Effects of simulated warming and decomposition interface on the litter decomposition rate of Zizania latifolia and its phyllospheric microbial community structure and function
Peng-Fei YAN1, Peng-Fei ZHAN1, De-Rong XIAO1, Yi WANG2, Rui YU1, Zhen-Ya LIU1, Hang WANG,1,*1 Southwest Forestry University National Plateau Wetlands Research Center/Wetlands College, Kunming 650224, China 2 College of Agronomy and Biotechnology, Yunnan Agricultural University, Kunming 650201, China
Supported by the National Natural Science Foundation of China(41877346) Supported by the National Natural Science Foundation of China(31500409) Supported by the National Natural Science Foundation of China(41867059)
Abstract Aims Litters of emergent plants are important components of material cycling in wetland ecosystems. To clarify the effects of climate warming and habitat difference on the litter decomposition processes and phyllospheric microorganisms of wetland emergent plants is of great significance for revealing the key material cycling processes in wetland ecosystems. MethodsZizania latifolia, a dominant emergent plant in typical wetlands of Northwestern Yunnan Plateau, was chosen for this study. Using litter bag methods, we studied mass remaining and the abundance, community structure and metabolic potential of phyllospheric microorganisms of the litter from Zizania latifolia under simulated warming (1.5-2.0 ℃) and under three habitats (air, water and soil interface). Important findings Simulated climatic warming and habitat difference significantly affected the litter decomposition rate. After one-year decomposition, the mass remaining of litter was 66.4% under the simulated warming treatment, while 77.7% under the control treatment. The decomposition constant (k) value was 1.64 times under warming compared to the control. The mass remaining of litter at the water and soil interface was 42.2% and 25.3%, and the k value at the water and soil interface was 3.63 and 5.25 times of that at the air interface respectively. These results indicate that habitat difference was the key factor controlling the decomposition of emergent plant litter in wetlands. Moreover, warming mainly changed the community composition of litter phyllospheric microorganisms, while decomposition interface mainly affected the abundance, community structure and metabolic potential of phyllospheric microorganisms. Notably, phyllospheric microorganisms of litter at soil interface had the highest metabolic potential and utilized alcohols as main carbon sources. The characteristics of phyllospheric microorganisms between different treatments were in good agreement with litter decomposition rate, which provides an important theoretical basis for revealing the microbial mechanisms driving the decomposition of wetland plant litter. Keywords:wetland ecosystem;litter decomposition;phyllospheric microorganisms;simulated warming;habitat difference
PDF (4044KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 闫鹏飞, 展鹏飞, 肖德荣, 王燚, 余瑞, 刘振亚, 王行. 模拟增温及分解界面对茭草凋落物分解速率及叶际微生物结构和功能的影响. 植物生态学报, 2019, 43(2): 107-118. DOI: 10.17521/cjpe.2018.0272 YAN Peng-Fei, ZHAN Peng-Fei, XIAO De-Rong, WANG Yi, YU Rui, LIU Zhen-Ya, WANG Hang. Effects of simulated warming and decomposition interface on the litter decomposition rate of Zizania latifolia and its phyllospheric microbial community structure and function. Chinese Journal of Plant Ecology, 2019, 43(2): 107-118. DOI: 10.17521/cjpe.2018.0272
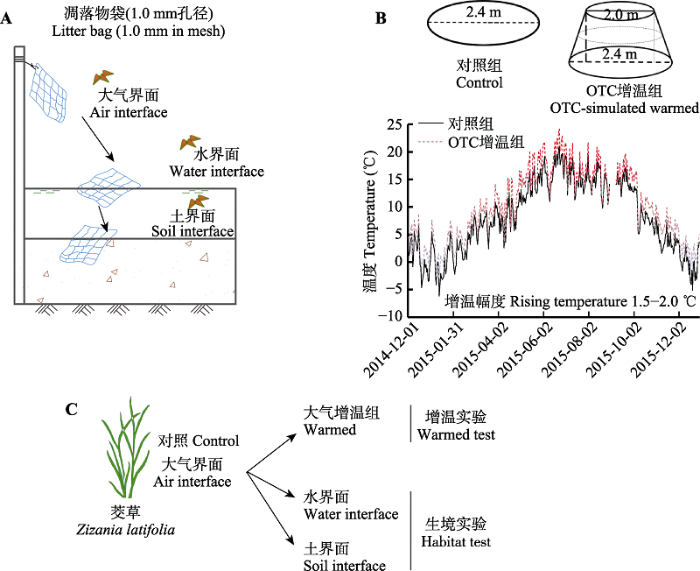
Fig. 1Experiment of simulated warming and habitat difference for litter decomposition of Zizania latifolia. A, Three habitats include air interface, water interface, and soil interface. Among them, litter bags under air decomposition were hang over the bamboo (1.2 m from the ground), litter bags under water decomposition were floated in the surface of water (with the aids of table tennis), and litter bags under soil decomposition were fixed by PVC tubes in the soils (5.0 cm in deep). B, The design and operation of Open-top Chamber (OTC). Among them, control group has no OTC devices, and OTC devices simulate rising temperature (warming group). The device was constructed by solar panels with 2.4 m base and 2.0 m opening in diameter. The temperatures between control and warming groups were recorded from December 2014 to December 2015 (once per hour). In warming treatment, the temperature has been raised by 1.5-2.0 ℃. C, The research object was a typical emergent wetland plant, Zizania latifolia. Its leaf litter was subjected to warming and habitat difference treatments.
新窗口打开|下载原图ZIP|生成PPT 图2茭草叶凋落物质量残留率的季节性动态(平均值±标准误差, n = 3)。利用Post Hoc Tests进行两两比较分析, 不同小写字母代表处理之间的差异达到显著水平(p < 0.05)。
Fig. 2Seasonal dynamics in mass remaining of leaf litter from Zizania latifolia (mean ± SE, n = 3). The different lowercase letters above error bars indicate significant differences between treatments by Post Hoc Tests (p < 0.05).
新窗口打开|下载原图ZIP|生成PPT 图4茭草凋落物叶际细菌群落多样性指数(Chao1)及细菌在属分类水平上的群落结构特征。图中误差线为标准误差(n = 3), 利用Post Hoc Tests进行两两比较分析, 不同小写字母代表处理之间的差异达到显著水平(p < 0.05)。
Fig. 4Diversity of bacterial community indicated by Chao1 index and the bacterial community composition at the genus level for leaf litter of Zizania latifolia. The error bars represent standard errors (n = 3), and the different lowercase letters above error bars indicate significant differences between treatments by Post Hoc Tests (p < 0.05).
Fig. 5Dynamics in average well color development (AWCD) value for carbon sources utilized by litter phyllospheric microorganisms of Zizania latifolia during an incubation period of 12-168 h. The error bars represent standard errors (n = 3), and the different lowercase letters above error bars indicate significant differences between treatments by Post Hoc Tests (p < 0.05).
Fig. 7Similarity analysis shows the contribution of different carbon sources to the dissimilarity between control vs. warming, air interface vs. water interface, air interface vs. soil interface, and water interface vs. soil interface, illustrated by heatmaps. The color (blue to red) represents the relative contribution of different carbon substrates (0-100%). The observations at 72-96 h incubation point were used for drawing the heatmaps.
BergB, McClaughertyC ( 1989). Nitrogen and phosphorus release from decomposing litter in relation to the disappearance of lignin Canadian Journal of Botany, 67, 1148-1156. DOI:10.1139/b89-150URL [本文引用: 1]
BonanomiG, CapodilupoM, IncertiG, MazzoleniS ( 2015). Litter quality and temperature modulate microbial diversity effects on decomposition in model experiments Community Ecology, 16, 167-177. DOI:10.1139/b89-150URL [本文引用: 1]
ChaudharyD, KumarR, SihagK, Rashmi & KumariA ( 2017). Phyllospheric microflora and its impact on plant growth: A review Agricultural Reviews, 38, 51-59. URL [本文引用: 1]
ChenC, XinK, LiuH, ChengJ, ShenXH, WangY, ZhangL ( 2017). Pantoea alhagi, a novel endophytic bacterium with ability to improve growth and drought tolerance in wheat Scientific Reports, 7, 41564. DOI: 10.1038/srep41564. DOI:10.1038/srep41564URL [本文引用: 1]
ChenYM, HeRL, DengCC, YangWQ, ZhangJ, YangL, LiuY ( 2015). Litter decomposition and lignocellulose enzyme activities of Actinothuidium hookeri and Cystopteris montana in alpine timberline ecotone of Western Sichuan, China. Chinese Journal of Applied Ecology, 26, 3251-3258. DOI:10.3321/j.issn:0412-1961.2006.02.002URLPMID:26915177 [本文引用: 1] 以川西高山林线交错带两种优势地被物锦丝藓和高山冷蕨为对象,对针叶林和林线中锦丝藓植物残体及高山冷蕨凋落叶分解的质量损失和木质纤维素酶活性特征进行研究.结果表明:锦丝藓和高山冷蕨的质量损失率在雪被期和生长季均表现为林线高于针叶林,而酶活性整体上表现为针叶林显著高于林线.两种地被物不同季节的质量损失有显著差异,雪被期林线和针叶林的锦丝藓质量损失率占全年的69.8%和83.0%;生长季林线和针叶林的高山冷蕨质量损失率分别占全年的82.6%和83.4%.高山冷蕨凋落叶在生长季节快速分解,与其生长季节末较高的纤维素酶活性相吻合,说明纤维素和半纤维素的酶解作用可能是凋落物前期质量损失的主要原因.多元线性回归分析表明,环境因子和凋落叶初始质量能共同解释酶活性变异的45.8%~85.1%,两种地被物分解过程中酶活性主要受到雪被期冻融循环的影响. [ 陈亚梅, 和润莲, 邓长春, 杨万勤, 张健, 杨林, 刘洋 ( 2015). 川西高山林线交错带两种地被物分解的木质纤维素酶活性特征 应用生态学报, 26, 3251-3258.] DOI:10.3321/j.issn:0412-1961.2006.02.002URLPMID:26915177 [本文引用: 1] 以川西高山林线交错带两种优势地被物锦丝藓和高山冷蕨为对象,对针叶林和林线中锦丝藓植物残体及高山冷蕨凋落叶分解的质量损失和木质纤维素酶活性特征进行研究.结果表明:锦丝藓和高山冷蕨的质量损失率在雪被期和生长季均表现为林线高于针叶林,而酶活性整体上表现为针叶林显著高于林线.两种地被物不同季节的质量损失有显著差异,雪被期林线和针叶林的锦丝藓质量损失率占全年的69.8%和83.0%;生长季林线和针叶林的高山冷蕨质量损失率分别占全年的82.6%和83.4%.高山冷蕨凋落叶在生长季节快速分解,与其生长季节末较高的纤维素酶活性相吻合,说明纤维素和半纤维素的酶解作用可能是凋落物前期质量损失的主要原因.多元线性回归分析表明,环境因子和凋落叶初始质量能共同解释酶活性变异的45.8%~85.1%,两种地被物分解过程中酶活性主要受到雪被期冻融循环的影响.
ChristofferB, MireiaBF, JaroneP, CatherineL ( 2018). Response of microbial communities to changing climate conditions during summer cyanobacterial blooms in the Baltic Sea Frontiers in Microbiology, 9, 1562. DOI: 10.3389/ fmicb.2018.01562. DOI:10.3389/fmicb.2018.01562URL [本文引用: 1] Frequencies and biomass of Baltic Sea cyanobacterial blooms are expected to be higher in future climate conditions, but also of longer duration as a result of increased sea surface temperature. Concurrently, climate predictions indicate a reduced salinity in the Baltic Sea. These climate-driven changes are expected to alter not solely the phytoplankton community but also the role of microbial communities for nutrient remineralization. Here, we present the response of summer plankton communities (filamentous cyanobacteria, picocyanobacteria, and heterotrophic bacteria) to the interplay of increasing temperature (from 16 to 18°C and 20°C) and reduced salinity (from salinity 6.9 to 5.9) in the Baltic Proper (NW Gotland Sea) using a microcosm approach. Warmer temperatures led to an earlier peak of cyanobacterial biomass, while yields were reduced. These conditions caused a decrease of nitrogen-fixers (Dolichospermum sp.) biomass, while non nitrogen-fixers (Pseudanabaena sp.) increased. Salinity reduction did not affect cyanobacterial growth nor community composition. Among heterotrophic bacteria, Actinobacteria showed preference for high temperature, while Gammaproteobacteria thrived at in situ temperature. Heterotrophic bacteria community changed drastically at lower salinity and resembled communities at high temperature. Picocyanobacteria and heterotrophic bacterial biomass had a pronounced increase associated with the decay of filamentous cyanobacteria. This suggests that shifts in community composition of heterotrophic bacteria are influenced both directly by abiotic factors (temperature and salinity) and potentially indirectly by cyanobacteria. Our findings suggest that at warmer temperature, lower yield of photosynthetic cyanobacteria combined with lower proportion of nitrogen-fixers in the community could result in lower carbon export to the marine food web with consequences for the decomposer community of heterotrophic bacteria.
FanZX, Br?uningA, ThomasA, LiJB, CaoKF ( 2011). Spatial and temporal temperature trends on the Yunnan Plateau (Southwest China) during 1961-2004 International Journal of Climatology, 31, 2078-2090. DOI:10.1002/joc.2214URL [本文引用: 1] Monthly mean (TEM), maximum (Tmax) and minimum (Tmin) surface air temperatures at 119 meteorological stations on the Yunnan Plateau (YP, Southwest China) were analysed for temporal trends and spatial variation patterns during the period 1961–2004. Linear trend analyses revealed that annual temperature over the YP increased at a rate of 0.3 °C/decade during the period 1961–2004, while warming trend of 0.33 °C/decade and 0.26 °C/decade was observed for winter and summer temperatures, respectively. Warming trends of nighttime minimum temperature are more pronounced than those of daytime maximum temperature, especially during winter season. Consequently, a decreasing trend of diurnal temperature ranges (DTR = Tmax ? Tmin) was observed. Five spatial patterns of temperature variability were objectively defined by rotated empirical orthogonal function (EOF) analysis, which are associated with distinct temporal temperature variations and geographical area over the YP. Annual temperature increases were found to be most pronounced in the southern and northwestern (high-elevation) parts of the YP, whereas the hot-dry valleys along the Yangtze and Red River basins experienced cooling during the past four decades. Copyright ? 2010 Royal Meteorological Society
FerreiraV, RaposeiroPM, PereiraA, CruzAM, CostaAC, GracaM, GoncalvesV ( 2016). Leaf litter decomposition in remote oceanic island streams is driven by microbes and depends on litter quality and environmental conditions Freshwater Biology, 61, 783-799. DOI:10.1111/fwb.12749URL [本文引用: 1] 1. Leaf litter decomposition is an important process in many streams. The flow of carbon and nutrients to higher trophic levels generally depends on litter characteristics and environmental conditions, and is driven by the activities of microbes and invertebrate shredders. However, little is known about what drives litter decomposition in oceanic islands, where invertebrate communities are species-poor. 2. In this study we assessed the relative importance of litter quality and environmental conditions on the biological colonization and decomposition of litter exposed to and protected from macroinvertebrates, in the Azores archipelago, North Atlantic Ocean. Three leaf litter species with distinct physical and chemical characteristics (Acacia melanoxylon, Clethra arborea and Pittosporum undulatum) were incubated in six streams with distinct water characteristics. Coarse and fine mesh bags were used to isolate the relative role of macroinvertebrates on litter decomposition. Incubation of litter took place in late spring – early summer and lasted for up to 56 days. 3. No significant differences in litter decomposition rates were found between coarse and fine mesh bags suggesting that microbes, especially aquatic hyphomycetes, are the key players in litter decomposition in these island streams. 4. Litter decomposition rates were inversely proportional to initial lignin concentration: A. melanoxylon 0.0080 d–1, C. arborea 0.0121 d–1, P. undulatum 0.0292 d–1, on average across streams. 5. Litter decomposition rates and associated decomposers differed among streams, suggesting that environmental conditions (e.g. nutrient concentration) may be important moderators of biological activities in these streams, as found for continental streams. 6. Species richness, fungal biomass and reproductive activity of aquatic hyphomycetes on decomposing litter were recorded in Atlantic islands for the first time and were at levels similar to those found for continental streams. 7. High microbial activities in Atlantic island streams ensure litter decomposition when detritivore abundance is low.
HeXB, SongFQ, ZhangP, LinYH, TianXJ, RenLL, ChenCL, XiaoN, TanHX ( 2007). Variation in litter decomposition- temperature relationships between coniferous and broadleaf forests in Huangshan Mountain, China Journal of Forestry Research, 18, 291-297. DOI:10.1007/s11676-007-0058-0URL A study was conducted to identify the differences in the decompositions of leaf litter, lignin and carbohydrate between coniferous forest and broadleaf forest at 20°C and 30°C in Huangshan Mountain, Anhui Province, China. Results showed that at 20°C mass loss of leaf litter driven by microbial decomposers was higher in broadleaf forest than that in coniferous forest, whereas the difference in mass loss of leaf litter was not significant at 30°C. The temperature increase did not affect the mass loss of leaf litter for coniferous forest treatment, but significantly reduced the decomposition rate for broadleaf forest treatment. The functional decomposers of microorganism in broadleaf for- est produced a higher lignin decomposition rate at 20°C, compared to that in coniferous forest, but the difference in lignin decomposition was not found between two forest types at 30°C. Improved temperature increased the lignin decomposition for both broadleaf and conifer- ous forest. Additionally, the functional group of microorganism from broadleaf forest showed marginally higher carbohydrate loss than that from coniferous forest at both temperatures. Temperature increase reduced the carbohydrate decomposition for broadleaf forest, while only a little reduce was found for coniferous forest. Remarkable differences occurred in responses between most enzymes (Phenoloxidase, peroxi- dase, β-glucosidase and endocellulase) and decomposition rate of leaf litter to forest type and temperature, although there exist strong rela- tionships between measured enzyme activities and decomposition rate in most cases. The reason is that more than one enzyme contribute to the mass loss of leaf litter and organic chemical components. In conclusion, at a community scale the coniferous and broadleaf forests dif- fered in their temperature-decomposition relationships.
HobaraS, OsonoT, HiroseD, NoroK, HirotaM, BennerR ( 2014). The roles of microorganisms in litter decomposition and soil formation Biogeochemistry, 118, 471-486.? DOI:10.1007/s10533-013-9912-7URL [本文引用: 1] Much has been learned about the microbial decomposition of plant litter, but relatively little is known about microbial contributions to litter and soil chemistry. We conducted a 3-year litterbag experiment and measured hydrolyzable amino acids (AA) and amino sugars (AS) to gain insights about microbial contributions to the chemical characteristics of decomposing litter and soil. Microscopic observations of hyphae were used to estimate fungal contributions to litter. The carbon (C)-normalized yields of AA and AS increased during decomposition along with nitrogen (N), indicating a shift in chemical characteristics from C-rich plant-derived biopolymers to N-rich, microbially-derived biochemicals. The contributions of fungal biomass to C and N were minor, but necromass of fungi as melanized and clamp-bearing hyphae increased during litter decomposition. Yields of glucosamine and galactosamine in litter approached those in microorganisms, particularly bacteria, suggesting major contributions of bacterial residues to litter during decomposition. The microbial contributions to decomposing litter were consistent with those observed in organic and mineral soils. Microorganisms play important roles in the organization and stabilization of soil organic matter as well as N immobilization and organic C preservation.
IPCC(Intergovernmental Panel on Climate Change)( 2013). Climate Change 2013: The Physical Science Basis. Cambridge University Press, Cambridge, UK. DOI:10.1007/BF00524943URL [本文引用: 2]
KuehnKA, SteinerD, GessnerMO ( 2004). Diel mineralization patterns of standing-dead plant litter: Implications for CO2 flux from wetlands Ecology, 85, 2504-2518. DOI:10.1890/03-4082URL [本文引用: 1] We examined the effects of environmental conditions on the microbially mediated CO2 evolution from standing-dead litter (leaf blades, leaf sheaths, and culms) of the common reed, Phragmites australis (Cav.) Trin. ex Steud., in two temperate littoral freshwater marshes. Water availability was the major factor affecting CO2 evolution rates. In the laboratory, microbial assemblages responded rapidly to controlled additions of water, with large increases in CO2 evolution occurring within five minutes after wetting of litter (e.g., leaf blades: 10-295 μ g CO2-C · (g ash-free dry mass [AFDM])-1· h-1. Under field conditions, CO2 evolution in the absence of precipitation exhibited a pronounced diel periodicity, with the highest rates occurring during periods of increased water availability resulting from a temperature-induced rise in relative humidity (>95%) and corresponding litter water potential (> -2.0 MPa) during nighttime. For example, in October, rates of CO2 evolution over a 24-h cycle ranged from 5 to 223 μ g CO2-C· (g AFDM)-1· h-1 for leaf blades and from 10 to 155 $\mug CO_2-C\cdot(g AFDM)^{-1}\cdot h^{-1}$ for leaf sheaths. Maximum rates of CO2 evolution from sheaths were consistently lower than those for leaf blades ($by \sim25\%$), but were typically an order of magnitude higher than those observed from culm litter (e.g., 1.0-18 μ g CO2-C· (g AFDM)-1· h-1 over a diel cycle in August) exposed to identical environmental conditions. Much of the differences in maximum CO2 evolution rates from different litter types were related (r2 = 0.72) to differences in litter associated fungal biomass (leaf blades 34-74 mg dry mass/g AFDM, leaf sheaths 16-67 mg dry mass/g AFDM, and culms 2-7 mg dry mass/g AFDM), which was estimated from litter ergosterol concentrations. Based on measured stocks of standing-dead plant litter, estimated daily CO2 flux from standing-dead shoots ranged between 51 and 570 mg C/m2 of wetland surface area. These values translate into a roughly estimated annual carbon mineralization equivalent to a mean of 8% (leaf blades), 29% (leaf sheaths), and 3% (culms) of net aboveground plant production. These data provide compelling evidence that microbial decomposition of plant litter in the aerial standing-dead phase can contribute appreciably to overall carbon flux from marshes to the atmosphere, even in cool temperate climates, where most wetlands occur.
KurtenGL, BarkohA ( 2016). Evaluation of community-level physiological profiling for monitoring microbial community function in aquaculture ponds North American Journal of Aquaculture, 78, 34-44. DOI:10.1080/15222055.2015.1079580URL [本文引用: 2] Microbial communities in aquaculture ponds play pivotal roles in pond productivity and fish production success. Nonetheless, culturists do not consider this variable in pond management decision making likely because of a lack of practical assessment tools. Therefore, we evaluated the community-level physiological profiling (CLPP) technique for characterizing microbial community function for use in aquaculture. Unfiltered water samples collected from various plastic-lined ponds over time were incubated on Biolog EcoPlates. Responses (substrate optical densities) were measured at 24-h intervals for up to 12 d to determine the best incubation interval and the number of required replicates needed for determining microbial community function in aquaculture ponds. The repeatability and within-pond variability of the CLPP metrics were also evaluated. During cooler months and in ponds with no fish and low apparent microbial activity, a 96-h incubation period was required to differentiate microbial community functional characteristics. During warmer months and when ponds were stocked with fish and had significant phytoplankton blooms, incubation periods of 72 h were sufficient. For routine monitoring, a single sample of pond water on one Biolog EcoPlate was adequate for assessing microbial community function. Multiple water samples from a single pond revealed low heterogeneity in the microbial community function. When multiple Biolog EcoPlates were inoculated with a single sample of pond water the dissimilarity of substrate responses was low (2.9%), indicating high repeatability of the CLPP technique. The CLPP method appears to be robust and allows for the assessment of functional characteristics of the heterotrophic microbial community such as relative diversity, similarity, and community functional activity. Therefore, this technique may be used by fish culturists to assess hatchery pond microbial community function, which would lead to a better understanding of the importance of the microbial community to the overall aquaculture pond ecosystem function. Such a tool would allow improved and more comprehensive management of aquaculture pond ecosystems.
LiQ, ZhouDW, ChenXY ( 2014). The accumulation, decomposition and ecological effects of above-ground litter in terrestrial ecosystem Acta Ecologica Sinica , 34, 3807-3819. [本文引用: 1]
LiuGF, CornwellWK, PanX, YeD, LiuFH, HuangZY, DongM, CornelissenJH ( 2015). Decomposition of 51 semidesert species from wide-ranging phylogeny is faster in standing and sand-buried than in surface leaf litters: Implications for carbon and nutrient dynamics Plant and Soil, 396, 175-187. [本文引用: 1]
MartyC, HouleD, GagnonC ( 2015). Variation in stocks and distribution of organic C in soils across 21 eastern Canadian temperate and boreal forests . Forest Ecology and Management, 345, 29-38. [本文引用: 1]
MoghadamFS, ZimmerM ( 2014). Effects of warming and nutrient enrichment on how grazing pressure affects leaf litter-colonizing bacteria Journal of Environment Quality, 43, 851-858. [本文引用: 1]
NewellSY ( 2001). Fungal biomass and productivity in? standing-decaying leaves of black needlerush (Juncus roemerianus) Marine & Freshwater Research, 52, 249-255. [本文引用: 1]
NiXY, YangWQ, LiH, XuLY, HeJ, TanB, WuFZ ( 2017). The responses of early foliar litter humification to reduced snow cover during winter in an alpine forest Canadian Journal of Science, 94, 453-461. [本文引用: 1]
ParkerTC, SandermanJ, HoldenRD, Blume-WerryG, SjogerstenS, LargeD, Castro-DiazM, StreetLE, SubkeJA, WookeyPA ( 2018). Exploring drivers of litter decomposition in a greening Arctic: Results from a transplant experiment across a tree-line Ecology, 99, 2284-2294. [本文引用: 1]
PengSL, LiuQ ( 2002). The dynamics of forest litter and its responses to global warming Acta Ecologica Sinica , 22, 1534-1544. [本文引用: 1]
SongXZ, JiangH, ZhangHL, YuSQ, ZhouGM, MaYD, ChangSX ( 2008). A review on the effects of global environment change on litter decomposition Acta Ecologica Sinica , 28, 4414-4423. [本文引用: 1]
SunZG, LiuJS ( 2007). Development in study of wetland litter decomposition and its responses to global change Acta Ecologica Sinica , 27, 1606-1618. [本文引用: 1]
VeronicaF, ChauvetE ( 2011). Future increase in temperature more than decrease in litter quality can affect microbial litter decomposition in streams Oecologia, 167, 279-291. [本文引用: 1]
WangFY ( 1989). Forest litter quality research review Advances in Ecology , 2, 82-89.
[ 王凤友 ( 1989). 森林凋落量研究综述 生态学进展, 2, 82-89.]
WangYH, GongJR, LiuM, HuangYM, YanX, ZhangZY, XuS, LuoQP ( 2015). Effects of grassland-use on soil respiration and litter decomposition Chinese Journal of Plant Ecology , 39, 239-248. [本文引用: 2]
WangZ, CaoGQ, ZhangYQ, ZhangHY, WangF, ChenAL ( 2017). Responses of metabolism diversity of topsoil microbial to the litterfall addition in Cunninghamia laneolata plantation. Journal of Forest and Environment, 37, 148-154. [本文引用: 1]
WarskowAL, JuniE ( 1972). Nutritional requirements of Acinetobacter strains isolated from soil, water, and sewage Journal of Bacteriology, 112, 1014-1016. [本文引用: 1]
WierzchosJ, CaseroMC, ArtiedaO, AscasoC ( 2018). Endolithic microbial habitats as refuges for life in polyextreme environment of the Atacama Desert Current Opinion in Microbiology, 43, 124-131. [本文引用: 1]
WuFZ, YangWQ, ZhangJ, DengRJ ( 2010). Litter decomposition in two subalpine forests during the freeze-thaw season Acta Oecologica, 36, 135-140. [本文引用: 1]
XuXF, TianHQ, WanSQ ( 2007). Climate warming impacts on carbon cycling in terrestrial ecosystems Journal of Plant Ecology (Chinese Version), 31, 175-188. [本文引用: 2]
XuZF, YinHJ, ZhaoCZ, CaoG, WanML, LiuQ ( 2009). A review of response of litter decomposition in terrestrial ecosystems to global warming Journal of Plant Ecology , 33, 1208-1219. [本文引用: 1]
YangWQ, DengRJ, ZhangJ ( 2007). Forest litter decomposition and its responses to global climate change Chinese Journal of Applied Ecology , 18, 2889-2895. [本文引用: 1]
YueK, YangWQ, PengY, HuangCP, ZhangC, WuFZ ( 2016). Effects of streams on lignin degradation during foliar litter decomposition in an alpine forest Chinese Journal of Plant Ecology , 40, 893-901. [本文引用: 1]
ZhangJ, ZhaiFF, ZhangJH, SunFB, ZhangHJ, MaoZG ( 2010). The behavior of anaerobic fermentation in the technique of alcohol fermentation cooperate with methane fermentation Journal of Food Science & Biotechnology , 29, 276-281. [本文引用: 1]
ZhangXN, LiuZY, LiLP, WangH, ZhangY, SunM, XiaoDR ( 2017). Effect of experimental warming on the decomposition of litter from dominant lakeside plants in a typical wetland of Northwestern Yunnan Plateau, China Acta Ecologica Sinica , 37, 7811-7820. [本文引用: 1]
ZhengH, OuyangZY, FangZG, ZhaoTQ ( 2004). Application of BIOLOG to study on soil microbial community functional diversity Acta Pedologica Sinica , 41, 456-461.
... 生境差异对凋落物的分解产生重要影响(Liu et al., 2015).有研究对比了溪流、河岸带以及林地三种区域生境下凋落物的分解特征, 发现溪流生境条件下凋落物中木质素残留质量最低(岳楷等, 2016); 而在干旱少雨地区, 处于立枯及掩埋状态的凋落物比地表的凋落物分解更快(Liu et al., 2015), 表明生境中的不同环境要素可深刻影响凋落物分解过程.不同于地形、坡度、朝向以及土壤水分等环境因素调控陆生植物凋落物分解过程, 对于湿地挺水植物而言, 其所存在的生境主要包括立枯阶段的大气生境、倒伏阶段的水生境和沉水阶段的土生境, 因此湿地挺水植物凋落物的分解速率主要由处于不同分解阶段的水文环境因素调控.在大气界面, 受水分条件限制, 同时缺乏大型底栖动物活动及冻融作用对凋落物的物理破碎过程, 导致凋落物分解速率较低.与大气界面相比, 水、土界面温暖而湿润的环境因素更有利于凋落物中酸不溶性组分等难分解的高分子结构体的快速降解(Wu et al., 2010), 并且土壤中含有大量的微生物, 通过与凋落物直接接触, 其对底物的利用效率比大气界面更高(Hobara et al., 2014).另外, 我们发现每到秋冬季节, 滇西北区域水路交错地带有明显的冻融现象, 冻结作用也可以直接破坏凋落物的物理结构从而加快处于水、土界面凋落物的分解速率(Ni et al., 2017).可见, 由这些生境所引起的环境因子差异是影响凋落物分解的关键(Parker et al., 2018). ...
... ); 而在干旱少雨地区, 处于立枯及掩埋状态的凋落物比地表的凋落物分解更快(Liu et al., 2015), 表明生境中的不同环境要素可深刻影响凋落物分解过程.不同于地形、坡度、朝向以及土壤水分等环境因素调控陆生植物凋落物分解过程, 对于湿地挺水植物而言, 其所存在的生境主要包括立枯阶段的大气生境、倒伏阶段的水生境和沉水阶段的土生境, 因此湿地挺水植物凋落物的分解速率主要由处于不同分解阶段的水文环境因素调控.在大气界面, 受水分条件限制, 同时缺乏大型底栖动物活动及冻融作用对凋落物的物理破碎过程, 导致凋落物分解速率较低.与大气界面相比, 水、土界面温暖而湿润的环境因素更有利于凋落物中酸不溶性组分等难分解的高分子结构体的快速降解(Wu et al., 2010), 并且土壤中含有大量的微生物, 通过与凋落物直接接触, 其对底物的利用效率比大气界面更高(Hobara et al., 2014).另外, 我们发现每到秋冬季节, 滇西北区域水路交错地带有明显的冻融现象, 冻结作用也可以直接破坏凋落物的物理结构从而加快处于水、土界面凋落物的分解速率(Ni et al., 2017).可见, 由这些生境所引起的环境因子差异是影响凋落物分解的关键(Parker et al., 2018). ...
凋落物分解过程中土壤微生物群落的变化 2 2016
... 生境差异对凋落物的分解产生重要影响(Liu et al., 2015).有研究对比了溪流、河岸带以及林地三种区域生境下凋落物的分解特征, 发现溪流生境条件下凋落物中木质素残留质量最低(岳楷等, 2016); 而在干旱少雨地区, 处于立枯及掩埋状态的凋落物比地表的凋落物分解更快(Liu et al., 2015), 表明生境中的不同环境要素可深刻影响凋落物分解过程.不同于地形、坡度、朝向以及土壤水分等环境因素调控陆生植物凋落物分解过程, 对于湿地挺水植物而言, 其所存在的生境主要包括立枯阶段的大气生境、倒伏阶段的水生境和沉水阶段的土生境, 因此湿地挺水植物凋落物的分解速率主要由处于不同分解阶段的水文环境因素调控.在大气界面, 受水分条件限制, 同时缺乏大型底栖动物活动及冻融作用对凋落物的物理破碎过程, 导致凋落物分解速率较低.与大气界面相比, 水、土界面温暖而湿润的环境因素更有利于凋落物中酸不溶性组分等难分解的高分子结构体的快速降解(Wu et al., 2010), 并且土壤中含有大量的微生物, 通过与凋落物直接接触, 其对底物的利用效率比大气界面更高(Hobara et al., 2014).另外, 我们发现每到秋冬季节, 滇西北区域水路交错地带有明显的冻融现象, 冻结作用也可以直接破坏凋落物的物理结构从而加快处于水、土界面凋落物的分解速率(Ni et al., 2017).可见, 由这些生境所引起的环境因子差异是影响凋落物分解的关键(Parker et al., 2018). ...
... ); 而在干旱少雨地区, 处于立枯及掩埋状态的凋落物比地表的凋落物分解更快(Liu et al., 2015), 表明生境中的不同环境要素可深刻影响凋落物分解过程.不同于地形、坡度、朝向以及土壤水分等环境因素调控陆生植物凋落物分解过程, 对于湿地挺水植物而言, 其所存在的生境主要包括立枯阶段的大气生境、倒伏阶段的水生境和沉水阶段的土生境, 因此湿地挺水植物凋落物的分解速率主要由处于不同分解阶段的水文环境因素调控.在大气界面, 受水分条件限制, 同时缺乏大型底栖动物活动及冻融作用对凋落物的物理破碎过程, 导致凋落物分解速率较低.与大气界面相比, 水、土界面温暖而湿润的环境因素更有利于凋落物中酸不溶性组分等难分解的高分子结构体的快速降解(Wu et al., 2010), 并且土壤中含有大量的微生物, 通过与凋落物直接接触, 其对底物的利用效率比大气界面更高(Hobara et al., 2014).另外, 我们发现每到秋冬季节, 滇西北区域水路交错地带有明显的冻融现象, 冻结作用也可以直接破坏凋落物的物理结构从而加快处于水、土界面凋落物的分解速率(Ni et al., 2017).可见, 由这些生境所引起的环境因子差异是影响凋落物分解的关键(Parker et al., 2018). ...
Decomposition of 51 semidesert species from wide-ranging phylogeny is faster in standing and sand-buried than in surface leaf litters: Implications for carbon and nutrient dynamics 1 2015
 ,1,*
,1,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT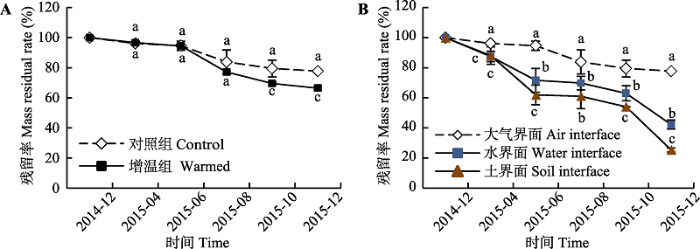 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT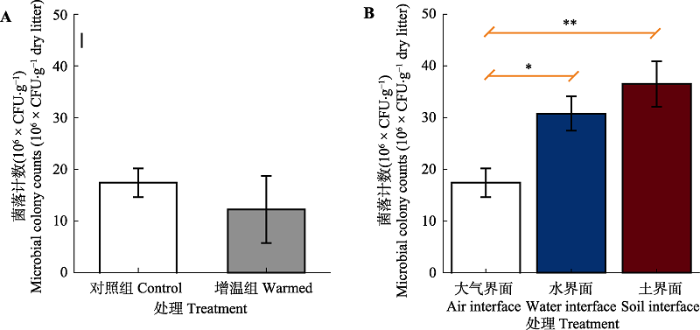 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT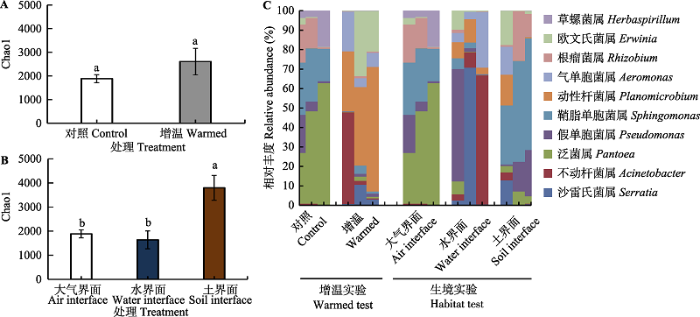 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT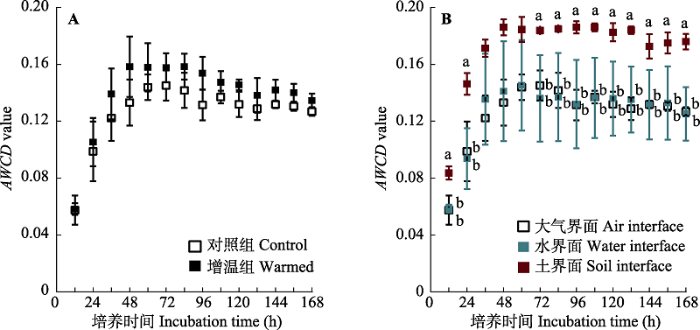 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT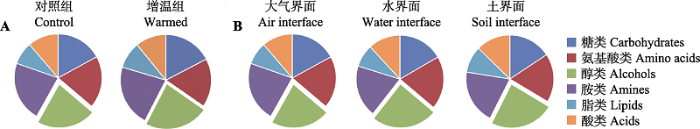 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT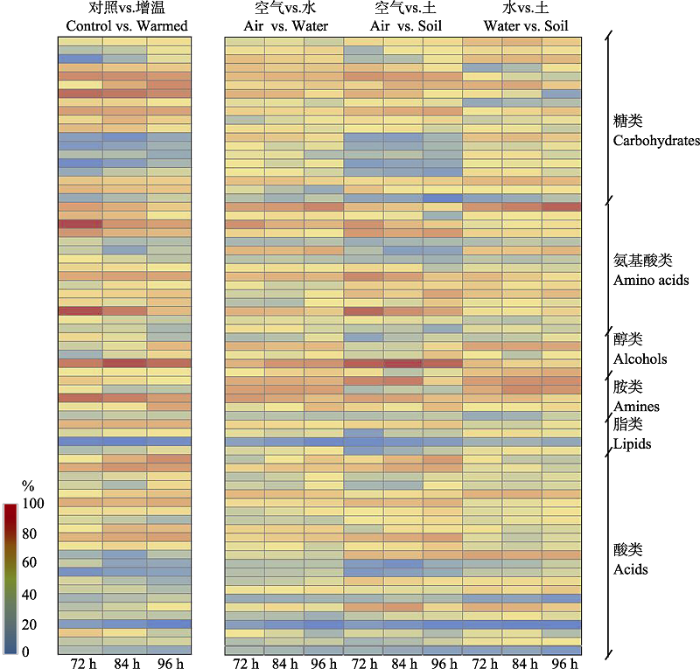 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}