Effects of forest canopy condition on the establishment of Castanopsis fargesii seedlings in a subtropical evergreen broad-leaved forest
WU Xiao-Qi,, YANG Sheng-He, HUANG Li, LI Xiao-Han, YANG Chao, QIAN Shen-Hua, YANG Yong-Chuan,,*Key Lab of Three Gorges Reservoir Region’s Eco-Environment, Ministry of Education, Chongqing University, Chongqing 400045, China
Supported by the National Research Basic Work Special Project.2015FY210200-16 Chongqing Technology Innovation and Application Demonstration Major Theme Special Project.cstc2018jszx-zdyfxmX0007 the Natural Science Foundation of Chongqing (CSTC2016jcyjA0379)..CSTC2016jcyjA0379
Abstract Aims The establishment of seedlings is a key life-history stage during the regeneration of plant populations. This study is to investigate the effects of forest canopy condition (large-sized gap: >150 m 2, medium-sized gap: 100-150 m 2, small-sized gap: 50-100 m 2, and understory as control) on the establishment of Castanopsis fargesii seedlings, and to guide in situ management and restoration of subtropical evergreen broad-leaved forests. Methods We conducted a 3-year in situ seed sowing experiment in the Jinyun Mountain National Nature Reserve under different canopy conditions, and monitored the growth and survival of C. fargesii seedlings for three years (2014-2016). Important findings We found that: (1) the emergence of the seedlings lasted from July to December, and the emergence of seedlings was delayed under large canopy gaps; (2) overall germination rate was (62.8 ± 2.0)%. The averaged survival rate during three growing seasons was (65.1 ± 2.2)%, with wither being the key type of seedling mortality; (3) there was no significant difference in the germination and survival rates under different canopy conditions in 2014, but significant difference in the survival rates in 2015 and 2016; (4) seedling growth among canopy conditions showed no difference in 2014, but distinct differences in 2015 and 2016. The seedling growth in large- and medium-sized gaps was significantly promoted compared with those growing under closed canopies in terms of total biomass, plant height, base diameter, root length and leaf number (except for the specific leaf area); (5) for each of the four types of canopy conditions, leaf mass ratio (LMR) and stem mass ratio (SMR) of seedlings growing increased within last three years, while root mass ratio (RMR) and root shoot ratio (RSR) decreased. The LMR in large-sized gaps was significantly higher than those in closed canopies, but not for the RMR and RSR. The results suggested that the initial survival and growth of C. fargesii seedlings may be supported by the nutrients stored in seeds, and thus less affected by canopy conditions. However, in late-establishment stages (i.e., the second and third years), gap size could become a key factor in affecting both the seedling survival and growth. And large- and medium-sized gaps could promote the seedling establishment success for C. fargesii. Keywords:canopy condition;seed germination;seedling establishment;growth;Castanopsis fargesii
PDF (1332KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 吴小琪, 杨圣贺, 黄力, 李笑寒, 杨超, 钱深华, 杨永川. 常绿阔叶林林冠环境对栲幼苗建成的影响. 植物生态学报[J], 2019, 43(1): 55-64 doi:10.17521/cjpe.2018.0275 WU Xiao-Qi. Effects of forest canopy condition on the establishment of Castanopsis fargesii seedlings in a subtropical evergreen broad-leaved forest. Chinese Journal of Plant Ecology[J], 2019, 43(1): 55-64 doi:10.17521/cjpe.2018.0275
Fig. 3Temporal dynamics of Castanopsis fargesii seedling survival rates (mean ± SE) under different canopy conditions. Different lowercase letters showed significant differences among seedling survival rate under different canopy conditions (p < 0.05).
Fig. 4Ratio of the causes of Castanopsis fargesii seedling death under different canopy conditions (mean ± SE). Different lowercase letters showed significant differences among the causes of seedling death under same canopy condition (p < 0.05).
新窗口打开|下载原图ZIP|生成PPT 图5不同林冠环境下栲幼苗生长指标的年际动态(平均值±标准误差)。*表示林冠环境对该阶段幼苗生长指标有着显著影响(**, p < 0.01; ***, p < 0.001)。
Fig. 5The interannual variation of the growth indicators for Castanopsis fargesii seedling under different canopy conditions (mean ± SE). * indicates a significant canopy effect on a specific seedling growth indicator at a given stage (**, p < 0.01; ***, p < 0.001).
新窗口打开|下载原图ZIP|生成PPT 图6不同林冠环境下栲幼苗生物量分配的年际动态(平均值±标准误差)。*表示林冠环境对该阶段幼苗生物量分配有着显著影响(**, p < 0.01; ***, p < 0.001)。
Fig. 6The interannual variation of the biomass allocation for Castanopsis fargesii seedlings under different canopy conditions (mean ± SE). LMR, leaf mass ratio; SMR, stem mass ratio; RMR, root mass ratio; RSR, root shoot ratio. * indicates a significant canopy effect on seedling biomass allocation at a given stage (**, p < 0.01; ***, p < 0.001).
AbeM, MiguchiH, NakashizukaT ( 2001). An interactive effect of simultaneous death of dwarf bamboo, canopy gap, and predatory rodents on beech regeneration Oecologia, 127, 281-286. DOI:10.1007/s004420000585URL [本文引用: 1]
BonalD, BurbanB, StahlC, WagnerF, HéraultB ( 2016). The response of tropical rainforests to drought—Lessons from recent research and future prospects Annals of Forest Science, 73, 27-44. DOI:10.1007/s13595-015-0522-5 [本文引用: 1]
ChanthornW, CaughlinT ( 2013). The relative importance of fungal infection, conspecific density and environmental heterogeneity for seedling survival in a dominant topical tree Biotropica, 45, 587-593. DOI:10.1111/btp.2013.45.issue-5URL [本文引用: 1]
ChenB, DaLJ, SongYC ( 2002). Seed germination and seedling growth of Castanopsis fargesii in evergreen broadleaved forest and in gaps. Journal of Tropical and Subtropical Botany, 10, 207-214. [本文引用: 2]
ChenGP, XianJR, CaoXW, LiuJQ, YangYH, LeiW ( 2016). Effects of canopy gap on the survival dynamics of Abies faxoniana seedlings in a subalpine coniferous forest. Acta Ecologica Sinica, 36, 6475-6486. [本文引用: 1]
ChenSB, SongAQ, LiZJ ( 2005). Research advance in response of forest seedling regeneration to light environmental heterogeneity Chinese Journal of Applied Ecology, 16, 365-370. [本文引用: 1]
ChmuraDJ, ModrzyńskiJ, ChmielarzP, TjoelkerMG ( 2017). Plasticity in seedling morphology, biomass allocation and physiology among ten temperate tree species in response to shade is related to shade tolerance and not leaf habit Plant Biology, 19, 172-182. DOI:10.1111/plb.12531URL [本文引用: 1]
ClarkCJ, PoulsenJR, LeveyDJ ( 2013). Roles of seed and establishment limitation in determining patterns of afrotropical tree recruitment PLOS ONE, 8, e63330. DOI: 10.1371/journal.pone.0063330. URL [本文引用: 1]
ComitaLS, HubbellSP ( 2009). Local neighborhood and species’ shade tolerance influence survival in a diverse seedling bank Ecology, 90, 328-334. DOI:10.1890/08-0451.1URL [本文引用: 1]
DingSY, LuXL ( 2007). Comparing ecophysiological characteristics of Castanopsis fargesii seedlings and saplings under different disturbance regimes: A case study of the Tiantong evergreen broad-leaved forest, Zhejiang Province of China. Acta Ecological Sinica, 27, 3892-3900. [本文引用: 1]
DongH, YangLX ( 2018). Effects of soil in larch plantations on the growth of Picea koraiensis and P. crassifolia seedlings. Chinese Journal of Applied Ecology, 29, 3199-3205. [本文引用: 1]
DonohueK, CasasRRD, BurghardtL, KovachK, WillisCG ( 2010). Germination, postgermination adaptation, and species ecological ranges Annual Review of Ecology & Systematics, 41, 293-319. [本文引用: 1]
DuYJ, PengSJ, XuGL, HuangZL, HuangYJ ( 2007). Study of distance-dependence on Castanopsis Chinensis seed in coniferous and broad-leaved mixed forest of Dinghushan, China. Journal of Plant Ecology (Chinese Version), 31, 998-1006. [本文引用: 1]
EngelbrechtBMJ, DallingJW, PearsonTRH, WolfRL, GálvezDA, KoehlerT, TyreeMT, KursarTA ( 2006). Short dry spells in the wet season increase mortality of tropical pioneer seedlings Oecologia, 148, 258-269. DOI:10.1007/s00442-006-0368-5URL [本文引用: 1]
FengDL, ZhangLN, HuangZH, ChenJH, ZhuXL ( 2013). Effects of gap size on the seed germination and seedling growth of Castanopsis carlesii and Castanopsis fargesii. Journal Northwest A&F University (Nature Science), 41(6), 61-66. [本文引用: 1]
FigueroaJA, LuskCH ( 2001). Germination requirements and seedling shade tolerance are not correlated in a Chilean temperate rain forest New Phytologist, 152, 483-489. DOI:10.1046/j.0028-646X.2001.00282.xURL [本文引用: 2]
GaoXY ( 2015). Castanopsis fargesii Decline and the Effect on Seedling Recruitment on Mt. Jinyun. Master degree dissertation, Chongqing University, Chongqing. [本文引用: 2]
GaoXY, HuangL, YangC, YangYC, YuanXZ ( 2016). Seed germination characteristics of the dominate tree Castanopsis fargesii in evergreen broadleaved forest on Mt. Jinyun. Journal of Chongqing Normal University (Nature Science), 33(2), 127-133. [本文引用: 2]
GongHD, YangGP, LuZY, LiuYH, CaoM ( 2011). Composition and spatio-temporal distribution of tree seedlings in an evergreen broad-leaved forest in the Ailao Mountain, Yunnan Biodiversity Science, 19, 151-157. [本文引用: 1]
GremerJR, KimballS, VenableDL ( 2016). Within- and among-year germination in Sonoran Desert winter annuals: Bet hedging and predictive germination in a variable environment Ecology Letters, 19, 1209-1218. DOI:10.1111/ele.12655URL [本文引用: 2]
HouZJ, ZhaoCZ, LiY, ZhangQ, MaXL ( 2014). Trade-off between height and branch numbers in Stellera chamaejasme on slopes of different aspects in a degraded alpine grassland. Chinese Journal of Plant Ecology, 38, 281-288. [本文引用: 1]
JinY, RussoSE, YuM ( 2018). Effects of light and topography on regeneration and coexistence of evergreen and deciduous tree species in a Chinese subtropical forest Journal of Ecology, 106, 1634-1645. DOI:10.1111/jec.2018.106.issue-4URL [本文引用: 1]
LarsonJE, SheleyRL, HardegreeSP, DoescherPS, JamesJJ ( 2015). Seed and seedling traits affecting critical life stage transitions and recruitment outcomes in dryland grasses Journal of Applied Ecology, 52, 199-209. DOI:10.1111/1365-2664.12350URL [本文引用: 1]
LarsonJE, SheleyRL, HardegreeSP, DoescherPS, JamesJJ ( 2016). Do key dimensions of seed and seedling functional trait variation capture variation in recruitment probability? Oecologia, 181, 39-53. DOI:10.1007/s00442-015-3430-3URL [本文引用: 1]
LiQK, MaKP ( 2003). Factors affecting establishment of Quercus liaotungensis Koidz. under mature mixed oak forest overstory and in shrubland. Forest Ecology & Management, 176, 133-146. [本文引用: 1]
LiYN, LiDS, XuZQ, YuXF ( 2014). The process of emergence and height growth of Qucercus species in different provenances. Journal of Northwest Forestry University, 29(4), 139-144. [本文引用: 1]
LuJM, JohnsonDJ, QiaoXJ, LuZJ, WangQG, JiangMX ( 2015). Density dependence and habitat preference shape seedling survival in a subtropical forest in central China Journal of Plant Ecology, 8, 568-577. [本文引用: 1]
MaLW, ZhangWH, ZhouJY, XueYQ, WuM, MaC ( 2013). Effects of forest gap size on the growth of Quercus variabilis seedlings on north slopes of the Qinling Mountains. Scientia Silvae Sinicae, 49(12), 43-50. DOI:10.11707/j.1001-7488.20131207Magsci [本文引用: 1] <p>栓皮栎是中国暖温带和亚热带林区的主要建群种和资源树种。为了明确林窗大小对栓皮栎实生幼苗生长发育的影响,通过设置固定样地,测定秦岭北坡5种不同面积林窗(面积分别为<50,50~100,100~150,150~200,>200 m<sup>2</sup>)中栓皮栎实生幼苗的存活数量、生长、干物质积累和分配等指标,分析栓皮栎实生幼苗的生长发育与环境因子之间的关系。结果表明:在幼苗期(1~5年生),随着年龄的增长,实生幼苗的数量呈现逐渐下降的趋势;在面积为150~200 m<sup>2</sup>的林窗中,栓皮栎实生幼苗在数量、生长指标和干物质积累量方面都优于其他林窗中的栓皮栎实生幼苗;林窗内的光照强度、灌木层盖度、草本层盖度、草本层高度、土壤温度、土壤养分条件与实生幼苗的存活呈正相关;空气湿度、枯枝落叶层厚度则与实生幼苗的生长呈负相关。在未来的经营过程中,抚育间伐创造面积为150~200 m<sup>2</sup>的林窗,林分郁闭度保持在0.75左右,能改善林地生境,促进种子萌芽和实生幼苗生长发育,为培育高质量的林木提供有利条件。</p> [ 马莉薇, 张文辉, 周建云, 薛瑶芹, 吴敏, 马闯 ( 2013). 秦岭北坡林窗大小对栓皮栎实生幼苗生长发育的影响 林业科学, 49(12), 43-50.] DOI:10.11707/j.1001-7488.20131207Magsci [本文引用: 1] <p>栓皮栎是中国暖温带和亚热带林区的主要建群种和资源树种。为了明确林窗大小对栓皮栎实生幼苗生长发育的影响,通过设置固定样地,测定秦岭北坡5种不同面积林窗(面积分别为<50,50~100,100~150,150~200,>200 m<sup>2</sup>)中栓皮栎实生幼苗的存活数量、生长、干物质积累和分配等指标,分析栓皮栎实生幼苗的生长发育与环境因子之间的关系。结果表明:在幼苗期(1~5年生),随着年龄的增长,实生幼苗的数量呈现逐渐下降的趋势;在面积为150~200 m<sup>2</sup>的林窗中,栓皮栎实生幼苗在数量、生长指标和干物质积累量方面都优于其他林窗中的栓皮栎实生幼苗;林窗内的光照强度、灌木层盖度、草本层盖度、草本层高度、土壤温度、土壤养分条件与实生幼苗的存活呈正相关;空气湿度、枯枝落叶层厚度则与实生幼苗的生长呈负相关。在未来的经营过程中,抚育间伐创造面积为150~200 m<sup>2</sup>的林窗,林分郁闭度保持在0.75左右,能改善林地生境,促进种子萌芽和实生幼苗生长发育,为培育高质量的林木提供有利条件。</p>
MoralesJ, SqueoFA, TracolY, ArmasC, GutiérrezJR ( 2015). Resource economics and coordination among above- and below-ground functional traits of three dominant shrubs from the Chilean coastal desert Journal of Plant Ecology, 8, 70-78. DOI:10.1093/jpe/rtu010URL [本文引用: 1]
OuJD, WuZZ, LuoN ( 2016). Effects of forest gap size on the growth and form quality of Taxus wallichina var. mairei in Cunninghamia lanceolata forest. Chinese Journal of Applied Ecology, 27, 3098– 3104. [本文引用: 1]
PiiroinenT, ValtonenA, RoininenH ( 2017). The seed-to-seedling transition is limited by ground vegetation and vertebrate herbivores in a selectively logged rainforest Forest Ecology & Management, 384, 137-146. [本文引用: 1]
ReddyKR, BrandD, WijewardanaC, GaoW ( 2017). Temperature effects on cotton seedling emergence, growth, and development Agronomy Journal, 109, 1379-1387. DOI:10.2134/agronj2016.07.0439URL [本文引用: 1]
TangJM, ZhaiMP ( 2006). Morphological response and biomass allocation of Schima superba seedlings at different habitats in gap. Journal of Huazhong Agricultural University, 25, 559-563. [本文引用: 1]
TianD, LiP, FangWJ, XuJ, LuoYK, YanZB, ZhuB, WangJJ, XuXN, FangJY ( 2017). Growth responses of trees and understory plants to nitrogen fertilization in a subtropical forest in China Biogeosciences, 14, 3461-3469. DOI:10.5194/bg-14-3461-2017URL [本文引用: 1]
WangBC, SmithTB ( 2002). Closing the seed dispersal loop Trends in Ecology & Evolution, 17, 379-386. [本文引用: 1]
XingZL, ZhangWH, HuangCZ, YuSC, ZhouJY ( 2017). Effects of forest gap size on the aboveground architecture of Pinus tabulaeformis seedlings. Acta Botanica Boreali-?Occidentalia Sinica, 37, 578-585. [本文引用: 1]
YanXF, CaoM ( 2008). Effects of forest gap on the seedling growth of a canopy tree species Pometia tomentosa in tropical rainforest of Xishuangbanna. Chinese Journal of Applied Ecology, 19, 238-244. [本文引用: 1]
YangYC, HuangL, QianSH, FukudaKJ ( 2015). Completing the life history of Castanopsis fargesii: Changes in the seed dispersal, seedling and sapling recruitment patterns. European Journal of Forest Research, 134, 1143-1154. [本文引用: 6]
ZhangK, BaskinJM, BaskinCC, YangX, HuangZ ( 2017). Effect of seed morph and light level on growth and reproduction of the amphicarpic plant Amphicarpaea edgeworthii(Fabaceae). Scientific Reports, 7, 39886. [本文引用: 1]
ZhengYL, SunWB, ZhouY, CoombsD ( 2009). Variation in seed and seedling traits among natural populations of Trigonobalanus doichangensis( A. Camus) Forman (Fagaceae), a rare and endangered plant in southwest China. New Forests, 37, 285-294. [本文引用: 1]
ZhouJH, ChengXR, YuMK, ZhugeJJ ( 2015). Responses of growth, leaf traits and biomass allocation of Phoebe sheareri seedlings to different habitats in forest gap. Journal of Northeast Forestry University, 43(12), 21-24, 50. [本文引用: 1]
ZhuJJ, GondaY, YuLZ, LiFQ, YanQL, SunYR ( 2012). Regeneration of a coastal pine (Pinus thunbergii Parl.) forest 11 years after thinning, Niigata, Japan. PLOS ONE, 7, e47593. DOI: 10.1371/journal.pone.0047593. [本文引用: 1]
ZhuJJ, MatsuzakiT, LeeFQ, GondaY ( 2003). Effect of gap size created by thinning on seedling emergency, survival and establishment in a coastal pine forest Forest Ecology & Management, 182, 339-354. [本文引用: 4]
An interactive effect of simultaneous death of dwarf bamboo, canopy gap, and predatory rodents on beech regeneration 1 2001
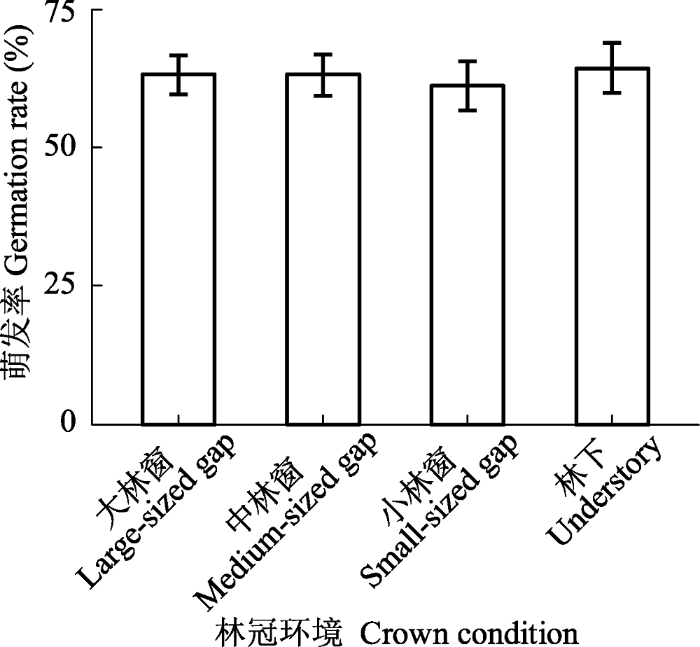
... 种子萌发是植物离开相对安全的种子阶段进入高度脆弱的幼苗期的关键过渡时段(Gremer et al., 2016), 林冠干扰所导致的林冠环境差异对植物种子萌发过程影响的研究相对较少.本研究发现, 栲种子在野外的平均萌发率为(62.8 ± 2.0)%, 略低于室内萌发率(75.1%, 高祥阳等, 2016), 但比壳斗科的大部分物种萌发率高(Abe et al., 2001; Li & Ma, 2003; Zheng et al., 2009).种子的野外萌发率是反映植物生殖潜力的关键性状, 栲种子较高的野外萌发率是维持种群自然更新和在群落中优势地位的基础.此外, 我们发现, 林冠环境对栲种子的萌发率无显著影响(图2), 与陈波等(2002)的研究结果一致.栲种子质量较大, 其萌发率主要取决于种子自身营养物质的含量, 而受光异质性影响较弱(Figueroa & Lusk, 2001; 陈圣宾等, 2005). ...
The response of tropical rainforests to drought—Lessons from recent research and future prospects 1 2016
The relative importance of fungal infection, conspecific density and environmental heterogeneity for seedling survival in a dominant topical tree 1 2013
... 冠层干扰是驱动森林中物种自然更新的一个决定性因素.冠层干扰所导致的林冠环境的差异直接影响着其下生境的环境条件(比如光照强度、土壤温湿度、土壤养分、地表稳定性、凋落物厚度、病原微生物)、资源组合方式以及资源的有效性(Zhu et al., 2003; 欧建德等, 2016; Chmura et al., 2017; Reddy et al., 2017; 董慧和杨立学, 2018), 从而对植物幼苗建成过程产生影响(Zhu et al., 2003, 2012; Zhang et al., 2017).相对而言, 林冠环境对植物种 子萌发阶段, 包括种子萌发率、萌发时间和萌发进程影响的相关研究较少, 尽管该阶段对后续幼苗命运、生长以及资源竞争都具有重要的间接作用(Donohue et al., 2010).相比种子萌发阶段, 木本植物在幼苗阶段受到的林冠环境影响更强, 相关研究也相对广泛(Figueroa & Lusk, 2001).林冠环境对植物幼苗的作用主要表现为对其存活率、生物量积累和生物量分配策略产生影响, 但对不同植物物种影响程度和方式有所差异.比如林冠干扰引起的光环境的差异会影响油松(Pinus tabuliformis)、栓皮栎(Quercus variabilis)、岷江冷杉(Abies fargesii var. faxoniana)和小红栲(Castanopsis carlesii)等物种幼苗的存活率和生长(冯大兰等, 2013; 马莉薇等, 2013; 陈国鹏等, 2016; 邢忠利等, 2017), 并且可使番龙眼(Pometia pinnata)、紫楠(Phoebe sheareri)和木荷(Schima superba)等幼苗生物量分配发生变化(汤景明和翟明普, 2006; 闫兴富和曹敏, 2008; 周俊宏等, 2015).尽管如此, 目前对植物幼苗建成过程中生物量累积动态和生物量分配方式出现分化的时间的研究总体较少, 尤其对具有一定耐阴性质的演替后期种. ...
林窗对岷江冷杉幼苗生存过程的影响 1 2016
... 冠层干扰是驱动森林中物种自然更新的一个决定性因素.冠层干扰所导致的林冠环境的差异直接影响着其下生境的环境条件(比如光照强度、土壤温湿度、土壤养分、地表稳定性、凋落物厚度、病原微生物)、资源组合方式以及资源的有效性(Zhu et al., 2003; 欧建德等, 2016; Chmura et al., 2017; Reddy et al., 2017; 董慧和杨立学, 2018), 从而对植物幼苗建成过程产生影响(Zhu et al., 2003, 2012; Zhang et al., 2017).相对而言, 林冠环境对植物种 子萌发阶段, 包括种子萌发率、萌发时间和萌发进程影响的相关研究较少, 尽管该阶段对后续幼苗命运、生长以及资源竞争都具有重要的间接作用(Donohue et al., 2010).相比种子萌发阶段, 木本植物在幼苗阶段受到的林冠环境影响更强, 相关研究也相对广泛(Figueroa & Lusk, 2001).林冠环境对植物幼苗的作用主要表现为对其存活率、生物量积累和生物量分配策略产生影响, 但对不同植物物种影响程度和方式有所差异.比如林冠干扰引起的光环境的差异会影响油松(Pinus tabuliformis)、栓皮栎(Quercus variabilis)、岷江冷杉(Abies fargesii var. faxoniana)和小红栲(Castanopsis carlesii)等物种幼苗的存活率和生长(冯大兰等, 2013; 马莉薇等, 2013; 陈国鹏等, 2016; 邢忠利等, 2017), 并且可使番龙眼(Pometia pinnata)、紫楠(Phoebe sheareri)和木荷(Schima superba)等幼苗生物量分配发生变化(汤景明和翟明普, 2006; 闫兴富和曹敏, 2008; 周俊宏等, 2015).尽管如此, 目前对植物幼苗建成过程中生物量累积动态和生物量分配方式出现分化的时间的研究总体较少, 尤其对具有一定耐阴性质的演替后期种. ...
森林幼苗更新对光环境异质性的响应研究进展 1 2005
... 种子萌发是植物离开相对安全的种子阶段进入高度脆弱的幼苗期的关键过渡时段(Gremer et al., 2016), 林冠干扰所导致的林冠环境差异对植物种子萌发过程影响的研究相对较少.本研究发现, 栲种子在野外的平均萌发率为(62.8 ± 2.0)%, 略低于室内萌发率(75.1%, 高祥阳等, 2016), 但比壳斗科的大部分物种萌发率高(Abe et al., 2001; Li & Ma, 2003; Zheng et al., 2009).种子的野外萌发率是反映植物生殖潜力的关键性状, 栲种子较高的野外萌发率是维持种群自然更新和在群落中优势地位的基础.此外, 我们发现, 林冠环境对栲种子的萌发率无显著影响(图2), 与陈波等(2002)的研究结果一致.栲种子质量较大, 其萌发率主要取决于种子自身营养物质的含量, 而受光异质性影响较弱(Figueroa & Lusk, 2001; 陈圣宾等, 2005). ...
森林幼苗更新对光环境异质性的响应研究进展 1 2005
... 种子萌发是植物离开相对安全的种子阶段进入高度脆弱的幼苗期的关键过渡时段(Gremer et al., 2016), 林冠干扰所导致的林冠环境差异对植物种子萌发过程影响的研究相对较少.本研究发现, 栲种子在野外的平均萌发率为(62.8 ± 2.0)%, 略低于室内萌发率(75.1%, 高祥阳等, 2016), 但比壳斗科的大部分物种萌发率高(Abe et al., 2001; Li & Ma, 2003; Zheng et al., 2009).种子的野外萌发率是反映植物生殖潜力的关键性状, 栲种子较高的野外萌发率是维持种群自然更新和在群落中优势地位的基础.此外, 我们发现, 林冠环境对栲种子的萌发率无显著影响(图2), 与陈波等(2002)的研究结果一致.栲种子质量较大, 其萌发率主要取决于种子自身营养物质的含量, 而受光异质性影响较弱(Figueroa & Lusk, 2001; 陈圣宾等, 2005). ...
Plasticity in seedling morphology, biomass allocation and physiology among ten temperate tree species in response to shade is related to shade tolerance and not leaf habit 1 2017
... 冠层干扰是驱动森林中物种自然更新的一个决定性因素.冠层干扰所导致的林冠环境的差异直接影响着其下生境的环境条件(比如光照强度、土壤温湿度、土壤养分、地表稳定性、凋落物厚度、病原微生物)、资源组合方式以及资源的有效性(Zhu et al., 2003; 欧建德等, 2016; Chmura et al., 2017; Reddy et al., 2017; 董慧和杨立学, 2018), 从而对植物幼苗建成过程产生影响(Zhu et al., 2003, 2012; Zhang et al., 2017).相对而言, 林冠环境对植物种 子萌发阶段, 包括种子萌发率、萌发时间和萌发进程影响的相关研究较少, 尽管该阶段对后续幼苗命运、生长以及资源竞争都具有重要的间接作用(Donohue et al., 2010).相比种子萌发阶段, 木本植物在幼苗阶段受到的林冠环境影响更强, 相关研究也相对广泛(Figueroa & Lusk, 2001).林冠环境对植物幼苗的作用主要表现为对其存活率、生物量积累和生物量分配策略产生影响, 但对不同植物物种影响程度和方式有所差异.比如林冠干扰引起的光环境的差异会影响油松(Pinus tabuliformis)、栓皮栎(Quercus variabilis)、岷江冷杉(Abies fargesii var. faxoniana)和小红栲(Castanopsis carlesii)等物种幼苗的存活率和生长(冯大兰等, 2013; 马莉薇等, 2013; 陈国鹏等, 2016; 邢忠利等, 2017), 并且可使番龙眼(Pometia pinnata)、紫楠(Phoebe sheareri)和木荷(Schima superba)等幼苗生物量分配发生变化(汤景明和翟明普, 2006; 闫兴富和曹敏, 2008; 周俊宏等, 2015).尽管如此, 目前对植物幼苗建成过程中生物量累积动态和生物量分配方式出现分化的时间的研究总体较少, 尤其对具有一定耐阴性质的演替后期种. ...
Roles of seed and establishment limitation in determining patterns of afrotropical tree recruitment 1 2013
Effects of light and topography on regeneration and coexistence of evergreen and deciduous tree species in a Chinese subtropical forest 1 2018
... 亚热带常绿阔叶林生物多样性丰富, 是仅次于热带雨林的一种具有大生物量和高生产力的生态系统, 在保护区域生态环境、实现人类可持续发展等方面均具有十分重要的生态价值(宋永昌, 2013).亚热带常绿阔叶林的建群种多为具有一定耐阴性质的演替后期种, 比如栲(Castanopsis fargesii)、润楠(Machilus nanmu)和短刺米槠(Castanopsis carlesii var. spinulosa)等, 该类森林的自然更新依赖于冠层干扰(巩合德等, 2011; Yang et al., 2015; Jin et al., 2018).近年来, 随着人类活动的增加, 常绿阔叶林出现生境破碎化和优势种衰退等问题(宋永昌, 2013), 导致林冠环境的剧烈变化, 该类森林的管理和恢复日益受到重视, 而其有效性应当建立在对常绿阔叶林优势种自然更新过程深入认识的基础上.因此, 研究冠层干扰所导致的环境异质性对常绿阔叶林优势种建成过程的影响能够为对该类森林的管理、保护和恢复提供科学参考. ...
Seed and seedling traits affecting critical life stage transitions and recruitment outcomes in dryland grasses 1 2015
... 植物幼苗建成阶段包括种子萌发、出苗、幼苗存活和生长等过程, 该阶段是植物种群自然更新的关键生活史阶段(Wang & Smith, 2002; Yang et al., 2015), 并决定着群落物种组成和相对多度(Larson et al., 2015).深入了解植物种群幼苗增补过程中种子和幼苗命运的影响因素和异质生境中幼苗生长和生物量分配策略, 一直以来是植物种群和群落生态学研究的重点和热点, 具有重要的理论和实践意义(Yang et al., 2015; Larson et al., 2016; Piiroinen et al., 2017). ...
Do key dimensions of seed and seedling functional trait variation capture variation in recruitment probability? 1 2016
... 植物幼苗建成阶段包括种子萌发、出苗、幼苗存活和生长等过程, 该阶段是植物种群自然更新的关键生活史阶段(Wang & Smith, 2002; Yang et al., 2015), 并决定着群落物种组成和相对多度(Larson et al., 2015).深入了解植物种群幼苗增补过程中种子和幼苗命运的影响因素和异质生境中幼苗生长和生物量分配策略, 一直以来是植物种群和群落生态学研究的重点和热点, 具有重要的理论和实践意义(Yang et al., 2015; Larson et al., 2016; Piiroinen et al., 2017). ...
Factors affecting establishment of Quercus liaotungensis Koidz. under mature mixed oak forest overstory and in shrubland. 1 2003
... 种子萌发是植物离开相对安全的种子阶段进入高度脆弱的幼苗期的关键过渡时段(Gremer et al., 2016), 林冠干扰所导致的林冠环境差异对植物种子萌发过程影响的研究相对较少.本研究发现, 栲种子在野外的平均萌发率为(62.8 ± 2.0)%, 略低于室内萌发率(75.1%, 高祥阳等, 2016), 但比壳斗科的大部分物种萌发率高(Abe et al., 2001; Li & Ma, 2003; Zheng et al., 2009).种子的野外萌发率是反映植物生殖潜力的关键性状, 栲种子较高的野外萌发率是维持种群自然更新和在群落中优势地位的基础.此外, 我们发现, 林冠环境对栲种子的萌发率无显著影响(图2), 与陈波等(2002)的研究结果一致.栲种子质量较大, 其萌发率主要取决于种子自身营养物质的含量, 而受光异质性影响较弱(Figueroa & Lusk, 2001; 陈圣宾等, 2005). ...
... 冠层干扰是驱动森林中物种自然更新的一个决定性因素.冠层干扰所导致的林冠环境的差异直接影响着其下生境的环境条件(比如光照强度、土壤温湿度、土壤养分、地表稳定性、凋落物厚度、病原微生物)、资源组合方式以及资源的有效性(Zhu et al., 2003; 欧建德等, 2016; Chmura et al., 2017; Reddy et al., 2017; 董慧和杨立学, 2018), 从而对植物幼苗建成过程产生影响(Zhu et al., 2003, 2012; Zhang et al., 2017).相对而言, 林冠环境对植物种 子萌发阶段, 包括种子萌发率、萌发时间和萌发进程影响的相关研究较少, 尽管该阶段对后续幼苗命运、生长以及资源竞争都具有重要的间接作用(Donohue et al., 2010).相比种子萌发阶段, 木本植物在幼苗阶段受到的林冠环境影响更强, 相关研究也相对广泛(Figueroa & Lusk, 2001).林冠环境对植物幼苗的作用主要表现为对其存活率、生物量积累和生物量分配策略产生影响, 但对不同植物物种影响程度和方式有所差异.比如林冠干扰引起的光环境的差异会影响油松(Pinus tabuliformis)、栓皮栎(Quercus variabilis)、岷江冷杉(Abies fargesii var. faxoniana)和小红栲(Castanopsis carlesii)等物种幼苗的存活率和生长(冯大兰等, 2013; 马莉薇等, 2013; 陈国鹏等, 2016; 邢忠利等, 2017), 并且可使番龙眼(Pometia pinnata)、紫楠(Phoebe sheareri)和木荷(Schima superba)等幼苗生物量分配发生变化(汤景明和翟明普, 2006; 闫兴富和曹敏, 2008; 周俊宏等, 2015).尽管如此, 目前对植物幼苗建成过程中生物量累积动态和生物量分配方式出现分化的时间的研究总体较少, 尤其对具有一定耐阴性质的演替后期种. ...
林窗大小对杉木林内南方红豆杉生长与形质的影响 1 2016
... 冠层干扰是驱动森林中物种自然更新的一个决定性因素.冠层干扰所导致的林冠环境的差异直接影响着其下生境的环境条件(比如光照强度、土壤温湿度、土壤养分、地表稳定性、凋落物厚度、病原微生物)、资源组合方式以及资源的有效性(Zhu et al., 2003; 欧建德等, 2016; Chmura et al., 2017; Reddy et al., 2017; 董慧和杨立学, 2018), 从而对植物幼苗建成过程产生影响(Zhu et al., 2003, 2012; Zhang et al., 2017).相对而言, 林冠环境对植物种 子萌发阶段, 包括种子萌发率、萌发时间和萌发进程影响的相关研究较少, 尽管该阶段对后续幼苗命运、生长以及资源竞争都具有重要的间接作用(Donohue et al., 2010).相比种子萌发阶段, 木本植物在幼苗阶段受到的林冠环境影响更强, 相关研究也相对广泛(Figueroa & Lusk, 2001).林冠环境对植物幼苗的作用主要表现为对其存活率、生物量积累和生物量分配策略产生影响, 但对不同植物物种影响程度和方式有所差异.比如林冠干扰引起的光环境的差异会影响油松(Pinus tabuliformis)、栓皮栎(Quercus variabilis)、岷江冷杉(Abies fargesii var. faxoniana)和小红栲(Castanopsis carlesii)等物种幼苗的存活率和生长(冯大兰等, 2013; 马莉薇等, 2013; 陈国鹏等, 2016; 邢忠利等, 2017), 并且可使番龙眼(Pometia pinnata)、紫楠(Phoebe sheareri)和木荷(Schima superba)等幼苗生物量分配发生变化(汤景明和翟明普, 2006; 闫兴富和曹敏, 2008; 周俊宏等, 2015).尽管如此, 目前对植物幼苗建成过程中生物量累积动态和生物量分配方式出现分化的时间的研究总体较少, 尤其对具有一定耐阴性质的演替后期种. ...
The seed-to-seedling transition is limited by ground vegetation and vertebrate herbivores in a selectively logged rainforest 1 2017
... 植物幼苗建成阶段包括种子萌发、出苗、幼苗存活和生长等过程, 该阶段是植物种群自然更新的关键生活史阶段(Wang & Smith, 2002; Yang et al., 2015), 并决定着群落物种组成和相对多度(Larson et al., 2015).深入了解植物种群幼苗增补过程中种子和幼苗命运的影响因素和异质生境中幼苗生长和生物量分配策略, 一直以来是植物种群和群落生态学研究的重点和热点, 具有重要的理论和实践意义(Yang et al., 2015; Larson et al., 2016; Piiroinen et al., 2017). ...
Temperature effects on cotton seedling emergence, growth, and development 1 2017
... 冠层干扰是驱动森林中物种自然更新的一个决定性因素.冠层干扰所导致的林冠环境的差异直接影响着其下生境的环境条件(比如光照强度、土壤温湿度、土壤养分、地表稳定性、凋落物厚度、病原微生物)、资源组合方式以及资源的有效性(Zhu et al., 2003; 欧建德等, 2016; Chmura et al., 2017; Reddy et al., 2017; 董慧和杨立学, 2018), 从而对植物幼苗建成过程产生影响(Zhu et al., 2003, 2012; Zhang et al., 2017).相对而言, 林冠环境对植物种 子萌发阶段, 包括种子萌发率、萌发时间和萌发进程影响的相关研究较少, 尽管该阶段对后续幼苗命运、生长以及资源竞争都具有重要的间接作用(Donohue et al., 2010).相比种子萌发阶段, 木本植物在幼苗阶段受到的林冠环境影响更强, 相关研究也相对广泛(Figueroa & Lusk, 2001).林冠环境对植物幼苗的作用主要表现为对其存活率、生物量积累和生物量分配策略产生影响, 但对不同植物物种影响程度和方式有所差异.比如林冠干扰引起的光环境的差异会影响油松(Pinus tabuliformis)、栓皮栎(Quercus variabilis)、岷江冷杉(Abies fargesii var. faxoniana)和小红栲(Castanopsis carlesii)等物种幼苗的存活率和生长(冯大兰等, 2013; 马莉薇等, 2013; 陈国鹏等, 2016; 邢忠利等, 2017), 并且可使番龙眼(Pometia pinnata)、紫楠(Phoebe sheareri)和木荷(Schima superba)等幼苗生物量分配发生变化(汤景明和翟明普, 2006; 闫兴富和曹敏, 2008; 周俊宏等, 2015).尽管如此, 目前对植物幼苗建成过程中生物量累积动态和生物量分配方式出现分化的时间的研究总体较少, 尤其对具有一定耐阴性质的演替后期种. ...
2 2013
... 亚热带常绿阔叶林生物多样性丰富, 是仅次于热带雨林的一种具有大生物量和高生产力的生态系统, 在保护区域生态环境、实现人类可持续发展等方面均具有十分重要的生态价值(宋永昌, 2013).亚热带常绿阔叶林的建群种多为具有一定耐阴性质的演替后期种, 比如栲(Castanopsis fargesii)、润楠(Machilus nanmu)和短刺米槠(Castanopsis carlesii var. spinulosa)等, 该类森林的自然更新依赖于冠层干扰(巩合德等, 2011; Yang et al., 2015; Jin et al., 2018).近年来, 随着人类活动的增加, 常绿阔叶林出现生境破碎化和优势种衰退等问题(宋永昌, 2013), 导致林冠环境的剧烈变化, 该类森林的管理和恢复日益受到重视, 而其有效性应当建立在对常绿阔叶林优势种自然更新过程深入认识的基础上.因此, 研究冠层干扰所导致的环境异质性对常绿阔叶林优势种建成过程的影响能够为对该类森林的管理、保护和恢复提供科学参考. ...
... 植物幼苗建成阶段包括种子萌发、出苗、幼苗存活和生长等过程, 该阶段是植物种群自然更新的关键生活史阶段(Wang & Smith, 2002; Yang et al., 2015), 并决定着群落物种组成和相对多度(Larson et al., 2015).深入了解植物种群幼苗增补过程中种子和幼苗命运的影响因素和异质生境中幼苗生长和生物量分配策略, 一直以来是植物种群和群落生态学研究的重点和热点, 具有重要的理论和实践意义(Yang et al., 2015; Larson et al., 2016; Piiroinen et al., 2017). ...
林窗大小对油松实生苗地上部分构型的影响 1 2017
... 冠层干扰是驱动森林中物种自然更新的一个决定性因素.冠层干扰所导致的林冠环境的差异直接影响着其下生境的环境条件(比如光照强度、土壤温湿度、土壤养分、地表稳定性、凋落物厚度、病原微生物)、资源组合方式以及资源的有效性(Zhu et al., 2003; 欧建德等, 2016; Chmura et al., 2017; Reddy et al., 2017; 董慧和杨立学, 2018), 从而对植物幼苗建成过程产生影响(Zhu et al., 2003, 2012; Zhang et al., 2017).相对而言, 林冠环境对植物种 子萌发阶段, 包括种子萌发率、萌发时间和萌发进程影响的相关研究较少, 尽管该阶段对后续幼苗命运、生长以及资源竞争都具有重要的间接作用(Donohue et al., 2010).相比种子萌发阶段, 木本植物在幼苗阶段受到的林冠环境影响更强, 相关研究也相对广泛(Figueroa & Lusk, 2001).林冠环境对植物幼苗的作用主要表现为对其存活率、生物量积累和生物量分配策略产生影响, 但对不同植物物种影响程度和方式有所差异.比如林冠干扰引起的光环境的差异会影响油松(Pinus tabuliformis)、栓皮栎(Quercus variabilis)、岷江冷杉(Abies fargesii var. faxoniana)和小红栲(Castanopsis carlesii)等物种幼苗的存活率和生长(冯大兰等, 2013; 马莉薇等, 2013; 陈国鹏等, 2016; 邢忠利等, 2017), 并且可使番龙眼(Pometia pinnata)、紫楠(Phoebe sheareri)和木荷(Schima superba)等幼苗生物量分配发生变化(汤景明和翟明普, 2006; 闫兴富和曹敏, 2008; 周俊宏等, 2015).尽管如此, 目前对植物幼苗建成过程中生物量累积动态和生物量分配方式出现分化的时间的研究总体较少, 尤其对具有一定耐阴性质的演替后期种. ...
林窗大小对油松实生苗地上部分构型的影响 1 2017
... 冠层干扰是驱动森林中物种自然更新的一个决定性因素.冠层干扰所导致的林冠环境的差异直接影响着其下生境的环境条件(比如光照强度、土壤温湿度、土壤养分、地表稳定性、凋落物厚度、病原微生物)、资源组合方式以及资源的有效性(Zhu et al., 2003; 欧建德等, 2016; Chmura et al., 2017; Reddy et al., 2017; 董慧和杨立学, 2018), 从而对植物幼苗建成过程产生影响(Zhu et al., 2003, 2012; Zhang et al., 2017).相对而言, 林冠环境对植物种 子萌发阶段, 包括种子萌发率、萌发时间和萌发进程影响的相关研究较少, 尽管该阶段对后续幼苗命运、生长以及资源竞争都具有重要的间接作用(Donohue et al., 2010).相比种子萌发阶段, 木本植物在幼苗阶段受到的林冠环境影响更强, 相关研究也相对广泛(Figueroa & Lusk, 2001).林冠环境对植物幼苗的作用主要表现为对其存活率、生物量积累和生物量分配策略产生影响, 但对不同植物物种影响程度和方式有所差异.比如林冠干扰引起的光环境的差异会影响油松(Pinus tabuliformis)、栓皮栎(Quercus variabilis)、岷江冷杉(Abies fargesii var. faxoniana)和小红栲(Castanopsis carlesii)等物种幼苗的存活率和生长(冯大兰等, 2013; 马莉薇等, 2013; 陈国鹏等, 2016; 邢忠利等, 2017), 并且可使番龙眼(Pometia pinnata)、紫楠(Phoebe sheareri)和木荷(Schima superba)等幼苗生物量分配发生变化(汤景明和翟明普, 2006; 闫兴富和曹敏, 2008; 周俊宏等, 2015).尽管如此, 目前对植物幼苗建成过程中生物量累积动态和生物量分配方式出现分化的时间的研究总体较少, 尤其对具有一定耐阴性质的演替后期种. ...
林窗对热带雨林冠层树种绒毛番龙眼幼苗生长的影响 1 2008
... 冠层干扰是驱动森林中物种自然更新的一个决定性因素.冠层干扰所导致的林冠环境的差异直接影响着其下生境的环境条件(比如光照强度、土壤温湿度、土壤养分、地表稳定性、凋落物厚度、病原微生物)、资源组合方式以及资源的有效性(Zhu et al., 2003; 欧建德等, 2016; Chmura et al., 2017; Reddy et al., 2017; 董慧和杨立学, 2018), 从而对植物幼苗建成过程产生影响(Zhu et al., 2003, 2012; Zhang et al., 2017).相对而言, 林冠环境对植物种 子萌发阶段, 包括种子萌发率、萌发时间和萌发进程影响的相关研究较少, 尽管该阶段对后续幼苗命运、生长以及资源竞争都具有重要的间接作用(Donohue et al., 2010).相比种子萌发阶段, 木本植物在幼苗阶段受到的林冠环境影响更强, 相关研究也相对广泛(Figueroa & Lusk, 2001).林冠环境对植物幼苗的作用主要表现为对其存活率、生物量积累和生物量分配策略产生影响, 但对不同植物物种影响程度和方式有所差异.比如林冠干扰引起的光环境的差异会影响油松(Pinus tabuliformis)、栓皮栎(Quercus variabilis)、岷江冷杉(Abies fargesii var. faxoniana)和小红栲(Castanopsis carlesii)等物种幼苗的存活率和生长(冯大兰等, 2013; 马莉薇等, 2013; 陈国鹏等, 2016; 邢忠利等, 2017), 并且可使番龙眼(Pometia pinnata)、紫楠(Phoebe sheareri)和木荷(Schima superba)等幼苗生物量分配发生变化(汤景明和翟明普, 2006; 闫兴富和曹敏, 2008; 周俊宏等, 2015).尽管如此, 目前对植物幼苗建成过程中生物量累积动态和生物量分配方式出现分化的时间的研究总体较少, 尤其对具有一定耐阴性质的演替后期种. ...
林窗对热带雨林冠层树种绒毛番龙眼幼苗生长的影响 1 2008
... 冠层干扰是驱动森林中物种自然更新的一个决定性因素.冠层干扰所导致的林冠环境的差异直接影响着其下生境的环境条件(比如光照强度、土壤温湿度、土壤养分、地表稳定性、凋落物厚度、病原微生物)、资源组合方式以及资源的有效性(Zhu et al., 2003; 欧建德等, 2016; Chmura et al., 2017; Reddy et al., 2017; 董慧和杨立学, 2018), 从而对植物幼苗建成过程产生影响(Zhu et al., 2003, 2012; Zhang et al., 2017).相对而言, 林冠环境对植物种 子萌发阶段, 包括种子萌发率、萌发时间和萌发进程影响的相关研究较少, 尽管该阶段对后续幼苗命运、生长以及资源竞争都具有重要的间接作用(Donohue et al., 2010).相比种子萌发阶段, 木本植物在幼苗阶段受到的林冠环境影响更强, 相关研究也相对广泛(Figueroa & Lusk, 2001).林冠环境对植物幼苗的作用主要表现为对其存活率、生物量积累和生物量分配策略产生影响, 但对不同植物物种影响程度和方式有所差异.比如林冠干扰引起的光环境的差异会影响油松(Pinus tabuliformis)、栓皮栎(Quercus variabilis)、岷江冷杉(Abies fargesii var. faxoniana)和小红栲(Castanopsis carlesii)等物种幼苗的存活率和生长(冯大兰等, 2013; 马莉薇等, 2013; 陈国鹏等, 2016; 邢忠利等, 2017), 并且可使番龙眼(Pometia pinnata)、紫楠(Phoebe sheareri)和木荷(Schima superba)等幼苗生物量分配发生变化(汤景明和翟明普, 2006; 闫兴富和曹敏, 2008; 周俊宏等, 2015).尽管如此, 目前对植物幼苗建成过程中生物量累积动态和生物量分配方式出现分化的时间的研究总体较少, 尤其对具有一定耐阴性质的演替后期种. ...
光照对辽东栎种子萌发和幼苗生长的影响 2 2011
... 种子的萌发时间对幼苗存活和生长都具有重要作用, 并且易受环境因子影响(Gremer et al., 2016).总体上看, 栲种子在野外萌发持续时间较长, 分散的出苗格局避免了偶然因素对新萌发幼苗带来的灾难性影响, 可能是栲等演替后期种的种群更新适应性策略.不同于种子萌发率, 本研究发现林冠环境对栲种子的萌发时间具有明显影响, 林下萌发最快, 并且大林窗对萌发具有迟滞作用, 与同为壳斗科植物的辽东栎(Quercus wutaishanica)相似(闫兴富等, 2011).大林窗中种子的延迟萌发可规避强光照射对新幼苗产生光伤害, 对幼苗在野外的早期生存具有积极作用(Zhu et al., 2003). ...
Completing the life history of Castanopsis fargesii: Changes in the seed dispersal, seedling and sapling recruitment patterns. 6 2015
... 植物幼苗建成阶段包括种子萌发、出苗、幼苗存活和生长等过程, 该阶段是植物种群自然更新的关键生活史阶段(Wang & Smith, 2002; Yang et al., 2015), 并决定着群落物种组成和相对多度(Larson et al., 2015).深入了解植物种群幼苗增补过程中种子和幼苗命运的影响因素和异质生境中幼苗生长和生物量分配策略, 一直以来是植物种群和群落生态学研究的重点和热点, 具有重要的理论和实践意义(Yang et al., 2015; Larson et al., 2016; Piiroinen et al., 2017). ...
... ).深入了解植物种群幼苗增补过程中种子和幼苗命运的影响因素和异质生境中幼苗生长和生物量分配策略, 一直以来是植物种群和群落生态学研究的重点和热点, 具有重要的理论和实践意义(Yang et al., 2015; Larson et al., 2016; Piiroinen et al., 2017). ...
... 亚热带常绿阔叶林生物多样性丰富, 是仅次于热带雨林的一种具有大生物量和高生产力的生态系统, 在保护区域生态环境、实现人类可持续发展等方面均具有十分重要的生态价值(宋永昌, 2013).亚热带常绿阔叶林的建群种多为具有一定耐阴性质的演替后期种, 比如栲(Castanopsis fargesii)、润楠(Machilus nanmu)和短刺米槠(Castanopsis carlesii var. spinulosa)等, 该类森林的自然更新依赖于冠层干扰(巩合德等, 2011; Yang et al., 2015; Jin et al., 2018).近年来, 随着人类活动的增加, 常绿阔叶林出现生境破碎化和优势种衰退等问题(宋永昌, 2013), 导致林冠环境的剧烈变化, 该类森林的管理和恢复日益受到重视, 而其有效性应当建立在对常绿阔叶林优势种自然更新过程深入认识的基础上.因此, 研究冠层干扰所导致的环境异质性对常绿阔叶林优势种建成过程的影响能够为对该类森林的管理、保护和恢复提供科学参考. ...
Effect of seed morph and light level on growth and reproduction of the amphicarpic plant Amphicarpaea edgeworthii(Fabaceae). 1 2017
... 冠层干扰是驱动森林中物种自然更新的一个决定性因素.冠层干扰所导致的林冠环境的差异直接影响着其下生境的环境条件(比如光照强度、土壤温湿度、土壤养分、地表稳定性、凋落物厚度、病原微生物)、资源组合方式以及资源的有效性(Zhu et al., 2003; 欧建德等, 2016; Chmura et al., 2017; Reddy et al., 2017; 董慧和杨立学, 2018), 从而对植物幼苗建成过程产生影响(Zhu et al., 2003, 2012; Zhang et al., 2017).相对而言, 林冠环境对植物种 子萌发阶段, 包括种子萌发率、萌发时间和萌发进程影响的相关研究较少, 尽管该阶段对后续幼苗命运、生长以及资源竞争都具有重要的间接作用(Donohue et al., 2010).相比种子萌发阶段, 木本植物在幼苗阶段受到的林冠环境影响更强, 相关研究也相对广泛(Figueroa & Lusk, 2001).林冠环境对植物幼苗的作用主要表现为对其存活率、生物量积累和生物量分配策略产生影响, 但对不同植物物种影响程度和方式有所差异.比如林冠干扰引起的光环境的差异会影响油松(Pinus tabuliformis)、栓皮栎(Quercus variabilis)、岷江冷杉(Abies fargesii var. faxoniana)和小红栲(Castanopsis carlesii)等物种幼苗的存活率和生长(冯大兰等, 2013; 马莉薇等, 2013; 陈国鹏等, 2016; 邢忠利等, 2017), 并且可使番龙眼(Pometia pinnata)、紫楠(Phoebe sheareri)和木荷(Schima superba)等幼苗生物量分配发生变化(汤景明和翟明普, 2006; 闫兴富和曹敏, 2008; 周俊宏等, 2015).尽管如此, 目前对植物幼苗建成过程中生物量累积动态和生物量分配方式出现分化的时间的研究总体较少, 尤其对具有一定耐阴性质的演替后期种. ...
Variation in seed and seedling traits among natural populations of Trigonobalanus doichangensis( A. Camus) Forman (Fagaceae), a rare and endangered plant in southwest China. 1 2009
... 种子萌发是植物离开相对安全的种子阶段进入高度脆弱的幼苗期的关键过渡时段(Gremer et al., 2016), 林冠干扰所导致的林冠环境差异对植物种子萌发过程影响的研究相对较少.本研究发现, 栲种子在野外的平均萌发率为(62.8 ± 2.0)%, 略低于室内萌发率(75.1%, 高祥阳等, 2016), 但比壳斗科的大部分物种萌发率高(Abe et al., 2001; Li & Ma, 2003; Zheng et al., 2009).种子的野外萌发率是反映植物生殖潜力的关键性状, 栲种子较高的野外萌发率是维持种群自然更新和在群落中优势地位的基础.此外, 我们发现, 林冠环境对栲种子的萌发率无显著影响(图2), 与陈波等(2002)的研究结果一致.栲种子质量较大, 其萌发率主要取决于种子自身营养物质的含量, 而受光异质性影响较弱(Figueroa & Lusk, 2001; 陈圣宾等, 2005). ...
紫楠幼苗生长、叶性状和生物量分配对林窗不同生境的响应 1 2015
... 冠层干扰是驱动森林中物种自然更新的一个决定性因素.冠层干扰所导致的林冠环境的差异直接影响着其下生境的环境条件(比如光照强度、土壤温湿度、土壤养分、地表稳定性、凋落物厚度、病原微生物)、资源组合方式以及资源的有效性(Zhu et al., 2003; 欧建德等, 2016; Chmura et al., 2017; Reddy et al., 2017; 董慧和杨立学, 2018), 从而对植物幼苗建成过程产生影响(Zhu et al., 2003, 2012; Zhang et al., 2017).相对而言, 林冠环境对植物种 子萌发阶段, 包括种子萌发率、萌发时间和萌发进程影响的相关研究较少, 尽管该阶段对后续幼苗命运、生长以及资源竞争都具有重要的间接作用(Donohue et al., 2010).相比种子萌发阶段, 木本植物在幼苗阶段受到的林冠环境影响更强, 相关研究也相对广泛(Figueroa & Lusk, 2001).林冠环境对植物幼苗的作用主要表现为对其存活率、生物量积累和生物量分配策略产生影响, 但对不同植物物种影响程度和方式有所差异.比如林冠干扰引起的光环境的差异会影响油松(Pinus tabuliformis)、栓皮栎(Quercus variabilis)、岷江冷杉(Abies fargesii var. faxoniana)和小红栲(Castanopsis carlesii)等物种幼苗的存活率和生长(冯大兰等, 2013; 马莉薇等, 2013; 陈国鹏等, 2016; 邢忠利等, 2017), 并且可使番龙眼(Pometia pinnata)、紫楠(Phoebe sheareri)和木荷(Schima superba)等幼苗生物量分配发生变化(汤景明和翟明普, 2006; 闫兴富和曹敏, 2008; 周俊宏等, 2015).尽管如此, 目前对植物幼苗建成过程中生物量累积动态和生物量分配方式出现分化的时间的研究总体较少, 尤其对具有一定耐阴性质的演替后期种. ...
紫楠幼苗生长、叶性状和生物量分配对林窗不同生境的响应 1 2015
... 冠层干扰是驱动森林中物种自然更新的一个决定性因素.冠层干扰所导致的林冠环境的差异直接影响着其下生境的环境条件(比如光照强度、土壤温湿度、土壤养分、地表稳定性、凋落物厚度、病原微生物)、资源组合方式以及资源的有效性(Zhu et al., 2003; 欧建德等, 2016; Chmura et al., 2017; Reddy et al., 2017; 董慧和杨立学, 2018), 从而对植物幼苗建成过程产生影响(Zhu et al., 2003, 2012; Zhang et al., 2017).相对而言, 林冠环境对植物种 子萌发阶段, 包括种子萌发率、萌发时间和萌发进程影响的相关研究较少, 尽管该阶段对后续幼苗命运、生长以及资源竞争都具有重要的间接作用(Donohue et al., 2010).相比种子萌发阶段, 木本植物在幼苗阶段受到的林冠环境影响更强, 相关研究也相对广泛(Figueroa & Lusk, 2001).林冠环境对植物幼苗的作用主要表现为对其存活率、生物量积累和生物量分配策略产生影响, 但对不同植物物种影响程度和方式有所差异.比如林冠干扰引起的光环境的差异会影响油松(Pinus tabuliformis)、栓皮栎(Quercus variabilis)、岷江冷杉(Abies fargesii var. faxoniana)和小红栲(Castanopsis carlesii)等物种幼苗的存活率和生长(冯大兰等, 2013; 马莉薇等, 2013; 陈国鹏等, 2016; 邢忠利等, 2017), 并且可使番龙眼(Pometia pinnata)、紫楠(Phoebe sheareri)和木荷(Schima superba)等幼苗生物量分配发生变化(汤景明和翟明普, 2006; 闫兴富和曹敏, 2008; 周俊宏等, 2015).尽管如此, 目前对植物幼苗建成过程中生物量累积动态和生物量分配方式出现分化的时间的研究总体较少, 尤其对具有一定耐阴性质的演替后期种. ...
Regeneration of a coastal pine (Pinus thunbergii Parl.) forest 11 years after thinning, Niigata, Japan. 1 2012
... 冠层干扰是驱动森林中物种自然更新的一个决定性因素.冠层干扰所导致的林冠环境的差异直接影响着其下生境的环境条件(比如光照强度、土壤温湿度、土壤养分、地表稳定性、凋落物厚度、病原微生物)、资源组合方式以及资源的有效性(Zhu et al., 2003; 欧建德等, 2016; Chmura et al., 2017; Reddy et al., 2017; 董慧和杨立学, 2018), 从而对植物幼苗建成过程产生影响(Zhu et al., 2003, 2012; Zhang et al., 2017).相对而言, 林冠环境对植物种 子萌发阶段, 包括种子萌发率、萌发时间和萌发进程影响的相关研究较少, 尽管该阶段对后续幼苗命运、生长以及资源竞争都具有重要的间接作用(Donohue et al., 2010).相比种子萌发阶段, 木本植物在幼苗阶段受到的林冠环境影响更强, 相关研究也相对广泛(Figueroa & Lusk, 2001).林冠环境对植物幼苗的作用主要表现为对其存活率、生物量积累和生物量分配策略产生影响, 但对不同植物物种影响程度和方式有所差异.比如林冠干扰引起的光环境的差异会影响油松(Pinus tabuliformis)、栓皮栎(Quercus variabilis)、岷江冷杉(Abies fargesii var. faxoniana)和小红栲(Castanopsis carlesii)等物种幼苗的存活率和生长(冯大兰等, 2013; 马莉薇等, 2013; 陈国鹏等, 2016; 邢忠利等, 2017), 并且可使番龙眼(Pometia pinnata)、紫楠(Phoebe sheareri)和木荷(Schima superba)等幼苗生物量分配发生变化(汤景明和翟明普, 2006; 闫兴富和曹敏, 2008; 周俊宏等, 2015).尽管如此, 目前对植物幼苗建成过程中生物量累积动态和生物量分配方式出现分化的时间的研究总体较少, 尤其对具有一定耐阴性质的演替后期种. ...
Effect of gap size created by thinning on seedling emergency, survival and establishment in a coastal pine forest 4 2003
... 冠层干扰是驱动森林中物种自然更新的一个决定性因素.冠层干扰所导致的林冠环境的差异直接影响着其下生境的环境条件(比如光照强度、土壤温湿度、土壤养分、地表稳定性、凋落物厚度、病原微生物)、资源组合方式以及资源的有效性(Zhu et al., 2003; 欧建德等, 2016; Chmura et al., 2017; Reddy et al., 2017; 董慧和杨立学, 2018), 从而对植物幼苗建成过程产生影响(Zhu et al., 2003, 2012; Zhang et al., 2017).相对而言, 林冠环境对植物种 子萌发阶段, 包括种子萌发率、萌发时间和萌发进程影响的相关研究较少, 尽管该阶段对后续幼苗命运、生长以及资源竞争都具有重要的间接作用(Donohue et al., 2010).相比种子萌发阶段, 木本植物在幼苗阶段受到的林冠环境影响更强, 相关研究也相对广泛(Figueroa & Lusk, 2001).林冠环境对植物幼苗的作用主要表现为对其存活率、生物量积累和生物量分配策略产生影响, 但对不同植物物种影响程度和方式有所差异.比如林冠干扰引起的光环境的差异会影响油松(Pinus tabuliformis)、栓皮栎(Quercus variabilis)、岷江冷杉(Abies fargesii var. faxoniana)和小红栲(Castanopsis carlesii)等物种幼苗的存活率和生长(冯大兰等, 2013; 马莉薇等, 2013; 陈国鹏等, 2016; 邢忠利等, 2017), 并且可使番龙眼(Pometia pinnata)、紫楠(Phoebe sheareri)和木荷(Schima superba)等幼苗生物量分配发生变化(汤景明和翟明普, 2006; 闫兴富和曹敏, 2008; 周俊宏等, 2015).尽管如此, 目前对植物幼苗建成过程中生物量累积动态和生物量分配方式出现分化的时间的研究总体较少, 尤其对具有一定耐阴性质的演替后期种. ...
 ,, 杨圣贺, 黄力, 李笑寒, 杨超, 钱深华, 杨永川
,, 杨圣贺, 黄力, 李笑寒, 杨超, 钱深华, 杨永川 ,*重庆大学三峡库区生态环境教育部重点实验室, 重庆 400045
,*重庆大学三峡库区生态环境教育部重点实验室, 重庆 400045
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT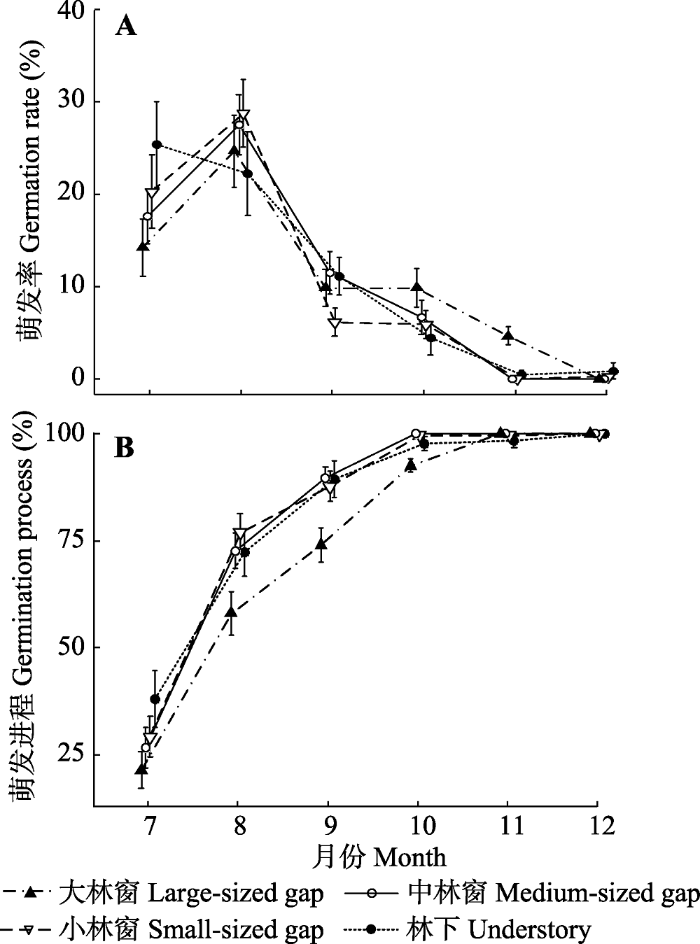 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT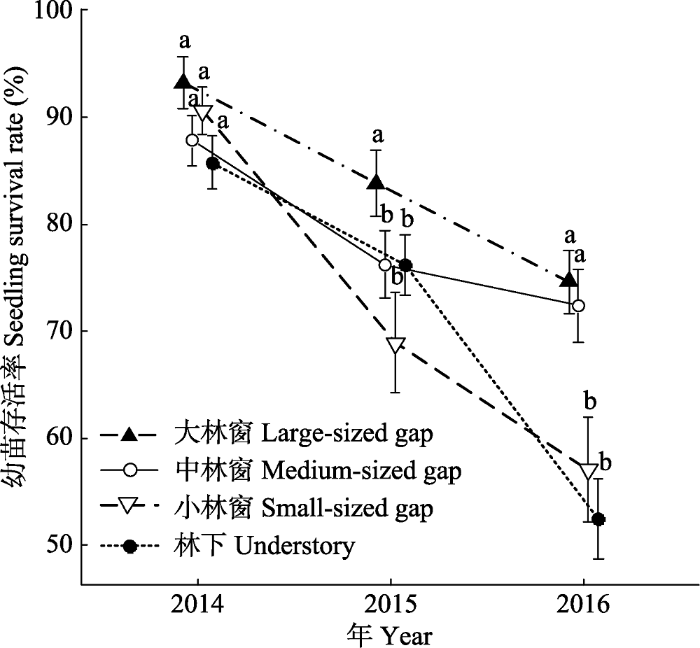 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT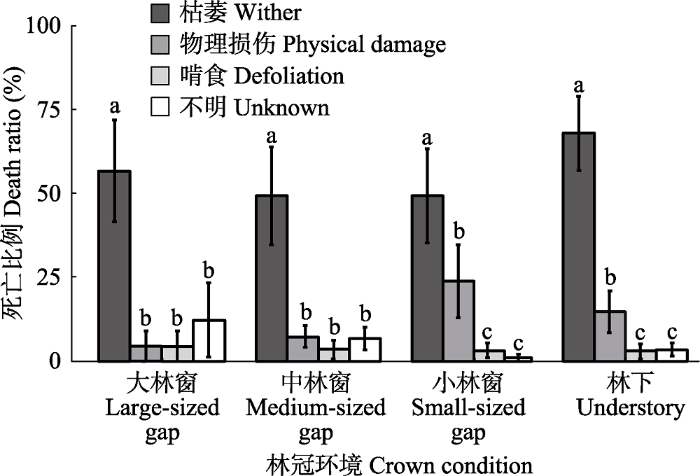 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT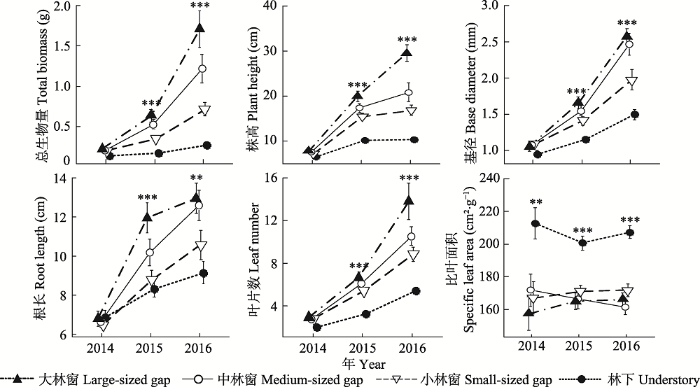 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT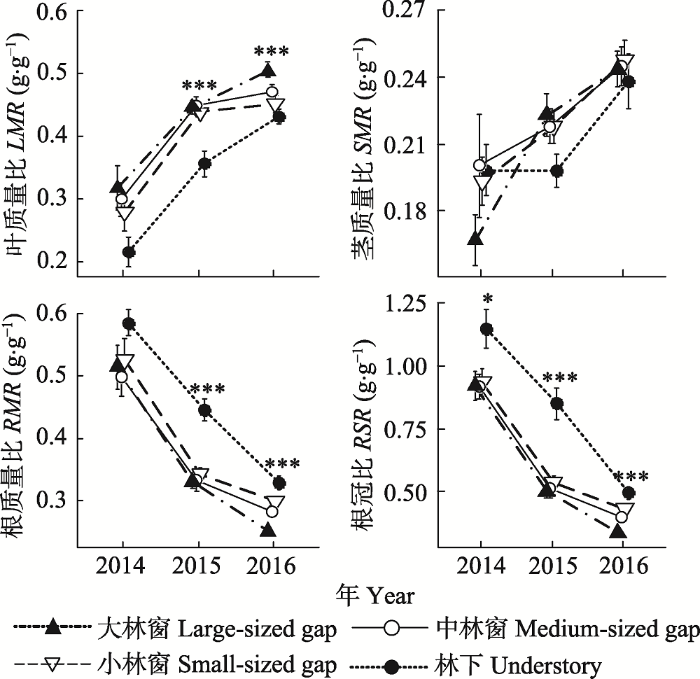 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}