶的N含量较低可能与其光合作用较弱有关, 而较高的P含量可能受到研究区土壤P含量的间接影响, 哀牢山地区森林土壤的全P和有效P含量分别为1.17 g·kg
-1 和3.40 mg·kg
-1 (
鲁志云等, 2017 ), 中国土壤全P和有效P含量分别为0.60 g·kg
-1 和2.39 mg·kg
-1 (
汪涛等, 2008 ), 哀牢山地区森林土壤全P含量显著高于全国水平(
鲁志云等, 2017 ).此外, 哀牢山地区山地森林表层土壤(0-20 cm)的C:N、C:P、N:P值分别为14.9、84.9、5.7, 高于中国陆地(0-10 cm)土壤的C:N、C:P、N:P水平(分别为12.3、52.7、3.9)(
Tian et al ., 2010 ), 更接近于全球陆地(0-10 cm)和全球森林土壤水平(分别为12.3、72.0、5.9和12.4、81.9、6.6)(
Cleveland & Liptzin, 2007 ).已有的研究认为我国陆地植物生长普遍受P限制(
Han et al ., 2005 ;
任书杰等, 2007 ), 但哀牢山地区较高的土壤P含量暗示该地区可能并不缺P.关于哀牢山地区森林寄生植物、寄主的生态化学计量特征与土壤化学元素计量的关系将是以后研究的重要内容. ...
... ).已有的研究认为我国陆地植物生长普遍受P限制(
Han et al ., 2005 ;
任书杰等, 2007 ), 但哀牢山地区较高的土壤P含量暗示该地区可能并不缺P.关于哀牢山地区森林寄生植物、寄主的生态化学计量特征与土壤化学元素计量的关系将是以后研究的重要内容. ...
Biogeography and variability of eleven mineral elements in plant leaves across gradients of climate, soil and plant functional type in China
1
2011
... 生态化学计量学是主要研究生态系统能量平衡和多重化学元素平衡的科学(
Sterner & Elser, 2002 ;
曾德慧和陈广生, 2005 ), 其核心问题是揭示生物体元素组成差异对其生态功能的影响(
Elser et al ., 2000 ;
蒋利玲等, 2014 ).生态化学计量学从元素比率的角度将不同尺度、不同生物群系以及不同领域的生物学研究联系起来(
Sterner & Elser, 2002 ;
Michaels, 2003 ), 成为生态学研究的主要方法和研究热点(
曾冬萍等, 2013 ;
刘文亭等, 2015 ).这一领域研究最多的化学元素是碳(C)、氮(N)、磷(P), 作为3种最重要的生命元素, C、N、P是所有生命过程的基础, 在植物的生化功能方面紧密耦合(
李威, 2016 ).基于不同生命形式, 系统发育以及光合作用碳同化途径的植物叶片生态化学计量特征及其变异模式的研究已取得重大进展(
Han et al ., 2005 ,
2011 ), 但这些研究的主要对象为非寄生植物.有关研究表明, 非寄生植物生态化学计量特征与土壤养分状况相关(
Reich & Oleksyn, 2004 ;
赵一娉等, 2017 ), 但寄生植物没有根系直接接触土壤, 其生命活动所需要的养分和水分主要来自于寄主植物.目前, 有关寄生植物生态化学计量特征的研究鲜有报道. ...
Diverse effects of parasites in ecosystems: Linking interdependent processes
1
2012
... 半寄生植物是寄生植物的主要类群, 其叶片具有叶绿体, 在生命周期的至少一个阶段可进行光合作用, 仅从寄主体内获取部分水和溶解的矿物质(
Nickrent et al ., 1998 ).在自然界中, 半寄生植物对植物群落结构和多样性具有显著影响(
Phoenix & Press, 2010 ;
Press & Phoenix, 2010 ;
Demey et al ., 2014 ), 而其营养丰富、可快速分解和释放养分的凋落物可能会提高森林土壤养分可利用率(
Demey et al ., 2015 ), 影响生态系统水平凋落物分解和养分循环(
Quested et al ., 2010 ).虽然半寄生植物的丰度和生物量相对较小, 但它们对生物地球化学循环、植物种群结构和多样性的影响是很明确的(
Nave et al ., 2017 ), 因而被描述为影响群落动态的生态系统工程师, 以及影响养分循环的关键物种(
Watson, 2010 ;
Hatcher et al ., 2012 ). ...
青藏高原东部窄叶鲜卑花灌丛土壤C、N、P生态化学计量学特征
3
2017
... 寄生植物不同的营养素富集程度也不同, 被认为是吸器选择性吸收离子的结果, 若摄取是通过蒸腾而被动吸收, 那么营养元素的比值应保持相同(
Gebauer et al ., 2018 ).本研究中, 椆树桑寄生吸器的C:N、C:P及N:P均与其寄主枝条存在显著差异, 柳叶钝果寄生吸器的C:N和C:P也与其寄主枝条存在显著差异, 这在一定程度上说明C、N和P元素并非简单的通过蒸腾作用而被转运至寄生植物体内, 但也不能排除离子通过被动运输进入吸器后, 由于吸器具有合成物质的功能而导致元素比值发生改变(
黄新亚等, 2011 ).植物不同器官C、N、P含量的差异不仅受植物基本生理过程需求的影响, 也受相应器官组织结构和功能分化的影响(
Minden & Kleyer, 2015 ).叶片是植物的同化器官与重要的养分储存器官, 因而寄生植物叶片的N和P含量高于寄主枝、吸器与寄生枝(
Niklas & Cobb, 2005 ;
贺合亮等, 2017 ).植物叶片中C、N、P含量的变异系数普遍存在C < N < P的规律(
Koerselman & Meuleman, 1996 ;
Güsewell & Koerselman, 2002 ;
姜沛沛等, 2017 ).本研究中两种半寄生植物的寄主枝、吸器、寄生枝与寄生叶的C、N、P含量皆符合这个规律, 这是因为C作为构成植物骨架的基本结构物质, 且为多种植物生理活动提供能源, 而N含量可被植物控制在较窄的范围内, 故C在植物体内含量高且变异小, N含量在各器官中的变异性小于P含量(
Vitousek, 1998 ;
贺合亮等, 2017 ).本研究中两种寄生植物C:N的变异性小于C:P,是因为植物光合作用中C的同化与营养元素的吸收并非同一途径, 而C含量高且变异小, C:N和C:P的变异主要受到N和P含量变化的影响(
Hedin, 2004 ;
曾昭霞等, 2015 ). ...
... ;
贺合亮等, 2017 ).本研究中两种寄生植物C:N的变异性小于C:P,是因为植物光合作用中C的同化与营养元素的吸收并非同一途径, 而C含量高且变异小, C:N和C:P的变异主要受到N和P含量变化的影响(
Hedin, 2004 ;
曾昭霞等, 2015 ). ...
... 植物体中C与N、P含量一般呈负相关关系是高等陆生植物C、N、P计量的普遍规律(
Sterner & Elser, 2002 ;
贺合亮等, 2017 ).在本研究中, 仅寄主枝符合此规律, 其C含量与N、P含量及C:N、C:P、N:P均存在极显著的相关关系.寄生植物吸器的C含量仅与N:P具有显著的相关关系, 枝条的C含量则与N、P含量及C:N、C:P、N:P均不存在极显著的相关关系.寄生植物叶片的C含量与N含量为正相关关系, 与P含量的相关性未达到显著, 也不同于此规律, 但该结果与荒漠植物叶片C、N、P生态化学计量特征具有一致性(
张珂等, 2014 ;
罗艳等, 2017 ).陆生植物的C主要来自大气中的CO
2 (
廖宗文等, 2014 ), 半寄生植物的C既可以来自自身光合作用, 也可以从寄主处获取(
Tě?itel et al ., 2015 ).规律不同说明半寄生植物在固C过程中对养分(N、P等)的利用策略有别于一般陆生植物. ...
青藏高原东部窄叶鲜卑花灌丛土壤C、N、P生态化学计量学特征
3
2017
... 寄生植物不同的营养素富集程度也不同, 被认为是吸器选择性吸收离子的结果, 若摄取是通过蒸腾而被动吸收, 那么营养元素的比值应保持相同(
Gebauer et al ., 2018 ).本研究中, 椆树桑寄生吸器的C:N、C:P及N:P均与其寄主枝条存在显著差异, 柳叶钝果寄生吸器的C:N和C:P也与其寄主枝条存在显著差异, 这在一定程度上说明C、N和P元素并非简单的通过蒸腾作用而被转运至寄生植物体内, 但也不能排除离子通过被动运输进入吸器后, 由于吸器具有合成物质的功能而导致元素比值发生改变(
黄新亚等, 2011 ).植物不同器官C、N、P含量的差异不仅受植物基本生理过程需求的影响, 也受相应器官组织结构和功能分化的影响(
Minden & Kleyer, 2015 ).叶片是植物的同化器官与重要的养分储存器官, 因而寄生植物叶片的N和P含量高于寄主枝、吸器与寄生枝(
Niklas & Cobb, 2005 ;
贺合亮等, 2017 ).植物叶片中C、N、P含量的变异系数普遍存在C < N < P的规律(
Koerselman & Meuleman, 1996 ;
Güsewell & Koerselman, 2002 ;
姜沛沛等, 2017 ).本研究中两种半寄生植物的寄主枝、吸器、寄生枝与寄生叶的C、N、P含量皆符合这个规律, 这是因为C作为构成植物骨架的基本结构物质, 且为多种植物生理活动提供能源, 而N含量可被植物控制在较窄的范围内, 故C在植物体内含量高且变异小, N含量在各器官中的变异性小于P含量(
Vitousek, 1998 ;
贺合亮等, 2017 ).本研究中两种寄生植物C:N的变异性小于C:P,是因为植物光合作用中C的同化与营养元素的吸收并非同一途径, 而C含量高且变异小, C:N和C:P的变异主要受到N和P含量变化的影响(
Hedin, 2004 ;
曾昭霞等, 2015 ). ...
... ;
贺合亮等, 2017 ).本研究中两种寄生植物C:N的变异性小于C:P,是因为植物光合作用中C的同化与营养元素的吸收并非同一途径, 而C含量高且变异小, C:N和C:P的变异主要受到N和P含量变化的影响(
Hedin, 2004 ;
曾昭霞等, 2015 ). ...
... 植物体中C与N、P含量一般呈负相关关系是高等陆生植物C、N、P计量的普遍规律(
Sterner & Elser, 2002 ;
贺合亮等, 2017 ).在本研究中, 仅寄主枝符合此规律, 其C含量与N、P含量及C:N、C:P、N:P均存在极显著的相关关系.寄生植物吸器的C含量仅与N:P具有显著的相关关系, 枝条的C含量则与N、P含量及C:N、C:P、N:P均不存在极显著的相关关系.寄生植物叶片的C含量与N含量为正相关关系, 与P含量的相关性未达到显著, 也不同于此规律, 但该结果与荒漠植物叶片C、N、P生态化学计量特征具有一致性(
张珂等, 2014 ;
罗艳等, 2017 ).陆生植物的C主要来自大气中的CO
2 (
廖宗文等, 2014 ), 半寄生植物的C既可以来自自身光合作用, 也可以从寄主处获取(
Tě?itel et al ., 2015 ).规律不同说明半寄生植物在固C过程中对养分(N、P等)的利用策略有别于一般陆生植物. ...
新温185号核桃叶片和果实矿质元素浓度时节变化及其相关性
1
2015
... 寄生植物与其寄主之间的养分相关性蕴含了重要信息, 这些信息尚未被充分理解(
Gebauer et al ., 2018 ).寄主植物中的元素浓度通常是用来预测半寄生植物体内元素浓度的最佳指标(
Glatzel & Geils, 2009 ;
Gebauer et al ., 2012 ).本研究中寄主枝与吸器之间的N、P含量以及C:N、C:P和N:P均呈极显著正相关关系, 说明寄生植物吸器与寄主枝存在非常密切的关系, 但寄主枝与寄生枝间仅N含量和C:N存在显著相关关系, 寄主枝与寄生叶间仅C、N含量显著相关.这说明寄主枝的C、N、P浓度并非同时适用于预测寄生植物不同器官的C、N、P浓度, 如寄主枝的C浓度可能仅适用于预测寄生叶的C浓度.此外, 寄主枝、吸器、寄生枝与寄生叶两两器官间, 仅寄主枝C含量与寄生叶C含量为极显著负相关关系, 说明寄主枝C含量对寄生叶的形成以及生理活动的正常进行可能有着不可或缺的作用.寄主枝与吸器、吸器与寄生枝间P含量极显著正相关, 说明寄主枝、吸器和寄生枝对P的吸收具有一致性(
何苗等, 2015 ). ...
新温185号核桃叶片和果实矿质元素浓度时节变化及其相关性
1
2015
... 寄生植物与其寄主之间的养分相关性蕴含了重要信息, 这些信息尚未被充分理解(
Gebauer et al ., 2018 ).寄主植物中的元素浓度通常是用来预测半寄生植物体内元素浓度的最佳指标(
Glatzel & Geils, 2009 ;
Gebauer et al ., 2012 ).本研究中寄主枝与吸器之间的N、P含量以及C:N、C:P和N:P均呈极显著正相关关系, 说明寄生植物吸器与寄主枝存在非常密切的关系, 但寄主枝与寄生枝间仅N含量和C:N存在显著相关关系, 寄主枝与寄生叶间仅C、N含量显著相关.这说明寄主枝的C、N、P浓度并非同时适用于预测寄生植物不同器官的C、N、P浓度, 如寄主枝的C浓度可能仅适用于预测寄生叶的C浓度.此外, 寄主枝、吸器、寄生枝与寄生叶两两器官间, 仅寄主枝C含量与寄生叶C含量为极显著负相关关系, 说明寄主枝C含量对寄生叶的形成以及生理活动的正常进行可能有着不可或缺的作用.寄主枝与吸器、吸器与寄生枝间P含量极显著正相关, 说明寄主枝、吸器和寄生枝对P的吸收具有一致性(
何苗等, 2015 ). ...
Global organization of terrestrial plant-nutrient interactions
1
2004
... 寄生植物不同的营养素富集程度也不同, 被认为是吸器选择性吸收离子的结果, 若摄取是通过蒸腾而被动吸收, 那么营养元素的比值应保持相同(
Gebauer et al ., 2018 ).本研究中, 椆树桑寄生吸器的C:N、C:P及N:P均与其寄主枝条存在显著差异, 柳叶钝果寄生吸器的C:N和C:P也与其寄主枝条存在显著差异, 这在一定程度上说明C、N和P元素并非简单的通过蒸腾作用而被转运至寄生植物体内, 但也不能排除离子通过被动运输进入吸器后, 由于吸器具有合成物质的功能而导致元素比值发生改变(
黄新亚等, 2011 ).植物不同器官C、N、P含量的差异不仅受植物基本生理过程需求的影响, 也受相应器官组织结构和功能分化的影响(
Minden & Kleyer, 2015 ).叶片是植物的同化器官与重要的养分储存器官, 因而寄生植物叶片的N和P含量高于寄主枝、吸器与寄生枝(
Niklas & Cobb, 2005 ;
贺合亮等, 2017 ).植物叶片中C、N、P含量的变异系数普遍存在C < N < P的规律(
Koerselman & Meuleman, 1996 ;
Güsewell & Koerselman, 2002 ;
姜沛沛等, 2017 ).本研究中两种半寄生植物的寄主枝、吸器、寄生枝与寄生叶的C、N、P含量皆符合这个规律, 这是因为C作为构成植物骨架的基本结构物质, 且为多种植物生理活动提供能源, 而N含量可被植物控制在较窄的范围内, 故C在植物体内含量高且变异小, N含量在各器官中的变异性小于P含量(
Vitousek, 1998 ;
贺合亮等, 2017 ).本研究中两种寄生植物C:N的变异性小于C:P,是因为植物光合作用中C的同化与营养元素的吸收并非同一途径, 而C含量高且变异小, C:N和C:P的变异主要受到N和P含量变化的影响(
Hedin, 2004 ;
曾昭霞等, 2015 ). ...
Physiological linkage in co-variation of foliar nitrogen and phosphorus in tropical tree species along a gradient of soil phosphorus availability
2
2015
... N和P在植物中为协同元素, 植物叶片N、P含量的正相关关系亦为高等陆生植物养分计量的普遍规律(
Sterner & Elser, 2002 ;
Wright et al ., 2005 ).植物N、P间的耦合关系很早就有文献记录(
Garten, 1976 ), 随后也得到许多研究的验证(
Li et al ., 2014 ;
Hidaka & Kitayama, 2015 ;
崔高阳等, 2015 ).对中国753种陆地植物叶片的研究(
Han et al ., 2005 )以及中国东部南北样带654种植物叶片的研究(
任书杰等, 2007 )表明, 不同生活型或不同系统发育植物的叶片N和P含量大部分也呈极显著相关关系.水生植物N、P含量间同样具有强线性关系, 这可能是大多数植物N、P含量的基本特征(
Duarte, 1992 ).
Hidaka和Kitayama (2015) 认为植物叶片N、P含量间的正相关关系可能部分受到土壤可利用性氮含量的影响, 而植物营养策略(N利用策略受到P利用策略的影响)以及植物细胞中的N和P生态化学计量特征(如蛋白质合成中N和P的生化关系)则可提供另一种解释, 但仍需要进一步分析以阐明相关性的生理背景.本研究中, N和P由寄主枝、吸器中的极显著正相关关系, 寄生枝中显著正相关关系, 到寄生叶中变为了相关关系不显著.
Garten (1976) 的研究中大型水生植物的N和P含量相关性也不显著, 这可能是大型水生植物的N含量范围较小, 或者是当环境P缺乏时对N的奢侈消费造成的.
Han等(2005) 和
任书杰等(2007) 的研究中, 裸子植物和蕨类植物的N、P含量相关性也不显著.本研究中寄生叶N、P间的不协同性可能与寄生植物不直接从土壤吸收养分、混合营养策略以及寄生植物叶片重吸收P但不重吸收N有关(
Suriyagoda et al ., 2017 ).受到N、P不耦合的影响, 寄生叶的N含量和C:P, P含量和C:N也不存在显著的相关关系. ...
... ).
Hidaka和Kitayama (2015) 认为植物叶片N、P含量间的正相关关系可能部分受到土壤可利用性氮含量的影响, 而植物营养策略(N利用策略受到P利用策略的影响)以及植物细胞中的N和P生态化学计量特征(如蛋白质合成中N和P的生化关系)则可提供另一种解释, 但仍需要进一步分析以阐明相关性的生理背景.本研究中, N和P由寄主枝、吸器中的极显著正相关关系, 寄生枝中显著正相关关系, 到寄生叶中变为了相关关系不显著.
Garten (1976) 的研究中大型水生植物的N和P含量相关性也不显著, 这可能是大型水生植物的N含量范围较小, 或者是当环境P缺乏时对N的奢侈消费造成的.
Han等(2005) 和
任书杰等(2007) 的研究中, 裸子植物和蕨类植物的N、P含量相关性也不显著.本研究中寄生叶N、P间的不协同性可能与寄生植物不直接从土壤吸收养分、混合营养策略以及寄生植物叶片重吸收P但不重吸收N有关(
Suriyagoda et al ., 2017 ).受到N、P不耦合的影响, 寄生叶的N含量和C:P, P含量和C:N也不存在显著的相关关系. ...
寄生植物对寄主的选择和影响
1
2004
... 大多数寄生植物具有广寄性, 而不同的寄生植物其寄主种类有很大的差异(
Norton & Lange, 1999 ;
胡飞和孔垂华, 2004 ).本次调查记录中椆树桑寄生的寄主均为壳斗科植物, 柳叶钝果寄生的寄主较为广泛, 但未发现与椆树桑寄生具有相同寄主.不同的寄主物种对同一寄生植物的影响可能不同, 有研究发现同一桑寄生植物的挥发性成分组成及含量、总黄酮含量、毒性等会因其寄主物种不同而不同(
李永华等, 2009 ,
2012 ;
周汉华等, 2013 ), 但对寄生植物生态化学计量特征与寄主物种的关系进行探讨的研究较少.在本研究中, 寄生于不同寄主的同一寄生植物的同一器官C、N、P生态化学计量特征基本没有显著的不同, 而对同一寄生植物的不同寄主情况下不同器官之间的生态化学计量特征进行双因素方差分析, 也发现寄主物种的影响不显著.这可能与同一寄生植物的不同寄主枝条本身生态化学计量特征差异不显著有关. ...
寄生植物对寄主的选择和影响
1
2004
... 大多数寄生植物具有广寄性, 而不同的寄生植物其寄主种类有很大的差异(
Norton & Lange, 1999 ;
胡飞和孔垂华, 2004 ).本次调查记录中椆树桑寄生的寄主均为壳斗科植物, 柳叶钝果寄生的寄主较为广泛, 但未发现与椆树桑寄生具有相同寄主.不同的寄主物种对同一寄生植物的影响可能不同, 有研究发现同一桑寄生植物的挥发性成分组成及含量、总黄酮含量、毒性等会因其寄主物种不同而不同(
李永华等, 2009 ,
2012 ;
周汉华等, 2013 ), 但对寄生植物生态化学计量特征与寄主物种的关系进行探讨的研究较少.在本研究中, 寄生于不同寄主的同一寄生植物的同一器官C、N、P生态化学计量特征基本没有显著的不同, 而对同一寄生植物的不同寄主情况下不同器官之间的生态化学计量特征进行双因素方差分析, 也发现寄主物种的影响不显著.这可能与同一寄生植物的不同寄主枝条本身生态化学计量特征差异不显著有关. ...
长白山森林不同演替阶段植物与土壤氮磷的化学计量特征
2
2014
... 大多数有关寄生植物的研究主要关注于植物的光合器官, 而对寄生植物不同器官与寄主植物间的关系研究较少.有关研究表明, 植物叶片N、P浓度与枝条N、P浓度具有相关性(
Kerkhoff et al ., 2006 ), 根和叶间的N浓度也具有显著的相关性(
Craine et al ., 2005 ).本研究寄生枝与寄生叶之间的P含量没有呈现出显著的相关性, 但N含量存在极显著正相关关系, 该结果与
杨梅等(2015) 、
胡耀升等(2014) 的研究果相一致.叶和茎间N含量的相关关系反映了叶、茎在生物、生化方面的功能需求, 这是由植株冠层结构所决定的(
Westoby et al ., 2002 ;
胡耀升等, 2014 ). ...
... ;
胡耀升等, 2014 ). ...
长白山森林不同演替阶段植物与土壤氮磷的化学计量特征
2
2014
... 大多数有关寄生植物的研究主要关注于植物的光合器官, 而对寄生植物不同器官与寄主植物间的关系研究较少.有关研究表明, 植物叶片N、P浓度与枝条N、P浓度具有相关性(
Kerkhoff et al ., 2006 ), 根和叶间的N浓度也具有显著的相关性(
Craine et al ., 2005 ).本研究寄生枝与寄生叶之间的P含量没有呈现出显著的相关性, 但N含量存在极显著正相关关系, 该结果与
杨梅等(2015) 、
胡耀升等(2014) 的研究果相一致.叶和茎间N含量的相关关系反映了叶、茎在生物、生化方面的功能需求, 这是由植株冠层结构所决定的(
Westoby et al ., 2002 ;
胡耀升等, 2014 ). ...
... ;
胡耀升等, 2014 ). ...
寄生植物的生物学特性及生态学效应
1
2011
... 寄生植物不同的营养素富集程度也不同, 被认为是吸器选择性吸收离子的结果, 若摄取是通过蒸腾而被动吸收, 那么营养元素的比值应保持相同(
Gebauer et al ., 2018 ).本研究中, 椆树桑寄生吸器的C:N、C:P及N:P均与其寄主枝条存在显著差异, 柳叶钝果寄生吸器的C:N和C:P也与其寄主枝条存在显著差异, 这在一定程度上说明C、N和P元素并非简单的通过蒸腾作用而被转运至寄生植物体内, 但也不能排除离子通过被动运输进入吸器后, 由于吸器具有合成物质的功能而导致元素比值发生改变(
黄新亚等, 2011 ).植物不同器官C、N、P含量的差异不仅受植物基本生理过程需求的影响, 也受相应器官组织结构和功能分化的影响(
Minden & Kleyer, 2015 ).叶片是植物的同化器官与重要的养分储存器官, 因而寄生植物叶片的N和P含量高于寄主枝、吸器与寄生枝(
Niklas & Cobb, 2005 ;
贺合亮等, 2017 ).植物叶片中C、N、P含量的变异系数普遍存在C < N < P的规律(
Koerselman & Meuleman, 1996 ;
Güsewell & Koerselman, 2002 ;
姜沛沛等, 2017 ).本研究中两种半寄生植物的寄主枝、吸器、寄生枝与寄生叶的C、N、P含量皆符合这个规律, 这是因为C作为构成植物骨架的基本结构物质, 且为多种植物生理活动提供能源, 而N含量可被植物控制在较窄的范围内, 故C在植物体内含量高且变异小, N含量在各器官中的变异性小于P含量(
Vitousek, 1998 ;
贺合亮等, 2017 ).本研究中两种寄生植物C:N的变异性小于C:P,是因为植物光合作用中C的同化与营养元素的吸收并非同一途径, 而C含量高且变异小, C:N和C:P的变异主要受到N和P含量变化的影响(
Hedin, 2004 ;
曾昭霞等, 2015 ). ...
寄生植物的生物学特性及生态学效应
1
2011
... 寄生植物不同的营养素富集程度也不同, 被认为是吸器选择性吸收离子的结果, 若摄取是通过蒸腾而被动吸收, 那么营养元素的比值应保持相同(
Gebauer et al ., 2018 ).本研究中, 椆树桑寄生吸器的C:N、C:P及N:P均与其寄主枝条存在显著差异, 柳叶钝果寄生吸器的C:N和C:P也与其寄主枝条存在显著差异, 这在一定程度上说明C、N和P元素并非简单的通过蒸腾作用而被转运至寄生植物体内, 但也不能排除离子通过被动运输进入吸器后, 由于吸器具有合成物质的功能而导致元素比值发生改变(
黄新亚等, 2011 ).植物不同器官C、N、P含量的差异不仅受植物基本生理过程需求的影响, 也受相应器官组织结构和功能分化的影响(
Minden & Kleyer, 2015 ).叶片是植物的同化器官与重要的养分储存器官, 因而寄生植物叶片的N和P含量高于寄主枝、吸器与寄生枝(
Niklas & Cobb, 2005 ;
贺合亮等, 2017 ).植物叶片中C、N、P含量的变异系数普遍存在C < N < P的规律(
Koerselman & Meuleman, 1996 ;
Güsewell & Koerselman, 2002 ;
姜沛沛等, 2017 ).本研究中两种半寄生植物的寄主枝、吸器、寄生枝与寄生叶的C、N、P含量皆符合这个规律, 这是因为C作为构成植物骨架的基本结构物质, 且为多种植物生理活动提供能源, 而N含量可被植物控制在较窄的范围内, 故C在植物体内含量高且变异小, N含量在各器官中的变异性小于P含量(
Vitousek, 1998 ;
贺合亮等, 2017 ).本研究中两种寄生植物C:N的变异性小于C:P,是因为植物光合作用中C的同化与营养元素的吸收并非同一途径, 而C含量高且变异小, C:N和C:P的变异主要受到N和P含量变化的影响(
Hedin, 2004 ;
曾昭霞等, 2015 ). ...
闽江河口湿地3种植物化学计量内稳性特征
1
2014
... 生态化学计量学是主要研究生态系统能量平衡和多重化学元素平衡的科学(
Sterner & Elser, 2002 ;
曾德慧和陈广生, 2005 ), 其核心问题是揭示生物体元素组成差异对其生态功能的影响(
Elser et al ., 2000 ;
蒋利玲等, 2014 ).生态化学计量学从元素比率的角度将不同尺度、不同生物群系以及不同领域的生物学研究联系起来(
Sterner & Elser, 2002 ;
Michaels, 2003 ), 成为生态学研究的主要方法和研究热点(
曾冬萍等, 2013 ;
刘文亭等, 2015 ).这一领域研究最多的化学元素是碳(C)、氮(N)、磷(P), 作为3种最重要的生命元素, C、N、P是所有生命过程的基础, 在植物的生化功能方面紧密耦合(
李威, 2016 ).基于不同生命形式, 系统发育以及光合作用碳同化途径的植物叶片生态化学计量特征及其变异模式的研究已取得重大进展(
Han et al ., 2005 ,
2011 ), 但这些研究的主要对象为非寄生植物.有关研究表明, 非寄生植物生态化学计量特征与土壤养分状况相关(
Reich & Oleksyn, 2004 ;
赵一娉等, 2017 ), 但寄生植物没有根系直接接触土壤, 其生命活动所需要的养分和水分主要来自于寄主植物.目前, 有关寄生植物生态化学计量特征的研究鲜有报道. ...
闽江河口湿地3种植物化学计量内稳性特征
1
2014
... 生态化学计量学是主要研究生态系统能量平衡和多重化学元素平衡的科学(
Sterner & Elser, 2002 ;
曾德慧和陈广生, 2005 ), 其核心问题是揭示生物体元素组成差异对其生态功能的影响(
Elser et al ., 2000 ;
蒋利玲等, 2014 ).生态化学计量学从元素比率的角度将不同尺度、不同生物群系以及不同领域的生物学研究联系起来(
Sterner & Elser, 2002 ;
Michaels, 2003 ), 成为生态学研究的主要方法和研究热点(
曾冬萍等, 2013 ;
刘文亭等, 2015 ).这一领域研究最多的化学元素是碳(C)、氮(N)、磷(P), 作为3种最重要的生命元素, C、N、P是所有生命过程的基础, 在植物的生化功能方面紧密耦合(
李威, 2016 ).基于不同生命形式, 系统发育以及光合作用碳同化途径的植物叶片生态化学计量特征及其变异模式的研究已取得重大进展(
Han et al ., 2005 ,
2011 ), 但这些研究的主要对象为非寄生植物.有关研究表明, 非寄生植物生态化学计量特征与土壤养分状况相关(
Reich & Oleksyn, 2004 ;
赵一娉等, 2017 ), 但寄生植物没有根系直接接触土壤, 其生命活动所需要的养分和水分主要来自于寄主植物.目前, 有关寄生植物生态化学计量特征的研究鲜有报道. ...
陕西省3种主要树种叶片、凋落物和土壤N、P化学计量特征
1
2017
... 寄生植物不同的营养素富集程度也不同, 被认为是吸器选择性吸收离子的结果, 若摄取是通过蒸腾而被动吸收, 那么营养元素的比值应保持相同(
Gebauer et al ., 2018 ).本研究中, 椆树桑寄生吸器的C:N、C:P及N:P均与其寄主枝条存在显著差异, 柳叶钝果寄生吸器的C:N和C:P也与其寄主枝条存在显著差异, 这在一定程度上说明C、N和P元素并非简单的通过蒸腾作用而被转运至寄生植物体内, 但也不能排除离子通过被动运输进入吸器后, 由于吸器具有合成物质的功能而导致元素比值发生改变(
黄新亚等, 2011 ).植物不同器官C、N、P含量的差异不仅受植物基本生理过程需求的影响, 也受相应器官组织结构和功能分化的影响(
Minden & Kleyer, 2015 ).叶片是植物的同化器官与重要的养分储存器官, 因而寄生植物叶片的N和P含量高于寄主枝、吸器与寄生枝(
Niklas & Cobb, 2005 ;
贺合亮等, 2017 ).植物叶片中C、N、P含量的变异系数普遍存在C < N < P的规律(
Koerselman & Meuleman, 1996 ;
Güsewell & Koerselman, 2002 ;
姜沛沛等, 2017 ).本研究中两种半寄生植物的寄主枝、吸器、寄生枝与寄生叶的C、N、P含量皆符合这个规律, 这是因为C作为构成植物骨架的基本结构物质, 且为多种植物生理活动提供能源, 而N含量可被植物控制在较窄的范围内, 故C在植物体内含量高且变异小, N含量在各器官中的变异性小于P含量(
Vitousek, 1998 ;
贺合亮等, 2017 ).本研究中两种寄生植物C:N的变异性小于C:P,是因为植物光合作用中C的同化与营养元素的吸收并非同一途径, 而C含量高且变异小, C:N和C:P的变异主要受到N和P含量变化的影响(
Hedin, 2004 ;
曾昭霞等, 2015 ). ...
陕西省3种主要树种叶片、凋落物和土壤N、P化学计量特征
1
2017
... 寄生植物不同的营养素富集程度也不同, 被认为是吸器选择性吸收离子的结果, 若摄取是通过蒸腾而被动吸收, 那么营养元素的比值应保持相同(
Gebauer et al ., 2018 ).本研究中, 椆树桑寄生吸器的C:N、C:P及N:P均与其寄主枝条存在显著差异, 柳叶钝果寄生吸器的C:N和C:P也与其寄主枝条存在显著差异, 这在一定程度上说明C、N和P元素并非简单的通过蒸腾作用而被转运至寄生植物体内, 但也不能排除离子通过被动运输进入吸器后, 由于吸器具有合成物质的功能而导致元素比值发生改变(
黄新亚等, 2011 ).植物不同器官C、N、P含量的差异不仅受植物基本生理过程需求的影响, 也受相应器官组织结构和功能分化的影响(
Minden & Kleyer, 2015 ).叶片是植物的同化器官与重要的养分储存器官, 因而寄生植物叶片的N和P含量高于寄主枝、吸器与寄生枝(
Niklas & Cobb, 2005 ;
贺合亮等, 2017 ).植物叶片中C、N、P含量的变异系数普遍存在C < N < P的规律(
Koerselman & Meuleman, 1996 ;
Güsewell & Koerselman, 2002 ;
姜沛沛等, 2017 ).本研究中两种半寄生植物的寄主枝、吸器、寄生枝与寄生叶的C、N、P含量皆符合这个规律, 这是因为C作为构成植物骨架的基本结构物质, 且为多种植物生理活动提供能源, 而N含量可被植物控制在较窄的范围内, 故C在植物体内含量高且变异小, N含量在各器官中的变异性小于P含量(
Vitousek, 1998 ;
贺合亮等, 2017 ).本研究中两种寄生植物C:N的变异性小于C:P,是因为植物光合作用中C的同化与营养元素的吸收并非同一途径, 而C含量高且变异小, C:N和C:P的变异主要受到N和P含量变化的影响(
Hedin, 2004 ;
曾昭霞等, 2015 ). ...
Phylogenetic and growth form variation in the scaling of nitrogen and phosphorus in the seed plants
1
2006
... 大多数有关寄生植物的研究主要关注于植物的光合器官, 而对寄生植物不同器官与寄主植物间的关系研究较少.有关研究表明, 植物叶片N、P浓度与枝条N、P浓度具有相关性(
Kerkhoff et al ., 2006 ), 根和叶间的N浓度也具有显著的相关性(
Craine et al ., 2005 ).本研究寄生枝与寄生叶之间的P含量没有呈现出显著的相关性, 但N含量存在极显著正相关关系, 该结果与
杨梅等(2015) 、
胡耀升等(2014) 的研究果相一致.叶和茎间N含量的相关关系反映了叶、茎在生物、生化方面的功能需求, 这是由植株冠层结构所决定的(
Westoby et al ., 2002 ;
胡耀升等, 2014 ). ...
The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation
1
1996
... 寄生植物不同的营养素富集程度也不同, 被认为是吸器选择性吸收离子的结果, 若摄取是通过蒸腾而被动吸收, 那么营养元素的比值应保持相同(
Gebauer et al ., 2018 ).本研究中, 椆树桑寄生吸器的C:N、C:P及N:P均与其寄主枝条存在显著差异, 柳叶钝果寄生吸器的C:N和C:P也与其寄主枝条存在显著差异, 这在一定程度上说明C、N和P元素并非简单的通过蒸腾作用而被转运至寄生植物体内, 但也不能排除离子通过被动运输进入吸器后, 由于吸器具有合成物质的功能而导致元素比值发生改变(
黄新亚等, 2011 ).植物不同器官C、N、P含量的差异不仅受植物基本生理过程需求的影响, 也受相应器官组织结构和功能分化的影响(
Minden & Kleyer, 2015 ).叶片是植物的同化器官与重要的养分储存器官, 因而寄生植物叶片的N和P含量高于寄主枝、吸器与寄生枝(
Niklas & Cobb, 2005 ;
贺合亮等, 2017 ).植物叶片中C、N、P含量的变异系数普遍存在C < N < P的规律(
Koerselman & Meuleman, 1996 ;
Güsewell & Koerselman, 2002 ;
姜沛沛等, 2017 ).本研究中两种半寄生植物的寄主枝、吸器、寄生枝与寄生叶的C、N、P含量皆符合这个规律, 这是因为C作为构成植物骨架的基本结构物质, 且为多种植物生理活动提供能源, 而N含量可被植物控制在较窄的范围内, 故C在植物体内含量高且变异小, N含量在各器官中的变异性小于P含量(
Vitousek, 1998 ;
贺合亮等, 2017 ).本研究中两种寄生植物C:N的变异性小于C:P,是因为植物光合作用中C的同化与营养元素的吸收并非同一途径, 而C含量高且变异小, C:N和C:P的变异主要受到N和P含量变化的影响(
Hedin, 2004 ;
曾昭霞等, 2015 ). ...
Root hemiparasites in productive communities should attack competitive host, and harm them to make regeneration gaps
1
2015
... 半寄生植物与寄主的复杂关系包括寄生和竞争(
Lep? & Tě?itel, 2015 ).作为混合营养策略的代表, 半寄生植物既能从自身的光合作用中获取碳同化物, 也能通过吸器从寄主体内获取碳水化合物(
Nave et al ., 2017 ;
Selosse et al ., 2017 ).半寄生植物自身的光合作用在同化寄生矿物资源, 并将矿物资源转换化成自身生长和适合度中起着至关重要的作用, 而将木质部所获得的资源转化为生长和适应的关键过程, 可能是其进化稳定性的基础(
Tě?itel et al ., 2015 ).尽管半寄生植物与寄主关系的研究开展较早, 不少****对半寄生植物与寄主之间的水分利用、光合作用以及矿物质营养等方面的关系进行了较为深入的研究(
Tennakoon et al ., 2011 ;
Chen et al ., 2013 ;
Suriyagoda et al ., 2017 ;
Gebauer et al ., 2018 ), 但营养物质从寄主到半寄生植物的转移机制仍未完全研究清楚(
Gebauer et al ., 2012 ,
2018 ).有关研究表明, 半寄生植物
Rhinanthus alectorolophus 中5%-40%的C来自寄主, 而寄主体内的N亦可被半寄生植物获取(
Tě?itel et al ., 2015 ;
Světlíková et al ., 2016 ).寄生植物的吸器同时连接寄主枝条与寄生枝条, 研究寄主枝条-吸器-寄生枝条-寄生叶片这一连续体的生态化学计量特征变化将有助于进一步理解寄生植物与寄主间的养分关系. ...
云南哀牢山原生林及次生林群落附生地衣物种多样性与分布
1
2007
... 哀牢山国家级自然保护区位于云南省中部哀牢山山脉中北段上部, 地处云贵高原、横断山和青藏高原南缘三大地理区域的结合部, 是生物多样性较丰富及植物区系地理成分荟萃之地, 该保护区内分布着我国目前保存最完整、面积最大的亚热带山地湿性常绿阔叶林(
朱华和闫丽春, 2009 ).本研究区位于哀牢山北段的徐家坝地区(23.35°-24.44° N, 100.54°-101.30° E), 该地区海拔2 000-2 750 m, 面积约5 100 hm
2 , 年降水量1 947 mm, 年蒸发量1 192 mm, 相对湿度85%, 年平均气温11.3 ℃ (
李苏等, 2007 ).该地区常绿阔叶林的土壤全C、全N与全P含量分别为98.65、6.53和1.17 mg·g
-1 , C:N、C:P、N:P为14.96、91.22和5.95 (
鲁志云等, 2017 ).徐家坝地区分布有椆树桑寄生(
Loranthus delavayi )、柳叶钝果寄生(
Taxillus delavayi )、卵叶梨果寄生(
Scurrula chingii )、五蕊寄生(
Dendrophthoe pentandra )、卵叶槲寄生(
Viscum album var.
meridianum )等寄生植物(
朱华和闫丽春, 2009 ). ...
云南哀牢山原生林及次生林群落附生地衣物种多样性与分布
1
2007
... 哀牢山国家级自然保护区位于云南省中部哀牢山山脉中北段上部, 地处云贵高原、横断山和青藏高原南缘三大地理区域的结合部, 是生物多样性较丰富及植物区系地理成分荟萃之地, 该保护区内分布着我国目前保存最完整、面积最大的亚热带山地湿性常绿阔叶林(
朱华和闫丽春, 2009 ).本研究区位于哀牢山北段的徐家坝地区(23.35°-24.44° N, 100.54°-101.30° E), 该地区海拔2 000-2 750 m, 面积约5 100 hm
2 , 年降水量1 947 mm, 年蒸发量1 192 mm, 相对湿度85%, 年平均气温11.3 ℃ (
李苏等, 2007 ).该地区常绿阔叶林的土壤全C、全N与全P含量分别为98.65、6.53和1.17 mg·g
-1 , C:N、C:P、N:P为14.96、91.22和5.95 (
鲁志云等, 2017 ).徐家坝地区分布有椆树桑寄生(
Loranthus delavayi )、柳叶钝果寄生(
Taxillus delavayi )、卵叶梨果寄生(
Scurrula chingii )、五蕊寄生(
Dendrophthoe pentandra )、卵叶槲寄生(
Viscum album var.
meridianum )等寄生植物(
朱华和闫丽春, 2009 ). ...
植物生态化学计量学的全球格局综述
1
2016
... 生态化学计量学是主要研究生态系统能量平衡和多重化学元素平衡的科学(
Sterner & Elser, 2002 ;
曾德慧和陈广生, 2005 ), 其核心问题是揭示生物体元素组成差异对其生态功能的影响(
Elser et al ., 2000 ;
蒋利玲等, 2014 ).生态化学计量学从元素比率的角度将不同尺度、不同生物群系以及不同领域的生物学研究联系起来(
Sterner & Elser, 2002 ;
Michaels, 2003 ), 成为生态学研究的主要方法和研究热点(
曾冬萍等, 2013 ;
刘文亭等, 2015 ).这一领域研究最多的化学元素是碳(C)、氮(N)、磷(P), 作为3种最重要的生命元素, C、N、P是所有生命过程的基础, 在植物的生化功能方面紧密耦合(
李威, 2016 ).基于不同生命形式, 系统发育以及光合作用碳同化途径的植物叶片生态化学计量特征及其变异模式的研究已取得重大进展(
Han et al ., 2005 ,
2011 ), 但这些研究的主要对象为非寄生植物.有关研究表明, 非寄生植物生态化学计量特征与土壤养分状况相关(
Reich & Oleksyn, 2004 ;
赵一娉等, 2017 ), 但寄生植物没有根系直接接触土壤, 其生命活动所需要的养分和水分主要来自于寄主植物.目前, 有关寄生植物生态化学计量特征的研究鲜有报道. ...
植物生态化学计量学的全球格局综述
1
2016
... 生态化学计量学是主要研究生态系统能量平衡和多重化学元素平衡的科学(
Sterner & Elser, 2002 ;
曾德慧和陈广生, 2005 ), 其核心问题是揭示生物体元素组成差异对其生态功能的影响(
Elser et al ., 2000 ;
蒋利玲等, 2014 ).生态化学计量学从元素比率的角度将不同尺度、不同生物群系以及不同领域的生物学研究联系起来(
Sterner & Elser, 2002 ;
Michaels, 2003 ), 成为生态学研究的主要方法和研究热点(
曾冬萍等, 2013 ;
刘文亭等, 2015 ).这一领域研究最多的化学元素是碳(C)、氮(N)、磷(P), 作为3种最重要的生命元素, C、N、P是所有生命过程的基础, 在植物的生化功能方面紧密耦合(
李威, 2016 ).基于不同生命形式, 系统发育以及光合作用碳同化途径的植物叶片生态化学计量特征及其变异模式的研究已取得重大进展(
Han et al ., 2005 ,
2011 ), 但这些研究的主要对象为非寄生植物.有关研究表明, 非寄生植物生态化学计量特征与土壤养分状况相关(
Reich & Oleksyn, 2004 ;
赵一娉等, 2017 ), 但寄生植物没有根系直接接触土壤, 其生命活动所需要的养分和水分主要来自于寄主植物.目前, 有关寄生植物生态化学计量特征的研究鲜有报道. ...
Variation in leaf nitrogen and phosphorus stoichiometry in the nitrogen-fixing Chinese sea-buckthorn (Hippophae rhamnoides L. subsp. sinensis Rousi) across northern China
1
2014
... N和P在植物中为协同元素, 植物叶片N、P含量的正相关关系亦为高等陆生植物养分计量的普遍规律(
Sterner & Elser, 2002 ;
Wright et al ., 2005 ).植物N、P间的耦合关系很早就有文献记录(
Garten, 1976 ), 随后也得到许多研究的验证(
Li et al ., 2014 ;
Hidaka & Kitayama, 2015 ;
崔高阳等, 2015 ).对中国753种陆地植物叶片的研究(
Han et al ., 2005 )以及中国东部南北样带654种植物叶片的研究(
任书杰等, 2007 )表明, 不同生活型或不同系统发育植物的叶片N和P含量大部分也呈极显著相关关系.水生植物N、P含量间同样具有强线性关系, 这可能是大多数植物N、P含量的基本特征(
Duarte, 1992 ).
Hidaka和Kitayama (2015) 认为植物叶片N、P含量间的正相关关系可能部分受到土壤可利用性氮含量的影响, 而植物营养策略(N利用策略受到P利用策略的影响)以及植物细胞中的N和P生态化学计量特征(如蛋白质合成中N和P的生化关系)则可提供另一种解释, 但仍需要进一步分析以阐明相关性的生理背景.本研究中, N和P由寄主枝、吸器中的极显著正相关关系, 寄生枝中显著正相关关系, 到寄生叶中变为了相关关系不显著.
Garten (1976) 的研究中大型水生植物的N和P含量相关性也不显著, 这可能是大型水生植物的N含量范围较小, 或者是当环境P缺乏时对N的奢侈消费造成的.
Han等(2005) 和
任书杰等(2007) 的研究中, 裸子植物和蕨类植物的N、P含量相关性也不显著.本研究中寄生叶N、P间的不协同性可能与寄生植物不直接从土壤吸收养分、混合营养策略以及寄生植物叶片重吸收P但不重吸收N有关(
Suriyagoda et al ., 2017 ).受到N、P不耦合的影响, 寄生叶的N含量和C:P, P含量和C:N也不存在显著的相关关系. ...
不同寄主植物桑寄生总黄酮含量研究
1
2009
... 大多数寄生植物具有广寄性, 而不同的寄生植物其寄主种类有很大的差异(
Norton & Lange, 1999 ;
胡飞和孔垂华, 2004 ).本次调查记录中椆树桑寄生的寄主均为壳斗科植物, 柳叶钝果寄生的寄主较为广泛, 但未发现与椆树桑寄生具有相同寄主.不同的寄主物种对同一寄生植物的影响可能不同, 有研究发现同一桑寄生植物的挥发性成分组成及含量、总黄酮含量、毒性等会因其寄主物种不同而不同(
李永华等, 2009 ,
2012 ;
周汉华等, 2013 ), 但对寄生植物生态化学计量特征与寄主物种的关系进行探讨的研究较少.在本研究中, 寄生于不同寄主的同一寄生植物的同一器官C、N、P生态化学计量特征基本没有显著的不同, 而对同一寄生植物的不同寄主情况下不同器官之间的生态化学计量特征进行双因素方差分析, 也发现寄主物种的影响不显著.这可能与同一寄生植物的不同寄主枝条本身生态化学计量特征差异不显著有关. ...
不同寄主植物桑寄生总黄酮含量研究
1
2009
... 大多数寄生植物具有广寄性, 而不同的寄生植物其寄主种类有很大的差异(
Norton & Lange, 1999 ;
胡飞和孔垂华, 2004 ).本次调查记录中椆树桑寄生的寄主均为壳斗科植物, 柳叶钝果寄生的寄主较为广泛, 但未发现与椆树桑寄生具有相同寄主.不同的寄主物种对同一寄生植物的影响可能不同, 有研究发现同一桑寄生植物的挥发性成分组成及含量、总黄酮含量、毒性等会因其寄主物种不同而不同(
李永华等, 2009 ,
2012 ;
周汉华等, 2013 ), 但对寄生植物生态化学计量特征与寄主物种的关系进行探讨的研究较少.在本研究中, 寄生于不同寄主的同一寄生植物的同一器官C、N、P生态化学计量特征基本没有显著的不同, 而对同一寄生植物的不同寄主情况下不同器官之间的生态化学计量特征进行双因素方差分析, 也发现寄主物种的影响不显著.这可能与同一寄生植物的不同寄主枝条本身生态化学计量特征差异不显著有关. ...
寄主植物对桑寄生药材挥发性成分的影响研究
1
2012
... 大多数寄生植物具有广寄性, 而不同的寄生植物其寄主种类有很大的差异(
Norton & Lange, 1999 ;
胡飞和孔垂华, 2004 ).本次调查记录中椆树桑寄生的寄主均为壳斗科植物, 柳叶钝果寄生的寄主较为广泛, 但未发现与椆树桑寄生具有相同寄主.不同的寄主物种对同一寄生植物的影响可能不同, 有研究发现同一桑寄生植物的挥发性成分组成及含量、总黄酮含量、毒性等会因其寄主物种不同而不同(
李永华等, 2009 ,
2012 ;
周汉华等, 2013 ), 但对寄生植物生态化学计量特征与寄主物种的关系进行探讨的研究较少.在本研究中, 寄生于不同寄主的同一寄生植物的同一器官C、N、P生态化学计量特征基本没有显著的不同, 而对同一寄生植物的不同寄主情况下不同器官之间的生态化学计量特征进行双因素方差分析, 也发现寄主物种的影响不显著.这可能与同一寄生植物的不同寄主枝条本身生态化学计量特征差异不显著有关. ...
寄主植物对桑寄生药材挥发性成分的影响研究
1
2012
... 大多数寄生植物具有广寄性, 而不同的寄生植物其寄主种类有很大的差异(
Norton & Lange, 1999 ;
胡飞和孔垂华, 2004 ).本次调查记录中椆树桑寄生的寄主均为壳斗科植物, 柳叶钝果寄生的寄主较为广泛, 但未发现与椆树桑寄生具有相同寄主.不同的寄主物种对同一寄生植物的影响可能不同, 有研究发现同一桑寄生植物的挥发性成分组成及含量、总黄酮含量、毒性等会因其寄主物种不同而不同(
李永华等, 2009 ,
2012 ;
周汉华等, 2013 ), 但对寄生植物生态化学计量特征与寄主物种的关系进行探讨的研究较少.在本研究中, 寄生于不同寄主的同一寄生植物的同一器官C、N、P生态化学计量特征基本没有显著的不同, 而对同一寄生植物的不同寄主情况下不同器官之间的生态化学计量特征进行双因素方差分析, 也发现寄主物种的影响不显著.这可能与同一寄生植物的不同寄主枝条本身生态化学计量特征差异不显著有关. ...
有机碳肥对养分平衡的作用初探——试析植物营养中的碳短板
1
2014
... 植物体中C与N、P含量一般呈负相关关系是高等陆生植物C、N、P计量的普遍规律(
Sterner & Elser, 2002 ;
贺合亮等, 2017 ).在本研究中, 仅寄主枝符合此规律, 其C含量与N、P含量及C:N、C:P、N:P均存在极显著的相关关系.寄生植物吸器的C含量仅与N:P具有显著的相关关系, 枝条的C含量则与N、P含量及C:N、C:P、N:P均不存在极显著的相关关系.寄生植物叶片的C含量与N含量为正相关关系, 与P含量的相关性未达到显著, 也不同于此规律, 但该结果与荒漠植物叶片C、N、P生态化学计量特征具有一致性(
张珂等, 2014 ;
罗艳等, 2017 ).陆生植物的C主要来自大气中的CO
2 (
廖宗文等, 2014 ), 半寄生植物的C既可以来自自身光合作用, 也可以从寄主处获取(
Tě?itel et al ., 2015 ).规律不同说明半寄生植物在固C过程中对养分(N、P等)的利用策略有别于一般陆生植物. ...
有机碳肥对养分平衡的作用初探——试析植物营养中的碳短板
1
2014
... 植物体中C与N、P含量一般呈负相关关系是高等陆生植物C、N、P计量的普遍规律(
Sterner & Elser, 2002 ;
贺合亮等, 2017 ).在本研究中, 仅寄主枝符合此规律, 其C含量与N、P含量及C:N、C:P、N:P均存在极显著的相关关系.寄生植物吸器的C含量仅与N:P具有显著的相关关系, 枝条的C含量则与N、P含量及C:N、C:P、N:P均不存在极显著的相关关系.寄生植物叶片的C含量与N含量为正相关关系, 与P含量的相关性未达到显著, 也不同于此规律, 但该结果与荒漠植物叶片C、N、P生态化学计量特征具有一致性(
张珂等, 2014 ;
罗艳等, 2017 ).陆生植物的C主要来自大气中的CO
2 (
廖宗文等, 2014 ), 半寄生植物的C既可以来自自身光合作用, 也可以从寄主处获取(
Tě?itel et al ., 2015 ).规律不同说明半寄生植物在固C过程中对养分(N、P等)的利用策略有别于一般陆生植物. ...
中国草原生态化学计量学研究进展
1
2015
... 生态化学计量学是主要研究生态系统能量平衡和多重化学元素平衡的科学(
Sterner & Elser, 2002 ;
曾德慧和陈广生, 2005 ), 其核心问题是揭示生物体元素组成差异对其生态功能的影响(
Elser et al ., 2000 ;
蒋利玲等, 2014 ).生态化学计量学从元素比率的角度将不同尺度、不同生物群系以及不同领域的生物学研究联系起来(
Sterner & Elser, 2002 ;
Michaels, 2003 ), 成为生态学研究的主要方法和研究热点(
曾冬萍等, 2013 ;
刘文亭等, 2015 ).这一领域研究最多的化学元素是碳(C)、氮(N)、磷(P), 作为3种最重要的生命元素, C、N、P是所有生命过程的基础, 在植物的生化功能方面紧密耦合(
李威, 2016 ).基于不同生命形式, 系统发育以及光合作用碳同化途径的植物叶片生态化学计量特征及其变异模式的研究已取得重大进展(
Han et al ., 2005 ,
2011 ), 但这些研究的主要对象为非寄生植物.有关研究表明, 非寄生植物生态化学计量特征与土壤养分状况相关(
Reich & Oleksyn, 2004 ;
赵一娉等, 2017 ), 但寄生植物没有根系直接接触土壤, 其生命活动所需要的养分和水分主要来自于寄主植物.目前, 有关寄生植物生态化学计量特征的研究鲜有报道. ...
中国草原生态化学计量学研究进展
1
2015
... 生态化学计量学是主要研究生态系统能量平衡和多重化学元素平衡的科学(
Sterner & Elser, 2002 ;
曾德慧和陈广生, 2005 ), 其核心问题是揭示生物体元素组成差异对其生态功能的影响(
Elser et al ., 2000 ;
蒋利玲等, 2014 ).生态化学计量学从元素比率的角度将不同尺度、不同生物群系以及不同领域的生物学研究联系起来(
Sterner & Elser, 2002 ;
Michaels, 2003 ), 成为生态学研究的主要方法和研究热点(
曾冬萍等, 2013 ;
刘文亭等, 2015 ).这一领域研究最多的化学元素是碳(C)、氮(N)、磷(P), 作为3种最重要的生命元素, C、N、P是所有生命过程的基础, 在植物的生化功能方面紧密耦合(
李威, 2016 ).基于不同生命形式, 系统发育以及光合作用碳同化途径的植物叶片生态化学计量特征及其变异模式的研究已取得重大进展(
Han et al ., 2005 ,
2011 ), 但这些研究的主要对象为非寄生植物.有关研究表明, 非寄生植物生态化学计量特征与土壤养分状况相关(
Reich & Oleksyn, 2004 ;
赵一娉等, 2017 ), 但寄生植物没有根系直接接触土壤, 其生命活动所需要的养分和水分主要来自于寄主植物.目前, 有关寄生植物生态化学计量特征的研究鲜有报道. ...
哀牢山森林凋落物与腐殖质及土壤的生态化学计量特征
3
2017
... 哀牢山国家级自然保护区位于云南省中部哀牢山山脉中北段上部, 地处云贵高原、横断山和青藏高原南缘三大地理区域的结合部, 是生物多样性较丰富及植物区系地理成分荟萃之地, 该保护区内分布着我国目前保存最完整、面积最大的亚热带山地湿性常绿阔叶林(
朱华和闫丽春, 2009 ).本研究区位于哀牢山北段的徐家坝地区(23.35°-24.44° N, 100.54°-101.30° E), 该地区海拔2 000-2 750 m, 面积约5 100 hm
2 , 年降水量1 947 mm, 年蒸发量1 192 mm, 相对湿度85%, 年平均气温11.3 ℃ (
李苏等, 2007 ).该地区常绿阔叶林的土壤全C、全N与全P含量分别为98.65、6.53和1.17 mg·g
-1 , C:N、C:P、N:P为14.96、91.22和5.95 (
鲁志云等, 2017 ).徐家坝地区分布有椆树桑寄生(
Loranthus delavayi )、柳叶钝果寄生(
Taxillus delavayi )、卵叶梨果寄生(
Scurrula chingii )、五蕊寄生(
Dendrophthoe pentandra )、卵叶槲寄生(
Viscum album var.
meridianum )等寄生植物(
朱华和闫丽春, 2009 ). ...
... 寄生植物是一类重要而独特的植物类群.本研究中两种半寄生植物叶片的平均N含量为12.08 mg·g
-1 , 明显低于全球陆生植物的N含量20.09 mg·g
-1 (
Reich & Oleksyn, 2004 )、中国陆生植物的N含量18.6 mg·g
-1 (
Han et al ., 2005 ); 而P含量3.06 mg·g
-1 则明显地高于全球和中国陆生植物的P含量, 分别为1.77 mg·g
-1 (
Reich & Oleksyn, 2004 )、1.21 mg·g
-1 (
Han et al ., 2005 ).寄生叶的N含量较低可能与其光合作用较弱有关, 而较高的P含量可能受到研究区土壤P含量的间接影响, 哀牢山地区森林土壤的全P和有效P含量分别为1.17 g·kg
-1 和3.40 mg·kg
-1 (
鲁志云等, 2017 ), 中国土壤全P和有效P含量分别为0.60 g·kg
-1 和2.39 mg·kg
-1 (
汪涛等, 2008 ), 哀牢山地区森林土壤全P含量显著高于全国水平(
鲁志云等, 2017 ).此外, 哀牢山地区山地森林表层土壤(0-20 cm)的C:N、C:P、N:P值分别为14.9、84.9、5.7, 高于中国陆地(0-10 cm)土壤的C:N、C:P、N:P水平(分别为12.3、52.7、3.9)(
Tian et al ., 2010 ), 更接近于全球陆地(0-10 cm)和全球森林土壤水平(分别为12.3、72.0、5.9和12.4、81.9、6.6)(
Cleveland & Liptzin, 2007 ).已有的研究认为我国陆地植物生长普遍受P限制(
Han et al ., 2005 ;
任书杰等, 2007 ), 但哀牢山地区较高的土壤P含量暗示该地区可能并不缺P.关于哀牢山地区森林寄生植物、寄主的生态化学计量特征与土壤化学元素计量的关系将是以后研究的重要内容. ...
... ), 哀牢山地区森林土壤全P含量显著高于全国水平(
鲁志云等, 2017 ).此外, 哀牢山地区山地森林表层土壤(0-20 cm)的C:N、C:P、N:P值分别为14.9、84.9、5.7, 高于中国陆地(0-10 cm)土壤的C:N、C:P、N:P水平(分别为12.3、52.7、3.9)(
Tian et al ., 2010 ), 更接近于全球陆地(0-10 cm)和全球森林土壤水平(分别为12.3、72.0、5.9和12.4、81.9、6.6)(
Cleveland & Liptzin, 2007 ).已有的研究认为我国陆地植物生长普遍受P限制(
Han et al ., 2005 ;
任书杰等, 2007 ), 但哀牢山地区较高的土壤P含量暗示该地区可能并不缺P.关于哀牢山地区森林寄生植物、寄主的生态化学计量特征与土壤化学元素计量的关系将是以后研究的重要内容. ...
哀牢山森林凋落物与腐殖质及土壤的生态化学计量特征
3
2017
... 哀牢山国家级自然保护区位于云南省中部哀牢山山脉中北段上部, 地处云贵高原、横断山和青藏高原南缘三大地理区域的结合部, 是生物多样性较丰富及植物区系地理成分荟萃之地, 该保护区内分布着我国目前保存最完整、面积最大的亚热带山地湿性常绿阔叶林(
朱华和闫丽春, 2009 ).本研究区位于哀牢山北段的徐家坝地区(23.35°-24.44° N, 100.54°-101.30° E), 该地区海拔2 000-2 750 m, 面积约5 100 hm
2 , 年降水量1 947 mm, 年蒸发量1 192 mm, 相对湿度85%, 年平均气温11.3 ℃ (
李苏等, 2007 ).该地区常绿阔叶林的土壤全C、全N与全P含量分别为98.65、6.53和1.17 mg·g
-1 , C:N、C:P、N:P为14.96、91.22和5.95 (
鲁志云等, 2017 ).徐家坝地区分布有椆树桑寄生(
Loranthus delavayi )、柳叶钝果寄生(
Taxillus delavayi )、卵叶梨果寄生(
Scurrula chingii )、五蕊寄生(
Dendrophthoe pentandra )、卵叶槲寄生(
Viscum album var.
meridianum )等寄生植物(
朱华和闫丽春, 2009 ). ...
... 寄生植物是一类重要而独特的植物类群.本研究中两种半寄生植物叶片的平均N含量为12.08 mg·g
-1 , 明显低于全球陆生植物的N含量20.09 mg·g
-1 (
Reich & Oleksyn, 2004 )、中国陆生植物的N含量18.6 mg·g
-1 (
Han et al ., 2005 ); 而P含量3.06 mg·g
-1 则明显地高于全球和中国陆生植物的P含量, 分别为1.77 mg·g
-1 (
Reich & Oleksyn, 2004 )、1.21 mg·g
-1 (
Han et al ., 2005 ).寄生叶的N含量较低可能与其光合作用较弱有关, 而较高的P含量可能受到研究区土壤P含量的间接影响, 哀牢山地区森林土壤的全P和有效P含量分别为1.17 g·kg
-1 和3.40 mg·kg
-1 (
鲁志云等, 2017 ), 中国土壤全P和有效P含量分别为0.60 g·kg
-1 和2.39 mg·kg
-1 (
汪涛等, 2008 ), 哀牢山地区森林土壤全P含量显著高于全国水平(
鲁志云等, 2017 ).此外, 哀牢山地区山地森林表层土壤(0-20 cm)的C:N、C:P、N:P值分别为14.9、84.9、5.7, 高于中国陆地(0-10 cm)土壤的C:N、C:P、N:P水平(分别为12.3、52.7、3.9)(
Tian et al ., 2010 ), 更接近于全球陆地(0-10 cm)和全球森林土壤水平(分别为12.3、72.0、5.9和12.4、81.9、6.6)(
Cleveland & Liptzin, 2007 ).已有的研究认为我国陆地植物生长普遍受P限制(
Han et al ., 2005 ;
任书杰等, 2007 ), 但哀牢山地区较高的土壤P含量暗示该地区可能并不缺P.关于哀牢山地区森林寄生植物、寄主的生态化学计量特征与土壤化学元素计量的关系将是以后研究的重要内容. ...
... ), 哀牢山地区森林土壤全P含量显著高于全国水平(
鲁志云等, 2017 ).此外, 哀牢山地区山地森林表层土壤(0-20 cm)的C:N、C:P、N:P值分别为14.9、84.9、5.7, 高于中国陆地(0-10 cm)土壤的C:N、C:P、N:P水平(分别为12.3、52.7、3.9)(
Tian et al ., 2010 ), 更接近于全球陆地(0-10 cm)和全球森林土壤水平(分别为12.3、72.0、5.9和12.4、81.9、6.6)(
Cleveland & Liptzin, 2007 ).已有的研究认为我国陆地植物生长普遍受P限制(
Han et al ., 2005 ;
任书杰等, 2007 ), 但哀牢山地区较高的土壤P含量暗示该地区可能并不缺P.关于哀牢山地区森林寄生植物、寄主的生态化学计量特征与土壤化学元素计量的关系将是以后研究的重要内容. ...
塔里木河上游荒漠区4种灌木植物叶片与土壤生态化学计量特征
1
2017
... 植物体中C与N、P含量一般呈负相关关系是高等陆生植物C、N、P计量的普遍规律(
Sterner & Elser, 2002 ;
贺合亮等, 2017 ).在本研究中, 仅寄主枝符合此规律, 其C含量与N、P含量及C:N、C:P、N:P均存在极显著的相关关系.寄生植物吸器的C含量仅与N:P具有显著的相关关系, 枝条的C含量则与N、P含量及C:N、C:P、N:P均不存在极显著的相关关系.寄生植物叶片的C含量与N含量为正相关关系, 与P含量的相关性未达到显著, 也不同于此规律, 但该结果与荒漠植物叶片C、N、P生态化学计量特征具有一致性(
张珂等, 2014 ;
罗艳等, 2017 ).陆生植物的C主要来自大气中的CO
2 (
廖宗文等, 2014 ), 半寄生植物的C既可以来自自身光合作用, 也可以从寄主处获取(
Tě?itel et al ., 2015 ).规律不同说明半寄生植物在固C过程中对养分(N、P等)的利用策略有别于一般陆生植物. ...
塔里木河上游荒漠区4种灌木植物叶片与土壤生态化学计量特征
1
2017
... 植物体中C与N、P含量一般呈负相关关系是高等陆生植物C、N、P计量的普遍规律(
Sterner & Elser, 2002 ;
贺合亮等, 2017 ).在本研究中, 仅寄主枝符合此规律, 其C含量与N、P含量及C:N、C:P、N:P均存在极显著的相关关系.寄生植物吸器的C含量仅与N:P具有显著的相关关系, 枝条的C含量则与N、P含量及C:N、C:P、N:P均不存在极显著的相关关系.寄生植物叶片的C含量与N含量为正相关关系, 与P含量的相关性未达到显著, 也不同于此规律, 但该结果与荒漠植物叶片C、N、P生态化学计量特征具有一致性(
张珂等, 2014 ;
罗艳等, 2017 ).陆生植物的C主要来自大气中的CO
2 (
廖宗文等, 2014 ), 半寄生植物的C既可以来自自身光合作用, 也可以从寄主处获取(
Tě?itel et al ., 2015 ).规律不同说明半寄生植物在固C过程中对养分(N、P等)的利用策略有别于一般陆生植物. ...
The ratios of life
1
2003
... 生态化学计量学是主要研究生态系统能量平衡和多重化学元素平衡的科学(
Sterner & Elser, 2002 ;
曾德慧和陈广生, 2005 ), 其核心问题是揭示生物体元素组成差异对其生态功能的影响(
Elser et al ., 2000 ;
蒋利玲等, 2014 ).生态化学计量学从元素比率的角度将不同尺度、不同生物群系以及不同领域的生物学研究联系起来(
Sterner & Elser, 2002 ;
Michaels, 2003 ), 成为生态学研究的主要方法和研究热点(
曾冬萍等, 2013 ;
刘文亭等, 2015 ).这一领域研究最多的化学元素是碳(C)、氮(N)、磷(P), 作为3种最重要的生命元素, C、N、P是所有生命过程的基础, 在植物的生化功能方面紧密耦合(
李威, 2016 ).基于不同生命形式, 系统发育以及光合作用碳同化途径的植物叶片生态化学计量特征及其变异模式的研究已取得重大进展(
Han et al ., 2005 ,
2011 ), 但这些研究的主要对象为非寄生植物.有关研究表明, 非寄生植物生态化学计量特征与土壤养分状况相关(
Reich & Oleksyn, 2004 ;
赵一娉等, 2017 ), 但寄生植物没有根系直接接触土壤, 其生命活动所需要的养分和水分主要来自于寄主植物.目前, 有关寄生植物生态化学计量特征的研究鲜有报道. ...
Internal and external regulation of plant organ stoichiometry
1
2015
... 寄生植物不同的营养素富集程度也不同, 被认为是吸器选择性吸收离子的结果, 若摄取是通过蒸腾而被动吸收, 那么营养元素的比值应保持相同(
Gebauer et al ., 2018 ).本研究中, 椆树桑寄生吸器的C:N、C:P及N:P均与其寄主枝条存在显著差异, 柳叶钝果寄生吸器的C:N和C:P也与其寄主枝条存在显著差异, 这在一定程度上说明C、N和P元素并非简单的通过蒸腾作用而被转运至寄生植物体内, 但也不能排除离子通过被动运输进入吸器后, 由于吸器具有合成物质的功能而导致元素比值发生改变(
黄新亚等, 2011 ).植物不同器官C、N、P含量的差异不仅受植物基本生理过程需求的影响, 也受相应器官组织结构和功能分化的影响(
Minden & Kleyer, 2015 ).叶片是植物的同化器官与重要的养分储存器官, 因而寄生植物叶片的N和P含量高于寄主枝、吸器与寄生枝(
Niklas & Cobb, 2005 ;
贺合亮等, 2017 ).植物叶片中C、N、P含量的变异系数普遍存在C < N < P的规律(
Koerselman & Meuleman, 1996 ;
Güsewell & Koerselman, 2002 ;
姜沛沛等, 2017 ).本研究中两种半寄生植物的寄主枝、吸器、寄生枝与寄生叶的C、N、P含量皆符合这个规律, 这是因为C作为构成植物骨架的基本结构物质, 且为多种植物生理活动提供能源, 而N含量可被植物控制在较窄的范围内, 故C在植物体内含量高且变异小, N含量在各器官中的变异性小于P含量(
Vitousek, 1998 ;
贺合亮等, 2017 ).本研究中两种寄生植物C:N的变异性小于C:P,是因为植物光合作用中C的同化与营养元素的吸收并非同一途径, 而C含量高且变异小, C:N和C:P的变异主要受到N和P含量变化的影响(
Hedin, 2004 ;
曾昭霞等, 2015 ). ...
Radiocarbon suggests the hemiparasitic annual Melampyrum lineare Desr. may acquire carbon from stressed hosts
2
2017
... 半寄生植物是寄生植物的主要类群, 其叶片具有叶绿体, 在生命周期的至少一个阶段可进行光合作用, 仅从寄主体内获取部分水和溶解的矿物质(
Nickrent et al ., 1998 ).在自然界中, 半寄生植物对植物群落结构和多样性具有显著影响(
Phoenix & Press, 2010 ;
Press & Phoenix, 2010 ;
Demey et al ., 2014 ), 而其营养丰富、可快速分解和释放养分的凋落物可能会提高森林土壤养分可利用率(
Demey et al ., 2015 ), 影响生态系统水平凋落物分解和养分循环(
Quested et al ., 2010 ).虽然半寄生植物的丰度和生物量相对较小, 但它们对生物地球化学循环、植物种群结构和多样性的影响是很明确的(
Nave et al ., 2017 ), 因而被描述为影响群落动态的生态系统工程师, 以及影响养分循环的关键物种(
Watson, 2010 ;
Hatcher et al ., 2012 ). ...
... 半寄生植物与寄主的复杂关系包括寄生和竞争(
Lep? & Tě?itel, 2015 ).作为混合营养策略的代表, 半寄生植物既能从自身的光合作用中获取碳同化物, 也能通过吸器从寄主体内获取碳水化合物(
Nave et al ., 2017 ;
Selosse et al ., 2017 ).半寄生植物自身的光合作用在同化寄生矿物资源, 并将矿物资源转换化成自身生长和适合度中起着至关重要的作用, 而将木质部所获得的资源转化为生长和适应的关键过程, 可能是其进化稳定性的基础(
Tě?itel et al ., 2015 ).尽管半寄生植物与寄主关系的研究开展较早, 不少****对半寄生植物与寄主之间的水分利用、光合作用以及矿物质营养等方面的关系进行了较为深入的研究(
Tennakoon et al ., 2011 ;
Chen et al ., 2013 ;
Suriyagoda et al ., 2017 ;
Gebauer et al ., 2018 ), 但营养物质从寄主到半寄生植物的转移机制仍未完全研究清楚(
Gebauer et al ., 2012 ,
2018 ).有关研究表明, 半寄生植物
Rhinanthus alectorolophus 中5%-40%的C来自寄主, 而寄主体内的N亦可被半寄生植物获取(
Tě?itel et al ., 2015 ;
Světlíková et al ., 2016 ).寄生植物的吸器同时连接寄主枝条与寄生枝条, 研究寄主枝条-吸器-寄生枝条-寄生叶片这一连续体的生态化学计量特征变化将有助于进一步理解寄生植物与寄主间的养分关系. ...
Molecular phylogenetic and evolutionary studies of parasitic plants. Molecular Systematics of Plants II
1
1998
... 半寄生植物是寄生植物的主要类群, 其叶片具有叶绿体, 在生命周期的至少一个阶段可进行光合作用, 仅从寄主体内获取部分水和溶解的矿物质(
Nickrent et al ., 1998 ).在自然界中, 半寄生植物对植物群落结构和多样性具有显著影响(
Phoenix & Press, 2010 ;
Press & Phoenix, 2010 ;
Demey et al ., 2014 ), 而其营养丰富、可快速分解和释放养分的凋落物可能会提高森林土壤养分可利用率(
Demey et al ., 2015 ), 影响生态系统水平凋落物分解和养分循环(
Quested et al ., 2010 ).虽然半寄生植物的丰度和生物量相对较小, 但它们对生物地球化学循环、植物种群结构和多样性的影响是很明确的(
Nave et al ., 2017 ), 因而被描述为影响群落动态的生态系统工程师, 以及影响养分循环的关键物种(
Watson, 2010 ;
Hatcher et al ., 2012 ). ...
Plant allometry, leaf nitrogen and phosphorus stoichiometry, and interspecific trends in annual growth rates
1
2006
... N、P在植物叶片中的关系可以通过幂函数
N =
βPα 进行量化(
Wright et al ., 2004 ;
Reich et al ., 2010 ;
Yan et al ., 2018 ).
Wright等(2004) 基于全球范围内广泛的叶片N和P浓度数据, 认为N对P的幂指数约为2/3,
Niklas (2006) 的文章中该指数为0.73, 接近3/4而不是2/3,
Reich等(2010) 收集了更为全面的叶片N、P浓度数据, 提出了叶片N、P浓度计量的2/3幂法则(
N =
αP 2/3 ).目前已有研究表明, 植物叶片N、P计量关系并非恒定, 而是受到植物功能群、纬度区、生态区、施肥类型与水平等多种因素影响(
Tian et al ., 2018 ;
Yan et al ., 2018 ).在本研究中, 寄主枝、吸器与寄生枝的N、P计量关系的回归斜率均小于3/4与2/3, 尽管叶片N、P的计量关系有一些趋势但没有统计学意义.以往的研究主要基于植物叶片的数据, 结果是否适用于不同器官有待验证. ...
N, P, and C stoichiometry of Eranthis hyemalis (Ranunculaceae) and the allometry of plant growth
1
2005
... 寄生植物不同的营养素富集程度也不同, 被认为是吸器选择性吸收离子的结果, 若摄取是通过蒸腾而被动吸收, 那么营养元素的比值应保持相同(
Gebauer et al ., 2018 ).本研究中, 椆树桑寄生吸器的C:N、C:P及N:P均与其寄主枝条存在显著差异, 柳叶钝果寄生吸器的C:N和C:P也与其寄主枝条存在显著差异, 这在一定程度上说明C、N和P元素并非简单的通过蒸腾作用而被转运至寄生植物体内, 但也不能排除离子通过被动运输进入吸器后, 由于吸器具有合成物质的功能而导致元素比值发生改变(
黄新亚等, 2011 ).植物不同器官C、N、P含量的差异不仅受植物基本生理过程需求的影响, 也受相应器官组织结构和功能分化的影响(
Minden & Kleyer, 2015 ).叶片是植物的同化器官与重要的养分储存器官, 因而寄生植物叶片的N和P含量高于寄主枝、吸器与寄生枝(
Niklas & Cobb, 2005 ;
贺合亮等, 2017 ).植物叶片中C、N、P含量的变异系数普遍存在C < N < P的规律(
Koerselman & Meuleman, 1996 ;
Güsewell & Koerselman, 2002 ;
姜沛沛等, 2017 ).本研究中两种半寄生植物的寄主枝、吸器、寄生枝与寄生叶的C、N、P含量皆符合这个规律, 这是因为C作为构成植物骨架的基本结构物质, 且为多种植物生理活动提供能源, 而N含量可被植物控制在较窄的范围内, 故C在植物体内含量高且变异小, N含量在各器官中的变异性小于P含量(
Vitousek, 1998 ;
贺合亮等, 2017 ).本研究中两种寄生植物C:N的变异性小于C:P,是因为植物光合作用中C的同化与营养元素的吸收并非同一途径, 而C含量高且变异小, C:N和C:P的变异主要受到N和P含量变化的影响(
Hedin, 2004 ;
曾昭霞等, 2015 ). ...
Host specificity in Parasitic mistletoes (Loranthaceae) in New Zealand
1
1999
... 大多数寄生植物具有广寄性, 而不同的寄生植物其寄主种类有很大的差异(
Norton & Lange, 1999 ;
胡飞和孔垂华, 2004 ).本次调查记录中椆树桑寄生的寄主均为壳斗科植物, 柳叶钝果寄生的寄主较为广泛, 但未发现与椆树桑寄生具有相同寄主.不同的寄主物种对同一寄生植物的影响可能不同, 有研究发现同一桑寄生植物的挥发性成分组成及含量、总黄酮含量、毒性等会因其寄主物种不同而不同(
李永华等, 2009 ,
2012 ;
周汉华等, 2013 ), 但对寄生植物生态化学计量特征与寄主物种的关系进行探讨的研究较少.在本研究中, 寄生于不同寄主的同一寄生植物的同一器官C、N、P生态化学计量特征基本没有显著的不同, 而对同一寄生植物的不同寄主情况下不同器官之间的生态化学计量特征进行双因素方差分析, 也发现寄主物种的影响不显著.这可能与同一寄生植物的不同寄主枝条本身生态化学计量特征差异不显著有关. ...
Linking physiological traits to impacts on community structure and function: The role of root hemiparasitic Orobanchaceae (ex-Scrophulariaceae)
1
2010
... 半寄生植物是寄生植物的主要类群, 其叶片具有叶绿体, 在生命周期的至少一个阶段可进行光合作用, 仅从寄主体内获取部分水和溶解的矿物质(
Nickrent et al ., 1998 ).在自然界中, 半寄生植物对植物群落结构和多样性具有显著影响(
Phoenix & Press, 2010 ;
Press & Phoenix, 2010 ;
Demey et al ., 2014 ), 而其营养丰富、可快速分解和释放养分的凋落物可能会提高森林土壤养分可利用率(
Demey et al ., 2015 ), 影响生态系统水平凋落物分解和养分循环(
Quested et al ., 2010 ).虽然半寄生植物的丰度和生物量相对较小, 但它们对生物地球化学循环、植物种群结构和多样性的影响是很明确的(
Nave et al ., 2017 ), 因而被描述为影响群落动态的生态系统工程师, 以及影响养分循环的关键物种(
Watson, 2010 ;
Hatcher et al ., 2012 ). ...
Impacts of parasitic plants on natural communities
1
2010
... 半寄生植物是寄生植物的主要类群, 其叶片具有叶绿体, 在生命周期的至少一个阶段可进行光合作用, 仅从寄主体内获取部分水和溶解的矿物质(
Nickrent et al ., 1998 ).在自然界中, 半寄生植物对植物群落结构和多样性具有显著影响(
Phoenix & Press, 2010 ;
Press & Phoenix, 2010 ;
Demey et al ., 2014 ), 而其营养丰富、可快速分解和释放养分的凋落物可能会提高森林土壤养分可利用率(
Demey et al ., 2015 ), 影响生态系统水平凋落物分解和养分循环(
Quested et al ., 2010 ).虽然半寄生植物的丰度和生物量相对较小, 但它们对生物地球化学循环、植物种群结构和多样性的影响是很明确的(
Nave et al ., 2017 ), 因而被描述为影响群落动态的生态系统工程师, 以及影响养分循环的关键物种(
Watson, 2010 ;
Hatcher et al ., 2012 ). ...
1
1998
... 哀牢山国家级自然保护区分布着我国目前保存最完整、面积最大的亚热带山地湿性常绿阔叶林, 林内附生植物丰富, 也有相当数量的寄生植物(
游承侠, 1983 ;
Young & Herwitz, 1995 ;
朱华和闫丽春, 2009 ), 不少****对该地区森林群落类型及结构、树种、附生和寄生植物多样性、生物量等方面进行了研究(
邱学忠和谢寿昌, 1998 ), 但对该区域不同类群植物包括寄生植物的生态化学计量特征及养分利用的了解甚少.本研究以哀牢山山地湿性常绿阔叶林中常见的半寄生植物为对象, 对寄生植物的寄主枝条-吸器-寄生枝条-寄生叶片生态化学计量特征及其动态进行研究, 以期进一步理解半寄生植物的养分利用策略, 为山地森林资源保护与管理提供科学依据. ...
1
1998
... 哀牢山国家级自然保护区分布着我国目前保存最完整、面积最大的亚热带山地湿性常绿阔叶林, 林内附生植物丰富, 也有相当数量的寄生植物(
游承侠, 1983 ;
Young & Herwitz, 1995 ;
朱华和闫丽春, 2009 ), 不少****对该地区森林群落类型及结构、树种、附生和寄生植物多样性、生物量等方面进行了研究(
邱学忠和谢寿昌, 1998 ), 但对该区域不同类群植物包括寄生植物的生态化学计量特征及养分利用的了解甚少.本研究以哀牢山山地湿性常绿阔叶林中常见的半寄生植物为对象, 对寄生植物的寄主枝条-吸器-寄生枝条-寄生叶片生态化学计量特征及其动态进行研究, 以期进一步理解半寄生植物的养分利用策略, 为山地森林资源保护与管理提供科学依据. ...
The impact of hemiparasitic plant litter on decomposition: Direct, seasonal and litter mixing effects
1
2010
... 半寄生植物是寄生植物的主要类群, 其叶片具有叶绿体, 在生命周期的至少一个阶段可进行光合作用, 仅从寄主体内获取部分水和溶解的矿物质(
Nickrent et al ., 1998 ).在自然界中, 半寄生植物对植物群落结构和多样性具有显著影响(
Phoenix & Press, 2010 ;
Press & Phoenix, 2010 ;
Demey et al ., 2014 ), 而其营养丰富、可快速分解和释放养分的凋落物可能会提高森林土壤养分可利用率(
Demey et al ., 2015 ), 影响生态系统水平凋落物分解和养分循环(
Quested et al ., 2010 ).虽然半寄生植物的丰度和生物量相对较小, 但它们对生物地球化学循环、植物种群结构和多样性的影响是很明确的(
Nave et al ., 2017 ), 因而被描述为影响群落动态的生态系统工程师, 以及影响养分循环的关键物种(
Watson, 2010 ;
Hatcher et al ., 2012 ). ...
Global patterns of plant leaf N and P in relation to temperature and latitude
3
2004
... 生态化学计量学是主要研究生态系统能量平衡和多重化学元素平衡的科学(
Sterner & Elser, 2002 ;
曾德慧和陈广生, 2005 ), 其核心问题是揭示生物体元素组成差异对其生态功能的影响(
Elser et al ., 2000 ;
蒋利玲等, 2014 ).生态化学计量学从元素比率的角度将不同尺度、不同生物群系以及不同领域的生物学研究联系起来(
Sterner & Elser, 2002 ;
Michaels, 2003 ), 成为生态学研究的主要方法和研究热点(
曾冬萍等, 2013 ;
刘文亭等, 2015 ).这一领域研究最多的化学元素是碳(C)、氮(N)、磷(P), 作为3种最重要的生命元素, C、N、P是所有生命过程的基础, 在植物的生化功能方面紧密耦合(
李威, 2016 ).基于不同生命形式, 系统发育以及光合作用碳同化途径的植物叶片生态化学计量特征及其变异模式的研究已取得重大进展(
Han et al ., 2005 ,
2011 ), 但这些研究的主要对象为非寄生植物.有关研究表明, 非寄生植物生态化学计量特征与土壤养分状况相关(
Reich & Oleksyn, 2004 ;
赵一娉等, 2017 ), 但寄生植物没有根系直接接触土壤, 其生命活动所需要的养分和水分主要来自于寄主植物.目前, 有关寄生植物生态化学计量特征的研究鲜有报道. ...
... 寄生植物是一类重要而独特的植物类群.本研究中两种半寄生植物叶片的平均N含量为12.08 mg·g
-1 , 明显低于全球陆生植物的N含量20.09 mg·g
-1 (
Reich & Oleksyn, 2004 )、中国陆生植物的N含量18.6 mg·g
-1 (
Han et al ., 2005 ); 而P含量3.06 mg·g
-1 则明显地高于全球和中国陆生植物的P含量, 分别为1.77 mg·g
-1 (
Reich & Oleksyn, 2004 )、1.21 mg·g
-1 (
Han et al ., 2005 ).寄生叶的N含量较低可能与其光合作用较弱有关, 而较高的P含量可能受到研究区土壤P含量的间接影响, 哀牢山地区森林土壤的全P和有效P含量分别为1.17 g·kg
-1 和3.40 mg·kg
-1 (
鲁志云等, 2017 ), 中国土壤全P和有效P含量分别为0.60 g·kg
-1 和2.39 mg·kg
-1 (
汪涛等, 2008 ), 哀牢山地区森林土壤全P含量显著高于全国水平(
鲁志云等, 2017 ).此外, 哀牢山地区山地森林表层土壤(0-20 cm)的C:N、C:P、N:P值分别为14.9、84.9、5.7, 高于中国陆地(0-10 cm)土壤的C:N、C:P、N:P水平(分别为12.3、52.7、3.9)(
Tian et al ., 2010 ), 更接近于全球陆地(0-10 cm)和全球森林土壤水平(分别为12.3、72.0、5.9和12.4、81.9、6.6)(
Cleveland & Liptzin, 2007 ).已有的研究认为我国陆地植物生长普遍受P限制(
Han et al ., 2005 ;
任书杰等, 2007 ), 但哀牢山地区较高的土壤P含量暗示该地区可能并不缺P.关于哀牢山地区森林寄生植物、寄主的生态化学计量特征与土壤化学元素计量的关系将是以后研究的重要内容. ...
... (
Reich & Oleksyn, 2004 )、1.21 mg·g
-1 (
Han et al ., 2005 ).寄生叶的N含量较低可能与其光合作用较弱有关, 而较高的P含量可能受到研究区土壤P含量的间接影响, 哀牢山地区森林土壤的全P和有效P含量分别为1.17 g·kg
-1 和3.40 mg·kg
-1 (
鲁志云等, 2017 ), 中国土壤全P和有效P含量分别为0.60 g·kg
-1 和2.39 mg·kg
-1 (
汪涛等, 2008 ), 哀牢山地区森林土壤全P含量显著高于全国水平(
鲁志云等, 2017 ).此外, 哀牢山地区山地森林表层土壤(0-20 cm)的C:N、C:P、N:P值分别为14.9、84.9、5.7, 高于中国陆地(0-10 cm)土壤的C:N、C:P、N:P水平(分别为12.3、52.7、3.9)(
Tian et al ., 2010 ), 更接近于全球陆地(0-10 cm)和全球森林土壤水平(分别为12.3、72.0、5.9和12.4、81.9、6.6)(
Cleveland & Liptzin, 2007 ).已有的研究认为我国陆地植物生长普遍受P限制(
Han et al ., 2005 ;
任书杰等, 2007 ), 但哀牢山地区较高的土壤P含量暗示该地区可能并不缺P.关于哀牢山地区森林寄生植物、寄主的生态化学计量特征与土壤化学元素计量的关系将是以后研究的重要内容. ...
Evidence of a General 2/3-power law of scaling leaf nitrogen to phosphorus among major plant groups and biomes
2
2010
... N、P在植物叶片中的关系可以通过幂函数
N =
βPα 进行量化(
Wright et al ., 2004 ;
Reich et al ., 2010 ;
Yan et al ., 2018 ).
Wright等(2004) 基于全球范围内广泛的叶片N和P浓度数据, 认为N对P的幂指数约为2/3,
Niklas (2006) 的文章中该指数为0.73, 接近3/4而不是2/3,
Reich等(2010) 收集了更为全面的叶片N、P浓度数据, 提出了叶片N、P浓度计量的2/3幂法则(
N =
αP 2/3 ).目前已有研究表明, 植物叶片N、P计量关系并非恒定, 而是受到植物功能群、纬度区、生态区、施肥类型与水平等多种因素影响(
Tian et al ., 2018 ;
Yan et al ., 2018 ).在本研究中, 寄主枝、吸器与寄生枝的N、P计量关系的回归斜率均小于3/4与2/3, 尽管叶片N、P的计量关系有一些趋势但没有统计学意义.以往的研究主要基于植物叶片的数据, 结果是否适用于不同器官有待验证. ...
... 的文章中该指数为0.73, 接近3/4而不是2/3,
Reich等(2010) 收集了更为全面的叶片N、P浓度数据, 提出了叶片N、P浓度计量的2/3幂法则(
N =
αP 2/3 ).目前已有研究表明, 植物叶片N、P计量关系并非恒定, 而是受到植物功能群、纬度区、生态区、施肥类型与水平等多种因素影响(
Tian et al ., 2018 ;
Yan et al ., 2018 ).在本研究中, 寄主枝、吸器与寄生枝的N、P计量关系的回归斜率均小于3/4与2/3, 尽管叶片N、P的计量关系有一些趋势但没有统计学意义.以往的研究主要基于植物叶片的数据, 结果是否适用于不同器官有待验证. ...
中国东部南北样带654种植物叶片氮和磷的化学计量学特征研究
3
2007
... N和P在植物中为协同元素, 植物叶片N、P含量的正相关关系亦为高等陆生植物养分计量的普遍规律(
Sterner & Elser, 2002 ;
Wright et al ., 2005 ).植物N、P间的耦合关系很早就有文献记录(
Garten, 1976 ), 随后也得到许多研究的验证(
Li et al ., 2014 ;
Hidaka & Kitayama, 2015 ;
崔高阳等, 2015 ).对中国753种陆地植物叶片的研究(
Han et al ., 2005 )以及中国东部南北样带654种植物叶片的研究(
任书杰等, 2007 )表明, 不同生活型或不同系统发育植物的叶片N和P含量大部分也呈极显著相关关系.水生植物N、P含量间同样具有强线性关系, 这可能是大多数植物N、P含量的基本特征(
Duarte, 1992 ).
Hidaka和Kitayama (2015) 认为植物叶片N、P含量间的正相关关系可能部分受到土壤可利用性氮含量的影响, 而植物营养策略(N利用策略受到P利用策略的影响)以及植物细胞中的N和P生态化学计量特征(如蛋白质合成中N和P的生化关系)则可提供另一种解释, 但仍需要进一步分析以阐明相关性的生理背景.本研究中, N和P由寄主枝、吸器中的极显著正相关关系, 寄生枝中显著正相关关系, 到寄生叶中变为了相关关系不显著.
Garten (1976) 的研究中大型水生植物的N和P含量相关性也不显著, 这可能是大型水生植物的N含量范围较小, 或者是当环境P缺乏时对N的奢侈消费造成的.
Han等(2005) 和
任书杰等(2007) 的研究中, 裸子植物和蕨类植物的N、P含量相关性也不显著.本研究中寄生叶N、P间的不协同性可能与寄生植物不直接从土壤吸收养分、混合营养策略以及寄生植物叶片重吸收P但不重吸收N有关(
Suriyagoda et al ., 2017 ).受到N、P不耦合的影响, 寄生叶的N含量和C:P, P含量和C:N也不存在显著的相关关系. ...
... 和
任书杰等(2007) 的研究中, 裸子植物和蕨类植物的N、P含量相关性也不显著.本研究中寄生叶N、P间的不协同性可能与寄生植物不直接从土壤吸收养分、混合营养策略以及寄生植物叶片重吸收P但不重吸收N有关(
Suriyagoda et al ., 2017 ).受到N、P不耦合的影响, 寄生叶的N含量和C:P, P含量和C:N也不存在显著的相关关系. ...
... 寄生植物是一类重要而独特的植物类群.本研究中两种半寄生植物叶片的平均N含量为12.08 mg·g
-1 , 明显低于全球陆生植物的N含量20.09 mg·g
-1 (
Reich & Oleksyn, 2004 )、中国陆生植物的N含量18.6 mg·g
-1 (
Han et al ., 2005 ); 而P含量3.06 mg·g
-1 则明显地高于全球和中国陆生植物的P含量, 分别为1.77 mg·g
-1 (
Reich & Oleksyn, 2004 )、1.21 mg·g
-1 (
Han et al ., 2005 ).寄生叶的N含量较低可能与其光合作用较弱有关, 而较高的P含量可能受到研究区土壤P含量的间接影响, 哀牢山地区森林土壤的全P和有效P含量分别为1.17 g·kg
-1 和3.40 mg·kg
-1 (
鲁志云等, 2017 ), 中国土壤全P和有效P含量分别为0.60 g·kg
-1 和2.39 mg·kg
-1 (
汪涛等, 2008 ), 哀牢山地区森林土壤全P含量显著高于全国水平(
鲁志云等, 2017 ).此外, 哀牢山地区山地森林表层土壤(0-20 cm)的C:N、C:P、N:P值分别为14.9、84.9、5.7, 高于中国陆地(0-10 cm)土壤的C:N、C:P、N:P水平(分别为12.3、52.7、3.9)(
Tian et al ., 2010 ), 更接近于全球陆地(0-10 cm)和全球森林土壤水平(分别为12.3、72.0、5.9和12.4、81.9、6.6)(
Cleveland & Liptzin, 2007 ).已有的研究认为我国陆地植物生长普遍受P限制(
Han et al ., 2005 ;
任书杰等, 2007 ), 但哀牢山地区较高的土壤P含量暗示该地区可能并不缺P.关于哀牢山地区森林寄生植物、寄主的生态化学计量特征与土壤化学元素计量的关系将是以后研究的重要内容. ...
中国东部南北样带654种植物叶片氮和磷的化学计量学特征研究
3
2007
... N和P在植物中为协同元素, 植物叶片N、P含量的正相关关系亦为高等陆生植物养分计量的普遍规律(
Sterner & Elser, 2002 ;
Wright et al ., 2005 ).植物N、P间的耦合关系很早就有文献记录(
Garten, 1976 ), 随后也得到许多研究的验证(
Li et al ., 2014 ;
Hidaka & Kitayama, 2015 ;
崔高阳等, 2015 ).对中国753种陆地植物叶片的研究(
Han et al ., 2005 )以及中国东部南北样带654种植物叶片的研究(
任书杰等, 2007 )表明, 不同生活型或不同系统发育植物的叶片N和P含量大部分也呈极显著相关关系.水生植物N、P含量间同样具有强线性关系, 这可能是大多数植物N、P含量的基本特征(
Duarte, 1992 ).
Hidaka和Kitayama (2015) 认为植物叶片N、P含量间的正相关关系可能部分受到土壤可利用性氮含量的影响, 而植物营养策略(N利用策略受到P利用策略的影响)以及植物细胞中的N和P生态化学计量特征(如蛋白质合成中N和P的生化关系)则可提供另一种解释, 但仍需要进一步分析以阐明相关性的生理背景.本研究中, N和P由寄主枝、吸器中的极显著正相关关系, 寄生枝中显著正相关关系, 到寄生叶中变为了相关关系不显著.
Garten (1976) 的研究中大型水生植物的N和P含量相关性也不显著, 这可能是大型水生植物的N含量范围较小, 或者是当环境P缺乏时对N的奢侈消费造成的.
Han等(2005) 和
任书杰等(2007) 的研究中, 裸子植物和蕨类植物的N、P含量相关性也不显著.本研究中寄生叶N、P间的不协同性可能与寄生植物不直接从土壤吸收养分、混合营养策略以及寄生植物叶片重吸收P但不重吸收N有关(
Suriyagoda et al ., 2017 ).受到N、P不耦合的影响, 寄生叶的N含量和C:P, P含量和C:N也不存在显著的相关关系. ...
... 和
任书杰等(2007) 的研究中, 裸子植物和蕨类植物的N、P含量相关性也不显著.本研究中寄生叶N、P间的不协同性可能与寄生植物不直接从土壤吸收养分、混合营养策略以及寄生植物叶片重吸收P但不重吸收N有关(
Suriyagoda et al ., 2017 ).受到N、P不耦合的影响, 寄生叶的N含量和C:P, P含量和C:N也不存在显著的相关关系. ...
... 寄生植物是一类重要而独特的植物类群.本研究中两种半寄生植物叶片的平均N含量为12.08 mg·g
-1 , 明显低于全球陆生植物的N含量20.09 mg·g
-1 (
Reich & Oleksyn, 2004 )、中国陆生植物的N含量18.6 mg·g
-1 (
Han et al ., 2005 ); 而P含量3.06 mg·g
-1 则明显地高于全球和中国陆生植物的P含量, 分别为1.77 mg·g
-1 (
Reich & Oleksyn, 2004 )、1.21 mg·g
-1 (
Han et al ., 2005 ).寄生叶的N含量较低可能与其光合作用较弱有关, 而较高的P含量可能受到研究区土壤P含量的间接影响, 哀牢山地区森林土壤的全P和有效P含量分别为1.17 g·kg
-1 和3.40 mg·kg
-1 (
鲁志云等, 2017 ), 中国土壤全P和有效P含量分别为0.60 g·kg
-1 和2.39 mg·kg
-1 (
汪涛等, 2008 ), 哀牢山地区森林土壤全P含量显著高于全国水平(
鲁志云等, 2017 ).此外, 哀牢山地区山地森林表层土壤(0-20 cm)的C:N、C:P、N:P值分别为14.9、84.9、5.7, 高于中国陆地(0-10 cm)土壤的C:N、C:P、N:P水平(分别为12.3、52.7、3.9)(
Tian et al ., 2010 ), 更接近于全球陆地(0-10 cm)和全球森林土壤水平(分别为12.3、72.0、5.9和12.4、81.9、6.6)(
Cleveland & Liptzin, 2007 ).已有的研究认为我国陆地植物生长普遍受P限制(
Han et al ., 2005 ;
任书杰等, 2007 ), 但哀牢山地区较高的土壤P含量暗示该地区可能并不缺P.关于哀牢山地区森林寄生植物、寄主的生态化学计量特征与土壤化学元素计量的关系将是以后研究的重要内容. ...
Mixotrophy everywhere on land and in water: The grand écart hypothesis
1
2017
... 半寄生植物与寄主的复杂关系包括寄生和竞争(
Lep? & Tě?itel, 2015 ).作为混合营养策略的代表, 半寄生植物既能从自身的光合作用中获取碳同化物, 也能通过吸器从寄主体内获取碳水化合物(
Nave et al ., 2017 ;
Selosse et al ., 2017 ).半寄生植物自身的光合作用在同化寄生矿物资源, 并将矿物资源转换化成自身生长和适合度中起着至关重要的作用, 而将木质部所获得的资源转化为生长和适应的关键过程, 可能是其进化稳定性的基础(
Tě?itel et al ., 2015 ).尽管半寄生植物与寄主关系的研究开展较早, 不少****对半寄生植物与寄主之间的水分利用、光合作用以及矿物质营养等方面的关系进行了较为深入的研究(
Tennakoon et al ., 2011 ;
Chen et al ., 2013 ;
Suriyagoda et al ., 2017 ;
Gebauer et al ., 2018 ), 但营养物质从寄主到半寄生植物的转移机制仍未完全研究清楚(
Gebauer et al ., 2012 ,
2018 ).有关研究表明, 半寄生植物
Rhinanthus alectorolophus 中5%-40%的C来自寄主, 而寄主体内的N亦可被半寄生植物获取(
Tě?itel et al ., 2015 ;
Světlíková et al ., 2016 ).寄生植物的吸器同时连接寄主枝条与寄生枝条, 研究寄主枝条-吸器-寄生枝条-寄生叶片这一连续体的生态化学计量特征变化将有助于进一步理解寄生植物与寄主间的养分关系. ...
Ecological Stoichiometry:The Biology of Elements From Molecules to the Biosphere
4
2002
... 生态化学计量学是主要研究生态系统能量平衡和多重化学元素平衡的科学(
Sterner & Elser, 2002 ;
曾德慧和陈广生, 2005 ), 其核心问题是揭示生物体元素组成差异对其生态功能的影响(
Elser et al ., 2000 ;
蒋利玲等, 2014 ).生态化学计量学从元素比率的角度将不同尺度、不同生物群系以及不同领域的生物学研究联系起来(
Sterner & Elser, 2002 ;
Michaels, 2003 ), 成为生态学研究的主要方法和研究热点(
曾冬萍等, 2013 ;
刘文亭等, 2015 ).这一领域研究最多的化学元素是碳(C)、氮(N)、磷(P), 作为3种最重要的生命元素, C、N、P是所有生命过程的基础, 在植物的生化功能方面紧密耦合(
李威, 2016 ).基于不同生命形式, 系统发育以及光合作用碳同化途径的植物叶片生态化学计量特征及其变异模式的研究已取得重大进展(
Han et al ., 2005 ,
2011 ), 但这些研究的主要对象为非寄生植物.有关研究表明, 非寄生植物生态化学计量特征与土壤养分状况相关(
Reich & Oleksyn, 2004 ;
赵一娉等, 2017 ), 但寄生植物没有根系直接接触土壤, 其生命活动所需要的养分和水分主要来自于寄主植物.目前, 有关寄生植物生态化学计量特征的研究鲜有报道. ...
... ).生态化学计量学从元素比率的角度将不同尺度、不同生物群系以及不同领域的生物学研究联系起来(
Sterner & Elser, 2002 ;
Michaels, 2003 ), 成为生态学研究的主要方法和研究热点(
曾冬萍等, 2013 ;
刘文亭等, 2015 ).这一领域研究最多的化学元素是碳(C)、氮(N)、磷(P), 作为3种最重要的生命元素, C、N、P是所有生命过程的基础, 在植物的生化功能方面紧密耦合(
李威, 2016 ).基于不同生命形式, 系统发育以及光合作用碳同化途径的植物叶片生态化学计量特征及其变异模式的研究已取得重大进展(
Han et al ., 2005 ,
2011 ), 但这些研究的主要对象为非寄生植物.有关研究表明, 非寄生植物生态化学计量特征与土壤养分状况相关(
Reich & Oleksyn, 2004 ;
赵一娉等, 2017 ), 但寄生植物没有根系直接接触土壤, 其生命活动所需要的养分和水分主要来自于寄主植物.目前, 有关寄生植物生态化学计量特征的研究鲜有报道. ...
... 植物体中C与N、P含量一般呈负相关关系是高等陆生植物C、N、P计量的普遍规律(
Sterner & Elser, 2002 ;
贺合亮等, 2017 ).在本研究中, 仅寄主枝符合此规律, 其C含量与N、P含量及C:N、C:P、N:P均存在极显著的相关关系.寄生植物吸器的C含量仅与N:P具有显著的相关关系, 枝条的C含量则与N、P含量及C:N、C:P、N:P均不存在极显著的相关关系.寄生植物叶片的C含量与N含量为正相关关系, 与P含量的相关性未达到显著, 也不同于此规律, 但该结果与荒漠植物叶片C、N、P生态化学计量特征具有一致性(
张珂等, 2014 ;
罗艳等, 2017 ).陆生植物的C主要来自大气中的CO
2 (
廖宗文等, 2014 ), 半寄生植物的C既可以来自自身光合作用, 也可以从寄主处获取(
Tě?itel et al ., 2015 ).规律不同说明半寄生植物在固C过程中对养分(N、P等)的利用策略有别于一般陆生植物. ...
... N和P在植物中为协同元素, 植物叶片N、P含量的正相关关系亦为高等陆生植物养分计量的普遍规律(
Sterner & Elser, 2002 ;
Wright et al ., 2005 ).植物N、P间的耦合关系很早就有文献记录(
Garten, 1976 ), 随后也得到许多研究的验证(
Li et al ., 2014 ;
Hidaka & Kitayama, 2015 ;
崔高阳等, 2015 ).对中国753种陆地植物叶片的研究(
Han et al ., 2005 )以及中国东部南北样带654种植物叶片的研究(
任书杰等, 2007 )表明, 不同生活型或不同系统发育植物的叶片N和P含量大部分也呈极显著相关关系.水生植物N、P含量间同样具有强线性关系, 这可能是大多数植物N、P含量的基本特征(
Duarte, 1992 ).
Hidaka和Kitayama (2015) 认为植物叶片N、P含量间的正相关关系可能部分受到土壤可利用性氮含量的影响, 而植物营养策略(N利用策略受到P利用策略的影响)以及植物细胞中的N和P生态化学计量特征(如蛋白质合成中N和P的生化关系)则可提供另一种解释, 但仍需要进一步分析以阐明相关性的生理背景.本研究中, N和P由寄主枝、吸器中的极显著正相关关系, 寄生枝中显著正相关关系, 到寄生叶中变为了相关关系不显著.
Garten (1976) 的研究中大型水生植物的N和P含量相关性也不显著, 这可能是大型水生植物的N含量范围较小, 或者是当环境P缺乏时对N的奢侈消费造成的.
Han等(2005) 和
任书杰等(2007) 的研究中, 裸子植物和蕨类植物的N、P含量相关性也不显著.本研究中寄生叶N、P间的不协同性可能与寄生植物不直接从土壤吸收养分、混合营养策略以及寄生植物叶片重吸收P但不重吸收N有关(
Suriyagoda et al ., 2017 ).受到N、P不耦合的影响, 寄生叶的N含量和C:P, P含量和C:N也不存在显著的相关关系. ...
Nutrient resorption from senescing leaves of epiphytes, hemiparasites and their hosts in tropical forests of Sri Lanka
2
2017
... 半寄生植物与寄主的复杂关系包括寄生和竞争(
Lep? & Tě?itel, 2015 ).作为混合营养策略的代表, 半寄生植物既能从自身的光合作用中获取碳同化物, 也能通过吸器从寄主体内获取碳水化合物(
Nave et al ., 2017 ;
Selosse et al ., 2017 ).半寄生植物自身的光合作用在同化寄生矿物资源, 并将矿物资源转换化成自身生长和适合度中起着至关重要的作用, 而将木质部所获得的资源转化为生长和适应的关键过程, 可能是其进化稳定性的基础(
Tě?itel et al ., 2015 ).尽管半寄生植物与寄主关系的研究开展较早, 不少****对半寄生植物与寄主之间的水分利用、光合作用以及矿物质营养等方面的关系进行了较为深入的研究(
Tennakoon et al ., 2011 ;
Chen et al ., 2013 ;
Suriyagoda et al ., 2017 ;
Gebauer et al ., 2018 ), 但营养物质从寄主到半寄生植物的转移机制仍未完全研究清楚(
Gebauer et al ., 2012 ,
2018 ).有关研究表明, 半寄生植物
Rhinanthus alectorolophus 中5%-40%的C来自寄主, 而寄主体内的N亦可被半寄生植物获取(
Tě?itel et al ., 2015 ;
Světlíková et al ., 2016 ).寄生植物的吸器同时连接寄主枝条与寄生枝条, 研究寄主枝条-吸器-寄生枝条-寄生叶片这一连续体的生态化学计量特征变化将有助于进一步理解寄生植物与寄主间的养分关系. ...
... N和P在植物中为协同元素, 植物叶片N、P含量的正相关关系亦为高等陆生植物养分计量的普遍规律(
Sterner & Elser, 2002 ;
Wright et al ., 2005 ).植物N、P间的耦合关系很早就有文献记录(
Garten, 1976 ), 随后也得到许多研究的验证(
Li et al ., 2014 ;
Hidaka & Kitayama, 2015 ;
崔高阳等, 2015 ).对中国753种陆地植物叶片的研究(
Han et al ., 2005 )以及中国东部南北样带654种植物叶片的研究(
任书杰等, 2007 )表明, 不同生活型或不同系统发育植物的叶片N和P含量大部分也呈极显著相关关系.水生植物N、P含量间同样具有强线性关系, 这可能是大多数植物N、P含量的基本特征(
Duarte, 1992 ).
Hidaka和Kitayama (2015) 认为植物叶片N、P含量间的正相关关系可能部分受到土壤可利用性氮含量的影响, 而植物营养策略(N利用策略受到P利用策略的影响)以及植物细胞中的N和P生态化学计量特征(如蛋白质合成中N和P的生化关系)则可提供另一种解释, 但仍需要进一步分析以阐明相关性的生理背景.本研究中, N和P由寄主枝、吸器中的极显著正相关关系, 寄生枝中显著正相关关系, 到寄生叶中变为了相关关系不显著.
Garten (1976) 的研究中大型水生植物的N和P含量相关性也不显著, 这可能是大型水生植物的N含量范围较小, 或者是当环境P缺乏时对N的奢侈消费造成的.
Han等(2005) 和
任书杰等(2007) 的研究中, 裸子植物和蕨类植物的N、P含量相关性也不显著.本研究中寄生叶N、P间的不协同性可能与寄生植物不直接从土壤吸收养分、混合营养策略以及寄生植物叶片重吸收P但不重吸收N有关(
Suriyagoda et al ., 2017 ).受到N、P不耦合的影响, 寄生叶的N含量和C:P, P含量和C:N也不存在显著的相关关系. ...
Tracing nitrogen flow in a root-hemiparasitic association by foliar stable-isotope labelling
1
2016
... 半寄生植物与寄主的复杂关系包括寄生和竞争(
Lep? & Tě?itel, 2015 ).作为混合营养策略的代表, 半寄生植物既能从自身的光合作用中获取碳同化物, 也能通过吸器从寄主体内获取碳水化合物(
Nave et al ., 2017 ;
Selosse et al ., 2017 ).半寄生植物自身的光合作用在同化寄生矿物资源, 并将矿物资源转换化成自身生长和适合度中起着至关重要的作用, 而将木质部所获得的资源转化为生长和适应的关键过程, 可能是其进化稳定性的基础(
Tě?itel et al ., 2015 ).尽管半寄生植物与寄主关系的研究开展较早, 不少****对半寄生植物与寄主之间的水分利用、光合作用以及矿物质营养等方面的关系进行了较为深入的研究(
Tennakoon et al ., 2011 ;
Chen et al ., 2013 ;
Suriyagoda et al ., 2017 ;
Gebauer et al ., 2018 ), 但营养物质从寄主到半寄生植物的转移机制仍未完全研究清楚(
Gebauer et al ., 2012 ,
2018 ).有关研究表明, 半寄生植物
Rhinanthus alectorolophus 中5%-40%的C来自寄主, 而寄主体内的N亦可被半寄生植物获取(
Tě?itel et al ., 2015 ;
Světlíková et al ., 2016 ).寄生植物的吸器同时连接寄主枝条与寄生枝条, 研究寄主枝条-吸器-寄生枝条-寄生叶片这一连续体的生态化学计量特征变化将有助于进一步理解寄生植物与寄主间的养分关系. ...
Nutritional and isotopic relationships of selected Bornean tropical mistletoe-?host associations in Brunei Darussalam
1
2011
... 半寄生植物与寄主的复杂关系包括寄生和竞争(
Lep? & Tě?itel, 2015 ).作为混合营养策略的代表, 半寄生植物既能从自身的光合作用中获取碳同化物, 也能通过吸器从寄主体内获取碳水化合物(
Nave et al ., 2017 ;
Selosse et al ., 2017 ).半寄生植物自身的光合作用在同化寄生矿物资源, 并将矿物资源转换化成自身生长和适合度中起着至关重要的作用, 而将木质部所获得的资源转化为生长和适应的关键过程, 可能是其进化稳定性的基础(
Tě?itel et al ., 2015 ).尽管半寄生植物与寄主关系的研究开展较早, 不少****对半寄生植物与寄主之间的水分利用、光合作用以及矿物质营养等方面的关系进行了较为深入的研究(
Tennakoon et al ., 2011 ;
Chen et al ., 2013 ;
Suriyagoda et al ., 2017 ;
Gebauer et al ., 2018 ), 但营养物质从寄主到半寄生植物的转移机制仍未完全研究清楚(
Gebauer et al ., 2012 ,
2018 ).有关研究表明, 半寄生植物
Rhinanthus alectorolophus 中5%-40%的C来自寄主, 而寄主体内的N亦可被半寄生植物获取(
Tě?itel et al ., 2015 ;
Světlíková et al ., 2016 ).寄生植物的吸器同时连接寄主枝条与寄生枝条, 研究寄主枝条-吸器-寄生枝条-寄生叶片这一连续体的生态化学计量特征变化将有助于进一步理解寄生植物与寄主间的养分关系. ...
Integrating ecology and physiology of root-hemiparasitic interaction: Interactive effects of abiotic resources shape the interplay between parasitism and autotrophy
3
2015
... 半寄生植物与寄主的复杂关系包括寄生和竞争(
Lep? & Tě?itel, 2015 ).作为混合营养策略的代表, 半寄生植物既能从自身的光合作用中获取碳同化物, 也能通过吸器从寄主体内获取碳水化合物(
Nave et al ., 2017 ;
Selosse et al ., 2017 ).半寄生植物自身的光合作用在同化寄生矿物资源, 并将矿物资源转换化成自身生长和适合度中起着至关重要的作用, 而将木质部所获得的资源转化为生长和适应的关键过程, 可能是其进化稳定性的基础(
Tě?itel et al ., 2015 ).尽管半寄生植物与寄主关系的研究开展较早, 不少****对半寄生植物与寄主之间的水分利用、光合作用以及矿物质营养等方面的关系进行了较为深入的研究(
Tennakoon et al ., 2011 ;
Chen et al ., 2013 ;
Suriyagoda et al ., 2017 ;
Gebauer et al ., 2018 ), 但营养物质从寄主到半寄生植物的转移机制仍未完全研究清楚(
Gebauer et al ., 2012 ,
2018 ).有关研究表明, 半寄生植物
Rhinanthus alectorolophus 中5%-40%的C来自寄主, 而寄主体内的N亦可被半寄生植物获取(
Tě?itel et al ., 2015 ;
Světlíková et al ., 2016 ).寄生植物的吸器同时连接寄主枝条与寄生枝条, 研究寄主枝条-吸器-寄生枝条-寄生叶片这一连续体的生态化学计量特征变化将有助于进一步理解寄生植物与寄主间的养分关系. ...
... 中5%-40%的C来自寄主, 而寄主体内的N亦可被半寄生植物获取(
Tě?itel et al ., 2015 ;
Světlíková et al ., 2016 ).寄生植物的吸器同时连接寄主枝条与寄生枝条, 研究寄主枝条-吸器-寄生枝条-寄生叶片这一连续体的生态化学计量特征变化将有助于进一步理解寄生植物与寄主间的养分关系. ...
... 植物体中C与N、P含量一般呈负相关关系是高等陆生植物C、N、P计量的普遍规律(
Sterner & Elser, 2002 ;
贺合亮等, 2017 ).在本研究中, 仅寄主枝符合此规律, 其C含量与N、P含量及C:N、C:P、N:P均存在极显著的相关关系.寄生植物吸器的C含量仅与N:P具有显著的相关关系, 枝条的C含量则与N、P含量及C:N、C:P、N:P均不存在极显著的相关关系.寄生植物叶片的C含量与N含量为正相关关系, 与P含量的相关性未达到显著, 也不同于此规律, 但该结果与荒漠植物叶片C、N、P生态化学计量特征具有一致性(
张珂等, 2014 ;
罗艳等, 2017 ).陆生植物的C主要来自大气中的CO
2 (
廖宗文等, 2014 ), 半寄生植物的C既可以来自自身光合作用, 也可以从寄主处获取(
Tě?itel et al ., 2015 ).规律不同说明半寄生植物在固C过程中对养分(N、P等)的利用策略有别于一般陆生植物. ...
Global leaf nitrogen and phosphorus stoichiometry and their scaling exponent
1
2018
... N、P在植物叶片中的关系可以通过幂函数
N =
βPα 进行量化(
Wright et al ., 2004 ;
Reich et al ., 2010 ;
Yan et al ., 2018 ).
Wright等(2004) 基于全球范围内广泛的叶片N和P浓度数据, 认为N对P的幂指数约为2/3,
Niklas (2006) 的文章中该指数为0.73, 接近3/4而不是2/3,
Reich等(2010) 收集了更为全面的叶片N、P浓度数据, 提出了叶片N、P浓度计量的2/3幂法则(
N =
αP 2/3 ).目前已有研究表明, 植物叶片N、P计量关系并非恒定, 而是受到植物功能群、纬度区、生态区、施肥类型与水平等多种因素影响(
Tian et al ., 2018 ;
Yan et al ., 2018 ).在本研究中, 寄主枝、吸器与寄生枝的N、P计量关系的回归斜率均小于3/4与2/3, 尽管叶片N、P的计量关系有一些趋势但没有统计学意义.以往的研究主要基于植物叶片的数据, 结果是否适用于不同器官有待验证. ...
Pattern and variation of C:N:P ratios in China’s soils: A synthesis of observational data
1
2010
... 寄生植物是一类重要而独特的植物类群.本研究中两种半寄生植物叶片的平均N含量为12.08 mg·g
-1 , 明显低于全球陆生植物的N含量20.09 mg·g
-1 (
Reich & Oleksyn, 2004 )、中国陆生植物的N含量18.6 mg·g
-1 (
Han et al ., 2005 ); 而P含量3.06 mg·g
-1 则明显地高于全球和中国陆生植物的P含量, 分别为1.77 mg·g
-1 (
Reich & Oleksyn, 2004 )、1.21 mg·g
-1 (
Han et al ., 2005 ).寄生叶的N含量较低可能与其光合作用较弱有关, 而较高的P含量可能受到研究区土壤P含量的间接影响, 哀牢山地区森林土壤的全P和有效P含量分别为1.17 g·kg
-1 和3.40 mg·kg
-1 (
鲁志云等, 2017 ), 中国土壤全P和有效P含量分别为0.60 g·kg
-1 和2.39 mg·kg
-1 (
汪涛等, 2008 ), 哀牢山地区森林土壤全P含量显著高于全国水平(
鲁志云等, 2017 ).此外, 哀牢山地区山地森林表层土壤(0-20 cm)的C:N、C:P、N:P值分别为14.9、84.9、5.7, 高于中国陆地(0-10 cm)土壤的C:N、C:P、N:P水平(分别为12.3、52.7、3.9)(
Tian et al ., 2010 ), 更接近于全球陆地(0-10 cm)和全球森林土壤水平(分别为12.3、72.0、5.9和12.4、81.9、6.6)(
Cleveland & Liptzin, 2007 ).已有的研究认为我国陆地植物生长普遍受P限制(
Han et al ., 2005 ;
任书杰等, 2007 ), 但哀牢山地区较高的土壤P含量暗示该地区可能并不缺P.关于哀牢山地区森林寄生植物、寄主的生态化学计量特征与土壤化学元素计量的关系将是以后研究的重要内容. ...
Foliar and litter nutrients, nutrient resorption, and decomposition in Hawaiian Metrosideros polymorpha
1
1998
... 寄生植物不同的营养素富集程度也不同, 被认为是吸器选择性吸收离子的结果, 若摄取是通过蒸腾而被动吸收, 那么营养元素的比值应保持相同(
Gebauer et al ., 2018 ).本研究中, 椆树桑寄生吸器的C:N、C:P及N:P均与其寄主枝条存在显著差异, 柳叶钝果寄生吸器的C:N和C:P也与其寄主枝条存在显著差异, 这在一定程度上说明C、N和P元素并非简单的通过蒸腾作用而被转运至寄生植物体内, 但也不能排除离子通过被动运输进入吸器后, 由于吸器具有合成物质的功能而导致元素比值发生改变(
黄新亚等, 2011 ).植物不同器官C、N、P含量的差异不仅受植物基本生理过程需求的影响, 也受相应器官组织结构和功能分化的影响(
Minden & Kleyer, 2015 ).叶片是植物的同化器官与重要的养分储存器官, 因而寄生植物叶片的N和P含量高于寄主枝、吸器与寄生枝(
Niklas & Cobb, 2005 ;
贺合亮等, 2017 ).植物叶片中C、N、P含量的变异系数普遍存在C < N < P的规律(
Koerselman & Meuleman, 1996 ;
Güsewell & Koerselman, 2002 ;
姜沛沛等, 2017 ).本研究中两种半寄生植物的寄主枝、吸器、寄生枝与寄生叶的C、N、P含量皆符合这个规律, 这是因为C作为构成植物骨架的基本结构物质, 且为多种植物生理活动提供能源, 而N含量可被植物控制在较窄的范围内, 故C在植物体内含量高且变异小, N含量在各器官中的变异性小于P含量(
Vitousek, 1998 ;
贺合亮等, 2017 ).本研究中两种寄生植物C:N的变异性小于C:P,是因为植物光合作用中C的同化与营养元素的吸收并非同一途径, 而C含量高且变异小, C:N和C:P的变异主要受到N和P含量变化的影响(
Hedin, 2004 ;
曾昭霞等, 2015 ). ...
中国土壤磷库的大小、分布及其影响因素
1
2008
... 寄生植物是一类重要而独特的植物类群.本研究中两种半寄生植物叶片的平均N含量为12.08 mg·g
-1 , 明显低于全球陆生植物的N含量20.09 mg·g
-1 (
Reich & Oleksyn, 2004 )、中国陆生植物的N含量18.6 mg·g
-1 (
Han et al ., 2005 ); 而P含量3.06 mg·g
-1 则明显地高于全球和中国陆生植物的P含量, 分别为1.77 mg·g
-1 (
Reich & Oleksyn, 2004 )、1.21 mg·g
-1 (
Han et al ., 2005 ).寄生叶的N含量较低可能与其光合作用较弱有关, 而较高的P含量可能受到研究区土壤P含量的间接影响, 哀牢山地区森林土壤的全P和有效P含量分别为1.17 g·kg
-1 和3.40 mg·kg
-1 (
鲁志云等, 2017 ), 中国土壤全P和有效P含量分别为0.60 g·kg
-1 和2.39 mg·kg
-1 (
汪涛等, 2008 ), 哀牢山地区森林土壤全P含量显著高于全国水平(
鲁志云等, 2017 ).此外, 哀牢山地区山地森林表层土壤(0-20 cm)的C:N、C:P、N:P值分别为14.9、84.9、5.7, 高于中国陆地(0-10 cm)土壤的C:N、C:P、N:P水平(分别为12.3、52.7、3.9)(
Tian et al ., 2010 ), 更接近于全球陆地(0-10 cm)和全球森林土壤水平(分别为12.3、72.0、5.9和12.4、81.9、6.6)(
Cleveland & Liptzin, 2007 ).已有的研究认为我国陆地植物生长普遍受P限制(
Han et al ., 2005 ;
任书杰等, 2007 ), 但哀牢山地区较高的土壤P含量暗示该地区可能并不缺P.关于哀牢山地区森林寄生植物、寄主的生态化学计量特征与土壤化学元素计量的关系将是以后研究的重要内容. ...
中国土壤磷库的大小、分布及其影响因素
1
2008
... 寄生植物是一类重要而独特的植物类群.本研究中两种半寄生植物叶片的平均N含量为12.08 mg·g
-1 , 明显低于全球陆生植物的N含量20.09 mg·g
-1 (
Reich & Oleksyn, 2004 )、中国陆生植物的N含量18.6 mg·g
-1 (
Han et al ., 2005 ); 而P含量3.06 mg·g
-1 则明显地高于全球和中国陆生植物的P含量, 分别为1.77 mg·g
-1 (
Reich & Oleksyn, 2004 )、1.21 mg·g
-1 (
Han et al ., 2005 ).寄生叶的N含量较低可能与其光合作用较弱有关, 而较高的P含量可能受到研究区土壤P含量的间接影响, 哀牢山地区森林土壤的全P和有效P含量分别为1.17 g·kg
-1 和3.40 mg·kg
-1 (
鲁志云等, 2017 ), 中国土壤全P和有效P含量分别为0.60 g·kg
-1 和2.39 mg·kg
-1 (
汪涛等, 2008 ), 哀牢山地区森林土壤全P含量显著高于全国水平(
鲁志云等, 2017 ).此外, 哀牢山地区山地森林表层土壤(0-20 cm)的C:N、C:P、N:P值分别为14.9、84.9、5.7, 高于中国陆地(0-10 cm)土壤的C:N、C:P、N:P水平(分别为12.3、52.7、3.9)(
Tian et al ., 2010 ), 更接近于全球陆地(0-10 cm)和全球森林土壤水平(分别为12.3、72.0、5.9和12.4、81.9、6.6)(
Cleveland & Liptzin, 2007 ).已有的研究认为我国陆地植物生长普遍受P限制(
Han et al ., 2005 ;
任书杰等, 2007 ), 但哀牢山地区较高的土壤P含量暗示该地区可能并不缺P.关于哀牢山地区森林寄生植物、寄主的生态化学计量特征与土壤化学元素计量的关系将是以后研究的重要内容. ...
西双版纳4种生境下的桑寄生与寄主植物多样性及分布特点
1
2017
... 2017年11月对哀牢山徐家坝地区的寄生植物进行初步调查, 在爱山亭、茶园、生态站、小坝子以及生态站通往小坝子、老周山、簸箕坝等的路旁, 因一定程度的干扰或处于林缘地带, 森林林冠较为开阔、阳光较充足的地方发现较多的寄生植物.通过采集标本进行鉴定, 这些寄生植物主要为椆树桑寄生与柳叶钝果寄生, 故选取这两种常见的半寄生植物作为研究对象.基于这两种半寄生植物的分布特点以及调查地实际情况, 本研究采用样带调查法, 在上述区域设置面积为10 m × 10 m-20 m × 4 000 m的样带进行采样(
王煊妮和张玲, 2017 ). ...
西双版纳4种生境下的桑寄生与寄主植物多样性及分布特点
1
2017
... 2017年11月对哀牢山徐家坝地区的寄生植物进行初步调查, 在爱山亭、茶园、生态站、小坝子以及生态站通往小坝子、老周山、簸箕坝等的路旁, 因一定程度的干扰或处于林缘地带, 森林林冠较为开阔、阳光较充足的地方发现较多的寄生植物.通过采集标本进行鉴定, 这些寄生植物主要为椆树桑寄生与柳叶钝果寄生, 故选取这两种常见的半寄生植物作为研究对象.基于这两种半寄生植物的分布特点以及调查地实际情况, 本研究采用样带调查法, 在上述区域设置面积为10 m × 10 m-20 m × 4 000 m的样带进行采样(
王煊妮和张玲, 2017 ). ...
Parasitic plants as facilitators: More dryad than dracula?
1
2010
... 半寄生植物是寄生植物的主要类群, 其叶片具有叶绿体, 在生命周期的至少一个阶段可进行光合作用, 仅从寄主体内获取部分水和溶解的矿物质(
Nickrent et al ., 1998 ).在自然界中, 半寄生植物对植物群落结构和多样性具有显著影响(
Phoenix & Press, 2010 ;
Press & Phoenix, 2010 ;
Demey et al ., 2014 ), 而其营养丰富、可快速分解和释放养分的凋落物可能会提高森林土壤养分可利用率(
Demey et al ., 2015 ), 影响生态系统水平凋落物分解和养分循环(
Quested et al ., 2010 ).虽然半寄生植物的丰度和生物量相对较小, 但它们对生物地球化学循环、植物种群结构和多样性的影响是很明确的(
Nave et al ., 2017 ), 因而被描述为影响群落动态的生态系统工程师, 以及影响养分循环的关键物种(
Watson, 2010 ;
Hatcher et al ., 2012 ). ...
Plant ecological strategies: Some leading dimensions of variation between species
1
2002
... 大多数有关寄生植物的研究主要关注于植物的光合器官, 而对寄生植物不同器官与寄主植物间的关系研究较少.有关研究表明, 植物叶片N、P浓度与枝条N、P浓度具有相关性(
Kerkhoff et al ., 2006 ), 根和叶间的N浓度也具有显著的相关性(
Craine et al ., 2005 ).本研究寄生枝与寄生叶之间的P含量没有呈现出显著的相关性, 但N含量存在极显著正相关关系, 该结果与
杨梅等(2015) 、
胡耀升等(2014) 的研究果相一致.叶和茎间N含量的相关关系反映了叶、茎在生物、生化方面的功能需求, 这是由植株冠层结构所决定的(
Westoby et al ., 2002 ;
胡耀升等, 2014 ). ...
Assessing the generality of global leaf trait relationships
1
2005
... N和P在植物中为协同元素, 植物叶片N、P含量的正相关关系亦为高等陆生植物养分计量的普遍规律(
Sterner & Elser, 2002 ;
Wright et al ., 2005 ).植物N、P间的耦合关系很早就有文献记录(
Garten, 1976 ), 随后也得到许多研究的验证(
Li et al ., 2014 ;
Hidaka & Kitayama, 2015 ;
崔高阳等, 2015 ).对中国753种陆地植物叶片的研究(
Han et al ., 2005 )以及中国东部南北样带654种植物叶片的研究(
任书杰等, 2007 )表明, 不同生活型或不同系统发育植物的叶片N和P含量大部分也呈极显著相关关系.水生植物N、P含量间同样具有强线性关系, 这可能是大多数植物N、P含量的基本特征(
Duarte, 1992 ).
Hidaka和Kitayama (2015) 认为植物叶片N、P含量间的正相关关系可能部分受到土壤可利用性氮含量的影响, 而植物营养策略(N利用策略受到P利用策略的影响)以及植物细胞中的N和P生态化学计量特征(如蛋白质合成中N和P的生化关系)则可提供另一种解释, 但仍需要进一步分析以阐明相关性的生理背景.本研究中, N和P由寄主枝、吸器中的极显著正相关关系, 寄生枝中显著正相关关系, 到寄生叶中变为了相关关系不显著.
Garten (1976) 的研究中大型水生植物的N和P含量相关性也不显著, 这可能是大型水生植物的N含量范围较小, 或者是当环境P缺乏时对N的奢侈消费造成的.
Han等(2005) 和
任书杰等(2007) 的研究中, 裸子植物和蕨类植物的N、P含量相关性也不显著.本研究中寄生叶N、P间的不协同性可能与寄生植物不直接从土壤吸收养分、混合营养策略以及寄生植物叶片重吸收P但不重吸收N有关(
Suriyagoda et al ., 2017 ).受到N、P不耦合的影响, 寄生叶的N含量和C:P, P含量和C:N也不存在显著的相关关系. ...
The world- wide leaf economics spectrum
2
2004
... N、P在植物叶片中的关系可以通过幂函数
N =
βPα 进行量化(
Wright et al ., 2004 ;
Reich et al ., 2010 ;
Yan et al ., 2018 ).
Wright等(2004) 基于全球范围内广泛的叶片N和P浓度数据, 认为N对P的幂指数约为2/3,
Niklas (2006) 的文章中该指数为0.73, 接近3/4而不是2/3,
Reich等(2010) 收集了更为全面的叶片N、P浓度数据, 提出了叶片N、P浓度计量的2/3幂法则(
N =
αP 2/3 ).目前已有研究表明, 植物叶片N、P计量关系并非恒定, 而是受到植物功能群、纬度区、生态区、施肥类型与水平等多种因素影响(
Tian et al ., 2018 ;
Yan et al ., 2018 ).在本研究中, 寄主枝、吸器与寄生枝的N、P计量关系的回归斜率均小于3/4与2/3, 尽管叶片N、P的计量关系有一些趋势但没有统计学意义.以往的研究主要基于植物叶片的数据, 结果是否适用于不同器官有待验证. ...
... ).
Wright等(2004) 基于全球范围内广泛的叶片N和P浓度数据, 认为N对P的幂指数约为2/3,
Niklas (2006) 的文章中该指数为0.73, 接近3/4而不是2/3,
Reich等(2010) 收集了更为全面的叶片N、P浓度数据, 提出了叶片N、P浓度计量的2/3幂法则(
N =
αP 2/3 ).目前已有研究表明, 植物叶片N、P计量关系并非恒定, 而是受到植物功能群、纬度区、生态区、施肥类型与水平等多种因素影响(
Tian et al ., 2018 ;
Yan et al ., 2018 ).在本研究中, 寄主枝、吸器与寄生枝的N、P计量关系的回归斜率均小于3/4与2/3, 尽管叶片N、P的计量关系有一些趋势但没有统计学意义.以往的研究主要基于植物叶片的数据, 结果是否适用于不同器官有待验证. ...
Nutrient addition affects scaling relationship of leaf nitrogen to phosphorus in Arabidopsis thaliana
2
2018
... N、P在植物叶片中的关系可以通过幂函数
N =
βPα 进行量化(
Wright et al ., 2004 ;
Reich et al ., 2010 ;
Yan et al ., 2018 ).
Wright等(2004) 基于全球范围内广泛的叶片N和P浓度数据, 认为N对P的幂指数约为2/3,
Niklas (2006) 的文章中该指数为0.73, 接近3/4而不是2/3,
Reich等(2010) 收集了更为全面的叶片N、P浓度数据, 提出了叶片N、P浓度计量的2/3幂法则(
N =
αP 2/3 ).目前已有研究表明, 植物叶片N、P计量关系并非恒定, 而是受到植物功能群、纬度区、生态区、施肥类型与水平等多种因素影响(
Tian et al ., 2018 ;
Yan et al ., 2018 ).在本研究中, 寄主枝、吸器与寄生枝的N、P计量关系的回归斜率均小于3/4与2/3, 尽管叶片N、P的计量关系有一些趋势但没有统计学意义.以往的研究主要基于植物叶片的数据, 结果是否适用于不同器官有待验证. ...
... ;
Yan et al ., 2018 ).在本研究中, 寄主枝、吸器与寄生枝的N、P计量关系的回归斜率均小于3/4与2/3, 尽管叶片N、P的计量关系有一些趋势但没有统计学意义.以往的研究主要基于植物叶片的数据, 结果是否适用于不同器官有待验证. ...
不同生长期烤烟各器官C、N、P生态化学计量学特征
1
2015
... 大多数有关寄生植物的研究主要关注于植物的光合器官, 而对寄生植物不同器官与寄主植物间的关系研究较少.有关研究表明, 植物叶片N、P浓度与枝条N、P浓度具有相关性(
Kerkhoff et al ., 2006 ), 根和叶间的N浓度也具有显著的相关性(
Craine et al ., 2005 ).本研究寄生枝与寄生叶之间的P含量没有呈现出显著的相关性, 但N含量存在极显著正相关关系, 该结果与
杨梅等(2015) 、
胡耀升等(2014) 的研究果相一致.叶和茎间N含量的相关关系反映了叶、茎在生物、生化方面的功能需求, 这是由植株冠层结构所决定的(
Westoby et al ., 2002 ;
胡耀升等, 2014 ). ...
不同生长期烤烟各器官C、N、P生态化学计量学特征
1
2015
... 大多数有关寄生植物的研究主要关注于植物的光合器官, 而对寄生植物不同器官与寄主植物间的关系研究较少.有关研究表明, 植物叶片N、P浓度与枝条N、P浓度具有相关性(
Kerkhoff et al ., 2006 ), 根和叶间的N浓度也具有显著的相关性(
Craine et al ., 2005 ).本研究寄生枝与寄生叶之间的P含量没有呈现出显著的相关性, 但N含量存在极显著正相关关系, 该结果与
杨梅等(2015) 、
胡耀升等(2014) 的研究果相一致.叶和茎间N含量的相关关系反映了叶、茎在生物、生化方面的功能需求, 这是由植株冠层结构所决定的(
Westoby et al ., 2002 ;
胡耀升等, 2014 ). ...
哀牢山徐家坝地区的植被分类
1
1983
... 哀牢山国家级自然保护区分布着我国目前保存最完整、面积最大的亚热带山地湿性常绿阔叶林, 林内附生植物丰富, 也有相当数量的寄生植物(
游承侠, 1983 ;
Young & Herwitz, 1995 ;
朱华和闫丽春, 2009 ), 不少****对该地区森林群落类型及结构、树种、附生和寄生植物多样性、生物量等方面进行了研究(
邱学忠和谢寿昌, 1998 ), 但对该区域不同类群植物包括寄生植物的生态化学计量特征及养分利用的了解甚少.本研究以哀牢山山地湿性常绿阔叶林中常见的半寄生植物为对象, 对寄生植物的寄主枝条-吸器-寄生枝条-寄生叶片生态化学计量特征及其动态进行研究, 以期进一步理解半寄生植物的养分利用策略, 为山地森林资源保护与管理提供科学依据. ...
哀牢山徐家坝地区的植被分类
1
1983
... 哀牢山国家级自然保护区分布着我国目前保存最完整、面积最大的亚热带山地湿性常绿阔叶林, 林内附生植物丰富, 也有相当数量的寄生植物(
游承侠, 1983 ;
Young & Herwitz, 1995 ;
朱华和闫丽春, 2009 ), 不少****对该地区森林群落类型及结构、树种、附生和寄生植物多样性、生物量等方面进行了研究(
邱学忠和谢寿昌, 1998 ), 但对该区域不同类群植物包括寄生植物的生态化学计量特征及养分利用的了解甚少.本研究以哀牢山山地湿性常绿阔叶林中常见的半寄生植物为对象, 对寄生植物的寄主枝条-吸器-寄生枝条-寄生叶片生态化学计量特征及其动态进行研究, 以期进一步理解半寄生植物的养分利用策略, 为山地森林资源保护与管理提供科学依据. ...
Floristic diversity and co-occurrences in a subtropical broad-leaved forest and two contrasting regrowth stands in central-west Yunnan Province, China
1
1995
... 哀牢山国家级自然保护区分布着我国目前保存最完整、面积最大的亚热带山地湿性常绿阔叶林, 林内附生植物丰富, 也有相当数量的寄生植物(
游承侠, 1983 ;
Young & Herwitz, 1995 ;
朱华和闫丽春, 2009 ), 不少****对该地区森林群落类型及结构、树种、附生和寄生植物多样性、生物量等方面进行了研究(
邱学忠和谢寿昌, 1998 ), 但对该区域不同类群植物包括寄生植物的生态化学计量特征及养分利用的了解甚少.本研究以哀牢山山地湿性常绿阔叶林中常见的半寄生植物为对象, 对寄生植物的寄主枝条-吸器-寄生枝条-寄生叶片生态化学计量特征及其动态进行研究, 以期进一步理解半寄生植物的养分利用策略, 为山地森林资源保护与管理提供科学依据. ...
生态化学计量学: 复杂生命系统奥秘的探索
1
2005
... 生态化学计量学是主要研究生态系统能量平衡和多重化学元素平衡的科学(
Sterner & Elser, 2002 ;
曾德慧和陈广生, 2005 ), 其核心问题是揭示生物体元素组成差异对其生态功能的影响(
Elser et al ., 2000 ;
蒋利玲等, 2014 ).生态化学计量学从元素比率的角度将不同尺度、不同生物群系以及不同领域的生物学研究联系起来(
Sterner & Elser, 2002 ;
Michaels, 2003 ), 成为生态学研究的主要方法和研究热点(
曾冬萍等, 2013 ;
刘文亭等, 2015 ).这一领域研究最多的化学元素是碳(C)、氮(N)、磷(P), 作为3种最重要的生命元素, C、N、P是所有生命过程的基础, 在植物的生化功能方面紧密耦合(
李威, 2016 ).基于不同生命形式, 系统发育以及光合作用碳同化途径的植物叶片生态化学计量特征及其变异模式的研究已取得重大进展(
Han et al ., 2005 ,
2011 ), 但这些研究的主要对象为非寄生植物.有关研究表明, 非寄生植物生态化学计量特征与土壤养分状况相关(
Reich & Oleksyn, 2004 ;
赵一娉等, 2017 ), 但寄生植物没有根系直接接触土壤, 其生命活动所需要的养分和水分主要来自于寄主植物.目前, 有关寄生植物生态化学计量特征的研究鲜有报道. ...
生态化学计量学: 复杂生命系统奥秘的探索
1
2005
... 生态化学计量学是主要研究生态系统能量平衡和多重化学元素平衡的科学(
Sterner & Elser, 2002 ;
曾德慧和陈广生, 2005 ), 其核心问题是揭示生物体元素组成差异对其生态功能的影响(
Elser et al ., 2000 ;
蒋利玲等, 2014 ).生态化学计量学从元素比率的角度将不同尺度、不同生物群系以及不同领域的生物学研究联系起来(
Sterner & Elser, 2002 ;
Michaels, 2003 ), 成为生态学研究的主要方法和研究热点(
曾冬萍等, 2013 ;
刘文亭等, 2015 ).这一领域研究最多的化学元素是碳(C)、氮(N)、磷(P), 作为3种最重要的生命元素, C、N、P是所有生命过程的基础, 在植物的生化功能方面紧密耦合(
李威, 2016 ).基于不同生命形式, 系统发育以及光合作用碳同化途径的植物叶片生态化学计量特征及其变异模式的研究已取得重大进展(
Han et al ., 2005 ,
2011 ), 但这些研究的主要对象为非寄生植物.有关研究表明, 非寄生植物生态化学计量特征与土壤养分状况相关(
Reich & Oleksyn, 2004 ;
赵一娉等, 2017 ), 但寄生植物没有根系直接接触土壤, 其生命活动所需要的养分和水分主要来自于寄主植物.目前, 有关寄生植物生态化学计量特征的研究鲜有报道. ...
生态化学计量学特征及其应用研究进展
1
2013
... 生态化学计量学是主要研究生态系统能量平衡和多重化学元素平衡的科学(
Sterner & Elser, 2002 ;
曾德慧和陈广生, 2005 ), 其核心问题是揭示生物体元素组成差异对其生态功能的影响(
Elser et al ., 2000 ;
蒋利玲等, 2014 ).生态化学计量学从元素比率的角度将不同尺度、不同生物群系以及不同领域的生物学研究联系起来(
Sterner & Elser, 2002 ;
Michaels, 2003 ), 成为生态学研究的主要方法和研究热点(
曾冬萍等, 2013 ;
刘文亭等, 2015 ).这一领域研究最多的化学元素是碳(C)、氮(N)、磷(P), 作为3种最重要的生命元素, C、N、P是所有生命过程的基础, 在植物的生化功能方面紧密耦合(
李威, 2016 ).基于不同生命形式, 系统发育以及光合作用碳同化途径的植物叶片生态化学计量特征及其变异模式的研究已取得重大进展(
Han et al ., 2005 ,
2011 ), 但这些研究的主要对象为非寄生植物.有关研究表明, 非寄生植物生态化学计量特征与土壤养分状况相关(
Reich & Oleksyn, 2004 ;
赵一娉等, 2017 ), 但寄生植物没有根系直接接触土壤, 其生命活动所需要的养分和水分主要来自于寄主植物.目前, 有关寄生植物生态化学计量特征的研究鲜有报道. ...
生态化学计量学特征及其应用研究进展
1
2013
... 生态化学计量学是主要研究生态系统能量平衡和多重化学元素平衡的科学(
Sterner & Elser, 2002 ;
曾德慧和陈广生, 2005 ), 其核心问题是揭示生物体元素组成差异对其生态功能的影响(
Elser et al ., 2000 ;
蒋利玲等, 2014 ).生态化学计量学从元素比率的角度将不同尺度、不同生物群系以及不同领域的生物学研究联系起来(
Sterner & Elser, 2002 ;
Michaels, 2003 ), 成为生态学研究的主要方法和研究热点(
曾冬萍等, 2013 ;
刘文亭等, 2015 ).这一领域研究最多的化学元素是碳(C)、氮(N)、磷(P), 作为3种最重要的生命元素, C、N、P是所有生命过程的基础, 在植物的生化功能方面紧密耦合(
李威, 2016 ).基于不同生命形式, 系统发育以及光合作用碳同化途径的植物叶片生态化学计量特征及其变异模式的研究已取得重大进展(
Han et al ., 2005 ,
2011 ), 但这些研究的主要对象为非寄生植物.有关研究表明, 非寄生植物生态化学计量特征与土壤养分状况相关(
Reich & Oleksyn, 2004 ;
赵一娉等, 2017 ), 但寄生植物没有根系直接接触土壤, 其生命活动所需要的养分和水分主要来自于寄主植物.目前, 有关寄生植物生态化学计量特征的研究鲜有报道. ...
桂西北喀斯特森林植物-凋落物-土壤生态化学计量特征
1
2015
... 寄生植物不同的营养素富集程度也不同, 被认为是吸器选择性吸收离子的结果, 若摄取是通过蒸腾而被动吸收, 那么营养元素的比值应保持相同(
Gebauer et al ., 2018 ).本研究中, 椆树桑寄生吸器的C:N、C:P及N:P均与其寄主枝条存在显著差异, 柳叶钝果寄生吸器的C:N和C:P也与其寄主枝条存在显著差异, 这在一定程度上说明C、N和P元素并非简单的通过蒸腾作用而被转运至寄生植物体内, 但也不能排除离子通过被动运输进入吸器后, 由于吸器具有合成物质的功能而导致元素比值发生改变(
黄新亚等, 2011 ).植物不同器官C、N、P含量的差异不仅受植物基本生理过程需求的影响, 也受相应器官组织结构和功能分化的影响(
Minden & Kleyer, 2015 ).叶片是植物的同化器官与重要的养分储存器官, 因而寄生植物叶片的N和P含量高于寄主枝、吸器与寄生枝(
Niklas & Cobb, 2005 ;
贺合亮等, 2017 ).植物叶片中C、N、P含量的变异系数普遍存在C < N < P的规律(
Koerselman & Meuleman, 1996 ;
Güsewell & Koerselman, 2002 ;
姜沛沛等, 2017 ).本研究中两种半寄生植物的寄主枝、吸器、寄生枝与寄生叶的C、N、P含量皆符合这个规律, 这是因为C作为构成植物骨架的基本结构物质, 且为多种植物生理活动提供能源, 而N含量可被植物控制在较窄的范围内, 故C在植物体内含量高且变异小, N含量在各器官中的变异性小于P含量(
Vitousek, 1998 ;
贺合亮等, 2017 ).本研究中两种寄生植物C:N的变异性小于C:P,是因为植物光合作用中C的同化与营养元素的吸收并非同一途径, 而C含量高且变异小, C:N和C:P的变异主要受到N和P含量变化的影响(
Hedin, 2004 ;
曾昭霞等, 2015 ). ...
桂西北喀斯特森林植物-凋落物-土壤生态化学计量特征
1
2015
... 寄生植物不同的营养素富集程度也不同, 被认为是吸器选择性吸收离子的结果, 若摄取是通过蒸腾而被动吸收, 那么营养元素的比值应保持相同(
Gebauer et al ., 2018 ).本研究中, 椆树桑寄生吸器的C:N、C:P及N:P均与其寄主枝条存在显著差异, 柳叶钝果寄生吸器的C:N和C:P也与其寄主枝条存在显著差异, 这在一定程度上说明C、N和P元素并非简单的通过蒸腾作用而被转运至寄生植物体内, 但也不能排除离子通过被动运输进入吸器后, 由于吸器具有合成物质的功能而导致元素比值发生改变(
黄新亚等, 2011 ).植物不同器官C、N、P含量的差异不仅受植物基本生理过程需求的影响, 也受相应器官组织结构和功能分化的影响(
Minden & Kleyer, 2015 ).叶片是植物的同化器官与重要的养分储存器官, 因而寄生植物叶片的N和P含量高于寄主枝、吸器与寄生枝(
Niklas & Cobb, 2005 ;
贺合亮等, 2017 ).植物叶片中C、N、P含量的变异系数普遍存在C < N < P的规律(
Koerselman & Meuleman, 1996 ;
Güsewell & Koerselman, 2002 ;
姜沛沛等, 2017 ).本研究中两种半寄生植物的寄主枝、吸器、寄生枝与寄生叶的C、N、P含量皆符合这个规律, 这是因为C作为构成植物骨架的基本结构物质, 且为多种植物生理活动提供能源, 而N含量可被植物控制在较窄的范围内, 故C在植物体内含量高且变异小, N含量在各器官中的变异性小于P含量(
Vitousek, 1998 ;
贺合亮等, 2017 ).本研究中两种寄生植物C:N的变异性小于C:P,是因为植物光合作用中C的同化与营养元素的吸收并非同一途径, 而C含量高且变异小, C:N和C:P的变异主要受到N和P含量变化的影响(
Hedin, 2004 ;
曾昭霞等, 2015 ). ...
阿拉善荒漠典型植物叶片碳、氮、磷化学计量特征
1
2014
... 植物体中C与N、P含量一般呈负相关关系是高等陆生植物C、N、P计量的普遍规律(
Sterner & Elser, 2002 ;
贺合亮等, 2017 ).在本研究中, 仅寄主枝符合此规律, 其C含量与N、P含量及C:N、C:P、N:P均存在极显著的相关关系.寄生植物吸器的C含量仅与N:P具有显著的相关关系, 枝条的C含量则与N、P含量及C:N、C:P、N:P均不存在极显著的相关关系.寄生植物叶片的C含量与N含量为正相关关系, 与P含量的相关性未达到显著, 也不同于此规律, 但该结果与荒漠植物叶片C、N、P生态化学计量特征具有一致性(
张珂等, 2014 ;
罗艳等, 2017 ).陆生植物的C主要来自大气中的CO
2 (
廖宗文等, 2014 ), 半寄生植物的C既可以来自自身光合作用, 也可以从寄主处获取(
Tě?itel et al ., 2015 ).规律不同说明半寄生植物在固C过程中对养分(N、P等)的利用策略有别于一般陆生植物. ...
阿拉善荒漠典型植物叶片碳、氮、磷化学计量特征
1
2014
... 植物体中C与N、P含量一般呈负相关关系是高等陆生植物C、N、P计量的普遍规律(
Sterner & Elser, 2002 ;
贺合亮等, 2017 ).在本研究中, 仅寄主枝符合此规律, 其C含量与N、P含量及C:N、C:P、N:P均存在极显著的相关关系.寄生植物吸器的C含量仅与N:P具有显著的相关关系, 枝条的C含量则与N、P含量及C:N、C:P、N:P均不存在极显著的相关关系.寄生植物叶片的C含量与N含量为正相关关系, 与P含量的相关性未达到显著, 也不同于此规律, 但该结果与荒漠植物叶片C、N、P生态化学计量特征具有一致性(
张珂等, 2014 ;
罗艳等, 2017 ).陆生植物的C主要来自大气中的CO
2 (
廖宗文等, 2014 ), 半寄生植物的C既可以来自自身光合作用, 也可以从寄主处获取(
Tě?itel et al ., 2015 ).规律不同说明半寄生植物在固C过程中对养分(N、P等)的利用策略有别于一般陆生植物. ...
黄土丘陵沟壑区森林生态系统生态化学计量特征
1
2017
... 生态化学计量学是主要研究生态系统能量平衡和多重化学元素平衡的科学(
Sterner & Elser, 2002 ;
曾德慧和陈广生, 2005 ), 其核心问题是揭示生物体元素组成差异对其生态功能的影响(
Elser et al ., 2000 ;
蒋利玲等, 2014 ).生态化学计量学从元素比率的角度将不同尺度、不同生物群系以及不同领域的生物学研究联系起来(
Sterner & Elser, 2002 ;
Michaels, 2003 ), 成为生态学研究的主要方法和研究热点(
曾冬萍等, 2013 ;
刘文亭等, 2015 ).这一领域研究最多的化学元素是碳(C)、氮(N)、磷(P), 作为3种最重要的生命元素, C、N、P是所有生命过程的基础, 在植物的生化功能方面紧密耦合(
李威, 2016 ).基于不同生命形式, 系统发育以及光合作用碳同化途径的植物叶片生态化学计量特征及其变异模式的研究已取得重大进展(
Han et al ., 2005 ,
2011 ), 但这些研究的主要对象为非寄生植物.有关研究表明, 非寄生植物生态化学计量特征与土壤养分状况相关(
Reich & Oleksyn, 2004 ;
赵一娉等, 2017 ), 但寄生植物没有根系直接接触土壤, 其生命活动所需要的养分和水分主要来自于寄主植物.目前, 有关寄生植物生态化学计量特征的研究鲜有报道. ...
黄土丘陵沟壑区森林生态系统生态化学计量特征
1
2017
... 生态化学计量学是主要研究生态系统能量平衡和多重化学元素平衡的科学(
Sterner & Elser, 2002 ;
曾德慧和陈广生, 2005 ), 其核心问题是揭示生物体元素组成差异对其生态功能的影响(
Elser et al ., 2000 ;
蒋利玲等, 2014 ).生态化学计量学从元素比率的角度将不同尺度、不同生物群系以及不同领域的生物学研究联系起来(
Sterner & Elser, 2002 ;
Michaels, 2003 ), 成为生态学研究的主要方法和研究热点(
曾冬萍等, 2013 ;
刘文亭等, 2015 ).这一领域研究最多的化学元素是碳(C)、氮(N)、磷(P), 作为3种最重要的生命元素, C、N、P是所有生命过程的基础, 在植物的生化功能方面紧密耦合(
李威, 2016 ).基于不同生命形式, 系统发育以及光合作用碳同化途径的植物叶片生态化学计量特征及其变异模式的研究已取得重大进展(
Han et al ., 2005 ,
2011 ), 但这些研究的主要对象为非寄生植物.有关研究表明, 非寄生植物生态化学计量特征与土壤养分状况相关(
Reich & Oleksyn, 2004 ;
赵一娉等, 2017 ), 但寄生植物没有根系直接接触土壤, 其生命活动所需要的养分和水分主要来自于寄主植物.目前, 有关寄生植物生态化学计量特征的研究鲜有报道. ...
不同寄主上的桑寄生药材毒性的比较研究
1
2013
... 大多数寄生植物具有广寄性, 而不同的寄生植物其寄主种类有很大的差异(
Norton & Lange, 1999 ;
胡飞和孔垂华, 2004 ).本次调查记录中椆树桑寄生的寄主均为壳斗科植物, 柳叶钝果寄生的寄主较为广泛, 但未发现与椆树桑寄生具有相同寄主.不同的寄主物种对同一寄生植物的影响可能不同, 有研究发现同一桑寄生植物的挥发性成分组成及含量、总黄酮含量、毒性等会因其寄主物种不同而不同(
李永华等, 2009 ,
2012 ;
周汉华等, 2013 ), 但对寄生植物生态化学计量特征与寄主物种的关系进行探讨的研究较少.在本研究中, 寄生于不同寄主的同一寄生植物的同一器官C、N、P生态化学计量特征基本没有显著的不同, 而对同一寄生植物的不同寄主情况下不同器官之间的生态化学计量特征进行双因素方差分析, 也发现寄主物种的影响不显著.这可能与同一寄生植物的不同寄主枝条本身生态化学计量特征差异不显著有关. ...
不同寄主上的桑寄生药材毒性的比较研究
1
2013
... 大多数寄生植物具有广寄性, 而不同的寄生植物其寄主种类有很大的差异(
Norton & Lange, 1999 ;
胡飞和孔垂华, 2004 ).本次调查记录中椆树桑寄生的寄主均为壳斗科植物, 柳叶钝果寄生的寄主较为广泛, 但未发现与椆树桑寄生具有相同寄主.不同的寄主物种对同一寄生植物的影响可能不同, 有研究发现同一桑寄生植物的挥发性成分组成及含量、总黄酮含量、毒性等会因其寄主物种不同而不同(
李永华等, 2009 ,
2012 ;
周汉华等, 2013 ), 但对寄生植物生态化学计量特征与寄主物种的关系进行探讨的研究较少.在本研究中, 寄生于不同寄主的同一寄生植物的同一器官C、N、P生态化学计量特征基本没有显著的不同, 而对同一寄生植物的不同寄主情况下不同器官之间的生态化学计量特征进行双因素方差分析, 也发现寄主物种的影响不显著.这可能与同一寄生植物的不同寄主枝条本身生态化学计量特征差异不显著有关. ...
3
2009
... 哀牢山国家级自然保护区分布着我国目前保存最完整、面积最大的亚热带山地湿性常绿阔叶林, 林内附生植物丰富, 也有相当数量的寄生植物(
游承侠, 1983 ;
Young & Herwitz, 1995 ;
朱华和闫丽春, 2009 ), 不少****对该地区森林群落类型及结构、树种、附生和寄生植物多样性、生物量等方面进行了研究(
邱学忠和谢寿昌, 1998 ), 但对该区域不同类群植物包括寄生植物的生态化学计量特征及养分利用的了解甚少.本研究以哀牢山山地湿性常绿阔叶林中常见的半寄生植物为对象, 对寄生植物的寄主枝条-吸器-寄生枝条-寄生叶片生态化学计量特征及其动态进行研究, 以期进一步理解半寄生植物的养分利用策略, 为山地森林资源保护与管理提供科学依据. ...
... 哀牢山国家级自然保护区位于云南省中部哀牢山山脉中北段上部, 地处云贵高原、横断山和青藏高原南缘三大地理区域的结合部, 是生物多样性较丰富及植物区系地理成分荟萃之地, 该保护区内分布着我国目前保存最完整、面积最大的亚热带山地湿性常绿阔叶林(
朱华和闫丽春, 2009 ).本研究区位于哀牢山北段的徐家坝地区(23.35°-24.44° N, 100.54°-101.30° E), 该地区海拔2 000-2 750 m, 面积约5 100 hm
2 , 年降水量1 947 mm, 年蒸发量1 192 mm, 相对湿度85%, 年平均气温11.3 ℃ (
李苏等, 2007 ).该地区常绿阔叶林的土壤全C、全N与全P含量分别为98.65、6.53和1.17 mg·g
-1 , C:N、C:P、N:P为14.96、91.22和5.95 (
鲁志云等, 2017 ).徐家坝地区分布有椆树桑寄生(
Loranthus delavayi )、柳叶钝果寄生(
Taxillus delavayi )、卵叶梨果寄生(
Scurrula chingii )、五蕊寄生(
Dendrophthoe pentandra )、卵叶槲寄生(
Viscum album var.
meridianum )等寄生植物(
朱华和闫丽春, 2009 ). ...
... )等寄生植物(
朱华和闫丽春, 2009 ). ...
3
2009
... 哀牢山国家级自然保护区分布着我国目前保存最完整、面积最大的亚热带山地湿性常绿阔叶林, 林内附生植物丰富, 也有相当数量的寄生植物(
游承侠, 1983 ;
Young & Herwitz, 1995 ;
朱华和闫丽春, 2009 ), 不少****对该地区森林群落类型及结构、树种、附生和寄生植物多样性、生物量等方面进行了研究(
邱学忠和谢寿昌, 1998 ), 但对该区域不同类群植物包括寄生植物的生态化学计量特征及养分利用的了解甚少.本研究以哀牢山山地湿性常绿阔叶林中常见的半寄生植物为对象, 对寄生植物的寄主枝条-吸器-寄生枝条-寄生叶片生态化学计量特征及其动态进行研究, 以期进一步理解半寄生植物的养分利用策略, 为山地森林资源保护与管理提供科学依据. ...
... 哀牢山国家级自然保护区位于云南省中部哀牢山山脉中北段上部, 地处云贵高原、横断山和青藏高原南缘三大地理区域的结合部, 是生物多样性较丰富及植物区系地理成分荟萃之地, 该保护区内分布着我国目前保存最完整、面积最大的亚热带山地湿性常绿阔叶林(
朱华和闫丽春, 2009 ).本研究区位于哀牢山北段的徐家坝地区(23.35°-24.44° N, 100.54°-101.30° E), 该地区海拔2 000-2 750 m, 面积约5 100 hm
2 , 年降水量1 947 mm, 年蒸发量1 192 mm, 相对湿度85%, 年平均气温11.3 ℃ (
李苏等, 2007 ).该地区常绿阔叶林的土壤全C、全N与全P含量分别为98.65、6.53和1.17 mg·g
-1 , C:N、C:P、N:P为14.96、91.22和5.95 (
鲁志云等, 2017 ).徐家坝地区分布有椆树桑寄生(
Loranthus delavayi )、柳叶钝果寄生(
Taxillus delavayi )、卵叶梨果寄生(
Scurrula chingii )、五蕊寄生(
Dendrophthoe pentandra )、卵叶槲寄生(
Viscum album var.
meridianum )等寄生植物(
朱华和闫丽春, 2009 ). ...
... )等寄生植物(
朱华和闫丽春, 2009 ). ...
 ,1,*, 胡涛1,2, 黄俊彪1,2, 张婷婷1,21
,1,*, 胡涛1,2, 黄俊彪1,2, 张婷婷1,21 
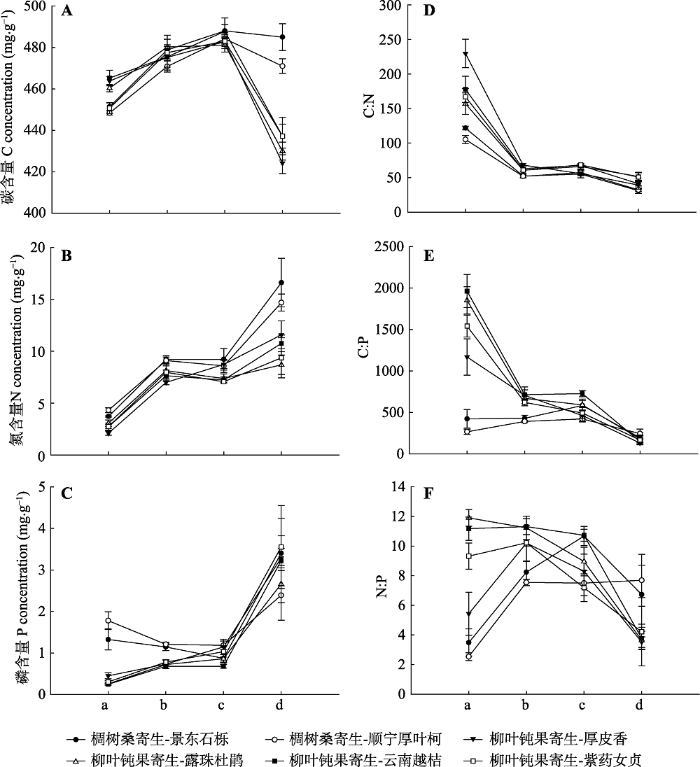 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT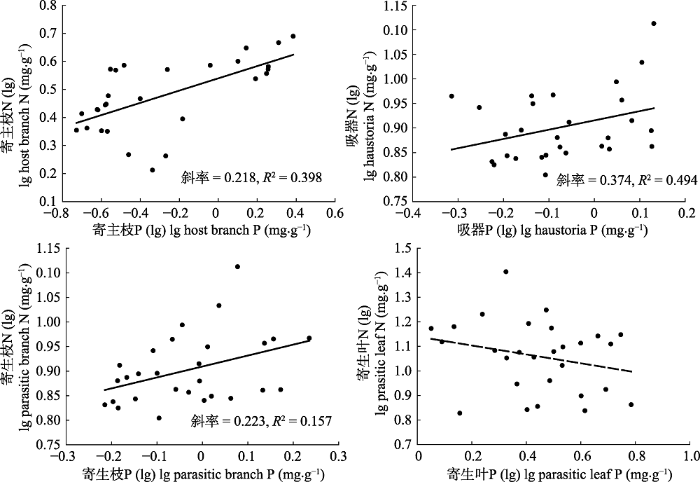 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}