Supported by the National Natural Science Foundation of China(U1702235) the Ministry and Province Joint Construction Project of Yunnan University(C176280109)
Abstract AimsPrevious studies have shown that Dendrobium plants form a specific symbiotic relationship with fungi at differentiation stages during natural seed germination. In order to explore the evolution and adaptation of this symbiotic relationship in interspecific hybrid progenies, this study was to understand whether the strong specificity with symbiotic fungi during seedling formation and differentiation was also an important factor limiting the formation of hybrid progenies in Dendrobium, and the relationship between hybrid progenies of orchids and symbiotic fungi during seed germination stage. MethodsThe effects of fungi on germination of interspecific hybrid seeds of D. officinale and D. tortile were studied using the highly specific fungi strains Tulasnella SSCDO-5 and Epulorhiza FDd1, which can effectively promote seed germination and seedling formation in D. officinale and D. devonianum, respectively. Important findings The results showed that both SSCDO-5 and FDd1 strains could effectively promote the protocorm and seedling formation of hybrid seeds after 68 days incubation with no significant difference. The SSCDO-5 strain from D. officinale did not show any advantages, and the seedling formation rate of hybrid was lower than that of FDd1 strain from D. devonianum. The seedling formation rate incubation with SSCDO-5 strain was (22.13 ± 6.62)% while with FDd1 strain was (29.53 ± 5.51)%. The specificity of SSCDO-5 strain with D. officinale at seedling formation and development stage was not inherited or expressed in hybrid progenies, indicating that hybridization broke the symbiotic relationship of this specificity, which enabled hybrid progenies to establish new symbiotic relationship with different fungi. Our results do not support the hypothesis that the specificity of symbiotic fungi is an important limiting factor for the formation of hybrid progenies in Dendrobium. We speculate that the symbiotic relationship between Dendrobium plants and fungi during seedling differentiation and development is formed and established in the process of adapting to specific ecological environment. Keywords:symbiosis specificity;mycorrhizal fungi;Orchidaceae;Dendrobium;hybrid speciation;symbiotic seed germination
PDF (1530KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 范紫腾, 毋钰灵, 王新菊, 李太强, 高江云. 共生真菌对兰科植物种间杂交后代种子萌发的效应. 植物生态学报, 2019, 43(4): 374-382. DOI: 10.17521/cjpe.2019.0048 FAN Zi-Teng, WU Yu-Ling, WANG Xin-Ju, LI Tai-Qiang, GAO Jiang-Yun. Effects of symbiotic fungi on seed germination of interspecific hybrid progenies in Orchidaceae. Chinese Journal of Plant Ecology, 2019, 43(4): 374-382. DOI: 10.17521/cjpe.2019.0048
Fig. 1Seed germination, protocorm and seedling formation of three Dendrobium species under different treatments at different stages. A, Germinating seed and formed protocorm of D. officinale after 30 days incubation with SSCDO-5 strain. B, A seeding of D. devonianum after 58 days incubation with FDd1 strain. C, Seedings of hybrid after 68 days incubation with FDd1 strain. D, Seedings of hybrid after 68 days incubation with SSCDO-5 strain.
Fig. 2Percentage of seed germination (A, D, G), protocorm (B, E, H) and seedling (C, F, I) formation of different treatments in three Dendrobium species (mean ± SE). Different lowercase letters above the bars represent significant differences between different treatments at the same stage (α = 0.05). OMA, oatmeal agar medium without fungal inoculation was used as nutrient-poor control treatment; MS, MS medium without fungal inoculation was used as nutrient-rich control treatment; SSCDO-5, treatment of fungal inoculation with SSCDO-5 strain on OMA medium; FDd1, treatment of fungal inoculation with FDd1 strain on OMA medium.
Table 1 表1 表1不同处理三种石斛种子萌发率、原球茎形成率和幼苗形成率的多重比较(均值差±标准误差) Table 1Multiple comparisons on seed germination rate, protocorms formation rate and seedlings formation rate of different treatments for three Dendrobium species (mean ± SE)
多重比较 Multiple comparisons
铁皮石斛 D. officinale
齿瓣石斛 D. devonianum
杂交种 Hybrid
种子萌发率 Seed germination rate
原球茎形成率 Protocorms formation rate
幼苗率 Seedlings formation rate
种子萌发率 Seed germination rate
原球茎形成率 Protocorms formation rate
幼苗率 Seedlings formation rate
种子萌发率 Seed germination rate
原球茎形成率 Protocorms formation rate
幼苗率 Seedlings formation rate
OMA-MS
-0.139 3 ± 0.058 2*
0.000 0 ± 0.042 7
0.000 0 ± 0.053 4
-0.044 4 ± 0.071 1
-0.065 5 ± 0.063 9
-0.021 4 ± 0.051 6
-0.012 0 ± 0.058 1
-0.039 3 ± 0.053 0
-0.017 2 ± 0.064 4
OMA-SSCDO-5
-0.235 3 ± 0.057 7***
-0.888 8 ± 0.042 3***
-0.696 6 ± 0.052 9***
0.312 1 ± 0.069 5***
-0.354 8 ± 0.062 4***
-0.096 8 ± 0.050 4
-0.036 9 ± 0.058 1
-0.704 0 ± 0.053 0***
-0.221 3 ± 0.064 4**
OMA-FDd1
-0.242 3 ± 0.058 2***
-0.065 5 ± 0.042 7
0.065 5 ± 0.053 4
-0.094 8 ± 0.070 5
-0.651 7 ± 0.063 3***
-0.436 8 ± 0.051 2***
-0.070 1 ± 0.059 1
-0.800 4 ± 0.053 9***
-0.295 3 ± 0.065 5***
MS-SSCDO-5
-0.096 0 ± 0.058 2
-0.888 8 ± 0.042 7***
-0.696 6 ± 0.053 4***
0.356 5 ± 0.068 1***
-0.289 4 ± 0.061 1***
-0.075 3 ± 0.049 4
-0.024 9 ± 0.055 3
-0.664 8 ± 0.050 4***
-0.204 0 ± 0.061 3**
MS- FDd1
-0.102 9 ± 0.058 7
-0.006 6 ± 0.043 1
0.065 5 ± 0.053 8
-0.050 4 ± 0.069 2
-0.586 2 ± 0.062 1***
-0.415 4 ± 0.050 2***
-0.058 1 ± 0.056 3
-0.761 1 ± 0.0513 4***
-0.278 0 ± 0.062 4***
SSCDO-5- FDd1
-0.007 0 ± 0.058 2
0.823 3 ± 0.042 7***
0.631 0 ± 0.053 4***
-0.406 9 ± 0.067 5***
-0.296 9 ± 0.060 6***
-0.340 0 ± 0.049 0***
-0.033 2 ± 0.056 3
-0.096 4 ± 0.051 3
-0.074 0 ± 0.062 4
OMA, oatmeal agar medium without fungal inoculation was used as nutrient-poor control treatment; MS, MS medium without fungal inoculation was used as nutrient-rich control treatment; SSCDO-5, treatment of fungal inoculation with SSCDO-5 strain on OMA medium; FDd1, treatment of fungal inoculation with FDd1 strain on OMA medium. *, p < 0.05; **, p < 0.01; ***, p < 0.001. OMA, 不接菌的燕麦琼脂培养基作为营养缺乏对照处理; MS, 不接菌的MS培养基作为营养丰富对照处理; SSCDO-5, 燕麦培养基上接种SSCDO-5菌株处理; FDd1, 燕麦培养基上接种FDd1菌株处理菌的处理。
AcetoS, CaputoP, CozzolinoS, GaudioL, MorettiA ( 1999). Phylogeny and evolution of Orchis and allied genera based on ITS DNA variation: Morphological gaps and molecular continuity Molecular Phylogenetics & Evolution, 13, 67-76. [本文引用: 1]
ArdittiJ ( 1967). Factors affecting the germination of orchid seeds The Botanical Review, 33, 1-97. [本文引用: 2]
ArdittiJ, GhaniAKA ( 2000). Numerical and physical properties of orchid seeds and their biological implications New Phytologist, 145, 367-421. [本文引用: 1]
BidartondoMI, ReadDJ (2008). Fungal specificity bottlenecks during orchid germination and development Molecular Ecology, 17, 3707-3716. [本文引用: 2]
BrundrettMC, ScadeA, BattyAL, DixonKW, SivasithamparamK ( 2003). Development of in situ and ex situ seed baiting techniques to detect mycorrhizal fungi from terrestrial orchid habitats Mycological Research, 107, 1210-1220. [本文引用: 1]
ChaseMW, CameronKM, FreudensteinJV, PridgeonAM, SalazarG, BergC, SchuitemanA ( 2015). An updated classification of Orchidaceae Botanical Journal of the Linnean Society, 177, 151-174. [本文引用: 1]
CozzolinoS, WidmerA ( 2005). Orchid diversity: An evolutionary consequence of deception? Trends in Ecology & Evolution, 20, 487-494. [本文引用: 1]
DafniA, IvriY (1979). Pollination ecology of, and hybridization between,Orchis coriophora L. and O. collina Sol. ex Russ.(Orchidaceae) in Israel New Phytologist, 83, 181-187. [本文引用: 1]
DearnaleyJWD, PerottoS, SelosseMA (2016). Structure and development of orchid mycorrhizas In: Martin F ed. Molecular Mycorrhizal Symbiosis. Springer, Berlin. 63-86. [本文引用: 1]
GaoJY, LiuQ, YuDL (2014). Orchids of Xishuangbanna: Diversity and Conservation. China Forestry Publishing House, Beijing. 15-31. [本文引用: 2]
HegartyMJ, HiscockSJ ( 2005). Hybrid speciation in plants: New insights from molecular studies New Phytologist, 165, 411-423. [本文引用: 1]
HollickPS, TaylorRJ, McCombJA, DixonKW ( 2005). If orchid mycorrhizal fungi are so specific, how do natural hybrids cope? Selbyana, 26, 159-170. [本文引用: 2]
HuangH, ZiXM, LinH, GaoJY ( 2018). Host-specificity of symbiotic mycorrhizal fungi for enhancing seed germination, protocorm formation and seedling development of over-collected medicinal orchid,Dendrobium devonianum Journal of Microbiology, 56, 42-48. [本文引用: 6]
KauthP, DutraD, JohnsonT (2008). Techniques and applications of in vitro orchid seed germination. In: Teixeira da Silva JA ed. Floriculture, Ornamental and Plant Biotechnology: Advances and Topical Issues Global Science Books, Iselworth,UK. 375-391. [本文引用: 2]
LeeYI ( 2007). The asymbiotic seed germination of six Paphiopedilum species in relation to the time of seed collection and seed pretreatment Acta Horticulturae, 755, 381-386. [本文引用: 1]
LiuHX, LuoYB, LiuH ( 2010). Studies of mycorrhizal fungi of Chinese orchids and their role in orchid conservation in China―A review The Botanic Review, 76, 241-262. [本文引用: 1]
LongB, NiemieraAX, ChengZY, LongCL ( 2010). In vitro propagation of four threatened Paphiopedilum species (Orchidaceae) Plant Cell Tissue and Organ Culture, 101, 151-162. [本文引用: 1]
MalletJ ( 2005). Hybridization as an invasion of the genome Trends in Ecology & Evolution, 20, 229-237. [本文引用: 1]
MartosF, FranciosM, PaillerT, KottkeI, CédricG, Marc- AndréSelosse ( 2012). The role of epiphytism in architecture and evolutionary constraint within mycorrhizal networks of tropical orchids Molecular Ecology, 21, 5098-5109. [本文引用: 1]
MasuharaG, KatsuyaK (1994). In situ and in vitro specificity between Rhizoctonia spp. and Spiranthes sinensis(Persoon) Ames. var. amoena 127, 711-718. [本文引用: 1]
McCormickMK, JacquemynH ( 2014). What constrains the distribution of orchid populations? New Phytologist, 202, 392-400. [本文引用: 1]
McKendrickSL, LeakeJR, ReadDJ ( 2000). Symbiotic germination and development of myco-heterotrophic plants in nature: Transfer of carbon from ectomycorrhizal Salix repens and Betula pendula to the orchid Corallorhiza trifida through shared hyphal connections New Phytologist, 145, 539-548. [本文引用: 1]
MengYY, ShaoSC, LiuSJ, GaoJY ( 2019). Do the fungi associated with roots of adult plants support seed germination? A case study on Dendrobium exile(Orchidaceae) Global Ecology and Conservation, 17, e00582. DOI: 10.1016/? j.gecco.2019.e00582. [本文引用: 2]
NontachaiyapoomS, SasiratS, ManochL ( 2010). Isolation and identification of Rhizoctonia-like fungi from roots of three orchid genera, Paphiopedilum, Dendrobium, and Cymbidium, collected in Chiang Rai and Chiang Mai provinces of Thailand Mycorrhiza, 20, 459-471. [本文引用: 1]
RamsayMM, StewartJ ( 1998). Re-establishment of the lady’s slipper orchid (Cypripedium Calceolus L.) in Britain Botanical Journal of the Linnean Society, 126, 173-181. [本文引用: 1]
RasmussenHN, DixonKW, JersakovaJ, TesitelovaT ( 2015). Germination and seedling establishment in orchids: A complex of requirements Annals of Botany, 116, 391-402. [本文引用: 3]
RobertsDL (2003). Pollination biology: The role of sexual reproduction in orchid conservation In: Dixon KW, Kell SP, Barrett RL, Cribb PJ eds. Orchid Conservation. Natural History Publications, Kota Kinabalu, Sabah. 113-136. [本文引用: 1]
SchatzB ( 2006). Fine scale distribution of pollinator explains the occurrence of the natural orchid hybrid × Orchis bergonii Ecoscience, 13, 111-118. [本文引用: 1]
SchatzB, GeoffroyA, DainatB, BessiereJM, BuatoisB, Hossaert-MckeyM, SelosseMA ( 2010). A case study of modified interactions with symbionts in a hybrid mediterranean orchid American Journal of Botany, 97, 1278-1288. [本文引用: 2]
ShengCL, LiYY, GaoJY ( 2012). Ex situ symbiotic seed germination, isolation and identification of effective symbiotic fungus in Cymbidium mannii(Orchidaceae) Chinese Journal of Plant Ecology, 36, 859-869. [本文引用: 3]
StewartSL, KaneME ( 2007). Symbiotic seed germination and evidence for in vitro mycobiont specificity in Spiranthes brevilabris(Orchidaceae) and its implications for species-?level conservation In Vitro Cellular & Developmental Biology-Plant, 43, 178-186. [本文引用: 1]
StimartDP, AscherPD ( 1981). In vitro germination of Paphiopedilum seed on a completely defined medium Scientia Horticulturae, 14, 165-170. [本文引用: 1]
SwartsND, DixonKW ( 2009). Terrestrial orchid conservation in the age of extinction Annals of Botany, 104, 543-556. [本文引用: 1]
VujanovicV, St-ArnaudM, BarabeD, ThibeaultG ( 2000). Viability testing of orchid seed and the promotion of colouration and germination Annals of Botany, 86, 79-86. [本文引用: 1]
YamazakiJ, MiyoshiK ( 2006). In vitro asymbiotic germination of immature seed and formation of protocorm by Cephalanthera falcata(Orchidaceae) Annals of Botany, 98, 1197-1206. [本文引用: 1]
ZelmerCD, CuthbertsonL, CurrahRS ( 1996). Fungi associated with terrestrial orchid mycorrhizas, seeds and protocorms Mycoscience, 37, 439-448. [本文引用: 1]
ZettlerLW, PiskinKA, StewartSL, HartsockJJ, BowlesML, BellTJ ( 2005). Protocorm mycobionts of the federally threatened eastern prairie fringed orchid,Platanthera leucophaea(Nutt.) Lindley, and a technique to prompt leaf elongation in seedlings Studies in Mycology, 53, 163-171. [本文引用: 1]
ZhouX, GaoJY ( 2016). Highly compatible Epa-01 strain promotes seed germination and protocorm development of Papilionanthe teres(Orchidaceae) Plant Cell Tissue and Organ Culture, 125, 479-493. [本文引用: 3]
ZiXM, ShengCL, GoodaleUM, ShaoSC, GaoJY ( 2014). In situ seed baiting to isolate germination-enhancing fungi for an epiphytic orchid, Dendrobium aphyllum(Orchidaceae) Mycorrhiza, 24, 487-499. [本文引用: 4]
ZotzG ( 2013). The systematic distribution of vascular epiphytes—A critical update Botanical Journal of the Linnean Society, 171, 453-481. [本文引用: 1]
Phylogeny and evolution of Orchis and allied genera based on ITS DNA variation: Morphological gaps and molecular continuity 1 1999
Host-specificity of symbiotic mycorrhizal fungi for enhancing seed germination, protocorm formation and seedling development of over-collected medicinal orchid,Dendrobium devonianum 6 2018
... 胶膜菌科真菌是兰科植物最主要的一种菌根真菌类群, 也是很多石斛属植物种子萌发的有效共生真菌(Liu et al., 2010; Dearnaley et al., 2012; Zi et al., 2014; Huang et al., 2018), 瘤菌根菌属(Epulorhiza)被认为是胶膜菌属(Tulasnella)的无性态属(anamorph/teleomorph pair; Dearnaley et al., 2012).本研究中, SSCDO-5菌株为胶膜菌属真菌, FDd1菌株为瘤菌根菌属真菌, 能分别有效地促进铁皮石斛和齿瓣石斛原球茎和幼苗形成, 在幼苗形成和发育阶段表现出了很强的专一性(图2), 这和我们先前的研究结果一致(Huang et al., 2018; Shao et al., unpublished data), 而这种专一性的共生关系是如何形成和维系的, 兰科植物种子和对应真菌是如何相互选择和识别的, 其机制并不清楚.在对不同类群兰科植物种子的非共生萌发研究中都发现, 未成熟的种子比成熟的种子更容易萌发, 因为成熟种子的种皮会导致种子透水性降低(Ramsay & Stewart, 1998; Yamazaki & Miyoshi, 2006; Lee, 2007; Kauth et al., 2008; Long et al., 2010), 而在本研究和我们先前的研究中都发现, 与作为对照的营养丰富培养基处理相比较, 在亲和真菌的接菌处理中, 幼苗形成所需要的时间显著较短, 同时具有显著高的幼苗形成率(Zhou & Gao, 2016), 这是否也暗示种子的种皮结构和真菌建立了特定的信号传导和识别机制, 一方面使得亲和的真菌能快速侵染种子, 促进种子萌发和幼苗形成, 另一方面可以防止不亲和真菌的侵染? ...
... ), 这和我们先前的研究结果一致(Huang et al., 2018; Shao et al., unpublished data), 而这种专一性的共生关系是如何形成和维系的, 兰科植物种子和对应真菌是如何相互选择和识别的, 其机制并不清楚.在对不同类群兰科植物种子的非共生萌发研究中都发现, 未成熟的种子比成熟的种子更容易萌发, 因为成熟种子的种皮会导致种子透水性降低(Ramsay & Stewart, 1998; Yamazaki & Miyoshi, 2006; Lee, 2007; Kauth et al., 2008; Long et al., 2010), 而在本研究和我们先前的研究中都发现, 与作为对照的营养丰富培养基处理相比较, 在亲和真菌的接菌处理中, 幼苗形成所需要的时间显著较短, 同时具有显著高的幼苗形成率(Zhou & Gao, 2016), 这是否也暗示种子的种皮结构和真菌建立了特定的信号传导和识别机制, 一方面使得亲和的真菌能快速侵染种子, 促进种子萌发和幼苗形成, 另一方面可以防止不亲和真菌的侵染? ...
Techniques and applications of in vitro orchid seed germination. In: Teixeira da Silva JA ed. Floriculture, Ornamental and Plant Biotechnology: Advances and Topical Issues 2 2008
Symbiotic germination and development of myco-heterotrophic plants in nature: Transfer of carbon from ectomycorrhizal Salix repens and Betula pendula to the orchid Corallorhiza trifida through shared hyphal connections 1 2000
... 在我们对不同石斛属植物的研究中, 也得到了相同的结果, 利用种子诱导技术从自然形成的原球茎或幼苗分化初期分离得到的真菌, 在幼苗形成阶段有着很强的专一性, 实验证明不同来源的真菌都能促进种子萌发和原球茎形成, 但却不能有效促进幼苗的形成和发育(Zi et al., 2014; Huang et al., 2018; Meng et al., 2019).例如, 本研究所用的FDd1的菌株, 在和齿瓣石斛种子共生培养50天时, 幼苗形成率为(72.36 ± 11.7)%, 很多幼苗已经有根的发育, 但作为对照处理的2种真菌, FDaI7和FCb4菌株, 分别为硬叶兰(Cymbidium mannii)和兜唇石斛(D. aphyllum)种子萌发的有效共生真菌(盛春玲等, 2012; Zi et al., 2014), 虽然都能有效促进齿瓣石斛种子萌发和原球茎的形成, 但FDaI7接菌处理的幼苗形成率仅为(0.74 ± 1.7)%, 而FCb4接菌处理没有观察到幼苗; 到了共生培养90天时, FDaI7接菌处理的幼苗形成率为(23.59 ± 24.52)%, FCb4接菌处理的幼苗形成率为(9.08 ± 12.51)%, 而FDd1接菌处理的幼苗形成率在共生培养50天时就已趋于饱和, FDd1真菌能快速促进齿瓣石斛种子萌发并有效形成幼苗, 对于附生兰科植物在自然条件下种群幼苗的成功建立至关重要(Huang et al., 2018). ...
Isolation and identification of Rhizoctonia-like fungi from roots of three orchid genera, Paphiopedilum, Dendrobium, and Cymbidium, collected in Chiang Rai and Chiang Mai provinces of Thailand 1 2010
... 长期以来, 用于兰科植物种子共生萌发相关研究的真菌多是从野生成年植株的根中分离和筛选获得(Masuhara & Katsuya, 1994; Zelmer et al., 1996; Brundrett et al., 2003; Stewart & Kane, 2007; Nontachaiyapoom et al., 2010), 由于野生植株根中可能存在作用未知的不同内生真菌, 得到的菌株往往是随机的, 更为重要的是, 成年植株根中的真菌对种子萌发是否有效并不确定(Zettler et al., 2005), 然而, 很多研究结果都表明所获得的真菌能有效促进种子萌发, 这样的结果在很大程度上是由于采用了Arditti (1967)的种子萌发标准, 即把种胚膨大并产生根状物即视为种子萌发, 并没有关注真菌对后续幼苗形成和发育的影响.而兰科植物在种子萌发后都要经历或短或长的非光合作用阶段, 完全依靠真菌提供有机碳(Rasmussen et al., 2015), 非亲和的真菌或许能刺激种子萌发, 但并不能支持后续的幼苗发育(Bidartondo & Read, 2008; Rasmussen et al., 2015). ...
Re-establishment of the lady’s slipper orchid (Cypripedium Calceolus L.) in Britain 1 1998
... 胶膜菌科真菌是兰科植物最主要的一种菌根真菌类群, 也是很多石斛属植物种子萌发的有效共生真菌(Liu et al., 2010; Dearnaley et al., 2012; Zi et al., 2014; Huang et al., 2018), 瘤菌根菌属(Epulorhiza)被认为是胶膜菌属(Tulasnella)的无性态属(anamorph/teleomorph pair; Dearnaley et al., 2012).本研究中, SSCDO-5菌株为胶膜菌属真菌, FDd1菌株为瘤菌根菌属真菌, 能分别有效地促进铁皮石斛和齿瓣石斛原球茎和幼苗形成, 在幼苗形成和发育阶段表现出了很强的专一性(图2), 这和我们先前的研究结果一致(Huang et al., 2018; Shao et al., unpublished data), 而这种专一性的共生关系是如何形成和维系的, 兰科植物种子和对应真菌是如何相互选择和识别的, 其机制并不清楚.在对不同类群兰科植物种子的非共生萌发研究中都发现, 未成熟的种子比成熟的种子更容易萌发, 因为成熟种子的种皮会导致种子透水性降低(Ramsay & Stewart, 1998; Yamazaki & Miyoshi, 2006; Lee, 2007; Kauth et al., 2008; Long et al., 2010), 而在本研究和我们先前的研究中都发现, 与作为对照的营养丰富培养基处理相比较, 在亲和真菌的接菌处理中, 幼苗形成所需要的时间显著较短, 同时具有显著高的幼苗形成率(Zhou & Gao, 2016), 这是否也暗示种子的种皮结构和真菌建立了特定的信号传导和识别机制, 一方面使得亲和的真菌能快速侵染种子, 促进种子萌发和幼苗形成, 另一方面可以防止不亲和真菌的侵染? ...
Germination and seedling establishment in orchids: A complex of requirements 3 2015
... 在我们对不同石斛属植物的研究中, 也得到了相同的结果, 利用种子诱导技术从自然形成的原球茎或幼苗分化初期分离得到的真菌, 在幼苗形成阶段有着很强的专一性, 实验证明不同来源的真菌都能促进种子萌发和原球茎形成, 但却不能有效促进幼苗的形成和发育(Zi et al., 2014; Huang et al., 2018; Meng et al., 2019).例如, 本研究所用的FDd1的菌株, 在和齿瓣石斛种子共生培养50天时, 幼苗形成率为(72.36 ± 11.7)%, 很多幼苗已经有根的发育, 但作为对照处理的2种真菌, FDaI7和FCb4菌株, 分别为硬叶兰(Cymbidium mannii)和兜唇石斛(D. aphyllum)种子萌发的有效共生真菌(盛春玲等, 2012; Zi et al., 2014), 虽然都能有效促进齿瓣石斛种子萌发和原球茎的形成, 但FDaI7接菌处理的幼苗形成率仅为(0.74 ± 1.7)%, 而FCb4接菌处理没有观察到幼苗; 到了共生培养90天时, FDaI7接菌处理的幼苗形成率为(23.59 ± 24.52)%, FCb4接菌处理的幼苗形成率为(9.08 ± 12.51)%, 而FDd1接菌处理的幼苗形成率在共生培养50天时就已趋于饱和, FDd1真菌能快速促进齿瓣石斛种子萌发并有效形成幼苗, 对于附生兰科植物在自然条件下种群幼苗的成功建立至关重要(Huang et al., 2018). ...
Symbiotic seed germination and evidence for in vitro mycobiont specificity in Spiranthes brevilabris(Orchidaceae) and its implications for species-?level conservation 1 2007
... 长期以来, 用于兰科植物种子共生萌发相关研究的真菌多是从野生成年植株的根中分离和筛选获得(Masuhara & Katsuya, 1994; Zelmer et al., 1996; Brundrett et al., 2003; Stewart & Kane, 2007; Nontachaiyapoom et al., 2010), 由于野生植株根中可能存在作用未知的不同内生真菌, 得到的菌株往往是随机的, 更为重要的是, 成年植株根中的真菌对种子萌发是否有效并不确定(Zettler et al., 2005), 然而, 很多研究结果都表明所获得的真菌能有效促进种子萌发, 这样的结果在很大程度上是由于采用了Arditti (1967)的种子萌发标准, 即把种胚膨大并产生根状物即视为种子萌发, 并没有关注真菌对后续幼苗形成和发育的影响.而兰科植物在种子萌发后都要经历或短或长的非光合作用阶段, 完全依靠真菌提供有机碳(Rasmussen et al., 2015), 非亲和的真菌或许能刺激种子萌发, 但并不能支持后续的幼苗发育(Bidartondo & Read, 2008; Rasmussen et al., 2015). ...
In vitro germination of Paphiopedilum seed on a completely defined medium 1 1981
In vitro asymbiotic germination of immature seed and formation of protocorm by Cephalanthera falcata(Orchidaceae) 1 2006
... 胶膜菌科真菌是兰科植物最主要的一种菌根真菌类群, 也是很多石斛属植物种子萌发的有效共生真菌(Liu et al., 2010; Dearnaley et al., 2012; Zi et al., 2014; Huang et al., 2018), 瘤菌根菌属(Epulorhiza)被认为是胶膜菌属(Tulasnella)的无性态属(anamorph/teleomorph pair; Dearnaley et al., 2012).本研究中, SSCDO-5菌株为胶膜菌属真菌, FDd1菌株为瘤菌根菌属真菌, 能分别有效地促进铁皮石斛和齿瓣石斛原球茎和幼苗形成, 在幼苗形成和发育阶段表现出了很强的专一性(图2), 这和我们先前的研究结果一致(Huang et al., 2018; Shao et al., unpublished data), 而这种专一性的共生关系是如何形成和维系的, 兰科植物种子和对应真菌是如何相互选择和识别的, 其机制并不清楚.在对不同类群兰科植物种子的非共生萌发研究中都发现, 未成熟的种子比成熟的种子更容易萌发, 因为成熟种子的种皮会导致种子透水性降低(Ramsay & Stewart, 1998; Yamazaki & Miyoshi, 2006; Lee, 2007; Kauth et al., 2008; Long et al., 2010), 而在本研究和我们先前的研究中都发现, 与作为对照的营养丰富培养基处理相比较, 在亲和真菌的接菌处理中, 幼苗形成所需要的时间显著较短, 同时具有显著高的幼苗形成率(Zhou & Gao, 2016), 这是否也暗示种子的种皮结构和真菌建立了特定的信号传导和识别机制, 一方面使得亲和的真菌能快速侵染种子, 促进种子萌发和幼苗形成, 另一方面可以防止不亲和真菌的侵染? ...
Fungi associated with terrestrial orchid mycorrhizas, seeds and protocorms 1 1996
... 长期以来, 用于兰科植物种子共生萌发相关研究的真菌多是从野生成年植株的根中分离和筛选获得(Masuhara & Katsuya, 1994; Zelmer et al., 1996; Brundrett et al., 2003; Stewart & Kane, 2007; Nontachaiyapoom et al., 2010), 由于野生植株根中可能存在作用未知的不同内生真菌, 得到的菌株往往是随机的, 更为重要的是, 成年植株根中的真菌对种子萌发是否有效并不确定(Zettler et al., 2005), 然而, 很多研究结果都表明所获得的真菌能有效促进种子萌发, 这样的结果在很大程度上是由于采用了Arditti (1967)的种子萌发标准, 即把种胚膨大并产生根状物即视为种子萌发, 并没有关注真菌对后续幼苗形成和发育的影响.而兰科植物在种子萌发后都要经历或短或长的非光合作用阶段, 完全依靠真菌提供有机碳(Rasmussen et al., 2015), 非亲和的真菌或许能刺激种子萌发, 但并不能支持后续的幼苗发育(Bidartondo & Read, 2008; Rasmussen et al., 2015). ...
Protocorm mycobionts of the federally threatened eastern prairie fringed orchid,Platanthera leucophaea(Nutt.) Lindley, and a technique to prompt leaf elongation in seedlings 1 2005
... 长期以来, 用于兰科植物种子共生萌发相关研究的真菌多是从野生成年植株的根中分离和筛选获得(Masuhara & Katsuya, 1994; Zelmer et al., 1996; Brundrett et al., 2003; Stewart & Kane, 2007; Nontachaiyapoom et al., 2010), 由于野生植株根中可能存在作用未知的不同内生真菌, 得到的菌株往往是随机的, 更为重要的是, 成年植株根中的真菌对种子萌发是否有效并不确定(Zettler et al., 2005), 然而, 很多研究结果都表明所获得的真菌能有效促进种子萌发, 这样的结果在很大程度上是由于采用了Arditti (1967)的种子萌发标准, 即把种胚膨大并产生根状物即视为种子萌发, 并没有关注真菌对后续幼苗形成和发育的影响.而兰科植物在种子萌发后都要经历或短或长的非光合作用阶段, 完全依靠真菌提供有机碳(Rasmussen et al., 2015), 非亲和的真菌或许能刺激种子萌发, 但并不能支持后续的幼苗发育(Bidartondo & Read, 2008; Rasmussen et al., 2015). ...
Highly compatible Epa-01 strain promotes seed germination and protocorm development of Papilionanthe teres(Orchidaceae) 3 2016
 ,
, ,*云南省植物繁殖适应与进化生态学重点实验室, 昆明 650500;云南大学生态学与进化生物学实验室, 昆明 650500
,*云南省植物繁殖适应与进化生态学重点实验室, 昆明 650500;云南大学生态学与进化生物学实验室, 昆明 650500
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT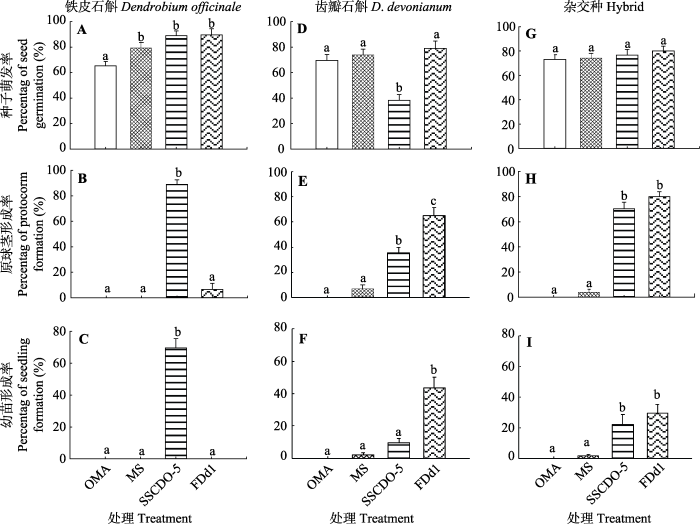 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}