Study on compositions of parenchyma in twigs of woody saplings in Dabie Mountains, Anhui, China
Xi-Jin ZHANG1, Kun SONG,1,2,3,*, Fa-Guang PU4, Zhi-Wen GAO1, Tian-Pin NI1, Xing-Hang CHU1, Ze-Ying WANG1, Kan-Kan SHANG5, Liang-Jun DA,1,2,3,*1 Zhejiang Tiantong Forest Ecosystem National Observation and Research Station, School of Ecological and Environmental Sciences, East China Normal University, Shanghai 200241, China 2 Shanghai Key Lab for Urban Ecological Process and Eco-Restoration, Shanghai 200241, China 3 Institute of Eco-Chongming, Shanghai 200062, China 4 Anhui Dabie Mountain Forest Ecosystem National Observation Station, Jinzhai, Anhui 237354, China 5 Shanghai Chenshan Botanical Garden (Shanghai Chenshan Plant Science Research Centre, Chinese Academy of Sciences), Shanghai 201602, China;
Supported by the National Natural Science Foundation of China(31500355) Supported by the National Natural Science Foundation of China(31670438) Supported by the National Natural Science Foundation of China(31600343)
Abstract Aims Axial parenchyma (AP) and ray parenchyma (RP) in secondary xylem have many important ecological functions, such as storage and translocation. Quantifying the compositions of parenchyma in secondary xylem will benefit the further research about their functions. However, the understanding of parenchyma compositions in current-year branches is still lacking. Methods Eighteen woody saplings in the Tianma National Nature Reserve were selected for sampling. The proportions of cross-sectional area occupied by AP and RP were measured. The variations in parenchyma among species and their phylogenetic signals were analyzed. Compared with the related dataset, the differences between twigs and trunks were tested. Important findings (1) The proportions of total parenchyma in twigs of the 18 woody saplings were 9.96%-18.56%, with the average of 14.80%; the proportions of RP (7.74%-15.45%) were higher than that of AP (1.13%-7.49%). (2) The total parenchyma (RAP) in twigs was lower than that in trunks, of which, RP in twigs was lower than that in trunks while AP showed an opposite pattern. The differences between twigs and trunks may be caused by the differences between different organs and different life history periods. (3) Significant phylogenetic signal was detected in the amount of AP in secondary xylem of twigs. This study primitively verified the phylogenetic signal of the secondary xylem parenchyma, and suggested that the difference between organs, and the difference between life history periods, had important effects on the variations of parenchyma. Keywords:parenchyma;phylogenetic signal;xylem;organ
PDF (1405KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 张希金, 宋坤, 蒲发光, 高志文, 倪田品, 褚兴行, 王泽英, 商侃侃, 达良俊. 安徽大别山木本植物幼树小枝薄壁组织组成特征初探. 植物生态学报, 2019, 43(3): 238-244. DOI: 10.17521/cjpe.2018.0316 ZHANG Xi-Jin, SONG Kun, PU Fa-Guang, GAO Zhi-Wen, NI Tian-Pin, CHU Xing-Hang, WANG Ze-Ying, SHANG Kan-Kan, DA Liang-Jun. Study on compositions of parenchyma in twigs of woody saplings in Dabie Mountains, Anhui, China. Chinese Journal of Plant Ecology, 2019, 43(3): 238-244. DOI: 10.17521/cjpe.2018.0316
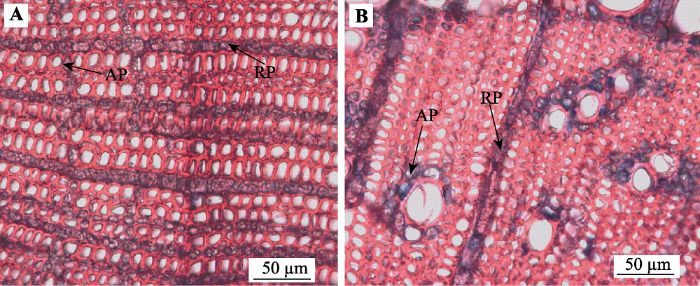
Fig. 1Dyed image of cross sections of twigs photographed by optical microscope. A, Carpinus cordata. B, Schoepfia jasminodora. Ray parenchyma cells arranged radially and were dyed blue to purple, while axial parenchyma cells arranged tangentially or paratracheally in the two pictures.
Table 1 表1 表1大别山木本植物采样信息汇总 Table 1Characteristics of woody samplings from the Dabie Mountains
Fig. 2Differences of the amount of parenchyma in sapling twigs and trunks of adult trees (mean ± SD, n = 8). The data of the amount of parenchyma in trunks were derived from Zheng & Martínez-Cabrera (2013). AP, axial parenchyma; RP, ray parenchyma; RAP, RP + AP. Different lowercase letters indicate significant differences (p < 0.05).
BelAJEV ( 1990). Xylem-phloem exchange via the rays: The undervalued route of transport Journal of Experimental Botany, 41, 631-644. [本文引用: 1]
BhatKM, BhatKV, DhamodaranTK ( 1985). Wood and bark properties of branches of selected tree species growing in Kerala KFRI Research Report, Kerala Forest Research Institute. https://pdfs.semanticscholar.org/6d17/?0853fce456bb543f7e060b3030b435dc9a9f.pdf. Cited: 2018-12-17. [本文引用: 2]
BlombergSP, GarlandT, IvesAR ( 2003). Testing for phylogenetic signal in comparative data: Behavioral traits are more labile Evolution, 57, 717-745. [本文引用: 3]
BurgertI, EcksteinD ( 2001). The tensile strength of isolated wood rays of beech (Fagus sylvatica L.) and its significance for the biomechanics of living trees Trees, 15, 168-170. [本文引用: 1]
CaoK, RaoMD, YuJZ, LiuXJ, MiXC, ChenJH ( 2013). The phylogenetic signal of functional traits and their effects on community structure in an evergreen broad-leaved forest Biodiversity Science, 21, 564-571. [本文引用: 2]
DuY, MaoL, QueenboroughSA, FreckletonRP, ChenB, MaK ( 2015). Phylogenetic constraints and trait correlates of flowering phenology in the angiosperm flora of China Global Ecology and Biogeography, 24, 928-938. [本文引用: 2]
HearnDJ ( 2009). Developmental patterns in anatomy are shared among separate evolutionary origins of stem succulent and storage root-bearing growth habits in Adenia (Passifloraceae) American Journal of Botany, 96, 1941-1956. [本文引用: 3]
HuY, YanL, LiH ( 2006). Studies on the anatomical characteristics of the stems of 14 desert plants Journal of Arid Land Resources and Environment, 20, 204-210. [本文引用: 1]
MorrisH, PlavcováL, CveckoP, FichtlerE, GillinghamMAF, Martínez-CabreraHI, McglinnDJ, WheelerE, ZhengJM, ZiemińskaK, JansenS ( 2016). A global analysis of parenchyma tissue fractions in secondary xylem of seed plants New Phytologist, 209, 1553-1565. [本文引用: 11]
OlanoJM, ArzacA, García-CervigónAI, von ArxG, RozasV ( 2013). New star on the stage: Amount of ray parenchyma in tree rings shows a link to climate New Phytologist, 198, 486-495. [本文引用: 2]
PlavcováL, HochG, MorrisH, GhiasiS, JansenS ( 2016). The amount of parenchyma and living fibers affects storage of nonstructural carbohydrates in young stems and roots of temperate trees American Journal of Botany, 103, 1-10. [本文引用: 5]
PlavcováL, JansenS ( 2015). The role of xylem parenchyma in the storage and utilization of Nonstructural Carbohydrates In: Hacke U ed. Functional and Ecological Xylem Anatomy. Springer, Cham. 209-234. [本文引用: 4]
PoorterL, McdonaldI, AlarcónA, FichtlerE, LiconaJC, Pe?a-ClarosM, SterckF, VillegasZ, Sass-KlaassenU ( 2010). The importance of wood traits and hydraulic conductance for the performance and life history strategies of 42 rainforest species New Phytologist, 185, 481-492. [本文引用: 4]
RungwattanaK, HietzP ( 2018). Radial variation of wood functional traits reflect size-related adaptations of tree mechanics and hydraulics Functional Ecology, 32, 260-272. [本文引用: 1]
SpicerR ( 2014). Symplasmic networks in secondary vascular tissues: parenchyma distribution and activity supporting long-distance transport Journal of Experimental Botany, 65, 1829-1848. [本文引用: 3]
TanHY, HaoBZ, WuJL ( 2000). Seasonal ultrastructural changes of secondary phloem parenchyma cells in a tropical tree, Dalbergia odorifera Acta Botanica Yunnanica, 22, 461-466. [本文引用: 1]
WheelerEA, BaasP, RodgersS ( 2007). Variations in dicot wood anatomy: A global analysis based on the insidewood database Iawa Journal, 28, 229-258. [本文引用: 1]
YangXD, JiPP, ReYS, LiHX ( 2018). Relationship between flowering phenology and phylogeny in 31 woody plants of Urumqi, Xinjiang Acta Ecologica Sinica, 38, 1003-1015. [本文引用: 1]
ZhangHY, WangCK, WangXC ( 2013). Comparison of concentrations of non-structural carbohydrates between new twigs and old branches for 12 temperate species Acta Ecologica Sinica, 33, 5675-5685. [本文引用: 1]
ZhangHY, WangCK, WangXC ( 2015). Within-crown variation in concentrations of non-structural carbohydrates of five temperate tree species Acta Ecologica Sinica, 35, 6496-6506. [本文引用: 1]
ZhengGQ, BaoH, YangJ, SuXL, HuZH ( 2015). Ultrastructure of phloem and the flesh sink-cells during fruit development of Lucium barbarum Acta Botanica Boreali-Occidentalia Sinica, 35, 2211-2218. [本文引用: 1]
ZhengJM, Martínez-CabreraHI ( 2013). Wood anatomical correlates with theoretical conductivity and wood density across China: Evolutionary evidence of the functional differentiation of axial and radial parenchyma Annals of Botany, 112, 927-935. [本文引用: 12]
ZiemińskaK, ButlerDW, GleasonSM, WrightIJ, WestobyM ( 2013). Fibre wall and lumen fractions drive wood density variation across 24 Australian angiosperms AoB Plants , 5, plt046. DOI: 101093/aobpla/plt046. [本文引用: 1]
The tensile strength of isolated wood rays of beech (Fagus sylvatica L.) and its significance for the biomechanics of living trees 1 2001
... 薄壁组织作为木本植物次生木质部的重要组成部分, 其总含量在被子植物中一般为20%-40%, 而在裸子植物中一般仅为5%-10% (Plavcová & Jansen, 2015).薄壁组织依据细胞排列方式和方向可分为射线薄壁组织(RP)和轴向薄壁组织(AP).薄壁组织执行着多种功能, 如物质存储(Plavcová & Jansen, 2015, Plavcová et al., 2016)和转运(Bel, 1990)、防御病原菌(Wheeler et al., 2007)以及机械支持作用(Burgert & Eckstein, 2001)等.木质部薄壁组织与导管、纤维等其他细胞类型存在含量上的权衡关系(Zheng & Martínez-Cabrera, 2013; Morris et al., 2016), 影响着木质部生理生态功能的发挥.目前国外对于木本植物薄壁组织含量、功能及其影响因素已有较多报道(Hearn, 2009; Olano et al., 2013; Spicer, 2014; Morris et al., 2016; Plavcová et al., 2016; Rungwattana & Hietz, 2018), 而国内对木本植物薄壁组织的研究还主要集中在植物器官解剖结构方面(谭海燕等, 2000; 胡云等, 2006; 卫星等, 2008; 郑国琦等, 2015), 但近年来国内也开始加强其含量和功能的研究.如郑国琦等(2010)曾报道宁夏枸杞(Lycium barbarum)根、茎中次生木质部薄壁组织含量与灌水量的关系; Zheng和Martínez- Cabrera (2013)用近800种国内树木的木质部性状(包括薄壁组织含量)数据研究了两种薄壁组织功能分化及其与支持、传导功能的权衡关系; Martínez- Cabrera等(2017)使用Zheng和Martínez-Cabrera (2013)的数据分析了近缘种薄壁组织含量变异性随进化时间的变化(Martínez-Cabrera et al., 2017). ...
古田山木本植物功能性状的系统发育信号及其对群落结构的影响 2 2013
... 虽然以往研究表明环境因素是薄壁组织含量变异的重要影响因素(郑国琦等, 2010; Olano et al., 2013; Morris et al., 2016), 但其含量同时也受系统发育的影响.Spicer (2014)和Morris等(2016)的研究均表明被子植物木质部薄壁组织含量高于针叶植物(Spicer, 2014, Morris et al., 2016).Hearn (2009)对西番莲科蒴莲属(Adenia)的52个物种分析表明薄壁组织含量与维管束协同进化且随进化而增多(Hearn, 2009).这都说明薄壁组织含量与系统发育有关.由于物种进化是影响性状的重要因素, 亲缘关系越近的物种间性状差异越小(即谱系信号), 且谱系信号在自然界中普遍存在(Blomberg et al., 2003; 曹科等, 2013; Du et al., 2015), 因此, 木本植物小枝中薄壁组织含量可能也具有显著的谱系信号. ...
... 本研究小枝中RP和RAP含量未检测到显著的谱系信号, 而AP含量检测到显著的谱系信号, 表明物种亲缘关系越近, 木质部AP含量越相近.RP含量未检测到显著的谱系信号, 可能与其近缘种RP含量差异性在整个进化历史上一直保持较高差异性有关(Martínez-Cabrera et al., 2017).以往研究发现很多木本植物性状(叶性状、木质密度、种子质量、开花物候等)都可检测到显著的谱系信号(曹科等, 2013; Du et al., 2015; 杨晓东等, 2018), 体现了谱系信号在自然界的普遍性. ...
古田山木本植物功能性状的系统发育信号及其对群落结构的影响 2 2013
... 虽然以往研究表明环境因素是薄壁组织含量变异的重要影响因素(郑国琦等, 2010; Olano et al., 2013; Morris et al., 2016), 但其含量同时也受系统发育的影响.Spicer (2014)和Morris等(2016)的研究均表明被子植物木质部薄壁组织含量高于针叶植物(Spicer, 2014, Morris et al., 2016).Hearn (2009)对西番莲科蒴莲属(Adenia)的52个物种分析表明薄壁组织含量与维管束协同进化且随进化而增多(Hearn, 2009).这都说明薄壁组织含量与系统发育有关.由于物种进化是影响性状的重要因素, 亲缘关系越近的物种间性状差异越小(即谱系信号), 且谱系信号在自然界中普遍存在(Blomberg et al., 2003; 曹科等, 2013; Du et al., 2015), 因此, 木本植物小枝中薄壁组织含量可能也具有显著的谱系信号. ...
... 本研究小枝中RP和RAP含量未检测到显著的谱系信号, 而AP含量检测到显著的谱系信号, 表明物种亲缘关系越近, 木质部AP含量越相近.RP含量未检测到显著的谱系信号, 可能与其近缘种RP含量差异性在整个进化历史上一直保持较高差异性有关(Martínez-Cabrera et al., 2017).以往研究发现很多木本植物性状(叶性状、木质密度、种子质量、开花物候等)都可检测到显著的谱系信号(曹科等, 2013; Du et al., 2015; 杨晓东等, 2018), 体现了谱系信号在自然界的普遍性. ...
Phylogenetic constraints and trait correlates of flowering phenology in the angiosperm flora of China 2 2015
... 虽然以往研究表明环境因素是薄壁组织含量变异的重要影响因素(郑国琦等, 2010; Olano et al., 2013; Morris et al., 2016), 但其含量同时也受系统发育的影响.Spicer (2014)和Morris等(2016)的研究均表明被子植物木质部薄壁组织含量高于针叶植物(Spicer, 2014, Morris et al., 2016).Hearn (2009)对西番莲科蒴莲属(Adenia)的52个物种分析表明薄壁组织含量与维管束协同进化且随进化而增多(Hearn, 2009).这都说明薄壁组织含量与系统发育有关.由于物种进化是影响性状的重要因素, 亲缘关系越近的物种间性状差异越小(即谱系信号), 且谱系信号在自然界中普遍存在(Blomberg et al., 2003; 曹科等, 2013; Du et al., 2015), 因此, 木本植物小枝中薄壁组织含量可能也具有显著的谱系信号. ...
... 本研究小枝中RP和RAP含量未检测到显著的谱系信号, 而AP含量检测到显著的谱系信号, 表明物种亲缘关系越近, 木质部AP含量越相近.RP含量未检测到显著的谱系信号, 可能与其近缘种RP含量差异性在整个进化历史上一直保持较高差异性有关(Martínez-Cabrera et al., 2017).以往研究发现很多木本植物性状(叶性状、木质密度、种子质量、开花物候等)都可检测到显著的谱系信号(曹科等, 2013; Du et al., 2015; 杨晓东等, 2018), 体现了谱系信号在自然界的普遍性. ...
Developmental patterns in anatomy are shared among separate evolutionary origins of stem succulent and storage root-bearing growth habits in Adenia (Passifloraceae) 3 2009
... 薄壁组织作为木本植物次生木质部的重要组成部分, 其总含量在被子植物中一般为20%-40%, 而在裸子植物中一般仅为5%-10% (Plavcová & Jansen, 2015).薄壁组织依据细胞排列方式和方向可分为射线薄壁组织(RP)和轴向薄壁组织(AP).薄壁组织执行着多种功能, 如物质存储(Plavcová & Jansen, 2015, Plavcová et al., 2016)和转运(Bel, 1990)、防御病原菌(Wheeler et al., 2007)以及机械支持作用(Burgert & Eckstein, 2001)等.木质部薄壁组织与导管、纤维等其他细胞类型存在含量上的权衡关系(Zheng & Martínez-Cabrera, 2013; Morris et al., 2016), 影响着木质部生理生态功能的发挥.目前国外对于木本植物薄壁组织含量、功能及其影响因素已有较多报道(Hearn, 2009; Olano et al., 2013; Spicer, 2014; Morris et al., 2016; Plavcová et al., 2016; Rungwattana & Hietz, 2018), 而国内对木本植物薄壁组织的研究还主要集中在植物器官解剖结构方面(谭海燕等, 2000; 胡云等, 2006; 卫星等, 2008; 郑国琦等, 2015), 但近年来国内也开始加强其含量和功能的研究.如郑国琦等(2010)曾报道宁夏枸杞(Lycium barbarum)根、茎中次生木质部薄壁组织含量与灌水量的关系; Zheng和Martínez- Cabrera (2013)用近800种国内树木的木质部性状(包括薄壁组织含量)数据研究了两种薄壁组织功能分化及其与支持、传导功能的权衡关系; Martínez- Cabrera等(2017)使用Zheng和Martínez-Cabrera (2013)的数据分析了近缘种薄壁组织含量变异性随进化时间的变化(Martínez-Cabrera et al., 2017). ...
... 虽然以往研究表明环境因素是薄壁组织含量变异的重要影响因素(郑国琦等, 2010; Olano et al., 2013; Morris et al., 2016), 但其含量同时也受系统发育的影响.Spicer (2014)和Morris等(2016)的研究均表明被子植物木质部薄壁组织含量高于针叶植物(Spicer, 2014, Morris et al., 2016).Hearn (2009)对西番莲科蒴莲属(Adenia)的52个物种分析表明薄壁组织含量与维管束协同进化且随进化而增多(Hearn, 2009).这都说明薄壁组织含量与系统发育有关.由于物种进化是影响性状的重要因素, 亲缘关系越近的物种间性状差异越小(即谱系信号), 且谱系信号在自然界中普遍存在(Blomberg et al., 2003; 曹科等, 2013; Du et al., 2015), 因此, 木本植物小枝中薄壁组织含量可能也具有显著的谱系信号. ...
... )的52个物种分析表明薄壁组织含量与维管束协同进化且随进化而增多(Hearn, 2009).这都说明薄壁组织含量与系统发育有关.由于物种进化是影响性状的重要因素, 亲缘关系越近的物种间性状差异越小(即谱系信号), 且谱系信号在自然界中普遍存在(Blomberg et al., 2003; 曹科等, 2013; Du et al., 2015), 因此, 木本植物小枝中薄壁组织含量可能也具有显著的谱系信号. ...
14种荒漠植物茎的解剖结构特征分析 1 2006
... 薄壁组织作为木本植物次生木质部的重要组成部分, 其总含量在被子植物中一般为20%-40%, 而在裸子植物中一般仅为5%-10% (Plavcová & Jansen, 2015).薄壁组织依据细胞排列方式和方向可分为射线薄壁组织(RP)和轴向薄壁组织(AP).薄壁组织执行着多种功能, 如物质存储(Plavcová & Jansen, 2015, Plavcová et al., 2016)和转运(Bel, 1990)、防御病原菌(Wheeler et al., 2007)以及机械支持作用(Burgert & Eckstein, 2001)等.木质部薄壁组织与导管、纤维等其他细胞类型存在含量上的权衡关系(Zheng & Martínez-Cabrera, 2013; Morris et al., 2016), 影响着木质部生理生态功能的发挥.目前国外对于木本植物薄壁组织含量、功能及其影响因素已有较多报道(Hearn, 2009; Olano et al., 2013; Spicer, 2014; Morris et al., 2016; Plavcová et al., 2016; Rungwattana & Hietz, 2018), 而国内对木本植物薄壁组织的研究还主要集中在植物器官解剖结构方面(谭海燕等, 2000; 胡云等, 2006; 卫星等, 2008; 郑国琦等, 2015), 但近年来国内也开始加强其含量和功能的研究.如郑国琦等(2010)曾报道宁夏枸杞(Lycium barbarum)根、茎中次生木质部薄壁组织含量与灌水量的关系; Zheng和Martínez- Cabrera (2013)用近800种国内树木的木质部性状(包括薄壁组织含量)数据研究了两种薄壁组织功能分化及其与支持、传导功能的权衡关系; Martínez- Cabrera等(2017)使用Zheng和Martínez-Cabrera (2013)的数据分析了近缘种薄壁组织含量变异性随进化时间的变化(Martínez-Cabrera et al., 2017). ...
14种荒漠植物茎的解剖结构特征分析 1 2006
... 薄壁组织作为木本植物次生木质部的重要组成部分, 其总含量在被子植物中一般为20%-40%, 而在裸子植物中一般仅为5%-10% (Plavcová & Jansen, 2015).薄壁组织依据细胞排列方式和方向可分为射线薄壁组织(RP)和轴向薄壁组织(AP).薄壁组织执行着多种功能, 如物质存储(Plavcová & Jansen, 2015, Plavcová et al., 2016)和转运(Bel, 1990)、防御病原菌(Wheeler et al., 2007)以及机械支持作用(Burgert & Eckstein, 2001)等.木质部薄壁组织与导管、纤维等其他细胞类型存在含量上的权衡关系(Zheng & Martínez-Cabrera, 2013; Morris et al., 2016), 影响着木质部生理生态功能的发挥.目前国外对于木本植物薄壁组织含量、功能及其影响因素已有较多报道(Hearn, 2009; Olano et al., 2013; Spicer, 2014; Morris et al., 2016; Plavcová et al., 2016; Rungwattana & Hietz, 2018), 而国内对木本植物薄壁组织的研究还主要集中在植物器官解剖结构方面(谭海燕等, 2000; 胡云等, 2006; 卫星等, 2008; 郑国琦等, 2015), 但近年来国内也开始加强其含量和功能的研究.如郑国琦等(2010)曾报道宁夏枸杞(Lycium barbarum)根、茎中次生木质部薄壁组织含量与灌水量的关系; Zheng和Martínez- Cabrera (2013)用近800种国内树木的木质部性状(包括薄壁组织含量)数据研究了两种薄壁组织功能分化及其与支持、传导功能的权衡关系; Martínez- Cabrera等(2017)使用Zheng和Martínez-Cabrera (2013)的数据分析了近缘种薄壁组织含量变异性随进化时间的变化(Martínez-Cabrera et al., 2017). ...
Wood anatomical correlates with theoretical conductivity and wood density across China: Evolutionary evidence of the functional differentiation of axial and radial parenchyma 12 2013
... 薄壁组织作为木本植物次生木质部的重要组成部分, 其总含量在被子植物中一般为20%-40%, 而在裸子植物中一般仅为5%-10% (Plavcová & Jansen, 2015).薄壁组织依据细胞排列方式和方向可分为射线薄壁组织(RP)和轴向薄壁组织(AP).薄壁组织执行着多种功能, 如物质存储(Plavcová & Jansen, 2015, Plavcová et al., 2016)和转运(Bel, 1990)、防御病原菌(Wheeler et al., 2007)以及机械支持作用(Burgert & Eckstein, 2001)等.木质部薄壁组织与导管、纤维等其他细胞类型存在含量上的权衡关系(Zheng & Martínez-Cabrera, 2013; Morris et al., 2016), 影响着木质部生理生态功能的发挥.目前国外对于木本植物薄壁组织含量、功能及其影响因素已有较多报道(Hearn, 2009; Olano et al., 2013; Spicer, 2014; Morris et al., 2016; Plavcová et al., 2016; Rungwattana & Hietz, 2018), 而国内对木本植物薄壁组织的研究还主要集中在植物器官解剖结构方面(谭海燕等, 2000; 胡云等, 2006; 卫星等, 2008; 郑国琦等, 2015), 但近年来国内也开始加强其含量和功能的研究.如郑国琦等(2010)曾报道宁夏枸杞(Lycium barbarum)根、茎中次生木质部薄壁组织含量与灌水量的关系; Zheng和Martínez- Cabrera (2013)用近800种国内树木的木质部性状(包括薄壁组织含量)数据研究了两种薄壁组织功能分化及其与支持、传导功能的权衡关系; Martínez- Cabrera等(2017)使用Zheng和Martínez-Cabrera (2013)的数据分析了近缘种薄壁组织含量变异性随进化时间的变化(Martínez-Cabrera et al., 2017). ...
... 对比Zheng和Martínez-Cabrera (2013)基于树干测算的薄壁组织含量数据, 发现相同物种幼树当年生小枝AP含量高于成熟个体树干(t = 3.15, p < 0.05), RP含量低于树干(t = -2.55, p < 0.05), RAP含量与树干无显著差异(t = -1.82, p > 0.05)(图2). ...
... = 8).树干数据来自Zheng和Martínez-Cabrera (2013).AP, 轴向薄壁组织; RP, 射线薄壁组织; RAP, 总薄壁组织.不同小写字母代表差异显著(p < 0.05). Differences of the amount of parenchyma in sapling twigs and trunks of adult trees (mean ± <i>SD, n</i> = 8). The data of the amount of parenchyma in trunks were derived from <xref ref-type="bibr" rid="b26">Zheng & Martínez-Cabrera (2013)</xref>. AP, axial parenchyma; RP, ray parenchyma; RAP, RP + AP. Different lowercase letters indicate significant differences (<i>p</i> < 0.05).Fig. 22.3 薄壁组织含量与谱系结构的关系 基于本次所研究18个物种, 用Blomberg等(2003)提出的K值法检测了RP、AP和RAP含量的谱系信号强度.结果表明: AP含量检测到显著的谱系信号(p < 0.01), RP和RAP含量均未检测到显著的谱系信号(表3). ...
... = 8). The data of the amount of parenchyma in trunks were derived from Zheng & Martínez-Cabrera (2013). AP, axial parenchyma; RP, ray parenchyma; RAP, RP + AP. Different lowercase letters indicate significant differences (p < 0.05). Fig. 22.3 薄壁组织含量与谱系结构的关系 基于本次所研究18个物种, 用Blomberg等(2003)提出的K值法检测了RP、AP和RAP含量的谱系信号强度.结果表明: AP含量检测到显著的谱系信号(p < 0.01), RP和RAP含量均未检测到显著的谱系信号(表3). ...
 ,1,2,3,*, 蒲发光4, 高志文1, 倪田品1, 褚兴行1, 王泽英1, 商侃侃5, 达良俊
,1,2,3,*, 蒲发光4, 高志文1, 倪田品1, 褚兴行1, 王泽英1, 商侃侃5, 达良俊
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT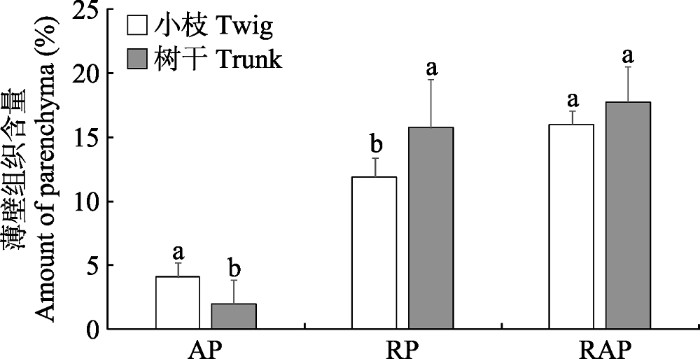 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}