杨文高
1 , 字洪标
1 , 陈科宇
1 , 阿的鲁骥
1 , 胡雷
1 , 王鑫
1 , 王根绪
2 , 王长庭
, 1 , * 1 西南民族大学青藏高原研究院, 成都 610041 2 中国科学院成都山地灾害与环境研究所中国科学院山地表生过程与生态调控重点实验室, 成都 610041 Ecological stoichiometric characteristics of shrubs and soils in different forest types in Qinghai, China Wen-Gao YANG
1 , Hong-Biao ZI
1 , Ke-Yu CHEN
1 , Lu-Ji ADE
1 , Lei HU
1 , Xin WANG
1 , Gen-Xu WANG
2 , Chang-Ting WANG
, 1 , * 1 Institute of Qinghai-Tibetan Plateau, Southwest University for Nationalities, Chengdu 610041, China 2 Key Laboratory of Mountain Surface Processes and Ecological Regulation of Chinese Academy of Sciences, Institute of Mountain Hazards and Environment, Chinese Academy of Sciences, Chengdu 610041, China 通讯作者: * wangct6@163.com 编委: 储诚进
责任编辑: 李敏
收稿日期: 2018-12-31
修回日期: 2019-04-2
网络出版日期: 2019-04-20
基金资助: 国家自然科学基金 (31870407 )国家自然科学基金 (31370542 )四川省重点研发项目 (2018SZ0333 )中央高校基本科研业务费专项资金 (2018NZD13 )
Corresponding authors: * wangct6@163.com Received: 2018-12-31
Revised: 2019-04-2
Online: 2019-04-20
Fund supported: Supported by the Natural National Science Foundation of China (31870407 )Supported by the Natural National Science Foundation of China (31370542 )the Key R&D Projects in Sichuan Province (2018SZ0333 )the Fundamental Research Funds for the Central Universities (2018NZD13 )
摘要 灌木层作为森林生态系统的重要组成部分, 了解其生态化学计量特征将有助于揭示森林生态系统物质周转和养分循环等生态功能。该研究选取青海省7种主要优势林分——白桦(
Betula platyphylla )林、毛白杨(
Populus tomentosa )林
、 红桦(
Betula albosinensis )林、青扦(
Picea wilsonii )林、山杨(
Populus davidiana )林、圆柏(
Sabina chinensis )林、云杉(
Picea asperata )林为研究对象, 采用野外取样和室内实验分析相结合的方法, 研究了不同林分林下灌木层不同器官(叶、枝干、根)及其表层(0-10 cm)土壤的碳(C)、氮(N)、磷(P)含量及其相关性。结果表明: 7种林分间灌木(叶、枝干、根) P含量、C:P均没有明显差异性; 山杨林、圆柏林、云杉林的林下灌木(叶、枝干、根) N含量、N:P高于白桦林、毛白杨林、红桦林和青扦林, C:N则相反。圆柏林的林下灌木生长受P限制, 其余6种林分的林下灌木生长受N限制。7种林分间土壤有机碳(SOC)和总氮(TN)含量呈现出明显差异性, 而总磷(TP)含量则差异不明显。相关性分析表明, 林下灌木(叶、枝干、根) N含量、C:N、N:P与土壤TN含量、C:N、N:P呈极显著相关性, 而P含量、C:P与土壤TP含量呈显著相关性。冗余分析表明, 林下灌木层植被C、N、P含量及生态化学计量特征受到土壤化学计量特征及各环境因子的共同影响, 其中土壤C:N、海拔、年平均气温、年降水量为主要影响因子。
关键词: 生态化学计量学 ;
森林生态系统 ;
林分类型 ;
灌木层 ;
环境因子 ;
青海 Abstract Aims As a dominant understory layer, shrubs is important in the material turnover and nutrient circulation of forested ecosystems. It is essential to explore stoichiometric characteristics of carbon (C), nitrogen (N) and phosphorus (P) of the shrubs and their driving factors, including microenvironments and soil nutrients.
Methods The leaves, branches, roots of the shrubs and the soils they rooted were sampled from seven dominant forest types of Qinghai, China, and the tissue contents of C, N and P were examined. One-way ANOVA was used to explored the difference of the shrubs and the soils among the forest types using, respectively. Redundancy analysis (RDA) was used to analyze the effects of soils and environmental factors on the stoichiometric characteristics of C, N and P of shrubs.
Important findings Our results showed that there were no significant differences in the P content and C:P of the leaves, branches and roots among all the seven dominant forest types, while the N content and N:P of shrubs in the
Populus davidiana, Sabina chinensis and
Picea asperata forests were significantly higher than those in
Betula platyphylla, Populus tomentosa, Betula albosinensis and
Picea wilsonii forests, while the C:N ratios were the other way around. The shrubs in
Sabina chinensis forest were limited by the soil P content, but that in the other six forest types was limited by the soil N content. The contents of soil organic C (SOC) and soil total N (TN) were significantly different among the seven forest types, while the soil total P (TP) was not. Correlation analysis showed that the N content, the C:N and N:P of understory shrub tissues (leaves, branches and roots) were significantly correlated with soil TN content, soil C:N and N:P, while tissue P contents and the C:P ratios were correlated with soil TP contents. Redundancy analysis (RDA) showed that the stoichiometric characteristics of C, N and P the understory shrub layer were synthetically affected by soils and environmental factors, of which the soil C:N, altitude, mean annual temperature and mean annual precipitation were the main influence factors.
Keywords: ecological stoichiometry ;
forest ecosystem ;
forest type ;
shrub layer ;
environmental factor ;
Qinghai PDF (7618KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 杨文高, 字洪标, 陈科宇, 阿的鲁骥, 胡雷, 王鑫, 王根绪, 王长庭. 青海森林生态系统中灌木层和土壤生态化学计量特征. 植物生态学报, 2019, 43(4): 352-364. DOI:
10.17521/cjpe.2018.0326 YANG Wen-Gao, ZI Hong-Biao, CHEN Ke-Yu, ADE Lu-Ji, HU Lei, WANG Xin, WANG Gen-Xu, WANG Chang-Ting.
Ecological stoichiometric characteristics of shrubs and soils in different forest types in Qinghai, China .
Chinese Journal of Plant Ecology , 2019, 43(4): 352-364. DOI:
10.17521/cjpe.2018.0326 碳(C)、氮(N)、磷(P)作为最重要的生命元素, 是维系地球上所有生命活动的基础(
贺金生和韩兴国, 2010 )。C作为植物有机体结构的重要组成元素, 是植物有机体内生理生化反应过程的能量来源之一(
Güsewell, 2004 ), N、P是植物的基本营养元素, 也是植物有机体内蛋白质、酶类、核酸和脂质的重要组成元素(
Yang & Wang, 2011 ;
戚德辉等, 2016 ;
Tang et al ., 2018 )。C、N、P含量及其化学计量比的差异显著影响植物的生态策略, 从而被广泛应用到生态系统在生物地球化学循环和生态过程耦合关系的研究中(
Elser et al ., 1996 )。截至目前, 国内外****已在区域乃至全球范围开展了许多陆地生态系统生态化学计量特征的研究(
Han et al ., 2005 ;
Elser et al ., 2007 ;
黄小波等, 2016 )。但主要集中在植物叶片和土壤生态化学计量特征方面, 针对植物枝干、根等重要器官方面的研究相对较少(
贺合亮等, 2017 )。枝干是植物连接地下吸收器官和地上同化组织的传导器官, 具有支撑植物体的结构性骨架作用(
李单凤等, 2015 )。根系作为吸收、存储、运输水分和养分的主要器官, 也是植物有机体对外界环境变化响应最为敏感的地下器官, 在森林生态系统物质循环过程中发挥关键作用(
Iversen et al ., 2017 ;
陈晓萍等, 2018 )。因此, 研究植物不同器官间生态化学计量特征以及与土壤组分在各生态过程间的互作关系, 对于揭示生态系统养分循环规律具有重要意义。
在森林生态系统中, 林下灌木层与乔木层和草本层共同维持着森林生态系统结构与功能的稳定, 在参与养分循环、改善土壤肥力、为林下生物提供栖息环境和提高生态系统多样性等方面起到重要作用, 是森林生态系统不可或缺的组成部分(
郑绍伟等, 2007 ;
卢振龙和龚孝生, 2009 )。灌木植被通常具有木质化茎干而主干不明显, 其茎干多分枝, 树冠矮小, 根系分布广而深(
李清河等, 2006 )。目前, 对森林的林下灌木层的研究主要聚焦于灌木生物量生长模型的建立(
万五星等, 2014 ), 物种多样性和种间联结性(
崔宁洁等, 2014 ), 生态位以及物种分布特征的分析(
康永祥等, 2008 )。对于林下灌木层生态化学计量特征的研究相对较少, 尤其是高寒地区不同林分类型中灌木层的研究尚鲜有报道。
青海省位于青藏高原的东北部, 深居内陆腹地, 是全球气候变化的敏感区和脆弱区, 也是我国乃至亚洲重要的生态安全屏障(
贾敬敦等, 2004 )。青海森林覆盖面积小, 但植被类型多样化, 其森林生态系统及林下灌木层植被在高寒区水土保持, 涵养水源, 防风固沙, 维系生态系统平衡与稳定等方面发挥着关键的生态功能(
张胜邦, 2005 )。青海地区森林凋落物层较厚, 矿质土壤层较薄, 同时受雪被和季节性冻融等特殊因素的综合影响, 其森林生态系统物质循环过程可能具有其独特性(
左巍等, 2016 )。由于特殊的地理位置, 气候寒冷而干旱, 地形地貌复杂多样, 使植被对气候变化异常敏感和脆弱(
王根绪等, 2006 )。因此, 研究青海森林的林下灌木层生态化学计量特征将有助于我们深入了解高寒区森林生态系统的稳定性以及养分循环机理, 揭示高寒环境下森林植被的生态适应策略。基于此, 本研究选取青海省7种主要优势林分——白桦(
Betula platyphylla )林、毛白杨(
Populus tomentosa )林
、 红桦(
Betula albosinensis )林、青扦(
Picea wilsonii )林、山杨(
Populus davidiana )林、圆柏(
Sabina chinensis )林、云杉(
Picea asperata )林的林下灌木层植被为研究对象, 通过分析不同林分林下灌木植被和土壤生态化学计量特征, 阐明林下灌木植被化学计量特征与土壤组分之间的耦合关系, 并探讨林下灌木植被的养分限制情况及其生态化学计量特征的影响因素, 旨在为研究林下灌木植被对高寒环境的响应与适应机制提供基础数据, 也为指导青海高寒区森林生态系统的保护、恢复与重建提供科学依据。
1 材料和方法 1.1 研究区概况 青海省地处青藏高原的东北部, 地理位置为31.65°-39.32° N, 89.58°-103.07° E。地形复杂多样, 高山、丘陵、河谷、盆地交错分布, 平均海拔3 000 m以上, 属于典型高原大陆性气候。常年平均气温-3.7-6.0 ℃, 年日照时间2 340-3 550 h, 年降水量16.7-776.1 mm (大部分400 mm以下), 年蒸发量1 118.4-3 536.2 mm (大部分1 500 mm以上), 植被地域跨青藏高原、温带荒漠和温带草原3个植被区, 具有高寒和旱生的特点。青海省森林资源贫乏, 森林覆盖率仅有4.57%, 森林植被主要分布于江河及其支流的河谷两岸, 海拔大多在2 000-4 000 m, 以寒温性针叶林为主, 其次为落叶阔叶林(
字洪标等, 2017 )。常见的针叶树种有云杉、圆柏等, 阔叶树种有杨树(
Populus spp.)、白桦等, 林下灌木有金露梅(
Potentilla fruticosa )、鲜卑花(
Sibiraea laevigata )、野蔷薇(
Rosa multiflora )
、 银露梅(
Potentilla glabra )
、 杯腺柳(
Salix cupularis )等, 主要的森林土壤类型有棕壤、黄棕壤、暗棕壤、褐棕壤、砂壤土等(
董旭, 2009 ;
胡雷等, 2015 )。
1.2 试验设计与取样方法 针对青海省森林生态系统分布状况, 并结合全省各森林类型分布面积与蓄积比重、林龄、起源等情况, 于植物生长旺盛季(7-8月)在全省21个县范围内选择80个标准样地(
图1 ), 同时记录该样地坡度、海拔、坡向、郁闭度等环境因子, 在每个标准样地中随机设置3块50 m × 20 m的乔木样方, 各样方间距大于100 m, 总计240块乔木样方。在上述乔木样方内采用对角线设置3个2 m × 2 m灌木样方, 记录灌木名称、株(丛)数、总盖度、平均高度和平均基径(附录I)。将样方内的灌木植被全部收获, 同时按灌木不同部位(叶、枝干、根)进行分类混合后带回实验室, 置于恒温烘箱中65 ℃烘干至恒质量, 样品预处理后测定C、N、P含量。在每个乔木调查样方内采用对角线设置3个1 m × 1 m土壤调查样方, 在每个土壤调查样方内用土钻法(内径5 cm)分5层(0-10、10-20、20-30、30-50、50-100 cm)分别钻取土壤样品(不够100 cm至基岩为止), 同一层土壤样品混合为1个土壤样品。土壤样品预处理后测定土壤有机碳(SOC)、全氮(TN)、全磷(TP)含量。由于森林生态系统表层土壤(0-10 cm)具有表聚性, 其SOC、TN、TP含量从表层向下层形成垂直递减的分布格局(
李婷等, 2015 ), 所以本研究选取该土层来探讨土壤化学计量特征。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1青海森林样地点位图。 Fig. 1Map of the sampling forests in Qinghai. 1.3 样品测定 所有植物样品和土壤样品均按照
生态系统固碳项目技术规范编写组(2015) 制定的统一测量方法测定, 其中植物C和SOC含量采用重铬酸钾——外加热法测定; 植物N和土壤TN含量采用凯氏定氮法测定; 植物P含量采用浓硫酸-过氧化氢消煮——钼锑抗分光光度法测定, 土壤TP含量采用浓硫酸-高氯酸消煮——钼锑抗分光光度法测定。
1.4 数据分析 采用SPSS 19.0软件对数据进行统计分析, 采用Origin 8.5进行作图。采用单因素方差分析法检验测定不同林分林下灌木各器官以及土壤C、N、P含量及化学计量比的差异, 若方差齐性, 用最小显著差异(
LSD )法进行显著性多重比较; 若方差非齐性, 则用Tamhanes T2法进行多重比较, 显著性水平为
α = 0.05。运用Pearson相关分析方法对林下灌木各器官和土壤C、N、P含量及化学计量比之间进行相关性分析, 显著性水平设为
α = 0.05。采用CANOCO (Version 4.5)对灌木各器官养分及环境因子进行冗余分析(RDA)排序, 并绘制二维排序图, 其中各采样点的气象数据(年降水量、年平均气温)从相关的文献整理得出。图表中数据为平均值±标准误差。
2 结果 2.1 灌木C、N、P含量特征 不同林分林下灌木叶、枝干、根C含量变化范围分别为434.5-467.8、454.45-468.00、429.22-462.34 mg·g
-1 , 平均值为449.00、466.69、450.14 mg·g
-1 ; N含量变化范围为10.13-21.98、2.93-5.80、2.76-5.43 mg·g
-1 , 平均值为19.46、4.62、4.03 mg·g
-1 ; P含量变化范围为1.36-1.88、0.43-0.59、0.38-0.54 mg·g
-1 , 平均值为1.69、0.56、0.50 mg·g
-1 。不同林分林下灌木相同器官间, 青扦林灌木叶片C含量显著高于毛白杨林(
p < 0.05), 青扦林灌木枝干C含量显著低于其他林分(
p < 0.05); 山杨林、圆柏林、云杉林灌木叶、枝干N含量显著高于其他林分(
p < 0.05), 圆柏林和云杉林灌木根N含量显著高于白桦林、毛白杨林、红桦林和青扦林(
p < 0.05); 云杉林叶P含量显著高于圆柏林(
p < 0.05), 而灌木枝干和根P含量在所有林分之间没有显著差异(
p > 0.05)。同一林分不同器官间, 毛白杨林、云杉林灌木枝干C含量显著高于叶和根C含量(
p < 0.05), 圆柏林和白桦林灌木叶、枝干与根之间C含量差异显著(
p < 0.05); 所有林分中灌木叶片N、P含量均分别显著高于枝干和根N、P含量(
p < 0.05)?(
图2 )。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2青海不同林分林下灌木C、N、P含量特征(平均值±标准误差)。不同大写字母表示不同林分林下灌木相同器官C、N、P含量差异显著(p < 0.05), 不同小写字母表示相同林分林下灌木不同器官C、N、P含量差异显著(p < 0.05)。 Fig. 2The C, N and P contents of shrubs of the different forest types in Qinghai (mean ± SE ). Different uppercase letters indicate significant differences of the C, N and P contents in the same organs of shrubs under different forest types (p < 0.05), and different lowercase letters indicate significant differences of the C, N and P contents in different organs of shrubs under the same forest type (p < 0.05). 2.2 灌木C、N、P化学计量比特征 不同林分林下灌木叶、枝干、根C:N变化范围为21.70-44.59、112.02-162.55、90.38-173.68, 平均值为28.80、121.62、128.89; C:P变化范围为256.14-349.14、905.65-1 108.43、1 045.53-1 212.60, 平均值为300.44、962.07、1 054.13; N:P变化范围为7.21-16.72、5.64-14.14、5.17-14.41, 平均值为12.24、9.16、9.15。不同林分林下灌木相同器官间, 山杨林、圆柏林、云杉林灌木叶和枝干C:N均显著低于其他林分(
p < 0.05), 白桦林、毛白杨林灌木根C:N显著高于山杨林、圆柏林和云杉林(
p < 0.05); 7种林分林下灌木叶、枝干和根C:P均无显著差异(
p > 0.05); 山杨林、圆柏林、云杉林灌木叶N:P显著高于白桦林、红桦林、毛白杨林和青扦林(
p < 0.05), 圆柏林灌木枝干和根N:P均显著高于其他林分(
p < 0.05)。同一林分不同器官间, 7种林分林下灌木枝干和根C:N显著高于叶C:N (
p < 0.05); 毛白杨林、红桦林、青扦林和圆柏林的林下灌木叶C:P显著低于枝干和根C:P (
p < 0.05), 白桦林、山杨林、云杉林灌木叶、枝干、根C:P三者间差异显著(
p < 0.05); 而白桦林、山杨林、云杉林灌木叶N:P显著高于枝干和根(
p < 0.05)?(
图3 )。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3青海不同林分林下灌木C:N、C:P、N:P特征(平均值±标准误差)。不同大写字母表示不同林分林下灌木相同器官C:N、C:P、N:P差异显著(p < 0.05), 不同小写字母表示相同林分林下灌木不同器官C:N、C:P、N:P差异显著(p < 0.05)。 Fig. 3The C:N, C:P and N:P ratios in shrubs of different forest types in Qinghai (mean ± SE ). Different uppercase letters indicate significant differences of the C:N、C:P、N:P in the same organs of shrubs under different forest types (p < 0.05), and different lowercase letters indicate significant differences of the C:N、C:P、N:P in different organs of shrubs under the same forest type (p < 0.05). 2.3 SOC、TN、TP含量及化学计量比特征 不同林分SOC、TN、TP含量变化范围为59.21- 133.81、2.95-5.86、0.71-0.87 mg·g
-1 , 平均值为95.11、5.17、0.82 mg·g
-1 ; C:N、C:P、N:P变化范围为15.70-28.39、82.02-121.75、3.99-7.67, 平均值为20.55、117.72、6.35。其中红桦林SOC含量显著高于白桦林、毛白杨林、圆柏林和山杨林(
p < 0.05); 圆柏林和云杉林土壤TN含量显著高于毛白杨林(
p < 0.05); 所有林分之间土壤TP含量无显著差异(
p > 0.05)。圆柏林和山杨林土壤C:N显著低于红桦林和青扦林(
p < 0.05); 红桦林土壤C:P显著高于白桦林、毛白杨林和山杨林(
p < 0.05); 毛白杨林土壤N:P显著低于圆柏林和云杉林(
p < 0.05)(
表1 )。
Table 1 表1 表1 青海不同林分土壤有机碳(SOC)、全氮(TN)、全磷(TP)含量及化学计量比特征(平均值±标准误差)
Table 1
Soil organic carbon (SOC), total nitrogen (TN), total phosphorus (TP) content and stoichiometry characteristics in different forest types in Qinghai (mean ± SE ) 林分类型 SOC含量 -1 ) TN含量 -1 ) TP含量 -1 ) C:N C:P N:P 白桦林 Betula platyphylla forest 91.43 ± 33.66bc 3.85 ± 1.43ab 0.84 ± 0.12a 24.39 ± 4.24ab 109.57 ± 39.04b 4.58 ± 1.56bc 毛白杨林 Populus tomentosa forest 72.80 ± 14.33bc 2.95 ± 0.52b 0.75 ± 0.08a 24.77 ± 2.83ab 99.34 ± 25.78b 3.99 ± 0.82c 红桦林 Betula albosinensis forest 133.81 ± 12.03a 4.90 ± 0.77ab 0.83 ± 0.02a 27.53±2.20a 161.04 ± 12.45a 5.89 ± 0.82abc 青扦林 Picea wilsonii forest 99.11 ± 24.60ab 3.49 ± 0.67ab 0.85 ± 0.18a 28.39 ± 4.14a 118.21 ± 25.62ab 4.23 ± 0.95bc 山杨林 Populus davidiana forest 59.21 ± 22.57c 3.60 ± 0.94ab 0.71 ± 0.16a 16.6 ± 5.09c 82.02 ± 18.60b 5.29 ± 1.94abc 圆柏林 Sabina chinensis forest 88.97 ± 49.70bc 5.86 ± 2.92a 0.72 ± 0.81a 15.70 ± 5.63c 120.54 ± 59.85ab 7.67 ± 3.53a 云杉林 Picea asperata forest 100.20 ± 37.05ab 5.85 ± 2.80a 0.87 ± 0.18a 19.50 ± 7.88bc 121.75 ± 44.38ab 6.97 ± 3.06ab 平均值 Average 95.11 ± 37.63 5.17 ± 2.60 0.82 ± 0.16 20.55 ± 7.35 117.72 ± 44.32 6.35 ± 2.98
Different lowercase letters indicate significant differences of SOC, TN, TP content and stoichiometry characteristics in different forest types (p < 0.05). 不同小写字母表示不同林分SOC、TN、TP含量及化学计量比差异显著(p < 0.05)。 新窗口打开 |
下载CSV 2.4 灌木各器官与土壤C、N、P及化学计量比之间的相关性分析 叶片C含量与SOC、TN、TP含量呈显著正相关关系(
p < 0.05); 根C含量与SOC含量、土壤C:P呈极显著正相关关系(
p < 0.01), 与土壤N:P呈显著正相关关系(
p < 0.05); 叶片N、P含量与土壤TN、TP含量呈极显著正相关关系(
p < 0.01), 与土壤C:N呈极显著负相关关系(
p < 0.01); 枝干和根N含量与土壤TN含量、N:P呈极显著正相关关系(
p < 0.01), 与土壤C:N呈极显著负相关关系(
p < 0.01); 枝干P含量与SOC含量、C:N呈显著负相关关系(
p < 0.05), 与土壤TP含量呈极显著正相关关系(
p < 0.01); 根P含量与土壤TP含量呈显著正相关关系(
p < 0.05), 枝干和根P含量与C:P呈极显著负相关关系(
p < 0.01); 叶片、枝干和根C:N与土壤TN含量、N:P呈极显著负相关关系(
p < 0.01), 与土壤C:N极显著正相关关系(
p < 0.01); 叶片C:P与土壤TP含量呈极显著负相关关系(
p < 0.01); 枝干和根C:P与土壤C:P呈极显著正相关关系(
p < 0.01), 与土壤TP含量呈显著负相关关系(
p < 0.05); 叶片、枝干和根N:P与土壤TN含量、N:P呈极显著正相关关系(
p < 0.01), 与土壤C:N极显著负相关关系(
p < 0.01)(
表2 )。
Table 2 表2 表2 青海不同林分灌木各器官与土壤C、N、P含量及化学计量比之间的相关性
Table 2
Correlation among the shrubs and soil C, N and P stoichiometric characteristics in different forest types in Qinghai 器官 Organ 项目 Item 土壤 Soil SOC TN TP C:N C:P N:P 叶 Leaf C 0.159* 0.166* 0.160* -0.078 0.100 0.106 N 0.063 0.614** 0.224** -0.759** -0.028 0.561** P 0.005 0.214** 0.427** -0.261** -0.185* 0.022 C:N -0.054 -0.524** -0.134 0.662** 0.001 -0.514** C:P -0.023 -0.116 -0.354** 0.106 0.139 0.048 N:P 0.067 0.496** -0.100 -0.617** 0.122 0.596** 枝干 Branch C 0.048 0.012 0.032 -0.019 0.063 0.011 N -0.083 0.457** 0.087 -0.742** -0.137 0.444** P -0.162* -0.005 0.230** -0.175* -0.277** -0.120 C:N 0.026 -0.514** -0.068 0.757** 0.072 -0.516** C:P 0.122 0.012 -0.190* 0.105 0.227** 0.110 N:P 0.018 0.328** -0.113 -0.453** 0.063 0.397** 根 Root C 0.239** 0.120 -0.084 0.142 0.301** 0.175* N -0.084 0.340** -0.076 -0.575** -0.052 0.413** P -0.132 -0.024 0.188* -0.107 -0.209** -0.106 C:N 0.058 -0.349** 0.008 0.573** 0.053 -0.396** C:P 0.135 0.116 -0.176* -0.016 0.202** 0.189* N:P -0.002 0.286** -0.207** -0.422** 0.077 0.397**
SOC, 土壤有机碳; TN, 全氮; TP, 全磷。*, p < 0.05; **, p < 0.01。 SOC, soil organic carbon; TN, total nitrogen; TP, total phosphorus. *, p < 0.05; **, p < 0.01. 新窗口打开 |
下载CSV 2.5 灌木各器官C、N、P含量及生态化学计量比与环境因子之间的关系 由灌木(叶片、枝干、根) C、N、P含量及生态化学计量比与环境因子之间的RDA结果(
图4 ;
表3 )可知, 海拔、年平均气温、年降水量、坡度、郁闭度和SOC含量、土壤TN含量、土壤TP含量、土壤C:N、土壤C:P、土壤N:P共11个环境因子解释了灌木叶片、枝干和根C、N、P含量及生态化学计量比特征变异的66.7%、54.9%、38.8%。由各环境因子解释量(
表3 )可以得出, 对叶片C、N、P含量及生态化学计量比特征影响的大小为: 土壤C:N (44.2%) >海拔(8.0%) >年降水量(7.2%) >年平均气温(2.5%) >土壤N:P (1.5%) >土壤TN含量(1.0%) > SOC含量(0.9%), 其中土壤C:N、海拔、年降水量、年平均气温和土壤N:P的影响达到极显著水平(
p < 0.01), 土壤TN和SOC含量的影响达到显著水平(
p < 0.05)。对枝干C、N、P含量及生态化学计量比特征影响的大小为: 土壤C:N (38.1%) >海拔(6.1%) >年降水量(3.9%) >年平均气温(2.1%) > SOC含量(1.5%) >土壤N:P (1.2%), 其中土壤C:N、海拔、年降水量、年平均气温和SOC含量的影响达到极显著水平(
p < 0.01), 土壤N:P的影响达到显著水平(
p < 0.05)。对根C、N、P含量及生态化学计量比特征影响的大小为: 土壤C:N (19.2%) >年平均气温(5.2%) >海拔(4.2%) >年降水量(3.2%) >土壤C:P (3.1%) >郁闭度(1.3%) >坡度(1.2%), 其中土壤C:N、年平均气温、海拔、年降水量和土壤C:P的影响达到极显著水平(
p < 0.01), 郁闭度和坡度的影响达到显著水平(
p < 0.05)。
图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4青海不同林分灌木叶片(A)、枝干(B)和根(C) C、N、P含量及化学计量比与环境因子之间的冗余分析二维排序图。Altitude, 海拔; Canopy, 郁闭度; Slope, 坡度; MAP, 年降水量; MAT, 年平均气温; SC, 土壤碳含量; SN, 土壤氮含量; SP, 土壤磷含量; SCN, 土壤C:N; SCP, 土壤C:P; SNP, 土壤N:P; LC, 叶片碳含量; LN, 叶片氮含量; LP, 叶片磷含量; BC, 枝干碳含量; BN, 枝干氮含量; BP, 枝干磷含量; RC, 根碳含量; RN, 根氮含量; RP, 根磷含量。 Fig. 4Two-dimensional sequence diagram of redundancy analysis between the C, N and P contents and their ratios in leaves (A), branches (B) and roots (C) of shrubs with environmental factors in different forest types in Qinghai. MAP, mean annual precipitation; MAT, mean annual air temperature; SC, soil carbon content; SN, soil nitrogen content; SP, soil phosphorus content; SCN, soil C:N; SCP, soil C:P; SNP, soil N:P; LC, leaf carbon content; LN, leaf nitrogen content; LP, leaf phosphorus content; BC, branch carbon content; BN, branch nitrogen content; BP, branch phosphorus content; RC, root carbon content; RN, root nitrogen content; RP, root phosphorus content. Table 3 表3 表3 青海不同林分灌木环境因子解释量与显著性检验
Table 3
Explained variance of environmental factors and significant test of shrubs in different forest types in Qinghai 器官 环境因子 环境因子解释量(%) p F 叶片 Leaf SCN 44.2 0.002 132.5 Altitude 8.0 0.002 32.68 MAP 7.2 0.002 24.68 MAT 2.5 0.002 10.94 SNP 1.5 0.006 6.62 SN 1.0 0.016 4.45 SC 0.9 0.014 3.96 SCP 0.7 0.052 3.22 SP 0.5 0.128 2.29 Slope 0.3 0.302 1.273 Canopy 0.0 0.876 0.16 枝干 Branch SCN 38.1 0.002 99.01 Altitude 6.1 0.002 17.47 MAP 3.9 0.002 12.09 MAT 2.1 0.002 6.68 SC 1.5 0.006 4.85 SNP 1.2 0.018 4.03 Canopy 0.7 0.104 2.25 Slope 0.6 0.124 2.04 SCP 0.4 0.288 1.19 SP 0.3 0.434 0.84 SN 0.1 0.638 0.37 根 Root SCN 19.2 0.002 39.26 MAT 5.2 0.002 12.31 Altitude 4.2 0.002 8.98 MAP 3.2 0.002 7.21 SCP 3.1 0.002 7.16 Canopy 1.3 0.046 3.14 Slope 1.2 0.034 3.14 SP 0.6 0.252 1.41 SN 0.3 0.446 0.88 SC 0.3 0.400 0.85 SNP 0.2 0.538 0.63
MAP, mean annual precipitation; MAT, mean annual air temperature; SC, soil carbon content; SN, soil nitrogen content; SP, soil phosphorus content; SCN, soil C:N; SCP, soil C:P; SNP, soil N:P.Altitude, 海拔; Canopy, 郁闭度; Slope, 坡度; MAP, 年降水量; MAT, 年平均气温; SC, 土壤碳含量; SN, 土壤氮含量; SP, 土壤磷含量; SCN, 土壤C:N; SCP, 土壤C:P; SNP, 土壤N:P。新窗口打开 |
下载CSV 3 讨论 3.1 林下灌木植被C、N、P含量及化学计量比特征 本研究中, 灌木叶片C含量平均值为449.00 mg·g
-1 , 低于全球陆生植物叶片C含量464.20 mg·g
-1 (
Elser et al ., 2000 )和中国东部南北样带植物叶片C含量480.10 mg·g
-1 (
任书杰等, 2012 ), 说明青海森林的林下灌木植被C储存能力相对较弱。灌木叶片N、P含量平均值分别为19.46、1.69 mg·g
-1 , 略低于全球陆生植物N、P含量(20.10、1.77 mg·g
-1 (
Reich & Oleksyn, 2004 )和20.60、1.99 mg·g
-1 (
Elser et al ., 2000 )), 略低于我国植物叶片N含量(20.24 mg·g
-1 ), 高于其叶片P含量(1.46 mg·g
-1 )(
Han et al ., 2005 ), 高于我国南方优势灌丛优势植被N、P含量(16.57、1.02 mg·g
-1 )(
李家湘等, 2017 )、陕西森林的林下灌木叶片N、P含量(13.79、1.11 mg·g
-1 )(
姜沛沛等, 2016 ), 说明青海森林的林下灌木叶片N、P含量较高。这可能与本研究区域地处高寒区, 纬度偏高, 年降水量少, 年均气温低, 昼夜温差大, 日照强度大等特殊的区域气候条件有一定的关系(
卢航等, 2013 )。温度-植物生理假说认为, 低温诱导植物生长周期缩短, 同时, 植物趋向于分配更多的营养元素于光合器官中, 通过丰富叶片N、P含量在有限的生长周期内保持高效的光合速率以及酶活性来维持植物机体内正常的新陈代谢活动(
Reich & Oleksyn, 2004 ), 因此高海拔地区植物叶片N、P含量较为丰富。这与
杨阔等(2010) 发现青藏高原草地植物群落叶片N、P含量较高的结果一致。
生态化学计量学“内稳态理论”认为, 在长期的进化过程中, 随外界环境的变化, 生物有机体能保持其化学组成的相对恒定性(
Sterner & Elser, 2002 )。本研究发现, 7种林分间灌木(叶、枝干、根) P含量、C:P均没有明显差异性, 该结果符合“内稳态理论”, 因为7种林分间土壤TP含量相对稳定。山杨林、圆柏林、云杉林的林下灌木(叶、枝干、根) N含量、N:P高于白桦林、毛白杨林、红桦林和青扦林, C:N则相反, 这可能是由土壤TN含量、土壤C:N的显著差异造成的。相关性分析表明, 林下灌木(叶、枝干、根) N含量、N:P、C:N受土壤TN含量、土壤C:N的极显著影响(
p < 0.01)(
表2 )。冗余分析也表明(
图4 ;
表3 ), 土壤C:N极显著影响林下灌木(叶、枝干、根) C、N、P化学计量特征(
p < 0.01), 即灌木C、N、P含量受土壤有效性N的重要调控。在7种林分中, 山杨林、圆柏林和云杉林土壤C:N最低(
表1 ), 说明土壤有效性N较高。因此, 通过林下灌木层植被根系对土壤N的吸收利用, 各器官呈现出较高N含量, 较低C:N的特征。
植物叶片N:P通常是判断植物养分限制状况的指标之一。研究表明, 当N:P < 14时, 植物生长受N限制; 当N:P > 16时, 植物生长主要受P限制; 当N:P介于14-16之间时, 植物生长受N和P的共同限制或者不受养分限制(
Koerselman, 1996 ;
Tessier & Raynal, 2003 )。本研究中, 圆柏林灌木叶片N:P (16.72) > 16, 其余6种林分叶片N:P表现为青扦林(6.22) <红桦林(7.21) <毛白杨林(7.47) <白桦林(7.48) <云杉林(12.6) <山杨林(13.7) < 14。圆柏林的林下灌木植被生长受P限制, 其余6种林分林下灌木植被生长受到N限制。这可能是由于林分差异导致其土壤N、P供应能力的不同。本研究中, 圆柏林的林下灌木叶片P含量最低(
图2 ), 圆柏林土壤TN含量最高、C:N最低、C:P较高(
表1 ), 圆柏林的林下土壤N含量丰富, N有效性较高, 而P有效性不足, 因此, 林下灌木植被在N供应较为充沛的条件下, 其生长更趋向于受P限制。其余6种林分林下灌木植被生长限制情况与我国北方森林植被普遍受到N限制的结果相一致(
Han et al ., 2005 )。
3.2 森林土壤C、N、P含量及化学计量比特征 本研究区森林SOC、TN、TP含量平均值为(95.11、5.17、0.82 mg·g
-1 ), 高于全国土壤C、N、P含量(24.56、1.88、0.78 mg·g
-1 )(
Tian et al ., 2010 ), 与青藏高原高寒植被带土壤N、P含量(5.66、0.75 mg·g
-1 )相接近(
卢同平等, 2017 )。土壤C:N、C:P (20.6、117.7)高于全球森林土壤C:N、C:P (12.4、81.9)(
Cleveland & Liptzin, 2007 )和全国土壤C:N、C:P (12.3、52.7)(
Tian et al ., 2010 )。研究表明, 土壤C:N、C:P是衡量土壤有机质分解矿化速率的重要指标, 通常土壤C:N、C:P与有机质分解矿化速率呈反比关系, 较高的C:N、C:P代表较慢的有机质分解速率, 以及较低的土壤N、P有效性(
Cleveland & Liptzin, 2007 )。由此可推断, 青海高寒区森林SOC、TN、TP储备丰富, 有机质分解和矿化速率较慢, N、P有效性较低。这可能与本研究区域自然环境和植被类型特性有关, 青海高寒地区年降水量偏少、年平均气温低, 森林地表积累了丰富的凋落物, 长期的腐殖化作用有利于土壤有机质的富集, 较低的土壤温度抑制了土壤微生物及土壤酶活性, 地表有机质的分解周期长, 系统累积量大于消耗量, 导致森林土壤N、P元素得以累积, 但缓慢的分解速率导致土壤N、P有效性较低(
高巧等, 2014 ;
王淑彬等, 2014 )。
本研究中, 7种林分间SOC和TN含量呈现出明显差异, 而TP含量则差异不明显。这可能是由于受到营养元素的积累方式以及林分类型的影响, 林下土壤养分状况差异较大。在自然森林生态系统中, 森林土壤有机碳来自地表凋落物以及根系凋亡分解的积累, 而土壤全氮主要通过有机质分解矿化的途径累积(
李丹维等, 2017 )。首先, 林分的差异会影响地表凋落物、地下根系以及根系分泌物的组成, 同时凋落物的类型、数量以及分解速率是影响森林土壤养分含量变化和实现系统养分循环的重要因素(
杨佳佳等, 2014 )。其次, 林分的差异会影响土壤微生物、土壤动物和土壤酶的组成与功能多样性(
庞圣江等, 2015 ;
字洪标等, 2017 ), 而在土壤生物化学循环过程中, 土壤微生物通过氨化、硝化和反硝化等作用将土壤内颗粒态有机氮转化成可溶性有机氮, 而土壤酶作为催化剂也参与了其中复杂的生化反应(
李喜霞等, 2018 )。土壤P作为一种沉积性元素, 主要来源于岩石母质的风化, 其过程稳定而漫长, 因而在自然生态系统中P迁移能力较弱, 空间异质性较小(
李红林等, 2015 )。因此不同林分之间TP含量差异不明显。
3.3 灌木C、N、P含量及化学计量比特征与土壤组分的相关性和环境因子分析 相关性分析发现, 林下灌木(叶、枝干、根) N含量、C:N、N:P与土壤TN含量、C:N、N:P呈极显著相关关系, 而P含量、C:P与土壤TP含量呈显著相关关系, 表明植物体内各器官元素分配时相互协调, 且N、P吸收效率与土壤组分紧密相关。这是因为植物通过叶片凋落物和根系凋亡形成土壤有机质, 土壤有机质分解矿化为植物正常代谢活动提供必要的营养元素(
毕建华等, 2017 )。植物根系吸收和运输作为土壤N、P输出的主要途径, 使得植物与土壤在养分需求和供应之间达到动态平衡, 形成生态系统N、P转化的有效循环过程(
李婷等, 2015 )。
依据RDA发现, 林下灌木C、N、P含量及化学计量特征主要受土壤C:N、海拔、年降水量和年平均气温的影响, 这表明青海高寒区森林的林下灌木受到土壤N有效性、海拔以及区域水热条件的重要调控。土壤作为植被获取营养元素的主要机质, 其N、P有效性将显著影响地上植被组成与结构, 生产力以及有机体内元素分配格局(
李丹维等, 2017 )。
McGroddy等(2004) 认为, 土壤N、P有效性是植物有机体内元素含量的主要驱动因素, 同时土壤N、P有效性容易受降水淋溶作用的影响。
张仁懿等(2014) 研究也表明青藏高原亚高寒草甸植物养分状况受土壤N有效性的显著影响, 土壤N有效性对高寒植物生长的影响比其他元素更为关键。海拔梯度综合了温度、降水、光照等水热条件的变化, 是影响植被生长的主导因子(
Guo et al ., 2010 )。本研究结果与木本植物叶片N、P元素含量受年平均气温和年降水量共同驱动作用的结果(
Chen et al ., 2013b )相符。因为:(1)海拔梯度上降水、气温的差异会影响植物的生长周期, 从而改变C、N、P在各器官间的分配(
Kerkhoff et al ., 2005 ); (2)海拔梯度上土壤温度和水分的变化将影响土壤微生物的群系特征及代谢效率, 间接影响土壤有机质的分解和矿化速率, 而且土壤母质的风化速率以及N、P元素的淋溶强度也受降水和气温等气候因子的显著影响(
刘颖等, 2018 ); (3)不同的海拔梯度上森林植被受到的人为干扰程度也有一定的差异, 放牧、旅游、耕种等活动也会影响植物的元素分配格局(
Nogués-Bravo et al ., 2008 )。相关研究表明, 伴随着全球气候变化, 青藏高原地区已出现气温剧烈升高, 降水量增加的暖湿化趋势(
Chen et al ., 2013a )。因此, 未来在全球气候变化和区域N沉降可能加剧的背景下, 青海森林生态系统灌木层C、N、P化学计量特征将受到强烈影响。
4 结论 青海森林生态系统中灌木P含量、C:P和土壤TP含量受林分差异的影响较小, 而灌木N含量、N:P、C:N和SOC、土壤TN含量受林分差异的影响较大。圆柏林的林下灌木层植被生长受P限制, 其余6种林分林下灌木层植被生长受到N限制。林下灌木C、N、P含量及化学计量比与土壤组分紧密关联。土壤C:N、海拔、年平均气温和年降水量是林下灌木层植被C、N、P含量及生态化学计量变化的主要影响因子。
致谢 感谢西南民族大学研究生“创新型科研项目”重点项目(CX2018SZ106)资助。
附录Ⅰ 青海不同林分林下灌木植被基本信息 Supplement Ⅰ Basic information of shrub vegetation under different forest types in Qinghai http://www.plant-ecology.com/fileup/PDF/cjpe.2018.0326-S1.pdf
[1] Bi JH Su BL Yu DP Wu J Cao LL Dai LM Zhou L 2017 ). Ecological stoichiometry of different forest types in mountainous region of eastern Liaoning ProvinceChinese Journal of Ecology , 36, 117 -123 . [本文引用: 1] [ 毕建华 , 苏宝玲 , 于大炮 , 吴健 , 曹琳琳 , 代力民 , 周莉 ( 2017 ). 辽东山区不同森林类型生态化学计量特征生态学杂志 , 36, 117 -123 .] [本文引用: 1] [2] Chen H Zhu Q Peng CH Wu N Wang YF Fang XQ Gao YH Zhu D Yang G Tian JQ Kang XM Piao SL Ouyang H Xiang WH Luo ZB Jiang H Song XZ Zhang Y Yu GR Zhao XQ Gong P Yao TD Wu JH 2013 a). The impacts of climate change and human activities on biogeochemical cycles on the Qinghai-Tibetan PlateauGlobal Change Biology , 19, 2940 -2955 . [本文引用: 1] [3] Chen XP Guo BQ Zhong QL Wang MT Li M Yang FC Cheng DL 2018 ). Response of fine root carbon, nitrogen, and phosphorus stoichiometry to soil nutrients in Pinus taiwanensis along an elevation gradient in the Wuyi MountainsActa Ecologica Sinica 234 -281 . [本文引用: 1] [ 陈晓萍 , 郭炳桥 , 钟全林 , 王满堂 , 李曼 , 杨福春 , 程栋梁 ( 2018 ). 武夷山不同海拔黄山松细根碳、氮、磷化学计量特征对土壤养分的适应生态学报 , 38, 234 -281 .] [本文引用: 1] [4] Chen YH Han WX Tang LY Tang ZY Fang JY 2013 b). Leaf nitrogen and phosphorus concentrations of woody plants differ in responses to climate, soil and plant growth formEcography , 36, 178 -184 . [本文引用: 1] [5] Cleveland CC Liptzin D 2007 ). C:N:P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomass?Biochemistry , 85, 235 -252 . [本文引用: 2] [6] Compilation Group of Technical Specifications for Ecosystem Carbon Sequestration Project ( 2015 ). Technical Specifications for Observation and Investigation of Ecosystem Carbon Sequestration . Science Press, Beijing. [本文引用: 1] [ 生态系统固碳项目技术规范编写组 ( 2015 ). 生态系统固碳观测与调查技术规范 . 科学出版社, 北京.] [本文引用: 1] [7] Cui NJ Chen XH Liu Y Zhang J Yang WQ 2014 ). Shrub and herb diversity at different ages of Pinus massoniana plantationActa Ecologica Sinica 4313 -4323 . [本文引用: 1] [ 崔宁洁 , 陈小红 , 刘洋 , 张健 , 杨万勤 ( 2014 ). 不同林龄马尾松人工林林下灌木和草本多样性生态学报 , 34, 4313 -4323 .] [本文引用: 1] [8] Dong X 2009 ). Evaluation of forest resources in Qinghai ProvinceJournal of Anhui Agricultural Sciences , 37, 5727 -5728 . [本文引用: 1] [ 董旭 ( 2009 ). 青海省森林资源评价安徽农业科学 , 37, 5727 -5728 .] [本文引用: 1] [9] Elser JJ Bracken MES Cleland EE Gruner DS Harpole WS Hillebrand H 2007 ). Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystemsEcology Letters , 10, 1135 -1142 . [本文引用: 1] [10] Elser JJ Dobberfuhl DR Mackay NA 1996 ). Organism size, life history, and N:P stoichiometryBioscience , 46, 674 -684 . [本文引用: 1] [11] Elser JJ Fagan WF Denno RF 2000 ). Nutritional constraints in terrestrial and freshwater food websNature , 408, 578 -580 . [本文引用: 2] [12] Gao Q Yang XC Yi CY Liu Q 2014 ). Estimation of biomass allocation and carbon density in alpine dwarf shrubs in Garzê Zangzu Autonomous Prefecture of Sichuan Province, ChinaChinese Journal of Plant Ecology , 38, 355 -365 . [本文引用: 1] [ 高巧 , 阳小成 , 尹春英 , 刘庆 ( 2014 ). 四川省甘孜藏族自治州高寒矮灌丛生物量分配及其碳密度的估算植物生态学报 , 38, 355 -365 .] [本文引用: 1] [13] Guo H Mazer SJ Du G 2010 ). Geographic variation in primary sex allocation per flower within and among 12 species of Pedicularis(Orobanchaceae): Proportional male investment increases with elevationAmerican Journal of Botany 1334 -1341 . [本文引用: 1] [14] Güsewell S 2004 ). N:P ratios in terrestrial plants: Variation and functional significanceNew Phytologist , 164, 243 -266 . [本文引用: 1] [15] Han WX Fang JY Guo DL Zhang Y 2005 ). Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in ChinaNew Phytologist , 168, 377 -385 . [本文引用: 3] [16] He HL Yang XC Li DD Yin CY Li YX Zhou GY Zhang L Liu Q 2017 ). Stoichiometric characteristics of carbon, nitrogen and phosphorus of Sibiraea angustata shrub on the Eastern Qinghai-Xizang PlateauChinese Journal of Plant Ecology 126 -135 . [本文引用: 1] [ 贺合亮 , 阳小成 , 李丹丹 , 尹春英 , 黎云祥 , 周国英 , 张林 , 刘庆 ( 2017 ). 青藏高原东部窄叶鲜卑花碳、氮、磷化学计量特征植物生态学报 , 41, 126 -135 .] [本文引用: 1] [17] He JS Han XG 2010 ). Ecological stoichiometry: Searching for unifying principles from individuals to ecosystemsChinese Journal of Plant Ecology , 34, 2 -6 . [本文引用: 1] [ 贺金生 , 韩兴国 ( 2010 ). 生态化学计量学: 探索从个体到生态系统的统一化理论植物生态学报 , 34, 2 -6 .] [本文引用: 1] [18] Hu L Wang CT Wang GX Liu W Ade LJ 2015 ). Carbon sequestration of forest ecosystem vegetation in Qinghai ProvinceSouthwest China Journal of Agricultural Sciences , 28, 826 -832 . [本文引用: 1] [ 胡雷 , 王长庭 , 王根绪 , 刘伟 , 阿的鲁骥 ( 2015 ). 青海省森林生态系统植被固碳现状研究西南农业学报 , 28, 826 -832 .] [本文引用: 1] [19] Huang XB Liu WD Su JR Li SF Lang XD 2016 ). Stoichiometry of leaf C, N and P across 152 woody species of a monsoon broad-leaved evergreen forest in Pu’er, Yunnan ProvinceChinese Journal of Ecology , 35, 567 -575 . [本文引用: 1] [ 黄小波 , 刘万德 , 苏建荣 , 李帅锋 , 郎学东 ( 2016 ). 云南普洱季风常绿阔叶林152种木本植物叶片C、N、P化学计量特征生态学杂志 , 35, 567 -575 .] [本文引用: 1] [20] Iversen CM Mccormack ML Powell AS Blackwood CB Freschet GT Jens K Catherine R Stover DB Cyrille V 2017 ). A global fine-root ecology database to address below-ground challenges in plant ecologyNew Phytologist , 215, 15 -26 . [本文引用: 1] [21] Jia JD Wu YQ Zhang DS Kang MY Shi PJ 2004 ). Environmental changes and spatial distribution of ecological conservation in Qinghai ProvinceResources Science , 26, 9 -16 . [本文引用: 1] [ 贾敬敦 , 伍永秋 , 张登山 , 康慕谊 , 史培军 ( 2004 ). 青海生态环境变化与生态建设的空间布局资源科学 , 26, 9 -16 .] [本文引用: 1] [22] Jiang PP Cao Y Chen YM 2016 ). C, N, P stoichiometric characteristics of tree, shrub, herb leaves and litter in forest community of Shaanxi Province, ChinaChinese Journal of Applied Ecology , 27, 365 -372 . [本文引用: 1] [ 姜沛沛 , 曹扬 , 陈云明 ( 2016 ). 陕西省森林群落乔灌草叶片和凋落物C、N、P生态化学计量特征应用生态学报 , 27, 365 -372 .] [本文引用: 1] [23] Kang YX Yue JW Lei RD Liang ZS Chen YJ 2008 ). Dominant species niche of Quercus liaotungensis communities in Huanglong Mt. of ShaanxiActa Botanica Boreali- Occidentalia Sinica 574 -581 . [本文引用: 1] [ 康永祥 , 岳军伟 , 雷瑞德 , 梁宗锁 , 陈永剑 ( 2008 ). 陕北黄龙山辽东栎群落优势种群生态位研究西北植物学报 , 28, 574 -581 .] [本文引用: 1] [24] Kerkhoff AJ Enquist BJ Elser JJ Fagan WF 2005 ). Plant allometry, stoichiometry and the temperature-dependence of primary productivityGlobal Ecology and Biogeography , 14, 585 -598 . [本文引用: 1] [25] Koerselman W 1996 ). The vegetation N:P ratio: A new tool to detect the nature of nutrient limitationJournal of Applied Ecology , 33, 1441 -1450 . [本文引用: 1] [26] Li DF Yu SL Wang GX Fang WW 2015 ). Environmental heterogeneity and mechanism of stoichiometry properties of vegetative organs in dominant shrub communities across the Loess PlateauChinese Journal of Plant Ecology , 39, 453 -465 . [本文引用: 1] [ 李单凤 , 于顺利 , 王国勋 , 方伟伟 ( 2015 ). 黄土高原优势灌丛营养器官化学计量特征的环境分异和机制植物生态学报 , 39, 453 -465 .] [本文引用: 1] [27] Li DW Wang ZQ Tian HX He WX Geng ZC 2017 ). Carbon, nitrogen and phosphorus contents in soils on Taibai Mountain and their ecological stoichiometry relative to elevationActa Pedologica Sinica , 54, 160 -170 . [本文引用: 2] [ 李丹维 , 王紫泉 , 田海霞 , 和文祥 , 耿增超 ( 2017 ). 太白山不同海拔土壤碳、氮、磷含量及生态化学计量特征土壤学报 , 54, 160 -170 .] [本文引用: 2] [28] Li HL Gong L Zhu ML Liu ZY Xie LN Hong Y 2015 ). Stoichiometric characteristics of soil in an oasis on northern edge of Tarim Basin, ChinaActa Pedologica Sinica , 52, 1345 -1355 . [本文引用: 1] [ 李红林 , 贡璐 , 朱美玲 , 刘曾媛 , 解丽娜 , 洪毅 ( 2015 ). 塔里木盆地北缘绿洲土壤化学计量特征土壤学报 , 52, 1345 -1355 .] [本文引用: 1] [29] Li JX Xu WT Xiong GM Wang Y Zhang CM Lu ZJ Li YL Xie ZQ 2017 ). Leaf nitrogen and phosphorus concentration and the empirical regulations in dominant woody plants of shrublands across southern ChinaChinese Journal of Plant Ecology , 41, 31 -42 . [本文引用: 1] [ 李家湘 , 徐文婷 , 熊高明 , 王杨 , 赵常明 , 卢志军 , 李跃林 , 谢宗强 ( 2017 ). 中国南方灌丛优势木本植物叶的氮、磷含量及其影响因素植物生态学报 , 41, 31 -42 .] [本文引用: 1] [30] Li QH Jiang ZP Zhang JB Zhao YM 2006 ). The progress and perspective of research on the ecological characteristics and benefits of shrubJournal of Arid Land Resources and Environment , 20, 159 -164 . [本文引用: 1] [ 李清河 , 江泽平 , 张景波 , 赵英铭 ( 2006 ). 灌木的生态特性与生态效能的研究与进展干旱区资源与环境 , 20, 159 -164 .] [本文引用: 1] [31] Li T Deng Q Yuan ZY Jiao F 2015 ). Latitudinal changes in plant stoichiometric and soil C, N, P stoichiometry in Loess PlateauEnvironmental Science , 36, 2988 -2996 . [本文引用: 2] [ 李婷 , 邓强 , 袁志友 , 焦峰 ( 2015 ). 黄土高原纬度梯度上的植物与土壤碳、氮、磷化学计量学特征环境科学 , 36, 2988 -2996 .] [本文引用: 2] [32] Li XX Du TY Wei YW Zhou YB 2018 ). Characteristics of ecological stoichiometry in broad-leaved and Korean pine mixed forest and its response to latitude gradient in Northeast ChinaActa Ecologica Sinica 3952 -3960 . [本文引用: 1] [ 李喜霞 , 杜天雨 , 魏亚伟 , 周永斌 ( 2018 ). 阔叶红松林生态化学计量学特征及其对纬度梯度的响应生态学报 , 38, 3952 -3960 .] [本文引用: 1] [33] Liu Y Gong YB Li Y Zhu DW Liu H Shuai W 2018 ). Soil stoichiometric characteristics of alpine shrub meadow at different elevations, Western SichuanJournal of Sichuan Agricultural University , 36, 167 -174 . [本文引用: 1] [ 刘颖 , 宫渊波 , 李瑶 , 朱德雯 , 刘韩 , 帅伟 ( 2018 ). 川西高寒灌丛草地不同海拔梯度土壤化学计量特征四川农业大学学报 , 36, 167 -174 .] [本文引用: 1] [34] Lu H Liu K Wu JH 2013 ). Change of carbon storage in forest vegetation and current situation analysis of Qinghai Province in recent 20 yearsResources and Environment in the Yangtze Basin , 22, 1333 -1338 . [本文引用: 1] [ 卢航 , 刘康 , 吴金鸿 ( 2013 ). 青海省近20年森林植被碳储量变化及其现状分析长江流域资源与环境 , 22, 1333 -1338 .] [本文引用: 1] [35] Lu TP Zhang WX Niu J Lin YJ Wu MJ 2017 ). Study on spatial variability and driving factors of stoichiometry of nitrogen and phosphorus in soils of typical natural zones of ChinaActa Pedologica Sinica , 54, 682 -692 . [本文引用: 1] [ 卢同平 , 张文翔 , 牛洁 , 林永静 , 武梦娟 ( 2017 ). 典型自然带土壤氮磷化学计量空间分异特征及其驱动因素研究土壤学报 , 54, 682 -692 .] [本文引用: 1] [36] Lu ZL Gong XS 2009 ). Progress on the research of shrub biomass estimationForest Inventory and Planning , 34(4 ), 37 -40 . [本文引用: 1] [ 卢振龙 , 龚孝生 ( 2009 ). 灌木生物量测定的研究进展林业调查规划 , 34(4 ), 37 -40 .] [本文引用: 1] [37] McGroddy ME Daufresne T Hedin LO 2004 ). Scaling of C:N:P stoichiometry in forest worldwide: Implications of terrestrial Redfield-type ratiosEcology , 85, 2390 -2401 . [本文引用: 1] [38] Nogués-Bravo D Araújo MB Romdal T Rahbek C 2008 ). Scale effects and human impact on the elevational species richness gradientsNature , 453, 216 -219 . [本文引用: 1] [39] Pang SJ Zhang P Jia HY Yang BG Deng SK Feng CL 2015 ). Research on soil ecological stoichiometry under different forest types in Northwest GuangxiChinese Agricultural Science Bulletin , 31, 17 -23 . [本文引用: 1] [ 庞圣江 , 张培 , 贾宏炎 , 杨保国 , 邓硕坤 , 冯昌林 ( 2015 ). 桂西北不同森林类型土壤生态化学计量特征中国农学通报 , 31, 17 -23 .] [本文引用: 1] [40] Qi DH Wen ZM Wang HX Guo R Yang SS 2016 ). Stoichiometry traits of carbon, nitrogen, and phosphorus in plants of different functional groups and their responses to micro-topographical variations in the hilly and gully region of the Loess Plateau, ChinaActa Ecologica Sinica , 36, 6420 -6430 . [本文引用: 1] [ 戚德辉 , 温仲明 , 王红霞 , 郭茹 , 杨士梭 ( 2016 ). 黄土丘陵区不同功能群植物碳氮磷生态化学计量特征及其对微地形的响应生态学报 , 36, 6420 -6430 .] [本文引用: 1] [41] Reich PB Oleksyn J 2004 ). Global patterns of plant leaf N and P in relation to temperature and latitudeProceedings of the National Academy of Sciences of the United States of America , 101, 11001 -11006 . [本文引用: 2] [42] Ren SJ Yu GR Jiang CM Fang HJ Sun XM 2012 ). Stoichiometric characteristics of leaf carbon, nitrogen, and phosphorus of 102 dominant species in forest ecosystems along the North-South Transect of East ChinaChinese Journal of Applied Ecology , 23, 581 -586 . [本文引用: 1] [ 任书杰 , 于贵瑞 , 姜春明 , 方华军 , 孙晓敏 ( 2012 ). 中国东部南北样带森林生态系统102个优势种叶片碳氮磷化学计量学统计特征应用生态学报 , 23, 581 -586 .] [本文引用: 1] [43] Sterner RW Elser JJ 2002 ). Ecological Stoichiometry: The Biology of Elements from Molecules to Biosphere . Princeton University Press, Princeton. [本文引用: 1] [44] Tang ZY Xu WT Zhou GS Bai YF Xie ZQ 2018 ). Patterns of plant carbon, nitrogen, and phosphorus concentration in relation to productivity in China’s terrestrial ecosystemsProceedings of the National Academy of Sciences of the United States of America , 115, 4033 -4038 . [本文引用: 1] [45] Tessier JT Raynal DJ 2003 ). Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturationJournal of Applied Ecology , 40, 523 -534 . [本文引用: 1] [46] Tian HQ Chen GS Zhang C 2010 ). Pattern and variation of C:N:P ratios in China?s soils: A synthesis of observational dataBiogeochemistry , 98, 139 -151 . [本文引用: 2] [47] Wan WX Wang XK Li DY Feng XQ Zhang QQ Yu XH 2014 ). Biomass allometric models for understory shrubs of warm temperate forest ecosystemActa Ecologica Sinica , 34, 6985 -6992 . [本文引用: 1] [ 万五星 , 王效科 , 李东义 , 冯学全 , 张千千 , 于小红 ( 2014 ). 暖温带森林生态系统林下灌木生物量相对生长模型生态学报 , 34, 6985 -6992 .] [本文引用: 1] [48] Wang GX Li YS Wu QB Wang YB 2006 ). Relationship between permafrost and vegetation in the Qinghai-Tibet Plateau tundra and its impact on the alpine ecosystemScience in China: Earth Science , 36, 743 -754 . [本文引用: 1] [ 王根绪 , 李元首 , 吴青柏 , 王一博 ( 2006 ). 青藏高原冻土区冻土与植被的关系及其对高寒生态系统的影响中国科学:地球科学 , 36, 743 -754 .] [本文引用: 1] [49] Wang SB Xu HF Song TQ Huang GQ Peng WX Du H 2014 ). Spatial heterogeneity of the main nutrients in Guangxi forest soilsActa Ecologica Sinica , 34, 5292 -5299 . [本文引用: 1] [ 王淑彬 , 徐慧芳 , 宋同清 , 黄国勤 , 彭晚霞 , 杜虎 ( 2014 ). 广西森林土壤主要养分的空间异质性生态学报 , 34, 5292 -5299 .] [本文引用: 1] [50] Yang HM Wang DM 2011 ). Advances in the study on ecological stoichiometry in grass-environment system and its response to environmental factorsActa Prataculturae Sinica , 5, 247 -264 . [本文引用: 1] [51] Yang JJ Zhang XR Chen YN Ma LS An SS Dang TH 2014 ). Ecological stoichiometric relationships between components of Robinia pseudoacacia forest in Loess PlateauActa Pedologica Sinica 133 -142 . [本文引用: 1] [ 杨佳佳 , 张向茹 , 陈亚南 , 马露莎 , 安韶山 , 党廷辉 ( 2014 ). 黄土高原刺槐林不同组分生态化学计量关系研究土壤学报 , 51, 133 -142 .] [本文引用: 1] [52] Yang K Huang JH Dong D Ma WH HE JS 2010 ). Canopy leaf N and P stoichiometry in grassland communities of Qinghai-Tibetan Plateau, ChinaChinese Journal of Plant Ecology , 34, 17 -22 . [本文引用: 1] [ 杨阔 , 黄建辉 , 董丹 , 马文红 , 贺金生 ( 2010 ). 青藏高原草地植物群落冠层叶片氮磷化学计量学分析植物生态学报 , 34, 17 -22 .] [本文引用: 1] [53] Zhang RY Xu DH Chen LY Wang G 2014 ). Plant N status in the alpine grassland of the Qinghai-Tibet Plateau: Base on the N:P stoichiometryEnvironmental Science , 35, 1131 -1137 . [本文引用: 1] [ 张仁懿 , 徐当会 , 陈凌云 , 王刚 ( 2014 ). 基于N:P化学计量特征的高寒草甸植物养分状况研究环境科学 , 35, 1131 -1137 .] [本文引用: 1] [54] Zhang SB 2005 ). On the Ecological Status of Qinghai Key Public Welfare Forests The First National Forestry Academic Conference, Hangzhou . 383 -387 . [本文引用: 1] [ 张胜邦 ( 2005 ). 论青海国家重点公益林的生态地位首届全国林业学术大会, 杭州 . 383 -387 .] [本文引用: 1] [55] Zheng SW Tang M Zou JH Mu CL 2007 ). Summary of research on shrub biomass in ChinaJournal of Chengdu University (Natural Science Edition )189 -192 . [本文引用: 1] [ 郑绍伟 , 唐敏 , 邹俊辉 , 慕长龙 ( 2007 ). 灌木群落及生物量研究综述成都大学学报(自然科学版) , 26, 189 -192 .] [本文引用: 1] [56] Zi HB Xiang ZY Wang GX Ade LJ Wang CT 2017 ). Profile of soil microbial community under different stand types in Qinghai ProvinceScientia Silvae Sinicae , 53(3 ), 21 -32 . [本文引用: 2] [ 字洪标 , 向泽宇 , 王根绪 , 阿的鲁骥 , 王长庭 ( 2017 ). 青海不同林分土壤微生物群落结构(PLFA)林业科学 , 53(3 ), 21 -32 .] [本文引用: 2] [57] Zuo W He KN Tian Y Wang WL 2016 ). Surface litter stoichiometry for five forest types in alpine region, Qinghai, ChinaChinese Journal of Ecology , 35, 2271 -2278 . [本文引用: 1] [ 左巍 , 贺康宁 , 田赟 , 王玮璐 ( 2016 ). 青海高寒区不同林分类型凋落物养分状况及化学计量特征生态学杂志 , 35, 2271 -2278 .] [本文引用: 1] 辽东山区不同森林类型生态化学计量特征
1
2017
... 相关性分析发现, 林下灌木(叶、枝干、根) N含量、C:N、N:P与土壤TN含量、C:N、N:P呈极显著相关关系, 而P含量、C:P与土壤TP含量呈显著相关关系, 表明植物体内各器官元素分配时相互协调, 且N、P吸收效率与土壤组分紧密相关.这是因为植物通过叶片凋落物和根系凋亡形成土壤有机质, 土壤有机质分解矿化为植物正常代谢活动提供必要的营养元素(
毕建华等, 2017 ).植物根系吸收和运输作为土壤N、P输出的主要途径, 使得植物与土壤在养分需求和供应之间达到动态平衡, 形成生态系统N、P转化的有效循环过程(
李婷等, 2015 ). ...
辽东山区不同森林类型生态化学计量特征
1
2017
... 相关性分析发现, 林下灌木(叶、枝干、根) N含量、C:N、N:P与土壤TN含量、C:N、N:P呈极显著相关关系, 而P含量、C:P与土壤TP含量呈显著相关关系, 表明植物体内各器官元素分配时相互协调, 且N、P吸收效率与土壤组分紧密相关.这是因为植物通过叶片凋落物和根系凋亡形成土壤有机质, 土壤有机质分解矿化为植物正常代谢活动提供必要的营养元素(
毕建华等, 2017 ).植物根系吸收和运输作为土壤N、P输出的主要途径, 使得植物与土壤在养分需求和供应之间达到动态平衡, 形成生态系统N、P转化的有效循环过程(
李婷等, 2015 ). ...
The impacts of climate change and human activities on biogeochemical cycles on the Qinghai-Tibetan Plateau
1
2013
... 依据RDA发现, 林下灌木C、N、P含量及化学计量特征主要受土壤C:N、海拔、年降水量和年平均气温的影响, 这表明青海高寒区森林的林下灌木受到土壤N有效性、海拔以及区域水热条件的重要调控.土壤作为植被获取营养元素的主要机质, 其N、P有效性将显著影响地上植被组成与结构, 生产力以及有机体内元素分配格局(
李丹维等, 2017 ).
McGroddy等(2004) 认为, 土壤N、P有效性是植物有机体内元素含量的主要驱动因素, 同时土壤N、P有效性容易受降水淋溶作用的影响.
张仁懿等(2014) 研究也表明青藏高原亚高寒草甸植物养分状况受土壤N有效性的显著影响, 土壤N有效性对高寒植物生长的影响比其他元素更为关键.海拔梯度综合了温度、降水、光照等水热条件的变化, 是影响植被生长的主导因子(
Guo et al ., 2010 ).本研究结果与木本植物叶片N、P元素含量受年平均气温和年降水量共同驱动作用的结果(
Chen et al ., 2013b )相符.因为:(1)海拔梯度上降水、气温的差异会影响植物的生长周期, 从而改变C、N、P在各器官间的分配(
Kerkhoff et al ., 2005 ); (2)海拔梯度上土壤温度和水分的变化将影响土壤微生物的群系特征及代谢效率, 间接影响土壤有机质的分解和矿化速率, 而且土壤母质的风化速率以及N、P元素的淋溶强度也受降水和气温等气候因子的显著影响(
刘颖等, 2018 ); (3)不同的海拔梯度上森林植被受到的人为干扰程度也有一定的差异, 放牧、旅游、耕种等活动也会影响植物的元素分配格局(
Nogués-Bravo et al ., 2008 ).相关研究表明, 伴随着全球气候变化, 青藏高原地区已出现气温剧烈升高, 降水量增加的暖湿化趋势(
Chen et al ., 2013a ).因此, 未来在全球气候变化和区域N沉降可能加剧的背景下, 青海森林生态系统灌木层C、N、P化学计量特征将受到强烈影响. ...
武夷山不同海拔黄山松细根碳、氮、磷化学计量特征对土壤养分的适应
1
2018
... 碳(C)、氮(N)、磷(P)作为最重要的生命元素, 是维系地球上所有生命活动的基础(
贺金生和韩兴国, 2010 ).C作为植物有机体结构的重要组成元素, 是植物有机体内生理生化反应过程的能量来源之一(
Güsewell, 2004 ), N、P是植物的基本营养元素, 也是植物有机体内蛋白质、酶类、核酸和脂质的重要组成元素(
Yang & Wang, 2011 ;
戚德辉等, 2016 ;
Tang et al ., 2018 ).C、N、P含量及其化学计量比的差异显著影响植物的生态策略, 从而被广泛应用到生态系统在生物地球化学循环和生态过程耦合关系的研究中(
Elser et al ., 1996 ).截至目前, 国内外****已在区域乃至全球范围开展了许多陆地生态系统生态化学计量特征的研究(
Han et al ., 2005 ;
Elser et al ., 2007 ;
黄小波等, 2016 ).但主要集中在植物叶片和土壤生态化学计量特征方面, 针对植物枝干、根等重要器官方面的研究相对较少(
贺合亮等, 2017 ).枝干是植物连接地下吸收器官和地上同化组织的传导器官, 具有支撑植物体的结构性骨架作用(
李单凤等, 2015 ).根系作为吸收、存储、运输水分和养分的主要器官, 也是植物有机体对外界环境变化响应最为敏感的地下器官, 在森林生态系统物质循环过程中发挥关键作用(
Iversen et al ., 2017 ;
陈晓萍等, 2018 ).因此, 研究植物不同器官间生态化学计量特征以及与土壤组分在各生态过程间的互作关系, 对于揭示生态系统养分循环规律具有重要意义. ...
武夷山不同海拔黄山松细根碳、氮、磷化学计量特征对土壤养分的适应
1
2018
... 碳(C)、氮(N)、磷(P)作为最重要的生命元素, 是维系地球上所有生命活动的基础(
贺金生和韩兴国, 2010 ).C作为植物有机体结构的重要组成元素, 是植物有机体内生理生化反应过程的能量来源之一(
Güsewell, 2004 ), N、P是植物的基本营养元素, 也是植物有机体内蛋白质、酶类、核酸和脂质的重要组成元素(
Yang & Wang, 2011 ;
戚德辉等, 2016 ;
Tang et al ., 2018 ).C、N、P含量及其化学计量比的差异显著影响植物的生态策略, 从而被广泛应用到生态系统在生物地球化学循环和生态过程耦合关系的研究中(
Elser et al ., 1996 ).截至目前, 国内外****已在区域乃至全球范围开展了许多陆地生态系统生态化学计量特征的研究(
Han et al ., 2005 ;
Elser et al ., 2007 ;
黄小波等, 2016 ).但主要集中在植物叶片和土壤生态化学计量特征方面, 针对植物枝干、根等重要器官方面的研究相对较少(
贺合亮等, 2017 ).枝干是植物连接地下吸收器官和地上同化组织的传导器官, 具有支撑植物体的结构性骨架作用(
李单凤等, 2015 ).根系作为吸收、存储、运输水分和养分的主要器官, 也是植物有机体对外界环境变化响应最为敏感的地下器官, 在森林生态系统物质循环过程中发挥关键作用(
Iversen et al ., 2017 ;
陈晓萍等, 2018 ).因此, 研究植物不同器官间生态化学计量特征以及与土壤组分在各生态过程间的互作关系, 对于揭示生态系统养分循环规律具有重要意义. ...
Leaf nitrogen and phosphorus concentrations of woody plants differ in responses to climate, soil and plant growth form
1
2013
... 依据RDA发现, 林下灌木C、N、P含量及化学计量特征主要受土壤C:N、海拔、年降水量和年平均气温的影响, 这表明青海高寒区森林的林下灌木受到土壤N有效性、海拔以及区域水热条件的重要调控.土壤作为植被获取营养元素的主要机质, 其N、P有效性将显著影响地上植被组成与结构, 生产力以及有机体内元素分配格局(
李丹维等, 2017 ).
McGroddy等(2004) 认为, 土壤N、P有效性是植物有机体内元素含量的主要驱动因素, 同时土壤N、P有效性容易受降水淋溶作用的影响.
张仁懿等(2014) 研究也表明青藏高原亚高寒草甸植物养分状况受土壤N有效性的显著影响, 土壤N有效性对高寒植物生长的影响比其他元素更为关键.海拔梯度综合了温度、降水、光照等水热条件的变化, 是影响植被生长的主导因子(
Guo et al ., 2010 ).本研究结果与木本植物叶片N、P元素含量受年平均气温和年降水量共同驱动作用的结果(
Chen et al ., 2013b )相符.因为:(1)海拔梯度上降水、气温的差异会影响植物的生长周期, 从而改变C、N、P在各器官间的分配(
Kerkhoff et al ., 2005 ); (2)海拔梯度上土壤温度和水分的变化将影响土壤微生物的群系特征及代谢效率, 间接影响土壤有机质的分解和矿化速率, 而且土壤母质的风化速率以及N、P元素的淋溶强度也受降水和气温等气候因子的显著影响(
刘颖等, 2018 ); (3)不同的海拔梯度上森林植被受到的人为干扰程度也有一定的差异, 放牧、旅游、耕种等活动也会影响植物的元素分配格局(
Nogués-Bravo et al ., 2008 ).相关研究表明, 伴随着全球气候变化, 青藏高原地区已出现气温剧烈升高, 降水量增加的暖湿化趋势(
Chen et al ., 2013a ).因此, 未来在全球气候变化和区域N沉降可能加剧的背景下, 青海森林生态系统灌木层C、N、P化学计量特征将受到强烈影响. ...
C:N:P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomass?
2
2007
... 本研究区森林SOC、TN、TP含量平均值为(95.11、5.17、0.82 mg·g
-1 ), 高于全国土壤C、N、P含量(24.56、1.88、0.78 mg·g
-1 )(
Tian et al ., 2010 ), 与青藏高原高寒植被带土壤N、P含量(5.66、0.75 mg·g
-1 )相接近(
卢同平等, 2017 ).土壤C:N、C:P (20.6、117.7)高于全球森林土壤C:N、C:P (12.4、81.9)(
Cleveland & Liptzin, 2007 )和全国土壤C:N、C:P (12.3、52.7)(
Tian et al ., 2010 ).研究表明, 土壤C:N、C:P是衡量土壤有机质分解矿化速率的重要指标, 通常土壤C:N、C:P与有机质分解矿化速率呈反比关系, 较高的C:N、C:P代表较慢的有机质分解速率, 以及较低的土壤N、P有效性(
Cleveland & Liptzin, 2007 ).由此可推断, 青海高寒区森林SOC、TN、TP储备丰富, 有机质分解和矿化速率较慢, N、P有效性较低.这可能与本研究区域自然环境和植被类型特性有关, 青海高寒地区年降水量偏少、年平均气温低, 森林地表积累了丰富的凋落物, 长期的腐殖化作用有利于土壤有机质的富集, 较低的土壤温度抑制了土壤微生物及土壤酶活性, 地表有机质的分解周期长, 系统累积量大于消耗量, 导致森林土壤N、P元素得以累积, 但缓慢的分解速率导致土壤N、P有效性较低(
高巧等, 2014 ;
王淑彬等, 2014 ). ...
... ).研究表明, 土壤C:N、C:P是衡量土壤有机质分解矿化速率的重要指标, 通常土壤C:N、C:P与有机质分解矿化速率呈反比关系, 较高的C:N、C:P代表较慢的有机质分解速率, 以及较低的土壤N、P有效性(
Cleveland & Liptzin, 2007 ).由此可推断, 青海高寒区森林SOC、TN、TP储备丰富, 有机质分解和矿化速率较慢, N、P有效性较低.这可能与本研究区域自然环境和植被类型特性有关, 青海高寒地区年降水量偏少、年平均气温低, 森林地表积累了丰富的凋落物, 长期的腐殖化作用有利于土壤有机质的富集, 较低的土壤温度抑制了土壤微生物及土壤酶活性, 地表有机质的分解周期长, 系统累积量大于消耗量, 导致森林土壤N、P元素得以累积, 但缓慢的分解速率导致土壤N、P有效性较低(
高巧等, 2014 ;
王淑彬等, 2014 ). ...
1
2015
... 所有植物样品和土壤样品均按照
生态系统固碳项目技术规范编写组(2015) 制定的统一测量方法测定, 其中植物C和SOC含量采用重铬酸钾——外加热法测定; 植物N和土壤TN含量采用凯氏定氮法测定; 植物P含量采用浓硫酸-过氧化氢消煮——钼锑抗分光光度法测定, 土壤TP含量采用浓硫酸-高氯酸消煮——钼锑抗分光光度法测定. ...
1
2015
... 所有植物样品和土壤样品均按照
生态系统固碳项目技术规范编写组(2015) 制定的统一测量方法测定, 其中植物C和SOC含量采用重铬酸钾——外加热法测定; 植物N和土壤TN含量采用凯氏定氮法测定; 植物P含量采用浓硫酸-过氧化氢消煮——钼锑抗分光光度法测定, 土壤TP含量采用浓硫酸-高氯酸消煮——钼锑抗分光光度法测定. ...
不同林龄马尾松人工林林下灌木和草本多样性
1
2014
... 在森林生态系统中, 林下灌木层与乔木层和草本层共同维持着森林生态系统结构与功能的稳定, 在参与养分循环、改善土壤肥力、为林下生物提供栖息环境和提高生态系统多样性等方面起到重要作用, 是森林生态系统不可或缺的组成部分(
郑绍伟等, 2007 ;
卢振龙和龚孝生, 2009 ).灌木植被通常具有木质化茎干而主干不明显, 其茎干多分枝, 树冠矮小, 根系分布广而深(
李清河等, 2006 ).目前, 对森林的林下灌木层的研究主要聚焦于灌木生物量生长模型的建立(
万五星等, 2014 ), 物种多样性和种间联结性(
崔宁洁等, 2014 ), 生态位以及物种分布特征的分析(
康永祥等, 2008 ).对于林下灌木层生态化学计量特征的研究相对较少, 尤其是高寒地区不同林分类型中灌木层的研究尚鲜有报道. ...
不同林龄马尾松人工林林下灌木和草本多样性
1
2014
... 在森林生态系统中, 林下灌木层与乔木层和草本层共同维持着森林生态系统结构与功能的稳定, 在参与养分循环、改善土壤肥力、为林下生物提供栖息环境和提高生态系统多样性等方面起到重要作用, 是森林生态系统不可或缺的组成部分(
郑绍伟等, 2007 ;
卢振龙和龚孝生, 2009 ).灌木植被通常具有木质化茎干而主干不明显, 其茎干多分枝, 树冠矮小, 根系分布广而深(
李清河等, 2006 ).目前, 对森林的林下灌木层的研究主要聚焦于灌木生物量生长模型的建立(
万五星等, 2014 ), 物种多样性和种间联结性(
崔宁洁等, 2014 ), 生态位以及物种分布特征的分析(
康永祥等, 2008 ).对于林下灌木层生态化学计量特征的研究相对较少, 尤其是高寒地区不同林分类型中灌木层的研究尚鲜有报道. ...
青海省森林资源评价
1
2009
... 青海省地处青藏高原的东北部, 地理位置为31.65°-39.32° N, 89.58°-103.07° E.地形复杂多样, 高山、丘陵、河谷、盆地交错分布, 平均海拔3 000 m以上, 属于典型高原大陆性气候.常年平均气温-3.7-6.0 ℃, 年日照时间2 340-3 550 h, 年降水量16.7-776.1 mm (大部分400 mm以下), 年蒸发量1 118.4-3 536.2 mm (大部分1 500 mm以上), 植被地域跨青藏高原、温带荒漠和温带草原3个植被区, 具有高寒和旱生的特点.青海省森林资源贫乏, 森林覆盖率仅有4.57%, 森林植被主要分布于江河及其支流的河谷两岸, 海拔大多在2 000-4 000 m, 以寒温性针叶林为主, 其次为落叶阔叶林(
字洪标等, 2017 ).常见的针叶树种有云杉、圆柏等, 阔叶树种有杨树(
Populus spp.)、白桦等, 林下灌木有金露梅(
Potentilla fruticosa )、鲜卑花(
Sibiraea laevigata )、野蔷薇(
Rosa multiflora )
、 银露梅(
Potentilla glabra )
、 杯腺柳(
Salix cupularis )等, 主要的森林土壤类型有棕壤、黄棕壤、暗棕壤、褐棕壤、砂壤土等(
董旭, 2009 ;
胡雷等, 2015 ). ...
青海省森林资源评价
1
2009
... 青海省地处青藏高原的东北部, 地理位置为31.65°-39.32° N, 89.58°-103.07° E.地形复杂多样, 高山、丘陵、河谷、盆地交错分布, 平均海拔3 000 m以上, 属于典型高原大陆性气候.常年平均气温-3.7-6.0 ℃, 年日照时间2 340-3 550 h, 年降水量16.7-776.1 mm (大部分400 mm以下), 年蒸发量1 118.4-3 536.2 mm (大部分1 500 mm以上), 植被地域跨青藏高原、温带荒漠和温带草原3个植被区, 具有高寒和旱生的特点.青海省森林资源贫乏, 森林覆盖率仅有4.57%, 森林植被主要分布于江河及其支流的河谷两岸, 海拔大多在2 000-4 000 m, 以寒温性针叶林为主, 其次为落叶阔叶林(
字洪标等, 2017 ).常见的针叶树种有云杉、圆柏等, 阔叶树种有杨树(
Populus spp.)、白桦等, 林下灌木有金露梅(
Potentilla fruticosa )、鲜卑花(
Sibiraea laevigata )、野蔷薇(
Rosa multiflora )
、 银露梅(
Potentilla glabra )
、 杯腺柳(
Salix cupularis )等, 主要的森林土壤类型有棕壤、黄棕壤、暗棕壤、褐棕壤、砂壤土等(
董旭, 2009 ;
胡雷等, 2015 ). ...
Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems
1
2007
... 碳(C)、氮(N)、磷(P)作为最重要的生命元素, 是维系地球上所有生命活动的基础(
贺金生和韩兴国, 2010 ).C作为植物有机体结构的重要组成元素, 是植物有机体内生理生化反应过程的能量来源之一(
Güsewell, 2004 ), N、P是植物的基本营养元素, 也是植物有机体内蛋白质、酶类、核酸和脂质的重要组成元素(
Yang & Wang, 2011 ;
戚德辉等, 2016 ;
Tang et al ., 2018 ).C、N、P含量及其化学计量比的差异显著影响植物的生态策略, 从而被广泛应用到生态系统在生物地球化学循环和生态过程耦合关系的研究中(
Elser et al ., 1996 ).截至目前, 国内外****已在区域乃至全球范围开展了许多陆地生态系统生态化学计量特征的研究(
Han et al ., 2005 ;
Elser et al ., 2007 ;
黄小波等, 2016 ).但主要集中在植物叶片和土壤生态化学计量特征方面, 针对植物枝干、根等重要器官方面的研究相对较少(
贺合亮等, 2017 ).枝干是植物连接地下吸收器官和地上同化组织的传导器官, 具有支撑植物体的结构性骨架作用(
李单凤等, 2015 ).根系作为吸收、存储、运输水分和养分的主要器官, 也是植物有机体对外界环境变化响应最为敏感的地下器官, 在森林生态系统物质循环过程中发挥关键作用(
Iversen et al ., 2017 ;
陈晓萍等, 2018 ).因此, 研究植物不同器官间生态化学计量特征以及与土壤组分在各生态过程间的互作关系, 对于揭示生态系统养分循环规律具有重要意义. ...
Organism size, life history, and N:P stoichiometry
1
1996
... 碳(C)、氮(N)、磷(P)作为最重要的生命元素, 是维系地球上所有生命活动的基础(
贺金生和韩兴国, 2010 ).C作为植物有机体结构的重要组成元素, 是植物有机体内生理生化反应过程的能量来源之一(
Güsewell, 2004 ), N、P是植物的基本营养元素, 也是植物有机体内蛋白质、酶类、核酸和脂质的重要组成元素(
Yang & Wang, 2011 ;
戚德辉等, 2016 ;
Tang et al ., 2018 ).C、N、P含量及其化学计量比的差异显著影响植物的生态策略, 从而被广泛应用到生态系统在生物地球化学循环和生态过程耦合关系的研究中(
Elser et al ., 1996 ).截至目前, 国内外****已在区域乃至全球范围开展了许多陆地生态系统生态化学计量特征的研究(
Han et al ., 2005 ;
Elser et al ., 2007 ;
黄小波等, 2016 ).但主要集中在植物叶片和土壤生态化学计量特征方面, 针对植物枝干、根等重要器官方面的研究相对较少(
贺合亮等, 2017 ).枝干是植物连接地下吸收器官和地上同化组织的传导器官, 具有支撑植物体的结构性骨架作用(
李单凤等, 2015 ).根系作为吸收、存储、运输水分和养分的主要器官, 也是植物有机体对外界环境变化响应最为敏感的地下器官, 在森林生态系统物质循环过程中发挥关键作用(
Iversen et al ., 2017 ;
陈晓萍等, 2018 ).因此, 研究植物不同器官间生态化学计量特征以及与土壤组分在各生态过程间的互作关系, 对于揭示生态系统养分循环规律具有重要意义. ...
Nutritional constraints in terrestrial and freshwater food webs
2
2000
... 本研究中, 灌木叶片C含量平均值为449.00 mg·g
-1 , 低于全球陆生植物叶片C含量464.20 mg·g
-1 (
Elser et al ., 2000 )和中国东部南北样带植物叶片C含量480.10 mg·g
-1 (
任书杰等, 2012 ), 说明青海森林的林下灌木植被C储存能力相对较弱.灌木叶片N、P含量平均值分别为19.46、1.69 mg·g
-1 , 略低于全球陆生植物N、P含量(20.10、1.77 mg·g
-1 (
Reich & Oleksyn, 2004 )和20.60、1.99 mg·g
-1 (
Elser et al ., 2000 )), 略低于我国植物叶片N含量(20.24 mg·g
-1 ), 高于其叶片P含量(1.46 mg·g
-1 )(
Han et al ., 2005 ), 高于我国南方优势灌丛优势植被N、P含量(16.57、1.02 mg·g
-1 )(
李家湘等, 2017 )、陕西森林的林下灌木叶片N、P含量(13.79、1.11 mg·g
-1 )(
姜沛沛等, 2016 ), 说明青海森林的林下灌木叶片N、P含量较高.这可能与本研究区域地处高寒区, 纬度偏高, 年降水量少, 年均气温低, 昼夜温差大, 日照强度大等特殊的区域气候条件有一定的关系(
卢航等, 2013 ).温度-植物生理假说认为, 低温诱导植物生长周期缩短, 同时, 植物趋向于分配更多的营养元素于光合器官中, 通过丰富叶片N、P含量在有限的生长周期内保持高效的光合速率以及酶活性来维持植物机体内正常的新陈代谢活动(
Reich & Oleksyn, 2004 ), 因此高海拔地区植物叶片N、P含量较为丰富.这与
杨阔等(2010) 发现青藏高原草地植物群落叶片N、P含量较高的结果一致. ...
... (
Elser et al ., 2000 )), 略低于我国植物叶片N含量(20.24 mg·g
-1 ), 高于其叶片P含量(1.46 mg·g
-1 )(
Han et al ., 2005 ), 高于我国南方优势灌丛优势植被N、P含量(16.57、1.02 mg·g
-1 )(
李家湘等, 2017 )、陕西森林的林下灌木叶片N、P含量(13.79、1.11 mg·g
-1 )(
姜沛沛等, 2016 ), 说明青海森林的林下灌木叶片N、P含量较高.这可能与本研究区域地处高寒区, 纬度偏高, 年降水量少, 年均气温低, 昼夜温差大, 日照强度大等特殊的区域气候条件有一定的关系(
卢航等, 2013 ).温度-植物生理假说认为, 低温诱导植物生长周期缩短, 同时, 植物趋向于分配更多的营养元素于光合器官中, 通过丰富叶片N、P含量在有限的生长周期内保持高效的光合速率以及酶活性来维持植物机体内正常的新陈代谢活动(
Reich & Oleksyn, 2004 ), 因此高海拔地区植物叶片N、P含量较为丰富.这与
杨阔等(2010) 发现青藏高原草地植物群落叶片N、P含量较高的结果一致. ...
四川省甘孜藏族自治州高寒矮灌丛生物量分配及其碳密度的估算
1
2014
... 本研究区森林SOC、TN、TP含量平均值为(95.11、5.17、0.82 mg·g
-1 ), 高于全国土壤C、N、P含量(24.56、1.88、0.78 mg·g
-1 )(
Tian et al ., 2010 ), 与青藏高原高寒植被带土壤N、P含量(5.66、0.75 mg·g
-1 )相接近(
卢同平等, 2017 ).土壤C:N、C:P (20.6、117.7)高于全球森林土壤C:N、C:P (12.4、81.9)(
Cleveland & Liptzin, 2007 )和全国土壤C:N、C:P (12.3、52.7)(
Tian et al ., 2010 ).研究表明, 土壤C:N、C:P是衡量土壤有机质分解矿化速率的重要指标, 通常土壤C:N、C:P与有机质分解矿化速率呈反比关系, 较高的C:N、C:P代表较慢的有机质分解速率, 以及较低的土壤N、P有效性(
Cleveland & Liptzin, 2007 ).由此可推断, 青海高寒区森林SOC、TN、TP储备丰富, 有机质分解和矿化速率较慢, N、P有效性较低.这可能与本研究区域自然环境和植被类型特性有关, 青海高寒地区年降水量偏少、年平均气温低, 森林地表积累了丰富的凋落物, 长期的腐殖化作用有利于土壤有机质的富集, 较低的土壤温度抑制了土壤微生物及土壤酶活性, 地表有机质的分解周期长, 系统累积量大于消耗量, 导致森林土壤N、P元素得以累积, 但缓慢的分解速率导致土壤N、P有效性较低(
高巧等, 2014 ;
王淑彬等, 2014 ). ...
四川省甘孜藏族自治州高寒矮灌丛生物量分配及其碳密度的估算
1
2014
... 本研究区森林SOC、TN、TP含量平均值为(95.11、5.17、0.82 mg·g
-1 ), 高于全国土壤C、N、P含量(24.56、1.88、0.78 mg·g
-1 )(
Tian et al ., 2010 ), 与青藏高原高寒植被带土壤N、P含量(5.66、0.75 mg·g
-1 )相接近(
卢同平等, 2017 ).土壤C:N、C:P (20.6、117.7)高于全球森林土壤C:N、C:P (12.4、81.9)(
Cleveland & Liptzin, 2007 )和全国土壤C:N、C:P (12.3、52.7)(
Tian et al ., 2010 ).研究表明, 土壤C:N、C:P是衡量土壤有机质分解矿化速率的重要指标, 通常土壤C:N、C:P与有机质分解矿化速率呈反比关系, 较高的C:N、C:P代表较慢的有机质分解速率, 以及较低的土壤N、P有效性(
Cleveland & Liptzin, 2007 ).由此可推断, 青海高寒区森林SOC、TN、TP储备丰富, 有机质分解和矿化速率较慢, N、P有效性较低.这可能与本研究区域自然环境和植被类型特性有关, 青海高寒地区年降水量偏少、年平均气温低, 森林地表积累了丰富的凋落物, 长期的腐殖化作用有利于土壤有机质的富集, 较低的土壤温度抑制了土壤微生物及土壤酶活性, 地表有机质的分解周期长, 系统累积量大于消耗量, 导致森林土壤N、P元素得以累积, 但缓慢的分解速率导致土壤N、P有效性较低(
高巧等, 2014 ;
王淑彬等, 2014 ). ...
Geographic variation in primary sex allocation per flower within and among 12 species of Pedicularis(Orobanchaceae): Proportional male investment increases with elevation
1
2010
... 依据RDA发现, 林下灌木C、N、P含量及化学计量特征主要受土壤C:N、海拔、年降水量和年平均气温的影响, 这表明青海高寒区森林的林下灌木受到土壤N有效性、海拔以及区域水热条件的重要调控.土壤作为植被获取营养元素的主要机质, 其N、P有效性将显著影响地上植被组成与结构, 生产力以及有机体内元素分配格局(
李丹维等, 2017 ).
McGroddy等(2004) 认为, 土壤N、P有效性是植物有机体内元素含量的主要驱动因素, 同时土壤N、P有效性容易受降水淋溶作用的影响.
张仁懿等(2014) 研究也表明青藏高原亚高寒草甸植物养分状况受土壤N有效性的显著影响, 土壤N有效性对高寒植物生长的影响比其他元素更为关键.海拔梯度综合了温度、降水、光照等水热条件的变化, 是影响植被生长的主导因子(
Guo et al ., 2010 ).本研究结果与木本植物叶片N、P元素含量受年平均气温和年降水量共同驱动作用的结果(
Chen et al ., 2013b )相符.因为:(1)海拔梯度上降水、气温的差异会影响植物的生长周期, 从而改变C、N、P在各器官间的分配(
Kerkhoff et al ., 2005 ); (2)海拔梯度上土壤温度和水分的变化将影响土壤微生物的群系特征及代谢效率, 间接影响土壤有机质的分解和矿化速率, 而且土壤母质的风化速率以及N、P元素的淋溶强度也受降水和气温等气候因子的显著影响(
刘颖等, 2018 ); (3)不同的海拔梯度上森林植被受到的人为干扰程度也有一定的差异, 放牧、旅游、耕种等活动也会影响植物的元素分配格局(
Nogués-Bravo et al ., 2008 ).相关研究表明, 伴随着全球气候变化, 青藏高原地区已出现气温剧烈升高, 降水量增加的暖湿化趋势(
Chen et al ., 2013a ).因此, 未来在全球气候变化和区域N沉降可能加剧的背景下, 青海森林生态系统灌木层C、N、P化学计量特征将受到强烈影响. ...
N:P ratios in terrestrial plants: Variation and functional significance
1
2004
... 碳(C)、氮(N)、磷(P)作为最重要的生命元素, 是维系地球上所有生命活动的基础(
贺金生和韩兴国, 2010 ).C作为植物有机体结构的重要组成元素, 是植物有机体内生理生化反应过程的能量来源之一(
Güsewell, 2004 ), N、P是植物的基本营养元素, 也是植物有机体内蛋白质、酶类、核酸和脂质的重要组成元素(
Yang & Wang, 2011 ;
戚德辉等, 2016 ;
Tang et al ., 2018 ).C、N、P含量及其化学计量比的差异显著影响植物的生态策略, 从而被广泛应用到生态系统在生物地球化学循环和生态过程耦合关系的研究中(
Elser et al ., 1996 ).截至目前, 国内外****已在区域乃至全球范围开展了许多陆地生态系统生态化学计量特征的研究(
Han et al ., 2005 ;
Elser et al ., 2007 ;
黄小波等, 2016 ).但主要集中在植物叶片和土壤生态化学计量特征方面, 针对植物枝干、根等重要器官方面的研究相对较少(
贺合亮等, 2017 ).枝干是植物连接地下吸收器官和地上同化组织的传导器官, 具有支撑植物体的结构性骨架作用(
李单凤等, 2015 ).根系作为吸收、存储、运输水分和养分的主要器官, 也是植物有机体对外界环境变化响应最为敏感的地下器官, 在森林生态系统物质循环过程中发挥关键作用(
Iversen et al ., 2017 ;
陈晓萍等, 2018 ).因此, 研究植物不同器官间生态化学计量特征以及与土壤组分在各生态过程间的互作关系, 对于揭示生态系统养分循环规律具有重要意义. ...
Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China
3
2005
... 碳(C)、氮(N)、磷(P)作为最重要的生命元素, 是维系地球上所有生命活动的基础(
贺金生和韩兴国, 2010 ).C作为植物有机体结构的重要组成元素, 是植物有机体内生理生化反应过程的能量来源之一(
Güsewell, 2004 ), N、P是植物的基本营养元素, 也是植物有机体内蛋白质、酶类、核酸和脂质的重要组成元素(
Yang & Wang, 2011 ;
戚德辉等, 2016 ;
Tang et al ., 2018 ).C、N、P含量及其化学计量比的差异显著影响植物的生态策略, 从而被广泛应用到生态系统在生物地球化学循环和生态过程耦合关系的研究中(
Elser et al ., 1996 ).截至目前, 国内外****已在区域乃至全球范围开展了许多陆地生态系统生态化学计量特征的研究(
Han et al ., 2005 ;
Elser et al ., 2007 ;
黄小波等, 2016 ).但主要集中在植物叶片和土壤生态化学计量特征方面, 针对植物枝干、根等重要器官方面的研究相对较少(
贺合亮等, 2017 ).枝干是植物连接地下吸收器官和地上同化组织的传导器官, 具有支撑植物体的结构性骨架作用(
李单凤等, 2015 ).根系作为吸收、存储、运输水分和养分的主要器官, 也是植物有机体对外界环境变化响应最为敏感的地下器官, 在森林生态系统物质循环过程中发挥关键作用(
Iversen et al ., 2017 ;
陈晓萍等, 2018 ).因此, 研究植物不同器官间生态化学计量特征以及与土壤组分在各生态过程间的互作关系, 对于揭示生态系统养分循环规律具有重要意义. ...
... 本研究中, 灌木叶片C含量平均值为449.00 mg·g
-1 , 低于全球陆生植物叶片C含量464.20 mg·g
-1 (
Elser et al ., 2000 )和中国东部南北样带植物叶片C含量480.10 mg·g
-1 (
任书杰等, 2012 ), 说明青海森林的林下灌木植被C储存能力相对较弱.灌木叶片N、P含量平均值分别为19.46、1.69 mg·g
-1 , 略低于全球陆生植物N、P含量(20.10、1.77 mg·g
-1 (
Reich & Oleksyn, 2004 )和20.60、1.99 mg·g
-1 (
Elser et al ., 2000 )), 略低于我国植物叶片N含量(20.24 mg·g
-1 ), 高于其叶片P含量(1.46 mg·g
-1 )(
Han et al ., 2005 ), 高于我国南方优势灌丛优势植被N、P含量(16.57、1.02 mg·g
-1 )(
李家湘等, 2017 )、陕西森林的林下灌木叶片N、P含量(13.79、1.11 mg·g
-1 )(
姜沛沛等, 2016 ), 说明青海森林的林下灌木叶片N、P含量较高.这可能与本研究区域地处高寒区, 纬度偏高, 年降水量少, 年均气温低, 昼夜温差大, 日照强度大等特殊的区域气候条件有一定的关系(
卢航等, 2013 ).温度-植物生理假说认为, 低温诱导植物生长周期缩短, 同时, 植物趋向于分配更多的营养元素于光合器官中, 通过丰富叶片N、P含量在有限的生长周期内保持高效的光合速率以及酶活性来维持植物机体内正常的新陈代谢活动(
Reich & Oleksyn, 2004 ), 因此高海拔地区植物叶片N、P含量较为丰富.这与
杨阔等(2010) 发现青藏高原草地植物群落叶片N、P含量较高的结果一致. ...
... 植物叶片N:P通常是判断植物养分限制状况的指标之一.研究表明, 当N:P < 14时, 植物生长受N限制; 当N:P > 16时, 植物生长主要受P限制; 当N:P介于14-16之间时, 植物生长受N和P的共同限制或者不受养分限制(
Koerselman, 1996 ;
Tessier & Raynal, 2003 ).本研究中, 圆柏林灌木叶片N:P (16.72) > 16, 其余6种林分叶片N:P表现为青扦林(6.22) <红桦林(7.21) <毛白杨林(7.47) <白桦林(7.48) <云杉林(12.6) <山杨林(13.7) < 14.圆柏林的林下灌木植被生长受P限制, 其余6种林分林下灌木植被生长受到N限制.这可能是由于林分差异导致其土壤N、P供应能力的不同.本研究中, 圆柏林的林下灌木叶片P含量最低(
图2 ), 圆柏林土壤TN含量最高、C:N最低、C:P较高(
表1 ), 圆柏林的林下土壤N含量丰富, N有效性较高, 而P有效性不足, 因此, 林下灌木植被在N供应较为充沛的条件下, 其生长更趋向于受P限制.其余6种林分林下灌木植被生长限制情况与我国北方森林植被普遍受到N限制的结果相一致(
Han et al ., 2005 ). ...
青藏高原东部窄叶鲜卑花碳、氮、磷化学计量特征
1
2017
... 碳(C)、氮(N)、磷(P)作为最重要的生命元素, 是维系地球上所有生命活动的基础(
贺金生和韩兴国, 2010 ).C作为植物有机体结构的重要组成元素, 是植物有机体内生理生化反应过程的能量来源之一(
Güsewell, 2004 ), N、P是植物的基本营养元素, 也是植物有机体内蛋白质、酶类、核酸和脂质的重要组成元素(
Yang & Wang, 2011 ;
戚德辉等, 2016 ;
Tang et al ., 2018 ).C、N、P含量及其化学计量比的差异显著影响植物的生态策略, 从而被广泛应用到生态系统在生物地球化学循环和生态过程耦合关系的研究中(
Elser et al ., 1996 ).截至目前, 国内外****已在区域乃至全球范围开展了许多陆地生态系统生态化学计量特征的研究(
Han et al ., 2005 ;
Elser et al ., 2007 ;
黄小波等, 2016 ).但主要集中在植物叶片和土壤生态化学计量特征方面, 针对植物枝干、根等重要器官方面的研究相对较少(
贺合亮等, 2017 ).枝干是植物连接地下吸收器官和地上同化组织的传导器官, 具有支撑植物体的结构性骨架作用(
李单凤等, 2015 ).根系作为吸收、存储、运输水分和养分的主要器官, 也是植物有机体对外界环境变化响应最为敏感的地下器官, 在森林生态系统物质循环过程中发挥关键作用(
Iversen et al ., 2017 ;
陈晓萍等, 2018 ).因此, 研究植物不同器官间生态化学计量特征以及与土壤组分在各生态过程间的互作关系, 对于揭示生态系统养分循环规律具有重要意义. ...
青藏高原东部窄叶鲜卑花碳、氮、磷化学计量特征
1
2017
... 碳(C)、氮(N)、磷(P)作为最重要的生命元素, 是维系地球上所有生命活动的基础(
贺金生和韩兴国, 2010 ).C作为植物有机体结构的重要组成元素, 是植物有机体内生理生化反应过程的能量来源之一(
Güsewell, 2004 ), N、P是植物的基本营养元素, 也是植物有机体内蛋白质、酶类、核酸和脂质的重要组成元素(
Yang & Wang, 2011 ;
戚德辉等, 2016 ;
Tang et al ., 2018 ).C、N、P含量及其化学计量比的差异显著影响植物的生态策略, 从而被广泛应用到生态系统在生物地球化学循环和生态过程耦合关系的研究中(
Elser et al ., 1996 ).截至目前, 国内外****已在区域乃至全球范围开展了许多陆地生态系统生态化学计量特征的研究(
Han et al ., 2005 ;
Elser et al ., 2007 ;
黄小波等, 2016 ).但主要集中在植物叶片和土壤生态化学计量特征方面, 针对植物枝干、根等重要器官方面的研究相对较少(
贺合亮等, 2017 ).枝干是植物连接地下吸收器官和地上同化组织的传导器官, 具有支撑植物体的结构性骨架作用(
李单凤等, 2015 ).根系作为吸收、存储、运输水分和养分的主要器官, 也是植物有机体对外界环境变化响应最为敏感的地下器官, 在森林生态系统物质循环过程中发挥关键作用(
Iversen et al ., 2017 ;
陈晓萍等, 2018 ).因此, 研究植物不同器官间生态化学计量特征以及与土壤组分在各生态过程间的互作关系, 对于揭示生态系统养分循环规律具有重要意义. ...
生态化学计量学: 探索从个体到生态系统的统一化理论
1
2010
... 碳(C)、氮(N)、磷(P)作为最重要的生命元素, 是维系地球上所有生命活动的基础(
贺金生和韩兴国, 2010 ).C作为植物有机体结构的重要组成元素, 是植物有机体内生理生化反应过程的能量来源之一(
Güsewell, 2004 ), N、P是植物的基本营养元素, 也是植物有机体内蛋白质、酶类、核酸和脂质的重要组成元素(
Yang & Wang, 2011 ;
戚德辉等, 2016 ;
Tang et al ., 2018 ).C、N、P含量及其化学计量比的差异显著影响植物的生态策略, 从而被广泛应用到生态系统在生物地球化学循环和生态过程耦合关系的研究中(
Elser et al ., 1996 ).截至目前, 国内外****已在区域乃至全球范围开展了许多陆地生态系统生态化学计量特征的研究(
Han et al ., 2005 ;
Elser et al ., 2007 ;
黄小波等, 2016 ).但主要集中在植物叶片和土壤生态化学计量特征方面, 针对植物枝干、根等重要器官方面的研究相对较少(
贺合亮等, 2017 ).枝干是植物连接地下吸收器官和地上同化组织的传导器官, 具有支撑植物体的结构性骨架作用(
李单凤等, 2015 ).根系作为吸收、存储、运输水分和养分的主要器官, 也是植物有机体对外界环境变化响应最为敏感的地下器官, 在森林生态系统物质循环过程中发挥关键作用(
Iversen et al ., 2017 ;
陈晓萍等, 2018 ).因此, 研究植物不同器官间生态化学计量特征以及与土壤组分在各生态过程间的互作关系, 对于揭示生态系统养分循环规律具有重要意义. ...
生态化学计量学: 探索从个体到生态系统的统一化理论
1
2010
... 碳(C)、氮(N)、磷(P)作为最重要的生命元素, 是维系地球上所有生命活动的基础(
贺金生和韩兴国, 2010 ).C作为植物有机体结构的重要组成元素, 是植物有机体内生理生化反应过程的能量来源之一(
Güsewell, 2004 ), N、P是植物的基本营养元素, 也是植物有机体内蛋白质、酶类、核酸和脂质的重要组成元素(
Yang & Wang, 2011 ;
戚德辉等, 2016 ;
Tang et al ., 2018 ).C、N、P含量及其化学计量比的差异显著影响植物的生态策略, 从而被广泛应用到生态系统在生物地球化学循环和生态过程耦合关系的研究中(
Elser et al ., 1996 ).截至目前, 国内外****已在区域乃至全球范围开展了许多陆地生态系统生态化学计量特征的研究(
Han et al ., 2005 ;
Elser et al ., 2007 ;
黄小波等, 2016 ).但主要集中在植物叶片和土壤生态化学计量特征方面, 针对植物枝干、根等重要器官方面的研究相对较少(
贺合亮等, 2017 ).枝干是植物连接地下吸收器官和地上同化组织的传导器官, 具有支撑植物体的结构性骨架作用(
李单凤等, 2015 ).根系作为吸收、存储、运输水分和养分的主要器官, 也是植物有机体对外界环境变化响应最为敏感的地下器官, 在森林生态系统物质循环过程中发挥关键作用(
Iversen et al ., 2017 ;
陈晓萍等, 2018 ).因此, 研究植物不同器官间生态化学计量特征以及与土壤组分在各生态过程间的互作关系, 对于揭示生态系统养分循环规律具有重要意义. ...
青海省森林生态系统植被固碳现状研究
1
2015
... 青海省地处青藏高原的东北部, 地理位置为31.65°-39.32° N, 89.58°-103.07° E.地形复杂多样, 高山、丘陵、河谷、盆地交错分布, 平均海拔3 000 m以上, 属于典型高原大陆性气候.常年平均气温-3.7-6.0 ℃, 年日照时间2 340-3 550 h, 年降水量16.7-776.1 mm (大部分400 mm以下), 年蒸发量1 118.4-3 536.2 mm (大部分1 500 mm以上), 植被地域跨青藏高原、温带荒漠和温带草原3个植被区, 具有高寒和旱生的特点.青海省森林资源贫乏, 森林覆盖率仅有4.57%, 森林植被主要分布于江河及其支流的河谷两岸, 海拔大多在2 000-4 000 m, 以寒温性针叶林为主, 其次为落叶阔叶林(
字洪标等, 2017 ).常见的针叶树种有云杉、圆柏等, 阔叶树种有杨树(
Populus spp.)、白桦等, 林下灌木有金露梅(
Potentilla fruticosa )、鲜卑花(
Sibiraea laevigata )、野蔷薇(
Rosa multiflora )
、 银露梅(
Potentilla glabra )
、 杯腺柳(
Salix cupularis )等, 主要的森林土壤类型有棕壤、黄棕壤、暗棕壤、褐棕壤、砂壤土等(
董旭, 2009 ;
胡雷等, 2015 ). ...
青海省森林生态系统植被固碳现状研究
1
2015
... 青海省地处青藏高原的东北部, 地理位置为31.65°-39.32° N, 89.58°-103.07° E.地形复杂多样, 高山、丘陵、河谷、盆地交错分布, 平均海拔3 000 m以上, 属于典型高原大陆性气候.常年平均气温-3.7-6.0 ℃, 年日照时间2 340-3 550 h, 年降水量16.7-776.1 mm (大部分400 mm以下), 年蒸发量1 118.4-3 536.2 mm (大部分1 500 mm以上), 植被地域跨青藏高原、温带荒漠和温带草原3个植被区, 具有高寒和旱生的特点.青海省森林资源贫乏, 森林覆盖率仅有4.57%, 森林植被主要分布于江河及其支流的河谷两岸, 海拔大多在2 000-4 000 m, 以寒温性针叶林为主, 其次为落叶阔叶林(
字洪标等, 2017 ).常见的针叶树种有云杉、圆柏等, 阔叶树种有杨树(
Populus spp.)、白桦等, 林下灌木有金露梅(
Potentilla fruticosa )、鲜卑花(
Sibiraea laevigata )、野蔷薇(
Rosa multiflora )
、 银露梅(
Potentilla glabra )
、 杯腺柳(
Salix cupularis )等, 主要的森林土壤类型有棕壤、黄棕壤、暗棕壤、褐棕壤、砂壤土等(
董旭, 2009 ;
胡雷等, 2015 ). ...
云南普洱季风常绿阔叶林152种木本植物叶片C、N、P化学计量特征
1
2016
... 碳(C)、氮(N)、磷(P)作为最重要的生命元素, 是维系地球上所有生命活动的基础(
贺金生和韩兴国, 2010 ).C作为植物有机体结构的重要组成元素, 是植物有机体内生理生化反应过程的能量来源之一(
Güsewell, 2004 ), N、P是植物的基本营养元素, 也是植物有机体内蛋白质、酶类、核酸和脂质的重要组成元素(
Yang & Wang, 2011 ;
戚德辉等, 2016 ;
Tang et al ., 2018 ).C、N、P含量及其化学计量比的差异显著影响植物的生态策略, 从而被广泛应用到生态系统在生物地球化学循环和生态过程耦合关系的研究中(
Elser et al ., 1996 ).截至目前, 国内外****已在区域乃至全球范围开展了许多陆地生态系统生态化学计量特征的研究(
Han et al ., 2005 ;
Elser et al ., 2007 ;
黄小波等, 2016 ).但主要集中在植物叶片和土壤生态化学计量特征方面, 针对植物枝干、根等重要器官方面的研究相对较少(
贺合亮等, 2017 ).枝干是植物连接地下吸收器官和地上同化组织的传导器官, 具有支撑植物体的结构性骨架作用(
李单凤等, 2015 ).根系作为吸收、存储、运输水分和养分的主要器官, 也是植物有机体对外界环境变化响应最为敏感的地下器官, 在森林生态系统物质循环过程中发挥关键作用(
Iversen et al ., 2017 ;
陈晓萍等, 2018 ).因此, 研究植物不同器官间生态化学计量特征以及与土壤组分在各生态过程间的互作关系, 对于揭示生态系统养分循环规律具有重要意义. ...
云南普洱季风常绿阔叶林152种木本植物叶片C、N、P化学计量特征
1
2016
... 碳(C)、氮(N)、磷(P)作为最重要的生命元素, 是维系地球上所有生命活动的基础(
贺金生和韩兴国, 2010 ).C作为植物有机体结构的重要组成元素, 是植物有机体内生理生化反应过程的能量来源之一(
Güsewell, 2004 ), N、P是植物的基本营养元素, 也是植物有机体内蛋白质、酶类、核酸和脂质的重要组成元素(
Yang & Wang, 2011 ;
戚德辉等, 2016 ;
Tang et al ., 2018 ).C、N、P含量及其化学计量比的差异显著影响植物的生态策略, 从而被广泛应用到生态系统在生物地球化学循环和生态过程耦合关系的研究中(
Elser et al ., 1996 ).截至目前, 国内外****已在区域乃至全球范围开展了许多陆地生态系统生态化学计量特征的研究(
Han et al ., 2005 ;
Elser et al ., 2007 ;
黄小波等, 2016 ).但主要集中在植物叶片和土壤生态化学计量特征方面, 针对植物枝干、根等重要器官方面的研究相对较少(
贺合亮等, 2017 ).枝干是植物连接地下吸收器官和地上同化组织的传导器官, 具有支撑植物体的结构性骨架作用(
李单凤等, 2015 ).根系作为吸收、存储、运输水分和养分的主要器官, 也是植物有机体对外界环境变化响应最为敏感的地下器官, 在森林生态系统物质循环过程中发挥关键作用(
Iversen et al ., 2017 ;
陈晓萍等, 2018 ).因此, 研究植物不同器官间生态化学计量特征以及与土壤组分在各生态过程间的互作关系, 对于揭示生态系统养分循环规律具有重要意义. ...
A global fine-root ecology database to address below-ground challenges in plant ecology
1
2017
... 碳(C)、氮(N)、磷(P)作为最重要的生命元素, 是维系地球上所有生命活动的基础(
贺金生和韩兴国, 2010 ).C作为植物有机体结构的重要组成元素, 是植物有机体内生理生化反应过程的能量来源之一(
Güsewell, 2004 ), N、P是植物的基本营养元素, 也是植物有机体内蛋白质、酶类、核酸和脂质的重要组成元素(
Yang & Wang, 2011 ;
戚德辉等, 2016 ;
Tang et al ., 2018 ).C、N、P含量及其化学计量比的差异显著影响植物的生态策略, 从而被广泛应用到生态系统在生物地球化学循环和生态过程耦合关系的研究中(
Elser et al ., 1996 ).截至目前, 国内外****已在区域乃至全球范围开展了许多陆地生态系统生态化学计量特征的研究(
Han et al ., 2005 ;
Elser et al ., 2007 ;
黄小波等, 2016 ).但主要集中在植物叶片和土壤生态化学计量特征方面, 针对植物枝干、根等重要器官方面的研究相对较少(
贺合亮等, 2017 ).枝干是植物连接地下吸收器官和地上同化组织的传导器官, 具有支撑植物体的结构性骨架作用(
李单凤等, 2015 ).根系作为吸收、存储、运输水分和养分的主要器官, 也是植物有机体对外界环境变化响应最为敏感的地下器官, 在森林生态系统物质循环过程中发挥关键作用(
Iversen et al ., 2017 ;
陈晓萍等, 2018 ).因此, 研究植物不同器官间生态化学计量特征以及与土壤组分在各生态过程间的互作关系, 对于揭示生态系统养分循环规律具有重要意义. ...
青海生态环境变化与生态建设的空间布局
1
2004
... 青海省位于青藏高原的东北部, 深居内陆腹地, 是全球气候变化的敏感区和脆弱区, 也是我国乃至亚洲重要的生态安全屏障(
贾敬敦等, 2004 ).青海森林覆盖面积小, 但植被类型多样化, 其森林生态系统及林下灌木层植被在高寒区水土保持, 涵养水源, 防风固沙, 维系生态系统平衡与稳定等方面发挥着关键的生态功能(
张胜邦, 2005 ).青海地区森林凋落物层较厚, 矿质土壤层较薄, 同时受雪被和季节性冻融等特殊因素的综合影响, 其森林生态系统物质循环过程可能具有其独特性(
左巍等, 2016 ).由于特殊的地理位置, 气候寒冷而干旱, 地形地貌复杂多样, 使植被对气候变化异常敏感和脆弱(
王根绪等, 2006 ).因此, 研究青海森林的林下灌木层生态化学计量特征将有助于我们深入了解高寒区森林生态系统的稳定性以及养分循环机理, 揭示高寒环境下森林植被的生态适应策略.基于此, 本研究选取青海省7种主要优势林分——白桦(
Betula platyphylla )林、毛白杨(
Populus tomentosa )林
、 红桦(
Betula albosinensis )林、青扦(
Picea wilsonii )林、山杨(
Populus davidiana )林、圆柏(
Sabina chinensis )林、云杉(
Picea asperata )林的林下灌木层植被为研究对象, 通过分析不同林分林下灌木植被和土壤生态化学计量特征, 阐明林下灌木植被化学计量特征与土壤组分之间的耦合关系, 并探讨林下灌木植被的养分限制情况及其生态化学计量特征的影响因素, 旨在为研究林下灌木植被对高寒环境的响应与适应机制提供基础数据, 也为指导青海高寒区森林生态系统的保护、恢复与重建提供科学依据. ...
青海生态环境变化与生态建设的空间布局
1
2004
... 青海省位于青藏高原的东北部, 深居内陆腹地, 是全球气候变化的敏感区和脆弱区, 也是我国乃至亚洲重要的生态安全屏障(
贾敬敦等, 2004 ).青海森林覆盖面积小, 但植被类型多样化, 其森林生态系统及林下灌木层植被在高寒区水土保持, 涵养水源, 防风固沙, 维系生态系统平衡与稳定等方面发挥着关键的生态功能(
张胜邦, 2005 ).青海地区森林凋落物层较厚, 矿质土壤层较薄, 同时受雪被和季节性冻融等特殊因素的综合影响, 其森林生态系统物质循环过程可能具有其独特性(
左巍等, 2016 ).由于特殊的地理位置, 气候寒冷而干旱, 地形地貌复杂多样, 使植被对气候变化异常敏感和脆弱(
王根绪等, 2006 ).因此, 研究青海森林的林下灌木层生态化学计量特征将有助于我们深入了解高寒区森林生态系统的稳定性以及养分循环机理, 揭示高寒环境下森林植被的生态适应策略.基于此, 本研究选取青海省7种主要优势林分——白桦(
Betula platyphylla )林、毛白杨(
Populus tomentosa )林
、 红桦(
Betula albosinensis )林、青扦(
Picea wilsonii )林、山杨(
Populus davidiana )林、圆柏(
Sabina chinensis )林、云杉(
Picea asperata )林的林下灌木层植被为研究对象, 通过分析不同林分林下灌木植被和土壤生态化学计量特征, 阐明林下灌木植被化学计量特征与土壤组分之间的耦合关系, 并探讨林下灌木植被的养分限制情况及其生态化学计量特征的影响因素, 旨在为研究林下灌木植被对高寒环境的响应与适应机制提供基础数据, 也为指导青海高寒区森林生态系统的保护、恢复与重建提供科学依据. ...
陕西省森林群落乔灌草叶片和凋落物C、N、P生态化学计量特征
1
2016
... 本研究中, 灌木叶片C含量平均值为449.00 mg·g
-1 , 低于全球陆生植物叶片C含量464.20 mg·g
-1 (
Elser et al ., 2000 )和中国东部南北样带植物叶片C含量480.10 mg·g
-1 (
任书杰等, 2012 ), 说明青海森林的林下灌木植被C储存能力相对较弱.灌木叶片N、P含量平均值分别为19.46、1.69 mg·g
-1 , 略低于全球陆生植物N、P含量(20.10、1.77 mg·g
-1 (
Reich & Oleksyn, 2004 )和20.60、1.99 mg·g
-1 (
Elser et al ., 2000 )), 略低于我国植物叶片N含量(20.24 mg·g
-1 ), 高于其叶片P含量(1.46 mg·g
-1 )(
Han et al ., 2005 ), 高于我国南方优势灌丛优势植被N、P含量(16.57、1.02 mg·g
-1 )(
李家湘等, 2017 )、陕西森林的林下灌木叶片N、P含量(13.79、1.11 mg·g
-1 )(
姜沛沛等, 2016 ), 说明青海森林的林下灌木叶片N、P含量较高.这可能与本研究区域地处高寒区, 纬度偏高, 年降水量少, 年均气温低, 昼夜温差大, 日照强度大等特殊的区域气候条件有一定的关系(
卢航等, 2013 ).温度-植物生理假说认为, 低温诱导植物生长周期缩短, 同时, 植物趋向于分配更多的营养元素于光合器官中, 通过丰富叶片N、P含量在有限的生长周期内保持高效的光合速率以及酶活性来维持植物机体内正常的新陈代谢活动(
Reich & Oleksyn, 2004 ), 因此高海拔地区植物叶片N、P含量较为丰富.这与
杨阔等(2010) 发现青藏高原草地植物群落叶片N、P含量较高的结果一致. ...
陕西省森林群落乔灌草叶片和凋落物C、N、P生态化学计量特征
1
2016
... 本研究中, 灌木叶片C含量平均值为449.00 mg·g
-1 , 低于全球陆生植物叶片C含量464.20 mg·g
-1 (
Elser et al ., 2000 )和中国东部南北样带植物叶片C含量480.10 mg·g
-1 (
任书杰等, 2012 ), 说明青海森林的林下灌木植被C储存能力相对较弱.灌木叶片N、P含量平均值分别为19.46、1.69 mg·g
-1 , 略低于全球陆生植物N、P含量(20.10、1.77 mg·g
-1 (
Reich & Oleksyn, 2004 )和20.60、1.99 mg·g
-1 (
Elser et al ., 2000 )), 略低于我国植物叶片N含量(20.24 mg·g
-1 ), 高于其叶片P含量(1.46 mg·g
-1 )(
Han et al ., 2005 ), 高于我国南方优势灌丛优势植被N、P含量(16.57、1.02 mg·g
-1 )(
李家湘等, 2017 )、陕西森林的林下灌木叶片N、P含量(13.79、1.11 mg·g
-1 )(
姜沛沛等, 2016 ), 说明青海森林的林下灌木叶片N、P含量较高.这可能与本研究区域地处高寒区, 纬度偏高, 年降水量少, 年均气温低, 昼夜温差大, 日照强度大等特殊的区域气候条件有一定的关系(
卢航等, 2013 ).温度-植物生理假说认为, 低温诱导植物生长周期缩短, 同时, 植物趋向于分配更多的营养元素于光合器官中, 通过丰富叶片N、P含量在有限的生长周期内保持高效的光合速率以及酶活性来维持植物机体内正常的新陈代谢活动(
Reich & Oleksyn, 2004 ), 因此高海拔地区植物叶片N、P含量较为丰富.这与
杨阔等(2010) 发现青藏高原草地植物群落叶片N、P含量较高的结果一致. ...
陕北黄龙山辽东栎群落优势种群生态位研究
1
2008
... 在森林生态系统中, 林下灌木层与乔木层和草本层共同维持着森林生态系统结构与功能的稳定, 在参与养分循环、改善土壤肥力、为林下生物提供栖息环境和提高生态系统多样性等方面起到重要作用, 是森林生态系统不可或缺的组成部分(
郑绍伟等, 2007 ;
卢振龙和龚孝生, 2009 ).灌木植被通常具有木质化茎干而主干不明显, 其茎干多分枝, 树冠矮小, 根系分布广而深(
李清河等, 2006 ).目前, 对森林的林下灌木层的研究主要聚焦于灌木生物量生长模型的建立(
万五星等, 2014 ), 物种多样性和种间联结性(
崔宁洁等, 2014 ), 生态位以及物种分布特征的分析(
康永祥等, 2008 ).对于林下灌木层生态化学计量特征的研究相对较少, 尤其是高寒地区不同林分类型中灌木层的研究尚鲜有报道. ...
陕北黄龙山辽东栎群落优势种群生态位研究
1
2008
... 在森林生态系统中, 林下灌木层与乔木层和草本层共同维持着森林生态系统结构与功能的稳定, 在参与养分循环、改善土壤肥力、为林下生物提供栖息环境和提高生态系统多样性等方面起到重要作用, 是森林生态系统不可或缺的组成部分(
郑绍伟等, 2007 ;
卢振龙和龚孝生, 2009 ).灌木植被通常具有木质化茎干而主干不明显, 其茎干多分枝, 树冠矮小, 根系分布广而深(
李清河等, 2006 ).目前, 对森林的林下灌木层的研究主要聚焦于灌木生物量生长模型的建立(
万五星等, 2014 ), 物种多样性和种间联结性(
崔宁洁等, 2014 ), 生态位以及物种分布特征的分析(
康永祥等, 2008 ).对于林下灌木层生态化学计量特征的研究相对较少, 尤其是高寒地区不同林分类型中灌木层的研究尚鲜有报道. ...
Plant allometry, stoichiometry and the temperature-dependence of primary productivity
1
2005
... 依据RDA发现, 林下灌木C、N、P含量及化学计量特征主要受土壤C:N、海拔、年降水量和年平均气温的影响, 这表明青海高寒区森林的林下灌木受到土壤N有效性、海拔以及区域水热条件的重要调控.土壤作为植被获取营养元素的主要机质, 其N、P有效性将显著影响地上植被组成与结构, 生产力以及有机体内元素分配格局(
李丹维等, 2017 ).
McGroddy等(2004) 认为, 土壤N、P有效性是植物有机体内元素含量的主要驱动因素, 同时土壤N、P有效性容易受降水淋溶作用的影响.
张仁懿等(2014) 研究也表明青藏高原亚高寒草甸植物养分状况受土壤N有效性的显著影响, 土壤N有效性对高寒植物生长的影响比其他元素更为关键.海拔梯度综合了温度、降水、光照等水热条件的变化, 是影响植被生长的主导因子(
Guo et al ., 2010 ).本研究结果与木本植物叶片N、P元素含量受年平均气温和年降水量共同驱动作用的结果(
Chen et al ., 2013b )相符.因为:(1)海拔梯度上降水、气温的差异会影响植物的生长周期, 从而改变C、N、P在各器官间的分配(
Kerkhoff et al ., 2005 ); (2)海拔梯度上土壤温度和水分的变化将影响土壤微生物的群系特征及代谢效率, 间接影响土壤有机质的分解和矿化速率, 而且土壤母质的风化速率以及N、P元素的淋溶强度也受降水和气温等气候因子的显著影响(
刘颖等, 2018 ); (3)不同的海拔梯度上森林植被受到的人为干扰程度也有一定的差异, 放牧、旅游、耕种等活动也会影响植物的元素分配格局(
Nogués-Bravo et al ., 2008 ).相关研究表明, 伴随着全球气候变化, 青藏高原地区已出现气温剧烈升高, 降水量增加的暖湿化趋势(
Chen et al ., 2013a ).因此, 未来在全球气候变化和区域N沉降可能加剧的背景下, 青海森林生态系统灌木层C、N、P化学计量特征将受到强烈影响. ...
The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation
1
1996
... 植物叶片N:P通常是判断植物养分限制状况的指标之一.研究表明, 当N:P < 14时, 植物生长受N限制; 当N:P > 16时, 植物生长主要受P限制; 当N:P介于14-16之间时, 植物生长受N和P的共同限制或者不受养分限制(
Koerselman, 1996 ;
Tessier & Raynal, 2003 ).本研究中, 圆柏林灌木叶片N:P (16.72) > 16, 其余6种林分叶片N:P表现为青扦林(6.22) <红桦林(7.21) <毛白杨林(7.47) <白桦林(7.48) <云杉林(12.6) <山杨林(13.7) < 14.圆柏林的林下灌木植被生长受P限制, 其余6种林分林下灌木植被生长受到N限制.这可能是由于林分差异导致其土壤N、P供应能力的不同.本研究中, 圆柏林的林下灌木叶片P含量最低(
图2 ), 圆柏林土壤TN含量最高、C:N最低、C:P较高(
表1 ), 圆柏林的林下土壤N含量丰富, N有效性较高, 而P有效性不足, 因此, 林下灌木植被在N供应较为充沛的条件下, 其生长更趋向于受P限制.其余6种林分林下灌木植被生长限制情况与我国北方森林植被普遍受到N限制的结果相一致(
Han et al ., 2005 ). ...
黄土高原优势灌丛营养器官化学计量特征的环境分异和机制
1
2015
... 碳(C)、氮(N)、磷(P)作为最重要的生命元素, 是维系地球上所有生命活动的基础(
贺金生和韩兴国, 2010 ).C作为植物有机体结构的重要组成元素, 是植物有机体内生理生化反应过程的能量来源之一(
Güsewell, 2004 ), N、P是植物的基本营养元素, 也是植物有机体内蛋白质、酶类、核酸和脂质的重要组成元素(
Yang & Wang, 2011 ;
戚德辉等, 2016 ;
Tang et al ., 2018 ).C、N、P含量及其化学计量比的差异显著影响植物的生态策略, 从而被广泛应用到生态系统在生物地球化学循环和生态过程耦合关系的研究中(
Elser et al ., 1996 ).截至目前, 国内外****已在区域乃至全球范围开展了许多陆地生态系统生态化学计量特征的研究(
Han et al ., 2005 ;
Elser et al ., 2007 ;
黄小波等, 2016 ).但主要集中在植物叶片和土壤生态化学计量特征方面, 针对植物枝干、根等重要器官方面的研究相对较少(
贺合亮等, 2017 ).枝干是植物连接地下吸收器官和地上同化组织的传导器官, 具有支撑植物体的结构性骨架作用(
李单凤等, 2015 ).根系作为吸收、存储、运输水分和养分的主要器官, 也是植物有机体对外界环境变化响应最为敏感的地下器官, 在森林生态系统物质循环过程中发挥关键作用(
Iversen et al ., 2017 ;
陈晓萍等, 2018 ).因此, 研究植物不同器官间生态化学计量特征以及与土壤组分在各生态过程间的互作关系, 对于揭示生态系统养分循环规律具有重要意义. ...
黄土高原优势灌丛营养器官化学计量特征的环境分异和机制
1
2015
... 碳(C)、氮(N)、磷(P)作为最重要的生命元素, 是维系地球上所有生命活动的基础(
贺金生和韩兴国, 2010 ).C作为植物有机体结构的重要组成元素, 是植物有机体内生理生化反应过程的能量来源之一(
Güsewell, 2004 ), N、P是植物的基本营养元素, 也是植物有机体内蛋白质、酶类、核酸和脂质的重要组成元素(
Yang & Wang, 2011 ;
戚德辉等, 2016 ;
Tang et al ., 2018 ).C、N、P含量及其化学计量比的差异显著影响植物的生态策略, 从而被广泛应用到生态系统在生物地球化学循环和生态过程耦合关系的研究中(
Elser et al ., 1996 ).截至目前, 国内外****已在区域乃至全球范围开展了许多陆地生态系统生态化学计量特征的研究(
Han et al ., 2005 ;
Elser et al ., 2007 ;
黄小波等, 2016 ).但主要集中在植物叶片和土壤生态化学计量特征方面, 针对植物枝干、根等重要器官方面的研究相对较少(
贺合亮等, 2017 ).枝干是植物连接地下吸收器官和地上同化组织的传导器官, 具有支撑植物体的结构性骨架作用(
李单凤等, 2015 ).根系作为吸收、存储、运输水分和养分的主要器官, 也是植物有机体对外界环境变化响应最为敏感的地下器官, 在森林生态系统物质循环过程中发挥关键作用(
Iversen et al ., 2017 ;
陈晓萍等, 2018 ).因此, 研究植物不同器官间生态化学计量特征以及与土壤组分在各生态过程间的互作关系, 对于揭示生态系统养分循环规律具有重要意义. ...
太白山不同海拔土壤碳、氮、磷含量及生态化学计量特征
2
2017
... 本研究中, 7种林分间SOC和TN含量呈现出明显差异, 而TP含量则差异不明显.这可能是由于受到营养元素的积累方式以及林分类型的影响, 林下土壤养分状况差异较大.在自然森林生态系统中, 森林土壤有机碳来自地表凋落物以及根系凋亡分解的积累, 而土壤全氮主要通过有机质分解矿化的途径累积(
李丹维等, 2017 ).首先, 林分的差异会影响地表凋落物、地下根系以及根系分泌物的组成, 同时凋落物的类型、数量以及分解速率是影响森林土壤养分含量变化和实现系统养分循环的重要因素(
杨佳佳等, 2014 ).其次, 林分的差异会影响土壤微生物、土壤动物和土壤酶的组成与功能多样性(
庞圣江等, 2015 ;
字洪标等, 2017 ), 而在土壤生物化学循环过程中, 土壤微生物通过氨化、硝化和反硝化等作用将土壤内颗粒态有机氮转化成可溶性有机氮, 而土壤酶作为催化剂也参与了其中复杂的生化反应(
李喜霞等, 2018 ).土壤P作为一种沉积性元素, 主要来源于岩石母质的风化, 其过程稳定而漫长, 因而在自然生态系统中P迁移能力较弱, 空间异质性较小(
李红林等, 2015 ).因此不同林分之间TP含量差异不明显. ...
... 依据RDA发现, 林下灌木C、N、P含量及化学计量特征主要受土壤C:N、海拔、年降水量和年平均气温的影响, 这表明青海高寒区森林的林下灌木受到土壤N有效性、海拔以及区域水热条件的重要调控.土壤作为植被获取营养元素的主要机质, 其N、P有效性将显著影响地上植被组成与结构, 生产力以及有机体内元素分配格局(
李丹维等, 2017 ).
McGroddy等(2004) 认为, 土壤N、P有效性是植物有机体内元素含量的主要驱动因素, 同时土壤N、P有效性容易受降水淋溶作用的影响.
张仁懿等(2014) 研究也表明青藏高原亚高寒草甸植物养分状况受土壤N有效性的显著影响, 土壤N有效性对高寒植物生长的影响比其他元素更为关键.海拔梯度综合了温度、降水、光照等水热条件的变化, 是影响植被生长的主导因子(
Guo et al ., 2010 ).本研究结果与木本植物叶片N、P元素含量受年平均气温和年降水量共同驱动作用的结果(
Chen et al ., 2013b )相符.因为:(1)海拔梯度上降水、气温的差异会影响植物的生长周期, 从而改变C、N、P在各器官间的分配(
Kerkhoff et al ., 2005 ); (2)海拔梯度上土壤温度和水分的变化将影响土壤微生物的群系特征及代谢效率, 间接影响土壤有机质的分解和矿化速率, 而且土壤母质的风化速率以及N、P元素的淋溶强度也受降水和气温等气候因子的显著影响(
刘颖等, 2018 ); (3)不同的海拔梯度上森林植被受到的人为干扰程度也有一定的差异, 放牧、旅游、耕种等活动也会影响植物的元素分配格局(
Nogués-Bravo et al ., 2008 ).相关研究表明, 伴随着全球气候变化, 青藏高原地区已出现气温剧烈升高, 降水量增加的暖湿化趋势(
Chen et al ., 2013a ).因此, 未来在全球气候变化和区域N沉降可能加剧的背景下, 青海森林生态系统灌木层C、N、P化学计量特征将受到强烈影响. ...
太白山不同海拔土壤碳、氮、磷含量及生态化学计量特征
2
2017
... 本研究中, 7种林分间SOC和TN含量呈现出明显差异, 而TP含量则差异不明显.这可能是由于受到营养元素的积累方式以及林分类型的影响, 林下土壤养分状况差异较大.在自然森林生态系统中, 森林土壤有机碳来自地表凋落物以及根系凋亡分解的积累, 而土壤全氮主要通过有机质分解矿化的途径累积(
李丹维等, 2017 ).首先, 林分的差异会影响地表凋落物、地下根系以及根系分泌物的组成, 同时凋落物的类型、数量以及分解速率是影响森林土壤养分含量变化和实现系统养分循环的重要因素(
杨佳佳等, 2014 ).其次, 林分的差异会影响土壤微生物、土壤动物和土壤酶的组成与功能多样性(
庞圣江等, 2015 ;
字洪标等, 2017 ), 而在土壤生物化学循环过程中, 土壤微生物通过氨化、硝化和反硝化等作用将土壤内颗粒态有机氮转化成可溶性有机氮, 而土壤酶作为催化剂也参与了其中复杂的生化反应(
李喜霞等, 2018 ).土壤P作为一种沉积性元素, 主要来源于岩石母质的风化, 其过程稳定而漫长, 因而在自然生态系统中P迁移能力较弱, 空间异质性较小(
李红林等, 2015 ).因此不同林分之间TP含量差异不明显. ...
... 依据RDA发现, 林下灌木C、N、P含量及化学计量特征主要受土壤C:N、海拔、年降水量和年平均气温的影响, 这表明青海高寒区森林的林下灌木受到土壤N有效性、海拔以及区域水热条件的重要调控.土壤作为植被获取营养元素的主要机质, 其N、P有效性将显著影响地上植被组成与结构, 生产力以及有机体内元素分配格局(
李丹维等, 2017 ).
McGroddy等(2004) 认为, 土壤N、P有效性是植物有机体内元素含量的主要驱动因素, 同时土壤N、P有效性容易受降水淋溶作用的影响.
张仁懿等(2014) 研究也表明青藏高原亚高寒草甸植物养分状况受土壤N有效性的显著影响, 土壤N有效性对高寒植物生长的影响比其他元素更为关键.海拔梯度综合了温度、降水、光照等水热条件的变化, 是影响植被生长的主导因子(
Guo et al ., 2010 ).本研究结果与木本植物叶片N、P元素含量受年平均气温和年降水量共同驱动作用的结果(
Chen et al ., 2013b )相符.因为:(1)海拔梯度上降水、气温的差异会影响植物的生长周期, 从而改变C、N、P在各器官间的分配(
Kerkhoff et al ., 2005 ); (2)海拔梯度上土壤温度和水分的变化将影响土壤微生物的群系特征及代谢效率, 间接影响土壤有机质的分解和矿化速率, 而且土壤母质的风化速率以及N、P元素的淋溶强度也受降水和气温等气候因子的显著影响(
刘颖等, 2018 ); (3)不同的海拔梯度上森林植被受到的人为干扰程度也有一定的差异, 放牧、旅游、耕种等活动也会影响植物的元素分配格局(
Nogués-Bravo et al ., 2008 ).相关研究表明, 伴随着全球气候变化, 青藏高原地区已出现气温剧烈升高, 降水量增加的暖湿化趋势(
Chen et al ., 2013a ).因此, 未来在全球气候变化和区域N沉降可能加剧的背景下, 青海森林生态系统灌木层C、N、P化学计量特征将受到强烈影响. ...
塔里木盆地北缘绿洲土壤化学计量特征
1
2015
... 本研究中, 7种林分间SOC和TN含量呈现出明显差异, 而TP含量则差异不明显.这可能是由于受到营养元素的积累方式以及林分类型的影响, 林下土壤养分状况差异较大.在自然森林生态系统中, 森林土壤有机碳来自地表凋落物以及根系凋亡分解的积累, 而土壤全氮主要通过有机质分解矿化的途径累积(
李丹维等, 2017 ).首先, 林分的差异会影响地表凋落物、地下根系以及根系分泌物的组成, 同时凋落物的类型、数量以及分解速率是影响森林土壤养分含量变化和实现系统养分循环的重要因素(
杨佳佳等, 2014 ).其次, 林分的差异会影响土壤微生物、土壤动物和土壤酶的组成与功能多样性(
庞圣江等, 2015 ;
字洪标等, 2017 ), 而在土壤生物化学循环过程中, 土壤微生物通过氨化、硝化和反硝化等作用将土壤内颗粒态有机氮转化成可溶性有机氮, 而土壤酶作为催化剂也参与了其中复杂的生化反应(
李喜霞等, 2018 ).土壤P作为一种沉积性元素, 主要来源于岩石母质的风化, 其过程稳定而漫长, 因而在自然生态系统中P迁移能力较弱, 空间异质性较小(
李红林等, 2015 ).因此不同林分之间TP含量差异不明显. ...
塔里木盆地北缘绿洲土壤化学计量特征
1
2015
... 本研究中, 7种林分间SOC和TN含量呈现出明显差异, 而TP含量则差异不明显.这可能是由于受到营养元素的积累方式以及林分类型的影响, 林下土壤养分状况差异较大.在自然森林生态系统中, 森林土壤有机碳来自地表凋落物以及根系凋亡分解的积累, 而土壤全氮主要通过有机质分解矿化的途径累积(
李丹维等, 2017 ).首先, 林分的差异会影响地表凋落物、地下根系以及根系分泌物的组成, 同时凋落物的类型、数量以及分解速率是影响森林土壤养分含量变化和实现系统养分循环的重要因素(
杨佳佳等, 2014 ).其次, 林分的差异会影响土壤微生物、土壤动物和土壤酶的组成与功能多样性(
庞圣江等, 2015 ;
字洪标等, 2017 ), 而在土壤生物化学循环过程中, 土壤微生物通过氨化、硝化和反硝化等作用将土壤内颗粒态有机氮转化成可溶性有机氮, 而土壤酶作为催化剂也参与了其中复杂的生化反应(
李喜霞等, 2018 ).土壤P作为一种沉积性元素, 主要来源于岩石母质的风化, 其过程稳定而漫长, 因而在自然生态系统中P迁移能力较弱, 空间异质性较小(
李红林等, 2015 ).因此不同林分之间TP含量差异不明显. ...
中国南方灌丛优势木本植物叶的氮、磷含量及其影响因素
1
2017
... 本研究中, 灌木叶片C含量平均值为449.00 mg·g
-1 , 低于全球陆生植物叶片C含量464.20 mg·g
-1 (
Elser et al ., 2000 )和中国东部南北样带植物叶片C含量480.10 mg·g
-1 (
任书杰等, 2012 ), 说明青海森林的林下灌木植被C储存能力相对较弱.灌木叶片N、P含量平均值分别为19.46、1.69 mg·g
-1 , 略低于全球陆生植物N、P含量(20.10、1.77 mg·g
-1 (
Reich & Oleksyn, 2004 )和20.60、1.99 mg·g
-1 (
Elser et al ., 2000 )), 略低于我国植物叶片N含量(20.24 mg·g
-1 ), 高于其叶片P含量(1.46 mg·g
-1 )(
Han et al ., 2005 ), 高于我国南方优势灌丛优势植被N、P含量(16.57、1.02 mg·g
-1 )(
李家湘等, 2017 )、陕西森林的林下灌木叶片N、P含量(13.79、1.11 mg·g
-1 )(
姜沛沛等, 2016 ), 说明青海森林的林下灌木叶片N、P含量较高.这可能与本研究区域地处高寒区, 纬度偏高, 年降水量少, 年均气温低, 昼夜温差大, 日照强度大等特殊的区域气候条件有一定的关系(
卢航等, 2013 ).温度-植物生理假说认为, 低温诱导植物生长周期缩短, 同时, 植物趋向于分配更多的营养元素于光合器官中, 通过丰富叶片N、P含量在有限的生长周期内保持高效的光合速率以及酶活性来维持植物机体内正常的新陈代谢活动(
Reich & Oleksyn, 2004 ), 因此高海拔地区植物叶片N、P含量较为丰富.这与
杨阔等(2010) 发现青藏高原草地植物群落叶片N、P含量较高的结果一致. ...
中国南方灌丛优势木本植物叶的氮、磷含量及其影响因素
1
2017
... 本研究中, 灌木叶片C含量平均值为449.00 mg·g
-1 , 低于全球陆生植物叶片C含量464.20 mg·g
-1 (
Elser et al ., 2000 )和中国东部南北样带植物叶片C含量480.10 mg·g
-1 (
任书杰等, 2012 ), 说明青海森林的林下灌木植被C储存能力相对较弱.灌木叶片N、P含量平均值分别为19.46、1.69 mg·g
-1 , 略低于全球陆生植物N、P含量(20.10、1.77 mg·g
-1 (
Reich & Oleksyn, 2004 )和20.60、1.99 mg·g
-1 (
Elser et al ., 2000 )), 略低于我国植物叶片N含量(20.24 mg·g
-1 ), 高于其叶片P含量(1.46 mg·g
-1 )(
Han et al ., 2005 ), 高于我国南方优势灌丛优势植被N、P含量(16.57、1.02 mg·g
-1 )(
李家湘等, 2017 )、陕西森林的林下灌木叶片N、P含量(13.79、1.11 mg·g
-1 )(
姜沛沛等, 2016 ), 说明青海森林的林下灌木叶片N、P含量较高.这可能与本研究区域地处高寒区, 纬度偏高, 年降水量少, 年均气温低, 昼夜温差大, 日照强度大等特殊的区域气候条件有一定的关系(
卢航等, 2013 ).温度-植物生理假说认为, 低温诱导植物生长周期缩短, 同时, 植物趋向于分配更多的营养元素于光合器官中, 通过丰富叶片N、P含量在有限的生长周期内保持高效的光合速率以及酶活性来维持植物机体内正常的新陈代谢活动(
Reich & Oleksyn, 2004 ), 因此高海拔地区植物叶片N、P含量较为丰富.这与
杨阔等(2010) 发现青藏高原草地植物群落叶片N、P含量较高的结果一致. ...
灌木的生态特性与生态效能的研究与进展
1
2006
... 在森林生态系统中, 林下灌木层与乔木层和草本层共同维持着森林生态系统结构与功能的稳定, 在参与养分循环、改善土壤肥力、为林下生物提供栖息环境和提高生态系统多样性等方面起到重要作用, 是森林生态系统不可或缺的组成部分(
郑绍伟等, 2007 ;
卢振龙和龚孝生, 2009 ).灌木植被通常具有木质化茎干而主干不明显, 其茎干多分枝, 树冠矮小, 根系分布广而深(
李清河等, 2006 ).目前, 对森林的林下灌木层的研究主要聚焦于灌木生物量生长模型的建立(
万五星等, 2014 ), 物种多样性和种间联结性(
崔宁洁等, 2014 ), 生态位以及物种分布特征的分析(
康永祥等, 2008 ).对于林下灌木层生态化学计量特征的研究相对较少, 尤其是高寒地区不同林分类型中灌木层的研究尚鲜有报道. ...
灌木的生态特性与生态效能的研究与进展
1
2006
... 在森林生态系统中, 林下灌木层与乔木层和草本层共同维持着森林生态系统结构与功能的稳定, 在参与养分循环、改善土壤肥力、为林下生物提供栖息环境和提高生态系统多样性等方面起到重要作用, 是森林生态系统不可或缺的组成部分(
郑绍伟等, 2007 ;
卢振龙和龚孝生, 2009 ).灌木植被通常具有木质化茎干而主干不明显, 其茎干多分枝, 树冠矮小, 根系分布广而深(
李清河等, 2006 ).目前, 对森林的林下灌木层的研究主要聚焦于灌木生物量生长模型的建立(
万五星等, 2014 ), 物种多样性和种间联结性(
崔宁洁等, 2014 ), 生态位以及物种分布特征的分析(
康永祥等, 2008 ).对于林下灌木层生态化学计量特征的研究相对较少, 尤其是高寒地区不同林分类型中灌木层的研究尚鲜有报道. ...
黄土高原纬度梯度上的植物与土壤碳、氮、磷化学计量学特征
2
2015
... 针对青海省森林生态系统分布状况, 并结合全省各森林类型分布面积与蓄积比重、林龄、起源等情况, 于植物生长旺盛季(7-8月)在全省21个县范围内选择80个标准样地(
图1 ), 同时记录该样地坡度、海拔、坡向、郁闭度等环境因子, 在每个标准样地中随机设置3块50 m × 20 m的乔木样方, 各样方间距大于100 m, 总计240块乔木样方.在上述乔木样方内采用对角线设置3个2 m × 2 m灌木样方, 记录灌木名称、株(丛)数、总盖度、平均高度和平均基径(附录I).将样方内的灌木植被全部收获, 同时按灌木不同部位(叶、枝干、根)进行分类混合后带回实验室, 置于恒温烘箱中65 ℃烘干至恒质量, 样品预处理后测定C、N、P含量.在每个乔木调查样方内采用对角线设置3个1 m × 1 m土壤调查样方, 在每个土壤调查样方内用土钻法(内径5 cm)分5层(0-10、10-20、20-30、30-50、50-100 cm)分别钻取土壤样品(不够100 cm至基岩为止), 同一层土壤样品混合为1个土壤样品.土壤样品预处理后测定土壤有机碳(SOC)、全氮(TN)、全磷(TP)含量.由于森林生态系统表层土壤(0-10 cm)具有表聚性, 其SOC、TN、TP含量从表层向下层形成垂直递减的分布格局(
李婷等, 2015 ), 所以本研究选取该土层来探讨土壤化学计量特征. ...
... 相关性分析发现, 林下灌木(叶、枝干、根) N含量、C:N、N:P与土壤TN含量、C:N、N:P呈极显著相关关系, 而P含量、C:P与土壤TP含量呈显著相关关系, 表明植物体内各器官元素分配时相互协调, 且N、P吸收效率与土壤组分紧密相关.这是因为植物通过叶片凋落物和根系凋亡形成土壤有机质, 土壤有机质分解矿化为植物正常代谢活动提供必要的营养元素(
毕建华等, 2017 ).植物根系吸收和运输作为土壤N、P输出的主要途径, 使得植物与土壤在养分需求和供应之间达到动态平衡, 形成生态系统N、P转化的有效循环过程(
李婷等, 2015 ). ...
黄土高原纬度梯度上的植物与土壤碳、氮、磷化学计量学特征
2
2015
... 针对青海省森林生态系统分布状况, 并结合全省各森林类型分布面积与蓄积比重、林龄、起源等情况, 于植物生长旺盛季(7-8月)在全省21个县范围内选择80个标准样地(
图1 ), 同时记录该样地坡度、海拔、坡向、郁闭度等环境因子, 在每个标准样地中随机设置3块50 m × 20 m的乔木样方, 各样方间距大于100 m, 总计240块乔木样方.在上述乔木样方内采用对角线设置3个2 m × 2 m灌木样方, 记录灌木名称、株(丛)数、总盖度、平均高度和平均基径(附录I).将样方内的灌木植被全部收获, 同时按灌木不同部位(叶、枝干、根)进行分类混合后带回实验室, 置于恒温烘箱中65 ℃烘干至恒质量, 样品预处理后测定C、N、P含量.在每个乔木调查样方内采用对角线设置3个1 m × 1 m土壤调查样方, 在每个土壤调查样方内用土钻法(内径5 cm)分5层(0-10、10-20、20-30、30-50、50-100 cm)分别钻取土壤样品(不够100 cm至基岩为止), 同一层土壤样品混合为1个土壤样品.土壤样品预处理后测定土壤有机碳(SOC)、全氮(TN)、全磷(TP)含量.由于森林生态系统表层土壤(0-10 cm)具有表聚性, 其SOC、TN、TP含量从表层向下层形成垂直递减的分布格局(
李婷等, 2015 ), 所以本研究选取该土层来探讨土壤化学计量特征. ...
... 相关性分析发现, 林下灌木(叶、枝干、根) N含量、C:N、N:P与土壤TN含量、C:N、N:P呈极显著相关关系, 而P含量、C:P与土壤TP含量呈显著相关关系, 表明植物体内各器官元素分配时相互协调, 且N、P吸收效率与土壤组分紧密相关.这是因为植物通过叶片凋落物和根系凋亡形成土壤有机质, 土壤有机质分解矿化为植物正常代谢活动提供必要的营养元素(
毕建华等, 2017 ).植物根系吸收和运输作为土壤N、P输出的主要途径, 使得植物与土壤在养分需求和供应之间达到动态平衡, 形成生态系统N、P转化的有效循环过程(
李婷等, 2015 ). ...
阔叶红松林生态化学计量学特征及其对纬度梯度的响应
1
2018
... 本研究中, 7种林分间SOC和TN含量呈现出明显差异, 而TP含量则差异不明显.这可能是由于受到营养元素的积累方式以及林分类型的影响, 林下土壤养分状况差异较大.在自然森林生态系统中, 森林土壤有机碳来自地表凋落物以及根系凋亡分解的积累, 而土壤全氮主要通过有机质分解矿化的途径累积(
李丹维等, 2017 ).首先, 林分的差异会影响地表凋落物、地下根系以及根系分泌物的组成, 同时凋落物的类型、数量以及分解速率是影响森林土壤养分含量变化和实现系统养分循环的重要因素(
杨佳佳等, 2014 ).其次, 林分的差异会影响土壤微生物、土壤动物和土壤酶的组成与功能多样性(
庞圣江等, 2015 ;
字洪标等, 2017 ), 而在土壤生物化学循环过程中, 土壤微生物通过氨化、硝化和反硝化等作用将土壤内颗粒态有机氮转化成可溶性有机氮, 而土壤酶作为催化剂也参与了其中复杂的生化反应(
李喜霞等, 2018 ).土壤P作为一种沉积性元素, 主要来源于岩石母质的风化, 其过程稳定而漫长, 因而在自然生态系统中P迁移能力较弱, 空间异质性较小(
李红林等, 2015 ).因此不同林分之间TP含量差异不明显. ...
阔叶红松林生态化学计量学特征及其对纬度梯度的响应
1
2018
... 本研究中, 7种林分间SOC和TN含量呈现出明显差异, 而TP含量则差异不明显.这可能是由于受到营养元素的积累方式以及林分类型的影响, 林下土壤养分状况差异较大.在自然森林生态系统中, 森林土壤有机碳来自地表凋落物以及根系凋亡分解的积累, 而土壤全氮主要通过有机质分解矿化的途径累积(
李丹维等, 2017 ).首先, 林分的差异会影响地表凋落物、地下根系以及根系分泌物的组成, 同时凋落物的类型、数量以及分解速率是影响森林土壤养分含量变化和实现系统养分循环的重要因素(
杨佳佳等, 2014 ).其次, 林分的差异会影响土壤微生物、土壤动物和土壤酶的组成与功能多样性(
庞圣江等, 2015 ;
字洪标等, 2017 ), 而在土壤生物化学循环过程中, 土壤微生物通过氨化、硝化和反硝化等作用将土壤内颗粒态有机氮转化成可溶性有机氮, 而土壤酶作为催化剂也参与了其中复杂的生化反应(
李喜霞等, 2018 ).土壤P作为一种沉积性元素, 主要来源于岩石母质的风化, 其过程稳定而漫长, 因而在自然生态系统中P迁移能力较弱, 空间异质性较小(
李红林等, 2015 ).因此不同林分之间TP含量差异不明显. ...
川西高寒灌丛草地不同海拔梯度土壤化学计量特征
1
2018
... 依据RDA发现, 林下灌木C、N、P含量及化学计量特征主要受土壤C:N、海拔、年降水量和年平均气温的影响, 这表明青海高寒区森林的林下灌木受到土壤N有效性、海拔以及区域水热条件的重要调控.土壤作为植被获取营养元素的主要机质, 其N、P有效性将显著影响地上植被组成与结构, 生产力以及有机体内元素分配格局(
李丹维等, 2017 ).
McGroddy等(2004) 认为, 土壤N、P有效性是植物有机体内元素含量的主要驱动因素, 同时土壤N、P有效性容易受降水淋溶作用的影响.
张仁懿等(2014) 研究也表明青藏高原亚高寒草甸植物养分状况受土壤N有效性的显著影响, 土壤N有效性对高寒植物生长的影响比其他元素更为关键.海拔梯度综合了温度、降水、光照等水热条件的变化, 是影响植被生长的主导因子(
Guo et al ., 2010 ).本研究结果与木本植物叶片N、P元素含量受年平均气温和年降水量共同驱动作用的结果(
Chen et al ., 2013b )相符.因为:(1)海拔梯度上降水、气温的差异会影响植物的生长周期, 从而改变C、N、P在各器官间的分配(
Kerkhoff et al ., 2005 ); (2)海拔梯度上土壤温度和水分的变化将影响土壤微生物的群系特征及代谢效率, 间接影响土壤有机质的分解和矿化速率, 而且土壤母质的风化速率以及N、P元素的淋溶强度也受降水和气温等气候因子的显著影响(
刘颖等, 2018 ); (3)不同的海拔梯度上森林植被受到的人为干扰程度也有一定的差异, 放牧、旅游、耕种等活动也会影响植物的元素分配格局(
Nogués-Bravo et al ., 2008 ).相关研究表明, 伴随着全球气候变化, 青藏高原地区已出现气温剧烈升高, 降水量增加的暖湿化趋势(
Chen et al ., 2013a ).因此, 未来在全球气候变化和区域N沉降可能加剧的背景下, 青海森林生态系统灌木层C、N、P化学计量特征将受到强烈影响. ...
川西高寒灌丛草地不同海拔梯度土壤化学计量特征
1
2018
... 依据RDA发现, 林下灌木C、N、P含量及化学计量特征主要受土壤C:N、海拔、年降水量和年平均气温的影响, 这表明青海高寒区森林的林下灌木受到土壤N有效性、海拔以及区域水热条件的重要调控.土壤作为植被获取营养元素的主要机质, 其N、P有效性将显著影响地上植被组成与结构, 生产力以及有机体内元素分配格局(
李丹维等, 2017 ).
McGroddy等(2004) 认为, 土壤N、P有效性是植物有机体内元素含量的主要驱动因素, 同时土壤N、P有效性容易受降水淋溶作用的影响.
张仁懿等(2014) 研究也表明青藏高原亚高寒草甸植物养分状况受土壤N有效性的显著影响, 土壤N有效性对高寒植物生长的影响比其他元素更为关键.海拔梯度综合了温度、降水、光照等水热条件的变化, 是影响植被生长的主导因子(
Guo et al ., 2010 ).本研究结果与木本植物叶片N、P元素含量受年平均气温和年降水量共同驱动作用的结果(
Chen et al ., 2013b )相符.因为:(1)海拔梯度上降水、气温的差异会影响植物的生长周期, 从而改变C、N、P在各器官间的分配(
Kerkhoff et al ., 2005 ); (2)海拔梯度上土壤温度和水分的变化将影响土壤微生物的群系特征及代谢效率, 间接影响土壤有机质的分解和矿化速率, 而且土壤母质的风化速率以及N、P元素的淋溶强度也受降水和气温等气候因子的显著影响(
刘颖等, 2018 ); (3)不同的海拔梯度上森林植被受到的人为干扰程度也有一定的差异, 放牧、旅游、耕种等活动也会影响植物的元素分配格局(
Nogués-Bravo et al ., 2008 ).相关研究表明, 伴随着全球气候变化, 青藏高原地区已出现气温剧烈升高, 降水量增加的暖湿化趋势(
Chen et al ., 2013a ).因此, 未来在全球气候变化和区域N沉降可能加剧的背景下, 青海森林生态系统灌木层C、N、P化学计量特征将受到强烈影响. ...
青海省近20年森林植被碳储量变化及其现状分析
1
2013
... 本研究中, 灌木叶片C含量平均值为449.00 mg·g
-1 , 低于全球陆生植物叶片C含量464.20 mg·g
-1 (
Elser et al ., 2000 )和中国东部南北样带植物叶片C含量480.10 mg·g
-1 (
任书杰等, 2012 ), 说明青海森林的林下灌木植被C储存能力相对较弱.灌木叶片N、P含量平均值分别为19.46、1.69 mg·g
-1 , 略低于全球陆生植物N、P含量(20.10、1.77 mg·g
-1 (
Reich & Oleksyn, 2004 )和20.60、1.99 mg·g
-1 (
Elser et al ., 2000 )), 略低于我国植物叶片N含量(20.24 mg·g
-1 ), 高于其叶片P含量(1.46 mg·g
-1 )(
Han et al ., 2005 ), 高于我国南方优势灌丛优势植被N、P含量(16.57、1.02 mg·g
-1 )(
李家湘等, 2017 )、陕西森林的林下灌木叶片N、P含量(13.79、1.11 mg·g
-1 )(
姜沛沛等, 2016 ), 说明青海森林的林下灌木叶片N、P含量较高.这可能与本研究区域地处高寒区, 纬度偏高, 年降水量少, 年均气温低, 昼夜温差大, 日照强度大等特殊的区域气候条件有一定的关系(
卢航等, 2013 ).温度-植物生理假说认为, 低温诱导植物生长周期缩短, 同时, 植物趋向于分配更多的营养元素于光合器官中, 通过丰富叶片N、P含量在有限的生长周期内保持高效的光合速率以及酶活性来维持植物机体内正常的新陈代谢活动(
Reich & Oleksyn, 2004 ), 因此高海拔地区植物叶片N、P含量较为丰富.这与
杨阔等(2010) 发现青藏高原草地植物群落叶片N、P含量较高的结果一致. ...
青海省近20年森林植被碳储量变化及其现状分析
1
2013
... 本研究中, 灌木叶片C含量平均值为449.00 mg·g
-1 , 低于全球陆生植物叶片C含量464.20 mg·g
-1 (
Elser et al ., 2000 )和中国东部南北样带植物叶片C含量480.10 mg·g
-1 (
任书杰等, 2012 ), 说明青海森林的林下灌木植被C储存能力相对较弱.灌木叶片N、P含量平均值分别为19.46、1.69 mg·g
-1 , 略低于全球陆生植物N、P含量(20.10、1.77 mg·g
-1 (
Reich & Oleksyn, 2004 )和20.60、1.99 mg·g
-1 (
Elser et al ., 2000 )), 略低于我国植物叶片N含量(20.24 mg·g
-1 ), 高于其叶片P含量(1.46 mg·g
-1 )(
Han et al ., 2005 ), 高于我国南方优势灌丛优势植被N、P含量(16.57、1.02 mg·g
-1 )(
李家湘等, 2017 )、陕西森林的林下灌木叶片N、P含量(13.79、1.11 mg·g
-1 )(
姜沛沛等, 2016 ), 说明青海森林的林下灌木叶片N、P含量较高.这可能与本研究区域地处高寒区, 纬度偏高, 年降水量少, 年均气温低, 昼夜温差大, 日照强度大等特殊的区域气候条件有一定的关系(
卢航等, 2013 ).温度-植物生理假说认为, 低温诱导植物生长周期缩短, 同时, 植物趋向于分配更多的营养元素于光合器官中, 通过丰富叶片N、P含量在有限的生长周期内保持高效的光合速率以及酶活性来维持植物机体内正常的新陈代谢活动(
Reich & Oleksyn, 2004 ), 因此高海拔地区植物叶片N、P含量较为丰富.这与
杨阔等(2010) 发现青藏高原草地植物群落叶片N、P含量较高的结果一致. ...
典型自然带土壤氮磷化学计量空间分异特征及其驱动因素研究
1
2017
... 本研究区森林SOC、TN、TP含量平均值为(95.11、5.17、0.82 mg·g
-1 ), 高于全国土壤C、N、P含量(24.56、1.88、0.78 mg·g
-1 )(
Tian et al ., 2010 ), 与青藏高原高寒植被带土壤N、P含量(5.66、0.75 mg·g
-1 )相接近(
卢同平等, 2017 ).土壤C:N、C:P (20.6、117.7)高于全球森林土壤C:N、C:P (12.4、81.9)(
Cleveland & Liptzin, 2007 )和全国土壤C:N、C:P (12.3、52.7)(
Tian et al ., 2010 ).研究表明, 土壤C:N、C:P是衡量土壤有机质分解矿化速率的重要指标, 通常土壤C:N、C:P与有机质分解矿化速率呈反比关系, 较高的C:N、C:P代表较慢的有机质分解速率, 以及较低的土壤N、P有效性(
Cleveland & Liptzin, 2007 ).由此可推断, 青海高寒区森林SOC、TN、TP储备丰富, 有机质分解和矿化速率较慢, N、P有效性较低.这可能与本研究区域自然环境和植被类型特性有关, 青海高寒地区年降水量偏少、年平均气温低, 森林地表积累了丰富的凋落物, 长期的腐殖化作用有利于土壤有机质的富集, 较低的土壤温度抑制了土壤微生物及土壤酶活性, 地表有机质的分解周期长, 系统累积量大于消耗量, 导致森林土壤N、P元素得以累积, 但缓慢的分解速率导致土壤N、P有效性较低(
高巧等, 2014 ;
王淑彬等, 2014 ). ...
典型自然带土壤氮磷化学计量空间分异特征及其驱动因素研究
1
2017
... 本研究区森林SOC、TN、TP含量平均值为(95.11、5.17、0.82 mg·g
-1 ), 高于全国土壤C、N、P含量(24.56、1.88、0.78 mg·g
-1 )(
Tian et al ., 2010 ), 与青藏高原高寒植被带土壤N、P含量(5.66、0.75 mg·g
-1 )相接近(
卢同平等, 2017 ).土壤C:N、C:P (20.6、117.7)高于全球森林土壤C:N、C:P (12.4、81.9)(
Cleveland & Liptzin, 2007 )和全国土壤C:N、C:P (12.3、52.7)(
Tian et al ., 2010 ).研究表明, 土壤C:N、C:P是衡量土壤有机质分解矿化速率的重要指标, 通常土壤C:N、C:P与有机质分解矿化速率呈反比关系, 较高的C:N、C:P代表较慢的有机质分解速率, 以及较低的土壤N、P有效性(
Cleveland & Liptzin, 2007 ).由此可推断, 青海高寒区森林SOC、TN、TP储备丰富, 有机质分解和矿化速率较慢, N、P有效性较低.这可能与本研究区域自然环境和植被类型特性有关, 青海高寒地区年降水量偏少、年平均气温低, 森林地表积累了丰富的凋落物, 长期的腐殖化作用有利于土壤有机质的富集, 较低的土壤温度抑制了土壤微生物及土壤酶活性, 地表有机质的分解周期长, 系统累积量大于消耗量, 导致森林土壤N、P元素得以累积, 但缓慢的分解速率导致土壤N、P有效性较低(
高巧等, 2014 ;
王淑彬等, 2014 ). ...
灌木生物量测定的研究进展
1
2009
... 在森林生态系统中, 林下灌木层与乔木层和草本层共同维持着森林生态系统结构与功能的稳定, 在参与养分循环、改善土壤肥力、为林下生物提供栖息环境和提高生态系统多样性等方面起到重要作用, 是森林生态系统不可或缺的组成部分(
郑绍伟等, 2007 ;
卢振龙和龚孝生, 2009 ).灌木植被通常具有木质化茎干而主干不明显, 其茎干多分枝, 树冠矮小, 根系分布广而深(
李清河等, 2006 ).目前, 对森林的林下灌木层的研究主要聚焦于灌木生物量生长模型的建立(
万五星等, 2014 ), 物种多样性和种间联结性(
崔宁洁等, 2014 ), 生态位以及物种分布特征的分析(
康永祥等, 2008 ).对于林下灌木层生态化学计量特征的研究相对较少, 尤其是高寒地区不同林分类型中灌木层的研究尚鲜有报道. ...
灌木生物量测定的研究进展
1
2009
... 在森林生态系统中, 林下灌木层与乔木层和草本层共同维持着森林生态系统结构与功能的稳定, 在参与养分循环、改善土壤肥力、为林下生物提供栖息环境和提高生态系统多样性等方面起到重要作用, 是森林生态系统不可或缺的组成部分(
郑绍伟等, 2007 ;
卢振龙和龚孝生, 2009 ).灌木植被通常具有木质化茎干而主干不明显, 其茎干多分枝, 树冠矮小, 根系分布广而深(
李清河等, 2006 ).目前, 对森林的林下灌木层的研究主要聚焦于灌木生物量生长模型的建立(
万五星等, 2014 ), 物种多样性和种间联结性(
崔宁洁等, 2014 ), 生态位以及物种分布特征的分析(
康永祥等, 2008 ).对于林下灌木层生态化学计量特征的研究相对较少, 尤其是高寒地区不同林分类型中灌木层的研究尚鲜有报道. ...
Scaling of C:N:P stoichiometry in forest worldwide: Implications of terrestrial Redfield-type ratios
1
2004
... 依据RDA发现, 林下灌木C、N、P含量及化学计量特征主要受土壤C:N、海拔、年降水量和年平均气温的影响, 这表明青海高寒区森林的林下灌木受到土壤N有效性、海拔以及区域水热条件的重要调控.土壤作为植被获取营养元素的主要机质, 其N、P有效性将显著影响地上植被组成与结构, 生产力以及有机体内元素分配格局(
李丹维等, 2017 ).
McGroddy等(2004) 认为, 土壤N、P有效性是植物有机体内元素含量的主要驱动因素, 同时土壤N、P有效性容易受降水淋溶作用的影响.
张仁懿等(2014) 研究也表明青藏高原亚高寒草甸植物养分状况受土壤N有效性的显著影响, 土壤N有效性对高寒植物生长的影响比其他元素更为关键.海拔梯度综合了温度、降水、光照等水热条件的变化, 是影响植被生长的主导因子(
Guo et al ., 2010 ).本研究结果与木本植物叶片N、P元素含量受年平均气温和年降水量共同驱动作用的结果(
Chen et al ., 2013b )相符.因为:(1)海拔梯度上降水、气温的差异会影响植物的生长周期, 从而改变C、N、P在各器官间的分配(
Kerkhoff et al ., 2005 ); (2)海拔梯度上土壤温度和水分的变化将影响土壤微生物的群系特征及代谢效率, 间接影响土壤有机质的分解和矿化速率, 而且土壤母质的风化速率以及N、P元素的淋溶强度也受降水和气温等气候因子的显著影响(
刘颖等, 2018 ); (3)不同的海拔梯度上森林植被受到的人为干扰程度也有一定的差异, 放牧、旅游、耕种等活动也会影响植物的元素分配格局(
Nogués-Bravo et al ., 2008 ).相关研究表明, 伴随着全球气候变化, 青藏高原地区已出现气温剧烈升高, 降水量增加的暖湿化趋势(
Chen et al ., 2013a ).因此, 未来在全球气候变化和区域N沉降可能加剧的背景下, 青海森林生态系统灌木层C、N、P化学计量特征将受到强烈影响. ...
Scale effects and human impact on the elevational species richness gradients
1
2008
... 依据RDA发现, 林下灌木C、N、P含量及化学计量特征主要受土壤C:N、海拔、年降水量和年平均气温的影响, 这表明青海高寒区森林的林下灌木受到土壤N有效性、海拔以及区域水热条件的重要调控.土壤作为植被获取营养元素的主要机质, 其N、P有效性将显著影响地上植被组成与结构, 生产力以及有机体内元素分配格局(
李丹维等, 2017 ).
McGroddy等(2004) 认为, 土壤N、P有效性是植物有机体内元素含量的主要驱动因素, 同时土壤N、P有效性容易受降水淋溶作用的影响.
张仁懿等(2014) 研究也表明青藏高原亚高寒草甸植物养分状况受土壤N有效性的显著影响, 土壤N有效性对高寒植物生长的影响比其他元素更为关键.海拔梯度综合了温度、降水、光照等水热条件的变化, 是影响植被生长的主导因子(
Guo et al ., 2010 ).本研究结果与木本植物叶片N、P元素含量受年平均气温和年降水量共同驱动作用的结果(
Chen et al ., 2013b )相符.因为:(1)海拔梯度上降水、气温的差异会影响植物的生长周期, 从而改变C、N、P在各器官间的分配(
Kerkhoff et al ., 2005 ); (2)海拔梯度上土壤温度和水分的变化将影响土壤微生物的群系特征及代谢效率, 间接影响土壤有机质的分解和矿化速率, 而且土壤母质的风化速率以及N、P元素的淋溶强度也受降水和气温等气候因子的显著影响(
刘颖等, 2018 ); (3)不同的海拔梯度上森林植被受到的人为干扰程度也有一定的差异, 放牧、旅游、耕种等活动也会影响植物的元素分配格局(
Nogués-Bravo et al ., 2008 ).相关研究表明, 伴随着全球气候变化, 青藏高原地区已出现气温剧烈升高, 降水量增加的暖湿化趋势(
Chen et al ., 2013a ).因此, 未来在全球气候变化和区域N沉降可能加剧的背景下, 青海森林生态系统灌木层C、N、P化学计量特征将受到强烈影响. ...
桂西北不同森林类型土壤生态化学计量特征
1
2015
... 本研究中, 7种林分间SOC和TN含量呈现出明显差异, 而TP含量则差异不明显.这可能是由于受到营养元素的积累方式以及林分类型的影响, 林下土壤养分状况差异较大.在自然森林生态系统中, 森林土壤有机碳来自地表凋落物以及根系凋亡分解的积累, 而土壤全氮主要通过有机质分解矿化的途径累积(
李丹维等, 2017 ).首先, 林分的差异会影响地表凋落物、地下根系以及根系分泌物的组成, 同时凋落物的类型、数量以及分解速率是影响森林土壤养分含量变化和实现系统养分循环的重要因素(
杨佳佳等, 2014 ).其次, 林分的差异会影响土壤微生物、土壤动物和土壤酶的组成与功能多样性(
庞圣江等, 2015 ;
字洪标等, 2017 ), 而在土壤生物化学循环过程中, 土壤微生物通过氨化、硝化和反硝化等作用将土壤内颗粒态有机氮转化成可溶性有机氮, 而土壤酶作为催化剂也参与了其中复杂的生化反应(
李喜霞等, 2018 ).土壤P作为一种沉积性元素, 主要来源于岩石母质的风化, 其过程稳定而漫长, 因而在自然生态系统中P迁移能力较弱, 空间异质性较小(
李红林等, 2015 ).因此不同林分之间TP含量差异不明显. ...
桂西北不同森林类型土壤生态化学计量特征
1
2015
... 本研究中, 7种林分间SOC和TN含量呈现出明显差异, 而TP含量则差异不明显.这可能是由于受到营养元素的积累方式以及林分类型的影响, 林下土壤养分状况差异较大.在自然森林生态系统中, 森林土壤有机碳来自地表凋落物以及根系凋亡分解的积累, 而土壤全氮主要通过有机质分解矿化的途径累积(
李丹维等, 2017 ).首先, 林分的差异会影响地表凋落物、地下根系以及根系分泌物的组成, 同时凋落物的类型、数量以及分解速率是影响森林土壤养分含量变化和实现系统养分循环的重要因素(
杨佳佳等, 2014 ).其次, 林分的差异会影响土壤微生物、土壤动物和土壤酶的组成与功能多样性(
庞圣江等, 2015 ;
字洪标等, 2017 ), 而在土壤生物化学循环过程中, 土壤微生物通过氨化、硝化和反硝化等作用将土壤内颗粒态有机氮转化成可溶性有机氮, 而土壤酶作为催化剂也参与了其中复杂的生化反应(
李喜霞等, 2018 ).土壤P作为一种沉积性元素, 主要来源于岩石母质的风化, 其过程稳定而漫长, 因而在自然生态系统中P迁移能力较弱, 空间异质性较小(
李红林等, 2015 ).因此不同林分之间TP含量差异不明显. ...
黄土丘陵区不同功能群植物碳氮磷生态化学计量特征及其对微地形的响应
1
2016
... 碳(C)、氮(N)、磷(P)作为最重要的生命元素, 是维系地球上所有生命活动的基础(
贺金生和韩兴国, 2010 ).C作为植物有机体结构的重要组成元素, 是植物有机体内生理生化反应过程的能量来源之一(
Güsewell, 2004 ), N、P是植物的基本营养元素, 也是植物有机体内蛋白质、酶类、核酸和脂质的重要组成元素(
Yang & Wang, 2011 ;
戚德辉等, 2016 ;
Tang et al ., 2018 ).C、N、P含量及其化学计量比的差异显著影响植物的生态策略, 从而被广泛应用到生态系统在生物地球化学循环和生态过程耦合关系的研究中(
Elser et al ., 1996 ).截至目前, 国内外****已在区域乃至全球范围开展了许多陆地生态系统生态化学计量特征的研究(
Han et al ., 2005 ;
Elser et al ., 2007 ;
黄小波等, 2016 ).但主要集中在植物叶片和土壤生态化学计量特征方面, 针对植物枝干、根等重要器官方面的研究相对较少(
贺合亮等, 2017 ).枝干是植物连接地下吸收器官和地上同化组织的传导器官, 具有支撑植物体的结构性骨架作用(
李单凤等, 2015 ).根系作为吸收、存储、运输水分和养分的主要器官, 也是植物有机体对外界环境变化响应最为敏感的地下器官, 在森林生态系统物质循环过程中发挥关键作用(
Iversen et al ., 2017 ;
陈晓萍等, 2018 ).因此, 研究植物不同器官间生态化学计量特征以及与土壤组分在各生态过程间的互作关系, 对于揭示生态系统养分循环规律具有重要意义. ...
黄土丘陵区不同功能群植物碳氮磷生态化学计量特征及其对微地形的响应
1
2016
... 碳(C)、氮(N)、磷(P)作为最重要的生命元素, 是维系地球上所有生命活动的基础(
贺金生和韩兴国, 2010 ).C作为植物有机体结构的重要组成元素, 是植物有机体内生理生化反应过程的能量来源之一(
Güsewell, 2004 ), N、P是植物的基本营养元素, 也是植物有机体内蛋白质、酶类、核酸和脂质的重要组成元素(
Yang & Wang, 2011 ;
戚德辉等, 2016 ;
Tang et al ., 2018 ).C、N、P含量及其化学计量比的差异显著影响植物的生态策略, 从而被广泛应用到生态系统在生物地球化学循环和生态过程耦合关系的研究中(
Elser et al ., 1996 ).截至目前, 国内外****已在区域乃至全球范围开展了许多陆地生态系统生态化学计量特征的研究(
Han et al ., 2005 ;
Elser et al ., 2007 ;
黄小波等, 2016 ).但主要集中在植物叶片和土壤生态化学计量特征方面, 针对植物枝干、根等重要器官方面的研究相对较少(
贺合亮等, 2017 ).枝干是植物连接地下吸收器官和地上同化组织的传导器官, 具有支撑植物体的结构性骨架作用(
李单凤等, 2015 ).根系作为吸收、存储、运输水分和养分的主要器官, 也是植物有机体对外界环境变化响应最为敏感的地下器官, 在森林生态系统物质循环过程中发挥关键作用(
Iversen et al ., 2017 ;
陈晓萍等, 2018 ).因此, 研究植物不同器官间生态化学计量特征以及与土壤组分在各生态过程间的互作关系, 对于揭示生态系统养分循环规律具有重要意义. ...
Global patterns of plant leaf N and P in relation to temperature and latitude
2
2004
... 本研究中, 灌木叶片C含量平均值为449.00 mg·g
-1 , 低于全球陆生植物叶片C含量464.20 mg·g
-1 (
Elser et al ., 2000 )和中国东部南北样带植物叶片C含量480.10 mg·g
-1 (
任书杰等, 2012 ), 说明青海森林的林下灌木植被C储存能力相对较弱.灌木叶片N、P含量平均值分别为19.46、1.69 mg·g
-1 , 略低于全球陆生植物N、P含量(20.10、1.77 mg·g
-1 (
Reich & Oleksyn, 2004 )和20.60、1.99 mg·g
-1 (
Elser et al ., 2000 )), 略低于我国植物叶片N含量(20.24 mg·g
-1 ), 高于其叶片P含量(1.46 mg·g
-1 )(
Han et al ., 2005 ), 高于我国南方优势灌丛优势植被N、P含量(16.57、1.02 mg·g
-1 )(
李家湘等, 2017 )、陕西森林的林下灌木叶片N、P含量(13.79、1.11 mg·g
-1 )(
姜沛沛等, 2016 ), 说明青海森林的林下灌木叶片N、P含量较高.这可能与本研究区域地处高寒区, 纬度偏高, 年降水量少, 年均气温低, 昼夜温差大, 日照强度大等特殊的区域气候条件有一定的关系(
卢航等, 2013 ).温度-植物生理假说认为, 低温诱导植物生长周期缩短, 同时, 植物趋向于分配更多的营养元素于光合器官中, 通过丰富叶片N、P含量在有限的生长周期内保持高效的光合速率以及酶活性来维持植物机体内正常的新陈代谢活动(
Reich & Oleksyn, 2004 ), 因此高海拔地区植物叶片N、P含量较为丰富.这与
杨阔等(2010) 发现青藏高原草地植物群落叶片N、P含量较高的结果一致. ...
... ).温度-植物生理假说认为, 低温诱导植物生长周期缩短, 同时, 植物趋向于分配更多的营养元素于光合器官中, 通过丰富叶片N、P含量在有限的生长周期内保持高效的光合速率以及酶活性来维持植物机体内正常的新陈代谢活动(
Reich & Oleksyn, 2004 ), 因此高海拔地区植物叶片N、P含量较为丰富.这与
杨阔等(2010) 发现青藏高原草地植物群落叶片N、P含量较高的结果一致. ...
中国东部南北样带森林生态系统102个优势种叶片碳氮磷化学计量学统计特征
1
2012
... 本研究中, 灌木叶片C含量平均值为449.00 mg·g
-1 , 低于全球陆生植物叶片C含量464.20 mg·g
-1 (
Elser et al ., 2000 )和中国东部南北样带植物叶片C含量480.10 mg·g
-1 (
任书杰等, 2012 ), 说明青海森林的林下灌木植被C储存能力相对较弱.灌木叶片N、P含量平均值分别为19.46、1.69 mg·g
-1 , 略低于全球陆生植物N、P含量(20.10、1.77 mg·g
-1 (
Reich & Oleksyn, 2004 )和20.60、1.99 mg·g
-1 (
Elser et al ., 2000 )), 略低于我国植物叶片N含量(20.24 mg·g
-1 ), 高于其叶片P含量(1.46 mg·g
-1 )(
Han et al ., 2005 ), 高于我国南方优势灌丛优势植被N、P含量(16.57、1.02 mg·g
-1 )(
李家湘等, 2017 )、陕西森林的林下灌木叶片N、P含量(13.79、1.11 mg·g
-1 )(
姜沛沛等, 2016 ), 说明青海森林的林下灌木叶片N、P含量较高.这可能与本研究区域地处高寒区, 纬度偏高, 年降水量少, 年均气温低, 昼夜温差大, 日照强度大等特殊的区域气候条件有一定的关系(
卢航等, 2013 ).温度-植物生理假说认为, 低温诱导植物生长周期缩短, 同时, 植物趋向于分配更多的营养元素于光合器官中, 通过丰富叶片N、P含量在有限的生长周期内保持高效的光合速率以及酶活性来维持植物机体内正常的新陈代谢活动(
Reich & Oleksyn, 2004 ), 因此高海拔地区植物叶片N、P含量较为丰富.这与
杨阔等(2010) 发现青藏高原草地植物群落叶片N、P含量较高的结果一致. ...
中国东部南北样带森林生态系统102个优势种叶片碳氮磷化学计量学统计特征
1
2012
... 本研究中, 灌木叶片C含量平均值为449.00 mg·g
-1 , 低于全球陆生植物叶片C含量464.20 mg·g
-1 (
Elser et al ., 2000 )和中国东部南北样带植物叶片C含量480.10 mg·g
-1 (
任书杰等, 2012 ), 说明青海森林的林下灌木植被C储存能力相对较弱.灌木叶片N、P含量平均值分别为19.46、1.69 mg·g
-1 , 略低于全球陆生植物N、P含量(20.10、1.77 mg·g
-1 (
Reich & Oleksyn, 2004 )和20.60、1.99 mg·g
-1 (
Elser et al ., 2000 )), 略低于我国植物叶片N含量(20.24 mg·g
-1 ), 高于其叶片P含量(1.46 mg·g
-1 )(
Han et al ., 2005 ), 高于我国南方优势灌丛优势植被N、P含量(16.57、1.02 mg·g
-1 )(
李家湘等, 2017 )、陕西森林的林下灌木叶片N、P含量(13.79、1.11 mg·g
-1 )(
姜沛沛等, 2016 ), 说明青海森林的林下灌木叶片N、P含量较高.这可能与本研究区域地处高寒区, 纬度偏高, 年降水量少, 年均气温低, 昼夜温差大, 日照强度大等特殊的区域气候条件有一定的关系(
卢航等, 2013 ).温度-植物生理假说认为, 低温诱导植物生长周期缩短, 同时, 植物趋向于分配更多的营养元素于光合器官中, 通过丰富叶片N、P含量在有限的生长周期内保持高效的光合速率以及酶活性来维持植物机体内正常的新陈代谢活动(
Reich & Oleksyn, 2004 ), 因此高海拔地区植物叶片N、P含量较为丰富.这与
杨阔等(2010) 发现青藏高原草地植物群落叶片N、P含量较高的结果一致. ...
1
2002
... 生态化学计量学“内稳态理论”认为, 在长期的进化过程中, 随外界环境的变化, 生物有机体能保持其化学组成的相对恒定性(
Sterner & Elser, 2002 ).本研究发现, 7种林分间灌木(叶、枝干、根) P含量、C:P均没有明显差异性, 该结果符合“内稳态理论”, 因为7种林分间土壤TP含量相对稳定.山杨林、圆柏林、云杉林的林下灌木(叶、枝干、根) N含量、N:P高于白桦林、毛白杨林、红桦林和青扦林, C:N则相反, 这可能是由土壤TN含量、土壤C:N的显著差异造成的.相关性分析表明, 林下灌木(叶、枝干、根) N含量、N:P、C:N受土壤TN含量、土壤C:N的极显著影响(
p < 0.01)(
表2 ).冗余分析也表明(
图4 ;
表3 ), 土壤C:N极显著影响林下灌木(叶、枝干、根) C、N、P化学计量特征(
p < 0.01), 即灌木C、N、P含量受土壤有效性N的重要调控.在7种林分中, 山杨林、圆柏林和云杉林土壤C:N最低(
表1 ), 说明土壤有效性N较高.因此, 通过林下灌木层植被根系对土壤N的吸收利用, 各器官呈现出较高N含量, 较低C:N的特征. ...
Patterns of plant carbon, nitrogen, and phosphorus concentration in relation to productivity in China’s terrestrial ecosystems
1
2018
... 碳(C)、氮(N)、磷(P)作为最重要的生命元素, 是维系地球上所有生命活动的基础(
贺金生和韩兴国, 2010 ).C作为植物有机体结构的重要组成元素, 是植物有机体内生理生化反应过程的能量来源之一(
Güsewell, 2004 ), N、P是植物的基本营养元素, 也是植物有机体内蛋白质、酶类、核酸和脂质的重要组成元素(
Yang & Wang, 2011 ;
戚德辉等, 2016 ;
Tang et al ., 2018 ).C、N、P含量及其化学计量比的差异显著影响植物的生态策略, 从而被广泛应用到生态系统在生物地球化学循环和生态过程耦合关系的研究中(
Elser et al ., 1996 ).截至目前, 国内外****已在区域乃至全球范围开展了许多陆地生态系统生态化学计量特征的研究(
Han et al ., 2005 ;
Elser et al ., 2007 ;
黄小波等, 2016 ).但主要集中在植物叶片和土壤生态化学计量特征方面, 针对植物枝干、根等重要器官方面的研究相对较少(
贺合亮等, 2017 ).枝干是植物连接地下吸收器官和地上同化组织的传导器官, 具有支撑植物体的结构性骨架作用(
李单凤等, 2015 ).根系作为吸收、存储、运输水分和养分的主要器官, 也是植物有机体对外界环境变化响应最为敏感的地下器官, 在森林生态系统物质循环过程中发挥关键作用(
Iversen et al ., 2017 ;
陈晓萍等, 2018 ).因此, 研究植物不同器官间生态化学计量特征以及与土壤组分在各生态过程间的互作关系, 对于揭示生态系统养分循环规律具有重要意义. ...
Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation
1
2003
... 植物叶片N:P通常是判断植物养分限制状况的指标之一.研究表明, 当N:P < 14时, 植物生长受N限制; 当N:P > 16时, 植物生长主要受P限制; 当N:P介于14-16之间时, 植物生长受N和P的共同限制或者不受养分限制(
Koerselman, 1996 ;
Tessier & Raynal, 2003 ).本研究中, 圆柏林灌木叶片N:P (16.72) > 16, 其余6种林分叶片N:P表现为青扦林(6.22) <红桦林(7.21) <毛白杨林(7.47) <白桦林(7.48) <云杉林(12.6) <山杨林(13.7) < 14.圆柏林的林下灌木植被生长受P限制, 其余6种林分林下灌木植被生长受到N限制.这可能是由于林分差异导致其土壤N、P供应能力的不同.本研究中, 圆柏林的林下灌木叶片P含量最低(
图2 ), 圆柏林土壤TN含量最高、C:N最低、C:P较高(
表1 ), 圆柏林的林下土壤N含量丰富, N有效性较高, 而P有效性不足, 因此, 林下灌木植被在N供应较为充沛的条件下, 其生长更趋向于受P限制.其余6种林分林下灌木植被生长限制情况与我国北方森林植被普遍受到N限制的结果相一致(
Han et al ., 2005 ). ...
Pattern and variation of C:N:P ratios in China?s soils: A synthesis of observational data
2
2010
... 本研究区森林SOC、TN、TP含量平均值为(95.11、5.17、0.82 mg·g
-1 ), 高于全国土壤C、N、P含量(24.56、1.88、0.78 mg·g
-1 )(
Tian et al ., 2010 ), 与青藏高原高寒植被带土壤N、P含量(5.66、0.75 mg·g
-1 )相接近(
卢同平等, 2017 ).土壤C:N、C:P (20.6、117.7)高于全球森林土壤C:N、C:P (12.4、81.9)(
Cleveland & Liptzin, 2007 )和全国土壤C:N、C:P (12.3、52.7)(
Tian et al ., 2010 ).研究表明, 土壤C:N、C:P是衡量土壤有机质分解矿化速率的重要指标, 通常土壤C:N、C:P与有机质分解矿化速率呈反比关系, 较高的C:N、C:P代表较慢的有机质分解速率, 以及较低的土壤N、P有效性(
Cleveland & Liptzin, 2007 ).由此可推断, 青海高寒区森林SOC、TN、TP储备丰富, 有机质分解和矿化速率较慢, N、P有效性较低.这可能与本研究区域自然环境和植被类型特性有关, 青海高寒地区年降水量偏少、年平均气温低, 森林地表积累了丰富的凋落物, 长期的腐殖化作用有利于土壤有机质的富集, 较低的土壤温度抑制了土壤微生物及土壤酶活性, 地表有机质的分解周期长, 系统累积量大于消耗量, 导致森林土壤N、P元素得以累积, 但缓慢的分解速率导致土壤N、P有效性较低(
高巧等, 2014 ;
王淑彬等, 2014 ). ...
... )和全国土壤C:N、C:P (12.3、52.7)(
Tian et al ., 2010 ).研究表明, 土壤C:N、C:P是衡量土壤有机质分解矿化速率的重要指标, 通常土壤C:N、C:P与有机质分解矿化速率呈反比关系, 较高的C:N、C:P代表较慢的有机质分解速率, 以及较低的土壤N、P有效性(
Cleveland & Liptzin, 2007 ).由此可推断, 青海高寒区森林SOC、TN、TP储备丰富, 有机质分解和矿化速率较慢, N、P有效性较低.这可能与本研究区域自然环境和植被类型特性有关, 青海高寒地区年降水量偏少、年平均气温低, 森林地表积累了丰富的凋落物, 长期的腐殖化作用有利于土壤有机质的富集, 较低的土壤温度抑制了土壤微生物及土壤酶活性, 地表有机质的分解周期长, 系统累积量大于消耗量, 导致森林土壤N、P元素得以累积, 但缓慢的分解速率导致土壤N、P有效性较低(
高巧等, 2014 ;
王淑彬等, 2014 ). ...
暖温带森林生态系统林下灌木生物量相对生长模型
1
2014
... 在森林生态系统中, 林下灌木层与乔木层和草本层共同维持着森林生态系统结构与功能的稳定, 在参与养分循环、改善土壤肥力、为林下生物提供栖息环境和提高生态系统多样性等方面起到重要作用, 是森林生态系统不可或缺的组成部分(
郑绍伟等, 2007 ;
卢振龙和龚孝生, 2009 ).灌木植被通常具有木质化茎干而主干不明显, 其茎干多分枝, 树冠矮小, 根系分布广而深(
李清河等, 2006 ).目前, 对森林的林下灌木层的研究主要聚焦于灌木生物量生长模型的建立(
万五星等, 2014 ), 物种多样性和种间联结性(
崔宁洁等, 2014 ), 生态位以及物种分布特征的分析(
康永祥等, 2008 ).对于林下灌木层生态化学计量特征的研究相对较少, 尤其是高寒地区不同林分类型中灌木层的研究尚鲜有报道. ...
暖温带森林生态系统林下灌木生物量相对生长模型
1
2014
... 在森林生态系统中, 林下灌木层与乔木层和草本层共同维持着森林生态系统结构与功能的稳定, 在参与养分循环、改善土壤肥力、为林下生物提供栖息环境和提高生态系统多样性等方面起到重要作用, 是森林生态系统不可或缺的组成部分(
郑绍伟等, 2007 ;
卢振龙和龚孝生, 2009 ).灌木植被通常具有木质化茎干而主干不明显, 其茎干多分枝, 树冠矮小, 根系分布广而深(
李清河等, 2006 ).目前, 对森林的林下灌木层的研究主要聚焦于灌木生物量生长模型的建立(
万五星等, 2014 ), 物种多样性和种间联结性(
崔宁洁等, 2014 ), 生态位以及物种分布特征的分析(
康永祥等, 2008 ).对于林下灌木层生态化学计量特征的研究相对较少, 尤其是高寒地区不同林分类型中灌木层的研究尚鲜有报道. ...
青藏高原冻土区冻土与植被的关系及其对高寒生态系统的影响
1
2006
... 青海省位于青藏高原的东北部, 深居内陆腹地, 是全球气候变化的敏感区和脆弱区, 也是我国乃至亚洲重要的生态安全屏障(
贾敬敦等, 2004 ).青海森林覆盖面积小, 但植被类型多样化, 其森林生态系统及林下灌木层植被在高寒区水土保持, 涵养水源, 防风固沙, 维系生态系统平衡与稳定等方面发挥着关键的生态功能(
张胜邦, 2005 ).青海地区森林凋落物层较厚, 矿质土壤层较薄, 同时受雪被和季节性冻融等特殊因素的综合影响, 其森林生态系统物质循环过程可能具有其独特性(
左巍等, 2016 ).由于特殊的地理位置, 气候寒冷而干旱, 地形地貌复杂多样, 使植被对气候变化异常敏感和脆弱(
王根绪等, 2006 ).因此, 研究青海森林的林下灌木层生态化学计量特征将有助于我们深入了解高寒区森林生态系统的稳定性以及养分循环机理, 揭示高寒环境下森林植被的生态适应策略.基于此, 本研究选取青海省7种主要优势林分——白桦(
Betula platyphylla )林、毛白杨(
Populus tomentosa )林
、 红桦(
Betula albosinensis )林、青扦(
Picea wilsonii )林、山杨(
Populus davidiana )林、圆柏(
Sabina chinensis )林、云杉(
Picea asperata )林的林下灌木层植被为研究对象, 通过分析不同林分林下灌木植被和土壤生态化学计量特征, 阐明林下灌木植被化学计量特征与土壤组分之间的耦合关系, 并探讨林下灌木植被的养分限制情况及其生态化学计量特征的影响因素, 旨在为研究林下灌木植被对高寒环境的响应与适应机制提供基础数据, 也为指导青海高寒区森林生态系统的保护、恢复与重建提供科学依据. ...
青藏高原冻土区冻土与植被的关系及其对高寒生态系统的影响
1
2006
... 青海省位于青藏高原的东北部, 深居内陆腹地, 是全球气候变化的敏感区和脆弱区, 也是我国乃至亚洲重要的生态安全屏障(
贾敬敦等, 2004 ).青海森林覆盖面积小, 但植被类型多样化, 其森林生态系统及林下灌木层植被在高寒区水土保持, 涵养水源, 防风固沙, 维系生态系统平衡与稳定等方面发挥着关键的生态功能(
张胜邦, 2005 ).青海地区森林凋落物层较厚, 矿质土壤层较薄, 同时受雪被和季节性冻融等特殊因素的综合影响, 其森林生态系统物质循环过程可能具有其独特性(
左巍等, 2016 ).由于特殊的地理位置, 气候寒冷而干旱, 地形地貌复杂多样, 使植被对气候变化异常敏感和脆弱(
王根绪等, 2006 ).因此, 研究青海森林的林下灌木层生态化学计量特征将有助于我们深入了解高寒区森林生态系统的稳定性以及养分循环机理, 揭示高寒环境下森林植被的生态适应策略.基于此, 本研究选取青海省7种主要优势林分——白桦(
Betula platyphylla )林、毛白杨(
Populus tomentosa )林
、 红桦(
Betula albosinensis )林、青扦(
Picea wilsonii )林、山杨(
Populus davidiana )林、圆柏(
Sabina chinensis )林、云杉(
Picea asperata )林的林下灌木层植被为研究对象, 通过分析不同林分林下灌木植被和土壤生态化学计量特征, 阐明林下灌木植被化学计量特征与土壤组分之间的耦合关系, 并探讨林下灌木植被的养分限制情况及其生态化学计量特征的影响因素, 旨在为研究林下灌木植被对高寒环境的响应与适应机制提供基础数据, 也为指导青海高寒区森林生态系统的保护、恢复与重建提供科学依据. ...
广西森林土壤主要养分的空间异质性
1
2014
... 本研究区森林SOC、TN、TP含量平均值为(95.11、5.17、0.82 mg·g
-1 ), 高于全国土壤C、N、P含量(24.56、1.88、0.78 mg·g
-1 )(
Tian et al ., 2010 ), 与青藏高原高寒植被带土壤N、P含量(5.66、0.75 mg·g
-1 )相接近(
卢同平等, 2017 ).土壤C:N、C:P (20.6、117.7)高于全球森林土壤C:N、C:P (12.4、81.9)(
Cleveland & Liptzin, 2007 )和全国土壤C:N、C:P (12.3、52.7)(
Tian et al ., 2010 ).研究表明, 土壤C:N、C:P是衡量土壤有机质分解矿化速率的重要指标, 通常土壤C:N、C:P与有机质分解矿化速率呈反比关系, 较高的C:N、C:P代表较慢的有机质分解速率, 以及较低的土壤N、P有效性(
Cleveland & Liptzin, 2007 ).由此可推断, 青海高寒区森林SOC、TN、TP储备丰富, 有机质分解和矿化速率较慢, N、P有效性较低.这可能与本研究区域自然环境和植被类型特性有关, 青海高寒地区年降水量偏少、年平均气温低, 森林地表积累了丰富的凋落物, 长期的腐殖化作用有利于土壤有机质的富集, 较低的土壤温度抑制了土壤微生物及土壤酶活性, 地表有机质的分解周期长, 系统累积量大于消耗量, 导致森林土壤N、P元素得以累积, 但缓慢的分解速率导致土壤N、P有效性较低(
高巧等, 2014 ;
王淑彬等, 2014 ). ...
广西森林土壤主要养分的空间异质性
1
2014
... 本研究区森林SOC、TN、TP含量平均值为(95.11、5.17、0.82 mg·g
-1 ), 高于全国土壤C、N、P含量(24.56、1.88、0.78 mg·g
-1 )(
Tian et al ., 2010 ), 与青藏高原高寒植被带土壤N、P含量(5.66、0.75 mg·g
-1 )相接近(
卢同平等, 2017 ).土壤C:N、C:P (20.6、117.7)高于全球森林土壤C:N、C:P (12.4、81.9)(
Cleveland & Liptzin, 2007 )和全国土壤C:N、C:P (12.3、52.7)(
Tian et al ., 2010 ).研究表明, 土壤C:N、C:P是衡量土壤有机质分解矿化速率的重要指标, 通常土壤C:N、C:P与有机质分解矿化速率呈反比关系, 较高的C:N、C:P代表较慢的有机质分解速率, 以及较低的土壤N、P有效性(
Cleveland & Liptzin, 2007 ).由此可推断, 青海高寒区森林SOC、TN、TP储备丰富, 有机质分解和矿化速率较慢, N、P有效性较低.这可能与本研究区域自然环境和植被类型特性有关, 青海高寒地区年降水量偏少、年平均气温低, 森林地表积累了丰富的凋落物, 长期的腐殖化作用有利于土壤有机质的富集, 较低的土壤温度抑制了土壤微生物及土壤酶活性, 地表有机质的分解周期长, 系统累积量大于消耗量, 导致森林土壤N、P元素得以累积, 但缓慢的分解速率导致土壤N、P有效性较低(
高巧等, 2014 ;
王淑彬等, 2014 ). ...
Advances in the study on ecological stoichiometry in grass-environment system and its response to environmental factors
1
2011
... 碳(C)、氮(N)、磷(P)作为最重要的生命元素, 是维系地球上所有生命活动的基础(
贺金生和韩兴国, 2010 ).C作为植物有机体结构的重要组成元素, 是植物有机体内生理生化反应过程的能量来源之一(
Güsewell, 2004 ), N、P是植物的基本营养元素, 也是植物有机体内蛋白质、酶类、核酸和脂质的重要组成元素(
Yang & Wang, 2011 ;
戚德辉等, 2016 ;
Tang et al ., 2018 ).C、N、P含量及其化学计量比的差异显著影响植物的生态策略, 从而被广泛应用到生态系统在生物地球化学循环和生态过程耦合关系的研究中(
Elser et al ., 1996 ).截至目前, 国内外****已在区域乃至全球范围开展了许多陆地生态系统生态化学计量特征的研究(
Han et al ., 2005 ;
Elser et al ., 2007 ;
黄小波等, 2016 ).但主要集中在植物叶片和土壤生态化学计量特征方面, 针对植物枝干、根等重要器官方面的研究相对较少(
贺合亮等, 2017 ).枝干是植物连接地下吸收器官和地上同化组织的传导器官, 具有支撑植物体的结构性骨架作用(
李单凤等, 2015 ).根系作为吸收、存储、运输水分和养分的主要器官, 也是植物有机体对外界环境变化响应最为敏感的地下器官, 在森林生态系统物质循环过程中发挥关键作用(
Iversen et al ., 2017 ;
陈晓萍等, 2018 ).因此, 研究植物不同器官间生态化学计量特征以及与土壤组分在各生态过程间的互作关系, 对于揭示生态系统养分循环规律具有重要意义. ...
黄土高原刺槐林不同组分生态化学计量关系研究
1
2014
... 本研究中, 7种林分间SOC和TN含量呈现出明显差异, 而TP含量则差异不明显.这可能是由于受到营养元素的积累方式以及林分类型的影响, 林下土壤养分状况差异较大.在自然森林生态系统中, 森林土壤有机碳来自地表凋落物以及根系凋亡分解的积累, 而土壤全氮主要通过有机质分解矿化的途径累积(
李丹维等, 2017 ).首先, 林分的差异会影响地表凋落物、地下根系以及根系分泌物的组成, 同时凋落物的类型、数量以及分解速率是影响森林土壤养分含量变化和实现系统养分循环的重要因素(
杨佳佳等, 2014 ).其次, 林分的差异会影响土壤微生物、土壤动物和土壤酶的组成与功能多样性(
庞圣江等, 2015 ;
字洪标等, 2017 ), 而在土壤生物化学循环过程中, 土壤微生物通过氨化、硝化和反硝化等作用将土壤内颗粒态有机氮转化成可溶性有机氮, 而土壤酶作为催化剂也参与了其中复杂的生化反应(
李喜霞等, 2018 ).土壤P作为一种沉积性元素, 主要来源于岩石母质的风化, 其过程稳定而漫长, 因而在自然生态系统中P迁移能力较弱, 空间异质性较小(
李红林等, 2015 ).因此不同林分之间TP含量差异不明显. ...
黄土高原刺槐林不同组分生态化学计量关系研究
1
2014
... 本研究中, 7种林分间SOC和TN含量呈现出明显差异, 而TP含量则差异不明显.这可能是由于受到营养元素的积累方式以及林分类型的影响, 林下土壤养分状况差异较大.在自然森林生态系统中, 森林土壤有机碳来自地表凋落物以及根系凋亡分解的积累, 而土壤全氮主要通过有机质分解矿化的途径累积(
李丹维等, 2017 ).首先, 林分的差异会影响地表凋落物、地下根系以及根系分泌物的组成, 同时凋落物的类型、数量以及分解速率是影响森林土壤养分含量变化和实现系统养分循环的重要因素(
杨佳佳等, 2014 ).其次, 林分的差异会影响土壤微生物、土壤动物和土壤酶的组成与功能多样性(
庞圣江等, 2015 ;
字洪标等, 2017 ), 而在土壤生物化学循环过程中, 土壤微生物通过氨化、硝化和反硝化等作用将土壤内颗粒态有机氮转化成可溶性有机氮, 而土壤酶作为催化剂也参与了其中复杂的生化反应(
李喜霞等, 2018 ).土壤P作为一种沉积性元素, 主要来源于岩石母质的风化, 其过程稳定而漫长, 因而在自然生态系统中P迁移能力较弱, 空间异质性较小(
李红林等, 2015 ).因此不同林分之间TP含量差异不明显. ...
青藏高原草地植物群落冠层叶片氮磷化学计量学分析
1
2010
... 本研究中, 灌木叶片C含量平均值为449.00 mg·g
-1 , 低于全球陆生植物叶片C含量464.20 mg·g
-1 (
Elser et al ., 2000 )和中国东部南北样带植物叶片C含量480.10 mg·g
-1 (
任书杰等, 2012 ), 说明青海森林的林下灌木植被C储存能力相对较弱.灌木叶片N、P含量平均值分别为19.46、1.69 mg·g
-1 , 略低于全球陆生植物N、P含量(20.10、1.77 mg·g
-1 (
Reich & Oleksyn, 2004 )和20.60、1.99 mg·g
-1 (
Elser et al ., 2000 )), 略低于我国植物叶片N含量(20.24 mg·g
-1 ), 高于其叶片P含量(1.46 mg·g
-1 )(
Han et al ., 2005 ), 高于我国南方优势灌丛优势植被N、P含量(16.57、1.02 mg·g
-1 )(
李家湘等, 2017 )、陕西森林的林下灌木叶片N、P含量(13.79、1.11 mg·g
-1 )(
姜沛沛等, 2016 ), 说明青海森林的林下灌木叶片N、P含量较高.这可能与本研究区域地处高寒区, 纬度偏高, 年降水量少, 年均气温低, 昼夜温差大, 日照强度大等特殊的区域气候条件有一定的关系(
卢航等, 2013 ).温度-植物生理假说认为, 低温诱导植物生长周期缩短, 同时, 植物趋向于分配更多的营养元素于光合器官中, 通过丰富叶片N、P含量在有限的生长周期内保持高效的光合速率以及酶活性来维持植物机体内正常的新陈代谢活动(
Reich & Oleksyn, 2004 ), 因此高海拔地区植物叶片N、P含量较为丰富.这与
杨阔等(2010) 发现青藏高原草地植物群落叶片N、P含量较高的结果一致. ...
青藏高原草地植物群落冠层叶片氮磷化学计量学分析
1
2010
... 本研究中, 灌木叶片C含量平均值为449.00 mg·g
-1 , 低于全球陆生植物叶片C含量464.20 mg·g
-1 (
Elser et al ., 2000 )和中国东部南北样带植物叶片C含量480.10 mg·g
-1 (
任书杰等, 2012 ), 说明青海森林的林下灌木植被C储存能力相对较弱.灌木叶片N、P含量平均值分别为19.46、1.69 mg·g
-1 , 略低于全球陆生植物N、P含量(20.10、1.77 mg·g
-1 (
Reich & Oleksyn, 2004 )和20.60、1.99 mg·g
-1 (
Elser et al ., 2000 )), 略低于我国植物叶片N含量(20.24 mg·g
-1 ), 高于其叶片P含量(1.46 mg·g
-1 )(
Han et al ., 2005 ), 高于我国南方优势灌丛优势植被N、P含量(16.57、1.02 mg·g
-1 )(
李家湘等, 2017 )、陕西森林的林下灌木叶片N、P含量(13.79、1.11 mg·g
-1 )(
姜沛沛等, 2016 ), 说明青海森林的林下灌木叶片N、P含量较高.这可能与本研究区域地处高寒区, 纬度偏高, 年降水量少, 年均气温低, 昼夜温差大, 日照强度大等特殊的区域气候条件有一定的关系(
卢航等, 2013 ).温度-植物生理假说认为, 低温诱导植物生长周期缩短, 同时, 植物趋向于分配更多的营养元素于光合器官中, 通过丰富叶片N、P含量在有限的生长周期内保持高效的光合速率以及酶活性来维持植物机体内正常的新陈代谢活动(
Reich & Oleksyn, 2004 ), 因此高海拔地区植物叶片N、P含量较为丰富.这与
杨阔等(2010) 发现青藏高原草地植物群落叶片N、P含量较高的结果一致. ...
基于N:P化学计量特征的高寒草甸植物养分状况研究
1
2014
... 依据RDA发现, 林下灌木C、N、P含量及化学计量特征主要受土壤C:N、海拔、年降水量和年平均气温的影响, 这表明青海高寒区森林的林下灌木受到土壤N有效性、海拔以及区域水热条件的重要调控.土壤作为植被获取营养元素的主要机质, 其N、P有效性将显著影响地上植被组成与结构, 生产力以及有机体内元素分配格局(
李丹维等, 2017 ).
McGroddy等(2004) 认为, 土壤N、P有效性是植物有机体内元素含量的主要驱动因素, 同时土壤N、P有效性容易受降水淋溶作用的影响.
张仁懿等(2014) 研究也表明青藏高原亚高寒草甸植物养分状况受土壤N有效性的显著影响, 土壤N有效性对高寒植物生长的影响比其他元素更为关键.海拔梯度综合了温度、降水、光照等水热条件的变化, 是影响植被生长的主导因子(
Guo et al ., 2010 ).本研究结果与木本植物叶片N、P元素含量受年平均气温和年降水量共同驱动作用的结果(
Chen et al ., 2013b )相符.因为:(1)海拔梯度上降水、气温的差异会影响植物的生长周期, 从而改变C、N、P在各器官间的分配(
Kerkhoff et al ., 2005 ); (2)海拔梯度上土壤温度和水分的变化将影响土壤微生物的群系特征及代谢效率, 间接影响土壤有机质的分解和矿化速率, 而且土壤母质的风化速率以及N、P元素的淋溶强度也受降水和气温等气候因子的显著影响(
刘颖等, 2018 ); (3)不同的海拔梯度上森林植被受到的人为干扰程度也有一定的差异, 放牧、旅游、耕种等活动也会影响植物的元素分配格局(
Nogués-Bravo et al ., 2008 ).相关研究表明, 伴随着全球气候变化, 青藏高原地区已出现气温剧烈升高, 降水量增加的暖湿化趋势(
Chen et al ., 2013a ).因此, 未来在全球气候变化和区域N沉降可能加剧的背景下, 青海森林生态系统灌木层C、N、P化学计量特征将受到强烈影响. ...
基于N:P化学计量特征的高寒草甸植物养分状况研究
1
2014
... 依据RDA发现, 林下灌木C、N、P含量及化学计量特征主要受土壤C:N、海拔、年降水量和年平均气温的影响, 这表明青海高寒区森林的林下灌木受到土壤N有效性、海拔以及区域水热条件的重要调控.土壤作为植被获取营养元素的主要机质, 其N、P有效性将显著影响地上植被组成与结构, 生产力以及有机体内元素分配格局(
李丹维等, 2017 ).
McGroddy等(2004) 认为, 土壤N、P有效性是植物有机体内元素含量的主要驱动因素, 同时土壤N、P有效性容易受降水淋溶作用的影响.
张仁懿等(2014) 研究也表明青藏高原亚高寒草甸植物养分状况受土壤N有效性的显著影响, 土壤N有效性对高寒植物生长的影响比其他元素更为关键.海拔梯度综合了温度、降水、光照等水热条件的变化, 是影响植被生长的主导因子(
Guo et al ., 2010 ).本研究结果与木本植物叶片N、P元素含量受年平均气温和年降水量共同驱动作用的结果(
Chen et al ., 2013b )相符.因为:(1)海拔梯度上降水、气温的差异会影响植物的生长周期, 从而改变C、N、P在各器官间的分配(
Kerkhoff et al ., 2005 ); (2)海拔梯度上土壤温度和水分的变化将影响土壤微生物的群系特征及代谢效率, 间接影响土壤有机质的分解和矿化速率, 而且土壤母质的风化速率以及N、P元素的淋溶强度也受降水和气温等气候因子的显著影响(
刘颖等, 2018 ); (3)不同的海拔梯度上森林植被受到的人为干扰程度也有一定的差异, 放牧、旅游、耕种等活动也会影响植物的元素分配格局(
Nogués-Bravo et al ., 2008 ).相关研究表明, 伴随着全球气候变化, 青藏高原地区已出现气温剧烈升高, 降水量增加的暖湿化趋势(
Chen et al ., 2013a ).因此, 未来在全球气候变化和区域N沉降可能加剧的背景下, 青海森林生态系统灌木层C、N、P化学计量特征将受到强烈影响. ...
论青海国家重点公益林的生态地位
1
2005
... 青海省位于青藏高原的东北部, 深居内陆腹地, 是全球气候变化的敏感区和脆弱区, 也是我国乃至亚洲重要的生态安全屏障(
贾敬敦等, 2004 ).青海森林覆盖面积小, 但植被类型多样化, 其森林生态系统及林下灌木层植被在高寒区水土保持, 涵养水源, 防风固沙, 维系生态系统平衡与稳定等方面发挥着关键的生态功能(
张胜邦, 2005 ).青海地区森林凋落物层较厚, 矿质土壤层较薄, 同时受雪被和季节性冻融等特殊因素的综合影响, 其森林生态系统物质循环过程可能具有其独特性(
左巍等, 2016 ).由于特殊的地理位置, 气候寒冷而干旱, 地形地貌复杂多样, 使植被对气候变化异常敏感和脆弱(
王根绪等, 2006 ).因此, 研究青海森林的林下灌木层生态化学计量特征将有助于我们深入了解高寒区森林生态系统的稳定性以及养分循环机理, 揭示高寒环境下森林植被的生态适应策略.基于此, 本研究选取青海省7种主要优势林分——白桦(
Betula platyphylla )林、毛白杨(
Populus tomentosa )林
、 红桦(
Betula albosinensis )林、青扦(
Picea wilsonii )林、山杨(
Populus davidiana )林、圆柏(
Sabina chinensis )林、云杉(
Picea asperata )林的林下灌木层植被为研究对象, 通过分析不同林分林下灌木植被和土壤生态化学计量特征, 阐明林下灌木植被化学计量特征与土壤组分之间的耦合关系, 并探讨林下灌木植被的养分限制情况及其生态化学计量特征的影响因素, 旨在为研究林下灌木植被对高寒环境的响应与适应机制提供基础数据, 也为指导青海高寒区森林生态系统的保护、恢复与重建提供科学依据. ...
论青海国家重点公益林的生态地位
1
2005
... 青海省位于青藏高原的东北部, 深居内陆腹地, 是全球气候变化的敏感区和脆弱区, 也是我国乃至亚洲重要的生态安全屏障(
贾敬敦等, 2004 ).青海森林覆盖面积小, 但植被类型多样化, 其森林生态系统及林下灌木层植被在高寒区水土保持, 涵养水源, 防风固沙, 维系生态系统平衡与稳定等方面发挥着关键的生态功能(
张胜邦, 2005 ).青海地区森林凋落物层较厚, 矿质土壤层较薄, 同时受雪被和季节性冻融等特殊因素的综合影响, 其森林生态系统物质循环过程可能具有其独特性(
左巍等, 2016 ).由于特殊的地理位置, 气候寒冷而干旱, 地形地貌复杂多样, 使植被对气候变化异常敏感和脆弱(
王根绪等, 2006 ).因此, 研究青海森林的林下灌木层生态化学计量特征将有助于我们深入了解高寒区森林生态系统的稳定性以及养分循环机理, 揭示高寒环境下森林植被的生态适应策略.基于此, 本研究选取青海省7种主要优势林分——白桦(
Betula platyphylla )林、毛白杨(
Populus tomentosa )林
、 红桦(
Betula albosinensis )林、青扦(
Picea wilsonii )林、山杨(
Populus davidiana )林、圆柏(
Sabina chinensis )林、云杉(
Picea asperata )林的林下灌木层植被为研究对象, 通过分析不同林分林下灌木植被和土壤生态化学计量特征, 阐明林下灌木植被化学计量特征与土壤组分之间的耦合关系, 并探讨林下灌木植被的养分限制情况及其生态化学计量特征的影响因素, 旨在为研究林下灌木植被对高寒环境的响应与适应机制提供基础数据, 也为指导青海高寒区森林生态系统的保护、恢复与重建提供科学依据. ...
灌木群落及生物量研究综述
1
2007
... 在森林生态系统中, 林下灌木层与乔木层和草本层共同维持着森林生态系统结构与功能的稳定, 在参与养分循环、改善土壤肥力、为林下生物提供栖息环境和提高生态系统多样性等方面起到重要作用, 是森林生态系统不可或缺的组成部分(
郑绍伟等, 2007 ;
卢振龙和龚孝生, 2009 ).灌木植被通常具有木质化茎干而主干不明显, 其茎干多分枝, 树冠矮小, 根系分布广而深(
李清河等, 2006 ).目前, 对森林的林下灌木层的研究主要聚焦于灌木生物量生长模型的建立(
万五星等, 2014 ), 物种多样性和种间联结性(
崔宁洁等, 2014 ), 生态位以及物种分布特征的分析(
康永祥等, 2008 ).对于林下灌木层生态化学计量特征的研究相对较少, 尤其是高寒地区不同林分类型中灌木层的研究尚鲜有报道. ...
灌木群落及生物量研究综述
1
2007
... 在森林生态系统中, 林下灌木层与乔木层和草本层共同维持着森林生态系统结构与功能的稳定, 在参与养分循环、改善土壤肥力、为林下生物提供栖息环境和提高生态系统多样性等方面起到重要作用, 是森林生态系统不可或缺的组成部分(
郑绍伟等, 2007 ;
卢振龙和龚孝生, 2009 ).灌木植被通常具有木质化茎干而主干不明显, 其茎干多分枝, 树冠矮小, 根系分布广而深(
李清河等, 2006 ).目前, 对森林的林下灌木层的研究主要聚焦于灌木生物量生长模型的建立(
万五星等, 2014 ), 物种多样性和种间联结性(
崔宁洁等, 2014 ), 生态位以及物种分布特征的分析(
康永祥等, 2008 ).对于林下灌木层生态化学计量特征的研究相对较少, 尤其是高寒地区不同林分类型中灌木层的研究尚鲜有报道. ...
青海不同林分土壤微生物群落结构(PLFA)
2
2017
... 青海省地处青藏高原的东北部, 地理位置为31.65°-39.32° N, 89.58°-103.07° E.地形复杂多样, 高山、丘陵、河谷、盆地交错分布, 平均海拔3 000 m以上, 属于典型高原大陆性气候.常年平均气温-3.7-6.0 ℃, 年日照时间2 340-3 550 h, 年降水量16.7-776.1 mm (大部分400 mm以下), 年蒸发量1 118.4-3 536.2 mm (大部分1 500 mm以上), 植被地域跨青藏高原、温带荒漠和温带草原3个植被区, 具有高寒和旱生的特点.青海省森林资源贫乏, 森林覆盖率仅有4.57%, 森林植被主要分布于江河及其支流的河谷两岸, 海拔大多在2 000-4 000 m, 以寒温性针叶林为主, 其次为落叶阔叶林(
字洪标等, 2017 ).常见的针叶树种有云杉、圆柏等, 阔叶树种有杨树(
Populus spp.)、白桦等, 林下灌木有金露梅(
Potentilla fruticosa )、鲜卑花(
Sibiraea laevigata )、野蔷薇(
Rosa multiflora )
、 银露梅(
Potentilla glabra )
、 杯腺柳(
Salix cupularis )等, 主要的森林土壤类型有棕壤、黄棕壤、暗棕壤、褐棕壤、砂壤土等(
董旭, 2009 ;
胡雷等, 2015 ). ...
... 本研究中, 7种林分间SOC和TN含量呈现出明显差异, 而TP含量则差异不明显.这可能是由于受到营养元素的积累方式以及林分类型的影响, 林下土壤养分状况差异较大.在自然森林生态系统中, 森林土壤有机碳来自地表凋落物以及根系凋亡分解的积累, 而土壤全氮主要通过有机质分解矿化的途径累积(
李丹维等, 2017 ).首先, 林分的差异会影响地表凋落物、地下根系以及根系分泌物的组成, 同时凋落物的类型、数量以及分解速率是影响森林土壤养分含量变化和实现系统养分循环的重要因素(
杨佳佳等, 2014 ).其次, 林分的差异会影响土壤微生物、土壤动物和土壤酶的组成与功能多样性(
庞圣江等, 2015 ;
字洪标等, 2017 ), 而在土壤生物化学循环过程中, 土壤微生物通过氨化、硝化和反硝化等作用将土壤内颗粒态有机氮转化成可溶性有机氮, 而土壤酶作为催化剂也参与了其中复杂的生化反应(
李喜霞等, 2018 ).土壤P作为一种沉积性元素, 主要来源于岩石母质的风化, 其过程稳定而漫长, 因而在自然生态系统中P迁移能力较弱, 空间异质性较小(
李红林等, 2015 ).因此不同林分之间TP含量差异不明显. ...
青海不同林分土壤微生物群落结构(PLFA)
2
2017
... 青海省地处青藏高原的东北部, 地理位置为31.65°-39.32° N, 89.58°-103.07° E.地形复杂多样, 高山、丘陵、河谷、盆地交错分布, 平均海拔3 000 m以上, 属于典型高原大陆性气候.常年平均气温-3.7-6.0 ℃, 年日照时间2 340-3 550 h, 年降水量16.7-776.1 mm (大部分400 mm以下), 年蒸发量1 118.4-3 536.2 mm (大部分1 500 mm以上), 植被地域跨青藏高原、温带荒漠和温带草原3个植被区, 具有高寒和旱生的特点.青海省森林资源贫乏, 森林覆盖率仅有4.57%, 森林植被主要分布于江河及其支流的河谷两岸, 海拔大多在2 000-4 000 m, 以寒温性针叶林为主, 其次为落叶阔叶林(
字洪标等, 2017 ).常见的针叶树种有云杉、圆柏等, 阔叶树种有杨树(
Populus spp.)、白桦等, 林下灌木有金露梅(
Potentilla fruticosa )、鲜卑花(
Sibiraea laevigata )、野蔷薇(
Rosa multiflora )
、 银露梅(
Potentilla glabra )
、 杯腺柳(
Salix cupularis )等, 主要的森林土壤类型有棕壤、黄棕壤、暗棕壤、褐棕壤、砂壤土等(
董旭, 2009 ;
胡雷等, 2015 ). ...
... 本研究中, 7种林分间SOC和TN含量呈现出明显差异, 而TP含量则差异不明显.这可能是由于受到营养元素的积累方式以及林分类型的影响, 林下土壤养分状况差异较大.在自然森林生态系统中, 森林土壤有机碳来自地表凋落物以及根系凋亡分解的积累, 而土壤全氮主要通过有机质分解矿化的途径累积(
李丹维等, 2017 ).首先, 林分的差异会影响地表凋落物、地下根系以及根系分泌物的组成, 同时凋落物的类型、数量以及分解速率是影响森林土壤养分含量变化和实现系统养分循环的重要因素(
杨佳佳等, 2014 ).其次, 林分的差异会影响土壤微生物、土壤动物和土壤酶的组成与功能多样性(
庞圣江等, 2015 ;
字洪标等, 2017 ), 而在土壤生物化学循环过程中, 土壤微生物通过氨化、硝化和反硝化等作用将土壤内颗粒态有机氮转化成可溶性有机氮, 而土壤酶作为催化剂也参与了其中复杂的生化反应(
李喜霞等, 2018 ).土壤P作为一种沉积性元素, 主要来源于岩石母质的风化, 其过程稳定而漫长, 因而在自然生态系统中P迁移能力较弱, 空间异质性较小(
李红林等, 2015 ).因此不同林分之间TP含量差异不明显. ...
青海高寒区不同林分类型凋落物养分状况及化学计量特征
1
2016
... 青海省位于青藏高原的东北部, 深居内陆腹地, 是全球气候变化的敏感区和脆弱区, 也是我国乃至亚洲重要的生态安全屏障(
贾敬敦等, 2004 ).青海森林覆盖面积小, 但植被类型多样化, 其森林生态系统及林下灌木层植被在高寒区水土保持, 涵养水源, 防风固沙, 维系生态系统平衡与稳定等方面发挥着关键的生态功能(
张胜邦, 2005 ).青海地区森林凋落物层较厚, 矿质土壤层较薄, 同时受雪被和季节性冻融等特殊因素的综合影响, 其森林生态系统物质循环过程可能具有其独特性(
左巍等, 2016 ).由于特殊的地理位置, 气候寒冷而干旱, 地形地貌复杂多样, 使植被对气候变化异常敏感和脆弱(
王根绪等, 2006 ).因此, 研究青海森林的林下灌木层生态化学计量特征将有助于我们深入了解高寒区森林生态系统的稳定性以及养分循环机理, 揭示高寒环境下森林植被的生态适应策略.基于此, 本研究选取青海省7种主要优势林分——白桦(
Betula platyphylla )林、毛白杨(
Populus tomentosa )林
、 红桦(
Betula albosinensis )林、青扦(
Picea wilsonii )林、山杨(
Populus davidiana )林、圆柏(
Sabina chinensis )林、云杉(
Picea asperata )林的林下灌木层植被为研究对象, 通过分析不同林分林下灌木植被和土壤生态化学计量特征, 阐明林下灌木植被化学计量特征与土壤组分之间的耦合关系, 并探讨林下灌木植被的养分限制情况及其生态化学计量特征的影响因素, 旨在为研究林下灌木植被对高寒环境的响应与适应机制提供基础数据, 也为指导青海高寒区森林生态系统的保护、恢复与重建提供科学依据. ...
青海高寒区不同林分类型凋落物养分状况及化学计量特征
1
2016
... 青海省位于青藏高原的东北部, 深居内陆腹地, 是全球气候变化的敏感区和脆弱区, 也是我国乃至亚洲重要的生态安全屏障(
贾敬敦等, 2004 ).青海森林覆盖面积小, 但植被类型多样化, 其森林生态系统及林下灌木层植被在高寒区水土保持, 涵养水源, 防风固沙, 维系生态系统平衡与稳定等方面发挥着关键的生态功能(
张胜邦, 2005 ).青海地区森林凋落物层较厚, 矿质土壤层较薄, 同时受雪被和季节性冻融等特殊因素的综合影响, 其森林生态系统物质循环过程可能具有其独特性(
左巍等, 2016 ).由于特殊的地理位置, 气候寒冷而干旱, 地形地貌复杂多样, 使植被对气候变化异常敏感和脆弱(
王根绪等, 2006 ).因此, 研究青海森林的林下灌木层生态化学计量特征将有助于我们深入了解高寒区森林生态系统的稳定性以及养分循环机理, 揭示高寒环境下森林植被的生态适应策略.基于此, 本研究选取青海省7种主要优势林分——白桦(
Betula platyphylla )林、毛白杨(
Populus tomentosa )林
、 红桦(
Betula albosinensis )林、青扦(
Picea wilsonii )林、山杨(
Populus davidiana )林、圆柏(
Sabina chinensis )林、云杉(
Picea asperata )林的林下灌木层植被为研究对象, 通过分析不同林分林下灌木植被和土壤生态化学计量特征, 阐明林下灌木植被化学计量特征与土壤组分之间的耦合关系, 并探讨林下灌木植被的养分限制情况及其生态化学计量特征的影响因素, 旨在为研究林下灌木植被对高寒环境的响应与适应机制提供基础数据, 也为指导青海高寒区森林生态系统的保护、恢复与重建提供科学依据. ...
 ,1,*
,1,*
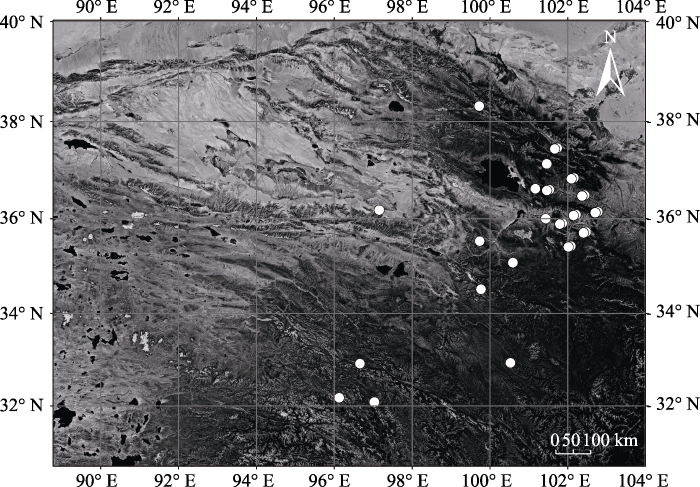 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT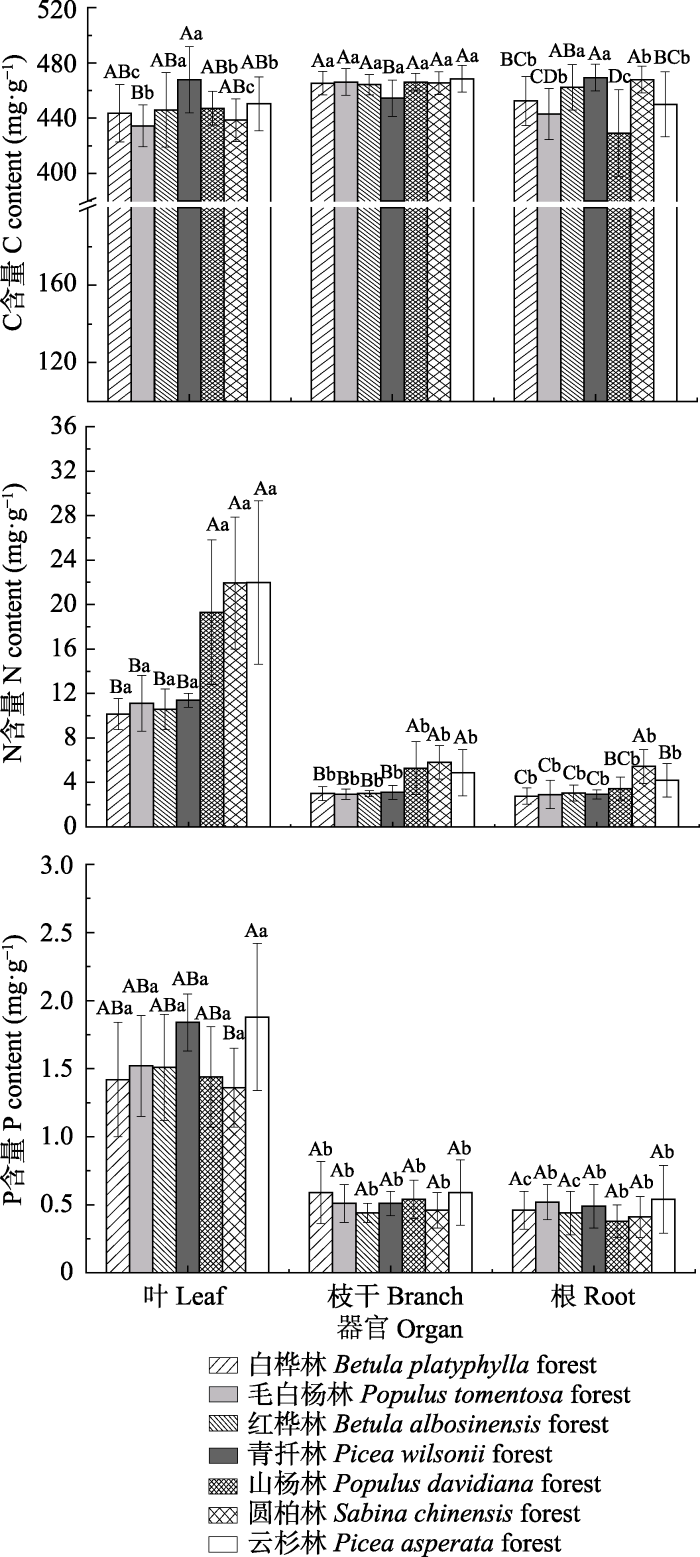 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT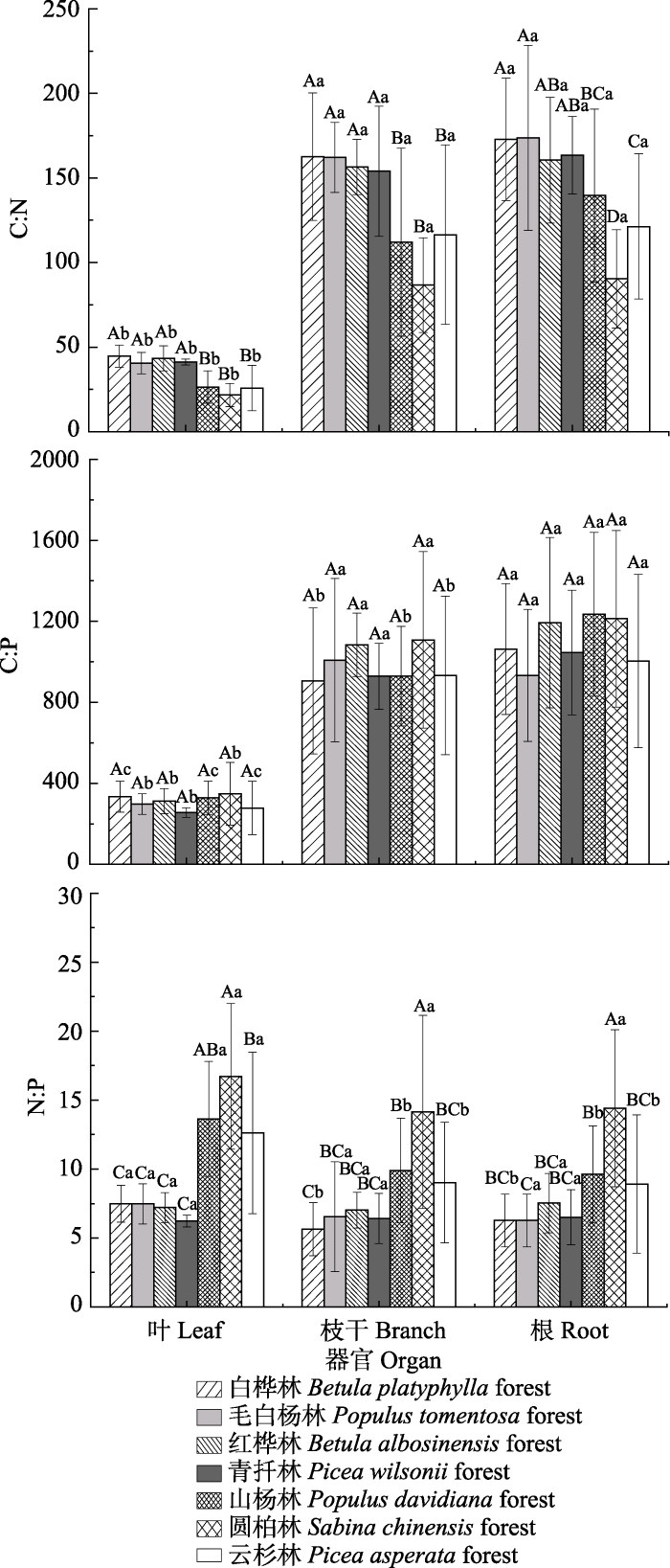 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT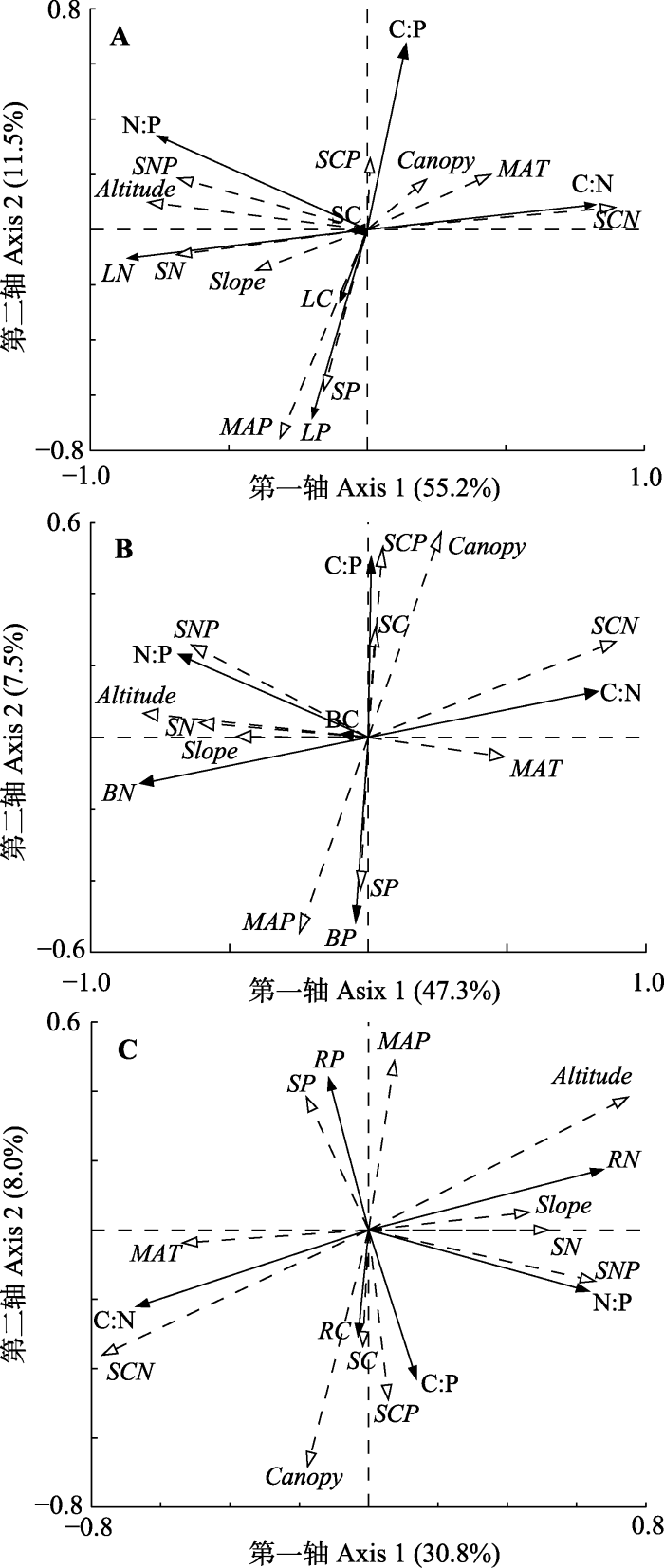 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}