Spatial variations of community functional traits at different successional stages in temperate forests of Changbai Mountains, Northeast China
Shu-Jun HAO, Xiao-Yu LI, Man-Man HOU, Xiu-Hai ZHAO,*Research Center of Forest Management Engineering of State Forestry and Grassland Administration, Beijing Forestry University, Beijing 10083, China
Supported by the National Key R&D Program of China(2017YFC0504005) The National Natural Science Foundation of China(31670643)
摘要 群落构建机制研究是生态学研究的热点。长白山自然保护区拥有完整的原始阔叶红松林生态系统, 近年来随着物种多样性丧失愈发严重, 对该地区开展群落构建机制研究显得尤为重要。该研究以长白山不同演替阶段的3块5.2 hm 2固定监测样地(次生杨桦林、次生针阔混交林、原始椴树红松林)为研究对象, 通过采集样地内主要树种的6个关键功能性状(叶面积、比叶面积、叶片厚度、叶氮含量、叶磷含量、最大树高), 分析不同空间尺度下(5 m × 5 m, 10 m × 10 m, 20 m × 20 m, 30 m × 30 m, 40 m × 40 m, 50 m × 50 m和60 m × 60 m)及不同演替阶段群落性状空间值的变化, 结合零模型的模拟结果对长白山温带森林演替过程中的群落构建机制进行讨论。结果表明: 种库大小对于研究结果具有重要影响。在较大的种库下, 环境过滤作用影响显著。而在样地水平进行研究时, 演替早期和中期, 群落性状空间值与零模型模拟值无显著差异, 在演替的晚期, 群落性状空间值显著高于零模型模拟值。结合多个群落功能多样性指数分析发现, 环境过滤和竞争作用共同决定该地区顶级群落的物种组成。在演替早期大量物种迁入, 群落内物种间存在强烈的资源竞争, 而随着演替进行, 部分物种逐渐被竞争排除出群落, 群落中的物种呈现明显的生态位分化, 竞争作用是维持物种共存的主要机制。 关键词:群落构建;功能性状;性状空间;空间尺度
Abstract Aims The community assembly mechanisms are among the focal topics in ecological studies. In Changbai Mountains Nature Reserve, there is an intact primary broadleaved-Korean pine forest ecosystem. With increasing loss of species diversity in recent years, study that explores the community assembly mechanisms in this region is particularly important. Methods This study was conducted in three large permanent plots, each of the size 5.2 hm 2, along suessional stages (secondary poplar and birch mixed forest, PBF; secondary mixed conifer and broad-leaved forest, CBF; and primary Tilia amurensis-Pinus koraiensis mixed forest, TKF) in Changbai Mountains. Six functional traits of major tree species were measured, including leaf area, specific leaf area, leaf thickness, leaf nitrogen content, leaf phosphorus content, and maximum tree height. Changes in the spatial values of community traints were analyzed at different spatial scales (5 m × 5 m, 10 m × 10 m, 20 m × 20 m, 30 m × 30 m, 40 m × 40 m, 50 m × 50 m and 60 m × 60 m). By comparing the observed values with expected values of null models, the community assembly mechanisms in temperate forests of Changbai Mountains were explored. Important findings Results show that the size of species pool has an important impact on the outcome; in a larger species pool, the environmental filtration has a significant impact. At the plot level and for early and intermediate stages of succession, the observed spatial values of community traits do not significantly differ from the expected values. At the late successional stage, the observed spatial values of community traits were greater than the expected values. The analysis of multiple community functional diversity indices shows that the combined processes of habitat filtring and competitive exclusion are the main determinants of the species composition of the climax community in this region. In the early successional stage, large numbers of species are immigrated, and there are strong resource competitions among the species within a community. With progressing succession, some species are excluded, species maintained in the community show significant niche differentiations, and competition is the main mechanism species coexistence. Keywords:community assembly;functional trait;trait space;spatial scale
PDF (1883KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 郝姝珺, 李晓宇, 侯嫚嫚, 赵秀海. 长白山温带森林不同演替阶段群落功能性状的空间变化. 植物生态学报, 2019, 43(3): 208-216. DOI: 10.17521/cjpe.2018.0295 HAO Shu-Jun, LI Xiao-Yu, HOU Man-Man, ZHAO Xiu-Hai. Spatial variations of community functional traits at different successional stages in temperate forests of Changbai Mountains, Northeast China. Chinese Journal of Plant Ecology, 2019, 43(3): 208-216. DOI: 10.17521/cjpe.2018.0295
Table 2 表2 表2功能性状及生态学意义 Table 2Functional traits and ecological meaning
功能性状 Functional trait
单位 Unit
生态学意义 Ecological meaning
叶面积 Leaf area
mm2
代表叶片对光的捕获能力, 与环境胁迫和干扰下的生态策略有关 Represents the ability to capture light; linked to ecological strategy with respect to environmental stress and disturbances
比叶面积 Specific leaf area
mm2·mg-1
与叶片光合能力正相关, 与叶片寿命负相关 Positively related to photosynthetic rate and negatively to leaf longevity
叶片厚度 Leaf thickness
mm
代表植物抗干扰和高投入的能力 Represents the ability to defy changes and high investment
叶氮含量 Leaf nitrogen concentration
mg·g-1
与光合作用蛋白和最大光合速率有关 Related to the proteins in photosynthetic machinery and maximum photosynthetic rate
叶磷含量 Leaf phosphorus concentration
mg·g-1
与植物最大光合速率以及营养级质量有关 Related to maximum photosynthetic rate and high nutritional quality in food webs
最大树高 Maximum tree height
m
代表植株在光垂直梯度所处的位置及竞争能力 Associated with position of the species in the vertical light gradient of plants and competitiveness
Fig. 2The weighted average (CWM) of individual functional traits at different successional stages in temperate forests of Changbai Mountains. PBF, secondary poplar and birch mixed forest; CBF, secondary mixed conifer and broad-leaved forest; TKF, primary Tilia amurensis-Pinus koraiensis mixed forest. The box plots show the results of repeated sampling for 20 times. The black triangle is values at plot level; different lowercase letters designate the results of multiple comparisons, and it indicate significant difference (p < 0.05).
Fig. 3Trends in functional diversity at different successional stages in temperate forests of Changbai Mountains. PBF, secondary poplar and birch mixed forest; CBF, secondary mixed conifer and broad-leaved forest; TKF, primary Tilia amurensis-Pinus koraiensis mixed forest. FEve, functional uniformity; FDiv, functional divergence; FDis, functional dispersion; RaoQ, functional entropy index. The box plots show the results of repeated sampling for 20 times. The black triangle is the value at plot level; different lowercase letters show the results of multiple comparisons, and it indicate significant difference (p < 0.05).
新窗口打开|下载原图ZIP|生成PPT 图4长白山温带森林不同演替阶段群落性状空间值随空间尺度的变化。A, D, 次生杨桦林。B, E, 次生针阔混交林。C, F, 原始椴树红松林。A, B, C, 种库一。D, E, F, 种库二。SES, 标准化效应值。***, p < 0.001, 表示性状空间观测值与零模型模拟值存在显著差异。
Fig. 4Changes in the spatial value of community traits at different succession stages with spatial scale in temperate forests of Changbai Mountains. A, D, Secondary poplar forest. B, E, Secondary coniferous and broad-leaved mixed forest. C, F, Primary Tilia amurensis-Pinus koraiensis mixed forest. A, B, C, Species pool 1. D, E, F, Species pool 2. SES, standardized effect size. ***, p < 0.01, it indicates that there is a significant difference between the eigenspace observations and the zero model simulation values.
BaralotoC, HardyOJ, PaineCT, DexterKG, CruaudC, DunningLT, ChaveJ ( 2012). Using functional traits and phylogenetic trees to examine the assembly of tropical tree communities Journal of Ecology, 100, 690-701. [本文引用: 1]
CornelissenJHC, LavorelS, GarnierE, DíazS, BuchmannN, GurvichDE ( 2003). A handbook of protocols for standardised and easy measurement of plant functional traits worldwide Australian Journal of Botany, 51, 335-380. [本文引用: 1]
CornwellWK, SchwilkDW, AckerlyDD ( 2006). A trait-based test for habitat filtering: Convex hull volume Ecology, 87, 1465-1471. [本文引用: 1]
DiamondJM ( 1975). Assembly of species communities In: Cody ML, Diamond JM eds. Ecology & Evolution of Communities. Harvard University Press, Cambridge, USA.
DíazS, QuétierF, CáceresDM, TrainorSF, Pérez-?HarguindeguyN, Bret-HarteMS, FineganB, Pe?a-ClarosM, PoorterL ( 2011). Linking functional diversity and social actor strategies in a framework for interdisciplinary analysis of nature’s benefits to society Proceedings of the National Academy of Sciences of the United States of America, 108, 895-902. [本文引用: 1]
EvinerVT, Chapin IIIFS ( 2003). Functional matrix: A conceptual framework for predicting multiple plant effects on ecosystem processes. Annual Review of Ecology, Evolution, and Systematics, 34, 455-485.
FangS, YuanZQ, LinF, YeJ, HaoZQ, WangXG ( 2014). Functional and phylogenetic structures of woody plants in broad-leaved Korean pine mixed forest in Changbai Mountains, Jilin, China Chinese Science Bulletin, 59, 2342-2348. [本文引用: 3]
GrimeJP ( 2006). Trait convergence and trait divergence in herbaceous plant communities: Mechanisms and consequences Journal of Vegetation Science, 17, 255-260.
HerbenT, GoldbergDE ( 2014). Community assembly by limiting similarity vs. competitive hierarchies: Testing the consequences of dispersion of individual traits Journal of Ecology, 102, 156-166.
HouMM, LiXY, WangJW, LiuS, ZhaoXH ( 2017). Phylogenetic development and functional structures during successional stages of conifer and broad-leaved mixed forest communities in Changbai Mountains, China Acta Ecologica Sinica, 37, 7503-7513. [本文引用: 3]
HutchinsonGE ( 1978). An Introduction to Population Ecology. Yale University Press, New Haven.
KraftN, AckerlyDD ( 2014). The assembly of plant communities In: Monson RK ed. The Plant Sciences—Ecology and the Environment. Springer, Berlin. 67-88.
KraftNJ, ValenciaR, AckerlyDD ( 2008). Functional traits and niche-based tree community assembly in an Amazonian forest Science, 322, 580-582.
LalibertéE, LegendreP ( 2010). A distance-based framework for measuring functional diversity from multiple traits Ecology, 91, 299-305. [本文引用: 1]
LessardJP, BelmakerJ, MyersJA, ChaseJM, RahbekC ( 2012 a). Inferring local ecological processes amid species pool influences Trends in Ecology & Evolution, 27, 600-607. [本文引用: 1]
LessardJP, BorregaardMK, FordyceJA, RahbekC, WeiserMD, DunnRR, SandersNJ ( 2012 b). Strong influence of regional species pools on continent-wide structuring of local communities Proceedings Biological Sciences, 279, 266-274. [本文引用: 1]
LiXY, LiaoJX, HouMM, FanXH ( 2016). Multi-scale analysis on community phylogenetic structure of secondary Populus davidiana-Betula platyphylla forest in Changbai Mountains, northeastern China Journal of Beijing Forestry University, 38(12), 14-20. [本文引用: 2]
LiY, ShipleyB, PriceJN, DantasVDL, TammeR, WestobyM, LaughlinDC ( 2017). Habitat filtering determines the functional niche occupancy of plant communities worldwide Journal of Ecology, 106, 1001-1009. [本文引用: 2]
LitvakMK, HansellRI ( 1990). A community perspective on the multi-dimensional niche Journal of Animal Ecology, 59, 931-940.
LohbeckM, PoorterL, Martínez-RamosM, BongersF ( 2015). Biomass is the main driver of changes in ecosystem process rates during tropical forest succession Ecology, 96, 1242-1252. [本文引用: 1]
MasonNW, de BelloF, Dole?alJ, Lep?J ( 2011). Niche overlap reveals the effects of competition, disturbance and contrasting assembly processes in experimental grassland communities Journal of Ecology, 99, 788-796.
MasonNW, MouillotD, LeeWG, WilsonJB ( 2005). Functional richness, functional evenness and functional divergence: The primary components of functional diversity Oikos, 111, 112-118. [本文引用: 2]
McGillBJ, EnquistBJ, WeiherE, WestobyM ( 2006). Rebuilding community ecology from functional traits Trends in Ecology & Evolution, 21, 178-185.
MouchetMA, VillégerS, MasonNW, MouillotD ( 2010). Functional diversity measures: An overview of their redundancy and their ability to discriminate community assembly rules Functional Ecology, 24, 867-876. [本文引用: 1]
MouillotD, StubbsW, FaureM, DumayO, TomasiniJA, WilsonJB, Do ChiT ( 2005). Niche overlap estimates based on quantitative functional traits: A new family of non-parametric indices Oecologia, 145, 345-353.
OstertagR, WarmanL, CordellS, VitousekPM ( 2015). Using plant functional traits to restore Hawaiian rainforest Journal of Applied Ecology, 52, 805-809. [本文引用: 1]
RosenfeldJS ( 2002). Functional redundancy in ecology and conservation Oikos, 98, 156-162
SatdichanhM, MilletJ, HeinimannA, NanthavongK, HarrisonRD ( 2015). Using plant functional traits and phylogenies to understand patterns of plant community assembly in a seasonal tropical forest in Lao PDR PLOS ONE, 10, e0130151. DOI: 10.1371/journal.pone.0130151. [本文引用: 1]
SongYT, WangP, ZhouDW ( 2011). Methods of measuring plant community functional diversity Chinese Journal of Ecology, 30, 2053-2059. [本文引用: 2]
StubbsWJ, WilsonJB ( 2004). Evidence for limiting similarity in a sand dune community Journal of Ecology, 92, 557-567.
SuterineM, EdwardsPJ ( 2013). Convergent succession of plant communities is linked to species’ functional traits Perspectives in Plant Ecology,Evolution and Systematics, 15, 217-225. [本文引用: 1]
SwensonNG ( 2013). The assembly of tropical tree communities—The advances and shortcomings of phylogenetic and functional trait analyses Ecography, 36, 264-276.
SwensonNG, WeiserMD ( 2014). On the packing and filling of functional space in eastern North American tree assemblages Ecography, 37, 1056-1062.
VillégerS, MasonNW, MouillotD ( 2008). New multidimensional functional diversity indices for a multifaceted framework in functional ecology Ecology, 89, 2290-2301. [本文引用: 1]
WangSP, TangZY, QiaoXJ, ShenZH, WangXP, ZhengCY, FangJY ( 2013). The influence of species pools and local processes on the community structure: A test case with woody plant communities in China’s mountains Ecography, 35, 1168-1175. [本文引用: 1]
WebbCO, AckerlyDD, McPeekMA, DonoghueMJ ( 2002). Phylogenies and community ecology Annual Review of Ecology and Systematics, 33, 475-505. [本文引用: 1]
WeiherE, ClarkeGP, KeddyPA ( 1998). Community assembly rules, morphological dispersion, and the coexistence of plant species Oikos, 81, 309-322.
WeiherE, FreundD, BuntonT, StefanskiA, LeeT, BentivengaS ( 2011). Advances, challenges and a developing synthesis of ecological community assembly theory Philosophical Transactions of the Royal Society of London B: Biological Sciences, 366, 2403-2413.
ZhouXG, LuWK, YeD, WenYG ( 2014). Assembly mechanism forest community based on phylogeny and functional traits Guangxi Science, 21, 525-533.
 ,*北京林业大学国家林业和草原局森林经营工程技术研究中心, 北京 100083
,*北京林业大学国家林业和草原局森林经营工程技术研究中心, 北京 100083
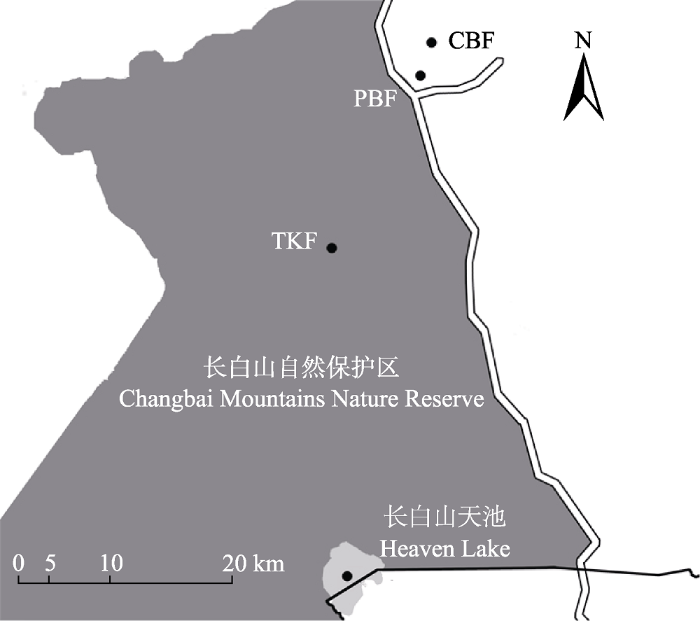 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT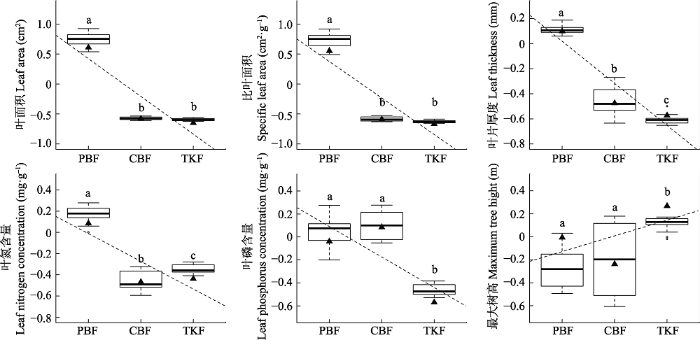 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT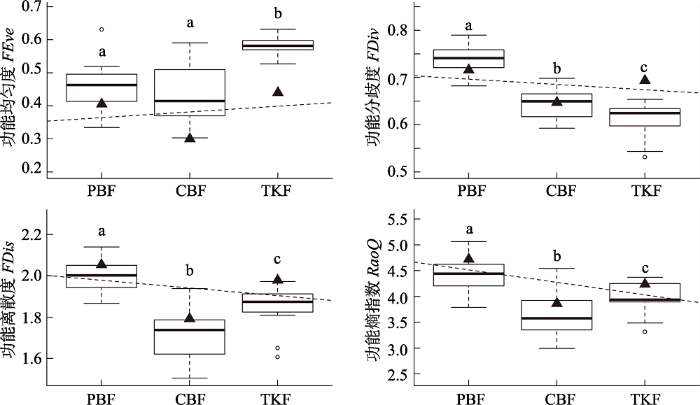 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT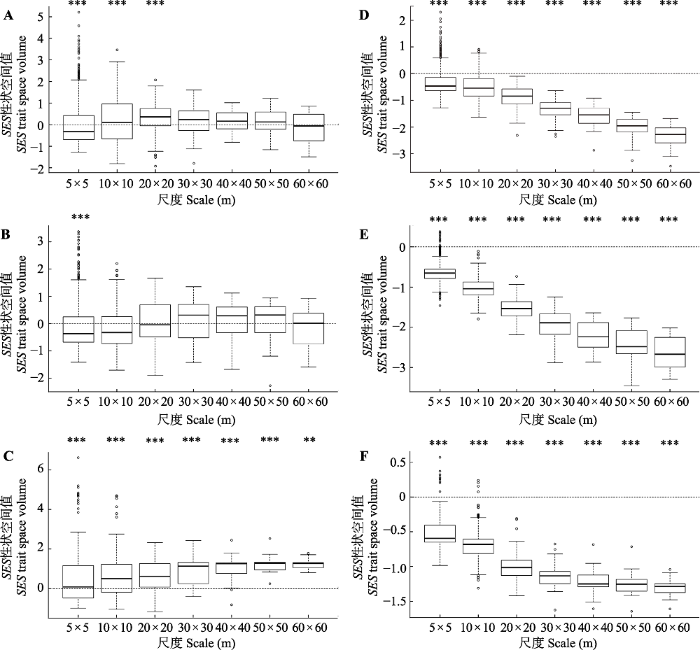 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}