周慧敏
1 , 2 , 李品
2 , 3 , 冯兆忠
, 3 , 4 , * , 张殷波
, 5 , * 1 山西大学黄土高原研究所, 太原 030006 2 中国科学院生态环境研究中心城市与区域生态国家重点实验室, 北京 100085 3 中国科学院大学资源环境学院, 北京 101408 4 南京信息工程大学生态研究院, 南京 210044 5 山西大学环境与资源学院, 太原 030006 Short-term effects of combined elevated ozone and limited irrigation on accumulation and allocation of non-structural carbohydrates in leaves and roots of poplar sapling Hui-Min ZHOU
1 , 2 , Pin LI
2 , 3 , Zhao-Zhong FENG
, 3 , 4 , * , Yin-Bo ZHANG
, 5 , * 1 Institute of Loess Plateau, Shanxi University, Taiyuan 030006, China 2 State Key Laboratory of Urban and Regional Ecology, Research Centre for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing 100085, China 3 College of Resources and Environment, University of Chinese Academy of Sciences, Beijing 101408, China 4 Institute of Ecology, Nanjing University of Information Sciences & Technology, Nanjing 210044, China; 5 College of Environmental Science and Resources, Shanxi University, Taiyuan 030006, China 通讯作者: * Feng ZZ:
zhaozhong.feng@nuist.edu.cn ;Zhang YB:
zhangyinbo@sxu.edu.cn 编委: 张守仁
责任编辑: 李敏
收稿日期: 2019-02-11
修回日期: 2019-04-17
网络出版日期: 2019-04-20
基金资助: 国家自然科学基金 (31870458 )国家自然科学基金 (41771034 )
Corresponding authors: * Feng ZZ:
zhaozhong.feng@nuist.edu.cn ;Zhang YB:
zhangyinbo@sxu.edu.cn Received: 2019-02-11
Revised: 2019-04-17
Online: 2019-04-20
Fund supported: Supported by the National Natural Science Foundation of China (31870458 )Supported by the National Natural Science Foundation of China (41771034 )
摘要 人类活动加剧和全球变化导致植物在生长季同时受到高浓度地表臭氧(O
3 )和干旱的双重胁迫。为了探究两者对植物非结构性碳水化合物(TNC)积累和分配的影响, 该实验采用开顶式气室研究了2种O
3 浓度(CF, 过滤空气; NF40, NF (未过滤空气) + 40 nmol·mol
-1 O
3 )和2个水分处理(对照, 充分灌溉; 干旱, 非充分灌溉)及其交互作用对杨树基因型‘546’ (
Populus deltoides cv. ‘55/56’ ×
P. deltoides cv. ‘Imperial’)叶片和细根中TNC及其组分(葡萄糖、果糖、蔗糖、多糖、总可溶性糖和淀粉)含量的影响。结果表明: O
3 浓度升高显著降低杨树叶片中淀粉和TNC的含量, 增加葡萄糖、果糖和总可溶性糖含量, 但对细根中淀粉和总可溶性糖含量的影响不显著。干旱胁迫显著增加细根中果糖和多糖含量, 降低蔗糖含量, 但对叶片中淀粉和总可溶性糖含量的影响不显著。充分灌溉下O
3 浓度升高显著增加了杨树叶片多糖和总可溶性糖含量, 而干旱下O
3 浓度升高显著增加了TNC含量的根叶比。该研究结果发现O
3 主要影响叶片中TNC及各组分的含量, 而干旱主要影响细根中TNC及各组分的含量。从杨树叶片TNC的响应来看, 适度的水分限制有助于减缓O
3 的负面伤害。
关键词: 臭氧 ;
干旱 ;
非结构性碳水化合物 ;
杨树 ;
细根 ;
叶片 Abstract Aims Intensified human activities has resulted in more extreme droughts and high ground-level ozone (O
3 ) concentration during plant growing seasons. To clarify the combined effects of elevated O
3 (E-O
3 ) and drought on plants, we investigated the effects of elevated O
3 , drought and their interaction on the accumulation and allocation of non-structural carbohydrates (TNC) in leaves and fine roots of poplar.
Methods The O
3 -sensitive poplar ‘546’ (
Populus deltoides cv. ‘55/56’ ×
P. deltoides cv. ‘Imperial’) saplings were exposed to two O
3 treatments (charcoal-filtered air (CF) and non-filtered ambient air (NF) + 40 nmol·mol
-1 O
3 (E-O
3 )), and two watering treatments (control, well-watered and drought, reduced watering) using open-top chambers (OTC
S ). The content of TNC and its components (glucose, fructose, sucrose, polysaccharides, total soluble sugars and starch) in leaves and fine roots were measured.
Important findings E-O
3 significantly decreased the content of starch and TNC, but increased the content of fructose, glucose and total soluble sugar in leaves. However, E-O
3 did not have significant effects on starch and soluble sugars in fine roots. In addition, drought significantly increased the content of fructose and polysaccharide, but decreased the content of sucrose in fine roots, although drought had no significant impacts on the content of starch and soluble sugars in leaves. There was significant interaction of O
3 and drought on the content of polysaccharides and total soluble sugar in leaves, as indicated by the significant increase by E-O
3 under well-watered irrigation, but no effects under drought. E-O
3 significantly increased the ratio of root to leaf in TNC content, but not under well water condition. In conclusion, E-O
3 affected the content of TNC and its components in leaves, while drought affected them in fine roots. From the response of TNC, moderate drought mitigates the negative effects of O
3 pollution on plants.
Keywords: O3 ;
drought ;
non-structural carbohydrate ;
poplar ;
fine root ;
leaf PDF (1211KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 周慧敏, 李品, 冯兆忠, 张殷波. 地表臭氧浓度升高与干旱交互作用对杨树非结构性碳水化合物积累和叶根分配的短期影响. 植物生态学报, 2019, 43(4): 296-304. DOI:
10.17521/cjpe.2019.0032 ZHOU Hui-Min, LI Pin, FENG Zhao-Zhong, ZHANG Yin-Bo.
Short-term effects of combined elevated ozone and limited irrigation on accumulation and allocation of non-structural carbohydrates in leaves and roots of poplar sapling .
Chinese Journal of Plant Ecology , 2019, 43(4): 296-304. DOI:
10.17521/cjpe.2019.0032 快速城市化进程导致地表臭氧(O
3 )浓度持续升高(
Cooper et al ., 2014 ), 并且高浓度O
3 主要发生在植物生长旺盛时期(5-9月)(
冯兆忠等, 2018 )。在世界上许多地方, 植物生长期间常伴随频繁的水分亏缺, 从而使得植物同时受到高浓度O
3 和干旱的双重胁迫(
Matyssek et al ., 2010 ;
高峰等, 2017 )。O
3 污染和干旱都会降低植物光饱和点光合速率, 影响植物的碳汇能力(
Zhang et al ., 2011 ;
Ainsworth et al ., 2012 ;
吴敏等, 2014 ;
Li et al ., 2018 ), 进而影响植物的生长发育(
高峰等, 2017 )。作为光合作用的主要产物, 碳水化合物主要包括结构性碳水化合物(SC)和非结构性碳水化合物(TNC)。TNC包括淀粉和总可溶性糖, 是树木生长代谢过程中重要的能量供应物质, 其浓度和含量的大小通常可以反映植物整体的碳供应状况, 体现植物阶段性碳获取和支出的平衡关系(
于丽敏等, 2011 ;
李娜妮等, 2016 )。其中, 淀粉主要用于储存物质和能量, 在植物生长、发育和繁殖等生命活动中起重要作用(
赵镭等, 2011 ); 总可溶性糖作为一种信号分子, 参与调节基因表达(
Rolland et al ., 2006 ), 并对植物体内糖代谢起到解耦联作用(
Pego et al ., 1999 )。总可溶性糖主要包括葡萄糖、果糖、蔗糖、多聚糖等。葡萄糖和果糖作为单糖直接供给植物能量, 但葡萄糖比果糖分解速度更快, 其能量释放也更快。蔗糖是植物光合作用的主要产物, 是同化产物由“源”向“库”运输的主要形式(
Farrar et al ., 2000 ), 蔗糖作为非还原性糖, 分解为果糖和葡萄糖后再向植物供应能量(
Quentin et al ., 2015 )。
近年来, TNC对环境因子胁迫的响应研究主要集中在高温(
李娜等, 2014 )、CO
2 浓度升高(
董彦红等, 2015 )、干旱(
杜尧等, 2014 )和氮磷添加(
王雪等, 2014 )等。大量文献报道了干旱胁迫对树木不同器官TNC含量的影响, 但结论并不一致。例如
Anacardium excelsum 、
Luehea seemannii 枝条的TNC含量在干旱季节大于湿润季节(
Newell et al ., 2002 );
Jacaratia mexicana 树干的TNC含量在湿润季节最高, 而
Spondias purpurea 树干的TNC含量在干旱与湿润季节无显著差异(
Bullock, 1992 )。另有研究表明, 高浓度O
3 减少了西黄松(
Pinus ponderosa )幼苗碳水化合物向根系的分配(
Andersen & Rygiewicz, 1995 )。但O
3 与干旱对树木TNC的复合影响却鲜有报道。
杨树(
Populus spp.)是我国北方平原地区防护林、用材林和城乡绿化的首选树种。目前, 我国杨树人工林面积已超1亿亩, 居世界之首。同时, 我国北方地区干燥少雨, 当前环境O
3 浓度已超过树木生长的受害临界水平(
Li et al ., 2018 )。本研究利用开顶式气室(OTCs), 研究O
3 和干旱对杨树叶片和细根TNC含量的复合影响, 以期为了解大气污染和气候变化下杨树的生长响应提供科学依据。
1 材料和方法 1.1 实验地点 实验地点位于北京市延庆区延庆镇唐家堡村(40.75° N, 116.62° E)。该地区属于温带大陆性季风气候, 年平均气温为10.2 ℃, 夏季平均气温为22-25 ℃, 年降水量为567 mm。
1.2 实验材料和实验设计 选择对O
3 敏感的一年生杨树幼苗‘546’ (
Populus deltoides cv. ‘55/56’
× P. deltoides cv. ‘Imperial’)(
Hu et al ., 2015 ;
Shang et al ., 2017 )为实验材料。2017年4月6日将育苗盒中长势较为一致的幼苗统一移入20 L的圆形花盆中, 用3 mm筛网过筛的当地棕色土壤与有机土等量充分混合作为栽培基质。5月7日, 将株高和基径基本一致的幼树移入OTCs中预适应10天后进行O
3 熏蒸。
OTC为高3.0 m, 边长为1.0 m的正八面柱体。实验设置2个O
3 处理: CF (活性炭过滤的空气, [O
3 ] < 40 nmol·mol
-1 )、NF40 (NF (非过滤的空气) + 40 nmol·mol
-1 O
3 )。每个O
3 处理设置3个OTC重复。每个OTC设置两个水分处理: 对照, 即充分灌溉(control, 叶片水势-1.3 MPa); 干旱, 即非充分灌溉(drought, 减水40%, 叶片水势-3.2 MPa)(
张婷等, 2016 ;
Shang et al ., 2019 ), 每个亚处理设有7-8株植株重复。O
3 熏蒸自2017年6月10日开始至9月22日结束, 每天熏蒸10 h (8:00-18:00), 阴雨天停止熏蒸。实验期间各OTC的O
3 浓度由美国热电公司49i臭氧分析仪监测(Model 49i, Thermo Scientific, Franklin, USA), OTC内部和外部的温度和湿度由温湿度传感器进行实时监测并利用CR1000数据采集器(Campbell Scientific, North Logan, USA)记录, 记录频次为5 min。实验期间CF和E-O
3 处理下的平均O
3 浓度分别为24 nmol·mol
-1 和81 nmol·mol
-1 , AOT40值(小时浓度超过40 nmol·mol
-1 时的累积O
3 浓度)分别为2.4 μmol·mol
-1 ·h和41.6 μmol·mol
-1 ·h。在O
3 熏蒸时间内, OTC内外的平均气温分别为22.4 ℃和20.5 ℃。
1.3 指标测定 于9月中下旬(生长末期), 每个OTC中随机选取4盆植株(2盆充分灌溉, 2盆干旱处理), 6个OTC共收集24株植物, 分叶片、茎、根收获, 并计算每株生物量。根用清水洗净后分为粗根(直径> 2 mm)和细根(直径< 2 mm)。叶片和细根在105 ℃下杀青15 min后, 在65 ℃下烘干24 h。将叶片和细根干样用球磨仪(GT200, 北京格瑞德曼仪器设备有限公司)粉碎后分别测定可溶性糖和淀粉含量, 总和即为TNC含量。
可溶性糖提取及测定: 分别称取0.5 g磨好的细根和叶片样品于锥形瓶中, 加入50 mL蒸馏水, 高压蒸煮2 h, 冷却至室温后于4 ℃, 15 000 r·min
-1 下离心10 min, 取上清液作为可溶性糖提取液。通过高效液相色谱法(HPLC)测定可溶性糖组分(葡萄糖、果糖、蔗糖、多糖), 其中测定系统使用Sugar-Pak 1色谱柱和折射率检测器(HPLC2695, Waters, Massachusetts, USA)。柱温为70 ℃, 蒸馏水用作移动液相(流速0.6 mL·min
-1 )(
Chen et al ., 2015 )。总可溶性糖含量为各个组分的总和。
淀粉提取及测定: 分别称取0.05 g和0.5 g叶片和细根样品置于10 mL离心管中, 加入1 mL蒸馏水和5 mL 80%乙醇溶液置于涡旋振荡器中混匀静置, 15 000 r·min
-1 下离心10 min, 弃上清液, 重复2次。于残留物中加入1 mL蒸馏水和6 mL 52%高氯酸溶液, 置于涡旋振荡器涡旋10 min, 15 000 r·min
-1 下离心10 min, 将上清液转入25 mL容量瓶, 重复2次, 用蒸馏水定容至25 mL。将该溶液进行过滤, 滤液转入50 mL容量瓶, 并用蒸馏水定容, 得到淀粉提取液, 采用高氯酸水解-蒽酮比色法测定淀粉含量(
田纪春, 2006 )。
1.4 数据分析 数据通过方差齐性及正态分布检验(
p > 0.05)。实验数据以OTC为重复单元(
n = 3)采用SPSS 18.0软件进行双因素(O
3 和干旱)方差分析, 检验两者主效应及交互效应的显著性。利用Tukey’s HSD进行多重比较,
p < 0.05则认为是处理间达到显著性差异。运用Origin 9.0软件绘图。
2 结果 2.1 臭氧与干旱对杨树TNC含量的影响 综合2个水分处理, O
3 浓度升高显著降低叶片TNC含量, 降低程度为6.67% (
p = 0.027;
表1 ), 而干旱对叶片TNC含量的影响不显著(
p > 0.05;
图1A )。O
3 和干旱对细根TNC含量均无显著影响(
p > 0.05;
图1B ;
表1 )。
Table 1 表1 表1 O
3 和干旱对杨树叶片和细根中非结构性碳水化合物(TNC)组分的方差分析结果
Table 1
Analysis of variance analysis of non-structural carbohydrates (TNC) components in leaves and fine roots of poplar by O3 and drought 臭氧3 干旱 臭氧×干旱3 × Drought 叶片 葡萄糖 Glucose 0.047 0.239 0.685 果糖 Fructose 0.004 0.563 0.394 蔗糖 Sucrose 0.217 0.384 0.995 多糖 Polysaccharide 0.128 0.227 0.039 总可溶性糖 0.003 0.449 0.009 淀粉 Starch 0.002 0.130 0.885 TNC 0.027 0.257 0.231 细根 葡萄糖 Glucose 0.546 0.282 0.980 果糖 Fructose 0.768 <0.001 0.459 蔗糖 Sucrose 0.970 0.002 0.455 多糖 Polysaccharide 0.575 0.025 0.245 总可溶性糖 0.494 0.168 0.157 淀粉 Starch 0.291 0.946 0.661 TNC 0.294 0.244 0.177 细根/叶片 总可溶性糖 0.193 0.427 0.013 淀粉 Starch 0.005 0.237 0.372 TNC 0.005 0.03 0.022
Data in the table are p value and bold number means p < 0.05. 表中数据为p 值,粗体表示p < 0.05。 新窗口打开 |
下载CSV 图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1臭氧浓度升高和干旱处理对杨树叶片(A)和细根(B)中非结构性碳水化合物(TNC)含量的影响(平均值±标准误差)。图中相同小写字母表示差异不显著(p > 0.05)。CF, 过滤空气; NF40, 未过滤空气+ 40 nmol·mol-1 O3 。 Fig. 1Effects of elevated ozone and drought on the total non-structural carbohydrates (TNC) content in leaves (A) and fine roots (B) of poplar saplings (mean ± SE ). The same lowercase letter in the figures stands for non-significant difference between treatments (p > 0.05). CF, charcoal-filtered air; NF40, non-filtered air + 40 nmol·mol-1 O3 . 2.2 臭氧与干旱对杨树叶片可溶性糖和淀粉含量的影响 O
3 浓度升高显著增加杨树叶片葡萄糖、果糖和总可溶性糖含量, 增加幅度分别为132.9%、83.8%和23% (
图2A 、2B、2E), 但显著降低叶片中的淀粉含量(
p = 0.002;
图2F ;
表1 )。与CF相比, NF40处理下的叶片淀粉含量降低了36.4%。干旱胁迫对叶片可溶性糖(
p = 0.449)和淀粉(
p = 0.130)含量的影响均不显著(
表1 )。O
3 与干旱对叶片多糖和总可溶性糖含量的影响有显著的交互作用(
p < 0.05), 灌溉下O
3 浓度升高显著增加了杨树叶片多糖(+18%)和总可溶性糖含量(+23%), 而在干旱胁迫下O
3 的影响不显著(
p > 0.05;
图2D 、2E)。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2臭氧浓度升高和干旱处理对叶片中葡萄糖、果糖、蔗糖、多糖、总可溶性糖和淀粉含量的影响(平均值±标准误差)。图中不同小写字母表示差异显著(p < 0.05)。CF, 过滤空气; NF40, 未过滤空气+ 40 nmol·mol-1 O3 。 Fig. 2Effects of ozone and drought on glucose, fructose, sucrose, polysaccharide, total soluble sugar and starch in leaves of poplar saplings (mean ± SE ). Different lowercase letters in the figures stand for the significant difference between treatments (p < 0.05). CF, charcoal-filtered air; NF40, non-filtered air + 40 nmol·mol-1 O3 . 2.3 臭氧与干旱对杨树细根可溶性糖和淀粉含量的影响 O
3 浓度升高对细根可溶性糖和淀粉含量均无显著性影响。干旱显著增加细根中果糖(
p < 0.001)和多糖含量(
p = 0.025;
表1 ), 但显著降低了蔗糖含量(
p = 0.002;
图3B -3D)。O
3 和干旱复合处理对杨树细根的可溶性糖(
p = 0.168)和淀粉(
p = 0.946)含量的影响不显著(
图3E 、3F)。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3臭氧浓度升高和干旱处理对细根中蔗糖、果糖、葡萄糖、多糖、总可溶性糖和淀粉含量的影响(平均值±标准误差)。图中不同小写字母表示差异显著(p < 0.05)。CF, 过滤空气; NF40, 未过滤空气+ 40 nmol·mol-1 O3 。 Fig. 3Effect of elevated ozone and drought on the glucose, fructose, sucrose, polysaccharide, total soluble sugar and starch in fine roots (mean ± SE ). Different letters in the figures stand for the significant difference between treatments (p < 0.05). CF, charcoal- filtered air; NF40, non-filtered air + 40 nmol·mol-1 O3 . 2.4 臭氧与干旱对杨树总糖、淀粉和TNC含量的根叶比的影响 O
3 浓度升高显著增加了杨树淀粉和TNC含量的根叶比(
表1 ;
图4B 、4C)。干旱显著增加TNC含量的根叶比。O
3 浓度升高和干旱对杨树总糖和TNC含量的根叶比存在显著的交互效应, 即充分灌溉下O
3 浓度升高显著降低总糖含量的根叶比, 但在干旱下无显著影响(
p = 0.013;
图4A ); 充分灌溉下O
3 浓度升高对淀粉含量的影响不显著, 但干旱胁迫下O
3 浓度升高显著增加了TNC含量的根叶比(
p = 0.022)。
图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4臭氧浓度升高和干旱处理对杨树叶片和细根总可溶性糖、淀粉和TNC含量的比值的影响(平均值±标准误差)。图中不同小写字母差异显著(p < 0.05)。CF, 过滤空气; NF40, 未过滤空气+ 40 nmol·mol-1 O3 。 Fig. 4Effects of elevated ozone and drought on the ratio of fine roots to leaves in total soluble sugar, starch and total non-structural carbohydrates of poplar saplings (mean ± SE ). Different letters in the figures stand for the significant difference between treatments (p < 0.05). CF, charcoal-filtered air; NF40, non-filtered air + 40 nmol·mol-1 O3 . 3 讨论 TNC是植物生长和代谢的主要能源物质, 其在植物体内含量的变化能反映植物的生理状况, 也反映植物对环境变化的动态响应。本实验结果表明, O
3 浓度升高显著降低了杨树叶片TNC含量, 这与之前O
3 对楠木(
Phoebe bournei )(
Chen et al ., 2015 )和春小麦(
Tritcium aestivum )(
Gelang et al ., 2011 )影响的结果一致, 可能是由于O
3 降低植物光合速率进而影响碳同化和TNC合成。
Topa等(2001) 研究发现, 经过3年的O
3 处理(比环境O
3 浓度高3倍), 五角枫(
Acer mono )叶片的TNC含量在春季显著降低, 但在夏季则受O
3 影响不显著, 表明在植物生长的不同阶段, O
3 对植物TNC含量的影响不同。本实验未发现干旱影响到杨树叶片的TNC含量, 这与之前的结果不一致, 即干旱有助于植物叶片内的TNC积累(
Tissue & Wright, 1995 ;
Pooter & Kitajima, 2007 ), 可能与本研究的实验设计有关。为了防止植物永久萎蔫, 实验期间采用非充分灌溉处理, 但显著降低了叶片水势(水势为-3.2 MPa), 表明本实验的植株的确受到了干旱的影响(
Shang et al ., 2019 )。同步进行的实验结果表明, 干旱胁迫降低了生物量(
Li et al ., 2019 ), 但没有降低植物的光合能力(
Shang et al ., 2019 ), 这可能是干旱胁迫未显著影响植物TNC含量的原因。
另一方面, 植物叶片可溶性糖和淀粉对于O
3 和干旱的响应不同。O
3 浓度升高显著增加杨树叶片中葡萄糖和果糖含量, 导致总可溶性糖含量升高, 但降低了淀粉含量, 这与对云杉(
Picea asperata )的研究结果一致(
Braun et al ., 2004 )。在高浓度的O
3 环境下, 植物会消耗体内大量的淀粉以抵御氧化胁迫, 因此导致储存的淀粉含量降低。葡萄糖是呼吸代谢的直接底物, 意味着高浓度的O
3 会促进叶片的呼吸代谢, 这与之前的实验结果一致(
Su et al ., 2017 )。干旱胁迫导致植物叶片中淀粉向可溶性糖转化, 调节植物叶片的渗透压, 增加叶片膨压(
Iannucci et al ., 2002 )。此外, 干旱胁迫下调了叶片淀粉合成能力, 导致淀粉含量降低(
Quick et al ., 1992 )。干旱胁迫对落叶松(
Larix gmelinii )影响的研究表明, 叶片内淀粉含量降低是因为淀粉水解成可溶性糖, 从而维持正常的生理活动(
杜尧等, 2014 )。但本研究结果显示, 干旱胁迫对植物叶片可溶性糖和淀粉含量均无显著影响, 可能是因为本研究采用长期轻度的干旱处理。为了避免植物产生极度干旱而导致水力结构去功能化, 因此在实验过程中给干旱植物提供节水灌溉, 从而形成了干旱复水的锻炼过程, 增加了叶片对干旱胁迫的抗性。O
3 和水分对植物叶片总可溶性糖和多糖含量存在显著的交互效应, 主要体现在干旱胁迫下O
3 对可溶性糖含量的影响不显著。干旱降低了气孔导度, 减少叶片的O
3 吸收通量, 从而减轻植物受到O
3 的伤害程度(
Gao et al ., 2017 )。
本研究结果还表明高浓度O
3 对杨树细根TNC含量的影响不显著。O
3 显著增加了楠木细根的TNC含量, 但对马尾松(
Pinus massoniana )细根的TNC含量没有影响(
Chen et al ., 2015 ), 说明不同植物的细根对O
3 胁迫的响应存在差异, 也说明不同植物应对O
3 胁迫的生存策略有所差异。在高浓度的O
3 环境下, 碳供应和碳需求之间的不协调和不平衡, 导致植物将光合产物转移用于抵御逆境的储备库而不是用于生长(
Galvez et al ., 2011 )。另一方面, O
3 和干旱胁迫对植物细根的淀粉含量无显著影响, 这与前人的研究结果有所差异。
Chen等(2015) 研究表明O
3 浓度升高显著增加了楠木细根中的淀粉含量;
Adams等(2013) 发现从干旱胁迫开始至树木死亡的过程中树木细根的淀粉含量在降低;
肖列等(2017) 发现随着干旱胁迫的加剧, 白羊草(
Bothriochloa ischaemum )根系淀粉含量显著下降。此外, 干旱胁迫显著增加了细根多糖和果糖的含量而降低蔗糖含量, 总体上增加了可溶性糖含量, 这与对黄瓜(
Cucumis sativus )、白羊草的研究结果是一致的(
董彦红等, 2015 ;
肖列等, 2017 )。通过可溶性糖的增加能增加渗透调节物质的浓度, 从而保持细胞中的水分不流失, 这对干旱胁迫下树木的生长来说是一种节省能量维持正常生理代谢的适应策略。由此可见, 不同树种细根的TNC组分对O
3 和干旱的响应不同, 可能与实验O
3 熏蒸的持续时间、累积的O
3 剂量, 干旱的胁迫程度以及树种不同有关。
O
3 浓度升高显著增加了淀粉和TNC含量的根叶比, 原因是其显著降低了叶片中的淀粉和TNC含量而对细根没有影响。为了抵御O
3 胁迫, 叶片中的淀粉转换成可溶性糖, 增加呼吸代谢。但是植物细根的TNC主要来源于叶片光合作用形成的碳水化合物, 因此O
3 浓度升高对细根TNC含量的影响是间接的。尽管高O
3 环境下, 通过韧皮部运输到细根的TNC含量是降低的, 但用于结构生长的碳含量也显著降低(
Li et al ., 2019 )。有趣的是, 只有在水分亏缺下O
3 浓度升高对TNC含量根叶比的影响是显著的。干旱胁迫未影响到叶片和细根中的TNC含量, 但显著增加了TNC含量的根叶比。干旱胁迫导致植物的根冠比升高, 将更多的碳分配到了地下, 但TNC参与根系结构生长。因此, 干旱增加了TNC含量的根叶比, 有助于根系生长, 并增加了根系与土壤的接触面积, 进而提高了水分吸收能力。
值得注意的是本研究是基于OTC的短期模拟实验观测O
3 浓度增加和干旱胁迫对杨树叶片和细根TNC及其组分含量的影响, 并不能囊括在更长时间胁迫下植物的响应情况。另外, 本研究发现虽然在干旱胁迫下, 杨树叶片和细根的TNC含量无明显变化, 但是可溶性糖含量整体增加而淀粉含量是下降的, 这可能是由可溶性糖和淀粉的相互转化引起的。在本试验中O
3 与干旱胁迫对总可溶性糖和淀粉无显著交互作用, 这可能与实验设置的O
3 浓度和干旱胁迫水平有关。因此, 在今后研究中, 设置多梯度胁迫实验对于观察杨树叶片和细根的响应动态是必需的。
综上所述, O
3 浓度升高和干旱胁迫对杨树TNC及其组分含量产生显著影响。O
3 浓度升高显著增加了杨树叶片葡萄糖、果糖和总糖含量, 降低了淀粉和TNC含量。干旱胁迫显著增加了细根中果糖和多糖的含量, 但显著降低了蔗糖的含量。O
3 污染显著增加了杨树细根与叶片淀粉和TNC含量的比值。在干旱胁迫下, O
3 浓度升高显著降低了总可溶性糖含量的根叶比, 但显著提高了TNC含量的根叶比。本研究结果有望为O
3 污染和干旱等共同胁迫作用影响植物生长发育过程提供科学依据。
[1] Adams HD Germino MJ Breshears DD Gafford GB Guardiola M Zou CB Huxman TE 2013 ). Nonstructural leaf carbohydrate dynamics of Pinus edulis during drought- induced tree mortality reveal role for carbon metabolism in mortality mechanismNew Phytologist 1142 -1151 . [本文引用: 1] [2] Ainsworth EA Yendrek CR Sitch S Collins WJ Emberson LD 2012 ). The effects of tropospheric ozone on net primary productivity and implications for climate changeAnnual Review of Plant Biology, 637 -661 . [本文引用: 1] [3] Andersen CP Rygiewicz PT 1995 ). Allocation of carbon and mychorrhizal Pinus ponderosa seedlings exposed to ozoneNew Phytologist, 471 -478 . [本文引用: 1] [4] Braun S Zugmaier U Thomas V Fluckiger W 2004 ). Carbohydrate concentrations in different plant parts of young beech and spruce along a gradient of ozone pollutionAtmospheric Environment , 38, 2399 -2407 . [本文引用: 1] [5] Bullock SH 1992 ). Seasonal differences in non-structural carbohydrates in two dioecious monsoon-climate treesBiotropica , 24, 140 -145 . [本文引用: 1] [6] Chen Z Shang H Cao J Yu H 2015 ). Effects of ambient ozone concentrations on contents of nonstructural carbohydrates in Phoebe bournei and Pinus massoniana seedlings in subtropical ChinaWater, Air, & Soil Pollution 310 . [本文引用: 4] [7] Cooper OR Parrish DD Ziemke J Balashov NV Cupeiro M Galbally IE Gilge S Horowitz L Jensen NR Lamarque JF Naik V Oltmans SJ Schwab J Shindell DT Thompson AM Thouret V Wang Y Zbinden RM 2014 ). Global distribution and trends of tropospheric ozone: An observation- based reviewElementa: Science of the Anthropocene , 2, 000029 . DOI: 10.12952/journal.elementa.000029 . [本文引用: 1] [8] Dong YH Liu BB Zhang X Liu XN Ai XZ Li QM 2015 ). Responses of non-structural carbohydrate metabolism of cucumber seedlings to drought stress and doubled CO2 concentrationChinese Journal of Applied Ecology , 26, 53 -60 . [本文引用: 2] [ 董彦红 , 刘彬彬 , 张旭 , 刘学娜 , 艾希珍 , 李清明 ( 2015 ). 黄瓜幼苗非结构性碳水化合物代谢对干旱胁迫与CO2 倍增的响应应用生态学报 , 26, 53 -60 .] [本文引用: 2] [9] Du Y Han Y Wang CK 2014 ). The influence of drought on non-structural carbohydrates in the needles and twigs of Larix gmelinii Acta Ecologica Sinica 6090 -6100 . [本文引用: 2] [ 杜尧 , 韩轶 , 王传宽 ( 2014 ). 干旱对兴安落叶松枝叶非结构性碳水化合物的影响生态学报 , 34, 6090 -6100 .] [本文引用: 2] [10] Farrar J Pollock C Gallagher J 2000 ). Sucrose and the integration of metabolism in vascular plantsPlant Science , 154, 1 -11 . [本文引用: 1] [11] Feng ZZ Li P Yuan XY Gao F Jiang LJ Dai LL 2018 ). Progress in ecological and environmental effects of ground- level O3 in ChinaActa Ecologica Sinica , 38, 1530 -1541 . [本文引用: 1] [ 冯兆忠 , 李品 , 袁相洋 , 高峰 , 姜立军 , 代碌碌 ( 2018 ). 我国地表臭氧的生态环境效应研究进展生态学报 , 38, 1530 -1541 .] [本文引用: 1] [12] Galvez DA Landh?usser SM Tyree MT 2011 ). Root carbon reserve dynamics in aspen seedlings: Does simulated drought induce reserve limitation?Tree Physiology , 31, 250 -257 . [本文引用: 1] [13] Gao F Catalayud V Paoletti E Hoshika Y Feng ZZ 2017 ). Water stress mitigates the negative effects of ozone on photosynthesis and biomass in poplar plantsEnvironmental Pollution , 230, 268 -279 . [本文引用: 1] [14] Gao F Li P Feng ZZ 2017 ). Interactive effects of ozone and drought stress on plants: A reviewChinese Journal of Plant Ecology , 41, 252 -268 . [本文引用: 2] [ 高峰 , 李品 , 冯兆忠 ( 2017 ). 臭氧与干旱对植物复合影响的研究进展植物生态学报 , 41, 252 -268 .] [本文引用: 2] [15] Gelang J Selldén G Younis S Pleijel H 2011 ). Effects of ozone on biomass, non-structural carbohydrates and nitrogen in spring wheat with artificially manipulated source/ sink ratioEnvironmental and Experimental Botany , 46, 155 -169 . [本文引用: 1] [16] Hu EZ Gao F Xin Y Jia HX Li KH Hu JJ Feng ZZ 2015 ). Concentration- and flux-based ozone dose-response relationships for five poplar clones grown in North ChinaEnvironmental Pollution , 207, 21 -30 . [本文引用: 1] [17] Iannucci A Russo M Arena L Di FN Martiniello P 2002 ). Water deficit effects on osmotic adjustment and solute accumulation in leaves of annual cloversEuropean Journal of Agronomy , 16, 111 -122 . [本文引用: 1] [18] Li N Sun T Mao ZJ 2014 ). Effects of long-term high- temperature stress on the biomass and non-structure carbohydrates of Pinus sylvestris varmongolica seedlings. Bulletin of Botanical Research 212 -218 . [本文引用: 1] [ 李娜 , 孙涛 , 毛子军 ( 2014 ). 长期极端高温胁迫对樟子松幼苗生物量及非结构性碳水化合物的影响植物研究 , 34, 212 -218 .] [本文引用: 1] [19] Li NN He NP Yu GR 2016 ). Evaluation of leaf non-structural carbohydrate contents in typical forest ecosystems in northeast ChinaActa Ecologica Sinica , 36, 430 -438 . [本文引用: 1] [ 李娜妮 , 何念鹏 , 于贵瑞 ( 2016 ). 中国东北典型森林生态系统植物叶片的非结构性碳水化合物研究生态学报 , 36, 430 -438 .] [本文引用: 1] [20] Li P de Marco A Feng ZZ Anav A Zhou DJ Paoletti E 2018 ). Nationwide ground-level ozone measurements in China suggest serious risks to forestsEnvironmental Pollution , 237, 803 -813 . [本文引用: 2] [21] Li P Zhou HM Xu YS Shang B Feng ZZ 2019 ). Effects of elevated ozone on biomass allocation and non-structural carbohydrates in poplar are strongly related with water and nitrogen availabilityScience of the Total Environment , 665, 929 -936 . [本文引用: 2] [22] Matyssek R Wieser G Ceulemans R Rennenberg H Pretzsch H Haberer K Loew M Nunn AJ Werner H Wipfler P Osswaldg W Nikolova P Hanke DE Kraigher H Tausz M Bahnweg G Kitao M Dieler J Sandermann H Herbinger K Grebenc T Blumenroether M Deckmyn G Grams TEE Heerdt C Leuchner M Fabian P Haeberle KH 2010 ). Enhanced ozone strongly reduces carbon sink strength of adult beech (Fagus sylvatica )—Resume from the free-air fumigation study at Kranzberg forestEnvironmental Pollution 2527 -2532 . [本文引用: 1] [23] Newell EA Mulkey SS Wright SJ 2002 ). Seasonal patterns of carbohydrate storage in four tropical tree speciesOecologia , 131, 333 -342 . [本文引用: 1] [24] Pego JV Weisbeek PJ Smeekens SC 1999 ). Mannose inhibits Arabidopsis germination via a hexokinase-mediated stepPlant Physiology 1017 -1024 . [本文引用: 1] [25] Poorter L Kitajima K 2007 ). Carbohydrate storage and light requirements of tropical moist and dry forest tree speciesEcology , 88, 1000 -1011 . [本文引用: 1] [26] Quentin AG Pinkard EA Ryan MG Tissue DT Baggett LS Admas HD Maillard P Marchand J 2015 ). Non-structural carbohydrates in woody plants compared among laboratoriesTree Physiology , 35, 1146 . [本文引用: 1] [27] Quick WP Chaves MM Wendler R David M Rodrigues ML Passaharinho JA Pereira JS Adcock MD Leegood RC Stitt M 1992 ). The effect of water stress on photosynthetic carbon metabolism in four species grown under field conditionsPlant, Cell & Environment , 15, 25 -35 . [本文引用: 1] [28] Rolland F Baena-Gonzalez E Sheen J 2006 ). Sugar sensing and signaling in plants: Conserved and novel mechanismsAnnual Review of Plant Biology , 57, 675 -709 . [本文引用: 1] [29] Shang B Feng ZZ Li P Yuan XY Xu YS Calatayud V 2017 ). Ozone exposure- and flux-based response relationships with photosynthesis, leaf morphology and biomass in two poplar clonesScience of the Total Environment 185 -195 . [本文引用: 1] [30] Shang B Yuan XY Li P Xu YS Feng ZZ 2019 ). Effects of elevated ozone and water deficit on poplar saplings: Changes in carbon and nitrogen stocks and their allocation to different organsForest Ecology and Management , 441, 89 -98 . [本文引用: 3] [31] Su B Zhou M Xu H Zhang X Li Y Su H Xiang B 2017 ). Photosynthesis and biochemical responses to elevated O3 in Plantago major and Sonchus oleraceus growing in a lowland habitat of northern ChinaJournal of Environmental Sciences 113 -121 . [本文引用: 1] [32] Tian JC 2006 ). Grain Quality Testing Theory and Methods . Science Press, Beijing. 71 -182 . [本文引用: 1] [ 田纪春 (2006 ). 谷物品质测试理论与方法 . 科学出版社, 北京. 71 -182 .] [本文引用: 1] [33] Tissue DT Wright SJ 1995 ). Effect of seasonal water availability on phenology and the annual shoot carbohydrate cycle of tropical forest shrubsFunctional Ecology , 9, 518 -527 . [本文引用: 1] [34] Topa MA Vanderklein DW Corbin A 2001 ). Effects of elevated ozone and low light on diurnal and seasonal carbon gain in sugar maplePlant, Cell & Environment , 24, 663 -677 . [本文引用: 1] [35] Wang X Luo WT Yu Q Yan CF Xu ZW Li MH Jiang Y 2014 ). Effects of nutrient addition on nitrogen, phosphorus and non-structural carbohydrates concentrations in leaves of dominant plant species in a semiarid steppeChinese Journal of Ecology , 33, 1795 -1802 . [本文引用: 1] [ 王雪 , 雒文涛 , 庾强 , 闫彩凤 , 徐柱文 , 李迈和 , 姜勇 (2014 ). 半干旱典型草原养分添加对优势物种叶片氮磷及非结构性碳水化合物含量的影响生态学杂志 , 33, 1795 -1802 .] [本文引用: 1] [36] Wu M Zhang WH Zhou JY Ma C Han WJ 2014 ). Effects of drought stress on growth, physiological and biochemical parameters in fine roots of Quercus variabilis Bl. seedlingsActa Ecologica Sinica 4223 -4233 . [本文引用: 1] [ 吴敏 , 张文辉 , 周建云 , 马闯 , 韩文娟 ( 2014 ). 干旱胁迫对栓皮栎幼苗细根的生长与生理生化指标的影响生态学报 , 34, 4223 -4233 .] [本文引用: 1] [37] Xiao L Liu GB Li P Xue S 2017 ). Responses of photosynthesis and non-structural carbohydrates of Bothriochloa ischaemum to doubled CO2 concentration and drought stressJournal of Plant Nutrition and Fertilizer 389 -397 . [本文引用: 2] [ 肖列 , 刘国彬 , 李鹏 , 薛萐 ( 2017 ). 白羊草光合特性及非结构性碳水化合物含量对CO2 浓度倍增和干旱胁迫的响应植物营养与肥料学报 , 23, 389 -397 .] [本文引用: 2] [38] Yu LM Wang CK Wang XC 2011 ). Allocation of nonstructural carbohydrates for three temperate tree species in Northeast ChinaChinese Journal of Plant Ecology , 35, 1245 -1255 . [本文引用: 1] [ 于丽敏 , 王传宽 , 王兴昌 ( 2011 ). 三种温带树种非结构性碳水化合物的分配植物生态学报 , 35, 1245 -1255 .] [本文引用: 1] [39] Zhao L Yang HB Wang DL Zhang N Wang XH 2011 ). Seedlings photosynthesis traits and non-structural carbohydrate storage of common species in Tiantong National Forest Park, Zhejiang ProvinceJournal of East China Normal University (Natural Science )4 ), 35 -44 . [本文引用: 1] [ 赵镭 , 杨海波 , 王达力 , 张娜 , 王希华 ( 2011 ). 浙江天童常见种幼苗的光合特性及非结构性碳水化合物储存华东师范大学学报(自然科学版) , 7(4 ), 35 -44 .] [本文引用: 1] [40] Zhang T Cao Y Chen YM Liu GB 2016 ). Effects of drought stress on non-structural carbohydrates of Robinia pseudoacacia saplings at the end of the growing seasonJournal of Soil and Water Conservation 297 -304 . [本文引用: 1] [ 张婷 , 曹扬 , 陈云明 , 刘国彬 ( 2016 ). 生长季末期干旱胁迫对刺槐幼苗非结构性碳水化合物的影响水土保持学报 , 30, 297 -304 .] [本文引用: 1] [41] Zhang YJ Xie ZK Wang YJ Su PX An LP Gao H 2011 ). Effect of water stress on leaf photosynthesis, chlorophyll content, and growth of oriental lilyRussian Journal of Plant Physiology , 58, 844 -850 . [本文引用: 1] Nonstructural leaf carbohydrate dynamics of
Pinus edulis during drought- induced tree mortality reveal role for carbon metabolism in mortality mechanism
1
2013
... 本研究结果还表明高浓度O
3 对杨树细根TNC含量的影响不显著.O
3 显著增加了楠木细根的TNC含量, 但对马尾松(
Pinus massoniana )细根的TNC含量没有影响(
Chen et al ., 2015 ), 说明不同植物的细根对O
3 胁迫的响应存在差异, 也说明不同植物应对O
3 胁迫的生存策略有所差异.在高浓度的O
3 环境下, 碳供应和碳需求之间的不协调和不平衡, 导致植物将光合产物转移用于抵御逆境的储备库而不是用于生长(
Galvez et al ., 2011 ).另一方面, O
3 和干旱胁迫对植物细根的淀粉含量无显著影响, 这与前人的研究结果有所差异.
Chen等(2015) 研究表明O
3 浓度升高显著增加了楠木细根中的淀粉含量;
Adams等(2013) 发现从干旱胁迫开始至树木死亡的过程中树木细根的淀粉含量在降低;
肖列等(2017) 发现随着干旱胁迫的加剧, 白羊草(
Bothriochloa ischaemum )根系淀粉含量显著下降.此外, 干旱胁迫显著增加了细根多糖和果糖的含量而降低蔗糖含量, 总体上增加了可溶性糖含量, 这与对黄瓜(
Cucumis sativus )、白羊草的研究结果是一致的(
董彦红等, 2015 ;
肖列等, 2017 ).通过可溶性糖的增加能增加渗透调节物质的浓度, 从而保持细胞中的水分不流失, 这对干旱胁迫下树木的生长来说是一种节省能量维持正常生理代谢的适应策略.由此可见, 不同树种细根的TNC组分对O
3 和干旱的响应不同, 可能与实验O
3 熏蒸的持续时间、累积的O
3 剂量, 干旱的胁迫程度以及树种不同有关. ...
The effects of tropospheric ozone on net primary productivity and implications for climate change
1
2012
... 快速城市化进程导致地表臭氧(O
3 )浓度持续升高(
Cooper et al ., 2014 ), 并且高浓度O
3 主要发生在植物生长旺盛时期(5-9月)(
冯兆忠等, 2018 ).在世界上许多地方, 植物生长期间常伴随频繁的水分亏缺, 从而使得植物同时受到高浓度O
3 和干旱的双重胁迫(
Matyssek et al ., 2010 ;
高峰等, 2017 ).O
3 污染和干旱都会降低植物光饱和点光合速率, 影响植物的碳汇能力(
Zhang et al ., 2011 ;
Ainsworth et al ., 2012 ;
吴敏等, 2014 ;
Li et al ., 2018 ), 进而影响植物的生长发育(
高峰等, 2017 ).作为光合作用的主要产物, 碳水化合物主要包括结构性碳水化合物(SC)和非结构性碳水化合物(TNC).TNC包括淀粉和总可溶性糖, 是树木生长代谢过程中重要的能量供应物质, 其浓度和含量的大小通常可以反映植物整体的碳供应状况, 体现植物阶段性碳获取和支出的平衡关系(
于丽敏等, 2011 ;
李娜妮等, 2016 ).其中, 淀粉主要用于储存物质和能量, 在植物生长、发育和繁殖等生命活动中起重要作用(
赵镭等, 2011 ); 总可溶性糖作为一种信号分子, 参与调节基因表达(
Rolland et al ., 2006 ), 并对植物体内糖代谢起到解耦联作用(
Pego et al ., 1999 ).总可溶性糖主要包括葡萄糖、果糖、蔗糖、多聚糖等.葡萄糖和果糖作为单糖直接供给植物能量, 但葡萄糖比果糖分解速度更快, 其能量释放也更快.蔗糖是植物光合作用的主要产物, 是同化产物由“源”向“库”运输的主要形式(
Farrar et al ., 2000 ), 蔗糖作为非还原性糖, 分解为果糖和葡萄糖后再向植物供应能量(
Quentin et al ., 2015 ). ...
Allocation of carbon and mychorrhizal
Pinus ponderosa seedlings exposed to ozone
1
1995
... 近年来, TNC对环境因子胁迫的响应研究主要集中在高温(
李娜等, 2014 )、CO
2 浓度升高(
董彦红等, 2015 )、干旱(
杜尧等, 2014 )和氮磷添加(
王雪等, 2014 )等.大量文献报道了干旱胁迫对树木不同器官TNC含量的影响, 但结论并不一致.例如
Anacardium excelsum 、
Luehea seemannii 枝条的TNC含量在干旱季节大于湿润季节(
Newell et al ., 2002 );
Jacaratia mexicana 树干的TNC含量在湿润季节最高, 而
Spondias purpurea 树干的TNC含量在干旱与湿润季节无显著差异(
Bullock, 1992 ).另有研究表明, 高浓度O
3 减少了西黄松(
Pinus ponderosa )幼苗碳水化合物向根系的分配(
Andersen & Rygiewicz, 1995 ).但O
3 与干旱对树木TNC的复合影响却鲜有报道. ...
Carbohydrate concentrations in different plant parts of young beech and spruce along a gradient of ozone pollution
1
2004
... 另一方面, 植物叶片可溶性糖和淀粉对于O
3 和干旱的响应不同.O
3 浓度升高显著增加杨树叶片中葡萄糖和果糖含量, 导致总可溶性糖含量升高, 但降低了淀粉含量, 这与对云杉(
Picea asperata )的研究结果一致(
Braun et al ., 2004 ).在高浓度的O
3 环境下, 植物会消耗体内大量的淀粉以抵御氧化胁迫, 因此导致储存的淀粉含量降低.葡萄糖是呼吸代谢的直接底物, 意味着高浓度的O
3 会促进叶片的呼吸代谢, 这与之前的实验结果一致(
Su et al ., 2017 ).干旱胁迫导致植物叶片中淀粉向可溶性糖转化, 调节植物叶片的渗透压, 增加叶片膨压(
Iannucci et al ., 2002 ).此外, 干旱胁迫下调了叶片淀粉合成能力, 导致淀粉含量降低(
Quick et al ., 1992 ).干旱胁迫对落叶松(
Larix gmelinii )影响的研究表明, 叶片内淀粉含量降低是因为淀粉水解成可溶性糖, 从而维持正常的生理活动(
杜尧等, 2014 ).但本研究结果显示, 干旱胁迫对植物叶片可溶性糖和淀粉含量均无显著影响, 可能是因为本研究采用长期轻度的干旱处理.为了避免植物产生极度干旱而导致水力结构去功能化, 因此在实验过程中给干旱植物提供节水灌溉, 从而形成了干旱复水的锻炼过程, 增加了叶片对干旱胁迫的抗性.O
3 和水分对植物叶片总可溶性糖和多糖含量存在显著的交互效应, 主要体现在干旱胁迫下O
3 对可溶性糖含量的影响不显著.干旱降低了气孔导度, 减少叶片的O
3 吸收通量, 从而减轻植物受到O
3 的伤害程度(
Gao et al ., 2017 ). ...
Seasonal differences in non-structural carbohydrates in two dioecious monsoon-climate trees
1
1992
... 近年来, TNC对环境因子胁迫的响应研究主要集中在高温(
李娜等, 2014 )、CO
2 浓度升高(
董彦红等, 2015 )、干旱(
杜尧等, 2014 )和氮磷添加(
王雪等, 2014 )等.大量文献报道了干旱胁迫对树木不同器官TNC含量的影响, 但结论并不一致.例如
Anacardium excelsum 、
Luehea seemannii 枝条的TNC含量在干旱季节大于湿润季节(
Newell et al ., 2002 );
Jacaratia mexicana 树干的TNC含量在湿润季节最高, 而
Spondias purpurea 树干的TNC含量在干旱与湿润季节无显著差异(
Bullock, 1992 ).另有研究表明, 高浓度O
3 减少了西黄松(
Pinus ponderosa )幼苗碳水化合物向根系的分配(
Andersen & Rygiewicz, 1995 ).但O
3 与干旱对树木TNC的复合影响却鲜有报道. ...
Effects of ambient ozone concentrations on contents of nonstructural carbohydrates in
Phoebe bournei and
Pinus massoniana seedlings in subtropical China
4
2015
... 可溶性糖提取及测定: 分别称取0.5 g磨好的细根和叶片样品于锥形瓶中, 加入50 mL蒸馏水, 高压蒸煮2 h, 冷却至室温后于4 ℃, 15 000 r·min
-1 下离心10 min, 取上清液作为可溶性糖提取液.通过高效液相色谱法(HPLC)测定可溶性糖组分(葡萄糖、果糖、蔗糖、多糖), 其中测定系统使用Sugar-Pak 1色谱柱和折射率检测器(HPLC2695, Waters, Massachusetts, USA).柱温为70 ℃, 蒸馏水用作移动液相(流速0.6 mL·min
-1 )(
Chen et al ., 2015 ).总可溶性糖含量为各个组分的总和. ...
... TNC是植物生长和代谢的主要能源物质, 其在植物体内含量的变化能反映植物的生理状况, 也反映植物对环境变化的动态响应.本实验结果表明, O
3 浓度升高显著降低了杨树叶片TNC含量, 这与之前O
3 对楠木(
Phoebe bournei )(
Chen et al ., 2015 )和春小麦(
Tritcium aestivum )(
Gelang et al ., 2011 )影响的结果一致, 可能是由于O
3 降低植物光合速率进而影响碳同化和TNC合成.
Topa等(2001) 研究发现, 经过3年的O
3 处理(比环境O
3 浓度高3倍), 五角枫(
Acer mono )叶片的TNC含量在春季显著降低, 但在夏季则受O
3 影响不显著, 表明在植物生长的不同阶段, O
3 对植物TNC含量的影响不同.本实验未发现干旱影响到杨树叶片的TNC含量, 这与之前的结果不一致, 即干旱有助于植物叶片内的TNC积累(
Tissue & Wright, 1995 ;
Pooter & Kitajima, 2007 ), 可能与本研究的实验设计有关.为了防止植物永久萎蔫, 实验期间采用非充分灌溉处理, 但显著降低了叶片水势(水势为-3.2 MPa), 表明本实验的植株的确受到了干旱的影响(
Shang et al ., 2019 ).同步进行的实验结果表明, 干旱胁迫降低了生物量(
Li et al ., 2019 ), 但没有降低植物的光合能力(
Shang et al ., 2019 ), 这可能是干旱胁迫未显著影响植物TNC含量的原因. ...
... 本研究结果还表明高浓度O
3 对杨树细根TNC含量的影响不显著.O
3 显著增加了楠木细根的TNC含量, 但对马尾松(
Pinus massoniana )细根的TNC含量没有影响(
Chen et al ., 2015 ), 说明不同植物的细根对O
3 胁迫的响应存在差异, 也说明不同植物应对O
3 胁迫的生存策略有所差异.在高浓度的O
3 环境下, 碳供应和碳需求之间的不协调和不平衡, 导致植物将光合产物转移用于抵御逆境的储备库而不是用于生长(
Galvez et al ., 2011 ).另一方面, O
3 和干旱胁迫对植物细根的淀粉含量无显著影响, 这与前人的研究结果有所差异.
Chen等(2015) 研究表明O
3 浓度升高显著增加了楠木细根中的淀粉含量;
Adams等(2013) 发现从干旱胁迫开始至树木死亡的过程中树木细根的淀粉含量在降低;
肖列等(2017) 发现随着干旱胁迫的加剧, 白羊草(
Bothriochloa ischaemum )根系淀粉含量显著下降.此外, 干旱胁迫显著增加了细根多糖和果糖的含量而降低蔗糖含量, 总体上增加了可溶性糖含量, 这与对黄瓜(
Cucumis sativus )、白羊草的研究结果是一致的(
董彦红等, 2015 ;
肖列等, 2017 ).通过可溶性糖的增加能增加渗透调节物质的浓度, 从而保持细胞中的水分不流失, 这对干旱胁迫下树木的生长来说是一种节省能量维持正常生理代谢的适应策略.由此可见, 不同树种细根的TNC组分对O
3 和干旱的响应不同, 可能与实验O
3 熏蒸的持续时间、累积的O
3 剂量, 干旱的胁迫程度以及树种不同有关. ...
... 和干旱胁迫对植物细根的淀粉含量无显著影响, 这与前人的研究结果有所差异.
Chen等(2015) 研究表明O
3 浓度升高显著增加了楠木细根中的淀粉含量;
Adams等(2013) 发现从干旱胁迫开始至树木死亡的过程中树木细根的淀粉含量在降低;
肖列等(2017) 发现随着干旱胁迫的加剧, 白羊草(
Bothriochloa ischaemum )根系淀粉含量显著下降.此外, 干旱胁迫显著增加了细根多糖和果糖的含量而降低蔗糖含量, 总体上增加了可溶性糖含量, 这与对黄瓜(
Cucumis sativus )、白羊草的研究结果是一致的(
董彦红等, 2015 ;
肖列等, 2017 ).通过可溶性糖的增加能增加渗透调节物质的浓度, 从而保持细胞中的水分不流失, 这对干旱胁迫下树木的生长来说是一种节省能量维持正常生理代谢的适应策略.由此可见, 不同树种细根的TNC组分对O
3 和干旱的响应不同, 可能与实验O
3 熏蒸的持续时间、累积的O
3 剂量, 干旱的胁迫程度以及树种不同有关. ...
Global distribution and trends of tropospheric ozone: An observation- based review
1
2014
... 快速城市化进程导致地表臭氧(O
3 )浓度持续升高(
Cooper et al ., 2014 ), 并且高浓度O
3 主要发生在植物生长旺盛时期(5-9月)(
冯兆忠等, 2018 ).在世界上许多地方, 植物生长期间常伴随频繁的水分亏缺, 从而使得植物同时受到高浓度O
3 和干旱的双重胁迫(
Matyssek et al ., 2010 ;
高峰等, 2017 ).O
3 污染和干旱都会降低植物光饱和点光合速率, 影响植物的碳汇能力(
Zhang et al ., 2011 ;
Ainsworth et al ., 2012 ;
吴敏等, 2014 ;
Li et al ., 2018 ), 进而影响植物的生长发育(
高峰等, 2017 ).作为光合作用的主要产物, 碳水化合物主要包括结构性碳水化合物(SC)和非结构性碳水化合物(TNC).TNC包括淀粉和总可溶性糖, 是树木生长代谢过程中重要的能量供应物质, 其浓度和含量的大小通常可以反映植物整体的碳供应状况, 体现植物阶段性碳获取和支出的平衡关系(
于丽敏等, 2011 ;
李娜妮等, 2016 ).其中, 淀粉主要用于储存物质和能量, 在植物生长、发育和繁殖等生命活动中起重要作用(
赵镭等, 2011 ); 总可溶性糖作为一种信号分子, 参与调节基因表达(
Rolland et al ., 2006 ), 并对植物体内糖代谢起到解耦联作用(
Pego et al ., 1999 ).总可溶性糖主要包括葡萄糖、果糖、蔗糖、多聚糖等.葡萄糖和果糖作为单糖直接供给植物能量, 但葡萄糖比果糖分解速度更快, 其能量释放也更快.蔗糖是植物光合作用的主要产物, 是同化产物由“源”向“库”运输的主要形式(
Farrar et al ., 2000 ), 蔗糖作为非还原性糖, 分解为果糖和葡萄糖后再向植物供应能量(
Quentin et al ., 2015 ). ...
黄瓜幼苗非结构性碳水化合物代谢对干旱胁迫与CO
2 倍增的响应
2
2015
... 近年来, TNC对环境因子胁迫的响应研究主要集中在高温(
李娜等, 2014 )、CO
2 浓度升高(
董彦红等, 2015 )、干旱(
杜尧等, 2014 )和氮磷添加(
王雪等, 2014 )等.大量文献报道了干旱胁迫对树木不同器官TNC含量的影响, 但结论并不一致.例如
Anacardium excelsum 、
Luehea seemannii 枝条的TNC含量在干旱季节大于湿润季节(
Newell et al ., 2002 );
Jacaratia mexicana 树干的TNC含量在湿润季节最高, 而
Spondias purpurea 树干的TNC含量在干旱与湿润季节无显著差异(
Bullock, 1992 ).另有研究表明, 高浓度O
3 减少了西黄松(
Pinus ponderosa )幼苗碳水化合物向根系的分配(
Andersen & Rygiewicz, 1995 ).但O
3 与干旱对树木TNC的复合影响却鲜有报道. ...
... 本研究结果还表明高浓度O
3 对杨树细根TNC含量的影响不显著.O
3 显著增加了楠木细根的TNC含量, 但对马尾松(
Pinus massoniana )细根的TNC含量没有影响(
Chen et al ., 2015 ), 说明不同植物的细根对O
3 胁迫的响应存在差异, 也说明不同植物应对O
3 胁迫的生存策略有所差异.在高浓度的O
3 环境下, 碳供应和碳需求之间的不协调和不平衡, 导致植物将光合产物转移用于抵御逆境的储备库而不是用于生长(
Galvez et al ., 2011 ).另一方面, O
3 和干旱胁迫对植物细根的淀粉含量无显著影响, 这与前人的研究结果有所差异.
Chen等(2015) 研究表明O
3 浓度升高显著增加了楠木细根中的淀粉含量;
Adams等(2013) 发现从干旱胁迫开始至树木死亡的过程中树木细根的淀粉含量在降低;
肖列等(2017) 发现随着干旱胁迫的加剧, 白羊草(
Bothriochloa ischaemum )根系淀粉含量显著下降.此外, 干旱胁迫显著增加了细根多糖和果糖的含量而降低蔗糖含量, 总体上增加了可溶性糖含量, 这与对黄瓜(
Cucumis sativus )、白羊草的研究结果是一致的(
董彦红等, 2015 ;
肖列等, 2017 ).通过可溶性糖的增加能增加渗透调节物质的浓度, 从而保持细胞中的水分不流失, 这对干旱胁迫下树木的生长来说是一种节省能量维持正常生理代谢的适应策略.由此可见, 不同树种细根的TNC组分对O
3 和干旱的响应不同, 可能与实验O
3 熏蒸的持续时间、累积的O
3 剂量, 干旱的胁迫程度以及树种不同有关. ...
黄瓜幼苗非结构性碳水化合物代谢对干旱胁迫与CO
2 倍增的响应
2
2015
... 近年来, TNC对环境因子胁迫的响应研究主要集中在高温(
李娜等, 2014 )、CO
2 浓度升高(
董彦红等, 2015 )、干旱(
杜尧等, 2014 )和氮磷添加(
王雪等, 2014 )等.大量文献报道了干旱胁迫对树木不同器官TNC含量的影响, 但结论并不一致.例如
Anacardium excelsum 、
Luehea seemannii 枝条的TNC含量在干旱季节大于湿润季节(
Newell et al ., 2002 );
Jacaratia mexicana 树干的TNC含量在湿润季节最高, 而
Spondias purpurea 树干的TNC含量在干旱与湿润季节无显著差异(
Bullock, 1992 ).另有研究表明, 高浓度O
3 减少了西黄松(
Pinus ponderosa )幼苗碳水化合物向根系的分配(
Andersen & Rygiewicz, 1995 ).但O
3 与干旱对树木TNC的复合影响却鲜有报道. ...
... 本研究结果还表明高浓度O
3 对杨树细根TNC含量的影响不显著.O
3 显著增加了楠木细根的TNC含量, 但对马尾松(
Pinus massoniana )细根的TNC含量没有影响(
Chen et al ., 2015 ), 说明不同植物的细根对O
3 胁迫的响应存在差异, 也说明不同植物应对O
3 胁迫的生存策略有所差异.在高浓度的O
3 环境下, 碳供应和碳需求之间的不协调和不平衡, 导致植物将光合产物转移用于抵御逆境的储备库而不是用于生长(
Galvez et al ., 2011 ).另一方面, O
3 和干旱胁迫对植物细根的淀粉含量无显著影响, 这与前人的研究结果有所差异.
Chen等(2015) 研究表明O
3 浓度升高显著增加了楠木细根中的淀粉含量;
Adams等(2013) 发现从干旱胁迫开始至树木死亡的过程中树木细根的淀粉含量在降低;
肖列等(2017) 发现随着干旱胁迫的加剧, 白羊草(
Bothriochloa ischaemum )根系淀粉含量显著下降.此外, 干旱胁迫显著增加了细根多糖和果糖的含量而降低蔗糖含量, 总体上增加了可溶性糖含量, 这与对黄瓜(
Cucumis sativus )、白羊草的研究结果是一致的(
董彦红等, 2015 ;
肖列等, 2017 ).通过可溶性糖的增加能增加渗透调节物质的浓度, 从而保持细胞中的水分不流失, 这对干旱胁迫下树木的生长来说是一种节省能量维持正常生理代谢的适应策略.由此可见, 不同树种细根的TNC组分对O
3 和干旱的响应不同, 可能与实验O
3 熏蒸的持续时间、累积的O
3 剂量, 干旱的胁迫程度以及树种不同有关. ...
干旱对兴安落叶松枝叶非结构性碳水化合物的影响
2
2014
... 近年来, TNC对环境因子胁迫的响应研究主要集中在高温(
李娜等, 2014 )、CO
2 浓度升高(
董彦红等, 2015 )、干旱(
杜尧等, 2014 )和氮磷添加(
王雪等, 2014 )等.大量文献报道了干旱胁迫对树木不同器官TNC含量的影响, 但结论并不一致.例如
Anacardium excelsum 、
Luehea seemannii 枝条的TNC含量在干旱季节大于湿润季节(
Newell et al ., 2002 );
Jacaratia mexicana 树干的TNC含量在湿润季节最高, 而
Spondias purpurea 树干的TNC含量在干旱与湿润季节无显著差异(
Bullock, 1992 ).另有研究表明, 高浓度O
3 减少了西黄松(
Pinus ponderosa )幼苗碳水化合物向根系的分配(
Andersen & Rygiewicz, 1995 ).但O
3 与干旱对树木TNC的复合影响却鲜有报道. ...
... 另一方面, 植物叶片可溶性糖和淀粉对于O
3 和干旱的响应不同.O
3 浓度升高显著增加杨树叶片中葡萄糖和果糖含量, 导致总可溶性糖含量升高, 但降低了淀粉含量, 这与对云杉(
Picea asperata )的研究结果一致(
Braun et al ., 2004 ).在高浓度的O
3 环境下, 植物会消耗体内大量的淀粉以抵御氧化胁迫, 因此导致储存的淀粉含量降低.葡萄糖是呼吸代谢的直接底物, 意味着高浓度的O
3 会促进叶片的呼吸代谢, 这与之前的实验结果一致(
Su et al ., 2017 ).干旱胁迫导致植物叶片中淀粉向可溶性糖转化, 调节植物叶片的渗透压, 增加叶片膨压(
Iannucci et al ., 2002 ).此外, 干旱胁迫下调了叶片淀粉合成能力, 导致淀粉含量降低(
Quick et al ., 1992 ).干旱胁迫对落叶松(
Larix gmelinii )影响的研究表明, 叶片内淀粉含量降低是因为淀粉水解成可溶性糖, 从而维持正常的生理活动(
杜尧等, 2014 ).但本研究结果显示, 干旱胁迫对植物叶片可溶性糖和淀粉含量均无显著影响, 可能是因为本研究采用长期轻度的干旱处理.为了避免植物产生极度干旱而导致水力结构去功能化, 因此在实验过程中给干旱植物提供节水灌溉, 从而形成了干旱复水的锻炼过程, 增加了叶片对干旱胁迫的抗性.O
3 和水分对植物叶片总可溶性糖和多糖含量存在显著的交互效应, 主要体现在干旱胁迫下O
3 对可溶性糖含量的影响不显著.干旱降低了气孔导度, 减少叶片的O
3 吸收通量, 从而减轻植物受到O
3 的伤害程度(
Gao et al ., 2017 ). ...
干旱对兴安落叶松枝叶非结构性碳水化合物的影响
2
2014
... 近年来, TNC对环境因子胁迫的响应研究主要集中在高温(
李娜等, 2014 )、CO
2 浓度升高(
董彦红等, 2015 )、干旱(
杜尧等, 2014 )和氮磷添加(
王雪等, 2014 )等.大量文献报道了干旱胁迫对树木不同器官TNC含量的影响, 但结论并不一致.例如
Anacardium excelsum 、
Luehea seemannii 枝条的TNC含量在干旱季节大于湿润季节(
Newell et al ., 2002 );
Jacaratia mexicana 树干的TNC含量在湿润季节最高, 而
Spondias purpurea 树干的TNC含量在干旱与湿润季节无显著差异(
Bullock, 1992 ).另有研究表明, 高浓度O
3 减少了西黄松(
Pinus ponderosa )幼苗碳水化合物向根系的分配(
Andersen & Rygiewicz, 1995 ).但O
3 与干旱对树木TNC的复合影响却鲜有报道. ...
... 另一方面, 植物叶片可溶性糖和淀粉对于O
3 和干旱的响应不同.O
3 浓度升高显著增加杨树叶片中葡萄糖和果糖含量, 导致总可溶性糖含量升高, 但降低了淀粉含量, 这与对云杉(
Picea asperata )的研究结果一致(
Braun et al ., 2004 ).在高浓度的O
3 环境下, 植物会消耗体内大量的淀粉以抵御氧化胁迫, 因此导致储存的淀粉含量降低.葡萄糖是呼吸代谢的直接底物, 意味着高浓度的O
3 会促进叶片的呼吸代谢, 这与之前的实验结果一致(
Su et al ., 2017 ).干旱胁迫导致植物叶片中淀粉向可溶性糖转化, 调节植物叶片的渗透压, 增加叶片膨压(
Iannucci et al ., 2002 ).此外, 干旱胁迫下调了叶片淀粉合成能力, 导致淀粉含量降低(
Quick et al ., 1992 ).干旱胁迫对落叶松(
Larix gmelinii )影响的研究表明, 叶片内淀粉含量降低是因为淀粉水解成可溶性糖, 从而维持正常的生理活动(
杜尧等, 2014 ).但本研究结果显示, 干旱胁迫对植物叶片可溶性糖和淀粉含量均无显著影响, 可能是因为本研究采用长期轻度的干旱处理.为了避免植物产生极度干旱而导致水力结构去功能化, 因此在实验过程中给干旱植物提供节水灌溉, 从而形成了干旱复水的锻炼过程, 增加了叶片对干旱胁迫的抗性.O
3 和水分对植物叶片总可溶性糖和多糖含量存在显著的交互效应, 主要体现在干旱胁迫下O
3 对可溶性糖含量的影响不显著.干旱降低了气孔导度, 减少叶片的O
3 吸收通量, 从而减轻植物受到O
3 的伤害程度(
Gao et al ., 2017 ). ...
Sucrose and the integration of metabolism in vascular plants
1
2000
... 快速城市化进程导致地表臭氧(O
3 )浓度持续升高(
Cooper et al ., 2014 ), 并且高浓度O
3 主要发生在植物生长旺盛时期(5-9月)(
冯兆忠等, 2018 ).在世界上许多地方, 植物生长期间常伴随频繁的水分亏缺, 从而使得植物同时受到高浓度O
3 和干旱的双重胁迫(
Matyssek et al ., 2010 ;
高峰等, 2017 ).O
3 污染和干旱都会降低植物光饱和点光合速率, 影响植物的碳汇能力(
Zhang et al ., 2011 ;
Ainsworth et al ., 2012 ;
吴敏等, 2014 ;
Li et al ., 2018 ), 进而影响植物的生长发育(
高峰等, 2017 ).作为光合作用的主要产物, 碳水化合物主要包括结构性碳水化合物(SC)和非结构性碳水化合物(TNC).TNC包括淀粉和总可溶性糖, 是树木生长代谢过程中重要的能量供应物质, 其浓度和含量的大小通常可以反映植物整体的碳供应状况, 体现植物阶段性碳获取和支出的平衡关系(
于丽敏等, 2011 ;
李娜妮等, 2016 ).其中, 淀粉主要用于储存物质和能量, 在植物生长、发育和繁殖等生命活动中起重要作用(
赵镭等, 2011 ); 总可溶性糖作为一种信号分子, 参与调节基因表达(
Rolland et al ., 2006 ), 并对植物体内糖代谢起到解耦联作用(
Pego et al ., 1999 ).总可溶性糖主要包括葡萄糖、果糖、蔗糖、多聚糖等.葡萄糖和果糖作为单糖直接供给植物能量, 但葡萄糖比果糖分解速度更快, 其能量释放也更快.蔗糖是植物光合作用的主要产物, 是同化产物由“源”向“库”运输的主要形式(
Farrar et al ., 2000 ), 蔗糖作为非还原性糖, 分解为果糖和葡萄糖后再向植物供应能量(
Quentin et al ., 2015 ). ...
我国地表臭氧的生态环境效应研究进展
1
2018
... 快速城市化进程导致地表臭氧(O
3 )浓度持续升高(
Cooper et al ., 2014 ), 并且高浓度O
3 主要发生在植物生长旺盛时期(5-9月)(
冯兆忠等, 2018 ).在世界上许多地方, 植物生长期间常伴随频繁的水分亏缺, 从而使得植物同时受到高浓度O
3 和干旱的双重胁迫(
Matyssek et al ., 2010 ;
高峰等, 2017 ).O
3 污染和干旱都会降低植物光饱和点光合速率, 影响植物的碳汇能力(
Zhang et al ., 2011 ;
Ainsworth et al ., 2012 ;
吴敏等, 2014 ;
Li et al ., 2018 ), 进而影响植物的生长发育(
高峰等, 2017 ).作为光合作用的主要产物, 碳水化合物主要包括结构性碳水化合物(SC)和非结构性碳水化合物(TNC).TNC包括淀粉和总可溶性糖, 是树木生长代谢过程中重要的能量供应物质, 其浓度和含量的大小通常可以反映植物整体的碳供应状况, 体现植物阶段性碳获取和支出的平衡关系(
于丽敏等, 2011 ;
李娜妮等, 2016 ).其中, 淀粉主要用于储存物质和能量, 在植物生长、发育和繁殖等生命活动中起重要作用(
赵镭等, 2011 ); 总可溶性糖作为一种信号分子, 参与调节基因表达(
Rolland et al ., 2006 ), 并对植物体内糖代谢起到解耦联作用(
Pego et al ., 1999 ).总可溶性糖主要包括葡萄糖、果糖、蔗糖、多聚糖等.葡萄糖和果糖作为单糖直接供给植物能量, 但葡萄糖比果糖分解速度更快, 其能量释放也更快.蔗糖是植物光合作用的主要产物, 是同化产物由“源”向“库”运输的主要形式(
Farrar et al ., 2000 ), 蔗糖作为非还原性糖, 分解为果糖和葡萄糖后再向植物供应能量(
Quentin et al ., 2015 ). ...
我国地表臭氧的生态环境效应研究进展
1
2018
... 快速城市化进程导致地表臭氧(O
3 )浓度持续升高(
Cooper et al ., 2014 ), 并且高浓度O
3 主要发生在植物生长旺盛时期(5-9月)(
冯兆忠等, 2018 ).在世界上许多地方, 植物生长期间常伴随频繁的水分亏缺, 从而使得植物同时受到高浓度O
3 和干旱的双重胁迫(
Matyssek et al ., 2010 ;
高峰等, 2017 ).O
3 污染和干旱都会降低植物光饱和点光合速率, 影响植物的碳汇能力(
Zhang et al ., 2011 ;
Ainsworth et al ., 2012 ;
吴敏等, 2014 ;
Li et al ., 2018 ), 进而影响植物的生长发育(
高峰等, 2017 ).作为光合作用的主要产物, 碳水化合物主要包括结构性碳水化合物(SC)和非结构性碳水化合物(TNC).TNC包括淀粉和总可溶性糖, 是树木生长代谢过程中重要的能量供应物质, 其浓度和含量的大小通常可以反映植物整体的碳供应状况, 体现植物阶段性碳获取和支出的平衡关系(
于丽敏等, 2011 ;
李娜妮等, 2016 ).其中, 淀粉主要用于储存物质和能量, 在植物生长、发育和繁殖等生命活动中起重要作用(
赵镭等, 2011 ); 总可溶性糖作为一种信号分子, 参与调节基因表达(
Rolland et al ., 2006 ), 并对植物体内糖代谢起到解耦联作用(
Pego et al ., 1999 ).总可溶性糖主要包括葡萄糖、果糖、蔗糖、多聚糖等.葡萄糖和果糖作为单糖直接供给植物能量, 但葡萄糖比果糖分解速度更快, 其能量释放也更快.蔗糖是植物光合作用的主要产物, 是同化产物由“源”向“库”运输的主要形式(
Farrar et al ., 2000 ), 蔗糖作为非还原性糖, 分解为果糖和葡萄糖后再向植物供应能量(
Quentin et al ., 2015 ). ...
Root carbon reserve dynamics in aspen seedlings: Does simulated drought induce reserve limitation?
1
2011
... 本研究结果还表明高浓度O
3 对杨树细根TNC含量的影响不显著.O
3 显著增加了楠木细根的TNC含量, 但对马尾松(
Pinus massoniana )细根的TNC含量没有影响(
Chen et al ., 2015 ), 说明不同植物的细根对O
3 胁迫的响应存在差异, 也说明不同植物应对O
3 胁迫的生存策略有所差异.在高浓度的O
3 环境下, 碳供应和碳需求之间的不协调和不平衡, 导致植物将光合产物转移用于抵御逆境的储备库而不是用于生长(
Galvez et al ., 2011 ).另一方面, O
3 和干旱胁迫对植物细根的淀粉含量无显著影响, 这与前人的研究结果有所差异.
Chen等(2015) 研究表明O
3 浓度升高显著增加了楠木细根中的淀粉含量;
Adams等(2013) 发现从干旱胁迫开始至树木死亡的过程中树木细根的淀粉含量在降低;
肖列等(2017) 发现随着干旱胁迫的加剧, 白羊草(
Bothriochloa ischaemum )根系淀粉含量显著下降.此外, 干旱胁迫显著增加了细根多糖和果糖的含量而降低蔗糖含量, 总体上增加了可溶性糖含量, 这与对黄瓜(
Cucumis sativus )、白羊草的研究结果是一致的(
董彦红等, 2015 ;
肖列等, 2017 ).通过可溶性糖的增加能增加渗透调节物质的浓度, 从而保持细胞中的水分不流失, 这对干旱胁迫下树木的生长来说是一种节省能量维持正常生理代谢的适应策略.由此可见, 不同树种细根的TNC组分对O
3 和干旱的响应不同, 可能与实验O
3 熏蒸的持续时间、累积的O
3 剂量, 干旱的胁迫程度以及树种不同有关. ...
Water stress mitigates the negative effects of ozone on photosynthesis and biomass in poplar plants
1
2017
... 另一方面, 植物叶片可溶性糖和淀粉对于O
3 和干旱的响应不同.O
3 浓度升高显著增加杨树叶片中葡萄糖和果糖含量, 导致总可溶性糖含量升高, 但降低了淀粉含量, 这与对云杉(
Picea asperata )的研究结果一致(
Braun et al ., 2004 ).在高浓度的O
3 环境下, 植物会消耗体内大量的淀粉以抵御氧化胁迫, 因此导致储存的淀粉含量降低.葡萄糖是呼吸代谢的直接底物, 意味着高浓度的O
3 会促进叶片的呼吸代谢, 这与之前的实验结果一致(
Su et al ., 2017 ).干旱胁迫导致植物叶片中淀粉向可溶性糖转化, 调节植物叶片的渗透压, 增加叶片膨压(
Iannucci et al ., 2002 ).此外, 干旱胁迫下调了叶片淀粉合成能力, 导致淀粉含量降低(
Quick et al ., 1992 ).干旱胁迫对落叶松(
Larix gmelinii )影响的研究表明, 叶片内淀粉含量降低是因为淀粉水解成可溶性糖, 从而维持正常的生理活动(
杜尧等, 2014 ).但本研究结果显示, 干旱胁迫对植物叶片可溶性糖和淀粉含量均无显著影响, 可能是因为本研究采用长期轻度的干旱处理.为了避免植物产生极度干旱而导致水力结构去功能化, 因此在实验过程中给干旱植物提供节水灌溉, 从而形成了干旱复水的锻炼过程, 增加了叶片对干旱胁迫的抗性.O
3 和水分对植物叶片总可溶性糖和多糖含量存在显著的交互效应, 主要体现在干旱胁迫下O
3 对可溶性糖含量的影响不显著.干旱降低了气孔导度, 减少叶片的O
3 吸收通量, 从而减轻植物受到O
3 的伤害程度(
Gao et al ., 2017 ). ...
臭氧与干旱对植物复合影响的研究进展
2
2017
... 快速城市化进程导致地表臭氧(O
3 )浓度持续升高(
Cooper et al ., 2014 ), 并且高浓度O
3 主要发生在植物生长旺盛时期(5-9月)(
冯兆忠等, 2018 ).在世界上许多地方, 植物生长期间常伴随频繁的水分亏缺, 从而使得植物同时受到高浓度O
3 和干旱的双重胁迫(
Matyssek et al ., 2010 ;
高峰等, 2017 ).O
3 污染和干旱都会降低植物光饱和点光合速率, 影响植物的碳汇能力(
Zhang et al ., 2011 ;
Ainsworth et al ., 2012 ;
吴敏等, 2014 ;
Li et al ., 2018 ), 进而影响植物的生长发育(
高峰等, 2017 ).作为光合作用的主要产物, 碳水化合物主要包括结构性碳水化合物(SC)和非结构性碳水化合物(TNC).TNC包括淀粉和总可溶性糖, 是树木生长代谢过程中重要的能量供应物质, 其浓度和含量的大小通常可以反映植物整体的碳供应状况, 体现植物阶段性碳获取和支出的平衡关系(
于丽敏等, 2011 ;
李娜妮等, 2016 ).其中, 淀粉主要用于储存物质和能量, 在植物生长、发育和繁殖等生命活动中起重要作用(
赵镭等, 2011 ); 总可溶性糖作为一种信号分子, 参与调节基因表达(
Rolland et al ., 2006 ), 并对植物体内糖代谢起到解耦联作用(
Pego et al ., 1999 ).总可溶性糖主要包括葡萄糖、果糖、蔗糖、多聚糖等.葡萄糖和果糖作为单糖直接供给植物能量, 但葡萄糖比果糖分解速度更快, 其能量释放也更快.蔗糖是植物光合作用的主要产物, 是同化产物由“源”向“库”运输的主要形式(
Farrar et al ., 2000 ), 蔗糖作为非还原性糖, 分解为果糖和葡萄糖后再向植物供应能量(
Quentin et al ., 2015 ). ...
... ), 进而影响植物的生长发育(
高峰等, 2017 ).作为光合作用的主要产物, 碳水化合物主要包括结构性碳水化合物(SC)和非结构性碳水化合物(TNC).TNC包括淀粉和总可溶性糖, 是树木生长代谢过程中重要的能量供应物质, 其浓度和含量的大小通常可以反映植物整体的碳供应状况, 体现植物阶段性碳获取和支出的平衡关系(
于丽敏等, 2011 ;
李娜妮等, 2016 ).其中, 淀粉主要用于储存物质和能量, 在植物生长、发育和繁殖等生命活动中起重要作用(
赵镭等, 2011 ); 总可溶性糖作为一种信号分子, 参与调节基因表达(
Rolland et al ., 2006 ), 并对植物体内糖代谢起到解耦联作用(
Pego et al ., 1999 ).总可溶性糖主要包括葡萄糖、果糖、蔗糖、多聚糖等.葡萄糖和果糖作为单糖直接供给植物能量, 但葡萄糖比果糖分解速度更快, 其能量释放也更快.蔗糖是植物光合作用的主要产物, 是同化产物由“源”向“库”运输的主要形式(
Farrar et al ., 2000 ), 蔗糖作为非还原性糖, 分解为果糖和葡萄糖后再向植物供应能量(
Quentin et al ., 2015 ). ...
臭氧与干旱对植物复合影响的研究进展
2
2017
... 快速城市化进程导致地表臭氧(O
3 )浓度持续升高(
Cooper et al ., 2014 ), 并且高浓度O
3 主要发生在植物生长旺盛时期(5-9月)(
冯兆忠等, 2018 ).在世界上许多地方, 植物生长期间常伴随频繁的水分亏缺, 从而使得植物同时受到高浓度O
3 和干旱的双重胁迫(
Matyssek et al ., 2010 ;
高峰等, 2017 ).O
3 污染和干旱都会降低植物光饱和点光合速率, 影响植物的碳汇能力(
Zhang et al ., 2011 ;
Ainsworth et al ., 2012 ;
吴敏等, 2014 ;
Li et al ., 2018 ), 进而影响植物的生长发育(
高峰等, 2017 ).作为光合作用的主要产物, 碳水化合物主要包括结构性碳水化合物(SC)和非结构性碳水化合物(TNC).TNC包括淀粉和总可溶性糖, 是树木生长代谢过程中重要的能量供应物质, 其浓度和含量的大小通常可以反映植物整体的碳供应状况, 体现植物阶段性碳获取和支出的平衡关系(
于丽敏等, 2011 ;
李娜妮等, 2016 ).其中, 淀粉主要用于储存物质和能量, 在植物生长、发育和繁殖等生命活动中起重要作用(
赵镭等, 2011 ); 总可溶性糖作为一种信号分子, 参与调节基因表达(
Rolland et al ., 2006 ), 并对植物体内糖代谢起到解耦联作用(
Pego et al ., 1999 ).总可溶性糖主要包括葡萄糖、果糖、蔗糖、多聚糖等.葡萄糖和果糖作为单糖直接供给植物能量, 但葡萄糖比果糖分解速度更快, 其能量释放也更快.蔗糖是植物光合作用的主要产物, 是同化产物由“源”向“库”运输的主要形式(
Farrar et al ., 2000 ), 蔗糖作为非还原性糖, 分解为果糖和葡萄糖后再向植物供应能量(
Quentin et al ., 2015 ). ...
... ), 进而影响植物的生长发育(
高峰等, 2017 ).作为光合作用的主要产物, 碳水化合物主要包括结构性碳水化合物(SC)和非结构性碳水化合物(TNC).TNC包括淀粉和总可溶性糖, 是树木生长代谢过程中重要的能量供应物质, 其浓度和含量的大小通常可以反映植物整体的碳供应状况, 体现植物阶段性碳获取和支出的平衡关系(
于丽敏等, 2011 ;
李娜妮等, 2016 ).其中, 淀粉主要用于储存物质和能量, 在植物生长、发育和繁殖等生命活动中起重要作用(
赵镭等, 2011 ); 总可溶性糖作为一种信号分子, 参与调节基因表达(
Rolland et al ., 2006 ), 并对植物体内糖代谢起到解耦联作用(
Pego et al ., 1999 ).总可溶性糖主要包括葡萄糖、果糖、蔗糖、多聚糖等.葡萄糖和果糖作为单糖直接供给植物能量, 但葡萄糖比果糖分解速度更快, 其能量释放也更快.蔗糖是植物光合作用的主要产物, 是同化产物由“源”向“库”运输的主要形式(
Farrar et al ., 2000 ), 蔗糖作为非还原性糖, 分解为果糖和葡萄糖后再向植物供应能量(
Quentin et al ., 2015 ). ...
Effects of ozone on biomass, non-structural carbohydrates and nitrogen in spring wheat with artificially manipulated source/ sink ratio
1
2011
... TNC是植物生长和代谢的主要能源物质, 其在植物体内含量的变化能反映植物的生理状况, 也反映植物对环境变化的动态响应.本实验结果表明, O
3 浓度升高显著降低了杨树叶片TNC含量, 这与之前O
3 对楠木(
Phoebe bournei )(
Chen et al ., 2015 )和春小麦(
Tritcium aestivum )(
Gelang et al ., 2011 )影响的结果一致, 可能是由于O
3 降低植物光合速率进而影响碳同化和TNC合成.
Topa等(2001) 研究发现, 经过3年的O
3 处理(比环境O
3 浓度高3倍), 五角枫(
Acer mono )叶片的TNC含量在春季显著降低, 但在夏季则受O
3 影响不显著, 表明在植物生长的不同阶段, O
3 对植物TNC含量的影响不同.本实验未发现干旱影响到杨树叶片的TNC含量, 这与之前的结果不一致, 即干旱有助于植物叶片内的TNC积累(
Tissue & Wright, 1995 ;
Pooter & Kitajima, 2007 ), 可能与本研究的实验设计有关.为了防止植物永久萎蔫, 实验期间采用非充分灌溉处理, 但显著降低了叶片水势(水势为-3.2 MPa), 表明本实验的植株的确受到了干旱的影响(
Shang et al ., 2019 ).同步进行的实验结果表明, 干旱胁迫降低了生物量(
Li et al ., 2019 ), 但没有降低植物的光合能力(
Shang et al ., 2019 ), 这可能是干旱胁迫未显著影响植物TNC含量的原因. ...
Concentration- and flux-based ozone dose-response relationships for five poplar clones grown in North China
1
2015
... 选择对O
3 敏感的一年生杨树幼苗‘546’ (
Populus deltoides cv. ‘55/56’
× P. deltoides cv. ‘Imperial’)(
Hu et al ., 2015 ;
Shang et al ., 2017 )为实验材料.2017年4月6日将育苗盒中长势较为一致的幼苗统一移入20 L的圆形花盆中, 用3 mm筛网过筛的当地棕色土壤与有机土等量充分混合作为栽培基质.5月7日, 将株高和基径基本一致的幼树移入OTCs中预适应10天后进行O
3 熏蒸. ...
Water deficit effects on osmotic adjustment and solute accumulation in leaves of annual clovers
1
2002
... 另一方面, 植物叶片可溶性糖和淀粉对于O
3 和干旱的响应不同.O
3 浓度升高显著增加杨树叶片中葡萄糖和果糖含量, 导致总可溶性糖含量升高, 但降低了淀粉含量, 这与对云杉(
Picea asperata )的研究结果一致(
Braun et al ., 2004 ).在高浓度的O
3 环境下, 植物会消耗体内大量的淀粉以抵御氧化胁迫, 因此导致储存的淀粉含量降低.葡萄糖是呼吸代谢的直接底物, 意味着高浓度的O
3 会促进叶片的呼吸代谢, 这与之前的实验结果一致(
Su et al ., 2017 ).干旱胁迫导致植物叶片中淀粉向可溶性糖转化, 调节植物叶片的渗透压, 增加叶片膨压(
Iannucci et al ., 2002 ).此外, 干旱胁迫下调了叶片淀粉合成能力, 导致淀粉含量降低(
Quick et al ., 1992 ).干旱胁迫对落叶松(
Larix gmelinii )影响的研究表明, 叶片内淀粉含量降低是因为淀粉水解成可溶性糖, 从而维持正常的生理活动(
杜尧等, 2014 ).但本研究结果显示, 干旱胁迫对植物叶片可溶性糖和淀粉含量均无显著影响, 可能是因为本研究采用长期轻度的干旱处理.为了避免植物产生极度干旱而导致水力结构去功能化, 因此在实验过程中给干旱植物提供节水灌溉, 从而形成了干旱复水的锻炼过程, 增加了叶片对干旱胁迫的抗性.O
3 和水分对植物叶片总可溶性糖和多糖含量存在显著的交互效应, 主要体现在干旱胁迫下O
3 对可溶性糖含量的影响不显著.干旱降低了气孔导度, 减少叶片的O
3 吸收通量, 从而减轻植物受到O
3 的伤害程度(
Gao et al ., 2017 ). ...
长期极端高温胁迫对樟子松幼苗生物量及非结构性碳水化合物的影响
1
2014
... 近年来, TNC对环境因子胁迫的响应研究主要集中在高温(
李娜等, 2014 )、CO
2 浓度升高(
董彦红等, 2015 )、干旱(
杜尧等, 2014 )和氮磷添加(
王雪等, 2014 )等.大量文献报道了干旱胁迫对树木不同器官TNC含量的影响, 但结论并不一致.例如
Anacardium excelsum 、
Luehea seemannii 枝条的TNC含量在干旱季节大于湿润季节(
Newell et al ., 2002 );
Jacaratia mexicana 树干的TNC含量在湿润季节最高, 而
Spondias purpurea 树干的TNC含量在干旱与湿润季节无显著差异(
Bullock, 1992 ).另有研究表明, 高浓度O
3 减少了西黄松(
Pinus ponderosa )幼苗碳水化合物向根系的分配(
Andersen & Rygiewicz, 1995 ).但O
3 与干旱对树木TNC的复合影响却鲜有报道. ...
长期极端高温胁迫对樟子松幼苗生物量及非结构性碳水化合物的影响
1
2014
... 近年来, TNC对环境因子胁迫的响应研究主要集中在高温(
李娜等, 2014 )、CO
2 浓度升高(
董彦红等, 2015 )、干旱(
杜尧等, 2014 )和氮磷添加(
王雪等, 2014 )等.大量文献报道了干旱胁迫对树木不同器官TNC含量的影响, 但结论并不一致.例如
Anacardium excelsum 、
Luehea seemannii 枝条的TNC含量在干旱季节大于湿润季节(
Newell et al ., 2002 );
Jacaratia mexicana 树干的TNC含量在湿润季节最高, 而
Spondias purpurea 树干的TNC含量在干旱与湿润季节无显著差异(
Bullock, 1992 ).另有研究表明, 高浓度O
3 减少了西黄松(
Pinus ponderosa )幼苗碳水化合物向根系的分配(
Andersen & Rygiewicz, 1995 ).但O
3 与干旱对树木TNC的复合影响却鲜有报道. ...
中国东北典型森林生态系统植物叶片的非结构性碳水化合物研究
1
2016
... 快速城市化进程导致地表臭氧(O
3 )浓度持续升高(
Cooper et al ., 2014 ), 并且高浓度O
3 主要发生在植物生长旺盛时期(5-9月)(
冯兆忠等, 2018 ).在世界上许多地方, 植物生长期间常伴随频繁的水分亏缺, 从而使得植物同时受到高浓度O
3 和干旱的双重胁迫(
Matyssek et al ., 2010 ;
高峰等, 2017 ).O
3 污染和干旱都会降低植物光饱和点光合速率, 影响植物的碳汇能力(
Zhang et al ., 2011 ;
Ainsworth et al ., 2012 ;
吴敏等, 2014 ;
Li et al ., 2018 ), 进而影响植物的生长发育(
高峰等, 2017 ).作为光合作用的主要产物, 碳水化合物主要包括结构性碳水化合物(SC)和非结构性碳水化合物(TNC).TNC包括淀粉和总可溶性糖, 是树木生长代谢过程中重要的能量供应物质, 其浓度和含量的大小通常可以反映植物整体的碳供应状况, 体现植物阶段性碳获取和支出的平衡关系(
于丽敏等, 2011 ;
李娜妮等, 2016 ).其中, 淀粉主要用于储存物质和能量, 在植物生长、发育和繁殖等生命活动中起重要作用(
赵镭等, 2011 ); 总可溶性糖作为一种信号分子, 参与调节基因表达(
Rolland et al ., 2006 ), 并对植物体内糖代谢起到解耦联作用(
Pego et al ., 1999 ).总可溶性糖主要包括葡萄糖、果糖、蔗糖、多聚糖等.葡萄糖和果糖作为单糖直接供给植物能量, 但葡萄糖比果糖分解速度更快, 其能量释放也更快.蔗糖是植物光合作用的主要产物, 是同化产物由“源”向“库”运输的主要形式(
Farrar et al ., 2000 ), 蔗糖作为非还原性糖, 分解为果糖和葡萄糖后再向植物供应能量(
Quentin et al ., 2015 ). ...
中国东北典型森林生态系统植物叶片的非结构性碳水化合物研究
1
2016
... 快速城市化进程导致地表臭氧(O
3 )浓度持续升高(
Cooper et al ., 2014 ), 并且高浓度O
3 主要发生在植物生长旺盛时期(5-9月)(
冯兆忠等, 2018 ).在世界上许多地方, 植物生长期间常伴随频繁的水分亏缺, 从而使得植物同时受到高浓度O
3 和干旱的双重胁迫(
Matyssek et al ., 2010 ;
高峰等, 2017 ).O
3 污染和干旱都会降低植物光饱和点光合速率, 影响植物的碳汇能力(
Zhang et al ., 2011 ;
Ainsworth et al ., 2012 ;
吴敏等, 2014 ;
Li et al ., 2018 ), 进而影响植物的生长发育(
高峰等, 2017 ).作为光合作用的主要产物, 碳水化合物主要包括结构性碳水化合物(SC)和非结构性碳水化合物(TNC).TNC包括淀粉和总可溶性糖, 是树木生长代谢过程中重要的能量供应物质, 其浓度和含量的大小通常可以反映植物整体的碳供应状况, 体现植物阶段性碳获取和支出的平衡关系(
于丽敏等, 2011 ;
李娜妮等, 2016 ).其中, 淀粉主要用于储存物质和能量, 在植物生长、发育和繁殖等生命活动中起重要作用(
赵镭等, 2011 ); 总可溶性糖作为一种信号分子, 参与调节基因表达(
Rolland et al ., 2006 ), 并对植物体内糖代谢起到解耦联作用(
Pego et al ., 1999 ).总可溶性糖主要包括葡萄糖、果糖、蔗糖、多聚糖等.葡萄糖和果糖作为单糖直接供给植物能量, 但葡萄糖比果糖分解速度更快, 其能量释放也更快.蔗糖是植物光合作用的主要产物, 是同化产物由“源”向“库”运输的主要形式(
Farrar et al ., 2000 ), 蔗糖作为非还原性糖, 分解为果糖和葡萄糖后再向植物供应能量(
Quentin et al ., 2015 ). ...
Nationwide ground-level ozone measurements in China suggest serious risks to forests
2
2018
... 快速城市化进程导致地表臭氧(O
3 )浓度持续升高(
Cooper et al ., 2014 ), 并且高浓度O
3 主要发生在植物生长旺盛时期(5-9月)(
冯兆忠等, 2018 ).在世界上许多地方, 植物生长期间常伴随频繁的水分亏缺, 从而使得植物同时受到高浓度O
3 和干旱的双重胁迫(
Matyssek et al ., 2010 ;
高峰等, 2017 ).O
3 污染和干旱都会降低植物光饱和点光合速率, 影响植物的碳汇能力(
Zhang et al ., 2011 ;
Ainsworth et al ., 2012 ;
吴敏等, 2014 ;
Li et al ., 2018 ), 进而影响植物的生长发育(
高峰等, 2017 ).作为光合作用的主要产物, 碳水化合物主要包括结构性碳水化合物(SC)和非结构性碳水化合物(TNC).TNC包括淀粉和总可溶性糖, 是树木生长代谢过程中重要的能量供应物质, 其浓度和含量的大小通常可以反映植物整体的碳供应状况, 体现植物阶段性碳获取和支出的平衡关系(
于丽敏等, 2011 ;
李娜妮等, 2016 ).其中, 淀粉主要用于储存物质和能量, 在植物生长、发育和繁殖等生命活动中起重要作用(
赵镭等, 2011 ); 总可溶性糖作为一种信号分子, 参与调节基因表达(
Rolland et al ., 2006 ), 并对植物体内糖代谢起到解耦联作用(
Pego et al ., 1999 ).总可溶性糖主要包括葡萄糖、果糖、蔗糖、多聚糖等.葡萄糖和果糖作为单糖直接供给植物能量, 但葡萄糖比果糖分解速度更快, 其能量释放也更快.蔗糖是植物光合作用的主要产物, 是同化产物由“源”向“库”运输的主要形式(
Farrar et al ., 2000 ), 蔗糖作为非还原性糖, 分解为果糖和葡萄糖后再向植物供应能量(
Quentin et al ., 2015 ). ...
... 杨树(
Populus spp.)是我国北方平原地区防护林、用材林和城乡绿化的首选树种.目前, 我国杨树人工林面积已超1亿亩, 居世界之首.同时, 我国北方地区干燥少雨, 当前环境O
3 浓度已超过树木生长的受害临界水平(
Li et al ., 2018 ).本研究利用开顶式气室(OTCs), 研究O
3 和干旱对杨树叶片和细根TNC含量的复合影响, 以期为了解大气污染和气候变化下杨树的生长响应提供科学依据. ...
Effects of elevated ozone on biomass allocation and non-structural carbohydrates in poplar are strongly related with water and nitrogen availability
2
2019
... TNC是植物生长和代谢的主要能源物质, 其在植物体内含量的变化能反映植物的生理状况, 也反映植物对环境变化的动态响应.本实验结果表明, O
3 浓度升高显著降低了杨树叶片TNC含量, 这与之前O
3 对楠木(
Phoebe bournei )(
Chen et al ., 2015 )和春小麦(
Tritcium aestivum )(
Gelang et al ., 2011 )影响的结果一致, 可能是由于O
3 降低植物光合速率进而影响碳同化和TNC合成.
Topa等(2001) 研究发现, 经过3年的O
3 处理(比环境O
3 浓度高3倍), 五角枫(
Acer mono )叶片的TNC含量在春季显著降低, 但在夏季则受O
3 影响不显著, 表明在植物生长的不同阶段, O
3 对植物TNC含量的影响不同.本实验未发现干旱影响到杨树叶片的TNC含量, 这与之前的结果不一致, 即干旱有助于植物叶片内的TNC积累(
Tissue & Wright, 1995 ;
Pooter & Kitajima, 2007 ), 可能与本研究的实验设计有关.为了防止植物永久萎蔫, 实验期间采用非充分灌溉处理, 但显著降低了叶片水势(水势为-3.2 MPa), 表明本实验的植株的确受到了干旱的影响(
Shang et al ., 2019 ).同步进行的实验结果表明, 干旱胁迫降低了生物量(
Li et al ., 2019 ), 但没有降低植物的光合能力(
Shang et al ., 2019 ), 这可能是干旱胁迫未显著影响植物TNC含量的原因. ...
... O
3 浓度升高显著增加了淀粉和TNC含量的根叶比, 原因是其显著降低了叶片中的淀粉和TNC含量而对细根没有影响.为了抵御O
3 胁迫, 叶片中的淀粉转换成可溶性糖, 增加呼吸代谢.但是植物细根的TNC主要来源于叶片光合作用形成的碳水化合物, 因此O
3 浓度升高对细根TNC含量的影响是间接的.尽管高O
3 环境下, 通过韧皮部运输到细根的TNC含量是降低的, 但用于结构生长的碳含量也显著降低(
Li et al ., 2019 ).有趣的是, 只有在水分亏缺下O
3 浓度升高对TNC含量根叶比的影响是显著的.干旱胁迫未影响到叶片和细根中的TNC含量, 但显著增加了TNC含量的根叶比.干旱胁迫导致植物的根冠比升高, 将更多的碳分配到了地下, 但TNC参与根系结构生长.因此, 干旱增加了TNC含量的根叶比, 有助于根系生长, 并增加了根系与土壤的接触面积, 进而提高了水分吸收能力. ...
Enhanced ozone strongly reduces carbon sink strength of adult beech (
Fagus sylvatica )—Resume from the free-air fumigation study at Kranzberg forest
1
2010
... 快速城市化进程导致地表臭氧(O
3 )浓度持续升高(
Cooper et al ., 2014 ), 并且高浓度O
3 主要发生在植物生长旺盛时期(5-9月)(
冯兆忠等, 2018 ).在世界上许多地方, 植物生长期间常伴随频繁的水分亏缺, 从而使得植物同时受到高浓度O
3 和干旱的双重胁迫(
Matyssek et al ., 2010 ;
高峰等, 2017 ).O
3 污染和干旱都会降低植物光饱和点光合速率, 影响植物的碳汇能力(
Zhang et al ., 2011 ;
Ainsworth et al ., 2012 ;
吴敏等, 2014 ;
Li et al ., 2018 ), 进而影响植物的生长发育(
高峰等, 2017 ).作为光合作用的主要产物, 碳水化合物主要包括结构性碳水化合物(SC)和非结构性碳水化合物(TNC).TNC包括淀粉和总可溶性糖, 是树木生长代谢过程中重要的能量供应物质, 其浓度和含量的大小通常可以反映植物整体的碳供应状况, 体现植物阶段性碳获取和支出的平衡关系(
于丽敏等, 2011 ;
李娜妮等, 2016 ).其中, 淀粉主要用于储存物质和能量, 在植物生长、发育和繁殖等生命活动中起重要作用(
赵镭等, 2011 ); 总可溶性糖作为一种信号分子, 参与调节基因表达(
Rolland et al ., 2006 ), 并对植物体内糖代谢起到解耦联作用(
Pego et al ., 1999 ).总可溶性糖主要包括葡萄糖、果糖、蔗糖、多聚糖等.葡萄糖和果糖作为单糖直接供给植物能量, 但葡萄糖比果糖分解速度更快, 其能量释放也更快.蔗糖是植物光合作用的主要产物, 是同化产物由“源”向“库”运输的主要形式(
Farrar et al ., 2000 ), 蔗糖作为非还原性糖, 分解为果糖和葡萄糖后再向植物供应能量(
Quentin et al ., 2015 ). ...
Seasonal patterns of carbohydrate storage in four tropical tree species
1
2002
... 近年来, TNC对环境因子胁迫的响应研究主要集中在高温(
李娜等, 2014 )、CO
2 浓度升高(
董彦红等, 2015 )、干旱(
杜尧等, 2014 )和氮磷添加(
王雪等, 2014 )等.大量文献报道了干旱胁迫对树木不同器官TNC含量的影响, 但结论并不一致.例如
Anacardium excelsum 、
Luehea seemannii 枝条的TNC含量在干旱季节大于湿润季节(
Newell et al ., 2002 );
Jacaratia mexicana 树干的TNC含量在湿润季节最高, 而
Spondias purpurea 树干的TNC含量在干旱与湿润季节无显著差异(
Bullock, 1992 ).另有研究表明, 高浓度O
3 减少了西黄松(
Pinus ponderosa )幼苗碳水化合物向根系的分配(
Andersen & Rygiewicz, 1995 ).但O
3 与干旱对树木TNC的复合影响却鲜有报道. ...
Mannose inhibits
Arabidopsis germination via a hexokinase-mediated step
1
1999
... 快速城市化进程导致地表臭氧(O
3 )浓度持续升高(
Cooper et al ., 2014 ), 并且高浓度O
3 主要发生在植物生长旺盛时期(5-9月)(
冯兆忠等, 2018 ).在世界上许多地方, 植物生长期间常伴随频繁的水分亏缺, 从而使得植物同时受到高浓度O
3 和干旱的双重胁迫(
Matyssek et al ., 2010 ;
高峰等, 2017 ).O
3 污染和干旱都会降低植物光饱和点光合速率, 影响植物的碳汇能力(
Zhang et al ., 2011 ;
Ainsworth et al ., 2012 ;
吴敏等, 2014 ;
Li et al ., 2018 ), 进而影响植物的生长发育(
高峰等, 2017 ).作为光合作用的主要产物, 碳水化合物主要包括结构性碳水化合物(SC)和非结构性碳水化合物(TNC).TNC包括淀粉和总可溶性糖, 是树木生长代谢过程中重要的能量供应物质, 其浓度和含量的大小通常可以反映植物整体的碳供应状况, 体现植物阶段性碳获取和支出的平衡关系(
于丽敏等, 2011 ;
李娜妮等, 2016 ).其中, 淀粉主要用于储存物质和能量, 在植物生长、发育和繁殖等生命活动中起重要作用(
赵镭等, 2011 ); 总可溶性糖作为一种信号分子, 参与调节基因表达(
Rolland et al ., 2006 ), 并对植物体内糖代谢起到解耦联作用(
Pego et al ., 1999 ).总可溶性糖主要包括葡萄糖、果糖、蔗糖、多聚糖等.葡萄糖和果糖作为单糖直接供给植物能量, 但葡萄糖比果糖分解速度更快, 其能量释放也更快.蔗糖是植物光合作用的主要产物, 是同化产物由“源”向“库”运输的主要形式(
Farrar et al ., 2000 ), 蔗糖作为非还原性糖, 分解为果糖和葡萄糖后再向植物供应能量(
Quentin et al ., 2015 ). ...
Carbohydrate storage and light requirements of tropical moist and dry forest tree species
1
2007
... TNC是植物生长和代谢的主要能源物质, 其在植物体内含量的变化能反映植物的生理状况, 也反映植物对环境变化的动态响应.本实验结果表明, O
3 浓度升高显著降低了杨树叶片TNC含量, 这与之前O
3 对楠木(
Phoebe bournei )(
Chen et al ., 2015 )和春小麦(
Tritcium aestivum )(
Gelang et al ., 2011 )影响的结果一致, 可能是由于O
3 降低植物光合速率进而影响碳同化和TNC合成.
Topa等(2001) 研究发现, 经过3年的O
3 处理(比环境O
3 浓度高3倍), 五角枫(
Acer mono )叶片的TNC含量在春季显著降低, 但在夏季则受O
3 影响不显著, 表明在植物生长的不同阶段, O
3 对植物TNC含量的影响不同.本实验未发现干旱影响到杨树叶片的TNC含量, 这与之前的结果不一致, 即干旱有助于植物叶片内的TNC积累(
Tissue & Wright, 1995 ;
Pooter & Kitajima, 2007 ), 可能与本研究的实验设计有关.为了防止植物永久萎蔫, 实验期间采用非充分灌溉处理, 但显著降低了叶片水势(水势为-3.2 MPa), 表明本实验的植株的确受到了干旱的影响(
Shang et al ., 2019 ).同步进行的实验结果表明, 干旱胁迫降低了生物量(
Li et al ., 2019 ), 但没有降低植物的光合能力(
Shang et al ., 2019 ), 这可能是干旱胁迫未显著影响植物TNC含量的原因. ...
Non-structural carbohydrates in woody plants compared among laboratories
1
2015
... 快速城市化进程导致地表臭氧(O
3 )浓度持续升高(
Cooper et al ., 2014 ), 并且高浓度O
3 主要发生在植物生长旺盛时期(5-9月)(
冯兆忠等, 2018 ).在世界上许多地方, 植物生长期间常伴随频繁的水分亏缺, 从而使得植物同时受到高浓度O
3 和干旱的双重胁迫(
Matyssek et al ., 2010 ;
高峰等, 2017 ).O
3 污染和干旱都会降低植物光饱和点光合速率, 影响植物的碳汇能力(
Zhang et al ., 2011 ;
Ainsworth et al ., 2012 ;
吴敏等, 2014 ;
Li et al ., 2018 ), 进而影响植物的生长发育(
高峰等, 2017 ).作为光合作用的主要产物, 碳水化合物主要包括结构性碳水化合物(SC)和非结构性碳水化合物(TNC).TNC包括淀粉和总可溶性糖, 是树木生长代谢过程中重要的能量供应物质, 其浓度和含量的大小通常可以反映植物整体的碳供应状况, 体现植物阶段性碳获取和支出的平衡关系(
于丽敏等, 2011 ;
李娜妮等, 2016 ).其中, 淀粉主要用于储存物质和能量, 在植物生长、发育和繁殖等生命活动中起重要作用(
赵镭等, 2011 ); 总可溶性糖作为一种信号分子, 参与调节基因表达(
Rolland et al ., 2006 ), 并对植物体内糖代谢起到解耦联作用(
Pego et al ., 1999 ).总可溶性糖主要包括葡萄糖、果糖、蔗糖、多聚糖等.葡萄糖和果糖作为单糖直接供给植物能量, 但葡萄糖比果糖分解速度更快, 其能量释放也更快.蔗糖是植物光合作用的主要产物, 是同化产物由“源”向“库”运输的主要形式(
Farrar et al ., 2000 ), 蔗糖作为非还原性糖, 分解为果糖和葡萄糖后再向植物供应能量(
Quentin et al ., 2015 ). ...
The effect of water stress on photosynthetic carbon metabolism in four species grown under field conditions
1
1992
... 另一方面, 植物叶片可溶性糖和淀粉对于O
3 和干旱的响应不同.O
3 浓度升高显著增加杨树叶片中葡萄糖和果糖含量, 导致总可溶性糖含量升高, 但降低了淀粉含量, 这与对云杉(
Picea asperata )的研究结果一致(
Braun et al ., 2004 ).在高浓度的O
3 环境下, 植物会消耗体内大量的淀粉以抵御氧化胁迫, 因此导致储存的淀粉含量降低.葡萄糖是呼吸代谢的直接底物, 意味着高浓度的O
3 会促进叶片的呼吸代谢, 这与之前的实验结果一致(
Su et al ., 2017 ).干旱胁迫导致植物叶片中淀粉向可溶性糖转化, 调节植物叶片的渗透压, 增加叶片膨压(
Iannucci et al ., 2002 ).此外, 干旱胁迫下调了叶片淀粉合成能力, 导致淀粉含量降低(
Quick et al ., 1992 ).干旱胁迫对落叶松(
Larix gmelinii )影响的研究表明, 叶片内淀粉含量降低是因为淀粉水解成可溶性糖, 从而维持正常的生理活动(
杜尧等, 2014 ).但本研究结果显示, 干旱胁迫对植物叶片可溶性糖和淀粉含量均无显著影响, 可能是因为本研究采用长期轻度的干旱处理.为了避免植物产生极度干旱而导致水力结构去功能化, 因此在实验过程中给干旱植物提供节水灌溉, 从而形成了干旱复水的锻炼过程, 增加了叶片对干旱胁迫的抗性.O
3 和水分对植物叶片总可溶性糖和多糖含量存在显著的交互效应, 主要体现在干旱胁迫下O
3 对可溶性糖含量的影响不显著.干旱降低了气孔导度, 减少叶片的O
3 吸收通量, 从而减轻植物受到O
3 的伤害程度(
Gao et al ., 2017 ). ...
Sugar sensing and signaling in plants: Conserved and novel mechanisms
1
2006
... 快速城市化进程导致地表臭氧(O
3 )浓度持续升高(
Cooper et al ., 2014 ), 并且高浓度O
3 主要发生在植物生长旺盛时期(5-9月)(
冯兆忠等, 2018 ).在世界上许多地方, 植物生长期间常伴随频繁的水分亏缺, 从而使得植物同时受到高浓度O
3 和干旱的双重胁迫(
Matyssek et al ., 2010 ;
高峰等, 2017 ).O
3 污染和干旱都会降低植物光饱和点光合速率, 影响植物的碳汇能力(
Zhang et al ., 2011 ;
Ainsworth et al ., 2012 ;
吴敏等, 2014 ;
Li et al ., 2018 ), 进而影响植物的生长发育(
高峰等, 2017 ).作为光合作用的主要产物, 碳水化合物主要包括结构性碳水化合物(SC)和非结构性碳水化合物(TNC).TNC包括淀粉和总可溶性糖, 是树木生长代谢过程中重要的能量供应物质, 其浓度和含量的大小通常可以反映植物整体的碳供应状况, 体现植物阶段性碳获取和支出的平衡关系(
于丽敏等, 2011 ;
李娜妮等, 2016 ).其中, 淀粉主要用于储存物质和能量, 在植物生长、发育和繁殖等生命活动中起重要作用(
赵镭等, 2011 ); 总可溶性糖作为一种信号分子, 参与调节基因表达(
Rolland et al ., 2006 ), 并对植物体内糖代谢起到解耦联作用(
Pego et al ., 1999 ).总可溶性糖主要包括葡萄糖、果糖、蔗糖、多聚糖等.葡萄糖和果糖作为单糖直接供给植物能量, 但葡萄糖比果糖分解速度更快, 其能量释放也更快.蔗糖是植物光合作用的主要产物, 是同化产物由“源”向“库”运输的主要形式(
Farrar et al ., 2000 ), 蔗糖作为非还原性糖, 分解为果糖和葡萄糖后再向植物供应能量(
Quentin et al ., 2015 ). ...
Ozone exposure- and flux-based response relationships with photosynthesis, leaf morphology and biomass in two poplar clones
1
2017
... 选择对O
3 敏感的一年生杨树幼苗‘546’ (
Populus deltoides cv. ‘55/56’
× P. deltoides cv. ‘Imperial’)(
Hu et al ., 2015 ;
Shang et al ., 2017 )为实验材料.2017年4月6日将育苗盒中长势较为一致的幼苗统一移入20 L的圆形花盆中, 用3 mm筛网过筛的当地棕色土壤与有机土等量充分混合作为栽培基质.5月7日, 将株高和基径基本一致的幼树移入OTCs中预适应10天后进行O
3 熏蒸. ...
Effects of elevated ozone and water deficit on poplar saplings: Changes in carbon and nitrogen stocks and their allocation to different organs
3
2019
... OTC为高3.0 m, 边长为1.0 m的正八面柱体.实验设置2个O
3 处理: CF (活性炭过滤的空气, [O
3 ] < 40 nmol·mol
-1 )、NF40 (NF (非过滤的空气) + 40 nmol·mol
-1 O
3 ).每个O
3 处理设置3个OTC重复.每个OTC设置两个水分处理: 对照, 即充分灌溉(control, 叶片水势-1.3 MPa); 干旱, 即非充分灌溉(drought, 减水40%, 叶片水势-3.2 MPa)(
张婷等, 2016 ;
Shang et al ., 2019 ), 每个亚处理设有7-8株植株重复.O
3 熏蒸自2017年6月10日开始至9月22日结束, 每天熏蒸10 h (8:00-18:00), 阴雨天停止熏蒸.实验期间各OTC的O
3 浓度由美国热电公司49i臭氧分析仪监测(Model 49i, Thermo Scientific, Franklin, USA), OTC内部和外部的温度和湿度由温湿度传感器进行实时监测并利用CR1000数据采集器(Campbell Scientific, North Logan, USA)记录, 记录频次为5 min.实验期间CF和E-O
3 处理下的平均O
3 浓度分别为24 nmol·mol
-1 和81 nmol·mol
-1 , AOT40值(小时浓度超过40 nmol·mol
-1 时的累积O
3 浓度)分别为2.4 μmol·mol
-1 ·h和41.6 μmol·mol
-1 ·h.在O
3 熏蒸时间内, OTC内外的平均气温分别为22.4 ℃和20.5 ℃. ...
... TNC是植物生长和代谢的主要能源物质, 其在植物体内含量的变化能反映植物的生理状况, 也反映植物对环境变化的动态响应.本实验结果表明, O
3 浓度升高显著降低了杨树叶片TNC含量, 这与之前O
3 对楠木(
Phoebe bournei )(
Chen et al ., 2015 )和春小麦(
Tritcium aestivum )(
Gelang et al ., 2011 )影响的结果一致, 可能是由于O
3 降低植物光合速率进而影响碳同化和TNC合成.
Topa等(2001) 研究发现, 经过3年的O
3 处理(比环境O
3 浓度高3倍), 五角枫(
Acer mono )叶片的TNC含量在春季显著降低, 但在夏季则受O
3 影响不显著, 表明在植物生长的不同阶段, O
3 对植物TNC含量的影响不同.本实验未发现干旱影响到杨树叶片的TNC含量, 这与之前的结果不一致, 即干旱有助于植物叶片内的TNC积累(
Tissue & Wright, 1995 ;
Pooter & Kitajima, 2007 ), 可能与本研究的实验设计有关.为了防止植物永久萎蔫, 实验期间采用非充分灌溉处理, 但显著降低了叶片水势(水势为-3.2 MPa), 表明本实验的植株的确受到了干旱的影响(
Shang et al ., 2019 ).同步进行的实验结果表明, 干旱胁迫降低了生物量(
Li et al ., 2019 ), 但没有降低植物的光合能力(
Shang et al ., 2019 ), 这可能是干旱胁迫未显著影响植物TNC含量的原因. ...
... ), 但没有降低植物的光合能力(
Shang et al ., 2019 ), 这可能是干旱胁迫未显著影响植物TNC含量的原因. ...
Photosynthesis and biochemical responses to elevated O
3 in
Plantago major and S
onchus oleraceus growing in a lowland habitat of northern China
1
2017
... 另一方面, 植物叶片可溶性糖和淀粉对于O
3 和干旱的响应不同.O
3 浓度升高显著增加杨树叶片中葡萄糖和果糖含量, 导致总可溶性糖含量升高, 但降低了淀粉含量, 这与对云杉(
Picea asperata )的研究结果一致(
Braun et al ., 2004 ).在高浓度的O
3 环境下, 植物会消耗体内大量的淀粉以抵御氧化胁迫, 因此导致储存的淀粉含量降低.葡萄糖是呼吸代谢的直接底物, 意味着高浓度的O
3 会促进叶片的呼吸代谢, 这与之前的实验结果一致(
Su et al ., 2017 ).干旱胁迫导致植物叶片中淀粉向可溶性糖转化, 调节植物叶片的渗透压, 增加叶片膨压(
Iannucci et al ., 2002 ).此外, 干旱胁迫下调了叶片淀粉合成能力, 导致淀粉含量降低(
Quick et al ., 1992 ).干旱胁迫对落叶松(
Larix gmelinii )影响的研究表明, 叶片内淀粉含量降低是因为淀粉水解成可溶性糖, 从而维持正常的生理活动(
杜尧等, 2014 ).但本研究结果显示, 干旱胁迫对植物叶片可溶性糖和淀粉含量均无显著影响, 可能是因为本研究采用长期轻度的干旱处理.为了避免植物产生极度干旱而导致水力结构去功能化, 因此在实验过程中给干旱植物提供节水灌溉, 从而形成了干旱复水的锻炼过程, 增加了叶片对干旱胁迫的抗性.O
3 和水分对植物叶片总可溶性糖和多糖含量存在显著的交互效应, 主要体现在干旱胁迫下O
3 对可溶性糖含量的影响不显著.干旱降低了气孔导度, 减少叶片的O
3 吸收通量, 从而减轻植物受到O
3 的伤害程度(
Gao et al ., 2017 ). ...
1
2006
... 淀粉提取及测定: 分别称取0.05 g和0.5 g叶片和细根样品置于10 mL离心管中, 加入1 mL蒸馏水和5 mL 80%乙醇溶液置于涡旋振荡器中混匀静置, 15 000 r·min
-1 下离心10 min, 弃上清液, 重复2次.于残留物中加入1 mL蒸馏水和6 mL 52%高氯酸溶液, 置于涡旋振荡器涡旋10 min, 15 000 r·min
-1 下离心10 min, 将上清液转入25 mL容量瓶, 重复2次, 用蒸馏水定容至25 mL.将该溶液进行过滤, 滤液转入50 mL容量瓶, 并用蒸馏水定容, 得到淀粉提取液, 采用高氯酸水解-蒽酮比色法测定淀粉含量(
田纪春, 2006 ). ...
1
2006
... 淀粉提取及测定: 分别称取0.05 g和0.5 g叶片和细根样品置于10 mL离心管中, 加入1 mL蒸馏水和5 mL 80%乙醇溶液置于涡旋振荡器中混匀静置, 15 000 r·min
-1 下离心10 min, 弃上清液, 重复2次.于残留物中加入1 mL蒸馏水和6 mL 52%高氯酸溶液, 置于涡旋振荡器涡旋10 min, 15 000 r·min
-1 下离心10 min, 将上清液转入25 mL容量瓶, 重复2次, 用蒸馏水定容至25 mL.将该溶液进行过滤, 滤液转入50 mL容量瓶, 并用蒸馏水定容, 得到淀粉提取液, 采用高氯酸水解-蒽酮比色法测定淀粉含量(
田纪春, 2006 ). ...
Effect of seasonal water availability on phenology and the annual shoot carbohydrate cycle of tropical forest shrubs
1
1995
... TNC是植物生长和代谢的主要能源物质, 其在植物体内含量的变化能反映植物的生理状况, 也反映植物对环境变化的动态响应.本实验结果表明, O
3 浓度升高显著降低了杨树叶片TNC含量, 这与之前O
3 对楠木(
Phoebe bournei )(
Chen et al ., 2015 )和春小麦(
Tritcium aestivum )(
Gelang et al ., 2011 )影响的结果一致, 可能是由于O
3 降低植物光合速率进而影响碳同化和TNC合成.
Topa等(2001) 研究发现, 经过3年的O
3 处理(比环境O
3 浓度高3倍), 五角枫(
Acer mono )叶片的TNC含量在春季显著降低, 但在夏季则受O
3 影响不显著, 表明在植物生长的不同阶段, O
3 对植物TNC含量的影响不同.本实验未发现干旱影响到杨树叶片的TNC含量, 这与之前的结果不一致, 即干旱有助于植物叶片内的TNC积累(
Tissue & Wright, 1995 ;
Pooter & Kitajima, 2007 ), 可能与本研究的实验设计有关.为了防止植物永久萎蔫, 实验期间采用非充分灌溉处理, 但显著降低了叶片水势(水势为-3.2 MPa), 表明本实验的植株的确受到了干旱的影响(
Shang et al ., 2019 ).同步进行的实验结果表明, 干旱胁迫降低了生物量(
Li et al ., 2019 ), 但没有降低植物的光合能力(
Shang et al ., 2019 ), 这可能是干旱胁迫未显著影响植物TNC含量的原因. ...
Effects of elevated ozone and low light on diurnal and seasonal carbon gain in sugar maple
1
2001
... TNC是植物生长和代谢的主要能源物质, 其在植物体内含量的变化能反映植物的生理状况, 也反映植物对环境变化的动态响应.本实验结果表明, O
3 浓度升高显著降低了杨树叶片TNC含量, 这与之前O
3 对楠木(
Phoebe bournei )(
Chen et al ., 2015 )和春小麦(
Tritcium aestivum )(
Gelang et al ., 2011 )影响的结果一致, 可能是由于O
3 降低植物光合速率进而影响碳同化和TNC合成.
Topa等(2001) 研究发现, 经过3年的O
3 处理(比环境O
3 浓度高3倍), 五角枫(
Acer mono )叶片的TNC含量在春季显著降低, 但在夏季则受O
3 影响不显著, 表明在植物生长的不同阶段, O
3 对植物TNC含量的影响不同.本实验未发现干旱影响到杨树叶片的TNC含量, 这与之前的结果不一致, 即干旱有助于植物叶片内的TNC积累(
Tissue & Wright, 1995 ;
Pooter & Kitajima, 2007 ), 可能与本研究的实验设计有关.为了防止植物永久萎蔫, 实验期间采用非充分灌溉处理, 但显著降低了叶片水势(水势为-3.2 MPa), 表明本实验的植株的确受到了干旱的影响(
Shang et al ., 2019 ).同步进行的实验结果表明, 干旱胁迫降低了生物量(
Li et al ., 2019 ), 但没有降低植物的光合能力(
Shang et al ., 2019 ), 这可能是干旱胁迫未显著影响植物TNC含量的原因. ...
半干旱典型草原养分添加对优势物种叶片氮磷及非结构性碳水化合物含量的影响
1
2014
... 近年来, TNC对环境因子胁迫的响应研究主要集中在高温(
李娜等, 2014 )、CO
2 浓度升高(
董彦红等, 2015 )、干旱(
杜尧等, 2014 )和氮磷添加(
王雪等, 2014 )等.大量文献报道了干旱胁迫对树木不同器官TNC含量的影响, 但结论并不一致.例如
Anacardium excelsum 、
Luehea seemannii 枝条的TNC含量在干旱季节大于湿润季节(
Newell et al ., 2002 );
Jacaratia mexicana 树干的TNC含量在湿润季节最高, 而
Spondias purpurea 树干的TNC含量在干旱与湿润季节无显著差异(
Bullock, 1992 ).另有研究表明, 高浓度O
3 减少了西黄松(
Pinus ponderosa )幼苗碳水化合物向根系的分配(
Andersen & Rygiewicz, 1995 ).但O
3 与干旱对树木TNC的复合影响却鲜有报道. ...
半干旱典型草原养分添加对优势物种叶片氮磷及非结构性碳水化合物含量的影响
1
2014
... 近年来, TNC对环境因子胁迫的响应研究主要集中在高温(
李娜等, 2014 )、CO
2 浓度升高(
董彦红等, 2015 )、干旱(
杜尧等, 2014 )和氮磷添加(
王雪等, 2014 )等.大量文献报道了干旱胁迫对树木不同器官TNC含量的影响, 但结论并不一致.例如
Anacardium excelsum 、
Luehea seemannii 枝条的TNC含量在干旱季节大于湿润季节(
Newell et al ., 2002 );
Jacaratia mexicana 树干的TNC含量在湿润季节最高, 而
Spondias purpurea 树干的TNC含量在干旱与湿润季节无显著差异(
Bullock, 1992 ).另有研究表明, 高浓度O
3 减少了西黄松(
Pinus ponderosa )幼苗碳水化合物向根系的分配(
Andersen & Rygiewicz, 1995 ).但O
3 与干旱对树木TNC的复合影响却鲜有报道. ...
干旱胁迫对栓皮栎幼苗细根的生长与生理生化指标的影响
1
2014
... 快速城市化进程导致地表臭氧(O
3 )浓度持续升高(
Cooper et al ., 2014 ), 并且高浓度O
3 主要发生在植物生长旺盛时期(5-9月)(
冯兆忠等, 2018 ).在世界上许多地方, 植物生长期间常伴随频繁的水分亏缺, 从而使得植物同时受到高浓度O
3 和干旱的双重胁迫(
Matyssek et al ., 2010 ;
高峰等, 2017 ).O
3 污染和干旱都会降低植物光饱和点光合速率, 影响植物的碳汇能力(
Zhang et al ., 2011 ;
Ainsworth et al ., 2012 ;
吴敏等, 2014 ;
Li et al ., 2018 ), 进而影响植物的生长发育(
高峰等, 2017 ).作为光合作用的主要产物, 碳水化合物主要包括结构性碳水化合物(SC)和非结构性碳水化合物(TNC).TNC包括淀粉和总可溶性糖, 是树木生长代谢过程中重要的能量供应物质, 其浓度和含量的大小通常可以反映植物整体的碳供应状况, 体现植物阶段性碳获取和支出的平衡关系(
于丽敏等, 2011 ;
李娜妮等, 2016 ).其中, 淀粉主要用于储存物质和能量, 在植物生长、发育和繁殖等生命活动中起重要作用(
赵镭等, 2011 ); 总可溶性糖作为一种信号分子, 参与调节基因表达(
Rolland et al ., 2006 ), 并对植物体内糖代谢起到解耦联作用(
Pego et al ., 1999 ).总可溶性糖主要包括葡萄糖、果糖、蔗糖、多聚糖等.葡萄糖和果糖作为单糖直接供给植物能量, 但葡萄糖比果糖分解速度更快, 其能量释放也更快.蔗糖是植物光合作用的主要产物, 是同化产物由“源”向“库”运输的主要形式(
Farrar et al ., 2000 ), 蔗糖作为非还原性糖, 分解为果糖和葡萄糖后再向植物供应能量(
Quentin et al ., 2015 ). ...
干旱胁迫对栓皮栎幼苗细根的生长与生理生化指标的影响
1
2014
... 快速城市化进程导致地表臭氧(O
3 )浓度持续升高(
Cooper et al ., 2014 ), 并且高浓度O
3 主要发生在植物生长旺盛时期(5-9月)(
冯兆忠等, 2018 ).在世界上许多地方, 植物生长期间常伴随频繁的水分亏缺, 从而使得植物同时受到高浓度O
3 和干旱的双重胁迫(
Matyssek et al ., 2010 ;
高峰等, 2017 ).O
3 污染和干旱都会降低植物光饱和点光合速率, 影响植物的碳汇能力(
Zhang et al ., 2011 ;
Ainsworth et al ., 2012 ;
吴敏等, 2014 ;
Li et al ., 2018 ), 进而影响植物的生长发育(
高峰等, 2017 ).作为光合作用的主要产物, 碳水化合物主要包括结构性碳水化合物(SC)和非结构性碳水化合物(TNC).TNC包括淀粉和总可溶性糖, 是树木生长代谢过程中重要的能量供应物质, 其浓度和含量的大小通常可以反映植物整体的碳供应状况, 体现植物阶段性碳获取和支出的平衡关系(
于丽敏等, 2011 ;
李娜妮等, 2016 ).其中, 淀粉主要用于储存物质和能量, 在植物生长、发育和繁殖等生命活动中起重要作用(
赵镭等, 2011 ); 总可溶性糖作为一种信号分子, 参与调节基因表达(
Rolland et al ., 2006 ), 并对植物体内糖代谢起到解耦联作用(
Pego et al ., 1999 ).总可溶性糖主要包括葡萄糖、果糖、蔗糖、多聚糖等.葡萄糖和果糖作为单糖直接供给植物能量, 但葡萄糖比果糖分解速度更快, 其能量释放也更快.蔗糖是植物光合作用的主要产物, 是同化产物由“源”向“库”运输的主要形式(
Farrar et al ., 2000 ), 蔗糖作为非还原性糖, 分解为果糖和葡萄糖后再向植物供应能量(
Quentin et al ., 2015 ). ...
白羊草光合特性及非结构性碳水化合物含量对CO
2 浓度倍增和干旱胁迫的响应
2
2017
... 本研究结果还表明高浓度O
3 对杨树细根TNC含量的影响不显著.O
3 显著增加了楠木细根的TNC含量, 但对马尾松(
Pinus massoniana )细根的TNC含量没有影响(
Chen et al ., 2015 ), 说明不同植物的细根对O
3 胁迫的响应存在差异, 也说明不同植物应对O
3 胁迫的生存策略有所差异.在高浓度的O
3 环境下, 碳供应和碳需求之间的不协调和不平衡, 导致植物将光合产物转移用于抵御逆境的储备库而不是用于生长(
Galvez et al ., 2011 ).另一方面, O
3 和干旱胁迫对植物细根的淀粉含量无显著影响, 这与前人的研究结果有所差异.
Chen等(2015) 研究表明O
3 浓度升高显著增加了楠木细根中的淀粉含量;
Adams等(2013) 发现从干旱胁迫开始至树木死亡的过程中树木细根的淀粉含量在降低;
肖列等(2017) 发现随着干旱胁迫的加剧, 白羊草(
Bothriochloa ischaemum )根系淀粉含量显著下降.此外, 干旱胁迫显著增加了细根多糖和果糖的含量而降低蔗糖含量, 总体上增加了可溶性糖含量, 这与对黄瓜(
Cucumis sativus )、白羊草的研究结果是一致的(
董彦红等, 2015 ;
肖列等, 2017 ).通过可溶性糖的增加能增加渗透调节物质的浓度, 从而保持细胞中的水分不流失, 这对干旱胁迫下树木的生长来说是一种节省能量维持正常生理代谢的适应策略.由此可见, 不同树种细根的TNC组分对O
3 和干旱的响应不同, 可能与实验O
3 熏蒸的持续时间、累积的O
3 剂量, 干旱的胁迫程度以及树种不同有关. ...
... ;
肖列等, 2017 ).通过可溶性糖的增加能增加渗透调节物质的浓度, 从而保持细胞中的水分不流失, 这对干旱胁迫下树木的生长来说是一种节省能量维持正常生理代谢的适应策略.由此可见, 不同树种细根的TNC组分对O
3 和干旱的响应不同, 可能与实验O
3 熏蒸的持续时间、累积的O
3 剂量, 干旱的胁迫程度以及树种不同有关. ...
白羊草光合特性及非结构性碳水化合物含量对CO
2 浓度倍增和干旱胁迫的响应
2
2017
... 本研究结果还表明高浓度O
3 对杨树细根TNC含量的影响不显著.O
3 显著增加了楠木细根的TNC含量, 但对马尾松(
Pinus massoniana )细根的TNC含量没有影响(
Chen et al ., 2015 ), 说明不同植物的细根对O
3 胁迫的响应存在差异, 也说明不同植物应对O
3 胁迫的生存策略有所差异.在高浓度的O
3 环境下, 碳供应和碳需求之间的不协调和不平衡, 导致植物将光合产物转移用于抵御逆境的储备库而不是用于生长(
Galvez et al ., 2011 ).另一方面, O
3 和干旱胁迫对植物细根的淀粉含量无显著影响, 这与前人的研究结果有所差异.
Chen等(2015) 研究表明O
3 浓度升高显著增加了楠木细根中的淀粉含量;
Adams等(2013) 发现从干旱胁迫开始至树木死亡的过程中树木细根的淀粉含量在降低;
肖列等(2017) 发现随着干旱胁迫的加剧, 白羊草(
Bothriochloa ischaemum )根系淀粉含量显著下降.此外, 干旱胁迫显著增加了细根多糖和果糖的含量而降低蔗糖含量, 总体上增加了可溶性糖含量, 这与对黄瓜(
Cucumis sativus )、白羊草的研究结果是一致的(
董彦红等, 2015 ;
肖列等, 2017 ).通过可溶性糖的增加能增加渗透调节物质的浓度, 从而保持细胞中的水分不流失, 这对干旱胁迫下树木的生长来说是一种节省能量维持正常生理代谢的适应策略.由此可见, 不同树种细根的TNC组分对O
3 和干旱的响应不同, 可能与实验O
3 熏蒸的持续时间、累积的O
3 剂量, 干旱的胁迫程度以及树种不同有关. ...
... ;
肖列等, 2017 ).通过可溶性糖的增加能增加渗透调节物质的浓度, 从而保持细胞中的水分不流失, 这对干旱胁迫下树木的生长来说是一种节省能量维持正常生理代谢的适应策略.由此可见, 不同树种细根的TNC组分对O
3 和干旱的响应不同, 可能与实验O
3 熏蒸的持续时间、累积的O
3 剂量, 干旱的胁迫程度以及树种不同有关. ...
三种温带树种非结构性碳水化合物的分配
1
2011
... 快速城市化进程导致地表臭氧(O
3 )浓度持续升高(
Cooper et al ., 2014 ), 并且高浓度O
3 主要发生在植物生长旺盛时期(5-9月)(
冯兆忠等, 2018 ).在世界上许多地方, 植物生长期间常伴随频繁的水分亏缺, 从而使得植物同时受到高浓度O
3 和干旱的双重胁迫(
Matyssek et al ., 2010 ;
高峰等, 2017 ).O
3 污染和干旱都会降低植物光饱和点光合速率, 影响植物的碳汇能力(
Zhang et al ., 2011 ;
Ainsworth et al ., 2012 ;
吴敏等, 2014 ;
Li et al ., 2018 ), 进而影响植物的生长发育(
高峰等, 2017 ).作为光合作用的主要产物, 碳水化合物主要包括结构性碳水化合物(SC)和非结构性碳水化合物(TNC).TNC包括淀粉和总可溶性糖, 是树木生长代谢过程中重要的能量供应物质, 其浓度和含量的大小通常可以反映植物整体的碳供应状况, 体现植物阶段性碳获取和支出的平衡关系(
于丽敏等, 2011 ;
李娜妮等, 2016 ).其中, 淀粉主要用于储存物质和能量, 在植物生长、发育和繁殖等生命活动中起重要作用(
赵镭等, 2011 ); 总可溶性糖作为一种信号分子, 参与调节基因表达(
Rolland et al ., 2006 ), 并对植物体内糖代谢起到解耦联作用(
Pego et al ., 1999 ).总可溶性糖主要包括葡萄糖、果糖、蔗糖、多聚糖等.葡萄糖和果糖作为单糖直接供给植物能量, 但葡萄糖比果糖分解速度更快, 其能量释放也更快.蔗糖是植物光合作用的主要产物, 是同化产物由“源”向“库”运输的主要形式(
Farrar et al ., 2000 ), 蔗糖作为非还原性糖, 分解为果糖和葡萄糖后再向植物供应能量(
Quentin et al ., 2015 ). ...
三种温带树种非结构性碳水化合物的分配
1
2011
... 快速城市化进程导致地表臭氧(O
3 )浓度持续升高(
Cooper et al ., 2014 ), 并且高浓度O
3 主要发生在植物生长旺盛时期(5-9月)(
冯兆忠等, 2018 ).在世界上许多地方, 植物生长期间常伴随频繁的水分亏缺, 从而使得植物同时受到高浓度O
3 和干旱的双重胁迫(
Matyssek et al ., 2010 ;
高峰等, 2017 ).O
3 污染和干旱都会降低植物光饱和点光合速率, 影响植物的碳汇能力(
Zhang et al ., 2011 ;
Ainsworth et al ., 2012 ;
吴敏等, 2014 ;
Li et al ., 2018 ), 进而影响植物的生长发育(
高峰等, 2017 ).作为光合作用的主要产物, 碳水化合物主要包括结构性碳水化合物(SC)和非结构性碳水化合物(TNC).TNC包括淀粉和总可溶性糖, 是树木生长代谢过程中重要的能量供应物质, 其浓度和含量的大小通常可以反映植物整体的碳供应状况, 体现植物阶段性碳获取和支出的平衡关系(
于丽敏等, 2011 ;
李娜妮等, 2016 ).其中, 淀粉主要用于储存物质和能量, 在植物生长、发育和繁殖等生命活动中起重要作用(
赵镭等, 2011 ); 总可溶性糖作为一种信号分子, 参与调节基因表达(
Rolland et al ., 2006 ), 并对植物体内糖代谢起到解耦联作用(
Pego et al ., 1999 ).总可溶性糖主要包括葡萄糖、果糖、蔗糖、多聚糖等.葡萄糖和果糖作为单糖直接供给植物能量, 但葡萄糖比果糖分解速度更快, 其能量释放也更快.蔗糖是植物光合作用的主要产物, 是同化产物由“源”向“库”运输的主要形式(
Farrar et al ., 2000 ), 蔗糖作为非还原性糖, 分解为果糖和葡萄糖后再向植物供应能量(
Quentin et al ., 2015 ). ...
浙江天童常见种幼苗的光合特性及非结构性碳水化合物储存
1
2011
... 快速城市化进程导致地表臭氧(O
3 )浓度持续升高(
Cooper et al ., 2014 ), 并且高浓度O
3 主要发生在植物生长旺盛时期(5-9月)(
冯兆忠等, 2018 ).在世界上许多地方, 植物生长期间常伴随频繁的水分亏缺, 从而使得植物同时受到高浓度O
3 和干旱的双重胁迫(
Matyssek et al ., 2010 ;
高峰等, 2017 ).O
3 污染和干旱都会降低植物光饱和点光合速率, 影响植物的碳汇能力(
Zhang et al ., 2011 ;
Ainsworth et al ., 2012 ;
吴敏等, 2014 ;
Li et al ., 2018 ), 进而影响植物的生长发育(
高峰等, 2017 ).作为光合作用的主要产物, 碳水化合物主要包括结构性碳水化合物(SC)和非结构性碳水化合物(TNC).TNC包括淀粉和总可溶性糖, 是树木生长代谢过程中重要的能量供应物质, 其浓度和含量的大小通常可以反映植物整体的碳供应状况, 体现植物阶段性碳获取和支出的平衡关系(
于丽敏等, 2011 ;
李娜妮等, 2016 ).其中, 淀粉主要用于储存物质和能量, 在植物生长、发育和繁殖等生命活动中起重要作用(
赵镭等, 2011 ); 总可溶性糖作为一种信号分子, 参与调节基因表达(
Rolland et al ., 2006 ), 并对植物体内糖代谢起到解耦联作用(
Pego et al ., 1999 ).总可溶性糖主要包括葡萄糖、果糖、蔗糖、多聚糖等.葡萄糖和果糖作为单糖直接供给植物能量, 但葡萄糖比果糖分解速度更快, 其能量释放也更快.蔗糖是植物光合作用的主要产物, 是同化产物由“源”向“库”运输的主要形式(
Farrar et al ., 2000 ), 蔗糖作为非还原性糖, 分解为果糖和葡萄糖后再向植物供应能量(
Quentin et al ., 2015 ). ...
浙江天童常见种幼苗的光合特性及非结构性碳水化合物储存
1
2011
... 快速城市化进程导致地表臭氧(O
3 )浓度持续升高(
Cooper et al ., 2014 ), 并且高浓度O
3 主要发生在植物生长旺盛时期(5-9月)(
冯兆忠等, 2018 ).在世界上许多地方, 植物生长期间常伴随频繁的水分亏缺, 从而使得植物同时受到高浓度O
3 和干旱的双重胁迫(
Matyssek et al ., 2010 ;
高峰等, 2017 ).O
3 污染和干旱都会降低植物光饱和点光合速率, 影响植物的碳汇能力(
Zhang et al ., 2011 ;
Ainsworth et al ., 2012 ;
吴敏等, 2014 ;
Li et al ., 2018 ), 进而影响植物的生长发育(
高峰等, 2017 ).作为光合作用的主要产物, 碳水化合物主要包括结构性碳水化合物(SC)和非结构性碳水化合物(TNC).TNC包括淀粉和总可溶性糖, 是树木生长代谢过程中重要的能量供应物质, 其浓度和含量的大小通常可以反映植物整体的碳供应状况, 体现植物阶段性碳获取和支出的平衡关系(
于丽敏等, 2011 ;
李娜妮等, 2016 ).其中, 淀粉主要用于储存物质和能量, 在植物生长、发育和繁殖等生命活动中起重要作用(
赵镭等, 2011 ); 总可溶性糖作为一种信号分子, 参与调节基因表达(
Rolland et al ., 2006 ), 并对植物体内糖代谢起到解耦联作用(
Pego et al ., 1999 ).总可溶性糖主要包括葡萄糖、果糖、蔗糖、多聚糖等.葡萄糖和果糖作为单糖直接供给植物能量, 但葡萄糖比果糖分解速度更快, 其能量释放也更快.蔗糖是植物光合作用的主要产物, 是同化产物由“源”向“库”运输的主要形式(
Farrar et al ., 2000 ), 蔗糖作为非还原性糖, 分解为果糖和葡萄糖后再向植物供应能量(
Quentin et al ., 2015 ). ...
生长季末期干旱胁迫对刺槐幼苗非结构性碳水化合物的影响
1
2016
... OTC为高3.0 m, 边长为1.0 m的正八面柱体.实验设置2个O
3 处理: CF (活性炭过滤的空气, [O
3 ] < 40 nmol·mol
-1 )、NF40 (NF (非过滤的空气) + 40 nmol·mol
-1 O
3 ).每个O
3 处理设置3个OTC重复.每个OTC设置两个水分处理: 对照, 即充分灌溉(control, 叶片水势-1.3 MPa); 干旱, 即非充分灌溉(drought, 减水40%, 叶片水势-3.2 MPa)(
张婷等, 2016 ;
Shang et al ., 2019 ), 每个亚处理设有7-8株植株重复.O
3 熏蒸自2017年6月10日开始至9月22日结束, 每天熏蒸10 h (8:00-18:00), 阴雨天停止熏蒸.实验期间各OTC的O
3 浓度由美国热电公司49i臭氧分析仪监测(Model 49i, Thermo Scientific, Franklin, USA), OTC内部和外部的温度和湿度由温湿度传感器进行实时监测并利用CR1000数据采集器(Campbell Scientific, North Logan, USA)记录, 记录频次为5 min.实验期间CF和E-O
3 处理下的平均O
3 浓度分别为24 nmol·mol
-1 和81 nmol·mol
-1 , AOT40值(小时浓度超过40 nmol·mol
-1 时的累积O
3 浓度)分别为2.4 μmol·mol
-1 ·h和41.6 μmol·mol
-1 ·h.在O
3 熏蒸时间内, OTC内外的平均气温分别为22.4 ℃和20.5 ℃. ...
生长季末期干旱胁迫对刺槐幼苗非结构性碳水化合物的影响
1
2016
... OTC为高3.0 m, 边长为1.0 m的正八面柱体.实验设置2个O
3 处理: CF (活性炭过滤的空气, [O
3 ] < 40 nmol·mol
-1 )、NF40 (NF (非过滤的空气) + 40 nmol·mol
-1 O
3 ).每个O
3 处理设置3个OTC重复.每个OTC设置两个水分处理: 对照, 即充分灌溉(control, 叶片水势-1.3 MPa); 干旱, 即非充分灌溉(drought, 减水40%, 叶片水势-3.2 MPa)(
张婷等, 2016 ;
Shang et al ., 2019 ), 每个亚处理设有7-8株植株重复.O
3 熏蒸自2017年6月10日开始至9月22日结束, 每天熏蒸10 h (8:00-18:00), 阴雨天停止熏蒸.实验期间各OTC的O
3 浓度由美国热电公司49i臭氧分析仪监测(Model 49i, Thermo Scientific, Franklin, USA), OTC内部和外部的温度和湿度由温湿度传感器进行实时监测并利用CR1000数据采集器(Campbell Scientific, North Logan, USA)记录, 记录频次为5 min.实验期间CF和E-O
3 处理下的平均O
3 浓度分别为24 nmol·mol
-1 和81 nmol·mol
-1 , AOT40值(小时浓度超过40 nmol·mol
-1 时的累积O
3 浓度)分别为2.4 μmol·mol
-1 ·h和41.6 μmol·mol
-1 ·h.在O
3 熏蒸时间内, OTC内外的平均气温分别为22.4 ℃和20.5 ℃. ...
Effect of water stress on leaf photosynthesis, chlorophyll content, and growth of oriental lily
1
2011
... 快速城市化进程导致地表臭氧(O
3 )浓度持续升高(
Cooper et al ., 2014 ), 并且高浓度O
3 主要发生在植物生长旺盛时期(5-9月)(
冯兆忠等, 2018 ).在世界上许多地方, 植物生长期间常伴随频繁的水分亏缺, 从而使得植物同时受到高浓度O
3 和干旱的双重胁迫(
Matyssek et al ., 2010 ;
高峰等, 2017 ).O
3 污染和干旱都会降低植物光饱和点光合速率, 影响植物的碳汇能力(
Zhang et al ., 2011 ;
Ainsworth et al ., 2012 ;
吴敏等, 2014 ;
Li et al ., 2018 ), 进而影响植物的生长发育(
高峰等, 2017 ).作为光合作用的主要产物, 碳水化合物主要包括结构性碳水化合物(SC)和非结构性碳水化合物(TNC).TNC包括淀粉和总可溶性糖, 是树木生长代谢过程中重要的能量供应物质, 其浓度和含量的大小通常可以反映植物整体的碳供应状况, 体现植物阶段性碳获取和支出的平衡关系(
于丽敏等, 2011 ;
李娜妮等, 2016 ).其中, 淀粉主要用于储存物质和能量, 在植物生长、发育和繁殖等生命活动中起重要作用(
赵镭等, 2011 ); 总可溶性糖作为一种信号分子, 参与调节基因表达(
Rolland et al ., 2006 ), 并对植物体内糖代谢起到解耦联作用(
Pego et al ., 1999 ).总可溶性糖主要包括葡萄糖、果糖、蔗糖、多聚糖等.葡萄糖和果糖作为单糖直接供给植物能量, 但葡萄糖比果糖分解速度更快, 其能量释放也更快.蔗糖是植物光合作用的主要产物, 是同化产物由“源”向“库”运输的主要形式(
Farrar et al ., 2000 ), 蔗糖作为非还原性糖, 分解为果糖和葡萄糖后再向植物供应能量(
Quentin et al ., 2015 ). ...
 ,3,4,*, 张殷波
,3,4,*, 张殷波
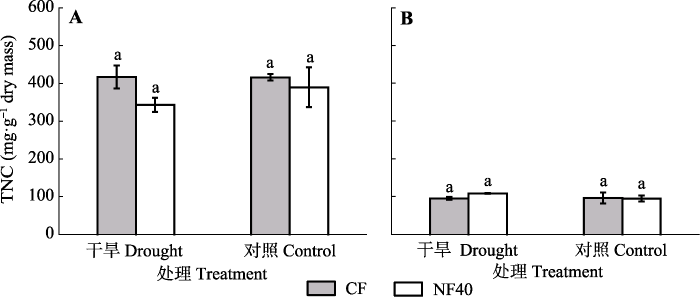 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT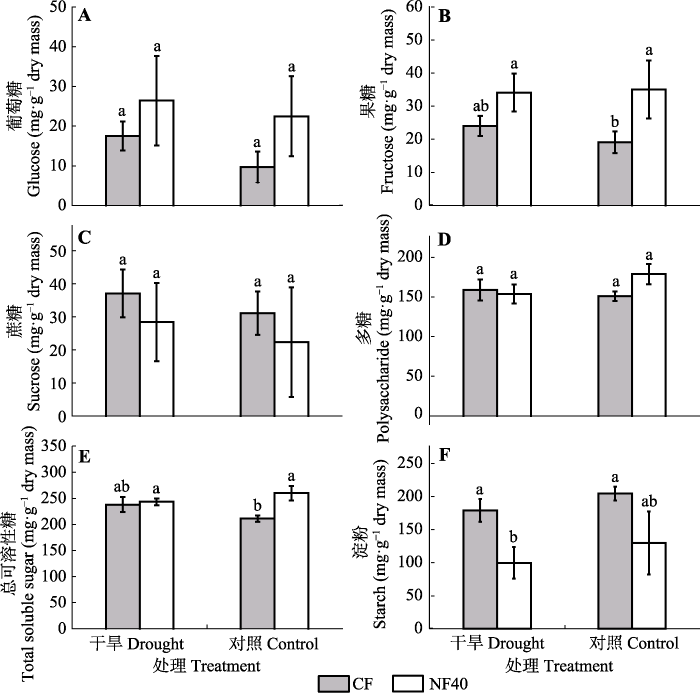 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT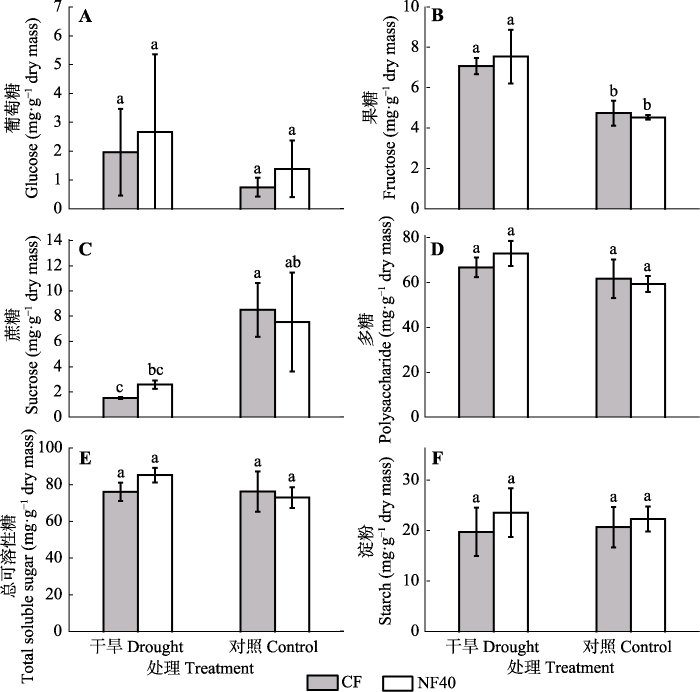 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT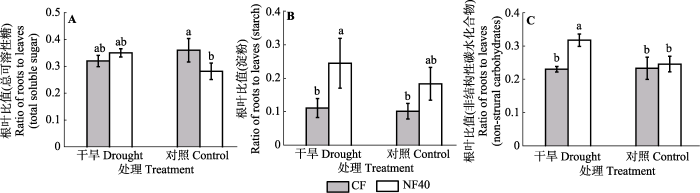 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}