Effects of collar size and buried depth on the measurement of soil respiration in a typical steppe
Jian-Jun LI1,2,*, Lian LIU1,2,*, Di-Ma CHEN1, Feng-Wei XU1,2, Jun-Hui CHENG3, Yong-Fei BAI,1,**1 State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China 2 College of Resources and Environment, University of Chinese Academy of Sciences, Beijing 100049, China 3 College of Grassland and Environment Science, Xingjiang Agricultural University, ürümqi 830052, China;
Abstract Aims Soil respiration plays an important role in carbon cycling in grassland ecosystems. However, the effects of collar size and buried depth during field measurement on soil respiration are rarely assessed. Methods We conducted a two-factor experiment to examine how soil collar depth (2 cm and 5 cm) and size (15 cm × 15 cm and 30 cm × 30 cm) affected the soil respiration (SR), post aboveground net primary productivity (post-ANPP), soil temperature (ST), and soil water content (SWC) in a semi-arid steppe. Important findings The results showed that the deep-inserted soil collar (5 cm soil depth) decreased the soil respiration by 8.0%-9.7% compared with the shallow-inserted soil collar (2 cm soil depth). The large-sized soil collar (30 cm × 30 cm) decreased the soil respiration by 9.1%-10.8% compared with the small-sized soil collar (15 cm × 15 cm). We also found that the deep-inserted and large-sized soil collars had higher ST but lower SWC compared with the shallow-depth and small-sized soil collars. Structural equation model indicated that the lower respiration in the deep-inserted and large-sized soil collars was due to the lower post-ANPP, ST, and SWC. Overall, we found that the soil collar size and buried depth can substantially alter the magnitude of soil respiration by changing plant biomass, ST, and SWC. These findings suggest that the influences of collar size and buried depth on soil respiration should be considered for better estimation and modeling of soil CO2 fluxes in terrestrial ecosystems. Keywords:collar;net primary productivity;soil temperature;soil water content;clonal integration;seasonal dynamics;CO2;structural equation model
PDF (1406KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 李建军, 刘恋, 陈迪马, 许丰伟, 程军回, 白永飞. 底座入土深度和面积对典型草原土壤呼吸测定结果的影响. 植物生态学报, 2019, 43(2): 152-164. DOI: 10.17521/cjpe.2017.0280 LI Jian-Jun, LIU Lian, CHEN Di-Ma, XU Feng-Wei, CHENG Jun-Hui, BAI Yong-Fei. Effects of collar size and buried depth on the measurement of soil respiration in a typical steppe. Chinese Journal of Plant Ecology, 2019, 43(2): 152-164. DOI: 10.17521/cjpe.2017.0280
本研究在中国科学院内蒙古草原生态系统定位研究站(IMGERS, 43.63° N, 116.70° E)综合观测场内进行。该区域海拔高度为1 200 m, 年降水量为346 mm (1982-2014), 降水量年内和年际间变异较大, 60%-80%的降水发生在生长季的5-9月。年平均气温0.3 ℃, 其中1月份平均气温-21.6 ℃, 7月份平均气温19.0 ℃。本区域地带性土壤属暗栗钙土。植物群落的优势植物包括: 羊草、大针茅(Stipa grandis)、冰草(Agropyron cristatum)、糙隐子草(Cleistogenes squarrosa)等(Chen et al., 2013)。
1.2 实验设计
草原生态系统中, 测定土壤呼吸的底座入土深度一般为2-5 cm, 底座直径一般为10-30 cm (Subke et al., 2006; Heinemeyer et al., 2011)。本实验于2013年的5月底至10月中旬进行。实验包括底座入土深度和底座面积2个因子, 其中底座入土深度包括2个水平: 2 cm (D2)和5 cm (D5), 底座入土深度不包括凋落物层厚度; 采用方形底座, 底座面积包括2个水平: 15 cm × 15 cm (S15)和30 cm × 30 cm (S30), 所有底座高度均为8 cm, 但由于入土深度不同, 因而露出地面的高度存在差异。实验采用随机区组设计, 4个处理, 每个处理6次重复, 共计24个1 m × 1 m的实验小区, 小区之间采用间距为2 m的过道进行隔离。底座的安装时间为2013年6月初, 在每个实验小区内, 将底座设置在小区的中央, 为减少底座安装和拆卸对土壤的干扰, 底座始终安置于样地直至实验结束(Xia et al., 2009)。土壤呼吸的测定从6月下旬开始至10月中旬结束, 每月月初对每个小区进行留茬5 cm的模拟放牧处理, 所收获的地上生物量(不包括底座内的部分)从系统中移除, 底座中的生物量单独进行测定。
1.3 土壤呼吸、土壤温度和土壤含水量的测定
综合考虑天气条件, 同时避免降雨脉冲对测定结果的影响(Chen et al., 2009), 本实验的土壤呼吸测定分别在2013年6月24日、7月6日、7月22日、8月14日、9月15日和10月16日的9:00-11:00进行。与底座面积配套, 土壤呼吸测定时将气室放置底座之上。实验时采用两个不透明方形气室(15 cm × 15 cm × 15 cm 和30 cm × 30 cm × 30 cm), 通过与红外气体分析仪(LI-840, LI-COR, Lincoln, USA)以及一个气泵相连的装置来进行测定。为增加测定结果的可靠性, 气室与底座间通过安装密封圈防止漏气, 同时在遮光气室中安装风扇混合气室中的气体, 使红外气体分析仪测得的CO2浓度是混合均匀的。测定的CO2浓度数据通过LI-840软件记录于电脑文档中。LI-840软件数据记录频率设置为1 s, 为保证CO2和H2O浓度在连续测量过程中有60 s的有效数据, 每个小区的测量时间设置为100 s。根据Jasoni等(2005)的分析方法, 本实验用LI-840装置获取的数据计算土壤呼吸。同时, 为保证数据的匹配, 在测定土壤CO2释放速率的同时测定底座内0-10 cm的土壤温度和土壤含水量。土壤温度的测定使用温度测定探针进行, 土壤含水量使用便携式土壤含水量测定仪TDR-200 (Spectrum Technologies, Plainfield, USA)测定。
1.4 处理前底座中地上生产力(pre-AGP)和处理后地上净初级生产力(post-ANPP)
依据以往研究, 本实验在测定土壤呼吸前一天, 将底座内所有绿色植物齐地收获, 目的是消除植物地上部分光合作用和呼吸作用对土壤呼吸的影响, 同时可以降低因植物损伤产生的扰动效应(Chen et al., 2009; Xia et al., 2009; Yan et al., 2010)。6次收获的植物地上部分均带回实验室(为减小生物量移除对结果的影响, 在收获处理后地上净初级生产力时, 从附近剪取相等质量的植物放于底座内), 置于65 ℃的烘箱中烘干至恒质量, 然后混合称量。第一次收获的地上干物质为pre-AGP, 其他5次收获的干物质总和为处理后底座中的post-ANPP。
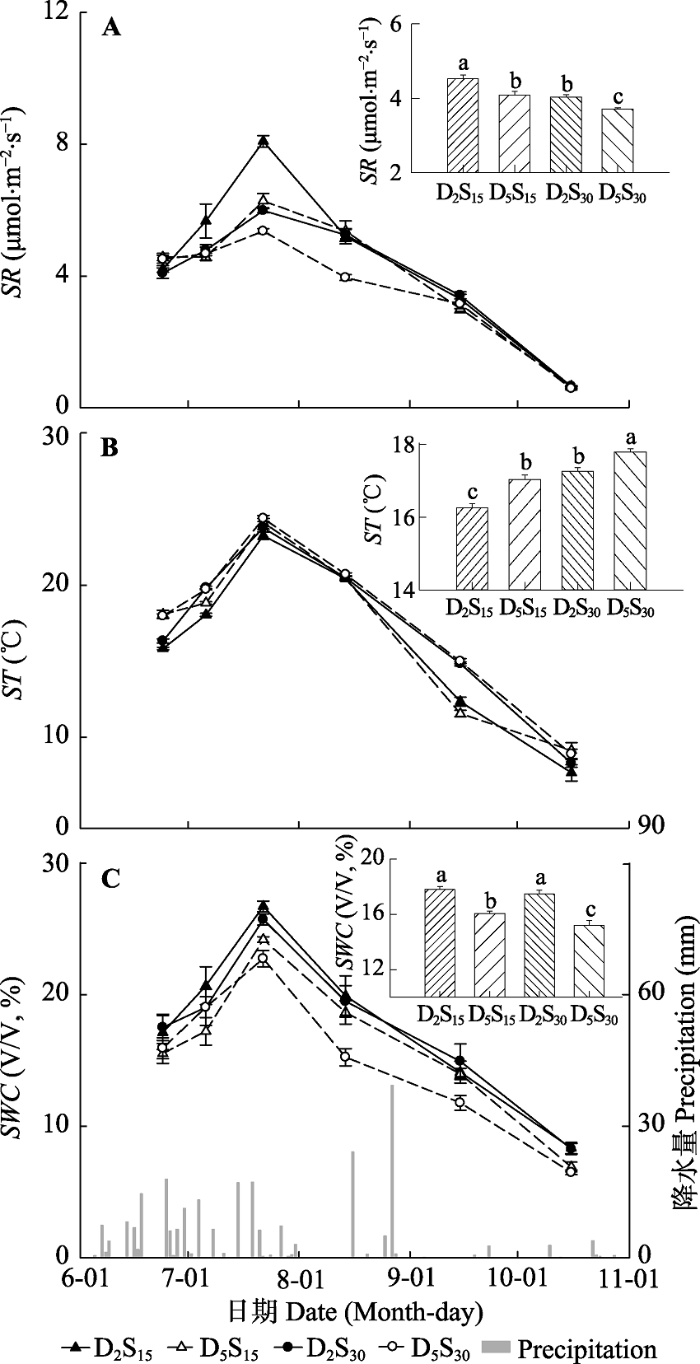
Fig. 1Effects of collar buried depth, collar size, and precipitation events on soil respiration rate (SR)(A), soil temperature (ST)(B), and soil water content (SWC)(C) in the growing season of 2013 (insets: mean ± SE). D2S15、D5S15、D2S30、D5S30 see Table 2. Different lowercase letters indicate significant difference among treatments (p < 0.05).
Table 1 表1 表1底座入土深度(D)、底座边长(S)和测定时间(t)对土壤呼吸速率(SR)、土壤温度(ST)和土壤含水量(SWC)影响的重复测量方差分析结果 Table 1Results of repeated measures ANOVA for the effects of collar buried depth (D), collar size (S), and time of measurement (t) on soil respiration (SR), soil temperature (ST), and soil water content (SWC)
处理 Treatment
响应变量 Response variable
SR
ST
SWC
t
506.6***
2293.6***
311.8***
D
20.5***
143.8***
6.2*
S
35.9***
13.2**
61.4***
t × D
15.1***
24.7***
0.8ns
t × S
6.2**
4.3*
5.5**
D × S
0.6ns
1.5ns
1.2ns
t × D × S
7.2***
2.5ns
0.2ns
ns, p > 0.05; *, p < 0.05; **, p < 0.01; ***, p < 0.001。
Table 2 表2 表2底座入土深度和底座边长对生长季土壤呼吸速率(SR)、土壤温度(ST)和土壤含水量(SWC)单次测量结果的单因素方差分析 Table 2The variables to collar buried depth and side length of the square collar on soil respiration rate (SR), soil temperature (ST), and soil water content (SWC) of each time depend on one-way ANOVA
日期 Date
处理 Treatment
SR
ST
SWC
2013-06-24
D2S15
4.22 ± 0.10ab
15.83 ± 0.08b
17.12 ± 1.30a
D5S15
4.58 ± 0.11a
18.08 ± 0.27a
15.52 ± 0.75a
D2S30
4.10 ± 0.16b
16.33 ± 0.14b
17.50 ± 1.00a
D5S30
4.52 ± 0.12a
17.98 ± 0.13a
15.92 ± 0.8a
2013-07-06
D2S15
5.68 ± 0.52a
18.05 ± 0.10c
20.67 ± 1.43a
D5S15
4.79 ± 0.16b
19.83 ± 0.11a
17.20 ± 1.05a
D2S30
4.58 ± 0.11b
18.83 ± 0.08b
19.02 ± 1.36a
D5S30
4.69 ± 0.20b
19.73 ± 0.05a
19.03 ± 0.80a
2013-07-22
D2S15
8.10 ± 0.18a
23.27 ± 0.24c
26.68 ± 0.4a
D5S15
6.00 ± 0.06b
23.78 ± 0.07bc
24.17 ± 0.21b
D2S30
6.29 ± 0.22b
24.13 ± 0.26ab
25.68 ± 0.43a
D5S30
5.37 ± 0.07c
24.40 ± 0.20a
22.72 ± 0.62c
2013-08-14
D2S15
5.21 ± 0.22a
20.43 ± 0.13a
19.88 ± 1.53a
D5S15
5.26 ± 0.20a
20.48 ± 0.08a
18.63 ± 0.88a
D2S30
5.38 ± 0.31a
20.43 ± 0.13a
19.47 ± 1.19a
D5S30
3.96 ± 0.09b
20.72 ± 0.09a
15.23 ± 0.65b
2013-09-15
D2S15
3.31 ± 0.15a
12.33 ± 0.29b
14.10 ± 0.83ab
D5S15
3.42 ± 0.11a
14.83 ± 0.13a
13.97 ± 0.43ab
D2S30
3.02 ± 0.13a
11.55 ± 0.22c
14.93 ± 1.3a
D5S30
3.17 ± 0.15a
15.00 ± 0.17a
11.77 ± 0.56b
2013-10-16
D2S15
0.63 ± 0.03a
7.65 ± 0.54b
8.33 ± 0.42a
D5S15
0.66 ± 0.02a
8.28 ± 0.27ab
6.90 ± 0.38b
D2S30
0.67 ± 0.02a
9.12 ± 0.52a
8.27 ± 0.41a
D5S30
0.60 ± 0.04a
8.88 ± 0.31ab
6.48 ± 0.17b
D and S denote the buried depth and length of the square soil collar, respectively. D2S15, D = 2 cm and S =15 cm; D5S15, D = 5 cm, S = 15 cm; D2S30, D = 2 cm, S = 30 cm; and D5S30, D = 5 cm, S = 30 cm. Different lowercase letters in the same column indicate significantly different effect (p < 0.05, n = 6). D2S15表示不锈钢方形底座入土深度为2 cm, 边长15 cm, D5S15表示深度为5 cm, 边长15 cm, D2S30表示深度为2 cm, 边长30 cm, D5S30表示深度为5 cm, 边长30 cm。同列的不同小写字母表示差异显著(p < 0.05, n = 6)。
Fig. 2Results of ANOVAs for aboveground biomass before treatments (pre-AGP)(A), the aboveground net primary productivity (post-ANPP)(B) in the growing season of 2013 (mean ± SE). Different lowercase letters indicate significant difference among treatments (p < 0.05).
新窗口打开|下载原图ZIP|生成PPT 图3不同底座处理下土壤呼吸速率(SR)与土壤温度(ST)之间的关系。D2S15表示不锈钢方形底座入土深度为2 cm, 边长15 cm, D5S15表示深度为5 cm, 边长15 cm, D2S30表示深度为2 cm, 边长30 cm, D5S30表示深度为5 cm, 边长30 cm。
Fig. 3Relationship between soil respiration rate (SR) and soil temperature (ST) at 10 cm soil depth under different treatments. D and S denote the buried depth and length of the square soil collar, respectively. D2S15, D = 2 cm and S =15 cm; D5S15, D = 5 cm, S = 15 cm; D2S30, D = 2 cm, S = 30 cm; and D5S30, D = 5 cm, S = 30 cm.
图4
新窗口打开|下载原图ZIP|生成PPT 图4不同底座处理下土壤呼吸速率(SR)与土壤含水量(SWC)的关系。D2S15表示不锈钢方形底座入土深度为2 cm, 边长15 cm, D5S15表示深度为5 cm, 边长15 cm, D2S30表示深度为2 cm, 边长30 cm, D5S30表示深度为5 cm, 边长30 cm。
Fig. 4Relationship between soil respiration rate (SR) and soil water content (SWC) at 10 cm soil depth under different treatments. D and S denote the buried depth and length of the square soil collar, respectively. D2S15, D = 2 cm and S =15 cm; D5S15, D = 5 cm, S = 15 cm; D2S30, D = 2 cm, S = 30 cm; and D5S30, D = 5 cm, S = 30 cm.
图5
新窗口打开|下载原图ZIP|生成PPT 图5不同底座处理下土壤呼吸速率(SR)与底座内地上净初级生产力(post-ANPP)之间的相关关系。D2S15表示不锈钢方形底座入土深度为2 cm, 边长15 cm, D5S15表示深度为5 cm, 边长15 cm, D2S30表示深度为2 cm, 边长30 cm, D5S30表示深度为5 cm, 边长30 cm。
Fig. 5Relationships between soil respiration rate (SR) and aboveground net primary productively (post-ANPP) under different treatments. D and S denote the buried depth and length of the square soil collar, respectively. D2S15, D = 2 cm and S =15 cm; D5S15, D = 5 cm, S = 15 cm; D2S30, D = 2 cm, S = 30 cm; and D5S30, D = 5 cm, S = 30 cm.
Fig. 6Structural equation modeling analysis for the effects of collar depth (D) and size (S) on soil respiration rate, via pathways of biotic and abiotic factors during the study period. Square boxes indicate variables included in the model. Results of model fitting: χ2 = 0.89, p = 0.874 > 0.085, df = 5, RMSEA = 0.000 < 0.05, AGFI = 0.900 > 0.90, GFI = 0.975 > 0.90 (which indicates a good fit of the model to the data). Black and gray solid arrows indicate significantly positive and negative effects, respectively and dashed arrows indicate insignificant effects (p > 0.05). Values associated with the arrows represent standardized path coefficients. R2 values associated with response variables indicate the proportion of variation explained by relationships with all other variables. RMSEA, root-mean-square error of approximation; AGFI, adjusted goodness-of-fit index; GFI, goodness-of-fit index. post-ANPP, post aboveground net primary productivity; SR, soil respiration; ST, soil temperature; SWC, soil water content. **, p < 0.01; ***, p < 0.001.
AtkinOK, EdwardsEJ, LoveysBR ( 2000). Response of root respiration to changes in temperature and its relevance to global warming New Phytologist, 147, 141-154. DOI:10.1046/j.1469-8137.2000.00683.xURL [本文引用: 1]
BahnM, RodeghieroM, Anderson-DunnM, DoreS, GimenoC, Dr?slerM, WilliamsM, AmmannC, BerningerF, FlechardC ( 2008). Soil respiration in European grasslands in relation to climate and assimilate supply Ecosystems, 11, 1352-1367. DOI:10.1007/s10021-008-9198-0URL [本文引用: 2]
BaiYF, HanXG, WuJG, ChenZZ, LiLH ( 2004). Ecosystem stability and compensatory effects in the Inner Mongolia grassland Nature, 431, 181-184. DOI:10.1038/nature02850 [本文引用: 1]
BaiYF, WuJG, XingQ, PanQM, HuangJH, YangDL, HanXG ( 2008). Primary production and rain use efficiency across a precipitation gradient on the Mongolia Plateau Ecology, 89, 2140-2153. DOI:10.1890/07-0992.1URL [本文引用: 1]
BatjesNH ( 1996). Total carbon and nitrogen in the soils of the world European Journal of Soil Science, 47, 151-163. DOI:10.1111/ejs.1996.47.issue-2URL [本文引用: 1]
BerryLJ, NorrisW ( 1949). Studies of onion root respiration I. Velocity of oxygen consumption in different segments of root at different temperatures as a function of partial pressure of oxygen Biochimica et Biophysica Acta, 3, 593-606. DOI:10.1016/0006-3002(49)90133-XURL [本文引用: 1]
BooneRD, NadelhofferKJ, CanaryJD, KayeJP ( 1998). Roots exert a strong influence on the temperature sensitivity of soil respiration Nature, 396, 570-572. DOI:10.1038/25119 [本文引用: 1]
CaoGM, TangYH, MoWH, WangYS, LiYN, ZhaoXQ ( 2004). Grazing intensity alters soil respiration in an alpine meadow on the Tibetan Plateau Soil Biology & Biochemistry, 36, 237-243. [本文引用: 1]
ChenD, ZhengS, ShanY, TaubeF, BaiY ( 2013). Vertebrate herbivore-induced changes in plants and soils: Linkages to ecosystem functioning in a semi-arid steppe Functional Ecology, 27, 273-281. DOI:10.1111/fec.2013.27.issue-1URL [本文引用: 1]
ChenDM, LiJJ, LanZC, HuSJ, BaiYF ( 2016). Soil acidification exerts a greater control on soil respiration than soil nitrogen availability in grasslands subjected to long-term nitrogen enrichment Functional Ecology, 30, 658-669. DOI:10.1111/1365-2435.12525URL [本文引用: 2]
ChenSP, LinGH, HuangJH, JeneretteGD ( 2009). Dependence of carbon sequestration on the differential responses of ecosystem photosynthesis and respiration to rain pulses in a semiarid steppe Global Change Biology, 15, 2450-2461. DOI:10.1111/gcb.2009.15.issue-10URL [本文引用: 5]
DavidsonEA, VerchotLV, CattanioJH, AckermanIL, CarvalhoJ ( 2000). Effects of soil water content on soil respiration in forests and cattle pastures of eastern Amazonia Biogeochemistry, 48, 53-69. DOI:10.1023/A:1006204113917URL [本文引用: 2]
DrewittGB, BlackTA, NesicZ, HumphreysER, JorkEM, SwansonR, EthierGJ, GriffisT, MorgensternK ( 2002). Measuring forest floor CO2 fluxes in a Douglas-fir forest Agricultural and Forest Meteorology, 110, 299-317. DOI:10.1016/S0168-1923(01)00294-5URL [本文引用: 1]
GuenetB, JuarezS, BardouxG, AbbadieL, ChenuC ( 2012). Evidence that stable C is as vulnerable to priming effect as is more labile C in soil Soil Biology & Biochemistry, 52, 43-48. [本文引用: 1]
HeinemeyerA, Di BeneC, LloydAR, TortorellaD, BaxterR, HuntleyB, GelsominoA, InesonP ( 2011). Soil respiration: Implications of the plant-soil continuum and respiration chamber collar-insertion depth on measurement and modelling of soil CO2 efflux rates in three ecosystems European Journal of Soil Science, 62, 82-94. DOI:10.1111/ejs.2011.62.issue-1URL [本文引用: 8]
H?gbergP, ReadDJ ( 2006). Towards a more plant physiological perspective on soil ecology Trends in Ecology & Evolution, 21, 548-554. [本文引用: 1]
HutchinsonGL, LivingstonGP ( 2001). Vents and seals in non-steady-state chambers used for measuring gas exchange between soil and the atmosphere European Journal of Soil Science, 52, 675-682. DOI:10.1046/j.1365-2389.2001.00415.xURL [本文引用: 1]
JasoniRL, SmithSD, ArnoneJA ( 2005). Net ecosystem CO2 exchange in Mojave Desert shrublands during the eighth year of exposure to elevated CO2 Global Change Biology, 11, 749-756. DOI:10.1111/gcb.2005.11.issue-5URL [本文引用: 2]
KirschbaumMUF ( 2006). The temperature dependence of organic-?matter decomposition—Still a topic of debate Soil Biology & Biochemistry, 38, 2510-2518. [本文引用: 1]
KutschWL, StaackA, W?tzelJ, MiddelhoffU, KappenL ( 2001). Field measurements of root respiration and total soil respiration in an alder forest New Phytologist, 150, 157-168. DOI:10.1046/j.1469-8137.2001.00071.xURL [本文引用: 2]
KuzyakovY ( 2010). Priming effects: Interactions between living and dead organic matter Soil Biology & Biochemistry, 42, 1363-1371. [本文引用: 1]
KuzyakovY, ChengW ( 2001). Photosynthesis controls of rhizosphere respiration and organic matter decomposition Soil Biology and Biochemistry, 33, 1915-1925. DOI:10.1016/S0038-0717(01)00117-1URL [本文引用: 1]
LiW, CaoWX, LiuHD, LiXL, XuCL, ShiSL, FengJ, ZhouCM ( 2015). Analysis of soil respiration under different grazing management patterns in the alpine meadow-steppe of the Qinghai-Tibet Plateau Acta Prataculturae Sinica, 24, 22-32. [本文引用: 1]
LloydJ, TaylorJA ( 1994). On the temperature dependence of soil respiration Functional Ecology, 8, 315-323. DOI:10.2307/2389824URL [本文引用: 2]
MikanCJ, SchimelJP, DoyleAP ( 2002). Temperature controls of microbial respiration in arctic tundra soils above and below freezing Soil Biology & Biochemistry, 34, 1785-1795. [本文引用: 1]
NaySM, MattsonKG, BormannBT ( 1994). Biases of chamber methods for measuring soil CO2 efflux demonstrated with a laboratory apparatus Ecology, 75, 2460-2463. DOI:10.2307/1940900URL [本文引用: 1]
NgaoJ, LongdozB, PerrinD, VincentG, EpronD, Le DantecV, SoudaniK, AubinetM, WillmF, GranierA ( 2006). Cross-calibration functions for soil CO2 efflux measurement systems Annals of Forest Science, 63, 477-484. DOI:10.1051/forest:2006028URL [本文引用: 1]
RaichJW, PotterCS, BhagawatiD ( 2002). Interannual variability in global soil respiration, 1980-94 Global Change Biology, 8, 800-812. DOI:10.1046/j.1365-2486.2002.00511.xURL [本文引用: 1]
RaichJW, SchlesingerWH ( 1992). The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate Tellus B, 44, 81-99. DOI:10.3402/tellusb.v44i2.15428URL [本文引用: 1]
ReichsteinM, ReyA, FreibauerA, TenhunenJ, ValentiniR, BanzaJ, CasalsP, ChengY, GrünzweigJM, IrvineJ ( 2003). Modeling temporal and large-scale spatial variability of soil respiration from soil water availability, temperature and vegetation productivity indices Global Biogeochemical Cycles, 17, 1104. [本文引用: 1]
SchlesingerWH, AndrewsJA ( 2000). Soil respiration and the global carbon cycle Biogeochemistry, 48, 7-20. DOI:10.1023/A:1006247623877URL [本文引用: 2]
SubkeJA, InglimaI, Francesca CotrufoM ( 2006). Trends and methodological impacts in soil CO2 efflux partitioning: A meta analytical review Global Change Biology, 12, 921-943. DOI:10.1111/gcb.2006.12.issue-6URL [本文引用: 5]
SubkeJA, MoodyCS, HillTC, VokeN, ToetS, InesonP, TehY ( 2018). Rhizosphere activity and atmospheric methane concentrations drive variations of methane fluxes in a temperate forest soil Soil Biology & Biochemistry, 116, 323-332. [本文引用: 1]
WanSQ, LuoYQ ( 2003). Substrate regulation of soil respiration in a tallgrass prairie: Results of a clipping and shading experiment Global Biogeochemical Cycles, 17, 1054. DOI: 10.1029/2002GB001971. [本文引用: 1]
WangN, YuFH, LiPX, HeWM, LiuJ, YuGL, SongYB, DongM ( 2009). Clonal integration supports the expansion from terrestrial to aquatic environments of the amphibious stoloniferous herb Alternanthera philoxeroides. Plant Biology, 11, 483-489. DOI:10.1111/plb.2009.11.issue-3URL [本文引用: 1]
WangWJ, ZuYG, WangHM, HiranoT, TakagiK, SasaK, KoikeT ( 2005). Effect of collar insertion on soil respiration in a larch forest measured with a LI-6400 soil CO2 flux system Journal of Forest Research, 10, 57-60. DOI:10.1007/s10310-004-0102-2URL [本文引用: 1]
WangY, LiuH, ChungH, YuL, MiZ, GengY, JingX, WangS, ZengH, CaoG, ZhaoX, HeJS ( 2014). Non-growing-season soil respiration is controlled by freezing and thawing processes in the summer monsoon-?dominated Tibetan alpine grassland Global Biogeochemical Cycles, 28, 1081-1095. DOI:10.1002/2013GB004760URL [本文引用: 1]
WardSE, SmartSM, QuirkH, TallowinJRB, MortimerSR, ShielRS, WilbyA, BardgettRD ( 2016). Legacy effects of grassland management on soil carbon to depth Global Change Biology, 22, 2929-2938. DOI:10.1111/gcb.2016.22.issue-8URL [本文引用: 2]
WidénB, MajdiH ( 2001). Soil CO2 efflux and root respiration at three sites in a mixed pine and spruce forest: Seasonal and diurnal variation Canadian Journal of Forest Research, 31, 786-796. DOI:10.1139/x01-012URL [本文引用: 1]
XiaJY, NiuSL, WanSQ ( 2009). Response of ecosystem carbon exchange to warming and nitrogen addition during two hydrologically contrasting growing seasons in a temperate steppe Global Change Biology, 15, 1544-1556. DOI:10.1111/gcb.2009.15.issue-6URL [本文引用: 2]
YanLM, ChenSQ, HuangJH, LinGH ( 2010). Differential responses of auto- and heterotrophic soil respiration to water and nitrogen addition in a semiarid temperate steppe Global Change Biology, 16, 2345-2357. [本文引用: 4]
ZhangXL, TanYL, ZhangBW, LiA, DaryantoS, WangLX, HuangJH ( 2017). The impacts of precipitation increase and nitrogen addition on soil respiration in a semiarid temperate steppe Ecosphere, 8, e01655. DOI: 10.1002/ecs2.1655. DOI:10.1002/ecs2.1655URL [本文引用: 2]
ZhouGY, ZhouXH, HeYH, ShaoJJ, HuZH, LiuRQ, ZhouHM, HosseinibaiS ( 2017). Grazing intensity significantly affects belowground carbon and nitrogen cycling in grassland ecosystems: A meta-analysis Global Change Biology, 23, 1167-1179. DOI:10.1111/gcb.2017.23.issue-3URL [本文引用: 1]
ZhouXH, SherryRA, AnY, WallaceLL, LuoYQ ( 2006). Main and interactive effects of warming, clipping, and doubled precipitation on soil CO2 efflux in a grassland ecosystem Global Biogeochemical Cycles, 20, GB1003. DOI:10.1029/2005GB002526. [本文引用: 1]
ZhouXH, WanSQ, LuoYQ ( 2007). Source components and interannual variability of soil CO2 efflux under experimental warming and clipping in a grassland ecosystem Global Change Biology, 13, 761-775. [本文引用: 1]
Response of root respiration to changes in temperature and its relevance to global warming 1 2000
... 尽管底座面积不同, 但是入土较深的底座处理中土壤温度均显著高于入土较浅的处理, 而土壤温度对土壤呼吸具有促进作用.相反, 入土较深的底座处理中土壤含水量则显著低于较浅的处理, “高温干旱”使较深的底座处理中土壤呼吸强度降低.在本实验中, 底座处理通过直接改变土壤温度和土壤含水量来间接地影响土壤呼吸, 且主要是基于土壤含水量降低对土壤呼吸产生的抑制效应.许多研究表明, 土壤温度和土壤含水量是影响土壤呼吸重要的环境因子(Lloyd & Taylor, 1994; Davidson et al., 2000), 尤其是在干旱半干旱草原, 土壤水分有效性强烈影响生态系统土壤呼吸过程(Chen et al., 2009).研究表明, 无论是植物根系呼吸(Berry & Norris, 1949; Atkin et al., 2000), 还是土壤微生物呼吸(Mikan et al., 2002), 在土壤温度较低时, 土壤呼吸速率随着温度的升高而呈指数增加.但在土壤含水量较低的干旱期, 土壤温度升高反而抑制土壤CO2的释放(Zhou et al., 2006).以往研究发现, 土壤呼吸不同组分的温度敏感性不同, 根系周围土壤的异养呼吸比根系和根际自养呼吸对温度的敏感性更低(Boone et al., 1998), 本研究中, 无论底座面积大小, 入土较深的处理, 其土壤呼吸对土壤温度的敏感性更低(Q10值小).入土较深的处理, 其根系被切断的总量更多, 使土壤呼吸中自养呼吸部分降低, 促使异养呼吸占总呼吸的比例增加, 这可能是底座入土较深的处理中土壤呼吸对温度的敏感性更低的原因. ...
Soil respiration in European grasslands in relation to climate and assimilate supply 2 2008
... 在极端降雨事件发生时, 底座内可能出现水涝现象(Heinemeyer et al., 2011), 进而使底座内的土壤温度和含水量发生变化.尤其是在干旱和半干旱地区, 水分是生态系统结构与功能的重要限制因子(Davidson et al., 2000; Bai et al., 2008), 土壤水分有效性的增加, 可促使底座区域的土壤呼吸增强.同时, 底座阻隔了底座内外植物间物质的横向运输, 底座内外土壤蒸发散的差异会影响表层土壤温度(Reichstein et al., 2003), 而土壤呼吸的变化受土壤温度(Lloyd & Taylor, 1994; Zhou et al., 2007)、土壤含水量(Yan et al., 2010)和养分供应(Bahn et al., 2008)的共同影响.因此, 底座设置方面的不足, 可能直接或者间接地影响以上因素, 最终对土壤呼吸的测定结果产生影响. ...
Ecosystem stability and compensatory effects in the Inner Mongolia grassland 1 2004
... 同时, 本研究还发现底座面积的增加显著降低了表层土壤湿度.与面积较小的底座相比较, 在面积的较大底座内, 地上净初级生产力的下降可能导致蒸腾作用下降, 使土壤损失的水分减少, 同时地表接受太阳辐射增强, 可能使表层土壤温度显著升高, 导致表层土壤含水量下降, 因此底座内土壤含水量的变化取决于处理对蒸发和蒸腾影响的相对大小.实验结果显示, 面积较大的底座内土壤含水量较低, 表明底座面积较大的处理对蒸发作用的正效应大于对蒸腾作用的负效应, 最终导致较大的底座处理区域更加“高温干旱”.以往研究表明, 在干旱半干旱草原区域, 水分是草原生态系统土壤呼吸和初级生产力重要的限制因子(Bai et al., 2004; Yan et al., 2010), 高温干旱的环境, 抑制了土壤呼吸, 最终使底座面积较大的处理其土壤呼吸测定值较小(Cao et al., 2004). ...
Primary production and rain use efficiency across a precipitation gradient on the Mongolia Plateau 1 2008
... 在极端降雨事件发生时, 底座内可能出现水涝现象(Heinemeyer et al., 2011), 进而使底座内的土壤温度和含水量发生变化.尤其是在干旱和半干旱地区, 水分是生态系统结构与功能的重要限制因子(Davidson et al., 2000; Bai et al., 2008), 土壤水分有效性的增加, 可促使底座区域的土壤呼吸增强.同时, 底座阻隔了底座内外植物间物质的横向运输, 底座内外土壤蒸发散的差异会影响表层土壤温度(Reichstein et al., 2003), 而土壤呼吸的变化受土壤温度(Lloyd & Taylor, 1994; Zhou et al., 2007)、土壤含水量(Yan et al., 2010)和养分供应(Bahn et al., 2008)的共同影响.因此, 底座设置方面的不足, 可能直接或者间接地影响以上因素, 最终对土壤呼吸的测定结果产生影响. ...
Total carbon and nitrogen in the soils of the world 1 1996
Studies of onion root respiration I. Velocity of oxygen consumption in different segments of root at different temperatures as a function of partial pressure of oxygen 1 1949
... 尽管底座面积不同, 但是入土较深的底座处理中土壤温度均显著高于入土较浅的处理, 而土壤温度对土壤呼吸具有促进作用.相反, 入土较深的底座处理中土壤含水量则显著低于较浅的处理, “高温干旱”使较深的底座处理中土壤呼吸强度降低.在本实验中, 底座处理通过直接改变土壤温度和土壤含水量来间接地影响土壤呼吸, 且主要是基于土壤含水量降低对土壤呼吸产生的抑制效应.许多研究表明, 土壤温度和土壤含水量是影响土壤呼吸重要的环境因子(Lloyd & Taylor, 1994; Davidson et al., 2000), 尤其是在干旱半干旱草原, 土壤水分有效性强烈影响生态系统土壤呼吸过程(Chen et al., 2009).研究表明, 无论是植物根系呼吸(Berry & Norris, 1949; Atkin et al., 2000), 还是土壤微生物呼吸(Mikan et al., 2002), 在土壤温度较低时, 土壤呼吸速率随着温度的升高而呈指数增加.但在土壤含水量较低的干旱期, 土壤温度升高反而抑制土壤CO2的释放(Zhou et al., 2006).以往研究发现, 土壤呼吸不同组分的温度敏感性不同, 根系周围土壤的异养呼吸比根系和根际自养呼吸对温度的敏感性更低(Boone et al., 1998), 本研究中, 无论底座面积大小, 入土较深的处理, 其土壤呼吸对土壤温度的敏感性更低(Q10值小).入土较深的处理, 其根系被切断的总量更多, 使土壤呼吸中自养呼吸部分降低, 促使异养呼吸占总呼吸的比例增加, 这可能是底座入土较深的处理中土壤呼吸对温度的敏感性更低的原因. ...
Roots exert a strong influence on the temperature sensitivity of soil respiration 1 1998
... 尽管底座面积不同, 但是入土较深的底座处理中土壤温度均显著高于入土较浅的处理, 而土壤温度对土壤呼吸具有促进作用.相反, 入土较深的底座处理中土壤含水量则显著低于较浅的处理, “高温干旱”使较深的底座处理中土壤呼吸强度降低.在本实验中, 底座处理通过直接改变土壤温度和土壤含水量来间接地影响土壤呼吸, 且主要是基于土壤含水量降低对土壤呼吸产生的抑制效应.许多研究表明, 土壤温度和土壤含水量是影响土壤呼吸重要的环境因子(Lloyd & Taylor, 1994; Davidson et al., 2000), 尤其是在干旱半干旱草原, 土壤水分有效性强烈影响生态系统土壤呼吸过程(Chen et al., 2009).研究表明, 无论是植物根系呼吸(Berry & Norris, 1949; Atkin et al., 2000), 还是土壤微生物呼吸(Mikan et al., 2002), 在土壤温度较低时, 土壤呼吸速率随着温度的升高而呈指数增加.但在土壤含水量较低的干旱期, 土壤温度升高反而抑制土壤CO2的释放(Zhou et al., 2006).以往研究发现, 土壤呼吸不同组分的温度敏感性不同, 根系周围土壤的异养呼吸比根系和根际自养呼吸对温度的敏感性更低(Boone et al., 1998), 本研究中, 无论底座面积大小, 入土较深的处理, 其土壤呼吸对土壤温度的敏感性更低(Q10值小).入土较深的处理, 其根系被切断的总量更多, 使土壤呼吸中自养呼吸部分降低, 促使异养呼吸占总呼吸的比例增加, 这可能是底座入土较深的处理中土壤呼吸对温度的敏感性更低的原因. ...
Biotic and abiotic factors controlling soil respiration rates in Picea abies stands. 1 2000
Grazing intensity alters soil respiration in an alpine meadow on the Tibetan Plateau 1 2004
... 同时, 本研究还发现底座面积的增加显著降低了表层土壤湿度.与面积较小的底座相比较, 在面积的较大底座内, 地上净初级生产力的下降可能导致蒸腾作用下降, 使土壤损失的水分减少, 同时地表接受太阳辐射增强, 可能使表层土壤温度显著升高, 导致表层土壤含水量下降, 因此底座内土壤含水量的变化取决于处理对蒸发和蒸腾影响的相对大小.实验结果显示, 面积较大的底座内土壤含水量较低, 表明底座面积较大的处理对蒸发作用的正效应大于对蒸腾作用的负效应, 最终导致较大的底座处理区域更加“高温干旱”.以往研究表明, 在干旱半干旱草原区域, 水分是草原生态系统土壤呼吸和初级生产力重要的限制因子(Bai et al., 2004; Yan et al., 2010), 高温干旱的环境, 抑制了土壤呼吸, 最终使底座面积较大的处理其土壤呼吸测定值较小(Cao et al., 2004). ...
Vertebrate herbivore-induced changes in plants and soils: Linkages to ecosystem functioning in a semi-arid steppe 1 2013
... 本研究在中国科学院内蒙古草原生态系统定位研究站(IMGERS, 43.63° N, 116.70° E)综合观测场内进行.该区域海拔高度为1 200 m, 年降水量为346 mm (1982-2014), 降水量年内和年际间变异较大, 60%-80%的降水发生在生长季的5-9月.年平均气温0.3 ℃, 其中1月份平均气温-21.6 ℃, 7月份平均气温19.0 ℃.本区域地带性土壤属暗栗钙土.植物群落的优势植物包括: 羊草、大针茅(Stipa grandis)、冰草(Agropyron cristatum)、糙隐子草(Cleistogenes squarrosa)等(Chen et al., 2013). ...
Soil acidification exerts a greater control on soil respiration than soil nitrogen availability in grasslands subjected to long-term nitrogen enrichment 2 2016
Dependence of carbon sequestration on the differential responses of ecosystem photosynthesis and respiration to rain pulses in a semiarid steppe 5 2009
... 综合考虑天气条件, 同时避免降雨脉冲对测定结果的影响(Chen et al., 2009), 本实验的土壤呼吸测定分别在2013年6月24日、7月6日、7月22日、8月14日、9月15日和10月16日的9:00-11:00进行.与底座面积配套, 土壤呼吸测定时将气室放置底座之上.实验时采用两个不透明方形气室(15 cm × 15 cm × 15 cm 和30 cm × 30 cm × 30 cm), 通过与红外气体分析仪(LI-840, LI-COR, Lincoln, USA)以及一个气泵相连的装置来进行测定.为增加测定结果的可靠性, 气室与底座间通过安装密封圈防止漏气, 同时在遮光气室中安装风扇混合气室中的气体, 使红外气体分析仪测得的CO2浓度是混合均匀的.测定的CO2浓度数据通过LI-840软件记录于电脑文档中.LI-840软件数据记录频率设置为1 s, 为保证CO2和H2O浓度在连续测量过程中有60 s的有效数据, 每个小区的测量时间设置为100 s.根据Jasoni等(2005)的分析方法, 本实验用LI-840装置获取的数据计算土壤呼吸.同时, 为保证数据的匹配, 在测定土壤CO2释放速率的同时测定底座内0-10 cm的土壤温度和土壤含水量.土壤温度的测定使用温度测定探针进行, 土壤含水量使用便携式土壤含水量测定仪TDR-200 (Spectrum Technologies, Plainfield, USA)测定. ...
... 依据以往研究, 本实验在测定土壤呼吸前一天, 将底座内所有绿色植物齐地收获, 目的是消除植物地上部分光合作用和呼吸作用对土壤呼吸的影响, 同时可以降低因植物损伤产生的扰动效应(Chen et al., 2009; Xia et al., 2009; Yan et al., 2010).6次收获的植物地上部分均带回实验室(为减小生物量移除对结果的影响, 在收获处理后地上净初级生产力时, 从附近剪取相等质量的植物放于底座内), 置于65 ℃的烘箱中烘干至恒质量, 然后混合称量.第一次收获的地上干物质为pre-AGP, 其他5次收获的干物质总和为处理后底座中的post-ANPP. ...
Effects of soil water content on soil respiration in forests and cattle pastures of eastern Amazonia 2 2000
... 在极端降雨事件发生时, 底座内可能出现水涝现象(Heinemeyer et al., 2011), 进而使底座内的土壤温度和含水量发生变化.尤其是在干旱和半干旱地区, 水分是生态系统结构与功能的重要限制因子(Davidson et al., 2000; Bai et al., 2008), 土壤水分有效性的增加, 可促使底座区域的土壤呼吸增强.同时, 底座阻隔了底座内外植物间物质的横向运输, 底座内外土壤蒸发散的差异会影响表层土壤温度(Reichstein et al., 2003), 而土壤呼吸的变化受土壤温度(Lloyd & Taylor, 1994; Zhou et al., 2007)、土壤含水量(Yan et al., 2010)和养分供应(Bahn et al., 2008)的共同影响.因此, 底座设置方面的不足, 可能直接或者间接地影响以上因素, 最终对土壤呼吸的测定结果产生影响. ...
... 尽管底座面积不同, 但是入土较深的底座处理中土壤温度均显著高于入土较浅的处理, 而土壤温度对土壤呼吸具有促进作用.相反, 入土较深的底座处理中土壤含水量则显著低于较浅的处理, “高温干旱”使较深的底座处理中土壤呼吸强度降低.在本实验中, 底座处理通过直接改变土壤温度和土壤含水量来间接地影响土壤呼吸, 且主要是基于土壤含水量降低对土壤呼吸产生的抑制效应.许多研究表明, 土壤温度和土壤含水量是影响土壤呼吸重要的环境因子(Lloyd & Taylor, 1994; Davidson et al., 2000), 尤其是在干旱半干旱草原, 土壤水分有效性强烈影响生态系统土壤呼吸过程(Chen et al., 2009).研究表明, 无论是植物根系呼吸(Berry & Norris, 1949; Atkin et al., 2000), 还是土壤微生物呼吸(Mikan et al., 2002), 在土壤温度较低时, 土壤呼吸速率随着温度的升高而呈指数增加.但在土壤含水量较低的干旱期, 土壤温度升高反而抑制土壤CO2的释放(Zhou et al., 2006).以往研究发现, 土壤呼吸不同组分的温度敏感性不同, 根系周围土壤的异养呼吸比根系和根际自养呼吸对温度的敏感性更低(Boone et al., 1998), 本研究中, 无论底座面积大小, 入土较深的处理, 其土壤呼吸对土壤温度的敏感性更低(Q10值小).入土较深的处理, 其根系被切断的总量更多, 使土壤呼吸中自养呼吸部分降低, 促使异养呼吸占总呼吸的比例增加, 这可能是底座入土较深的处理中土壤呼吸对温度的敏感性更低的原因. ...
Measuring forest floor CO2 fluxes in a Douglas-fir forest 1 2002
Soil respiration: Implications of the plant-soil continuum and respiration chamber collar-insertion depth on measurement and modelling of soil CO2 efflux rates in three ecosystems 8 2011
... ), 如草原生态系统生长季土壤呼吸测定结果被低估15%, 特别是在生长季旺期甚至被低估30%-50% (Heinemeyer et al., 2011).该研究表明, 底座入土越深, 土壤呼吸测定值就越小.因为底座切断植物根系和菌根真菌的菌丝, 使土壤呼吸中自养呼吸部分降低.底座入土越深, 切断根系和菌丝的量就越多, 土壤呼吸下降程度就越明显.但在湿润区域的研究发现, 增加底座入土深度, 生态系统的土壤呼吸并没有发生显著改变, 可能的原因是该区域土壤表层拥有较厚的苔藓层, 底座入土深度增加, 可能没有使根系的切断量随之增加(Drewitt et al., 2002).还有研究认为, 根系的“伤口”可能分泌大量易分解有机质, 这不仅可以增加土壤微生物呼吸, 还能通过“激发效应”加速土壤有机质的分解, 促进土壤呼吸(Kuzyakov, 2010; Guenet et al., 2012; Hopkins et al., 2013).因此在底座安置的初期, 土壤可能释放更多的CO2. ...
... 在极端降雨事件发生时, 底座内可能出现水涝现象(Heinemeyer et al., 2011), 进而使底座内的土壤温度和含水量发生变化.尤其是在干旱和半干旱地区, 水分是生态系统结构与功能的重要限制因子(Davidson et al., 2000; Bai et al., 2008), 土壤水分有效性的增加, 可促使底座区域的土壤呼吸增强.同时, 底座阻隔了底座内外植物间物质的横向运输, 底座内外土壤蒸发散的差异会影响表层土壤温度(Reichstein et al., 2003), 而土壤呼吸的变化受土壤温度(Lloyd & Taylor, 1994; Zhou et al., 2007)、土壤含水量(Yan et al., 2010)和养分供应(Bahn et al., 2008)的共同影响.因此, 底座设置方面的不足, 可能直接或者间接地影响以上因素, 最终对土壤呼吸的测定结果产生影响. ...
... 草原生态系统中, 测定土壤呼吸的底座入土深度一般为2-5 cm, 底座直径一般为10-30 cm (Subke et al., 2006; Heinemeyer et al., 2011).本实验于2013年的5月底至10月中旬进行.实验包括底座入土深度和底座面积2个因子, 其中底座入土深度包括2个水平: 2 cm (D2)和5 cm (D5), 底座入土深度不包括凋落物层厚度; 采用方形底座, 底座面积包括2个水平: 15 cm × 15 cm (S15)和30 cm × 30 cm (S30), 所有底座高度均为8 cm, 但由于入土深度不同, 因而露出地面的高度存在差异.实验采用随机区组设计, 4个处理, 每个处理6次重复, 共计24个1 m × 1 m的实验小区, 小区之间采用间距为2 m的过道进行隔离.底座的安装时间为2013年6月初, 在每个实验小区内, 将底座设置在小区的中央, 为减少底座安装和拆卸对土壤的干扰, 底座始终安置于样地直至实验结束(Xia et al., 2009).土壤呼吸的测定从6月下旬开始至10月中旬结束, 每月月初对每个小区进行留茬5 cm的模拟放牧处理, 所收获的地上生物量(不包括底座内的部分)从系统中移除, 底座中的生物量单独进行测定. ...
Analysis of soil respiration under different grazing management patterns in the alpine meadow-steppe of the Qinghai-Tibet Plateau 1 2015
... 2013年6月1日至10月31日期间, 实验区内自然降水共计46次, 累积降水量为244.2 mm.其中, <2 mm的降水24次, 2-5 mm的降水6次, 5-10 mm的降水8次, >10 mm的降水8次, 以小降水事件为主, 降水时间变异大, 主要发生在6-7月, 两个月降水量为162.1 mm, 占整个实验期间降水总量的66.4%.6-7月降水次数多达30次, >2 mm的有效降水多达16次, >10 mm的降水事件6次;8月1-15日期间没有降水, 但在8月16-27日发生了本年度极端降雨事件, 降水量分别为24.1 mm和39.3 mm; 9月和10月属于干旱期, 有效降水只有3次, 且降水量均<5 mm (图1C).与降水密切相关的土壤含水量的季节变化呈现“单峰”形, 7月22日土壤含水量达到峰值, 但仍然没有达到草原生态系统土壤含水量的阈值(30%) (Li et al., 2015)(图1C). ...
On the temperature dependence of soil respiration 2 1994
... 在极端降雨事件发生时, 底座内可能出现水涝现象(Heinemeyer et al., 2011), 进而使底座内的土壤温度和含水量发生变化.尤其是在干旱和半干旱地区, 水分是生态系统结构与功能的重要限制因子(Davidson et al., 2000; Bai et al., 2008), 土壤水分有效性的增加, 可促使底座区域的土壤呼吸增强.同时, 底座阻隔了底座内外植物间物质的横向运输, 底座内外土壤蒸发散的差异会影响表层土壤温度(Reichstein et al., 2003), 而土壤呼吸的变化受土壤温度(Lloyd & Taylor, 1994; Zhou et al., 2007)、土壤含水量(Yan et al., 2010)和养分供应(Bahn et al., 2008)的共同影响.因此, 底座设置方面的不足, 可能直接或者间接地影响以上因素, 最终对土壤呼吸的测定结果产生影响. ...
... 尽管底座面积不同, 但是入土较深的底座处理中土壤温度均显著高于入土较浅的处理, 而土壤温度对土壤呼吸具有促进作用.相反, 入土较深的底座处理中土壤含水量则显著低于较浅的处理, “高温干旱”使较深的底座处理中土壤呼吸强度降低.在本实验中, 底座处理通过直接改变土壤温度和土壤含水量来间接地影响土壤呼吸, 且主要是基于土壤含水量降低对土壤呼吸产生的抑制效应.许多研究表明, 土壤温度和土壤含水量是影响土壤呼吸重要的环境因子(Lloyd & Taylor, 1994; Davidson et al., 2000), 尤其是在干旱半干旱草原, 土壤水分有效性强烈影响生态系统土壤呼吸过程(Chen et al., 2009).研究表明, 无论是植物根系呼吸(Berry & Norris, 1949; Atkin et al., 2000), 还是土壤微生物呼吸(Mikan et al., 2002), 在土壤温度较低时, 土壤呼吸速率随着温度的升高而呈指数增加.但在土壤含水量较低的干旱期, 土壤温度升高反而抑制土壤CO2的释放(Zhou et al., 2006).以往研究发现, 土壤呼吸不同组分的温度敏感性不同, 根系周围土壤的异养呼吸比根系和根际自养呼吸对温度的敏感性更低(Boone et al., 1998), 本研究中, 无论底座面积大小, 入土较深的处理, 其土壤呼吸对土壤温度的敏感性更低(Q10值小).入土较深的处理, 其根系被切断的总量更多, 使土壤呼吸中自养呼吸部分降低, 促使异养呼吸占总呼吸的比例增加, 这可能是底座入土较深的处理中土壤呼吸对温度的敏感性更低的原因. ...
Temperature controls of microbial respiration in arctic tundra soils above and below freezing 1 2002
... 尽管底座面积不同, 但是入土较深的底座处理中土壤温度均显著高于入土较浅的处理, 而土壤温度对土壤呼吸具有促进作用.相反, 入土较深的底座处理中土壤含水量则显著低于较浅的处理, “高温干旱”使较深的底座处理中土壤呼吸强度降低.在本实验中, 底座处理通过直接改变土壤温度和土壤含水量来间接地影响土壤呼吸, 且主要是基于土壤含水量降低对土壤呼吸产生的抑制效应.许多研究表明, 土壤温度和土壤含水量是影响土壤呼吸重要的环境因子(Lloyd & Taylor, 1994; Davidson et al., 2000), 尤其是在干旱半干旱草原, 土壤水分有效性强烈影响生态系统土壤呼吸过程(Chen et al., 2009).研究表明, 无论是植物根系呼吸(Berry & Norris, 1949; Atkin et al., 2000), 还是土壤微生物呼吸(Mikan et al., 2002), 在土壤温度较低时, 土壤呼吸速率随着温度的升高而呈指数增加.但在土壤含水量较低的干旱期, 土壤温度升高反而抑制土壤CO2的释放(Zhou et al., 2006).以往研究发现, 土壤呼吸不同组分的温度敏感性不同, 根系周围土壤的异养呼吸比根系和根际自养呼吸对温度的敏感性更低(Boone et al., 1998), 本研究中, 无论底座面积大小, 入土较深的处理, 其土壤呼吸对土壤温度的敏感性更低(Q10值小).入土较深的处理, 其根系被切断的总量更多, 使土壤呼吸中自养呼吸部分降低, 促使异养呼吸占总呼吸的比例增加, 这可能是底座入土较深的处理中土壤呼吸对温度的敏感性更低的原因. ...
Biases of chamber methods for measuring soil CO2 efflux demonstrated with a laboratory apparatus 1 1994
Modeling temporal and large-scale spatial variability of soil respiration from soil water availability, temperature and vegetation productivity indices 1 2003
... 在极端降雨事件发生时, 底座内可能出现水涝现象(Heinemeyer et al., 2011), 进而使底座内的土壤温度和含水量发生变化.尤其是在干旱和半干旱地区, 水分是生态系统结构与功能的重要限制因子(Davidson et al., 2000; Bai et al., 2008), 土壤水分有效性的增加, 可促使底座区域的土壤呼吸增强.同时, 底座阻隔了底座内外植物间物质的横向运输, 底座内外土壤蒸发散的差异会影响表层土壤温度(Reichstein et al., 2003), 而土壤呼吸的变化受土壤温度(Lloyd & Taylor, 1994; Zhou et al., 2007)、土壤含水量(Yan et al., 2010)和养分供应(Bahn et al., 2008)的共同影响.因此, 底座设置方面的不足, 可能直接或者间接地影响以上因素, 最终对土壤呼吸的测定结果产生影响. ...
... 草原生态系统中, 测定土壤呼吸的底座入土深度一般为2-5 cm, 底座直径一般为10-30 cm (Subke et al., 2006; Heinemeyer et al., 2011).本实验于2013年的5月底至10月中旬进行.实验包括底座入土深度和底座面积2个因子, 其中底座入土深度包括2个水平: 2 cm (D2)和5 cm (D5), 底座入土深度不包括凋落物层厚度; 采用方形底座, 底座面积包括2个水平: 15 cm × 15 cm (S15)和30 cm × 30 cm (S30), 所有底座高度均为8 cm, 但由于入土深度不同, 因而露出地面的高度存在差异.实验采用随机区组设计, 4个处理, 每个处理6次重复, 共计24个1 m × 1 m的实验小区, 小区之间采用间距为2 m的过道进行隔离.底座的安装时间为2013年6月初, 在每个实验小区内, 将底座设置在小区的中央, 为减少底座安装和拆卸对土壤的干扰, 底座始终安置于样地直至实验结束(Xia et al., 2009).土壤呼吸的测定从6月下旬开始至10月中旬结束, 每月月初对每个小区进行留茬5 cm的模拟放牧处理, 所收获的地上生物量(不包括底座内的部分)从系统中移除, 底座中的生物量单独进行测定. ...
Clonal integration supports the expansion from terrestrial to aquatic environments of the amphibious stoloniferous herb Alternanthera philoxeroides. 1 2009
Response of ecosystem carbon exchange to warming and nitrogen addition during two hydrologically contrasting growing seasons in a temperate steppe 2 2009
... 草原生态系统中, 测定土壤呼吸的底座入土深度一般为2-5 cm, 底座直径一般为10-30 cm (Subke et al., 2006; Heinemeyer et al., 2011).本实验于2013年的5月底至10月中旬进行.实验包括底座入土深度和底座面积2个因子, 其中底座入土深度包括2个水平: 2 cm (D2)和5 cm (D5), 底座入土深度不包括凋落物层厚度; 采用方形底座, 底座面积包括2个水平: 15 cm × 15 cm (S15)和30 cm × 30 cm (S30), 所有底座高度均为8 cm, 但由于入土深度不同, 因而露出地面的高度存在差异.实验采用随机区组设计, 4个处理, 每个处理6次重复, 共计24个1 m × 1 m的实验小区, 小区之间采用间距为2 m的过道进行隔离.底座的安装时间为2013年6月初, 在每个实验小区内, 将底座设置在小区的中央, 为减少底座安装和拆卸对土壤的干扰, 底座始终安置于样地直至实验结束(Xia et al., 2009).土壤呼吸的测定从6月下旬开始至10月中旬结束, 每月月初对每个小区进行留茬5 cm的模拟放牧处理, 所收获的地上生物量(不包括底座内的部分)从系统中移除, 底座中的生物量单独进行测定. ...
... 依据以往研究, 本实验在测定土壤呼吸前一天, 将底座内所有绿色植物齐地收获, 目的是消除植物地上部分光合作用和呼吸作用对土壤呼吸的影响, 同时可以降低因植物损伤产生的扰动效应(Chen et al., 2009; Xia et al., 2009; Yan et al., 2010).6次收获的植物地上部分均带回实验室(为减小生物量移除对结果的影响, 在收获处理后地上净初级生产力时, 从附近剪取相等质量的植物放于底座内), 置于65 ℃的烘箱中烘干至恒质量, 然后混合称量.第一次收获的地上干物质为pre-AGP, 其他5次收获的干物质总和为处理后底座中的post-ANPP. ...
Differential responses of auto- and heterotrophic soil respiration to water and nitrogen addition in a semiarid temperate steppe 4 2010
... 在极端降雨事件发生时, 底座内可能出现水涝现象(Heinemeyer et al., 2011), 进而使底座内的土壤温度和含水量发生变化.尤其是在干旱和半干旱地区, 水分是生态系统结构与功能的重要限制因子(Davidson et al., 2000; Bai et al., 2008), 土壤水分有效性的增加, 可促使底座区域的土壤呼吸增强.同时, 底座阻隔了底座内外植物间物质的横向运输, 底座内外土壤蒸发散的差异会影响表层土壤温度(Reichstein et al., 2003), 而土壤呼吸的变化受土壤温度(Lloyd & Taylor, 1994; Zhou et al., 2007)、土壤含水量(Yan et al., 2010)和养分供应(Bahn et al., 2008)的共同影响.因此, 底座设置方面的不足, 可能直接或者间接地影响以上因素, 最终对土壤呼吸的测定结果产生影响. ...
... 依据以往研究, 本实验在测定土壤呼吸前一天, 将底座内所有绿色植物齐地收获, 目的是消除植物地上部分光合作用和呼吸作用对土壤呼吸的影响, 同时可以降低因植物损伤产生的扰动效应(Chen et al., 2009; Xia et al., 2009; Yan et al., 2010).6次收获的植物地上部分均带回实验室(为减小生物量移除对结果的影响, 在收获处理后地上净初级生产力时, 从附近剪取相等质量的植物放于底座内), 置于65 ℃的烘箱中烘干至恒质量, 然后混合称量.第一次收获的地上干物质为pre-AGP, 其他5次收获的干物质总和为处理后底座中的post-ANPP. ...
Main and interactive effects of warming, clipping, and doubled precipitation on soil CO2 efflux in a grassland ecosystem 1 2006
... 尽管底座面积不同, 但是入土较深的底座处理中土壤温度均显著高于入土较浅的处理, 而土壤温度对土壤呼吸具有促进作用.相反, 入土较深的底座处理中土壤含水量则显著低于较浅的处理, “高温干旱”使较深的底座处理中土壤呼吸强度降低.在本实验中, 底座处理通过直接改变土壤温度和土壤含水量来间接地影响土壤呼吸, 且主要是基于土壤含水量降低对土壤呼吸产生的抑制效应.许多研究表明, 土壤温度和土壤含水量是影响土壤呼吸重要的环境因子(Lloyd & Taylor, 1994; Davidson et al., 2000), 尤其是在干旱半干旱草原, 土壤水分有效性强烈影响生态系统土壤呼吸过程(Chen et al., 2009).研究表明, 无论是植物根系呼吸(Berry & Norris, 1949; Atkin et al., 2000), 还是土壤微生物呼吸(Mikan et al., 2002), 在土壤温度较低时, 土壤呼吸速率随着温度的升高而呈指数增加.但在土壤含水量较低的干旱期, 土壤温度升高反而抑制土壤CO2的释放(Zhou et al., 2006).以往研究发现, 土壤呼吸不同组分的温度敏感性不同, 根系周围土壤的异养呼吸比根系和根际自养呼吸对温度的敏感性更低(Boone et al., 1998), 本研究中, 无论底座面积大小, 入土较深的处理, 其土壤呼吸对土壤温度的敏感性更低(Q10值小).入土较深的处理, 其根系被切断的总量更多, 使土壤呼吸中自养呼吸部分降低, 促使异养呼吸占总呼吸的比例增加, 这可能是底座入土较深的处理中土壤呼吸对温度的敏感性更低的原因. ...
Source components and interannual variability of soil CO2 efflux under experimental warming and clipping in a grassland ecosystem 1 2007
... 在极端降雨事件发生时, 底座内可能出现水涝现象(Heinemeyer et al., 2011), 进而使底座内的土壤温度和含水量发生变化.尤其是在干旱和半干旱地区, 水分是生态系统结构与功能的重要限制因子(Davidson et al., 2000; Bai et al., 2008), 土壤水分有效性的增加, 可促使底座区域的土壤呼吸增强.同时, 底座阻隔了底座内外植物间物质的横向运输, 底座内外土壤蒸发散的差异会影响表层土壤温度(Reichstein et al., 2003), 而土壤呼吸的变化受土壤温度(Lloyd & Taylor, 1994; Zhou et al., 2007)、土壤含水量(Yan et al., 2010)和养分供应(Bahn et al., 2008)的共同影响.因此, 底座设置方面的不足, 可能直接或者间接地影响以上因素, 最终对土壤呼吸的测定结果产生影响. ...
 ,1,**
,1,**
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT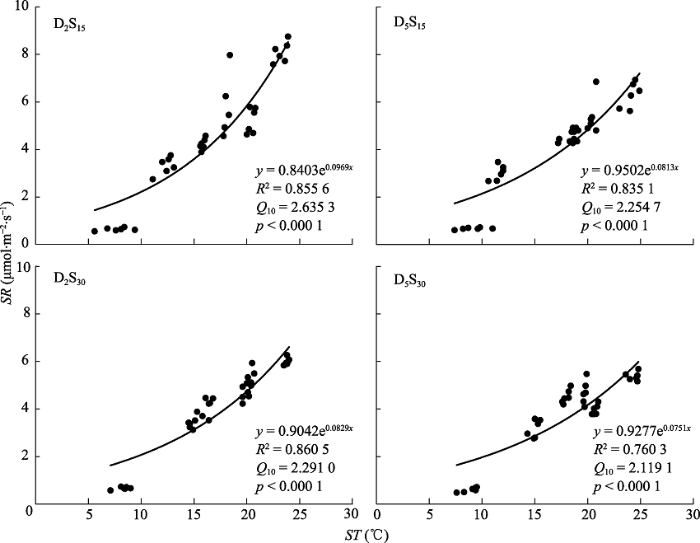 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT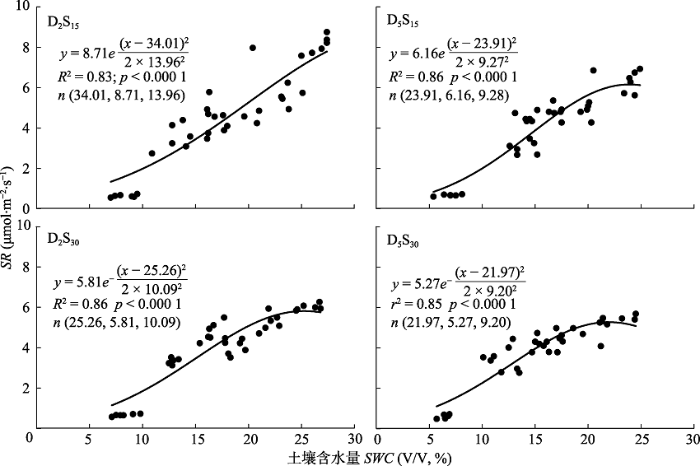 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT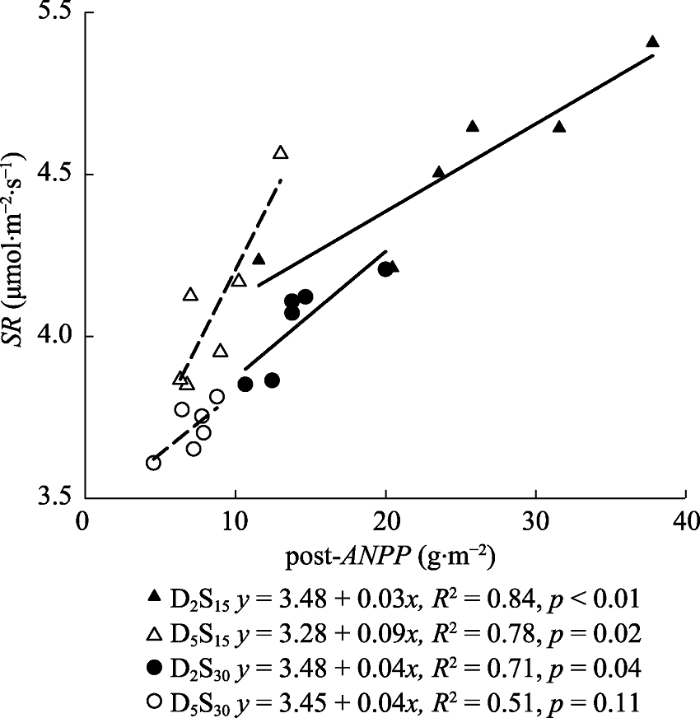 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT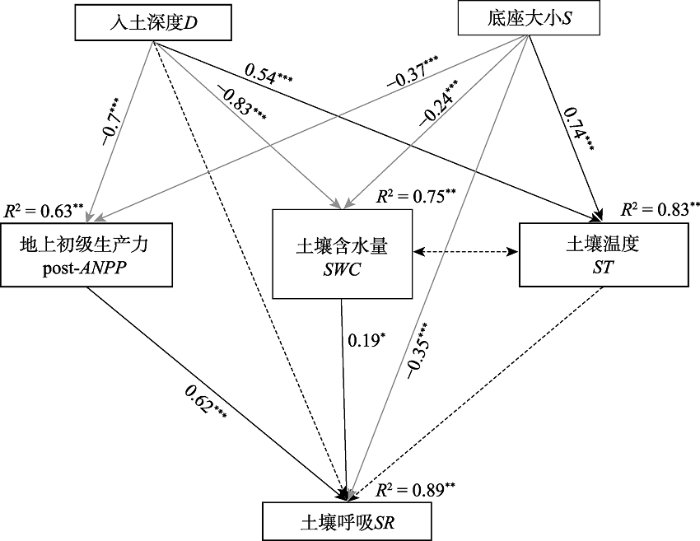 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}