 ,, 刘润红, 荣春艳, 常斌, 姜勇
,, 刘润红, 荣春艳, 常斌, 姜勇 ,*广西师范大学珍稀濒危动植物生态与环境保护教育部重点实验室/广西师范大学生命科学学院, 广西桂林 541006
,*广西师范大学珍稀濒危动植物生态与环境保护教育部重点实验室/广西师范大学生命科学学院, 广西桂林 541006Variation and correlation of plant functional traits in the riparian zone of the Lijiang River, Guilin, Southwest China
LIANG Shi-Chu,, LIU Run-Hong, RONG Chun-Yan, CHANG Bin, JIANG Yong,*Key Laboratory of Ecology of Rare and Endangered Species and Environmental Protection, Ministry of Education, Guangxi Normal University/College of Life Science, Guangxi Normal University, Guilin, Guangxi 541006, China通讯作者:
编委: 储诚进
责任编辑: 李敏
收稿日期:2018-05-20接受日期:2018-12-08网络出版日期:2019-01-20
| 基金资助: |
Received:2018-05-20Accepted:2018-12-08Online:2019-01-20
| Fund supported: |

摘要
关键词:
Abstract
Methods We investigated species composition of riparian plant communities in 36 plots along the longitudinal gradient (represented by upstream, midstream, downstream) of the Lijiang River, Guilin, Southwest China. We measured three functional traits for 42 woody plant species: leaf area (LA), specific leaf area (SLA), and wood density (WD). For each plant community, we calculated 1) species abundance-weighted mean community trait values, and 2) species-level mean trait values. For each of these calculations, we used trait-gradient analysis to partition the three traits into alpha and beta components. We then conducted Pearson correlations to analyze the relationships among the three traits along the longitudinal gradient. Finally we tested the strength of environmental filtering using a null model that generates randomly assembled communities with species richness given by observed values.
Important findings The species abundance-weighted mean community value of LA was lowest in the midstream communities, which was significantly different from that in the downstream communities. The mean community value of WD for midstream and upstream communities was significantly higher than that for downstream communities. Mean community value of SLA was significantly different among the three reaches. The beta components of the three functional traits significantly differed among the three reaches and had observed values that are, on average, lower than simulated values. However, alpha components for all three traits were not significantly different among the three reaches and had consistently lower variation than beta components. This implies that the variation in the mean community trait value across plots was greater than trait variation between species within plots. The observed and simulated values of the alpha components for both LA and SLA were weakly correlated with each other within and among communities, which suggests that there are independent axes of differentiation among coexisting species. On the other hand, comparisons between observed and simulated values indicated that significantly negative correlations between SLA and WD were largely the result of strong environmental filters. Finally, these results imply that variation of plant functional traits is greater among communities than within communities.
Keywords:
PDF (3109KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
梁士楚, 刘润红, 荣春艳, 常斌, 姜勇. 漓江河岸带植物功能性状变异与关联. 植物生态学报[J], 2019, 43(1): 16-26 doi:10.17521/cjpe.2018.0119
LIANG Shi-Chu.
1 材料和方法
1.1 研究区概况
漓江处于广西壮族自治区东北部, 位于珠江流域西江水系一级支流——桂江的上游河段, 地理坐标为109.75°-110.67° E, 24.30°-25.68° N。根据降水、地形、径流等特征, 可将漓江划分为上游、中游、下游3个河段, 其中桂林水文站至源头为上游, 桂林水文站至阳朔水文站为中游, 阳朔水文站至恭城河口为下游(刘金荣等, 2003)。漓江流域属中亚热带湿润季风气候区, 全年光照充足, 气候温和, 四季分明, 雨量充沛, 雨热基本同期, 平均气温17.8-19.1℃, 年降水量1 814-1 941 mm, 年蒸发量1 377-1 857 mm, 无霜期长达300余天。漓江属于雨源型山区河流, 径流为降雨所形成, 年径流量比较丰富, 但年内和年际分布极不均匀, 汛期为3-8月, 径流占年径流量的近80%, 其中5-6月占年径流量的40%, 枯水期为9月至翌年2月, 最枯月1月径流量仅占年径流量的2% (段文军等, 2014)。漓江河岸带土壤类型以山地黄壤为主, 经过江水的持续冲刷和沉积, 具有石砾含量高、土层浅薄、土层厚度分布不均等特点(李青山等, 2014)。漓江河岸带常见乔木有枫杨(Pterocarya stenoptera)、乌桕(Sapium sebiferum)、朴树(Celtis sinensis)、阴香(Cinnamomum burmannii)、樟(Cinnamomum camphora)等; 主要灌木有石榕树(Ficus abelii)、牡荆(Vitex negundo var. cannabifolia)、白饭树(Flueggea virosa)、萝芙木(Rauvolfia verticillata)、细叶水团花(Adina rubella); 主要草本有狗牙根(Cynodon dactylon)、辣蓼(Polygonum hydropiper)、喜旱莲子草(Alternanthera philoxeroides)、竹叶草(Oplismenus compositus)、麦冬(Ophiopogon japonicus)、红花酢浆草(Oxalis corymbosa)、酢浆草(Oxalis corniculata)、葎草(Humulus scandens)、皱叶狗尾草(Setaria plicata)等(信忠保等, 2014; 李扬等, 2015)。1.2 群落学调查
经过现场踏查, 于2016年9-10月和2017年7-9月, 在漓江河岸带上游、中游、下游各建立了12个大小为20 m × 20 m的样方, 合计36个样方, 各样方的基本情况如表1所示。将每个样方划分成4个10 m × 10 m的小样方, 对各样方进行常规的群落学调查, 内容主要包括: 对乔木层植株进行每木检尺, 记录种名、胸径等指标; 灌木层植株记录种名、高度、基径等指标。同时, 记录各样方的土壤类型、海拔、距河距离、砾石裸露率、砾石直径、干扰程度等生境特征。土壤采样以5 m × 5 m小样方为基本单位, 采用“梅花五点法”, 在每个5 m × 5 m小样方的4个顶点和中心位置, 清除表层枯落物和腐殖质层后, 用直径10 cm的土钻钻取0-20 cm土层土样, 运用四分法混合均匀后、放置于室内阴凉通风处自然干燥。待风干后, 剔除其中的根系、石块、钙核及动植物残体等杂物后倒入研钵中研细, 过不同孔径(0.15、0.25、1.4 mm)的土壤筛, 对预处理后的土样进行土壤pH值、有机质含量、全氮含量、速效氮含量、全磷含量、速效磷含量、全钾含量、速效钾含量8个土壤化学性质指标的测定, 测定方法参照《土壤农业化学常规分析方法》(中国土壤学会农业化学专业委员会, 1983)。每个土壤样品重复测定3次后取其平均值作为本研究分析的数据。Table 1
表1
表1漓江河岸带不同河段植物群落样方基本概况
Table 1
| 河段 Reach | 样方数 Plot number | 海拔 Elevation (m) | 温度 Temperature (℃) | 降水量 Precipitation (mm) | 干扰强度 Disturbance intensity | 群落类型 Community type |
|---|---|---|---|---|---|---|
| 上游 Upstream | 3 | 154 | 24.7 | 1 941 | 轻度 Light | 枫杨-石榕树群落 Pterocarya stenoptera-Ficus abelii communities |
| 3 | 148 | 23.0 | 1 941 | 轻度 Light | 枫杨+朴树-萝芙木群落 Pterocarya stenoptera + Celtis sinensis-?Rauvolfia verticillata communities | |
| 6 | 144 | 23.7 | 1 941 | 轻度 Light | 枫杨+阴香-石榕树群落 Pterocarya stenoptera + Cinnamomum burmannii-Ficus abelii communities | |
| 中游 Midstream | 4 | 138 | 26.0 | 1 900 | 重度 High | 枫杨+乌桕-细叶水团花群落 Pterocarya stenoptera + Sapium sebiferum-Adina rubella communities |
| 5 | 134 | 25.0 | 1 900 | 重度 High | 阴香群落 Cinnamomum burmannii communities | |
| 3 | 104 | 24.0 | 1 900 | 重度 High | 枫杨-萝芙木群落Pterocarya stenoptera-?Rauvolfia verticillata communities | |
| 下游 Downstream | 8 | 111 | 30.1 | 1 900 | 中度 Middle | 乌桕+朴树-牡荆群落Sapium sebiferum + Celtis sinensis-Vitex negundo var. cannabifolia communities |
| 4 | 105 | 26.8 | 1 900 | 中度 Middle | 乌桕-木槿群落 Sapium sebiferum + Hibiscus syriacus communities |
新窗口打开|下载CSV
1.3 功能性状的选取及测定
选取叶面积(LA)、比叶面积(SLA)和木材密度(WD) 3个重要功能性状进行测量。其中, LA (cm2)反映叶片与外界环境接触面积的大小, 其会影响到植物体与外界环境的气体、能量等交换的平衡, 一般生在阴蔽、湿润、资源丰富地区的植物具有较大的叶片(Craine & Lee, 2003; Sun et al., 2006)。SLA是新鲜叶片面积与叶片干质量的比值(cm2·g-1), 反映植物投入单位质量的干物质所获得的捕光面积, 与潜在相对生长速率或最大光合速率正相关, 物种具有较大的SLA意味着生长速率较高, 养分利用效率较低, “防御性”资源投入较少, 叶片寿命相对较短(Cornelissen et al., 2003)。WD是枝条干质量与体积的比值(g·cm-3), 其与植物竖向生长的结构性支撑力、植物的寿命、树干的防御功能(如病虫害、可食性和物理性防御等)及碳储量有关, 可以反映植物的结构性投入和生长死亡的权衡关系(卜文圣等, 2013)。测定样方内所有胸径≥1 cm的木本植物植株个体的功能性状(包括乔、灌木和木质藤本)。在野外群落学调查过程中, 每个植物个体选取冠层顶端3片完全展开的健康成熟的阳生叶及3根长10 cm左右, 直径为1-2 cm的非当年生枝条放入密封袋内, 置于阴暗环境下并迅速带回实验室进行叶和茎功能性状测定。对于叶性状的测定, 利用叶面积仪(Yaxin-1241, 北京雅欣理仪科技有限公司, 北京)扫描每片叶片的叶面积, 然后将叶片样品编号后置于80 ℃烘箱中烘干至恒质量后, 称量并记录叶干质量。对于木材密度的测定, 本文利用测定的枝条密度代替木材密度, 将枝条削去表面的树皮后, 使用电子天平测定其鲜质量, 接着用量筒排水法测定枝条的体积, 然后将其编号并置于80 ℃烘箱中烘干至恒质量后, 测定枝条干质量。
1.4 统计与分析
1.4.1 冗余分析首先, 采用单因素方差分析对不同河段间的8个土壤化学性质(pH值、全氮含量、有机质含量、速效氮含量、全磷含量、速效磷含量、全钾含量、速效钾含量)指标进行统计检验, 根据分析结果排除部分不显著因子; 再结合土壤类型、海拔、距河距离、砾石裸露率、干扰程度、气候因子(温度和降水量)构建样地×环境变量矩阵。其次, 基于样地×多度矩阵、样地×性状矩阵、样地×环境变量矩阵, 利用R 3.4.3软件中Vegan包对矩阵进行冗余分析(RDA)。
1.4.2 层级方差分解
运用R 3.4.3 软件“lme”包中限制最大似然法(REML), 利用广义线性模型和“varcomp”函数对种内、种间、群落间、河段间和河段内与河段间植物功能性状的分异大小分别进行方差分解。
1.4.3 性状梯度分析
根据Ackerly和Cornwell (2007)提出的性状梯度分析法, 将植物群落(数量为n)内的物种(数量为s)按照该物种多度($\alpha_{ij}$)加权计算出其群落j的平均性状值(pj), 再根据群落平均性状值由小到大的顺序进行排序, 构建性状梯度, 然后利用该性状梯度将物种性状值(ti)累加降解为$\beta$和$\alpha$两个组分。其中, $\beta$组分($\beta_{i}$)为物种i在该性状梯度上所处的位置, 体现物种i在不同群落间的变异, 反映物种i对群落间环境变化的响应; 而$\alpha$组分($\alpha_i$)是某一物种i性状值(ti)与其所处群落平均性状值($\beta_{i}$)的差值, 体现了群落内物种i与其共存物种的性状值差异, 反映了对同一群落内种间交互作用等非生物环境适应策略的差异。性状梯度分析法主要计算公式如下
式中, pj为群落平均性状值, ti为物种平均性状值, aij为物种i在群落j中的多度或权重, tij为物种i在群落j中的性状值, n表示调查的群落总数, s表示调查出现的物种总数。本研究中n = 36, s = 42, 每个群落有3-25个物种。性状梯度分析之前对不满足正态分布的数据(除木材密度外)进行lg转换。利用单因素方差分析检验漓江河岸带不同河段间的功能性状指标(物种平均性状、α组分、β组分和群落平均性状)是否存在显著差异。如果存在显著差异, 则采用多重比较方法检测两两河段之间的功能性状值是否存在差异。然后分别对功能性状指标进行Pearson相关性分析, 相关性大小用Student’s t检验。
为了直观地解释性状梯度分析法, 现以常绿乔木阴香和落叶灌木牡荆的SLA为例来介绍α和β两个组分的生态学意义(图1)。根据上述公式计算得出阴香的β组分相对较小(lgβi = 2.37), 位于由群落平均SLA构建的性状梯度的下半部分, 说明阴香常见于平均SLA较低的群落中且阴香的SLA小于群落的平均值(lgαi = -0.28), 表明与其共存物种相比, 其SLA相对较低。相反, 牡荆的β组分相对较大(lgβi = 2.53), 位于性状梯度的上半部分, 说明牡荆常见于高SLA的群落中且牡荆的SLA大于群落的平均值(lgαi = 0.09), 说明与其共存物种相比, 其SLA相对较高。
图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1漓江河岸带植物阴香和牡荆物种平均比叶面积(lgSLAs)和群落平均比叶面积(lgSLAp)散点图。灰色圆点代表群落中的各个物种, 绿色实心圆点和橙色三角形分别表示阴香和牡荆, 在某pj处被矩形框起来的一列点表示群落j内所有共生物种。每个物种空心的图形点对应的横坐标代表该物种在样地中所处的平均位置(即该物种性状的β组分βi), 纵坐标是物种的平均性状值ti, 二者的差值即空心的图形点到y = x的距离(因为αi = ti - βi)就是物种功能性状的α组分αi, 代表群落中物种i平均比叶面积与共存物种平均比叶面积的差值, bi为物种i平均比叶面积(ti)对群落平均SLA (pj)的回归直线的斜率, 反映物种平均SLA沿群落平均性状梯度在种内的变化大小。
Fig. 1The scatterplot between species mean specific leaf area (i.e. lgSLAs, cm2·g-1) vs. plot mean specific leaf area (i.e. lgSLAp, cm2·g-1) between Cinnamomum burmannii and Vitex negundo in the riparian zone of the Lijiang River. Each grey point represents a species in a specific plot; the green solid points and the orange solid triangles represent Cinnamomum burmannii and Vitex negundo respectively, and a column of grey points in a black rectangle represent all the species within community. For each species, the abscissa values of the large open point show the mean position of occupied plots (i.e., the beta component of the species trait value, βi), while the ordinate values of the solid symbols are their mean species trait value (ti). The difference between βi and ti, or the distance from the y = x line is αi (because αi = ti - βi ). Regression line shows abundance-weighted least squares regression of species trait values relative to plot mean trait values, with slope bi. bi is the slope of each species’ s regression line of species mean trait values (ti) relative to plot mean trait values (pj), it reflects the intraspecific variation of the species mean specific leaf area along a gradient defined by community-level mean trait values.
1.4.4 零模型分析
采用零模型的方法计算随机期望下的不同性状$\alpha$/$\beta$组分协变强度和不同物种$\beta$组分的离散度。其中, $\alpha$/$\beta$组分协变强度的零模型是根据Ackerly和Cornwell (2007)自带的源程序进行计算; $\beta$组分的离散度零模型是基于Jung等(2010)提出的环境筛检验。根据实际观测建立一个河岸带不同河段间的植物物种库, 然后随机抽取与之不同河段间相同数目的物种, 进行不放回式999次随机抽样, 计算其功能性状的分布范围。与随机模拟群落相比, 如存在环境筛效应, 群落中的某些物种被过滤出, 导致落入植物功能性状值的范围变小。根据随机模拟获取不同河段间的功能性状β组分分布范围的次数, 计算其次数的5%和95%的置信区间, 若在置信区间之内, 表明不同河段间植物功能性状$\beta$组分与随机群落无显著差异; 若次数有5%小于或95%大于不同河段间的实际功能性状$\beta$组分分布范围, 表明不同河段间的实际植物功能性状显著大于或小于随机群落。
2 结果
2.1 不同河段间植物功能性状与环境因子的RDA排序
RDA分析结果如图2和表2所示: 上中下游河段被群落平均功能性状(LA, SLA和WD)显著分开, 同时展示了在每个梯度上环境因子对功能性状的显著影响。WD和SLA主要与RDA第一排序轴相关, 处于第一轴相对位置; LA主要与第二轴相关。LA主要受土壤有机质(SOM)含量、干扰强度(DI)、土壤pH值、土壤全氮(TN)含量、降水量(Pre)显著正影响; SLA主要受海拔(ELe)的显著正影响; WD主要受土壤有效氮(AN)含量、温度(Tem)、河段(Rea)、距离河岸距离(Dis)的显著正影响。图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2漓江河岸带植物群落水平功能性状与环境因子的冗余分析(RDA)排序图。AN, 有效氮含量; DI, 干扰强度; Dis, 距离河岸距离; Ele, 海拔; LA, 叶面积; pH, 土壤pH值; Pre, 降水量; Rea, 河段; SLA, 比叶面积; SOM, 有机质含量; Tem, 温度; TN, 全氮含量; WD, 木材密度。plot 1-12, 上游; plot 12-24, 中游; plot 25-36, 下游。
Fig. 2Redundancy analysis (RDA) ordination diagram showing the relationships between the three abundance weighted functional traits and 10 selected environmental factors of the riparian plant of the Lijiang River. AN, soil available nitrogen; DI, disturbance intensity; Dis, distance; Ele, elevation; LA, leaf area; pH, soil pH value; Pre, precipitation; Rea, reach; SLA, specific leaf area; SOM, soil organic matter content; Tem, temperature; TN, soil total nitrogen content; WD, wood density. plot 1-12, upstream; plot 12-24, midstream; plot 25-36, downstream.
Table 2
表2
表2漓江河岸带群落水平功能性状与环境因子冗余分析(RDA)中的前2轴环境变量解释方差和显著性检验
Table 2
| 环境因子 Environmental factor | RDA1 | RDA2 | R2 | p |
|---|---|---|---|---|
| 有机质 Soil organic matter (g·kg-1) | 0.40 | 0.91 | 0.34 | 0.002** |
| 全氮 Soil total nitrogen (g·kg-1) | -0.45 | 0.88 | 0.19 | 0.032* |
| 有效氮 Soil available nitrogen (mg·kg-1) | -0.74 | -0.66 | 0.57 | 0.001*** |
| pH | -0.26 | 0.96 | 0.21 | 0.015* |
| 干扰强度Disturbance intensity | -0.16 | 0.98 | 0.63 | 0.001*** |
| 距离河岸距离 Distance (m) | -0.85 | 0.51 | 0.29 | 0.004** |
| 降水量 Precipitation (mm) | -0.66 | 0.74 | 0.79 | 0.001*** |
| 温度 Temperature (℃) | -0.97 | 0.22 | 0.50 | 0.001*** |
| 海拔 Elevation (m) | 0.92 | -0.38 | 0.57 | 0.001*** |
| 河段 Reach | -0.90 | 0.41 | 0.88 | 0.001*** |
新窗口打开|下载CSV
2.2 不同河段植物功能性状的变化规律
统计分析如表3所示: 对于物种性状, WD是上游与下游差异性显著。对于β组分, LA、SLA和WD 3个性状在两两河段中均存在显著性差异, 而在α组分中3个性状在两两河段中均无显著性差异。对于群落性状, LA是中游和下游差异显著; SLA是两两河段中均差异性显著; WD是上游和中游显著大于下游。Table 3
表3
表3漓江不同河段功能性状参数值(平均值±标准偏差)
Table 3
| 河段 Reach | 功能性状 Functional trait | 性状参数 Functional trait parameter | |||
|---|---|---|---|---|---|
| 物种性状值 ti | β 组分 βi | α 组分 αi | 群落性状值 pj | ||
| 上游 Upstream | 叶面积 LA (cm2) | 1.30 ± 0.35a | 1.36 ± 0.08a | -0.06 ± 0.34a | 1.34 ± 0.14ab |
| 比叶面积 SLA (cm2·g-1) | 2.40 ± 0.13a | 2.40 ± 0.02a | -0.00 ± 0.13a | 2.39 ± 0.03a | |
| 木材密度 WD (g·cm-3) | 0.47 ± 0.10a | 0.44 ± 0.02a | 0.03 ± 0.10a | 0.44 ± 0.04a | |
| 中游 Midstream | 叶面积 LA (cm2) | 1.16 ± 0.32a | 1.24 ± 0.08b | -0.08 ± 0.27a | 1.26 ± 0.10a |
| 比叶面积 SLA (cm2·g-1) | 2.47 ± 0.21a | 2.47 ± 0.06b | 0.01 ± 0.21a | 2.46 ± 0.07b | |
| 木材密度 WD (g·cm-3) | 0.42 ± 0.12ab | 0.47 ± 0.01b | -0.04 ± 0.12a | 0.47 ± 0.02a | |
| 下游 Downstream | 叶面积 LA (cm2) | 1.24 ± 0.32a | 1.45 ± 0.05c | -0.20 ± 0.30a | 1.46 ± 0.07b |
| 比叶面积 SLA (cm2·g-1) | 2.48 ± 0.13a | 2.53 ± 0.02c | -0.06 ± 0.12a | 2.55 ± 0.03c | |
| 木材密度 WD (g·cm-3) | 0.36 ± 0.10b | 0.34 ± 0.01c | 0.02 ± 0.10a | 0.34 ± 0.02b | |
新窗口打开|下载CSV
2.3 样方平均性状、物种功能性状及其α组分、β组分的相关性
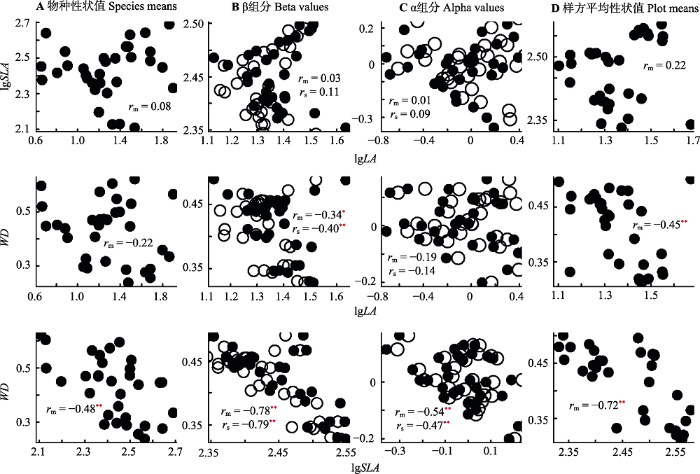
3个功能性状相关性如图3所示: 在物种水平上, SLA和WD之间呈显著负相关关系(图3A); 在样方尺度上, LA和SLA分别与WD呈极显著负相关关系(p < 0.01)(图3D); 在β组分上, LA和SLA分别与WD在真实观测值和随机模拟值均呈现极显著负相关关系(p < 0.01)(图3B); 在α组分上, SLA与WD在真实观测值和随机模拟值均呈现极显著负相关关系(p < 0.01) (图3C)。图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3漓江河岸带植物叶面积(LA)、比叶面积(SLA)和木材密度(WD)在物种性状值(A)、β组分(B)、α组分(C)、样方平均性状值(D)水平上的相关性散点图以及Pearson相关系数(r)。黑色实心点和rm分别代表实际测量数据和实测相关系数, 黑色圆圈和rs分别代表零模型随机模拟999次数据和模拟相关系数。*, p < 0.05; **, p < 0.01。
Fig. 3Scatterplots showing relationships between leaf area (LA), specific leaf area (SLA) and wood density (WD) for species trait values (A), beta components (B), alpha components (C), and plot mean trait values (D) of the riparian plant of the Lijiang River. The Pearson correlation coefficient (r) of these relationships are shown in each figure. Black solid dots and rm respectively represent observed values and observed correlation coefficient; black open circles and rs respectively represent random simulation values and simulation coefficient. *, p < 0.05; **, p < 0.01.
2.4 不同河段植物功能性状β组分的分布范围与零模型的比较分析
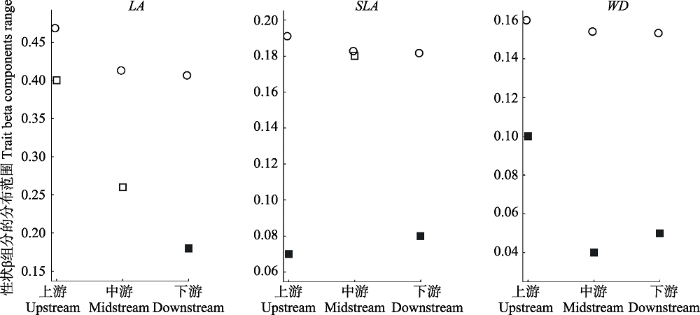
检测发现3个功能性状β组分实际观测值均小于零模型, 表明了功能性状β组分分布范围在3个不同河段受到环境的筛选作用且与零模型差异是否显著随河段的变化而变化。LA实际观测值在下游与零模型差异显著(p < 0.05); SLA实际观测值在上游和下游均与零模型差异显著(p < 0.05); WD实际观测值在上、中、下游均与零模型差异显著(p < 0.05)(图4)。图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4漓江不同河段植物功能性状β组分的分布范围与零模型比较。圆圈代表零模型预测值; 空心正方形代表实际观测值; 实心正方形代表实际观测值与零模型存在显著差异(p < 0.05)。
Fig. 4Difference on beta components ranges in the observed and simulated values of the three functional traits at the three reaches in Lijiang River. Circles and squares represent simulated and observed values respectively. Filled squares indicate that the observed values differ significantly from the simulated values.
2.5 不同水平上的植物功能性状的方差分异
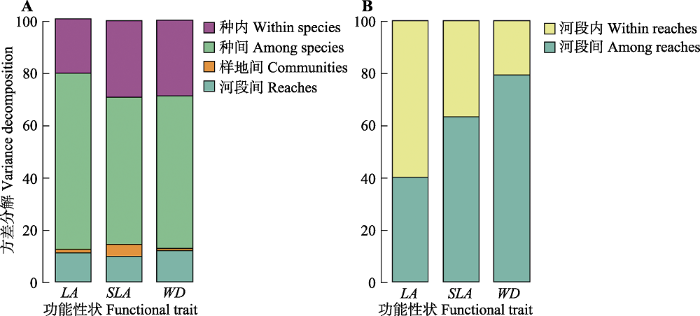
三个功能性状值(LA、SLA和WD)均体现群落水平性状变异最小, 其次是河段, 再次是种内, 种间性状变异最大, 即种间>种内>河段间>群落间(图5A)。SLA和WD在河段内和河段间的变异规律较一致, 即河段间>河段内; 但是LA相反, 表现为河段内>河段间(图5B)。图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5漓江河岸带植物种内、种间、群落间与河段间(A)及河段内和河段间(B)植物功能性状的方差分解。LA, 叶面积; SLA, 比叶面积; WD, 木材密度。
Fig. 5Partitioning of the variance in plant functional traits explained by four scales (i.e. within-specie, among-species, communities and reaches) (A) and by two scales (i.e. within and among reaches) (B) of the riparian plant of the Lijiang River. LA, leaf area; SLA, specific leaf area; WD, wood density.
3 讨论
3.1 群落平均性状α、β组分间沿河岸带不同河段变化规律
河岸带植物群落的生态特征受空间位置、干扰强度、气候、地形、土壤养分以及淹水的深度、持续时间和频率等环境因子的显著影响, 呈现随着环境梯度变化其梯度变化同样明显的特征(Vannote et al., 1980; 孙荣等, 2010)。本研究通过RDA排序结果展示了上中下游河段间不同环境因子组合(包括干扰强度强弱、海拔梯度、土壤养分含量等)对功能性状(LA、SLA和WD)产生了显著影响。结合图5B中的SLA和WD在河段间>河段内的性状变异规律进一步印证了不同河段间的环境梯度主导功能性状变异。图5B中LA功能性状分布变异呈现相反的变化规律, 即河段内>河段间, 但二者变异比例相差不大, 出现这样的变异规律是很出乎意料的, 同时也是值得深思的。这可能与采样误差及性状的选择有关。因此, 我们希望通过改进实验方案和尝试其他植物功能性状包括数值型性状和类别性状获取进一步验证。针对不同梯度下生境的高度异质性, 发现漓江河岸带群落平均性状LA、SLA和WD在上、中、下游河段之间均存在着显著的差异(表2; 图2), 表明植物群落为了适应河岸带不同河段的生境条件采取了不同的生态策略。例如, 群落平均SLA随着纵向梯度变化逐渐增大且差异性显著, 暗示了物种面对生境的变化而采取的适应性策略不同。即从上游物种光合速率低、叶寿命长且防御能力较强, 常绿物种比例较高逐渐转变为提高生长速率而降低养分利用效率的落叶灌木物种占主导。群落平均WD是中、上游高于下游, 表现出与SLA相反的性状梯度, 根据植物经济谱学说, 恶劣环境下选择慢收益型策略, 即物种具有SLA更小WD更高的性状特征。结合RDA排序结果和表1发现上流河段的环境资源比较匮乏, 这与上游生境特征相关, 即海拔和地势相对较高, 坡度较陡峭, 土壤层较薄, 砾石含量较高, 取样点距离河水距离较近, 导致淋溶作用强烈, 土壤中的营养成分流失严重, 尽管干扰最轻。下游相对地势平坦, 土壤层较厚, 土壤中养分含量随着地表水由上游流向下游地段积累, 土壤养分含量相对较高且温度适宜, 采样地点距离河水距离较远及干扰适度(中度干扰), 对比来讲下流河段生境较好。群落平均LA是中游小于下游且差异显著, 主要受干扰强度影响, 其次受土壤有机质含量、土壤pH值、土壤全氮含量、降水量等环境因子显著影响(图2; 表2)。实地调查发现中游距离市区最近, 旅游业盛行, 岸边过度垦荒, 人为干扰严重, 导致了中游物种种类偏少且叶面积大的物种比例较低, 结合图5A发现植物功能性状(LA、SLA和WD)均为种间变异贡献最大, 说明不同河段间的物种组成差异显著, 印证了面对干扰严重的恶劣生境LA采取了慢收益保守型策略。此外, 本研究发现3个功能性状的β组分在河岸带不同河段间的统计检验均差异性显著, 且河岸带不同河段的3个植物功能性状β组分的观测值均小于随机模拟的零模型分布(图3), 表明了功能性状β组分分布范围在河岸带不同河段受到环境筛作用的限制作用较为明显; 但α组分均差异性不显著(表3)且在河岸带不同河段3个功能性状α组分的分布范围均小于β组分, 说明在河岸带不同河段的群落构建过程中环境因素的影响要大于群落内种间的相互作用, 则性状梯度很大程度上取决于环境梯度。3.2 植物功能性状及其α、β组分间的相关性
植物在其漫长的历史进化和发展过程中, 受物理、生理等因素的综合作用和影响, 功能性状之间往往会呈现出一定的协变和权衡关系(Wright et al., 2007; Hu et al., 2015)。Fonseca等(2000)和Ackerly等(2002)发现LA与SLA在全球和区域尺度上均呈显著正相关关系。但是, Shipley (1995)和Grubb (1998)发现同一群落内植物的LA与SLA之间呈现不相关或负相关关系。这表明在地理尺度上LA与SLA均受某种选择因素(一般而言是气候因素)的限制, 但是在局域尺度上的适应可能会导致这两者从属于不同生态策略维度。本研究发现, 漓江河岸带木本植物LA与SLA在物种功能性状值、样方功能性状值及其β组分值相关性很弱(r分别为0.08、0.22和0.03), 尤其是α组分值几乎不存在相关关系(r = 0.01)。这一结果基本与习新强等(2011)对黔中喀斯特山区植物功能性状变异与关联的研究结果一致。SLA与LA在物种尺度没有显著相关性, 这主要是由αSLA与αLA之间没有显著相关关系造成的。此外, 群落水平和β组分二者均相关性不显著。Baraloto等(2010)分析热带雨林的叶和茎经济型谱发现, LA和SLA呈正交, 即两个性状属于不同的策略维度。由此可见, 在局域尺度上LA和SLA各自代表了植物在不同生态策略上的维度, 在本研究中得到了很好的佐证。WD与LA两性状在物种水平和β组分相关性存在显著差异(p<0.01), 但α组分相关性较弱且零模型的变化趋势与观测值变化一致。造成这一格局的原因可能是, 群落环境梯度导致优势物种从喜阳的落叶灌木物种(大LA, 小WD)逐渐变为耐阴的常绿乔木(小LA, 大WD), 反映在性状上就是这两个性状的β组分呈明显的负相关关系。而在群落内部, 不同的物种产生不同的性状组合来分化生态位(Cornwell et al., 2006; Kraft et al., 2008; Kunstler et al., 2016)形成大致的竞争均衡, 导致二者的α组分协变关系很低(图3C)。此外, 本研究中WD与SLA在各水平上均呈现出显著负相关关系且零模型检测与此变化一致, 暗示了植物功能性状SLA和WD对于环境筛选表现出较高的整体趋同适应性, 导致二者的α、β组分均具有很强的协变关系, 而不同物种β组分的离散度很大(表2)。这进一步反映了漓江上中下游的主导性环境梯度对于群落性状变异的强限制作用。
致谢 感谢2017年广西高校优秀教师出国项目、广西硕士研究生创新项目(YCSW2017079)和2017年度自治区级大学生创新创业训练计划项目(201710602236和201710602247)资助。

参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1111/ele.2007.10.issue-2URL [本文引用: 2]
DOI:10.1007/s004420100805URLPMID:28547053 [本文引用: 1]

We examined variation in leaf size and specific leaf area (SLA) in relation to the distribution of 22 chaparral shrub species on small-scale gradients of aspect and elevation. Potential incident solar radiation (insolation) was estimated from a geographic information system to quantify microclimate affinities of these species across north- and south-facing slopes. At the community level, leaf size and SLA both declined with increasing insolation, based on average trait values for the species found in plots along the gradient. However, leaf size and SLA were not significantly correlated across species, suggesting that these two traits are decoupled and associated with different aspects of performance along this environmental gradient. For individual species, SLA was negatively correlated with species distributions along the insolation gradient, and was significantly lower in evergreen versus deciduous species. Leaf size exhibited a negative but non-significant trend in relation to insolation distribution of individual species. At the community level, variance in leaf size increased with increasing insolation. For individual species, there was a greater range of leaf size on south-facing slopes, while there was an absence of small-leaved species on north-facing slopes. These results demonstrate that analyses of plant functional traits along environmental gradients based on community level averages may obscure important aspects of trait variation and distribution among the constituent species.
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/j.1461-0248.2010.01517.xURLPMID:20807232 [本文引用: 1]
Cross-species analyses of plant functional traits have shed light on factors contributing to differences in performance and distribution, but to date most studies have focused on either leaves or stems. We extend these tissue-specific analyses of functional strategy towards a whole-plant approach by integrating data on functional traits for 13 448 leaves and wood tissues from 4672 trees representing 668 species of Neotropical trees. Strong correlations amongst traits previously defined as the leaf economics spectrum reflect a tradeoff between investments in productive leaves with rapid turnover vs. costly physical leaf structure with a long revenue stream. A second axis of variation, the ‘stem economics spectrum’, defines a similar tradeoff at the stem level: dense wood vs. high wood water content and thick bark. Most importantly, these two axes are orthogonal, suggesting that tradeoffs operate independently at the leaf and at the stem levels. By simplifying the multivariate ecological strategies of tropical trees into positions along these two spectra, our results provide a basis to improve global vegetation models predicting responses of tropical forests to global change.
DOI:10.3724/SP.J.1003.2013.10012URL [本文引用: 1]
We explored how plant functional traits respond to environmental factors and examined the underlying mechanism driving the ecological strategies of plant species in a community. Experiments were conducted at four randomly selected tropical lowland rainforests at different successional stages:15-, 30- and 60-year-old secondary forests and one old-growth forest, each in the Bawangling Nature Reserve of Hainan Island. A total of 200 plots (20 m 20 m) were sampled. Individuals of tree and shrub species with diameter at breast height (DBH) 1 cm were identified and both their functional traits and the environmental factors in each plot were measured. ANOVA showed that canopy openness, level of soil nutrients, specific leaf area, and content of total organic carbon, nitrogen and phosphorus within the leaf decreased at the community level during the successional process. Meanwhile, leaf dry matter content, wood density and potential maximum height increased. However, soil water content and leaf potassium content, changed non-significantly. Multiple regression analysis demonstrated that environmental factors driving functional traits at the community level varied during the successional process. The key environmental factors were soil organic matter and pH value in the 15-year-old secondary forest; canopy openness and soil total phosphorus content in the 30-year-old secondary forest; available phosphorus and total potassium content of the soil in the 60-year-old secondary forest; and soil phosphorus content and organic matter content in the old-growth forest. Our results indicate that at different successional stages, the same functional traits respond to different environmental factors, enabling adaptation to specific environmental conditions.
DOI:10.3724/SP.J.1003.2013.10012URL [本文引用: 1]
We explored how plant functional traits respond to environmental factors and examined the underlying mechanism driving the ecological strategies of plant species in a community. Experiments were conducted at four randomly selected tropical lowland rainforests at different successional stages:15-, 30- and 60-year-old secondary forests and one old-growth forest, each in the Bawangling Nature Reserve of Hainan Island. A total of 200 plots (20 m 20 m) were sampled. Individuals of tree and shrub species with diameter at breast height (DBH) 1 cm were identified and both their functional traits and the environmental factors in each plot were measured. ANOVA showed that canopy openness, level of soil nutrients, specific leaf area, and content of total organic carbon, nitrogen and phosphorus within the leaf decreased at the community level during the successional process. Meanwhile, leaf dry matter content, wood density and potential maximum height increased. However, soil water content and leaf potassium content, changed non-significantly. Multiple regression analysis demonstrated that environmental factors driving functional traits at the community level varied during the successional process. The key environmental factors were soil organic matter and pH value in the 15-year-old secondary forest; canopy openness and soil total phosphorus content in the 30-year-old secondary forest; available phosphorus and total potassium content of the soil in the 60-year-old secondary forest; and soil phosphorus content and organic matter content in the old-growth forest. Our results indicate that at different successional stages, the same functional traits respond to different environmental factors, enabling adaptation to specific environmental conditions.
URL
探讨植物功能性状(plant functional trait)受进化历史的影响及其与群落结构之间的关系是近来生态学研究的热点话题之一。植物功能性状,指的是能够响应自然环境的变化,对生态系统功能与过程以及植物的生长和繁殖有一定影响的植物性状,主要包括:比叶面积、叶寿命、叶片氮磷含量、光合效率与呼吸速率、种子重量和扩散模式、木材密度等形态、结构及生理等各方面的性状。植物的功能性状不仅受到环境变化的影响,同时还受进化历史的影响。 本研究以古田山24公顷永久性监测样地中的156种木本植物为对象,收集了叶片氮含量(Nmass)、磷含量(Pmass)、叶面积(Leaf area, LA)、木质密度(Wood density, WD)、比叶面积(Specific leaf area, SLA)、种子重量(Seed mass)和最大树高(Maximum height, Hmax)7种功能性状的数据,检验了各个功能性状的系统发育信号(phylogeny signal),并运用系统独立比较(phylogenetic independent contrasts, PIC)的方法结合Pearson线性回归分析各个性状之间的关系以及功能性状与多度的关系。我们发现:所有7种功能性状都表现出较强的系统发育信号,表明植物系统发育历史显著地影响所有检验的功能性状;另一方面,叶氮含量与叶磷含量、叶氮含量与比叶面积、叶磷含量与比叶面积、叶面积与最大树高呈现极为显著的正相关关系,叶面积与木质密度有极为显著的负相关性,比叶面积与种子重量也有负相关关系,但是程度较弱,而其他性状之间无相关性;另外,叶氮含量、叶磷含量、叶面积、木质密度和比叶面积与物种多度有显著相关性,而种子重量和最大树高与物种多度没有显著的相关性,表明群落内不同物种的资源获取方式可以显著地影响群落的结构。
URL
探讨植物功能性状(plant functional trait)受进化历史的影响及其与群落结构之间的关系是近来生态学研究的热点话题之一。植物功能性状,指的是能够响应自然环境的变化,对生态系统功能与过程以及植物的生长和繁殖有一定影响的植物性状,主要包括:比叶面积、叶寿命、叶片氮磷含量、光合效率与呼吸速率、种子重量和扩散模式、木材密度等形态、结构及生理等各方面的性状。植物的功能性状不仅受到环境变化的影响,同时还受进化历史的影响。 本研究以古田山24公顷永久性监测样地中的156种木本植物为对象,收集了叶片氮含量(Nmass)、磷含量(Pmass)、叶面积(Leaf area, LA)、木质密度(Wood density, WD)、比叶面积(Specific leaf area, SLA)、种子重量(Seed mass)和最大树高(Maximum height, Hmax)7种功能性状的数据,检验了各个功能性状的系统发育信号(phylogeny signal),并运用系统独立比较(phylogenetic independent contrasts, PIC)的方法结合Pearson线性回归分析各个性状之间的关系以及功能性状与多度的关系。我们发现:所有7种功能性状都表现出较强的系统发育信号,表明植物系统发育历史显著地影响所有检验的功能性状;另一方面,叶氮含量与叶磷含量、叶氮含量与比叶面积、叶磷含量与比叶面积、叶面积与最大树高呈现极为显著的正相关关系,叶面积与木质密度有极为显著的负相关性,比叶面积与种子重量也有负相关关系,但是程度较弱,而其他性状之间无相关性;另外,叶氮含量、叶磷含量、叶面积、木质密度和比叶面积与物种多度有显著相关性,而种子重量和最大树高与物种多度没有显著的相关性,表明群落内不同物种的资源获取方式可以显著地影响群落的结构。
DOI:10.1071/bt02124URL [本文引用: 1]
There is growing recognition that classifying terrestrial plant species on the basis of their function ( into 'functional types') rather than their higher taxonomic identity, is a promising way forward for tackling important ecological questions at the scale of ecosystems, landscapes or biomes. These questions include those on vegetation responses to and vegetation effects on, environmental changes ( e. g. changes in climate, atmospheric chemistry, land use or other disturbances). There is also growing consensus about a shortlist of plant traits that should underlie such functional plant classifications, because they have strong predictive power of important ecosystem responses to environmental change and/or they themselves have strong impacts on ecosystem processes. The most favoured traits are those that are also relatively easy and inexpensive to measure for large numbers of plant species. Large international research efforts, promoted by the IGBP-GCTE Programme, are underway to screen predominant plant species in various ecosystems and biomes worldwide for such traits. This paper provides an international methodological protocol aimed at standardising this research effort, based on consensus among a broad group of scientists in this field. It features a practical handbook with step-by-step recipes, with relatively brief information about the ecological context, for 28 functional traits recognised as critical for tackling large-scale ecological questions.
DOI:10.1890/0012-9658(2006)87[1465:ATTFHF]2.0.CO;2URLPMID:16869422 [本文引用: 1]
Community assembly theory suggests that two processes affect the distribution of trait values within communities: competition and habitat filtering. Within a local community, competition leads to ecological differentiation of coexisting species, while habitat filtering reduces the spread of trait values, reflecting shared ecological tolerances. Many statistical tests for the effects of competition exist in the literature, but measures of habitat filtering are less well-developed. Here, we present convex hull volume, a construct from computational geometry, which provides an n-dimensional measure of the volume of trait space occupied by species in a community. Combined with ecological null models, this measure offers a useful test for habitat filtering. We use convex hull volume and a null model to analyze California woody-plant trait and community data. Our results show that observed plant communities occupy less trait space than expected from random assembly, a result consistent with habitat filtering.
DOI:10.1007/s00442-002-1155-6URLPMID:12647118 [本文引用: 1]
Across 30 grassland sites in New Zealand that ranged from native alpine grasslands to low elevation improved pastures, there were consistent patterns of leaf and root traits and significant differences between native and non-native grasses. Plants of high altitude sites have low N concentrations in both their leaves and roots, have thick leaves and roots, yet no differences in tissue density or photosynthetic water use efficiency when compared to plants of low altitude sites. Both the leaves and roots of the low altitude plants were enriched in15N relative to the plants of higher altitude, indicating that the low-N set of traits is associated with a more closed N cycle at high altitude. A second independent set of correlations shows that plants of wetter habitats have lower photosynthetic water use efficiency (more negative$\partial {}^{13}\text{C}$) and lower leaf and root tissue density than the plants of drier sites. For both leaves and roots, plants of native species consistently had traits associated with lower resource availability: lower N concentrations, denser tissues, more negative$\partial {}^{15}\text{N}$, and more positive$\partial {}^{13}\text{C}$than non-native species. If root %N is correlated with root longevity as has been shown in other systems, root longevity may be able to be predicted from simple measurements of leaf %N, though a hysteresis in the relationship between leaf and root N concentrations may make prediction of high longevity roots difficult.
DOI:10.1016/j.tree.2010.11.011URLPMID:21196061
The worldwide leaf economic spectrum (WLES) is a strikingly consistent pattern of correlations among leaf traits. Although the WLES effectively summarizes variation in plant ecological strategies, little is known about its evolution. We reviewed estimates of natural selection and genetic variation for leaf traits to test whether the evolution of the WLES was limited by selection against unfit trait combinations or by genetic constraints. There was significant selection for leaf traits on both ends of the WLES spectrum, as well as significant genetic variation for these traits. In addition, genetic correlations between WLES traits were variable in strength and direction. These data suggest that genetic constraints have had a smaller role than selection in the evolution of the WLES.
DOI:10.3969/j.issn.1000-0852.2014.05.016URL [本文引用: 1]
利用漓江流域及周边地区25个降水观测点50年的监测数据,通过 Mapgis 6.7计算流域年降水量和月降水量.在此基础上,采用变差系数法、5年滑动平均法和Mann-Kendall检验法,分析漓江流域1960~2010年降 水变化规律.结果表明:(1)1960~2010年,漓江流域降水年际动态为波动增加,年降水量每10a增加23.1mm;其中1970~2010年间, 漓江流域年降水变化表现为波动减少,年降水量每10a减少14.6mm;但增减趋势都没有通过显著性检验;(2)漓江流域年内降水具有典型的雨季和旱季交 替特征,雨季降水占全年降水的76.26%;(3)漓江流域月降水量随时间变化具有规律性的增减,其中1月和6月降水量随着时间推移增加,4月降水量随着 时间推移而减少,1、4、6月的这种增减趋势通过99%的显著性检验;(4)漓江流域降水年内变化幅度比年际明显,年降水量变差系数小于0.18,月降水 量变差系数大于0.61.
DOI:10.3969/j.issn.1000-0852.2014.05.016URL [本文引用: 1]
利用漓江流域及周边地区25个降水观测点50年的监测数据,通过 Mapgis 6.7计算流域年降水量和月降水量.在此基础上,采用变差系数法、5年滑动平均法和Mann-Kendall检验法,分析漓江流域1960~2010年降 水变化规律.结果表明:(1)1960~2010年,漓江流域降水年际动态为波动增加,年降水量每10a增加23.1mm;其中1970~2010年间, 漓江流域年降水变化表现为波动减少,年降水量每10a减少14.6mm;但增减趋势都没有通过显著性检验;(2)漓江流域年内降水具有典型的雨季和旱季交 替特征,雨季降水占全年降水的76.26%;(3)漓江流域月降水量随时间变化具有规律性的增减,其中1月和6月降水量随着时间推移增加,4月降水量随着 时间推移而减少,1、4、6月的这种增减趋势通过99%的显著性检验;(4)漓江流域降水年内变化幅度比年际明显,年降水量变差系数小于0.18,月降水 量变差系数大于0.61.
DOI:10.2307/2648405URL [本文引用: 1]
1 If different factors inhibiting plant growth, e.g. low rainfall or low soil nutrients, were to select for species that have similar constellations of traits, then the unfavourable factors might usefully be grouped together as 'stress'.
DOI:10.1371/journal.pbio.0040278URLPMID:16895443
...
DOI:10.1111/j.1654-1103.2006.tb02444.xURL
In landscapes subject to intensive agriculture, both soil fertility and vegetation disturbance are capable of impacting strongly, evenly and simultaneously on the herbaceous plant cover and each tends to impose uniformity on the traits of constituent species. In more natural and ancient grasslands greater spatial and temporal variation in both productivity and disturbance occurs and both factors have been implicated in the maintenance of species-richness in herbaceous communities. However, empirical data suggest that disturbance is the more potent driver of trait differentiation and species co-existence at a local scale. This may arise from the great diversity in opportunities for establishment, growth or reproduction that arise when the intensity of competition is reduced by damage to the vegetation.
DOI:10.1078/1433-8319-00049URL [本文引用: 1]
An attempt is made to review the whole range of plants able to cope with shortages of mineral nutrients, water or light for at least a substantial part of their lives. In the past insufficient attention has been paid to quantitative measurements of the tolerance of various species for lack of resources. Growth rate at a low level of supply and mere survival are the variables to be considered. Problems in defining tolerance arise where a collection of species shows a negative correlation between survival at a low level of supply and growth rate under these conditions; in at least some circumstances a balance between survival and growth rate is important. Plants coping with lack of major resources display one of three strategies: ‘low-flexibility’, ‘switching’ or ‘gearing-down’. The essential features of the low-flexibility strategy are long-lived leaves, low maximum relative growth rates, and inflexibility of form and of gas exchange rates when resource-shortage is relieved, both in seedlings and in adults. Plants which show the switching strategy display the low-flexibility strategy as young seedlings, but are flexibile in form as older plants, and commonly have high relative growth rates then. The gearing-down strategy is based on an ability to reduce strongly the respiration rate when resources are in short supply, both as seedlings and as adult. In some cases this involves shedding of parts which would otherwise consume respiratory substrate. Plants showing this strategy have some characteristics which are the opposite of those shown by plants with the low-flexibility strategy: short-lived leaves, and high flexibility in form and in rates of gas exchange. All three strategies are represented among plants tolerant of the most extreme shortages of nutrients, water and light. It is concluded that while ecologists should attempt to reduce the complexities of nature to an oligo-dimensional framework of generalizing ideas, it is not reasonable to expect that any very simple scheme – such as might be represented by three or four reference points in one plane – will have generality on a world scale.
DOI:10.3969/j.issn.1674-5906.2013.05.026URL
河岸带是河流—陆地生态系统之间进行物质、能量、信息交换的重要生态过渡区,具有独特的生态系统结构和服务功能,近年来成为国内外生态学和环境科学的研究热点之一。在阐述河岸带结构与功能基础上,分析了河岸带的影响因素与其退化机制,退化河岸带的生态恢复理论、基本原则、生态重建技术和发展方向。认为影响河岸带结构与功能的主要因素可归纳为水文与地貌过程、植被与人为干扰4个方面;识别影响河岸带生态系统的生物和物理作用过程及其退化成因是关键,指出了生态恢复应遵循的原则与宜采用的生态重建技术。在此基础上,提出河岸带生态恢复应在景观或流域尺度上借助"3S"技术和多学科协作从微观、中观和宏观不同层次开展研究,甄别生态退化的主导因素,采用植被重建与水文调控技术尽可能恢复与重建原有自然景观。从系统生态学与景观生态学的角度,提出河岸带生态学未来研究方向。建议今后应加强对河岸带生态系统结构、生态过程与功能及生态重建技术的研究成果进行系统整合,建立能够预测河岸带结构与功能动态的数量模型和评价体系,为实现河岸带重建与高效管理提供科学指导。
DOI:10.3969/j.issn.1674-5906.2013.05.026URL
河岸带是河流—陆地生态系统之间进行物质、能量、信息交换的重要生态过渡区,具有独特的生态系统结构和服务功能,近年来成为国内外生态学和环境科学的研究热点之一。在阐述河岸带结构与功能基础上,分析了河岸带的影响因素与其退化机制,退化河岸带的生态恢复理论、基本原则、生态重建技术和发展方向。认为影响河岸带结构与功能的主要因素可归纳为水文与地貌过程、植被与人为干扰4个方面;识别影响河岸带生态系统的生物和物理作用过程及其退化成因是关键,指出了生态恢复应遵循的原则与宜采用的生态重建技术。在此基础上,提出河岸带生态恢复应在景观或流域尺度上借助"3S"技术和多学科协作从微观、中观和宏观不同层次开展研究,甄别生态退化的主导因素,采用植被重建与水文调控技术尽可能恢复与重建原有自然景观。从系统生态学与景观生态学的角度,提出河岸带生态学未来研究方向。建议今后应加强对河岸带生态系统结构、生态过程与功能及生态重建技术的研究成果进行系统整合,建立能够预测河岸带结构与功能动态的数量模型和评价体系,为实现河岸带重建与高效管理提供科学指导。
URLPMID:4620397 [本文引用: 1]
Leaf economics spectrum (LES), characterizing covariation among a suite of leaf traits relevant to carbon and nutrient economics, has been examined largely among species but hardly within species. In addition, very little attempt has been made to examine whether the existence of LES depends on spatial scales. To address these questions, we quantified the variation and covariation of four leaf economic traits (specific leaf area, leaf dry matter content, leaf nitrogen and phosphorus contents) in a cosmopolitan wetland species (Phragmites australis) at three spatial (inter-regional, regional, and site) scales across most of the species range in China. The species expressed large intraspecific variation in the leaf economic traits at all of the three spatial scales. It also showed strong covariation among the four leaf economic traits across the species range. The coordination among leaf economic traits resulted in LES at all three scales and the environmental variables determining variation in leaf economic traits were different among the spatial scales. Our results provide novel evidence for within-species LES at multiple spatial scales, indicating that resource trade-off could also constrain intraspecific trait variation mainly driven by climatic and/or edaphic differences.
DOI:10.5846/stxb201508281789URL
以漓江水陆交错带为研究区,分两个条带分别量测了适生植物的5个叶性状指标:最大净光合速率(Amax)、比叶重(LMA)、单位质量叶片全氮含量(Nmaxx)、单位质量叶片全磷含量(Pmaxa)、单位质量叶片全钾含量(Kmass)。研究重度淹没带与微度淹没带不同功能型植物叶性状间的差异,分析并讨论重度淹没带叶性状间的关系与全球尺度是否存在差异,探究重度淹没带植物对水淹生境的生理响应机制。结果如下:(1)重度淹没带植物叶片的Amax、Nmaxx、Pmass显著高于微度淹没带。(2)乔木、灌木叶片的LMA均显著高于草本植物,而Amass。、PPUE均显著低于草本植物。(3)重度淹没带草本叶性状指标的Nmass、Pmass、PNUE均显著高于微度微度淹没带,而乔木、灌木的叶性状在两个条带的差异则不显著。(4)重度淹没带植物叶性状关系与全球尺度基本一致,其植物叶片具有低LMA,高Amass、Nmass、Pmass。分析可知,重度淹没带植物在出露期提高叶片光合效率及相关营养水平可能是其适应水淹胁迫特殊生境的关键策略之一;不同功能型植物对同一环境的适应能力存在一定的差异,草本对于水淹环境的响应更为积极,适应能力更好;重度淹没带也存在叶经济谱,其植物在经济谱中属于“快速投资一收益”型物种。
DOI:10.5846/stxb201508281789URL
以漓江水陆交错带为研究区,分两个条带分别量测了适生植物的5个叶性状指标:最大净光合速率(Amax)、比叶重(LMA)、单位质量叶片全氮含量(Nmaxx)、单位质量叶片全磷含量(Pmaxa)、单位质量叶片全钾含量(Kmass)。研究重度淹没带与微度淹没带不同功能型植物叶性状间的差异,分析并讨论重度淹没带叶性状间的关系与全球尺度是否存在差异,探究重度淹没带植物对水淹生境的生理响应机制。结果如下:(1)重度淹没带植物叶片的Amax、Nmaxx、Pmass显著高于微度淹没带。(2)乔木、灌木叶片的LMA均显著高于草本植物,而Amass。、PPUE均显著低于草本植物。(3)重度淹没带草本叶性状指标的Nmass、Pmass、PNUE均显著高于微度微度淹没带,而乔木、灌木的叶性状在两个条带的差异则不显著。(4)重度淹没带植物叶性状关系与全球尺度基本一致,其植物叶片具有低LMA,高Amass、Nmass、Pmass。分析可知,重度淹没带植物在出露期提高叶片光合效率及相关营养水平可能是其适应水淹胁迫特殊生境的关键策略之一;不同功能型植物对同一环境的适应能力存在一定的差异,草本对于水淹环境的响应更为积极,适应能力更好;重度淹没带也存在叶经济谱,其植物在经济谱中属于“快速投资一收益”型物种。
DOI:10.3969/j:issn.2095-1191.2013.10.1700URL
【目的】分析漓江河流形态、水质状况、河岸植被群落结构,为漓江及其沿岸生态景观的修复提供参考依据。【方法】采用普查、重点样地调查、问卷调查结合查阅相关文献的方法,对漓江沿岸河岸形态、水质状况、植物景观现状进行分析,提出相应的生态修复技术。【结果】漓江沿岸由于人类活动的频繁,特别是自然河道的裁弯取直、渠道化、修筑硬质驳岸等人为因素的影响,自然植被受到严重干扰,阻碍了其生态服务功能的发挥。应用生态砾石床水质净化技术、植物浮岛技术、增加河岸植被缓冲带等一系列生态修复技术,使漓江沿岸驳岸渠道化现象得以改善,恢复了河流的蜿蜒曲折状态;沿岸残缺的植被得以修复,植物数量和种类有所增加,植物景观色彩和层次更加丰富,物种多样性和植物群落结构已基本恢复。【结论】应用生态砾石床水质净化技术、植物浮岛技术、增加河岸植被缓冲带等一系列生态修复技术,可促进漓江旅游生态系统的可持续发展。
DOI:10.3969/j:issn.2095-1191.2013.10.1700URL
【目的】分析漓江河流形态、水质状况、河岸植被群落结构,为漓江及其沿岸生态景观的修复提供参考依据。【方法】采用普查、重点样地调查、问卷调查结合查阅相关文献的方法,对漓江沿岸河岸形态、水质状况、植物景观现状进行分析,提出相应的生态修复技术。【结果】漓江沿岸由于人类活动的频繁,特别是自然河道的裁弯取直、渠道化、修筑硬质驳岸等人为因素的影响,自然植被受到严重干扰,阻碍了其生态服务功能的发挥。应用生态砾石床水质净化技术、植物浮岛技术、增加河岸植被缓冲带等一系列生态修复技术,使漓江沿岸驳岸渠道化现象得以改善,恢复了河流的蜿蜒曲折状态;沿岸残缺的植被得以修复,植物数量和种类有所增加,植物景观色彩和层次更加丰富,物种多样性和植物群落结构已基本恢复。【结论】应用生态砾石床水质净化技术、植物浮岛技术、增加河岸植被缓冲带等一系列生态修复技术,可促进漓江旅游生态系统的可持续发展。
DOI:10.1111/j.1365-2745.2010.01687.xURL [本文引用: 1]
1. Trait-based approaches applied to community ecology have led to a considerable advance in understanding the effect of environmental filters on species assembly. Although plant traits are known to vary both between and within species, little is known about the role of intraspecific trait variability in the non-random assembly mechanisms controlling the coexistence of species, including habitat filtering and niche differentiation.2. We investigate the role of intraspecific variability in three key functional traits – specific leaf area (SLA), leaf dry matter content (LDMC) and height – in structuring grassland communities distributed along a flooding gradient. We quantified the contribution of intraspecific variability relative to interspecific differences in the trait–gradient relationship, and we used a null model approach to detect patterns of habitat filtering and niche differentiation, with and without intraspecific variability.3. Community mean SLA and height varied significantly along the flooding gradient and intraspecific variability accounted for 44% and 32%, respectively, of these trait–gradient relationships. LDMC did not vary along the gradient, with and without accounting for intraspecific variability. Our null model approach revealed significant patterns of habitat filtering and niche differentiation for SLA and height, but not for LDMC. More strikingly, considering intraspecific trait variability greatly increased the detection of habitat filtering and was necessary to detect niche differentiation processes.4. Synthesis. Our study provides evidence for a strong role of intraspecific trait variability in community assembly. Our findings suggest that intraspecific trait variability promotes species coexistence, by enabling species to pass through both abiotic and biotic filters. We argue that community ecology would benefit from more attention to intraspecific variability.
DOI:10.1126/science.1160662URLPMID:19460986 [本文引用: 1]
It is debated whether species-level differences in ecological strategy, which play a key role in much of coexistence theory, are important in structuring highly diverse communities. We examined the co-occurrence patterns of over 1100 tree species in a 25-hectare Amazonian forest plot in relation to field-measured functional traits. Using a null model approach, we show that co-occurring trees are often less ecologically similar than a niche-free (neutral) model predicts. Furthermore, we find evidence for processes that simultaneously drive convergence and divergence in key aspects of plant strategy, suggesting that at least two distinct niche-based processes are occurring. Our results show that strategy differentiation among species contributes to the maintenance of diversity in one of the most diverse tropical forests in the world.
DOI:10.1038/nature16476URLPMID:26700807 [本文引用: 1]
Abstract Phenotypic traits and their associated trade-offs have been shown to have globally consistent effects on individual plant physiological functions, but how these effects scale up to influence competition, a key driver of community assembly in terrestrial vegetation, has remained unclear. Here we use growth data from more than 3 million trees in over 140,000 plots across the world to show how three key functional traits--wood density, specific leaf area and maximum height--consistently influence competitive interactions. Fast maximum growth of a species was correlated negatively with its wood density in all biomes, and positively with its specific leaf area in most biomes. Low wood density was also correlated with a low ability to tolerate competition and a low competitive effect on neighbours, while high specific leaf area was correlated with a low competitive effect. Thus, traits generate trade-offs between performance with competition versus performance without competition, a fundamental ingredient in the classical hypothesis that the coexistence of plant species is enabled via differentiation in their successional strategies. Competition within species was stronger than between species, but an increase in trait dissimilarity between species had little influence in weakening competition. No benefit of dissimilarity was detected for specific leaf area or wood density, and only a weak benefit for maximum height. Our trait-based approach to modelling competition makes generalization possible across the forest ecosystems of the world and their highly diverse species composition.
DOI:10.5846/stxb201303130407URL [本文引用: 1]
研究根系与土壤关系是发掘河岸带生态退化等问题内在原因的重要途径。在漓江流域水陆交错带选取缓坡、陡坡、江心洲、人工岸坡4种典型立地类型,对不同土层深度的根长密度、根系生物量、比根长,以及根系特征与土壤有机质、全氮、有效磷的关系进行了研究,旨在为漓江流域生态修复过程中植被恢复、植被配置、快速绿化材料选取提供科学依据。结果表明:(1)同一立地类型0—10cm土层和10—20cm土层比根长差异性不显著。0—10cm到10—20cm土层,各立地类型根长密度和根系生物量密度均减小,但不同立地类型根长密度和根系生物量密度的差异程度逐渐缩小,表明地形、地表植物类型及生长状况对根长密度分布的影响也随土层深度的增加而逐渐减小。细根根长和生物量随着土壤深度的增加而减小。(2)土壤有机质含量差异性显著,分布规律为人工岸坡〉陡坡〉江心洲〉缓坡;土壤全氮含量从大到小依次是人工岸坡、陡坡、缓坡、江心洲,其值分别为:3.12、2.33、1.56、1.32g/kg;土壤全氮与土壤有机质呈显著正相关。江心洲和缓坡有效磷含量远远大于人工岸坡和陡坡,原因是漓江水长期受人为洗漱影响,导致受江水干扰大的立地类型有效磷含量高。(3)根长密度、比根长、根系生物量与有机质、全氮含量呈正相关,与有效磷含量呈负相关,说明土壤根系越丰富,越有利于增加土壤有机质和全氮含量,但遏制了土壤有效磷。细根长度、生物量与根长密度在0.01水平(双侧)上显著正相关,与根系生物量密度呈负相关。
DOI:10.5846/stxb201303130407URL [本文引用: 1]
研究根系与土壤关系是发掘河岸带生态退化等问题内在原因的重要途径。在漓江流域水陆交错带选取缓坡、陡坡、江心洲、人工岸坡4种典型立地类型,对不同土层深度的根长密度、根系生物量、比根长,以及根系特征与土壤有机质、全氮、有效磷的关系进行了研究,旨在为漓江流域生态修复过程中植被恢复、植被配置、快速绿化材料选取提供科学依据。结果表明:(1)同一立地类型0—10cm土层和10—20cm土层比根长差异性不显著。0—10cm到10—20cm土层,各立地类型根长密度和根系生物量密度均减小,但不同立地类型根长密度和根系生物量密度的差异程度逐渐缩小,表明地形、地表植物类型及生长状况对根长密度分布的影响也随土层深度的增加而逐渐减小。细根根长和生物量随着土壤深度的增加而减小。(2)土壤有机质含量差异性显著,分布规律为人工岸坡〉陡坡〉江心洲〉缓坡;土壤全氮含量从大到小依次是人工岸坡、陡坡、缓坡、江心洲,其值分别为:3.12、2.33、1.56、1.32g/kg;土壤全氮与土壤有机质呈显著正相关。江心洲和缓坡有效磷含量远远大于人工岸坡和陡坡,原因是漓江水长期受人为洗漱影响,导致受江水干扰大的立地类型有效磷含量高。(3)根长密度、比根长、根系生物量与有机质、全氮含量呈正相关,与有效磷含量呈负相关,说明土壤根系越丰富,越有利于增加土壤有机质和全氮含量,但遏制了土壤有效磷。细根长度、生物量与根长密度在0.01水平(双侧)上显著正相关,与根系生物量密度呈负相关。
DOI:10.5846/stxb201312172967URL [本文引用: 1]
针对旅游区水陆交错带出现植被退化、砾石裸露导致景观观赏度降低的问题,以广西桂林市大圩古镇水陆交错带为研究区域,按照水陆交错带淹没程度将其划分不同淹没区,并在详细调查基础上研究了各淹没区植被指标与土壤理化特征分布特征。结果表明:研究区内水陆交错带物种组成以一年生草本植物为主,物种多样性在各个不同淹没区基本呈现为先增后减的趋势,具体为轻度淹没区中度淹没区重度淹没区微度淹没区;随着水文作用的减弱,土壤砂粒含量呈现逐渐减小的趋势,粉粒和黏粒含量呈现逐渐增加趋势,土壤容重、土壤有机质含量和土壤全磷含量呈现先减后增的趋势;全氮含量呈现先增后减趋势,全钾含量在各淹没区差异不明显;研究表明大圩古镇水陆交错带土壤理化性质与植被多样性强烈地受以水位为主导的水文过程的影响与控制。扁蓄可以作为一种指示水淹时间的物种;此外,大圩古镇水陆交错带除受水文作用影响较大外,高强度旅游干扰也是重要的因素之一,植被恢复工作应兼顾生态和旅游,提升大圩古镇景观游憩度。
DOI:10.5846/stxb201312172967URL [本文引用: 1]
针对旅游区水陆交错带出现植被退化、砾石裸露导致景观观赏度降低的问题,以广西桂林市大圩古镇水陆交错带为研究区域,按照水陆交错带淹没程度将其划分不同淹没区,并在详细调查基础上研究了各淹没区植被指标与土壤理化特征分布特征。结果表明:研究区内水陆交错带物种组成以一年生草本植物为主,物种多样性在各个不同淹没区基本呈现为先增后减的趋势,具体为轻度淹没区中度淹没区重度淹没区微度淹没区;随着水文作用的减弱,土壤砂粒含量呈现逐渐减小的趋势,粉粒和黏粒含量呈现逐渐增加趋势,土壤容重、土壤有机质含量和土壤全磷含量呈现先减后增的趋势;全氮含量呈现先增后减趋势,全钾含量在各淹没区差异不明显;研究表明大圩古镇水陆交错带土壤理化性质与植被多样性强烈地受以水位为主导的水文过程的影响与控制。扁蓄可以作为一种指示水淹时间的物种;此外,大圩古镇水陆交错带除受水文作用影响较大外,高强度旅游干扰也是重要的因素之一,植被恢复工作应兼顾生态和旅游,提升大圩古镇景观游憩度。
DOI:10.3969/j.issn.1001-4810.2003.01.013URL [本文引用: 1]
漓江从战国时期就有记载一直至今.历史上漓江(桂江)曾分别被称丽水、(丽鹿水)、离水、漓水.从这些称谓中可看出,2000多年以前古人就认为漓江是一条非常美丽神奇的水系,这与今天人们对它的评价是一致的.
DOI:10.3969/j.issn.1001-4810.2003.01.013URL [本文引用: 1]
漓江从战国时期就有记载一直至今.历史上漓江(桂江)曾分别被称丽水、(丽鹿水)、离水、漓水.从这些称谓中可看出,2000多年以前古人就认为漓江是一条非常美丽神奇的水系,这与今天人们对它的评价是一致的.
DOI:10.1360/N052014-00244URL
Plant functional traits are plant core attributes closely related to plant colonization, survival, growth and mortality. These core attributes could significantly affect ecosystem functions and reflect the response of vegetation to environmental change. There is growing recognition that many ecological questions at various levels can be addressed effectively with key functional traits. In this paper, we review key research progress over the last 10 years addressing important ecological issues based on functional traits, including global distribution patterns of functional traits and their associations, variation of traits distribution along environmental gradients, functional diversity and its application, mechanisms of community assembly and dynamics regarding functional traits, the relationship between traits and phylogeny, the effect and response of functional traits to ecosystem functions and disturbance. In addition, we also provide a future outlook, including research directions, methods and applications based on functional traits in plant science. Specially, we propose that functional traits can also be used as indicators or reference for biodiversity conservation and management of ecosystems under the current global climate change. China has a huge territory, which brings great natural recourses of geography and plants, providing a huge natural database for research on functional traits.
DOI:10.1360/N052014-00244URL
Plant functional traits are plant core attributes closely related to plant colonization, survival, growth and mortality. These core attributes could significantly affect ecosystem functions and reflect the response of vegetation to environmental change. There is growing recognition that many ecological questions at various levels can be addressed effectively with key functional traits. In this paper, we review key research progress over the last 10 years addressing important ecological issues based on functional traits, including global distribution patterns of functional traits and their associations, variation of traits distribution along environmental gradients, functional diversity and its application, mechanisms of community assembly and dynamics regarding functional traits, the relationship between traits and phylogeny, the effect and response of functional traits to ecosystem functions and disturbance. In addition, we also provide a future outlook, including research directions, methods and applications based on functional traits in plant science. Specially, we propose that functional traits can also be used as indicators or reference for biodiversity conservation and management of ecosystems under the current global climate change. China has a huge territory, which brings great natural recourses of geography and plants, providing a huge natural database for research on functional traits.
DOI:10.1016/j.tree.2006.02.002URLPMID:16701083
There is considerable debate about whether community ecology will ever produce general principles. We suggest here that this can be achieved but that community ecology has lost its way by focusing on pairwise species interactions independent of the environment. We assert that community ecology should return to an emphasis on four themes that are tied together by a two-step process: how the fundamental niche is governed by functional traits within the context of abiotic environmental gradients; and how the interaction between traits and fundamental niches maps onto the realized niche in the context of a biotic interaction milieu. We suggest this approach can create a more quantitative and predictive science that can more readily address issues of global change.
DOI:10.17521/cjpe.2007.0019URL
植物性状反映了植物对生长环境的响应和适应,将环境、植物个体和生态系统结构、过程与功能联系起来(所谓的“植物功能性状”)。该文介绍了植物功能性状的分类体系,综述了国内外植物功能性状与气候(包括气温、降水、光照)、地理空间变异(包括地形地貌、生态梯度、海拔)、营养、干扰(包括火灾、放牧、生物入侵、土地利用)等环境因素,以及与生态系统功能之间关系的研究进展,探讨了全球变化(气候变化和CO2浓度升高)对个体和群落植物功能性状的影响。植物功能性状的研究已经取得很多成果,并应用于全球变化、古植被恢复和古气候定量重建、环境监测与评价、生态保护和恢复等研究中,但大尺度、多生境因子下的植物功能性状研究仍有待于加强,同时需要改进性状的测量手段;我国的植物功能性状研究还需要更加明朗化和系统化。
DOI:10.17521/cjpe.2007.0019URL
植物性状反映了植物对生长环境的响应和适应,将环境、植物个体和生态系统结构、过程与功能联系起来(所谓的“植物功能性状”)。该文介绍了植物功能性状的分类体系,综述了国内外植物功能性状与气候(包括气温、降水、光照)、地理空间变异(包括地形地貌、生态梯度、海拔)、营养、干扰(包括火灾、放牧、生物入侵、土地利用)等环境因素,以及与生态系统功能之间关系的研究进展,探讨了全球变化(气候变化和CO2浓度升高)对个体和群落植物功能性状的影响。植物功能性状的研究已经取得很多成果,并应用于全球变化、古植被恢复和古气候定量重建、环境监测与评价、生态保护和恢复等研究中,但大尺度、多生境因子下的植物功能性状研究仍有待于加强,同时需要改进性状的测量手段;我国的植物功能性状研究还需要更加明朗化和系统化。
DOI:10.1641/0006-3568(2000)050[0783:AORECB]2.0.CO;2URL
DOI:10.2307/2390579URL [本文引用: 1]
1. Specific leaf area is a variable implicated in a number of functional aspects of plant ecology, including gas exchange, relative growth rate and palatability. 2. Six leaf attributes (water content, lamina thickness, protruding vein thickness, dry weight, lamina area and specific leaf area) were determined on a collection of 572 leaves. The data set was derived from 194 individual plants, 34 species, 32 genera, 18 families and both classes of angiosperms. All species except one (a shrub) were herbaceous. All species were from sunny habitats. 3. Structured relationships between these six variables were obtained. These relationships consisted of path analyses, multiple regressions and major axis (allometric) regressions. 4. There was an interspecific allometric trend for leaf dry weight to increase more rapidly than leaf surface area, resulting in larger leaves having a lower specific leaf area. This trend did not exist at an intraspecific level. 5. Path analysis showed that the most important direct effect on specific leaf area was the water content of the leaf. Increasing water content resulted in a larger specific leaf area. The direct effect of lamina thickness was to decrease specific leaf area but lamina thickness itself was affected by leaf water content. 6. These effects on specific leaf area were mediated through variation in leaf dry weight. Leaf surface area appeared to be independent of the other five variables. 7. These results help to explain two unresolved general patterns in plant comparative ecology: (1) why net photosynthetic rate is not positively correlated with maximum relative growth rates and (2) why there is a negative correlation between maximum relative growth rate and seed size.
DOI:10.1111/j.0022-0477.2004.00898.xURL
1 We sought evidence among the plant species of a New Zealand sand dune community that limiting similarity controls the ability of species to coexist. Sampling was at four spatial scales, from a single point up to a scale of 50 m2. Twenty-three functional characters were measured on each of the species, covering the morphology of the shoot and root systems and nutrient status, and intended to represent modes of resource acquisition. 2 Patterns of association between plant species at the four scales were examined for any tendency for plants with similar functional characters to coexist less often than expected at random (e.g. if a point has three species, do they have notably different characters?) The observed results were compared with the patterns expected under a null model using a range of test statistics. 3 A test over all characters found that the mean dissimilarity between nearest-neighbour species in functional space, and the minimum dissimilarity, were greater than expected under the null model at the 0.5 0.5 m scale. This supports the MacArthur & Levins model, although the actual community did not show an even spread of species over functional space. 4 Limiting similarity effects were seen even more consistently in separate characters when within-species variation was taken into account to calculate measures of overlap. The characters involved were mainly those related to rooting patterns and leaf water control, and thus perhaps reflecting the acquisition of nutrients and/or water. 5 Our results seem to be amongst the most convincing support for the theory of limiting similarity, and the only example involving vegetative processes in plant communities. The characters involved suggest that species can more readily coexist if they differ in their water-use pattern, reducing competition between them.
DOI:10.1111/j.1469-8137.2008.02650.xURLPMID:19138225
Field studies with transgenic Arabidopsis lines have been performed over 8 yr, to better understand the influence that certain genes have on plant performance. Many (if not most) plant phenotypes cannot be observed under the near constant, low-stress conditions in growth chambers, making field experiments necessary. However, there are challenges in performing such experiments: permission must be obtained and regulations obeyed, the profound influence of uncontrollable biotic and abiotic factors has to be considered, and experimental design has to be strictly controlled. The aim here is to provide inspiration and guidelines for researchers who are not used to setting up such experiments, allowing others to learn from our mistakes. This is believed to be the first example of a 'manual' for field experiments with transgenic Arabidopsis plants. Many of the challenges encountered are common for all field experiments, and many researchers from ecological backgrounds are skilled in such methods. There is huge potential in combining the detailed mechanistic understanding of molecular biologists with ecologists' expertise in examining plant performance under field conditions, and it is suggested that more interdisciplinary collaborations will open up new scientific avenues to aid analyses of the roles of genetic and physiological variation in natural systems.
DOI:10.1631/jzus.A1000244URL [本文引用: 1]
通过样地调查和Spearman相关分析,研究了澎溪河消落带植物群落物种丰富度格局.结果表明:沿河流纵向梯度,总物种丰富度、灌木层和草本层物种丰富度均表现出抛物线型,乔木层物种丰富度表现出直线型;沿河流侧向梯度,总物种丰富度和草本层物种丰富度随高程的增加先升高后降低;乔木层和灌木层物种丰富度随高程的升高而增加.在纵向梯度上,底质类型与物种丰富度显著相关,距河口的空间距离对物种丰富度有重要的控制作用;在侧向梯度上,淹水时间、土壤含水量、底质异质性对群落空间格局有重要影响.总体来说,消落带植物物种丰富度格局仍受原河流地貌及洪水格局的影响,但由于水位季节性变动,已形成与水位变动相适应的分布格局.
DOI:10.1631/jzus.A1000244URL [本文引用: 1]
通过样地调查和Spearman相关分析,研究了澎溪河消落带植物群落物种丰富度格局.结果表明:沿河流纵向梯度,总物种丰富度、灌木层和草本层物种丰富度均表现出抛物线型,乔木层物种丰富度表现出直线型;沿河流侧向梯度,总物种丰富度和草本层物种丰富度随高程的增加先升高后降低;乔木层和灌木层物种丰富度随高程的升高而增加.在纵向梯度上,底质类型与物种丰富度显著相关,距河口的空间距离对物种丰富度有重要的控制作用;在侧向梯度上,淹水时间、土壤含水量、底质异质性对群落空间格局有重要影响.总体来说,消落带植物物种丰富度格局仍受原河流地貌及洪水格局的影响,但由于水位季节性变动,已形成与水位变动相适应的分布格局.
DOI:10.1093/aob/mcj004URLPMID:2803375 [本文引用: 1]
61 Background and Aims The leaf size-twig size spectrum is one of the leading dimensions of plant ecological variation, and now it is under development. The purpose of this study was to test whether the relationship between leaf size and twig size is isometric or allometric, and to examine the relationship between plant allometric growth and life history strategies in the spectrum. 61 Methods Leaf and stem characters—including leaf and stem mass, total leaf area, individual leaf area, stem crosssectional area, leaf number and stem length—at the twig level for 59 woody species were investigated along an altitudinal gradient on Changbaishan Mountain in the temperate zone of China. The environmental gradient ranges from temperate broad-leaved mixed forest at low altitude, to conifer forest at middle altitude, and to sub-alpine birch forest at high altitude. The scaling relationships between stem cross-sectional area and stem mass, stem mass and leaf mass, and leaf mass and leaf area at the twig level were simultaneously determined. 61 Key Results Twig cross-sectional area was found to have invariant allometric scaling relationships with the stem mass, leaf mass, total leaf area and individual leaf area, all with common slopes being significantly larger than 1, for three altitudinal-zoned vegetation types under investigation. However, leaf mass was found to be isometrically related to stem mass and leaf area along the environmental gradient. Based on the predictions of previous models, the exponent value of the relationship between twig cross-sectional area and total leaf area can be inferred to be 1.5, which falls between the confidence intervals of the relationship at each altitude, and between the confidence intervals of the common slope value (1.17-1.56) of this study. This invariant scaling relationship is assumed to result from the fracturai network and/or developmental constraints of plants. The allometric constants (y-intercepts) of the relationships between the stem cross-sectional area and leaf area (both total leaf area and individual leaf area) were found to decrease significantly along the altitudinal gradient. This suggests that the species would support less leaf area at a given twig cross-sectional area with increasing environmental stress. 61 Conclusions This study demonstrated that plants respond to the environmental gradient by changing the y-intercepts of the relationship between leaf size-twig size, while keeping the exponent value of the allometric relationship as an invariant constant. The allometric growth in the twig size-leaf size spectrum is related to many other components of plant life history strategy, including the well established life history trade-off between efficiency and safety in the hydraulic transport of water.
DOI:10.1139/f80-017URL [本文引用: 1]
DOI:10.1093/aob/mcl066URLPMID:28029050663 [本文引用: 1]
61 Background and Aims When ecologically important plant traits are correlated they may be said to constitute an ecological 'strategy' dimension. Through identifying these dimensions and understanding their inter-relationships we gain insight into why particular trait combinations are favoured over others and into the implications of trait differences among species. Here we investigated relationships among several traits, and thus the strategy dimensions they represented, across 2134 woody species from seven Neotropical forests. 61 Methods Six traits were studied: specific leaf area (SLA), the average size of leaves, seed and fruit, typical maximum plant height, and wood density (WD). Trait relationships were quantified across species at each individual forest as well as across the dataset as a whole. 'Phylogenetic' analyses were used to test for correlations among evolutionary trait-divergences and to ascertain whether interspecific relationships were biased by strong taxonomic patterning in the traits. 61 Key Results The interspecific and phylogenetic analyses yielded congruent results. Seed and fruit size were expected, and confirmed, to be tightly related. As expected, plant height was correlated with each of seed and fruit size, albeit weakly. Weak support was found for an expected positive relationship between leaf and fruit size. The prediction that SLA and WD would be negatively correlated was not supported. Otherwise the traits were predicted to be largely unrelated, being representatives of putatively independent strategy dimensions. This was indeed the case, although WD was consistently, negatively related to leaf size. 61 Conclusions The dimensions represented by SLA, seed/fruit size and leaf size were essentially independent and thus conveyed largely independent information about plant strategies. To a lesser extent the same was true for plant height and WD. Our tentative explanation for negative WD-leaf size relationships, now also known from other habitats, is that the traits are indirectly linked via plant hydraulics.
DOI:10.3724/SP.J.1258.2011.01000URL [本文引用: 1]
认识植物功能性状随演替进展的变化规律和不同性状之间的关系,有助于从功能生态学的角度来理解群落演替。该文调查了位于贵州省中部的普定县喀斯特山区26个样方的物种组成,这些样方分别处于灌丛、落叶阔叶林、落叶常绿混交林3个演替阶段;测量了分布于该区域的82种木本植物的3个功能性状值(叶面积、比叶面积、最大高度);根据物种在样方中的多度加权计算得到26个样方的性状平均值,在此基础上分析了随演替进展植物功能性状在群落水平上的变异格局和不同性状之间的相关性;采用性状梯度分析法分析了各阶段优势物种的功能性状在群落间(β组分)和群落内(α组分)的变异格局及相关性。结果表明:1)沿灌丛→落叶阔叶林→落叶常绿混交林这一演替顺序,群落平均叶面积和平均高度逐渐增加,而群落平均比叶面积则逐渐变小;2)就群落平均性状值而言,叶面积与最大高度呈较强的正相关关系,最大高度与比叶面积、叶面积与比叶面积均呈较强的负相关关系;3)物种功能性状的α组分之间没有显著的相关关系,而β组分之间相关性显著。这说明:随演替的进展,群落优势物种对环境的适应策略由高速生长转向提高资源利用效率,而同一群落内共存的物种采取不同的性状组合来适应共同的群落环境。
DOI:10.3724/SP.J.1258.2011.01000URL [本文引用: 1]
认识植物功能性状随演替进展的变化规律和不同性状之间的关系,有助于从功能生态学的角度来理解群落演替。该文调查了位于贵州省中部的普定县喀斯特山区26个样方的物种组成,这些样方分别处于灌丛、落叶阔叶林、落叶常绿混交林3个演替阶段;测量了分布于该区域的82种木本植物的3个功能性状值(叶面积、比叶面积、最大高度);根据物种在样方中的多度加权计算得到26个样方的性状平均值,在此基础上分析了随演替进展植物功能性状在群落水平上的变异格局和不同性状之间的相关性;采用性状梯度分析法分析了各阶段优势物种的功能性状在群落间(β组分)和群落内(α组分)的变异格局及相关性。结果表明:1)沿灌丛→落叶阔叶林→落叶常绿混交林这一演替顺序,群落平均叶面积和平均高度逐渐增加,而群落平均比叶面积则逐渐变小;2)就群落平均性状值而言,叶面积与最大高度呈较强的正相关关系,最大高度与比叶面积、叶面积与比叶面积均呈较强的负相关关系;3)物种功能性状的α组分之间没有显著的相关关系,而β组分之间相关性显著。这说明:随演替的进展,群落优势物种对环境的适应策略由高速生长转向提高资源利用效率,而同一群落内共存的物种采取不同的性状组合来适应共同的群落环境。
URL [本文引用: 1]
河岸带植被结构与组成等配置状况直接影响其稳定河岸、净化水质、生物多样性、生物栖息地以及旅游景观等重要功能。查明漓江河岸带主要植被配置类型及其各自退化表现、原因与恢复途径,是开展河岸带退化植被恢复的基础与前提。该研究通过野外实地调查与测量,对漓江桂林-阳朔河段51个河岸带断面的地形、土壤、植被等进行了系统调查,分析了河岸带植被退化的主要表现、主导干扰因素,并通过流域与河岸带尺度综合的角度,提出了漓江河岸带植被恢复的思路。从岸坡地形与植被组合出发,按照自然、近自然、人工等干扰程度,广西漓江河岸带可划分为江心洲宽滩地疏林灌草自然型、丘陵陡坡林灌草自然型、平原农田缓坡灌草近自然型、平原滩地林草近自然型、聚落阶地疏灌林草人工-自然混合型、城镇岸坡林草人工型等6种典型植被配置模式。漓江河岸带植被退化主要表现为多样性差、景观破碎、生态功能弱,而城镇、交通、旅游等不合理开发建设是主导性的干扰因素。从生态稳定和景观美观的角度,提出了漓江河岸带植被恢复与优化的思路,以期为广西漓江河岸带生态修复提供科学依据。
URL [本文引用: 1]
河岸带植被结构与组成等配置状况直接影响其稳定河岸、净化水质、生物多样性、生物栖息地以及旅游景观等重要功能。查明漓江河岸带主要植被配置类型及其各自退化表现、原因与恢复途径,是开展河岸带退化植被恢复的基础与前提。该研究通过野外实地调查与测量,对漓江桂林-阳朔河段51个河岸带断面的地形、土壤、植被等进行了系统调查,分析了河岸带植被退化的主要表现、主导干扰因素,并通过流域与河岸带尺度综合的角度,提出了漓江河岸带植被恢复的思路。从岸坡地形与植被组合出发,按照自然、近自然、人工等干扰程度,广西漓江河岸带可划分为江心洲宽滩地疏林灌草自然型、丘陵陡坡林灌草自然型、平原农田缓坡灌草近自然型、平原滩地林草近自然型、聚落阶地疏灌林草人工-自然混合型、城镇岸坡林草人工型等6种典型植被配置模式。漓江河岸带植被退化主要表现为多样性差、景观破碎、生态功能弱,而城镇、交通、旅游等不合理开发建设是主导性的干扰因素。从生态稳定和景观美观的角度,提出了漓江河岸带植被恢复与优化的思路,以期为广西漓江河岸带生态修复提供科学依据。
A trait-based approach to community assembly: Partitioning of species trait values into within- and among-community components
2
2007
... 根据
... 采用零模型的方法计算随机期望下的不同性状$\alpha$/$\beta$组分协变强度和不同物种$\beta$组分的离散度.其中, $\alpha$/$\beta$组分协变强度的零模型是根据
Leaf size, specific leaf area and microhabitat distribution of chaparral woody plants: Contrasting patterns in species level and community level analyses
1
2002
... 植物在其漫长的历史进化和发展过程中, 受物理、生理等因素的综合作用和影响, 功能性状之间往往会呈现出一定的协变和权衡关系(
1
1983
... 经过现场踏查, 于2016年9-10月和2017年7-9月, 在漓江河岸带上游、中游、下游各建立了12个大小为20 m × 20 m的样方, 合计36个样方, 各样方的基本情况如
1
1983
... 经过现场踏查, 于2016年9-10月和2017年7-9月, 在漓江河岸带上游、中游、下游各建立了12个大小为20 m × 20 m的样方, 合计36个样方, 各样方的基本情况如
Decoupled leaf and stem economics in rain forest trees
1
2010
... 植物在其漫长的历史进化和发展过程中, 受物理、生理等因素的综合作用和影响, 功能性状之间往往会呈现出一定的协变和权衡关系(
海南岛热带低地雨林群落水平植物功能性状与环境因子相关性随演替阶段的变化
1
2013
... 选取叶面积(LA)、比叶面积(SLA)和木材密度(WD) 3个重要功能性状进行测量.其中, LA (cm2)反映叶片与外界环境接触面积的大小, 其会影响到植物体与外界环境的气体、能量等交换的平衡, 一般生在阴蔽、湿润、资源丰富地区的植物具有较大的叶片(
海南岛热带低地雨林群落水平植物功能性状与环境因子相关性随演替阶段的变化
1
2013
... 选取叶面积(LA)、比叶面积(SLA)和木材密度(WD) 3个重要功能性状进行测量.其中, LA (cm2)反映叶片与外界环境接触面积的大小, 其会影响到植物体与外界环境的气体、能量等交换的平衡, 一般生在阴蔽、湿润、资源丰富地区的植物具有较大的叶片(
古田山植物功能性状的系统发育信号、不同性状之间的关系及其对群落结构的影响
0
2014
古田山植物功能性状的系统发育信号、不同性状之间的关系及其对群落结构的影响
0
2014
A handbook of protocols for standardized and easy measurement of plant functional traits worldwide
1
2003
... 选取叶面积(LA)、比叶面积(SLA)和木材密度(WD) 3个重要功能性状进行测量.其中, LA (cm2)反映叶片与外界环境接触面积的大小, 其会影响到植物体与外界环境的气体、能量等交换的平衡, 一般生在阴蔽、湿润、资源丰富地区的植物具有较大的叶片(
A trait-based test for habitat filtering: Convex hull volume
1
2006
... WD与LA两性状在物种水平和β组分相关性存在显著差异(p<0.01), 但α组分相关性较弱且零模型的变化趋势与观测值变化一致.造成这一格局的原因可能是, 群落环境梯度导致优势物种从喜阳的落叶灌木物种(大LA, 小WD)逐渐变为耐阴的常绿乔木(小LA, 大WD), 反映在性状上就是这两个性状的β组分呈明显的负相关关系.而在群落内部, 不同的物种产生不同的性状组合来分化生态位(
Covariation in leaf and root traits for native and non-native grasses along an altitudinal gradient in New Zealand
1
2003
... 选取叶面积(LA)、比叶面积(SLA)和木材密度(WD) 3个重要功能性状进行测量.其中, LA (cm2)反映叶片与外界环境接触面积的大小, 其会影响到植物体与外界环境的气体、能量等交换的平衡, 一般生在阴蔽、湿润、资源丰富地区的植物具有较大的叶片(
The evolution of the worldwide leaf economics spectrum
0
2011
1960~2010年漓江流域降水变化特征研究
1
2014
... 漓江处于广西壮族自治区东北部, 位于珠江流域西江水系一级支流——桂江的上游河段, 地理坐标为109.75°-110.67° E, 24.30°-25.68° N.根据降水、地形、径流等特征, 可将漓江划分为上游、中游、下游3个河段, 其中桂林水文站至源头为上游, 桂林水文站至阳朔水文站为中游, 阳朔水文站至恭城河口为下游(
1960~2010年漓江流域降水变化特征研究
1
2014
... 漓江处于广西壮族自治区东北部, 位于珠江流域西江水系一级支流——桂江的上游河段, 地理坐标为109.75°-110.67° E, 24.30°-25.68° N.根据降水、地形、径流等特征, 可将漓江划分为上游、中游、下游3个河段, 其中桂林水文站至源头为上游, 桂林水文站至阳朔水文站为中游, 阳朔水文站至恭城河口为下游(
Shifts in trait-combinations along rainfall and phosphorus gradients
1
2000
... 植物在其漫长的历史进化和发展过程中, 受物理、生理等因素的综合作用和影响, 功能性状之间往往会呈现出一定的协变和权衡关系(
Beyond neutrality—Ecology finds its niche
0
2006
Trait convergence and trait divergence in herbaceous plant communities: Mechanisms and consequences
0
2006
A reassessment of the strategies of plants which cope with shortages of resources
1
1998
... 植物在其漫长的历史进化和发展过程中, 受物理、生理等因素的综合作用和影响, 功能性状之间往往会呈现出一定的协变和权衡关系(
河岸带生态学研究进展与展望
0
2013
河岸带生态学研究进展与展望
0
2013
Novel evidence for within-species leaf economics spectrum at multiple spatial scales
1
2015
... 植物在其漫长的历史进化和发展过程中, 受物理、生理等因素的综合作用和影响, 功能性状之间往往会呈现出一定的协变和权衡关系(
漓江水陆交错带植物叶性状对水淹胁迫的响应及经济谱分析
0
2017
漓江水陆交错带植物叶性状对水淹胁迫的响应及经济谱分析
0
2017
漓江沿岸水陆交错带景观调查与生态修复技术
0
2013
漓江沿岸水陆交错带景观调查与生态修复技术
0
2013
Intraspecific variability and trait-based community assembly
1
2010
... 采用零模型的方法计算随机期望下的不同性状$\alpha$/$\beta$组分协变强度和不同物种$\beta$组分的离散度.其中, $\alpha$/$\beta$组分协变强度的零模型是根据
Functional traits and niche-based tree community, assembly in an Amazonian forest
1
2008
... WD与LA两性状在物种水平和β组分相关性存在显著差异(p<0.01), 但α组分相关性较弱且零模型的变化趋势与观测值变化一致.造成这一格局的原因可能是, 群落环境梯度导致优势物种从喜阳的落叶灌木物种(大LA, 小WD)逐渐变为耐阴的常绿乔木(小LA, 大WD), 反映在性状上就是这两个性状的β组分呈明显的负相关关系.而在群落内部, 不同的物种产生不同的性状组合来分化生态位(
Plant functional traits have globally consistent effects on competition
1
2016
... WD与LA两性状在物种水平和β组分相关性存在显著差异(p<0.01), 但α组分相关性较弱且零模型的变化趋势与观测值变化一致.造成这一格局的原因可能是, 群落环境梯度导致优势物种从喜阳的落叶灌木物种(大LA, 小WD)逐渐变为耐阴的常绿乔木(小LA, 大WD), 反映在性状上就是这两个性状的β组分呈明显的负相关关系.而在群落内部, 不同的物种产生不同的性状组合来分化生态位(
漓江水陆交错带典型立地根系分布与土壤性质的关系
1
2014
... 漓江处于广西壮族自治区东北部, 位于珠江流域西江水系一级支流——桂江的上游河段, 地理坐标为109.75°-110.67° E, 24.30°-25.68° N.根据降水、地形、径流等特征, 可将漓江划分为上游、中游、下游3个河段, 其中桂林水文站至源头为上游, 桂林水文站至阳朔水文站为中游, 阳朔水文站至恭城河口为下游(
漓江水陆交错带典型立地根系分布与土壤性质的关系
1
2014
... 漓江处于广西壮族自治区东北部, 位于珠江流域西江水系一级支流——桂江的上游河段, 地理坐标为109.75°-110.67° E, 24.30°-25.68° N.根据降水、地形、径流等特征, 可将漓江划分为上游、中游、下游3个河段, 其中桂林水文站至源头为上游, 桂林水文站至阳朔水文站为中游, 阳朔水文站至恭城河口为下游(
漓江水陆交错带不同淹没区植物多样性与土壤特征
1
2015
... 漓江处于广西壮族自治区东北部, 位于珠江流域西江水系一级支流——桂江的上游河段, 地理坐标为109.75°-110.67° E, 24.30°-25.68° N.根据降水、地形、径流等特征, 可将漓江划分为上游、中游、下游3个河段, 其中桂林水文站至源头为上游, 桂林水文站至阳朔水文站为中游, 阳朔水文站至恭城河口为下游(
漓江水陆交错带不同淹没区植物多样性与土壤特征
1
2015
... 漓江处于广西壮族自治区东北部, 位于珠江流域西江水系一级支流——桂江的上游河段, 地理坐标为109.75°-110.67° E, 24.30°-25.68° N.根据降水、地形、径流等特征, 可将漓江划分为上游、中游、下游3个河段, 其中桂林水文站至源头为上游, 桂林水文站至阳朔水文站为中游, 阳朔水文站至恭城河口为下游(
历史上漓江(桂江)水系名称的变化浅议
1
2003
... 漓江处于广西壮族自治区东北部, 位于珠江流域西江水系一级支流——桂江的上游河段, 地理坐标为109.75°-110.67° E, 24.30°-25.68° N.根据降水、地形、径流等特征, 可将漓江划分为上游、中游、下游3个河段, 其中桂林水文站至源头为上游, 桂林水文站至阳朔水文站为中游, 阳朔水文站至恭城河口为下游(
历史上漓江(桂江)水系名称的变化浅议
1
2003
... 漓江处于广西壮族自治区东北部, 位于珠江流域西江水系一级支流——桂江的上游河段, 地理坐标为109.75°-110.67° E, 24.30°-25.68° N.根据降水、地形、径流等特征, 可将漓江划分为上游、中游、下游3个河段, 其中桂林水文站至源头为上游, 桂林水文站至阳朔水文站为中游, 阳朔水文站至恭城河口为下游(
植物功能性状研究进展
0
2015
植物功能性状研究进展
0
2015
Rebuilding community ecology from functional traits
0
2006
植物功能性状与环境和生态系统功能
0
2007
植物功能性状与环境和生态系统功能
0
2007
Alterations of riparian ecosystems caused by river regulation
0
2000
Structured interspecific determinants of specific leaf area in 34 species of herbaceous angiosperms
1
1995
... 植物在其漫长的历史进化和发展过程中, 受物理、生理等因素的综合作用和影响, 功能性状之间往往会呈现出一定的协变和权衡关系(
Evidence for limiting similarity in a sand dune community
0
2004
Testing the Holy Grail framework: Using functional traits to predict ecosystem change
0
2008
三峡水库澎溪河消落带植物群落物种丰富度格局
1
2010
... 河岸带植物群落的生态特征受空间位置、干扰强度、气候、地形、土壤养分以及淹水的深度、持续时间和频率等环境因子的显著影响, 呈现随着环境梯度变化其梯度变化同样明显的特征(
三峡水库澎溪河消落带植物群落物种丰富度格局
1
2010
... 河岸带植物群落的生态特征受空间位置、干扰强度、气候、地形、土壤养分以及淹水的深度、持续时间和频率等环境因子的显著影响, 呈现随着环境梯度变化其梯度变化同样明显的特征(
The leaf size-twig size spectrum of temperate woody species along an altitudinal gradient: An invariant allometric scaling relationship
1
2006
... 选取叶面积(LA)、比叶面积(SLA)和木材密度(WD) 3个重要功能性状进行测量.其中, LA (cm2)反映叶片与外界环境接触面积的大小, 其会影响到植物体与外界环境的气体、能量等交换的平衡, 一般生在阴蔽、湿润、资源丰富地区的植物具有较大的叶片(
The river continuum concept
1
1980
... 河岸带植物群落的生态特征受空间位置、干扰强度、气候、地形、土壤养分以及淹水的深度、持续时间和频率等环境因子的显著影响, 呈现随着环境梯度变化其梯度变化同样明显的特征(
Relationships among ecologically important dimensions of plant trait variation in seven Neotropical forests
1
2007
... 植物在其漫长的历史进化和发展过程中, 受物理、生理等因素的综合作用和影响, 功能性状之间往往会呈现出一定的协变和权衡关系(
黔中喀斯特山区植物功能性状的变异与关联
1
2011
... 植物在其漫长的历史进化和发展过程中, 受物理、生理等因素的综合作用和影响, 功能性状之间往往会呈现出一定的协变和权衡关系(
黔中喀斯特山区植物功能性状的变异与关联
1
2011
... 植物在其漫长的历史进化和发展过程中, 受物理、生理等因素的综合作用和影响, 功能性状之间往往会呈现出一定的协变和权衡关系(
广西桂林漓江河岸带植被配置类型与退化机制研究
1
2014
... 漓江处于广西壮族自治区东北部, 位于珠江流域西江水系一级支流——桂江的上游河段, 地理坐标为109.75°-110.67° E, 24.30°-25.68° N.根据降水、地形、径流等特征, 可将漓江划分为上游、中游、下游3个河段, 其中桂林水文站至源头为上游, 桂林水文站至阳朔水文站为中游, 阳朔水文站至恭城河口为下游(
广西桂林漓江河岸带植被配置类型与退化机制研究
1
2014
... 漓江处于广西壮族自治区东北部, 位于珠江流域西江水系一级支流——桂江的上游河段, 地理坐标为109.75°-110.67° E, 24.30°-25.68° N.根据降水、地形、径流等特征, 可将漓江划分为上游、中游、下游3个河段, 其中桂林水文站至源头为上游, 桂林水文站至阳朔水文站为中游, 阳朔水文站至恭城河口为下游(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}