Influences of stand, soil and space factors on spatial heterogeneity of leaf area index in a spruce-fir valley forest in Xiao Hinggan Ling, China
Huan-Ying YANG1, Jian-Da SONG1, Tao ZHOU1, Guang-Ze JIN2,3, Feng JIANG2, Zhi-Li LIU,2,3,*1 School of Forestry, Northeast Forestry University, Harbin 150040, China 2 Center for Ecological Research, Northeast Forestry University, Harbin 150040, China 3 Key Laboratory of Sustainable Forest Ecosystem Management-Ministry of Education, Northeast Forestry University, Harbin 150040, China
Supported by the National Natural Science Foundation of China(31600587) the China Postdoctoral Science Foundation(2016M590271) the Heilongjiang Postdoctoral Foundation(LBH-TZ1802)
Abstract AimsSpatial heterogeneity of leaf area index (LAI) is very important for exploring the growth and spatial distributions of plants, as well as response strategy of plants to climate changes. Many previous studies have shown that biotic and abiotic factors had significant influences on spatial heterogeneity of LAI. However, few studies have been conducted to show the relative contributions of different influencing factors to the total variations of LAI. Our aim was to quantify the relative contributions of stand, soil and space factors to the total spatial variations of LAI in a spruce-fir valley forest in northeast China. MethodsWe relied on a 9.12 hm 2 (380 m × 240 m) spruce-fir valley forest plot in Xiao Hinggan Ling, China, which was divided into 228 subplots (20 m × 20 m). First, we measured LAI for each subplot by using the LAI-2200 plant canopy analyzer and then analyzed the spatial heterogeneity of LAI using geo-statistic methods (semivariogram and Kriging interpolation). Second, we measured 28 stand factors and 10 soil factors for each subplot, and quantified space factors using principal coordinates of neighbor matrices (PCNM). Finally, we quantified the relative contributions of stand, soil and space factors to the total spatial variations of LAI using the variance partitioning method. Important findings The results showed that strong spatial autocorrelations of LAI values within 37 m distances in the spruce-fir valley forest, and the LAI presented different spatial patterns along distinct directions. The stand, soil and space factors totally explained 50.4% of the total spatial variations of LAI in the forest plot. The space factors explained greater spatial variations of LAI in relative to stand and soil factors, and solely explained 25.5% of the total spatial variations. The density of middle tree group (5 cm < diameter at breast height ≤ 10 cm) and basal area of major tree groups (including Abies nephrolepis and Picea spp.) were both significantly and positively correlated with LAI; and soil mass moisture content was significantly and negatively correlated with LAI. These results generally suggest that the spatial autocorrelation is more important than stand factor and soil factor for determining spatial heterogeneity of LAI of the spruce-fir valley forest in Xiao Hinggan Ling, China. Keywords:spruce-fir valley forest;leaf area index;spatial heterogeneity;geostatistical analysis
PDF (2590KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 杨焕莹, 宋建达, 周焘, 金光泽, 姜峰, 刘志理. 林分、土壤及空间因子对谷地云冷杉林叶面积指数空间异质性的影响. 植物生态学报, 2019, 43(4): 342-351. DOI: 10.17521/cjpe.2018.0310 YANG Huan-Ying, SONG Jian-Da, ZHOU Tao, JIN Guang-Ze, JIANG Feng, LIU Zhi-Li. Influences of stand, soil and space factors on spatial heterogeneity of leaf area index in a spruce-fir valley forest in Xiao Hinggan Ling, China. Chinese Journal of Plant Ecology, 2019, 43(4): 342-351. DOI: 10.17521/cjpe.2018.0310
Table 1 表1 表1小兴安岭谷地云冷杉林叶面积指数(LAI)变异函数的理论模型及参数 Table 1Theory semivariogram theoretical models and fitted parameters for leaf area index (LAI) values in a spruce-fir valley forest in Xiao Hinggan Ling, China
Fig. 3Anisotropic semivariograms at four directions (east- west (0°), south-north (90°), northeast-southwest (45°), and northwest-southeast (135°)) of leaf area index (LAI) in a spruce- fir valley forest in Xiao Hinggan Ling, China.
图4
新窗口打开|下载原图ZIP|生成PPT 图4小兴安岭谷地云冷杉林叶面积指数(LAI)在4个方向上(0° vs 90°和45° vs 135°)的各向异性比。
Fig. 4Anisotropy ratios of semivariograms between two directions (0° vs 90° and 45° vs 135°) of leaf area index (LAI) in a spruce-fir valley forest in Xiao Hinggan Ling, China.
新窗口打开|下载原图ZIP|生成PPT 图5林分、土壤和空间因子对小兴安岭谷地云冷杉林叶面积指数(LAI)变异的相对贡献率(%)。***, p < 0.001。
Fig. 5Relative contribution (%) of stand, soil and space factors to variations of leaf area index (LAI) in a spruce-fir valley forest in Xiao Hinggan Ling, China. ***, p < 0.001.
Table 2 表2 表2小兴安岭谷地云冷杉林叶面积指数(LAI)空间变异的显著影响因子统计分析 Table 2Summary statistics of significant influence factors of the spatial distribution of leaf area index (LAI) in a spruce-fir valley forest in Xiao Hinggan Ling, China.
因子 Factor
调整R2 Adjusted R2
F
p
a中等树密度 aDensitymid (N·hm-2)
0.17
46.77
0.000 1
a主要树种胸高断面积 aBAmajor (m2·hm-2)
0.05
14.03
0.000 4
a全部树阔针比 aPBBNtotal (%)
0.02
6.36
0.013 0
b容重 bBulk density (g·cm-3)
0.04
9.89
0.036 5
b质量含水率 bMass moisture content (g·g-1)
0.02
4.55
0.002 3
a, stand factors; b, soil factors. Densitymid, the density of middle tree group (5 cm < DBH ≤ 10 cm); BAmajor, basal area of major trees; PBBNtotal, the proportion of basal area of broadleaf species to needleleaf species for total trees in a plot. a, 林分因子; b, 土壤因子。
Table 3 表3 表3小兴安岭谷地云冷杉林叶面积指数(LAI)空间变异的显著影响因子统计信息 Table 3Descriptive statistical characteristics of the significant influence factors of the spatial distribution of leaf area index (LAI) in a spruce-fir valley forest in Xiao Hinggan Ling, China
参数 Parameter
林分因子 Stand factor
土壤因子 Soil factor
中等树密度 Densitymid (N·hm-2)
主要树种胸高断面积 BAmajor (m2·hm-2)
全部树阔针比 PBBNtotal
容重 Bulk density (g·cm-3)
质量含水率 Mass moisture content (g·g-1)
最大值 Maximum
2 750
37.8
2.70
0.63
5.53
最小值 Minimum
50
2.9
0.00
0.12
0.53
平均值 Mean
1 006
15.9
0.26
0.27
2.97
标准偏差 SD
553
6.3
0.31
0.11
1.15
变异系数 CV (%)
55
40
118
40
39
Densitymid, the density of middle tree group (5 cm < DBH ≤ 10 cm); BAmajor, basal area of major trees; PBBNtotal, the proportion of basal area of broadleaf species to needleleaf species for total trees in a plot.
Fig. 6Relationships between five significant influence factors and leaf area index (LAI) in a spruce-fir valley forest in Xiao Hinggan Ling, China. Densitymid, the density of middle tree group (5 cm < DBH ≤ 10 cm); BAmajor, basal area of major trees; PBBNtotal, the proportion of basal area of broadleaf species to needleleaf species for total trees in a plot.
3 讨论
3.1 LAI的空间异质性
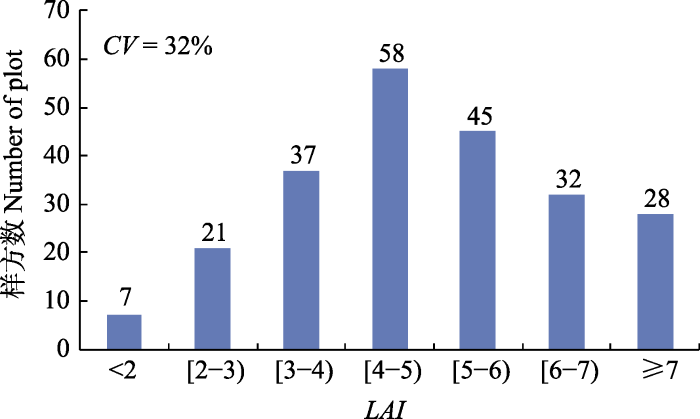
LAI能够很好地反映植物的生长状况及分布格局(Liu et al., 2015; Zhu et al., 2016), 因此, 揭示谷地云冷杉林LAI的空间异质性及其影响因素对于探究目前谷地云冷杉林处于衰退状态的原因具有重要指示作用, 还可为确定最优的调查样方面积提供科学依据。调查样方面积大, 更具代表性, 且调查结果更可靠, 但需耗费更多的人力、物力和财力; 样方面积小, 虽然降低了工作量, 但缺乏代表性, 调查结果的可靠性较差, 因此, 确定最优的调查样方面积, 对于群落生态学研究至关重要(Bequet et al., 2012b)。本研究发现谷地云冷杉林的LAI在37 m (变程)范围内具有强烈的空间自相关性, 因此, 为避免空间自相关对调查结果的影响, 测定谷地云冷杉林LAI的最优样方面积为37 m × 37 m, 这对日后研究具有重要参考价值。其他****得到类似结论, 如Zhu等(2016)通过分析7月份亚热带常绿阔叶林和落叶阔叶林内LAI的空间异质性, 得到变程(A)分别为18 m和23 m。然而也有其他****得到的变程远大于本文结果, 如姚丹丹等(2015)利用半球摄影法分析了吉林省谷地云冷杉林LAI的空间异质性, 得到平均变程为66 m; Zhu等(2016)得到7月份亚热带针阔混交林内的变程为93 m; Liu等(2018)报道小兴安岭阔叶红松林中的变程为92 m。这些不同结论可能主要源于森林类型、调查时期以及LAI测定方法的差异。此外, LAI的空间异质性不仅与尺度相关, 还受方向的影响, 方向不同, LAI呈现不同的空间分布格局(图3, 图4), 其他****也得到相同的结论(王政权等, 2000; Liu et al., 2018), 表明在日后研究LAI或其他变量的空间异质性时应给予方向性更多关注。
AndradeA, WolfeDW, FereresE ( 1993). Leaf expansion, photosynthesis, and water relations of sunflower plants grown on compacted soil Plant and Soil, 149, 175-184. [本文引用: 1]
BequetR, CampioliM, KintV, MuysB, BogaertJ, CeulemansR ( 2012a). Spatial variability of leaf area index in homogeneous forests relates to local variation in tree characteristics Forest Science, 58, 633-640. [本文引用: 3]
BequetR, KintV, CampioliM, VansteenkisteD, MuysB, CeulemansR ( 2012b). Influence of stand, site and meteorological variables on the maximum leaf area index of beech, oak and Scots pine European Journal of Forest Research, 131, 283-295. [本文引用: 4]
BivandRS, PebesmaEJ, Gómez-RubioV (2013). Applied Spatial Data Analysis with R Springer,New York. [本文引用: 1]
BorcardD, LegendreP ( 2002). All-scale spatial analysis of ecological data by means of principal coordinates of neighbour matrices Ecological Modelling, 153, 51-68. [本文引用: 1]
ChenJM, BlackTA ( 1992). Defining leaf area index for non-flat leaves Plant, Cell & Environment, 15, 421-429. [本文引用: 1]
ChenJM, RichPM, GowerST, NormanJM, PlummerS ( 1997). Leaf area index of boreal forests: Theory, techniques, and measurements Journal of Geophysical Research, 102, 29429-29443. [本文引用: 2]
DingYH, SunY ( 2006). New progress in international climate change research Advances in Climate Change Research, 2, 161-167. [本文引用: 1]
HanM, ShiB, JinG ( 2019). Spatial patterns of soil respiration in a spruce-fir valley forest, Northeast China Journal of Soils and Sediments, 19, 10-22. [本文引用: 2]
JonckheereI, FleckS, NackaertsK, MuysB, CoppinP, WeissM, BaretF ( 2004). Review of methods for in situ leaf area index determination: Part I. Theories, sensors and hemispherical photography Agricultural and Forest Meteorology, 121, 19-35. [本文引用: 1]
JonesMM, TuomistoH, BorcardD, LegendreP, ClarkDB, OlivasPC ( 2008). Explaining variation in tropical plant community composition: Influence of environmental and spatial data quality Oecologia, 155, 593-604. [本文引用: 2]
LeblancSG, ChenJM, FernandesR, DeeringDW, ConleyA (2005). Methodology comparison for canopy structure parameters extraction from digital hemispherical photography in boreal forests Agricultural and Forest Meteorology, 129, 187-207. [本文引用: 1]
LegendreP, MiXC, RenHB, MaKP, YuMJ, SunI, HeFL (2009). Partitioning beta diversity in a subtropical broad-leaved forest of China Ecology, 90, 663-674. [本文引用: 1]
LiH, ReynoldsJ (1995). On definition and quantification of heterogeneity Oikos, 73, 280-284. [本文引用: 2]
LiHB, WangZQ, WangQC (1998). Theory and methodology of spatial heterogeneity quantification Chinese Journal of Applied Ecology, 9, 651-657. [本文引用: 3]
LiWH ( 1980). Study on community structure and succession of spruce-fir valley forest in the Xiaoxing’an Mountains, China Natural Resources, 4, 17-29. [本文引用: 2]
LiuZL, ChenJM, JinGZ, QiYJ ( 2015). Estimating seasonal variations of leaf area index using litterfall collection and optical methods in four mixed evergreen-deciduous forests Agricultural and Forest Meteorology, 209, 36-48. [本文引用: 2]
LiuZL, JiangF, ZhuY, LiF, JinGZ (2018). Spatial heterogeneity of leaf area index in a temperate old-growth forest: Spatial autocorrelation dominates over biotic and abiotic factors Science of the Total Environment, 634, 287-295. [本文引用: 10]
LiuZL, JinGZ (2014). Estimation of seasonal changes in leaf area index based on optical methods in spruce-fir valley forest Chinese Journal of Applied Ecology, 25, 3420-3428. [本文引用: 1]
LiuZL, JinGZ ( 2015). Using litter collection and tree data to model leaf area index in the mixed broadleaved-Korean pine forest in the Xiaoxing’an Mountains, China Acta Ecologica Sinica, 35, 3190-3198. [本文引用: 1]
MajasalmiT, StenbergP, RautiainenM ( 2017). Comparison of ground and satellite-based methods for estimating stand-level fPAR in a boreal forest Agricultural and Forest Meteorology, 232, 422-432. [本文引用: 1]
NaithaniKJ, BaldwinDC, GainesKP, LinH, EissenstatDM (2013). Spatial distribution of tree species governs the spatio-?temporal interaction of leaf area index and soil moisture across a forested landscape PLOS ONE, 8, e58704. DOI: 10.1371/journal.pone.0058704. [本文引用: 1]
NaithaniKJ, EwersBE, PendallE (2012). Sap flux-scaled transpiration and stomatal conductance response to soil and atmospheric drought in a semi-arid sagebrush ecosystem Journal of Hydrology, 464-465, 176-185. [本文引用: 1]
OussibleM, CrookstonRK, LarsonWE ( 1992). Subsurface compaction reduces the root and shoot growth and grain yield of wheat Agronomy Journal, 84, 34-38. [本文引用: 1]
?ztürkM, Bolat?, ErgünA ( 2015). Influence of air-soil temperature on leaf expansion and LAI of Carpinus betulus trees in a temperate urban forest patch Agricultural and Forest Meteorology, 200, 185-191. [本文引用: 2]
PiaoTF, ComitaLS, JinGZ, KimJH (2013). Density dependence across multiple life stages in a temperate old-growth forest of northeast China Oecologia, 172, 207-217. [本文引用: 1]
R CoreTeam ( 2018). R: A language and environment for statistical computing R Foundation for Statistical Computing, Vienna, Austria. . URL [本文引用: 1]
RossiRE, MullaDJ, JournelAG, FranzEH ( 1992). Geostatistical tools for modeling and interpreting ecological spatial dependence Ecological Monographs, 62, 277-314.
SchleppiP, ThimonierA, WalthertL ( 2011). Estimating leaf area index of mature temperate forests using regressions on site and vegetation data Forest Ecology and Management, 261, 601-610. [本文引用: 5]
SpadavecchiaLW, Mathew, BellR, StoyPC, HuntleyB, Van WijkMT ( 2008). Topographic controls on the leaf area index and plant functional type of a tundra ecosystem Journal of Ecology, 96, 1238-1251. [本文引用: 1]
SuHX, BaiF, LiGQ ( 2012). Seasonal dynamics in leaf area index in three typical temperate montane forests of China: A comparison of multi-observation methods Chinese Journal of Plant Ecology, 36, 231-242. [本文引用: 1]
Tor-ngernP, OrenR, WardEJ, PalmrothS, McCarthyHR, DomecJC ( 2015). Increases in atmospheric CO2 have little influence on transpiration of a temperate forest canopy New Phytologist, 205, 518-525. [本文引用: 1]
von ArxG, PannatierEG, ThimonierA, RebetezM ( 2013). Microclimate in forests with varying leaf area index and soil moisture: Potential implications for seedling establishment in a changing climate Journal of Ecology, 101, 1201-1213. [本文引用: 2]
WangQG ( 2004). Study on Spruce Forest Decline Mechanism in Low-lying Land in Eastern Mountain Area in Heilongjiang Province PhD dissertation, Northeast Forestry University, Harbin.
WangZQ, WangQC, LiHB ( 2000). Characteristics and comparison of spatial heterogeneity of the main species of Korean pine in old growth forests Acta Phytoecological Sinica, 24, 718-723. [本文引用: 1]
WoodgateW, ArmstonJD, DisneyM, JonesSD, SuarezL, HillMJ, WilkesP, Soto-BerelovM ( 2016). Quantifying the impact of woody material on leaf area index estimation from hemispherical photography using 3D canopy simulations Agricultural and Forest Meteorology, 226-227, 1-12. [本文引用: 1]
WuJ, AlbertLP, LopesAP, Restrepo-CoupeN, HayekM, WiedemannKT, GuanK, StarkSC, ChristoffersenB, ProhaskaN, TavaresJV, MarosticaS, KobayashiH, FerreiraML, CamposKS, da SilvaR, BrandoPM, DyeDG, HuxmanTE, HueteAR, NelsonBW, SaleskaSR (2016). Leaf development and demography explain photosynthetic seasonality in Amazon evergreen forests Science, 351, 972-976. [本文引用: 1]
XuY, FranklinSB, WangQ, ShiZ, LuoY, LuZ, ZhangJ, QiaoX, JiangM (2015). Topographic and biotic factors determine forest biomass spatial distribution in a subtropical mountain moist forest Forest Ecology and Management, 357, 95-103. [本文引用: 2]
YangY, WangH, HarrisonSP, PrenticeIC, WrightIJ, PengC, LinG (2018). Quantifying leaf-trait covariation and its controls across climates and biomes New Phytologist, 221, 155-168. [本文引用: 3]
YaoDD, LeiXD, YuL, LuJ, FuLY, YuRG (2015). Spatial heterogeneity of leaf area index of mixed spruce-fir- deciduous stands in northeast China Acta Ecologica Sinica, 35, 71-79. [本文引用: 2]
ZhengJY, ShaoMA, ZhangXC ( 2004). Spatial variation of surface soil’s bulk density and saturated hydraulic conductivity on slope in loess region Journal of Soil and Water Conservation, 18, 53-56. [本文引用: 4]
ZhuW, XiangW, PanQ, ZengY, OuyangS, LeiP, DengX, FangX, PengC ( 2016). Spatial and seasonal variations of leaf area index (LAI) in subtropical secondary forests related to floristic composition and stand characters Biogeosciences, 13, 3819-3831. [本文引用: 6]
Within-species patterns challenge our understanding of the leaf economics spectrum 1 2018
... 空间因子(空间自相关)可以通过影响环境变量及群落组成进而影响LAI的分布(Spadavecchia et al., 2008; Liu et al., 2018).然而, 关于解析不同因子对LAI空间变异相对贡献率的报道尚少.主轴邻距法(PCNM)是能够有效量化空间结构特性的有效手段, 只是该方法以往常用于群落生态学研究中(Borcard & Legendre, 2002; Jones et al., 2008).方差分解的方法因能有效区分不同因素对某特征值变异的相对贡献率而备受关注(Xu et al., 2015; Anderegg et al., 2018; Yang et al., 2018).结合这两种方法, 即可有效区分生物因子、非生物因子和空间自相关对解释LAI空间异质性的相对贡献率.Liu等(2018)报道在小兴安岭9 hm2阔叶红松(Pinus koraiensis)林内, 相对于生物因子和非生物因子, 空间自相关对LAI的变异具有更加显著的决定作用, 然而这种结论是否具有普适性应进行进一步验证. ...
Leaf expansion, photosynthesis, and water relations of sunflower plants grown on compacted soil 1 1993
Spatial variability of leaf area index in homogeneous forests relates to local variation in tree characteristics 3 2012
... 森林生态系统的LAI通常具有较高的空间异质性(Zhu et al., 2016; Liu et al., 2018), 主要源于其具有高大、复杂的林冠结构.LAI受多种因素的影响, 如Naithani等(2013)表明LAI随海拔的升高呈增大趋势; Bequet等(2012a)报道林分密度和优势木的树高是Fagus sylvatica林LAI的主要预测因子, 而林分年龄和树木胸径(DBH)增长量对夏栎(Quercus robur)林的LAI具有决定作用; Schleppi等(2011)表明树高及乔木层、灌木层及草本层的覆盖率能显著影响LAI的变异; Zhu等(2016)报道不同林分内针叶和阔叶胸高断面积(BA)的比例对其LAI的变异具有显著影响; 姚丹丹等(2015)报道在云冷杉针阔混交林内, 林分平均DBH和平均高度与LAI显著负相关, 而林分密度和树种数与LAI显著正相关.可见, LAI空间异质性的影响因素可概括为2类: (1)能够决定植物对环境变化响应规律的生物因子, 如林分因子(DBH, BA和密度等)(Bequet et al., 2012a; Zhu et al., 2016); (2)能够影响植物生长的非生物因子, 主要包括土壤因子(土壤含水率、容重和土壤养分等)和地形因子(海拔、坡度和坡向等), 其中地形因子不仅可以直接影响植物的生长, 也可通过影响土壤因素的分布而间接影响植物的生长(Bequet et al., 2012b; Liu et al., 2018). ...
... 和密度等)(Bequet et al., 2012a; Zhu et al., 2016); (2)能够影响植物生长的非生物因子, 主要包括土壤因子(土壤含水率、容重和土壤养分等)和地形因子(海拔、坡度和坡向等), 其中地形因子不仅可以直接影响植物的生长, 也可通过影响土壤因素的分布而间接影响植物的生长(Bequet et al., 2012b; Liu et al., 2018). ...
All-scale spatial analysis of ecological data by means of principal coordinates of neighbour matrices 1 2002
... 空间因子(空间自相关)可以通过影响环境变量及群落组成进而影响LAI的分布(Spadavecchia et al., 2008; Liu et al., 2018).然而, 关于解析不同因子对LAI空间变异相对贡献率的报道尚少.主轴邻距法(PCNM)是能够有效量化空间结构特性的有效手段, 只是该方法以往常用于群落生态学研究中(Borcard & Legendre, 2002; Jones et al., 2008).方差分解的方法因能有效区分不同因素对某特征值变异的相对贡献率而备受关注(Xu et al., 2015; Anderegg et al., 2018; Yang et al., 2018).结合这两种方法, 即可有效区分生物因子、非生物因子和空间自相关对解释LAI空间异质性的相对贡献率.Liu等(2018)报道在小兴安岭9 hm2阔叶红松(Pinus koraiensis)林内, 相对于生物因子和非生物因子, 空间自相关对LAI的变异具有更加显著的决定作用, 然而这种结论是否具有普适性应进行进一步验证. ...
Defining leaf area index for non-flat leaves 1 1992
... 叶面积指数(LAI)通常定义为单位地面面积上总绿叶表面积的一半(Chen & Black, 1992).LAI对植物的光合作用、蒸腾作用及呼吸作用等许多植物生理生态过程具有调控作用(Tor-ngern et al., 2015; Wu et al., 2016), 也是常用于模拟森林生态系统碳-水循环及森林生产力的关键参数(Majasalmi et al., 2017).此外, LAI的空间异质性是植物与环境相互作用的综合体现(Spadavecchia et al., 2008; Zhu et al., 2016), 因此, 准确地量化LAI的空间异质性及其影响因素有助于了解植被的分布格局及其对全球气候变化的响应机制. ...
Leaf area index of boreal forests: Theory, techniques, and measurements 2 1997
Spatial patterns of soil respiration in a spruce-fir valley forest, Northeast China 2 2019
... 以每个20 m × 20 m的小样方为采样单位, 为捕捉细微尺度上LAI的空间变异, 除在每个样方的原点进行采样外, 另在每个原点附近随机选择一个方向(东、南、西、北、东南、东北、西南和西北), 添加距离原点2、5、8 m处的任意2个采样点, 在每个采样点取表层(0-10 cm)土样品(每个样方3个采样点) (Han et al., 2019), 并进行土壤因子的测定, 包括有机碳含量、全氮含量、水解氮含量、全磷含量、有效磷含量、有效钾含量、pH、容重、质量含水率和体积含水率.采用Kriging插值法对土壤数据进行插值, 获得5 m × 5 m网格的数据后进行分析. ...
... 地形因子也是影响LAI空间分布的一个重要非生物因子(Spadavecchia et al., 2008; Naithani et al., 2012; Liu et al., 2018), 但该区域的谷地云冷杉林处于沟谷、地势较平坦, 海拔高差小于5 m (Han et al., 2019), 因此, 本研究未考虑地形因子对LAI空间异质性的影响.近年来, 空间因子对LAI空间异质性的影响逐渐引起大家的关注(Spadavecchia et al., 2008; Liu et al., 2018).本研究结果表明空间自相关对LAI空间变异的贡献率最大, 解释了LAI总变异的46.2%, 与以往结论相符, 如Xu等(2015)表明空间自相关比地形和生物因子更能影响生物量的空间变异; Liu等(2018)也发现在阔叶红松林内空间自相关比林分、地形和土壤因子解释了更多的LAI空间变异. ...
Review of methods for in situ leaf area index determination: Part I. Theories, sensors and hemispherical photography 1 2004
Explaining variation in tropical plant community composition: Influence of environmental and spatial data quality 2 2008
... 空间因子(空间自相关)可以通过影响环境变量及群落组成进而影响LAI的分布(Spadavecchia et al., 2008; Liu et al., 2018).然而, 关于解析不同因子对LAI空间变异相对贡献率的报道尚少.主轴邻距法(PCNM)是能够有效量化空间结构特性的有效手段, 只是该方法以往常用于群落生态学研究中(Borcard & Legendre, 2002; Jones et al., 2008).方差分解的方法因能有效区分不同因素对某特征值变异的相对贡献率而备受关注(Xu et al., 2015; Anderegg et al., 2018; Yang et al., 2018).结合这两种方法, 即可有效区分生物因子、非生物因子和空间自相关对解释LAI空间异质性的相对贡献率.Liu等(2018)报道在小兴安岭9 hm2阔叶红松(Pinus koraiensis)林内, 相对于生物因子和非生物因子, 空间自相关对LAI的变异具有更加显著的决定作用, 然而这种结论是否具有普适性应进行进一步验证. ...
Estimating seasonal variations of leaf area index using litterfall collection and optical methods in four mixed evergreen-deciduous forests 2 2015
... 于2016年7-8月, 采用LAI-2200植物冠层分析仪测定每个小样方的LAI值, 包括0°-13°、16°-28°、32°-43°、47°-58°和61°-74° 5个天顶角范围内的LAI值.采集点在每个小样方的中心处, 采集时间在阴天或日出、日落前后.采集数据时确保感应探头离地面1.3 m, 且保持水平, 并使用90°顶盖.每次测量的初始值和结束值, 即天空空白值均采自样地附近的空地处.该林分的平均冠层高度(height)为18 m, 而LAI-2200测量LAI的水平覆盖面是半径为tag (30°) × (height - 1.3) m (约9.5 m)的圆形.因此, 为避免因两个样方的LAI被重复测量而影响结果, 本文采用前2环(约0°-30°天顶角) LAI值.此外, LAI-2200测定值并非真实LAI, 通常受木质部及集聚效应的影响(苏宏新等, 2012; Woodgate et al., 2016), 为提高LAI-2200测量精度, 采用校正系数1.3 (Liu et al., 2015)对其测量值进行校正, 即文中LAI均为校正后的值(图1). ...
... LAI能够很好地反映植物的生长状况及分布格局(Liu et al., 2015; Zhu et al., 2016), 因此, 揭示谷地云冷杉林LAI的空间异质性及其影响因素对于探究目前谷地云冷杉林处于衰退状态的原因具有重要指示作用, 还可为确定最优的调查样方面积提供科学依据.调查样方面积大, 更具代表性, 且调查结果更可靠, 但需耗费更多的人力、物力和财力; 样方面积小, 虽然降低了工作量, 但缺乏代表性, 调查结果的可靠性较差, 因此, 确定最优的调查样方面积, 对于群落生态学研究至关重要(Bequet et al., 2012b).本研究发现谷地云冷杉林的LAI在37 m (变程)范围内具有强烈的空间自相关性, 因此, 为避免空间自相关对调查结果的影响, 测定谷地云冷杉林LAI的最优样方面积为37 m × 37 m, 这对日后研究具有重要参考价值.其他****得到类似结论, 如Zhu等(2016)通过分析7月份亚热带常绿阔叶林和落叶阔叶林内LAI的空间异质性, 得到变程(A)分别为18 m和23 m.然而也有其他****得到的变程远大于本文结果, 如姚丹丹等(2015)利用半球摄影法分析了吉林省谷地云冷杉林LAI的空间异质性, 得到平均变程为66 m; Zhu等(2016)得到7月份亚热带针阔混交林内的变程为93 m; Liu等(2018)报道小兴安岭阔叶红松林中的变程为92 m.这些不同结论可能主要源于森林类型、调查时期以及LAI测定方法的差异.此外, LAI的空间异质性不仅与尺度相关, 还受方向的影响, 方向不同, LAI呈现不同的空间分布格局(图3, 图4), 其他****也得到相同的结论(王政权等, 2000; Liu et al., 2018), 表明在日后研究LAI或其他变量的空间异质性时应给予方向性更多关注. ...
Spatial heterogeneity of leaf area index in a temperate old-growth forest: Spatial autocorrelation dominates over biotic and abiotic factors 10 2018
... 森林生态系统的LAI通常具有较高的空间异质性(Zhu et al., 2016; Liu et al., 2018), 主要源于其具有高大、复杂的林冠结构.LAI受多种因素的影响, 如Naithani等(2013)表明LAI随海拔的升高呈增大趋势; Bequet等(2012a)报道林分密度和优势木的树高是Fagus sylvatica林LAI的主要预测因子, 而林分年龄和树木胸径(DBH)增长量对夏栎(Quercus robur)林的LAI具有决定作用; Schleppi等(2011)表明树高及乔木层、灌木层及草本层的覆盖率能显著影响LAI的变异; Zhu等(2016)报道不同林分内针叶和阔叶胸高断面积(BA)的比例对其LAI的变异具有显著影响; 姚丹丹等(2015)报道在云冷杉针阔混交林内, 林分平均DBH和平均高度与LAI显著负相关, 而林分密度和树种数与LAI显著正相关.可见, LAI空间异质性的影响因素可概括为2类: (1)能够决定植物对环境变化响应规律的生物因子, 如林分因子(DBH, BA和密度等)(Bequet et al., 2012a; Zhu et al., 2016); (2)能够影响植物生长的非生物因子, 主要包括土壤因子(土壤含水率、容重和土壤养分等)和地形因子(海拔、坡度和坡向等), 其中地形因子不仅可以直接影响植物的生长, 也可通过影响土壤因素的分布而间接影响植物的生长(Bequet et al., 2012b; Liu et al., 2018). ...
... ; Liu et al., 2018). ...
... 空间因子(空间自相关)可以通过影响环境变量及群落组成进而影响LAI的分布(Spadavecchia et al., 2008; Liu et al., 2018).然而, 关于解析不同因子对LAI空间变异相对贡献率的报道尚少.主轴邻距法(PCNM)是能够有效量化空间结构特性的有效手段, 只是该方法以往常用于群落生态学研究中(Borcard & Legendre, 2002; Jones et al., 2008).方差分解的方法因能有效区分不同因素对某特征值变异的相对贡献率而备受关注(Xu et al., 2015; Anderegg et al., 2018; Yang et al., 2018).结合这两种方法, 即可有效区分生物因子、非生物因子和空间自相关对解释LAI空间异质性的相对贡献率.Liu等(2018)报道在小兴安岭9 hm2阔叶红松(Pinus koraiensis)林内, 相对于生物因子和非生物因子, 空间自相关对LAI的变异具有更加显著的决定作用, 然而这种结论是否具有普适性应进行进一步验证. ...
... 地形因子也是影响LAI空间分布的一个重要非生物因子(Spadavecchia et al., 2008; Naithani et al., 2012; Liu et al., 2018), 但该区域的谷地云冷杉林处于沟谷、地势较平坦, 海拔高差小于5 m (Han et al., 2019), 因此, 本研究未考虑地形因子对LAI空间异质性的影响.近年来, 空间因子对LAI空间异质性的影响逐渐引起大家的关注(Spadavecchia et al., 2008; Liu et al., 2018).本研究结果表明空间自相关对LAI空间变异的贡献率最大, 解释了LAI总变异的46.2%, 与以往结论相符, 如Xu等(2015)表明空间自相关比地形和生物因子更能影响生物量的空间变异; Liu等(2018)也发现在阔叶红松林内空间自相关比林分、地形和土壤因子解释了更多的LAI空间变异. ...
... ; Liu et al., 2018).本研究结果表明空间自相关对LAI空间变异的贡献率最大, 解释了LAI总变异的46.2%, 与以往结论相符, 如Xu等(2015)表明空间自相关比地形和生物因子更能影响生物量的空间变异; Liu等(2018)也发现在阔叶红松林内空间自相关比林分、地形和土壤因子解释了更多的LAI空间变异. ...
Comparison of ground and satellite-based methods for estimating stand-level fPAR in a boreal forest 1 2017
... 叶面积指数(LAI)通常定义为单位地面面积上总绿叶表面积的一半(Chen & Black, 1992).LAI对植物的光合作用、蒸腾作用及呼吸作用等许多植物生理生态过程具有调控作用(Tor-ngern et al., 2015; Wu et al., 2016), 也是常用于模拟森林生态系统碳-水循环及森林生产力的关键参数(Majasalmi et al., 2017).此外, LAI的空间异质性是植物与环境相互作用的综合体现(Spadavecchia et al., 2008; Zhu et al., 2016), 因此, 准确地量化LAI的空间异质性及其影响因素有助于了解植被的分布格局及其对全球气候变化的响应机制. ...
Spatial distribution of tree species governs the spatio-?temporal interaction of leaf area index and soil moisture across a forested landscape 1 2013
... 森林生态系统的LAI通常具有较高的空间异质性(Zhu et al., 2016; Liu et al., 2018), 主要源于其具有高大、复杂的林冠结构.LAI受多种因素的影响, 如Naithani等(2013)表明LAI随海拔的升高呈增大趋势; Bequet等(2012a)报道林分密度和优势木的树高是Fagus sylvatica林LAI的主要预测因子, 而林分年龄和树木胸径(DBH)增长量对夏栎(Quercus robur)林的LAI具有决定作用; Schleppi等(2011)表明树高及乔木层、灌木层及草本层的覆盖率能显著影响LAI的变异; Zhu等(2016)报道不同林分内针叶和阔叶胸高断面积(BA)的比例对其LAI的变异具有显著影响; 姚丹丹等(2015)报道在云冷杉针阔混交林内, 林分平均DBH和平均高度与LAI显著负相关, 而林分密度和树种数与LAI显著正相关.可见, LAI空间异质性的影响因素可概括为2类: (1)能够决定植物对环境变化响应规律的生物因子, 如林分因子(DBH, BA和密度等)(Bequet et al., 2012a; Zhu et al., 2016); (2)能够影响植物生长的非生物因子, 主要包括土壤因子(土壤含水率、容重和土壤养分等)和地形因子(海拔、坡度和坡向等), 其中地形因子不仅可以直接影响植物的生长, 也可通过影响土壤因素的分布而间接影响植物的生长(Bequet et al., 2012b; Liu et al., 2018). ...
Sap flux-scaled transpiration and stomatal conductance response to soil and atmospheric drought in a semi-arid sagebrush ecosystem 1 2012
... 地形因子也是影响LAI空间分布的一个重要非生物因子(Spadavecchia et al., 2008; Naithani et al., 2012; Liu et al., 2018), 但该区域的谷地云冷杉林处于沟谷、地势较平坦, 海拔高差小于5 m (Han et al., 2019), 因此, 本研究未考虑地形因子对LAI空间异质性的影响.近年来, 空间因子对LAI空间异质性的影响逐渐引起大家的关注(Spadavecchia et al., 2008; Liu et al., 2018).本研究结果表明空间自相关对LAI空间变异的贡献率最大, 解释了LAI总变异的46.2%, 与以往结论相符, 如Xu等(2015)表明空间自相关比地形和生物因子更能影响生物量的空间变异; Liu等(2018)也发现在阔叶红松林内空间自相关比林分、地形和土壤因子解释了更多的LAI空间变异. ...
Geostatistical tools for modeling and interpreting ecological spatial dependence 1992
Estimating leaf area index of mature temperate forests using regressions on site and vegetation data 5 2011
... 叶面积指数(LAI)通常定义为单位地面面积上总绿叶表面积的一半(Chen & Black, 1992).LAI对植物的光合作用、蒸腾作用及呼吸作用等许多植物生理生态过程具有调控作用(Tor-ngern et al., 2015; Wu et al., 2016), 也是常用于模拟森林生态系统碳-水循环及森林生产力的关键参数(Majasalmi et al., 2017).此外, LAI的空间异质性是植物与环境相互作用的综合体现(Spadavecchia et al., 2008; Zhu et al., 2016), 因此, 准确地量化LAI的空间异质性及其影响因素有助于了解植被的分布格局及其对全球气候变化的响应机制. ...
... 森林生态系统的LAI通常具有较高的空间异质性(Zhu et al., 2016; Liu et al., 2018), 主要源于其具有高大、复杂的林冠结构.LAI受多种因素的影响, 如Naithani等(2013)表明LAI随海拔的升高呈增大趋势; Bequet等(2012a)报道林分密度和优势木的树高是Fagus sylvatica林LAI的主要预测因子, 而林分年龄和树木胸径(DBH)增长量对夏栎(Quercus robur)林的LAI具有决定作用; Schleppi等(2011)表明树高及乔木层、灌木层及草本层的覆盖率能显著影响LAI的变异; Zhu等(2016)报道不同林分内针叶和阔叶胸高断面积(BA)的比例对其LAI的变异具有显著影响; 姚丹丹等(2015)报道在云冷杉针阔混交林内, 林分平均DBH和平均高度与LAI显著负相关, 而林分密度和树种数与LAI显著正相关.可见, LAI空间异质性的影响因素可概括为2类: (1)能够决定植物对环境变化响应规律的生物因子, 如林分因子(DBH, BA和密度等)(Bequet et al., 2012a; Zhu et al., 2016); (2)能够影响植物生长的非生物因子, 主要包括土壤因子(土壤含水率、容重和土壤养分等)和地形因子(海拔、坡度和坡向等), 其中地形因子不仅可以直接影响植物的生长, 也可通过影响土壤因素的分布而间接影响植物的生长(Bequet et al., 2012b; Liu et al., 2018). ...
... 空间因子(空间自相关)可以通过影响环境变量及群落组成进而影响LAI的分布(Spadavecchia et al., 2008; Liu et al., 2018).然而, 关于解析不同因子对LAI空间变异相对贡献率的报道尚少.主轴邻距法(PCNM)是能够有效量化空间结构特性的有效手段, 只是该方法以往常用于群落生态学研究中(Borcard & Legendre, 2002; Jones et al., 2008).方差分解的方法因能有效区分不同因素对某特征值变异的相对贡献率而备受关注(Xu et al., 2015; Anderegg et al., 2018; Yang et al., 2018).结合这两种方法, 即可有效区分生物因子、非生物因子和空间自相关对解释LAI空间异质性的相对贡献率.Liu等(2018)报道在小兴安岭9 hm2阔叶红松(Pinus koraiensis)林内, 相对于生物因子和非生物因子, 空间自相关对LAI的变异具有更加显著的决定作用, 然而这种结论是否具有普适性应进行进一步验证. ...
... 地形因子也是影响LAI空间分布的一个重要非生物因子(Spadavecchia et al., 2008; Naithani et al., 2012; Liu et al., 2018), 但该区域的谷地云冷杉林处于沟谷、地势较平坦, 海拔高差小于5 m (Han et al., 2019), 因此, 本研究未考虑地形因子对LAI空间异质性的影响.近年来, 空间因子对LAI空间异质性的影响逐渐引起大家的关注(Spadavecchia et al., 2008; Liu et al., 2018).本研究结果表明空间自相关对LAI空间变异的贡献率最大, 解释了LAI总变异的46.2%, 与以往结论相符, 如Xu等(2015)表明空间自相关比地形和生物因子更能影响生物量的空间变异; Liu等(2018)也发现在阔叶红松林内空间自相关比林分、地形和土壤因子解释了更多的LAI空间变异. ...
... 空间异质性的影响逐渐引起大家的关注(Spadavecchia et al., 2008; Liu et al., 2018).本研究结果表明空间自相关对LAI空间变异的贡献率最大, 解释了LAI总变异的46.2%, 与以往结论相符, 如Xu等(2015)表明空间自相关比地形和生物因子更能影响生物量的空间变异; Liu等(2018)也发现在阔叶红松林内空间自相关比林分、地形和土壤因子解释了更多的LAI空间变异. ...
Topographic controls on the leaf area index and plant functional type of a tundra ecosystem 1 2008
... 于2016年7-8月, 采用LAI-2200植物冠层分析仪测定每个小样方的LAI值, 包括0°-13°、16°-28°、32°-43°、47°-58°和61°-74° 5个天顶角范围内的LAI值.采集点在每个小样方的中心处, 采集时间在阴天或日出、日落前后.采集数据时确保感应探头离地面1.3 m, 且保持水平, 并使用90°顶盖.每次测量的初始值和结束值, 即天空空白值均采自样地附近的空地处.该林分的平均冠层高度(height)为18 m, 而LAI-2200测量LAI的水平覆盖面是半径为tag (30°) × (height - 1.3) m (约9.5 m)的圆形.因此, 为避免因两个样方的LAI被重复测量而影响结果, 本文采用前2环(约0°-30°天顶角) LAI值.此外, LAI-2200测定值并非真实LAI, 通常受木质部及集聚效应的影响(苏宏新等, 2012; Woodgate et al., 2016), 为提高LAI-2200测量精度, 采用校正系数1.3 (Liu et al., 2015)对其测量值进行校正, 即文中LAI均为校正后的值(图1). ...
3类典型温带山地森林的叶面积指数的季节动态: 多种监测方法比较 1 2012
... 叶面积指数(LAI)通常定义为单位地面面积上总绿叶表面积的一半(Chen & Black, 1992).LAI对植物的光合作用、蒸腾作用及呼吸作用等许多植物生理生态过程具有调控作用(Tor-ngern et al., 2015; Wu et al., 2016), 也是常用于模拟森林生态系统碳-水循环及森林生产力的关键参数(Majasalmi et al., 2017).此外, LAI的空间异质性是植物与环境相互作用的综合体现(Spadavecchia et al., 2008; Zhu et al., 2016), 因此, 准确地量化LAI的空间异质性及其影响因素有助于了解植被的分布格局及其对全球气候变化的响应机制. ...
3类典型温带山地森林的叶面积指数的季节动态: 多种监测方法比较 1 2012
... 叶面积指数(LAI)通常定义为单位地面面积上总绿叶表面积的一半(Chen & Black, 1992).LAI对植物的光合作用、蒸腾作用及呼吸作用等许多植物生理生态过程具有调控作用(Tor-ngern et al., 2015; Wu et al., 2016), 也是常用于模拟森林生态系统碳-水循环及森林生产力的关键参数(Majasalmi et al., 2017).此外, LAI的空间异质性是植物与环境相互作用的综合体现(Spadavecchia et al., 2008; Zhu et al., 2016), 因此, 准确地量化LAI的空间异质性及其影响因素有助于了解植被的分布格局及其对全球气候变化的响应机制. ...
Increases in atmospheric CO2 have little influence on transpiration of a temperate forest canopy 1 2015
... LAI能够很好地反映植物的生长状况及分布格局(Liu et al., 2015; Zhu et al., 2016), 因此, 揭示谷地云冷杉林LAI的空间异质性及其影响因素对于探究目前谷地云冷杉林处于衰退状态的原因具有重要指示作用, 还可为确定最优的调查样方面积提供科学依据.调查样方面积大, 更具代表性, 且调查结果更可靠, 但需耗费更多的人力、物力和财力; 样方面积小, 虽然降低了工作量, 但缺乏代表性, 调查结果的可靠性较差, 因此, 确定最优的调查样方面积, 对于群落生态学研究至关重要(Bequet et al., 2012b).本研究发现谷地云冷杉林的LAI在37 m (变程)范围内具有强烈的空间自相关性, 因此, 为避免空间自相关对调查结果的影响, 测定谷地云冷杉林LAI的最优样方面积为37 m × 37 m, 这对日后研究具有重要参考价值.其他****得到类似结论, 如Zhu等(2016)通过分析7月份亚热带常绿阔叶林和落叶阔叶林内LAI的空间异质性, 得到变程(A)分别为18 m和23 m.然而也有其他****得到的变程远大于本文结果, 如姚丹丹等(2015)利用半球摄影法分析了吉林省谷地云冷杉林LAI的空间异质性, 得到平均变程为66 m; Zhu等(2016)得到7月份亚热带针阔混交林内的变程为93 m; Liu等(2018)报道小兴安岭阔叶红松林中的变程为92 m.这些不同结论可能主要源于森林类型、调查时期以及LAI测定方法的差异.此外, LAI的空间异质性不仅与尺度相关, 还受方向的影响, 方向不同, LAI呈现不同的空间分布格局(图3, 图4), 其他****也得到相同的结论(王政权等, 2000; Liu et al., 2018), 表明在日后研究LAI或其他变量的空间异质性时应给予方向性更多关注. ...
红松老龄林主要树种的空间异质性特征与比较的定量研究 1 2000
... 于2016年7-8月, 采用LAI-2200植物冠层分析仪测定每个小样方的LAI值, 包括0°-13°、16°-28°、32°-43°、47°-58°和61°-74° 5个天顶角范围内的LAI值.采集点在每个小样方的中心处, 采集时间在阴天或日出、日落前后.采集数据时确保感应探头离地面1.3 m, 且保持水平, 并使用90°顶盖.每次测量的初始值和结束值, 即天空空白值均采自样地附近的空地处.该林分的平均冠层高度(height)为18 m, 而LAI-2200测量LAI的水平覆盖面是半径为tag (30°) × (height - 1.3) m (约9.5 m)的圆形.因此, 为避免因两个样方的LAI被重复测量而影响结果, 本文采用前2环(约0°-30°天顶角) LAI值.此外, LAI-2200测定值并非真实LAI, 通常受木质部及集聚效应的影响(苏宏新等, 2012; Woodgate et al., 2016), 为提高LAI-2200测量精度, 采用校正系数1.3 (Liu et al., 2015)对其测量值进行校正, 即文中LAI均为校正后的值(图1). ...
红松老龄林主要树种的空间异质性特征与比较的定量研究 1 2000
... 于2016年7-8月, 采用LAI-2200植物冠层分析仪测定每个小样方的LAI值, 包括0°-13°、16°-28°、32°-43°、47°-58°和61°-74° 5个天顶角范围内的LAI值.采集点在每个小样方的中心处, 采集时间在阴天或日出、日落前后.采集数据时确保感应探头离地面1.3 m, 且保持水平, 并使用90°顶盖.每次测量的初始值和结束值, 即天空空白值均采自样地附近的空地处.该林分的平均冠层高度(height)为18 m, 而LAI-2200测量LAI的水平覆盖面是半径为tag (30°) × (height - 1.3) m (约9.5 m)的圆形.因此, 为避免因两个样方的LAI被重复测量而影响结果, 本文采用前2环(约0°-30°天顶角) LAI值.此外, LAI-2200测定值并非真实LAI, 通常受木质部及集聚效应的影响(苏宏新等, 2012; Woodgate et al., 2016), 为提高LAI-2200测量精度, 采用校正系数1.3 (Liu et al., 2015)对其测量值进行校正, 即文中LAI均为校正后的值(图1). ...
Quantifying the impact of woody material on leaf area index estimation from hemispherical photography using 3D canopy simulations 1 2016
... 叶面积指数(LAI)通常定义为单位地面面积上总绿叶表面积的一半(Chen & Black, 1992).LAI对植物的光合作用、蒸腾作用及呼吸作用等许多植物生理生态过程具有调控作用(Tor-ngern et al., 2015; Wu et al., 2016), 也是常用于模拟森林生态系统碳-水循环及森林生产力的关键参数(Majasalmi et al., 2017).此外, LAI的空间异质性是植物与环境相互作用的综合体现(Spadavecchia et al., 2008; Zhu et al., 2016), 因此, 准确地量化LAI的空间异质性及其影响因素有助于了解植被的分布格局及其对全球气候变化的响应机制. ...
Leaf development and demography explain photosynthetic seasonality in Amazon evergreen forests 1 2016
... 空间因子(空间自相关)可以通过影响环境变量及群落组成进而影响LAI的分布(Spadavecchia et al., 2008; Liu et al., 2018).然而, 关于解析不同因子对LAI空间变异相对贡献率的报道尚少.主轴邻距法(PCNM)是能够有效量化空间结构特性的有效手段, 只是该方法以往常用于群落生态学研究中(Borcard & Legendre, 2002; Jones et al., 2008).方差分解的方法因能有效区分不同因素对某特征值变异的相对贡献率而备受关注(Xu et al., 2015; Anderegg et al., 2018; Yang et al., 2018).结合这两种方法, 即可有效区分生物因子、非生物因子和空间自相关对解释LAI空间异质性的相对贡献率.Liu等(2018)报道在小兴安岭9 hm2阔叶红松(Pinus koraiensis)林内, 相对于生物因子和非生物因子, 空间自相关对LAI的变异具有更加显著的决定作用, 然而这种结论是否具有普适性应进行进一步验证. ...
Topographic and biotic factors determine forest biomass spatial distribution in a subtropical mountain moist forest 2 2015
... 空间因子(空间自相关)可以通过影响环境变量及群落组成进而影响LAI的分布(Spadavecchia et al., 2008; Liu et al., 2018).然而, 关于解析不同因子对LAI空间变异相对贡献率的报道尚少.主轴邻距法(PCNM)是能够有效量化空间结构特性的有效手段, 只是该方法以往常用于群落生态学研究中(Borcard & Legendre, 2002; Jones et al., 2008).方差分解的方法因能有效区分不同因素对某特征值变异的相对贡献率而备受关注(Xu et al., 2015; Anderegg et al., 2018; Yang et al., 2018).结合这两种方法, 即可有效区分生物因子、非生物因子和空间自相关对解释LAI空间异质性的相对贡献率.Liu等(2018)报道在小兴安岭9 hm2阔叶红松(Pinus koraiensis)林内, 相对于生物因子和非生物因子, 空间自相关对LAI的变异具有更加显著的决定作用, 然而这种结论是否具有普适性应进行进一步验证. ...
... 地形因子也是影响LAI空间分布的一个重要非生物因子(Spadavecchia et al., 2008; Naithani et al., 2012; Liu et al., 2018), 但该区域的谷地云冷杉林处于沟谷、地势较平坦, 海拔高差小于5 m (Han et al., 2019), 因此, 本研究未考虑地形因子对LAI空间异质性的影响.近年来, 空间因子对LAI空间异质性的影响逐渐引起大家的关注(Spadavecchia et al., 2008; Liu et al., 2018).本研究结果表明空间自相关对LAI空间变异的贡献率最大, 解释了LAI总变异的46.2%, 与以往结论相符, 如Xu等(2015)表明空间自相关比地形和生物因子更能影响生物量的空间变异; Liu等(2018)也发现在阔叶红松林内空间自相关比林分、地形和土壤因子解释了更多的LAI空间变异. ...
Quantifying leaf-trait covariation and its controls across climates and biomes 3 2018
... LAI能够很好地反映植物的生长状况及分布格局(Liu et al., 2015; Zhu et al., 2016), 因此, 揭示谷地云冷杉林LAI的空间异质性及其影响因素对于探究目前谷地云冷杉林处于衰退状态的原因具有重要指示作用, 还可为确定最优的调查样方面积提供科学依据.调查样方面积大, 更具代表性, 且调查结果更可靠, 但需耗费更多的人力、物力和财力; 样方面积小, 虽然降低了工作量, 但缺乏代表性, 调查结果的可靠性较差, 因此, 确定最优的调查样方面积, 对于群落生态学研究至关重要(Bequet et al., 2012b).本研究发现谷地云冷杉林的LAI在37 m (变程)范围内具有强烈的空间自相关性, 因此, 为避免空间自相关对调查结果的影响, 测定谷地云冷杉林LAI的最优样方面积为37 m × 37 m, 这对日后研究具有重要参考价值.其他****得到类似结论, 如Zhu等(2016)通过分析7月份亚热带常绿阔叶林和落叶阔叶林内LAI的空间异质性, 得到变程(A)分别为18 m和23 m.然而也有其他****得到的变程远大于本文结果, 如姚丹丹等(2015)利用半球摄影法分析了吉林省谷地云冷杉林LAI的空间异质性, 得到平均变程为66 m; Zhu等(2016)得到7月份亚热带针阔混交林内的变程为93 m; Liu等(2018)报道小兴安岭阔叶红松林中的变程为92 m.这些不同结论可能主要源于森林类型、调查时期以及LAI测定方法的差异.此外, LAI的空间异质性不仅与尺度相关, 还受方向的影响, 方向不同, LAI呈现不同的空间分布格局(图3, 图4), 其他****也得到相同的结论(王政权等, 2000; Liu et al., 2018), 表明在日后研究LAI或其他变量的空间异质性时应给予方向性更多关注. ...
Spatial and seasonal variations of leaf area index (LAI) in subtropical secondary forests related to floristic composition and stand characters 6 2016
... 森林生态系统的LAI通常具有较高的空间异质性(Zhu et al., 2016; Liu et al., 2018), 主要源于其具有高大、复杂的林冠结构.LAI受多种因素的影响, 如Naithani等(2013)表明LAI随海拔的升高呈增大趋势; Bequet等(2012a)报道林分密度和优势木的树高是Fagus sylvatica林LAI的主要预测因子, 而林分年龄和树木胸径(DBH)增长量对夏栎(Quercus robur)林的LAI具有决定作用; Schleppi等(2011)表明树高及乔木层、灌木层及草本层的覆盖率能显著影响LAI的变异; Zhu等(2016)报道不同林分内针叶和阔叶胸高断面积(BA)的比例对其LAI的变异具有显著影响; 姚丹丹等(2015)报道在云冷杉针阔混交林内, 林分平均DBH和平均高度与LAI显著负相关, 而林分密度和树种数与LAI显著正相关.可见, LAI空间异质性的影响因素可概括为2类: (1)能够决定植物对环境变化响应规律的生物因子, 如林分因子(DBH, BA和密度等)(Bequet et al., 2012a; Zhu et al., 2016); (2)能够影响植物生长的非生物因子, 主要包括土壤因子(土壤含水率、容重和土壤养分等)和地形因子(海拔、坡度和坡向等), 其中地形因子不仅可以直接影响植物的生长, 也可通过影响土壤因素的分布而间接影响植物的生长(Bequet et al., 2012b; Liu et al., 2018). ...
... ; Zhu et al., 2016); (2)能够影响植物生长的非生物因子, 主要包括土壤因子(土壤含水率、容重和土壤养分等)和地形因子(海拔、坡度和坡向等), 其中地形因子不仅可以直接影响植物的生长, 也可通过影响土壤因素的分布而间接影响植物的生长(Bequet et al., 2012b; Liu et al., 2018). ...
... LAI能够很好地反映植物的生长状况及分布格局(Liu et al., 2015; Zhu et al., 2016), 因此, 揭示谷地云冷杉林LAI的空间异质性及其影响因素对于探究目前谷地云冷杉林处于衰退状态的原因具有重要指示作用, 还可为确定最优的调查样方面积提供科学依据.调查样方面积大, 更具代表性, 且调查结果更可靠, 但需耗费更多的人力、物力和财力; 样方面积小, 虽然降低了工作量, 但缺乏代表性, 调查结果的可靠性较差, 因此, 确定最优的调查样方面积, 对于群落生态学研究至关重要(Bequet et al., 2012b).本研究发现谷地云冷杉林的LAI在37 m (变程)范围内具有强烈的空间自相关性, 因此, 为避免空间自相关对调查结果的影响, 测定谷地云冷杉林LAI的最优样方面积为37 m × 37 m, 这对日后研究具有重要参考价值.其他****得到类似结论, 如Zhu等(2016)通过分析7月份亚热带常绿阔叶林和落叶阔叶林内LAI的空间异质性, 得到变程(A)分别为18 m和23 m.然而也有其他****得到的变程远大于本文结果, 如姚丹丹等(2015)利用半球摄影法分析了吉林省谷地云冷杉林LAI的空间异质性, 得到平均变程为66 m; Zhu等(2016)得到7月份亚热带针阔混交林内的变程为93 m; Liu等(2018)报道小兴安岭阔叶红松林中的变程为92 m.这些不同结论可能主要源于森林类型、调查时期以及LAI测定方法的差异.此外, LAI的空间异质性不仅与尺度相关, 还受方向的影响, 方向不同, LAI呈现不同的空间分布格局(图3, 图4), 其他****也得到相同的结论(王政权等, 2000; Liu et al., 2018), 表明在日后研究LAI或其他变量的空间异质性时应给予方向性更多关注. ...
... 的空间异质性, 得到平均变程为66 m; Zhu等(2016)得到7月份亚热带针阔混交林内的变程为93 m; Liu等(2018)报道小兴安岭阔叶红松林中的变程为92 m.这些不同结论可能主要源于森林类型、调查时期以及LAI测定方法的差异.此外, LAI的空间异质性不仅与尺度相关, 还受方向的影响, 方向不同, LAI呈现不同的空间分布格局(图3, 图4), 其他****也得到相同的结论(王政权等, 2000; Liu et al., 2018), 表明在日后研究LAI或其他变量的空间异质性时应给予方向性更多关注. ...
 ,2,3,*
,2,3,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT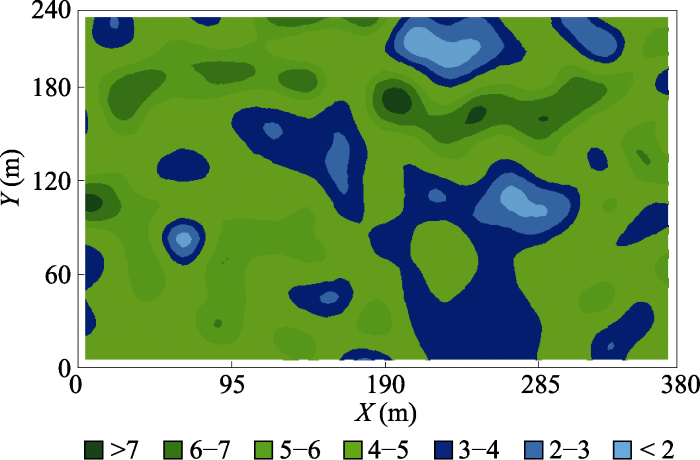 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT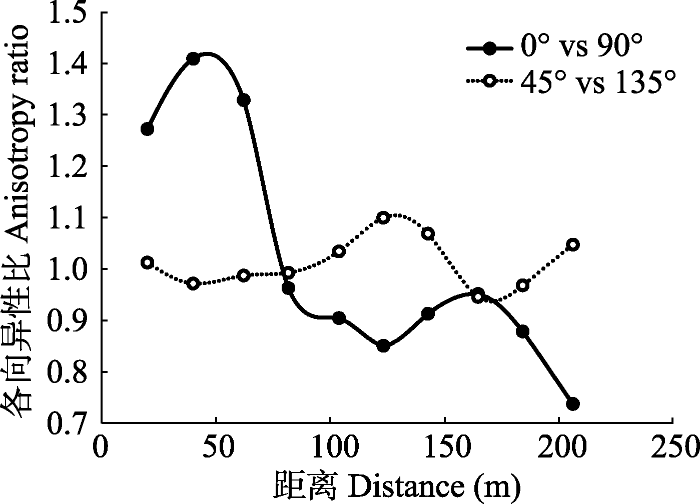 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT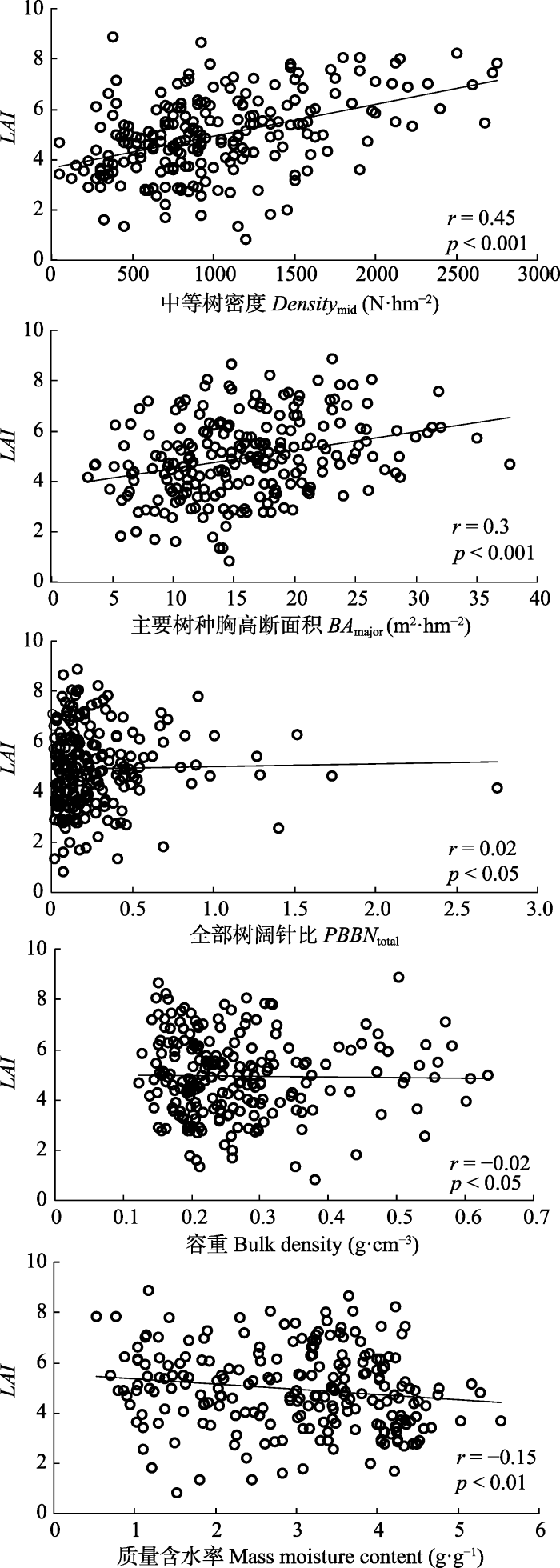 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}