闫雅楠
1 , 2 , 叶小齐
, 1 , * , 吴明
1 , 闫明
2 , 张昕丽
1 1 中国林业科学研究院亚热带林业研究所, 杭州湾湿地生态系统定位观测研究站, 浙江富阳 311400 2 山西师范大学生命科学学院, 山西临汾 041000 Diversity and potassium-solubilizing activity of rhizosphere potassium-solubilizing bacteria of invasive Solidago canadensis Ya-Nan YAN
1 , 2 , Xiao-Qi YE
, 1 , * , Ming WU
1 , Ming YAN
2 , Xin-Li ZHANG
1 1 Research Institute of Subtropical Forestry, Chinese Academy of Forestry, Wetland Ecosystem Research Station of Hangzhou Bay, Fuyang, Zhejiang 311400, China 2 School of Life Science, Shanxi Normal University, Linfen, Shanxi 041000, China 通讯作者: * mengxqi@126.com 编委: 于飞海
责任编辑: 李敏, 赵航(实习)
收稿日期: 2019-02-28
修回日期: 2019-05-29
网络出版日期: 2019-06-20
Corresponding authors: * mengxqi@126.com Received: 2019-02-28
Revised: 2019-05-29
Online: 2019-06-20
Fund supported: Supported by the National Natural Science Foundation of China (31770578 )
摘要 入侵植物加拿大一枝黄花(
Solidago canadensis )具有较强的钾(K)富集能力, 这可能和其对土壤微生物群落的改变有关。根际解钾菌能够将植物难以利用的矿物态钾转化为植物可以利用的可溶性钾, 而加拿大一枝黄花如何影响根际解钾菌多样性和解钾活性尚未明了。该研究以浙江省杭州湾湿地围垦区内自然生长的加拿大一枝黄花和其伴生本地植物白茅(
Imperata cylindrica )为研究对象, 比较了加拿大一枝黄花和白茅体内及土壤中的钾含量水平, 钾供给水平对生物量积累的影响, 以及根际解钾菌的数量、多样性和解钾活性的差异。结果表明, 加拿大一枝黄花茎、叶中的钾含量均显著高于白茅, 分别是白茅的1.59和7.33倍; 加拿大一枝黄花和白茅的土壤全钾含量差异不显著, 速效钾含量在0-10 cm土层中差异显著、在10-20 cm土层中差异不显著。随着钾供应水平提高, 加拿大一枝黄花和白茅的生物量均显著增加。利用解钾培养基计数培养后发现, 加拿大一枝黄花根际解钾菌的数量是白茅的3.51倍。分离培养后将出现解钾圈的菌株进行鉴定, 利用解钾液体培养实验测定其解钾量, 发现从加拿大一枝黄花根际土中分离得到的15个解钾菌株中, 有9个具有高效解钾能力, 其处理液中K
+ 含量较空白对照高出85.11%-192.54%, 其中菌株H2-20解钾能力最强, 解钾量为10.657 mg·L
-1 。加拿大一枝黄花根际解钾菌解钾作用显著高于白茅。经16S rDNA鉴定发现, 加拿大一枝黄花15个根际解钾菌株分属11个属, 其中有6个属已经被报道证实具有明显解钾能力。这些结果表明加拿大一枝黄花根际解钾菌数量较为丰富, 且大多具有较高解钾活性, 可能对其钾富集具有重要贡献。
关键词: 入侵植物 ;
根际解钾菌 ;
解钾能力 ;
入侵机制 ;
系统发育树 Abstract Aims Solidago canadensis, an invasive herbaceous species, has a strong capacity of potassium enrichment, that may relate to its influence on soil microbial community. Rhizosphere potassium-soluble bacteria can convert mineral potassium into soluble forms being able to be used by plants. It is not known how invasion of
S. canadensis may affect diversity and potassium-solubilizing activity of the potassium-solubilizing bacteria.
Methods We compared
S. canadensis and its coexisting native plant
Imperata cylindrica in the reclaimed Hangzhou Bay wetland, Zhejiang Province. We compared the potassium contents of soil and the plant tissues of
S. canadensis and
Imperata cylindrica which coexists with the invasive species, the effect of potassium supply level on biomass accumulation of plants, and the quantity, diversity and potassium-soluble activity of the rhizosphere potassium-solubilizing bacteria.
Important findings The potassium contents in stem and leaf of
S. canadensis were significantly higher (1.59 and 7.33 times respectively) than that of
I. cylindrica, the contents of available potassium in the 0-10 cm soil layer where the two species grew were significantly different, but not in the 10-20 cm soil layer. Potassium application experiments showed significant biomass increase in both
S. canadensis and
I. cylindrica, and tissue potassium concentrations as well. Potassium-dissolving medium culture results showed that the number of potassium-?solubilizing bacteria of
S. canadensis rhizosphere was 2.51 times higher than that of
I. cylindrica . The strains with potassium-dissolving rings were identified, and the amount of released potassium was determined. Among the 15 strains of potassium-solubilizing bacteria isolated from the rhizosphere soil of
S. canadensis , nine efficiently dissolved potassium, and the content of K
+ in the treatment solution was 85.11%-192.54% higher than that in the control. Strain H2-20 had the strongest ability with the dissolved K
+ of 10.657 mg·L
-1 . The potassium- solubilizing effect of rhizosphere potassium-solubilizing bacteria of
S. canadensis was significantly higher than that of
I. cylindrica . According to 16S rDNA identification, the 15 strains of bacteria associated with
S. canadensis were of 11 genera, and 6 of them had been reported to have the potassium-solubilizing ability. Our results suggest that potassium-solubilizing bacteria in the rhizosphere of
S. canadensis is abundant, and may play an important role in potassium enrichment.
Keywords: invasive plant ;
rhizosphere potassium-solubilizing bacteria ;
potassium capacity ;
intrusion mechanism ;
phylogenetic tree PDF (1756KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 闫雅楠, 叶小齐, 吴明, 闫明, 张昕丽. 入侵植物加拿大一枝黄花根际解钾菌多样性及解钾活性. 植物生态学报, 2019, 43(6): 543-556. DOI:
10.17521/cjpe.2019.0045 YAN Ya-Nan, YE Xiao-Qi, WU Ming, YAN Ming, ZHANG Xin-Li.
Diversity and potassium-solubilizing activity of rhizosphere potassium-solubilizing bacteria of invasive Solidago canadensis .
Chinese Journal of Plant Ecology , 2019, 43(6): 543-556. DOI:
10.17521/cjpe.2019.0045 外来植物入侵不仅导致入侵地农林生产经济损失, 而且对生态系统和生物多样性造成了严重的威胁(
鞠瑞亭等, 2012 )。据调查, 我国外来入侵植物多达670余种, 分布范围主要集中在东南沿海地区, 且入侵速度正在逐年加快、数量逐年增多, 若不加以防治, 则直接影响我国经济发展和物种栖息地保护(
闫小玲等, 2012 )。了解外来植物的入侵机制是制定科学合理的防控方案的关键。为了解释外来植物的入侵机制, 研究者提出了很多研究假说, 例如天敌逃逸假说(
Keane et al ., 2002 )、竞争能力增强进化假说(
Blossey & N?tzold, 1995 ), 但这些假说主要针对入侵植物本身的进化, 而没有考虑入侵植物与土壤微生物的相互作用。随着研究的不断深入, 越来越多的****意识到植物入侵过程与地下生态存在紧密联系(
de la Pe?a et al ., 2010 ;
Ren & Zhang, 2009 ), 入侵植物与土壤微生物之间的互作在入侵过程中发挥重要作用(
Wardle et al ., 2004 )。
外来植物在入侵过程中可能通过释放根系分泌物和改变微生物碳源等方式影响土壤微生物结构和功能, 而土壤微生物群落的改变反过来会影响外来植物的进一步入侵(
杨星等, 2012 ;
Uddin et al ., 2014 ;
Zhao et al ., 2014 ;
Martin et al ., 2017 )。以往研究表明, 紫茉莉(
Mirabilis jalapa )、破坏草(紫茎泽兰)(
Ageratina adenophora )、互花米草(
Spartina alterniflora )可以改变入侵地土壤微生物多样性(
牛红榜, 2007 ;
Zhao et al ., 2014 ;
廖丹等, 2018 ); 银荆(
Acacia dealbata )入侵西班牙北部地区后使当地土壤中细菌丰富度显著增加, 真菌的丰富度和多样性显著减少(
Lorenzo et al ., 2010 ); 旱雀麦(
Bromus tectorum )使入侵地的细菌数量显著增多, 真菌数量显著减少(
Belnap & Phillips, 2001 ); 葱芥(
Alliaria petiolata )入侵后导致土壤中丛枝菌根真菌数量显著减少(
Roberts & Anderson, 2001 )。这些结果说明, 外来植物入侵能够导致土壤微生物多样性发生明显改变。而另一方面, 土壤微生物的改变可能促进外来植物的入侵, 甚至可能抑制本地植物的生长。例如, 土壤微生物显著提高了入侵植物加拿大一枝黄花(
Solidago canadensis )的出芽率和资源竞争力, 而降低了本土植物长芒草(
Stipa bungeana )的竞争力(
Sun & He, 2010 )。因此, 探究入侵植物对土壤微生物的影响可能是了解植物入侵机理的关键所在。
加拿大一枝黄花是菊科一枝黄花属植物, 多年生草本, 具有发达根状茎, 原产于北美地区, 最初作为庭园花卉引入我国上海、南京等地, 现已广泛逸生为恶性外来种(
中国科学院中国植物志编辑委员会, 1985 ;
郭水良和方芳, 2003 )。加拿大一枝黄花繁殖能力极强, 既能利用种子进行有性繁殖, 也可通过地下根状茎进行无性繁殖, 一般情况下, 每株植株可产约7万粒种子, 且种子具有较高萌发率, 此外, 植株根部具有上百个根茎结构, 每个根茎在第2年可萌发形成独立的个体, 如此高的繁殖能力使其在入侵后能够迅速扩散形成单优势种群落(
王玉良等, 2009 ;
张海亮等, 2015a )。此外, 加拿大一枝黄花根部还会产生一些化感物质影响周围土著植物生长, 对入侵地生物多样性造成了一定的影响(
方芳等, 2004 )。而目前加拿大一枝黄花在我国入侵范围逐渐扩大, 入侵速度逐渐加快, 对我国生态环境、生物多样性、农业和畜牧业均造成了不同程度的影响(
董梅等, 2006 ;
陆慧明等, 2006 ), 因此亟须加强加拿大一枝黄花入侵机理与防控技术方面的研究。
目前国内外对加拿大一枝黄花入侵机制的研究主要集中在繁殖特性(
黄华和郭水良, 2005 ;
郝建华等, 2009 ;
张海亮等, 2015b )、化感作用(
梅玲笑, 2005 ;
郝建华等, 2009 )、生理生态特征与环境适应性(
郭水良和方芳, 2003 ;
杜乐山, 2014 )以及土壤微生物响应(
Zhang et al ., 2005 ;
沈荔花等, 2007 )等方面。研究表明, 在不同程度的入侵地中, 加拿大一枝黄花体内钾含量均高于伴生植物, 尤其在生长期和开花期差异最为显著, 且植物吸钾量与生物量呈正相关关系(
周振荣, 2010 )。在微生物响应方面, 研究发现加拿大一枝黄花入侵后能改变土壤微生物的群落数量、结构和功能多样性, 可能有助于加拿大一枝黄花进一步入侵(
Liao et al ., 2013 ;
Wang et al ., 2018 )。根际解钾菌能将土壤中难溶性钾转化为可被植物直接吸收的有效性钾, 进而促进植物钾吸收, 增强植物抗逆性(
李元芳, 1994 ;
郭勋斌等, 2001 )。加拿大一枝黄花入侵是否改变了根际解钾菌数量、结构和解钾活性等问题还有待深入研究。因此, 本研究以入侵植物加拿大一枝黄花和伴生本地植物白茅(
Imperata cylindrica )为研究对象, 比较加拿大一枝黄花与白茅的根际解钾菌数量、多样性与解钾能力差异, 旨在初步探讨根际解钾菌在加拿大一枝黄花入侵过程中发挥的作用, 为深入研究加拿大一枝黄花入侵机制提供理论依据。
1 材料和方法 1.1 样品采集 采样点位于杭州湾国家湿地公园生态保育区内(121.17° E, 30.32° N)。该区域地处浙江省宁波慈溪市西北部, 属北亚热带季风气候, 地势平坦, 生境类型一致, 远离城市和人为干扰, 植被自然生长, 土壤pH值8-9, 含盐量在0.2%左右, 植物物种多样性相对较低, 主要为单优势草本群落, 典型的有加拿大一枝黄花群落、白茅群落、芦苇(
Phragmites australis )群落等。入侵植物加拿大一枝黄花和伴生本地植物白茅均为多年生草本植物, 具有发达的根状茎, 同期发育, 8月份生长趋于稳定(
梁雷等, 2016 )。
植物和土壤样品采集于8月份, 分别选取以加拿大一枝黄花和白茅为优势种的5个不同样地, 各样地间距超过200 m, 每个样地设置1 m × 1 m小样方, 采集加拿大一枝黄花和白茅新鲜的茎和叶, 用于测定植物茎和叶中的氮磷钾含量; 按照四分法采集0-10 cm、10-20 cm土层的新鲜土壤样品, 用于测定土壤中的养分含量; 同时采集植物根系表面不易抖落的土壤, 即根际土, 将采集好的根际土壤样品装于无菌袋密封低温保存, 用于分离各植物的根际解钾菌。加拿大一枝黄花和白茅的种子样品分别采集于12月和5月, 选取自然成熟的种子进行采集, 用于植物的室内培养试验。将各样品编号, 带回实验室。
1.2 样品处理及氮磷钾含量测定 植物样品用蒸馏水清洗干净, 于105 ℃杀青2 h, 60 ℃烘干至恒质量, 冷却后剪碎, 用HNO
3 -H
2 O
2 消化后, 用火焰光度计法测植物氮磷钾含量。不同土层的土壤样品风干、敲碎、挑除杂质, 研磨至过100目网筛, 用半微量开氏法检测土壤全氮含量、碱解扩散法测定土壤碱解氮含量、0.03 mol·L
-1 NH
4 F-0.025 mol·L
-1 HCl法测定土壤有效磷含量、HClO
4 -H
2 SO
4 法检测土壤全磷含量、NaOH熔融-火焰光度计法测土壤全钾含量、NH
4 OAc浸提-火焰光度计法测定土壤速效钾含量(
鲍士旦, 2000 )。样品检测委托国家林业局经济林产品质量检验检测中心 进行。
1.3 施加钾培养 分别取加拿大一枝黄花和白茅的种子约100粒, 播种于铺有滤纸的培养皿中, 加蒸馏水至种子完全浸没, 将培养皿带盖置于培养箱中培养4-5天, 设置光照、黑暗环境各12 h, 温度分别设置28 ℃和22 ℃。待植物子叶展开后, 挑选大小均等的幼苗移栽入装有石英砂的花盆中, 每盆移栽5株, 置于温室进行培养, 前期浇灌蒸馏水, 3天后开始浇灌营养液, 营养液以无钾霍格兰营养液为基础液(
蔡庆生, 2013 ), 钾添加量设置3个水平: 0 mol·L
-1 (缺钾)、0.1 × 10
-3 mol·L
-1 (低钾)、6 × 10
-3 mol·L
-1 (高钾), 每个处理重复15次。10天后进行间苗, 每盆保留一株。营养液处理90天后结束培养。
培养结束后将植物连根取出, 表面清洗干净后, 置于105 ℃下杀青15 min, 再于60 ℃下烘干至干物质质量不再发生改变后, 称量并记录各处理下植物的总生物量。
1.4 解钾菌的计数培养 无菌条件下, 取1 g植物根际土, 溶于装有99 mL无菌水的三角瓶中, 配制成体积质量为1 × 10
-2 g·mL
-1 的土壤悬浮液, 于28 ℃、120 r·min
-1 条件下振荡培养30 min, 使微生物均匀分散。将土壤悬浮液依次稀释为1 × 10
-3 、1 × 10
-4 、1 × 10
-5 g·mL
-1 3个不同体积质量, 取0.1 mL涂布于解钾培养基(配方见
表1 )表面, 每个体积质量重复3次, 接种后将培养皿倒置, 置于28 ℃恒温环境下培养3天后进行计数(
林先贵, 2010 )。
Table 1 表1 表1 解钾培养基配方
Table 1
Formula of culture medium for potassium-solubilizing bacteria used in this experiment 成分 含量 -1 ) 成分 含量 -1 ) 蔗糖 Sucrose 5.0 Na2 HPO4 2.0 MgSO4 0.5 FeCl3 0.005 CaCO3 0.1 土壤矿物 1.0 琼脂 Agar 18.0 蒸馏水 Distilled water 1 000
新窗口打开 |
下载CSV 1.5 解钾菌的筛选与鉴定 计数完成后, 继续培养2-3天, 待菌落生长旺盛时, 挑选菌斑较大, 凸起较高, 黏稠有弹性, 且有明显解钾圈的菌落进行划线分离培养, 反复划线3次, 直至得到纯种菌株, 记录菌落形态特征、解钾圈半径(
R )、菌落半径(
r ), 计算根际解钾菌解钾圈直径与菌落直径比(
R /
r ), 以表征该种解钾菌解钾能力的大小。并将纯种菌株送至浙江天科高新技术发展有限公司进行16S基因序列的PCR扩增、序列测定和分析。测序结果提交至GenBank获取基因登录号, 同时利用BLAST进行同源性分析, 挑选出同源性较高的相邻种、属16S rDNA序列, 用CLUSTALX 1.8和Phylip软件构建系统发育树。
1.6 解钾菌的解钾能力测定 1.6.1 配制菌悬液
配制Luria-Bertani液体培养基(以下简称LB液体培养基), 用于菌种的扩增培养, 每瓶分装50 mL, 将筛选出的20株根际解钾菌分别接种一环至LB培养基中, 28 ℃, 120 r·min
-1 摇床培养3天后, 取1 mL菌液加入无菌水中制成1 × 10
-7 的解钾菌悬浮液简称菌悬液, 充分摇匀, 取100 μL菌悬液于解钾培养基中, 30 ℃恒温培养24 h, 对菌斑进行计数, 检验菌悬液密度是否达到1 × 10
8 CFU·mL
-1 (CFU, 菌落形成单位), 达到后备用。分别从20株解钾菌菌悬液(活菌数约为1 × 10
8 CFU·mL
-1 )中取5 mL菌悬液, 混合, 121 ℃灭菌30 min, 备用。LB液体培养基配方: 酵母膏5 g、蛋白胨10 g、NaCl 10 g, 溶解于1 000 g蒸馏水中。
1.6.2 解钾能力测定
配制解钾液体培养基, 每瓶分装100 mL, 取5 mL菌悬液(活菌数约为1 × 10
8 CFU·mL
-1 )加入解钾液体培养基中, 以混合灭菌后的菌悬液为对照, 每个处理重复5次, 28 ℃、120 r·min
-1 摇床培养7天后, 将菌悬液4 500 r·min
-1 离心20 min, 收集上清液, 加2 mL 6% H
2 O
2 消煮1 h, 再离心, 取上清液, 用电感耦合等离子体光谱法测定上清液中水溶性钾含量, 将接种了解钾菌菌悬液的处理与对照处理进行比较, 计算出20株解钾菌的解钾量。
(1) $解钾菌的解钾量(mg·L^{-1})=菌悬液中K^{+}含量-对照液中K^{+}含量$ 1.7 数据处理 数据处理和分析软件使用Microsoft Excel 2013和SPSS 17.0。加拿大一枝黄花和白茅茎、叶中的氮、磷和钾含量, 加拿大一枝黄花和白茅根际解钾菌的数量、
R /
r 值以及解钾作用差异, 以上数据均进行独立样本
t 检验, 显著性水平均设为
α = 0.05; 各植物样地不同土层的土壤中养分含量差异和施钾培养后加拿大一枝黄花和白茅的生物量差异采用单因素方差分析处理, 显著性水平设为
α = 0.05。系统发育树用Figtree version 1.4.2进行修饰, 作图软件使用Origin 8.0。
2 结果 2.1 加拿大一枝黄花和白茅植物及土壤中氮磷钾含量水平 在叶片中, 加拿大一枝黄花的氮磷钾含量均极显著多于白茅(
p < 0.01), 分别是白茅含量的3.41、4.28和7.33倍; 在茎中, 加拿大一枝黄花的氮含量极显著少于白茅(
p < 0.01), 约为白茅的0.60倍, 磷含量与白茅无显著差异, 钾含量极显著高于白茅(
p < 0.01), 且约为白茅的1.59倍(
图1 )。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1加拿大一枝黄花和白茅叶和茎中的氮磷钾含量(平均值±标准误差)。**, p < 0.01; ns, 差异不显著。 Fig. 1Contents of nitrogen, phosphorus and potassium in leaves and stems of Solidago canadensis and Imperata cylindrica (mean ± SE ). **, p < 0.01; ns, no significant difference. 在0-10 cm和10-20 cm两个土层中, 加拿大一枝黄花的全氮含量和全钾含量与白茅差异不显著(
p > 0.05), 而其碱解氮含量、全磷含量和有效磷含量均显著多于白茅(
p < 0.05;
图2 )。其中, 在0-10 cm和10-20 cm两个土层中, 加拿大一枝黄花的碱解氮含量分别是白茅的2.59和2.48倍, 全磷含量分别是白茅的1.19和1.21倍, 有效磷含量分别是白茅的1.69和1.70倍(
图2 )。在0-10 cm土层中加拿大一枝黄花的有效钾含量显著高于白茅(
p < 0.05), 而在10-20 cm土层中其差异不显著(
p > 0.05;
图2 )。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2加拿大一枝黄花和白茅群落土壤不同土层中的养分含量(平均值±标准误差)。不同小写字母表示不同处理间差异显著(p < 0.05); ns, 差异不显著(p > 0.05)。 Fig. 2Total N, total P and total K contents in different soil layers where Solidago canadensis and Imperata cylindrica grew (mean ± SE ). Different lowercase letters indicate significant differences among the treatments (p < 0.05); ns, no significant difference (p > 0.05). 2.2 钾对加拿大一枝黄花干物质积累的影响 加拿大一枝黄花和白茅的生物量积累均随钾供应水平提高而增加(
p < 0.05;
图3 )。不同钾供应水平条件下, 白茅生物量和加拿大一枝黄花生物量没有显著差异(
p > 0.05;
图3 ), 但在高钾供应水平条件下, 加拿大一枝黄花生物量略高于白茅。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3不同钾处理下加拿大一枝黄花和白茅生物量对比(平均值±标准误差)。不同小写字母表示各植物不同处理间差异显著(p < 0.05)。 Fig. 3A biomass comparison of Solidago canadensis and Imperata cylindrica under different potassium treatments (mean ± SE ). Different lowercase letters indicate significant differences among the treatments of different plants (p < 0.05). 2.3 不同植物根际解钾菌数量比较 加拿大一枝黄花根际解钾菌数量显著高于白茅(
p < 0.05;
图4 )。5个样地依次高出140.66%、62.46%、235.69%、283.67%和533.33%, 平均约超出3.51倍。
图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4加拿大一枝黄花和白茅根际解钾菌数量对比(平均值±标准误差)。*, p < 0.05。 Fig. 4A comparison of potassium-solubilizing bacteria numbers in rhizosphere of Solidago canadensis and Imperata cylindrica (mean ± SE ). *, p < 0.05. 2.4 解钾菌初筛 加拿大一枝黄花根际土中初步筛选出15株解钾菌, 白茅根际土中初步筛选出5株解钾菌。分别记录菌落形态特征、解钾圈直径(
R )及菌落直径(
r )(
表2 ,
表3 ), 加拿大一枝黄花根际解钾菌解钾圈直径与菌落直径比(
R /
r )均值为1.396, 大于1.5的有6株, 分别是H3-8、H2-14、H2-16、H2-17、H2-20和H1-21, 最大值为1.68 (H1-21); 白茅根际解钾菌的
R /
r 均值为1.252, 全部小于1.5, 其中最大值为1.34 (B1-6)。两种植物根际解钾菌的
R /
r 值差异显著(
p < 0.05)。
Table 2 表2 表2 加拿大一枝黄花15株根际解钾菌菌落特征和解钾能力
Table 2
Colony characteristics and potassium-solubilizing ability of 15 strains of rhizosphere potassium-solubilizing bacteria from Solidago canadensis 菌株编号 Strain No. 菌落形态特征 解钾圈半径 R )(cm) 菌落半径 r )(cm) R /r H5-1 菌斑小且不规则, 白色 Plaque small and irregular, white 0.29 0.26 1.14 H1-3 菌斑大, 凸起低, 色素淡且均匀 Plaque large, low protuberance, light and uniform pigmentation 1.08 0.93 1.16 H1-4 不规则凸起, 透明, 较少色素 Irregular protuberance, transparency, less pigmentation 0.56 0.48 1.17 H6-5 小液滴状, 边缘规则, 色素呈放射状 Small droplets with regular edges and radial pigments 0.47 0.45 1.04 H3-8 菌斑小且不规则, 色素深呈放射状 plaque small and irregular, dark and radial pigments 0.42 0.25 1.68 H5-10 不规则凸起, 菌斑乳白色, 边缘黄色沉着 Irregular protuberance, milky white plaque, yellow edge 0.66 0.53 1.26 H1-12 不规则液滴状, 边缘模糊, 中央色素沉着 Irregular droplet shape, blurred edge, central pigmentation 0.78 0.66 1.18 H2-14 液滴状, 凸起高, 色素呈圆形 Droplet-shaped, tall protuberance, rounded pigments 0.84 0.54 1.56 H1-15 菌斑小, 边缘不规则, 色素呈伞状 Plaque small, irregular margin, umbrella-shaped pigments 0.56 0.38 1.46 H2-16 不规则液滴状, 浑浊, 中央色素沉着 Irregular droplet, turbid, central pigmentation 0.62 0.40 1.55 H2-17 规则小液滴凸起, 色素少 Regular small droplets protruding, less pigmentation 0.52 0.32 1.63 H5-18 液滴状, 透明, 中央色素沉着 Droplet-like, transparent, central pigmentation 0.72 0.52 1.38 H1-19 边缘锯齿形, 液滴状, 色素呈同心圆状 Edge serrated, droplet-shaped, concentric circle of pigments 1.10 0.74 1.49 H2-20 菌斑较大, 规则液滴状, 色素均匀 Plaque large, regular droplet shape, uniform pigmentation 1.00 0.64 1.56 H1-21 规则液滴状, 凸起高, 透明, 色素少 Regular droplet shape, high protrusion, transparent, less pigmentation 0.94 0.56 1.68
新窗口打开 |
下载CSV Table 3 表3 表3 白茅5株根际解钾菌菌落形态特征和解钾能力
Table 3
Colony characteristics and potassium-solubilizing ability of 5 strains of rhizosphere potassium-solubilizing bacteria from Imperata cylindrica 菌株编号 菌落形态特征 解钾圈半径R )(cm) 菌落半径r )(cm) R /r B1-6 凸起低, 浑浊, 中央色素呈同心圆状 Low protrusion, turbid, concentric circle of central pigments 0.74 0.55 1.34 B6-7 不规则凸起, 浑浊 Irregular bulge, turbidity 0.65 0.55 1.18 B2-9 菌斑小且不规则, 白色, 无凸起 Plaque small and irregular, white, without protuberance 0.92 0.70 1.31 B4-11 规则液滴状, 凸起较高, 色素淡且均匀 Regular droplet shape, higher protuberance, light and uniform 0.50 0.41 1.22 B6-13 液滴状, 凸起高, 色素均匀沉着 Droplet-like, protruding, uniformly pigmented 0.56 0.46 1.21
新窗口打开 |
下载CSV 2.5 解钾菌16S rDNA鉴定 利用细菌16S rDNA基因通用引物扩增获得的DNA片段长约1 500 bp, 同源性分析结果见
表4 、
表5 。
Table 4 表4 表4 分离自加拿大一枝黄花的根际解钾菌系统发育分析
Table 4
Phylogenetic analysis of rhizosphere potassium-solubilizing bacteria from Solidago canadensis. 菌株编号 登录号 最相近菌株(登录号) 序列相似性 H5-1 MH490984 Pseudoflavitalea soli KIS20-3 (NR_148655)96% H2-3 MH490985 Mitsuaria sp. SS48 (HQ891978)99% H1-4 MH490986 Rhizobium sp. KMM 9576 (LC126306)100% H6-5 MH490987 Microbacterium sp. 3B2 (MG763154)99% H3-8 MH490990 Streptomyces variabilis SD22 (MH244336)98% H5-10 MH490992 Azotobacter chroococcum YCYS (JQ692178)99% H1-12 MH490994 Stenotrophomonas panacihumi 5-III (KP969077)99% H2-14 MH490996 Pseudomonas sp. EA_S_32 (KJ642336)98% H1-15 MH490997 Cupriavidus sp. FZ96 (KF803333)99% H2-16 MH490998 Ensifer adhaerens WJB133 (KU877667)100% H2-17 MH490999 Rhizobium taeanense PSB 2-6 (DQ114473)99% H5-18 MH491000 Pseudoflavitalea soli KIS20-3 (NR_148655)96% H1-19 MH491001 Filimonas endophytica SR 2-06 (KJ572396)99% H2-20 MH491002 Lysobacter niastensis GH41-7 (NR_043868)99% H1-21 MH491003 Siphonobacter aquaeclarae HPG59 (JQ291601)99%
新窗口打开 |
下载CSV Table 5 表5 表5 分离自白茅的根际解钾菌系统发育分析
Table 5
Phylogenetic analysis of rhizosphere potassium-solubilizing bacteria from Imperata cylindrica 菌株编号 登录号 最相近菌株(登录号) 序列相似性 B1-6 MH490988 Microbacterium imperiale (JN585685)99% B6-7 MH490989 Enterobacter sp. PRd5 (KY203970)99% B2-9 MH490991 Alloactinosynnema album 03-9939 (NR_116323)99% B4-11 MH490993 Chryseolinea sp. SDU1-6 (MG662377)95% B6-13 MH490995 Pseudoflavitalea soli KIS20-3 (NR_148655)96%
新窗口打开 |
下载CSV 加拿大一枝黄花15株根际解钾菌主要分为5大类群: 变形菌门α亚群(
Alphaproteobacteria )、变形菌门β亚群(
Betaproteobacteria )、变形菌门γ亚群(
Gammaproteobacteria )、拟杆菌门(
Bacteroidetes )和放线菌门(
Actinobacteria ), 白茅5株根际解钾菌分为3大类群: 变形菌门γ亚群(
Gammaproteobacteria )、拟杆菌门(
Bacteroidetes )和放线菌门(
Actinobacteria )。利用邻接法(NJ法)建立系统发育树, 发现加拿大一枝黄花15株根际解钾菌属于11个属, 白茅5株根际解钾菌均为不同属(
图5 ,
图6 )。
图5 新窗口打开 |
下载原图ZIP |
生成PPT 图5加拿大一枝黄花15株根际解钾菌的系统发育树。图中数字为bootstrap值, 用于检验进化树分支可信度, 数值越大可信度越高。 Fig. 5Phylogenetic tree of 15 strains rhizosphere potassium-solubilizing bacteria of Solidago canadensis . The numbers in the figure refer the bootstrap values for testing the reliability of branch of evolutionary tree, the greater the value, the higher the reliability. 图6 新窗口打开 |
下载原图ZIP |
生成PPT 图6白茅5株根际解钾菌的系统发育树。图中数字为bootstrap值, 用于检验进化树分支可信度, 数值越大可信度越高。 Fig. 6Phylogenetic tree of 5 strains rhizospheric potassium-solubilizing bacteria from Imperata cylindrica . The numbers in the figure refer the bootstrap values for testing the reliability of branch of evolutionary tree, the greater the value, the higher the reliability. 2.6 解钾菌解钾能力比较 加拿大一枝黄花与白茅的根际解钾菌均具有不同程度的解钾作用, 对照组中钾离子含量为5.535 mg·L
-1 , 与对照组相比, 实验组中的K
+ 含量均有所增加, 两种植物的解钾量均为正值。加拿大一枝黄花根际解钾菌解钾量均值为5.025 mg·L
-1 , 其中解钾量大于5 mg·L
-1 的菌株共有9株, 占比为60%, 解钾能力最强的菌株为H2-20, 解钾量为10.657 mg·L
-1 , 相比于对照组增加了192.54%。白茅根际解钾菌解钾量均值为2.723 mg·L
-1 , 其中解钾量大于5 mg·L
-1 的菌株仅有一株(B1-6), 占全部菌株的20%, 其解钾量为5.053 mg·L
-1 , 较对照组增加了91.29%。
t 检验结果显示加拿大一枝黄花根际解钾菌解钾量显著多于白茅根际解钾菌(
p < 0.05;
图7 )。
图7 新窗口打开 |
下载原图ZIP |
生成PPT 图7加拿大一枝黄花和白茅根际解钾菌解钾量均值及单一值对比(平均值±标准误差)。AVG, 不同根际解钾菌解钾量均值。 Fig. 7A comparison of means and single potassium solubilization amounts of potassium-solubilizing bacteria in rhizosphere of Solidago canadensis and Imperata cylindrica (mean ± SE ). AVG, mean potassium dissolution amount of different rhizosphere potassium-solubilizing bacteria. 3 讨论 入侵植物对营养元素的吸收、积累和释放对其生长、入侵能力及其入侵的生态学效应都具有重要作用。钾是植物生长发育所需要的主要营养元素之一, 在植物生长繁育过程中发挥必不可少的作用, 钾的增加不仅能够提高植物光合速率和酶的活化速度、增加植物糖和蛋白质含量、增强植物对氮的吸收, 还能够提高作物的产量、提高植物的抗逆性(
Graham & Ulrich, 1972 ;
胡笃敬等, 1993 )。本研究发现, 野外自然生长的加拿大一枝黄花叶片和茎中的钾含量显著高于伴生植物白茅, 尤其叶片中的钾含量是白茅的7.33倍, 说明在相同生境下, 加拿大一枝黄花吸收钾的能力远高于本地植物, 且加拿大一枝黄花能够将更多的钾分配到叶片中, 而叶片是光合作用最主要的场所, 叶片中养分含量增多可能增强植物光合速率和生长代谢速率(
Aerts, 2000 ;
王丽芳等, 2013 )。此外, 加拿大一枝黄花叶片中的氮磷含量明显高于茎, 更加证实了加拿大一枝黄花将更多养分分配到叶片中的生长策略。人为提高钾供给水平均能促进两种植物生物量积累增加(
图3 ), 但是我们发现加拿大一枝黄花入侵地土壤0-10 cm及10-20 cm土层中全氮、全磷、全钾及碱解氮、有效磷和速效钾含量均多于对照植物白茅, 尤其土壤中速效养分的含量显著多于白茅, 这与
梁雷等(2016) 的研究结果相一致。
陈彤等(2012) 发现加拿大一枝黄花能显著提高土壤中硝态氮含量, 并且其较高的氮积累能力可能和入侵后本地植物种群的动态变化格局紧密相关 这些研究说明加拿大一枝黄花存在一种能够增强土壤养分积累的机制, 而这种机制反过来可能促进其进一步入侵。外来植物对入侵地土壤养分含量的影响被大量研究。一些研究认为外来植物入侵往往能够通过影响入侵地土壤养分而促进自身成功入侵(
Holmgren et al ., 1997 )。许多外来植物入侵后通过改变土壤肥力影响其与本地种的竞争能力, 例如破坏草、黄顶菊(
Flaveria bidentis )、盐生草(
Halogeton glomeratus )等植物入侵时均会导致土壤养分含量向有利于入侵植物的方向发生改变(
Duda et al ., 2003 ;
蒋智林等, 2008 ;
张天瑞等, 2010 )。
植物与土壤微生物的相互作用在植物入侵过程中起着至关重要的作用(
Bowen et al ., 2017 )。研究表明外来植物通过改变入侵地土壤微生物结构和功能, 进而改善土壤养分供应, 促进植物生长, 增强植物抗逆性, 促使植物成功入侵(
Kourtev et al ., 2002 ;
Wardle et al ., 2004 ;
Nijjer et al ., 2008 ;
Coats et al ., 2014 )。例如, 紫茎泽兰入侵后改变了入侵地土壤微生物的群落结构, 同时提高了入侵地土壤肥力, 进而增强了紫茎泽兰的竞争能力(
牛红榜等, 2007 )。本研究发现加拿大一枝黄花根际土壤中解钾菌数量、群落结构和解钾活性都和伴生植物白茅有着较大的差异, 加拿大一枝黄花的根际解钾菌数量远多于对照, 且其中多数根际解钾菌均具有高效的解钾能力, 同时加拿大一枝黄花茎叶中的钾含量及入侵地土壤中全钾和速效钾的含量均高于对照。这说明, 加拿大一枝黄花的富钾作用可能与其较多的高效解钾菌有关。相关研究也表明加拿大一枝黄花入侵会显著改变土壤微生物群落结构和多样性(
沈荔花等, 2007 ;
陈晨等, 2009 ;
李国庆, 2009 ), 这些转变可能会显著地影响土壤中钾的有效性和供给水平。在其他入侵植物的相关研究中也发现, 入侵植物对土壤养分含量的影响也和其土壤微生物群落的改变紧密相关。例如, 黄顶菊根际耐寒短杆菌能增加土壤磷含量, 对病原菌的抑制作用较对照植物更强(
石青, 2018 ); 破坏草通过促进土壤中解磷菌的生长使其土壤中有效磷含量增加(
杨国庆等, 2014 );
Xanthium italicum 水提液不仅使土壤中细菌和真菌数量明显增多, 同时其土壤速效钾、速效氮等营养成分也显著增多(
邰凤姣等, 2016 )。
本研究发现的加拿大一枝黄花和白茅根际解钾菌的种类和此前报道的研究具有一定的相似性。国际公认的解钾菌菌种仅3种: 环状芽孢杆菌(
Bacillus circulans )、胶质芽孢杆菌(
Bacillus mucilaginosus )、土壤芽孢杆菌(
Bacillus edaphicus ), 但近年来解钾菌的种属鉴定结果越来越丰富, 新增的种属有假单胞菌属(
Pseudomonas )(
吴凡等, 2010 ;
李新新等, 2014 ;
葛红莲和纪秀娥, 2017 )、根瘤菌属(
Rhizobium )、肠杆菌属(
Enterobacteriaceae )(
吴凡等, 2010 )、微杆菌属(
Microbacterium )、固氮菌属(
Azotobacter )(
易浪波等, 2012 ;
李新新等, 2014 )、中华根瘤菌属(
Sinorhizobium )、慢生根瘤菌属(
Mesorhizobium )、屈挠杆菌属(
Flexibacter )(
鞠伟, 2016 ;
李春钢等, 2017 )等。在加拿大一枝黄花和白茅根际土中分离获得的20株解钾菌中, H1-4、B6-7与
吴凡等(2010) 的研究结果一致, H6-5和H1-6、H5-10、H2-14、H2-17与
李新新等(2014) 的研究结果相同, H3-8与
张妙宜等(2016) 筛选出的解钾菌同属, 以上菌株均被证明具有一定的解钾能力。此外, 本研究发现的解钾能力较强但未被报道的菌株有: H1-15, 属于含铜菌属(
Cupriavidus ); H5-18, 属于
Pseudoflavitalea; H2-20, 属于溶杆菌属(
Lysobacter ); H1-21, 属于
Siphonobacter, 这一研究结果有助于进一步丰富解钾菌的种属类别。
加拿大一枝黄花的入侵之所以更够改变根际解钾菌多样性可能与其根系分泌物有关。已有研究表明入侵植物能够通过根系分泌次生代谢物对土壤微生物群落结构造成一定的影响, 加拿大一枝黄花能够通过根系分泌物改变土壤的理化性质, 对不同微生物群落产生一定程度的增强或抑制作用(
沈荔花等, 2007 ;
Zhang et al ., 2007 ,
2009 )。本研究中加拿大一枝黄花根际解钾菌数量与解钾能力与对照植物存在明显差异, 这可能是由于加拿大一枝黄花能够通过特异性的根系分泌物化学组分影响根际解钾菌的数量、种类以及分布, 同时, 加拿大一枝黄花较大的生物量积累也可能使其释放更多的根系分泌物, 增加碳源供给, 从而增强了土壤微生物活性, 使得矿物质态的钾能够更快向水溶性钾转化, 从而导致土壤速效养分的提高。因此关于加拿大一枝黄花根系分泌物释放量及化学组分是如何影响根际功能微生物群落的结构和功能的还需要进一步深入研究。本研究只设置了一种本地伴生植物为对照, 实验结果有一定的局限, 后续研究将扩大研究范围, 增加不同功能类群的伴生物种作为对照, 以期更加深入全面地了解加拿大一枝黄花入侵后对入侵地土壤生态系统的影响及其机理。
致谢 感谢中国林业科学研究院中央级公益性科研院所基金科研业务费专项资金(CAFYBB2016SY010)资助。
[1] Aerts R Chapin III FS 2000 ). The mineral nutrition of wild plants revisited: A re-evaluation of processes and patternsAdvances in Ecological Research, 30, 1 -67 . [本文引用: 1] [2] Bao SD 2000 ). Soil Agricultural Chemistry Analysis. 3rd edn . China Agriculture Press, Beijing. 70 -109 . [本文引用: 1] [ 鲍士旦 (2000 ). 土壤农化分析(第三版) . 中国农业出版社, 北京. 70 -109 .] [本文引用: 1] [3] Belnap J Phillips SL 2001 ). Soil biota in an ungrazed grassland: Response to annual grass (Bromus tectorum ) invasionEcological Applications, 1261 -1275 . [本文引用: 1] [4] Blossey B N?tzold R 1995 ). Evolution of increased competitive ability in invasive nonindigenous plants: A hypothesisJournal of Ecology, 83, 887 -889 . [本文引用: 1] [5] Bowen JL Kearns PJ Byrnes JEK Wigginton S Allen WJ Greenwood M Tran K Yu J Cronin JT Meyerson LA 2017 ). Lineage overwhelms environmental conditions in determining rhizosphere bacterial community structure in a cosmopolitan invasive plantNature Communications, 8, 433 . DOI: 10.1038/s41467-017-00626-0 . [本文引用: 1] [6] Cai QS 2013 ). Plant Physiology Experiments . China Agricultural University Press, Beijing. 8 -9 . [本文引用: 1] [ 蔡庆生 (2013 ). 植物生理学实验 . 中国农业大学出版社, 北京. 8 -9 .] [本文引用: 1] [7] Chen C Fu YY Huang L Wang YJ He XL Xiao M 2009 ). The analysis of the diversity of the partial rhizosphere bacteria of Solidago canadensis L. on Chongming Island, Shanghai Journal of Shanghai Normal University (Natural Sciences )516 -521 . [本文引用: 1] [ 陈晨 , 傅盈盈 , 黄璐 , 王云菁 , 何小丽 , 肖明 (2009 ). 上海崇明东滩加拿大一枝黄花根际部分可培养细菌的多样性分析上海师范大学学报(自然科学版) , 38, 516 -521 .] [本文引用: 1] [8] Chen T Liu WL Zhang CB Wang J 2012 ). Effects of Solidago canadensis invadation on dynamics of native plant communities and their mechanismsChinese Journal of Plant Ecology 253 -261 . [本文引用: 1] [ 陈彤 , 刘文莉 , 张崇邦 , 王江 (2012 ). 加拿大一枝黄花入侵对本土植物群落动态的影响及其机制植物生态学报 , 36, 253 -261 .] [本文引用: 1] [9] Coats VC Pelletreau KN Rumpho ME 2014 ). Amplicon pyrosequencing reveals the soil microbial diversity associated with invasive Japanese barberry ( Berberis thunbergii DC.)Molecular Ecology 1318 -1332 . [本文引用: 1] [10] de la Pe?a E de Clercq N Bonte D Roiloa S Rodríguez- Echeverría S Freitas H 2010 ). Plant-soil feedback as a mechanism of invasion by Carpobrotus edulis Biological Invasions, 3637 -3648 . [本文引用: 1] [11] Dong M Lu JZ Zhang WJ Chen JK Li B 2006 ). Canada goldenrod ( Solidago canadensis ): An invasive alien weed rapidly spreading in ChinaActa Phytotaxonomica Sinica 72 -85 . [本文引用: 1] [ 董梅 , 陆建忠 , 张文驹 , 陈家宽 , 李博 (2006 ). 加拿大一枝黄花——一种正在迅速扩张的外来入侵植物植物分类学报 , 44, 72 -85 .] [本文引用: 1] [12] Du YS 2014 ). Phenotypic Plasticity and Adaptation of Solidago canadensis. Beijing Forestry University, Beijing . 60 -62 . [本文引用: 1] [ 杜乐山 (2014 ). 加拿大一枝黄花的表型可塑性与适应性研究北京林业大学, 北京 . 60 -62 .] [本文引用: 1] [13] Duda JJ Freeman DC Emlen JM Belnap J Kitchen SG Zak JC Sobek E Tracy M Montante J 2003 ). Differences in native soil ecology associated with invasion of the exotic annual chenopod, Halogeton glomeratus Biology and Fertility of Soils, 72 -77 . [本文引用: 1] [14] Fang F Guo SL Huang LB 2004 ). Allelopathic effects of the invasive plant Solidago canadensis Ecologic Science 331 -334 . [本文引用: 1] [ 方芳 , 郭水良 , 黄林兵 (2004 ). 入侵杂草加拿大一枝黄花的化感作用生态科学 , 23, 331 -334 .] [本文引用: 1] [15] Ge HL Ji XE 2017 ). Screening, identification and promoting effects of potassium-solubilizing bacteria in rhizosphere of cucumberNorthern Horticulture, (13 ), 21 -25 . [本文引用: 1] [ 葛红莲 , 纪秀娥 (2017 ). 黄瓜根际解钾细菌的分离筛选、鉴定及其促生效果北方园艺 , (13 ), 21 -25 .] [本文引用: 1] [16] Graham RD Ulrich A 1972 ). Potassium deficiency-induced changes in stomatal behavior, leaf water potentials, and root system permeability in Beta vulgaris LPlant Physiology 105 -109 . [本文引用: 1] [17] Guo SL Fang F 2003 ). Physiological adaptation of the invasive plant Solidago canadensis to environmentsActa Phytoecologica Sinica 47 -52 . [本文引用: 2] [ 郭水良 , 方芳 (2003 ). 入侵植物加拿大一枝黄花对环境的生理适应性研究植物生态学报 , 27, 47 -52 .] [本文引用: 2] [18] Guo XB Wu HS Liu HA Chen JH Shi GH 2001 ). Effect of Bacillus mucilaginosus preparation on the growth and development of rice plantActa Agriculturae Universitatis Jiangxiensis 447 -449 . [本文引用: 1] [ 郭勋斌 , 吴洪生 , 刘怀阿 , 陈佳宏 , 石光辉 (2001 ). 钾细菌制剂对水稻生长发育的影响江西农业大学学报 , 23, 447 -449 .] [本文引用: 1] [19] Hao JH Qian HJ Jiang W Shen ZG 2009 ). Traits of sexual reproduction in Solidago canadensis LEcology and Environmental Sciences 2278 -2282 . [本文引用: 2] [ 郝建华 , 钱海军 , 姜雯 , 沈宗根 (2009 ). 加拿大一枝黄花有性生殖特征研究生态环境学报 , 18, 2278 -2282 .] [本文引用: 2] [20] Holmgren M Scheffer M Huston MA 1997 ). The interplay of facilitation and competition in plant communitiesEcology, 78, 1966 -1975 . [本文引用: 1] [21] Hu DJ Dong RR Ge DZ 1993 ). Theory and Practice of Plant Potassium Nutrition . Hunan Science & Technology Press, Changsha. 58 -109 . [本文引用: 1] [ 胡笃敬 , 董任瑞 , 葛旦之 (1993 ). 植物钾营养的理论与实践 . 湖南科学技术出版社, 长沙. 58 -109 .] [本文引用: 1] [22] Huang H Guo SL 2005 ). Study on reproductive biology of the invasive plant Solidago canadensis Acta Ecologica Sinica 2795 -2803 . [本文引用: 1] [ 黄华 , 郭水良 (2005 ). 外来入侵植物加拿大一枝黄花繁殖生物学研究生态学报 , 25, 2795 -2803 .] [本文引用: 1] [23] Jiang ZL Liu WX Wan FH Li ZY 2008 ). Differences in soil enzymatic activities and soil nutrients of Ageratina adenophora and native plants communities at the rhizosphere zonesJournal of Agro-Environment Science 660 -664 . [本文引用: 1] [ 蒋智林 , 刘万学 , 万方浩 , 李正跃 (2008 ). 紫茎泽兰与本地植物群落根际土壤酶活性和土壤肥力的差异农业环境科学学报 , 27, 660 -664 .] [本文引用: 1] [24] Ju RT Li H Shi ZR Li B 2012 ). Progress of biological invasions research in China over the last decadeBiodiversity Science, 20, 581 -611 . [本文引用: 1] [ 鞠瑞亭 , 李慧 , 石正人 , 李博 (2012 ). 近十年中国生物入侵研究进展生物多样性 , 20, 581 -611 .] [本文引用: 1] [25] Ju W 2016 ). Study on Isolation and Identification of High Efficient Potassium Bacteria from Poplar Rhizospheric Soil Nanjing Forestry University, Nanjing . 31 -33 . [本文引用: 1] [ 鞠伟 (2016 ). 杨树根际高效解钾细菌的分离筛选与鉴定南京林业大学, 南京 . 31 -33 .] [本文引用: 1] [26] Keane RM Crawley MJ 2002 ). Exotic plant invasions and the enemy release hypothesisTrends in Ecology & Evolution, 17, 164 -170 . [本文引用: 1] [27] Kourtev PS Ehrenfeld JG H?ggblom M 2002 ). Exotic plant species alter the microbial community structure and function in the soilEcology, 83, 3152 -3166 . [本文引用: 1] [28] Li CG Zhong Y Li XX Jiang X Xie CW 2017 ). Screening and identification of a new type of potassium bacteriaJournal of Guizhou University (Natural Sciences )132 -135 . [本文引用: 1] [ 李春钢 , 钟艳 , 李夏夏 , 姜雄 , 谢承卫 (2017 ). 一种新型解钾菌的筛选及鉴定贵州大学学报(自然科学版) , 34, 132 -135 . ] [本文引用: 1] [29] Li GQ 2009 ). Effects of Invasive Plant Solidago canadensis L. on Microbial Community Diversity in Rhizosphere Soil Fujian Agriculture and Forestry University, Fuzhou . 44 -46 . [本文引用: 1] [ 李国庆 (2009 ). 入侵植物加拿大一枝黄花对根际土壤微生物群落多样性的影响研究福建农林大学, 福州 . 44 -46 .] [本文引用: 1] [30] Li XX Gao XX Chen X Lu WH Dong CX Cui ZL Cao H 2014 ). Isolation and identification of an efficient strain of potassium-dissolving bacteria and opatimization of its incubation conditionActa Pedologica Sinica, 51, 381 -388 . [本文引用: 3] [ 李新新 , 高新新 , 陈星 , 卢维浩 , 董彩霞 , 崔中利 , 曹慧 (2014 ). 一株高效解钾菌的筛选、鉴定及发酵条件的优化土壤学报 , 51, 381 -388 .] [本文引用: 3] [31] Li YF 1994 ). Characteristic and function of silicate bacterial fertilizerSoil and Fertilizer Sciences in China, 2, 48 -49 . [本文引用: 1] [ 李元芳 (1994 ). 硅酸盐细菌肥料的特性和作用土壤肥料 , 2, 48 -49 .] [本文引用: 1] [32] Liang L Ye XQ Wu M Shao XX Li CM 2016 ). Invasion effects of Solidago canadensis on soil nutrients and active organic carbon components in reclamation district of Hangzhou Bay wetlandSoils 680 -685 . [本文引用: 2] [ 梁雷 , 叶小齐 , 吴明 , 邵学新 , 李长明 (2016 ). 加拿大一枝黄花入侵对杭州湾湿地围垦区土壤养分及活性有机碳组分的影响土壤 , 48, 680 -685 .] [本文引用: 2] [33] Liao D Huang HB Zhuang SX Hong YW 2018 ). Effects of exotic Spartina alterniflora on rhizosphere and endophytic bacterial community structures and diversity in roots of native mangrovesChinese Journal of Applied and Environmental Biology 269 -275 . [本文引用: 1] [ 廖丹 , 黄华斌 , 庄峙厦 , 洪有为 (2018 ). 互花米草入侵对红树秋茄根际与根内细菌群落结构与多样性的影响应用与环境生物学报 , 24, 269 -275 .] [本文引用: 1] [34] Liao M Xie XM Peng Y Chai JJ Chen N 2013 ). Characteristics of soil microbial community functional and structure diversity with coverage of Solidago canadensis LJournal of Central South University 749 -756 . [本文引用: 1] [35] Ling XG 2010 ). Principles and Methods of Soil Microbiology Research . Higher Education Press, Beijing. 323 -326 . [本文引用: 1] [ 林先贵 (2010 ). 土壤微生物研究原理与方法 . 高等教育出版社, 北京. 323 -326 .] [本文引用: 1] [36] Lorenzo P Rodríguez-Echeverría S González L Freitas H 2010 ). Effect of invasive Acacia dealbata Link. on soil microorganisms as determined by PCR-DGGEApplied Soil Ecology 245 -251 . [本文引用: 1] [37] Lu HM Ruan HG Tang GM Cai YC Guo ZX Wang J 2006 ). Evaluation of harmfulness and utility on Canada Goldenrod (Solidago canadensis )Journal of Shanghai Jiaotong University (Agricultural Science )402 -406 . [本文引用: 1] [ 陆慧明 , 阮海根 , 汤根妹 , 蔡云彩 , 郭志霞 , 王坚 (2006 ). 加拿大一枝黄花利害分析上海交通大学学报(农业科学版) , 24, 402 -406 .] [本文引用: 1] [38] Martin PA Newton AC Bullock JM 2017 ). Impacts of invasive plants on carbon pools depend on both species’ traits and local climateEcology 1026 -1035 . [本文引用: 1] [39] Mei LX Chen X Tang JJ 2005 ). Allelopathic effects of invasive weed Solidago canadensis on native plantsChinese Journal of Applied Ecology 2379 -2382 . [本文引用: 1] [ 梅玲笑 , 陈欣 , 唐建军 (2005 ). 外来杂草加拿大一枝黄花对入侵地植物的化感效应应用生态学报 , 16, 2379 -2382 .] [本文引用: 1] [40] Nijjer S Rogers WE Lee CTA Siemann E 2008 ). The effects of soil biota and fertilization on the success of Sapium sebiferum Applied Soil Ecology 1 -11 . [本文引用: 1] [41] Niu HB Liu WX Wan FH 2007 ). Invasive effects of Ageratina adenophora Sprengel (Asteraceae) on soil microbial community and physical and chemical properties Acta Ecologica Sinica, 3051 -3060 . [本文引用: 2] [ 牛红榜 , 刘万学 , 万方浩 (2007 ). 紫茎泽兰(Ageratina adenophora )入侵对土壤微生物群落和理化性质的影响生态学报 , 27, 3051 -3060 . [本文引用: 2] [42] Ren MX Zhang QG 2009 ). The relative generality of plant invasion mechanisms and predicting future invasive plantsWeed Research, 49, 449 -460 . [本文引用: 1] [43] Roberts KJ Anderson RC 2001 ). Effect of garlic mustard [ Alliaria petiolata(Beib. Cavara & Grande)] extracts on plants and arbuscular mycorrhizal (AM) fungiThe American Midland Naturalist 146 -152 . [本文引用: 1] [44] Shen LH Guo QX Lin WX Chen Y Huang Z 2007 ). Impacts of invasive alien weed Solidago canadensis L. on microbial population in the root soilChinese Agricultural Science Bulletin 323 -327 . [本文引用: 3] [ 沈荔花 , 郭琼霞 , 林文雄 , 陈颖 , 黄振 (2007 ). 加拿大一枝黄花对土壤微生物区系的影响研究中国农学通报 , 23, 323 -327 .] [本文引用: 3] [45] Shi Q 2018 ). The Effect of Brevibaterium frigoritolerans and Bacillus megaterium in the Invasion of Flaveria bidentis Hebei University, Baoding , Hebei. 20 -22 . [本文引用: 1] [ 石青 (2018 ). 耐寒短杆菌和巨大芽孢杆菌在黄顶菊入侵过程中的作用河北大学, 河北保定 . 20 -22 .] [本文引用: 1] [46] Sun ZK He WM 2010 ). Evidence for enhanced mutualism hypothesis: Solidago canadensis plants from regular soils perform betterPLOS ONE e15418 . DOI: 10.1371/journal.?pone.0015418 . [本文引用: 1] [47] Tai FJ Zhu XZ Han CX Zhang C Shao H 2016 ). Effects of aqueous extract of the invasive plant Xanthium italicum Moretti on soil microbial community, soil enzyme activity and soil nutrient Ecological Science, 71 -78 . [本文引用: 1] [ 邰凤姣 , 朱珣之 , 韩彩霞 , 张弛 , 邵华 (2016 ). 外来入侵植物意大利苍耳对土壤微生物群落、土壤酶活性和土壤养分的影响生态科学 , 35, 71 -78 .] [本文引用: 1] [48] The Editorial Board of Flora of China, Chinese Academy of Sciences (1985 ). Flora of China 76 . [本文引用: 1] [ 中国科学院中国植物志编辑委员会 (1985 ). 中国植物志 . 科学出版社, 北京. 74, 76 .] [本文引用: 1] [49] Uddin MN Robinson RW Caridi D Al Harun MAY 2014 ). Suppression of native Melaleuca ericifolia by the invasive Phragmites australis through allelopathic root exudatesAmerican Journal of Botany 479 -487 . [本文引用: 1] [50] Wang C Zhou J Liu J Jiang K Xiao H Du D 2018 ). Responses of the soil fungal communities to the co-invasion of two invasive species with different cover classesPlant Biology, 20, 151 -159 . [本文引用: 1] [51] Wang LF Wang DX Shangguan ZP 2013 ). Structural characters and nutrient contents of leaves as well as nitrogen distribution among different organs of big\headed wheatActa Ecologica Sinica, 33, 5219 -5227 . [本文引用: 1] [ 王丽芳 , 王德轩 , 上官周平 (2013 ). 大穗型小麦叶片性状、养分含量及氮素分配特征生态学报 , 33, 5219 -5227 .] [本文引用: 1] [52] Wang YL Gao RR Yu JY 2009 ). Invasive structural foundation of exotic plant Canada goldenrodActa Ecologica Sinica, 29, 108 -119 . [本文引用: 1] [ 王玉良 , 高瑞如 , 余玖银 (2009 ). 外来植物加拿大一枝黄花(Solidago canadensis )入侵的结构基础生态学报 , 29, 108 -119 . ] [本文引用: 1] [53] Wardle DA Bardgett RD Klironomos JN Set?l? H van der Putten WH Wall DH 2004 ). Ecological linkages between aboveground and belowground biotaScience, 304, 1629 -1633 . [本文引用: 2] [54] Wu F Liu XL Zhang N Zhang SS Guo H Zhang BF Qiu NQ 2010 ). Isolation and identification of mulberry rhizospheric silicate bacteria and determination of their potassium- releasing activitiesScience of Sericulture, 36, 323 -329 . [本文引用: 3] [ 吴凡 , 刘训理 , 张楠 , 张莎莎 , 国辉 , 张本峰 , 仇念全 (2010 ). 桑树根际硅酸盐细菌的分离鉴定及解钾能力测定蚕业科学 , 36, 323 -329 .] [本文引用: 3] [55] Yan XL Shou HY Ma JS 2012 ). The problem and status of the alien invasive plants in ChinaPlant Diversity and Resources, 34, 287 -313 . [本文引用: 1] [ 闫小玲 , 寿海洋 , 马金双 (2012 ). 中国外来入侵植物研究现状及存在的问题植物分类与资源学报 , 34, 287 -313 .] [本文引用: 1] [56] Yang GQ Guo J Gui FR 2014 ). Effects of allelochemicals from Eupatorium adenophorum on soil available phosphorus content and growth of Bacillus megaterium Jiangsu Agricultural Sciences 12 ), 137 -140 . [本文引用: 1] [ 杨国庆 , 郭娇 , 桂富荣 (2014 ). 紫茎泽兰的化感物质对土壤有效磷含量和巨大芽孢杆菌生长的影响江苏农业科学 , 42(12 ), 137 -140 .] [本文引用: 1] [57] Yang X Zhang LH Zheng C Zhang JL Han JM Dong JG 2012 ). Effects of Flaveria bidentis invasion on soil microbial communities, enzyme activities and nutrientsPlant Nutrition and Fertilizer Science 907 -914 . [本文引用: 1] [ 杨星 , 张利辉 , 郑超 , 张金林 , 韩建民 , 董金皋 (2012 ). 黄顶菊入侵对土壤微生物、土壤酶活性及土壤养分的影响植物营养与肥料学报 , 18, 907 -914 .] [本文引用: 1] [58] Yi LB Peng QZ He QZ Peng QJ 2012 ). Isolation and identification of potash feldspar-solubilizing bacteria and their potassium-releasing activitiesChinese Journal of Microecology, 24, 773 -776, 785 . [本文引用: 1] [ 易浪波 , 彭清忠 , 何齐庄 , 彭清静 (2012 ). 高效钾长石分解菌株的筛选、鉴定及解钾活性研究中国微生态学杂志 , 24, 773 -776, 785 .] [本文引用: 1] [59] Zhang HL Zhu M Li GJ 2015a ). Impact of reproductive traits on the invasive ability of Solidago canadensis LJournal of China University of Metrology 324 -330 . [本文引用: 1] [ 张海亮 , 朱敏 , 李干金 (2015a ). 加拿大一枝黄花繁殖性状对其入侵性的影响中国计量学院学报 , 26, 324 -330 .] [本文引用: 1] [60] Zhang HL Zhu M Li GJ 2015b ). Factors influencing the nonrandom abscission of Solidago canadensis seedsChinese Journal of Plant Ecology 258 -263 . [本文引用: 1] [ 张海亮 , 朱敏 , 李干金 (2015b ). 影响加拿大一枝黄花种子非随机脱落的因素植物生态学报 , 39, 258 -263 .] [本文引用: 1] [61] Zhang MY Chen YF Zhou DB Qi DF Gao ZF Zhang XY 2016 ). Isolation, identification and fermentation conditions optimization of potassium bacteria in Castor rhizosphere soilChinese Journal of Tropical Crops 2268 -2275 . [本文引用: 1] [ 张妙宜 , 陈宇丰 , 周登博 , 起登凤 , 高祝芬 , 张锡炎 (2016 ). 蓖麻根际土壤解钾菌的筛选鉴定及发酵条件的优化热带作物学报 , 37, 2268 -2275 .] [本文引用: 1] [62] Zhang Q Yao LJ Yang RY Yang XY Tang JJ Chen X 2007 ). Potential allelopathic effects of an invasive species Solidago canadensis on the mycorrhizae of native plant speciesAllelopathy Journal, 71 -77 . [本文引用: 1] [63] Zhang SS Jin YL Tang JJ Chen X 2009 ). The invasive plant Solidago canadensis L. suppresses local soil pathogens through allelopathy Applied Soil Ecology, 215 -222 . [本文引用: 1] [64] Zhang TR Huangfu CH Bai XM Yang DL Li G Lai Y Zhao JN 2010 ). Effects of Flaveria bidentis invasion on soil nutrient contents and enzyme activities Chinese Journal of Ecology, 1353 -1358 . [本文引用: 1] [ 张天瑞 , 皇甫超河 , 白小明 , 杨殿林 , 李刚 , 赖欣 , 赵建宁 (2010 ). 黄顶菊入侵对土壤养分和酶活性的影响生态学杂志 , 29, 1353 -1358 .] [本文引用: 1] [65] Zhang WJ Rui WY Tu C Diab HG Louws FJ Mueller JP Creamer N Bell M Wagger MG Hu S 2005 ). Responses of soil microbial community structure and diversity to agricultural deintensificationPedosphere, 15, 440 -447 . [本文引用: 1] [66] Zhao JL Cheng CQ Gu XY Liu B 2014 ). Effects of root exudates from invasive plant (Mirabilis jalapa ) on soil microenviroment under different land-use typesAdvanced Materials Research 1419 -1424 . [本文引用: 2] [67] Zhou ZR 2010 ). On Influence of Solidago canadensis Invasion on Rhizosphere Micro-Environment of Soil. Nanjing Agricultural University, Nanjing . 33 -36 . [本文引用: 1] [ 周振荣 (2010 ). 外来入侵植物加拿大一枝黄花对根际土壤微环境的影响研究南京农业大学, 南京 . 33 -36 .] [本文引用: 1] The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns
1
2000
... 入侵植物对营养元素的吸收、积累和释放对其生长、入侵能力及其入侵的生态学效应都具有重要作用.钾是植物生长发育所需要的主要营养元素之一, 在植物生长繁育过程中发挥必不可少的作用, 钾的增加不仅能够提高植物光合速率和酶的活化速度、增加植物糖和蛋白质含量、增强植物对氮的吸收, 还能够提高作物的产量、提高植物的抗逆性(
Graham & Ulrich, 1972 ;
胡笃敬等, 1993 ).本研究发现, 野外自然生长的加拿大一枝黄花叶片和茎中的钾含量显著高于伴生植物白茅, 尤其叶片中的钾含量是白茅的7.33倍, 说明在相同生境下, 加拿大一枝黄花吸收钾的能力远高于本地植物, 且加拿大一枝黄花能够将更多的钾分配到叶片中, 而叶片是光合作用最主要的场所, 叶片中养分含量增多可能增强植物光合速率和生长代谢速率(
Aerts, 2000 ;
王丽芳等, 2013 ).此外, 加拿大一枝黄花叶片中的氮磷含量明显高于茎, 更加证实了加拿大一枝黄花将更多养分分配到叶片中的生长策略.人为提高钾供给水平均能促进两种植物生物量积累增加(
图3 ), 但是我们发现加拿大一枝黄花入侵地土壤0-10 cm及10-20 cm土层中全氮、全磷、全钾及碱解氮、有效磷和速效钾含量均多于对照植物白茅, 尤其土壤中速效养分的含量显著多于白茅, 这与
梁雷等(2016) 的研究结果相一致.
陈彤等(2012) 发现加拿大一枝黄花能显著提高土壤中硝态氮含量, 并且其较高的氮积累能力可能和入侵后本地植物种群的动态变化格局紧密相关 这些研究说明加拿大一枝黄花存在一种能够增强土壤养分积累的机制, 而这种机制反过来可能促进其进一步入侵.外来植物对入侵地土壤养分含量的影响被大量研究.一些研究认为外来植物入侵往往能够通过影响入侵地土壤养分而促进自身成功入侵(
Holmgren et al ., 1997 ).许多外来植物入侵后通过改变土壤肥力影响其与本地种的竞争能力, 例如破坏草、黄顶菊(
Flaveria bidentis )、盐生草(
Halogeton glomeratus )等植物入侵时均会导致土壤养分含量向有利于入侵植物的方向发生改变(
Duda et al ., 2003 ;
蒋智林等, 2008 ;
张天瑞等, 2010 ). ...
1
2000
... 植物样品用蒸馏水清洗干净, 于105 ℃杀青2 h, 60 ℃烘干至恒质量, 冷却后剪碎, 用HNO
3 -H
2 O
2 消化后, 用火焰光度计法测植物氮磷钾含量.不同土层的土壤样品风干、敲碎、挑除杂质, 研磨至过100目网筛, 用半微量开氏法检测土壤全氮含量、碱解扩散法测定土壤碱解氮含量、0.03 mol·L
-1 NH
4 F-0.025 mol·L
-1 HCl法测定土壤有效磷含量、HClO
4 -H
2 SO
4 法检测土壤全磷含量、NaOH熔融-火焰光度计法测土壤全钾含量、NH
4 OAc浸提-火焰光度计法测定土壤速效钾含量(
鲍士旦, 2000 ).样品检测委托国家林业局经济林产品质量检验检测中心 进行. ...
1
2000
... 植物样品用蒸馏水清洗干净, 于105 ℃杀青2 h, 60 ℃烘干至恒质量, 冷却后剪碎, 用HNO
3 -H
2 O
2 消化后, 用火焰光度计法测植物氮磷钾含量.不同土层的土壤样品风干、敲碎、挑除杂质, 研磨至过100目网筛, 用半微量开氏法检测土壤全氮含量、碱解扩散法测定土壤碱解氮含量、0.03 mol·L
-1 NH
4 F-0.025 mol·L
-1 HCl法测定土壤有效磷含量、HClO
4 -H
2 SO
4 法检测土壤全磷含量、NaOH熔融-火焰光度计法测土壤全钾含量、NH
4 OAc浸提-火焰光度计法测定土壤速效钾含量(
鲍士旦, 2000 ).样品检测委托国家林业局经济林产品质量检验检测中心 进行. ...
Soil biota in an ungrazed grassland: Response to annual grass (
Bromus tectorum ) invasion
1
2001
... 外来植物在入侵过程中可能通过释放根系分泌物和改变微生物碳源等方式影响土壤微生物结构和功能, 而土壤微生物群落的改变反过来会影响外来植物的进一步入侵(
杨星等, 2012 ;
Uddin et al ., 2014 ;
Zhao et al ., 2014 ;
Martin et al ., 2017 ).以往研究表明, 紫茉莉(
Mirabilis jalapa )、破坏草(紫茎泽兰)(
Ageratina adenophora )、互花米草(
Spartina alterniflora )可以改变入侵地土壤微生物多样性(
牛红榜, 2007 ;
Zhao et al ., 2014 ;
廖丹等, 2018 ); 银荆(
Acacia dealbata )入侵西班牙北部地区后使当地土壤中细菌丰富度显著增加, 真菌的丰富度和多样性显著减少(
Lorenzo et al ., 2010 ); 旱雀麦(
Bromus tectorum )使入侵地的细菌数量显著增多, 真菌数量显著减少(
Belnap & Phillips, 2001 ); 葱芥(
Alliaria petiolata )入侵后导致土壤中丛枝菌根真菌数量显著减少(
Roberts & Anderson, 2001 ).这些结果说明, 外来植物入侵能够导致土壤微生物多样性发生明显改变.而另一方面, 土壤微生物的改变可能促进外来植物的入侵, 甚至可能抑制本地植物的生长.例如, 土壤微生物显著提高了入侵植物加拿大一枝黄花(
Solidago canadensis )的出芽率和资源竞争力, 而降低了本土植物长芒草(
Stipa bungeana )的竞争力(
Sun & He, 2010 ).因此, 探究入侵植物对土壤微生物的影响可能是了解植物入侵机理的关键所在. ...
Evolution of increased competitive ability in invasive nonindigenous plants: A hypothesis
1
1995
... 外来植物入侵不仅导致入侵地农林生产经济损失, 而且对生态系统和生物多样性造成了严重的威胁(
鞠瑞亭等, 2012 ).据调查, 我国外来入侵植物多达670余种, 分布范围主要集中在东南沿海地区, 且入侵速度正在逐年加快、数量逐年增多, 若不加以防治, 则直接影响我国经济发展和物种栖息地保护(
闫小玲等, 2012 ).了解外来植物的入侵机制是制定科学合理的防控方案的关键.为了解释外来植物的入侵机制, 研究者提出了很多研究假说, 例如天敌逃逸假说(
Keane et al ., 2002 )、竞争能力增强进化假说(
Blossey & N?tzold, 1995 ), 但这些假说主要针对入侵植物本身的进化, 而没有考虑入侵植物与土壤微生物的相互作用.随着研究的不断深入, 越来越多的****意识到植物入侵过程与地下生态存在紧密联系(
de la Pe?a et al ., 2010 ;
Ren & Zhang, 2009 ), 入侵植物与土壤微生物之间的互作在入侵过程中发挥重要作用(
Wardle et al ., 2004 ). ...
Lineage overwhelms environmental conditions in determining rhizosphere bacterial community structure in a cosmopolitan invasive plant
1
2017
... 植物与土壤微生物的相互作用在植物入侵过程中起着至关重要的作用(
Bowen et al ., 2017 ).研究表明外来植物通过改变入侵地土壤微生物结构和功能, 进而改善土壤养分供应, 促进植物生长, 增强植物抗逆性, 促使植物成功入侵(
Kourtev et al ., 2002 ;
Wardle et al ., 2004 ;
Nijjer et al ., 2008 ;
Coats et al ., 2014 ).例如, 紫茎泽兰入侵后改变了入侵地土壤微生物的群落结构, 同时提高了入侵地土壤肥力, 进而增强了紫茎泽兰的竞争能力(
牛红榜等, 2007 ).本研究发现加拿大一枝黄花根际土壤中解钾菌数量、群落结构和解钾活性都和伴生植物白茅有着较大的差异, 加拿大一枝黄花的根际解钾菌数量远多于对照, 且其中多数根际解钾菌均具有高效的解钾能力, 同时加拿大一枝黄花茎叶中的钾含量及入侵地土壤中全钾和速效钾的含量均高于对照.这说明, 加拿大一枝黄花的富钾作用可能与其较多的高效解钾菌有关.相关研究也表明加拿大一枝黄花入侵会显著改变土壤微生物群落结构和多样性(
沈荔花等, 2007 ;
陈晨等, 2009 ;
李国庆, 2009 ), 这些转变可能会显著地影响土壤中钾的有效性和供给水平.在其他入侵植物的相关研究中也发现, 入侵植物对土壤养分含量的影响也和其土壤微生物群落的改变紧密相关.例如, 黄顶菊根际耐寒短杆菌能增加土壤磷含量, 对病原菌的抑制作用较对照植物更强(
石青, 2018 ); 破坏草通过促进土壤中解磷菌的生长使其土壤中有效磷含量增加(
杨国庆等, 2014 );
Xanthium italicum 水提液不仅使土壤中细菌和真菌数量明显增多, 同时其土壤速效钾、速效氮等营养成分也显著增多(
邰凤姣等, 2016 ). ...
1
2013
... 分别取加拿大一枝黄花和白茅的种子约100粒, 播种于铺有滤纸的培养皿中, 加蒸馏水至种子完全浸没, 将培养皿带盖置于培养箱中培养4-5天, 设置光照、黑暗环境各12 h, 温度分别设置28 ℃和22 ℃.待植物子叶展开后, 挑选大小均等的幼苗移栽入装有石英砂的花盆中, 每盆移栽5株, 置于温室进行培养, 前期浇灌蒸馏水, 3天后开始浇灌营养液, 营养液以无钾霍格兰营养液为基础液(
蔡庆生, 2013 ), 钾添加量设置3个水平: 0 mol·L
-1 (缺钾)、0.1 × 10
-3 mol·L
-1 (低钾)、6 × 10
-3 mol·L
-1 (高钾), 每个处理重复15次.10天后进行间苗, 每盆保留一株.营养液处理90天后结束培养. ...
1
2013
... 分别取加拿大一枝黄花和白茅的种子约100粒, 播种于铺有滤纸的培养皿中, 加蒸馏水至种子完全浸没, 将培养皿带盖置于培养箱中培养4-5天, 设置光照、黑暗环境各12 h, 温度分别设置28 ℃和22 ℃.待植物子叶展开后, 挑选大小均等的幼苗移栽入装有石英砂的花盆中, 每盆移栽5株, 置于温室进行培养, 前期浇灌蒸馏水, 3天后开始浇灌营养液, 营养液以无钾霍格兰营养液为基础液(
蔡庆生, 2013 ), 钾添加量设置3个水平: 0 mol·L
-1 (缺钾)、0.1 × 10
-3 mol·L
-1 (低钾)、6 × 10
-3 mol·L
-1 (高钾), 每个处理重复15次.10天后进行间苗, 每盆保留一株.营养液处理90天后结束培养. ...
上海崇明东滩加拿大一枝黄花根际部分可培养细菌的多样性分析
1
2009
... 植物与土壤微生物的相互作用在植物入侵过程中起着至关重要的作用(
Bowen et al ., 2017 ).研究表明外来植物通过改变入侵地土壤微生物结构和功能, 进而改善土壤养分供应, 促进植物生长, 增强植物抗逆性, 促使植物成功入侵(
Kourtev et al ., 2002 ;
Wardle et al ., 2004 ;
Nijjer et al ., 2008 ;
Coats et al ., 2014 ).例如, 紫茎泽兰入侵后改变了入侵地土壤微生物的群落结构, 同时提高了入侵地土壤肥力, 进而增强了紫茎泽兰的竞争能力(
牛红榜等, 2007 ).本研究发现加拿大一枝黄花根际土壤中解钾菌数量、群落结构和解钾活性都和伴生植物白茅有着较大的差异, 加拿大一枝黄花的根际解钾菌数量远多于对照, 且其中多数根际解钾菌均具有高效的解钾能力, 同时加拿大一枝黄花茎叶中的钾含量及入侵地土壤中全钾和速效钾的含量均高于对照.这说明, 加拿大一枝黄花的富钾作用可能与其较多的高效解钾菌有关.相关研究也表明加拿大一枝黄花入侵会显著改变土壤微生物群落结构和多样性(
沈荔花等, 2007 ;
陈晨等, 2009 ;
李国庆, 2009 ), 这些转变可能会显著地影响土壤中钾的有效性和供给水平.在其他入侵植物的相关研究中也发现, 入侵植物对土壤养分含量的影响也和其土壤微生物群落的改变紧密相关.例如, 黄顶菊根际耐寒短杆菌能增加土壤磷含量, 对病原菌的抑制作用较对照植物更强(
石青, 2018 ); 破坏草通过促进土壤中解磷菌的生长使其土壤中有效磷含量增加(
杨国庆等, 2014 );
Xanthium italicum 水提液不仅使土壤中细菌和真菌数量明显增多, 同时其土壤速效钾、速效氮等营养成分也显著增多(
邰凤姣等, 2016 ). ...
上海崇明东滩加拿大一枝黄花根际部分可培养细菌的多样性分析
1
2009
... 植物与土壤微生物的相互作用在植物入侵过程中起着至关重要的作用(
Bowen et al ., 2017 ).研究表明外来植物通过改变入侵地土壤微生物结构和功能, 进而改善土壤养分供应, 促进植物生长, 增强植物抗逆性, 促使植物成功入侵(
Kourtev et al ., 2002 ;
Wardle et al ., 2004 ;
Nijjer et al ., 2008 ;
Coats et al ., 2014 ).例如, 紫茎泽兰入侵后改变了入侵地土壤微生物的群落结构, 同时提高了入侵地土壤肥力, 进而增强了紫茎泽兰的竞争能力(
牛红榜等, 2007 ).本研究发现加拿大一枝黄花根际土壤中解钾菌数量、群落结构和解钾活性都和伴生植物白茅有着较大的差异, 加拿大一枝黄花的根际解钾菌数量远多于对照, 且其中多数根际解钾菌均具有高效的解钾能力, 同时加拿大一枝黄花茎叶中的钾含量及入侵地土壤中全钾和速效钾的含量均高于对照.这说明, 加拿大一枝黄花的富钾作用可能与其较多的高效解钾菌有关.相关研究也表明加拿大一枝黄花入侵会显著改变土壤微生物群落结构和多样性(
沈荔花等, 2007 ;
陈晨等, 2009 ;
李国庆, 2009 ), 这些转变可能会显著地影响土壤中钾的有效性和供给水平.在其他入侵植物的相关研究中也发现, 入侵植物对土壤养分含量的影响也和其土壤微生物群落的改变紧密相关.例如, 黄顶菊根际耐寒短杆菌能增加土壤磷含量, 对病原菌的抑制作用较对照植物更强(
石青, 2018 ); 破坏草通过促进土壤中解磷菌的生长使其土壤中有效磷含量增加(
杨国庆等, 2014 );
Xanthium italicum 水提液不仅使土壤中细菌和真菌数量明显增多, 同时其土壤速效钾、速效氮等营养成分也显著增多(
邰凤姣等, 2016 ). ...
加拿大一枝黄花入侵对本土植物群落动态的影响及其机制
1
2012
... 入侵植物对营养元素的吸收、积累和释放对其生长、入侵能力及其入侵的生态学效应都具有重要作用.钾是植物生长发育所需要的主要营养元素之一, 在植物生长繁育过程中发挥必不可少的作用, 钾的增加不仅能够提高植物光合速率和酶的活化速度、增加植物糖和蛋白质含量、增强植物对氮的吸收, 还能够提高作物的产量、提高植物的抗逆性(
Graham & Ulrich, 1972 ;
胡笃敬等, 1993 ).本研究发现, 野外自然生长的加拿大一枝黄花叶片和茎中的钾含量显著高于伴生植物白茅, 尤其叶片中的钾含量是白茅的7.33倍, 说明在相同生境下, 加拿大一枝黄花吸收钾的能力远高于本地植物, 且加拿大一枝黄花能够将更多的钾分配到叶片中, 而叶片是光合作用最主要的场所, 叶片中养分含量增多可能增强植物光合速率和生长代谢速率(
Aerts, 2000 ;
王丽芳等, 2013 ).此外, 加拿大一枝黄花叶片中的氮磷含量明显高于茎, 更加证实了加拿大一枝黄花将更多养分分配到叶片中的生长策略.人为提高钾供给水平均能促进两种植物生物量积累增加(
图3 ), 但是我们发现加拿大一枝黄花入侵地土壤0-10 cm及10-20 cm土层中全氮、全磷、全钾及碱解氮、有效磷和速效钾含量均多于对照植物白茅, 尤其土壤中速效养分的含量显著多于白茅, 这与
梁雷等(2016) 的研究结果相一致.
陈彤等(2012) 发现加拿大一枝黄花能显著提高土壤中硝态氮含量, 并且其较高的氮积累能力可能和入侵后本地植物种群的动态变化格局紧密相关 这些研究说明加拿大一枝黄花存在一种能够增强土壤养分积累的机制, 而这种机制反过来可能促进其进一步入侵.外来植物对入侵地土壤养分含量的影响被大量研究.一些研究认为外来植物入侵往往能够通过影响入侵地土壤养分而促进自身成功入侵(
Holmgren et al ., 1997 ).许多外来植物入侵后通过改变土壤肥力影响其与本地种的竞争能力, 例如破坏草、黄顶菊(
Flaveria bidentis )、盐生草(
Halogeton glomeratus )等植物入侵时均会导致土壤养分含量向有利于入侵植物的方向发生改变(
Duda et al ., 2003 ;
蒋智林等, 2008 ;
张天瑞等, 2010 ). ...
加拿大一枝黄花入侵对本土植物群落动态的影响及其机制
1
2012
... 入侵植物对营养元素的吸收、积累和释放对其生长、入侵能力及其入侵的生态学效应都具有重要作用.钾是植物生长发育所需要的主要营养元素之一, 在植物生长繁育过程中发挥必不可少的作用, 钾的增加不仅能够提高植物光合速率和酶的活化速度、增加植物糖和蛋白质含量、增强植物对氮的吸收, 还能够提高作物的产量、提高植物的抗逆性(
Graham & Ulrich, 1972 ;
胡笃敬等, 1993 ).本研究发现, 野外自然生长的加拿大一枝黄花叶片和茎中的钾含量显著高于伴生植物白茅, 尤其叶片中的钾含量是白茅的7.33倍, 说明在相同生境下, 加拿大一枝黄花吸收钾的能力远高于本地植物, 且加拿大一枝黄花能够将更多的钾分配到叶片中, 而叶片是光合作用最主要的场所, 叶片中养分含量增多可能增强植物光合速率和生长代谢速率(
Aerts, 2000 ;
王丽芳等, 2013 ).此外, 加拿大一枝黄花叶片中的氮磷含量明显高于茎, 更加证实了加拿大一枝黄花将更多养分分配到叶片中的生长策略.人为提高钾供给水平均能促进两种植物生物量积累增加(
图3 ), 但是我们发现加拿大一枝黄花入侵地土壤0-10 cm及10-20 cm土层中全氮、全磷、全钾及碱解氮、有效磷和速效钾含量均多于对照植物白茅, 尤其土壤中速效养分的含量显著多于白茅, 这与
梁雷等(2016) 的研究结果相一致.
陈彤等(2012) 发现加拿大一枝黄花能显著提高土壤中硝态氮含量, 并且其较高的氮积累能力可能和入侵后本地植物种群的动态变化格局紧密相关 这些研究说明加拿大一枝黄花存在一种能够增强土壤养分积累的机制, 而这种机制反过来可能促进其进一步入侵.外来植物对入侵地土壤养分含量的影响被大量研究.一些研究认为外来植物入侵往往能够通过影响入侵地土壤养分而促进自身成功入侵(
Holmgren et al ., 1997 ).许多外来植物入侵后通过改变土壤肥力影响其与本地种的竞争能力, 例如破坏草、黄顶菊(
Flaveria bidentis )、盐生草(
Halogeton glomeratus )等植物入侵时均会导致土壤养分含量向有利于入侵植物的方向发生改变(
Duda et al ., 2003 ;
蒋智林等, 2008 ;
张天瑞等, 2010 ). ...
Amplicon pyrosequencing reveals the soil microbial diversity associated with invasive Japanese barberry (
Berberis thunbergii DC.)
1
2014
... 植物与土壤微生物的相互作用在植物入侵过程中起着至关重要的作用(
Bowen et al ., 2017 ).研究表明外来植物通过改变入侵地土壤微生物结构和功能, 进而改善土壤养分供应, 促进植物生长, 增强植物抗逆性, 促使植物成功入侵(
Kourtev et al ., 2002 ;
Wardle et al ., 2004 ;
Nijjer et al ., 2008 ;
Coats et al ., 2014 ).例如, 紫茎泽兰入侵后改变了入侵地土壤微生物的群落结构, 同时提高了入侵地土壤肥力, 进而增强了紫茎泽兰的竞争能力(
牛红榜等, 2007 ).本研究发现加拿大一枝黄花根际土壤中解钾菌数量、群落结构和解钾活性都和伴生植物白茅有着较大的差异, 加拿大一枝黄花的根际解钾菌数量远多于对照, 且其中多数根际解钾菌均具有高效的解钾能力, 同时加拿大一枝黄花茎叶中的钾含量及入侵地土壤中全钾和速效钾的含量均高于对照.这说明, 加拿大一枝黄花的富钾作用可能与其较多的高效解钾菌有关.相关研究也表明加拿大一枝黄花入侵会显著改变土壤微生物群落结构和多样性(
沈荔花等, 2007 ;
陈晨等, 2009 ;
李国庆, 2009 ), 这些转变可能会显著地影响土壤中钾的有效性和供给水平.在其他入侵植物的相关研究中也发现, 入侵植物对土壤养分含量的影响也和其土壤微生物群落的改变紧密相关.例如, 黄顶菊根际耐寒短杆菌能增加土壤磷含量, 对病原菌的抑制作用较对照植物更强(
石青, 2018 ); 破坏草通过促进土壤中解磷菌的生长使其土壤中有效磷含量增加(
杨国庆等, 2014 );
Xanthium italicum 水提液不仅使土壤中细菌和真菌数量明显增多, 同时其土壤速效钾、速效氮等营养成分也显著增多(
邰凤姣等, 2016 ). ...
Plant-soil feedback as a mechanism of invasion by
Carpobrotus edulis 1
2010
... 外来植物入侵不仅导致入侵地农林生产经济损失, 而且对生态系统和生物多样性造成了严重的威胁(
鞠瑞亭等, 2012 ).据调查, 我国外来入侵植物多达670余种, 分布范围主要集中在东南沿海地区, 且入侵速度正在逐年加快、数量逐年增多, 若不加以防治, 则直接影响我国经济发展和物种栖息地保护(
闫小玲等, 2012 ).了解外来植物的入侵机制是制定科学合理的防控方案的关键.为了解释外来植物的入侵机制, 研究者提出了很多研究假说, 例如天敌逃逸假说(
Keane et al ., 2002 )、竞争能力增强进化假说(
Blossey & N?tzold, 1995 ), 但这些假说主要针对入侵植物本身的进化, 而没有考虑入侵植物与土壤微生物的相互作用.随着研究的不断深入, 越来越多的****意识到植物入侵过程与地下生态存在紧密联系(
de la Pe?a et al ., 2010 ;
Ren & Zhang, 2009 ), 入侵植物与土壤微生物之间的互作在入侵过程中发挥重要作用(
Wardle et al ., 2004 ). ...
加拿大一枝黄花——一种正在迅速扩张的外来入侵植物
1
2006
... 加拿大一枝黄花是菊科一枝黄花属植物, 多年生草本, 具有发达根状茎, 原产于北美地区, 最初作为庭园花卉引入我国上海、南京等地, 现已广泛逸生为恶性外来种(
中国科学院中国植物志编辑委员会, 1985 ;
郭水良和方芳, 2003 ).加拿大一枝黄花繁殖能力极强, 既能利用种子进行有性繁殖, 也可通过地下根状茎进行无性繁殖, 一般情况下, 每株植株可产约7万粒种子, 且种子具有较高萌发率, 此外, 植株根部具有上百个根茎结构, 每个根茎在第2年可萌发形成独立的个体, 如此高的繁殖能力使其在入侵后能够迅速扩散形成单优势种群落(
王玉良等, 2009 ;
张海亮等, 2015a ).此外, 加拿大一枝黄花根部还会产生一些化感物质影响周围土著植物生长, 对入侵地生物多样性造成了一定的影响(
方芳等, 2004 ).而目前加拿大一枝黄花在我国入侵范围逐渐扩大, 入侵速度逐渐加快, 对我国生态环境、生物多样性、农业和畜牧业均造成了不同程度的影响(
董梅等, 2006 ;
陆慧明等, 2006 ), 因此亟须加强加拿大一枝黄花入侵机理与防控技术方面的研究. ...
加拿大一枝黄花——一种正在迅速扩张的外来入侵植物
1
2006
... 加拿大一枝黄花是菊科一枝黄花属植物, 多年生草本, 具有发达根状茎, 原产于北美地区, 最初作为庭园花卉引入我国上海、南京等地, 现已广泛逸生为恶性外来种(
中国科学院中国植物志编辑委员会, 1985 ;
郭水良和方芳, 2003 ).加拿大一枝黄花繁殖能力极强, 既能利用种子进行有性繁殖, 也可通过地下根状茎进行无性繁殖, 一般情况下, 每株植株可产约7万粒种子, 且种子具有较高萌发率, 此外, 植株根部具有上百个根茎结构, 每个根茎在第2年可萌发形成独立的个体, 如此高的繁殖能力使其在入侵后能够迅速扩散形成单优势种群落(
王玉良等, 2009 ;
张海亮等, 2015a ).此外, 加拿大一枝黄花根部还会产生一些化感物质影响周围土著植物生长, 对入侵地生物多样性造成了一定的影响(
方芳等, 2004 ).而目前加拿大一枝黄花在我国入侵范围逐渐扩大, 入侵速度逐渐加快, 对我国生态环境、生物多样性、农业和畜牧业均造成了不同程度的影响(
董梅等, 2006 ;
陆慧明等, 2006 ), 因此亟须加强加拿大一枝黄花入侵机理与防控技术方面的研究. ...
加拿大一枝黄花的表型可塑性与适应性研究
1
2014
... 目前国内外对加拿大一枝黄花入侵机制的研究主要集中在繁殖特性(
黄华和郭水良, 2005 ;
郝建华等, 2009 ;
张海亮等, 2015b )、化感作用(
梅玲笑, 2005 ;
郝建华等, 2009 )、生理生态特征与环境适应性(
郭水良和方芳, 2003 ;
杜乐山, 2014 )以及土壤微生物响应(
Zhang et al ., 2005 ;
沈荔花等, 2007 )等方面.研究表明, 在不同程度的入侵地中, 加拿大一枝黄花体内钾含量均高于伴生植物, 尤其在生长期和开花期差异最为显著, 且植物吸钾量与生物量呈正相关关系(
周振荣, 2010 ).在微生物响应方面, 研究发现加拿大一枝黄花入侵后能改变土壤微生物的群落数量、结构和功能多样性, 可能有助于加拿大一枝黄花进一步入侵(
Liao et al ., 2013 ;
Wang et al ., 2018 ).根际解钾菌能将土壤中难溶性钾转化为可被植物直接吸收的有效性钾, 进而促进植物钾吸收, 增强植物抗逆性(
李元芳, 1994 ;
郭勋斌等, 2001 ).加拿大一枝黄花入侵是否改变了根际解钾菌数量、结构和解钾活性等问题还有待深入研究.因此, 本研究以入侵植物加拿大一枝黄花和伴生本地植物白茅(
Imperata cylindrica )为研究对象, 比较加拿大一枝黄花与白茅的根际解钾菌数量、多样性与解钾能力差异, 旨在初步探讨根际解钾菌在加拿大一枝黄花入侵过程中发挥的作用, 为深入研究加拿大一枝黄花入侵机制提供理论依据. ...
加拿大一枝黄花的表型可塑性与适应性研究
1
2014
... 目前国内外对加拿大一枝黄花入侵机制的研究主要集中在繁殖特性(
黄华和郭水良, 2005 ;
郝建华等, 2009 ;
张海亮等, 2015b )、化感作用(
梅玲笑, 2005 ;
郝建华等, 2009 )、生理生态特征与环境适应性(
郭水良和方芳, 2003 ;
杜乐山, 2014 )以及土壤微生物响应(
Zhang et al ., 2005 ;
沈荔花等, 2007 )等方面.研究表明, 在不同程度的入侵地中, 加拿大一枝黄花体内钾含量均高于伴生植物, 尤其在生长期和开花期差异最为显著, 且植物吸钾量与生物量呈正相关关系(
周振荣, 2010 ).在微生物响应方面, 研究发现加拿大一枝黄花入侵后能改变土壤微生物的群落数量、结构和功能多样性, 可能有助于加拿大一枝黄花进一步入侵(
Liao et al ., 2013 ;
Wang et al ., 2018 ).根际解钾菌能将土壤中难溶性钾转化为可被植物直接吸收的有效性钾, 进而促进植物钾吸收, 增强植物抗逆性(
李元芳, 1994 ;
郭勋斌等, 2001 ).加拿大一枝黄花入侵是否改变了根际解钾菌数量、结构和解钾活性等问题还有待深入研究.因此, 本研究以入侵植物加拿大一枝黄花和伴生本地植物白茅(
Imperata cylindrica )为研究对象, 比较加拿大一枝黄花与白茅的根际解钾菌数量、多样性与解钾能力差异, 旨在初步探讨根际解钾菌在加拿大一枝黄花入侵过程中发挥的作用, 为深入研究加拿大一枝黄花入侵机制提供理论依据. ...
Differences in native soil ecology associated with invasion of the exotic annual chenopod,
Halogeton glomeratus 1
2003
... 入侵植物对营养元素的吸收、积累和释放对其生长、入侵能力及其入侵的生态学效应都具有重要作用.钾是植物生长发育所需要的主要营养元素之一, 在植物生长繁育过程中发挥必不可少的作用, 钾的增加不仅能够提高植物光合速率和酶的活化速度、增加植物糖和蛋白质含量、增强植物对氮的吸收, 还能够提高作物的产量、提高植物的抗逆性(
Graham & Ulrich, 1972 ;
胡笃敬等, 1993 ).本研究发现, 野外自然生长的加拿大一枝黄花叶片和茎中的钾含量显著高于伴生植物白茅, 尤其叶片中的钾含量是白茅的7.33倍, 说明在相同生境下, 加拿大一枝黄花吸收钾的能力远高于本地植物, 且加拿大一枝黄花能够将更多的钾分配到叶片中, 而叶片是光合作用最主要的场所, 叶片中养分含量增多可能增强植物光合速率和生长代谢速率(
Aerts, 2000 ;
王丽芳等, 2013 ).此外, 加拿大一枝黄花叶片中的氮磷含量明显高于茎, 更加证实了加拿大一枝黄花将更多养分分配到叶片中的生长策略.人为提高钾供给水平均能促进两种植物生物量积累增加(
图3 ), 但是我们发现加拿大一枝黄花入侵地土壤0-10 cm及10-20 cm土层中全氮、全磷、全钾及碱解氮、有效磷和速效钾含量均多于对照植物白茅, 尤其土壤中速效养分的含量显著多于白茅, 这与
梁雷等(2016) 的研究结果相一致.
陈彤等(2012) 发现加拿大一枝黄花能显著提高土壤中硝态氮含量, 并且其较高的氮积累能力可能和入侵后本地植物种群的动态变化格局紧密相关 这些研究说明加拿大一枝黄花存在一种能够增强土壤养分积累的机制, 而这种机制反过来可能促进其进一步入侵.外来植物对入侵地土壤养分含量的影响被大量研究.一些研究认为外来植物入侵往往能够通过影响入侵地土壤养分而促进自身成功入侵(
Holmgren et al ., 1997 ).许多外来植物入侵后通过改变土壤肥力影响其与本地种的竞争能力, 例如破坏草、黄顶菊(
Flaveria bidentis )、盐生草(
Halogeton glomeratus )等植物入侵时均会导致土壤养分含量向有利于入侵植物的方向发生改变(
Duda et al ., 2003 ;
蒋智林等, 2008 ;
张天瑞等, 2010 ). ...
入侵杂草加拿大一枝黄花的化感作用
1
2004
... 加拿大一枝黄花是菊科一枝黄花属植物, 多年生草本, 具有发达根状茎, 原产于北美地区, 最初作为庭园花卉引入我国上海、南京等地, 现已广泛逸生为恶性外来种(
中国科学院中国植物志编辑委员会, 1985 ;
郭水良和方芳, 2003 ).加拿大一枝黄花繁殖能力极强, 既能利用种子进行有性繁殖, 也可通过地下根状茎进行无性繁殖, 一般情况下, 每株植株可产约7万粒种子, 且种子具有较高萌发率, 此外, 植株根部具有上百个根茎结构, 每个根茎在第2年可萌发形成独立的个体, 如此高的繁殖能力使其在入侵后能够迅速扩散形成单优势种群落(
王玉良等, 2009 ;
张海亮等, 2015a ).此外, 加拿大一枝黄花根部还会产生一些化感物质影响周围土著植物生长, 对入侵地生物多样性造成了一定的影响(
方芳等, 2004 ).而目前加拿大一枝黄花在我国入侵范围逐渐扩大, 入侵速度逐渐加快, 对我国生态环境、生物多样性、农业和畜牧业均造成了不同程度的影响(
董梅等, 2006 ;
陆慧明等, 2006 ), 因此亟须加强加拿大一枝黄花入侵机理与防控技术方面的研究. ...
入侵杂草加拿大一枝黄花的化感作用
1
2004
... 加拿大一枝黄花是菊科一枝黄花属植物, 多年生草本, 具有发达根状茎, 原产于北美地区, 最初作为庭园花卉引入我国上海、南京等地, 现已广泛逸生为恶性外来种(
中国科学院中国植物志编辑委员会, 1985 ;
郭水良和方芳, 2003 ).加拿大一枝黄花繁殖能力极强, 既能利用种子进行有性繁殖, 也可通过地下根状茎进行无性繁殖, 一般情况下, 每株植株可产约7万粒种子, 且种子具有较高萌发率, 此外, 植株根部具有上百个根茎结构, 每个根茎在第2年可萌发形成独立的个体, 如此高的繁殖能力使其在入侵后能够迅速扩散形成单优势种群落(
王玉良等, 2009 ;
张海亮等, 2015a ).此外, 加拿大一枝黄花根部还会产生一些化感物质影响周围土著植物生长, 对入侵地生物多样性造成了一定的影响(
方芳等, 2004 ).而目前加拿大一枝黄花在我国入侵范围逐渐扩大, 入侵速度逐渐加快, 对我国生态环境、生物多样性、农业和畜牧业均造成了不同程度的影响(
董梅等, 2006 ;
陆慧明等, 2006 ), 因此亟须加强加拿大一枝黄花入侵机理与防控技术方面的研究. ...
黄瓜根际解钾细菌的分离筛选、鉴定及其促生效果
1
2017
... 本研究发现的加拿大一枝黄花和白茅根际解钾菌的种类和此前报道的研究具有一定的相似性.国际公认的解钾菌菌种仅3种: 环状芽孢杆菌(
Bacillus circulans )、胶质芽孢杆菌(
Bacillus mucilaginosus )、土壤芽孢杆菌(
Bacillus edaphicus ), 但近年来解钾菌的种属鉴定结果越来越丰富, 新增的种属有假单胞菌属(
Pseudomonas )(
吴凡等, 2010 ;
李新新等, 2014 ;
葛红莲和纪秀娥, 2017 )、根瘤菌属(
Rhizobium )、肠杆菌属(
Enterobacteriaceae )(
吴凡等, 2010 )、微杆菌属(
Microbacterium )、固氮菌属(
Azotobacter )(
易浪波等, 2012 ;
李新新等, 2014 )、中华根瘤菌属(
Sinorhizobium )、慢生根瘤菌属(
Mesorhizobium )、屈挠杆菌属(
Flexibacter )(
鞠伟, 2016 ;
李春钢等, 2017 )等.在加拿大一枝黄花和白茅根际土中分离获得的20株解钾菌中, H1-4、B6-7与
吴凡等(2010) 的研究结果一致, H6-5和H1-6、H5-10、H2-14、H2-17与
李新新等(2014) 的研究结果相同, H3-8与
张妙宜等(2016) 筛选出的解钾菌同属, 以上菌株均被证明具有一定的解钾能力.此外, 本研究发现的解钾能力较强但未被报道的菌株有: H1-15, 属于含铜菌属(
Cupriavidus ); H5-18, 属于
Pseudoflavitalea; H2-20, 属于溶杆菌属(
Lysobacter ); H1-21, 属于
Siphonobacter, 这一研究结果有助于进一步丰富解钾菌的种属类别. ...
黄瓜根际解钾细菌的分离筛选、鉴定及其促生效果
1
2017
... 本研究发现的加拿大一枝黄花和白茅根际解钾菌的种类和此前报道的研究具有一定的相似性.国际公认的解钾菌菌种仅3种: 环状芽孢杆菌(
Bacillus circulans )、胶质芽孢杆菌(
Bacillus mucilaginosus )、土壤芽孢杆菌(
Bacillus edaphicus ), 但近年来解钾菌的种属鉴定结果越来越丰富, 新增的种属有假单胞菌属(
Pseudomonas )(
吴凡等, 2010 ;
李新新等, 2014 ;
葛红莲和纪秀娥, 2017 )、根瘤菌属(
Rhizobium )、肠杆菌属(
Enterobacteriaceae )(
吴凡等, 2010 )、微杆菌属(
Microbacterium )、固氮菌属(
Azotobacter )(
易浪波等, 2012 ;
李新新等, 2014 )、中华根瘤菌属(
Sinorhizobium )、慢生根瘤菌属(
Mesorhizobium )、屈挠杆菌属(
Flexibacter )(
鞠伟, 2016 ;
李春钢等, 2017 )等.在加拿大一枝黄花和白茅根际土中分离获得的20株解钾菌中, H1-4、B6-7与
吴凡等(2010) 的研究结果一致, H6-5和H1-6、H5-10、H2-14、H2-17与
李新新等(2014) 的研究结果相同, H3-8与
张妙宜等(2016) 筛选出的解钾菌同属, 以上菌株均被证明具有一定的解钾能力.此外, 本研究发现的解钾能力较强但未被报道的菌株有: H1-15, 属于含铜菌属(
Cupriavidus ); H5-18, 属于
Pseudoflavitalea; H2-20, 属于溶杆菌属(
Lysobacter ); H1-21, 属于
Siphonobacter, 这一研究结果有助于进一步丰富解钾菌的种属类别. ...
Potassium deficiency-induced changes in stomatal behavior, leaf water potentials, and root system permeability in
Beta vulgaris L
1
1972
... 入侵植物对营养元素的吸收、积累和释放对其生长、入侵能力及其入侵的生态学效应都具有重要作用.钾是植物生长发育所需要的主要营养元素之一, 在植物生长繁育过程中发挥必不可少的作用, 钾的增加不仅能够提高植物光合速率和酶的活化速度、增加植物糖和蛋白质含量、增强植物对氮的吸收, 还能够提高作物的产量、提高植物的抗逆性(
Graham & Ulrich, 1972 ;
胡笃敬等, 1993 ).本研究发现, 野外自然生长的加拿大一枝黄花叶片和茎中的钾含量显著高于伴生植物白茅, 尤其叶片中的钾含量是白茅的7.33倍, 说明在相同生境下, 加拿大一枝黄花吸收钾的能力远高于本地植物, 且加拿大一枝黄花能够将更多的钾分配到叶片中, 而叶片是光合作用最主要的场所, 叶片中养分含量增多可能增强植物光合速率和生长代谢速率(
Aerts, 2000 ;
王丽芳等, 2013 ).此外, 加拿大一枝黄花叶片中的氮磷含量明显高于茎, 更加证实了加拿大一枝黄花将更多养分分配到叶片中的生长策略.人为提高钾供给水平均能促进两种植物生物量积累增加(
图3 ), 但是我们发现加拿大一枝黄花入侵地土壤0-10 cm及10-20 cm土层中全氮、全磷、全钾及碱解氮、有效磷和速效钾含量均多于对照植物白茅, 尤其土壤中速效养分的含量显著多于白茅, 这与
梁雷等(2016) 的研究结果相一致.
陈彤等(2012) 发现加拿大一枝黄花能显著提高土壤中硝态氮含量, 并且其较高的氮积累能力可能和入侵后本地植物种群的动态变化格局紧密相关 这些研究说明加拿大一枝黄花存在一种能够增强土壤养分积累的机制, 而这种机制反过来可能促进其进一步入侵.外来植物对入侵地土壤养分含量的影响被大量研究.一些研究认为外来植物入侵往往能够通过影响入侵地土壤养分而促进自身成功入侵(
Holmgren et al ., 1997 ).许多外来植物入侵后通过改变土壤肥力影响其与本地种的竞争能力, 例如破坏草、黄顶菊(
Flaveria bidentis )、盐生草(
Halogeton glomeratus )等植物入侵时均会导致土壤养分含量向有利于入侵植物的方向发生改变(
Duda et al ., 2003 ;
蒋智林等, 2008 ;
张天瑞等, 2010 ). ...
入侵植物加拿大一枝黄花对环境的生理适应性研究
2
2003
... 加拿大一枝黄花是菊科一枝黄花属植物, 多年生草本, 具有发达根状茎, 原产于北美地区, 最初作为庭园花卉引入我国上海、南京等地, 现已广泛逸生为恶性外来种(
中国科学院中国植物志编辑委员会, 1985 ;
郭水良和方芳, 2003 ).加拿大一枝黄花繁殖能力极强, 既能利用种子进行有性繁殖, 也可通过地下根状茎进行无性繁殖, 一般情况下, 每株植株可产约7万粒种子, 且种子具有较高萌发率, 此外, 植株根部具有上百个根茎结构, 每个根茎在第2年可萌发形成独立的个体, 如此高的繁殖能力使其在入侵后能够迅速扩散形成单优势种群落(
王玉良等, 2009 ;
张海亮等, 2015a ).此外, 加拿大一枝黄花根部还会产生一些化感物质影响周围土著植物生长, 对入侵地生物多样性造成了一定的影响(
方芳等, 2004 ).而目前加拿大一枝黄花在我国入侵范围逐渐扩大, 入侵速度逐渐加快, 对我国生态环境、生物多样性、农业和畜牧业均造成了不同程度的影响(
董梅等, 2006 ;
陆慧明等, 2006 ), 因此亟须加强加拿大一枝黄花入侵机理与防控技术方面的研究. ...
... 目前国内外对加拿大一枝黄花入侵机制的研究主要集中在繁殖特性(
黄华和郭水良, 2005 ;
郝建华等, 2009 ;
张海亮等, 2015b )、化感作用(
梅玲笑, 2005 ;
郝建华等, 2009 )、生理生态特征与环境适应性(
郭水良和方芳, 2003 ;
杜乐山, 2014 )以及土壤微生物响应(
Zhang et al ., 2005 ;
沈荔花等, 2007 )等方面.研究表明, 在不同程度的入侵地中, 加拿大一枝黄花体内钾含量均高于伴生植物, 尤其在生长期和开花期差异最为显著, 且植物吸钾量与生物量呈正相关关系(
周振荣, 2010 ).在微生物响应方面, 研究发现加拿大一枝黄花入侵后能改变土壤微生物的群落数量、结构和功能多样性, 可能有助于加拿大一枝黄花进一步入侵(
Liao et al ., 2013 ;
Wang et al ., 2018 ).根际解钾菌能将土壤中难溶性钾转化为可被植物直接吸收的有效性钾, 进而促进植物钾吸收, 增强植物抗逆性(
李元芳, 1994 ;
郭勋斌等, 2001 ).加拿大一枝黄花入侵是否改变了根际解钾菌数量、结构和解钾活性等问题还有待深入研究.因此, 本研究以入侵植物加拿大一枝黄花和伴生本地植物白茅(
Imperata cylindrica )为研究对象, 比较加拿大一枝黄花与白茅的根际解钾菌数量、多样性与解钾能力差异, 旨在初步探讨根际解钾菌在加拿大一枝黄花入侵过程中发挥的作用, 为深入研究加拿大一枝黄花入侵机制提供理论依据. ...
入侵植物加拿大一枝黄花对环境的生理适应性研究
2
2003
... 加拿大一枝黄花是菊科一枝黄花属植物, 多年生草本, 具有发达根状茎, 原产于北美地区, 最初作为庭园花卉引入我国上海、南京等地, 现已广泛逸生为恶性外来种(
中国科学院中国植物志编辑委员会, 1985 ;
郭水良和方芳, 2003 ).加拿大一枝黄花繁殖能力极强, 既能利用种子进行有性繁殖, 也可通过地下根状茎进行无性繁殖, 一般情况下, 每株植株可产约7万粒种子, 且种子具有较高萌发率, 此外, 植株根部具有上百个根茎结构, 每个根茎在第2年可萌发形成独立的个体, 如此高的繁殖能力使其在入侵后能够迅速扩散形成单优势种群落(
王玉良等, 2009 ;
张海亮等, 2015a ).此外, 加拿大一枝黄花根部还会产生一些化感物质影响周围土著植物生长, 对入侵地生物多样性造成了一定的影响(
方芳等, 2004 ).而目前加拿大一枝黄花在我国入侵范围逐渐扩大, 入侵速度逐渐加快, 对我国生态环境、生物多样性、农业和畜牧业均造成了不同程度的影响(
董梅等, 2006 ;
陆慧明等, 2006 ), 因此亟须加强加拿大一枝黄花入侵机理与防控技术方面的研究. ...
... 目前国内外对加拿大一枝黄花入侵机制的研究主要集中在繁殖特性(
黄华和郭水良, 2005 ;
郝建华等, 2009 ;
张海亮等, 2015b )、化感作用(
梅玲笑, 2005 ;
郝建华等, 2009 )、生理生态特征与环境适应性(
郭水良和方芳, 2003 ;
杜乐山, 2014 )以及土壤微生物响应(
Zhang et al ., 2005 ;
沈荔花等, 2007 )等方面.研究表明, 在不同程度的入侵地中, 加拿大一枝黄花体内钾含量均高于伴生植物, 尤其在生长期和开花期差异最为显著, 且植物吸钾量与生物量呈正相关关系(
周振荣, 2010 ).在微生物响应方面, 研究发现加拿大一枝黄花入侵后能改变土壤微生物的群落数量、结构和功能多样性, 可能有助于加拿大一枝黄花进一步入侵(
Liao et al ., 2013 ;
Wang et al ., 2018 ).根际解钾菌能将土壤中难溶性钾转化为可被植物直接吸收的有效性钾, 进而促进植物钾吸收, 增强植物抗逆性(
李元芳, 1994 ;
郭勋斌等, 2001 ).加拿大一枝黄花入侵是否改变了根际解钾菌数量、结构和解钾活性等问题还有待深入研究.因此, 本研究以入侵植物加拿大一枝黄花和伴生本地植物白茅(
Imperata cylindrica )为研究对象, 比较加拿大一枝黄花与白茅的根际解钾菌数量、多样性与解钾能力差异, 旨在初步探讨根际解钾菌在加拿大一枝黄花入侵过程中发挥的作用, 为深入研究加拿大一枝黄花入侵机制提供理论依据. ...
钾细菌制剂对水稻生长发育的影响
1
2001
... 目前国内外对加拿大一枝黄花入侵机制的研究主要集中在繁殖特性(
黄华和郭水良, 2005 ;
郝建华等, 2009 ;
张海亮等, 2015b )、化感作用(
梅玲笑, 2005 ;
郝建华等, 2009 )、生理生态特征与环境适应性(
郭水良和方芳, 2003 ;
杜乐山, 2014 )以及土壤微生物响应(
Zhang et al ., 2005 ;
沈荔花等, 2007 )等方面.研究表明, 在不同程度的入侵地中, 加拿大一枝黄花体内钾含量均高于伴生植物, 尤其在生长期和开花期差异最为显著, 且植物吸钾量与生物量呈正相关关系(
周振荣, 2010 ).在微生物响应方面, 研究发现加拿大一枝黄花入侵后能改变土壤微生物的群落数量、结构和功能多样性, 可能有助于加拿大一枝黄花进一步入侵(
Liao et al ., 2013 ;
Wang et al ., 2018 ).根际解钾菌能将土壤中难溶性钾转化为可被植物直接吸收的有效性钾, 进而促进植物钾吸收, 增强植物抗逆性(
李元芳, 1994 ;
郭勋斌等, 2001 ).加拿大一枝黄花入侵是否改变了根际解钾菌数量、结构和解钾活性等问题还有待深入研究.因此, 本研究以入侵植物加拿大一枝黄花和伴生本地植物白茅(
Imperata cylindrica )为研究对象, 比较加拿大一枝黄花与白茅的根际解钾菌数量、多样性与解钾能力差异, 旨在初步探讨根际解钾菌在加拿大一枝黄花入侵过程中发挥的作用, 为深入研究加拿大一枝黄花入侵机制提供理论依据. ...
钾细菌制剂对水稻生长发育的影响
1
2001
... 目前国内外对加拿大一枝黄花入侵机制的研究主要集中在繁殖特性(
黄华和郭水良, 2005 ;
郝建华等, 2009 ;
张海亮等, 2015b )、化感作用(
梅玲笑, 2005 ;
郝建华等, 2009 )、生理生态特征与环境适应性(
郭水良和方芳, 2003 ;
杜乐山, 2014 )以及土壤微生物响应(
Zhang et al ., 2005 ;
沈荔花等, 2007 )等方面.研究表明, 在不同程度的入侵地中, 加拿大一枝黄花体内钾含量均高于伴生植物, 尤其在生长期和开花期差异最为显著, 且植物吸钾量与生物量呈正相关关系(
周振荣, 2010 ).在微生物响应方面, 研究发现加拿大一枝黄花入侵后能改变土壤微生物的群落数量、结构和功能多样性, 可能有助于加拿大一枝黄花进一步入侵(
Liao et al ., 2013 ;
Wang et al ., 2018 ).根际解钾菌能将土壤中难溶性钾转化为可被植物直接吸收的有效性钾, 进而促进植物钾吸收, 增强植物抗逆性(
李元芳, 1994 ;
郭勋斌等, 2001 ).加拿大一枝黄花入侵是否改变了根际解钾菌数量、结构和解钾活性等问题还有待深入研究.因此, 本研究以入侵植物加拿大一枝黄花和伴生本地植物白茅(
Imperata cylindrica )为研究对象, 比较加拿大一枝黄花与白茅的根际解钾菌数量、多样性与解钾能力差异, 旨在初步探讨根际解钾菌在加拿大一枝黄花入侵过程中发挥的作用, 为深入研究加拿大一枝黄花入侵机制提供理论依据. ...
加拿大一枝黄花有性生殖特征研究
2
2009
... 目前国内外对加拿大一枝黄花入侵机制的研究主要集中在繁殖特性(
黄华和郭水良, 2005 ;
郝建华等, 2009 ;
张海亮等, 2015b )、化感作用(
梅玲笑, 2005 ;
郝建华等, 2009 )、生理生态特征与环境适应性(
郭水良和方芳, 2003 ;
杜乐山, 2014 )以及土壤微生物响应(
Zhang et al ., 2005 ;
沈荔花等, 2007 )等方面.研究表明, 在不同程度的入侵地中, 加拿大一枝黄花体内钾含量均高于伴生植物, 尤其在生长期和开花期差异最为显著, 且植物吸钾量与生物量呈正相关关系(
周振荣, 2010 ).在微生物响应方面, 研究发现加拿大一枝黄花入侵后能改变土壤微生物的群落数量、结构和功能多样性, 可能有助于加拿大一枝黄花进一步入侵(
Liao et al ., 2013 ;
Wang et al ., 2018 ).根际解钾菌能将土壤中难溶性钾转化为可被植物直接吸收的有效性钾, 进而促进植物钾吸收, 增强植物抗逆性(
李元芳, 1994 ;
郭勋斌等, 2001 ).加拿大一枝黄花入侵是否改变了根际解钾菌数量、结构和解钾活性等问题还有待深入研究.因此, 本研究以入侵植物加拿大一枝黄花和伴生本地植物白茅(
Imperata cylindrica )为研究对象, 比较加拿大一枝黄花与白茅的根际解钾菌数量、多样性与解钾能力差异, 旨在初步探讨根际解钾菌在加拿大一枝黄花入侵过程中发挥的作用, 为深入研究加拿大一枝黄花入侵机制提供理论依据. ...
... ;
郝建华等, 2009 )、生理生态特征与环境适应性(
郭水良和方芳, 2003 ;
杜乐山, 2014 )以及土壤微生物响应(
Zhang et al ., 2005 ;
沈荔花等, 2007 )等方面.研究表明, 在不同程度的入侵地中, 加拿大一枝黄花体内钾含量均高于伴生植物, 尤其在生长期和开花期差异最为显著, 且植物吸钾量与生物量呈正相关关系(
周振荣, 2010 ).在微生物响应方面, 研究发现加拿大一枝黄花入侵后能改变土壤微生物的群落数量、结构和功能多样性, 可能有助于加拿大一枝黄花进一步入侵(
Liao et al ., 2013 ;
Wang et al ., 2018 ).根际解钾菌能将土壤中难溶性钾转化为可被植物直接吸收的有效性钾, 进而促进植物钾吸收, 增强植物抗逆性(
李元芳, 1994 ;
郭勋斌等, 2001 ).加拿大一枝黄花入侵是否改变了根际解钾菌数量、结构和解钾活性等问题还有待深入研究.因此, 本研究以入侵植物加拿大一枝黄花和伴生本地植物白茅(
Imperata cylindrica )为研究对象, 比较加拿大一枝黄花与白茅的根际解钾菌数量、多样性与解钾能力差异, 旨在初步探讨根际解钾菌在加拿大一枝黄花入侵过程中发挥的作用, 为深入研究加拿大一枝黄花入侵机制提供理论依据. ...
加拿大一枝黄花有性生殖特征研究
2
2009
... 目前国内外对加拿大一枝黄花入侵机制的研究主要集中在繁殖特性(
黄华和郭水良, 2005 ;
郝建华等, 2009 ;
张海亮等, 2015b )、化感作用(
梅玲笑, 2005 ;
郝建华等, 2009 )、生理生态特征与环境适应性(
郭水良和方芳, 2003 ;
杜乐山, 2014 )以及土壤微生物响应(
Zhang et al ., 2005 ;
沈荔花等, 2007 )等方面.研究表明, 在不同程度的入侵地中, 加拿大一枝黄花体内钾含量均高于伴生植物, 尤其在生长期和开花期差异最为显著, 且植物吸钾量与生物量呈正相关关系(
周振荣, 2010 ).在微生物响应方面, 研究发现加拿大一枝黄花入侵后能改变土壤微生物的群落数量、结构和功能多样性, 可能有助于加拿大一枝黄花进一步入侵(
Liao et al ., 2013 ;
Wang et al ., 2018 ).根际解钾菌能将土壤中难溶性钾转化为可被植物直接吸收的有效性钾, 进而促进植物钾吸收, 增强植物抗逆性(
李元芳, 1994 ;
郭勋斌等, 2001 ).加拿大一枝黄花入侵是否改变了根际解钾菌数量、结构和解钾活性等问题还有待深入研究.因此, 本研究以入侵植物加拿大一枝黄花和伴生本地植物白茅(
Imperata cylindrica )为研究对象, 比较加拿大一枝黄花与白茅的根际解钾菌数量、多样性与解钾能力差异, 旨在初步探讨根际解钾菌在加拿大一枝黄花入侵过程中发挥的作用, 为深入研究加拿大一枝黄花入侵机制提供理论依据. ...
... ;
郝建华等, 2009 )、生理生态特征与环境适应性(
郭水良和方芳, 2003 ;
杜乐山, 2014 )以及土壤微生物响应(
Zhang et al ., 2005 ;
沈荔花等, 2007 )等方面.研究表明, 在不同程度的入侵地中, 加拿大一枝黄花体内钾含量均高于伴生植物, 尤其在生长期和开花期差异最为显著, 且植物吸钾量与生物量呈正相关关系(
周振荣, 2010 ).在微生物响应方面, 研究发现加拿大一枝黄花入侵后能改变土壤微生物的群落数量、结构和功能多样性, 可能有助于加拿大一枝黄花进一步入侵(
Liao et al ., 2013 ;
Wang et al ., 2018 ).根际解钾菌能将土壤中难溶性钾转化为可被植物直接吸收的有效性钾, 进而促进植物钾吸收, 增强植物抗逆性(
李元芳, 1994 ;
郭勋斌等, 2001 ).加拿大一枝黄花入侵是否改变了根际解钾菌数量、结构和解钾活性等问题还有待深入研究.因此, 本研究以入侵植物加拿大一枝黄花和伴生本地植物白茅(
Imperata cylindrica )为研究对象, 比较加拿大一枝黄花与白茅的根际解钾菌数量、多样性与解钾能力差异, 旨在初步探讨根际解钾菌在加拿大一枝黄花入侵过程中发挥的作用, 为深入研究加拿大一枝黄花入侵机制提供理论依据. ...
The interplay of facilitation and competition in plant communities
1
1997
... 入侵植物对营养元素的吸收、积累和释放对其生长、入侵能力及其入侵的生态学效应都具有重要作用.钾是植物生长发育所需要的主要营养元素之一, 在植物生长繁育过程中发挥必不可少的作用, 钾的增加不仅能够提高植物光合速率和酶的活化速度、增加植物糖和蛋白质含量、增强植物对氮的吸收, 还能够提高作物的产量、提高植物的抗逆性(
Graham & Ulrich, 1972 ;
胡笃敬等, 1993 ).本研究发现, 野外自然生长的加拿大一枝黄花叶片和茎中的钾含量显著高于伴生植物白茅, 尤其叶片中的钾含量是白茅的7.33倍, 说明在相同生境下, 加拿大一枝黄花吸收钾的能力远高于本地植物, 且加拿大一枝黄花能够将更多的钾分配到叶片中, 而叶片是光合作用最主要的场所, 叶片中养分含量增多可能增强植物光合速率和生长代谢速率(
Aerts, 2000 ;
王丽芳等, 2013 ).此外, 加拿大一枝黄花叶片中的氮磷含量明显高于茎, 更加证实了加拿大一枝黄花将更多养分分配到叶片中的生长策略.人为提高钾供给水平均能促进两种植物生物量积累增加(
图3 ), 但是我们发现加拿大一枝黄花入侵地土壤0-10 cm及10-20 cm土层中全氮、全磷、全钾及碱解氮、有效磷和速效钾含量均多于对照植物白茅, 尤其土壤中速效养分的含量显著多于白茅, 这与
梁雷等(2016) 的研究结果相一致.
陈彤等(2012) 发现加拿大一枝黄花能显著提高土壤中硝态氮含量, 并且其较高的氮积累能力可能和入侵后本地植物种群的动态变化格局紧密相关 这些研究说明加拿大一枝黄花存在一种能够增强土壤养分积累的机制, 而这种机制反过来可能促进其进一步入侵.外来植物对入侵地土壤养分含量的影响被大量研究.一些研究认为外来植物入侵往往能够通过影响入侵地土壤养分而促进自身成功入侵(
Holmgren et al ., 1997 ).许多外来植物入侵后通过改变土壤肥力影响其与本地种的竞争能力, 例如破坏草、黄顶菊(
Flaveria bidentis )、盐生草(
Halogeton glomeratus )等植物入侵时均会导致土壤养分含量向有利于入侵植物的方向发生改变(
Duda et al ., 2003 ;
蒋智林等, 2008 ;
张天瑞等, 2010 ). ...
1
1993
... 入侵植物对营养元素的吸收、积累和释放对其生长、入侵能力及其入侵的生态学效应都具有重要作用.钾是植物生长发育所需要的主要营养元素之一, 在植物生长繁育过程中发挥必不可少的作用, 钾的增加不仅能够提高植物光合速率和酶的活化速度、增加植物糖和蛋白质含量、增强植物对氮的吸收, 还能够提高作物的产量、提高植物的抗逆性(
Graham & Ulrich, 1972 ;
胡笃敬等, 1993 ).本研究发现, 野外自然生长的加拿大一枝黄花叶片和茎中的钾含量显著高于伴生植物白茅, 尤其叶片中的钾含量是白茅的7.33倍, 说明在相同生境下, 加拿大一枝黄花吸收钾的能力远高于本地植物, 且加拿大一枝黄花能够将更多的钾分配到叶片中, 而叶片是光合作用最主要的场所, 叶片中养分含量增多可能增强植物光合速率和生长代谢速率(
Aerts, 2000 ;
王丽芳等, 2013 ).此外, 加拿大一枝黄花叶片中的氮磷含量明显高于茎, 更加证实了加拿大一枝黄花将更多养分分配到叶片中的生长策略.人为提高钾供给水平均能促进两种植物生物量积累增加(
图3 ), 但是我们发现加拿大一枝黄花入侵地土壤0-10 cm及10-20 cm土层中全氮、全磷、全钾及碱解氮、有效磷和速效钾含量均多于对照植物白茅, 尤其土壤中速效养分的含量显著多于白茅, 这与
梁雷等(2016) 的研究结果相一致.
陈彤等(2012) 发现加拿大一枝黄花能显著提高土壤中硝态氮含量, 并且其较高的氮积累能力可能和入侵后本地植物种群的动态变化格局紧密相关 这些研究说明加拿大一枝黄花存在一种能够增强土壤养分积累的机制, 而这种机制反过来可能促进其进一步入侵.外来植物对入侵地土壤养分含量的影响被大量研究.一些研究认为外来植物入侵往往能够通过影响入侵地土壤养分而促进自身成功入侵(
Holmgren et al ., 1997 ).许多外来植物入侵后通过改变土壤肥力影响其与本地种的竞争能力, 例如破坏草、黄顶菊(
Flaveria bidentis )、盐生草(
Halogeton glomeratus )等植物入侵时均会导致土壤养分含量向有利于入侵植物的方向发生改变(
Duda et al ., 2003 ;
蒋智林等, 2008 ;
张天瑞等, 2010 ). ...
1
1993
... 入侵植物对营养元素的吸收、积累和释放对其生长、入侵能力及其入侵的生态学效应都具有重要作用.钾是植物生长发育所需要的主要营养元素之一, 在植物生长繁育过程中发挥必不可少的作用, 钾的增加不仅能够提高植物光合速率和酶的活化速度、增加植物糖和蛋白质含量、增强植物对氮的吸收, 还能够提高作物的产量、提高植物的抗逆性(
Graham & Ulrich, 1972 ;
胡笃敬等, 1993 ).本研究发现, 野外自然生长的加拿大一枝黄花叶片和茎中的钾含量显著高于伴生植物白茅, 尤其叶片中的钾含量是白茅的7.33倍, 说明在相同生境下, 加拿大一枝黄花吸收钾的能力远高于本地植物, 且加拿大一枝黄花能够将更多的钾分配到叶片中, 而叶片是光合作用最主要的场所, 叶片中养分含量增多可能增强植物光合速率和生长代谢速率(
Aerts, 2000 ;
王丽芳等, 2013 ).此外, 加拿大一枝黄花叶片中的氮磷含量明显高于茎, 更加证实了加拿大一枝黄花将更多养分分配到叶片中的生长策略.人为提高钾供给水平均能促进两种植物生物量积累增加(
图3 ), 但是我们发现加拿大一枝黄花入侵地土壤0-10 cm及10-20 cm土层中全氮、全磷、全钾及碱解氮、有效磷和速效钾含量均多于对照植物白茅, 尤其土壤中速效养分的含量显著多于白茅, 这与
梁雷等(2016) 的研究结果相一致.
陈彤等(2012) 发现加拿大一枝黄花能显著提高土壤中硝态氮含量, 并且其较高的氮积累能力可能和入侵后本地植物种群的动态变化格局紧密相关 这些研究说明加拿大一枝黄花存在一种能够增强土壤养分积累的机制, 而这种机制反过来可能促进其进一步入侵.外来植物对入侵地土壤养分含量的影响被大量研究.一些研究认为外来植物入侵往往能够通过影响入侵地土壤养分而促进自身成功入侵(
Holmgren et al ., 1997 ).许多外来植物入侵后通过改变土壤肥力影响其与本地种的竞争能力, 例如破坏草、黄顶菊(
Flaveria bidentis )、盐生草(
Halogeton glomeratus )等植物入侵时均会导致土壤养分含量向有利于入侵植物的方向发生改变(
Duda et al ., 2003 ;
蒋智林等, 2008 ;
张天瑞等, 2010 ). ...
外来入侵植物加拿大一枝黄花繁殖生物学研究
1
2005
... 目前国内外对加拿大一枝黄花入侵机制的研究主要集中在繁殖特性(
黄华和郭水良, 2005 ;
郝建华等, 2009 ;
张海亮等, 2015b )、化感作用(
梅玲笑, 2005 ;
郝建华等, 2009 )、生理生态特征与环境适应性(
郭水良和方芳, 2003 ;
杜乐山, 2014 )以及土壤微生物响应(
Zhang et al ., 2005 ;
沈荔花等, 2007 )等方面.研究表明, 在不同程度的入侵地中, 加拿大一枝黄花体内钾含量均高于伴生植物, 尤其在生长期和开花期差异最为显著, 且植物吸钾量与生物量呈正相关关系(
周振荣, 2010 ).在微生物响应方面, 研究发现加拿大一枝黄花入侵后能改变土壤微生物的群落数量、结构和功能多样性, 可能有助于加拿大一枝黄花进一步入侵(
Liao et al ., 2013 ;
Wang et al ., 2018 ).根际解钾菌能将土壤中难溶性钾转化为可被植物直接吸收的有效性钾, 进而促进植物钾吸收, 增强植物抗逆性(
李元芳, 1994 ;
郭勋斌等, 2001 ).加拿大一枝黄花入侵是否改变了根际解钾菌数量、结构和解钾活性等问题还有待深入研究.因此, 本研究以入侵植物加拿大一枝黄花和伴生本地植物白茅(
Imperata cylindrica )为研究对象, 比较加拿大一枝黄花与白茅的根际解钾菌数量、多样性与解钾能力差异, 旨在初步探讨根际解钾菌在加拿大一枝黄花入侵过程中发挥的作用, 为深入研究加拿大一枝黄花入侵机制提供理论依据. ...
外来入侵植物加拿大一枝黄花繁殖生物学研究
1
2005
... 目前国内外对加拿大一枝黄花入侵机制的研究主要集中在繁殖特性(
黄华和郭水良, 2005 ;
郝建华等, 2009 ;
张海亮等, 2015b )、化感作用(
梅玲笑, 2005 ;
郝建华等, 2009 )、生理生态特征与环境适应性(
郭水良和方芳, 2003 ;
杜乐山, 2014 )以及土壤微生物响应(
Zhang et al ., 2005 ;
沈荔花等, 2007 )等方面.研究表明, 在不同程度的入侵地中, 加拿大一枝黄花体内钾含量均高于伴生植物, 尤其在生长期和开花期差异最为显著, 且植物吸钾量与生物量呈正相关关系(
周振荣, 2010 ).在微生物响应方面, 研究发现加拿大一枝黄花入侵后能改变土壤微生物的群落数量、结构和功能多样性, 可能有助于加拿大一枝黄花进一步入侵(
Liao et al ., 2013 ;
Wang et al ., 2018 ).根际解钾菌能将土壤中难溶性钾转化为可被植物直接吸收的有效性钾, 进而促进植物钾吸收, 增强植物抗逆性(
李元芳, 1994 ;
郭勋斌等, 2001 ).加拿大一枝黄花入侵是否改变了根际解钾菌数量、结构和解钾活性等问题还有待深入研究.因此, 本研究以入侵植物加拿大一枝黄花和伴生本地植物白茅(
Imperata cylindrica )为研究对象, 比较加拿大一枝黄花与白茅的根际解钾菌数量、多样性与解钾能力差异, 旨在初步探讨根际解钾菌在加拿大一枝黄花入侵过程中发挥的作用, 为深入研究加拿大一枝黄花入侵机制提供理论依据. ...
紫茎泽兰与本地植物群落根际土壤酶活性和土壤肥力的差异
1
2008
... 入侵植物对营养元素的吸收、积累和释放对其生长、入侵能力及其入侵的生态学效应都具有重要作用.钾是植物生长发育所需要的主要营养元素之一, 在植物生长繁育过程中发挥必不可少的作用, 钾的增加不仅能够提高植物光合速率和酶的活化速度、增加植物糖和蛋白质含量、增强植物对氮的吸收, 还能够提高作物的产量、提高植物的抗逆性(
Graham & Ulrich, 1972 ;
胡笃敬等, 1993 ).本研究发现, 野外自然生长的加拿大一枝黄花叶片和茎中的钾含量显著高于伴生植物白茅, 尤其叶片中的钾含量是白茅的7.33倍, 说明在相同生境下, 加拿大一枝黄花吸收钾的能力远高于本地植物, 且加拿大一枝黄花能够将更多的钾分配到叶片中, 而叶片是光合作用最主要的场所, 叶片中养分含量增多可能增强植物光合速率和生长代谢速率(
Aerts, 2000 ;
王丽芳等, 2013 ).此外, 加拿大一枝黄花叶片中的氮磷含量明显高于茎, 更加证实了加拿大一枝黄花将更多养分分配到叶片中的生长策略.人为提高钾供给水平均能促进两种植物生物量积累增加(
图3 ), 但是我们发现加拿大一枝黄花入侵地土壤0-10 cm及10-20 cm土层中全氮、全磷、全钾及碱解氮、有效磷和速效钾含量均多于对照植物白茅, 尤其土壤中速效养分的含量显著多于白茅, 这与
梁雷等(2016) 的研究结果相一致.
陈彤等(2012) 发现加拿大一枝黄花能显著提高土壤中硝态氮含量, 并且其较高的氮积累能力可能和入侵后本地植物种群的动态变化格局紧密相关 这些研究说明加拿大一枝黄花存在一种能够增强土壤养分积累的机制, 而这种机制反过来可能促进其进一步入侵.外来植物对入侵地土壤养分含量的影响被大量研究.一些研究认为外来植物入侵往往能够通过影响入侵地土壤养分而促进自身成功入侵(
Holmgren et al ., 1997 ).许多外来植物入侵后通过改变土壤肥力影响其与本地种的竞争能力, 例如破坏草、黄顶菊(
Flaveria bidentis )、盐生草(
Halogeton glomeratus )等植物入侵时均会导致土壤养分含量向有利于入侵植物的方向发生改变(
Duda et al ., 2003 ;
蒋智林等, 2008 ;
张天瑞等, 2010 ). ...
紫茎泽兰与本地植物群落根际土壤酶活性和土壤肥力的差异
1
2008
... 入侵植物对营养元素的吸收、积累和释放对其生长、入侵能力及其入侵的生态学效应都具有重要作用.钾是植物生长发育所需要的主要营养元素之一, 在植物生长繁育过程中发挥必不可少的作用, 钾的增加不仅能够提高植物光合速率和酶的活化速度、增加植物糖和蛋白质含量、增强植物对氮的吸收, 还能够提高作物的产量、提高植物的抗逆性(
Graham & Ulrich, 1972 ;
胡笃敬等, 1993 ).本研究发现, 野外自然生长的加拿大一枝黄花叶片和茎中的钾含量显著高于伴生植物白茅, 尤其叶片中的钾含量是白茅的7.33倍, 说明在相同生境下, 加拿大一枝黄花吸收钾的能力远高于本地植物, 且加拿大一枝黄花能够将更多的钾分配到叶片中, 而叶片是光合作用最主要的场所, 叶片中养分含量增多可能增强植物光合速率和生长代谢速率(
Aerts, 2000 ;
王丽芳等, 2013 ).此外, 加拿大一枝黄花叶片中的氮磷含量明显高于茎, 更加证实了加拿大一枝黄花将更多养分分配到叶片中的生长策略.人为提高钾供给水平均能促进两种植物生物量积累增加(
图3 ), 但是我们发现加拿大一枝黄花入侵地土壤0-10 cm及10-20 cm土层中全氮、全磷、全钾及碱解氮、有效磷和速效钾含量均多于对照植物白茅, 尤其土壤中速效养分的含量显著多于白茅, 这与
梁雷等(2016) 的研究结果相一致.
陈彤等(2012) 发现加拿大一枝黄花能显著提高土壤中硝态氮含量, 并且其较高的氮积累能力可能和入侵后本地植物种群的动态变化格局紧密相关 这些研究说明加拿大一枝黄花存在一种能够增强土壤养分积累的机制, 而这种机制反过来可能促进其进一步入侵.外来植物对入侵地土壤养分含量的影响被大量研究.一些研究认为外来植物入侵往往能够通过影响入侵地土壤养分而促进自身成功入侵(
Holmgren et al ., 1997 ).许多外来植物入侵后通过改变土壤肥力影响其与本地种的竞争能力, 例如破坏草、黄顶菊(
Flaveria bidentis )、盐生草(
Halogeton glomeratus )等植物入侵时均会导致土壤养分含量向有利于入侵植物的方向发生改变(
Duda et al ., 2003 ;
蒋智林等, 2008 ;
张天瑞等, 2010 ). ...
近十年中国生物入侵研究进展
1
2012
... 外来植物入侵不仅导致入侵地农林生产经济损失, 而且对生态系统和生物多样性造成了严重的威胁(
鞠瑞亭等, 2012 ).据调查, 我国外来入侵植物多达670余种, 分布范围主要集中在东南沿海地区, 且入侵速度正在逐年加快、数量逐年增多, 若不加以防治, 则直接影响我国经济发展和物种栖息地保护(
闫小玲等, 2012 ).了解外来植物的入侵机制是制定科学合理的防控方案的关键.为了解释外来植物的入侵机制, 研究者提出了很多研究假说, 例如天敌逃逸假说(
Keane et al ., 2002 )、竞争能力增强进化假说(
Blossey & N?tzold, 1995 ), 但这些假说主要针对入侵植物本身的进化, 而没有考虑入侵植物与土壤微生物的相互作用.随着研究的不断深入, 越来越多的****意识到植物入侵过程与地下生态存在紧密联系(
de la Pe?a et al ., 2010 ;
Ren & Zhang, 2009 ), 入侵植物与土壤微生物之间的互作在入侵过程中发挥重要作用(
Wardle et al ., 2004 ). ...
近十年中国生物入侵研究进展
1
2012
... 外来植物入侵不仅导致入侵地农林生产经济损失, 而且对生态系统和生物多样性造成了严重的威胁(
鞠瑞亭等, 2012 ).据调查, 我国外来入侵植物多达670余种, 分布范围主要集中在东南沿海地区, 且入侵速度正在逐年加快、数量逐年增多, 若不加以防治, 则直接影响我国经济发展和物种栖息地保护(
闫小玲等, 2012 ).了解外来植物的入侵机制是制定科学合理的防控方案的关键.为了解释外来植物的入侵机制, 研究者提出了很多研究假说, 例如天敌逃逸假说(
Keane et al ., 2002 )、竞争能力增强进化假说(
Blossey & N?tzold, 1995 ), 但这些假说主要针对入侵植物本身的进化, 而没有考虑入侵植物与土壤微生物的相互作用.随着研究的不断深入, 越来越多的****意识到植物入侵过程与地下生态存在紧密联系(
de la Pe?a et al ., 2010 ;
Ren & Zhang, 2009 ), 入侵植物与土壤微生物之间的互作在入侵过程中发挥重要作用(
Wardle et al ., 2004 ). ...
杨树根际高效解钾细菌的分离筛选与鉴定
1
2016
... 本研究发现的加拿大一枝黄花和白茅根际解钾菌的种类和此前报道的研究具有一定的相似性.国际公认的解钾菌菌种仅3种: 环状芽孢杆菌(
Bacillus circulans )、胶质芽孢杆菌(
Bacillus mucilaginosus )、土壤芽孢杆菌(
Bacillus edaphicus ), 但近年来解钾菌的种属鉴定结果越来越丰富, 新增的种属有假单胞菌属(
Pseudomonas )(
吴凡等, 2010 ;
李新新等, 2014 ;
葛红莲和纪秀娥, 2017 )、根瘤菌属(
Rhizobium )、肠杆菌属(
Enterobacteriaceae )(
吴凡等, 2010 )、微杆菌属(
Microbacterium )、固氮菌属(
Azotobacter )(
易浪波等, 2012 ;
李新新等, 2014 )、中华根瘤菌属(
Sinorhizobium )、慢生根瘤菌属(
Mesorhizobium )、屈挠杆菌属(
Flexibacter )(
鞠伟, 2016 ;
李春钢等, 2017 )等.在加拿大一枝黄花和白茅根际土中分离获得的20株解钾菌中, H1-4、B6-7与
吴凡等(2010) 的研究结果一致, H6-5和H1-6、H5-10、H2-14、H2-17与
李新新等(2014) 的研究结果相同, H3-8与
张妙宜等(2016) 筛选出的解钾菌同属, 以上菌株均被证明具有一定的解钾能力.此外, 本研究发现的解钾能力较强但未被报道的菌株有: H1-15, 属于含铜菌属(
Cupriavidus ); H5-18, 属于
Pseudoflavitalea; H2-20, 属于溶杆菌属(
Lysobacter ); H1-21, 属于
Siphonobacter, 这一研究结果有助于进一步丰富解钾菌的种属类别. ...
杨树根际高效解钾细菌的分离筛选与鉴定
1
2016
... 本研究发现的加拿大一枝黄花和白茅根际解钾菌的种类和此前报道的研究具有一定的相似性.国际公认的解钾菌菌种仅3种: 环状芽孢杆菌(
Bacillus circulans )、胶质芽孢杆菌(
Bacillus mucilaginosus )、土壤芽孢杆菌(
Bacillus edaphicus ), 但近年来解钾菌的种属鉴定结果越来越丰富, 新增的种属有假单胞菌属(
Pseudomonas )(
吴凡等, 2010 ;
李新新等, 2014 ;
葛红莲和纪秀娥, 2017 )、根瘤菌属(
Rhizobium )、肠杆菌属(
Enterobacteriaceae )(
吴凡等, 2010 )、微杆菌属(
Microbacterium )、固氮菌属(
Azotobacter )(
易浪波等, 2012 ;
李新新等, 2014 )、中华根瘤菌属(
Sinorhizobium )、慢生根瘤菌属(
Mesorhizobium )、屈挠杆菌属(
Flexibacter )(
鞠伟, 2016 ;
李春钢等, 2017 )等.在加拿大一枝黄花和白茅根际土中分离获得的20株解钾菌中, H1-4、B6-7与
吴凡等(2010) 的研究结果一致, H6-5和H1-6、H5-10、H2-14、H2-17与
李新新等(2014) 的研究结果相同, H3-8与
张妙宜等(2016) 筛选出的解钾菌同属, 以上菌株均被证明具有一定的解钾能力.此外, 本研究发现的解钾能力较强但未被报道的菌株有: H1-15, 属于含铜菌属(
Cupriavidus ); H5-18, 属于
Pseudoflavitalea; H2-20, 属于溶杆菌属(
Lysobacter ); H1-21, 属于
Siphonobacter, 这一研究结果有助于进一步丰富解钾菌的种属类别. ...
Exotic plant invasions and the enemy release hypothesis
1
2002
... 外来植物入侵不仅导致入侵地农林生产经济损失, 而且对生态系统和生物多样性造成了严重的威胁(
鞠瑞亭等, 2012 ).据调查, 我国外来入侵植物多达670余种, 分布范围主要集中在东南沿海地区, 且入侵速度正在逐年加快、数量逐年增多, 若不加以防治, 则直接影响我国经济发展和物种栖息地保护(
闫小玲等, 2012 ).了解外来植物的入侵机制是制定科学合理的防控方案的关键.为了解释外来植物的入侵机制, 研究者提出了很多研究假说, 例如天敌逃逸假说(
Keane et al ., 2002 )、竞争能力增强进化假说(
Blossey & N?tzold, 1995 ), 但这些假说主要针对入侵植物本身的进化, 而没有考虑入侵植物与土壤微生物的相互作用.随着研究的不断深入, 越来越多的****意识到植物入侵过程与地下生态存在紧密联系(
de la Pe?a et al ., 2010 ;
Ren & Zhang, 2009 ), 入侵植物与土壤微生物之间的互作在入侵过程中发挥重要作用(
Wardle et al ., 2004 ). ...
Exotic plant species alter the microbial community structure and function in the soil
1
2002
... 植物与土壤微生物的相互作用在植物入侵过程中起着至关重要的作用(
Bowen et al ., 2017 ).研究表明外来植物通过改变入侵地土壤微生物结构和功能, 进而改善土壤养分供应, 促进植物生长, 增强植物抗逆性, 促使植物成功入侵(
Kourtev et al ., 2002 ;
Wardle et al ., 2004 ;
Nijjer et al ., 2008 ;
Coats et al ., 2014 ).例如, 紫茎泽兰入侵后改变了入侵地土壤微生物的群落结构, 同时提高了入侵地土壤肥力, 进而增强了紫茎泽兰的竞争能力(
牛红榜等, 2007 ).本研究发现加拿大一枝黄花根际土壤中解钾菌数量、群落结构和解钾活性都和伴生植物白茅有着较大的差异, 加拿大一枝黄花的根际解钾菌数量远多于对照, 且其中多数根际解钾菌均具有高效的解钾能力, 同时加拿大一枝黄花茎叶中的钾含量及入侵地土壤中全钾和速效钾的含量均高于对照.这说明, 加拿大一枝黄花的富钾作用可能与其较多的高效解钾菌有关.相关研究也表明加拿大一枝黄花入侵会显著改变土壤微生物群落结构和多样性(
沈荔花等, 2007 ;
陈晨等, 2009 ;
李国庆, 2009 ), 这些转变可能会显著地影响土壤中钾的有效性和供给水平.在其他入侵植物的相关研究中也发现, 入侵植物对土壤养分含量的影响也和其土壤微生物群落的改变紧密相关.例如, 黄顶菊根际耐寒短杆菌能增加土壤磷含量, 对病原菌的抑制作用较对照植物更强(
石青, 2018 ); 破坏草通过促进土壤中解磷菌的生长使其土壤中有效磷含量增加(
杨国庆等, 2014 );
Xanthium italicum 水提液不仅使土壤中细菌和真菌数量明显增多, 同时其土壤速效钾、速效氮等营养成分也显著增多(
邰凤姣等, 2016 ). ...
一种新型解钾菌的筛选及鉴定
1
2017
... 本研究发现的加拿大一枝黄花和白茅根际解钾菌的种类和此前报道的研究具有一定的相似性.国际公认的解钾菌菌种仅3种: 环状芽孢杆菌(
Bacillus circulans )、胶质芽孢杆菌(
Bacillus mucilaginosus )、土壤芽孢杆菌(
Bacillus edaphicus ), 但近年来解钾菌的种属鉴定结果越来越丰富, 新增的种属有假单胞菌属(
Pseudomonas )(
吴凡等, 2010 ;
李新新等, 2014 ;
葛红莲和纪秀娥, 2017 )、根瘤菌属(
Rhizobium )、肠杆菌属(
Enterobacteriaceae )(
吴凡等, 2010 )、微杆菌属(
Microbacterium )、固氮菌属(
Azotobacter )(
易浪波等, 2012 ;
李新新等, 2014 )、中华根瘤菌属(
Sinorhizobium )、慢生根瘤菌属(
Mesorhizobium )、屈挠杆菌属(
Flexibacter )(
鞠伟, 2016 ;
李春钢等, 2017 )等.在加拿大一枝黄花和白茅根际土中分离获得的20株解钾菌中, H1-4、B6-7与
吴凡等(2010) 的研究结果一致, H6-5和H1-6、H5-10、H2-14、H2-17与
李新新等(2014) 的研究结果相同, H3-8与
张妙宜等(2016) 筛选出的解钾菌同属, 以上菌株均被证明具有一定的解钾能力.此外, 本研究发现的解钾能力较强但未被报道的菌株有: H1-15, 属于含铜菌属(
Cupriavidus ); H5-18, 属于
Pseudoflavitalea; H2-20, 属于溶杆菌属(
Lysobacter ); H1-21, 属于
Siphonobacter, 这一研究结果有助于进一步丰富解钾菌的种属类别. ...
一种新型解钾菌的筛选及鉴定
1
2017
... 本研究发现的加拿大一枝黄花和白茅根际解钾菌的种类和此前报道的研究具有一定的相似性.国际公认的解钾菌菌种仅3种: 环状芽孢杆菌(
Bacillus circulans )、胶质芽孢杆菌(
Bacillus mucilaginosus )、土壤芽孢杆菌(
Bacillus edaphicus ), 但近年来解钾菌的种属鉴定结果越来越丰富, 新增的种属有假单胞菌属(
Pseudomonas )(
吴凡等, 2010 ;
李新新等, 2014 ;
葛红莲和纪秀娥, 2017 )、根瘤菌属(
Rhizobium )、肠杆菌属(
Enterobacteriaceae )(
吴凡等, 2010 )、微杆菌属(
Microbacterium )、固氮菌属(
Azotobacter )(
易浪波等, 2012 ;
李新新等, 2014 )、中华根瘤菌属(
Sinorhizobium )、慢生根瘤菌属(
Mesorhizobium )、屈挠杆菌属(
Flexibacter )(
鞠伟, 2016 ;
李春钢等, 2017 )等.在加拿大一枝黄花和白茅根际土中分离获得的20株解钾菌中, H1-4、B6-7与
吴凡等(2010) 的研究结果一致, H6-5和H1-6、H5-10、H2-14、H2-17与
李新新等(2014) 的研究结果相同, H3-8与
张妙宜等(2016) 筛选出的解钾菌同属, 以上菌株均被证明具有一定的解钾能力.此外, 本研究发现的解钾能力较强但未被报道的菌株有: H1-15, 属于含铜菌属(
Cupriavidus ); H5-18, 属于
Pseudoflavitalea; H2-20, 属于溶杆菌属(
Lysobacter ); H1-21, 属于
Siphonobacter, 这一研究结果有助于进一步丰富解钾菌的种属类别. ...
入侵植物加拿大一枝黄花对根际土壤微生物群落多样性的影响研究
1
2009
... 植物与土壤微生物的相互作用在植物入侵过程中起着至关重要的作用(
Bowen et al ., 2017 ).研究表明外来植物通过改变入侵地土壤微生物结构和功能, 进而改善土壤养分供应, 促进植物生长, 增强植物抗逆性, 促使植物成功入侵(
Kourtev et al ., 2002 ;
Wardle et al ., 2004 ;
Nijjer et al ., 2008 ;
Coats et al ., 2014 ).例如, 紫茎泽兰入侵后改变了入侵地土壤微生物的群落结构, 同时提高了入侵地土壤肥力, 进而增强了紫茎泽兰的竞争能力(
牛红榜等, 2007 ).本研究发现加拿大一枝黄花根际土壤中解钾菌数量、群落结构和解钾活性都和伴生植物白茅有着较大的差异, 加拿大一枝黄花的根际解钾菌数量远多于对照, 且其中多数根际解钾菌均具有高效的解钾能力, 同时加拿大一枝黄花茎叶中的钾含量及入侵地土壤中全钾和速效钾的含量均高于对照.这说明, 加拿大一枝黄花的富钾作用可能与其较多的高效解钾菌有关.相关研究也表明加拿大一枝黄花入侵会显著改变土壤微生物群落结构和多样性(
沈荔花等, 2007 ;
陈晨等, 2009 ;
李国庆, 2009 ), 这些转变可能会显著地影响土壤中钾的有效性和供给水平.在其他入侵植物的相关研究中也发现, 入侵植物对土壤养分含量的影响也和其土壤微生物群落的改变紧密相关.例如, 黄顶菊根际耐寒短杆菌能增加土壤磷含量, 对病原菌的抑制作用较对照植物更强(
石青, 2018 ); 破坏草通过促进土壤中解磷菌的生长使其土壤中有效磷含量增加(
杨国庆等, 2014 );
Xanthium italicum 水提液不仅使土壤中细菌和真菌数量明显增多, 同时其土壤速效钾、速效氮等营养成分也显著增多(
邰凤姣等, 2016 ). ...
入侵植物加拿大一枝黄花对根际土壤微生物群落多样性的影响研究
1
2009
... 植物与土壤微生物的相互作用在植物入侵过程中起着至关重要的作用(
Bowen et al ., 2017 ).研究表明外来植物通过改变入侵地土壤微生物结构和功能, 进而改善土壤养分供应, 促进植物生长, 增强植物抗逆性, 促使植物成功入侵(
Kourtev et al ., 2002 ;
Wardle et al ., 2004 ;
Nijjer et al ., 2008 ;
Coats et al ., 2014 ).例如, 紫茎泽兰入侵后改变了入侵地土壤微生物的群落结构, 同时提高了入侵地土壤肥力, 进而增强了紫茎泽兰的竞争能力(
牛红榜等, 2007 ).本研究发现加拿大一枝黄花根际土壤中解钾菌数量、群落结构和解钾活性都和伴生植物白茅有着较大的差异, 加拿大一枝黄花的根际解钾菌数量远多于对照, 且其中多数根际解钾菌均具有高效的解钾能力, 同时加拿大一枝黄花茎叶中的钾含量及入侵地土壤中全钾和速效钾的含量均高于对照.这说明, 加拿大一枝黄花的富钾作用可能与其较多的高效解钾菌有关.相关研究也表明加拿大一枝黄花入侵会显著改变土壤微生物群落结构和多样性(
沈荔花等, 2007 ;
陈晨等, 2009 ;
李国庆, 2009 ), 这些转变可能会显著地影响土壤中钾的有效性和供给水平.在其他入侵植物的相关研究中也发现, 入侵植物对土壤养分含量的影响也和其土壤微生物群落的改变紧密相关.例如, 黄顶菊根际耐寒短杆菌能增加土壤磷含量, 对病原菌的抑制作用较对照植物更强(
石青, 2018 ); 破坏草通过促进土壤中解磷菌的生长使其土壤中有效磷含量增加(
杨国庆等, 2014 );
Xanthium italicum 水提液不仅使土壤中细菌和真菌数量明显增多, 同时其土壤速效钾、速效氮等营养成分也显著增多(
邰凤姣等, 2016 ). ...
一株高效解钾菌的筛选、鉴定及发酵条件的优化
3
2014
... 本研究发现的加拿大一枝黄花和白茅根际解钾菌的种类和此前报道的研究具有一定的相似性.国际公认的解钾菌菌种仅3种: 环状芽孢杆菌(
Bacillus circulans )、胶质芽孢杆菌(
Bacillus mucilaginosus )、土壤芽孢杆菌(
Bacillus edaphicus ), 但近年来解钾菌的种属鉴定结果越来越丰富, 新增的种属有假单胞菌属(
Pseudomonas )(
吴凡等, 2010 ;
李新新等, 2014 ;
葛红莲和纪秀娥, 2017 )、根瘤菌属(
Rhizobium )、肠杆菌属(
Enterobacteriaceae )(
吴凡等, 2010 )、微杆菌属(
Microbacterium )、固氮菌属(
Azotobacter )(
易浪波等, 2012 ;
李新新等, 2014 )、中华根瘤菌属(
Sinorhizobium )、慢生根瘤菌属(
Mesorhizobium )、屈挠杆菌属(
Flexibacter )(
鞠伟, 2016 ;
李春钢等, 2017 )等.在加拿大一枝黄花和白茅根际土中分离获得的20株解钾菌中, H1-4、B6-7与
吴凡等(2010) 的研究结果一致, H6-5和H1-6、H5-10、H2-14、H2-17与
李新新等(2014) 的研究结果相同, H3-8与
张妙宜等(2016) 筛选出的解钾菌同属, 以上菌株均被证明具有一定的解钾能力.此外, 本研究发现的解钾能力较强但未被报道的菌株有: H1-15, 属于含铜菌属(
Cupriavidus ); H5-18, 属于
Pseudoflavitalea; H2-20, 属于溶杆菌属(
Lysobacter ); H1-21, 属于
Siphonobacter, 这一研究结果有助于进一步丰富解钾菌的种属类别. ...
... ;
李新新等, 2014 )、中华根瘤菌属(
Sinorhizobium )、慢生根瘤菌属(
Mesorhizobium )、屈挠杆菌属(
Flexibacter )(
鞠伟, 2016 ;
李春钢等, 2017 )等.在加拿大一枝黄花和白茅根际土中分离获得的20株解钾菌中, H1-4、B6-7与
吴凡等(2010) 的研究结果一致, H6-5和H1-6、H5-10、H2-14、H2-17与
李新新等(2014) 的研究结果相同, H3-8与
张妙宜等(2016) 筛选出的解钾菌同属, 以上菌株均被证明具有一定的解钾能力.此外, 本研究发现的解钾能力较强但未被报道的菌株有: H1-15, 属于含铜菌属(
Cupriavidus ); H5-18, 属于
Pseudoflavitalea; H2-20, 属于溶杆菌属(
Lysobacter ); H1-21, 属于
Siphonobacter, 这一研究结果有助于进一步丰富解钾菌的种属类别. ...
... 的研究结果一致, H6-5和H1-6、H5-10、H2-14、H2-17与
李新新等(2014) 的研究结果相同, H3-8与
张妙宜等(2016) 筛选出的解钾菌同属, 以上菌株均被证明具有一定的解钾能力.此外, 本研究发现的解钾能力较强但未被报道的菌株有: H1-15, 属于含铜菌属(
Cupriavidus ); H5-18, 属于
Pseudoflavitalea; H2-20, 属于溶杆菌属(
Lysobacter ); H1-21, 属于
Siphonobacter, 这一研究结果有助于进一步丰富解钾菌的种属类别. ...
一株高效解钾菌的筛选、鉴定及发酵条件的优化
3
2014
... 本研究发现的加拿大一枝黄花和白茅根际解钾菌的种类和此前报道的研究具有一定的相似性.国际公认的解钾菌菌种仅3种: 环状芽孢杆菌(
Bacillus circulans )、胶质芽孢杆菌(
Bacillus mucilaginosus )、土壤芽孢杆菌(
Bacillus edaphicus ), 但近年来解钾菌的种属鉴定结果越来越丰富, 新增的种属有假单胞菌属(
Pseudomonas )(
吴凡等, 2010 ;
李新新等, 2014 ;
葛红莲和纪秀娥, 2017 )、根瘤菌属(
Rhizobium )、肠杆菌属(
Enterobacteriaceae )(
吴凡等, 2010 )、微杆菌属(
Microbacterium )、固氮菌属(
Azotobacter )(
易浪波等, 2012 ;
李新新等, 2014 )、中华根瘤菌属(
Sinorhizobium )、慢生根瘤菌属(
Mesorhizobium )、屈挠杆菌属(
Flexibacter )(
鞠伟, 2016 ;
李春钢等, 2017 )等.在加拿大一枝黄花和白茅根际土中分离获得的20株解钾菌中, H1-4、B6-7与
吴凡等(2010) 的研究结果一致, H6-5和H1-6、H5-10、H2-14、H2-17与
李新新等(2014) 的研究结果相同, H3-8与
张妙宜等(2016) 筛选出的解钾菌同属, 以上菌株均被证明具有一定的解钾能力.此外, 本研究发现的解钾能力较强但未被报道的菌株有: H1-15, 属于含铜菌属(
Cupriavidus ); H5-18, 属于
Pseudoflavitalea; H2-20, 属于溶杆菌属(
Lysobacter ); H1-21, 属于
Siphonobacter, 这一研究结果有助于进一步丰富解钾菌的种属类别. ...
... ;
李新新等, 2014 )、中华根瘤菌属(
Sinorhizobium )、慢生根瘤菌属(
Mesorhizobium )、屈挠杆菌属(
Flexibacter )(
鞠伟, 2016 ;
李春钢等, 2017 )等.在加拿大一枝黄花和白茅根际土中分离获得的20株解钾菌中, H1-4、B6-7与
吴凡等(2010) 的研究结果一致, H6-5和H1-6、H5-10、H2-14、H2-17与
李新新等(2014) 的研究结果相同, H3-8与
张妙宜等(2016) 筛选出的解钾菌同属, 以上菌株均被证明具有一定的解钾能力.此外, 本研究发现的解钾能力较强但未被报道的菌株有: H1-15, 属于含铜菌属(
Cupriavidus ); H5-18, 属于
Pseudoflavitalea; H2-20, 属于溶杆菌属(
Lysobacter ); H1-21, 属于
Siphonobacter, 这一研究结果有助于进一步丰富解钾菌的种属类别. ...
... 的研究结果一致, H6-5和H1-6、H5-10、H2-14、H2-17与
李新新等(2014) 的研究结果相同, H3-8与
张妙宜等(2016) 筛选出的解钾菌同属, 以上菌株均被证明具有一定的解钾能力.此外, 本研究发现的解钾能力较强但未被报道的菌株有: H1-15, 属于含铜菌属(
Cupriavidus ); H5-18, 属于
Pseudoflavitalea; H2-20, 属于溶杆菌属(
Lysobacter ); H1-21, 属于
Siphonobacter, 这一研究结果有助于进一步丰富解钾菌的种属类别. ...
硅酸盐细菌肥料的特性和作用
1
1994
... 目前国内外对加拿大一枝黄花入侵机制的研究主要集中在繁殖特性(
黄华和郭水良, 2005 ;
郝建华等, 2009 ;
张海亮等, 2015b )、化感作用(
梅玲笑, 2005 ;
郝建华等, 2009 )、生理生态特征与环境适应性(
郭水良和方芳, 2003 ;
杜乐山, 2014 )以及土壤微生物响应(
Zhang et al ., 2005 ;
沈荔花等, 2007 )等方面.研究表明, 在不同程度的入侵地中, 加拿大一枝黄花体内钾含量均高于伴生植物, 尤其在生长期和开花期差异最为显著, 且植物吸钾量与生物量呈正相关关系(
周振荣, 2010 ).在微生物响应方面, 研究发现加拿大一枝黄花入侵后能改变土壤微生物的群落数量、结构和功能多样性, 可能有助于加拿大一枝黄花进一步入侵(
Liao et al ., 2013 ;
Wang et al ., 2018 ).根际解钾菌能将土壤中难溶性钾转化为可被植物直接吸收的有效性钾, 进而促进植物钾吸收, 增强植物抗逆性(
李元芳, 1994 ;
郭勋斌等, 2001 ).加拿大一枝黄花入侵是否改变了根际解钾菌数量、结构和解钾活性等问题还有待深入研究.因此, 本研究以入侵植物加拿大一枝黄花和伴生本地植物白茅(
Imperata cylindrica )为研究对象, 比较加拿大一枝黄花与白茅的根际解钾菌数量、多样性与解钾能力差异, 旨在初步探讨根际解钾菌在加拿大一枝黄花入侵过程中发挥的作用, 为深入研究加拿大一枝黄花入侵机制提供理论依据. ...
硅酸盐细菌肥料的特性和作用
1
1994
... 目前国内外对加拿大一枝黄花入侵机制的研究主要集中在繁殖特性(
黄华和郭水良, 2005 ;
郝建华等, 2009 ;
张海亮等, 2015b )、化感作用(
梅玲笑, 2005 ;
郝建华等, 2009 )、生理生态特征与环境适应性(
郭水良和方芳, 2003 ;
杜乐山, 2014 )以及土壤微生物响应(
Zhang et al ., 2005 ;
沈荔花等, 2007 )等方面.研究表明, 在不同程度的入侵地中, 加拿大一枝黄花体内钾含量均高于伴生植物, 尤其在生长期和开花期差异最为显著, 且植物吸钾量与生物量呈正相关关系(
周振荣, 2010 ).在微生物响应方面, 研究发现加拿大一枝黄花入侵后能改变土壤微生物的群落数量、结构和功能多样性, 可能有助于加拿大一枝黄花进一步入侵(
Liao et al ., 2013 ;
Wang et al ., 2018 ).根际解钾菌能将土壤中难溶性钾转化为可被植物直接吸收的有效性钾, 进而促进植物钾吸收, 增强植物抗逆性(
李元芳, 1994 ;
郭勋斌等, 2001 ).加拿大一枝黄花入侵是否改变了根际解钾菌数量、结构和解钾活性等问题还有待深入研究.因此, 本研究以入侵植物加拿大一枝黄花和伴生本地植物白茅(
Imperata cylindrica )为研究对象, 比较加拿大一枝黄花与白茅的根际解钾菌数量、多样性与解钾能力差异, 旨在初步探讨根际解钾菌在加拿大一枝黄花入侵过程中发挥的作用, 为深入研究加拿大一枝黄花入侵机制提供理论依据. ...
加拿大一枝黄花入侵对杭州湾湿地围垦区土壤养分及活性有机碳组分的影响
2
2016
... 采样点位于杭州湾国家湿地公园生态保育区内(121.17° E, 30.32° N).该区域地处浙江省宁波慈溪市西北部, 属北亚热带季风气候, 地势平坦, 生境类型一致, 远离城市和人为干扰, 植被自然生长, 土壤pH值8-9, 含盐量在0.2%左右, 植物物种多样性相对较低, 主要为单优势草本群落, 典型的有加拿大一枝黄花群落、白茅群落、芦苇(
Phragmites australis )群落等.入侵植物加拿大一枝黄花和伴生本地植物白茅均为多年生草本植物, 具有发达的根状茎, 同期发育, 8月份生长趋于稳定(
梁雷等, 2016 ). ...
... 入侵植物对营养元素的吸收、积累和释放对其生长、入侵能力及其入侵的生态学效应都具有重要作用.钾是植物生长发育所需要的主要营养元素之一, 在植物生长繁育过程中发挥必不可少的作用, 钾的增加不仅能够提高植物光合速率和酶的活化速度、增加植物糖和蛋白质含量、增强植物对氮的吸收, 还能够提高作物的产量、提高植物的抗逆性(
Graham & Ulrich, 1972 ;
胡笃敬等, 1993 ).本研究发现, 野外自然生长的加拿大一枝黄花叶片和茎中的钾含量显著高于伴生植物白茅, 尤其叶片中的钾含量是白茅的7.33倍, 说明在相同生境下, 加拿大一枝黄花吸收钾的能力远高于本地植物, 且加拿大一枝黄花能够将更多的钾分配到叶片中, 而叶片是光合作用最主要的场所, 叶片中养分含量增多可能增强植物光合速率和生长代谢速率(
Aerts, 2000 ;
王丽芳等, 2013 ).此外, 加拿大一枝黄花叶片中的氮磷含量明显高于茎, 更加证实了加拿大一枝黄花将更多养分分配到叶片中的生长策略.人为提高钾供给水平均能促进两种植物生物量积累增加(
图3 ), 但是我们发现加拿大一枝黄花入侵地土壤0-10 cm及10-20 cm土层中全氮、全磷、全钾及碱解氮、有效磷和速效钾含量均多于对照植物白茅, 尤其土壤中速效养分的含量显著多于白茅, 这与
梁雷等(2016) 的研究结果相一致.
陈彤等(2012) 发现加拿大一枝黄花能显著提高土壤中硝态氮含量, 并且其较高的氮积累能力可能和入侵后本地植物种群的动态变化格局紧密相关 这些研究说明加拿大一枝黄花存在一种能够增强土壤养分积累的机制, 而这种机制反过来可能促进其进一步入侵.外来植物对入侵地土壤养分含量的影响被大量研究.一些研究认为外来植物入侵往往能够通过影响入侵地土壤养分而促进自身成功入侵(
Holmgren et al ., 1997 ).许多外来植物入侵后通过改变土壤肥力影响其与本地种的竞争能力, 例如破坏草、黄顶菊(
Flaveria bidentis )、盐生草(
Halogeton glomeratus )等植物入侵时均会导致土壤养分含量向有利于入侵植物的方向发生改变(
Duda et al ., 2003 ;
蒋智林等, 2008 ;
张天瑞等, 2010 ). ...
加拿大一枝黄花入侵对杭州湾湿地围垦区土壤养分及活性有机碳组分的影响
2
2016
... 采样点位于杭州湾国家湿地公园生态保育区内(121.17° E, 30.32° N).该区域地处浙江省宁波慈溪市西北部, 属北亚热带季风气候, 地势平坦, 生境类型一致, 远离城市和人为干扰, 植被自然生长, 土壤pH值8-9, 含盐量在0.2%左右, 植物物种多样性相对较低, 主要为单优势草本群落, 典型的有加拿大一枝黄花群落、白茅群落、芦苇(
Phragmites australis )群落等.入侵植物加拿大一枝黄花和伴生本地植物白茅均为多年生草本植物, 具有发达的根状茎, 同期发育, 8月份生长趋于稳定(
梁雷等, 2016 ). ...
... 入侵植物对营养元素的吸收、积累和释放对其生长、入侵能力及其入侵的生态学效应都具有重要作用.钾是植物生长发育所需要的主要营养元素之一, 在植物生长繁育过程中发挥必不可少的作用, 钾的增加不仅能够提高植物光合速率和酶的活化速度、增加植物糖和蛋白质含量、增强植物对氮的吸收, 还能够提高作物的产量、提高植物的抗逆性(
Graham & Ulrich, 1972 ;
胡笃敬等, 1993 ).本研究发现, 野外自然生长的加拿大一枝黄花叶片和茎中的钾含量显著高于伴生植物白茅, 尤其叶片中的钾含量是白茅的7.33倍, 说明在相同生境下, 加拿大一枝黄花吸收钾的能力远高于本地植物, 且加拿大一枝黄花能够将更多的钾分配到叶片中, 而叶片是光合作用最主要的场所, 叶片中养分含量增多可能增强植物光合速率和生长代谢速率(
Aerts, 2000 ;
王丽芳等, 2013 ).此外, 加拿大一枝黄花叶片中的氮磷含量明显高于茎, 更加证实了加拿大一枝黄花将更多养分分配到叶片中的生长策略.人为提高钾供给水平均能促进两种植物生物量积累增加(
图3 ), 但是我们发现加拿大一枝黄花入侵地土壤0-10 cm及10-20 cm土层中全氮、全磷、全钾及碱解氮、有效磷和速效钾含量均多于对照植物白茅, 尤其土壤中速效养分的含量显著多于白茅, 这与
梁雷等(2016) 的研究结果相一致.
陈彤等(2012) 发现加拿大一枝黄花能显著提高土壤中硝态氮含量, 并且其较高的氮积累能力可能和入侵后本地植物种群的动态变化格局紧密相关 这些研究说明加拿大一枝黄花存在一种能够增强土壤养分积累的机制, 而这种机制反过来可能促进其进一步入侵.外来植物对入侵地土壤养分含量的影响被大量研究.一些研究认为外来植物入侵往往能够通过影响入侵地土壤养分而促进自身成功入侵(
Holmgren et al ., 1997 ).许多外来植物入侵后通过改变土壤肥力影响其与本地种的竞争能力, 例如破坏草、黄顶菊(
Flaveria bidentis )、盐生草(
Halogeton glomeratus )等植物入侵时均会导致土壤养分含量向有利于入侵植物的方向发生改变(
Duda et al ., 2003 ;
蒋智林等, 2008 ;
张天瑞等, 2010 ). ...
互花米草入侵对红树秋茄根际与根内细菌群落结构与多样性的影响
1
2018
... 外来植物在入侵过程中可能通过释放根系分泌物和改变微生物碳源等方式影响土壤微生物结构和功能, 而土壤微生物群落的改变反过来会影响外来植物的进一步入侵(
杨星等, 2012 ;
Uddin et al ., 2014 ;
Zhao et al ., 2014 ;
Martin et al ., 2017 ).以往研究表明, 紫茉莉(
Mirabilis jalapa )、破坏草(紫茎泽兰)(
Ageratina adenophora )、互花米草(
Spartina alterniflora )可以改变入侵地土壤微生物多样性(
牛红榜, 2007 ;
Zhao et al ., 2014 ;
廖丹等, 2018 ); 银荆(
Acacia dealbata )入侵西班牙北部地区后使当地土壤中细菌丰富度显著增加, 真菌的丰富度和多样性显著减少(
Lorenzo et al ., 2010 ); 旱雀麦(
Bromus tectorum )使入侵地的细菌数量显著增多, 真菌数量显著减少(
Belnap & Phillips, 2001 ); 葱芥(
Alliaria petiolata )入侵后导致土壤中丛枝菌根真菌数量显著减少(
Roberts & Anderson, 2001 ).这些结果说明, 外来植物入侵能够导致土壤微生物多样性发生明显改变.而另一方面, 土壤微生物的改变可能促进外来植物的入侵, 甚至可能抑制本地植物的生长.例如, 土壤微生物显著提高了入侵植物加拿大一枝黄花(
Solidago canadensis )的出芽率和资源竞争力, 而降低了本土植物长芒草(
Stipa bungeana )的竞争力(
Sun & He, 2010 ).因此, 探究入侵植物对土壤微生物的影响可能是了解植物入侵机理的关键所在. ...
互花米草入侵对红树秋茄根际与根内细菌群落结构与多样性的影响
1
2018
... 外来植物在入侵过程中可能通过释放根系分泌物和改变微生物碳源等方式影响土壤微生物结构和功能, 而土壤微生物群落的改变反过来会影响外来植物的进一步入侵(
杨星等, 2012 ;
Uddin et al ., 2014 ;
Zhao et al ., 2014 ;
Martin et al ., 2017 ).以往研究表明, 紫茉莉(
Mirabilis jalapa )、破坏草(紫茎泽兰)(
Ageratina adenophora )、互花米草(
Spartina alterniflora )可以改变入侵地土壤微生物多样性(
牛红榜, 2007 ;
Zhao et al ., 2014 ;
廖丹等, 2018 ); 银荆(
Acacia dealbata )入侵西班牙北部地区后使当地土壤中细菌丰富度显著增加, 真菌的丰富度和多样性显著减少(
Lorenzo et al ., 2010 ); 旱雀麦(
Bromus tectorum )使入侵地的细菌数量显著增多, 真菌数量显著减少(
Belnap & Phillips, 2001 ); 葱芥(
Alliaria petiolata )入侵后导致土壤中丛枝菌根真菌数量显著减少(
Roberts & Anderson, 2001 ).这些结果说明, 外来植物入侵能够导致土壤微生物多样性发生明显改变.而另一方面, 土壤微生物的改变可能促进外来植物的入侵, 甚至可能抑制本地植物的生长.例如, 土壤微生物显著提高了入侵植物加拿大一枝黄花(
Solidago canadensis )的出芽率和资源竞争力, 而降低了本土植物长芒草(
Stipa bungeana )的竞争力(
Sun & He, 2010 ).因此, 探究入侵植物对土壤微生物的影响可能是了解植物入侵机理的关键所在. ...
Characteristics of soil microbial community functional and structure diversity with coverage of
Solidago canadensis L
1
2013
... 目前国内外对加拿大一枝黄花入侵机制的研究主要集中在繁殖特性(
黄华和郭水良, 2005 ;
郝建华等, 2009 ;
张海亮等, 2015b )、化感作用(
梅玲笑, 2005 ;
郝建华等, 2009 )、生理生态特征与环境适应性(
郭水良和方芳, 2003 ;
杜乐山, 2014 )以及土壤微生物响应(
Zhang et al ., 2005 ;
沈荔花等, 2007 )等方面.研究表明, 在不同程度的入侵地中, 加拿大一枝黄花体内钾含量均高于伴生植物, 尤其在生长期和开花期差异最为显著, 且植物吸钾量与生物量呈正相关关系(
周振荣, 2010 ).在微生物响应方面, 研究发现加拿大一枝黄花入侵后能改变土壤微生物的群落数量、结构和功能多样性, 可能有助于加拿大一枝黄花进一步入侵(
Liao et al ., 2013 ;
Wang et al ., 2018 ).根际解钾菌能将土壤中难溶性钾转化为可被植物直接吸收的有效性钾, 进而促进植物钾吸收, 增强植物抗逆性(
李元芳, 1994 ;
郭勋斌等, 2001 ).加拿大一枝黄花入侵是否改变了根际解钾菌数量、结构和解钾活性等问题还有待深入研究.因此, 本研究以入侵植物加拿大一枝黄花和伴生本地植物白茅(
Imperata cylindrica )为研究对象, 比较加拿大一枝黄花与白茅的根际解钾菌数量、多样性与解钾能力差异, 旨在初步探讨根际解钾菌在加拿大一枝黄花入侵过程中发挥的作用, 为深入研究加拿大一枝黄花入侵机制提供理论依据. ...
1
2010
... 无菌条件下, 取1 g植物根际土, 溶于装有99 mL无菌水的三角瓶中, 配制成体积质量为1 × 10
-2 g·mL
-1 的土壤悬浮液, 于28 ℃、120 r·min
-1 条件下振荡培养30 min, 使微生物均匀分散.将土壤悬浮液依次稀释为1 × 10
-3 、1 × 10
-4 、1 × 10
-5 g·mL
-1 3个不同体积质量, 取0.1 mL涂布于解钾培养基(配方见
表1 )表面, 每个体积质量重复3次, 接种后将培养皿倒置, 置于28 ℃恒温环境下培养3天后进行计数(
林先贵, 2010 ). ...
1
2010
... 无菌条件下, 取1 g植物根际土, 溶于装有99 mL无菌水的三角瓶中, 配制成体积质量为1 × 10
-2 g·mL
-1 的土壤悬浮液, 于28 ℃、120 r·min
-1 条件下振荡培养30 min, 使微生物均匀分散.将土壤悬浮液依次稀释为1 × 10
-3 、1 × 10
-4 、1 × 10
-5 g·mL
-1 3个不同体积质量, 取0.1 mL涂布于解钾培养基(配方见
表1 )表面, 每个体积质量重复3次, 接种后将培养皿倒置, 置于28 ℃恒温环境下培养3天后进行计数(
林先贵, 2010 ). ...
Effect of invasive
Acacia dealbata Link. on soil microorganisms as determined by PCR-DGGE
1
2010
... 外来植物在入侵过程中可能通过释放根系分泌物和改变微生物碳源等方式影响土壤微生物结构和功能, 而土壤微生物群落的改变反过来会影响外来植物的进一步入侵(
杨星等, 2012 ;
Uddin et al ., 2014 ;
Zhao et al ., 2014 ;
Martin et al ., 2017 ).以往研究表明, 紫茉莉(
Mirabilis jalapa )、破坏草(紫茎泽兰)(
Ageratina adenophora )、互花米草(
Spartina alterniflora )可以改变入侵地土壤微生物多样性(
牛红榜, 2007 ;
Zhao et al ., 2014 ;
廖丹等, 2018 ); 银荆(
Acacia dealbata )入侵西班牙北部地区后使当地土壤中细菌丰富度显著增加, 真菌的丰富度和多样性显著减少(
Lorenzo et al ., 2010 ); 旱雀麦(
Bromus tectorum )使入侵地的细菌数量显著增多, 真菌数量显著减少(
Belnap & Phillips, 2001 ); 葱芥(
Alliaria petiolata )入侵后导致土壤中丛枝菌根真菌数量显著减少(
Roberts & Anderson, 2001 ).这些结果说明, 外来植物入侵能够导致土壤微生物多样性发生明显改变.而另一方面, 土壤微生物的改变可能促进外来植物的入侵, 甚至可能抑制本地植物的生长.例如, 土壤微生物显著提高了入侵植物加拿大一枝黄花(
Solidago canadensis )的出芽率和资源竞争力, 而降低了本土植物长芒草(
Stipa bungeana )的竞争力(
Sun & He, 2010 ).因此, 探究入侵植物对土壤微生物的影响可能是了解植物入侵机理的关键所在. ...
加拿大一枝黄花利害分析
1
2006
... 加拿大一枝黄花是菊科一枝黄花属植物, 多年生草本, 具有发达根状茎, 原产于北美地区, 最初作为庭园花卉引入我国上海、南京等地, 现已广泛逸生为恶性外来种(
中国科学院中国植物志编辑委员会, 1985 ;
郭水良和方芳, 2003 ).加拿大一枝黄花繁殖能力极强, 既能利用种子进行有性繁殖, 也可通过地下根状茎进行无性繁殖, 一般情况下, 每株植株可产约7万粒种子, 且种子具有较高萌发率, 此外, 植株根部具有上百个根茎结构, 每个根茎在第2年可萌发形成独立的个体, 如此高的繁殖能力使其在入侵后能够迅速扩散形成单优势种群落(
王玉良等, 2009 ;
张海亮等, 2015a ).此外, 加拿大一枝黄花根部还会产生一些化感物质影响周围土著植物生长, 对入侵地生物多样性造成了一定的影响(
方芳等, 2004 ).而目前加拿大一枝黄花在我国入侵范围逐渐扩大, 入侵速度逐渐加快, 对我国生态环境、生物多样性、农业和畜牧业均造成了不同程度的影响(
董梅等, 2006 ;
陆慧明等, 2006 ), 因此亟须加强加拿大一枝黄花入侵机理与防控技术方面的研究. ...
加拿大一枝黄花利害分析
1
2006
... 加拿大一枝黄花是菊科一枝黄花属植物, 多年生草本, 具有发达根状茎, 原产于北美地区, 最初作为庭园花卉引入我国上海、南京等地, 现已广泛逸生为恶性外来种(
中国科学院中国植物志编辑委员会, 1985 ;
郭水良和方芳, 2003 ).加拿大一枝黄花繁殖能力极强, 既能利用种子进行有性繁殖, 也可通过地下根状茎进行无性繁殖, 一般情况下, 每株植株可产约7万粒种子, 且种子具有较高萌发率, 此外, 植株根部具有上百个根茎结构, 每个根茎在第2年可萌发形成独立的个体, 如此高的繁殖能力使其在入侵后能够迅速扩散形成单优势种群落(
王玉良等, 2009 ;
张海亮等, 2015a ).此外, 加拿大一枝黄花根部还会产生一些化感物质影响周围土著植物生长, 对入侵地生物多样性造成了一定的影响(
方芳等, 2004 ).而目前加拿大一枝黄花在我国入侵范围逐渐扩大, 入侵速度逐渐加快, 对我国生态环境、生物多样性、农业和畜牧业均造成了不同程度的影响(
董梅等, 2006 ;
陆慧明等, 2006 ), 因此亟须加强加拿大一枝黄花入侵机理与防控技术方面的研究. ...
Impacts of invasive plants on carbon pools depend on both species’ traits and local climate
1
2017
... 外来植物在入侵过程中可能通过释放根系分泌物和改变微生物碳源等方式影响土壤微生物结构和功能, 而土壤微生物群落的改变反过来会影响外来植物的进一步入侵(
杨星等, 2012 ;
Uddin et al ., 2014 ;
Zhao et al ., 2014 ;
Martin et al ., 2017 ).以往研究表明, 紫茉莉(
Mirabilis jalapa )、破坏草(紫茎泽兰)(
Ageratina adenophora )、互花米草(
Spartina alterniflora )可以改变入侵地土壤微生物多样性(
牛红榜, 2007 ;
Zhao et al ., 2014 ;
廖丹等, 2018 ); 银荆(
Acacia dealbata )入侵西班牙北部地区后使当地土壤中细菌丰富度显著增加, 真菌的丰富度和多样性显著减少(
Lorenzo et al ., 2010 ); 旱雀麦(
Bromus tectorum )使入侵地的细菌数量显著增多, 真菌数量显著减少(
Belnap & Phillips, 2001 ); 葱芥(
Alliaria petiolata )入侵后导致土壤中丛枝菌根真菌数量显著减少(
Roberts & Anderson, 2001 ).这些结果说明, 外来植物入侵能够导致土壤微生物多样性发生明显改变.而另一方面, 土壤微生物的改变可能促进外来植物的入侵, 甚至可能抑制本地植物的生长.例如, 土壤微生物显著提高了入侵植物加拿大一枝黄花(
Solidago canadensis )的出芽率和资源竞争力, 而降低了本土植物长芒草(
Stipa bungeana )的竞争力(
Sun & He, 2010 ).因此, 探究入侵植物对土壤微生物的影响可能是了解植物入侵机理的关键所在. ...
外来杂草加拿大一枝黄花对入侵地植物的化感效应
1
2005
... 目前国内外对加拿大一枝黄花入侵机制的研究主要集中在繁殖特性(
黄华和郭水良, 2005 ;
郝建华等, 2009 ;
张海亮等, 2015b )、化感作用(
梅玲笑, 2005 ;
郝建华等, 2009 )、生理生态特征与环境适应性(
郭水良和方芳, 2003 ;
杜乐山, 2014 )以及土壤微生物响应(
Zhang et al ., 2005 ;
沈荔花等, 2007 )等方面.研究表明, 在不同程度的入侵地中, 加拿大一枝黄花体内钾含量均高于伴生植物, 尤其在生长期和开花期差异最为显著, 且植物吸钾量与生物量呈正相关关系(
周振荣, 2010 ).在微生物响应方面, 研究发现加拿大一枝黄花入侵后能改变土壤微生物的群落数量、结构和功能多样性, 可能有助于加拿大一枝黄花进一步入侵(
Liao et al ., 2013 ;
Wang et al ., 2018 ).根际解钾菌能将土壤中难溶性钾转化为可被植物直接吸收的有效性钾, 进而促进植物钾吸收, 增强植物抗逆性(
李元芳, 1994 ;
郭勋斌等, 2001 ).加拿大一枝黄花入侵是否改变了根际解钾菌数量、结构和解钾活性等问题还有待深入研究.因此, 本研究以入侵植物加拿大一枝黄花和伴生本地植物白茅(
Imperata cylindrica )为研究对象, 比较加拿大一枝黄花与白茅的根际解钾菌数量、多样性与解钾能力差异, 旨在初步探讨根际解钾菌在加拿大一枝黄花入侵过程中发挥的作用, 为深入研究加拿大一枝黄花入侵机制提供理论依据. ...
外来杂草加拿大一枝黄花对入侵地植物的化感效应
1
2005
... 目前国内外对加拿大一枝黄花入侵机制的研究主要集中在繁殖特性(
黄华和郭水良, 2005 ;
郝建华等, 2009 ;
张海亮等, 2015b )、化感作用(
梅玲笑, 2005 ;
郝建华等, 2009 )、生理生态特征与环境适应性(
郭水良和方芳, 2003 ;
杜乐山, 2014 )以及土壤微生物响应(
Zhang et al ., 2005 ;
沈荔花等, 2007 )等方面.研究表明, 在不同程度的入侵地中, 加拿大一枝黄花体内钾含量均高于伴生植物, 尤其在生长期和开花期差异最为显著, 且植物吸钾量与生物量呈正相关关系(
周振荣, 2010 ).在微生物响应方面, 研究发现加拿大一枝黄花入侵后能改变土壤微生物的群落数量、结构和功能多样性, 可能有助于加拿大一枝黄花进一步入侵(
Liao et al ., 2013 ;
Wang et al ., 2018 ).根际解钾菌能将土壤中难溶性钾转化为可被植物直接吸收的有效性钾, 进而促进植物钾吸收, 增强植物抗逆性(
李元芳, 1994 ;
郭勋斌等, 2001 ).加拿大一枝黄花入侵是否改变了根际解钾菌数量、结构和解钾活性等问题还有待深入研究.因此, 本研究以入侵植物加拿大一枝黄花和伴生本地植物白茅(
Imperata cylindrica )为研究对象, 比较加拿大一枝黄花与白茅的根际解钾菌数量、多样性与解钾能力差异, 旨在初步探讨根际解钾菌在加拿大一枝黄花入侵过程中发挥的作用, 为深入研究加拿大一枝黄花入侵机制提供理论依据. ...
The effects of soil biota and fertilization on the success of
Sapium sebiferum 1
2008
... 植物与土壤微生物的相互作用在植物入侵过程中起着至关重要的作用(
Bowen et al ., 2017 ).研究表明外来植物通过改变入侵地土壤微生物结构和功能, 进而改善土壤养分供应, 促进植物生长, 增强植物抗逆性, 促使植物成功入侵(
Kourtev et al ., 2002 ;
Wardle et al ., 2004 ;
Nijjer et al ., 2008 ;
Coats et al ., 2014 ).例如, 紫茎泽兰入侵后改变了入侵地土壤微生物的群落结构, 同时提高了入侵地土壤肥力, 进而增强了紫茎泽兰的竞争能力(
牛红榜等, 2007 ).本研究发现加拿大一枝黄花根际土壤中解钾菌数量、群落结构和解钾活性都和伴生植物白茅有着较大的差异, 加拿大一枝黄花的根际解钾菌数量远多于对照, 且其中多数根际解钾菌均具有高效的解钾能力, 同时加拿大一枝黄花茎叶中的钾含量及入侵地土壤中全钾和速效钾的含量均高于对照.这说明, 加拿大一枝黄花的富钾作用可能与其较多的高效解钾菌有关.相关研究也表明加拿大一枝黄花入侵会显著改变土壤微生物群落结构和多样性(
沈荔花等, 2007 ;
陈晨等, 2009 ;
李国庆, 2009 ), 这些转变可能会显著地影响土壤中钾的有效性和供给水平.在其他入侵植物的相关研究中也发现, 入侵植物对土壤养分含量的影响也和其土壤微生物群落的改变紧密相关.例如, 黄顶菊根际耐寒短杆菌能增加土壤磷含量, 对病原菌的抑制作用较对照植物更强(
石青, 2018 ); 破坏草通过促进土壤中解磷菌的生长使其土壤中有效磷含量增加(
杨国庆等, 2014 );
Xanthium italicum 水提液不仅使土壤中细菌和真菌数量明显增多, 同时其土壤速效钾、速效氮等营养成分也显著增多(
邰凤姣等, 2016 ). ...
紫茎泽兰(
Ageratina adenophora )入侵对土壤微生物群落和理化性质的影响
2
2007
... 外来植物在入侵过程中可能通过释放根系分泌物和改变微生物碳源等方式影响土壤微生物结构和功能, 而土壤微生物群落的改变反过来会影响外来植物的进一步入侵(
杨星等, 2012 ;
Uddin et al ., 2014 ;
Zhao et al ., 2014 ;
Martin et al ., 2017 ).以往研究表明, 紫茉莉(
Mirabilis jalapa )、破坏草(紫茎泽兰)(
Ageratina adenophora )、互花米草(
Spartina alterniflora )可以改变入侵地土壤微生物多样性(
牛红榜, 2007 ;
Zhao et al ., 2014 ;
廖丹等, 2018 ); 银荆(
Acacia dealbata )入侵西班牙北部地区后使当地土壤中细菌丰富度显著增加, 真菌的丰富度和多样性显著减少(
Lorenzo et al ., 2010 ); 旱雀麦(
Bromus tectorum )使入侵地的细菌数量显著增多, 真菌数量显著减少(
Belnap & Phillips, 2001 ); 葱芥(
Alliaria petiolata )入侵后导致土壤中丛枝菌根真菌数量显著减少(
Roberts & Anderson, 2001 ).这些结果说明, 外来植物入侵能够导致土壤微生物多样性发生明显改变.而另一方面, 土壤微生物的改变可能促进外来植物的入侵, 甚至可能抑制本地植物的生长.例如, 土壤微生物显著提高了入侵植物加拿大一枝黄花(
Solidago canadensis )的出芽率和资源竞争力, 而降低了本土植物长芒草(
Stipa bungeana )的竞争力(
Sun & He, 2010 ).因此, 探究入侵植物对土壤微生物的影响可能是了解植物入侵机理的关键所在. ...
... 植物与土壤微生物的相互作用在植物入侵过程中起着至关重要的作用(
Bowen et al ., 2017 ).研究表明外来植物通过改变入侵地土壤微生物结构和功能, 进而改善土壤养分供应, 促进植物生长, 增强植物抗逆性, 促使植物成功入侵(
Kourtev et al ., 2002 ;
Wardle et al ., 2004 ;
Nijjer et al ., 2008 ;
Coats et al ., 2014 ).例如, 紫茎泽兰入侵后改变了入侵地土壤微生物的群落结构, 同时提高了入侵地土壤肥力, 进而增强了紫茎泽兰的竞争能力(
牛红榜等, 2007 ).本研究发现加拿大一枝黄花根际土壤中解钾菌数量、群落结构和解钾活性都和伴生植物白茅有着较大的差异, 加拿大一枝黄花的根际解钾菌数量远多于对照, 且其中多数根际解钾菌均具有高效的解钾能力, 同时加拿大一枝黄花茎叶中的钾含量及入侵地土壤中全钾和速效钾的含量均高于对照.这说明, 加拿大一枝黄花的富钾作用可能与其较多的高效解钾菌有关.相关研究也表明加拿大一枝黄花入侵会显著改变土壤微生物群落结构和多样性(
沈荔花等, 2007 ;
陈晨等, 2009 ;
李国庆, 2009 ), 这些转变可能会显著地影响土壤中钾的有效性和供给水平.在其他入侵植物的相关研究中也发现, 入侵植物对土壤养分含量的影响也和其土壤微生物群落的改变紧密相关.例如, 黄顶菊根际耐寒短杆菌能增加土壤磷含量, 对病原菌的抑制作用较对照植物更强(
石青, 2018 ); 破坏草通过促进土壤中解磷菌的生长使其土壤中有效磷含量增加(
杨国庆等, 2014 );
Xanthium italicum 水提液不仅使土壤中细菌和真菌数量明显增多, 同时其土壤速效钾、速效氮等营养成分也显著增多(
邰凤姣等, 2016 ). ...
紫茎泽兰(
Ageratina adenophora )入侵对土壤微生物群落和理化性质的影响
2
2007
... 外来植物在入侵过程中可能通过释放根系分泌物和改变微生物碳源等方式影响土壤微生物结构和功能, 而土壤微生物群落的改变反过来会影响外来植物的进一步入侵(
杨星等, 2012 ;
Uddin et al ., 2014 ;
Zhao et al ., 2014 ;
Martin et al ., 2017 ).以往研究表明, 紫茉莉(
Mirabilis jalapa )、破坏草(紫茎泽兰)(
Ageratina adenophora )、互花米草(
Spartina alterniflora )可以改变入侵地土壤微生物多样性(
牛红榜, 2007 ;
Zhao et al ., 2014 ;
廖丹等, 2018 ); 银荆(
Acacia dealbata )入侵西班牙北部地区后使当地土壤中细菌丰富度显著增加, 真菌的丰富度和多样性显著减少(
Lorenzo et al ., 2010 ); 旱雀麦(
Bromus tectorum )使入侵地的细菌数量显著增多, 真菌数量显著减少(
Belnap & Phillips, 2001 ); 葱芥(
Alliaria petiolata )入侵后导致土壤中丛枝菌根真菌数量显著减少(
Roberts & Anderson, 2001 ).这些结果说明, 外来植物入侵能够导致土壤微生物多样性发生明显改变.而另一方面, 土壤微生物的改变可能促进外来植物的入侵, 甚至可能抑制本地植物的生长.例如, 土壤微生物显著提高了入侵植物加拿大一枝黄花(
Solidago canadensis )的出芽率和资源竞争力, 而降低了本土植物长芒草(
Stipa bungeana )的竞争力(
Sun & He, 2010 ).因此, 探究入侵植物对土壤微生物的影响可能是了解植物入侵机理的关键所在. ...
... 植物与土壤微生物的相互作用在植物入侵过程中起着至关重要的作用(
Bowen et al ., 2017 ).研究表明外来植物通过改变入侵地土壤微生物结构和功能, 进而改善土壤养分供应, 促进植物生长, 增强植物抗逆性, 促使植物成功入侵(
Kourtev et al ., 2002 ;
Wardle et al ., 2004 ;
Nijjer et al ., 2008 ;
Coats et al ., 2014 ).例如, 紫茎泽兰入侵后改变了入侵地土壤微生物的群落结构, 同时提高了入侵地土壤肥力, 进而增强了紫茎泽兰的竞争能力(
牛红榜等, 2007 ).本研究发现加拿大一枝黄花根际土壤中解钾菌数量、群落结构和解钾活性都和伴生植物白茅有着较大的差异, 加拿大一枝黄花的根际解钾菌数量远多于对照, 且其中多数根际解钾菌均具有高效的解钾能力, 同时加拿大一枝黄花茎叶中的钾含量及入侵地土壤中全钾和速效钾的含量均高于对照.这说明, 加拿大一枝黄花的富钾作用可能与其较多的高效解钾菌有关.相关研究也表明加拿大一枝黄花入侵会显著改变土壤微生物群落结构和多样性(
沈荔花等, 2007 ;
陈晨等, 2009 ;
李国庆, 2009 ), 这些转变可能会显著地影响土壤中钾的有效性和供给水平.在其他入侵植物的相关研究中也发现, 入侵植物对土壤养分含量的影响也和其土壤微生物群落的改变紧密相关.例如, 黄顶菊根际耐寒短杆菌能增加土壤磷含量, 对病原菌的抑制作用较对照植物更强(
石青, 2018 ); 破坏草通过促进土壤中解磷菌的生长使其土壤中有效磷含量增加(
杨国庆等, 2014 );
Xanthium italicum 水提液不仅使土壤中细菌和真菌数量明显增多, 同时其土壤速效钾、速效氮等营养成分也显著增多(
邰凤姣等, 2016 ). ...
The relative generality of plant invasion mechanisms and predicting future invasive plants
1
2009
... 外来植物入侵不仅导致入侵地农林生产经济损失, 而且对生态系统和生物多样性造成了严重的威胁(
鞠瑞亭等, 2012 ).据调查, 我国外来入侵植物多达670余种, 分布范围主要集中在东南沿海地区, 且入侵速度正在逐年加快、数量逐年增多, 若不加以防治, 则直接影响我国经济发展和物种栖息地保护(
闫小玲等, 2012 ).了解外来植物的入侵机制是制定科学合理的防控方案的关键.为了解释外来植物的入侵机制, 研究者提出了很多研究假说, 例如天敌逃逸假说(
Keane et al ., 2002 )、竞争能力增强进化假说(
Blossey & N?tzold, 1995 ), 但这些假说主要针对入侵植物本身的进化, 而没有考虑入侵植物与土壤微生物的相互作用.随着研究的不断深入, 越来越多的****意识到植物入侵过程与地下生态存在紧密联系(
de la Pe?a et al ., 2010 ;
Ren & Zhang, 2009 ), 入侵植物与土壤微生物之间的互作在入侵过程中发挥重要作用(
Wardle et al ., 2004 ). ...
Effect of garlic mustard [ Alliaria petiolata(Beib. Cavara & Grande)] extracts on plants and arbuscular mycorrhizal (AM) fungi
1
2001
... 外来植物在入侵过程中可能通过释放根系分泌物和改变微生物碳源等方式影响土壤微生物结构和功能, 而土壤微生物群落的改变反过来会影响外来植物的进一步入侵(
杨星等, 2012 ;
Uddin et al ., 2014 ;
Zhao et al ., 2014 ;
Martin et al ., 2017 ).以往研究表明, 紫茉莉(
Mirabilis jalapa )、破坏草(紫茎泽兰)(
Ageratina adenophora )、互花米草(
Spartina alterniflora )可以改变入侵地土壤微生物多样性(
牛红榜, 2007 ;
Zhao et al ., 2014 ;
廖丹等, 2018 ); 银荆(
Acacia dealbata )入侵西班牙北部地区后使当地土壤中细菌丰富度显著增加, 真菌的丰富度和多样性显著减少(
Lorenzo et al ., 2010 ); 旱雀麦(
Bromus tectorum )使入侵地的细菌数量显著增多, 真菌数量显著减少(
Belnap & Phillips, 2001 ); 葱芥(
Alliaria petiolata )入侵后导致土壤中丛枝菌根真菌数量显著减少(
Roberts & Anderson, 2001 ).这些结果说明, 外来植物入侵能够导致土壤微生物多样性发生明显改变.而另一方面, 土壤微生物的改变可能促进外来植物的入侵, 甚至可能抑制本地植物的生长.例如, 土壤微生物显著提高了入侵植物加拿大一枝黄花(
Solidago canadensis )的出芽率和资源竞争力, 而降低了本土植物长芒草(
Stipa bungeana )的竞争力(
Sun & He, 2010 ).因此, 探究入侵植物对土壤微生物的影响可能是了解植物入侵机理的关键所在. ...
加拿大一枝黄花对土壤微生物区系的影响研究
3
2007
... 目前国内外对加拿大一枝黄花入侵机制的研究主要集中在繁殖特性(
黄华和郭水良, 2005 ;
郝建华等, 2009 ;
张海亮等, 2015b )、化感作用(
梅玲笑, 2005 ;
郝建华等, 2009 )、生理生态特征与环境适应性(
郭水良和方芳, 2003 ;
杜乐山, 2014 )以及土壤微生物响应(
Zhang et al ., 2005 ;
沈荔花等, 2007 )等方面.研究表明, 在不同程度的入侵地中, 加拿大一枝黄花体内钾含量均高于伴生植物, 尤其在生长期和开花期差异最为显著, 且植物吸钾量与生物量呈正相关关系(
周振荣, 2010 ).在微生物响应方面, 研究发现加拿大一枝黄花入侵后能改变土壤微生物的群落数量、结构和功能多样性, 可能有助于加拿大一枝黄花进一步入侵(
Liao et al ., 2013 ;
Wang et al ., 2018 ).根际解钾菌能将土壤中难溶性钾转化为可被植物直接吸收的有效性钾, 进而促进植物钾吸收, 增强植物抗逆性(
李元芳, 1994 ;
郭勋斌等, 2001 ).加拿大一枝黄花入侵是否改变了根际解钾菌数量、结构和解钾活性等问题还有待深入研究.因此, 本研究以入侵植物加拿大一枝黄花和伴生本地植物白茅(
Imperata cylindrica )为研究对象, 比较加拿大一枝黄花与白茅的根际解钾菌数量、多样性与解钾能力差异, 旨在初步探讨根际解钾菌在加拿大一枝黄花入侵过程中发挥的作用, 为深入研究加拿大一枝黄花入侵机制提供理论依据. ...
... 植物与土壤微生物的相互作用在植物入侵过程中起着至关重要的作用(
Bowen et al ., 2017 ).研究表明外来植物通过改变入侵地土壤微生物结构和功能, 进而改善土壤养分供应, 促进植物生长, 增强植物抗逆性, 促使植物成功入侵(
Kourtev et al ., 2002 ;
Wardle et al ., 2004 ;
Nijjer et al ., 2008 ;
Coats et al ., 2014 ).例如, 紫茎泽兰入侵后改变了入侵地土壤微生物的群落结构, 同时提高了入侵地土壤肥力, 进而增强了紫茎泽兰的竞争能力(
牛红榜等, 2007 ).本研究发现加拿大一枝黄花根际土壤中解钾菌数量、群落结构和解钾活性都和伴生植物白茅有着较大的差异, 加拿大一枝黄花的根际解钾菌数量远多于对照, 且其中多数根际解钾菌均具有高效的解钾能力, 同时加拿大一枝黄花茎叶中的钾含量及入侵地土壤中全钾和速效钾的含量均高于对照.这说明, 加拿大一枝黄花的富钾作用可能与其较多的高效解钾菌有关.相关研究也表明加拿大一枝黄花入侵会显著改变土壤微生物群落结构和多样性(
沈荔花等, 2007 ;
陈晨等, 2009 ;
李国庆, 2009 ), 这些转变可能会显著地影响土壤中钾的有效性和供给水平.在其他入侵植物的相关研究中也发现, 入侵植物对土壤养分含量的影响也和其土壤微生物群落的改变紧密相关.例如, 黄顶菊根际耐寒短杆菌能增加土壤磷含量, 对病原菌的抑制作用较对照植物更强(
石青, 2018 ); 破坏草通过促进土壤中解磷菌的生长使其土壤中有效磷含量增加(
杨国庆等, 2014 );
Xanthium italicum 水提液不仅使土壤中细菌和真菌数量明显增多, 同时其土壤速效钾、速效氮等营养成分也显著增多(
邰凤姣等, 2016 ). ...
... 加拿大一枝黄花的入侵之所以更够改变根际解钾菌多样性可能与其根系分泌物有关.已有研究表明入侵植物能够通过根系分泌次生代谢物对土壤微生物群落结构造成一定的影响, 加拿大一枝黄花能够通过根系分泌物改变土壤的理化性质, 对不同微生物群落产生一定程度的增强或抑制作用(
沈荔花等, 2007 ;
Zhang et al ., 2007 ,
2009 ).本研究中加拿大一枝黄花根际解钾菌数量与解钾能力与对照植物存在明显差异, 这可能是由于加拿大一枝黄花能够通过特异性的根系分泌物化学组分影响根际解钾菌的数量、种类以及分布, 同时, 加拿大一枝黄花较大的生物量积累也可能使其释放更多的根系分泌物, 增加碳源供给, 从而增强了土壤微生物活性, 使得矿物质态的钾能够更快向水溶性钾转化, 从而导致土壤速效养分的提高.因此关于加拿大一枝黄花根系分泌物释放量及化学组分是如何影响根际功能微生物群落的结构和功能的还需要进一步深入研究.本研究只设置了一种本地伴生植物为对照, 实验结果有一定的局限, 后续研究将扩大研究范围, 增加不同功能类群的伴生物种作为对照, 以期更加深入全面地了解加拿大一枝黄花入侵后对入侵地土壤生态系统的影响及其机理. ...
加拿大一枝黄花对土壤微生物区系的影响研究
3
2007
... 目前国内外对加拿大一枝黄花入侵机制的研究主要集中在繁殖特性(
黄华和郭水良, 2005 ;
郝建华等, 2009 ;
张海亮等, 2015b )、化感作用(
梅玲笑, 2005 ;
郝建华等, 2009 )、生理生态特征与环境适应性(
郭水良和方芳, 2003 ;
杜乐山, 2014 )以及土壤微生物响应(
Zhang et al ., 2005 ;
沈荔花等, 2007 )等方面.研究表明, 在不同程度的入侵地中, 加拿大一枝黄花体内钾含量均高于伴生植物, 尤其在生长期和开花期差异最为显著, 且植物吸钾量与生物量呈正相关关系(
周振荣, 2010 ).在微生物响应方面, 研究发现加拿大一枝黄花入侵后能改变土壤微生物的群落数量、结构和功能多样性, 可能有助于加拿大一枝黄花进一步入侵(
Liao et al ., 2013 ;
Wang et al ., 2018 ).根际解钾菌能将土壤中难溶性钾转化为可被植物直接吸收的有效性钾, 进而促进植物钾吸收, 增强植物抗逆性(
李元芳, 1994 ;
郭勋斌等, 2001 ).加拿大一枝黄花入侵是否改变了根际解钾菌数量、结构和解钾活性等问题还有待深入研究.因此, 本研究以入侵植物加拿大一枝黄花和伴生本地植物白茅(
Imperata cylindrica )为研究对象, 比较加拿大一枝黄花与白茅的根际解钾菌数量、多样性与解钾能力差异, 旨在初步探讨根际解钾菌在加拿大一枝黄花入侵过程中发挥的作用, 为深入研究加拿大一枝黄花入侵机制提供理论依据. ...
... 植物与土壤微生物的相互作用在植物入侵过程中起着至关重要的作用(
Bowen et al ., 2017 ).研究表明外来植物通过改变入侵地土壤微生物结构和功能, 进而改善土壤养分供应, 促进植物生长, 增强植物抗逆性, 促使植物成功入侵(
Kourtev et al ., 2002 ;
Wardle et al ., 2004 ;
Nijjer et al ., 2008 ;
Coats et al ., 2014 ).例如, 紫茎泽兰入侵后改变了入侵地土壤微生物的群落结构, 同时提高了入侵地土壤肥力, 进而增强了紫茎泽兰的竞争能力(
牛红榜等, 2007 ).本研究发现加拿大一枝黄花根际土壤中解钾菌数量、群落结构和解钾活性都和伴生植物白茅有着较大的差异, 加拿大一枝黄花的根际解钾菌数量远多于对照, 且其中多数根际解钾菌均具有高效的解钾能力, 同时加拿大一枝黄花茎叶中的钾含量及入侵地土壤中全钾和速效钾的含量均高于对照.这说明, 加拿大一枝黄花的富钾作用可能与其较多的高效解钾菌有关.相关研究也表明加拿大一枝黄花入侵会显著改变土壤微生物群落结构和多样性(
沈荔花等, 2007 ;
陈晨等, 2009 ;
李国庆, 2009 ), 这些转变可能会显著地影响土壤中钾的有效性和供给水平.在其他入侵植物的相关研究中也发现, 入侵植物对土壤养分含量的影响也和其土壤微生物群落的改变紧密相关.例如, 黄顶菊根际耐寒短杆菌能增加土壤磷含量, 对病原菌的抑制作用较对照植物更强(
石青, 2018 ); 破坏草通过促进土壤中解磷菌的生长使其土壤中有效磷含量增加(
杨国庆等, 2014 );
Xanthium italicum 水提液不仅使土壤中细菌和真菌数量明显增多, 同时其土壤速效钾、速效氮等营养成分也显著增多(
邰凤姣等, 2016 ). ...
... 加拿大一枝黄花的入侵之所以更够改变根际解钾菌多样性可能与其根系分泌物有关.已有研究表明入侵植物能够通过根系分泌次生代谢物对土壤微生物群落结构造成一定的影响, 加拿大一枝黄花能够通过根系分泌物改变土壤的理化性质, 对不同微生物群落产生一定程度的增强或抑制作用(
沈荔花等, 2007 ;
Zhang et al ., 2007 ,
2009 ).本研究中加拿大一枝黄花根际解钾菌数量与解钾能力与对照植物存在明显差异, 这可能是由于加拿大一枝黄花能够通过特异性的根系分泌物化学组分影响根际解钾菌的数量、种类以及分布, 同时, 加拿大一枝黄花较大的生物量积累也可能使其释放更多的根系分泌物, 增加碳源供给, 从而增强了土壤微生物活性, 使得矿物质态的钾能够更快向水溶性钾转化, 从而导致土壤速效养分的提高.因此关于加拿大一枝黄花根系分泌物释放量及化学组分是如何影响根际功能微生物群落的结构和功能的还需要进一步深入研究.本研究只设置了一种本地伴生植物为对照, 实验结果有一定的局限, 后续研究将扩大研究范围, 增加不同功能类群的伴生物种作为对照, 以期更加深入全面地了解加拿大一枝黄花入侵后对入侵地土壤生态系统的影响及其机理. ...
耐寒短杆菌和巨大芽孢杆菌在黄顶菊入侵过程中的作用
1
2018
... 植物与土壤微生物的相互作用在植物入侵过程中起着至关重要的作用(
Bowen et al ., 2017 ).研究表明外来植物通过改变入侵地土壤微生物结构和功能, 进而改善土壤养分供应, 促进植物生长, 增强植物抗逆性, 促使植物成功入侵(
Kourtev et al ., 2002 ;
Wardle et al ., 2004 ;
Nijjer et al ., 2008 ;
Coats et al ., 2014 ).例如, 紫茎泽兰入侵后改变了入侵地土壤微生物的群落结构, 同时提高了入侵地土壤肥力, 进而增强了紫茎泽兰的竞争能力(
牛红榜等, 2007 ).本研究发现加拿大一枝黄花根际土壤中解钾菌数量、群落结构和解钾活性都和伴生植物白茅有着较大的差异, 加拿大一枝黄花的根际解钾菌数量远多于对照, 且其中多数根际解钾菌均具有高效的解钾能力, 同时加拿大一枝黄花茎叶中的钾含量及入侵地土壤中全钾和速效钾的含量均高于对照.这说明, 加拿大一枝黄花的富钾作用可能与其较多的高效解钾菌有关.相关研究也表明加拿大一枝黄花入侵会显著改变土壤微生物群落结构和多样性(
沈荔花等, 2007 ;
陈晨等, 2009 ;
李国庆, 2009 ), 这些转变可能会显著地影响土壤中钾的有效性和供给水平.在其他入侵植物的相关研究中也发现, 入侵植物对土壤养分含量的影响也和其土壤微生物群落的改变紧密相关.例如, 黄顶菊根际耐寒短杆菌能增加土壤磷含量, 对病原菌的抑制作用较对照植物更强(
石青, 2018 ); 破坏草通过促进土壤中解磷菌的生长使其土壤中有效磷含量增加(
杨国庆等, 2014 );
Xanthium italicum 水提液不仅使土壤中细菌和真菌数量明显增多, 同时其土壤速效钾、速效氮等营养成分也显著增多(
邰凤姣等, 2016 ). ...
耐寒短杆菌和巨大芽孢杆菌在黄顶菊入侵过程中的作用
1
2018
... 植物与土壤微生物的相互作用在植物入侵过程中起着至关重要的作用(
Bowen et al ., 2017 ).研究表明外来植物通过改变入侵地土壤微生物结构和功能, 进而改善土壤养分供应, 促进植物生长, 增强植物抗逆性, 促使植物成功入侵(
Kourtev et al ., 2002 ;
Wardle et al ., 2004 ;
Nijjer et al ., 2008 ;
Coats et al ., 2014 ).例如, 紫茎泽兰入侵后改变了入侵地土壤微生物的群落结构, 同时提高了入侵地土壤肥力, 进而增强了紫茎泽兰的竞争能力(
牛红榜等, 2007 ).本研究发现加拿大一枝黄花根际土壤中解钾菌数量、群落结构和解钾活性都和伴生植物白茅有着较大的差异, 加拿大一枝黄花的根际解钾菌数量远多于对照, 且其中多数根际解钾菌均具有高效的解钾能力, 同时加拿大一枝黄花茎叶中的钾含量及入侵地土壤中全钾和速效钾的含量均高于对照.这说明, 加拿大一枝黄花的富钾作用可能与其较多的高效解钾菌有关.相关研究也表明加拿大一枝黄花入侵会显著改变土壤微生物群落结构和多样性(
沈荔花等, 2007 ;
陈晨等, 2009 ;
李国庆, 2009 ), 这些转变可能会显著地影响土壤中钾的有效性和供给水平.在其他入侵植物的相关研究中也发现, 入侵植物对土壤养分含量的影响也和其土壤微生物群落的改变紧密相关.例如, 黄顶菊根际耐寒短杆菌能增加土壤磷含量, 对病原菌的抑制作用较对照植物更强(
石青, 2018 ); 破坏草通过促进土壤中解磷菌的生长使其土壤中有效磷含量增加(
杨国庆等, 2014 );
Xanthium italicum 水提液不仅使土壤中细菌和真菌数量明显增多, 同时其土壤速效钾、速效氮等营养成分也显著增多(
邰凤姣等, 2016 ). ...
Evidence for enhanced mutualism hypothesis:
Solidago canadensis plants from regular soils perform better
1
2010
... 外来植物在入侵过程中可能通过释放根系分泌物和改变微生物碳源等方式影响土壤微生物结构和功能, 而土壤微生物群落的改变反过来会影响外来植物的进一步入侵(
杨星等, 2012 ;
Uddin et al ., 2014 ;
Zhao et al ., 2014 ;
Martin et al ., 2017 ).以往研究表明, 紫茉莉(
Mirabilis jalapa )、破坏草(紫茎泽兰)(
Ageratina adenophora )、互花米草(
Spartina alterniflora )可以改变入侵地土壤微生物多样性(
牛红榜, 2007 ;
Zhao et al ., 2014 ;
廖丹等, 2018 ); 银荆(
Acacia dealbata )入侵西班牙北部地区后使当地土壤中细菌丰富度显著增加, 真菌的丰富度和多样性显著减少(
Lorenzo et al ., 2010 ); 旱雀麦(
Bromus tectorum )使入侵地的细菌数量显著增多, 真菌数量显著减少(
Belnap & Phillips, 2001 ); 葱芥(
Alliaria petiolata )入侵后导致土壤中丛枝菌根真菌数量显著减少(
Roberts & Anderson, 2001 ).这些结果说明, 外来植物入侵能够导致土壤微生物多样性发生明显改变.而另一方面, 土壤微生物的改变可能促进外来植物的入侵, 甚至可能抑制本地植物的生长.例如, 土壤微生物显著提高了入侵植物加拿大一枝黄花(
Solidago canadensis )的出芽率和资源竞争力, 而降低了本土植物长芒草(
Stipa bungeana )的竞争力(
Sun & He, 2010 ).因此, 探究入侵植物对土壤微生物的影响可能是了解植物入侵机理的关键所在. ...
外来入侵植物意大利苍耳对土壤微生物群落、土壤酶活性和土壤养分的影响
1
2016
... 植物与土壤微生物的相互作用在植物入侵过程中起着至关重要的作用(
Bowen et al ., 2017 ).研究表明外来植物通过改变入侵地土壤微生物结构和功能, 进而改善土壤养分供应, 促进植物生长, 增强植物抗逆性, 促使植物成功入侵(
Kourtev et al ., 2002 ;
Wardle et al ., 2004 ;
Nijjer et al ., 2008 ;
Coats et al ., 2014 ).例如, 紫茎泽兰入侵后改变了入侵地土壤微生物的群落结构, 同时提高了入侵地土壤肥力, 进而增强了紫茎泽兰的竞争能力(
牛红榜等, 2007 ).本研究发现加拿大一枝黄花根际土壤中解钾菌数量、群落结构和解钾活性都和伴生植物白茅有着较大的差异, 加拿大一枝黄花的根际解钾菌数量远多于对照, 且其中多数根际解钾菌均具有高效的解钾能力, 同时加拿大一枝黄花茎叶中的钾含量及入侵地土壤中全钾和速效钾的含量均高于对照.这说明, 加拿大一枝黄花的富钾作用可能与其较多的高效解钾菌有关.相关研究也表明加拿大一枝黄花入侵会显著改变土壤微生物群落结构和多样性(
沈荔花等, 2007 ;
陈晨等, 2009 ;
李国庆, 2009 ), 这些转变可能会显著地影响土壤中钾的有效性和供给水平.在其他入侵植物的相关研究中也发现, 入侵植物对土壤养分含量的影响也和其土壤微生物群落的改变紧密相关.例如, 黄顶菊根际耐寒短杆菌能增加土壤磷含量, 对病原菌的抑制作用较对照植物更强(
石青, 2018 ); 破坏草通过促进土壤中解磷菌的生长使其土壤中有效磷含量增加(
杨国庆等, 2014 );
Xanthium italicum 水提液不仅使土壤中细菌和真菌数量明显增多, 同时其土壤速效钾、速效氮等营养成分也显著增多(
邰凤姣等, 2016 ). ...
外来入侵植物意大利苍耳对土壤微生物群落、土壤酶活性和土壤养分的影响
1
2016
... 植物与土壤微生物的相互作用在植物入侵过程中起着至关重要的作用(
Bowen et al ., 2017 ).研究表明外来植物通过改变入侵地土壤微生物结构和功能, 进而改善土壤养分供应, 促进植物生长, 增强植物抗逆性, 促使植物成功入侵(
Kourtev et al ., 2002 ;
Wardle et al ., 2004 ;
Nijjer et al ., 2008 ;
Coats et al ., 2014 ).例如, 紫茎泽兰入侵后改变了入侵地土壤微生物的群落结构, 同时提高了入侵地土壤肥力, 进而增强了紫茎泽兰的竞争能力(
牛红榜等, 2007 ).本研究发现加拿大一枝黄花根际土壤中解钾菌数量、群落结构和解钾活性都和伴生植物白茅有着较大的差异, 加拿大一枝黄花的根际解钾菌数量远多于对照, 且其中多数根际解钾菌均具有高效的解钾能力, 同时加拿大一枝黄花茎叶中的钾含量及入侵地土壤中全钾和速效钾的含量均高于对照.这说明, 加拿大一枝黄花的富钾作用可能与其较多的高效解钾菌有关.相关研究也表明加拿大一枝黄花入侵会显著改变土壤微生物群落结构和多样性(
沈荔花等, 2007 ;
陈晨等, 2009 ;
李国庆, 2009 ), 这些转变可能会显著地影响土壤中钾的有效性和供给水平.在其他入侵植物的相关研究中也发现, 入侵植物对土壤养分含量的影响也和其土壤微生物群落的改变紧密相关.例如, 黄顶菊根际耐寒短杆菌能增加土壤磷含量, 对病原菌的抑制作用较对照植物更强(
石青, 2018 ); 破坏草通过促进土壤中解磷菌的生长使其土壤中有效磷含量增加(
杨国庆等, 2014 );
Xanthium italicum 水提液不仅使土壤中细菌和真菌数量明显增多, 同时其土壤速效钾、速效氮等营养成分也显著增多(
邰凤姣等, 2016 ). ...
1
1985
... 加拿大一枝黄花是菊科一枝黄花属植物, 多年生草本, 具有发达根状茎, 原产于北美地区, 最初作为庭园花卉引入我国上海、南京等地, 现已广泛逸生为恶性外来种(
中国科学院中国植物志编辑委员会, 1985 ;
郭水良和方芳, 2003 ).加拿大一枝黄花繁殖能力极强, 既能利用种子进行有性繁殖, 也可通过地下根状茎进行无性繁殖, 一般情况下, 每株植株可产约7万粒种子, 且种子具有较高萌发率, 此外, 植株根部具有上百个根茎结构, 每个根茎在第2年可萌发形成独立的个体, 如此高的繁殖能力使其在入侵后能够迅速扩散形成单优势种群落(
王玉良等, 2009 ;
张海亮等, 2015a ).此外, 加拿大一枝黄花根部还会产生一些化感物质影响周围土著植物生长, 对入侵地生物多样性造成了一定的影响(
方芳等, 2004 ).而目前加拿大一枝黄花在我国入侵范围逐渐扩大, 入侵速度逐渐加快, 对我国生态环境、生物多样性、农业和畜牧业均造成了不同程度的影响(
董梅等, 2006 ;
陆慧明等, 2006 ), 因此亟须加强加拿大一枝黄花入侵机理与防控技术方面的研究. ...
1
1985
... 加拿大一枝黄花是菊科一枝黄花属植物, 多年生草本, 具有发达根状茎, 原产于北美地区, 最初作为庭园花卉引入我国上海、南京等地, 现已广泛逸生为恶性外来种(
中国科学院中国植物志编辑委员会, 1985 ;
郭水良和方芳, 2003 ).加拿大一枝黄花繁殖能力极强, 既能利用种子进行有性繁殖, 也可通过地下根状茎进行无性繁殖, 一般情况下, 每株植株可产约7万粒种子, 且种子具有较高萌发率, 此外, 植株根部具有上百个根茎结构, 每个根茎在第2年可萌发形成独立的个体, 如此高的繁殖能力使其在入侵后能够迅速扩散形成单优势种群落(
王玉良等, 2009 ;
张海亮等, 2015a ).此外, 加拿大一枝黄花根部还会产生一些化感物质影响周围土著植物生长, 对入侵地生物多样性造成了一定的影响(
方芳等, 2004 ).而目前加拿大一枝黄花在我国入侵范围逐渐扩大, 入侵速度逐渐加快, 对我国生态环境、生物多样性、农业和畜牧业均造成了不同程度的影响(
董梅等, 2006 ;
陆慧明等, 2006 ), 因此亟须加强加拿大一枝黄花入侵机理与防控技术方面的研究. ...
Suppression of native
Melaleuca ericifolia by the invasive
Phragmites australis through allelopathic root exudates
1
2014
... 外来植物在入侵过程中可能通过释放根系分泌物和改变微生物碳源等方式影响土壤微生物结构和功能, 而土壤微生物群落的改变反过来会影响外来植物的进一步入侵(
杨星等, 2012 ;
Uddin et al ., 2014 ;
Zhao et al ., 2014 ;
Martin et al ., 2017 ).以往研究表明, 紫茉莉(
Mirabilis jalapa )、破坏草(紫茎泽兰)(
Ageratina adenophora )、互花米草(
Spartina alterniflora )可以改变入侵地土壤微生物多样性(
牛红榜, 2007 ;
Zhao et al ., 2014 ;
廖丹等, 2018 ); 银荆(
Acacia dealbata )入侵西班牙北部地区后使当地土壤中细菌丰富度显著增加, 真菌的丰富度和多样性显著减少(
Lorenzo et al ., 2010 ); 旱雀麦(
Bromus tectorum )使入侵地的细菌数量显著增多, 真菌数量显著减少(
Belnap & Phillips, 2001 ); 葱芥(
Alliaria petiolata )入侵后导致土壤中丛枝菌根真菌数量显著减少(
Roberts & Anderson, 2001 ).这些结果说明, 外来植物入侵能够导致土壤微生物多样性发生明显改变.而另一方面, 土壤微生物的改变可能促进外来植物的入侵, 甚至可能抑制本地植物的生长.例如, 土壤微生物显著提高了入侵植物加拿大一枝黄花(
Solidago canadensis )的出芽率和资源竞争力, 而降低了本土植物长芒草(
Stipa bungeana )的竞争力(
Sun & He, 2010 ).因此, 探究入侵植物对土壤微生物的影响可能是了解植物入侵机理的关键所在. ...
Responses of the soil fungal communities to the co-invasion of two invasive species with different cover classes
1
2018
... 目前国内外对加拿大一枝黄花入侵机制的研究主要集中在繁殖特性(
黄华和郭水良, 2005 ;
郝建华等, 2009 ;
张海亮等, 2015b )、化感作用(
梅玲笑, 2005 ;
郝建华等, 2009 )、生理生态特征与环境适应性(
郭水良和方芳, 2003 ;
杜乐山, 2014 )以及土壤微生物响应(
Zhang et al ., 2005 ;
沈荔花等, 2007 )等方面.研究表明, 在不同程度的入侵地中, 加拿大一枝黄花体内钾含量均高于伴生植物, 尤其在生长期和开花期差异最为显著, 且植物吸钾量与生物量呈正相关关系(
周振荣, 2010 ).在微生物响应方面, 研究发现加拿大一枝黄花入侵后能改变土壤微生物的群落数量、结构和功能多样性, 可能有助于加拿大一枝黄花进一步入侵(
Liao et al ., 2013 ;
Wang et al ., 2018 ).根际解钾菌能将土壤中难溶性钾转化为可被植物直接吸收的有效性钾, 进而促进植物钾吸收, 增强植物抗逆性(
李元芳, 1994 ;
郭勋斌等, 2001 ).加拿大一枝黄花入侵是否改变了根际解钾菌数量、结构和解钾活性等问题还有待深入研究.因此, 本研究以入侵植物加拿大一枝黄花和伴生本地植物白茅(
Imperata cylindrica )为研究对象, 比较加拿大一枝黄花与白茅的根际解钾菌数量、多样性与解钾能力差异, 旨在初步探讨根际解钾菌在加拿大一枝黄花入侵过程中发挥的作用, 为深入研究加拿大一枝黄花入侵机制提供理论依据. ...
大穗型小麦叶片性状、养分含量及氮素分配特征
1
2013
... 入侵植物对营养元素的吸收、积累和释放对其生长、入侵能力及其入侵的生态学效应都具有重要作用.钾是植物生长发育所需要的主要营养元素之一, 在植物生长繁育过程中发挥必不可少的作用, 钾的增加不仅能够提高植物光合速率和酶的活化速度、增加植物糖和蛋白质含量、增强植物对氮的吸收, 还能够提高作物的产量、提高植物的抗逆性(
Graham & Ulrich, 1972 ;
胡笃敬等, 1993 ).本研究发现, 野外自然生长的加拿大一枝黄花叶片和茎中的钾含量显著高于伴生植物白茅, 尤其叶片中的钾含量是白茅的7.33倍, 说明在相同生境下, 加拿大一枝黄花吸收钾的能力远高于本地植物, 且加拿大一枝黄花能够将更多的钾分配到叶片中, 而叶片是光合作用最主要的场所, 叶片中养分含量增多可能增强植物光合速率和生长代谢速率(
Aerts, 2000 ;
王丽芳等, 2013 ).此外, 加拿大一枝黄花叶片中的氮磷含量明显高于茎, 更加证实了加拿大一枝黄花将更多养分分配到叶片中的生长策略.人为提高钾供给水平均能促进两种植物生物量积累增加(
图3 ), 但是我们发现加拿大一枝黄花入侵地土壤0-10 cm及10-20 cm土层中全氮、全磷、全钾及碱解氮、有效磷和速效钾含量均多于对照植物白茅, 尤其土壤中速效养分的含量显著多于白茅, 这与
梁雷等(2016) 的研究结果相一致.
陈彤等(2012) 发现加拿大一枝黄花能显著提高土壤中硝态氮含量, 并且其较高的氮积累能力可能和入侵后本地植物种群的动态变化格局紧密相关 这些研究说明加拿大一枝黄花存在一种能够增强土壤养分积累的机制, 而这种机制反过来可能促进其进一步入侵.外来植物对入侵地土壤养分含量的影响被大量研究.一些研究认为外来植物入侵往往能够通过影响入侵地土壤养分而促进自身成功入侵(
Holmgren et al ., 1997 ).许多外来植物入侵后通过改变土壤肥力影响其与本地种的竞争能力, 例如破坏草、黄顶菊(
Flaveria bidentis )、盐生草(
Halogeton glomeratus )等植物入侵时均会导致土壤养分含量向有利于入侵植物的方向发生改变(
Duda et al ., 2003 ;
蒋智林等, 2008 ;
张天瑞等, 2010 ). ...
大穗型小麦叶片性状、养分含量及氮素分配特征
1
2013
... 入侵植物对营养元素的吸收、积累和释放对其生长、入侵能力及其入侵的生态学效应都具有重要作用.钾是植物生长发育所需要的主要营养元素之一, 在植物生长繁育过程中发挥必不可少的作用, 钾的增加不仅能够提高植物光合速率和酶的活化速度、增加植物糖和蛋白质含量、增强植物对氮的吸收, 还能够提高作物的产量、提高植物的抗逆性(
Graham & Ulrich, 1972 ;
胡笃敬等, 1993 ).本研究发现, 野外自然生长的加拿大一枝黄花叶片和茎中的钾含量显著高于伴生植物白茅, 尤其叶片中的钾含量是白茅的7.33倍, 说明在相同生境下, 加拿大一枝黄花吸收钾的能力远高于本地植物, 且加拿大一枝黄花能够将更多的钾分配到叶片中, 而叶片是光合作用最主要的场所, 叶片中养分含量增多可能增强植物光合速率和生长代谢速率(
Aerts, 2000 ;
王丽芳等, 2013 ).此外, 加拿大一枝黄花叶片中的氮磷含量明显高于茎, 更加证实了加拿大一枝黄花将更多养分分配到叶片中的生长策略.人为提高钾供给水平均能促进两种植物生物量积累增加(
图3 ), 但是我们发现加拿大一枝黄花入侵地土壤0-10 cm及10-20 cm土层中全氮、全磷、全钾及碱解氮、有效磷和速效钾含量均多于对照植物白茅, 尤其土壤中速效养分的含量显著多于白茅, 这与
梁雷等(2016) 的研究结果相一致.
陈彤等(2012) 发现加拿大一枝黄花能显著提高土壤中硝态氮含量, 并且其较高的氮积累能力可能和入侵后本地植物种群的动态变化格局紧密相关 这些研究说明加拿大一枝黄花存在一种能够增强土壤养分积累的机制, 而这种机制反过来可能促进其进一步入侵.外来植物对入侵地土壤养分含量的影响被大量研究.一些研究认为外来植物入侵往往能够通过影响入侵地土壤养分而促进自身成功入侵(
Holmgren et al ., 1997 ).许多外来植物入侵后通过改变土壤肥力影响其与本地种的竞争能力, 例如破坏草、黄顶菊(
Flaveria bidentis )、盐生草(
Halogeton glomeratus )等植物入侵时均会导致土壤养分含量向有利于入侵植物的方向发生改变(
Duda et al ., 2003 ;
蒋智林等, 2008 ;
张天瑞等, 2010 ). ...
外来植物加拿大一枝黄花(
Solidago canadensis )入侵的结构基础
1
2009
... 加拿大一枝黄花是菊科一枝黄花属植物, 多年生草本, 具有发达根状茎, 原产于北美地区, 最初作为庭园花卉引入我国上海、南京等地, 现已广泛逸生为恶性外来种(
中国科学院中国植物志编辑委员会, 1985 ;
郭水良和方芳, 2003 ).加拿大一枝黄花繁殖能力极强, 既能利用种子进行有性繁殖, 也可通过地下根状茎进行无性繁殖, 一般情况下, 每株植株可产约7万粒种子, 且种子具有较高萌发率, 此外, 植株根部具有上百个根茎结构, 每个根茎在第2年可萌发形成独立的个体, 如此高的繁殖能力使其在入侵后能够迅速扩散形成单优势种群落(
王玉良等, 2009 ;
张海亮等, 2015a ).此外, 加拿大一枝黄花根部还会产生一些化感物质影响周围土著植物生长, 对入侵地生物多样性造成了一定的影响(
方芳等, 2004 ).而目前加拿大一枝黄花在我国入侵范围逐渐扩大, 入侵速度逐渐加快, 对我国生态环境、生物多样性、农业和畜牧业均造成了不同程度的影响(
董梅等, 2006 ;
陆慧明等, 2006 ), 因此亟须加强加拿大一枝黄花入侵机理与防控技术方面的研究. ...
外来植物加拿大一枝黄花(
Solidago canadensis )入侵的结构基础
1
2009
... 加拿大一枝黄花是菊科一枝黄花属植物, 多年生草本, 具有发达根状茎, 原产于北美地区, 最初作为庭园花卉引入我国上海、南京等地, 现已广泛逸生为恶性外来种(
中国科学院中国植物志编辑委员会, 1985 ;
郭水良和方芳, 2003 ).加拿大一枝黄花繁殖能力极强, 既能利用种子进行有性繁殖, 也可通过地下根状茎进行无性繁殖, 一般情况下, 每株植株可产约7万粒种子, 且种子具有较高萌发率, 此外, 植株根部具有上百个根茎结构, 每个根茎在第2年可萌发形成独立的个体, 如此高的繁殖能力使其在入侵后能够迅速扩散形成单优势种群落(
王玉良等, 2009 ;
张海亮等, 2015a ).此外, 加拿大一枝黄花根部还会产生一些化感物质影响周围土著植物生长, 对入侵地生物多样性造成了一定的影响(
方芳等, 2004 ).而目前加拿大一枝黄花在我国入侵范围逐渐扩大, 入侵速度逐渐加快, 对我国生态环境、生物多样性、农业和畜牧业均造成了不同程度的影响(
董梅等, 2006 ;
陆慧明等, 2006 ), 因此亟须加强加拿大一枝黄花入侵机理与防控技术方面的研究. ...
Ecological linkages between aboveground and belowground biota
2
2004
... 外来植物入侵不仅导致入侵地农林生产经济损失, 而且对生态系统和生物多样性造成了严重的威胁(
鞠瑞亭等, 2012 ).据调查, 我国外来入侵植物多达670余种, 分布范围主要集中在东南沿海地区, 且入侵速度正在逐年加快、数量逐年增多, 若不加以防治, 则直接影响我国经济发展和物种栖息地保护(
闫小玲等, 2012 ).了解外来植物的入侵机制是制定科学合理的防控方案的关键.为了解释外来植物的入侵机制, 研究者提出了很多研究假说, 例如天敌逃逸假说(
Keane et al ., 2002 )、竞争能力增强进化假说(
Blossey & N?tzold, 1995 ), 但这些假说主要针对入侵植物本身的进化, 而没有考虑入侵植物与土壤微生物的相互作用.随着研究的不断深入, 越来越多的****意识到植物入侵过程与地下生态存在紧密联系(
de la Pe?a et al ., 2010 ;
Ren & Zhang, 2009 ), 入侵植物与土壤微生物之间的互作在入侵过程中发挥重要作用(
Wardle et al ., 2004 ). ...
... 植物与土壤微生物的相互作用在植物入侵过程中起着至关重要的作用(
Bowen et al ., 2017 ).研究表明外来植物通过改变入侵地土壤微生物结构和功能, 进而改善土壤养分供应, 促进植物生长, 增强植物抗逆性, 促使植物成功入侵(
Kourtev et al ., 2002 ;
Wardle et al ., 2004 ;
Nijjer et al ., 2008 ;
Coats et al ., 2014 ).例如, 紫茎泽兰入侵后改变了入侵地土壤微生物的群落结构, 同时提高了入侵地土壤肥力, 进而增强了紫茎泽兰的竞争能力(
牛红榜等, 2007 ).本研究发现加拿大一枝黄花根际土壤中解钾菌数量、群落结构和解钾活性都和伴生植物白茅有着较大的差异, 加拿大一枝黄花的根际解钾菌数量远多于对照, 且其中多数根际解钾菌均具有高效的解钾能力, 同时加拿大一枝黄花茎叶中的钾含量及入侵地土壤中全钾和速效钾的含量均高于对照.这说明, 加拿大一枝黄花的富钾作用可能与其较多的高效解钾菌有关.相关研究也表明加拿大一枝黄花入侵会显著改变土壤微生物群落结构和多样性(
沈荔花等, 2007 ;
陈晨等, 2009 ;
李国庆, 2009 ), 这些转变可能会显著地影响土壤中钾的有效性和供给水平.在其他入侵植物的相关研究中也发现, 入侵植物对土壤养分含量的影响也和其土壤微生物群落的改变紧密相关.例如, 黄顶菊根际耐寒短杆菌能增加土壤磷含量, 对病原菌的抑制作用较对照植物更强(
石青, 2018 ); 破坏草通过促进土壤中解磷菌的生长使其土壤中有效磷含量增加(
杨国庆等, 2014 );
Xanthium italicum 水提液不仅使土壤中细菌和真菌数量明显增多, 同时其土壤速效钾、速效氮等营养成分也显著增多(
邰凤姣等, 2016 ). ...
桑树根际硅酸盐细菌的分离鉴定及解钾能力测定
3
2010
... 本研究发现的加拿大一枝黄花和白茅根际解钾菌的种类和此前报道的研究具有一定的相似性.国际公认的解钾菌菌种仅3种: 环状芽孢杆菌(
Bacillus circulans )、胶质芽孢杆菌(
Bacillus mucilaginosus )、土壤芽孢杆菌(
Bacillus edaphicus ), 但近年来解钾菌的种属鉴定结果越来越丰富, 新增的种属有假单胞菌属(
Pseudomonas )(
吴凡等, 2010 ;
李新新等, 2014 ;
葛红莲和纪秀娥, 2017 )、根瘤菌属(
Rhizobium )、肠杆菌属(
Enterobacteriaceae )(
吴凡等, 2010 )、微杆菌属(
Microbacterium )、固氮菌属(
Azotobacter )(
易浪波等, 2012 ;
李新新等, 2014 )、中华根瘤菌属(
Sinorhizobium )、慢生根瘤菌属(
Mesorhizobium )、屈挠杆菌属(
Flexibacter )(
鞠伟, 2016 ;
李春钢等, 2017 )等.在加拿大一枝黄花和白茅根际土中分离获得的20株解钾菌中, H1-4、B6-7与
吴凡等(2010) 的研究结果一致, H6-5和H1-6、H5-10、H2-14、H2-17与
李新新等(2014) 的研究结果相同, H3-8与
张妙宜等(2016) 筛选出的解钾菌同属, 以上菌株均被证明具有一定的解钾能力.此外, 本研究发现的解钾能力较强但未被报道的菌株有: H1-15, 属于含铜菌属(
Cupriavidus ); H5-18, 属于
Pseudoflavitalea; H2-20, 属于溶杆菌属(
Lysobacter ); H1-21, 属于
Siphonobacter, 这一研究结果有助于进一步丰富解钾菌的种属类别. ...
... )(
吴凡等, 2010 )、微杆菌属(
Microbacterium )、固氮菌属(
Azotobacter )(
易浪波等, 2012 ;
李新新等, 2014 )、中华根瘤菌属(
Sinorhizobium )、慢生根瘤菌属(
Mesorhizobium )、屈挠杆菌属(
Flexibacter )(
鞠伟, 2016 ;
李春钢等, 2017 )等.在加拿大一枝黄花和白茅根际土中分离获得的20株解钾菌中, H1-4、B6-7与
吴凡等(2010) 的研究结果一致, H6-5和H1-6、H5-10、H2-14、H2-17与
李新新等(2014) 的研究结果相同, H3-8与
张妙宜等(2016) 筛选出的解钾菌同属, 以上菌株均被证明具有一定的解钾能力.此外, 本研究发现的解钾能力较强但未被报道的菌株有: H1-15, 属于含铜菌属(
Cupriavidus ); H5-18, 属于
Pseudoflavitalea; H2-20, 属于溶杆菌属(
Lysobacter ); H1-21, 属于
Siphonobacter, 这一研究结果有助于进一步丰富解钾菌的种属类别. ...
... )等.在加拿大一枝黄花和白茅根际土中分离获得的20株解钾菌中, H1-4、B6-7与
吴凡等(2010) 的研究结果一致, H6-5和H1-6、H5-10、H2-14、H2-17与
李新新等(2014) 的研究结果相同, H3-8与
张妙宜等(2016) 筛选出的解钾菌同属, 以上菌株均被证明具有一定的解钾能力.此外, 本研究发现的解钾能力较强但未被报道的菌株有: H1-15, 属于含铜菌属(
Cupriavidus ); H5-18, 属于
Pseudoflavitalea; H2-20, 属于溶杆菌属(
Lysobacter ); H1-21, 属于
Siphonobacter, 这一研究结果有助于进一步丰富解钾菌的种属类别. ...
桑树根际硅酸盐细菌的分离鉴定及解钾能力测定
3
2010
... 本研究发现的加拿大一枝黄花和白茅根际解钾菌的种类和此前报道的研究具有一定的相似性.国际公认的解钾菌菌种仅3种: 环状芽孢杆菌(
Bacillus circulans )、胶质芽孢杆菌(
Bacillus mucilaginosus )、土壤芽孢杆菌(
Bacillus edaphicus ), 但近年来解钾菌的种属鉴定结果越来越丰富, 新增的种属有假单胞菌属(
Pseudomonas )(
吴凡等, 2010 ;
李新新等, 2014 ;
葛红莲和纪秀娥, 2017 )、根瘤菌属(
Rhizobium )、肠杆菌属(
Enterobacteriaceae )(
吴凡等, 2010 )、微杆菌属(
Microbacterium )、固氮菌属(
Azotobacter )(
易浪波等, 2012 ;
李新新等, 2014 )、中华根瘤菌属(
Sinorhizobium )、慢生根瘤菌属(
Mesorhizobium )、屈挠杆菌属(
Flexibacter )(
鞠伟, 2016 ;
李春钢等, 2017 )等.在加拿大一枝黄花和白茅根际土中分离获得的20株解钾菌中, H1-4、B6-7与
吴凡等(2010) 的研究结果一致, H6-5和H1-6、H5-10、H2-14、H2-17与
李新新等(2014) 的研究结果相同, H3-8与
张妙宜等(2016) 筛选出的解钾菌同属, 以上菌株均被证明具有一定的解钾能力.此外, 本研究发现的解钾能力较强但未被报道的菌株有: H1-15, 属于含铜菌属(
Cupriavidus ); H5-18, 属于
Pseudoflavitalea; H2-20, 属于溶杆菌属(
Lysobacter ); H1-21, 属于
Siphonobacter, 这一研究结果有助于进一步丰富解钾菌的种属类别. ...
... )(
吴凡等, 2010 )、微杆菌属(
Microbacterium )、固氮菌属(
Azotobacter )(
易浪波等, 2012 ;
李新新等, 2014 )、中华根瘤菌属(
Sinorhizobium )、慢生根瘤菌属(
Mesorhizobium )、屈挠杆菌属(
Flexibacter )(
鞠伟, 2016 ;
李春钢等, 2017 )等.在加拿大一枝黄花和白茅根际土中分离获得的20株解钾菌中, H1-4、B6-7与
吴凡等(2010) 的研究结果一致, H6-5和H1-6、H5-10、H2-14、H2-17与
李新新等(2014) 的研究结果相同, H3-8与
张妙宜等(2016) 筛选出的解钾菌同属, 以上菌株均被证明具有一定的解钾能力.此外, 本研究发现的解钾能力较强但未被报道的菌株有: H1-15, 属于含铜菌属(
Cupriavidus ); H5-18, 属于
Pseudoflavitalea; H2-20, 属于溶杆菌属(
Lysobacter ); H1-21, 属于
Siphonobacter, 这一研究结果有助于进一步丰富解钾菌的种属类别. ...
... )等.在加拿大一枝黄花和白茅根际土中分离获得的20株解钾菌中, H1-4、B6-7与
吴凡等(2010) 的研究结果一致, H6-5和H1-6、H5-10、H2-14、H2-17与
李新新等(2014) 的研究结果相同, H3-8与
张妙宜等(2016) 筛选出的解钾菌同属, 以上菌株均被证明具有一定的解钾能力.此外, 本研究发现的解钾能力较强但未被报道的菌株有: H1-15, 属于含铜菌属(
Cupriavidus ); H5-18, 属于
Pseudoflavitalea; H2-20, 属于溶杆菌属(
Lysobacter ); H1-21, 属于
Siphonobacter, 这一研究结果有助于进一步丰富解钾菌的种属类别. ...
中国外来入侵植物研究现状及存在的问题
1
2012
... 外来植物入侵不仅导致入侵地农林生产经济损失, 而且对生态系统和生物多样性造成了严重的威胁(
鞠瑞亭等, 2012 ).据调查, 我国外来入侵植物多达670余种, 分布范围主要集中在东南沿海地区, 且入侵速度正在逐年加快、数量逐年增多, 若不加以防治, 则直接影响我国经济发展和物种栖息地保护(
闫小玲等, 2012 ).了解外来植物的入侵机制是制定科学合理的防控方案的关键.为了解释外来植物的入侵机制, 研究者提出了很多研究假说, 例如天敌逃逸假说(
Keane et al ., 2002 )、竞争能力增强进化假说(
Blossey & N?tzold, 1995 ), 但这些假说主要针对入侵植物本身的进化, 而没有考虑入侵植物与土壤微生物的相互作用.随着研究的不断深入, 越来越多的****意识到植物入侵过程与地下生态存在紧密联系(
de la Pe?a et al ., 2010 ;
Ren & Zhang, 2009 ), 入侵植物与土壤微生物之间的互作在入侵过程中发挥重要作用(
Wardle et al ., 2004 ). ...
中国外来入侵植物研究现状及存在的问题
1
2012
... 外来植物入侵不仅导致入侵地农林生产经济损失, 而且对生态系统和生物多样性造成了严重的威胁(
鞠瑞亭等, 2012 ).据调查, 我国外来入侵植物多达670余种, 分布范围主要集中在东南沿海地区, 且入侵速度正在逐年加快、数量逐年增多, 若不加以防治, 则直接影响我国经济发展和物种栖息地保护(
闫小玲等, 2012 ).了解外来植物的入侵机制是制定科学合理的防控方案的关键.为了解释外来植物的入侵机制, 研究者提出了很多研究假说, 例如天敌逃逸假说(
Keane et al ., 2002 )、竞争能力增强进化假说(
Blossey & N?tzold, 1995 ), 但这些假说主要针对入侵植物本身的进化, 而没有考虑入侵植物与土壤微生物的相互作用.随着研究的不断深入, 越来越多的****意识到植物入侵过程与地下生态存在紧密联系(
de la Pe?a et al ., 2010 ;
Ren & Zhang, 2009 ), 入侵植物与土壤微生物之间的互作在入侵过程中发挥重要作用(
Wardle et al ., 2004 ). ...
紫茎泽兰的化感物质对土壤有效磷含量和巨大芽孢杆菌生长的影响
1
2014
... 植物与土壤微生物的相互作用在植物入侵过程中起着至关重要的作用(
Bowen et al ., 2017 ).研究表明外来植物通过改变入侵地土壤微生物结构和功能, 进而改善土壤养分供应, 促进植物生长, 增强植物抗逆性, 促使植物成功入侵(
Kourtev et al ., 2002 ;
Wardle et al ., 2004 ;
Nijjer et al ., 2008 ;
Coats et al ., 2014 ).例如, 紫茎泽兰入侵后改变了入侵地土壤微生物的群落结构, 同时提高了入侵地土壤肥力, 进而增强了紫茎泽兰的竞争能力(
牛红榜等, 2007 ).本研究发现加拿大一枝黄花根际土壤中解钾菌数量、群落结构和解钾活性都和伴生植物白茅有着较大的差异, 加拿大一枝黄花的根际解钾菌数量远多于对照, 且其中多数根际解钾菌均具有高效的解钾能力, 同时加拿大一枝黄花茎叶中的钾含量及入侵地土壤中全钾和速效钾的含量均高于对照.这说明, 加拿大一枝黄花的富钾作用可能与其较多的高效解钾菌有关.相关研究也表明加拿大一枝黄花入侵会显著改变土壤微生物群落结构和多样性(
沈荔花等, 2007 ;
陈晨等, 2009 ;
李国庆, 2009 ), 这些转变可能会显著地影响土壤中钾的有效性和供给水平.在其他入侵植物的相关研究中也发现, 入侵植物对土壤养分含量的影响也和其土壤微生物群落的改变紧密相关.例如, 黄顶菊根际耐寒短杆菌能增加土壤磷含量, 对病原菌的抑制作用较对照植物更强(
石青, 2018 ); 破坏草通过促进土壤中解磷菌的生长使其土壤中有效磷含量增加(
杨国庆等, 2014 );
Xanthium italicum 水提液不仅使土壤中细菌和真菌数量明显增多, 同时其土壤速效钾、速效氮等营养成分也显著增多(
邰凤姣等, 2016 ). ...
紫茎泽兰的化感物质对土壤有效磷含量和巨大芽孢杆菌生长的影响
1
2014
... 植物与土壤微生物的相互作用在植物入侵过程中起着至关重要的作用(
Bowen et al ., 2017 ).研究表明外来植物通过改变入侵地土壤微生物结构和功能, 进而改善土壤养分供应, 促进植物生长, 增强植物抗逆性, 促使植物成功入侵(
Kourtev et al ., 2002 ;
Wardle et al ., 2004 ;
Nijjer et al ., 2008 ;
Coats et al ., 2014 ).例如, 紫茎泽兰入侵后改变了入侵地土壤微生物的群落结构, 同时提高了入侵地土壤肥力, 进而增强了紫茎泽兰的竞争能力(
牛红榜等, 2007 ).本研究发现加拿大一枝黄花根际土壤中解钾菌数量、群落结构和解钾活性都和伴生植物白茅有着较大的差异, 加拿大一枝黄花的根际解钾菌数量远多于对照, 且其中多数根际解钾菌均具有高效的解钾能力, 同时加拿大一枝黄花茎叶中的钾含量及入侵地土壤中全钾和速效钾的含量均高于对照.这说明, 加拿大一枝黄花的富钾作用可能与其较多的高效解钾菌有关.相关研究也表明加拿大一枝黄花入侵会显著改变土壤微生物群落结构和多样性(
沈荔花等, 2007 ;
陈晨等, 2009 ;
李国庆, 2009 ), 这些转变可能会显著地影响土壤中钾的有效性和供给水平.在其他入侵植物的相关研究中也发现, 入侵植物对土壤养分含量的影响也和其土壤微生物群落的改变紧密相关.例如, 黄顶菊根际耐寒短杆菌能增加土壤磷含量, 对病原菌的抑制作用较对照植物更强(
石青, 2018 ); 破坏草通过促进土壤中解磷菌的生长使其土壤中有效磷含量增加(
杨国庆等, 2014 );
Xanthium italicum 水提液不仅使土壤中细菌和真菌数量明显增多, 同时其土壤速效钾、速效氮等营养成分也显著增多(
邰凤姣等, 2016 ). ...
黄顶菊入侵对土壤微生物、土壤酶活性及土壤养分的影响
1
2012
... 外来植物在入侵过程中可能通过释放根系分泌物和改变微生物碳源等方式影响土壤微生物结构和功能, 而土壤微生物群落的改变反过来会影响外来植物的进一步入侵(
杨星等, 2012 ;
Uddin et al ., 2014 ;
Zhao et al ., 2014 ;
Martin et al ., 2017 ).以往研究表明, 紫茉莉(
Mirabilis jalapa )、破坏草(紫茎泽兰)(
Ageratina adenophora )、互花米草(
Spartina alterniflora )可以改变入侵地土壤微生物多样性(
牛红榜, 2007 ;
Zhao et al ., 2014 ;
廖丹等, 2018 ); 银荆(
Acacia dealbata )入侵西班牙北部地区后使当地土壤中细菌丰富度显著增加, 真菌的丰富度和多样性显著减少(
Lorenzo et al ., 2010 ); 旱雀麦(
Bromus tectorum )使入侵地的细菌数量显著增多, 真菌数量显著减少(
Belnap & Phillips, 2001 ); 葱芥(
Alliaria petiolata )入侵后导致土壤中丛枝菌根真菌数量显著减少(
Roberts & Anderson, 2001 ).这些结果说明, 外来植物入侵能够导致土壤微生物多样性发生明显改变.而另一方面, 土壤微生物的改变可能促进外来植物的入侵, 甚至可能抑制本地植物的生长.例如, 土壤微生物显著提高了入侵植物加拿大一枝黄花(
Solidago canadensis )的出芽率和资源竞争力, 而降低了本土植物长芒草(
Stipa bungeana )的竞争力(
Sun & He, 2010 ).因此, 探究入侵植物对土壤微生物的影响可能是了解植物入侵机理的关键所在. ...
黄顶菊入侵对土壤微生物、土壤酶活性及土壤养分的影响
1
2012
... 外来植物在入侵过程中可能通过释放根系分泌物和改变微生物碳源等方式影响土壤微生物结构和功能, 而土壤微生物群落的改变反过来会影响外来植物的进一步入侵(
杨星等, 2012 ;
Uddin et al ., 2014 ;
Zhao et al ., 2014 ;
Martin et al ., 2017 ).以往研究表明, 紫茉莉(
Mirabilis jalapa )、破坏草(紫茎泽兰)(
Ageratina adenophora )、互花米草(
Spartina alterniflora )可以改变入侵地土壤微生物多样性(
牛红榜, 2007 ;
Zhao et al ., 2014 ;
廖丹等, 2018 ); 银荆(
Acacia dealbata )入侵西班牙北部地区后使当地土壤中细菌丰富度显著增加, 真菌的丰富度和多样性显著减少(
Lorenzo et al ., 2010 ); 旱雀麦(
Bromus tectorum )使入侵地的细菌数量显著增多, 真菌数量显著减少(
Belnap & Phillips, 2001 ); 葱芥(
Alliaria petiolata )入侵后导致土壤中丛枝菌根真菌数量显著减少(
Roberts & Anderson, 2001 ).这些结果说明, 外来植物入侵能够导致土壤微生物多样性发生明显改变.而另一方面, 土壤微生物的改变可能促进外来植物的入侵, 甚至可能抑制本地植物的生长.例如, 土壤微生物显著提高了入侵植物加拿大一枝黄花(
Solidago canadensis )的出芽率和资源竞争力, 而降低了本土植物长芒草(
Stipa bungeana )的竞争力(
Sun & He, 2010 ).因此, 探究入侵植物对土壤微生物的影响可能是了解植物入侵机理的关键所在. ...
高效钾长石分解菌株的筛选、鉴定及解钾活性研究
1
2012
... 本研究发现的加拿大一枝黄花和白茅根际解钾菌的种类和此前报道的研究具有一定的相似性.国际公认的解钾菌菌种仅3种: 环状芽孢杆菌(
Bacillus circulans )、胶质芽孢杆菌(
Bacillus mucilaginosus )、土壤芽孢杆菌(
Bacillus edaphicus ), 但近年来解钾菌的种属鉴定结果越来越丰富, 新增的种属有假单胞菌属(
Pseudomonas )(
吴凡等, 2010 ;
李新新等, 2014 ;
葛红莲和纪秀娥, 2017 )、根瘤菌属(
Rhizobium )、肠杆菌属(
Enterobacteriaceae )(
吴凡等, 2010 )、微杆菌属(
Microbacterium )、固氮菌属(
Azotobacter )(
易浪波等, 2012 ;
李新新等, 2014 )、中华根瘤菌属(
Sinorhizobium )、慢生根瘤菌属(
Mesorhizobium )、屈挠杆菌属(
Flexibacter )(
鞠伟, 2016 ;
李春钢等, 2017 )等.在加拿大一枝黄花和白茅根际土中分离获得的20株解钾菌中, H1-4、B6-7与
吴凡等(2010) 的研究结果一致, H6-5和H1-6、H5-10、H2-14、H2-17与
李新新等(2014) 的研究结果相同, H3-8与
张妙宜等(2016) 筛选出的解钾菌同属, 以上菌株均被证明具有一定的解钾能力.此外, 本研究发现的解钾能力较强但未被报道的菌株有: H1-15, 属于含铜菌属(
Cupriavidus ); H5-18, 属于
Pseudoflavitalea; H2-20, 属于溶杆菌属(
Lysobacter ); H1-21, 属于
Siphonobacter, 这一研究结果有助于进一步丰富解钾菌的种属类别. ...
高效钾长石分解菌株的筛选、鉴定及解钾活性研究
1
2012
... 本研究发现的加拿大一枝黄花和白茅根际解钾菌的种类和此前报道的研究具有一定的相似性.国际公认的解钾菌菌种仅3种: 环状芽孢杆菌(
Bacillus circulans )、胶质芽孢杆菌(
Bacillus mucilaginosus )、土壤芽孢杆菌(
Bacillus edaphicus ), 但近年来解钾菌的种属鉴定结果越来越丰富, 新增的种属有假单胞菌属(
Pseudomonas )(
吴凡等, 2010 ;
李新新等, 2014 ;
葛红莲和纪秀娥, 2017 )、根瘤菌属(
Rhizobium )、肠杆菌属(
Enterobacteriaceae )(
吴凡等, 2010 )、微杆菌属(
Microbacterium )、固氮菌属(
Azotobacter )(
易浪波等, 2012 ;
李新新等, 2014 )、中华根瘤菌属(
Sinorhizobium )、慢生根瘤菌属(
Mesorhizobium )、屈挠杆菌属(
Flexibacter )(
鞠伟, 2016 ;
李春钢等, 2017 )等.在加拿大一枝黄花和白茅根际土中分离获得的20株解钾菌中, H1-4、B6-7与
吴凡等(2010) 的研究结果一致, H6-5和H1-6、H5-10、H2-14、H2-17与
李新新等(2014) 的研究结果相同, H3-8与
张妙宜等(2016) 筛选出的解钾菌同属, 以上菌株均被证明具有一定的解钾能力.此外, 本研究发现的解钾能力较强但未被报道的菌株有: H1-15, 属于含铜菌属(
Cupriavidus ); H5-18, 属于
Pseudoflavitalea; H2-20, 属于溶杆菌属(
Lysobacter ); H1-21, 属于
Siphonobacter, 这一研究结果有助于进一步丰富解钾菌的种属类别. ...
加拿大一枝黄花繁殖性状对其入侵性的影响
1
2015a
... 加拿大一枝黄花是菊科一枝黄花属植物, 多年生草本, 具有发达根状茎, 原产于北美地区, 最初作为庭园花卉引入我国上海、南京等地, 现已广泛逸生为恶性外来种(
中国科学院中国植物志编辑委员会, 1985 ;
郭水良和方芳, 2003 ).加拿大一枝黄花繁殖能力极强, 既能利用种子进行有性繁殖, 也可通过地下根状茎进行无性繁殖, 一般情况下, 每株植株可产约7万粒种子, 且种子具有较高萌发率, 此外, 植株根部具有上百个根茎结构, 每个根茎在第2年可萌发形成独立的个体, 如此高的繁殖能力使其在入侵后能够迅速扩散形成单优势种群落(
王玉良等, 2009 ;
张海亮等, 2015a ).此外, 加拿大一枝黄花根部还会产生一些化感物质影响周围土著植物生长, 对入侵地生物多样性造成了一定的影响(
方芳等, 2004 ).而目前加拿大一枝黄花在我国入侵范围逐渐扩大, 入侵速度逐渐加快, 对我国生态环境、生物多样性、农业和畜牧业均造成了不同程度的影响(
董梅等, 2006 ;
陆慧明等, 2006 ), 因此亟须加强加拿大一枝黄花入侵机理与防控技术方面的研究. ...
加拿大一枝黄花繁殖性状对其入侵性的影响
1
2015a
... 加拿大一枝黄花是菊科一枝黄花属植物, 多年生草本, 具有发达根状茎, 原产于北美地区, 最初作为庭园花卉引入我国上海、南京等地, 现已广泛逸生为恶性外来种(
中国科学院中国植物志编辑委员会, 1985 ;
郭水良和方芳, 2003 ).加拿大一枝黄花繁殖能力极强, 既能利用种子进行有性繁殖, 也可通过地下根状茎进行无性繁殖, 一般情况下, 每株植株可产约7万粒种子, 且种子具有较高萌发率, 此外, 植株根部具有上百个根茎结构, 每个根茎在第2年可萌发形成独立的个体, 如此高的繁殖能力使其在入侵后能够迅速扩散形成单优势种群落(
王玉良等, 2009 ;
张海亮等, 2015a ).此外, 加拿大一枝黄花根部还会产生一些化感物质影响周围土著植物生长, 对入侵地生物多样性造成了一定的影响(
方芳等, 2004 ).而目前加拿大一枝黄花在我国入侵范围逐渐扩大, 入侵速度逐渐加快, 对我国生态环境、生物多样性、农业和畜牧业均造成了不同程度的影响(
董梅等, 2006 ;
陆慧明等, 2006 ), 因此亟须加强加拿大一枝黄花入侵机理与防控技术方面的研究. ...
影响加拿大一枝黄花种子非随机脱落的因素
1
2015b
... 目前国内外对加拿大一枝黄花入侵机制的研究主要集中在繁殖特性(
黄华和郭水良, 2005 ;
郝建华等, 2009 ;
张海亮等, 2015b )、化感作用(
梅玲笑, 2005 ;
郝建华等, 2009 )、生理生态特征与环境适应性(
郭水良和方芳, 2003 ;
杜乐山, 2014 )以及土壤微生物响应(
Zhang et al ., 2005 ;
沈荔花等, 2007 )等方面.研究表明, 在不同程度的入侵地中, 加拿大一枝黄花体内钾含量均高于伴生植物, 尤其在生长期和开花期差异最为显著, 且植物吸钾量与生物量呈正相关关系(
周振荣, 2010 ).在微生物响应方面, 研究发现加拿大一枝黄花入侵后能改变土壤微生物的群落数量、结构和功能多样性, 可能有助于加拿大一枝黄花进一步入侵(
Liao et al ., 2013 ;
Wang et al ., 2018 ).根际解钾菌能将土壤中难溶性钾转化为可被植物直接吸收的有效性钾, 进而促进植物钾吸收, 增强植物抗逆性(
李元芳, 1994 ;
郭勋斌等, 2001 ).加拿大一枝黄花入侵是否改变了根际解钾菌数量、结构和解钾活性等问题还有待深入研究.因此, 本研究以入侵植物加拿大一枝黄花和伴生本地植物白茅(
Imperata cylindrica )为研究对象, 比较加拿大一枝黄花与白茅的根际解钾菌数量、多样性与解钾能力差异, 旨在初步探讨根际解钾菌在加拿大一枝黄花入侵过程中发挥的作用, 为深入研究加拿大一枝黄花入侵机制提供理论依据. ...
影响加拿大一枝黄花种子非随机脱落的因素
1
2015b
... 目前国内外对加拿大一枝黄花入侵机制的研究主要集中在繁殖特性(
黄华和郭水良, 2005 ;
郝建华等, 2009 ;
张海亮等, 2015b )、化感作用(
梅玲笑, 2005 ;
郝建华等, 2009 )、生理生态特征与环境适应性(
郭水良和方芳, 2003 ;
杜乐山, 2014 )以及土壤微生物响应(
Zhang et al ., 2005 ;
沈荔花等, 2007 )等方面.研究表明, 在不同程度的入侵地中, 加拿大一枝黄花体内钾含量均高于伴生植物, 尤其在生长期和开花期差异最为显著, 且植物吸钾量与生物量呈正相关关系(
周振荣, 2010 ).在微生物响应方面, 研究发现加拿大一枝黄花入侵后能改变土壤微生物的群落数量、结构和功能多样性, 可能有助于加拿大一枝黄花进一步入侵(
Liao et al ., 2013 ;
Wang et al ., 2018 ).根际解钾菌能将土壤中难溶性钾转化为可被植物直接吸收的有效性钾, 进而促进植物钾吸收, 增强植物抗逆性(
李元芳, 1994 ;
郭勋斌等, 2001 ).加拿大一枝黄花入侵是否改变了根际解钾菌数量、结构和解钾活性等问题还有待深入研究.因此, 本研究以入侵植物加拿大一枝黄花和伴生本地植物白茅(
Imperata cylindrica )为研究对象, 比较加拿大一枝黄花与白茅的根际解钾菌数量、多样性与解钾能力差异, 旨在初步探讨根际解钾菌在加拿大一枝黄花入侵过程中发挥的作用, 为深入研究加拿大一枝黄花入侵机制提供理论依据. ...
蓖麻根际土壤解钾菌的筛选鉴定及发酵条件的优化
1
2016
... 本研究发现的加拿大一枝黄花和白茅根际解钾菌的种类和此前报道的研究具有一定的相似性.国际公认的解钾菌菌种仅3种: 环状芽孢杆菌(
Bacillus circulans )、胶质芽孢杆菌(
Bacillus mucilaginosus )、土壤芽孢杆菌(
Bacillus edaphicus ), 但近年来解钾菌的种属鉴定结果越来越丰富, 新增的种属有假单胞菌属(
Pseudomonas )(
吴凡等, 2010 ;
李新新等, 2014 ;
葛红莲和纪秀娥, 2017 )、根瘤菌属(
Rhizobium )、肠杆菌属(
Enterobacteriaceae )(
吴凡等, 2010 )、微杆菌属(
Microbacterium )、固氮菌属(
Azotobacter )(
易浪波等, 2012 ;
李新新等, 2014 )、中华根瘤菌属(
Sinorhizobium )、慢生根瘤菌属(
Mesorhizobium )、屈挠杆菌属(
Flexibacter )(
鞠伟, 2016 ;
李春钢等, 2017 )等.在加拿大一枝黄花和白茅根际土中分离获得的20株解钾菌中, H1-4、B6-7与
吴凡等(2010) 的研究结果一致, H6-5和H1-6、H5-10、H2-14、H2-17与
李新新等(2014) 的研究结果相同, H3-8与
张妙宜等(2016) 筛选出的解钾菌同属, 以上菌株均被证明具有一定的解钾能力.此外, 本研究发现的解钾能力较强但未被报道的菌株有: H1-15, 属于含铜菌属(
Cupriavidus ); H5-18, 属于
Pseudoflavitalea; H2-20, 属于溶杆菌属(
Lysobacter ); H1-21, 属于
Siphonobacter, 这一研究结果有助于进一步丰富解钾菌的种属类别. ...
蓖麻根际土壤解钾菌的筛选鉴定及发酵条件的优化
1
2016
... 本研究发现的加拿大一枝黄花和白茅根际解钾菌的种类和此前报道的研究具有一定的相似性.国际公认的解钾菌菌种仅3种: 环状芽孢杆菌(
Bacillus circulans )、胶质芽孢杆菌(
Bacillus mucilaginosus )、土壤芽孢杆菌(
Bacillus edaphicus ), 但近年来解钾菌的种属鉴定结果越来越丰富, 新增的种属有假单胞菌属(
Pseudomonas )(
吴凡等, 2010 ;
李新新等, 2014 ;
葛红莲和纪秀娥, 2017 )、根瘤菌属(
Rhizobium )、肠杆菌属(
Enterobacteriaceae )(
吴凡等, 2010 )、微杆菌属(
Microbacterium )、固氮菌属(
Azotobacter )(
易浪波等, 2012 ;
李新新等, 2014 )、中华根瘤菌属(
Sinorhizobium )、慢生根瘤菌属(
Mesorhizobium )、屈挠杆菌属(
Flexibacter )(
鞠伟, 2016 ;
李春钢等, 2017 )等.在加拿大一枝黄花和白茅根际土中分离获得的20株解钾菌中, H1-4、B6-7与
吴凡等(2010) 的研究结果一致, H6-5和H1-6、H5-10、H2-14、H2-17与
李新新等(2014) 的研究结果相同, H3-8与
张妙宜等(2016) 筛选出的解钾菌同属, 以上菌株均被证明具有一定的解钾能力.此外, 本研究发现的解钾能力较强但未被报道的菌株有: H1-15, 属于含铜菌属(
Cupriavidus ); H5-18, 属于
Pseudoflavitalea; H2-20, 属于溶杆菌属(
Lysobacter ); H1-21, 属于
Siphonobacter, 这一研究结果有助于进一步丰富解钾菌的种属类别. ...
Potential allelopathic effects of an invasive species
Solidago canadensis on the mycorrhizae of native plant species
1
2007
... 加拿大一枝黄花的入侵之所以更够改变根际解钾菌多样性可能与其根系分泌物有关.已有研究表明入侵植物能够通过根系分泌次生代谢物对土壤微生物群落结构造成一定的影响, 加拿大一枝黄花能够通过根系分泌物改变土壤的理化性质, 对不同微生物群落产生一定程度的增强或抑制作用(
沈荔花等, 2007 ;
Zhang et al ., 2007 ,
2009 ).本研究中加拿大一枝黄花根际解钾菌数量与解钾能力与对照植物存在明显差异, 这可能是由于加拿大一枝黄花能够通过特异性的根系分泌物化学组分影响根际解钾菌的数量、种类以及分布, 同时, 加拿大一枝黄花较大的生物量积累也可能使其释放更多的根系分泌物, 增加碳源供给, 从而增强了土壤微生物活性, 使得矿物质态的钾能够更快向水溶性钾转化, 从而导致土壤速效养分的提高.因此关于加拿大一枝黄花根系分泌物释放量及化学组分是如何影响根际功能微生物群落的结构和功能的还需要进一步深入研究.本研究只设置了一种本地伴生植物为对照, 实验结果有一定的局限, 后续研究将扩大研究范围, 增加不同功能类群的伴生物种作为对照, 以期更加深入全面地了解加拿大一枝黄花入侵后对入侵地土壤生态系统的影响及其机理. ...
The invasive plant
Solidago canadensis L. suppresses local soil pathogens through allelopathy
1
2009
... 加拿大一枝黄花的入侵之所以更够改变根际解钾菌多样性可能与其根系分泌物有关.已有研究表明入侵植物能够通过根系分泌次生代谢物对土壤微生物群落结构造成一定的影响, 加拿大一枝黄花能够通过根系分泌物改变土壤的理化性质, 对不同微生物群落产生一定程度的增强或抑制作用(
沈荔花等, 2007 ;
Zhang et al ., 2007 ,
2009 ).本研究中加拿大一枝黄花根际解钾菌数量与解钾能力与对照植物存在明显差异, 这可能是由于加拿大一枝黄花能够通过特异性的根系分泌物化学组分影响根际解钾菌的数量、种类以及分布, 同时, 加拿大一枝黄花较大的生物量积累也可能使其释放更多的根系分泌物, 增加碳源供给, 从而增强了土壤微生物活性, 使得矿物质态的钾能够更快向水溶性钾转化, 从而导致土壤速效养分的提高.因此关于加拿大一枝黄花根系分泌物释放量及化学组分是如何影响根际功能微生物群落的结构和功能的还需要进一步深入研究.本研究只设置了一种本地伴生植物为对照, 实验结果有一定的局限, 后续研究将扩大研究范围, 增加不同功能类群的伴生物种作为对照, 以期更加深入全面地了解加拿大一枝黄花入侵后对入侵地土壤生态系统的影响及其机理. ...
黄顶菊入侵对土壤养分和酶活性的影响
1
2010
... 入侵植物对营养元素的吸收、积累和释放对其生长、入侵能力及其入侵的生态学效应都具有重要作用.钾是植物生长发育所需要的主要营养元素之一, 在植物生长繁育过程中发挥必不可少的作用, 钾的增加不仅能够提高植物光合速率和酶的活化速度、增加植物糖和蛋白质含量、增强植物对氮的吸收, 还能够提高作物的产量、提高植物的抗逆性(
Graham & Ulrich, 1972 ;
胡笃敬等, 1993 ).本研究发现, 野外自然生长的加拿大一枝黄花叶片和茎中的钾含量显著高于伴生植物白茅, 尤其叶片中的钾含量是白茅的7.33倍, 说明在相同生境下, 加拿大一枝黄花吸收钾的能力远高于本地植物, 且加拿大一枝黄花能够将更多的钾分配到叶片中, 而叶片是光合作用最主要的场所, 叶片中养分含量增多可能增强植物光合速率和生长代谢速率(
Aerts, 2000 ;
王丽芳等, 2013 ).此外, 加拿大一枝黄花叶片中的氮磷含量明显高于茎, 更加证实了加拿大一枝黄花将更多养分分配到叶片中的生长策略.人为提高钾供给水平均能促进两种植物生物量积累增加(
图3 ), 但是我们发现加拿大一枝黄花入侵地土壤0-10 cm及10-20 cm土层中全氮、全磷、全钾及碱解氮、有效磷和速效钾含量均多于对照植物白茅, 尤其土壤中速效养分的含量显著多于白茅, 这与
梁雷等(2016) 的研究结果相一致.
陈彤等(2012) 发现加拿大一枝黄花能显著提高土壤中硝态氮含量, 并且其较高的氮积累能力可能和入侵后本地植物种群的动态变化格局紧密相关 这些研究说明加拿大一枝黄花存在一种能够增强土壤养分积累的机制, 而这种机制反过来可能促进其进一步入侵.外来植物对入侵地土壤养分含量的影响被大量研究.一些研究认为外来植物入侵往往能够通过影响入侵地土壤养分而促进自身成功入侵(
Holmgren et al ., 1997 ).许多外来植物入侵后通过改变土壤肥力影响其与本地种的竞争能力, 例如破坏草、黄顶菊(
Flaveria bidentis )、盐生草(
Halogeton glomeratus )等植物入侵时均会导致土壤养分含量向有利于入侵植物的方向发生改变(
Duda et al ., 2003 ;
蒋智林等, 2008 ;
张天瑞等, 2010 ). ...
黄顶菊入侵对土壤养分和酶活性的影响
1
2010
... 入侵植物对营养元素的吸收、积累和释放对其生长、入侵能力及其入侵的生态学效应都具有重要作用.钾是植物生长发育所需要的主要营养元素之一, 在植物生长繁育过程中发挥必不可少的作用, 钾的增加不仅能够提高植物光合速率和酶的活化速度、增加植物糖和蛋白质含量、增强植物对氮的吸收, 还能够提高作物的产量、提高植物的抗逆性(
Graham & Ulrich, 1972 ;
胡笃敬等, 1993 ).本研究发现, 野外自然生长的加拿大一枝黄花叶片和茎中的钾含量显著高于伴生植物白茅, 尤其叶片中的钾含量是白茅的7.33倍, 说明在相同生境下, 加拿大一枝黄花吸收钾的能力远高于本地植物, 且加拿大一枝黄花能够将更多的钾分配到叶片中, 而叶片是光合作用最主要的场所, 叶片中养分含量增多可能增强植物光合速率和生长代谢速率(
Aerts, 2000 ;
王丽芳等, 2013 ).此外, 加拿大一枝黄花叶片中的氮磷含量明显高于茎, 更加证实了加拿大一枝黄花将更多养分分配到叶片中的生长策略.人为提高钾供给水平均能促进两种植物生物量积累增加(
图3 ), 但是我们发现加拿大一枝黄花入侵地土壤0-10 cm及10-20 cm土层中全氮、全磷、全钾及碱解氮、有效磷和速效钾含量均多于对照植物白茅, 尤其土壤中速效养分的含量显著多于白茅, 这与
梁雷等(2016) 的研究结果相一致.
陈彤等(2012) 发现加拿大一枝黄花能显著提高土壤中硝态氮含量, 并且其较高的氮积累能力可能和入侵后本地植物种群的动态变化格局紧密相关 这些研究说明加拿大一枝黄花存在一种能够增强土壤养分积累的机制, 而这种机制反过来可能促进其进一步入侵.外来植物对入侵地土壤养分含量的影响被大量研究.一些研究认为外来植物入侵往往能够通过影响入侵地土壤养分而促进自身成功入侵(
Holmgren et al ., 1997 ).许多外来植物入侵后通过改变土壤肥力影响其与本地种的竞争能力, 例如破坏草、黄顶菊(
Flaveria bidentis )、盐生草(
Halogeton glomeratus )等植物入侵时均会导致土壤养分含量向有利于入侵植物的方向发生改变(
Duda et al ., 2003 ;
蒋智林等, 2008 ;
张天瑞等, 2010 ). ...
Responses of soil microbial community structure and diversity to agricultural deintensification
1
2005
... 目前国内外对加拿大一枝黄花入侵机制的研究主要集中在繁殖特性(
黄华和郭水良, 2005 ;
郝建华等, 2009 ;
张海亮等, 2015b )、化感作用(
梅玲笑, 2005 ;
郝建华等, 2009 )、生理生态特征与环境适应性(
郭水良和方芳, 2003 ;
杜乐山, 2014 )以及土壤微生物响应(
Zhang et al ., 2005 ;
沈荔花等, 2007 )等方面.研究表明, 在不同程度的入侵地中, 加拿大一枝黄花体内钾含量均高于伴生植物, 尤其在生长期和开花期差异最为显著, 且植物吸钾量与生物量呈正相关关系(
周振荣, 2010 ).在微生物响应方面, 研究发现加拿大一枝黄花入侵后能改变土壤微生物的群落数量、结构和功能多样性, 可能有助于加拿大一枝黄花进一步入侵(
Liao et al ., 2013 ;
Wang et al ., 2018 ).根际解钾菌能将土壤中难溶性钾转化为可被植物直接吸收的有效性钾, 进而促进植物钾吸收, 增强植物抗逆性(
李元芳, 1994 ;
郭勋斌等, 2001 ).加拿大一枝黄花入侵是否改变了根际解钾菌数量、结构和解钾活性等问题还有待深入研究.因此, 本研究以入侵植物加拿大一枝黄花和伴生本地植物白茅(
Imperata cylindrica )为研究对象, 比较加拿大一枝黄花与白茅的根际解钾菌数量、多样性与解钾能力差异, 旨在初步探讨根际解钾菌在加拿大一枝黄花入侵过程中发挥的作用, 为深入研究加拿大一枝黄花入侵机制提供理论依据. ...
Effects of root exudates from invasive plant (
Mirabilis jalapa ) on soil microenviroment under different land-use types
2
2014
... 外来植物在入侵过程中可能通过释放根系分泌物和改变微生物碳源等方式影响土壤微生物结构和功能, 而土壤微生物群落的改变反过来会影响外来植物的进一步入侵(
杨星等, 2012 ;
Uddin et al ., 2014 ;
Zhao et al ., 2014 ;
Martin et al ., 2017 ).以往研究表明, 紫茉莉(
Mirabilis jalapa )、破坏草(紫茎泽兰)(
Ageratina adenophora )、互花米草(
Spartina alterniflora )可以改变入侵地土壤微生物多样性(
牛红榜, 2007 ;
Zhao et al ., 2014 ;
廖丹等, 2018 ); 银荆(
Acacia dealbata )入侵西班牙北部地区后使当地土壤中细菌丰富度显著增加, 真菌的丰富度和多样性显著减少(
Lorenzo et al ., 2010 ); 旱雀麦(
Bromus tectorum )使入侵地的细菌数量显著增多, 真菌数量显著减少(
Belnap & Phillips, 2001 ); 葱芥(
Alliaria petiolata )入侵后导致土壤中丛枝菌根真菌数量显著减少(
Roberts & Anderson, 2001 ).这些结果说明, 外来植物入侵能够导致土壤微生物多样性发生明显改变.而另一方面, 土壤微生物的改变可能促进外来植物的入侵, 甚至可能抑制本地植物的生长.例如, 土壤微生物显著提高了入侵植物加拿大一枝黄花(
Solidago canadensis )的出芽率和资源竞争力, 而降低了本土植物长芒草(
Stipa bungeana )的竞争力(
Sun & He, 2010 ).因此, 探究入侵植物对土壤微生物的影响可能是了解植物入侵机理的关键所在. ...
... ;
Zhao et al ., 2014 ;
廖丹等, 2018 ); 银荆(
Acacia dealbata )入侵西班牙北部地区后使当地土壤中细菌丰富度显著增加, 真菌的丰富度和多样性显著减少(
Lorenzo et al ., 2010 ); 旱雀麦(
Bromus tectorum )使入侵地的细菌数量显著增多, 真菌数量显著减少(
Belnap & Phillips, 2001 ); 葱芥(
Alliaria petiolata )入侵后导致土壤中丛枝菌根真菌数量显著减少(
Roberts & Anderson, 2001 ).这些结果说明, 外来植物入侵能够导致土壤微生物多样性发生明显改变.而另一方面, 土壤微生物的改变可能促进外来植物的入侵, 甚至可能抑制本地植物的生长.例如, 土壤微生物显著提高了入侵植物加拿大一枝黄花(
Solidago canadensis )的出芽率和资源竞争力, 而降低了本土植物长芒草(
Stipa bungeana )的竞争力(
Sun & He, 2010 ).因此, 探究入侵植物对土壤微生物的影响可能是了解植物入侵机理的关键所在. ...
外来入侵植物加拿大一枝黄花对根际土壤微环境的影响研究
1
2010
... 目前国内外对加拿大一枝黄花入侵机制的研究主要集中在繁殖特性(
黄华和郭水良, 2005 ;
郝建华等, 2009 ;
张海亮等, 2015b )、化感作用(
梅玲笑, 2005 ;
郝建华等, 2009 )、生理生态特征与环境适应性(
郭水良和方芳, 2003 ;
杜乐山, 2014 )以及土壤微生物响应(
Zhang et al ., 2005 ;
沈荔花等, 2007 )等方面.研究表明, 在不同程度的入侵地中, 加拿大一枝黄花体内钾含量均高于伴生植物, 尤其在生长期和开花期差异最为显著, 且植物吸钾量与生物量呈正相关关系(
周振荣, 2010 ).在微生物响应方面, 研究发现加拿大一枝黄花入侵后能改变土壤微生物的群落数量、结构和功能多样性, 可能有助于加拿大一枝黄花进一步入侵(
Liao et al ., 2013 ;
Wang et al ., 2018 ).根际解钾菌能将土壤中难溶性钾转化为可被植物直接吸收的有效性钾, 进而促进植物钾吸收, 增强植物抗逆性(
李元芳, 1994 ;
郭勋斌等, 2001 ).加拿大一枝黄花入侵是否改变了根际解钾菌数量、结构和解钾活性等问题还有待深入研究.因此, 本研究以入侵植物加拿大一枝黄花和伴生本地植物白茅(
Imperata cylindrica )为研究对象, 比较加拿大一枝黄花与白茅的根际解钾菌数量、多样性与解钾能力差异, 旨在初步探讨根际解钾菌在加拿大一枝黄花入侵过程中发挥的作用, 为深入研究加拿大一枝黄花入侵机制提供理论依据. ...
外来入侵植物加拿大一枝黄花对根际土壤微环境的影响研究
1
2010
... 目前国内外对加拿大一枝黄花入侵机制的研究主要集中在繁殖特性(
黄华和郭水良, 2005 ;
郝建华等, 2009 ;
张海亮等, 2015b )、化感作用(
梅玲笑, 2005 ;
郝建华等, 2009 )、生理生态特征与环境适应性(
郭水良和方芳, 2003 ;
杜乐山, 2014 )以及土壤微生物响应(
Zhang et al ., 2005 ;
沈荔花等, 2007 )等方面.研究表明, 在不同程度的入侵地中, 加拿大一枝黄花体内钾含量均高于伴生植物, 尤其在生长期和开花期差异最为显著, 且植物吸钾量与生物量呈正相关关系(
周振荣, 2010 ).在微生物响应方面, 研究发现加拿大一枝黄花入侵后能改变土壤微生物的群落数量、结构和功能多样性, 可能有助于加拿大一枝黄花进一步入侵(
Liao et al ., 2013 ;
Wang et al ., 2018 ).根际解钾菌能将土壤中难溶性钾转化为可被植物直接吸收的有效性钾, 进而促进植物钾吸收, 增强植物抗逆性(
李元芳, 1994 ;
郭勋斌等, 2001 ).加拿大一枝黄花入侵是否改变了根际解钾菌数量、结构和解钾活性等问题还有待深入研究.因此, 本研究以入侵植物加拿大一枝黄花和伴生本地植物白茅(
Imperata cylindrica )为研究对象, 比较加拿大一枝黄花与白茅的根际解钾菌数量、多样性与解钾能力差异, 旨在初步探讨根际解钾菌在加拿大一枝黄花入侵过程中发挥的作用, 为深入研究加拿大一枝黄花入侵机制提供理论依据. ...
 ,1,*, 吴明1, 闫明2, 张昕丽1
,1,*, 吴明1, 闫明2, 张昕丽1
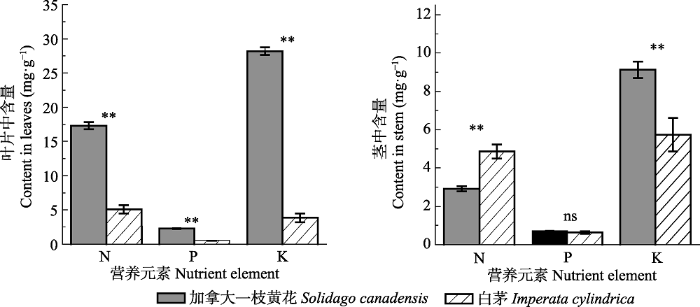 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT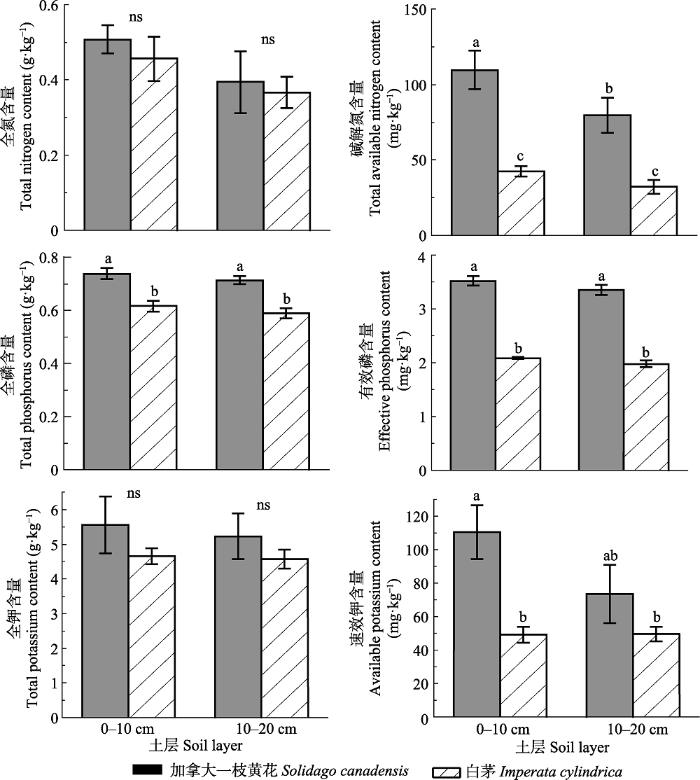 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT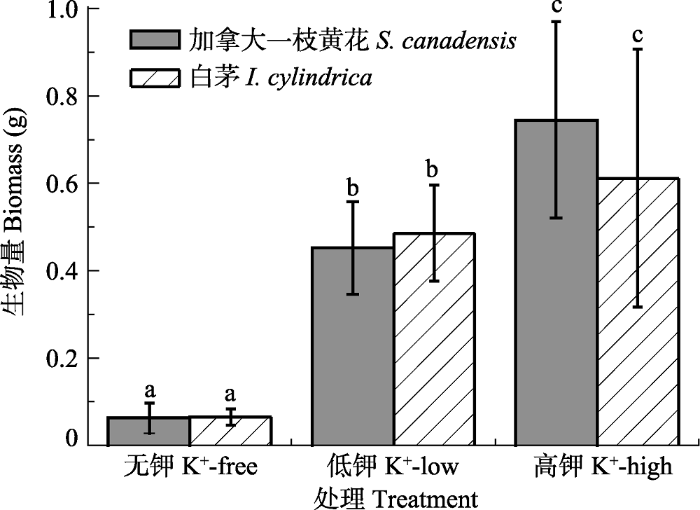 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT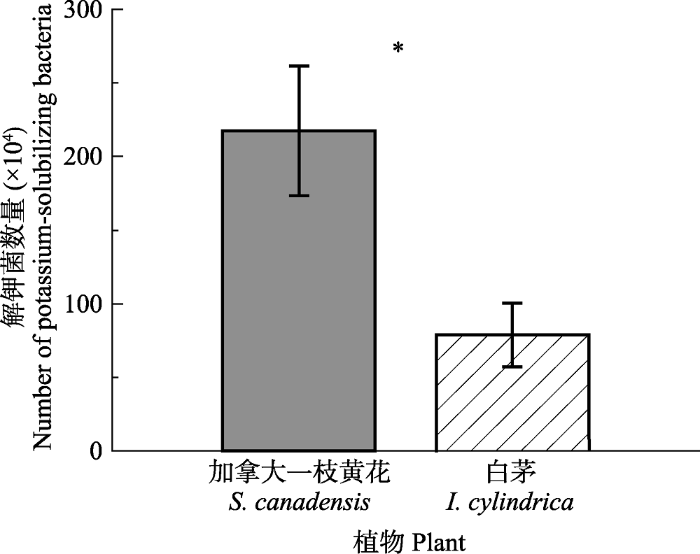 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT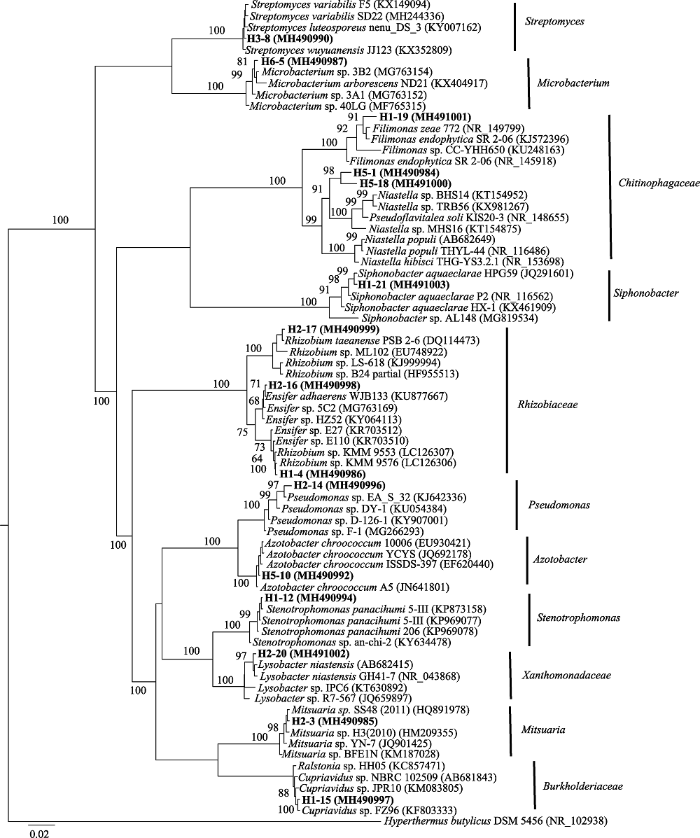 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT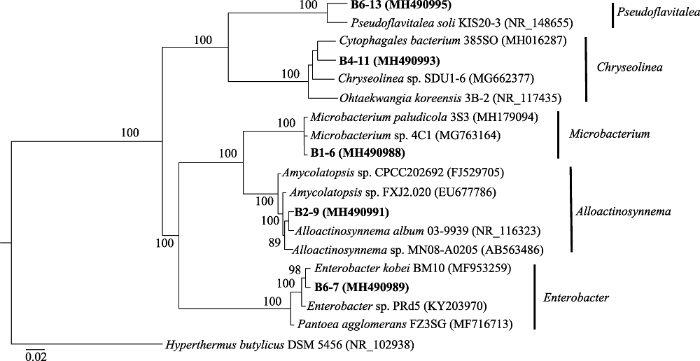 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT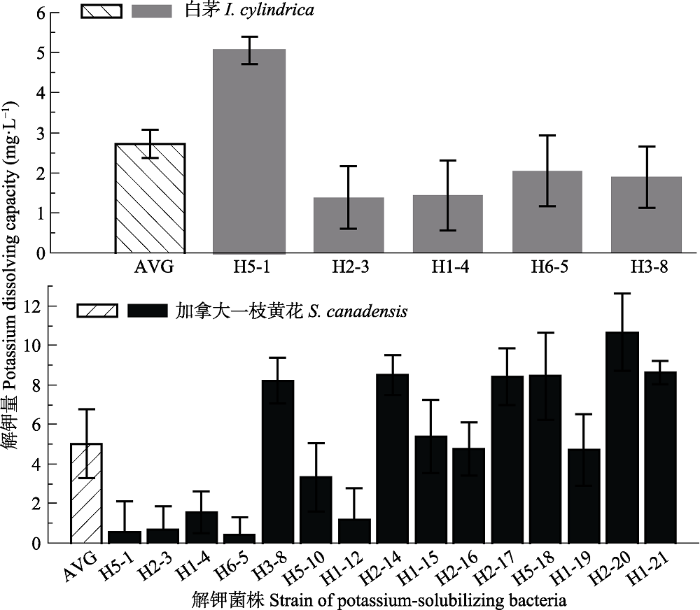 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}