Effects of simulated nitrogen deposition on growth and photosynthetic characteristics of Quercus wutaishanica and Acer pictum subsp. mono in a warm-temperate deciduous broad- leaved forest
Xiao-Ming LIU1,2, Xiao-Fang YANG1,2, Xuan WANG1,2, Shou-Ren ZHANG,1,*1 State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China 2 University of Chinese Academy of Sciences, Beijing 100049, China
Abstract Aims Warm-temperate deciduous broad-leaved forest plays an important role in maintaining regional ecosystem function and balance. To explore the growth and eco-physiological response to nitrogen (N) addition will deepen our understanding of its dynamic development under the scenario of global N deposition. Methods A simulated N deposition experiment was established with four N addition plots (100 kg·hm -2·a -1) and four control plots in a deciduous broad-leaved forest in Dongling Mountain, Beijing. The responses of branch growth, photosynthesis, chlorophyll fluorescence and seed mass of dominant species Quercus wutaishanica and associate species Acer pictum subsp. mono to N addition were investigated. Important findings Net photosynthetic rate, transpiration rate, and chlorophyll content were enhanced by the N addition. Furthermore, the N addition also enlarged the electron transport pool in photosystem II reaction center and increased the turnover number of the primary quinone acceptor (QA) reduction expressed by fast chlorophyll fluorescence method of JIP test in both species. The N addition also increased the length and biomass of the current year branches of the two tree species and improved the seed quality of Q. wutaishanica to a certain extent. In general, we observed more pronounced responses of photosynthetic nitrogen use efficiency, and branch biomass formation to N addition in Q. wutaishanica compared with A. pictum subsp. mono. Our results suggested that the dominant position of Q. wutaishanica should be further highlighted under increased N deposition scenario in the warm-temperate deciduous broad-leaved forest in Dongling Mountain. Keywords:nitrogen deposition;warm-temperate deciduous broad-leaved forest;photosynthesis;chlorophyll fluorescence;growth
PDF (1275KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 刘校铭, 杨晓芳, 王璇, 张守仁. 暖温带落叶阔叶林辽东栎和五角枫生长和光合生理生态特征对模拟氮沉降的响应. 植物生态学报, 2019, 43(3): 197-207. DOI: 10.17521/cjpe.2018.0303 LIU Xiao-Ming, YANG Xiao-Fang, WANG Xuan, ZHANG Shou-Ren. Effects of simulated nitrogen deposition on growth and photosynthetic characteristics of Quercus wutaishanica and Acer pictum subsp. mono in a warm-temperate deciduous broad- leaved forest. Chinese Journal of Plant Ecology, 2019, 43(3): 197-207. DOI: 10.17521/cjpe.2018.0303
从工业革命爆发以来, 化石燃料大量燃烧, 土地利用发生大规模改变, 农业、畜牧业快速发展。活跃的工农业生产活动使全球大气中的氮化物浓度不断增加(Vitousek et al., 1997)。在全球范围内, 由人类活动导致的氮沉降增加主要集中于北美、欧洲、东南亚三大地区(Galloway et al., 2003), 中国经济发达的东部地区处于全球三大高氮沉降区之一的东南亚地区。随着大规模的工业化生产和农业施肥活动, 进入陆地生态系统的氮不断增加, 经济发达的温带地区最为明显(莫江明等, 2006)。根据Galloway等(2002)的预测, 未来数十年内, 随着经济的进一步发展, 这种状况会不断加剧。氮富集及氮驱动力的预测模型表明, 到21世纪中叶, 全球活性氮的排放速度将达到267 Tg·a-1, 亚洲地区由于人为活动产生的活性氮增长速度可能高达105.3 Tg·a-1 (Holland et al., 1999; Zheng et al., 2002; Galloway et al., 2004, 2008)。近年来京津冀地区高浓度的氮氧化合物和其他可溶性气溶胶形成的雾霾引起人们的广泛关注, 该地区属全球典型的高氮沉降区(Zhang et al., 2012)。其氮湿沉降速率平均值为38 kg·hm-2·a-1, 无机氮沉降速率约为27 kg·hm-2·a-1, 有机氮沉降速率约为7 kg·hm-2·a-1, 氮的干沉降速率也高达50 kg·hm-2·a-1。Shen等(2009)研究表明, 未来北京地区总的大气氮沉降速率可超过100 kg·hm-2·a-1。
Table 1 Table 1Effects of nitrogen (N) treatment, tree species and their interaction on some eco-physiological and growth traits of Quercus wutaishanica and Acer pictum subsp. Mono in Dongling Mountain
性状 Trait
氮 N
树种 Species
氮 × 树种 N × Species
净光合速率 Net photosynthetic rate (Pn)
***
***
***
气孔导度 Stomatal conductance (Gs)
***
***
***
蒸腾速率 Transpiration rate (Tr)
***
***
***
水分利用效率 Water use Efficiency (WUE)
ns
ns
ns
叶绿素含量 Chlorophyll contents (Chl)
***
***
ns
光系统II最大光化学效率 Maximal PSII efficiency (PHI(Po))
ns
ns
ns
光系统II电子传递链电子受体库大小 PSII size of QA pool (Sm)
***
ns
ns
初级醌受体被还原周转次数 PSII QA reduced turn over number (N)
***
**
ns
光合氮利用效率 Photosynthetic nitrogen use efficiency (PNUE)
ns
ns
**
当年生枝长度 Length of current year branch
***
*
ns
当年生枝生物量 Biomass of current year branch
*
***
ns
辽东栎种子质量 Quercus wutaishanica seed mass
ns
-
-
-, 不存在。 ***, p < 0.001; **, p < 0.01; *, p < 0.05; ns, p > 0.05. -, not applicable.
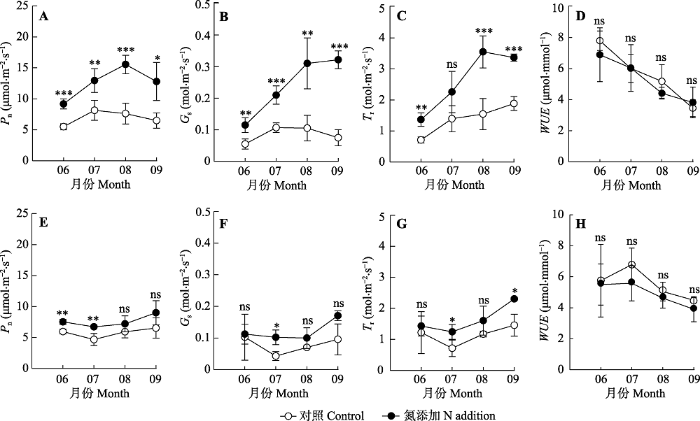
新窗口打开|下载原图ZIP|生成PPT 图1氮处理随季节变化对东灵山辽东栎(A-D)和五角枫(E-H)光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)和水分利用效率(WUE)的影响(平均值±标准偏差)。***, p < 0.001; **, p < 0.01; *, p < 0.05; ns, p > 0.05。
Fig. 1Seasonal changes of net photosynthetic rate (Pn), stomatal conductance (Gs), transpiration rate (Tr), and water use efficiency (WUE) of Quercus wutaishanica (A-D) and Acer pictum subsp. mono (E-H) in Dongling Mountain in response to nitrogen treatment (mean ± SD). ***, p < 0.001; **, p < 0.01; *, p < 0.05; ns, p > 0.05.
新窗口打开|下载原图ZIP|生成PPT 图2氮处理对辽东栎(A)和五角枫(B)叶绿素含量(Chl)的季节性影响(平均值±标准偏差)。**, p < 0.01; *, p < 0.05; ns, p > 0.05。
Fig. 2Seasonal changes of chlorophyll content (Chl) of Quercus wutaishanica (A) and Acer pictum subsp. mono (B) in response to nitrogen treatment (mean ± SD). **, p < 0.01; *, p < 0.05; ns, p > 0.05.
新窗口打开|下载原图ZIP|生成PPT 图3氮处理对辽东栎(A-C)和五角枫(D-F)叶绿素荧光性状光系统II最大光化学效率(PHI(Po))、 电子受体库大小(Sm)和初级醌受体被还原周转次数(N)的季节性影响(平均值±标准偏差)。**, p < 0.01; *, p < 0.05; ns, p > 0.05。由于仪器出现故障7月份数据缺失。
Fig. 3Seasonal changes of maximal PSII efficiency (PHI(Po)), size of QA pool (Sm) and the QA reduced turnover number (N) of Quercus wutaishanica (A-C) and Acer pictum subsp. mono (D-F) in response to nitrogen treatment (mean ± SD). **, p < 0.01; *, p < 0.05; ns, p > 0.05. Data measured in July were missed due to instrument disorder.
Fig. 4Effects of nitrogen addition, tree species and their interaction on photosynthetic nitrogen efficiency (PNUE)(A), length (B) and biomass (C) of the current-year branch of the two tree species, and effect of nitrogen addition on single seed mass (D) of Quercus wutaishanica (mean ± SD). Different lowercase letters indicate significant difference (p < 0.05).
AnderssonF, Fagerstr?mT, NilssonSI ( 1980). Forest ecosystem responses to acid deposition hydrogen ion budget and nitrogen/tree growth model approaches. In: Hutchinson TC, Havas M eds Effects of Acid Precipitation on Terrestrial Ecosystems. Springer,Boston. 319-334. [本文引用: 1]
BauerGA, BerntsonGM, BazzazFA ( 2010). Regenerating temperate forests under elevated CO2 and nitrogen deposition: Comparing biochemical and stomatal limitation of photosynthesis New Phytologist, 152, 249-266. [本文引用: 1]
BedisonJE, McneilBE ( 2009). Is the growth of temperate forest trees enhanced along an ambient nitrogen deposition gradient? Ecology, 90, 1736-1742. [本文引用: 1]
ChenDK, ShiJC, WangYH, ChenXQ ( 1984). Forest site classification and forest productivity classification and evaluation of secondary forest in Maoer Mountains Journal of Northeast Forestry University, 12(1), 1-18. [本文引用: 2]
ChenLZ ( 1997). The importance of Donglin Mountain region of warm temperate deciduous broad-leaved forest In: Chen LZ ed. The Study on Structure and Function of Forest in Warm Temperate Zone. Science Press, Beijing. 1-9. [本文引用: 2]
FangYT, MoJM, ZhouGY, XueJH ( 2005). Responses of diameter at breast height increment to N addition in forests of Dinghushan Biosphere Reserve Journal of Tropical and Subtropical Botany, 13, 198-204. [本文引用: 1]
GallowayJN, TownsendAR, Jan WillemE, MateeteB, ZucongC, FreneyJR, MartinelliLA, SeitzingerSP, SuttonMA ( 2008). Transformation of the nitrogen cycle: Recent trends, questions, and potential solutions Science, 320, 889-892. [本文引用: 1]
GarciaMO, OvasapyanT, GreasM, TresederKK ( 2008). Mycorrhizal dynamics under elevated CO2 and nitrogen fertilization in a warm temperate forest Plant and Soil, 303, 301-310. [本文引用: 1]
GüsewellS ( 2010). High nitrogen: Phosphorus ratios reduce nutrient retention and second-year growth of wetland sedges New Phytologist, 166, 537-550. [本文引用: 1]
HikosakaK, ShigenoA ( 2009). The role of Rubisco and cell walls in the interspecific variation in photosynthetic capacity Oecologia, 160, 443-451. [本文引用: 1]
H?gbergP, FanH, QuistM, BinkleyD, TammCO ( 2010). Tree growth and soil acidification in response to 30 years of experimental nitrogen loading on boreal forest Global Change Biology, 12, 489-499. [本文引用: 1]
HollandEA, DentenerFJ, BraswellBH, SulzmanJM ( 1999). Contemporary and pre-industrial global reactive nitrogen budgets Biogeochemistry, 46, 7-43. [本文引用: 1]
JiaS, WangZ, LiX, ZhangX, MclaughlinNB ( 2011). Effect of nitrogen fertilizer, root branch order and temperature on respiration and tissue N concentration of fine roots in Larix gmelinii and Fraxinus mandshurica Tree Physiology, 31, 718-726. [本文引用: 1]
JohnsonGN, ScholesJD, HortonP, YoungAJ ( 1993). Relationships between carotenoid composition and growth habit in British plant species. Plant, Cell & Environment, 16, 681-686. [本文引用: 1]
LebauerDS, TresederKK ( 2008). Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed Ecology, 89, 371-379. [本文引用: 1]
LiJX, LiF ( 2012). Characterization of chemical compounds of rainfall samples collected in a sand-storm process in Beijing China Environmental Science, 32, 2149-2154. [本文引用: 1]
LiPM, GaoHY, StrasserRJ ( 2005). Application of the fast chlorophyll fluorescence induction dynamics analysis in photosynthesis study Journal of Plant Physiology and Molecular Biology, 31, 559-566. [本文引用: 1]
LiQJ, YinLM, WangLH, LiuY ( 2010). Effects of simulated nitrogen deposition under different soil moistures on fruiting of Xanthoceras sorbifolia Liaoning Forestry Science and Technology, ( 3), 1-3. [本文引用: 1]
LiuXZ, WangGA, LiJZ, WangQ ( 2009). Nitrogen isotope composition of modern plants in Dongling Mountain area of Beijing and its response to altitude gradient Science in China Series D-Earth Sciences, 10, 1347-1359. [本文引用: 1]
LiuYC, YuGR, WangQF, ZhangYJ , ( 2012). Huge carbon sequestration potential in global forests Journal of Resources and Ecology, 3(3), 193-201. [本文引用: 1]
LuXK, MaoQG, GilliamFS, LuoY, MoJM ( 2014). Nitrogen deposition contributes to soil acidification in tropical ecosystems Global Change Biology, 20, 3790-3801. [本文引用: 1]
LuXK, MoJM, GilliamFS, YuG, WeiZ, FangYT, HuangJ ( 2011). Effects of experimental nitrogen additions on plant diversity in tropical forests of contrasting disturbance regimes in southern China Environmental Pollution, 159, 2228-2235. [本文引用: 1]
LuXK, MoJM, GilliamFS, ZhouGY, FangYT ( 2010). Effects of experimental nitrogen additions on plant diversity in an old-growth tropical forest Global Change Biology, 16, 2688-2700. [本文引用: 1]
LuoXS, LiuP, TangAH, LiuJY, ZongXY, ZhangQ, KouCL, ZhangLJ, FowlerD, FangmeierA, ChristieP, ZhangFS, LiuXJ ( 2013). An evaluation of atmospheric Nr pollution and deposition in North China after the Beijing Olympics Atmospheric Environment, 74, 209-216. [本文引用: 1]
MaSY, VerheyenK, PropsR, WasofS, VanhellemontM, BoeckxP, BoonN, de Frenne1P ( 2018). Plant and soil microbe responses to light, warming and nitrogen addition in a temperate forest Functional Ecology, 32, 1293-1303. [本文引用: 1]
MarianaV, CampanelloPI, BucciSJ, GuillermoG ( 2013). Functional relationships between leaf hydraulics and leaf economic traits in response to nutrient addition in subtropical tree species Tree physiology, 33, 1308-1318. [本文引用: 1]
MatsubaraS, KrauseGH, ArandaJ, VirgoA, BeiselKG, JahnsP, WinterK ( 2009). Sun-shade patterns of leaf carotenoid composition in 86 species of neotropical forest plants Functional Plant Biology, 36, 20-36. [本文引用: 1]
MoJM, BrownS, PengS, KongG ( 2003). Nitrogen availability in disturbed, rehabilitated and mature forests of tropical China Forest Ecology and Management, 175, 573-583. [本文引用: 1]
MoJM, FangYT, LinED, LiYE ( 2006). Soil N2O emission and its response to simulated N deposition in the main forests of Dinghushan in subtropical China Journal of Plant Ecology (Chinese version), 30, 901-910. [本文引用: 1]
MoJM, XueJH, FangYT ( 2004). Litter decomposition and its responses to simulated N deposition for the major plants of Dinghushan forests in subtropical China Acta Ecologica Sinica, 24, 1413-1420. [本文引用: 1]
NaveLE, GoughCM, MaurerKD, BohrerG, HardimanBS, MoineL ( 2015). Disturbance and the resilience of coupled carbon and nitrogen cycling in a north temperate forest Journal of Geophysical Research Biogeosciences, 116, 332-335. [本文引用: 1]
PalmrothS, BachLH, NordinA, PalmqvistK ( 2014). Nitrogen-?addition effects on leaf traits and photosynthetic carbon gain of boreal forest understory shrubs Oecologia, 175, 457-470. [本文引用: 1]
PanYP, WangYS, TanGQ, WuD ( 2012). Spatial distribution and seasonal variations of atmospheric sulfur deposition over northern China Atmospheric Chemistry and Physics Discussions, 12, 23645-23677. [本文引用: 1]
PengLQ, JingZX, WangQ ( 2014). Effects of simulated nitrogen deposition on the eco-physiological characteristics of Sinocalycanthus chinensis seedlings Chinese Journal of Ecology, 33, 989-995. [本文引用: 1]
Pérez-SobaM, StulenI, EerdenLJMVD ( 2010). Effect of atmospheric ammonia on the nitrogen metabolism of Scots pine (Pinus sylvestris) needles Physiologia Plantarum, 90, 629-636. [本文引用: 1]
QuanQ, ZhangZ, HeNP, SuHX, WenXF, SunXM ( 2015). Short-term effect of nitrogen addition on soil respiration of three temperate forests in Dongling Mountain Chinese Journal of Ecology, 34, 797-804 [本文引用: 3]
ShenJL, TangAH, LiuXJ, FangmeierA, GouldingKTW, ZhangFS ( 2009). High concentrations and dry deposition of reactive nitrogen species at two sites in the North China Plain Environmental Pollution, 157, 3106-3113. [本文引用: 1]
ShenYG, YeJY ( 1991). Relationship between dynamics of Thylakoid Membrane and energy transduction Plant Physiology Journal, 17, 109-112. [本文引用: 1]
StrasserRJ, SrivastavaA , Govindjee ( 1995). Polyphasic chlorophyll a fluorescence transient in plants and cyanobacteria photochem photobiol Photochemistry and Photobiology, 61, 32-42. [本文引用: 2]
StrasserRJ, Tsimilli-MichaelM, SrivastavaA ( 2004). Analysis of the chlorophyll a fluorescence transient In: Papageorgiou GC, Govindjee eds. Chlorophyll a Fluorescence Advances in Photosynthesis and Respiration. Springer, Dordrecht. 321-326. [本文引用: 2]
SunSZ ( 1997). The characteristics of the geology, geomorphology, and soils in Dongling Mountain region In: Chen LZ ed. The Study on Structure and Function of Forest in Warm Temperate Zone. Science Press,Beijing. 10-27. [本文引用: 1]
SunXL, XuYF, MaLY, ZhouH ( 2010). A review of acclimation of photosynthetic pigment composition in plant leaves to shade environment Chinese Journal of Plant Ecology, 34, 989-999. [本文引用: 1]
VitousekPM, AberJD, HowarthRW, LikensGE, MatsonPA, SchindlerDW, SchlesingerWH, TilmanD ( 1997). Human alteration of the global nitrogen cycle: Sources and consequences Ecological Applications, 7, 737-750. [本文引用: 1]
WangDZ, NieLS, LiJY ( 2006). Transfer characteristics of nutrient elements through hydrological process of Pinus tabulaeformis stand in Beijing Xishan area Acta Ecologica Sinica, 26, 2101-2107. [本文引用: 1]
WangM, ShiS, LinF, HaoZQ, JiangP, DaiGH ( 2012). Effects of soil water and nitrogen on growth and photosynthetic response of Manchurian ash (Fraxinus mandshurica) seedlings in northeastern China PLOS ONE, 7, e30754. DOI: 10.1371/journal.pone.0030754. [本文引用: 1]
WangXA, WangZG, XiaoYP ( 2004). Studies on the reproductive strategy of Larix chinensis I fecundity and seed weight Journal of Lanzhou University (Natural Sciences), 40, 72-75. [本文引用: 1]
YangWQ, LiuSH, MiaoM, GaoXD, ChenXY ( 2016). Spatial and temporal variation of dissolved inorganic nitrogen deposition along an urban to rural transect in Beijing Acta Scientiae Circumstantiae, 36, 1530-1538. [本文引用: 1]
ZhangJH, TangZY, ShenHH, FangJY ( 2017). Responses of growth and litterfall production to nitrogen addition treatments from common shrublands in Mt. Dongling, Beijing, China Chinese Journal of Plant Ecology, 41, 71-80. [本文引用: 1]
ZhengXH, FuCB, XuXK, YanXD, HuangY, HanSH, HuF, ChenGX ( 2002). The Asian nitrogen cycle case study Ambio, 31, 79-87. [本文引用: 1]
ZhuFF, YohM, GilliamFS, LuXK, MoJM ( 2013). Nutrient limitation in three lowland tropical forests in southern China receiving high nitrogen deposition: Insights from fine root responses to nutrient additions PLOS ONE, 8, e82661. DOI: 10.1371/journal.pone.0082661. [本文引用: 1]
Nitrogen saturation in temperate forest ecosystems hypotheses revisited 1 1998
... 森林生态系统是陆地生态系统的重要组成部分, 无论是在群落组成、更新演替还是生态环境保护、能源、经济方面都发挥着十分重要的作用.长时间持续的氮沉降会影响森林生态系统的功能(Güsewell, 2010; Liu et al., 2013).树木生长对氮沉降产生响应所导致的森林结构的改变, 又反过来影响全球气候(Aber et al., 1998; H?gberg et al., 2010). ...
Forest ecosystem responses to acid deposition hydrogen ion budget and nitrogen/tree growth model approaches. In: Hutchinson TC, Havas M eds 1 1980
... 氮沉降对温带森林影响研究从20世纪中叶开始, 已超过30年(Andersson et al., 1980; Garcia et al., 2008; Nave et al., 2015; Ma et al., 2018), 已经取得了不少成果, 但对于目前大气氮沉降如何长期影响树木生长和植被动态变化的认知程度依然十分有限(Bedison & Mcneil, 2009).近20年来国内陆续开展了关于氮沉降量增加对森林生态系统影响的研究(Mo et al., 2003; 方运霆等, 2004; 莫江明等, 2004;方华和莫江明, 2006; Lu et al., 2010; Lu et al., 2011; Zhu et al., 2013).由于在野外自然条件下进行控制实验的复杂性和高难度, 已有的氮沉降控制实验的研究对象大多以幼苗为主, 以自然森林成年大树为实验对象的研究较少(方运霆等, 2005), 基于幼苗或幼树对氮沉降的响应结果推测自然成熟森林的情况, 其代表性存在很大局限性. ...
Regenerating temperate forests under elevated CO2 and nitrogen deposition: Comparing biochemical and stomatal limitation of photosynthesis 1 2010
... 光合作用是绿色植物转化太阳能, 合成同化物质的重要生理过程, 是植物维持生长发育的基础.氮添加的浓度和时间会对植物的叶绿素合成产生不同的影响.本研究中氮添加处理提高了辽东栎和五角枫植物的叶片氮含量, 促进了叶片的叶绿素生成, 与前人研究结果(Bauer et al., 2010; Pérez-Soba et al.,2010; Jia et al., 2011; Wang et al., 2012; 郑翔等, 2018)一致.需要说明的是, 两树种间的净光合速率和叶绿素含量均存在显著差异, 辽东栎的光合速率显著高于五角枫, 而五角枫的叶绿素含量显著高于辽东栎.此外, 氮添加对叶片叶绿素含量的促进作用随着处理时间延长表现出明显的累积效应, 五角枫比辽东栎更为明显.氮添加对光合速率、气孔导度和蒸腾速率的促进作用也随着处理时间延长表现出明显的累积效应, 辽东栎比五角枫更明显.在本研究的实验样地中, 辽东栎树体高大, 占据上层空间, 树冠上中层可以接收到充足的太阳辐射, 是群落的建群种和优势种; 而样地内的五角枫树体矮小, 处于被遮阴的下层, 可接受光照少.不同的光照环境会导致树木同化组织在结构和功能方面的极大差异, 一般阳生植物比阴生植物具有更高的羧化酶活性, 而阴生植物为了尽可能地获得林下的散射光, 会提高叶绿素含量, 形成发达的捕获色素系统, 以截获更多的光能(Shen & Ye, 1991; Johnson et al., 1993; 张守仁等, 2004; Matsubara et al., 2009; 孙小玲等, 2010; 向芬等, 2018).叶片光合作用气体交换过程主要受气孔因素和非气孔因素(主要是Rubisco羧化酶活性)影响, 在饱和光下辽东栎的羧化速率(Vcmax)和气孔导度远高于五角枫(未发表数据), 这也解释了在饱和光下辽东栎光合速率高于五角枫, 而五角枫叶绿素含量高于辽东栎的原因. ...
Is the growth of temperate forest trees enhanced along an ambient nitrogen deposition gradient? 1 2009
... 氮沉降对温带森林影响研究从20世纪中叶开始, 已超过30年(Andersson et al., 1980; Garcia et al., 2008; Nave et al., 2015; Ma et al., 2018), 已经取得了不少成果, 但对于目前大气氮沉降如何长期影响树木生长和植被动态变化的认知程度依然十分有限(Bedison & Mcneil, 2009).近20年来国内陆续开展了关于氮沉降量增加对森林生态系统影响的研究(Mo et al., 2003; 方运霆等, 2004; 莫江明等, 2004;方华和莫江明, 2006; Lu et al., 2010; Lu et al., 2011; Zhu et al., 2013).由于在野外自然条件下进行控制实验的复杂性和高难度, 已有的氮沉降控制实验的研究对象大多以幼苗为主, 以自然森林成年大树为实验对象的研究较少(方运霆等, 2005), 基于幼苗或幼树对氮沉降的响应结果推测自然成熟森林的情况, 其代表性存在很大局限性. ...
... 氮沉降对温带森林影响研究从20世纪中叶开始, 已超过30年(Andersson et al., 1980; Garcia et al., 2008; Nave et al., 2015; Ma et al., 2018), 已经取得了不少成果, 但对于目前大气氮沉降如何长期影响树木生长和植被动态变化的认知程度依然十分有限(Bedison & Mcneil, 2009).近20年来国内陆续开展了关于氮沉降量增加对森林生态系统影响的研究(Mo et al., 2003; 方运霆等, 2004; 莫江明等, 2004;方华和莫江明, 2006; Lu et al., 2010; Lu et al., 2011; Zhu et al., 2013).由于在野外自然条件下进行控制实验的复杂性和高难度, 已有的氮沉降控制实验的研究对象大多以幼苗为主, 以自然森林成年大树为实验对象的研究较少(方运霆等, 2005), 基于幼苗或幼树对氮沉降的响应结果推测自然成熟森林的情况, 其代表性存在很大局限性. ...
氮沉降对森林凋落物分解的影响 1 2006
... 氮沉降对温带森林影响研究从20世纪中叶开始, 已超过30年(Andersson et al., 1980; Garcia et al., 2008; Nave et al., 2015; Ma et al., 2018), 已经取得了不少成果, 但对于目前大气氮沉降如何长期影响树木生长和植被动态变化的认知程度依然十分有限(Bedison & Mcneil, 2009).近20年来国内陆续开展了关于氮沉降量增加对森林生态系统影响的研究(Mo et al., 2003; 方运霆等, 2004; 莫江明等, 2004;方华和莫江明, 2006; Lu et al., 2010; Lu et al., 2011; Zhu et al., 2013).由于在野外自然条件下进行控制实验的复杂性和高难度, 已有的氮沉降控制实验的研究对象大多以幼苗为主, 以自然森林成年大树为实验对象的研究较少(方运霆等, 2005), 基于幼苗或幼树对氮沉降的响应结果推测自然成熟森林的情况, 其代表性存在很大局限性. ...
森林土壤氮素转换及其对氮沉降的响应 1 2004
... 氮沉降对温带森林影响研究从20世纪中叶开始, 已超过30年(Andersson et al., 1980; Garcia et al., 2008; Nave et al., 2015; Ma et al., 2018), 已经取得了不少成果, 但对于目前大气氮沉降如何长期影响树木生长和植被动态变化的认知程度依然十分有限(Bedison & Mcneil, 2009).近20年来国内陆续开展了关于氮沉降量增加对森林生态系统影响的研究(Mo et al., 2003; 方运霆等, 2004; 莫江明等, 2004;方华和莫江明, 2006; Lu et al., 2010; Lu et al., 2011; Zhu et al., 2013).由于在野外自然条件下进行控制实验的复杂性和高难度, 已有的氮沉降控制实验的研究对象大多以幼苗为主, 以自然森林成年大树为实验对象的研究较少(方运霆等, 2005), 基于幼苗或幼树对氮沉降的响应结果推测自然成熟森林的情况, 其代表性存在很大局限性. ...
森林土壤氮素转换及其对氮沉降的响应 1 2004
... 氮沉降对温带森林影响研究从20世纪中叶开始, 已超过30年(Andersson et al., 1980; Garcia et al., 2008; Nave et al., 2015; Ma et al., 2018), 已经取得了不少成果, 但对于目前大气氮沉降如何长期影响树木生长和植被动态变化的认知程度依然十分有限(Bedison & Mcneil, 2009).近20年来国内陆续开展了关于氮沉降量增加对森林生态系统影响的研究(Mo et al., 2003; 方运霆等, 2004; 莫江明等, 2004;方华和莫江明, 2006; Lu et al., 2010; Lu et al., 2011; Zhu et al., 2013).由于在野外自然条件下进行控制实验的复杂性和高难度, 已有的氮沉降控制实验的研究对象大多以幼苗为主, 以自然森林成年大树为实验对象的研究较少(方运霆等, 2005), 基于幼苗或幼树对氮沉降的响应结果推测自然成熟森林的情况, 其代表性存在很大局限性. ...
鼎湖山主要森林类型植物胸径生长对氮沉降增加的初期响应 1 2005
... 氮沉降对温带森林影响研究从20世纪中叶开始, 已超过30年(Andersson et al., 1980; Garcia et al., 2008; Nave et al., 2015; Ma et al., 2018), 已经取得了不少成果, 但对于目前大气氮沉降如何长期影响树木生长和植被动态变化的认知程度依然十分有限(Bedison & Mcneil, 2009).近20年来国内陆续开展了关于氮沉降量增加对森林生态系统影响的研究(Mo et al., 2003; 方运霆等, 2004; 莫江明等, 2004;方华和莫江明, 2006; Lu et al., 2010; Lu et al., 2011; Zhu et al., 2013).由于在野外自然条件下进行控制实验的复杂性和高难度, 已有的氮沉降控制实验的研究对象大多以幼苗为主, 以自然森林成年大树为实验对象的研究较少(方运霆等, 2005), 基于幼苗或幼树对氮沉降的响应结果推测自然成熟森林的情况, 其代表性存在很大局限性. ...
鼎湖山主要森林类型植物胸径生长对氮沉降增加的初期响应 1 2005
... 氮沉降对温带森林影响研究从20世纪中叶开始, 已超过30年(Andersson et al., 1980; Garcia et al., 2008; Nave et al., 2015; Ma et al., 2018), 已经取得了不少成果, 但对于目前大气氮沉降如何长期影响树木生长和植被动态变化的认知程度依然十分有限(Bedison & Mcneil, 2009).近20年来国内陆续开展了关于氮沉降量增加对森林生态系统影响的研究(Mo et al., 2003; 方运霆等, 2004; 莫江明等, 2004;方华和莫江明, 2006; Lu et al., 2010; Lu et al., 2011; Zhu et al., 2013).由于在野外自然条件下进行控制实验的复杂性和高难度, 已有的氮沉降控制实验的研究对象大多以幼苗为主, 以自然森林成年大树为实验对象的研究较少(方运霆等, 2005), 基于幼苗或幼树对氮沉降的响应结果推测自然成熟森林的情况, 其代表性存在很大局限性. ...
... 北京东灵山(39.92°-40.03° N, 115.48°-115.98° E)位于北京市西郊的门头沟区西部, 毗邻河北省, 距市区约115 km.东灵山位于小五台山东部余脉, 属太行山系.山峰海拔高度多高于1 000 m, 最高峰为2 303 m (冯云等, 2007).该地区属于暖温带半湿润大陆性季风气候, 春季干旱, 夏季暖热多雨, 冬季寒冷干燥(全权等, 2015).年平均气温5-10 ℃, 无霜期约195天, 年降水量500-650 mm, 且多集中于夏季, 夏季降水约占全年降水量的75% (冯云等, 2007; 全权等, 2015).该地处于华北地区中部, 地质构造上位于燕山沉降带西部, 地貌以山地侵蚀结构为主, 地势险峻, 谷深坡陡, 河流下切严重, 土壤主要为肥沃的褐色土和棕色的森林土(孙世洲, 1997).植被类型较丰富, 以辽东栎为主要树种, 五角枫、白蜡树(Fraxinus chinensis)、黄荆(Vitex negundo)、山杏(Armeniaca sibirica)和绣线菊(Spiraea salicifolia)为伴生树种构成典型的辽东栎林(刘贤赵等, 2009; 张建华等, 2017).根据北京城区和河北多个观测点近10年的干湿氮沉降数据, 北京南部和东部大气氮沉降速率可以高达80 kg·hm-2·a-1, 北京西部地区略低, 但高峰月份也可以达到50 kg·hm-2·a-1 (王登芝等, 2006; 李金香和李峰, 2012; Pan et al., 2012; Luo et al., 2013; Xu et al., 2015; 杨文琴等, 2016; 尹兴等, 2017). ...
... ).年平均气温5-10 ℃, 无霜期约195天, 年降水量500-650 mm, 且多集中于夏季, 夏季降水约占全年降水量的75% (冯云等, 2007; 全权等, 2015).该地处于华北地区中部, 地质构造上位于燕山沉降带西部, 地貌以山地侵蚀结构为主, 地势险峻, 谷深坡陡, 河流下切严重, 土壤主要为肥沃的褐色土和棕色的森林土(孙世洲, 1997).植被类型较丰富, 以辽东栎为主要树种, 五角枫、白蜡树(Fraxinus chinensis)、黄荆(Vitex negundo)、山杏(Armeniaca sibirica)和绣线菊(Spiraea salicifolia)为伴生树种构成典型的辽东栎林(刘贤赵等, 2009; 张建华等, 2017).根据北京城区和河北多个观测点近10年的干湿氮沉降数据, 北京南部和东部大气氮沉降速率可以高达80 kg·hm-2·a-1, 北京西部地区略低, 但高峰月份也可以达到50 kg·hm-2·a-1 (王登芝等, 2006; 李金香和李峰, 2012; Pan et al., 2012; Luo et al., 2013; Xu et al., 2015; 杨文琴等, 2016; 尹兴等, 2017). ...
The nitrogen cascade 1 2003
... 从工业革命爆发以来, 化石燃料大量燃烧, 土地利用发生大规模改变, 农业、畜牧业快速发展.活跃的工农业生产活动使全球大气中的氮化物浓度不断增加(Vitousek et al., 1997).在全球范围内, 由人类活动导致的氮沉降增加主要集中于北美、欧洲、东南亚三大地区(Galloway et al., 2003), 中国经济发达的东部地区处于全球三大高氮沉降区之一的东南亚地区.随着大规模的工业化生产和农业施肥活动, 进入陆地生态系统的氮不断增加, 经济发达的温带地区最为明显(莫江明等, 2006).根据Galloway等(2002)的预测, 未来数十年内, 随着经济的进一步发展, 这种状况会不断加剧.氮富集及氮驱动力的预测模型表明, 到21世纪中叶, 全球活性氮的排放速度将达到267 Tg·a-1, 亚洲地区由于人为活动产生的活性氮增长速度可能高达105.3 Tg·a-1 (Holland et al., 1999; Zheng et al., 2002; Galloway et al., 2004, 2008).近年来京津冀地区高浓度的氮氧化合物和其他可溶性气溶胶形成的雾霾引起人们的广泛关注, 该地区属全球典型的高氮沉降区(Zhang et al., 2012).其氮湿沉降速率平均值为38 kg·hm-2·a-1, 无机氮沉降速率约为27 kg·hm-2·a-1, 有机氮沉降速率约为7 kg·hm-2·a-1, 氮的干沉降速率也高达50 kg·hm-2·a-1.Shen等(2009)研究表明, 未来北京地区总的大气氮沉降速率可超过100 kg·hm-2·a-1. ...
Reactive nitrogen: Too much of a good thing? 1 2002
... 从工业革命爆发以来, 化石燃料大量燃烧, 土地利用发生大规模改变, 农业、畜牧业快速发展.活跃的工农业生产活动使全球大气中的氮化物浓度不断增加(Vitousek et al., 1997).在全球范围内, 由人类活动导致的氮沉降增加主要集中于北美、欧洲、东南亚三大地区(Galloway et al., 2003), 中国经济发达的东部地区处于全球三大高氮沉降区之一的东南亚地区.随着大规模的工业化生产和农业施肥活动, 进入陆地生态系统的氮不断增加, 经济发达的温带地区最为明显(莫江明等, 2006).根据Galloway等(2002)的预测, 未来数十年内, 随着经济的进一步发展, 这种状况会不断加剧.氮富集及氮驱动力的预测模型表明, 到21世纪中叶, 全球活性氮的排放速度将达到267 Tg·a-1, 亚洲地区由于人为活动产生的活性氮增长速度可能高达105.3 Tg·a-1 (Holland et al., 1999; Zheng et al., 2002; Galloway et al., 2004, 2008).近年来京津冀地区高浓度的氮氧化合物和其他可溶性气溶胶形成的雾霾引起人们的广泛关注, 该地区属全球典型的高氮沉降区(Zhang et al., 2012).其氮湿沉降速率平均值为38 kg·hm-2·a-1, 无机氮沉降速率约为27 kg·hm-2·a-1, 有机氮沉降速率约为7 kg·hm-2·a-1, 氮的干沉降速率也高达50 kg·hm-2·a-1.Shen等(2009)研究表明, 未来北京地区总的大气氮沉降速率可超过100 kg·hm-2·a-1. ...
Nitrogen cycles: Past, present, and future 1 2004
... 从工业革命爆发以来, 化石燃料大量燃烧, 土地利用发生大规模改变, 农业、畜牧业快速发展.活跃的工农业生产活动使全球大气中的氮化物浓度不断增加(Vitousek et al., 1997).在全球范围内, 由人类活动导致的氮沉降增加主要集中于北美、欧洲、东南亚三大地区(Galloway et al., 2003), 中国经济发达的东部地区处于全球三大高氮沉降区之一的东南亚地区.随着大规模的工业化生产和农业施肥活动, 进入陆地生态系统的氮不断增加, 经济发达的温带地区最为明显(莫江明等, 2006).根据Galloway等(2002)的预测, 未来数十年内, 随着经济的进一步发展, 这种状况会不断加剧.氮富集及氮驱动力的预测模型表明, 到21世纪中叶, 全球活性氮的排放速度将达到267 Tg·a-1, 亚洲地区由于人为活动产生的活性氮增长速度可能高达105.3 Tg·a-1 (Holland et al., 1999; Zheng et al., 2002; Galloway et al., 2004, 2008).近年来京津冀地区高浓度的氮氧化合物和其他可溶性气溶胶形成的雾霾引起人们的广泛关注, 该地区属全球典型的高氮沉降区(Zhang et al., 2012).其氮湿沉降速率平均值为38 kg·hm-2·a-1, 无机氮沉降速率约为27 kg·hm-2·a-1, 有机氮沉降速率约为7 kg·hm-2·a-1, 氮的干沉降速率也高达50 kg·hm-2·a-1.Shen等(2009)研究表明, 未来北京地区总的大气氮沉降速率可超过100 kg·hm-2·a-1. ...
Transformation of the nitrogen cycle: Recent trends, questions, and potential solutions 1 2008
... 从工业革命爆发以来, 化石燃料大量燃烧, 土地利用发生大规模改变, 农业、畜牧业快速发展.活跃的工农业生产活动使全球大气中的氮化物浓度不断增加(Vitousek et al., 1997).在全球范围内, 由人类活动导致的氮沉降增加主要集中于北美、欧洲、东南亚三大地区(Galloway et al., 2003), 中国经济发达的东部地区处于全球三大高氮沉降区之一的东南亚地区.随着大规模的工业化生产和农业施肥活动, 进入陆地生态系统的氮不断增加, 经济发达的温带地区最为明显(莫江明等, 2006).根据Galloway等(2002)的预测, 未来数十年内, 随着经济的进一步发展, 这种状况会不断加剧.氮富集及氮驱动力的预测模型表明, 到21世纪中叶, 全球活性氮的排放速度将达到267 Tg·a-1, 亚洲地区由于人为活动产生的活性氮增长速度可能高达105.3 Tg·a-1 (Holland et al., 1999; Zheng et al., 2002; Galloway et al., 2004, 2008).近年来京津冀地区高浓度的氮氧化合物和其他可溶性气溶胶形成的雾霾引起人们的广泛关注, 该地区属全球典型的高氮沉降区(Zhang et al., 2012).其氮湿沉降速率平均值为38 kg·hm-2·a-1, 无机氮沉降速率约为27 kg·hm-2·a-1, 有机氮沉降速率约为7 kg·hm-2·a-1, 氮的干沉降速率也高达50 kg·hm-2·a-1.Shen等(2009)研究表明, 未来北京地区总的大气氮沉降速率可超过100 kg·hm-2·a-1. ...
Mycorrhizal dynamics under elevated CO2 and nitrogen fertilization in a warm temperate forest 1 2008
... 氮沉降对温带森林影响研究从20世纪中叶开始, 已超过30年(Andersson et al., 1980; Garcia et al., 2008; Nave et al., 2015; Ma et al., 2018), 已经取得了不少成果, 但对于目前大气氮沉降如何长期影响树木生长和植被动态变化的认知程度依然十分有限(Bedison & Mcneil, 2009).近20年来国内陆续开展了关于氮沉降量增加对森林生态系统影响的研究(Mo et al., 2003; 方运霆等, 2004; 莫江明等, 2004;方华和莫江明, 2006; Lu et al., 2010; Lu et al., 2011; Zhu et al., 2013).由于在野外自然条件下进行控制实验的复杂性和高难度, 已有的氮沉降控制实验的研究对象大多以幼苗为主, 以自然森林成年大树为实验对象的研究较少(方运霆等, 2005), 基于幼苗或幼树对氮沉降的响应结果推测自然成熟森林的情况, 其代表性存在很大局限性. ...
Atmospheric reactive nitrogen in China: Sources, recent trends, and damage costs 1 2012
High nitrogen: Phosphorus ratios reduce nutrient retention and second-year growth of wetland sedges 1 2010
... 森林生态系统是陆地生态系统的重要组成部分, 无论是在群落组成、更新演替还是生态环境保护、能源、经济方面都发挥着十分重要的作用.长时间持续的氮沉降会影响森林生态系统的功能(Güsewell, 2010; Liu et al., 2013).树木生长对氮沉降产生响应所导致的森林结构的改变, 又反过来影响全球气候(Aber et al., 1998; H?gberg et al., 2010). ...
The role of Rubisco and cell walls in the interspecific variation in photosynthetic capacity 1 2009
... 温带北方森林常年受到氮限制, 土壤氮增加会促进木本植物生长, 增加森林的固碳潜力(Liu et al., 2012).氮是植物合成叶片光合作用羧化酶——核酮糖-1,5-双磷酸羧化酶/加氧酶(Rubisco)和光合作用重要色素叶绿素的重要组分, 叶片氮含量的变化会直接引起光合作用酶含量和叶绿素含量的改变, 所以施氮会直接提高植物光合作用能力(Hikosaka & Shigeno, 2009; Palmroth et al., 2014).此外, 有关氮沉降影响净初级生产力的经验模型也指出增加的大气氮沉降量会导致木本植物的碳固持增加, 提高森林的初级生产力(Kurts et al., 2008; Lu et al., 2014). ...
Tree growth and soil acidification in response to 30 years of experimental nitrogen loading on boreal forest 1 2010
... 森林生态系统是陆地生态系统的重要组成部分, 无论是在群落组成、更新演替还是生态环境保护、能源、经济方面都发挥着十分重要的作用.长时间持续的氮沉降会影响森林生态系统的功能(Güsewell, 2010; Liu et al., 2013).树木生长对氮沉降产生响应所导致的森林结构的改变, 又反过来影响全球气候(Aber et al., 1998; H?gberg et al., 2010). ...
Contemporary and pre-industrial global reactive nitrogen budgets 1 1999
... 从工业革命爆发以来, 化石燃料大量燃烧, 土地利用发生大规模改变, 农业、畜牧业快速发展.活跃的工农业生产活动使全球大气中的氮化物浓度不断增加(Vitousek et al., 1997).在全球范围内, 由人类活动导致的氮沉降增加主要集中于北美、欧洲、东南亚三大地区(Galloway et al., 2003), 中国经济发达的东部地区处于全球三大高氮沉降区之一的东南亚地区.随着大规模的工业化生产和农业施肥活动, 进入陆地生态系统的氮不断增加, 经济发达的温带地区最为明显(莫江明等, 2006).根据Galloway等(2002)的预测, 未来数十年内, 随着经济的进一步发展, 这种状况会不断加剧.氮富集及氮驱动力的预测模型表明, 到21世纪中叶, 全球活性氮的排放速度将达到267 Tg·a-1, 亚洲地区由于人为活动产生的活性氮增长速度可能高达105.3 Tg·a-1 (Holland et al., 1999; Zheng et al., 2002; Galloway et al., 2004, 2008).近年来京津冀地区高浓度的氮氧化合物和其他可溶性气溶胶形成的雾霾引起人们的广泛关注, 该地区属全球典型的高氮沉降区(Zhang et al., 2012).其氮湿沉降速率平均值为38 kg·hm-2·a-1, 无机氮沉降速率约为27 kg·hm-2·a-1, 有机氮沉降速率约为7 kg·hm-2·a-1, 氮的干沉降速率也高达50 kg·hm-2·a-1.Shen等(2009)研究表明, 未来北京地区总的大气氮沉降速率可超过100 kg·hm-2·a-1. ...
Effect of nitrogen fertilizer, root branch order and temperature on respiration and tissue N concentration of fine roots in Larix gmelinii and Fraxinus mandshurica 1 2011
... 光合作用是绿色植物转化太阳能, 合成同化物质的重要生理过程, 是植物维持生长发育的基础.氮添加的浓度和时间会对植物的叶绿素合成产生不同的影响.本研究中氮添加处理提高了辽东栎和五角枫植物的叶片氮含量, 促进了叶片的叶绿素生成, 与前人研究结果(Bauer et al., 2010; Pérez-Soba et al.,2010; Jia et al., 2011; Wang et al., 2012; 郑翔等, 2018)一致.需要说明的是, 两树种间的净光合速率和叶绿素含量均存在显著差异, 辽东栎的光合速率显著高于五角枫, 而五角枫的叶绿素含量显著高于辽东栎.此外, 氮添加对叶片叶绿素含量的促进作用随着处理时间延长表现出明显的累积效应, 五角枫比辽东栎更为明显.氮添加对光合速率、气孔导度和蒸腾速率的促进作用也随着处理时间延长表现出明显的累积效应, 辽东栎比五角枫更明显.在本研究的实验样地中, 辽东栎树体高大, 占据上层空间, 树冠上中层可以接收到充足的太阳辐射, 是群落的建群种和优势种; 而样地内的五角枫树体矮小, 处于被遮阴的下层, 可接受光照少.不同的光照环境会导致树木同化组织在结构和功能方面的极大差异, 一般阳生植物比阴生植物具有更高的羧化酶活性, 而阴生植物为了尽可能地获得林下的散射光, 会提高叶绿素含量, 形成发达的捕获色素系统, 以截获更多的光能(Shen & Ye, 1991; Johnson et al., 1993; 张守仁等, 2004; Matsubara et al., 2009; 孙小玲等, 2010; 向芬等, 2018).叶片光合作用气体交换过程主要受气孔因素和非气孔因素(主要是Rubisco羧化酶活性)影响, 在饱和光下辽东栎的羧化速率(Vcmax)和气孔导度远高于五角枫(未发表数据), 这也解释了在饱和光下辽东栎光合速率高于五角枫, 而五角枫叶绿素含量高于辽东栎的原因. ...
Relationships between carotenoid composition and growth habit in British plant species. Plant, 1 1993
... 光合作用是绿色植物转化太阳能, 合成同化物质的重要生理过程, 是植物维持生长发育的基础.氮添加的浓度和时间会对植物的叶绿素合成产生不同的影响.本研究中氮添加处理提高了辽东栎和五角枫植物的叶片氮含量, 促进了叶片的叶绿素生成, 与前人研究结果(Bauer et al., 2010; Pérez-Soba et al.,2010; Jia et al., 2011; Wang et al., 2012; 郑翔等, 2018)一致.需要说明的是, 两树种间的净光合速率和叶绿素含量均存在显著差异, 辽东栎的光合速率显著高于五角枫, 而五角枫的叶绿素含量显著高于辽东栎.此外, 氮添加对叶片叶绿素含量的促进作用随着处理时间延长表现出明显的累积效应, 五角枫比辽东栎更为明显.氮添加对光合速率、气孔导度和蒸腾速率的促进作用也随着处理时间延长表现出明显的累积效应, 辽东栎比五角枫更明显.在本研究的实验样地中, 辽东栎树体高大, 占据上层空间, 树冠上中层可以接收到充足的太阳辐射, 是群落的建群种和优势种; 而样地内的五角枫树体矮小, 处于被遮阴的下层, 可接受光照少.不同的光照环境会导致树木同化组织在结构和功能方面的极大差异, 一般阳生植物比阴生植物具有更高的羧化酶活性, 而阴生植物为了尽可能地获得林下的散射光, 会提高叶绿素含量, 形成发达的捕获色素系统, 以截获更多的光能(Shen & Ye, 1991; Johnson et al., 1993; 张守仁等, 2004; Matsubara et al., 2009; 孙小玲等, 2010; 向芬等, 2018).叶片光合作用气体交换过程主要受气孔因素和非气孔因素(主要是Rubisco羧化酶活性)影响, 在饱和光下辽东栎的羧化速率(Vcmax)和气孔导度远高于五角枫(未发表数据), 这也解释了在饱和光下辽东栎光合速率高于五角枫, 而五角枫叶绿素含量高于辽东栎的原因. ...
... 森林生态系统是陆地生态系统的重要组成部分, 无论是在群落组成、更新演替还是生态环境保护、能源、经济方面都发挥着十分重要的作用.长时间持续的氮沉降会影响森林生态系统的功能(Güsewell, 2010; Liu et al., 2013).树木生长对氮沉降产生响应所导致的森林结构的改变, 又反过来影响全球气候(Aber et al., 1998; H?gberg et al., 2010). ...
北京东灵山地区现代植物氮同位素组成及其对海拔梯度的响应 1 2009
... 北京东灵山(39.92°-40.03° N, 115.48°-115.98° E)位于北京市西郊的门头沟区西部, 毗邻河北省, 距市区约115 km.东灵山位于小五台山东部余脉, 属太行山系.山峰海拔高度多高于1 000 m, 最高峰为2 303 m (冯云等, 2007).该地区属于暖温带半湿润大陆性季风气候, 春季干旱, 夏季暖热多雨, 冬季寒冷干燥(全权等, 2015).年平均气温5-10 ℃, 无霜期约195天, 年降水量500-650 mm, 且多集中于夏季, 夏季降水约占全年降水量的75% (冯云等, 2007; 全权等, 2015).该地处于华北地区中部, 地质构造上位于燕山沉降带西部, 地貌以山地侵蚀结构为主, 地势险峻, 谷深坡陡, 河流下切严重, 土壤主要为肥沃的褐色土和棕色的森林土(孙世洲, 1997).植被类型较丰富, 以辽东栎为主要树种, 五角枫、白蜡树(Fraxinus chinensis)、黄荆(Vitex negundo)、山杏(Armeniaca sibirica)和绣线菊(Spiraea salicifolia)为伴生树种构成典型的辽东栎林(刘贤赵等, 2009; 张建华等, 2017).根据北京城区和河北多个观测点近10年的干湿氮沉降数据, 北京南部和东部大气氮沉降速率可以高达80 kg·hm-2·a-1, 北京西部地区略低, 但高峰月份也可以达到50 kg·hm-2·a-1 (王登芝等, 2006; 李金香和李峰, 2012; Pan et al., 2012; Luo et al., 2013; Xu et al., 2015; 杨文琴等, 2016; 尹兴等, 2017). ...
北京东灵山地区现代植物氮同位素组成及其对海拔梯度的响应 1 2009
... 北京东灵山(39.92°-40.03° N, 115.48°-115.98° E)位于北京市西郊的门头沟区西部, 毗邻河北省, 距市区约115 km.东灵山位于小五台山东部余脉, 属太行山系.山峰海拔高度多高于1 000 m, 最高峰为2 303 m (冯云等, 2007).该地区属于暖温带半湿润大陆性季风气候, 春季干旱, 夏季暖热多雨, 冬季寒冷干燥(全权等, 2015).年平均气温5-10 ℃, 无霜期约195天, 年降水量500-650 mm, 且多集中于夏季, 夏季降水约占全年降水量的75% (冯云等, 2007; 全权等, 2015).该地处于华北地区中部, 地质构造上位于燕山沉降带西部, 地貌以山地侵蚀结构为主, 地势险峻, 谷深坡陡, 河流下切严重, 土壤主要为肥沃的褐色土和棕色的森林土(孙世洲, 1997).植被类型较丰富, 以辽东栎为主要树种, 五角枫、白蜡树(Fraxinus chinensis)、黄荆(Vitex negundo)、山杏(Armeniaca sibirica)和绣线菊(Spiraea salicifolia)为伴生树种构成典型的辽东栎林(刘贤赵等, 2009; 张建华等, 2017).根据北京城区和河北多个观测点近10年的干湿氮沉降数据, 北京南部和东部大气氮沉降速率可以高达80 kg·hm-2·a-1, 北京西部地区略低, 但高峰月份也可以达到50 kg·hm-2·a-1 (王登芝等, 2006; 李金香和李峰, 2012; Pan et al., 2012; Luo et al., 2013; Xu et al., 2015; 杨文琴等, 2016; 尹兴等, 2017). ...
Huge carbon sequestration potential in global forests 1 2012
... 温带北方森林常年受到氮限制, 土壤氮增加会促进木本植物生长, 增加森林的固碳潜力(Liu et al., 2012).氮是植物合成叶片光合作用羧化酶——核酮糖-1,5-双磷酸羧化酶/加氧酶(Rubisco)和光合作用重要色素叶绿素的重要组分, 叶片氮含量的变化会直接引起光合作用酶含量和叶绿素含量的改变, 所以施氮会直接提高植物光合作用能力(Hikosaka & Shigeno, 2009; Palmroth et al., 2014).此外, 有关氮沉降影响净初级生产力的经验模型也指出增加的大气氮沉降量会导致木本植物的碳固持增加, 提高森林的初级生产力(Kurts et al., 2008; Lu et al., 2014). ...
Nitrogen deposition contributes to soil acidification in tropical ecosystems 1 2014
... 温带北方森林常年受到氮限制, 土壤氮增加会促进木本植物生长, 增加森林的固碳潜力(Liu et al., 2012).氮是植物合成叶片光合作用羧化酶——核酮糖-1,5-双磷酸羧化酶/加氧酶(Rubisco)和光合作用重要色素叶绿素的重要组分, 叶片氮含量的变化会直接引起光合作用酶含量和叶绿素含量的改变, 所以施氮会直接提高植物光合作用能力(Hikosaka & Shigeno, 2009; Palmroth et al., 2014).此外, 有关氮沉降影响净初级生产力的经验模型也指出增加的大气氮沉降量会导致木本植物的碳固持增加, 提高森林的初级生产力(Kurts et al., 2008; Lu et al., 2014). ...
Effects of experimental nitrogen additions on plant diversity in tropical forests of contrasting disturbance regimes in southern China 1 2011
... 氮沉降对温带森林影响研究从20世纪中叶开始, 已超过30年(Andersson et al., 1980; Garcia et al., 2008; Nave et al., 2015; Ma et al., 2018), 已经取得了不少成果, 但对于目前大气氮沉降如何长期影响树木生长和植被动态变化的认知程度依然十分有限(Bedison & Mcneil, 2009).近20年来国内陆续开展了关于氮沉降量增加对森林生态系统影响的研究(Mo et al., 2003; 方运霆等, 2004; 莫江明等, 2004;方华和莫江明, 2006; Lu et al., 2010; Lu et al., 2011; Zhu et al., 2013).由于在野外自然条件下进行控制实验的复杂性和高难度, 已有的氮沉降控制实验的研究对象大多以幼苗为主, 以自然森林成年大树为实验对象的研究较少(方运霆等, 2005), 基于幼苗或幼树对氮沉降的响应结果推测自然成熟森林的情况, 其代表性存在很大局限性. ...
Effects of experimental nitrogen additions on plant diversity in an old-growth tropical forest 1 2010
... 氮沉降对温带森林影响研究从20世纪中叶开始, 已超过30年(Andersson et al., 1980; Garcia et al., 2008; Nave et al., 2015; Ma et al., 2018), 已经取得了不少成果, 但对于目前大气氮沉降如何长期影响树木生长和植被动态变化的认知程度依然十分有限(Bedison & Mcneil, 2009).近20年来国内陆续开展了关于氮沉降量增加对森林生态系统影响的研究(Mo et al., 2003; 方运霆等, 2004; 莫江明等, 2004;方华和莫江明, 2006; Lu et al., 2010; Lu et al., 2011; Zhu et al., 2013).由于在野外自然条件下进行控制实验的复杂性和高难度, 已有的氮沉降控制实验的研究对象大多以幼苗为主, 以自然森林成年大树为实验对象的研究较少(方运霆等, 2005), 基于幼苗或幼树对氮沉降的响应结果推测自然成熟森林的情况, 其代表性存在很大局限性. ...
An evaluation of atmospheric Nr pollution and deposition in North China after the Beijing Olympics 1 2013
... 北京东灵山(39.92°-40.03° N, 115.48°-115.98° E)位于北京市西郊的门头沟区西部, 毗邻河北省, 距市区约115 km.东灵山位于小五台山东部余脉, 属太行山系.山峰海拔高度多高于1 000 m, 最高峰为2 303 m (冯云等, 2007).该地区属于暖温带半湿润大陆性季风气候, 春季干旱, 夏季暖热多雨, 冬季寒冷干燥(全权等, 2015).年平均气温5-10 ℃, 无霜期约195天, 年降水量500-650 mm, 且多集中于夏季, 夏季降水约占全年降水量的75% (冯云等, 2007; 全权等, 2015).该地处于华北地区中部, 地质构造上位于燕山沉降带西部, 地貌以山地侵蚀结构为主, 地势险峻, 谷深坡陡, 河流下切严重, 土壤主要为肥沃的褐色土和棕色的森林土(孙世洲, 1997).植被类型较丰富, 以辽东栎为主要树种, 五角枫、白蜡树(Fraxinus chinensis)、黄荆(Vitex negundo)、山杏(Armeniaca sibirica)和绣线菊(Spiraea salicifolia)为伴生树种构成典型的辽东栎林(刘贤赵等, 2009; 张建华等, 2017).根据北京城区和河北多个观测点近10年的干湿氮沉降数据, 北京南部和东部大气氮沉降速率可以高达80 kg·hm-2·a-1, 北京西部地区略低, 但高峰月份也可以达到50 kg·hm-2·a-1 (王登芝等, 2006; 李金香和李峰, 2012; Pan et al., 2012; Luo et al., 2013; Xu et al., 2015; 杨文琴等, 2016; 尹兴等, 2017). ...
Plant and soil microbe responses to light, warming and nitrogen addition in a temperate forest 1 2018
... 氮沉降对温带森林影响研究从20世纪中叶开始, 已超过30年(Andersson et al., 1980; Garcia et al., 2008; Nave et al., 2015; Ma et al., 2018), 已经取得了不少成果, 但对于目前大气氮沉降如何长期影响树木生长和植被动态变化的认知程度依然十分有限(Bedison & Mcneil, 2009).近20年来国内陆续开展了关于氮沉降量增加对森林生态系统影响的研究(Mo et al., 2003; 方运霆等, 2004; 莫江明等, 2004;方华和莫江明, 2006; Lu et al., 2010; Lu et al., 2011; Zhu et al., 2013).由于在野外自然条件下进行控制实验的复杂性和高难度, 已有的氮沉降控制实验的研究对象大多以幼苗为主, 以自然森林成年大树为实验对象的研究较少(方运霆等, 2005), 基于幼苗或幼树对氮沉降的响应结果推测自然成熟森林的情况, 其代表性存在很大局限性. ...
Functional relationships between leaf hydraulics and leaf economic traits in response to nutrient addition in subtropical tree species 1 2013
Sun-shade patterns of leaf carotenoid composition in 86 species of neotropical forest plants 1 2009
... 光合作用是绿色植物转化太阳能, 合成同化物质的重要生理过程, 是植物维持生长发育的基础.氮添加的浓度和时间会对植物的叶绿素合成产生不同的影响.本研究中氮添加处理提高了辽东栎和五角枫植物的叶片氮含量, 促进了叶片的叶绿素生成, 与前人研究结果(Bauer et al., 2010; Pérez-Soba et al.,2010; Jia et al., 2011; Wang et al., 2012; 郑翔等, 2018)一致.需要说明的是, 两树种间的净光合速率和叶绿素含量均存在显著差异, 辽东栎的光合速率显著高于五角枫, 而五角枫的叶绿素含量显著高于辽东栎.此外, 氮添加对叶片叶绿素含量的促进作用随着处理时间延长表现出明显的累积效应, 五角枫比辽东栎更为明显.氮添加对光合速率、气孔导度和蒸腾速率的促进作用也随着处理时间延长表现出明显的累积效应, 辽东栎比五角枫更明显.在本研究的实验样地中, 辽东栎树体高大, 占据上层空间, 树冠上中层可以接收到充足的太阳辐射, 是群落的建群种和优势种; 而样地内的五角枫树体矮小, 处于被遮阴的下层, 可接受光照少.不同的光照环境会导致树木同化组织在结构和功能方面的极大差异, 一般阳生植物比阴生植物具有更高的羧化酶活性, 而阴生植物为了尽可能地获得林下的散射光, 会提高叶绿素含量, 形成发达的捕获色素系统, 以截获更多的光能(Shen & Ye, 1991; Johnson et al., 1993; 张守仁等, 2004; Matsubara et al., 2009; 孙小玲等, 2010; 向芬等, 2018).叶片光合作用气体交换过程主要受气孔因素和非气孔因素(主要是Rubisco羧化酶活性)影响, 在饱和光下辽东栎的羧化速率(Vcmax)和气孔导度远高于五角枫(未发表数据), 这也解释了在饱和光下辽东栎光合速率高于五角枫, 而五角枫叶绿素含量高于辽东栎的原因. ...
Nitrogen availability in disturbed, rehabilitated and mature forests of tropical China 1 2003
... 氮沉降对温带森林影响研究从20世纪中叶开始, 已超过30年(Andersson et al., 1980; Garcia et al., 2008; Nave et al., 2015; Ma et al., 2018), 已经取得了不少成果, 但对于目前大气氮沉降如何长期影响树木生长和植被动态变化的认知程度依然十分有限(Bedison & Mcneil, 2009).近20年来国内陆续开展了关于氮沉降量增加对森林生态系统影响的研究(Mo et al., 2003; 方运霆等, 2004; 莫江明等, 2004;方华和莫江明, 2006; Lu et al., 2010; Lu et al., 2011; Zhu et al., 2013).由于在野外自然条件下进行控制实验的复杂性和高难度, 已有的氮沉降控制实验的研究对象大多以幼苗为主, 以自然森林成年大树为实验对象的研究较少(方运霆等, 2005), 基于幼苗或幼树对氮沉降的响应结果推测自然成熟森林的情况, 其代表性存在很大局限性. ...
鼎湖山主要森林土壤N2O排放及其对模拟N沉降的响应 1 2006
... 从工业革命爆发以来, 化石燃料大量燃烧, 土地利用发生大规模改变, 农业、畜牧业快速发展.活跃的工农业生产活动使全球大气中的氮化物浓度不断增加(Vitousek et al., 1997).在全球范围内, 由人类活动导致的氮沉降增加主要集中于北美、欧洲、东南亚三大地区(Galloway et al., 2003), 中国经济发达的东部地区处于全球三大高氮沉降区之一的东南亚地区.随着大规模的工业化生产和农业施肥活动, 进入陆地生态系统的氮不断增加, 经济发达的温带地区最为明显(莫江明等, 2006).根据Galloway等(2002)的预测, 未来数十年内, 随着经济的进一步发展, 这种状况会不断加剧.氮富集及氮驱动力的预测模型表明, 到21世纪中叶, 全球活性氮的排放速度将达到267 Tg·a-1, 亚洲地区由于人为活动产生的活性氮增长速度可能高达105.3 Tg·a-1 (Holland et al., 1999; Zheng et al., 2002; Galloway et al., 2004, 2008).近年来京津冀地区高浓度的氮氧化合物和其他可溶性气溶胶形成的雾霾引起人们的广泛关注, 该地区属全球典型的高氮沉降区(Zhang et al., 2012).其氮湿沉降速率平均值为38 kg·hm-2·a-1, 无机氮沉降速率约为27 kg·hm-2·a-1, 有机氮沉降速率约为7 kg·hm-2·a-1, 氮的干沉降速率也高达50 kg·hm-2·a-1.Shen等(2009)研究表明, 未来北京地区总的大气氮沉降速率可超过100 kg·hm-2·a-1. ...
鼎湖山主要森林土壤N2O排放及其对模拟N沉降的响应 1 2006
... 从工业革命爆发以来, 化石燃料大量燃烧, 土地利用发生大规模改变, 农业、畜牧业快速发展.活跃的工农业生产活动使全球大气中的氮化物浓度不断增加(Vitousek et al., 1997).在全球范围内, 由人类活动导致的氮沉降增加主要集中于北美、欧洲、东南亚三大地区(Galloway et al., 2003), 中国经济发达的东部地区处于全球三大高氮沉降区之一的东南亚地区.随着大规模的工业化生产和农业施肥活动, 进入陆地生态系统的氮不断增加, 经济发达的温带地区最为明显(莫江明等, 2006).根据Galloway等(2002)的预测, 未来数十年内, 随着经济的进一步发展, 这种状况会不断加剧.氮富集及氮驱动力的预测模型表明, 到21世纪中叶, 全球活性氮的排放速度将达到267 Tg·a-1, 亚洲地区由于人为活动产生的活性氮增长速度可能高达105.3 Tg·a-1 (Holland et al., 1999; Zheng et al., 2002; Galloway et al., 2004, 2008).近年来京津冀地区高浓度的氮氧化合物和其他可溶性气溶胶形成的雾霾引起人们的广泛关注, 该地区属全球典型的高氮沉降区(Zhang et al., 2012).其氮湿沉降速率平均值为38 kg·hm-2·a-1, 无机氮沉降速率约为27 kg·hm-2·a-1, 有机氮沉降速率约为7 kg·hm-2·a-1, 氮的干沉降速率也高达50 kg·hm-2·a-1.Shen等(2009)研究表明, 未来北京地区总的大气氮沉降速率可超过100 kg·hm-2·a-1. ...
鼎湖山主要森林植物凋落物分解及其对N沉降的响应 1 2004
... 氮沉降对温带森林影响研究从20世纪中叶开始, 已超过30年(Andersson et al., 1980; Garcia et al., 2008; Nave et al., 2015; Ma et al., 2018), 已经取得了不少成果, 但对于目前大气氮沉降如何长期影响树木生长和植被动态变化的认知程度依然十分有限(Bedison & Mcneil, 2009).近20年来国内陆续开展了关于氮沉降量增加对森林生态系统影响的研究(Mo et al., 2003; 方运霆等, 2004; 莫江明等, 2004;方华和莫江明, 2006; Lu et al., 2010; Lu et al., 2011; Zhu et al., 2013).由于在野外自然条件下进行控制实验的复杂性和高难度, 已有的氮沉降控制实验的研究对象大多以幼苗为主, 以自然森林成年大树为实验对象的研究较少(方运霆等, 2005), 基于幼苗或幼树对氮沉降的响应结果推测自然成熟森林的情况, 其代表性存在很大局限性. ...
鼎湖山主要森林植物凋落物分解及其对N沉降的响应 1 2004
... 氮沉降对温带森林影响研究从20世纪中叶开始, 已超过30年(Andersson et al., 1980; Garcia et al., 2008; Nave et al., 2015; Ma et al., 2018), 已经取得了不少成果, 但对于目前大气氮沉降如何长期影响树木生长和植被动态变化的认知程度依然十分有限(Bedison & Mcneil, 2009).近20年来国内陆续开展了关于氮沉降量增加对森林生态系统影响的研究(Mo et al., 2003; 方运霆等, 2004; 莫江明等, 2004;方华和莫江明, 2006; Lu et al., 2010; Lu et al., 2011; Zhu et al., 2013).由于在野外自然条件下进行控制实验的复杂性和高难度, 已有的氮沉降控制实验的研究对象大多以幼苗为主, 以自然森林成年大树为实验对象的研究较少(方运霆等, 2005), 基于幼苗或幼树对氮沉降的响应结果推测自然成熟森林的情况, 其代表性存在很大局限性. ...
Disturbance and the resilience of coupled carbon and nitrogen cycling in a north temperate forest 1 2015
... 氮沉降对温带森林影响研究从20世纪中叶开始, 已超过30年(Andersson et al., 1980; Garcia et al., 2008; Nave et al., 2015; Ma et al., 2018), 已经取得了不少成果, 但对于目前大气氮沉降如何长期影响树木生长和植被动态变化的认知程度依然十分有限(Bedison & Mcneil, 2009).近20年来国内陆续开展了关于氮沉降量增加对森林生态系统影响的研究(Mo et al., 2003; 方运霆等, 2004; 莫江明等, 2004;方华和莫江明, 2006; Lu et al., 2010; Lu et al., 2011; Zhu et al., 2013).由于在野外自然条件下进行控制实验的复杂性和高难度, 已有的氮沉降控制实验的研究对象大多以幼苗为主, 以自然森林成年大树为实验对象的研究较少(方运霆等, 2005), 基于幼苗或幼树对氮沉降的响应结果推测自然成熟森林的情况, 其代表性存在很大局限性. ...
Nitrogen-?addition effects on leaf traits and photosynthetic carbon gain of boreal forest understory shrubs 1 2014
... 温带北方森林常年受到氮限制, 土壤氮增加会促进木本植物生长, 增加森林的固碳潜力(Liu et al., 2012).氮是植物合成叶片光合作用羧化酶——核酮糖-1,5-双磷酸羧化酶/加氧酶(Rubisco)和光合作用重要色素叶绿素的重要组分, 叶片氮含量的变化会直接引起光合作用酶含量和叶绿素含量的改变, 所以施氮会直接提高植物光合作用能力(Hikosaka & Shigeno, 2009; Palmroth et al., 2014).此外, 有关氮沉降影响净初级生产力的经验模型也指出增加的大气氮沉降量会导致木本植物的碳固持增加, 提高森林的初级生产力(Kurts et al., 2008; Lu et al., 2014). ...
Spatial distribution and seasonal variations of atmospheric sulfur deposition over northern China 1 2012
... 北京东灵山(39.92°-40.03° N, 115.48°-115.98° E)位于北京市西郊的门头沟区西部, 毗邻河北省, 距市区约115 km.东灵山位于小五台山东部余脉, 属太行山系.山峰海拔高度多高于1 000 m, 最高峰为2 303 m (冯云等, 2007).该地区属于暖温带半湿润大陆性季风气候, 春季干旱, 夏季暖热多雨, 冬季寒冷干燥(全权等, 2015).年平均气温5-10 ℃, 无霜期约195天, 年降水量500-650 mm, 且多集中于夏季, 夏季降水约占全年降水量的75% (冯云等, 2007; 全权等, 2015).该地处于华北地区中部, 地质构造上位于燕山沉降带西部, 地貌以山地侵蚀结构为主, 地势险峻, 谷深坡陡, 河流下切严重, 土壤主要为肥沃的褐色土和棕色的森林土(孙世洲, 1997).植被类型较丰富, 以辽东栎为主要树种, 五角枫、白蜡树(Fraxinus chinensis)、黄荆(Vitex negundo)、山杏(Armeniaca sibirica)和绣线菊(Spiraea salicifolia)为伴生树种构成典型的辽东栎林(刘贤赵等, 2009; 张建华等, 2017).根据北京城区和河北多个观测点近10年的干湿氮沉降数据, 北京南部和东部大气氮沉降速率可以高达80 kg·hm-2·a-1, 北京西部地区略低, 但高峰月份也可以达到50 kg·hm-2·a-1 (王登芝等, 2006; 李金香和李峰, 2012; Pan et al., 2012; Luo et al., 2013; Xu et al., 2015; 杨文琴等, 2016; 尹兴等, 2017). ...
... 北京东灵山(39.92°-40.03° N, 115.48°-115.98° E)位于北京市西郊的门头沟区西部, 毗邻河北省, 距市区约115 km.东灵山位于小五台山东部余脉, 属太行山系.山峰海拔高度多高于1 000 m, 最高峰为2 303 m (冯云等, 2007).该地区属于暖温带半湿润大陆性季风气候, 春季干旱, 夏季暖热多雨, 冬季寒冷干燥(全权等, 2015).年平均气温5-10 ℃, 无霜期约195天, 年降水量500-650 mm, 且多集中于夏季, 夏季降水约占全年降水量的75% (冯云等, 2007; 全权等, 2015).该地处于华北地区中部, 地质构造上位于燕山沉降带西部, 地貌以山地侵蚀结构为主, 地势险峻, 谷深坡陡, 河流下切严重, 土壤主要为肥沃的褐色土和棕色的森林土(孙世洲, 1997).植被类型较丰富, 以辽东栎为主要树种, 五角枫、白蜡树(Fraxinus chinensis)、黄荆(Vitex negundo)、山杏(Armeniaca sibirica)和绣线菊(Spiraea salicifolia)为伴生树种构成典型的辽东栎林(刘贤赵等, 2009; 张建华等, 2017).根据北京城区和河北多个观测点近10年的干湿氮沉降数据, 北京南部和东部大气氮沉降速率可以高达80 kg·hm-2·a-1, 北京西部地区略低, 但高峰月份也可以达到50 kg·hm-2·a-1 (王登芝等, 2006; 李金香和李峰, 2012; Pan et al., 2012; Luo et al., 2013; Xu et al., 2015; 杨文琴等, 2016; 尹兴等, 2017). ...
东灵山地区的地质、地貌和土壤 1 1997
... 北京东灵山(39.92°-40.03° N, 115.48°-115.98° E)位于北京市西郊的门头沟区西部, 毗邻河北省, 距市区约115 km.东灵山位于小五台山东部余脉, 属太行山系.山峰海拔高度多高于1 000 m, 最高峰为2 303 m (冯云等, 2007).该地区属于暖温带半湿润大陆性季风气候, 春季干旱, 夏季暖热多雨, 冬季寒冷干燥(全权等, 2015).年平均气温5-10 ℃, 无霜期约195天, 年降水量500-650 mm, 且多集中于夏季, 夏季降水约占全年降水量的75% (冯云等, 2007; 全权等, 2015).该地处于华北地区中部, 地质构造上位于燕山沉降带西部, 地貌以山地侵蚀结构为主, 地势险峻, 谷深坡陡, 河流下切严重, 土壤主要为肥沃的褐色土和棕色的森林土(孙世洲, 1997).植被类型较丰富, 以辽东栎为主要树种, 五角枫、白蜡树(Fraxinus chinensis)、黄荆(Vitex negundo)、山杏(Armeniaca sibirica)和绣线菊(Spiraea salicifolia)为伴生树种构成典型的辽东栎林(刘贤赵等, 2009; 张建华等, 2017).根据北京城区和河北多个观测点近10年的干湿氮沉降数据, 北京南部和东部大气氮沉降速率可以高达80 kg·hm-2·a-1, 北京西部地区略低, 但高峰月份也可以达到50 kg·hm-2·a-1 (王登芝等, 2006; 李金香和李峰, 2012; Pan et al., 2012; Luo et al., 2013; Xu et al., 2015; 杨文琴等, 2016; 尹兴等, 2017). ...
植株叶片的光合色素构成对遮阴的响应 1 2010
... 光合作用是绿色植物转化太阳能, 合成同化物质的重要生理过程, 是植物维持生长发育的基础.氮添加的浓度和时间会对植物的叶绿素合成产生不同的影响.本研究中氮添加处理提高了辽东栎和五角枫植物的叶片氮含量, 促进了叶片的叶绿素生成, 与前人研究结果(Bauer et al., 2010; Pérez-Soba et al.,2010; Jia et al., 2011; Wang et al., 2012; 郑翔等, 2018)一致.需要说明的是, 两树种间的净光合速率和叶绿素含量均存在显著差异, 辽东栎的光合速率显著高于五角枫, 而五角枫的叶绿素含量显著高于辽东栎.此外, 氮添加对叶片叶绿素含量的促进作用随着处理时间延长表现出明显的累积效应, 五角枫比辽东栎更为明显.氮添加对光合速率、气孔导度和蒸腾速率的促进作用也随着处理时间延长表现出明显的累积效应, 辽东栎比五角枫更明显.在本研究的实验样地中, 辽东栎树体高大, 占据上层空间, 树冠上中层可以接收到充足的太阳辐射, 是群落的建群种和优势种; 而样地内的五角枫树体矮小, 处于被遮阴的下层, 可接受光照少.不同的光照环境会导致树木同化组织在结构和功能方面的极大差异, 一般阳生植物比阴生植物具有更高的羧化酶活性, 而阴生植物为了尽可能地获得林下的散射光, 会提高叶绿素含量, 形成发达的捕获色素系统, 以截获更多的光能(Shen & Ye, 1991; Johnson et al., 1993; 张守仁等, 2004; Matsubara et al., 2009; 孙小玲等, 2010; 向芬等, 2018).叶片光合作用气体交换过程主要受气孔因素和非气孔因素(主要是Rubisco羧化酶活性)影响, 在饱和光下辽东栎的羧化速率(Vcmax)和气孔导度远高于五角枫(未发表数据), 这也解释了在饱和光下辽东栎光合速率高于五角枫, 而五角枫叶绿素含量高于辽东栎的原因. ...
植株叶片的光合色素构成对遮阴的响应 1 2010
... 光合作用是绿色植物转化太阳能, 合成同化物质的重要生理过程, 是植物维持生长发育的基础.氮添加的浓度和时间会对植物的叶绿素合成产生不同的影响.本研究中氮添加处理提高了辽东栎和五角枫植物的叶片氮含量, 促进了叶片的叶绿素生成, 与前人研究结果(Bauer et al., 2010; Pérez-Soba et al.,2010; Jia et al., 2011; Wang et al., 2012; 郑翔等, 2018)一致.需要说明的是, 两树种间的净光合速率和叶绿素含量均存在显著差异, 辽东栎的光合速率显著高于五角枫, 而五角枫的叶绿素含量显著高于辽东栎.此外, 氮添加对叶片叶绿素含量的促进作用随着处理时间延长表现出明显的累积效应, 五角枫比辽东栎更为明显.氮添加对光合速率、气孔导度和蒸腾速率的促进作用也随着处理时间延长表现出明显的累积效应, 辽东栎比五角枫更明显.在本研究的实验样地中, 辽东栎树体高大, 占据上层空间, 树冠上中层可以接收到充足的太阳辐射, 是群落的建群种和优势种; 而样地内的五角枫树体矮小, 处于被遮阴的下层, 可接受光照少.不同的光照环境会导致树木同化组织在结构和功能方面的极大差异, 一般阳生植物比阴生植物具有更高的羧化酶活性, 而阴生植物为了尽可能地获得林下的散射光, 会提高叶绿素含量, 形成发达的捕获色素系统, 以截获更多的光能(Shen & Ye, 1991; Johnson et al., 1993; 张守仁等, 2004; Matsubara et al., 2009; 孙小玲等, 2010; 向芬等, 2018).叶片光合作用气体交换过程主要受气孔因素和非气孔因素(主要是Rubisco羧化酶活性)影响, 在饱和光下辽东栎的羧化速率(Vcmax)和气孔导度远高于五角枫(未发表数据), 这也解释了在饱和光下辽东栎光合速率高于五角枫, 而五角枫叶绿素含量高于辽东栎的原因. ...
Human alteration of the global nitrogen cycle: Sources and consequences 1 1997
... 从工业革命爆发以来, 化石燃料大量燃烧, 土地利用发生大规模改变, 农业、畜牧业快速发展.活跃的工农业生产活动使全球大气中的氮化物浓度不断增加(Vitousek et al., 1997).在全球范围内, 由人类活动导致的氮沉降增加主要集中于北美、欧洲、东南亚三大地区(Galloway et al., 2003), 中国经济发达的东部地区处于全球三大高氮沉降区之一的东南亚地区.随着大规模的工业化生产和农业施肥活动, 进入陆地生态系统的氮不断增加, 经济发达的温带地区最为明显(莫江明等, 2006).根据Galloway等(2002)的预测, 未来数十年内, 随着经济的进一步发展, 这种状况会不断加剧.氮富集及氮驱动力的预测模型表明, 到21世纪中叶, 全球活性氮的排放速度将达到267 Tg·a-1, 亚洲地区由于人为活动产生的活性氮增长速度可能高达105.3 Tg·a-1 (Holland et al., 1999; Zheng et al., 2002; Galloway et al., 2004, 2008).近年来京津冀地区高浓度的氮氧化合物和其他可溶性气溶胶形成的雾霾引起人们的广泛关注, 该地区属全球典型的高氮沉降区(Zhang et al., 2012).其氮湿沉降速率平均值为38 kg·hm-2·a-1, 无机氮沉降速率约为27 kg·hm-2·a-1, 有机氮沉降速率约为7 kg·hm-2·a-1, 氮的干沉降速率也高达50 kg·hm-2·a-1.Shen等(2009)研究表明, 未来北京地区总的大气氮沉降速率可超过100 kg·hm-2·a-1. ...
北京西山地区油松林水文过程中营养元素迁移特征 1 2006
... 北京东灵山(39.92°-40.03° N, 115.48°-115.98° E)位于北京市西郊的门头沟区西部, 毗邻河北省, 距市区约115 km.东灵山位于小五台山东部余脉, 属太行山系.山峰海拔高度多高于1 000 m, 最高峰为2 303 m (冯云等, 2007).该地区属于暖温带半湿润大陆性季风气候, 春季干旱, 夏季暖热多雨, 冬季寒冷干燥(全权等, 2015).年平均气温5-10 ℃, 无霜期约195天, 年降水量500-650 mm, 且多集中于夏季, 夏季降水约占全年降水量的75% (冯云等, 2007; 全权等, 2015).该地处于华北地区中部, 地质构造上位于燕山沉降带西部, 地貌以山地侵蚀结构为主, 地势险峻, 谷深坡陡, 河流下切严重, 土壤主要为肥沃的褐色土和棕色的森林土(孙世洲, 1997).植被类型较丰富, 以辽东栎为主要树种, 五角枫、白蜡树(Fraxinus chinensis)、黄荆(Vitex negundo)、山杏(Armeniaca sibirica)和绣线菊(Spiraea salicifolia)为伴生树种构成典型的辽东栎林(刘贤赵等, 2009; 张建华等, 2017).根据北京城区和河北多个观测点近10年的干湿氮沉降数据, 北京南部和东部大气氮沉降速率可以高达80 kg·hm-2·a-1, 北京西部地区略低, 但高峰月份也可以达到50 kg·hm-2·a-1 (王登芝等, 2006; 李金香和李峰, 2012; Pan et al., 2012; Luo et al., 2013; Xu et al., 2015; 杨文琴等, 2016; 尹兴等, 2017). ...
北京西山地区油松林水文过程中营养元素迁移特征 1 2006
... 北京东灵山(39.92°-40.03° N, 115.48°-115.98° E)位于北京市西郊的门头沟区西部, 毗邻河北省, 距市区约115 km.东灵山位于小五台山东部余脉, 属太行山系.山峰海拔高度多高于1 000 m, 最高峰为2 303 m (冯云等, 2007).该地区属于暖温带半湿润大陆性季风气候, 春季干旱, 夏季暖热多雨, 冬季寒冷干燥(全权等, 2015).年平均气温5-10 ℃, 无霜期约195天, 年降水量500-650 mm, 且多集中于夏季, 夏季降水约占全年降水量的75% (冯云等, 2007; 全权等, 2015).该地处于华北地区中部, 地质构造上位于燕山沉降带西部, 地貌以山地侵蚀结构为主, 地势险峻, 谷深坡陡, 河流下切严重, 土壤主要为肥沃的褐色土和棕色的森林土(孙世洲, 1997).植被类型较丰富, 以辽东栎为主要树种, 五角枫、白蜡树(Fraxinus chinensis)、黄荆(Vitex negundo)、山杏(Armeniaca sibirica)和绣线菊(Spiraea salicifolia)为伴生树种构成典型的辽东栎林(刘贤赵等, 2009; 张建华等, 2017).根据北京城区和河北多个观测点近10年的干湿氮沉降数据, 北京南部和东部大气氮沉降速率可以高达80 kg·hm-2·a-1, 北京西部地区略低, 但高峰月份也可以达到50 kg·hm-2·a-1 (王登芝等, 2006; 李金香和李峰, 2012; Pan et al., 2012; Luo et al., 2013; Xu et al., 2015; 杨文琴等, 2016; 尹兴等, 2017). ...
Effects of soil water and nitrogen on growth and photosynthetic response of Manchurian ash (Fraxinus mandshurica) seedlings in northeastern China 1 2012
... 光合作用是绿色植物转化太阳能, 合成同化物质的重要生理过程, 是植物维持生长发育的基础.氮添加的浓度和时间会对植物的叶绿素合成产生不同的影响.本研究中氮添加处理提高了辽东栎和五角枫植物的叶片氮含量, 促进了叶片的叶绿素生成, 与前人研究结果(Bauer et al., 2010; Pérez-Soba et al.,2010; Jia et al., 2011; Wang et al., 2012; 郑翔等, 2018)一致.需要说明的是, 两树种间的净光合速率和叶绿素含量均存在显著差异, 辽东栎的光合速率显著高于五角枫, 而五角枫的叶绿素含量显著高于辽东栎.此外, 氮添加对叶片叶绿素含量的促进作用随着处理时间延长表现出明显的累积效应, 五角枫比辽东栎更为明显.氮添加对光合速率、气孔导度和蒸腾速率的促进作用也随着处理时间延长表现出明显的累积效应, 辽东栎比五角枫更明显.在本研究的实验样地中, 辽东栎树体高大, 占据上层空间, 树冠上中层可以接收到充足的太阳辐射, 是群落的建群种和优势种; 而样地内的五角枫树体矮小, 处于被遮阴的下层, 可接受光照少.不同的光照环境会导致树木同化组织在结构和功能方面的极大差异, 一般阳生植物比阴生植物具有更高的羧化酶活性, 而阴生植物为了尽可能地获得林下的散射光, 会提高叶绿素含量, 形成发达的捕获色素系统, 以截获更多的光能(Shen & Ye, 1991; Johnson et al., 1993; 张守仁等, 2004; Matsubara et al., 2009; 孙小玲等, 2010; 向芬等, 2018).叶片光合作用气体交换过程主要受气孔因素和非气孔因素(主要是Rubisco羧化酶活性)影响, 在饱和光下辽东栎的羧化速率(Vcmax)和气孔导度远高于五角枫(未发表数据), 这也解释了在饱和光下辽东栎光合速率高于五角枫, 而五角枫叶绿素含量高于辽东栎的原因. ...
... 北京东灵山(39.92°-40.03° N, 115.48°-115.98° E)位于北京市西郊的门头沟区西部, 毗邻河北省, 距市区约115 km.东灵山位于小五台山东部余脉, 属太行山系.山峰海拔高度多高于1 000 m, 最高峰为2 303 m (冯云等, 2007).该地区属于暖温带半湿润大陆性季风气候, 春季干旱, 夏季暖热多雨, 冬季寒冷干燥(全权等, 2015).年平均气温5-10 ℃, 无霜期约195天, 年降水量500-650 mm, 且多集中于夏季, 夏季降水约占全年降水量的75% (冯云等, 2007; 全权等, 2015).该地处于华北地区中部, 地质构造上位于燕山沉降带西部, 地貌以山地侵蚀结构为主, 地势险峻, 谷深坡陡, 河流下切严重, 土壤主要为肥沃的褐色土和棕色的森林土(孙世洲, 1997).植被类型较丰富, 以辽东栎为主要树种, 五角枫、白蜡树(Fraxinus chinensis)、黄荆(Vitex negundo)、山杏(Armeniaca sibirica)和绣线菊(Spiraea salicifolia)为伴生树种构成典型的辽东栎林(刘贤赵等, 2009; 张建华等, 2017).根据北京城区和河北多个观测点近10年的干湿氮沉降数据, 北京南部和东部大气氮沉降速率可以高达80 kg·hm-2·a-1, 北京西部地区略低, 但高峰月份也可以达到50 kg·hm-2·a-1 (王登芝等, 2006; 李金香和李峰, 2012; Pan et al., 2012; Luo et al., 2013; Xu et al., 2015; 杨文琴等, 2016; 尹兴等, 2017). ...
北京东灵山地区常见灌丛生长及凋落物生产对氮添加的响应 1 2017
... 北京东灵山(39.92°-40.03° N, 115.48°-115.98° E)位于北京市西郊的门头沟区西部, 毗邻河北省, 距市区约115 km.东灵山位于小五台山东部余脉, 属太行山系.山峰海拔高度多高于1 000 m, 最高峰为2 303 m (冯云等, 2007).该地区属于暖温带半湿润大陆性季风气候, 春季干旱, 夏季暖热多雨, 冬季寒冷干燥(全权等, 2015).年平均气温5-10 ℃, 无霜期约195天, 年降水量500-650 mm, 且多集中于夏季, 夏季降水约占全年降水量的75% (冯云等, 2007; 全权等, 2015).该地处于华北地区中部, 地质构造上位于燕山沉降带西部, 地貌以山地侵蚀结构为主, 地势险峻, 谷深坡陡, 河流下切严重, 土壤主要为肥沃的褐色土和棕色的森林土(孙世洲, 1997).植被类型较丰富, 以辽东栎为主要树种, 五角枫、白蜡树(Fraxinus chinensis)、黄荆(Vitex negundo)、山杏(Armeniaca sibirica)和绣线菊(Spiraea salicifolia)为伴生树种构成典型的辽东栎林(刘贤赵等, 2009; 张建华等, 2017).根据北京城区和河北多个观测点近10年的干湿氮沉降数据, 北京南部和东部大气氮沉降速率可以高达80 kg·hm-2·a-1, 北京西部地区略低, 但高峰月份也可以达到50 kg·hm-2·a-1 (王登芝等, 2006; 李金香和李峰, 2012; Pan et al., 2012; Luo et al., 2013; Xu et al., 2015; 杨文琴等, 2016; 尹兴等, 2017). ...
北京东灵山地区常见灌丛生长及凋落物生产对氮添加的响应 1 2017
... 北京东灵山(39.92°-40.03° N, 115.48°-115.98° E)位于北京市西郊的门头沟区西部, 毗邻河北省, 距市区约115 km.东灵山位于小五台山东部余脉, 属太行山系.山峰海拔高度多高于1 000 m, 最高峰为2 303 m (冯云等, 2007).该地区属于暖温带半湿润大陆性季风气候, 春季干旱, 夏季暖热多雨, 冬季寒冷干燥(全权等, 2015).年平均气温5-10 ℃, 无霜期约195天, 年降水量500-650 mm, 且多集中于夏季, 夏季降水约占全年降水量的75% (冯云等, 2007; 全权等, 2015).该地处于华北地区中部, 地质构造上位于燕山沉降带西部, 地貌以山地侵蚀结构为主, 地势险峻, 谷深坡陡, 河流下切严重, 土壤主要为肥沃的褐色土和棕色的森林土(孙世洲, 1997).植被类型较丰富, 以辽东栎为主要树种, 五角枫、白蜡树(Fraxinus chinensis)、黄荆(Vitex negundo)、山杏(Armeniaca sibirica)和绣线菊(Spiraea salicifolia)为伴生树种构成典型的辽东栎林(刘贤赵等, 2009; 张建华等, 2017).根据北京城区和河北多个观测点近10年的干湿氮沉降数据, 北京南部和东部大气氮沉降速率可以高达80 kg·hm-2·a-1, 北京西部地区略低, 但高峰月份也可以达到50 kg·hm-2·a-1 (王登芝等, 2006; 李金香和李峰, 2012; Pan et al., 2012; Luo et al., 2013; Xu et al., 2015; 杨文琴等, 2016; 尹兴等, 2017). ...
杂种杨无性系的光系统Ⅱ放氧活性、光合色素及叶绿体超微结构对光胁迫的响应 1 2004
... 光合作用是绿色植物转化太阳能, 合成同化物质的重要生理过程, 是植物维持生长发育的基础.氮添加的浓度和时间会对植物的叶绿素合成产生不同的影响.本研究中氮添加处理提高了辽东栎和五角枫植物的叶片氮含量, 促进了叶片的叶绿素生成, 与前人研究结果(Bauer et al., 2010; Pérez-Soba et al.,2010; Jia et al., 2011; Wang et al., 2012; 郑翔等, 2018)一致.需要说明的是, 两树种间的净光合速率和叶绿素含量均存在显著差异, 辽东栎的光合速率显著高于五角枫, 而五角枫的叶绿素含量显著高于辽东栎.此外, 氮添加对叶片叶绿素含量的促进作用随着处理时间延长表现出明显的累积效应, 五角枫比辽东栎更为明显.氮添加对光合速率、气孔导度和蒸腾速率的促进作用也随着处理时间延长表现出明显的累积效应, 辽东栎比五角枫更明显.在本研究的实验样地中, 辽东栎树体高大, 占据上层空间, 树冠上中层可以接收到充足的太阳辐射, 是群落的建群种和优势种; 而样地内的五角枫树体矮小, 处于被遮阴的下层, 可接受光照少.不同的光照环境会导致树木同化组织在结构和功能方面的极大差异, 一般阳生植物比阴生植物具有更高的羧化酶活性, 而阴生植物为了尽可能地获得林下的散射光, 会提高叶绿素含量, 形成发达的捕获色素系统, 以截获更多的光能(Shen & Ye, 1991; Johnson et al., 1993; 张守仁等, 2004; Matsubara et al., 2009; 孙小玲等, 2010; 向芬等, 2018).叶片光合作用气体交换过程主要受气孔因素和非气孔因素(主要是Rubisco羧化酶活性)影响, 在饱和光下辽东栎的羧化速率(Vcmax)和气孔导度远高于五角枫(未发表数据), 这也解释了在饱和光下辽东栎光合速率高于五角枫, 而五角枫叶绿素含量高于辽东栎的原因. ...
杂种杨无性系的光系统Ⅱ放氧活性、光合色素及叶绿体超微结构对光胁迫的响应 1 2004
... 光合作用是绿色植物转化太阳能, 合成同化物质的重要生理过程, 是植物维持生长发育的基础.氮添加的浓度和时间会对植物的叶绿素合成产生不同的影响.本研究中氮添加处理提高了辽东栎和五角枫植物的叶片氮含量, 促进了叶片的叶绿素生成, 与前人研究结果(Bauer et al., 2010; Pérez-Soba et al.,2010; Jia et al., 2011; Wang et al., 2012; 郑翔等, 2018)一致.需要说明的是, 两树种间的净光合速率和叶绿素含量均存在显著差异, 辽东栎的光合速率显著高于五角枫, 而五角枫的叶绿素含量显著高于辽东栎.此外, 氮添加对叶片叶绿素含量的促进作用随着处理时间延长表现出明显的累积效应, 五角枫比辽东栎更为明显.氮添加对光合速率、气孔导度和蒸腾速率的促进作用也随着处理时间延长表现出明显的累积效应, 辽东栎比五角枫更明显.在本研究的实验样地中, 辽东栎树体高大, 占据上层空间, 树冠上中层可以接收到充足的太阳辐射, 是群落的建群种和优势种; 而样地内的五角枫树体矮小, 处于被遮阴的下层, 可接受光照少.不同的光照环境会导致树木同化组织在结构和功能方面的极大差异, 一般阳生植物比阴生植物具有更高的羧化酶活性, 而阴生植物为了尽可能地获得林下的散射光, 会提高叶绿素含量, 形成发达的捕获色素系统, 以截获更多的光能(Shen & Ye, 1991; Johnson et al., 1993; 张守仁等, 2004; Matsubara et al., 2009; 孙小玲等, 2010; 向芬等, 2018).叶片光合作用气体交换过程主要受气孔因素和非气孔因素(主要是Rubisco羧化酶活性)影响, 在饱和光下辽东栎的羧化速率(Vcmax)和气孔导度远高于五角枫(未发表数据), 这也解释了在饱和光下辽东栎光合速率高于五角枫, 而五角枫叶绿素含量高于辽东栎的原因. ...
Atmospheric organic nitrogen deposition in China 1 2012
... 从工业革命爆发以来, 化石燃料大量燃烧, 土地利用发生大规模改变, 农业、畜牧业快速发展.活跃的工农业生产活动使全球大气中的氮化物浓度不断增加(Vitousek et al., 1997).在全球范围内, 由人类活动导致的氮沉降增加主要集中于北美、欧洲、东南亚三大地区(Galloway et al., 2003), 中国经济发达的东部地区处于全球三大高氮沉降区之一的东南亚地区.随着大规模的工业化生产和农业施肥活动, 进入陆地生态系统的氮不断增加, 经济发达的温带地区最为明显(莫江明等, 2006).根据Galloway等(2002)的预测, 未来数十年内, 随着经济的进一步发展, 这种状况会不断加剧.氮富集及氮驱动力的预测模型表明, 到21世纪中叶, 全球活性氮的排放速度将达到267 Tg·a-1, 亚洲地区由于人为活动产生的活性氮增长速度可能高达105.3 Tg·a-1 (Holland et al., 1999; Zheng et al., 2002; Galloway et al., 2004, 2008).近年来京津冀地区高浓度的氮氧化合物和其他可溶性气溶胶形成的雾霾引起人们的广泛关注, 该地区属全球典型的高氮沉降区(Zhang et al., 2012).其氮湿沉降速率平均值为38 kg·hm-2·a-1, 无机氮沉降速率约为27 kg·hm-2·a-1, 有机氮沉降速率约为7 kg·hm-2·a-1, 氮的干沉降速率也高达50 kg·hm-2·a-1.Shen等(2009)研究表明, 未来北京地区总的大气氮沉降速率可超过100 kg·hm-2·a-1. ...
UV-B辐射增强和氮沉降对不同种源地乌桕叶绿素荧光参数的影响 1 2018
... 光合作用是绿色植物转化太阳能, 合成同化物质的重要生理过程, 是植物维持生长发育的基础.氮添加的浓度和时间会对植物的叶绿素合成产生不同的影响.本研究中氮添加处理提高了辽东栎和五角枫植物的叶片氮含量, 促进了叶片的叶绿素生成, 与前人研究结果(Bauer et al., 2010; Pérez-Soba et al.,2010; Jia et al., 2011; Wang et al., 2012; 郑翔等, 2018)一致.需要说明的是, 两树种间的净光合速率和叶绿素含量均存在显著差异, 辽东栎的光合速率显著高于五角枫, 而五角枫的叶绿素含量显著高于辽东栎.此外, 氮添加对叶片叶绿素含量的促进作用随着处理时间延长表现出明显的累积效应, 五角枫比辽东栎更为明显.氮添加对光合速率、气孔导度和蒸腾速率的促进作用也随着处理时间延长表现出明显的累积效应, 辽东栎比五角枫更明显.在本研究的实验样地中, 辽东栎树体高大, 占据上层空间, 树冠上中层可以接收到充足的太阳辐射, 是群落的建群种和优势种; 而样地内的五角枫树体矮小, 处于被遮阴的下层, 可接受光照少.不同的光照环境会导致树木同化组织在结构和功能方面的极大差异, 一般阳生植物比阴生植物具有更高的羧化酶活性, 而阴生植物为了尽可能地获得林下的散射光, 会提高叶绿素含量, 形成发达的捕获色素系统, 以截获更多的光能(Shen & Ye, 1991; Johnson et al., 1993; 张守仁等, 2004; Matsubara et al., 2009; 孙小玲等, 2010; 向芬等, 2018).叶片光合作用气体交换过程主要受气孔因素和非气孔因素(主要是Rubisco羧化酶活性)影响, 在饱和光下辽东栎的羧化速率(Vcmax)和气孔导度远高于五角枫(未发表数据), 这也解释了在饱和光下辽东栎光合速率高于五角枫, 而五角枫叶绿素含量高于辽东栎的原因. ...
UV-B辐射增强和氮沉降对不同种源地乌桕叶绿素荧光参数的影响 1 2018
... 光合作用是绿色植物转化太阳能, 合成同化物质的重要生理过程, 是植物维持生长发育的基础.氮添加的浓度和时间会对植物的叶绿素合成产生不同的影响.本研究中氮添加处理提高了辽东栎和五角枫植物的叶片氮含量, 促进了叶片的叶绿素生成, 与前人研究结果(Bauer et al., 2010; Pérez-Soba et al.,2010; Jia et al., 2011; Wang et al., 2012; 郑翔等, 2018)一致.需要说明的是, 两树种间的净光合速率和叶绿素含量均存在显著差异, 辽东栎的光合速率显著高于五角枫, 而五角枫的叶绿素含量显著高于辽东栎.此外, 氮添加对叶片叶绿素含量的促进作用随着处理时间延长表现出明显的累积效应, 五角枫比辽东栎更为明显.氮添加对光合速率、气孔导度和蒸腾速率的促进作用也随着处理时间延长表现出明显的累积效应, 辽东栎比五角枫更明显.在本研究的实验样地中, 辽东栎树体高大, 占据上层空间, 树冠上中层可以接收到充足的太阳辐射, 是群落的建群种和优势种; 而样地内的五角枫树体矮小, 处于被遮阴的下层, 可接受光照少.不同的光照环境会导致树木同化组织在结构和功能方面的极大差异, 一般阳生植物比阴生植物具有更高的羧化酶活性, 而阴生植物为了尽可能地获得林下的散射光, 会提高叶绿素含量, 形成发达的捕获色素系统, 以截获更多的光能(Shen & Ye, 1991; Johnson et al., 1993; 张守仁等, 2004; Matsubara et al., 2009; 孙小玲等, 2010; 向芬等, 2018).叶片光合作用气体交换过程主要受气孔因素和非气孔因素(主要是Rubisco羧化酶活性)影响, 在饱和光下辽东栎的羧化速率(Vcmax)和气孔导度远高于五角枫(未发表数据), 这也解释了在饱和光下辽东栎光合速率高于五角枫, 而五角枫叶绿素含量高于辽东栎的原因. ...
The Asian nitrogen cycle case study 1 2002
... 从工业革命爆发以来, 化石燃料大量燃烧, 土地利用发生大规模改变, 农业、畜牧业快速发展.活跃的工农业生产活动使全球大气中的氮化物浓度不断增加(Vitousek et al., 1997).在全球范围内, 由人类活动导致的氮沉降增加主要集中于北美、欧洲、东南亚三大地区(Galloway et al., 2003), 中国经济发达的东部地区处于全球三大高氮沉降区之一的东南亚地区.随着大规模的工业化生产和农业施肥活动, 进入陆地生态系统的氮不断增加, 经济发达的温带地区最为明显(莫江明等, 2006).根据Galloway等(2002)的预测, 未来数十年内, 随着经济的进一步发展, 这种状况会不断加剧.氮富集及氮驱动力的预测模型表明, 到21世纪中叶, 全球活性氮的排放速度将达到267 Tg·a-1, 亚洲地区由于人为活动产生的活性氮增长速度可能高达105.3 Tg·a-1 (Holland et al., 1999; Zheng et al., 2002; Galloway et al., 2004, 2008).近年来京津冀地区高浓度的氮氧化合物和其他可溶性气溶胶形成的雾霾引起人们的广泛关注, 该地区属全球典型的高氮沉降区(Zhang et al., 2012).其氮湿沉降速率平均值为38 kg·hm-2·a-1, 无机氮沉降速率约为27 kg·hm-2·a-1, 有机氮沉降速率约为7 kg·hm-2·a-1, 氮的干沉降速率也高达50 kg·hm-2·a-1.Shen等(2009)研究表明, 未来北京地区总的大气氮沉降速率可超过100 kg·hm-2·a-1. ...
Nutrient limitation in three lowland tropical forests in southern China receiving high nitrogen deposition: Insights from fine root responses to nutrient additions 1 2013
... 氮沉降对温带森林影响研究从20世纪中叶开始, 已超过30年(Andersson et al., 1980; Garcia et al., 2008; Nave et al., 2015; Ma et al., 2018), 已经取得了不少成果, 但对于目前大气氮沉降如何长期影响树木生长和植被动态变化的认知程度依然十分有限(Bedison & Mcneil, 2009).近20年来国内陆续开展了关于氮沉降量增加对森林生态系统影响的研究(Mo et al., 2003; 方运霆等, 2004; 莫江明等, 2004;方华和莫江明, 2006; Lu et al., 2010; Lu et al., 2011; Zhu et al., 2013).由于在野外自然条件下进行控制实验的复杂性和高难度, 已有的氮沉降控制实验的研究对象大多以幼苗为主, 以自然森林成年大树为实验对象的研究较少(方运霆等, 2005), 基于幼苗或幼树对氮沉降的响应结果推测自然成熟森林的情况, 其代表性存在很大局限性. ...
 ,1,*1
,1,*1 
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT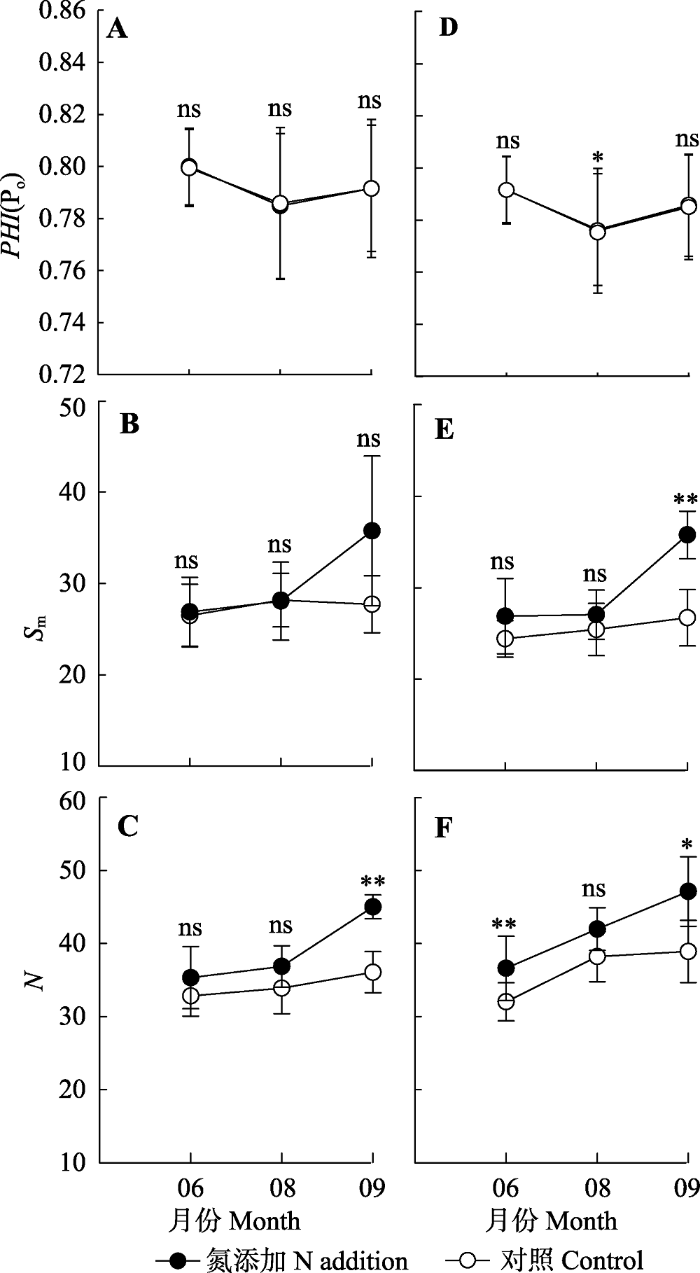 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT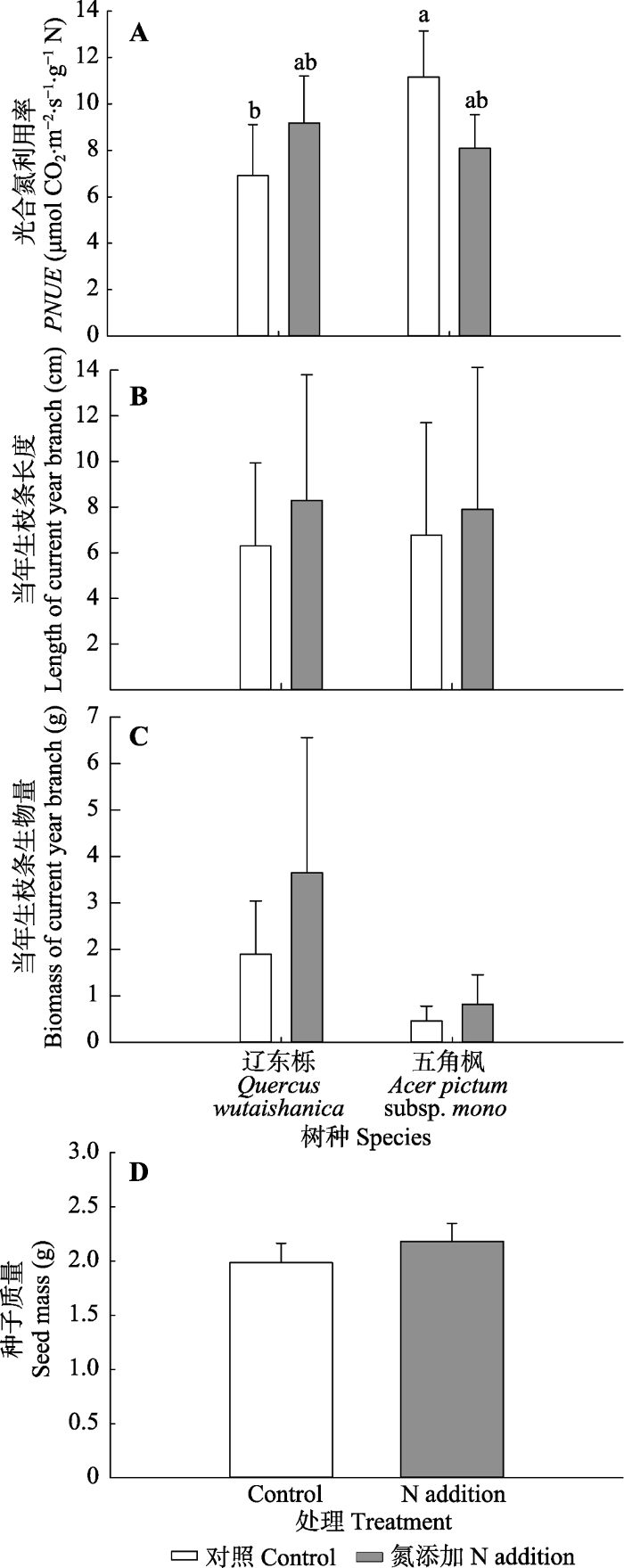 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}