蒙文萍
1 , 2 , 戴全厚
, 1 , * , 冉景丞
3 1.
贵州大学林学院, 贵阳 550025 2.
贵州省植物园, 贵阳 550001 3.
贵州省野生动物和森林植物管理站, 贵阳 550001 A review on the process of bryophyte karstification Wen-Ping MENG
1 , 2 , Quan-Hou DAI
, 1 , * , Jing-Cheng RAN
3 1.
College of Forestry, Guizhou University, Guiyang 550025, China 2.
Guizhou Botanical Garden, Guiyang 550001, China 3.
Guizhou Wildlife and Forest Plant Management Station, Guiyang 550001, China 通讯作者: *(
qhdairiver@163.com )
编委: 郭柯
责任编辑: 李敏, 实习编辑: 赵航
收稿日期: 2019-09-22
接受日期: 2019-05-6
网络出版日期: 2019-05-20
基金资助: 国家自然科学基金 (41671275 )国家重点研发计划 (2016YFC0502604 )国家自然科学基金委员会-贵州人民政府喀斯特科学研究中心项目 (U1812401 )
Editorial board: Guo Ke Editor: LI Min, ZHAO Hang Received: 2019-09-22
Accepted: 2019-05-6
Online: 2019-05-20
Fund supported: Supported by the National Natural Science Foundation of China (41671275 )The National Key R&D Program of China (2016YFC0502604 )The National Nature Science Foundation of China and the Karst Science Research Center of Guizhou Province (U1812401 )
摘要 苔藓植物参与岩溶作用的过程是生物岩溶的重要内容。开展苔藓植物岩溶作用研究可为石漠化裸岩的生态恢复及其综合治理提供理论依据和技术支撑。该文系统论述了苔藓植物岩溶作用的过程(溶蚀与沉积)、机理及其与生境互作机制的最新研究进展。苔藓植物及其生物结皮在干湿交替下发生膨胀、卷缩、冻融等物理机械力, 以及其代谢分泌物和因呼吸作用而产生的H
2 CO
3 与岩石中矿物反应, 破坏矿物晶体结构, 导致矿物裂解、使岩面崩解脱落而改变岩面形态, 塑造岩溶地貌, 形成原始土壤。苔藓植物岩溶作用驱动力的大小与植物功能性状、基岩性质与生存环境密切相关。此外, 生物岩溶作用需要长时间尺度的监测, 研究周期长, 建议建立野外长期监测基地, 加强对其内在机制与过程以及与生境间互作机制, 苔藓植物的生理代谢过程及其生物酶等与岩溶作用的相互关系, 岩溶地区苔藓植物环境适应性特征及其生物多样性维持机制等方面的研究。
关键词: 苔藓植物 ;
岩溶作用 ;
过程 ;
机理 ;
生境 ;
互作机制 Abstract Bryophyte participate in karstification is an important part of biokarst process. Numerous studies on bryophyte karstification provide theoretical and technical foundation for restoration and comprehensive management of bare rock in the rocky desertification area. This article systematically reviewed the process (dissolution and sedimentation), mechanism and interaction relationship between bryophyte karstification and habitats. Bryophyte and its biological crusts emerge physical forces such as expansion, curling, freezing and thawing when they are under alternating wet or dry conditions can destroy rock. In addition, their metabolic secretions and H
2 CO
3 formed by respiration, which react with minerals resulting in destruction of the crystal structure, pyrolysis the minerals, further the rock surface disintegrated and the surface morphology changed, the karst landform and the original soil formed. The driving force of bryophyte karstification closely related to plant functional traits, rock properties and habitat. Studies on biokarst need long-term monitoring and long research period. It is recommended to establish a long-term monitoring sites for strengthening examinations on process, internal mechanisms, and interaction relationship with habitat of bryophyte karstification. At the same time, physiological metabolic processes of bryophytes and the relationship with bryophyte karstification should be emphasized. The environmental adaptability of bryophyte and the maintenance mechanism of biodiversity in karst areas need research attention as well.
Keywords: bryophytes ;
karstification ;
process ;
mechanism ;
habitat ;
interaction mechanism PDF (2088KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 蒙文萍, 戴全厚, 冉景丞. 苔藓植物岩溶作用研究进展. 植物生态学报, 2019, 43(5): 396-407. DOI:
10.17521/cjpe.2019.0020 MENG Wen-Ping, DAI Quan-Hou, RAN Jing-Cheng.
A review on the process of bryophyte karstification .
Chinese Journal of Plant Ecology , 2019, 43(5): 396-407. DOI:
10.17521/cjpe.2019.0020 生物岩溶是指由生物引起的岩溶作用现象及 其产物, 即生物溶蚀、破坏可溶岩和沉淀形成次生化学沉积物及其所遗留或形成的形态(
Schneider, 1976 )。生物岩溶作用(溶蚀作用和沉积作用)激活了岩溶生态系统基质中的物质和能量, 使各个圈层间信息交流更加频繁与复杂。苔藓植物作为生态系统演替过程中的先锋植物(
薛建辉, 2013 ), 通过生物岩溶作用改善裸岩生境, 为草本和灌木的出现奠定了物质基础(
Harper & Belnap, 2001 ;
Jackson, 2015 ), 对岩溶生态系统的维持与发展具有重要意义(
Li et al ., 2006 ;
Costa et al ., 2018 )。苔藓植物岩溶作用已成为岩溶生态学研究的热点。因此, 了解和掌握岩面苔藓植物功能性状特征、岩溶作用机理及其与环境间的互作机制对研究岩溶地区的生物多样性、石漠化生态恢复、岩溶碳汇以及丰富生物岩溶理论等意义重大。然而, 苔藓植物岩溶作用研究因涉及学科众多、问题复杂, 还未进入系统化研究阶段。为了破除各学科间的壁垒, 促进苔藓植物岩溶作用研究更进一步发展, 本文根据近年来有关苔藓植物岩溶作用的文献资料, 从以下方面综述苔藓植物岩溶作用的研究进展。
1 岩面苔藓植物功能性状特征 苔藓植物常以丛集型、交织型、匍匐型等着生于岩面(
图1 ), 假根根系不发达, 只起固着作用(
娄玉霞, 2013 ), 其特殊的叶表面结构和细胞特征(
娄红祥, 2012 ), 使苔藓具有耐高温、抗旱、蓄水能力强、保湿固土等功能(
Proctor & Pence, 2002 ;
Krol et al ., 2003 ;
Zhang et al ., 2017 )。
曹建华和袁道先(1999) 研究发现, 当岩石表面有苔藓植物时, 其蒸发失水和主动吸水时间比裸岩分别延长了48%和57%, 持水量提高了16.6倍, 分析认为苔藓植物可以延缓岩面水分蒸发时间, 增加岩石水化学作用时间, 但是苔藓植物的这种蓄水保湿能力存在明显的种间差异(
申家琛等, 2017 ;
张显强等, 2018a )。苔藓植物参与岩溶作用的另一大优势, 是其叶表面角质层不发育, 且阳离子交换位点多(
吴玉环等, 2001 ), 能够吸附多种阳离子, 这些阳离子在苔藓植物体内被转化为无机盐、有机盐以及其他形式的化合物储存于组织中。苔藓植物除了以生物化学方式对岩石进行溶蚀之外, 还可以通过物理机械作用破坏岩石。例如, 苔藓植物的假根可以伸入岩石裂隙, 对岩石有一定的破坏作用(
Jackson, 2015 )。再者, 当气候干旱时, 苔藓植物在卷缩过程中会拔起岩面的一些小颗粒, 从而加速了岩石被破坏和成土速度(
连宾等, 2008 )。此外, 苔藓植物常与藻类、地衣、微生物等共生形成生物结皮, 它们之间的相互关系以及因气候因素引起的干湿交替和冻融作用对岩石也具有一定的破坏性(
郑云普等, 2009 )。例如, 菌类的菌丝可以伸入岩石晶体结构中破坏岩石, 细菌与真菌形成的配合基能够络合三价铁离子, 细菌分泌的柠檬酸、乙酸以及地衣分泌的地衣酸(
连宾等, 2011 ), 可以溶解岩石中的矿物(
Won-Pyo et al ., 2016 ;
Wang et al ., 2017 )。因此在研究苔藓植物岩溶作用时, 应明确苔藓植物自身及其与其他生物形成的生物结皮两者之间的关系。目前, 有关岩面苔藓植物功能性状的研究主要集中在小范围内特定生境中的苔藓植物研究, 无法代表岩溶生态系统苔藓植物的整体情况。对于大尺度空间上, 因生境异质性高而造成相同物种在形态结构、光合特征及其钙(Ca)、碳(C)、氮(N)、磷(P) 等元素的代谢调节机制等方面的差异性的研究还未引起人们的广泛关注。岩溶生态系统的物质基础相对贫乏, 具有“缺氮少磷钙丰富”的特征。因此, 研究Ca、C、N、P等元素在苔藓植物体内的代谢机制对揭示岩溶生态系统苔藓植物和其他植物的生物多样性维持机制, 及其岩溶作用特征具有重要的科学 意义。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1贵州普定喀斯特生态监测站岩面苔藓植物。A中的苔藓植物主要是多枝缩叶藓、卷叶湿地藓。B中的苔藓植物主要是平蒴藓、皱叶牛舌藓、狭网真藓。 Fig. 1Bryophytes on rock surface at Puding karst ecological monitoring station in Guizhou. A, mosses of Ptychomitriaceae gardenri and Hyophila acutifolia . B, mosses of Plagiobryum zierii, Bryum anagustirete and Anomodon rugelii . 2 苔藓植物对岩石的溶蚀作用 2.1 苔藓植物溶蚀作用原理与过程 苔藓植物对岩石的溶蚀作用原理与过程实质是岩石中矿物与苔藓植物之间的作用过程(
图2 )。苔藓植物因呼吸作用形成的H
2 CO
3 和代谢分泌物与岩石中的矿物发生酸化、碱化、氧化还原、螯合等反应, 随着反应过程不断推进, 岩石矿物成分发生变化, Ca、镁(Mg)、钠(Na)、钾(K)、铁(Fe)、铝(Al)、硅(Si)等从碳酸盐、硅酸盐、铝酸盐、硫化物、氧化物中溶出, 破坏原岩晶体结构, 达到溶蚀岩石, 促使风化的目的(
Street et al ., 2013 ;
Li et al ., 2015 ;
Lammers et al ., 2017 )。这种岩溶驱动力的大小因植物形态特征、岩石性质与生存环境改变而发生变化。有研究表明, 碳酸盐岩溶蚀过程中, 生物成因的CO
2 占溶蚀强度的49.26%, 有机酸占37.11% (
张楷燕, 2017 )。苔藓植物溶蚀作用过程释放的矿物离子和其他元素部分被苔藓植物拦截形成岩溶沉积物; 部分被苔藓植物吸收利用, 转化为其他物质存储于植物体内; 其余则随降雨冲刷而流失, 进入其他生态系统。关于苔藓植物呼吸作用和其分泌物引起的岩溶作用驱动力、效率及其与苔藓植物生活型、种间差异、生境特征之间的相互关系仍未引起人们的关注, 对苔藓植物溶蚀作用过程和机理、苔藓植物对岩石中矿物营养元素释放规律的影响, 以及这些矿物质在岩溶沉积过程中的形态转化过程及其与生境间的相互作用关系等的研究还未见报道, 这也正是揭示苔藓植物岩溶作用机理的关键点。岩石的溶解速率与有机酸的酸性和溶蚀能力密切相关(
刘胜亮等, 2017 ), 然而有机酸的理化特性取决于其官能团的属性(
赵志曼等, 2018 )。因此, 今后需结合有机化学、气相色谱质谱(GC-MS)、核磁共振(NMR)、液相色谱(HPLC)等技术手段深入研究苔藓植物的溶蚀过程与机理。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2苔藓植物岩溶作用过程示意图。岩石表面的苔藓植物通过物理机械力与生物化学作用力破坏和溶蚀岩石, 从而改变了岩面形态, 塑造了岩溶微地貌, 溶蚀产物的沉积形成了原始土壤。 Fig. 2A schematic flowchart showing the bryophyte karstification process. The moss on rock surface through physical and biochemical action to destroy and corrosion the rock, change the rock surface morphology and form the karst microtopography. At the same time, the dissolution products are deposited to form the original soil. 此外, 苔藓植物体内的碳酸酐酶(CA)对岩溶作用也有促进作用(
Smith & Griffiths, 2000 ), 它主要催化CO
2 的水合反应: CO
2 + H
2 O $\rightleftharpoons $ HCO
3 - + H
+ (
吴雁雯和张金池, 2015 )。
李为等(2005) 研究发现无CA催化时上述可逆反应达到平衡的时间是有CA催化时的6 × 10
7 倍。生物体内均含有CA, 且CA活性因物种的不同而存在显著差异, 细菌、真菌、放线菌内均含有CA, 且微生物细胞内CA活性不同于细胞外CA, 细菌的CA对碳酸盐岩系统碳储存贡献在20%以上(
Li et al ., 2006 ), 真菌的CA对白云岩的溶蚀能力强于对石灰岩(
申泰铭等, 2014 ;
申泰铭, 2018 )。那么苔藓植物体内CA对岩溶作用的贡献是多少?各组织内的CA活性是否也存在这种差异?对此以及关于苔藓植物体内CA代谢的调节机制及原理仍不得而知。目前, 关于苔藓植物CA的研究, 主要集中在各类苔藓植物CA含量、活性及其对岩石溶蚀过程的促进作用等方面。例如,
刘天雷等(2017) 研究发现, 虽然苔藓植物都会分泌CA, 但CA分泌量、速率和活性存在种间差异。这种差异性可能是因生活型或者生长环境的不同而导致苔藓植物生理代谢与功能性状的差异引起的。
2.2 苔藓植物溶蚀产物分析 岩石经苔藓植物溶蚀后, 表面常常出现一些凹凸不平和微溶孔, 这是由于岩石成分不均匀, 导致生物钻孔力的差异性(
王福星等, 1998 ), 以及溶蚀产物流失而形成溶蚀微地貌。
王福星等(1993) 将这些溶蚀微地貌, 按照口面形态、直径大小、深度等, 把短径小于1 mm的命名为微溶孔, 把短径为1-20 mm的命名为溶孔, 把短径为20-100 mm的命名为溶坑。
付兰和张朝晖(2010) 将连续分布的苔藓植物溶蚀微地貌, 按测算结果, 把长7.7-14.4 cm的命名为溶丝; 长0.8-19.0 cm, 宽0.6-7.2 cm的命名为溶斑; 长6.0-24.5 cm, 宽1.2-2.2 cm的命名为溶带。苔藓植物溶蚀过程很难通过宏观手段观测, 导致这两种命名法在大小和形态方面存在明显差异。然而,
Haward等(2011) 借助原子力显微镜(AFM)观察到黑云母表面浅层溶蚀坑的形成过程, 通过分析图像计算质量损失, 从而确定溶蚀过程中黑云母表面的活化能大小。研究表明, 通过分析活化能得到的溶坑大小与测量其周长所得的数据一致。因此, 在研究溶蚀微地貌时, 应采用高精度仪器, 统一测量标准, 分析微地貌是苔藓植物溶蚀作用产生, 还是由于淋溶风化而形成, 有利于后期岩溶微地貌之间的对比分析和进一步深入研究。这些微地貌的形态特征与生物类型和发育生境密切相关(
蔡胡霖, 2017 ), 在温湿度优越的地区发育更为广泛(
Liang & Xu, 2014 ;
Liu et al ., 2016 )。例如, 溶孔、溶锥多发生在低湿度、强光照、高温环境中, 而溶沟多是在高温、强光和pH较高的生境内发育(
申家琛等, 2018a )。
曹建华和王福星(1996) 认为苔藓植物岩溶微地貌与岩溶地区的宏观地貌形态之间存在一定的相关性, 然而目前这种关联性以及苔藓植物微地貌之间的关系和发育方向尚未得到科学揭示。
在岩石溶蚀过程中, 有些岩石因产状、矿物成分、气候、生物等因素利于溶蚀产物残留, 沉积物较多, 进而为沉积作用做贡献; 然而有些岩石可溶性矿物含量多, 则溶蚀残留物较少。例如, H
2 CO
3 与碳酸盐岩中的CaCO
3 反应生成Ca(HCO
3 )
2 随水溶失, 残留物较少。有机酸常常与岩石中钙等矿物结合生成有机酸钙等难溶化合物(
张大庚等, 2015 ), 赋存于残积物中, 进而影响土壤的矿物组分与含量。由于生物岩溶作用研究需要长时间尺度的监测, 研究难度较大, 所以对岩面因苔藓植物溶蚀作用而形成的溶蚀残积物中的物质种类、形态特征以及苔藓植物岩溶作用与生境之间关系的研究仍然很欠缺。
3 苔藓植物对岩溶沉积贡献 岩溶地区土壤的形成和发育是碳酸岩长期溶蚀、风化与生物富集过程的结果, 其一方面受岩石结构、性质影响, 另一方面也与植被作用密切相关(
张美良和邓自强, 1994 )。研究表明, 碳酸盐岩的成土速率与岩石溶蚀速率成正比(
曹建华等, 2004 ), 与酸不溶物含量密切相关(
曹沁智等, 2018 )。苔藓植物殖居于岩面, 假根系以及植物体的干湿交替冻融现象所产生的物理机械力作用于岩石, 产生岩石碎屑。同时, 有机物与矿物离子络合常常形成难溶物, 这些难溶物与苔藓及其生物结皮死亡残体黏结在一起, 不仅增加了土壤沉积量, 而且促进有机质积累, 增加土壤养分(
朱显谟, 1995 ;
刘天雷等, 2017 )。苔藓植物通过影响下垫面土壤湿度, 促进土壤对岩石的岩溶作用, 间接地影响沉积过程。同时, 苔藓植物的吸附拦截功能对防治岩石表面水土流失具有重要作用(
李军峰等, 2013 ;
贾少华等, 2014 ;
张显强等, 2018b )。苔藓植物的溶蚀作用为岩面土壤形成提供了原始物质, 关于苔藓对土壤沉积的贡献率与苔藓植物作用下岩面土壤的形成过程, 以及在此过程中岩石的物质、能量转化和土壤养分的变化等还未见报道。
苔藓植物沉积作用的另一表现是石灰华或钙华(
张捷, 1993 ;
Kleinteich et al ., 2017 )。钙华形成的物理化学过程实质是CaCO
3 的沉淀与CO
2 的脱气。苔藓植物在钙华形成过程中为CaCO
3 晶体提供了稳定的生长发育基础, 并且苔藓植物的形态结构控制着沉积物的形态变化(
Mili?a et al ., 2006 ), 且它们的发育形态随沉积环境而变化(
张朝晖等, 2004 ;
郭云等, 2018 )。如在洞穴弱光带一般形成洞穴石刺、向光钟乳石、洞口叠层石等, 这类形态有很强的方向性(
王福星等, 1998 )。苔藓植物在水生环境中也常常形成多样的沉积形态, 如凤尾藓(
Fissidens )钙华、石灰藓(
Hydrogonium )钙华形成的溪流瀑布沉积, 泽藓(
Philonotis )钙华、水灰藓(
Hygrohypnum )钙华形成的河流瀑布沉积, 地钱-真藓(
Marchantia-Bryum )钙华等形成的石灰华坝沉积, 以及净口藓(
Gymnostomum )钙华等形成的瀑布洞穴沉积(
张朝晖和陈家宽, 2007 )。同时, 也有研究发现, 水流环境也是影响苔藓钙华沉积的重要因素, 苔藓与钙华层之间的水流速度与钙华沉积速率呈正相关关系(
Wang et al ., 2014 )。钙华作为大陆古气候的潜在档案,
Yan等(2017) 利用碳氧稳定同位素比率(
δ 13 C和
δ 18 O), 研究钙华在沉积过程中碳、氧的循环规律, 分析认为, 钙华沉积与季节更替引起的气温、降水等的变化密切相关。目前对于苔藓植物沉积作用的研究主要集中在沉积形态以及成土量方面, 对于苔藓植物沉积速率, 对岩溶地区成土过程中的贡献, 以及岩溶沉积作用对生境的改造和环境对岩溶沉积作用的影响等方面的研究还未见报道。
4 影响苔藓植物岩溶作用的因素及其作用机理 4.1 湿度、温度、光照及其综合因子 苔藓植物群落分布特征与生境条件密切相关(
Heino & Virtanen, 2006 ;
刘艳等, 2015 ;
Mod et al ., 2016 ;
李泽科等, 2018 ;
庞嘉鹏等, 2018 ), 温度和水分是影响苔藓植物生存和分布的主要决定性因素, 尤其是空气湿度(
娄红祥, 2012 ;
Guo & Zhao, 2018 )。
党成强等(2018) 研究表明, 南亚毛灰藓(
Homomallium simlaense )在干旱和高温条件下, 各项生理指标均与体内相对含水量呈正比。苔藓植物在干旱缺水的情况下会停止生理代谢进入休眠状态, 当水分充足时再进行生命活动。而且苔藓植物生物量与相对生长速率随光照增强而增加, 随光照时间的延长, 其长度、pH值降低能力逐渐提高(
St. Martin & Mallik, 2017 )。苔藓植物虽然能够忍受高温, 但是温度高于25 ℃时却很少有净光合产物(
Proctor, 2000 ;
Hanson et al ., 2014 )。同时, 温度也是影响酶活性的关键因素, 温度超过60 ℃时会严重影响CA的活性(
洪子茜, 2018 )。然而, CA参与苔藓植物岩溶作用的最佳酶促反应温度还不得而知。同时, 温度也影响有机酸的反应性、络合能力和岩石中矿物自身的溶解程度(
闫志为等, 2009 ;
Benbouzid et al ., 2012 ), 进而影响岩石的溶蚀强度(
陈传平等, 2008 )。而且有机酸与矿物反应生成有机酸盐的存在形态也随温度发生变化(
Peters et al ., 2018 )。
因此, 湿度、温度、光照及其综合因子通过对苔藓植物生理代谢过程、基因型以及其他功能性状的影响(
Pharo & Zartman, 2006 ;
de la Rosa, 2016 ;
解梦, 2018 ), 而直接或间接地影响苔藓植物岩溶作用过程与效率。例如, 当温度在17.5-21.4 ℃, 湿度57.7%-72.6%, 光照2 700-5 900 lx条件下, 苔藓植物对碳酸盐岩的塑造作用最强(
申家琛等, 2018a )。但是目前对温湿度及其综合因子对苔藓植物岩溶作用的影响的关注度仍不够, 很多规律仍不清楚, 然而这些规律的发现是揭示苔藓植物岩溶机理的关键, 也是指导岩溶生态问题治理的重要科学依据。
4.2 pH与其他营养元素 Xie和Lou (2009) 认为, 生物岩溶作用随pH值降低而增强, 如果单纯从H
+ + CaCO
3 $\rightleftharpoons $ CO
2 + H
2 O + Ca
2+ 角度分析pH对岩溶作用的影响, pH值越低溶蚀速率越快。然而岩石的最佳溶解pH由矿物自身属性决定(
Bray et al ., 2015 ), 同时有机酸的溶蚀能力也受pH影响(
Pachana et al ., 2012 ;
Chen et al ., 2016 ), 而且每类苔藓植物的最适生长pH也不尽相同。如泥炭藓属(
Sphagnum )植物适宜偏酸性环境, 细叶小羽藓(
Haplocladium microphyllum )配子体的最适生长pH值为7 (
陈文佳等, 2012 )。作为苔藓植物岩溶作用的重要参与者CA, 它的活性也受pH影响,
洪子茜(2018) 研究表明, 当pH在7-9条件下CA的活性最高。pH由外而内对苔藓植物岩溶作用的过程、速率、对象、各级参与者都产生不同程度的促进或抑制作用。目前我们对pH影响苔藓植物岩溶的规律以及调节机制还不得而知, 今后还需综合全面地进行深入研究。
由于苔藓植物属于贫营养型, 它所需要的营养物质是维管束植物的1/10, 但是对N、P、K、Ca、Mg、Fe、硫(S)等元素的需求仍必不可少(
娄红祥, 2012 )。Ca
2+ 作为生物的第二信使, 具有调节生物抗逆性等多种功能, 对苔藓植物的生长有显著影响(
Amao et al ., 2010 ;
Kolupaev et al ., 2014 )。当苔藓植物缺Ca时会导致生长缓慢, 假根分化少, 原丝体不发育, 叶状体形状改变等(
娄红祥, 2012 )。而且CA活性也受Fe
2+ 、锌离子(Zn
2+ )、硝酸根(NO
- 3 )、氯离子(Cl
- )、溴离子(Br
- )、有机碳、交换性钙等的显著影响(
洪子茜, 2018 )。例如,
黄芬等(2015) 研究表明, Ca对CA的活性有很强的促进作用。生物与环境协同进化致使某些苔藓出现钙依赖性, 这种对钙的依赖性是否表现在基因水平, 从而影响岩溶地区苔藓植物生物多样性、空间分布特征、功能性状等还有待进一步研究。因此在研究苔藓植物岩溶作用与pH及营养元素之间的关系时, 应综合考虑苔藓植物种类、酶活性、逆境胁迫、水环境、基岩性质等因素。
4.3 岩石性质 岩石中所含矿物种类及其晶体结构、产状、透水性等决定了岩石的溶解能力(
Voinot et al ., 2013 ;
黄奇波等, 2015 ), 例如, 一般情况下泥质含量高的碳酸盐岩溶蚀速度快, 岩石中酸不溶物含量越低, 越易被溶解(
王世杰等, 1999 )。同时, 岩石中酸不溶物含量对碳酸盐岩的成土速率具有重要影响, 酸不溶物含量越高岩石风化后可残留物越多, 成土物质越丰富(
陈洪松等, 2018 )。例如, 白云岩中酸不溶物含量高, 发育的土层较石灰岩地区土层厚(
陈洪松等, 2013 )。岩石中所含矿物的性质影响各元素的释放速率, Andrew等(2015)在25 ℃、pH = 2.6的开放试验系统与封闭试验系统内发现黑云母中的元素释放按照Fe > Mg > Al > Si的顺序进行, 且溶解速率符合 ${{r}_{i}}={{k}_{\text{H},i}}\times a_{{{\text{H}}^{\text{+}}}}^{xi}$,
k H 为表面速率常数((2.15-30.6) × 10
-10 mole s
biotite ·m
-2 ·s
-1 ),
a H+ 为H
+ 活性,
xi 为矿物与H
+ 的反应级数(0.31-0.58); 分析认为, 岩石中各矿物的溶解具有空间各向异性, 其边缘的反应效率强于基面, 而且各个元素的释放具有一定的规律性。苔藓植物种类的空间分布特征与岩性密切相关(
Gorozhankina & Konstantinov, 2001 ), 而且岩性和有机酸种类也会影响有机酸与岩石的反应速率(
Cama & Ganor, 2006 ;
Ghoorah et al ., 2014 ;
Mwangi et al ., 2018 ), 从而导致它们岩溶作用的差异性。
5 研究不足和展望 苔藓植物岩溶作用是苔藓植物与岩石相互博弈的动态过程。近年来苔藓植物岩溶作用取得了不少研究进展, 主要集中在苔藓持水特性与岩溶作用的关系, 生物酶对溶蚀作用的影响, 苔藓植物及其生物结皮的成土作用、钙华沉积以及苔藓植物溶蚀微地貌及其与生境间的相互关系探讨方面。但也存在不足之处, 以下4个方面更为突出。
5.1 苔藓植物岩溶作用内在机制与过程 目前, 苔藓植物岩溶作用以定性描述为主, 溶蚀作用发生的机理、条件, 哪些物质参与溶蚀?这些物质与岩石的反应过程、化学键的断裂与重组方式、能量释放、产物的定性与定量分析, 以及它们的溶蚀速率与成土速率如何?不同生活型或不同物种之间是否存在岩溶作用的差异性?这些基础而又关键的问题仍不清楚。因此, 今后需要加强苔藓植物岩溶作用效率、驱动机制及其动态变化等内在机制与过程的研究。
5.2 苔藓植物岩溶作用与生境间互作机制 苔藓植物岩溶作用与环境之间的三向循环互作机制如
图3 所示, 苔藓、环境因子、岩石三者之间相互联系、相互作用。
申家琛等(2018b) 阐释了苔藓植物溶蚀微地貌发育及其分布特征与生境(温湿度、光照等)的关系, 但是对苔藓植物岩溶作用对生境的改造, 以及生态因子对苔藓植物岩溶作用的影响还未揭示。因此, 今后需要在这方面开展以下研究工作: (1)温度、湿度、光照、pH、地质背景等因子对苔藓植物生物多样性、生存繁殖, 生理调节机制、功能性状等的影响; (2)环境因子对苔藓植物溶蚀过程、效率、物质能量转化过程等的影响; (3)苔藓植物沉积过程对岩面微环境的温湿度、地貌塑造、土壤养分、水动力特征等的影响; (4)苔藓植物与其内生菌和藻类的共生关系以及与其他植物之间的化感作用对岩溶作用的影响。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3苔藓植物岩溶作用与环境、岩石间的三向循环互作机制图。苔藓、环境因子、岩石在岩溶作用过程中三者既相互促进又相互制约。环境因子与岩石通过控制苔藓植物的群落特征、形态、生理过程、基因等影响岩溶过程。反之基岩的岩性、组成、产状, 以及苔藓植物对岩面生境(温度、湿度、光照、土壤肥力、微生物)的改善与岩溶速率密切相关。 Fig. 3The three-way circulation interaction mechanism among bryophyte karstification, environment and rock. Bryophyte, environmental factors and rock in the karstification process promote and restraint each other. Environmental factors and rock affect karstification process and efficiency by controlling the community characteristics, morphology, physiological processes, genes, etc. of bryophyte. The lithology, composition, occurrence of rock, and the improvement of bryophyte on rock surface habitat (temperature, humidity, light, soil fertility, microbes) are closely related to the rate of karstification. 5.3 苔藓植物的生理代谢过程及其生物酶等与岩溶作用的相互关系 苔藓植物体内的CA对岩溶作用有促进作用, 但是CA活性存在显著的种间差异, 如何通过植物生理或者基因学调控CA代谢过程, 提高酶活性?苔藓植物呼吸作用产生的H
2 CO
3 与代谢分泌物的溶蚀作用差异性及相互关系?分泌物的代谢调控机制?这些科学问题的阐释对揭示苔藓植物岩溶作用机理、调控岩溶过程等具有重要的理论指导意义。
5.4 岩溶地区苔藓植物环境适应性特征及其生物多样性维持机制 土层浅薄、贫瘠、钙丰富是岩溶生态系统的典型特征。苔藓植物与岩溶生境在长期协同进化过程中形成了与环境相适应的功能性状特征。目前, 对岩溶地区苔藓植物环境适应性特征与生态系统过程和功能关系的认识还十分有限, 无法揭示岩溶生态系统生物多样性维持机制。因此, 需要开展苔藓植物形态结构特征、生理调节机制(光合、呼吸、抗逆性、物质代谢等)、养分需求及利用方式、繁殖方式、群落组成结构特征、物种多样性、遗传多样性等与岩溶生态系统结构、功能、生物多样性之间的互作机制。同时, 植物功能性状与生态系统间的耦合效应会成为今后植物生态学研究的热点问题。
此外, 建议在苔藓植物岩溶作用研究时建立野外长期监测基地, 监测苔藓植物对全球变化(增温、降水格局、氮沉降等)响应及其对岩溶作用的影响。同时, 通过同位素示踪法研究苔藓植物岩溶作用过程中碳循环路径, 建立大气-水环境-生物-土壤-基岩“五位一体”的碳循环研究模式, 进而为研究岩溶生态系统碳循环、CO
2 收支与排放量、碳汇问题、全球变暖等提供新思路、新方法。
致谢 感谢贵州省林业厅攻关项目([2016]12号)和贵州科学院青年重点基金([2020]03号)资助。
[1] Amao Y Takai K Ohashi A 2010 ). Effect of manganese and calcium ions on the photoinduced water oxidation with photosynthesis organ grana from green plantApplied Catalysis B: Environmental , 97, 36 -40 . [本文引用: 1] [2] Benbouzid H Le Floch S Stephan L Olier R Privat M 2012 ). Combined effects of salinity and temperature on the solubility of organic compoundsThe Journal of Chemical Thermodynamics , 48, 54 -64 . [本文引用: 1] [3] Bray AW Oelkers EH Bonneville S Wolff-Boenisch D Potts NJ Fones G Benning LG 2015 ). The effect of pH, grain size, and organic ligands on biotite weathering ratesGeochimica et Cosmochimica Acta , 164, 127 -145 . [本文引用: 1] [4] Cai HL 2017 ). Surface Dissolved Forms of Carbonate Rocks and Its Influencing Factors Master degree dissertation, Yunnan Normal University, Kunming . [ 蔡胡霖 (2017 ). 碳酸盐岩表面溶蚀形态及其影响因素硕士学位论文, 云南师范大学, 昆明 .] [5] Cama J Ganor J 2006 ). The effects of organic acids on the dissolution of silicate minerals: A case study of oxalate catalysis of kaolinite dissolutionGeochimica et Cosmochimica Acta , 70, 2191 -2209 . [本文引用: 1] [6] Cao JH Wang FX 1996 ). Observations on fractal characters between karst macrophology and biokarst micromorphology in the Guilin areaJournal of Nanjing University (Natural Science )146 -147 . [ 曹建华 , 王福星 (1996 ). 桂林地区生物岩溶微观形态与岩溶地貌宏观形态间的分形特征南京大学学报(自然科学版) , 32, 146 -147 .] [7] Cao JH Yuan DX 1999 ). Relationship between water-holding of carbonate rock and saxicolous algae, lichen and moss and its ecological significanceGeochimica, 28, 248 -256 . [本文引用: 1] [ 曹建华 , 袁道先 (1999 ). 石生藻类、地衣、苔藓与碳酸盐岩持水性及生态意义地球化学 , 28, 248 -256 .] [本文引用: 1] [8] Cao JH Yuan DX Zhang C Jiang ZC 2004 ). Karst ecosystem constrained by geological conditions in southwest ChinaEarth and Environment, 1, 1 -8 . [本文引用: 2] [ 曹建华 , 袁道先 , 章程 , 蒋忠诚 (2004 ). 受地质条件制约的中国西南岩溶生态系统地球与环境 , 1, 1 -8 .] [本文引用: 2] [9] Cao QZ Deng M Huang B Chen B 2018 ). Influence of acid-insoluble residue on dedolomite reaction in dolomitic rocksJournal of Nanjing Tech University (Natural Science Edition )89 -94 . [本文引用: 1] [ 曹沁智 , 邓敏 , 黄蓓 , 陈碧 (2018 ). 白云质岩石中酸不溶物对碱白云石反应的影响南京工业大学学报(自然科学版) , 40, 89 -94 .] [本文引用: 1] [10] Chen CP Gu X Zhou SM Liu JP 2008 ). Experimental research on dissolution dynamics of main minerals in several aqueous organic acid solutionsActa Geologica Sinica, 7, 1007 -1012 . [本文引用: 1] [ 陈传平 , 固旭 , 周苏闽 , 刘建平 (2008 ). 不同有机酸对矿物溶解的动力学实验研究地质学报 , 7, 1007 -1012 .] [本文引用: 1] [11] Chen HS Fu ZY Zhang W Nie YP 2018 ). Soil water processes and vegetation restoration in karst region of southwest ChinaChinese Journal of Nature, 40, 41 -46 . [本文引用: 1] [ 陈洪松 , 付智勇 , 张伟 , 聂云鹏 (2018 ). 西南喀斯特地区水土过程与植被恢复重建自然杂志 , 40, 41 -46 .] [本文引用: 1] [12] Chen HS Nie YP Wang KL 2013 ). Spatio-temporal heterogeneity of water and plant adaptation mechanisms in karst regions: A reviewActa Ecologica Sinica, 33, 317 -326 . [本文引用: 1] [ 陈洪松 , 聂云鹏 , 王克林 (2013 ). 岩溶山区水分时空异质性及植物适应机理研究进展生态学报 , 33, 317 -326 .] [本文引用: 1] [13] Chen WJ Zhang N Hang LL Wang Y Ji MC 2012 ). Effects of different medium and pH value on the growth of gametophyte in Haplocladium microphyllum Practical Forestry Technology , 4 ), 20 -23 . [本文引用: 1] [ 陈文佳 , 张楠 , 杭璐璐 , 王媛 , 季梦成 (2012 ). 不同培养基及pH值对细叶小羽藓配子体生长的影响林业实用技术 , (4 ), 20 -23 .] [本文引用: 1] [14] Chen ZY Liu F Bu TD Liu YS Zhu J 2016 ). Effects of organic acids on dissolution of Fe and Mn from weathering coal gangueActa Geochimica , 3, 316 -328 . [本文引用: 1] [15] Costa DP Amado-Filho GM Pereira RC Paradas WC Miyataka H Okamoto Y Asakawa Y 2018 ). Diversity of secondary metabolites in the liverwort Syzygiella rubricaulis(Nees) Stephani (Jamesoniellaceae, Marchantiophyta) from neotropical high mountainsChemistry & Biodiversity , 15, e1800239 . DOI: 10.1002/cbdv.201800239 . [本文引用: 1] [16] Dang CQ Li ZF Chen M Gao T Huang HM Liu JC Tao JP 2018 ). Physiological and biochemical characteristics of epilithic moss Homomallium simlaense(Mitt.) Broth. Mitt under high temperature and drought stressPlant Science Journal 393 -401 . [本文引用: 1] [ 党成强 , 李宗峰 , 陈淼 , 高婷 , 黄慧敏 , 刘锦春 , 陶建平 (2018 ). 石生南亚毛灰藓在不同温度和干旱条件下的生理生化特性植物科学学报 , 36, 393 -401 .] [本文引用: 1] [17] de la Rosa JPM 2016 ). The Burren: A glacial, karstic and biokarstic expression of a limestone plateau in western IrelandEarth Surface Processes and Landforms , 41, 1614 -1628 . [本文引用: 1] [18] Fu L Zhang ZH 2010 ). Preliminary study on biokarst erosion of bryophytes in Guiyang cityJournal of Guizhou Normal University (Nature Science )4 ), 140 -143 . [本文引用: 1] [ 付兰 , 张朝晖 (2010 ). 贵阳市苔藓植物的生物岩溶溶蚀初探贵州师范大学学报(自然科学版) , 28(4 ), 140 -143 .] [本文引用: 1] [19] Ghoorah M Dlugogorski BZ Balucan RD Kennedy EM 2014 ). Selection of acid for weak acid processing of wollastonite for mineralisation of CO2 Fuel , 122, 277 -286 . [本文引用: 1] [20] Gorozhankina SM Konstantinov VD 2001 ). Comparative ecocenotic characteristics of mosses in the taiga zone of West SiberiaRussian Journal of Ecology , 32, 386 -392 . [本文引用: 1] [21] Guo Y Wang ZH Zhang ZH 2018 ). Study on community characteristics of bryophytes in dolomite cave twilight zones: An example of the Shuidong Cave in Suiyang county, Guizhou ProvinceCarsologica Sinica, 37, 388 -399 . [本文引用: 1] [ 郭云 , 王智慧 , 张朝晖 (2018 ). 白云岩洞穴洞口弱光带的苔藓群落特征——以绥阳水洞为例中国岩溶 , 37, 388 -399 .] [本文引用: 1] [22] Guo YW Zhao YG 2018 ). Effects of storage temperature on the physiological characteristics and vegetative propagation of desiccation-tolerant mossesBiogeoscience , 15, 797 -808 . [本文引用: 1] [23] Hanson DT Renzaglia K Villarreal JC 2014 ). Diffusion limitation and CO2 concentrating mechanisms in bryophytes. In: Hanson DT, Rice SK eds. Photosynthesis in Bryophytes and Early Land PlantsSpringer, Dordrecht, Netherlands . 95 -111 . [本文引用: 1] [24] Harper KT Belnap J 2001 ). The influence of biological soil crusts on mineral uptake by associated vascular plantsJournal of Arid Environments , 47, 347 -357 . [本文引用: 1] [25] Haward SJ Smits MM Ragnarsdóttir KV Leake JR Banwart SA McMaster TJ 2011 ). In situ atomic force microscopy measurements of biotite basal plane reactivity in the presence of oxalic acidGeochimica et Cosmochimica Acta , 75, 6870 -6881 . [本文引用: 1] [26] Heino J Virtanen R 2006 ). Relationships between distribution and abundance vary with spatial scale and ecological group in stream bryophytesFreshwater Biology , 51, 1879 -1889 . [本文引用: 1] [27] Hong ZX 2018 ). Study on the characteristics of microbial carbonic anhydraseBiological Chemical Engineering, (4 ), 63 -66 . [本文引用: 3] [ 洪子茜 (2018 ). 微生物碳酸酐酶特性研究生物化工 , (4 ), 63 -66 .] [本文引用: 3] [28] Huang F Huang YM Gao X Cao JH 2015 ). Effects of karst environmental factors on activity of soil microorganic extracellular carbonic anhydrase of karst area in Maocun village, GuilinJournal of Southern Agriculture, 46, 1792 -1797 . [本文引用: 1] [ 黄芬 , 黄艳梅 , 高喜 , 曹建华 (2015 ). 岩溶环境因子对桂林毛村岩溶区土壤微生物胞外碳酸酐酶活性的影响南方农业学报 , 46, 1792 -1797 .] [本文引用: 1] [29] Huang QB Qin XQ Liu PY Lan FN Zhang LK 2015 ). Dissolution rate and it’s significance of different lithological tabletsEarth and Environment, 43, 379 -385 . [本文引用: 1] [ 黄奇波 , 覃小群 , 刘朋雨 , 蓝芙宁 , 张连凯 (2015 ). 不同岩性试片溶蚀速率差异及意义地球与环境 , 43, 379 -385 .] [本文引用: 1] [30] Jackson TA 2015 ). Weathering, secondary mineral genesis, and soil formation caused by lichens and mosses growing on granitic gneiss in a boreal forest environmentGeoderma , 251, 78 -91 . [本文引用: 2] [31] Jia SH Li JF Wang ZH Zhang ZH 2014 ). Ecological function of bryophyte on karst rocky desertification slopes along mountainous roadsChinese Journal of Ecology, 33, 1928 -1934 . [本文引用: 1] [ 贾少华 , 李军峰 , 王智慧 , 张朝晖 (2014 ). 岩溶山区公路石漠化边坡苔藓生态功能生态学杂志 , 33, 1928 -1934 .] [本文引用: 1] [32] Kleinteich J Golubic S Pessi IS Velázquez D Storme J-Y Darchambeau F Borges AV Compère P Radtke G Lee S-J Javaux EJ Wilmotte A 2017 ). Cyanobacterial contribution to travertine deposition in the Hoyoux River system, BelgiumMicrobial Ecology , 74, 33 -53 . [本文引用: 1] [33] Kolupaev YE Vayner AA Yastreb TO Oboznyi AI Khripach VA 2014 ). The role of reactive oxygen species and calcium ions in the implementation of the stress protective effect of brassinosteroids on plant cellsApplied Biochemistry and Microbiology , 50, 658 -663 . [本文引用: 1] [34] Krol E Dziubinska H Trebacz K 2003 ). Low-temperature induced transmembrane potential changes in the liverwort Conocephalum conicum Plant & Cell Physiology 527 -533 . [本文引用: 1] [35] Lammers K Smith MM Carroll SA 2017 ). Muscovite dissolution kinetics as a function of pH at elevated temperatureChemical Geology , 466, 149 -158 . [本文引用: 1] [36] Li JF Wang ZH Zhang ZH 2013 ). Bryophyte diversity and the effect of soil formation along with water conservation in karst rocky desertification regionResearch of Environmental Sciences, 26, 759 -764 . [本文引用: 1] [ 李军峰 , 王智慧 , 张朝晖 (2013 ). 喀斯特石漠化山区苔藓多样性及水土保持研究环境科学研究 , 26, 759 -764 .] [本文引用: 1] [37] Li T Wang HY Zhou ZJ Chen XQ Zhou JM 2015 ). A nano-scale study of the mechanisms of non-exchangeable potassium release from micasApplied Clay Science , 118, 131 -137 . [本文引用: 1] [38] Li W Yu LJ Yu JF Jia LP Wu Y 2005 ). Effects of karst environmental factors on expression and activity of bacterial extracellular carbonic anhydraseMicrobiology China, 5, 35 -39 . [本文引用: 1] [ 李为 , 余龙江 , 余俊峰 , 贾丽萍 , 吴云 (2005 ). 岩溶环境因子对细菌胞外碳酸酐酶表达及活性的影响微生物学通报 , 5, 35 -39 .] [本文引用: 1] [39] Li W Yu LT Wu Y Jia LP Yuan DX 2006 ). Enhancement of Ca 2 + release from limestone by microbical extracelluar carbonic anhydrase Bioresource Technology , 98, 950 -953 . [本文引用: 2] [40] Li ZK Wu QM Wang ZH Zhang ZH 2018 ). Diversity and ecological characteristics of bryophytes from peak cluster under the background of karst urbanBulletin of Botanical Research, 38, 433 -443 . [本文引用: 1] [ 李泽科 , 吴启美 , 王智慧 , 张朝晖 (2018 ). 岩溶城市背景下贵阳市峰丛苔藓植物多样性分布及其生态特征植物研究 , 38, 433 -443 .] [本文引用: 1] [41] Lian B Chen Y Zhu LJ Yang RD 2008 ). Progress in the study of the weathering of carbonate rock by microbesEarth Science Frontiers, 15, 90 -99 . [本文引用: 1] [ 连宾 , 陈烨 , 朱立军 , 杨瑞东 (2008 ). 微生物对碳酸盐岩的风化作用地学前缘 , 15, 90 -99 .] [本文引用: 1] [42] Lian B Yuan DX Liu ZH 2011 ). Effect of microbes on karstification in karst ecosystemsChinese Science Bulletin, 56, 2158 -2161 . [本文引用: 1] [ 连宾 , 袁道先 , 刘再华 (2011 ). 岩溶生态系统中微生物对岩溶作用影响的认识科学通报 , 56, 2158 -2161 .] [本文引用: 1] [43] Liang FY Xu B 2014 ). Discrimination of tower, cockpit, and non-karst landforms in Guilin, Southern China, based on morphometric characteristicsGeomorphology 42 -48 .? [本文引用: 1] [44] Liu MX Xu XL Wang DB Alexander YS Wang KL 2016 ). Karst catchments exhibited higher degradation stress from climate change than the non-karst catchments in southwest China: An ecohydrological perspectiveJournal of Hydrology , 535, 173 -180 . [本文引用: 1] [45] Liu SL Zhu SL Li J Yang Y Li JG 2017 ). A study on the ability of different organic acids to dissolve tricalcium phosphateActa Agriculturae Universitatis Jiangxiensis, 39, 1010 -1016 . [本文引用: 1] [ 刘胜亮 , 朱舒亮 , 李静 , 杨越 , 李建贵 (2017 ). 不同有机酸对磷酸三钙溶解能力的研究江西农业大学学报 , 39, 1010 -1016 .] [本文引用: 1] [46] Liu TL Cong CL Hu D Wang SJ Zhang XQ 2017 ). Carbonic anhydrase activity of six epilithic mosses and their underlying soil in the Puding karst area, Guizhou ProvinceCarsologica Sinica, 36, 187 -192 . [本文引用: 2] [ 刘天雷 , 从春蕾 , 胡丹 , 王世杰 , 张显强 (2017 ). 贵州普定6种喀斯特石生植物及其土壤的碳酸酐酶活性中国岩溶 , 36, 187 -192 .] [本文引用: 2] [47] Liu Y Pi CY Tian S 2015 ). Relationships between characteristics of ground bryophyte communities and environmental factors in urban area of Chongqing, ChinaChinese Journal of Applied Ecology, 26, 3145 -3152 . [本文引用: 1] [ 刘艳 , 皮春燕 , 田尚 (2015 ). 重庆主城区地面苔藓植物群落特征及其与环境的关系应用生态学报 , 26, 3145 -3152 .] [本文引用: 1] [48] Lou HX 2012 ). Bryophyte Chemistry and BiologyScience Press, Beijing . [本文引用: 4] [ 娄红祥 (2012 ). 苔藓化学与生物学 . 科学出版社, 北京.] [本文引用: 4] [49] Lou YX 2013 ). Study on Response Mechanism and Biological Indicators of Bryophytes to Heavy Metal PollutionPhD dissertation, Shanghai Normal University, Shanghai . [本文引用: 1] 娄玉霞 (2013 ). 苔藓植物对重金属污染的响应机理和生物指示的研究博士学位论文, 上海师范大学, 上海 .] [本文引用: 1] [50] Mili?a M Habdija I Primc-Habdija B Radanovi? I Kep?ija RM 2006 ). The role of flow velocity in the vertical distribution of particulate organic matter on moss-covered travertine barriers of the Plitvice Lakes (Croatia)Hydrobiologia , 553, 231 -243 . [本文引用: 1] [51] Mod HK Heikkinen RK le Roux PC V?re H Luoto M 2016 ). Contrasting effects of biotic interactions on richness and distribution of vascular plants, bryophytes and lichens in an arctic-alpine landscapePolar Biology , 39, 649 -657 . [本文引用: 1] [52] Mwangi P Brady PV Radonjic M Thyne G 2018 ). The effect of organic acids on wettability of sandstone and carbonate rocksJournal of Petroleum Science & Engineering , 165, 428 -435 . [本文引用: 1] [53] Pachana K Zuddas P Censi P 2012 ). Influence of pH and temperature on the early stage of mica alterationApplied Geochemistry , 27, 1738 -1744 . [本文引用: 1] [54] Pang JP Wang ZH Zhang ZH 2018 ). Characteristics of bryophyte communities and their successional patterns in different habitats from karst dolomite rocky desertification areasEcological Science, 37, 59 -66 . [本文引用: 1] [ 庞嘉鹏 , 王智慧 , 张朝晖 (2018 ). 喀斯特白云岩石漠化区域不同生境条件下苔藓植物群落特征及演替模式研究生态科学 , 37, 59 -66 .] [本文引用: 1] [55] Peters K Gorzolka K Bruelheide H Neumann S 2018 ). Seasonal variation of secondary metabolites in nine different bryophytesEcology and Evolution , 8, 9105 -9117 . [本文引用: 1] [56] Pharo EJ Zartman CE 2006 ). Bryophytes in a changing landscape: The hierarchical effects of habitat fragmentation on ecological and evolutionary processesBiological Conservation , 135, 315 -325 . [本文引用: 1] [57] Proctor MCF 2000 ). The bryophyte paradox: Tolerance of desiccation, evasion of droughtPlant Ecology , 151, 41 -49 . [本文引用: 1] [58] Proctor MCF Pence VC 2002 ). Vegetative tissues: Bryophytes vascular “resurrection plants” and vegetative propagules. In: Pritchard H, Back M eds. Desiccation and Plant SurvivalCABI Publishing, Wallingford, UK . 207 -237 . [59] Schneider J 1976 ). Biological and inorganic factors in the destruction of limestone coastsContributions to Sedimentology , 6, 112 -116 . [本文引用: 1] [60] Shen JC Zhang ZH Wang HH Huang H Wang ZH 2017 ). Water retention capacity of autumn mosses in south stone forest of Guiyang karst parkJournal of Ecology and Rural Environment, 33, 907 -912 . [本文引用: 1] [ 申家琛 , 张朝晖 , 王慧慧 , 黄欢 , 王智慧 (2017 ). 贵阳喀斯特公园南石林秋季藓类植物的持水特性生态与农村环境学报 , 33, 907 -912 .] [本文引用: 1] [61] Shen JC Zhang ZH Wang HH Huang H Wang ZH 2018a ). Corrosion effects and environmental correlation of bryophytes on limestone in Guiyang karst parkCarsologica Sinica, 37, 175 -184 . [本文引用: 2] [ 申家琛 , 张朝晖 , 王慧慧 , 黄欢 , 王智慧 (2018a ). 苔藓植物对石灰岩的溶蚀作用及环境相关性研究中国岩溶 , 37, 175 -184 .] [本文引用: 2] [62] Shen JC Zhang ZH Wang ZH 2018b ). The effects of rocky desertification degree on bryophyte diversity and soil chemical properties of crustsActa Ecologica Sinica, 38, 6043 -6054 . [本文引用: 1] [ 申家琛 , 张朝晖 , 王智慧 (2018b ). 石漠化程度对苔藓植物多样性及其结皮土壤化学性质的影响生态学报 , 38, 6043 -6054 .] [本文引用: 1] [63] Shen TM 2018 ). Contribution and Mechanism of Microbes and Carbonic Anhydrase to Carbon Storage Soil EcosystemsPhD dissertation, Huazhong University of Science and Technology, Wuhan . [ 申泰铭 (2018 ). 微生物及碳酸酐酶对岩溶土壤生态系统碳储存的贡献及其机制博士学位论文, 华中科技大学, 武汉 .] [64] Shen TM Xin BG Li W Yu LJ 2014 ). Characteristics of carbonate rock corrosion by different kinds of microbea and their carbonic anhydras in the CO2 -H2 O-carbonate systemBulletin of Mineralogy, Petrology and Geochemistry, 33, 797 -800 . [本文引用: 1] [ 申泰铭 , 邢必果 , 李为 , 余龙江 (2014 ). 不同种类微生物及其碳酸酐酶对CO2 -H2 O-碳酸盐系统中碳酸盐岩的溶蚀作用矿物岩石地球化学通报 , 33, 797 -800 .] [本文引用: 1] [65] Smith EC Griffiths H 2000 ). The role of carbonic anhydrase in photosynthesis and the activity of the carbon- concentrating-mechanism in bryophytes of the class anthocerotaeNew Phytologist , 145, 29 -37 . [本文引用: 1] [66] St. Martin P Mallik AU 2017 ). The status of non-vascular plants in trait-based ecosystem function studiesPerspectives in Plant Ecology, Evolution and Systematics , 27, 1 -8 . [本文引用: 1] [67] Street LE Subke J-A Sommerkorn M Sloan V Ducrotoy H Phoenix GK Williams M 2013 ). The role of mosses in carbon uptake and partitioning in arctic vegetationNew Phytologist , 199, 163 -175 . [本文引用: 1] [68] Voinot A Lemarchand D Collignon C Granet M Chabaux F Turpault M-P 2013 ). Experimental dissolution vs. transformation of micas under acidic soil conditions: Clues from boron isotopesGeochimica et Cosmochimica Acta , 117, 144 -160 . [本文引用: 1] [69] Wang FX Cao JH Huang JF 1998 ). Biokarst in the cave twilight zonesCarsologica Sinica, 17, 42 -48 . [本文引用: 2] [ 王福星 , 曹建华 , 黄俊发 (1998 ). 洞穴弱光带的生物岩溶中国岩溶 , 17, 42 -48 .] [本文引用: 2] [70] Wang FX Cao JH Huang JF Jiang LD Huang JF Wang J 1993 ). Biokarst . Geological Publishing House, Beijing. [本文引用: 1] [ 王福星 , 曹建华 , 黄俊发 , 江利登 , 黄基富 , 王晶 (1993 ). 生物岩溶 . 地质出版社, 北京.] [本文引用: 1] [71] Wang GM Xiong ZH Zhang J Zhang B 2017 ). Dissolution experiment and transformation condition analysis of Paleogene aragonite in the Jiyang Depression, ChinaAustralian Journal of Earth Sciences , 64, 343 -352 . [本文引用: 1] [72] Wang HJ Yan H Liu ZH 2014 ). Contrasts in variations of the carbon and oxygen isotopic composition of travertines formed in pools and a ramp stream at Huanglong Ravine, China: Implications for paleoclimatic interpretationsGeochimica et Cosmochimica Acta , 125, 34 -48 . [本文引用: 1] [73] Wang SJ Ji HB Ouyang ZY Zhou DQ Zheng LP Li TY 1999 ). Preliminary study on weathering and soil formation of carbonate rocksScience in China (Series D )441 -449 . [本文引用: 1] [ 王世杰 , 季宏兵 , 欧阳自远 , 周德全 , 郑乐平 , 黎廷宇 (1999 ). 碳酸盐岩风化成土作用的初步研究中国科学(D辑: 地球科学 ), 29, 441 -449 .] [本文引用: 1] [74] Won-Pyo P Bon-Jun K Chang AC Ferko TE Parker JR Ward TH Lara SV Nguyen CM 2016 ). Dissolution of metals from biosolid-treated soils by organic acid mixturesApplied and Environmental Soil Science , 2016 , 1 -15 . [本文引用: 1] [75] Wu YH Huang GH Gao Q Cao T 2001 ). Research advance in response and adaptation of bryophytes to environmental changeChinese Journal of Applied Ecology, 12, 943 -946 . [本文引用: 1] [ 吴玉环 , 黄国宏 , 高谦 , 曹同 (2001 ). 苔藓植物对环境变化的响应及适应性研究进展应用生态学报 , 12, 943 -946 .] [本文引用: 1] [76] Wu YW Zhang JC 2015 ). Microbial carbonic anhydrase action and application on carbon cycling in karst dynamic system: A reviewJournal of Biology, 32, 78 -83 . [本文引用: 1] [ 吴雁雯 , 张金池 (2015 ). 微生物碳酸酐酶在岩溶系统碳循环中的作用与应用研究进展生物学杂志 , 32, 78 -83 .] [本文引用: 1] [77] Xie CF Lou HX 2009 ). Secondary metabolites in bryophytes: An ecological aspectChemistry & Biodiversity , 6, 303 -312 . [本文引用: 1] [78] Xie M 2018 ). Determination of DNA C-values and Analysis of Variation Characteristics of BryophytesMaster degree dissertation, Shanghai Normal University, Shanghai . [本文引用: 1] [ 解梦 (2018 ). 苔藓植物DNA C-值测定及其变异特点分析硕士学位论文, 上海师范大学, 上海 .] [本文引用: 1] [79] Xue JH 2013 ). Forest Ecology . China Forestry Publishing House, Beijing. [ 薛建辉(2013 ). 森林生态学 . 中国林业出版社, 北京.] [80] Yan H Liu ZH Sun HL 2017 ). Effect of in-stream physicochemical processes on the seasonal variations in δ 13 C and δ 18 O values in laminated travertine deposits in a mountain stream channel Geochimica et Cosmochimica Acta , 202, 179 -189 . [本文引用: 1] [81] Yan ZW Liu HL Zhang ZW 2009 ). Influences of temperature and PCO2 on the solubility of calcite and dolomiteCarsologica Sinica, 28, 7 -10 . [本文引用: 1] [ 闫志为 , 刘辉利 , 张志卫 (2009 ). 温度及CO2 对方解石、白云石溶解度影响特征分析中国岩溶 , 28, 7 -10 .] [本文引用: 1] [82] Zhang DG Li J Liu H 2015 ). Effect of exogenous low molecular weight organic acids on calcium migration in soilJournal of Soil and Water Conservation, 29, 152 -155 . [本文引用: 1] [ 张大庚 , 栗杰 , 刘慧 (2015 ). 外源低分子量有机酸对土壤中钙素迁移特征的影响水土保持学报 , 29, 152 -155 .] [本文引用: 1] [83] Zhang J 1993 ). Biological role in the formation of karst barrier lakes in Mianshan limestone area, northwest SichuanJournal of Lake Sciences, 5, 33 -39 . [本文引用: 1] [ 张捷 (1993 ). 川西北眠山灰岩区岩溶堰塞湖形成中的生物作用湖泊科学 , 5, 33 -39 .] [本文引用: 1] [84] Zhang KY 2017 ). The Dissolution of Limestone by Some of the Mosses and the Microorganism in Karst Area in Guizhou Province Master degree dissertation, Southwestern University, Chongqing . [本文引用: 2] [ 张楷燕 (2017 ). 贵州喀斯特几种石生苔藓和土壤微生物对石灰岩的溶蚀作用硕士学位论文, 西南大学, 重庆 .] [本文引用: 2] [85] Zhang ML Deng ZQ 1994 ). The soil and soil-forming processes in karst area of South ChinaJournal Guizhou Institute of Technology, 23, 67 -75 . [本文引用: 1] [ 张美良 , 邓自强 (1994 ). 我国南方岩溶地区的土壤及其形成贵州工学院学报 , 23, 67 -75 .] [本文引用: 1] [86] Zhang XQ Liu TL Cong CL 2018a ). Study on soil conservation and pedogenic function of five bryophytes in the karst areas of Guizhou ProvinceCarsologica Sinica, 37, 708 -713 . [ 张显强 , 刘天雷 , 从春蕾 (2018 a). 贵州5种喀斯特石生藓类成土及保土生态功能研究中国岩溶 , 37, 708 -713 .] [87] Zhang XQ Long HY Liu TL Cong CL 2018 b). Comparison of water absorption characteristic and water retention capacity of five epilithic mosses in the karst areas of Guizhou ProvinceCarsologica Sinica, 37, 835 -841 . [ 张显强 , 龙华英 , 刘天雷 , 从春蕾 (2018 b). 贵州喀斯特地区5种石生藓类的持水性能及吸水特征比较中国岩溶 , 37, 835 -841 .] [88] Zhang XQ ZhaoYZ , Wang SJ 2017 ). Responses of antioxidant defense system of epilithic mosses to drought stress in karst rock desertified areasActa Geochimica , 36, 205 -212 . [89] Zhang ZH Chen JK 2007 ). Floristic characteristics of aquatic bryophytes and their biokarst deposition types at waterfalls in central Guizhou, ChinaCarsologica Sinica, 26, 171 -177 . [本文引用: 1] [ 张朝晖 , 陈家宽 (2007 ). 黔中瀑布水生苔藓植物区系及其生物喀斯特沉积生态类型研究中国岩溶 , 26, 171 -177 .] [本文引用: 1] [90] Zhang ZH Peng T Li XN Zhao CH 2004 ). Bryophytes in karst cave entrance zone of Kunming, ChinaCarsologica Sinica, 23, 230 -233 . [本文引用: 1] [ 张朝晖 , 彭涛 , 李晓娜 , 赵传海 (2004 ). 中国昆明地区岩溶洞穴洞口带苔藓植物研究中国岩溶 , 23, 230 -233 .] [本文引用: 1] [91] Zhao ZM Luan Y Quan SC Wu JL Liang Y Zeng Z 2018 ). Effect of functional groups for the organic acid on the crystal transition of phosphogypsum crystalJournal of Building Materials, 21, 247 -252 . [本文引用: 1] [ 赵志曼 , 栾扬 , 全思臣 , 吴佳丽 , 梁祎 , 曾众 (2018 ). 含辅助官能团类有机酸对磷石膏晶体的影响建筑材料学报 , 21, 247 -252 .] [本文引用: 1] [92] Zheng YP Zhao JC Zhang BC Li L Zhang YM 2009 ). Research progress of algae and bryophytes in desert biological crustsJournal of Integrative Plant Biology, 44, 371 -378 . [本文引用: 1] [ 郑云普 , 赵建成 , 张丙昌 , 李琳 , 张元明 (2009 ). 荒漠生物结皮中藻类和苔藓植物研究进展植物学报 , 44, 371 -378 .] [本文引用: 1] [93] Zhu XM 1995 ). The process of the original soil formationResearch of Water and Soil Conservation, 4, 83 -89 . [本文引用: 1] [ 朱显谟 (1995 ). 论原始土壤的成土过程水土保持研究 , 4, 83 -89 .] [本文引用: 1] Effect of manganese and calcium ions on the photoinduced water oxidation with photosynthesis organ grana from green plant
1
2010
... 由于苔藓植物属于贫营养型, 它所需要的营养物质是维管束植物的1/10, 但是对N、P、K、Ca、Mg、Fe、硫(S)等元素的需求仍必不可少(
娄红祥, 2012 ).Ca
2+ 作为生物的第二信使, 具有调节生物抗逆性等多种功能, 对苔藓植物的生长有显著影响(
Amao et al ., 2010 ;
Kolupaev et al ., 2014 ).当苔藓植物缺Ca时会导致生长缓慢, 假根分化少, 原丝体不发育, 叶状体形状改变等(
娄红祥, 2012 ).而且CA活性也受Fe
2+ 、锌离子(Zn
2+ )、硝酸根(NO
- 3 )、氯离子(Cl
- )、溴离子(Br
- )、有机碳、交换性钙等的显著影响(
洪子茜, 2018 ).例如,
黄芬等(2015) 研究表明, Ca对CA的活性有很强的促进作用.生物与环境协同进化致使某些苔藓出现钙依赖性, 这种对钙的依赖性是否表现在基因水平, 从而影响岩溶地区苔藓植物生物多样性、空间分布特征、功能性状等还有待进一步研究.因此在研究苔藓植物岩溶作用与pH及营养元素之间的关系时, 应综合考虑苔藓植物种类、酶活性、逆境胁迫、水环境、基岩性质等因素. ...
Combined effects of salinity and temperature on the solubility of organic compounds
1
2012
... 苔藓植物群落分布特征与生境条件密切相关(
Heino & Virtanen, 2006 ;
刘艳等, 2015 ;
Mod et al ., 2016 ;
李泽科等, 2018 ;
庞嘉鹏等, 2018 ), 温度和水分是影响苔藓植物生存和分布的主要决定性因素, 尤其是空气湿度(
娄红祥, 2012 ;
Guo & Zhao, 2018 ).
党成强等(2018) 研究表明, 南亚毛灰藓(
Homomallium simlaense )在干旱和高温条件下, 各项生理指标均与体内相对含水量呈正比.苔藓植物在干旱缺水的情况下会停止生理代谢进入休眠状态, 当水分充足时再进行生命活动.而且苔藓植物生物量与相对生长速率随光照增强而增加, 随光照时间的延长, 其长度、pH值降低能力逐渐提高(
St. Martin & Mallik, 2017 ).苔藓植物虽然能够忍受高温, 但是温度高于25 ℃时却很少有净光合产物(
Proctor, 2000 ;
Hanson et al ., 2014 ).同时, 温度也是影响酶活性的关键因素, 温度超过60 ℃时会严重影响CA的活性(
洪子茜, 2018 ).然而, CA参与苔藓植物岩溶作用的最佳酶促反应温度还不得而知.同时, 温度也影响有机酸的反应性、络合能力和岩石中矿物自身的溶解程度(
闫志为等, 2009 ;
Benbouzid et al ., 2012 ), 进而影响岩石的溶蚀强度(
陈传平等, 2008 ).而且有机酸与矿物反应生成有机酸盐的存在形态也随温度发生变化(
Peters et al ., 2018 ). ...
The effect of pH, grain size, and organic ligands on biotite weathering rates
1
2015
...
Xie和Lou (2009) 认为, 生物岩溶作用随pH值降低而增强, 如果单纯从H
+ + CaCO
3 $\rightleftharpoons $ CO
2 + H
2 O + Ca
2+ 角度分析pH对岩溶作用的影响, pH值越低溶蚀速率越快.然而岩石的最佳溶解pH由矿物自身属性决定(
Bray et al ., 2015 ), 同时有机酸的溶蚀能力也受pH影响(
Pachana et al ., 2012 ;
Chen et al ., 2016 ), 而且每类苔藓植物的最适生长pH也不尽相同.如泥炭藓属(
Sphagnum )植物适宜偏酸性环境, 细叶小羽藓(
Haplocladium microphyllum )配子体的最适生长pH值为7 (
陈文佳等, 2012 ).作为苔藓植物岩溶作用的重要参与者CA, 它的活性也受pH影响,
洪子茜(2018) 研究表明, 当pH在7-9条件下CA的活性最高.pH由外而内对苔藓植物岩溶作用的过程、速率、对象、各级参与者都产生不同程度的促进或抑制作用.目前我们对pH影响苔藓植物岩溶的规律以及调节机制还不得而知, 今后还需综合全面地进行深入研究. ...
碳酸盐岩表面溶蚀形态及其影响因素
2017
碳酸盐岩表面溶蚀形态及其影响因素
2017
The effects of organic acids on the dissolution of silicate minerals: A case study of oxalate catalysis of kaolinite dissolution
1
2006
... 岩石中所含矿物种类及其晶体结构、产状、透水性等决定了岩石的溶解能力(
Voinot et al ., 2013 ;
黄奇波等, 2015 ), 例如, 一般情况下泥质含量高的碳酸盐岩溶蚀速度快, 岩石中酸不溶物含量越低, 越易被溶解(
王世杰等, 1999 ).同时, 岩石中酸不溶物含量对碳酸盐岩的成土速率具有重要影响, 酸不溶物含量越高岩石风化后可残留物越多, 成土物质越丰富(
陈洪松等, 2018 ).例如, 白云岩中酸不溶物含量高, 发育的土层较石灰岩地区土层厚(
陈洪松等, 2013 ).岩石中所含矿物的性质影响各元素的释放速率, Andrew等(2015)在25 ℃、pH = 2.6的开放试验系统与封闭试验系统内发现黑云母中的元素释放按照Fe > Mg > Al > Si的顺序进行, 且溶解速率符合 ${{r}_{i}}={{k}_{\text{H},i}}\times a_{{{\text{H}}^{\text{+}}}}^{xi}$,
k H 为表面速率常数((2.15-30.6) × 10
-10 mole s
biotite ·m
-2 ·s
-1 ),
a H+ 为H
+ 活性,
xi 为矿物与H
+ 的反应级数(0.31-0.58); 分析认为, 岩石中各矿物的溶解具有空间各向异性, 其边缘的反应效率强于基面, 而且各个元素的释放具有一定的规律性.苔藓植物种类的空间分布特征与岩性密切相关(
Gorozhankina & Konstantinov, 2001 ), 而且岩性和有机酸种类也会影响有机酸与岩石的反应速率(
Cama & Ganor, 2006 ;
Ghoorah et al ., 2014 ;
Mwangi et al ., 2018 ), 从而导致它们岩溶作用的差异性. ...
桂林地区生物岩溶微观形态与岩溶地貌宏观形态间的分形特征
1996
桂林地区生物岩溶微观形态与岩溶地貌宏观形态间的分形特征
1996
石生藻类、地衣、苔藓与碳酸盐岩持水性及生态意义
1
1999
... 苔藓植物常以丛集型、交织型、匍匐型等着生于岩面(
图1 ), 假根根系不发达, 只起固着作用(
娄玉霞, 2013 ), 其特殊的叶表面结构和细胞特征(
娄红祥, 2012 ), 使苔藓具有耐高温、抗旱、蓄水能力强、保湿固土等功能(
Proctor & Pence, 2002 ;
Krol et al ., 2003 ;
Zhang et al ., 2017 ).
曹建华和袁道先(1999) 研究发现, 当岩石表面有苔藓植物时, 其蒸发失水和主动吸水时间比裸岩分别延长了48%和57%, 持水量提高了16.6倍, 分析认为苔藓植物可以延缓岩面水分蒸发时间, 增加岩石水化学作用时间, 但是苔藓植物的这种蓄水保湿能力存在明显的种间差异(
申家琛等, 2017 ;
张显强等, 2018a ).苔藓植物参与岩溶作用的另一大优势, 是其叶表面角质层不发育, 且阳离子交换位点多(
吴玉环等, 2001 ), 能够吸附多种阳离子, 这些阳离子在苔藓植物体内被转化为无机盐、有机盐以及其他形式的化合物储存于组织中.苔藓植物除了以生物化学方式对岩石进行溶蚀之外, 还可以通过物理机械作用破坏岩石.例如, 苔藓植物的假根可以伸入岩石裂隙, 对岩石有一定的破坏作用(
Jackson, 2015 ).再者, 当气候干旱时, 苔藓植物在卷缩过程中会拔起岩面的一些小颗粒, 从而加速了岩石被破坏和成土速度(
连宾等, 2008 ).此外, 苔藓植物常与藻类、地衣、微生物等共生形成生物结皮, 它们之间的相互关系以及因气候因素引起的干湿交替和冻融作用对岩石也具有一定的破坏性(
郑云普等, 2009 ).例如, 菌类的菌丝可以伸入岩石晶体结构中破坏岩石, 细菌与真菌形成的配合基能够络合三价铁离子, 细菌分泌的柠檬酸、乙酸以及地衣分泌的地衣酸(
连宾等, 2011 ), 可以溶解岩石中的矿物(
Won-Pyo et al ., 2016 ;
Wang et al ., 2017 ).因此在研究苔藓植物岩溶作用时, 应明确苔藓植物自身及其与其他生物形成的生物结皮两者之间的关系.目前, 有关岩面苔藓植物功能性状的研究主要集中在小范围内特定生境中的苔藓植物研究, 无法代表岩溶生态系统苔藓植物的整体情况.对于大尺度空间上, 因生境异质性高而造成相同物种在形态结构、光合特征及其钙(Ca)、碳(C)、氮(N)、磷(P) 等元素的代谢调节机制等方面的差异性的研究还未引起人们的广泛关注.岩溶生态系统的物质基础相对贫乏, 具有“缺氮少磷钙丰富”的特征.因此, 研究Ca、C、N、P等元素在苔藓植物体内的代谢机制对揭示岩溶生态系统苔藓植物和其他植物的生物多样性维持机制, 及其岩溶作用特征具有重要的科学 意义. ...
石生藻类、地衣、苔藓与碳酸盐岩持水性及生态意义
1
1999
... 苔藓植物常以丛集型、交织型、匍匐型等着生于岩面(
图1 ), 假根根系不发达, 只起固着作用(
娄玉霞, 2013 ), 其特殊的叶表面结构和细胞特征(
娄红祥, 2012 ), 使苔藓具有耐高温、抗旱、蓄水能力强、保湿固土等功能(
Proctor & Pence, 2002 ;
Krol et al ., 2003 ;
Zhang et al ., 2017 ).
曹建华和袁道先(1999) 研究发现, 当岩石表面有苔藓植物时, 其蒸发失水和主动吸水时间比裸岩分别延长了48%和57%, 持水量提高了16.6倍, 分析认为苔藓植物可以延缓岩面水分蒸发时间, 增加岩石水化学作用时间, 但是苔藓植物的这种蓄水保湿能力存在明显的种间差异(
申家琛等, 2017 ;
张显强等, 2018a ).苔藓植物参与岩溶作用的另一大优势, 是其叶表面角质层不发育, 且阳离子交换位点多(
吴玉环等, 2001 ), 能够吸附多种阳离子, 这些阳离子在苔藓植物体内被转化为无机盐、有机盐以及其他形式的化合物储存于组织中.苔藓植物除了以生物化学方式对岩石进行溶蚀之外, 还可以通过物理机械作用破坏岩石.例如, 苔藓植物的假根可以伸入岩石裂隙, 对岩石有一定的破坏作用(
Jackson, 2015 ).再者, 当气候干旱时, 苔藓植物在卷缩过程中会拔起岩面的一些小颗粒, 从而加速了岩石被破坏和成土速度(
连宾等, 2008 ).此外, 苔藓植物常与藻类、地衣、微生物等共生形成生物结皮, 它们之间的相互关系以及因气候因素引起的干湿交替和冻融作用对岩石也具有一定的破坏性(
郑云普等, 2009 ).例如, 菌类的菌丝可以伸入岩石晶体结构中破坏岩石, 细菌与真菌形成的配合基能够络合三价铁离子, 细菌分泌的柠檬酸、乙酸以及地衣分泌的地衣酸(
连宾等, 2011 ), 可以溶解岩石中的矿物(
Won-Pyo et al ., 2016 ;
Wang et al ., 2017 ).因此在研究苔藓植物岩溶作用时, 应明确苔藓植物自身及其与其他生物形成的生物结皮两者之间的关系.目前, 有关岩面苔藓植物功能性状的研究主要集中在小范围内特定生境中的苔藓植物研究, 无法代表岩溶生态系统苔藓植物的整体情况.对于大尺度空间上, 因生境异质性高而造成相同物种在形态结构、光合特征及其钙(Ca)、碳(C)、氮(N)、磷(P) 等元素的代谢调节机制等方面的差异性的研究还未引起人们的广泛关注.岩溶生态系统的物质基础相对贫乏, 具有“缺氮少磷钙丰富”的特征.因此, 研究Ca、C、N、P等元素在苔藓植物体内的代谢机制对揭示岩溶生态系统苔藓植物和其他植物的生物多样性维持机制, 及其岩溶作用特征具有重要的科学 意义. ...
受地质条件制约的中国西南岩溶生态系统
2
2004
... 岩石经苔藓植物溶蚀后, 表面常常出现一些凹凸不平和微溶孔, 这是由于岩石成分不均匀, 导致生物钻孔力的差异性(
王福星等, 1998 ), 以及溶蚀产物流失而形成溶蚀微地貌.
王福星等(1993) 将这些溶蚀微地貌, 按照口面形态、直径大小、深度等, 把短径小于1 mm的命名为微溶孔, 把短径为1-20 mm的命名为溶孔, 把短径为20-100 mm的命名为溶坑.
付兰和张朝晖(2010) 将连续分布的苔藓植物溶蚀微地貌, 按测算结果, 把长7.7-14.4 cm的命名为溶丝; 长0.8-19.0 cm, 宽0.6-7.2 cm的命名为溶斑; 长6.0-24.5 cm, 宽1.2-2.2 cm的命名为溶带.苔藓植物溶蚀过程很难通过宏观手段观测, 导致这两种命名法在大小和形态方面存在明显差异.然而,
Haward等(2011) 借助原子力显微镜(AFM)观察到黑云母表面浅层溶蚀坑的形成过程, 通过分析图像计算质量损失, 从而确定溶蚀过程中黑云母表面的活化能大小.研究表明, 通过分析活化能得到的溶坑大小与测量其周长所得的数据一致.因此, 在研究溶蚀微地貌时, 应采用高精度仪器, 统一测量标准, 分析微地貌是苔藓植物溶蚀作用产生, 还是由于淋溶风化而形成, 有利于后期岩溶微地貌之间的对比分析和进一步深入研究.这些微地貌的形态特征与生物类型和发育生境密切相关(
蔡胡霖, 2017 ), 在温湿度优越的地区发育更为广泛(
Liang & Xu, 2014 ;
Liu et al ., 2016 ).例如, 溶孔、溶锥多发生在低湿度、强光照、高温环境中, 而溶沟多是在高温、强光和pH较高的生境内发育(
申家琛等, 2018a ).
曹建华和王福星(1996) 认为苔藓植物岩溶微地貌与岩溶地区的宏观地貌形态之间存在一定的相关性, 然而目前这种关联性以及苔藓植物微地貌之间的关系和发育方向尚未得到科学揭示. ...
... 岩溶地区土壤的形成和发育是碳酸岩长期溶蚀、风化与生物富集过程的结果, 其一方面受岩石结构、性质影响, 另一方面也与植被作用密切相关(
张美良和邓自强, 1994 ).研究表明, 碳酸盐岩的成土速率与岩石溶蚀速率成正比(
曹建华等, 2004 ), 与酸不溶物含量密切相关(
曹沁智等, 2018 ).苔藓植物殖居于岩面, 假根系以及植物体的干湿交替冻融现象所产生的物理机械力作用于岩石, 产生岩石碎屑.同时, 有机物与矿物离子络合常常形成难溶物, 这些难溶物与苔藓及其生物结皮死亡残体黏结在一起, 不仅增加了土壤沉积量, 而且促进有机质积累, 增加土壤养分(
朱显谟, 1995 ;
刘天雷等, 2017 ).苔藓植物通过影响下垫面土壤湿度, 促进土壤对岩石的岩溶作用, 间接地影响沉积过程.同时, 苔藓植物的吸附拦截功能对防治岩石表面水土流失具有重要作用(
李军峰等, 2013 ;
贾少华等, 2014 ;
张显强等, 2018b ).苔藓植物的溶蚀作用为岩面土壤形成提供了原始物质, 关于苔藓对土壤沉积的贡献率与苔藓植物作用下岩面土壤的形成过程, 以及在此过程中岩石的物质、能量转化和土壤养分的变化等还未见报道. ...
受地质条件制约的中国西南岩溶生态系统
2
2004
... 岩石经苔藓植物溶蚀后, 表面常常出现一些凹凸不平和微溶孔, 这是由于岩石成分不均匀, 导致生物钻孔力的差异性(
王福星等, 1998 ), 以及溶蚀产物流失而形成溶蚀微地貌.
王福星等(1993) 将这些溶蚀微地貌, 按照口面形态、直径大小、深度等, 把短径小于1 mm的命名为微溶孔, 把短径为1-20 mm的命名为溶孔, 把短径为20-100 mm的命名为溶坑.
付兰和张朝晖(2010) 将连续分布的苔藓植物溶蚀微地貌, 按测算结果, 把长7.7-14.4 cm的命名为溶丝; 长0.8-19.0 cm, 宽0.6-7.2 cm的命名为溶斑; 长6.0-24.5 cm, 宽1.2-2.2 cm的命名为溶带.苔藓植物溶蚀过程很难通过宏观手段观测, 导致这两种命名法在大小和形态方面存在明显差异.然而,
Haward等(2011) 借助原子力显微镜(AFM)观察到黑云母表面浅层溶蚀坑的形成过程, 通过分析图像计算质量损失, 从而确定溶蚀过程中黑云母表面的活化能大小.研究表明, 通过分析活化能得到的溶坑大小与测量其周长所得的数据一致.因此, 在研究溶蚀微地貌时, 应采用高精度仪器, 统一测量标准, 分析微地貌是苔藓植物溶蚀作用产生, 还是由于淋溶风化而形成, 有利于后期岩溶微地貌之间的对比分析和进一步深入研究.这些微地貌的形态特征与生物类型和发育生境密切相关(
蔡胡霖, 2017 ), 在温湿度优越的地区发育更为广泛(
Liang & Xu, 2014 ;
Liu et al ., 2016 ).例如, 溶孔、溶锥多发生在低湿度、强光照、高温环境中, 而溶沟多是在高温、强光和pH较高的生境内发育(
申家琛等, 2018a ).
曹建华和王福星(1996) 认为苔藓植物岩溶微地貌与岩溶地区的宏观地貌形态之间存在一定的相关性, 然而目前这种关联性以及苔藓植物微地貌之间的关系和发育方向尚未得到科学揭示. ...
... 岩溶地区土壤的形成和发育是碳酸岩长期溶蚀、风化与生物富集过程的结果, 其一方面受岩石结构、性质影响, 另一方面也与植被作用密切相关(
张美良和邓自强, 1994 ).研究表明, 碳酸盐岩的成土速率与岩石溶蚀速率成正比(
曹建华等, 2004 ), 与酸不溶物含量密切相关(
曹沁智等, 2018 ).苔藓植物殖居于岩面, 假根系以及植物体的干湿交替冻融现象所产生的物理机械力作用于岩石, 产生岩石碎屑.同时, 有机物与矿物离子络合常常形成难溶物, 这些难溶物与苔藓及其生物结皮死亡残体黏结在一起, 不仅增加了土壤沉积量, 而且促进有机质积累, 增加土壤养分(
朱显谟, 1995 ;
刘天雷等, 2017 ).苔藓植物通过影响下垫面土壤湿度, 促进土壤对岩石的岩溶作用, 间接地影响沉积过程.同时, 苔藓植物的吸附拦截功能对防治岩石表面水土流失具有重要作用(
李军峰等, 2013 ;
贾少华等, 2014 ;
张显强等, 2018b ).苔藓植物的溶蚀作用为岩面土壤形成提供了原始物质, 关于苔藓对土壤沉积的贡献率与苔藓植物作用下岩面土壤的形成过程, 以及在此过程中岩石的物质、能量转化和土壤养分的变化等还未见报道. ...
白云质岩石中酸不溶物对碱白云石反应的影响
1
2018
... 岩溶地区土壤的形成和发育是碳酸岩长期溶蚀、风化与生物富集过程的结果, 其一方面受岩石结构、性质影响, 另一方面也与植被作用密切相关(
张美良和邓自强, 1994 ).研究表明, 碳酸盐岩的成土速率与岩石溶蚀速率成正比(
曹建华等, 2004 ), 与酸不溶物含量密切相关(
曹沁智等, 2018 ).苔藓植物殖居于岩面, 假根系以及植物体的干湿交替冻融现象所产生的物理机械力作用于岩石, 产生岩石碎屑.同时, 有机物与矿物离子络合常常形成难溶物, 这些难溶物与苔藓及其生物结皮死亡残体黏结在一起, 不仅增加了土壤沉积量, 而且促进有机质积累, 增加土壤养分(
朱显谟, 1995 ;
刘天雷等, 2017 ).苔藓植物通过影响下垫面土壤湿度, 促进土壤对岩石的岩溶作用, 间接地影响沉积过程.同时, 苔藓植物的吸附拦截功能对防治岩石表面水土流失具有重要作用(
李军峰等, 2013 ;
贾少华等, 2014 ;
张显强等, 2018b ).苔藓植物的溶蚀作用为岩面土壤形成提供了原始物质, 关于苔藓对土壤沉积的贡献率与苔藓植物作用下岩面土壤的形成过程, 以及在此过程中岩石的物质、能量转化和土壤养分的变化等还未见报道. ...
白云质岩石中酸不溶物对碱白云石反应的影响
1
2018
... 岩溶地区土壤的形成和发育是碳酸岩长期溶蚀、风化与生物富集过程的结果, 其一方面受岩石结构、性质影响, 另一方面也与植被作用密切相关(
张美良和邓自强, 1994 ).研究表明, 碳酸盐岩的成土速率与岩石溶蚀速率成正比(
曹建华等, 2004 ), 与酸不溶物含量密切相关(
曹沁智等, 2018 ).苔藓植物殖居于岩面, 假根系以及植物体的干湿交替冻融现象所产生的物理机械力作用于岩石, 产生岩石碎屑.同时, 有机物与矿物离子络合常常形成难溶物, 这些难溶物与苔藓及其生物结皮死亡残体黏结在一起, 不仅增加了土壤沉积量, 而且促进有机质积累, 增加土壤养分(
朱显谟, 1995 ;
刘天雷等, 2017 ).苔藓植物通过影响下垫面土壤湿度, 促进土壤对岩石的岩溶作用, 间接地影响沉积过程.同时, 苔藓植物的吸附拦截功能对防治岩石表面水土流失具有重要作用(
李军峰等, 2013 ;
贾少华等, 2014 ;
张显强等, 2018b ).苔藓植物的溶蚀作用为岩面土壤形成提供了原始物质, 关于苔藓对土壤沉积的贡献率与苔藓植物作用下岩面土壤的形成过程, 以及在此过程中岩石的物质、能量转化和土壤养分的变化等还未见报道. ...
不同有机酸对矿物溶解的动力学实验研究
1
2008
... 苔藓植物群落分布特征与生境条件密切相关(
Heino & Virtanen, 2006 ;
刘艳等, 2015 ;
Mod et al ., 2016 ;
李泽科等, 2018 ;
庞嘉鹏等, 2018 ), 温度和水分是影响苔藓植物生存和分布的主要决定性因素, 尤其是空气湿度(
娄红祥, 2012 ;
Guo & Zhao, 2018 ).
党成强等(2018) 研究表明, 南亚毛灰藓(
Homomallium simlaense )在干旱和高温条件下, 各项生理指标均与体内相对含水量呈正比.苔藓植物在干旱缺水的情况下会停止生理代谢进入休眠状态, 当水分充足时再进行生命活动.而且苔藓植物生物量与相对生长速率随光照增强而增加, 随光照时间的延长, 其长度、pH值降低能力逐渐提高(
St. Martin & Mallik, 2017 ).苔藓植物虽然能够忍受高温, 但是温度高于25 ℃时却很少有净光合产物(
Proctor, 2000 ;
Hanson et al ., 2014 ).同时, 温度也是影响酶活性的关键因素, 温度超过60 ℃时会严重影响CA的活性(
洪子茜, 2018 ).然而, CA参与苔藓植物岩溶作用的最佳酶促反应温度还不得而知.同时, 温度也影响有机酸的反应性、络合能力和岩石中矿物自身的溶解程度(
闫志为等, 2009 ;
Benbouzid et al ., 2012 ), 进而影响岩石的溶蚀强度(
陈传平等, 2008 ).而且有机酸与矿物反应生成有机酸盐的存在形态也随温度发生变化(
Peters et al ., 2018 ). ...
不同有机酸对矿物溶解的动力学实验研究
1
2008
... 苔藓植物群落分布特征与生境条件密切相关(
Heino & Virtanen, 2006 ;
刘艳等, 2015 ;
Mod et al ., 2016 ;
李泽科等, 2018 ;
庞嘉鹏等, 2018 ), 温度和水分是影响苔藓植物生存和分布的主要决定性因素, 尤其是空气湿度(
娄红祥, 2012 ;
Guo & Zhao, 2018 ).
党成强等(2018) 研究表明, 南亚毛灰藓(
Homomallium simlaense )在干旱和高温条件下, 各项生理指标均与体内相对含水量呈正比.苔藓植物在干旱缺水的情况下会停止生理代谢进入休眠状态, 当水分充足时再进行生命活动.而且苔藓植物生物量与相对生长速率随光照增强而增加, 随光照时间的延长, 其长度、pH值降低能力逐渐提高(
St. Martin & Mallik, 2017 ).苔藓植物虽然能够忍受高温, 但是温度高于25 ℃时却很少有净光合产物(
Proctor, 2000 ;
Hanson et al ., 2014 ).同时, 温度也是影响酶活性的关键因素, 温度超过60 ℃时会严重影响CA的活性(
洪子茜, 2018 ).然而, CA参与苔藓植物岩溶作用的最佳酶促反应温度还不得而知.同时, 温度也影响有机酸的反应性、络合能力和岩石中矿物自身的溶解程度(
闫志为等, 2009 ;
Benbouzid et al ., 2012 ), 进而影响岩石的溶蚀强度(
陈传平等, 2008 ).而且有机酸与矿物反应生成有机酸盐的存在形态也随温度发生变化(
Peters et al ., 2018 ). ...
西南喀斯特地区水土过程与植被恢复重建
1
2018
... 岩石中所含矿物种类及其晶体结构、产状、透水性等决定了岩石的溶解能力(
Voinot et al ., 2013 ;
黄奇波等, 2015 ), 例如, 一般情况下泥质含量高的碳酸盐岩溶蚀速度快, 岩石中酸不溶物含量越低, 越易被溶解(
王世杰等, 1999 ).同时, 岩石中酸不溶物含量对碳酸盐岩的成土速率具有重要影响, 酸不溶物含量越高岩石风化后可残留物越多, 成土物质越丰富(
陈洪松等, 2018 ).例如, 白云岩中酸不溶物含量高, 发育的土层较石灰岩地区土层厚(
陈洪松等, 2013 ).岩石中所含矿物的性质影响各元素的释放速率, Andrew等(2015)在25 ℃、pH = 2.6的开放试验系统与封闭试验系统内发现黑云母中的元素释放按照Fe > Mg > Al > Si的顺序进行, 且溶解速率符合 ${{r}_{i}}={{k}_{\text{H},i}}\times a_{{{\text{H}}^{\text{+}}}}^{xi}$,
k H 为表面速率常数((2.15-30.6) × 10
-10 mole s
biotite ·m
-2 ·s
-1 ),
a H+ 为H
+ 活性,
xi 为矿物与H
+ 的反应级数(0.31-0.58); 分析认为, 岩石中各矿物的溶解具有空间各向异性, 其边缘的反应效率强于基面, 而且各个元素的释放具有一定的规律性.苔藓植物种类的空间分布特征与岩性密切相关(
Gorozhankina & Konstantinov, 2001 ), 而且岩性和有机酸种类也会影响有机酸与岩石的反应速率(
Cama & Ganor, 2006 ;
Ghoorah et al ., 2014 ;
Mwangi et al ., 2018 ), 从而导致它们岩溶作用的差异性. ...
西南喀斯特地区水土过程与植被恢复重建
1
2018
... 岩石中所含矿物种类及其晶体结构、产状、透水性等决定了岩石的溶解能力(
Voinot et al ., 2013 ;
黄奇波等, 2015 ), 例如, 一般情况下泥质含量高的碳酸盐岩溶蚀速度快, 岩石中酸不溶物含量越低, 越易被溶解(
王世杰等, 1999 ).同时, 岩石中酸不溶物含量对碳酸盐岩的成土速率具有重要影响, 酸不溶物含量越高岩石风化后可残留物越多, 成土物质越丰富(
陈洪松等, 2018 ).例如, 白云岩中酸不溶物含量高, 发育的土层较石灰岩地区土层厚(
陈洪松等, 2013 ).岩石中所含矿物的性质影响各元素的释放速率, Andrew等(2015)在25 ℃、pH = 2.6的开放试验系统与封闭试验系统内发现黑云母中的元素释放按照Fe > Mg > Al > Si的顺序进行, 且溶解速率符合 ${{r}_{i}}={{k}_{\text{H},i}}\times a_{{{\text{H}}^{\text{+}}}}^{xi}$,
k H 为表面速率常数((2.15-30.6) × 10
-10 mole s
biotite ·m
-2 ·s
-1 ),
a H+ 为H
+ 活性,
xi 为矿物与H
+ 的反应级数(0.31-0.58); 分析认为, 岩石中各矿物的溶解具有空间各向异性, 其边缘的反应效率强于基面, 而且各个元素的释放具有一定的规律性.苔藓植物种类的空间分布特征与岩性密切相关(
Gorozhankina & Konstantinov, 2001 ), 而且岩性和有机酸种类也会影响有机酸与岩石的反应速率(
Cama & Ganor, 2006 ;
Ghoorah et al ., 2014 ;
Mwangi et al ., 2018 ), 从而导致它们岩溶作用的差异性. ...
岩溶山区水分时空异质性及植物适应机理研究进展
1
2013
... 岩石中所含矿物种类及其晶体结构、产状、透水性等决定了岩石的溶解能力(
Voinot et al ., 2013 ;
黄奇波等, 2015 ), 例如, 一般情况下泥质含量高的碳酸盐岩溶蚀速度快, 岩石中酸不溶物含量越低, 越易被溶解(
王世杰等, 1999 ).同时, 岩石中酸不溶物含量对碳酸盐岩的成土速率具有重要影响, 酸不溶物含量越高岩石风化后可残留物越多, 成土物质越丰富(
陈洪松等, 2018 ).例如, 白云岩中酸不溶物含量高, 发育的土层较石灰岩地区土层厚(
陈洪松等, 2013 ).岩石中所含矿物的性质影响各元素的释放速率, Andrew等(2015)在25 ℃、pH = 2.6的开放试验系统与封闭试验系统内发现黑云母中的元素释放按照Fe > Mg > Al > Si的顺序进行, 且溶解速率符合 ${{r}_{i}}={{k}_{\text{H},i}}\times a_{{{\text{H}}^{\text{+}}}}^{xi}$,
k H 为表面速率常数((2.15-30.6) × 10
-10 mole s
biotite ·m
-2 ·s
-1 ),
a H+ 为H
+ 活性,
xi 为矿物与H
+ 的反应级数(0.31-0.58); 分析认为, 岩石中各矿物的溶解具有空间各向异性, 其边缘的反应效率强于基面, 而且各个元素的释放具有一定的规律性.苔藓植物种类的空间分布特征与岩性密切相关(
Gorozhankina & Konstantinov, 2001 ), 而且岩性和有机酸种类也会影响有机酸与岩石的反应速率(
Cama & Ganor, 2006 ;
Ghoorah et al ., 2014 ;
Mwangi et al ., 2018 ), 从而导致它们岩溶作用的差异性. ...
岩溶山区水分时空异质性及植物适应机理研究进展
1
2013
... 岩石中所含矿物种类及其晶体结构、产状、透水性等决定了岩石的溶解能力(
Voinot et al ., 2013 ;
黄奇波等, 2015 ), 例如, 一般情况下泥质含量高的碳酸盐岩溶蚀速度快, 岩石中酸不溶物含量越低, 越易被溶解(
王世杰等, 1999 ).同时, 岩石中酸不溶物含量对碳酸盐岩的成土速率具有重要影响, 酸不溶物含量越高岩石风化后可残留物越多, 成土物质越丰富(
陈洪松等, 2018 ).例如, 白云岩中酸不溶物含量高, 发育的土层较石灰岩地区土层厚(
陈洪松等, 2013 ).岩石中所含矿物的性质影响各元素的释放速率, Andrew等(2015)在25 ℃、pH = 2.6的开放试验系统与封闭试验系统内发现黑云母中的元素释放按照Fe > Mg > Al > Si的顺序进行, 且溶解速率符合 ${{r}_{i}}={{k}_{\text{H},i}}\times a_{{{\text{H}}^{\text{+}}}}^{xi}$,
k H 为表面速率常数((2.15-30.6) × 10
-10 mole s
biotite ·m
-2 ·s
-1 ),
a H+ 为H
+ 活性,
xi 为矿物与H
+ 的反应级数(0.31-0.58); 分析认为, 岩石中各矿物的溶解具有空间各向异性, 其边缘的反应效率强于基面, 而且各个元素的释放具有一定的规律性.苔藓植物种类的空间分布特征与岩性密切相关(
Gorozhankina & Konstantinov, 2001 ), 而且岩性和有机酸种类也会影响有机酸与岩石的反应速率(
Cama & Ganor, 2006 ;
Ghoorah et al ., 2014 ;
Mwangi et al ., 2018 ), 从而导致它们岩溶作用的差异性. ...
不同培养基及pH值对细叶小羽藓配子体生长的影响
1
2012
...
Xie和Lou (2009) 认为, 生物岩溶作用随pH值降低而增强, 如果单纯从H
+ + CaCO
3 $\rightleftharpoons $ CO
2 + H
2 O + Ca
2+ 角度分析pH对岩溶作用的影响, pH值越低溶蚀速率越快.然而岩石的最佳溶解pH由矿物自身属性决定(
Bray et al ., 2015 ), 同时有机酸的溶蚀能力也受pH影响(
Pachana et al ., 2012 ;
Chen et al ., 2016 ), 而且每类苔藓植物的最适生长pH也不尽相同.如泥炭藓属(
Sphagnum )植物适宜偏酸性环境, 细叶小羽藓(
Haplocladium microphyllum )配子体的最适生长pH值为7 (
陈文佳等, 2012 ).作为苔藓植物岩溶作用的重要参与者CA, 它的活性也受pH影响,
洪子茜(2018) 研究表明, 当pH在7-9条件下CA的活性最高.pH由外而内对苔藓植物岩溶作用的过程、速率、对象、各级参与者都产生不同程度的促进或抑制作用.目前我们对pH影响苔藓植物岩溶的规律以及调节机制还不得而知, 今后还需综合全面地进行深入研究. ...
不同培养基及pH值对细叶小羽藓配子体生长的影响
1
2012
...
Xie和Lou (2009) 认为, 生物岩溶作用随pH值降低而增强, 如果单纯从H
+ + CaCO
3 $\rightleftharpoons $ CO
2 + H
2 O + Ca
2+ 角度分析pH对岩溶作用的影响, pH值越低溶蚀速率越快.然而岩石的最佳溶解pH由矿物自身属性决定(
Bray et al ., 2015 ), 同时有机酸的溶蚀能力也受pH影响(
Pachana et al ., 2012 ;
Chen et al ., 2016 ), 而且每类苔藓植物的最适生长pH也不尽相同.如泥炭藓属(
Sphagnum )植物适宜偏酸性环境, 细叶小羽藓(
Haplocladium microphyllum )配子体的最适生长pH值为7 (
陈文佳等, 2012 ).作为苔藓植物岩溶作用的重要参与者CA, 它的活性也受pH影响,
洪子茜(2018) 研究表明, 当pH在7-9条件下CA的活性最高.pH由外而内对苔藓植物岩溶作用的过程、速率、对象、各级参与者都产生不同程度的促进或抑制作用.目前我们对pH影响苔藓植物岩溶的规律以及调节机制还不得而知, 今后还需综合全面地进行深入研究. ...
Effects of organic acids on dissolution of Fe and Mn from weathering coal gangue
1
2016
...
Xie和Lou (2009) 认为, 生物岩溶作用随pH值降低而增强, 如果单纯从H
+ + CaCO
3 $\rightleftharpoons $ CO
2 + H
2 O + Ca
2+ 角度分析pH对岩溶作用的影响, pH值越低溶蚀速率越快.然而岩石的最佳溶解pH由矿物自身属性决定(
Bray et al ., 2015 ), 同时有机酸的溶蚀能力也受pH影响(
Pachana et al ., 2012 ;
Chen et al ., 2016 ), 而且每类苔藓植物的最适生长pH也不尽相同.如泥炭藓属(
Sphagnum )植物适宜偏酸性环境, 细叶小羽藓(
Haplocladium microphyllum )配子体的最适生长pH值为7 (
陈文佳等, 2012 ).作为苔藓植物岩溶作用的重要参与者CA, 它的活性也受pH影响,
洪子茜(2018) 研究表明, 当pH在7-9条件下CA的活性最高.pH由外而内对苔藓植物岩溶作用的过程、速率、对象、各级参与者都产生不同程度的促进或抑制作用.目前我们对pH影响苔藓植物岩溶的规律以及调节机制还不得而知, 今后还需综合全面地进行深入研究. ...
Diversity of secondary metabolites in the liverwort Syzygiella rubricaulis(Nees) Stephani (Jamesoniellaceae, Marchantiophyta) from neotropical high mountains
1
2018
... 生物岩溶是指由生物引起的岩溶作用现象及 其产物, 即生物溶蚀、破坏可溶岩和沉淀形成次生化学沉积物及其所遗留或形成的形态(
Schneider, 1976 ).生物岩溶作用(溶蚀作用和沉积作用)激活了岩溶生态系统基质中的物质和能量, 使各个圈层间信息交流更加频繁与复杂.苔藓植物作为生态系统演替过程中的先锋植物(
薛建辉, 2013 ), 通过生物岩溶作用改善裸岩生境, 为草本和灌木的出现奠定了物质基础(
Harper & Belnap, 2001 ;
Jackson, 2015 ), 对岩溶生态系统的维持与发展具有重要意义(
Li et al ., 2006 ;
Costa et al ., 2018 ).苔藓植物岩溶作用已成为岩溶生态学研究的热点.因此, 了解和掌握岩面苔藓植物功能性状特征、岩溶作用机理及其与环境间的互作机制对研究岩溶地区的生物多样性、石漠化生态恢复、岩溶碳汇以及丰富生物岩溶理论等意义重大.然而, 苔藓植物岩溶作用研究因涉及学科众多、问题复杂, 还未进入系统化研究阶段.为了破除各学科间的壁垒, 促进苔藓植物岩溶作用研究更进一步发展, 本文根据近年来有关苔藓植物岩溶作用的文献资料, 从以下方面综述苔藓植物岩溶作用的研究进展. ...
石生南亚毛灰藓在不同温度和干旱条件下的生理生化特性
1
2018
... 苔藓植物群落分布特征与生境条件密切相关(
Heino & Virtanen, 2006 ;
刘艳等, 2015 ;
Mod et al ., 2016 ;
李泽科等, 2018 ;
庞嘉鹏等, 2018 ), 温度和水分是影响苔藓植物生存和分布的主要决定性因素, 尤其是空气湿度(
娄红祥, 2012 ;
Guo & Zhao, 2018 ).
党成强等(2018) 研究表明, 南亚毛灰藓(
Homomallium simlaense )在干旱和高温条件下, 各项生理指标均与体内相对含水量呈正比.苔藓植物在干旱缺水的情况下会停止生理代谢进入休眠状态, 当水分充足时再进行生命活动.而且苔藓植物生物量与相对生长速率随光照增强而增加, 随光照时间的延长, 其长度、pH值降低能力逐渐提高(
St. Martin & Mallik, 2017 ).苔藓植物虽然能够忍受高温, 但是温度高于25 ℃时却很少有净光合产物(
Proctor, 2000 ;
Hanson et al ., 2014 ).同时, 温度也是影响酶活性的关键因素, 温度超过60 ℃时会严重影响CA的活性(
洪子茜, 2018 ).然而, CA参与苔藓植物岩溶作用的最佳酶促反应温度还不得而知.同时, 温度也影响有机酸的反应性、络合能力和岩石中矿物自身的溶解程度(
闫志为等, 2009 ;
Benbouzid et al ., 2012 ), 进而影响岩石的溶蚀强度(
陈传平等, 2008 ).而且有机酸与矿物反应生成有机酸盐的存在形态也随温度发生变化(
Peters et al ., 2018 ). ...
石生南亚毛灰藓在不同温度和干旱条件下的生理生化特性
1
2018
... 苔藓植物群落分布特征与生境条件密切相关(
Heino & Virtanen, 2006 ;
刘艳等, 2015 ;
Mod et al ., 2016 ;
李泽科等, 2018 ;
庞嘉鹏等, 2018 ), 温度和水分是影响苔藓植物生存和分布的主要决定性因素, 尤其是空气湿度(
娄红祥, 2012 ;
Guo & Zhao, 2018 ).
党成强等(2018) 研究表明, 南亚毛灰藓(
Homomallium simlaense )在干旱和高温条件下, 各项生理指标均与体内相对含水量呈正比.苔藓植物在干旱缺水的情况下会停止生理代谢进入休眠状态, 当水分充足时再进行生命活动.而且苔藓植物生物量与相对生长速率随光照增强而增加, 随光照时间的延长, 其长度、pH值降低能力逐渐提高(
St. Martin & Mallik, 2017 ).苔藓植物虽然能够忍受高温, 但是温度高于25 ℃时却很少有净光合产物(
Proctor, 2000 ;
Hanson et al ., 2014 ).同时, 温度也是影响酶活性的关键因素, 温度超过60 ℃时会严重影响CA的活性(
洪子茜, 2018 ).然而, CA参与苔藓植物岩溶作用的最佳酶促反应温度还不得而知.同时, 温度也影响有机酸的反应性、络合能力和岩石中矿物自身的溶解程度(
闫志为等, 2009 ;
Benbouzid et al ., 2012 ), 进而影响岩石的溶蚀强度(
陈传平等, 2008 ).而且有机酸与矿物反应生成有机酸盐的存在形态也随温度发生变化(
Peters et al ., 2018 ). ...
The Burren: A glacial, karstic and biokarstic expression of a limestone plateau in western Ireland
1
2016
... 因此, 湿度、温度、光照及其综合因子通过对苔藓植物生理代谢过程、基因型以及其他功能性状的影响(
Pharo & Zartman, 2006 ;
de la Rosa, 2016 ;
解梦, 2018 ), 而直接或间接地影响苔藓植物岩溶作用过程与效率.例如, 当温度在17.5-21.4 ℃, 湿度57.7%-72.6%, 光照2 700-5 900 lx条件下, 苔藓植物对碳酸盐岩的塑造作用最强(
申家琛等, 2018a ).但是目前对温湿度及其综合因子对苔藓植物岩溶作用的影响的关注度仍不够, 很多规律仍不清楚, 然而这些规律的发现是揭示苔藓植物岩溶机理的关键, 也是指导岩溶生态问题治理的重要科学依据. ...
贵阳市苔藓植物的生物岩溶溶蚀初探
1
2010
... 岩石经苔藓植物溶蚀后, 表面常常出现一些凹凸不平和微溶孔, 这是由于岩石成分不均匀, 导致生物钻孔力的差异性(
王福星等, 1998 ), 以及溶蚀产物流失而形成溶蚀微地貌.
王福星等(1993) 将这些溶蚀微地貌, 按照口面形态、直径大小、深度等, 把短径小于1 mm的命名为微溶孔, 把短径为1-20 mm的命名为溶孔, 把短径为20-100 mm的命名为溶坑.
付兰和张朝晖(2010) 将连续分布的苔藓植物溶蚀微地貌, 按测算结果, 把长7.7-14.4 cm的命名为溶丝; 长0.8-19.0 cm, 宽0.6-7.2 cm的命名为溶斑; 长6.0-24.5 cm, 宽1.2-2.2 cm的命名为溶带.苔藓植物溶蚀过程很难通过宏观手段观测, 导致这两种命名法在大小和形态方面存在明显差异.然而,
Haward等(2011) 借助原子力显微镜(AFM)观察到黑云母表面浅层溶蚀坑的形成过程, 通过分析图像计算质量损失, 从而确定溶蚀过程中黑云母表面的活化能大小.研究表明, 通过分析活化能得到的溶坑大小与测量其周长所得的数据一致.因此, 在研究溶蚀微地貌时, 应采用高精度仪器, 统一测量标准, 分析微地貌是苔藓植物溶蚀作用产生, 还是由于淋溶风化而形成, 有利于后期岩溶微地貌之间的对比分析和进一步深入研究.这些微地貌的形态特征与生物类型和发育生境密切相关(
蔡胡霖, 2017 ), 在温湿度优越的地区发育更为广泛(
Liang & Xu, 2014 ;
Liu et al ., 2016 ).例如, 溶孔、溶锥多发生在低湿度、强光照、高温环境中, 而溶沟多是在高温、强光和pH较高的生境内发育(
申家琛等, 2018a ).
曹建华和王福星(1996) 认为苔藓植物岩溶微地貌与岩溶地区的宏观地貌形态之间存在一定的相关性, 然而目前这种关联性以及苔藓植物微地貌之间的关系和发育方向尚未得到科学揭示. ...
贵阳市苔藓植物的生物岩溶溶蚀初探
1
2010
... 岩石经苔藓植物溶蚀后, 表面常常出现一些凹凸不平和微溶孔, 这是由于岩石成分不均匀, 导致生物钻孔力的差异性(
王福星等, 1998 ), 以及溶蚀产物流失而形成溶蚀微地貌.
王福星等(1993) 将这些溶蚀微地貌, 按照口面形态、直径大小、深度等, 把短径小于1 mm的命名为微溶孔, 把短径为1-20 mm的命名为溶孔, 把短径为20-100 mm的命名为溶坑.
付兰和张朝晖(2010) 将连续分布的苔藓植物溶蚀微地貌, 按测算结果, 把长7.7-14.4 cm的命名为溶丝; 长0.8-19.0 cm, 宽0.6-7.2 cm的命名为溶斑; 长6.0-24.5 cm, 宽1.2-2.2 cm的命名为溶带.苔藓植物溶蚀过程很难通过宏观手段观测, 导致这两种命名法在大小和形态方面存在明显差异.然而,
Haward等(2011) 借助原子力显微镜(AFM)观察到黑云母表面浅层溶蚀坑的形成过程, 通过分析图像计算质量损失, 从而确定溶蚀过程中黑云母表面的活化能大小.研究表明, 通过分析活化能得到的溶坑大小与测量其周长所得的数据一致.因此, 在研究溶蚀微地貌时, 应采用高精度仪器, 统一测量标准, 分析微地貌是苔藓植物溶蚀作用产生, 还是由于淋溶风化而形成, 有利于后期岩溶微地貌之间的对比分析和进一步深入研究.这些微地貌的形态特征与生物类型和发育生境密切相关(
蔡胡霖, 2017 ), 在温湿度优越的地区发育更为广泛(
Liang & Xu, 2014 ;
Liu et al ., 2016 ).例如, 溶孔、溶锥多发生在低湿度、强光照、高温环境中, 而溶沟多是在高温、强光和pH较高的生境内发育(
申家琛等, 2018a ).
曹建华和王福星(1996) 认为苔藓植物岩溶微地貌与岩溶地区的宏观地貌形态之间存在一定的相关性, 然而目前这种关联性以及苔藓植物微地貌之间的关系和发育方向尚未得到科学揭示. ...
Selection of acid for weak acid processing of wollastonite for mineralisation of CO
2 1
2014
... 岩石中所含矿物种类及其晶体结构、产状、透水性等决定了岩石的溶解能力(
Voinot et al ., 2013 ;
黄奇波等, 2015 ), 例如, 一般情况下泥质含量高的碳酸盐岩溶蚀速度快, 岩石中酸不溶物含量越低, 越易被溶解(
王世杰等, 1999 ).同时, 岩石中酸不溶物含量对碳酸盐岩的成土速率具有重要影响, 酸不溶物含量越高岩石风化后可残留物越多, 成土物质越丰富(
陈洪松等, 2018 ).例如, 白云岩中酸不溶物含量高, 发育的土层较石灰岩地区土层厚(
陈洪松等, 2013 ).岩石中所含矿物的性质影响各元素的释放速率, Andrew等(2015)在25 ℃、pH = 2.6的开放试验系统与封闭试验系统内发现黑云母中的元素释放按照Fe > Mg > Al > Si的顺序进行, 且溶解速率符合 ${{r}_{i}}={{k}_{\text{H},i}}\times a_{{{\text{H}}^{\text{+}}}}^{xi}$,
k H 为表面速率常数((2.15-30.6) × 10
-10 mole s
biotite ·m
-2 ·s
-1 ),
a H+ 为H
+ 活性,
xi 为矿物与H
+ 的反应级数(0.31-0.58); 分析认为, 岩石中各矿物的溶解具有空间各向异性, 其边缘的反应效率强于基面, 而且各个元素的释放具有一定的规律性.苔藓植物种类的空间分布特征与岩性密切相关(
Gorozhankina & Konstantinov, 2001 ), 而且岩性和有机酸种类也会影响有机酸与岩石的反应速率(
Cama & Ganor, 2006 ;
Ghoorah et al ., 2014 ;
Mwangi et al ., 2018 ), 从而导致它们岩溶作用的差异性. ...
Comparative ecocenotic characteristics of mosses in the taiga zone of West Siberia
1
2001
... 岩石中所含矿物种类及其晶体结构、产状、透水性等决定了岩石的溶解能力(
Voinot et al ., 2013 ;
黄奇波等, 2015 ), 例如, 一般情况下泥质含量高的碳酸盐岩溶蚀速度快, 岩石中酸不溶物含量越低, 越易被溶解(
王世杰等, 1999 ).同时, 岩石中酸不溶物含量对碳酸盐岩的成土速率具有重要影响, 酸不溶物含量越高岩石风化后可残留物越多, 成土物质越丰富(
陈洪松等, 2018 ).例如, 白云岩中酸不溶物含量高, 发育的土层较石灰岩地区土层厚(
陈洪松等, 2013 ).岩石中所含矿物的性质影响各元素的释放速率, Andrew等(2015)在25 ℃、pH = 2.6的开放试验系统与封闭试验系统内发现黑云母中的元素释放按照Fe > Mg > Al > Si的顺序进行, 且溶解速率符合 ${{r}_{i}}={{k}_{\text{H},i}}\times a_{{{\text{H}}^{\text{+}}}}^{xi}$,
k H 为表面速率常数((2.15-30.6) × 10
-10 mole s
biotite ·m
-2 ·s
-1 ),
a H+ 为H
+ 活性,
xi 为矿物与H
+ 的反应级数(0.31-0.58); 分析认为, 岩石中各矿物的溶解具有空间各向异性, 其边缘的反应效率强于基面, 而且各个元素的释放具有一定的规律性.苔藓植物种类的空间分布特征与岩性密切相关(
Gorozhankina & Konstantinov, 2001 ), 而且岩性和有机酸种类也会影响有机酸与岩石的反应速率(
Cama & Ganor, 2006 ;
Ghoorah et al ., 2014 ;
Mwangi et al ., 2018 ), 从而导致它们岩溶作用的差异性. ...
白云岩洞穴洞口弱光带的苔藓群落特征——以绥阳水洞为例
1
2018
... 苔藓植物沉积作用的另一表现是石灰华或钙华(
张捷, 1993 ;
Kleinteich et al ., 2017 ).钙华形成的物理化学过程实质是CaCO
3 的沉淀与CO
2 的脱气.苔藓植物在钙华形成过程中为CaCO
3 晶体提供了稳定的生长发育基础, 并且苔藓植物的形态结构控制着沉积物的形态变化(
Mili?a et al ., 2006 ), 且它们的发育形态随沉积环境而变化(
张朝晖等, 2004 ;
郭云等, 2018 ).如在洞穴弱光带一般形成洞穴石刺、向光钟乳石、洞口叠层石等, 这类形态有很强的方向性(
王福星等, 1998 ).苔藓植物在水生环境中也常常形成多样的沉积形态, 如凤尾藓(
Fissidens )钙华、石灰藓(
Hydrogonium )钙华形成的溪流瀑布沉积, 泽藓(
Philonotis )钙华、水灰藓(
Hygrohypnum )钙华形成的河流瀑布沉积, 地钱-真藓(
Marchantia-Bryum )钙华等形成的石灰华坝沉积, 以及净口藓(
Gymnostomum )钙华等形成的瀑布洞穴沉积(
张朝晖和陈家宽, 2007 ).同时, 也有研究发现, 水流环境也是影响苔藓钙华沉积的重要因素, 苔藓与钙华层之间的水流速度与钙华沉积速率呈正相关关系(
Wang et al ., 2014 ).钙华作为大陆古气候的潜在档案,
Yan等(2017) 利用碳氧稳定同位素比率(
δ 13 C和
δ 18 O), 研究钙华在沉积过程中碳、氧的循环规律, 分析认为, 钙华沉积与季节更替引起的气温、降水等的变化密切相关.目前对于苔藓植物沉积作用的研究主要集中在沉积形态以及成土量方面, 对于苔藓植物沉积速率, 对岩溶地区成土过程中的贡献, 以及岩溶沉积作用对生境的改造和环境对岩溶沉积作用的影响等方面的研究还未见报道. ...
白云岩洞穴洞口弱光带的苔藓群落特征——以绥阳水洞为例
1
2018
... 苔藓植物沉积作用的另一表现是石灰华或钙华(
张捷, 1993 ;
Kleinteich et al ., 2017 ).钙华形成的物理化学过程实质是CaCO
3 的沉淀与CO
2 的脱气.苔藓植物在钙华形成过程中为CaCO
3 晶体提供了稳定的生长发育基础, 并且苔藓植物的形态结构控制着沉积物的形态变化(
Mili?a et al ., 2006 ), 且它们的发育形态随沉积环境而变化(
张朝晖等, 2004 ;
郭云等, 2018 ).如在洞穴弱光带一般形成洞穴石刺、向光钟乳石、洞口叠层石等, 这类形态有很强的方向性(
王福星等, 1998 ).苔藓植物在水生环境中也常常形成多样的沉积形态, 如凤尾藓(
Fissidens )钙华、石灰藓(
Hydrogonium )钙华形成的溪流瀑布沉积, 泽藓(
Philonotis )钙华、水灰藓(
Hygrohypnum )钙华形成的河流瀑布沉积, 地钱-真藓(
Marchantia-Bryum )钙华等形成的石灰华坝沉积, 以及净口藓(
Gymnostomum )钙华等形成的瀑布洞穴沉积(
张朝晖和陈家宽, 2007 ).同时, 也有研究发现, 水流环境也是影响苔藓钙华沉积的重要因素, 苔藓与钙华层之间的水流速度与钙华沉积速率呈正相关关系(
Wang et al ., 2014 ).钙华作为大陆古气候的潜在档案,
Yan等(2017) 利用碳氧稳定同位素比率(
δ 13 C和
δ 18 O), 研究钙华在沉积过程中碳、氧的循环规律, 分析认为, 钙华沉积与季节更替引起的气温、降水等的变化密切相关.目前对于苔藓植物沉积作用的研究主要集中在沉积形态以及成土量方面, 对于苔藓植物沉积速率, 对岩溶地区成土过程中的贡献, 以及岩溶沉积作用对生境的改造和环境对岩溶沉积作用的影响等方面的研究还未见报道. ...
Effects of storage temperature on the physiological characteristics and vegetative propagation of desiccation-tolerant mosses
1
2018
... 苔藓植物群落分布特征与生境条件密切相关(
Heino & Virtanen, 2006 ;
刘艳等, 2015 ;
Mod et al ., 2016 ;
李泽科等, 2018 ;
庞嘉鹏等, 2018 ), 温度和水分是影响苔藓植物生存和分布的主要决定性因素, 尤其是空气湿度(
娄红祥, 2012 ;
Guo & Zhao, 2018 ).
党成强等(2018) 研究表明, 南亚毛灰藓(
Homomallium simlaense )在干旱和高温条件下, 各项生理指标均与体内相对含水量呈正比.苔藓植物在干旱缺水的情况下会停止生理代谢进入休眠状态, 当水分充足时再进行生命活动.而且苔藓植物生物量与相对生长速率随光照增强而增加, 随光照时间的延长, 其长度、pH值降低能力逐渐提高(
St. Martin & Mallik, 2017 ).苔藓植物虽然能够忍受高温, 但是温度高于25 ℃时却很少有净光合产物(
Proctor, 2000 ;
Hanson et al ., 2014 ).同时, 温度也是影响酶活性的关键因素, 温度超过60 ℃时会严重影响CA的活性(
洪子茜, 2018 ).然而, CA参与苔藓植物岩溶作用的最佳酶促反应温度还不得而知.同时, 温度也影响有机酸的反应性、络合能力和岩石中矿物自身的溶解程度(
闫志为等, 2009 ;
Benbouzid et al ., 2012 ), 进而影响岩石的溶蚀强度(
陈传平等, 2008 ).而且有机酸与矿物反应生成有机酸盐的存在形态也随温度发生变化(
Peters et al ., 2018 ). ...
Diffusion limitation and CO2 concentrating mechanisms in bryophytes. In: Hanson DT, Rice SK eds. Photosynthesis in Bryophytes and Early Land Plants
1
2014
... 苔藓植物群落分布特征与生境条件密切相关(
Heino & Virtanen, 2006 ;
刘艳等, 2015 ;
Mod et al ., 2016 ;
李泽科等, 2018 ;
庞嘉鹏等, 2018 ), 温度和水分是影响苔藓植物生存和分布的主要决定性因素, 尤其是空气湿度(
娄红祥, 2012 ;
Guo & Zhao, 2018 ).
党成强等(2018) 研究表明, 南亚毛灰藓(
Homomallium simlaense )在干旱和高温条件下, 各项生理指标均与体内相对含水量呈正比.苔藓植物在干旱缺水的情况下会停止生理代谢进入休眠状态, 当水分充足时再进行生命活动.而且苔藓植物生物量与相对生长速率随光照增强而增加, 随光照时间的延长, 其长度、pH值降低能力逐渐提高(
St. Martin & Mallik, 2017 ).苔藓植物虽然能够忍受高温, 但是温度高于25 ℃时却很少有净光合产物(
Proctor, 2000 ;
Hanson et al ., 2014 ).同时, 温度也是影响酶活性的关键因素, 温度超过60 ℃时会严重影响CA的活性(
洪子茜, 2018 ).然而, CA参与苔藓植物岩溶作用的最佳酶促反应温度还不得而知.同时, 温度也影响有机酸的反应性、络合能力和岩石中矿物自身的溶解程度(
闫志为等, 2009 ;
Benbouzid et al ., 2012 ), 进而影响岩石的溶蚀强度(
陈传平等, 2008 ).而且有机酸与矿物反应生成有机酸盐的存在形态也随温度发生变化(
Peters et al ., 2018 ). ...
The influence of biological soil crusts on mineral uptake by associated vascular plants
1
2001
... 生物岩溶是指由生物引起的岩溶作用现象及 其产物, 即生物溶蚀、破坏可溶岩和沉淀形成次生化学沉积物及其所遗留或形成的形态(
Schneider, 1976 ).生物岩溶作用(溶蚀作用和沉积作用)激活了岩溶生态系统基质中的物质和能量, 使各个圈层间信息交流更加频繁与复杂.苔藓植物作为生态系统演替过程中的先锋植物(
薛建辉, 2013 ), 通过生物岩溶作用改善裸岩生境, 为草本和灌木的出现奠定了物质基础(
Harper & Belnap, 2001 ;
Jackson, 2015 ), 对岩溶生态系统的维持与发展具有重要意义(
Li et al ., 2006 ;
Costa et al ., 2018 ).苔藓植物岩溶作用已成为岩溶生态学研究的热点.因此, 了解和掌握岩面苔藓植物功能性状特征、岩溶作用机理及其与环境间的互作机制对研究岩溶地区的生物多样性、石漠化生态恢复、岩溶碳汇以及丰富生物岩溶理论等意义重大.然而, 苔藓植物岩溶作用研究因涉及学科众多、问题复杂, 还未进入系统化研究阶段.为了破除各学科间的壁垒, 促进苔藓植物岩溶作用研究更进一步发展, 本文根据近年来有关苔藓植物岩溶作用的文献资料, 从以下方面综述苔藓植物岩溶作用的研究进展. ...
In situ atomic force microscopy measurements of biotite basal plane reactivity in the presence of oxalic acid
1
2011
... 岩石经苔藓植物溶蚀后, 表面常常出现一些凹凸不平和微溶孔, 这是由于岩石成分不均匀, 导致生物钻孔力的差异性(
王福星等, 1998 ), 以及溶蚀产物流失而形成溶蚀微地貌.
王福星等(1993) 将这些溶蚀微地貌, 按照口面形态、直径大小、深度等, 把短径小于1 mm的命名为微溶孔, 把短径为1-20 mm的命名为溶孔, 把短径为20-100 mm的命名为溶坑.
付兰和张朝晖(2010) 将连续分布的苔藓植物溶蚀微地貌, 按测算结果, 把长7.7-14.4 cm的命名为溶丝; 长0.8-19.0 cm, 宽0.6-7.2 cm的命名为溶斑; 长6.0-24.5 cm, 宽1.2-2.2 cm的命名为溶带.苔藓植物溶蚀过程很难通过宏观手段观测, 导致这两种命名法在大小和形态方面存在明显差异.然而,
Haward等(2011) 借助原子力显微镜(AFM)观察到黑云母表面浅层溶蚀坑的形成过程, 通过分析图像计算质量损失, 从而确定溶蚀过程中黑云母表面的活化能大小.研究表明, 通过分析活化能得到的溶坑大小与测量其周长所得的数据一致.因此, 在研究溶蚀微地貌时, 应采用高精度仪器, 统一测量标准, 分析微地貌是苔藓植物溶蚀作用产生, 还是由于淋溶风化而形成, 有利于后期岩溶微地貌之间的对比分析和进一步深入研究.这些微地貌的形态特征与生物类型和发育生境密切相关(
蔡胡霖, 2017 ), 在温湿度优越的地区发育更为广泛(
Liang & Xu, 2014 ;
Liu et al ., 2016 ).例如, 溶孔、溶锥多发生在低湿度、强光照、高温环境中, 而溶沟多是在高温、强光和pH较高的生境内发育(
申家琛等, 2018a ).
曹建华和王福星(1996) 认为苔藓植物岩溶微地貌与岩溶地区的宏观地貌形态之间存在一定的相关性, 然而目前这种关联性以及苔藓植物微地貌之间的关系和发育方向尚未得到科学揭示. ...
Relationships between distribution and abundance vary with spatial scale and ecological group in stream bryophytes
1
2006
... 苔藓植物群落分布特征与生境条件密切相关(
Heino & Virtanen, 2006 ;
刘艳等, 2015 ;
Mod et al ., 2016 ;
李泽科等, 2018 ;
庞嘉鹏等, 2018 ), 温度和水分是影响苔藓植物生存和分布的主要决定性因素, 尤其是空气湿度(
娄红祥, 2012 ;
Guo & Zhao, 2018 ).
党成强等(2018) 研究表明, 南亚毛灰藓(
Homomallium simlaense )在干旱和高温条件下, 各项生理指标均与体内相对含水量呈正比.苔藓植物在干旱缺水的情况下会停止生理代谢进入休眠状态, 当水分充足时再进行生命活动.而且苔藓植物生物量与相对生长速率随光照增强而增加, 随光照时间的延长, 其长度、pH值降低能力逐渐提高(
St. Martin & Mallik, 2017 ).苔藓植物虽然能够忍受高温, 但是温度高于25 ℃时却很少有净光合产物(
Proctor, 2000 ;
Hanson et al ., 2014 ).同时, 温度也是影响酶活性的关键因素, 温度超过60 ℃时会严重影响CA的活性(
洪子茜, 2018 ).然而, CA参与苔藓植物岩溶作用的最佳酶促反应温度还不得而知.同时, 温度也影响有机酸的反应性、络合能力和岩石中矿物自身的溶解程度(
闫志为等, 2009 ;
Benbouzid et al ., 2012 ), 进而影响岩石的溶蚀强度(
陈传平等, 2008 ).而且有机酸与矿物反应生成有机酸盐的存在形态也随温度发生变化(
Peters et al ., 2018 ). ...
微生物碳酸酐酶特性研究
3
2018
... 苔藓植物群落分布特征与生境条件密切相关(
Heino & Virtanen, 2006 ;
刘艳等, 2015 ;
Mod et al ., 2016 ;
李泽科等, 2018 ;
庞嘉鹏等, 2018 ), 温度和水分是影响苔藓植物生存和分布的主要决定性因素, 尤其是空气湿度(
娄红祥, 2012 ;
Guo & Zhao, 2018 ).
党成强等(2018) 研究表明, 南亚毛灰藓(
Homomallium simlaense )在干旱和高温条件下, 各项生理指标均与体内相对含水量呈正比.苔藓植物在干旱缺水的情况下会停止生理代谢进入休眠状态, 当水分充足时再进行生命活动.而且苔藓植物生物量与相对生长速率随光照增强而增加, 随光照时间的延长, 其长度、pH值降低能力逐渐提高(
St. Martin & Mallik, 2017 ).苔藓植物虽然能够忍受高温, 但是温度高于25 ℃时却很少有净光合产物(
Proctor, 2000 ;
Hanson et al ., 2014 ).同时, 温度也是影响酶活性的关键因素, 温度超过60 ℃时会严重影响CA的活性(
洪子茜, 2018 ).然而, CA参与苔藓植物岩溶作用的最佳酶促反应温度还不得而知.同时, 温度也影响有机酸的反应性、络合能力和岩石中矿物自身的溶解程度(
闫志为等, 2009 ;
Benbouzid et al ., 2012 ), 进而影响岩石的溶蚀强度(
陈传平等, 2008 ).而且有机酸与矿物反应生成有机酸盐的存在形态也随温度发生变化(
Peters et al ., 2018 ). ...
...
Xie和Lou (2009) 认为, 生物岩溶作用随pH值降低而增强, 如果单纯从H
+ + CaCO
3 $\rightleftharpoons $ CO
2 + H
2 O + Ca
2+ 角度分析pH对岩溶作用的影响, pH值越低溶蚀速率越快.然而岩石的最佳溶解pH由矿物自身属性决定(
Bray et al ., 2015 ), 同时有机酸的溶蚀能力也受pH影响(
Pachana et al ., 2012 ;
Chen et al ., 2016 ), 而且每类苔藓植物的最适生长pH也不尽相同.如泥炭藓属(
Sphagnum )植物适宜偏酸性环境, 细叶小羽藓(
Haplocladium microphyllum )配子体的最适生长pH值为7 (
陈文佳等, 2012 ).作为苔藓植物岩溶作用的重要参与者CA, 它的活性也受pH影响,
洪子茜(2018) 研究表明, 当pH在7-9条件下CA的活性最高.pH由外而内对苔藓植物岩溶作用的过程、速率、对象、各级参与者都产生不同程度的促进或抑制作用.目前我们对pH影响苔藓植物岩溶的规律以及调节机制还不得而知, 今后还需综合全面地进行深入研究. ...
... 由于苔藓植物属于贫营养型, 它所需要的营养物质是维管束植物的1/10, 但是对N、P、K、Ca、Mg、Fe、硫(S)等元素的需求仍必不可少(
娄红祥, 2012 ).Ca
2+ 作为生物的第二信使, 具有调节生物抗逆性等多种功能, 对苔藓植物的生长有显著影响(
Amao et al ., 2010 ;
Kolupaev et al ., 2014 ).当苔藓植物缺Ca时会导致生长缓慢, 假根分化少, 原丝体不发育, 叶状体形状改变等(
娄红祥, 2012 ).而且CA活性也受Fe
2+ 、锌离子(Zn
2+ )、硝酸根(NO
- 3 )、氯离子(Cl
- )、溴离子(Br
- )、有机碳、交换性钙等的显著影响(
洪子茜, 2018 ).例如,
黄芬等(2015) 研究表明, Ca对CA的活性有很强的促进作用.生物与环境协同进化致使某些苔藓出现钙依赖性, 这种对钙的依赖性是否表现在基因水平, 从而影响岩溶地区苔藓植物生物多样性、空间分布特征、功能性状等还有待进一步研究.因此在研究苔藓植物岩溶作用与pH及营养元素之间的关系时, 应综合考虑苔藓植物种类、酶活性、逆境胁迫、水环境、基岩性质等因素. ...
微生物碳酸酐酶特性研究
3
2018
... 苔藓植物群落分布特征与生境条件密切相关(
Heino & Virtanen, 2006 ;
刘艳等, 2015 ;
Mod et al ., 2016 ;
李泽科等, 2018 ;
庞嘉鹏等, 2018 ), 温度和水分是影响苔藓植物生存和分布的主要决定性因素, 尤其是空气湿度(
娄红祥, 2012 ;
Guo & Zhao, 2018 ).
党成强等(2018) 研究表明, 南亚毛灰藓(
Homomallium simlaense )在干旱和高温条件下, 各项生理指标均与体内相对含水量呈正比.苔藓植物在干旱缺水的情况下会停止生理代谢进入休眠状态, 当水分充足时再进行生命活动.而且苔藓植物生物量与相对生长速率随光照增强而增加, 随光照时间的延长, 其长度、pH值降低能力逐渐提高(
St. Martin & Mallik, 2017 ).苔藓植物虽然能够忍受高温, 但是温度高于25 ℃时却很少有净光合产物(
Proctor, 2000 ;
Hanson et al ., 2014 ).同时, 温度也是影响酶活性的关键因素, 温度超过60 ℃时会严重影响CA的活性(
洪子茜, 2018 ).然而, CA参与苔藓植物岩溶作用的最佳酶促反应温度还不得而知.同时, 温度也影响有机酸的反应性、络合能力和岩石中矿物自身的溶解程度(
闫志为等, 2009 ;
Benbouzid et al ., 2012 ), 进而影响岩石的溶蚀强度(
陈传平等, 2008 ).而且有机酸与矿物反应生成有机酸盐的存在形态也随温度发生变化(
Peters et al ., 2018 ). ...
...
Xie和Lou (2009) 认为, 生物岩溶作用随pH值降低而增强, 如果单纯从H
+ + CaCO
3 $\rightleftharpoons $ CO
2 + H
2 O + Ca
2+ 角度分析pH对岩溶作用的影响, pH值越低溶蚀速率越快.然而岩石的最佳溶解pH由矿物自身属性决定(
Bray et al ., 2015 ), 同时有机酸的溶蚀能力也受pH影响(
Pachana et al ., 2012 ;
Chen et al ., 2016 ), 而且每类苔藓植物的最适生长pH也不尽相同.如泥炭藓属(
Sphagnum )植物适宜偏酸性环境, 细叶小羽藓(
Haplocladium microphyllum )配子体的最适生长pH值为7 (
陈文佳等, 2012 ).作为苔藓植物岩溶作用的重要参与者CA, 它的活性也受pH影响,
洪子茜(2018) 研究表明, 当pH在7-9条件下CA的活性最高.pH由外而内对苔藓植物岩溶作用的过程、速率、对象、各级参与者都产生不同程度的促进或抑制作用.目前我们对pH影响苔藓植物岩溶的规律以及调节机制还不得而知, 今后还需综合全面地进行深入研究. ...
... 由于苔藓植物属于贫营养型, 它所需要的营养物质是维管束植物的1/10, 但是对N、P、K、Ca、Mg、Fe、硫(S)等元素的需求仍必不可少(
娄红祥, 2012 ).Ca
2+ 作为生物的第二信使, 具有调节生物抗逆性等多种功能, 对苔藓植物的生长有显著影响(
Amao et al ., 2010 ;
Kolupaev et al ., 2014 ).当苔藓植物缺Ca时会导致生长缓慢, 假根分化少, 原丝体不发育, 叶状体形状改变等(
娄红祥, 2012 ).而且CA活性也受Fe
2+ 、锌离子(Zn
2+ )、硝酸根(NO
- 3 )、氯离子(Cl
- )、溴离子(Br
- )、有机碳、交换性钙等的显著影响(
洪子茜, 2018 ).例如,
黄芬等(2015) 研究表明, Ca对CA的活性有很强的促进作用.生物与环境协同进化致使某些苔藓出现钙依赖性, 这种对钙的依赖性是否表现在基因水平, 从而影响岩溶地区苔藓植物生物多样性、空间分布特征、功能性状等还有待进一步研究.因此在研究苔藓植物岩溶作用与pH及营养元素之间的关系时, 应综合考虑苔藓植物种类、酶活性、逆境胁迫、水环境、基岩性质等因素. ...
岩溶环境因子对桂林毛村岩溶区土壤微生物胞外碳酸酐酶活性的影响
1
2015
... 由于苔藓植物属于贫营养型, 它所需要的营养物质是维管束植物的1/10, 但是对N、P、K、Ca、Mg、Fe、硫(S)等元素的需求仍必不可少(
娄红祥, 2012 ).Ca
2+ 作为生物的第二信使, 具有调节生物抗逆性等多种功能, 对苔藓植物的生长有显著影响(
Amao et al ., 2010 ;
Kolupaev et al ., 2014 ).当苔藓植物缺Ca时会导致生长缓慢, 假根分化少, 原丝体不发育, 叶状体形状改变等(
娄红祥, 2012 ).而且CA活性也受Fe
2+ 、锌离子(Zn
2+ )、硝酸根(NO
- 3 )、氯离子(Cl
- )、溴离子(Br
- )、有机碳、交换性钙等的显著影响(
洪子茜, 2018 ).例如,
黄芬等(2015) 研究表明, Ca对CA的活性有很强的促进作用.生物与环境协同进化致使某些苔藓出现钙依赖性, 这种对钙的依赖性是否表现在基因水平, 从而影响岩溶地区苔藓植物生物多样性、空间分布特征、功能性状等还有待进一步研究.因此在研究苔藓植物岩溶作用与pH及营养元素之间的关系时, 应综合考虑苔藓植物种类、酶活性、逆境胁迫、水环境、基岩性质等因素. ...
岩溶环境因子对桂林毛村岩溶区土壤微生物胞外碳酸酐酶活性的影响
1
2015
... 由于苔藓植物属于贫营养型, 它所需要的营养物质是维管束植物的1/10, 但是对N、P、K、Ca、Mg、Fe、硫(S)等元素的需求仍必不可少(
娄红祥, 2012 ).Ca
2+ 作为生物的第二信使, 具有调节生物抗逆性等多种功能, 对苔藓植物的生长有显著影响(
Amao et al ., 2010 ;
Kolupaev et al ., 2014 ).当苔藓植物缺Ca时会导致生长缓慢, 假根分化少, 原丝体不发育, 叶状体形状改变等(
娄红祥, 2012 ).而且CA活性也受Fe
2+ 、锌离子(Zn
2+ )、硝酸根(NO
- 3 )、氯离子(Cl
- )、溴离子(Br
- )、有机碳、交换性钙等的显著影响(
洪子茜, 2018 ).例如,
黄芬等(2015) 研究表明, Ca对CA的活性有很强的促进作用.生物与环境协同进化致使某些苔藓出现钙依赖性, 这种对钙的依赖性是否表现在基因水平, 从而影响岩溶地区苔藓植物生物多样性、空间分布特征、功能性状等还有待进一步研究.因此在研究苔藓植物岩溶作用与pH及营养元素之间的关系时, 应综合考虑苔藓植物种类、酶活性、逆境胁迫、水环境、基岩性质等因素. ...
不同岩性试片溶蚀速率差异及意义
1
2015
... 岩石中所含矿物种类及其晶体结构、产状、透水性等决定了岩石的溶解能力(
Voinot et al ., 2013 ;
黄奇波等, 2015 ), 例如, 一般情况下泥质含量高的碳酸盐岩溶蚀速度快, 岩石中酸不溶物含量越低, 越易被溶解(
王世杰等, 1999 ).同时, 岩石中酸不溶物含量对碳酸盐岩的成土速率具有重要影响, 酸不溶物含量越高岩石风化后可残留物越多, 成土物质越丰富(
陈洪松等, 2018 ).例如, 白云岩中酸不溶物含量高, 发育的土层较石灰岩地区土层厚(
陈洪松等, 2013 ).岩石中所含矿物的性质影响各元素的释放速率, Andrew等(2015)在25 ℃、pH = 2.6的开放试验系统与封闭试验系统内发现黑云母中的元素释放按照Fe > Mg > Al > Si的顺序进行, 且溶解速率符合 ${{r}_{i}}={{k}_{\text{H},i}}\times a_{{{\text{H}}^{\text{+}}}}^{xi}$,
k H 为表面速率常数((2.15-30.6) × 10
-10 mole s
biotite ·m
-2 ·s
-1 ),
a H+ 为H
+ 活性,
xi 为矿物与H
+ 的反应级数(0.31-0.58); 分析认为, 岩石中各矿物的溶解具有空间各向异性, 其边缘的反应效率强于基面, 而且各个元素的释放具有一定的规律性.苔藓植物种类的空间分布特征与岩性密切相关(
Gorozhankina & Konstantinov, 2001 ), 而且岩性和有机酸种类也会影响有机酸与岩石的反应速率(
Cama & Ganor, 2006 ;
Ghoorah et al ., 2014 ;
Mwangi et al ., 2018 ), 从而导致它们岩溶作用的差异性. ...
不同岩性试片溶蚀速率差异及意义
1
2015
... 岩石中所含矿物种类及其晶体结构、产状、透水性等决定了岩石的溶解能力(
Voinot et al ., 2013 ;
黄奇波等, 2015 ), 例如, 一般情况下泥质含量高的碳酸盐岩溶蚀速度快, 岩石中酸不溶物含量越低, 越易被溶解(
王世杰等, 1999 ).同时, 岩石中酸不溶物含量对碳酸盐岩的成土速率具有重要影响, 酸不溶物含量越高岩石风化后可残留物越多, 成土物质越丰富(
陈洪松等, 2018 ).例如, 白云岩中酸不溶物含量高, 发育的土层较石灰岩地区土层厚(
陈洪松等, 2013 ).岩石中所含矿物的性质影响各元素的释放速率, Andrew等(2015)在25 ℃、pH = 2.6的开放试验系统与封闭试验系统内发现黑云母中的元素释放按照Fe > Mg > Al > Si的顺序进行, 且溶解速率符合 ${{r}_{i}}={{k}_{\text{H},i}}\times a_{{{\text{H}}^{\text{+}}}}^{xi}$,
k H 为表面速率常数((2.15-30.6) × 10
-10 mole s
biotite ·m
-2 ·s
-1 ),
a H+ 为H
+ 活性,
xi 为矿物与H
+ 的反应级数(0.31-0.58); 分析认为, 岩石中各矿物的溶解具有空间各向异性, 其边缘的反应效率强于基面, 而且各个元素的释放具有一定的规律性.苔藓植物种类的空间分布特征与岩性密切相关(
Gorozhankina & Konstantinov, 2001 ), 而且岩性和有机酸种类也会影响有机酸与岩石的反应速率(
Cama & Ganor, 2006 ;
Ghoorah et al ., 2014 ;
Mwangi et al ., 2018 ), 从而导致它们岩溶作用的差异性. ...
Weathering, secondary mineral genesis, and soil formation caused by lichens and mosses growing on granitic gneiss in a boreal forest environment
2
2015
... 生物岩溶是指由生物引起的岩溶作用现象及 其产物, 即生物溶蚀、破坏可溶岩和沉淀形成次生化学沉积物及其所遗留或形成的形态(
Schneider, 1976 ).生物岩溶作用(溶蚀作用和沉积作用)激活了岩溶生态系统基质中的物质和能量, 使各个圈层间信息交流更加频繁与复杂.苔藓植物作为生态系统演替过程中的先锋植物(
薛建辉, 2013 ), 通过生物岩溶作用改善裸岩生境, 为草本和灌木的出现奠定了物质基础(
Harper & Belnap, 2001 ;
Jackson, 2015 ), 对岩溶生态系统的维持与发展具有重要意义(
Li et al ., 2006 ;
Costa et al ., 2018 ).苔藓植物岩溶作用已成为岩溶生态学研究的热点.因此, 了解和掌握岩面苔藓植物功能性状特征、岩溶作用机理及其与环境间的互作机制对研究岩溶地区的生物多样性、石漠化生态恢复、岩溶碳汇以及丰富生物岩溶理论等意义重大.然而, 苔藓植物岩溶作用研究因涉及学科众多、问题复杂, 还未进入系统化研究阶段.为了破除各学科间的壁垒, 促进苔藓植物岩溶作用研究更进一步发展, 本文根据近年来有关苔藓植物岩溶作用的文献资料, 从以下方面综述苔藓植物岩溶作用的研究进展. ...
... 苔藓植物常以丛集型、交织型、匍匐型等着生于岩面(
图1 ), 假根根系不发达, 只起固着作用(
娄玉霞, 2013 ), 其特殊的叶表面结构和细胞特征(
娄红祥, 2012 ), 使苔藓具有耐高温、抗旱、蓄水能力强、保湿固土等功能(
Proctor & Pence, 2002 ;
Krol et al ., 2003 ;
Zhang et al ., 2017 ).
曹建华和袁道先(1999) 研究发现, 当岩石表面有苔藓植物时, 其蒸发失水和主动吸水时间比裸岩分别延长了48%和57%, 持水量提高了16.6倍, 分析认为苔藓植物可以延缓岩面水分蒸发时间, 增加岩石水化学作用时间, 但是苔藓植物的这种蓄水保湿能力存在明显的种间差异(
申家琛等, 2017 ;
张显强等, 2018a ).苔藓植物参与岩溶作用的另一大优势, 是其叶表面角质层不发育, 且阳离子交换位点多(
吴玉环等, 2001 ), 能够吸附多种阳离子, 这些阳离子在苔藓植物体内被转化为无机盐、有机盐以及其他形式的化合物储存于组织中.苔藓植物除了以生物化学方式对岩石进行溶蚀之外, 还可以通过物理机械作用破坏岩石.例如, 苔藓植物的假根可以伸入岩石裂隙, 对岩石有一定的破坏作用(
Jackson, 2015 ).再者, 当气候干旱时, 苔藓植物在卷缩过程中会拔起岩面的一些小颗粒, 从而加速了岩石被破坏和成土速度(
连宾等, 2008 ).此外, 苔藓植物常与藻类、地衣、微生物等共生形成生物结皮, 它们之间的相互关系以及因气候因素引起的干湿交替和冻融作用对岩石也具有一定的破坏性(
郑云普等, 2009 ).例如, 菌类的菌丝可以伸入岩石晶体结构中破坏岩石, 细菌与真菌形成的配合基能够络合三价铁离子, 细菌分泌的柠檬酸、乙酸以及地衣分泌的地衣酸(
连宾等, 2011 ), 可以溶解岩石中的矿物(
Won-Pyo et al ., 2016 ;
Wang et al ., 2017 ).因此在研究苔藓植物岩溶作用时, 应明确苔藓植物自身及其与其他生物形成的生物结皮两者之间的关系.目前, 有关岩面苔藓植物功能性状的研究主要集中在小范围内特定生境中的苔藓植物研究, 无法代表岩溶生态系统苔藓植物的整体情况.对于大尺度空间上, 因生境异质性高而造成相同物种在形态结构、光合特征及其钙(Ca)、碳(C)、氮(N)、磷(P) 等元素的代谢调节机制等方面的差异性的研究还未引起人们的广泛关注.岩溶生态系统的物质基础相对贫乏, 具有“缺氮少磷钙丰富”的特征.因此, 研究Ca、C、N、P等元素在苔藓植物体内的代谢机制对揭示岩溶生态系统苔藓植物和其他植物的生物多样性维持机制, 及其岩溶作用特征具有重要的科学 意义. ...
岩溶山区公路石漠化边坡苔藓生态功能
1
2014
... 岩溶地区土壤的形成和发育是碳酸岩长期溶蚀、风化与生物富集过程的结果, 其一方面受岩石结构、性质影响, 另一方面也与植被作用密切相关(
张美良和邓自强, 1994 ).研究表明, 碳酸盐岩的成土速率与岩石溶蚀速率成正比(
曹建华等, 2004 ), 与酸不溶物含量密切相关(
曹沁智等, 2018 ).苔藓植物殖居于岩面, 假根系以及植物体的干湿交替冻融现象所产生的物理机械力作用于岩石, 产生岩石碎屑.同时, 有机物与矿物离子络合常常形成难溶物, 这些难溶物与苔藓及其生物结皮死亡残体黏结在一起, 不仅增加了土壤沉积量, 而且促进有机质积累, 增加土壤养分(
朱显谟, 1995 ;
刘天雷等, 2017 ).苔藓植物通过影响下垫面土壤湿度, 促进土壤对岩石的岩溶作用, 间接地影响沉积过程.同时, 苔藓植物的吸附拦截功能对防治岩石表面水土流失具有重要作用(
李军峰等, 2013 ;
贾少华等, 2014 ;
张显强等, 2018b ).苔藓植物的溶蚀作用为岩面土壤形成提供了原始物质, 关于苔藓对土壤沉积的贡献率与苔藓植物作用下岩面土壤的形成过程, 以及在此过程中岩石的物质、能量转化和土壤养分的变化等还未见报道. ...
岩溶山区公路石漠化边坡苔藓生态功能
1
2014
... 岩溶地区土壤的形成和发育是碳酸岩长期溶蚀、风化与生物富集过程的结果, 其一方面受岩石结构、性质影响, 另一方面也与植被作用密切相关(
张美良和邓自强, 1994 ).研究表明, 碳酸盐岩的成土速率与岩石溶蚀速率成正比(
曹建华等, 2004 ), 与酸不溶物含量密切相关(
曹沁智等, 2018 ).苔藓植物殖居于岩面, 假根系以及植物体的干湿交替冻融现象所产生的物理机械力作用于岩石, 产生岩石碎屑.同时, 有机物与矿物离子络合常常形成难溶物, 这些难溶物与苔藓及其生物结皮死亡残体黏结在一起, 不仅增加了土壤沉积量, 而且促进有机质积累, 增加土壤养分(
朱显谟, 1995 ;
刘天雷等, 2017 ).苔藓植物通过影响下垫面土壤湿度, 促进土壤对岩石的岩溶作用, 间接地影响沉积过程.同时, 苔藓植物的吸附拦截功能对防治岩石表面水土流失具有重要作用(
李军峰等, 2013 ;
贾少华等, 2014 ;
张显强等, 2018b ).苔藓植物的溶蚀作用为岩面土壤形成提供了原始物质, 关于苔藓对土壤沉积的贡献率与苔藓植物作用下岩面土壤的形成过程, 以及在此过程中岩石的物质、能量转化和土壤养分的变化等还未见报道. ...
Cyanobacterial contribution to travertine deposition in the Hoyoux River system, Belgium
1
2017
... 苔藓植物沉积作用的另一表现是石灰华或钙华(
张捷, 1993 ;
Kleinteich et al ., 2017 ).钙华形成的物理化学过程实质是CaCO
3 的沉淀与CO
2 的脱气.苔藓植物在钙华形成过程中为CaCO
3 晶体提供了稳定的生长发育基础, 并且苔藓植物的形态结构控制着沉积物的形态变化(
Mili?a et al ., 2006 ), 且它们的发育形态随沉积环境而变化(
张朝晖等, 2004 ;
郭云等, 2018 ).如在洞穴弱光带一般形成洞穴石刺、向光钟乳石、洞口叠层石等, 这类形态有很强的方向性(
王福星等, 1998 ).苔藓植物在水生环境中也常常形成多样的沉积形态, 如凤尾藓(
Fissidens )钙华、石灰藓(
Hydrogonium )钙华形成的溪流瀑布沉积, 泽藓(
Philonotis )钙华、水灰藓(
Hygrohypnum )钙华形成的河流瀑布沉积, 地钱-真藓(
Marchantia-Bryum )钙华等形成的石灰华坝沉积, 以及净口藓(
Gymnostomum )钙华等形成的瀑布洞穴沉积(
张朝晖和陈家宽, 2007 ).同时, 也有研究发现, 水流环境也是影响苔藓钙华沉积的重要因素, 苔藓与钙华层之间的水流速度与钙华沉积速率呈正相关关系(
Wang et al ., 2014 ).钙华作为大陆古气候的潜在档案,
Yan等(2017) 利用碳氧稳定同位素比率(
δ 13 C和
δ 18 O), 研究钙华在沉积过程中碳、氧的循环规律, 分析认为, 钙华沉积与季节更替引起的气温、降水等的变化密切相关.目前对于苔藓植物沉积作用的研究主要集中在沉积形态以及成土量方面, 对于苔藓植物沉积速率, 对岩溶地区成土过程中的贡献, 以及岩溶沉积作用对生境的改造和环境对岩溶沉积作用的影响等方面的研究还未见报道. ...
The role of reactive oxygen species and calcium ions in the implementation of the stress protective effect of brassinosteroids on plant cells
1
2014
... 由于苔藓植物属于贫营养型, 它所需要的营养物质是维管束植物的1/10, 但是对N、P、K、Ca、Mg、Fe、硫(S)等元素的需求仍必不可少(
娄红祥, 2012 ).Ca
2+ 作为生物的第二信使, 具有调节生物抗逆性等多种功能, 对苔藓植物的生长有显著影响(
Amao et al ., 2010 ;
Kolupaev et al ., 2014 ).当苔藓植物缺Ca时会导致生长缓慢, 假根分化少, 原丝体不发育, 叶状体形状改变等(
娄红祥, 2012 ).而且CA活性也受Fe
2+ 、锌离子(Zn
2+ )、硝酸根(NO
- 3 )、氯离子(Cl
- )、溴离子(Br
- )、有机碳、交换性钙等的显著影响(
洪子茜, 2018 ).例如,
黄芬等(2015) 研究表明, Ca对CA的活性有很强的促进作用.生物与环境协同进化致使某些苔藓出现钙依赖性, 这种对钙的依赖性是否表现在基因水平, 从而影响岩溶地区苔藓植物生物多样性、空间分布特征、功能性状等还有待进一步研究.因此在研究苔藓植物岩溶作用与pH及营养元素之间的关系时, 应综合考虑苔藓植物种类、酶活性、逆境胁迫、水环境、基岩性质等因素. ...
Low-temperature induced transmembrane potential changes in the liverwort
Conocephalum conicum 1
2003
... 苔藓植物常以丛集型、交织型、匍匐型等着生于岩面(
图1 ), 假根根系不发达, 只起固着作用(
娄玉霞, 2013 ), 其特殊的叶表面结构和细胞特征(
娄红祥, 2012 ), 使苔藓具有耐高温、抗旱、蓄水能力强、保湿固土等功能(
Proctor & Pence, 2002 ;
Krol et al ., 2003 ;
Zhang et al ., 2017 ).
曹建华和袁道先(1999) 研究发现, 当岩石表面有苔藓植物时, 其蒸发失水和主动吸水时间比裸岩分别延长了48%和57%, 持水量提高了16.6倍, 分析认为苔藓植物可以延缓岩面水分蒸发时间, 增加岩石水化学作用时间, 但是苔藓植物的这种蓄水保湿能力存在明显的种间差异(
申家琛等, 2017 ;
张显强等, 2018a ).苔藓植物参与岩溶作用的另一大优势, 是其叶表面角质层不发育, 且阳离子交换位点多(
吴玉环等, 2001 ), 能够吸附多种阳离子, 这些阳离子在苔藓植物体内被转化为无机盐、有机盐以及其他形式的化合物储存于组织中.苔藓植物除了以生物化学方式对岩石进行溶蚀之外, 还可以通过物理机械作用破坏岩石.例如, 苔藓植物的假根可以伸入岩石裂隙, 对岩石有一定的破坏作用(
Jackson, 2015 ).再者, 当气候干旱时, 苔藓植物在卷缩过程中会拔起岩面的一些小颗粒, 从而加速了岩石被破坏和成土速度(
连宾等, 2008 ).此外, 苔藓植物常与藻类、地衣、微生物等共生形成生物结皮, 它们之间的相互关系以及因气候因素引起的干湿交替和冻融作用对岩石也具有一定的破坏性(
郑云普等, 2009 ).例如, 菌类的菌丝可以伸入岩石晶体结构中破坏岩石, 细菌与真菌形成的配合基能够络合三价铁离子, 细菌分泌的柠檬酸、乙酸以及地衣分泌的地衣酸(
连宾等, 2011 ), 可以溶解岩石中的矿物(
Won-Pyo et al ., 2016 ;
Wang et al ., 2017 ).因此在研究苔藓植物岩溶作用时, 应明确苔藓植物自身及其与其他生物形成的生物结皮两者之间的关系.目前, 有关岩面苔藓植物功能性状的研究主要集中在小范围内特定生境中的苔藓植物研究, 无法代表岩溶生态系统苔藓植物的整体情况.对于大尺度空间上, 因生境异质性高而造成相同物种在形态结构、光合特征及其钙(Ca)、碳(C)、氮(N)、磷(P) 等元素的代谢调节机制等方面的差异性的研究还未引起人们的广泛关注.岩溶生态系统的物质基础相对贫乏, 具有“缺氮少磷钙丰富”的特征.因此, 研究Ca、C、N、P等元素在苔藓植物体内的代谢机制对揭示岩溶生态系统苔藓植物和其他植物的生物多样性维持机制, 及其岩溶作用特征具有重要的科学 意义. ...
Muscovite dissolution kinetics as a function of pH at elevated temperature
1
2017
... 苔藓植物对岩石的溶蚀作用原理与过程实质是岩石中矿物与苔藓植物之间的作用过程(
图2 ).苔藓植物因呼吸作用形成的H
2 CO
3 和代谢分泌物与岩石中的矿物发生酸化、碱化、氧化还原、螯合等反应, 随着反应过程不断推进, 岩石矿物成分发生变化, Ca、镁(Mg)、钠(Na)、钾(K)、铁(Fe)、铝(Al)、硅(Si)等从碳酸盐、硅酸盐、铝酸盐、硫化物、氧化物中溶出, 破坏原岩晶体结构, 达到溶蚀岩石, 促使风化的目的(
Street et al ., 2013 ;
Li et al ., 2015 ;
Lammers et al ., 2017 ).这种岩溶驱动力的大小因植物形态特征、岩石性质与生存环境改变而发生变化.有研究表明, 碳酸盐岩溶蚀过程中, 生物成因的CO
2 占溶蚀强度的49.26%, 有机酸占37.11% (
张楷燕, 2017 ).苔藓植物溶蚀作用过程释放的矿物离子和其他元素部分被苔藓植物拦截形成岩溶沉积物; 部分被苔藓植物吸收利用, 转化为其他物质存储于植物体内; 其余则随降雨冲刷而流失, 进入其他生态系统.关于苔藓植物呼吸作用和其分泌物引起的岩溶作用驱动力、效率及其与苔藓植物生活型、种间差异、生境特征之间的相互关系仍未引起人们的关注, 对苔藓植物溶蚀作用过程和机理、苔藓植物对岩石中矿物营养元素释放规律的影响, 以及这些矿物质在岩溶沉积过程中的形态转化过程及其与生境间的相互作用关系等的研究还未见报道, 这也正是揭示苔藓植物岩溶作用机理的关键点.岩石的溶解速率与有机酸的酸性和溶蚀能力密切相关(
刘胜亮等, 2017 ), 然而有机酸的理化特性取决于其官能团的属性(
赵志曼等, 2018 ).因此, 今后需结合有机化学、气相色谱质谱(GC-MS)、核磁共振(NMR)、液相色谱(HPLC)等技术手段深入研究苔藓植物的溶蚀过程与机理. ...
喀斯特石漠化山区苔藓多样性及水土保持研究
1
2013
... 岩溶地区土壤的形成和发育是碳酸岩长期溶蚀、风化与生物富集过程的结果, 其一方面受岩石结构、性质影响, 另一方面也与植被作用密切相关(
张美良和邓自强, 1994 ).研究表明, 碳酸盐岩的成土速率与岩石溶蚀速率成正比(
曹建华等, 2004 ), 与酸不溶物含量密切相关(
曹沁智等, 2018 ).苔藓植物殖居于岩面, 假根系以及植物体的干湿交替冻融现象所产生的物理机械力作用于岩石, 产生岩石碎屑.同时, 有机物与矿物离子络合常常形成难溶物, 这些难溶物与苔藓及其生物结皮死亡残体黏结在一起, 不仅增加了土壤沉积量, 而且促进有机质积累, 增加土壤养分(
朱显谟, 1995 ;
刘天雷等, 2017 ).苔藓植物通过影响下垫面土壤湿度, 促进土壤对岩石的岩溶作用, 间接地影响沉积过程.同时, 苔藓植物的吸附拦截功能对防治岩石表面水土流失具有重要作用(
李军峰等, 2013 ;
贾少华等, 2014 ;
张显强等, 2018b ).苔藓植物的溶蚀作用为岩面土壤形成提供了原始物质, 关于苔藓对土壤沉积的贡献率与苔藓植物作用下岩面土壤的形成过程, 以及在此过程中岩石的物质、能量转化和土壤养分的变化等还未见报道. ...
喀斯特石漠化山区苔藓多样性及水土保持研究
1
2013
... 岩溶地区土壤的形成和发育是碳酸岩长期溶蚀、风化与生物富集过程的结果, 其一方面受岩石结构、性质影响, 另一方面也与植被作用密切相关(
张美良和邓自强, 1994 ).研究表明, 碳酸盐岩的成土速率与岩石溶蚀速率成正比(
曹建华等, 2004 ), 与酸不溶物含量密切相关(
曹沁智等, 2018 ).苔藓植物殖居于岩面, 假根系以及植物体的干湿交替冻融现象所产生的物理机械力作用于岩石, 产生岩石碎屑.同时, 有机物与矿物离子络合常常形成难溶物, 这些难溶物与苔藓及其生物结皮死亡残体黏结在一起, 不仅增加了土壤沉积量, 而且促进有机质积累, 增加土壤养分(
朱显谟, 1995 ;
刘天雷等, 2017 ).苔藓植物通过影响下垫面土壤湿度, 促进土壤对岩石的岩溶作用, 间接地影响沉积过程.同时, 苔藓植物的吸附拦截功能对防治岩石表面水土流失具有重要作用(
李军峰等, 2013 ;
贾少华等, 2014 ;
张显强等, 2018b ).苔藓植物的溶蚀作用为岩面土壤形成提供了原始物质, 关于苔藓对土壤沉积的贡献率与苔藓植物作用下岩面土壤的形成过程, 以及在此过程中岩石的物质、能量转化和土壤养分的变化等还未见报道. ...
A nano-scale study of the mechanisms of non-exchangeable potassium release from micas
1
2015
... 苔藓植物对岩石的溶蚀作用原理与过程实质是岩石中矿物与苔藓植物之间的作用过程(
图2 ).苔藓植物因呼吸作用形成的H
2 CO
3 和代谢分泌物与岩石中的矿物发生酸化、碱化、氧化还原、螯合等反应, 随着反应过程不断推进, 岩石矿物成分发生变化, Ca、镁(Mg)、钠(Na)、钾(K)、铁(Fe)、铝(Al)、硅(Si)等从碳酸盐、硅酸盐、铝酸盐、硫化物、氧化物中溶出, 破坏原岩晶体结构, 达到溶蚀岩石, 促使风化的目的(
Street et al ., 2013 ;
Li et al ., 2015 ;
Lammers et al ., 2017 ).这种岩溶驱动力的大小因植物形态特征、岩石性质与生存环境改变而发生变化.有研究表明, 碳酸盐岩溶蚀过程中, 生物成因的CO
2 占溶蚀强度的49.26%, 有机酸占37.11% (
张楷燕, 2017 ).苔藓植物溶蚀作用过程释放的矿物离子和其他元素部分被苔藓植物拦截形成岩溶沉积物; 部分被苔藓植物吸收利用, 转化为其他物质存储于植物体内; 其余则随降雨冲刷而流失, 进入其他生态系统.关于苔藓植物呼吸作用和其分泌物引起的岩溶作用驱动力、效率及其与苔藓植物生活型、种间差异、生境特征之间的相互关系仍未引起人们的关注, 对苔藓植物溶蚀作用过程和机理、苔藓植物对岩石中矿物营养元素释放规律的影响, 以及这些矿物质在岩溶沉积过程中的形态转化过程及其与生境间的相互作用关系等的研究还未见报道, 这也正是揭示苔藓植物岩溶作用机理的关键点.岩石的溶解速率与有机酸的酸性和溶蚀能力密切相关(
刘胜亮等, 2017 ), 然而有机酸的理化特性取决于其官能团的属性(
赵志曼等, 2018 ).因此, 今后需结合有机化学、气相色谱质谱(GC-MS)、核磁共振(NMR)、液相色谱(HPLC)等技术手段深入研究苔藓植物的溶蚀过程与机理. ...
岩溶环境因子对细菌胞外碳酸酐酶表达及活性的影响
1
2005
... 此外, 苔藓植物体内的碳酸酐酶(CA)对岩溶作用也有促进作用(
Smith & Griffiths, 2000 ), 它主要催化CO
2 的水合反应: CO
2 + H
2 O $\rightleftharpoons $ HCO
3 - + H
+ (
吴雁雯和张金池, 2015 ).
李为等(2005) 研究发现无CA催化时上述可逆反应达到平衡的时间是有CA催化时的6 × 10
7 倍.生物体内均含有CA, 且CA活性因物种的不同而存在显著差异, 细菌、真菌、放线菌内均含有CA, 且微生物细胞内CA活性不同于细胞外CA, 细菌的CA对碳酸盐岩系统碳储存贡献在20%以上(
Li et al ., 2006 ), 真菌的CA对白云岩的溶蚀能力强于对石灰岩(
申泰铭等, 2014 ;
申泰铭, 2018 ).那么苔藓植物体内CA对岩溶作用的贡献是多少?各组织内的CA活性是否也存在这种差异?对此以及关于苔藓植物体内CA代谢的调节机制及原理仍不得而知.目前, 关于苔藓植物CA的研究, 主要集中在各类苔藓植物CA含量、活性及其对岩石溶蚀过程的促进作用等方面.例如,
刘天雷等(2017) 研究发现, 虽然苔藓植物都会分泌CA, 但CA分泌量、速率和活性存在种间差异.这种差异性可能是因生活型或者生长环境的不同而导致苔藓植物生理代谢与功能性状的差异引起的. ...
岩溶环境因子对细菌胞外碳酸酐酶表达及活性的影响
1
2005
... 此外, 苔藓植物体内的碳酸酐酶(CA)对岩溶作用也有促进作用(
Smith & Griffiths, 2000 ), 它主要催化CO
2 的水合反应: CO
2 + H
2 O $\rightleftharpoons $ HCO
3 - + H
+ (
吴雁雯和张金池, 2015 ).
李为等(2005) 研究发现无CA催化时上述可逆反应达到平衡的时间是有CA催化时的6 × 10
7 倍.生物体内均含有CA, 且CA活性因物种的不同而存在显著差异, 细菌、真菌、放线菌内均含有CA, 且微生物细胞内CA活性不同于细胞外CA, 细菌的CA对碳酸盐岩系统碳储存贡献在20%以上(
Li et al ., 2006 ), 真菌的CA对白云岩的溶蚀能力强于对石灰岩(
申泰铭等, 2014 ;
申泰铭, 2018 ).那么苔藓植物体内CA对岩溶作用的贡献是多少?各组织内的CA活性是否也存在这种差异?对此以及关于苔藓植物体内CA代谢的调节机制及原理仍不得而知.目前, 关于苔藓植物CA的研究, 主要集中在各类苔藓植物CA含量、活性及其对岩石溶蚀过程的促进作用等方面.例如,
刘天雷等(2017) 研究发现, 虽然苔藓植物都会分泌CA, 但CA分泌量、速率和活性存在种间差异.这种差异性可能是因生活型或者生长环境的不同而导致苔藓植物生理代谢与功能性状的差异引起的. ...
Enhancement of Ca
2 + release from limestone by microbical extracelluar carbonic anhydrase
2
2006
... 生物岩溶是指由生物引起的岩溶作用现象及 其产物, 即生物溶蚀、破坏可溶岩和沉淀形成次生化学沉积物及其所遗留或形成的形态(
Schneider, 1976 ).生物岩溶作用(溶蚀作用和沉积作用)激活了岩溶生态系统基质中的物质和能量, 使各个圈层间信息交流更加频繁与复杂.苔藓植物作为生态系统演替过程中的先锋植物(
薛建辉, 2013 ), 通过生物岩溶作用改善裸岩生境, 为草本和灌木的出现奠定了物质基础(
Harper & Belnap, 2001 ;
Jackson, 2015 ), 对岩溶生态系统的维持与发展具有重要意义(
Li et al ., 2006 ;
Costa et al ., 2018 ).苔藓植物岩溶作用已成为岩溶生态学研究的热点.因此, 了解和掌握岩面苔藓植物功能性状特征、岩溶作用机理及其与环境间的互作机制对研究岩溶地区的生物多样性、石漠化生态恢复、岩溶碳汇以及丰富生物岩溶理论等意义重大.然而, 苔藓植物岩溶作用研究因涉及学科众多、问题复杂, 还未进入系统化研究阶段.为了破除各学科间的壁垒, 促进苔藓植物岩溶作用研究更进一步发展, 本文根据近年来有关苔藓植物岩溶作用的文献资料, 从以下方面综述苔藓植物岩溶作用的研究进展. ...
... 此外, 苔藓植物体内的碳酸酐酶(CA)对岩溶作用也有促进作用(
Smith & Griffiths, 2000 ), 它主要催化CO
2 的水合反应: CO
2 + H
2 O $\rightleftharpoons $ HCO
3 - + H
+ (
吴雁雯和张金池, 2015 ).
李为等(2005) 研究发现无CA催化时上述可逆反应达到平衡的时间是有CA催化时的6 × 10
7 倍.生物体内均含有CA, 且CA活性因物种的不同而存在显著差异, 细菌、真菌、放线菌内均含有CA, 且微生物细胞内CA活性不同于细胞外CA, 细菌的CA对碳酸盐岩系统碳储存贡献在20%以上(
Li et al ., 2006 ), 真菌的CA对白云岩的溶蚀能力强于对石灰岩(
申泰铭等, 2014 ;
申泰铭, 2018 ).那么苔藓植物体内CA对岩溶作用的贡献是多少?各组织内的CA活性是否也存在这种差异?对此以及关于苔藓植物体内CA代谢的调节机制及原理仍不得而知.目前, 关于苔藓植物CA的研究, 主要集中在各类苔藓植物CA含量、活性及其对岩石溶蚀过程的促进作用等方面.例如,
刘天雷等(2017) 研究发现, 虽然苔藓植物都会分泌CA, 但CA分泌量、速率和活性存在种间差异.这种差异性可能是因生活型或者生长环境的不同而导致苔藓植物生理代谢与功能性状的差异引起的. ...
岩溶城市背景下贵阳市峰丛苔藓植物多样性分布及其生态特征
1
2018
... 苔藓植物群落分布特征与生境条件密切相关(
Heino & Virtanen, 2006 ;
刘艳等, 2015 ;
Mod et al ., 2016 ;
李泽科等, 2018 ;
庞嘉鹏等, 2018 ), 温度和水分是影响苔藓植物生存和分布的主要决定性因素, 尤其是空气湿度(
娄红祥, 2012 ;
Guo & Zhao, 2018 ).
党成强等(2018) 研究表明, 南亚毛灰藓(
Homomallium simlaense )在干旱和高温条件下, 各项生理指标均与体内相对含水量呈正比.苔藓植物在干旱缺水的情况下会停止生理代谢进入休眠状态, 当水分充足时再进行生命活动.而且苔藓植物生物量与相对生长速率随光照增强而增加, 随光照时间的延长, 其长度、pH值降低能力逐渐提高(
St. Martin & Mallik, 2017 ).苔藓植物虽然能够忍受高温, 但是温度高于25 ℃时却很少有净光合产物(
Proctor, 2000 ;
Hanson et al ., 2014 ).同时, 温度也是影响酶活性的关键因素, 温度超过60 ℃时会严重影响CA的活性(
洪子茜, 2018 ).然而, CA参与苔藓植物岩溶作用的最佳酶促反应温度还不得而知.同时, 温度也影响有机酸的反应性、络合能力和岩石中矿物自身的溶解程度(
闫志为等, 2009 ;
Benbouzid et al ., 2012 ), 进而影响岩石的溶蚀强度(
陈传平等, 2008 ).而且有机酸与矿物反应生成有机酸盐的存在形态也随温度发生变化(
Peters et al ., 2018 ). ...
岩溶城市背景下贵阳市峰丛苔藓植物多样性分布及其生态特征
1
2018
... 苔藓植物群落分布特征与生境条件密切相关(
Heino & Virtanen, 2006 ;
刘艳等, 2015 ;
Mod et al ., 2016 ;
李泽科等, 2018 ;
庞嘉鹏等, 2018 ), 温度和水分是影响苔藓植物生存和分布的主要决定性因素, 尤其是空气湿度(
娄红祥, 2012 ;
Guo & Zhao, 2018 ).
党成强等(2018) 研究表明, 南亚毛灰藓(
Homomallium simlaense )在干旱和高温条件下, 各项生理指标均与体内相对含水量呈正比.苔藓植物在干旱缺水的情况下会停止生理代谢进入休眠状态, 当水分充足时再进行生命活动.而且苔藓植物生物量与相对生长速率随光照增强而增加, 随光照时间的延长, 其长度、pH值降低能力逐渐提高(
St. Martin & Mallik, 2017 ).苔藓植物虽然能够忍受高温, 但是温度高于25 ℃时却很少有净光合产物(
Proctor, 2000 ;
Hanson et al ., 2014 ).同时, 温度也是影响酶活性的关键因素, 温度超过60 ℃时会严重影响CA的活性(
洪子茜, 2018 ).然而, CA参与苔藓植物岩溶作用的最佳酶促反应温度还不得而知.同时, 温度也影响有机酸的反应性、络合能力和岩石中矿物自身的溶解程度(
闫志为等, 2009 ;
Benbouzid et al ., 2012 ), 进而影响岩石的溶蚀强度(
陈传平等, 2008 ).而且有机酸与矿物反应生成有机酸盐的存在形态也随温度发生变化(
Peters et al ., 2018 ). ...
微生物对碳酸盐岩的风化作用
1
2008
... 苔藓植物常以丛集型、交织型、匍匐型等着生于岩面(
图1 ), 假根根系不发达, 只起固着作用(
娄玉霞, 2013 ), 其特殊的叶表面结构和细胞特征(
娄红祥, 2012 ), 使苔藓具有耐高温、抗旱、蓄水能力强、保湿固土等功能(
Proctor & Pence, 2002 ;
Krol et al ., 2003 ;
Zhang et al ., 2017 ).
曹建华和袁道先(1999) 研究发现, 当岩石表面有苔藓植物时, 其蒸发失水和主动吸水时间比裸岩分别延长了48%和57%, 持水量提高了16.6倍, 分析认为苔藓植物可以延缓岩面水分蒸发时间, 增加岩石水化学作用时间, 但是苔藓植物的这种蓄水保湿能力存在明显的种间差异(
申家琛等, 2017 ;
张显强等, 2018a ).苔藓植物参与岩溶作用的另一大优势, 是其叶表面角质层不发育, 且阳离子交换位点多(
吴玉环等, 2001 ), 能够吸附多种阳离子, 这些阳离子在苔藓植物体内被转化为无机盐、有机盐以及其他形式的化合物储存于组织中.苔藓植物除了以生物化学方式对岩石进行溶蚀之外, 还可以通过物理机械作用破坏岩石.例如, 苔藓植物的假根可以伸入岩石裂隙, 对岩石有一定的破坏作用(
Jackson, 2015 ).再者, 当气候干旱时, 苔藓植物在卷缩过程中会拔起岩面的一些小颗粒, 从而加速了岩石被破坏和成土速度(
连宾等, 2008 ).此外, 苔藓植物常与藻类、地衣、微生物等共生形成生物结皮, 它们之间的相互关系以及因气候因素引起的干湿交替和冻融作用对岩石也具有一定的破坏性(
郑云普等, 2009 ).例如, 菌类的菌丝可以伸入岩石晶体结构中破坏岩石, 细菌与真菌形成的配合基能够络合三价铁离子, 细菌分泌的柠檬酸、乙酸以及地衣分泌的地衣酸(
连宾等, 2011 ), 可以溶解岩石中的矿物(
Won-Pyo et al ., 2016 ;
Wang et al ., 2017 ).因此在研究苔藓植物岩溶作用时, 应明确苔藓植物自身及其与其他生物形成的生物结皮两者之间的关系.目前, 有关岩面苔藓植物功能性状的研究主要集中在小范围内特定生境中的苔藓植物研究, 无法代表岩溶生态系统苔藓植物的整体情况.对于大尺度空间上, 因生境异质性高而造成相同物种在形态结构、光合特征及其钙(Ca)、碳(C)、氮(N)、磷(P) 等元素的代谢调节机制等方面的差异性的研究还未引起人们的广泛关注.岩溶生态系统的物质基础相对贫乏, 具有“缺氮少磷钙丰富”的特征.因此, 研究Ca、C、N、P等元素在苔藓植物体内的代谢机制对揭示岩溶生态系统苔藓植物和其他植物的生物多样性维持机制, 及其岩溶作用特征具有重要的科学 意义. ...
微生物对碳酸盐岩的风化作用
1
2008
... 苔藓植物常以丛集型、交织型、匍匐型等着生于岩面(
图1 ), 假根根系不发达, 只起固着作用(
娄玉霞, 2013 ), 其特殊的叶表面结构和细胞特征(
娄红祥, 2012 ), 使苔藓具有耐高温、抗旱、蓄水能力强、保湿固土等功能(
Proctor & Pence, 2002 ;
Krol et al ., 2003 ;
Zhang et al ., 2017 ).
曹建华和袁道先(1999) 研究发现, 当岩石表面有苔藓植物时, 其蒸发失水和主动吸水时间比裸岩分别延长了48%和57%, 持水量提高了16.6倍, 分析认为苔藓植物可以延缓岩面水分蒸发时间, 增加岩石水化学作用时间, 但是苔藓植物的这种蓄水保湿能力存在明显的种间差异(
申家琛等, 2017 ;
张显强等, 2018a ).苔藓植物参与岩溶作用的另一大优势, 是其叶表面角质层不发育, 且阳离子交换位点多(
吴玉环等, 2001 ), 能够吸附多种阳离子, 这些阳离子在苔藓植物体内被转化为无机盐、有机盐以及其他形式的化合物储存于组织中.苔藓植物除了以生物化学方式对岩石进行溶蚀之外, 还可以通过物理机械作用破坏岩石.例如, 苔藓植物的假根可以伸入岩石裂隙, 对岩石有一定的破坏作用(
Jackson, 2015 ).再者, 当气候干旱时, 苔藓植物在卷缩过程中会拔起岩面的一些小颗粒, 从而加速了岩石被破坏和成土速度(
连宾等, 2008 ).此外, 苔藓植物常与藻类、地衣、微生物等共生形成生物结皮, 它们之间的相互关系以及因气候因素引起的干湿交替和冻融作用对岩石也具有一定的破坏性(
郑云普等, 2009 ).例如, 菌类的菌丝可以伸入岩石晶体结构中破坏岩石, 细菌与真菌形成的配合基能够络合三价铁离子, 细菌分泌的柠檬酸、乙酸以及地衣分泌的地衣酸(
连宾等, 2011 ), 可以溶解岩石中的矿物(
Won-Pyo et al ., 2016 ;
Wang et al ., 2017 ).因此在研究苔藓植物岩溶作用时, 应明确苔藓植物自身及其与其他生物形成的生物结皮两者之间的关系.目前, 有关岩面苔藓植物功能性状的研究主要集中在小范围内特定生境中的苔藓植物研究, 无法代表岩溶生态系统苔藓植物的整体情况.对于大尺度空间上, 因生境异质性高而造成相同物种在形态结构、光合特征及其钙(Ca)、碳(C)、氮(N)、磷(P) 等元素的代谢调节机制等方面的差异性的研究还未引起人们的广泛关注.岩溶生态系统的物质基础相对贫乏, 具有“缺氮少磷钙丰富”的特征.因此, 研究Ca、C、N、P等元素在苔藓植物体内的代谢机制对揭示岩溶生态系统苔藓植物和其他植物的生物多样性维持机制, 及其岩溶作用特征具有重要的科学 意义. ...
岩溶生态系统中微生物对岩溶作用影响的认识
1
2011
... 苔藓植物常以丛集型、交织型、匍匐型等着生于岩面(
图1 ), 假根根系不发达, 只起固着作用(
娄玉霞, 2013 ), 其特殊的叶表面结构和细胞特征(
娄红祥, 2012 ), 使苔藓具有耐高温、抗旱、蓄水能力强、保湿固土等功能(
Proctor & Pence, 2002 ;
Krol et al ., 2003 ;
Zhang et al ., 2017 ).
曹建华和袁道先(1999) 研究发现, 当岩石表面有苔藓植物时, 其蒸发失水和主动吸水时间比裸岩分别延长了48%和57%, 持水量提高了16.6倍, 分析认为苔藓植物可以延缓岩面水分蒸发时间, 增加岩石水化学作用时间, 但是苔藓植物的这种蓄水保湿能力存在明显的种间差异(
申家琛等, 2017 ;
张显强等, 2018a ).苔藓植物参与岩溶作用的另一大优势, 是其叶表面角质层不发育, 且阳离子交换位点多(
吴玉环等, 2001 ), 能够吸附多种阳离子, 这些阳离子在苔藓植物体内被转化为无机盐、有机盐以及其他形式的化合物储存于组织中.苔藓植物除了以生物化学方式对岩石进行溶蚀之外, 还可以通过物理机械作用破坏岩石.例如, 苔藓植物的假根可以伸入岩石裂隙, 对岩石有一定的破坏作用(
Jackson, 2015 ).再者, 当气候干旱时, 苔藓植物在卷缩过程中会拔起岩面的一些小颗粒, 从而加速了岩石被破坏和成土速度(
连宾等, 2008 ).此外, 苔藓植物常与藻类、地衣、微生物等共生形成生物结皮, 它们之间的相互关系以及因气候因素引起的干湿交替和冻融作用对岩石也具有一定的破坏性(
郑云普等, 2009 ).例如, 菌类的菌丝可以伸入岩石晶体结构中破坏岩石, 细菌与真菌形成的配合基能够络合三价铁离子, 细菌分泌的柠檬酸、乙酸以及地衣分泌的地衣酸(
连宾等, 2011 ), 可以溶解岩石中的矿物(
Won-Pyo et al ., 2016 ;
Wang et al ., 2017 ).因此在研究苔藓植物岩溶作用时, 应明确苔藓植物自身及其与其他生物形成的生物结皮两者之间的关系.目前, 有关岩面苔藓植物功能性状的研究主要集中在小范围内特定生境中的苔藓植物研究, 无法代表岩溶生态系统苔藓植物的整体情况.对于大尺度空间上, 因生境异质性高而造成相同物种在形态结构、光合特征及其钙(Ca)、碳(C)、氮(N)、磷(P) 等元素的代谢调节机制等方面的差异性的研究还未引起人们的广泛关注.岩溶生态系统的物质基础相对贫乏, 具有“缺氮少磷钙丰富”的特征.因此, 研究Ca、C、N、P等元素在苔藓植物体内的代谢机制对揭示岩溶生态系统苔藓植物和其他植物的生物多样性维持机制, 及其岩溶作用特征具有重要的科学 意义. ...
岩溶生态系统中微生物对岩溶作用影响的认识
1
2011
... 苔藓植物常以丛集型、交织型、匍匐型等着生于岩面(
图1 ), 假根根系不发达, 只起固着作用(
娄玉霞, 2013 ), 其特殊的叶表面结构和细胞特征(
娄红祥, 2012 ), 使苔藓具有耐高温、抗旱、蓄水能力强、保湿固土等功能(
Proctor & Pence, 2002 ;
Krol et al ., 2003 ;
Zhang et al ., 2017 ).
曹建华和袁道先(1999) 研究发现, 当岩石表面有苔藓植物时, 其蒸发失水和主动吸水时间比裸岩分别延长了48%和57%, 持水量提高了16.6倍, 分析认为苔藓植物可以延缓岩面水分蒸发时间, 增加岩石水化学作用时间, 但是苔藓植物的这种蓄水保湿能力存在明显的种间差异(
申家琛等, 2017 ;
张显强等, 2018a ).苔藓植物参与岩溶作用的另一大优势, 是其叶表面角质层不发育, 且阳离子交换位点多(
吴玉环等, 2001 ), 能够吸附多种阳离子, 这些阳离子在苔藓植物体内被转化为无机盐、有机盐以及其他形式的化合物储存于组织中.苔藓植物除了以生物化学方式对岩石进行溶蚀之外, 还可以通过物理机械作用破坏岩石.例如, 苔藓植物的假根可以伸入岩石裂隙, 对岩石有一定的破坏作用(
Jackson, 2015 ).再者, 当气候干旱时, 苔藓植物在卷缩过程中会拔起岩面的一些小颗粒, 从而加速了岩石被破坏和成土速度(
连宾等, 2008 ).此外, 苔藓植物常与藻类、地衣、微生物等共生形成生物结皮, 它们之间的相互关系以及因气候因素引起的干湿交替和冻融作用对岩石也具有一定的破坏性(
郑云普等, 2009 ).例如, 菌类的菌丝可以伸入岩石晶体结构中破坏岩石, 细菌与真菌形成的配合基能够络合三价铁离子, 细菌分泌的柠檬酸、乙酸以及地衣分泌的地衣酸(
连宾等, 2011 ), 可以溶解岩石中的矿物(
Won-Pyo et al ., 2016 ;
Wang et al ., 2017 ).因此在研究苔藓植物岩溶作用时, 应明确苔藓植物自身及其与其他生物形成的生物结皮两者之间的关系.目前, 有关岩面苔藓植物功能性状的研究主要集中在小范围内特定生境中的苔藓植物研究, 无法代表岩溶生态系统苔藓植物的整体情况.对于大尺度空间上, 因生境异质性高而造成相同物种在形态结构、光合特征及其钙(Ca)、碳(C)、氮(N)、磷(P) 等元素的代谢调节机制等方面的差异性的研究还未引起人们的广泛关注.岩溶生态系统的物质基础相对贫乏, 具有“缺氮少磷钙丰富”的特征.因此, 研究Ca、C、N、P等元素在苔藓植物体内的代谢机制对揭示岩溶生态系统苔藓植物和其他植物的生物多样性维持机制, 及其岩溶作用特征具有重要的科学 意义. ...
Discrimination of tower, cockpit, and non-karst landforms in Guilin, Southern China, based on morphometric characteristics
1
2014
... 岩石经苔藓植物溶蚀后, 表面常常出现一些凹凸不平和微溶孔, 这是由于岩石成分不均匀, 导致生物钻孔力的差异性(
王福星等, 1998 ), 以及溶蚀产物流失而形成溶蚀微地貌.
王福星等(1993) 将这些溶蚀微地貌, 按照口面形态、直径大小、深度等, 把短径小于1 mm的命名为微溶孔, 把短径为1-20 mm的命名为溶孔, 把短径为20-100 mm的命名为溶坑.
付兰和张朝晖(2010) 将连续分布的苔藓植物溶蚀微地貌, 按测算结果, 把长7.7-14.4 cm的命名为溶丝; 长0.8-19.0 cm, 宽0.6-7.2 cm的命名为溶斑; 长6.0-24.5 cm, 宽1.2-2.2 cm的命名为溶带.苔藓植物溶蚀过程很难通过宏观手段观测, 导致这两种命名法在大小和形态方面存在明显差异.然而,
Haward等(2011) 借助原子力显微镜(AFM)观察到黑云母表面浅层溶蚀坑的形成过程, 通过分析图像计算质量损失, 从而确定溶蚀过程中黑云母表面的活化能大小.研究表明, 通过分析活化能得到的溶坑大小与测量其周长所得的数据一致.因此, 在研究溶蚀微地貌时, 应采用高精度仪器, 统一测量标准, 分析微地貌是苔藓植物溶蚀作用产生, 还是由于淋溶风化而形成, 有利于后期岩溶微地貌之间的对比分析和进一步深入研究.这些微地貌的形态特征与生物类型和发育生境密切相关(
蔡胡霖, 2017 ), 在温湿度优越的地区发育更为广泛(
Liang & Xu, 2014 ;
Liu et al ., 2016 ).例如, 溶孔、溶锥多发生在低湿度、强光照、高温环境中, 而溶沟多是在高温、强光和pH较高的生境内发育(
申家琛等, 2018a ).
曹建华和王福星(1996) 认为苔藓植物岩溶微地貌与岩溶地区的宏观地貌形态之间存在一定的相关性, 然而目前这种关联性以及苔藓植物微地貌之间的关系和发育方向尚未得到科学揭示. ...
Karst catchments exhibited higher degradation stress from climate change than the non-karst catchments in southwest China: An ecohydrological perspective
1
2016
... 岩石经苔藓植物溶蚀后, 表面常常出现一些凹凸不平和微溶孔, 这是由于岩石成分不均匀, 导致生物钻孔力的差异性(
王福星等, 1998 ), 以及溶蚀产物流失而形成溶蚀微地貌.
王福星等(1993) 将这些溶蚀微地貌, 按照口面形态、直径大小、深度等, 把短径小于1 mm的命名为微溶孔, 把短径为1-20 mm的命名为溶孔, 把短径为20-100 mm的命名为溶坑.
付兰和张朝晖(2010) 将连续分布的苔藓植物溶蚀微地貌, 按测算结果, 把长7.7-14.4 cm的命名为溶丝; 长0.8-19.0 cm, 宽0.6-7.2 cm的命名为溶斑; 长6.0-24.5 cm, 宽1.2-2.2 cm的命名为溶带.苔藓植物溶蚀过程很难通过宏观手段观测, 导致这两种命名法在大小和形态方面存在明显差异.然而,
Haward等(2011) 借助原子力显微镜(AFM)观察到黑云母表面浅层溶蚀坑的形成过程, 通过分析图像计算质量损失, 从而确定溶蚀过程中黑云母表面的活化能大小.研究表明, 通过分析活化能得到的溶坑大小与测量其周长所得的数据一致.因此, 在研究溶蚀微地貌时, 应采用高精度仪器, 统一测量标准, 分析微地貌是苔藓植物溶蚀作用产生, 还是由于淋溶风化而形成, 有利于后期岩溶微地貌之间的对比分析和进一步深入研究.这些微地貌的形态特征与生物类型和发育生境密切相关(
蔡胡霖, 2017 ), 在温湿度优越的地区发育更为广泛(
Liang & Xu, 2014 ;
Liu et al ., 2016 ).例如, 溶孔、溶锥多发生在低湿度、强光照、高温环境中, 而溶沟多是在高温、强光和pH较高的生境内发育(
申家琛等, 2018a ).
曹建华和王福星(1996) 认为苔藓植物岩溶微地貌与岩溶地区的宏观地貌形态之间存在一定的相关性, 然而目前这种关联性以及苔藓植物微地貌之间的关系和发育方向尚未得到科学揭示. ...
不同有机酸对磷酸三钙溶解能力的研究
1
2017
... 苔藓植物对岩石的溶蚀作用原理与过程实质是岩石中矿物与苔藓植物之间的作用过程(
图2 ).苔藓植物因呼吸作用形成的H
2 CO
3 和代谢分泌物与岩石中的矿物发生酸化、碱化、氧化还原、螯合等反应, 随着反应过程不断推进, 岩石矿物成分发生变化, Ca、镁(Mg)、钠(Na)、钾(K)、铁(Fe)、铝(Al)、硅(Si)等从碳酸盐、硅酸盐、铝酸盐、硫化物、氧化物中溶出, 破坏原岩晶体结构, 达到溶蚀岩石, 促使风化的目的(
Street et al ., 2013 ;
Li et al ., 2015 ;
Lammers et al ., 2017 ).这种岩溶驱动力的大小因植物形态特征、岩石性质与生存环境改变而发生变化.有研究表明, 碳酸盐岩溶蚀过程中, 生物成因的CO
2 占溶蚀强度的49.26%, 有机酸占37.11% (
张楷燕, 2017 ).苔藓植物溶蚀作用过程释放的矿物离子和其他元素部分被苔藓植物拦截形成岩溶沉积物; 部分被苔藓植物吸收利用, 转化为其他物质存储于植物体内; 其余则随降雨冲刷而流失, 进入其他生态系统.关于苔藓植物呼吸作用和其分泌物引起的岩溶作用驱动力、效率及其与苔藓植物生活型、种间差异、生境特征之间的相互关系仍未引起人们的关注, 对苔藓植物溶蚀作用过程和机理、苔藓植物对岩石中矿物营养元素释放规律的影响, 以及这些矿物质在岩溶沉积过程中的形态转化过程及其与生境间的相互作用关系等的研究还未见报道, 这也正是揭示苔藓植物岩溶作用机理的关键点.岩石的溶解速率与有机酸的酸性和溶蚀能力密切相关(
刘胜亮等, 2017 ), 然而有机酸的理化特性取决于其官能团的属性(
赵志曼等, 2018 ).因此, 今后需结合有机化学、气相色谱质谱(GC-MS)、核磁共振(NMR)、液相色谱(HPLC)等技术手段深入研究苔藓植物的溶蚀过程与机理. ...
不同有机酸对磷酸三钙溶解能力的研究
1
2017
... 苔藓植物对岩石的溶蚀作用原理与过程实质是岩石中矿物与苔藓植物之间的作用过程(
图2 ).苔藓植物因呼吸作用形成的H
2 CO
3 和代谢分泌物与岩石中的矿物发生酸化、碱化、氧化还原、螯合等反应, 随着反应过程不断推进, 岩石矿物成分发生变化, Ca、镁(Mg)、钠(Na)、钾(K)、铁(Fe)、铝(Al)、硅(Si)等从碳酸盐、硅酸盐、铝酸盐、硫化物、氧化物中溶出, 破坏原岩晶体结构, 达到溶蚀岩石, 促使风化的目的(
Street et al ., 2013 ;
Li et al ., 2015 ;
Lammers et al ., 2017 ).这种岩溶驱动力的大小因植物形态特征、岩石性质与生存环境改变而发生变化.有研究表明, 碳酸盐岩溶蚀过程中, 生物成因的CO
2 占溶蚀强度的49.26%, 有机酸占37.11% (
张楷燕, 2017 ).苔藓植物溶蚀作用过程释放的矿物离子和其他元素部分被苔藓植物拦截形成岩溶沉积物; 部分被苔藓植物吸收利用, 转化为其他物质存储于植物体内; 其余则随降雨冲刷而流失, 进入其他生态系统.关于苔藓植物呼吸作用和其分泌物引起的岩溶作用驱动力、效率及其与苔藓植物生活型、种间差异、生境特征之间的相互关系仍未引起人们的关注, 对苔藓植物溶蚀作用过程和机理、苔藓植物对岩石中矿物营养元素释放规律的影响, 以及这些矿物质在岩溶沉积过程中的形态转化过程及其与生境间的相互作用关系等的研究还未见报道, 这也正是揭示苔藓植物岩溶作用机理的关键点.岩石的溶解速率与有机酸的酸性和溶蚀能力密切相关(
刘胜亮等, 2017 ), 然而有机酸的理化特性取决于其官能团的属性(
赵志曼等, 2018 ).因此, 今后需结合有机化学、气相色谱质谱(GC-MS)、核磁共振(NMR)、液相色谱(HPLC)等技术手段深入研究苔藓植物的溶蚀过程与机理. ...
贵州普定6种喀斯特石生植物及其土壤的碳酸酐酶活性
2
2017
... 此外, 苔藓植物体内的碳酸酐酶(CA)对岩溶作用也有促进作用(
Smith & Griffiths, 2000 ), 它主要催化CO
2 的水合反应: CO
2 + H
2 O $\rightleftharpoons $ HCO
3 - + H
+ (
吴雁雯和张金池, 2015 ).
李为等(2005) 研究发现无CA催化时上述可逆反应达到平衡的时间是有CA催化时的6 × 10
7 倍.生物体内均含有CA, 且CA活性因物种的不同而存在显著差异, 细菌、真菌、放线菌内均含有CA, 且微生物细胞内CA活性不同于细胞外CA, 细菌的CA对碳酸盐岩系统碳储存贡献在20%以上(
Li et al ., 2006 ), 真菌的CA对白云岩的溶蚀能力强于对石灰岩(
申泰铭等, 2014 ;
申泰铭, 2018 ).那么苔藓植物体内CA对岩溶作用的贡献是多少?各组织内的CA活性是否也存在这种差异?对此以及关于苔藓植物体内CA代谢的调节机制及原理仍不得而知.目前, 关于苔藓植物CA的研究, 主要集中在各类苔藓植物CA含量、活性及其对岩石溶蚀过程的促进作用等方面.例如,
刘天雷等(2017) 研究发现, 虽然苔藓植物都会分泌CA, 但CA分泌量、速率和活性存在种间差异.这种差异性可能是因生活型或者生长环境的不同而导致苔藓植物生理代谢与功能性状的差异引起的. ...
... 岩溶地区土壤的形成和发育是碳酸岩长期溶蚀、风化与生物富集过程的结果, 其一方面受岩石结构、性质影响, 另一方面也与植被作用密切相关(
张美良和邓自强, 1994 ).研究表明, 碳酸盐岩的成土速率与岩石溶蚀速率成正比(
曹建华等, 2004 ), 与酸不溶物含量密切相关(
曹沁智等, 2018 ).苔藓植物殖居于岩面, 假根系以及植物体的干湿交替冻融现象所产生的物理机械力作用于岩石, 产生岩石碎屑.同时, 有机物与矿物离子络合常常形成难溶物, 这些难溶物与苔藓及其生物结皮死亡残体黏结在一起, 不仅增加了土壤沉积量, 而且促进有机质积累, 增加土壤养分(
朱显谟, 1995 ;
刘天雷等, 2017 ).苔藓植物通过影响下垫面土壤湿度, 促进土壤对岩石的岩溶作用, 间接地影响沉积过程.同时, 苔藓植物的吸附拦截功能对防治岩石表面水土流失具有重要作用(
李军峰等, 2013 ;
贾少华等, 2014 ;
张显强等, 2018b ).苔藓植物的溶蚀作用为岩面土壤形成提供了原始物质, 关于苔藓对土壤沉积的贡献率与苔藓植物作用下岩面土壤的形成过程, 以及在此过程中岩石的物质、能量转化和土壤养分的变化等还未见报道. ...
贵州普定6种喀斯特石生植物及其土壤的碳酸酐酶活性
2
2017
... 此外, 苔藓植物体内的碳酸酐酶(CA)对岩溶作用也有促进作用(
Smith & Griffiths, 2000 ), 它主要催化CO
2 的水合反应: CO
2 + H
2 O $\rightleftharpoons $ HCO
3 - + H
+ (
吴雁雯和张金池, 2015 ).
李为等(2005) 研究发现无CA催化时上述可逆反应达到平衡的时间是有CA催化时的6 × 10
7 倍.生物体内均含有CA, 且CA活性因物种的不同而存在显著差异, 细菌、真菌、放线菌内均含有CA, 且微生物细胞内CA活性不同于细胞外CA, 细菌的CA对碳酸盐岩系统碳储存贡献在20%以上(
Li et al ., 2006 ), 真菌的CA对白云岩的溶蚀能力强于对石灰岩(
申泰铭等, 2014 ;
申泰铭, 2018 ).那么苔藓植物体内CA对岩溶作用的贡献是多少?各组织内的CA活性是否也存在这种差异?对此以及关于苔藓植物体内CA代谢的调节机制及原理仍不得而知.目前, 关于苔藓植物CA的研究, 主要集中在各类苔藓植物CA含量、活性及其对岩石溶蚀过程的促进作用等方面.例如,
刘天雷等(2017) 研究发现, 虽然苔藓植物都会分泌CA, 但CA分泌量、速率和活性存在种间差异.这种差异性可能是因生活型或者生长环境的不同而导致苔藓植物生理代谢与功能性状的差异引起的. ...
... 岩溶地区土壤的形成和发育是碳酸岩长期溶蚀、风化与生物富集过程的结果, 其一方面受岩石结构、性质影响, 另一方面也与植被作用密切相关(
张美良和邓自强, 1994 ).研究表明, 碳酸盐岩的成土速率与岩石溶蚀速率成正比(
曹建华等, 2004 ), 与酸不溶物含量密切相关(
曹沁智等, 2018 ).苔藓植物殖居于岩面, 假根系以及植物体的干湿交替冻融现象所产生的物理机械力作用于岩石, 产生岩石碎屑.同时, 有机物与矿物离子络合常常形成难溶物, 这些难溶物与苔藓及其生物结皮死亡残体黏结在一起, 不仅增加了土壤沉积量, 而且促进有机质积累, 增加土壤养分(
朱显谟, 1995 ;
刘天雷等, 2017 ).苔藓植物通过影响下垫面土壤湿度, 促进土壤对岩石的岩溶作用, 间接地影响沉积过程.同时, 苔藓植物的吸附拦截功能对防治岩石表面水土流失具有重要作用(
李军峰等, 2013 ;
贾少华等, 2014 ;
张显强等, 2018b ).苔藓植物的溶蚀作用为岩面土壤形成提供了原始物质, 关于苔藓对土壤沉积的贡献率与苔藓植物作用下岩面土壤的形成过程, 以及在此过程中岩石的物质、能量转化和土壤养分的变化等还未见报道. ...
重庆主城区地面苔藓植物群落特征及其与环境的关系
1
2015
... 苔藓植物群落分布特征与生境条件密切相关(
Heino & Virtanen, 2006 ;
刘艳等, 2015 ;
Mod et al ., 2016 ;
李泽科等, 2018 ;
庞嘉鹏等, 2018 ), 温度和水分是影响苔藓植物生存和分布的主要决定性因素, 尤其是空气湿度(
娄红祥, 2012 ;
Guo & Zhao, 2018 ).
党成强等(2018) 研究表明, 南亚毛灰藓(
Homomallium simlaense )在干旱和高温条件下, 各项生理指标均与体内相对含水量呈正比.苔藓植物在干旱缺水的情况下会停止生理代谢进入休眠状态, 当水分充足时再进行生命活动.而且苔藓植物生物量与相对生长速率随光照增强而增加, 随光照时间的延长, 其长度、pH值降低能力逐渐提高(
St. Martin & Mallik, 2017 ).苔藓植物虽然能够忍受高温, 但是温度高于25 ℃时却很少有净光合产物(
Proctor, 2000 ;
Hanson et al ., 2014 ).同时, 温度也是影响酶活性的关键因素, 温度超过60 ℃时会严重影响CA的活性(
洪子茜, 2018 ).然而, CA参与苔藓植物岩溶作用的最佳酶促反应温度还不得而知.同时, 温度也影响有机酸的反应性、络合能力和岩石中矿物自身的溶解程度(
闫志为等, 2009 ;
Benbouzid et al ., 2012 ), 进而影响岩石的溶蚀强度(
陈传平等, 2008 ).而且有机酸与矿物反应生成有机酸盐的存在形态也随温度发生变化(
Peters et al ., 2018 ). ...
重庆主城区地面苔藓植物群落特征及其与环境的关系
1
2015
... 苔藓植物群落分布特征与生境条件密切相关(
Heino & Virtanen, 2006 ;
刘艳等, 2015 ;
Mod et al ., 2016 ;
李泽科等, 2018 ;
庞嘉鹏等, 2018 ), 温度和水分是影响苔藓植物生存和分布的主要决定性因素, 尤其是空气湿度(
娄红祥, 2012 ;
Guo & Zhao, 2018 ).
党成强等(2018) 研究表明, 南亚毛灰藓(
Homomallium simlaense )在干旱和高温条件下, 各项生理指标均与体内相对含水量呈正比.苔藓植物在干旱缺水的情况下会停止生理代谢进入休眠状态, 当水分充足时再进行生命活动.而且苔藓植物生物量与相对生长速率随光照增强而增加, 随光照时间的延长, 其长度、pH值降低能力逐渐提高(
St. Martin & Mallik, 2017 ).苔藓植物虽然能够忍受高温, 但是温度高于25 ℃时却很少有净光合产物(
Proctor, 2000 ;
Hanson et al ., 2014 ).同时, 温度也是影响酶活性的关键因素, 温度超过60 ℃时会严重影响CA的活性(
洪子茜, 2018 ).然而, CA参与苔藓植物岩溶作用的最佳酶促反应温度还不得而知.同时, 温度也影响有机酸的反应性、络合能力和岩石中矿物自身的溶解程度(
闫志为等, 2009 ;
Benbouzid et al ., 2012 ), 进而影响岩石的溶蚀强度(
陈传平等, 2008 ).而且有机酸与矿物反应生成有机酸盐的存在形态也随温度发生变化(
Peters et al ., 2018 ). ...
Bryophyte Chemistry and Biology
4
2012
... 苔藓植物常以丛集型、交织型、匍匐型等着生于岩面(
图1 ), 假根根系不发达, 只起固着作用(
娄玉霞, 2013 ), 其特殊的叶表面结构和细胞特征(
娄红祥, 2012 ), 使苔藓具有耐高温、抗旱、蓄水能力强、保湿固土等功能(
Proctor & Pence, 2002 ;
Krol et al ., 2003 ;
Zhang et al ., 2017 ).
曹建华和袁道先(1999) 研究发现, 当岩石表面有苔藓植物时, 其蒸发失水和主动吸水时间比裸岩分别延长了48%和57%, 持水量提高了16.6倍, 分析认为苔藓植物可以延缓岩面水分蒸发时间, 增加岩石水化学作用时间, 但是苔藓植物的这种蓄水保湿能力存在明显的种间差异(
申家琛等, 2017 ;
张显强等, 2018a ).苔藓植物参与岩溶作用的另一大优势, 是其叶表面角质层不发育, 且阳离子交换位点多(
吴玉环等, 2001 ), 能够吸附多种阳离子, 这些阳离子在苔藓植物体内被转化为无机盐、有机盐以及其他形式的化合物储存于组织中.苔藓植物除了以生物化学方式对岩石进行溶蚀之外, 还可以通过物理机械作用破坏岩石.例如, 苔藓植物的假根可以伸入岩石裂隙, 对岩石有一定的破坏作用(
Jackson, 2015 ).再者, 当气候干旱时, 苔藓植物在卷缩过程中会拔起岩面的一些小颗粒, 从而加速了岩石被破坏和成土速度(
连宾等, 2008 ).此外, 苔藓植物常与藻类、地衣、微生物等共生形成生物结皮, 它们之间的相互关系以及因气候因素引起的干湿交替和冻融作用对岩石也具有一定的破坏性(
郑云普等, 2009 ).例如, 菌类的菌丝可以伸入岩石晶体结构中破坏岩石, 细菌与真菌形成的配合基能够络合三价铁离子, 细菌分泌的柠檬酸、乙酸以及地衣分泌的地衣酸(
连宾等, 2011 ), 可以溶解岩石中的矿物(
Won-Pyo et al ., 2016 ;
Wang et al ., 2017 ).因此在研究苔藓植物岩溶作用时, 应明确苔藓植物自身及其与其他生物形成的生物结皮两者之间的关系.目前, 有关岩面苔藓植物功能性状的研究主要集中在小范围内特定生境中的苔藓植物研究, 无法代表岩溶生态系统苔藓植物的整体情况.对于大尺度空间上, 因生境异质性高而造成相同物种在形态结构、光合特征及其钙(Ca)、碳(C)、氮(N)、磷(P) 等元素的代谢调节机制等方面的差异性的研究还未引起人们的广泛关注.岩溶生态系统的物质基础相对贫乏, 具有“缺氮少磷钙丰富”的特征.因此, 研究Ca、C、N、P等元素在苔藓植物体内的代谢机制对揭示岩溶生态系统苔藓植物和其他植物的生物多样性维持机制, 及其岩溶作用特征具有重要的科学 意义. ...
... 苔藓植物群落分布特征与生境条件密切相关(
Heino & Virtanen, 2006 ;
刘艳等, 2015 ;
Mod et al ., 2016 ;
李泽科等, 2018 ;
庞嘉鹏等, 2018 ), 温度和水分是影响苔藓植物生存和分布的主要决定性因素, 尤其是空气湿度(
娄红祥, 2012 ;
Guo & Zhao, 2018 ).
党成强等(2018) 研究表明, 南亚毛灰藓(
Homomallium simlaense )在干旱和高温条件下, 各项生理指标均与体内相对含水量呈正比.苔藓植物在干旱缺水的情况下会停止生理代谢进入休眠状态, 当水分充足时再进行生命活动.而且苔藓植物生物量与相对生长速率随光照增强而增加, 随光照时间的延长, 其长度、pH值降低能力逐渐提高(
St. Martin & Mallik, 2017 ).苔藓植物虽然能够忍受高温, 但是温度高于25 ℃时却很少有净光合产物(
Proctor, 2000 ;
Hanson et al ., 2014 ).同时, 温度也是影响酶活性的关键因素, 温度超过60 ℃时会严重影响CA的活性(
洪子茜, 2018 ).然而, CA参与苔藓植物岩溶作用的最佳酶促反应温度还不得而知.同时, 温度也影响有机酸的反应性、络合能力和岩石中矿物自身的溶解程度(
闫志为等, 2009 ;
Benbouzid et al ., 2012 ), 进而影响岩石的溶蚀强度(
陈传平等, 2008 ).而且有机酸与矿物反应生成有机酸盐的存在形态也随温度发生变化(
Peters et al ., 2018 ). ...
... 由于苔藓植物属于贫营养型, 它所需要的营养物质是维管束植物的1/10, 但是对N、P、K、Ca、Mg、Fe、硫(S)等元素的需求仍必不可少(
娄红祥, 2012 ).Ca
2+ 作为生物的第二信使, 具有调节生物抗逆性等多种功能, 对苔藓植物的生长有显著影响(
Amao et al ., 2010 ;
Kolupaev et al ., 2014 ).当苔藓植物缺Ca时会导致生长缓慢, 假根分化少, 原丝体不发育, 叶状体形状改变等(
娄红祥, 2012 ).而且CA活性也受Fe
2+ 、锌离子(Zn
2+ )、硝酸根(NO
- 3 )、氯离子(Cl
- )、溴离子(Br
- )、有机碳、交换性钙等的显著影响(
洪子茜, 2018 ).例如,
黄芬等(2015) 研究表明, Ca对CA的活性有很强的促进作用.生物与环境协同进化致使某些苔藓出现钙依赖性, 这种对钙的依赖性是否表现在基因水平, 从而影响岩溶地区苔藓植物生物多样性、空间分布特征、功能性状等还有待进一步研究.因此在研究苔藓植物岩溶作用与pH及营养元素之间的关系时, 应综合考虑苔藓植物种类、酶活性、逆境胁迫、水环境、基岩性质等因素. ...
... ).当苔藓植物缺Ca时会导致生长缓慢, 假根分化少, 原丝体不发育, 叶状体形状改变等(
娄红祥, 2012 ).而且CA活性也受Fe
2+ 、锌离子(Zn
2+ )、硝酸根(NO
- 3 )、氯离子(Cl
- )、溴离子(Br
- )、有机碳、交换性钙等的显著影响(
洪子茜, 2018 ).例如,
黄芬等(2015) 研究表明, Ca对CA的活性有很强的促进作用.生物与环境协同进化致使某些苔藓出现钙依赖性, 这种对钙的依赖性是否表现在基因水平, 从而影响岩溶地区苔藓植物生物多样性、空间分布特征、功能性状等还有待进一步研究.因此在研究苔藓植物岩溶作用与pH及营养元素之间的关系时, 应综合考虑苔藓植物种类、酶活性、逆境胁迫、水环境、基岩性质等因素. ...
Bryophyte Chemistry and Biology
4
2012
... 苔藓植物常以丛集型、交织型、匍匐型等着生于岩面(
图1 ), 假根根系不发达, 只起固着作用(
娄玉霞, 2013 ), 其特殊的叶表面结构和细胞特征(
娄红祥, 2012 ), 使苔藓具有耐高温、抗旱、蓄水能力强、保湿固土等功能(
Proctor & Pence, 2002 ;
Krol et al ., 2003 ;
Zhang et al ., 2017 ).
曹建华和袁道先(1999) 研究发现, 当岩石表面有苔藓植物时, 其蒸发失水和主动吸水时间比裸岩分别延长了48%和57%, 持水量提高了16.6倍, 分析认为苔藓植物可以延缓岩面水分蒸发时间, 增加岩石水化学作用时间, 但是苔藓植物的这种蓄水保湿能力存在明显的种间差异(
申家琛等, 2017 ;
张显强等, 2018a ).苔藓植物参与岩溶作用的另一大优势, 是其叶表面角质层不发育, 且阳离子交换位点多(
吴玉环等, 2001 ), 能够吸附多种阳离子, 这些阳离子在苔藓植物体内被转化为无机盐、有机盐以及其他形式的化合物储存于组织中.苔藓植物除了以生物化学方式对岩石进行溶蚀之外, 还可以通过物理机械作用破坏岩石.例如, 苔藓植物的假根可以伸入岩石裂隙, 对岩石有一定的破坏作用(
Jackson, 2015 ).再者, 当气候干旱时, 苔藓植物在卷缩过程中会拔起岩面的一些小颗粒, 从而加速了岩石被破坏和成土速度(
连宾等, 2008 ).此外, 苔藓植物常与藻类、地衣、微生物等共生形成生物结皮, 它们之间的相互关系以及因气候因素引起的干湿交替和冻融作用对岩石也具有一定的破坏性(
郑云普等, 2009 ).例如, 菌类的菌丝可以伸入岩石晶体结构中破坏岩石, 细菌与真菌形成的配合基能够络合三价铁离子, 细菌分泌的柠檬酸、乙酸以及地衣分泌的地衣酸(
连宾等, 2011 ), 可以溶解岩石中的矿物(
Won-Pyo et al ., 2016 ;
Wang et al ., 2017 ).因此在研究苔藓植物岩溶作用时, 应明确苔藓植物自身及其与其他生物形成的生物结皮两者之间的关系.目前, 有关岩面苔藓植物功能性状的研究主要集中在小范围内特定生境中的苔藓植物研究, 无法代表岩溶生态系统苔藓植物的整体情况.对于大尺度空间上, 因生境异质性高而造成相同物种在形态结构、光合特征及其钙(Ca)、碳(C)、氮(N)、磷(P) 等元素的代谢调节机制等方面的差异性的研究还未引起人们的广泛关注.岩溶生态系统的物质基础相对贫乏, 具有“缺氮少磷钙丰富”的特征.因此, 研究Ca、C、N、P等元素在苔藓植物体内的代谢机制对揭示岩溶生态系统苔藓植物和其他植物的生物多样性维持机制, 及其岩溶作用特征具有重要的科学 意义. ...
... 苔藓植物群落分布特征与生境条件密切相关(
Heino & Virtanen, 2006 ;
刘艳等, 2015 ;
Mod et al ., 2016 ;
李泽科等, 2018 ;
庞嘉鹏等, 2018 ), 温度和水分是影响苔藓植物生存和分布的主要决定性因素, 尤其是空气湿度(
娄红祥, 2012 ;
Guo & Zhao, 2018 ).
党成强等(2018) 研究表明, 南亚毛灰藓(
Homomallium simlaense )在干旱和高温条件下, 各项生理指标均与体内相对含水量呈正比.苔藓植物在干旱缺水的情况下会停止生理代谢进入休眠状态, 当水分充足时再进行生命活动.而且苔藓植物生物量与相对生长速率随光照增强而增加, 随光照时间的延长, 其长度、pH值降低能力逐渐提高(
St. Martin & Mallik, 2017 ).苔藓植物虽然能够忍受高温, 但是温度高于25 ℃时却很少有净光合产物(
Proctor, 2000 ;
Hanson et al ., 2014 ).同时, 温度也是影响酶活性的关键因素, 温度超过60 ℃时会严重影响CA的活性(
洪子茜, 2018 ).然而, CA参与苔藓植物岩溶作用的最佳酶促反应温度还不得而知.同时, 温度也影响有机酸的反应性、络合能力和岩石中矿物自身的溶解程度(
闫志为等, 2009 ;
Benbouzid et al ., 2012 ), 进而影响岩石的溶蚀强度(
陈传平等, 2008 ).而且有机酸与矿物反应生成有机酸盐的存在形态也随温度发生变化(
Peters et al ., 2018 ). ...
... 由于苔藓植物属于贫营养型, 它所需要的营养物质是维管束植物的1/10, 但是对N、P、K、Ca、Mg、Fe、硫(S)等元素的需求仍必不可少(
娄红祥, 2012 ).Ca
2+ 作为生物的第二信使, 具有调节生物抗逆性等多种功能, 对苔藓植物的生长有显著影响(
Amao et al ., 2010 ;
Kolupaev et al ., 2014 ).当苔藓植物缺Ca时会导致生长缓慢, 假根分化少, 原丝体不发育, 叶状体形状改变等(
娄红祥, 2012 ).而且CA活性也受Fe
2+ 、锌离子(Zn
2+ )、硝酸根(NO
- 3 )、氯离子(Cl
- )、溴离子(Br
- )、有机碳、交换性钙等的显著影响(
洪子茜, 2018 ).例如,
黄芬等(2015) 研究表明, Ca对CA的活性有很强的促进作用.生物与环境协同进化致使某些苔藓出现钙依赖性, 这种对钙的依赖性是否表现在基因水平, 从而影响岩溶地区苔藓植物生物多样性、空间分布特征、功能性状等还有待进一步研究.因此在研究苔藓植物岩溶作用与pH及营养元素之间的关系时, 应综合考虑苔藓植物种类、酶活性、逆境胁迫、水环境、基岩性质等因素. ...
... ).当苔藓植物缺Ca时会导致生长缓慢, 假根分化少, 原丝体不发育, 叶状体形状改变等(
娄红祥, 2012 ).而且CA活性也受Fe
2+ 、锌离子(Zn
2+ )、硝酸根(NO
- 3 )、氯离子(Cl
- )、溴离子(Br
- )、有机碳、交换性钙等的显著影响(
洪子茜, 2018 ).例如,
黄芬等(2015) 研究表明, Ca对CA的活性有很强的促进作用.生物与环境协同进化致使某些苔藓出现钙依赖性, 这种对钙的依赖性是否表现在基因水平, 从而影响岩溶地区苔藓植物生物多样性、空间分布特征、功能性状等还有待进一步研究.因此在研究苔藓植物岩溶作用与pH及营养元素之间的关系时, 应综合考虑苔藓植物种类、酶活性、逆境胁迫、水环境、基岩性质等因素. ...
苔藓植物对重金属污染的响应机理和生物指示的研究
1
2013
... 苔藓植物常以丛集型、交织型、匍匐型等着生于岩面(
图1 ), 假根根系不发达, 只起固着作用(
娄玉霞, 2013 ), 其特殊的叶表面结构和细胞特征(
娄红祥, 2012 ), 使苔藓具有耐高温、抗旱、蓄水能力强、保湿固土等功能(
Proctor & Pence, 2002 ;
Krol et al ., 2003 ;
Zhang et al ., 2017 ).
曹建华和袁道先(1999) 研究发现, 当岩石表面有苔藓植物时, 其蒸发失水和主动吸水时间比裸岩分别延长了48%和57%, 持水量提高了16.6倍, 分析认为苔藓植物可以延缓岩面水分蒸发时间, 增加岩石水化学作用时间, 但是苔藓植物的这种蓄水保湿能力存在明显的种间差异(
申家琛等, 2017 ;
张显强等, 2018a ).苔藓植物参与岩溶作用的另一大优势, 是其叶表面角质层不发育, 且阳离子交换位点多(
吴玉环等, 2001 ), 能够吸附多种阳离子, 这些阳离子在苔藓植物体内被转化为无机盐、有机盐以及其他形式的化合物储存于组织中.苔藓植物除了以生物化学方式对岩石进行溶蚀之外, 还可以通过物理机械作用破坏岩石.例如, 苔藓植物的假根可以伸入岩石裂隙, 对岩石有一定的破坏作用(
Jackson, 2015 ).再者, 当气候干旱时, 苔藓植物在卷缩过程中会拔起岩面的一些小颗粒, 从而加速了岩石被破坏和成土速度(
连宾等, 2008 ).此外, 苔藓植物常与藻类、地衣、微生物等共生形成生物结皮, 它们之间的相互关系以及因气候因素引起的干湿交替和冻融作用对岩石也具有一定的破坏性(
郑云普等, 2009 ).例如, 菌类的菌丝可以伸入岩石晶体结构中破坏岩石, 细菌与真菌形成的配合基能够络合三价铁离子, 细菌分泌的柠檬酸、乙酸以及地衣分泌的地衣酸(
连宾等, 2011 ), 可以溶解岩石中的矿物(
Won-Pyo et al ., 2016 ;
Wang et al ., 2017 ).因此在研究苔藓植物岩溶作用时, 应明确苔藓植物自身及其与其他生物形成的生物结皮两者之间的关系.目前, 有关岩面苔藓植物功能性状的研究主要集中在小范围内特定生境中的苔藓植物研究, 无法代表岩溶生态系统苔藓植物的整体情况.对于大尺度空间上, 因生境异质性高而造成相同物种在形态结构、光合特征及其钙(Ca)、碳(C)、氮(N)、磷(P) 等元素的代谢调节机制等方面的差异性的研究还未引起人们的广泛关注.岩溶生态系统的物质基础相对贫乏, 具有“缺氮少磷钙丰富”的特征.因此, 研究Ca、C、N、P等元素在苔藓植物体内的代谢机制对揭示岩溶生态系统苔藓植物和其他植物的生物多样性维持机制, 及其岩溶作用特征具有重要的科学 意义. ...
苔藓植物对重金属污染的响应机理和生物指示的研究
1
2013
... 苔藓植物常以丛集型、交织型、匍匐型等着生于岩面(
图1 ), 假根根系不发达, 只起固着作用(
娄玉霞, 2013 ), 其特殊的叶表面结构和细胞特征(
娄红祥, 2012 ), 使苔藓具有耐高温、抗旱、蓄水能力强、保湿固土等功能(
Proctor & Pence, 2002 ;
Krol et al ., 2003 ;
Zhang et al ., 2017 ).
曹建华和袁道先(1999) 研究发现, 当岩石表面有苔藓植物时, 其蒸发失水和主动吸水时间比裸岩分别延长了48%和57%, 持水量提高了16.6倍, 分析认为苔藓植物可以延缓岩面水分蒸发时间, 增加岩石水化学作用时间, 但是苔藓植物的这种蓄水保湿能力存在明显的种间差异(
申家琛等, 2017 ;
张显强等, 2018a ).苔藓植物参与岩溶作用的另一大优势, 是其叶表面角质层不发育, 且阳离子交换位点多(
吴玉环等, 2001 ), 能够吸附多种阳离子, 这些阳离子在苔藓植物体内被转化为无机盐、有机盐以及其他形式的化合物储存于组织中.苔藓植物除了以生物化学方式对岩石进行溶蚀之外, 还可以通过物理机械作用破坏岩石.例如, 苔藓植物的假根可以伸入岩石裂隙, 对岩石有一定的破坏作用(
Jackson, 2015 ).再者, 当气候干旱时, 苔藓植物在卷缩过程中会拔起岩面的一些小颗粒, 从而加速了岩石被破坏和成土速度(
连宾等, 2008 ).此外, 苔藓植物常与藻类、地衣、微生物等共生形成生物结皮, 它们之间的相互关系以及因气候因素引起的干湿交替和冻融作用对岩石也具有一定的破坏性(
郑云普等, 2009 ).例如, 菌类的菌丝可以伸入岩石晶体结构中破坏岩石, 细菌与真菌形成的配合基能够络合三价铁离子, 细菌分泌的柠檬酸、乙酸以及地衣分泌的地衣酸(
连宾等, 2011 ), 可以溶解岩石中的矿物(
Won-Pyo et al ., 2016 ;
Wang et al ., 2017 ).因此在研究苔藓植物岩溶作用时, 应明确苔藓植物自身及其与其他生物形成的生物结皮两者之间的关系.目前, 有关岩面苔藓植物功能性状的研究主要集中在小范围内特定生境中的苔藓植物研究, 无法代表岩溶生态系统苔藓植物的整体情况.对于大尺度空间上, 因生境异质性高而造成相同物种在形态结构、光合特征及其钙(Ca)、碳(C)、氮(N)、磷(P) 等元素的代谢调节机制等方面的差异性的研究还未引起人们的广泛关注.岩溶生态系统的物质基础相对贫乏, 具有“缺氮少磷钙丰富”的特征.因此, 研究Ca、C、N、P等元素在苔藓植物体内的代谢机制对揭示岩溶生态系统苔藓植物和其他植物的生物多样性维持机制, 及其岩溶作用特征具有重要的科学 意义. ...
The role of flow velocity in the vertical distribution of particulate organic matter on moss-covered travertine barriers of the Plitvice Lakes (Croatia)
1
2006
... 苔藓植物沉积作用的另一表现是石灰华或钙华(
张捷, 1993 ;
Kleinteich et al ., 2017 ).钙华形成的物理化学过程实质是CaCO
3 的沉淀与CO
2 的脱气.苔藓植物在钙华形成过程中为CaCO
3 晶体提供了稳定的生长发育基础, 并且苔藓植物的形态结构控制着沉积物的形态变化(
Mili?a et al ., 2006 ), 且它们的发育形态随沉积环境而变化(
张朝晖等, 2004 ;
郭云等, 2018 ).如在洞穴弱光带一般形成洞穴石刺、向光钟乳石、洞口叠层石等, 这类形态有很强的方向性(
王福星等, 1998 ).苔藓植物在水生环境中也常常形成多样的沉积形态, 如凤尾藓(
Fissidens )钙华、石灰藓(
Hydrogonium )钙华形成的溪流瀑布沉积, 泽藓(
Philonotis )钙华、水灰藓(
Hygrohypnum )钙华形成的河流瀑布沉积, 地钱-真藓(
Marchantia-Bryum )钙华等形成的石灰华坝沉积, 以及净口藓(
Gymnostomum )钙华等形成的瀑布洞穴沉积(
张朝晖和陈家宽, 2007 ).同时, 也有研究发现, 水流环境也是影响苔藓钙华沉积的重要因素, 苔藓与钙华层之间的水流速度与钙华沉积速率呈正相关关系(
Wang et al ., 2014 ).钙华作为大陆古气候的潜在档案,
Yan等(2017) 利用碳氧稳定同位素比率(
δ 13 C和
δ 18 O), 研究钙华在沉积过程中碳、氧的循环规律, 分析认为, 钙华沉积与季节更替引起的气温、降水等的变化密切相关.目前对于苔藓植物沉积作用的研究主要集中在沉积形态以及成土量方面, 对于苔藓植物沉积速率, 对岩溶地区成土过程中的贡献, 以及岩溶沉积作用对生境的改造和环境对岩溶沉积作用的影响等方面的研究还未见报道. ...
Contrasting effects of biotic interactions on richness and distribution of vascular plants, bryophytes and lichens in an arctic-alpine landscape
1
2016
... 苔藓植物群落分布特征与生境条件密切相关(
Heino & Virtanen, 2006 ;
刘艳等, 2015 ;
Mod et al ., 2016 ;
李泽科等, 2018 ;
庞嘉鹏等, 2018 ), 温度和水分是影响苔藓植物生存和分布的主要决定性因素, 尤其是空气湿度(
娄红祥, 2012 ;
Guo & Zhao, 2018 ).
党成强等(2018) 研究表明, 南亚毛灰藓(
Homomallium simlaense )在干旱和高温条件下, 各项生理指标均与体内相对含水量呈正比.苔藓植物在干旱缺水的情况下会停止生理代谢进入休眠状态, 当水分充足时再进行生命活动.而且苔藓植物生物量与相对生长速率随光照增强而增加, 随光照时间的延长, 其长度、pH值降低能力逐渐提高(
St. Martin & Mallik, 2017 ).苔藓植物虽然能够忍受高温, 但是温度高于25 ℃时却很少有净光合产物(
Proctor, 2000 ;
Hanson et al ., 2014 ).同时, 温度也是影响酶活性的关键因素, 温度超过60 ℃时会严重影响CA的活性(
洪子茜, 2018 ).然而, CA参与苔藓植物岩溶作用的最佳酶促反应温度还不得而知.同时, 温度也影响有机酸的反应性、络合能力和岩石中矿物自身的溶解程度(
闫志为等, 2009 ;
Benbouzid et al ., 2012 ), 进而影响岩石的溶蚀强度(
陈传平等, 2008 ).而且有机酸与矿物反应生成有机酸盐的存在形态也随温度发生变化(
Peters et al ., 2018 ). ...
The effect of organic acids on wettability of sandstone and carbonate rocks
1
2018
... 岩石中所含矿物种类及其晶体结构、产状、透水性等决定了岩石的溶解能力(
Voinot et al ., 2013 ;
黄奇波等, 2015 ), 例如, 一般情况下泥质含量高的碳酸盐岩溶蚀速度快, 岩石中酸不溶物含量越低, 越易被溶解(
王世杰等, 1999 ).同时, 岩石中酸不溶物含量对碳酸盐岩的成土速率具有重要影响, 酸不溶物含量越高岩石风化后可残留物越多, 成土物质越丰富(
陈洪松等, 2018 ).例如, 白云岩中酸不溶物含量高, 发育的土层较石灰岩地区土层厚(
陈洪松等, 2013 ).岩石中所含矿物的性质影响各元素的释放速率, Andrew等(2015)在25 ℃、pH = 2.6的开放试验系统与封闭试验系统内发现黑云母中的元素释放按照Fe > Mg > Al > Si的顺序进行, 且溶解速率符合 ${{r}_{i}}={{k}_{\text{H},i}}\times a_{{{\text{H}}^{\text{+}}}}^{xi}$,
k H 为表面速率常数((2.15-30.6) × 10
-10 mole s
biotite ·m
-2 ·s
-1 ),
a H+ 为H
+ 活性,
xi 为矿物与H
+ 的反应级数(0.31-0.58); 分析认为, 岩石中各矿物的溶解具有空间各向异性, 其边缘的反应效率强于基面, 而且各个元素的释放具有一定的规律性.苔藓植物种类的空间分布特征与岩性密切相关(
Gorozhankina & Konstantinov, 2001 ), 而且岩性和有机酸种类也会影响有机酸与岩石的反应速率(
Cama & Ganor, 2006 ;
Ghoorah et al ., 2014 ;
Mwangi et al ., 2018 ), 从而导致它们岩溶作用的差异性. ...
Influence of pH and temperature on the early stage of mica alteration
1
2012
...
Xie和Lou (2009) 认为, 生物岩溶作用随pH值降低而增强, 如果单纯从H
+ + CaCO
3 $\rightleftharpoons $ CO
2 + H
2 O + Ca
2+ 角度分析pH对岩溶作用的影响, pH值越低溶蚀速率越快.然而岩石的最佳溶解pH由矿物自身属性决定(
Bray et al ., 2015 ), 同时有机酸的溶蚀能力也受pH影响(
Pachana et al ., 2012 ;
Chen et al ., 2016 ), 而且每类苔藓植物的最适生长pH也不尽相同.如泥炭藓属(
Sphagnum )植物适宜偏酸性环境, 细叶小羽藓(
Haplocladium microphyllum )配子体的最适生长pH值为7 (
陈文佳等, 2012 ).作为苔藓植物岩溶作用的重要参与者CA, 它的活性也受pH影响,
洪子茜(2018) 研究表明, 当pH在7-9条件下CA的活性最高.pH由外而内对苔藓植物岩溶作用的过程、速率、对象、各级参与者都产生不同程度的促进或抑制作用.目前我们对pH影响苔藓植物岩溶的规律以及调节机制还不得而知, 今后还需综合全面地进行深入研究. ...
喀斯特白云岩石漠化区域不同生境条件下苔藓植物群落特征及演替模式研究
1
2018
... 苔藓植物群落分布特征与生境条件密切相关(
Heino & Virtanen, 2006 ;
刘艳等, 2015 ;
Mod et al ., 2016 ;
李泽科等, 2018 ;
庞嘉鹏等, 2018 ), 温度和水分是影响苔藓植物生存和分布的主要决定性因素, 尤其是空气湿度(
娄红祥, 2012 ;
Guo & Zhao, 2018 ).
党成强等(2018) 研究表明, 南亚毛灰藓(
Homomallium simlaense )在干旱和高温条件下, 各项生理指标均与体内相对含水量呈正比.苔藓植物在干旱缺水的情况下会停止生理代谢进入休眠状态, 当水分充足时再进行生命活动.而且苔藓植物生物量与相对生长速率随光照增强而增加, 随光照时间的延长, 其长度、pH值降低能力逐渐提高(
St. Martin & Mallik, 2017 ).苔藓植物虽然能够忍受高温, 但是温度高于25 ℃时却很少有净光合产物(
Proctor, 2000 ;
Hanson et al ., 2014 ).同时, 温度也是影响酶活性的关键因素, 温度超过60 ℃时会严重影响CA的活性(
洪子茜, 2018 ).然而, CA参与苔藓植物岩溶作用的最佳酶促反应温度还不得而知.同时, 温度也影响有机酸的反应性、络合能力和岩石中矿物自身的溶解程度(
闫志为等, 2009 ;
Benbouzid et al ., 2012 ), 进而影响岩石的溶蚀强度(
陈传平等, 2008 ).而且有机酸与矿物反应生成有机酸盐的存在形态也随温度发生变化(
Peters et al ., 2018 ). ...
喀斯特白云岩石漠化区域不同生境条件下苔藓植物群落特征及演替模式研究
1
2018
... 苔藓植物群落分布特征与生境条件密切相关(
Heino & Virtanen, 2006 ;
刘艳等, 2015 ;
Mod et al ., 2016 ;
李泽科等, 2018 ;
庞嘉鹏等, 2018 ), 温度和水分是影响苔藓植物生存和分布的主要决定性因素, 尤其是空气湿度(
娄红祥, 2012 ;
Guo & Zhao, 2018 ).
党成强等(2018) 研究表明, 南亚毛灰藓(
Homomallium simlaense )在干旱和高温条件下, 各项生理指标均与体内相对含水量呈正比.苔藓植物在干旱缺水的情况下会停止生理代谢进入休眠状态, 当水分充足时再进行生命活动.而且苔藓植物生物量与相对生长速率随光照增强而增加, 随光照时间的延长, 其长度、pH值降低能力逐渐提高(
St. Martin & Mallik, 2017 ).苔藓植物虽然能够忍受高温, 但是温度高于25 ℃时却很少有净光合产物(
Proctor, 2000 ;
Hanson et al ., 2014 ).同时, 温度也是影响酶活性的关键因素, 温度超过60 ℃时会严重影响CA的活性(
洪子茜, 2018 ).然而, CA参与苔藓植物岩溶作用的最佳酶促反应温度还不得而知.同时, 温度也影响有机酸的反应性、络合能力和岩石中矿物自身的溶解程度(
闫志为等, 2009 ;
Benbouzid et al ., 2012 ), 进而影响岩石的溶蚀强度(
陈传平等, 2008 ).而且有机酸与矿物反应生成有机酸盐的存在形态也随温度发生变化(
Peters et al ., 2018 ). ...
Seasonal variation of secondary metabolites in nine different bryophytes
1
2018
... 苔藓植物群落分布特征与生境条件密切相关(
Heino & Virtanen, 2006 ;
刘艳等, 2015 ;
Mod et al ., 2016 ;
李泽科等, 2018 ;
庞嘉鹏等, 2018 ), 温度和水分是影响苔藓植物生存和分布的主要决定性因素, 尤其是空气湿度(
娄红祥, 2012 ;
Guo & Zhao, 2018 ).
党成强等(2018) 研究表明, 南亚毛灰藓(
Homomallium simlaense )在干旱和高温条件下, 各项生理指标均与体内相对含水量呈正比.苔藓植物在干旱缺水的情况下会停止生理代谢进入休眠状态, 当水分充足时再进行生命活动.而且苔藓植物生物量与相对生长速率随光照增强而增加, 随光照时间的延长, 其长度、pH值降低能力逐渐提高(
St. Martin & Mallik, 2017 ).苔藓植物虽然能够忍受高温, 但是温度高于25 ℃时却很少有净光合产物(
Proctor, 2000 ;
Hanson et al ., 2014 ).同时, 温度也是影响酶活性的关键因素, 温度超过60 ℃时会严重影响CA的活性(
洪子茜, 2018 ).然而, CA参与苔藓植物岩溶作用的最佳酶促反应温度还不得而知.同时, 温度也影响有机酸的反应性、络合能力和岩石中矿物自身的溶解程度(
闫志为等, 2009 ;
Benbouzid et al ., 2012 ), 进而影响岩石的溶蚀强度(
陈传平等, 2008 ).而且有机酸与矿物反应生成有机酸盐的存在形态也随温度发生变化(
Peters et al ., 2018 ). ...
Bryophytes in a changing landscape: The hierarchical effects of habitat fragmentation on ecological and evolutionary processes
1
2006
... 因此, 湿度、温度、光照及其综合因子通过对苔藓植物生理代谢过程、基因型以及其他功能性状的影响(
Pharo & Zartman, 2006 ;
de la Rosa, 2016 ;
解梦, 2018 ), 而直接或间接地影响苔藓植物岩溶作用过程与效率.例如, 当温度在17.5-21.4 ℃, 湿度57.7%-72.6%, 光照2 700-5 900 lx条件下, 苔藓植物对碳酸盐岩的塑造作用最强(
申家琛等, 2018a ).但是目前对温湿度及其综合因子对苔藓植物岩溶作用的影响的关注度仍不够, 很多规律仍不清楚, 然而这些规律的发现是揭示苔藓植物岩溶机理的关键, 也是指导岩溶生态问题治理的重要科学依据. ...
The bryophyte paradox: Tolerance of desiccation, evasion of drought
1
2000
... 苔藓植物群落分布特征与生境条件密切相关(
Heino & Virtanen, 2006 ;
刘艳等, 2015 ;
Mod et al ., 2016 ;
李泽科等, 2018 ;
庞嘉鹏等, 2018 ), 温度和水分是影响苔藓植物生存和分布的主要决定性因素, 尤其是空气湿度(
娄红祥, 2012 ;
Guo & Zhao, 2018 ).
党成强等(2018) 研究表明, 南亚毛灰藓(
Homomallium simlaense )在干旱和高温条件下, 各项生理指标均与体内相对含水量呈正比.苔藓植物在干旱缺水的情况下会停止生理代谢进入休眠状态, 当水分充足时再进行生命活动.而且苔藓植物生物量与相对生长速率随光照增强而增加, 随光照时间的延长, 其长度、pH值降低能力逐渐提高(
St. Martin & Mallik, 2017 ).苔藓植物虽然能够忍受高温, 但是温度高于25 ℃时却很少有净光合产物(
Proctor, 2000 ;
Hanson et al ., 2014 ).同时, 温度也是影响酶活性的关键因素, 温度超过60 ℃时会严重影响CA的活性(
洪子茜, 2018 ).然而, CA参与苔藓植物岩溶作用的最佳酶促反应温度还不得而知.同时, 温度也影响有机酸的反应性、络合能力和岩石中矿物自身的溶解程度(
闫志为等, 2009 ;
Benbouzid et al ., 2012 ), 进而影响岩石的溶蚀强度(
陈传平等, 2008 ).而且有机酸与矿物反应生成有机酸盐的存在形态也随温度发生变化(
Peters et al ., 2018 ). ...
Vegetative tissues: Bryophytes vascular “resurrection plants” and vegetative propagules. In: Pritchard H, Back M eds. Desiccation and Plant Survival
2002
Biological and inorganic factors in the destruction of limestone coasts
1
1976
... 生物岩溶是指由生物引起的岩溶作用现象及 其产物, 即生物溶蚀、破坏可溶岩和沉淀形成次生化学沉积物及其所遗留或形成的形态(
Schneider, 1976 ).生物岩溶作用(溶蚀作用和沉积作用)激活了岩溶生态系统基质中的物质和能量, 使各个圈层间信息交流更加频繁与复杂.苔藓植物作为生态系统演替过程中的先锋植物(
薛建辉, 2013 ), 通过生物岩溶作用改善裸岩生境, 为草本和灌木的出现奠定了物质基础(
Harper & Belnap, 2001 ;
Jackson, 2015 ), 对岩溶生态系统的维持与发展具有重要意义(
Li et al ., 2006 ;
Costa et al ., 2018 ).苔藓植物岩溶作用已成为岩溶生态学研究的热点.因此, 了解和掌握岩面苔藓植物功能性状特征、岩溶作用机理及其与环境间的互作机制对研究岩溶地区的生物多样性、石漠化生态恢复、岩溶碳汇以及丰富生物岩溶理论等意义重大.然而, 苔藓植物岩溶作用研究因涉及学科众多、问题复杂, 还未进入系统化研究阶段.为了破除各学科间的壁垒, 促进苔藓植物岩溶作用研究更进一步发展, 本文根据近年来有关苔藓植物岩溶作用的文献资料, 从以下方面综述苔藓植物岩溶作用的研究进展. ...
贵阳喀斯特公园南石林秋季藓类植物的持水特性
1
2017
... 苔藓植物常以丛集型、交织型、匍匐型等着生于岩面(
图1 ), 假根根系不发达, 只起固着作用(
娄玉霞, 2013 ), 其特殊的叶表面结构和细胞特征(
娄红祥, 2012 ), 使苔藓具有耐高温、抗旱、蓄水能力强、保湿固土等功能(
Proctor & Pence, 2002 ;
Krol et al ., 2003 ;
Zhang et al ., 2017 ).
曹建华和袁道先(1999) 研究发现, 当岩石表面有苔藓植物时, 其蒸发失水和主动吸水时间比裸岩分别延长了48%和57%, 持水量提高了16.6倍, 分析认为苔藓植物可以延缓岩面水分蒸发时间, 增加岩石水化学作用时间, 但是苔藓植物的这种蓄水保湿能力存在明显的种间差异(
申家琛等, 2017 ;
张显强等, 2018a ).苔藓植物参与岩溶作用的另一大优势, 是其叶表面角质层不发育, 且阳离子交换位点多(
吴玉环等, 2001 ), 能够吸附多种阳离子, 这些阳离子在苔藓植物体内被转化为无机盐、有机盐以及其他形式的化合物储存于组织中.苔藓植物除了以生物化学方式对岩石进行溶蚀之外, 还可以通过物理机械作用破坏岩石.例如, 苔藓植物的假根可以伸入岩石裂隙, 对岩石有一定的破坏作用(
Jackson, 2015 ).再者, 当气候干旱时, 苔藓植物在卷缩过程中会拔起岩面的一些小颗粒, 从而加速了岩石被破坏和成土速度(
连宾等, 2008 ).此外, 苔藓植物常与藻类、地衣、微生物等共生形成生物结皮, 它们之间的相互关系以及因气候因素引起的干湿交替和冻融作用对岩石也具有一定的破坏性(
郑云普等, 2009 ).例如, 菌类的菌丝可以伸入岩石晶体结构中破坏岩石, 细菌与真菌形成的配合基能够络合三价铁离子, 细菌分泌的柠檬酸、乙酸以及地衣分泌的地衣酸(
连宾等, 2011 ), 可以溶解岩石中的矿物(
Won-Pyo et al ., 2016 ;
Wang et al ., 2017 ).因此在研究苔藓植物岩溶作用时, 应明确苔藓植物自身及其与其他生物形成的生物结皮两者之间的关系.目前, 有关岩面苔藓植物功能性状的研究主要集中在小范围内特定生境中的苔藓植物研究, 无法代表岩溶生态系统苔藓植物的整体情况.对于大尺度空间上, 因生境异质性高而造成相同物种在形态结构、光合特征及其钙(Ca)、碳(C)、氮(N)、磷(P) 等元素的代谢调节机制等方面的差异性的研究还未引起人们的广泛关注.岩溶生态系统的物质基础相对贫乏, 具有“缺氮少磷钙丰富”的特征.因此, 研究Ca、C、N、P等元素在苔藓植物体内的代谢机制对揭示岩溶生态系统苔藓植物和其他植物的生物多样性维持机制, 及其岩溶作用特征具有重要的科学 意义. ...
贵阳喀斯特公园南石林秋季藓类植物的持水特性
1
2017
... 苔藓植物常以丛集型、交织型、匍匐型等着生于岩面(
图1 ), 假根根系不发达, 只起固着作用(
娄玉霞, 2013 ), 其特殊的叶表面结构和细胞特征(
娄红祥, 2012 ), 使苔藓具有耐高温、抗旱、蓄水能力强、保湿固土等功能(
Proctor & Pence, 2002 ;
Krol et al ., 2003 ;
Zhang et al ., 2017 ).
曹建华和袁道先(1999) 研究发现, 当岩石表面有苔藓植物时, 其蒸发失水和主动吸水时间比裸岩分别延长了48%和57%, 持水量提高了16.6倍, 分析认为苔藓植物可以延缓岩面水分蒸发时间, 增加岩石水化学作用时间, 但是苔藓植物的这种蓄水保湿能力存在明显的种间差异(
申家琛等, 2017 ;
张显强等, 2018a ).苔藓植物参与岩溶作用的另一大优势, 是其叶表面角质层不发育, 且阳离子交换位点多(
吴玉环等, 2001 ), 能够吸附多种阳离子, 这些阳离子在苔藓植物体内被转化为无机盐、有机盐以及其他形式的化合物储存于组织中.苔藓植物除了以生物化学方式对岩石进行溶蚀之外, 还可以通过物理机械作用破坏岩石.例如, 苔藓植物的假根可以伸入岩石裂隙, 对岩石有一定的破坏作用(
Jackson, 2015 ).再者, 当气候干旱时, 苔藓植物在卷缩过程中会拔起岩面的一些小颗粒, 从而加速了岩石被破坏和成土速度(
连宾等, 2008 ).此外, 苔藓植物常与藻类、地衣、微生物等共生形成生物结皮, 它们之间的相互关系以及因气候因素引起的干湿交替和冻融作用对岩石也具有一定的破坏性(
郑云普等, 2009 ).例如, 菌类的菌丝可以伸入岩石晶体结构中破坏岩石, 细菌与真菌形成的配合基能够络合三价铁离子, 细菌分泌的柠檬酸、乙酸以及地衣分泌的地衣酸(
连宾等, 2011 ), 可以溶解岩石中的矿物(
Won-Pyo et al ., 2016 ;
Wang et al ., 2017 ).因此在研究苔藓植物岩溶作用时, 应明确苔藓植物自身及其与其他生物形成的生物结皮两者之间的关系.目前, 有关岩面苔藓植物功能性状的研究主要集中在小范围内特定生境中的苔藓植物研究, 无法代表岩溶生态系统苔藓植物的整体情况.对于大尺度空间上, 因生境异质性高而造成相同物种在形态结构、光合特征及其钙(Ca)、碳(C)、氮(N)、磷(P) 等元素的代谢调节机制等方面的差异性的研究还未引起人们的广泛关注.岩溶生态系统的物质基础相对贫乏, 具有“缺氮少磷钙丰富”的特征.因此, 研究Ca、C、N、P等元素在苔藓植物体内的代谢机制对揭示岩溶生态系统苔藓植物和其他植物的生物多样性维持机制, 及其岩溶作用特征具有重要的科学 意义. ...
苔藓植物对石灰岩的溶蚀作用及环境相关性研究
2
2018a
... 岩石经苔藓植物溶蚀后, 表面常常出现一些凹凸不平和微溶孔, 这是由于岩石成分不均匀, 导致生物钻孔力的差异性(
王福星等, 1998 ), 以及溶蚀产物流失而形成溶蚀微地貌.
王福星等(1993) 将这些溶蚀微地貌, 按照口面形态、直径大小、深度等, 把短径小于1 mm的命名为微溶孔, 把短径为1-20 mm的命名为溶孔, 把短径为20-100 mm的命名为溶坑.
付兰和张朝晖(2010) 将连续分布的苔藓植物溶蚀微地貌, 按测算结果, 把长7.7-14.4 cm的命名为溶丝; 长0.8-19.0 cm, 宽0.6-7.2 cm的命名为溶斑; 长6.0-24.5 cm, 宽1.2-2.2 cm的命名为溶带.苔藓植物溶蚀过程很难通过宏观手段观测, 导致这两种命名法在大小和形态方面存在明显差异.然而,
Haward等(2011) 借助原子力显微镜(AFM)观察到黑云母表面浅层溶蚀坑的形成过程, 通过分析图像计算质量损失, 从而确定溶蚀过程中黑云母表面的活化能大小.研究表明, 通过分析活化能得到的溶坑大小与测量其周长所得的数据一致.因此, 在研究溶蚀微地貌时, 应采用高精度仪器, 统一测量标准, 分析微地貌是苔藓植物溶蚀作用产生, 还是由于淋溶风化而形成, 有利于后期岩溶微地貌之间的对比分析和进一步深入研究.这些微地貌的形态特征与生物类型和发育生境密切相关(
蔡胡霖, 2017 ), 在温湿度优越的地区发育更为广泛(
Liang & Xu, 2014 ;
Liu et al ., 2016 ).例如, 溶孔、溶锥多发生在低湿度、强光照、高温环境中, 而溶沟多是在高温、强光和pH较高的生境内发育(
申家琛等, 2018a ).
曹建华和王福星(1996) 认为苔藓植物岩溶微地貌与岩溶地区的宏观地貌形态之间存在一定的相关性, 然而目前这种关联性以及苔藓植物微地貌之间的关系和发育方向尚未得到科学揭示. ...
... 因此, 湿度、温度、光照及其综合因子通过对苔藓植物生理代谢过程、基因型以及其他功能性状的影响(
Pharo & Zartman, 2006 ;
de la Rosa, 2016 ;
解梦, 2018 ), 而直接或间接地影响苔藓植物岩溶作用过程与效率.例如, 当温度在17.5-21.4 ℃, 湿度57.7%-72.6%, 光照2 700-5 900 lx条件下, 苔藓植物对碳酸盐岩的塑造作用最强(
申家琛等, 2018a ).但是目前对温湿度及其综合因子对苔藓植物岩溶作用的影响的关注度仍不够, 很多规律仍不清楚, 然而这些规律的发现是揭示苔藓植物岩溶机理的关键, 也是指导岩溶生态问题治理的重要科学依据. ...
苔藓植物对石灰岩的溶蚀作用及环境相关性研究
2
2018a
... 岩石经苔藓植物溶蚀后, 表面常常出现一些凹凸不平和微溶孔, 这是由于岩石成分不均匀, 导致生物钻孔力的差异性(
王福星等, 1998 ), 以及溶蚀产物流失而形成溶蚀微地貌.
王福星等(1993) 将这些溶蚀微地貌, 按照口面形态、直径大小、深度等, 把短径小于1 mm的命名为微溶孔, 把短径为1-20 mm的命名为溶孔, 把短径为20-100 mm的命名为溶坑.
付兰和张朝晖(2010) 将连续分布的苔藓植物溶蚀微地貌, 按测算结果, 把长7.7-14.4 cm的命名为溶丝; 长0.8-19.0 cm, 宽0.6-7.2 cm的命名为溶斑; 长6.0-24.5 cm, 宽1.2-2.2 cm的命名为溶带.苔藓植物溶蚀过程很难通过宏观手段观测, 导致这两种命名法在大小和形态方面存在明显差异.然而,
Haward等(2011) 借助原子力显微镜(AFM)观察到黑云母表面浅层溶蚀坑的形成过程, 通过分析图像计算质量损失, 从而确定溶蚀过程中黑云母表面的活化能大小.研究表明, 通过分析活化能得到的溶坑大小与测量其周长所得的数据一致.因此, 在研究溶蚀微地貌时, 应采用高精度仪器, 统一测量标准, 分析微地貌是苔藓植物溶蚀作用产生, 还是由于淋溶风化而形成, 有利于后期岩溶微地貌之间的对比分析和进一步深入研究.这些微地貌的形态特征与生物类型和发育生境密切相关(
蔡胡霖, 2017 ), 在温湿度优越的地区发育更为广泛(
Liang & Xu, 2014 ;
Liu et al ., 2016 ).例如, 溶孔、溶锥多发生在低湿度、强光照、高温环境中, 而溶沟多是在高温、强光和pH较高的生境内发育(
申家琛等, 2018a ).
曹建华和王福星(1996) 认为苔藓植物岩溶微地貌与岩溶地区的宏观地貌形态之间存在一定的相关性, 然而目前这种关联性以及苔藓植物微地貌之间的关系和发育方向尚未得到科学揭示. ...
... 因此, 湿度、温度、光照及其综合因子通过对苔藓植物生理代谢过程、基因型以及其他功能性状的影响(
Pharo & Zartman, 2006 ;
de la Rosa, 2016 ;
解梦, 2018 ), 而直接或间接地影响苔藓植物岩溶作用过程与效率.例如, 当温度在17.5-21.4 ℃, 湿度57.7%-72.6%, 光照2 700-5 900 lx条件下, 苔藓植物对碳酸盐岩的塑造作用最强(
申家琛等, 2018a ).但是目前对温湿度及其综合因子对苔藓植物岩溶作用的影响的关注度仍不够, 很多规律仍不清楚, 然而这些规律的发现是揭示苔藓植物岩溶机理的关键, 也是指导岩溶生态问题治理的重要科学依据. ...
石漠化程度对苔藓植物多样性及其结皮土壤化学性质的影响
1
2018b
... 苔藓植物岩溶作用与环境之间的三向循环互作机制如
图3 所示, 苔藓、环境因子、岩石三者之间相互联系、相互作用.
申家琛等(2018b) 阐释了苔藓植物溶蚀微地貌发育及其分布特征与生境(温湿度、光照等)的关系, 但是对苔藓植物岩溶作用对生境的改造, 以及生态因子对苔藓植物岩溶作用的影响还未揭示.因此, 今后需要在这方面开展以下研究工作: (1)温度、湿度、光照、pH、地质背景等因子对苔藓植物生物多样性、生存繁殖, 生理调节机制、功能性状等的影响; (2)环境因子对苔藓植物溶蚀过程、效率、物质能量转化过程等的影响; (3)苔藓植物沉积过程对岩面微环境的温湿度、地貌塑造、土壤养分、水动力特征等的影响; (4)苔藓植物与其内生菌和藻类的共生关系以及与其他植物之间的化感作用对岩溶作用的影响. ...
石漠化程度对苔藓植物多样性及其结皮土壤化学性质的影响
1
2018b
... 苔藓植物岩溶作用与环境之间的三向循环互作机制如
图3 所示, 苔藓、环境因子、岩石三者之间相互联系、相互作用.
申家琛等(2018b) 阐释了苔藓植物溶蚀微地貌发育及其分布特征与生境(温湿度、光照等)的关系, 但是对苔藓植物岩溶作用对生境的改造, 以及生态因子对苔藓植物岩溶作用的影响还未揭示.因此, 今后需要在这方面开展以下研究工作: (1)温度、湿度、光照、pH、地质背景等因子对苔藓植物生物多样性、生存繁殖, 生理调节机制、功能性状等的影响; (2)环境因子对苔藓植物溶蚀过程、效率、物质能量转化过程等的影响; (3)苔藓植物沉积过程对岩面微环境的温湿度、地貌塑造、土壤养分、水动力特征等的影响; (4)苔藓植物与其内生菌和藻类的共生关系以及与其他植物之间的化感作用对岩溶作用的影响. ...
微生物及碳酸酐酶对岩溶土壤生态系统碳储存的贡献及其机制
2018
微生物及碳酸酐酶对岩溶土壤生态系统碳储存的贡献及其机制
2018
不同种类微生物及其碳酸酐酶对CO
2 -H
2 O-碳酸盐系统中碳酸盐岩的溶蚀作用
1
2014
... 此外, 苔藓植物体内的碳酸酐酶(CA)对岩溶作用也有促进作用(
Smith & Griffiths, 2000 ), 它主要催化CO
2 的水合反应: CO
2 + H
2 O $\rightleftharpoons $ HCO
3 - + H
+ (
吴雁雯和张金池, 2015 ).
李为等(2005) 研究发现无CA催化时上述可逆反应达到平衡的时间是有CA催化时的6 × 10
7 倍.生物体内均含有CA, 且CA活性因物种的不同而存在显著差异, 细菌、真菌、放线菌内均含有CA, 且微生物细胞内CA活性不同于细胞外CA, 细菌的CA对碳酸盐岩系统碳储存贡献在20%以上(
Li et al ., 2006 ), 真菌的CA对白云岩的溶蚀能力强于对石灰岩(
申泰铭等, 2014 ;
申泰铭, 2018 ).那么苔藓植物体内CA对岩溶作用的贡献是多少?各组织内的CA活性是否也存在这种差异?对此以及关于苔藓植物体内CA代谢的调节机制及原理仍不得而知.目前, 关于苔藓植物CA的研究, 主要集中在各类苔藓植物CA含量、活性及其对岩石溶蚀过程的促进作用等方面.例如,
刘天雷等(2017) 研究发现, 虽然苔藓植物都会分泌CA, 但CA分泌量、速率和活性存在种间差异.这种差异性可能是因生活型或者生长环境的不同而导致苔藓植物生理代谢与功能性状的差异引起的. ...
不同种类微生物及其碳酸酐酶对CO
2 -H
2 O-碳酸盐系统中碳酸盐岩的溶蚀作用
1
2014
... 此外, 苔藓植物体内的碳酸酐酶(CA)对岩溶作用也有促进作用(
Smith & Griffiths, 2000 ), 它主要催化CO
2 的水合反应: CO
2 + H
2 O $\rightleftharpoons $ HCO
3 - + H
+ (
吴雁雯和张金池, 2015 ).
李为等(2005) 研究发现无CA催化时上述可逆反应达到平衡的时间是有CA催化时的6 × 10
7 倍.生物体内均含有CA, 且CA活性因物种的不同而存在显著差异, 细菌、真菌、放线菌内均含有CA, 且微生物细胞内CA活性不同于细胞外CA, 细菌的CA对碳酸盐岩系统碳储存贡献在20%以上(
Li et al ., 2006 ), 真菌的CA对白云岩的溶蚀能力强于对石灰岩(
申泰铭等, 2014 ;
申泰铭, 2018 ).那么苔藓植物体内CA对岩溶作用的贡献是多少?各组织内的CA活性是否也存在这种差异?对此以及关于苔藓植物体内CA代谢的调节机制及原理仍不得而知.目前, 关于苔藓植物CA的研究, 主要集中在各类苔藓植物CA含量、活性及其对岩石溶蚀过程的促进作用等方面.例如,
刘天雷等(2017) 研究发现, 虽然苔藓植物都会分泌CA, 但CA分泌量、速率和活性存在种间差异.这种差异性可能是因生活型或者生长环境的不同而导致苔藓植物生理代谢与功能性状的差异引起的. ...
The role of carbonic anhydrase in photosynthesis and the activity of the carbon- concentrating-mechanism in bryophytes of the class anthocerotae
1
2000
... 此外, 苔藓植物体内的碳酸酐酶(CA)对岩溶作用也有促进作用(
Smith & Griffiths, 2000 ), 它主要催化CO
2 的水合反应: CO
2 + H
2 O $\rightleftharpoons $ HCO
3 - + H
+ (
吴雁雯和张金池, 2015 ).
李为等(2005) 研究发现无CA催化时上述可逆反应达到平衡的时间是有CA催化时的6 × 10
7 倍.生物体内均含有CA, 且CA活性因物种的不同而存在显著差异, 细菌、真菌、放线菌内均含有CA, 且微生物细胞内CA活性不同于细胞外CA, 细菌的CA对碳酸盐岩系统碳储存贡献在20%以上(
Li et al ., 2006 ), 真菌的CA对白云岩的溶蚀能力强于对石灰岩(
申泰铭等, 2014 ;
申泰铭, 2018 ).那么苔藓植物体内CA对岩溶作用的贡献是多少?各组织内的CA活性是否也存在这种差异?对此以及关于苔藓植物体内CA代谢的调节机制及原理仍不得而知.目前, 关于苔藓植物CA的研究, 主要集中在各类苔藓植物CA含量、活性及其对岩石溶蚀过程的促进作用等方面.例如,
刘天雷等(2017) 研究发现, 虽然苔藓植物都会分泌CA, 但CA分泌量、速率和活性存在种间差异.这种差异性可能是因生活型或者生长环境的不同而导致苔藓植物生理代谢与功能性状的差异引起的. ...
The status of non-vascular plants in trait-based ecosystem function studies
1
2017
... 苔藓植物群落分布特征与生境条件密切相关(
Heino & Virtanen, 2006 ;
刘艳等, 2015 ;
Mod et al ., 2016 ;
李泽科等, 2018 ;
庞嘉鹏等, 2018 ), 温度和水分是影响苔藓植物生存和分布的主要决定性因素, 尤其是空气湿度(
娄红祥, 2012 ;
Guo & Zhao, 2018 ).
党成强等(2018) 研究表明, 南亚毛灰藓(
Homomallium simlaense )在干旱和高温条件下, 各项生理指标均与体内相对含水量呈正比.苔藓植物在干旱缺水的情况下会停止生理代谢进入休眠状态, 当水分充足时再进行生命活动.而且苔藓植物生物量与相对生长速率随光照增强而增加, 随光照时间的延长, 其长度、pH值降低能力逐渐提高(
St. Martin & Mallik, 2017 ).苔藓植物虽然能够忍受高温, 但是温度高于25 ℃时却很少有净光合产物(
Proctor, 2000 ;
Hanson et al ., 2014 ).同时, 温度也是影响酶活性的关键因素, 温度超过60 ℃时会严重影响CA的活性(
洪子茜, 2018 ).然而, CA参与苔藓植物岩溶作用的最佳酶促反应温度还不得而知.同时, 温度也影响有机酸的反应性、络合能力和岩石中矿物自身的溶解程度(
闫志为等, 2009 ;
Benbouzid et al ., 2012 ), 进而影响岩石的溶蚀强度(
陈传平等, 2008 ).而且有机酸与矿物反应生成有机酸盐的存在形态也随温度发生变化(
Peters et al ., 2018 ). ...
The role of mosses in carbon uptake and partitioning in arctic vegetation
1
2013
... 苔藓植物对岩石的溶蚀作用原理与过程实质是岩石中矿物与苔藓植物之间的作用过程(
图2 ).苔藓植物因呼吸作用形成的H
2 CO
3 和代谢分泌物与岩石中的矿物发生酸化、碱化、氧化还原、螯合等反应, 随着反应过程不断推进, 岩石矿物成分发生变化, Ca、镁(Mg)、钠(Na)、钾(K)、铁(Fe)、铝(Al)、硅(Si)等从碳酸盐、硅酸盐、铝酸盐、硫化物、氧化物中溶出, 破坏原岩晶体结构, 达到溶蚀岩石, 促使风化的目的(
Street et al ., 2013 ;
Li et al ., 2015 ;
Lammers et al ., 2017 ).这种岩溶驱动力的大小因植物形态特征、岩石性质与生存环境改变而发生变化.有研究表明, 碳酸盐岩溶蚀过程中, 生物成因的CO
2 占溶蚀强度的49.26%, 有机酸占37.11% (
张楷燕, 2017 ).苔藓植物溶蚀作用过程释放的矿物离子和其他元素部分被苔藓植物拦截形成岩溶沉积物; 部分被苔藓植物吸收利用, 转化为其他物质存储于植物体内; 其余则随降雨冲刷而流失, 进入其他生态系统.关于苔藓植物呼吸作用和其分泌物引起的岩溶作用驱动力、效率及其与苔藓植物生活型、种间差异、生境特征之间的相互关系仍未引起人们的关注, 对苔藓植物溶蚀作用过程和机理、苔藓植物对岩石中矿物营养元素释放规律的影响, 以及这些矿物质在岩溶沉积过程中的形态转化过程及其与生境间的相互作用关系等的研究还未见报道, 这也正是揭示苔藓植物岩溶作用机理的关键点.岩石的溶解速率与有机酸的酸性和溶蚀能力密切相关(
刘胜亮等, 2017 ), 然而有机酸的理化特性取决于其官能团的属性(
赵志曼等, 2018 ).因此, 今后需结合有机化学、气相色谱质谱(GC-MS)、核磁共振(NMR)、液相色谱(HPLC)等技术手段深入研究苔藓植物的溶蚀过程与机理. ...
Experimental dissolution vs. transformation of micas under acidic soil conditions: Clues from boron isotopes
1
2013
... 岩石中所含矿物种类及其晶体结构、产状、透水性等决定了岩石的溶解能力(
Voinot et al ., 2013 ;
黄奇波等, 2015 ), 例如, 一般情况下泥质含量高的碳酸盐岩溶蚀速度快, 岩石中酸不溶物含量越低, 越易被溶解(
王世杰等, 1999 ).同时, 岩石中酸不溶物含量对碳酸盐岩的成土速率具有重要影响, 酸不溶物含量越高岩石风化后可残留物越多, 成土物质越丰富(
陈洪松等, 2018 ).例如, 白云岩中酸不溶物含量高, 发育的土层较石灰岩地区土层厚(
陈洪松等, 2013 ).岩石中所含矿物的性质影响各元素的释放速率, Andrew等(2015)在25 ℃、pH = 2.6的开放试验系统与封闭试验系统内发现黑云母中的元素释放按照Fe > Mg > Al > Si的顺序进行, 且溶解速率符合 ${{r}_{i}}={{k}_{\text{H},i}}\times a_{{{\text{H}}^{\text{+}}}}^{xi}$,
k H 为表面速率常数((2.15-30.6) × 10
-10 mole s
biotite ·m
-2 ·s
-1 ),
a H+ 为H
+ 活性,
xi 为矿物与H
+ 的反应级数(0.31-0.58); 分析认为, 岩石中各矿物的溶解具有空间各向异性, 其边缘的反应效率强于基面, 而且各个元素的释放具有一定的规律性.苔藓植物种类的空间分布特征与岩性密切相关(
Gorozhankina & Konstantinov, 2001 ), 而且岩性和有机酸种类也会影响有机酸与岩石的反应速率(
Cama & Ganor, 2006 ;
Ghoorah et al ., 2014 ;
Mwangi et al ., 2018 ), 从而导致它们岩溶作用的差异性. ...
洞穴弱光带的生物岩溶
2
1998
... 岩石经苔藓植物溶蚀后, 表面常常出现一些凹凸不平和微溶孔, 这是由于岩石成分不均匀, 导致生物钻孔力的差异性(
王福星等, 1998 ), 以及溶蚀产物流失而形成溶蚀微地貌.
王福星等(1993) 将这些溶蚀微地貌, 按照口面形态、直径大小、深度等, 把短径小于1 mm的命名为微溶孔, 把短径为1-20 mm的命名为溶孔, 把短径为20-100 mm的命名为溶坑.
付兰和张朝晖(2010) 将连续分布的苔藓植物溶蚀微地貌, 按测算结果, 把长7.7-14.4 cm的命名为溶丝; 长0.8-19.0 cm, 宽0.6-7.2 cm的命名为溶斑; 长6.0-24.5 cm, 宽1.2-2.2 cm的命名为溶带.苔藓植物溶蚀过程很难通过宏观手段观测, 导致这两种命名法在大小和形态方面存在明显差异.然而,
Haward等(2011) 借助原子力显微镜(AFM)观察到黑云母表面浅层溶蚀坑的形成过程, 通过分析图像计算质量损失, 从而确定溶蚀过程中黑云母表面的活化能大小.研究表明, 通过分析活化能得到的溶坑大小与测量其周长所得的数据一致.因此, 在研究溶蚀微地貌时, 应采用高精度仪器, 统一测量标准, 分析微地貌是苔藓植物溶蚀作用产生, 还是由于淋溶风化而形成, 有利于后期岩溶微地貌之间的对比分析和进一步深入研究.这些微地貌的形态特征与生物类型和发育生境密切相关(
蔡胡霖, 2017 ), 在温湿度优越的地区发育更为广泛(
Liang & Xu, 2014 ;
Liu et al ., 2016 ).例如, 溶孔、溶锥多发生在低湿度、强光照、高温环境中, 而溶沟多是在高温、强光和pH较高的生境内发育(
申家琛等, 2018a ).
曹建华和王福星(1996) 认为苔藓植物岩溶微地貌与岩溶地区的宏观地貌形态之间存在一定的相关性, 然而目前这种关联性以及苔藓植物微地貌之间的关系和发育方向尚未得到科学揭示. ...
... 苔藓植物沉积作用的另一表现是石灰华或钙华(
张捷, 1993 ;
Kleinteich et al ., 2017 ).钙华形成的物理化学过程实质是CaCO
3 的沉淀与CO
2 的脱气.苔藓植物在钙华形成过程中为CaCO
3 晶体提供了稳定的生长发育基础, 并且苔藓植物的形态结构控制着沉积物的形态变化(
Mili?a et al ., 2006 ), 且它们的发育形态随沉积环境而变化(
张朝晖等, 2004 ;
郭云等, 2018 ).如在洞穴弱光带一般形成洞穴石刺、向光钟乳石、洞口叠层石等, 这类形态有很强的方向性(
王福星等, 1998 ).苔藓植物在水生环境中也常常形成多样的沉积形态, 如凤尾藓(
Fissidens )钙华、石灰藓(
Hydrogonium )钙华形成的溪流瀑布沉积, 泽藓(
Philonotis )钙华、水灰藓(
Hygrohypnum )钙华形成的河流瀑布沉积, 地钱-真藓(
Marchantia-Bryum )钙华等形成的石灰华坝沉积, 以及净口藓(
Gymnostomum )钙华等形成的瀑布洞穴沉积(
张朝晖和陈家宽, 2007 ).同时, 也有研究发现, 水流环境也是影响苔藓钙华沉积的重要因素, 苔藓与钙华层之间的水流速度与钙华沉积速率呈正相关关系(
Wang et al ., 2014 ).钙华作为大陆古气候的潜在档案,
Yan等(2017) 利用碳氧稳定同位素比率(
δ 13 C和
δ 18 O), 研究钙华在沉积过程中碳、氧的循环规律, 分析认为, 钙华沉积与季节更替引起的气温、降水等的变化密切相关.目前对于苔藓植物沉积作用的研究主要集中在沉积形态以及成土量方面, 对于苔藓植物沉积速率, 对岩溶地区成土过程中的贡献, 以及岩溶沉积作用对生境的改造和环境对岩溶沉积作用的影响等方面的研究还未见报道. ...
洞穴弱光带的生物岩溶
2
1998
... 岩石经苔藓植物溶蚀后, 表面常常出现一些凹凸不平和微溶孔, 这是由于岩石成分不均匀, 导致生物钻孔力的差异性(
王福星等, 1998 ), 以及溶蚀产物流失而形成溶蚀微地貌.
王福星等(1993) 将这些溶蚀微地貌, 按照口面形态、直径大小、深度等, 把短径小于1 mm的命名为微溶孔, 把短径为1-20 mm的命名为溶孔, 把短径为20-100 mm的命名为溶坑.
付兰和张朝晖(2010) 将连续分布的苔藓植物溶蚀微地貌, 按测算结果, 把长7.7-14.4 cm的命名为溶丝; 长0.8-19.0 cm, 宽0.6-7.2 cm的命名为溶斑; 长6.0-24.5 cm, 宽1.2-2.2 cm的命名为溶带.苔藓植物溶蚀过程很难通过宏观手段观测, 导致这两种命名法在大小和形态方面存在明显差异.然而,
Haward等(2011) 借助原子力显微镜(AFM)观察到黑云母表面浅层溶蚀坑的形成过程, 通过分析图像计算质量损失, 从而确定溶蚀过程中黑云母表面的活化能大小.研究表明, 通过分析活化能得到的溶坑大小与测量其周长所得的数据一致.因此, 在研究溶蚀微地貌时, 应采用高精度仪器, 统一测量标准, 分析微地貌是苔藓植物溶蚀作用产生, 还是由于淋溶风化而形成, 有利于后期岩溶微地貌之间的对比分析和进一步深入研究.这些微地貌的形态特征与生物类型和发育生境密切相关(
蔡胡霖, 2017 ), 在温湿度优越的地区发育更为广泛(
Liang & Xu, 2014 ;
Liu et al ., 2016 ).例如, 溶孔、溶锥多发生在低湿度、强光照、高温环境中, 而溶沟多是在高温、强光和pH较高的生境内发育(
申家琛等, 2018a ).
曹建华和王福星(1996) 认为苔藓植物岩溶微地貌与岩溶地区的宏观地貌形态之间存在一定的相关性, 然而目前这种关联性以及苔藓植物微地貌之间的关系和发育方向尚未得到科学揭示. ...
... 苔藓植物沉积作用的另一表现是石灰华或钙华(
张捷, 1993 ;
Kleinteich et al ., 2017 ).钙华形成的物理化学过程实质是CaCO
3 的沉淀与CO
2 的脱气.苔藓植物在钙华形成过程中为CaCO
3 晶体提供了稳定的生长发育基础, 并且苔藓植物的形态结构控制着沉积物的形态变化(
Mili?a et al ., 2006 ), 且它们的发育形态随沉积环境而变化(
张朝晖等, 2004 ;
郭云等, 2018 ).如在洞穴弱光带一般形成洞穴石刺、向光钟乳石、洞口叠层石等, 这类形态有很强的方向性(
王福星等, 1998 ).苔藓植物在水生环境中也常常形成多样的沉积形态, 如凤尾藓(
Fissidens )钙华、石灰藓(
Hydrogonium )钙华形成的溪流瀑布沉积, 泽藓(
Philonotis )钙华、水灰藓(
Hygrohypnum )钙华形成的河流瀑布沉积, 地钱-真藓(
Marchantia-Bryum )钙华等形成的石灰华坝沉积, 以及净口藓(
Gymnostomum )钙华等形成的瀑布洞穴沉积(
张朝晖和陈家宽, 2007 ).同时, 也有研究发现, 水流环境也是影响苔藓钙华沉积的重要因素, 苔藓与钙华层之间的水流速度与钙华沉积速率呈正相关关系(
Wang et al ., 2014 ).钙华作为大陆古气候的潜在档案,
Yan等(2017) 利用碳氧稳定同位素比率(
δ 13 C和
δ 18 O), 研究钙华在沉积过程中碳、氧的循环规律, 分析认为, 钙华沉积与季节更替引起的气温、降水等的变化密切相关.目前对于苔藓植物沉积作用的研究主要集中在沉积形态以及成土量方面, 对于苔藓植物沉积速率, 对岩溶地区成土过程中的贡献, 以及岩溶沉积作用对生境的改造和环境对岩溶沉积作用的影响等方面的研究还未见报道. ...
1
1993
... 岩石经苔藓植物溶蚀后, 表面常常出现一些凹凸不平和微溶孔, 这是由于岩石成分不均匀, 导致生物钻孔力的差异性(
王福星等, 1998 ), 以及溶蚀产物流失而形成溶蚀微地貌.
王福星等(1993) 将这些溶蚀微地貌, 按照口面形态、直径大小、深度等, 把短径小于1 mm的命名为微溶孔, 把短径为1-20 mm的命名为溶孔, 把短径为20-100 mm的命名为溶坑.
付兰和张朝晖(2010) 将连续分布的苔藓植物溶蚀微地貌, 按测算结果, 把长7.7-14.4 cm的命名为溶丝; 长0.8-19.0 cm, 宽0.6-7.2 cm的命名为溶斑; 长6.0-24.5 cm, 宽1.2-2.2 cm的命名为溶带.苔藓植物溶蚀过程很难通过宏观手段观测, 导致这两种命名法在大小和形态方面存在明显差异.然而,
Haward等(2011) 借助原子力显微镜(AFM)观察到黑云母表面浅层溶蚀坑的形成过程, 通过分析图像计算质量损失, 从而确定溶蚀过程中黑云母表面的活化能大小.研究表明, 通过分析活化能得到的溶坑大小与测量其周长所得的数据一致.因此, 在研究溶蚀微地貌时, 应采用高精度仪器, 统一测量标准, 分析微地貌是苔藓植物溶蚀作用产生, 还是由于淋溶风化而形成, 有利于后期岩溶微地貌之间的对比分析和进一步深入研究.这些微地貌的形态特征与生物类型和发育生境密切相关(
蔡胡霖, 2017 ), 在温湿度优越的地区发育更为广泛(
Liang & Xu, 2014 ;
Liu et al ., 2016 ).例如, 溶孔、溶锥多发生在低湿度、强光照、高温环境中, 而溶沟多是在高温、强光和pH较高的生境内发育(
申家琛等, 2018a ).
曹建华和王福星(1996) 认为苔藓植物岩溶微地貌与岩溶地区的宏观地貌形态之间存在一定的相关性, 然而目前这种关联性以及苔藓植物微地貌之间的关系和发育方向尚未得到科学揭示. ...
1
1993
... 岩石经苔藓植物溶蚀后, 表面常常出现一些凹凸不平和微溶孔, 这是由于岩石成分不均匀, 导致生物钻孔力的差异性(
王福星等, 1998 ), 以及溶蚀产物流失而形成溶蚀微地貌.
王福星等(1993) 将这些溶蚀微地貌, 按照口面形态、直径大小、深度等, 把短径小于1 mm的命名为微溶孔, 把短径为1-20 mm的命名为溶孔, 把短径为20-100 mm的命名为溶坑.
付兰和张朝晖(2010) 将连续分布的苔藓植物溶蚀微地貌, 按测算结果, 把长7.7-14.4 cm的命名为溶丝; 长0.8-19.0 cm, 宽0.6-7.2 cm的命名为溶斑; 长6.0-24.5 cm, 宽1.2-2.2 cm的命名为溶带.苔藓植物溶蚀过程很难通过宏观手段观测, 导致这两种命名法在大小和形态方面存在明显差异.然而,
Haward等(2011) 借助原子力显微镜(AFM)观察到黑云母表面浅层溶蚀坑的形成过程, 通过分析图像计算质量损失, 从而确定溶蚀过程中黑云母表面的活化能大小.研究表明, 通过分析活化能得到的溶坑大小与测量其周长所得的数据一致.因此, 在研究溶蚀微地貌时, 应采用高精度仪器, 统一测量标准, 分析微地貌是苔藓植物溶蚀作用产生, 还是由于淋溶风化而形成, 有利于后期岩溶微地貌之间的对比分析和进一步深入研究.这些微地貌的形态特征与生物类型和发育生境密切相关(
蔡胡霖, 2017 ), 在温湿度优越的地区发育更为广泛(
Liang & Xu, 2014 ;
Liu et al ., 2016 ).例如, 溶孔、溶锥多发生在低湿度、强光照、高温环境中, 而溶沟多是在高温、强光和pH较高的生境内发育(
申家琛等, 2018a ).
曹建华和王福星(1996) 认为苔藓植物岩溶微地貌与岩溶地区的宏观地貌形态之间存在一定的相关性, 然而目前这种关联性以及苔藓植物微地貌之间的关系和发育方向尚未得到科学揭示. ...
Dissolution experiment and transformation condition analysis of Paleogene aragonite in the Jiyang Depression, China
1
2017
... 苔藓植物常以丛集型、交织型、匍匐型等着生于岩面(
图1 ), 假根根系不发达, 只起固着作用(
娄玉霞, 2013 ), 其特殊的叶表面结构和细胞特征(
娄红祥, 2012 ), 使苔藓具有耐高温、抗旱、蓄水能力强、保湿固土等功能(
Proctor & Pence, 2002 ;
Krol et al ., 2003 ;
Zhang et al ., 2017 ).
曹建华和袁道先(1999) 研究发现, 当岩石表面有苔藓植物时, 其蒸发失水和主动吸水时间比裸岩分别延长了48%和57%, 持水量提高了16.6倍, 分析认为苔藓植物可以延缓岩面水分蒸发时间, 增加岩石水化学作用时间, 但是苔藓植物的这种蓄水保湿能力存在明显的种间差异(
申家琛等, 2017 ;
张显强等, 2018a ).苔藓植物参与岩溶作用的另一大优势, 是其叶表面角质层不发育, 且阳离子交换位点多(
吴玉环等, 2001 ), 能够吸附多种阳离子, 这些阳离子在苔藓植物体内被转化为无机盐、有机盐以及其他形式的化合物储存于组织中.苔藓植物除了以生物化学方式对岩石进行溶蚀之外, 还可以通过物理机械作用破坏岩石.例如, 苔藓植物的假根可以伸入岩石裂隙, 对岩石有一定的破坏作用(
Jackson, 2015 ).再者, 当气候干旱时, 苔藓植物在卷缩过程中会拔起岩面的一些小颗粒, 从而加速了岩石被破坏和成土速度(
连宾等, 2008 ).此外, 苔藓植物常与藻类、地衣、微生物等共生形成生物结皮, 它们之间的相互关系以及因气候因素引起的干湿交替和冻融作用对岩石也具有一定的破坏性(
郑云普等, 2009 ).例如, 菌类的菌丝可以伸入岩石晶体结构中破坏岩石, 细菌与真菌形成的配合基能够络合三价铁离子, 细菌分泌的柠檬酸、乙酸以及地衣分泌的地衣酸(
连宾等, 2011 ), 可以溶解岩石中的矿物(
Won-Pyo et al ., 2016 ;
Wang et al ., 2017 ).因此在研究苔藓植物岩溶作用时, 应明确苔藓植物自身及其与其他生物形成的生物结皮两者之间的关系.目前, 有关岩面苔藓植物功能性状的研究主要集中在小范围内特定生境中的苔藓植物研究, 无法代表岩溶生态系统苔藓植物的整体情况.对于大尺度空间上, 因生境异质性高而造成相同物种在形态结构、光合特征及其钙(Ca)、碳(C)、氮(N)、磷(P) 等元素的代谢调节机制等方面的差异性的研究还未引起人们的广泛关注.岩溶生态系统的物质基础相对贫乏, 具有“缺氮少磷钙丰富”的特征.因此, 研究Ca、C、N、P等元素在苔藓植物体内的代谢机制对揭示岩溶生态系统苔藓植物和其他植物的生物多样性维持机制, 及其岩溶作用特征具有重要的科学 意义. ...
Contrasts in variations of the carbon and oxygen isotopic composition of travertines formed in pools and a ramp stream at Huanglong Ravine, China: Implications for paleoclimatic interpretations
1
2014
... 苔藓植物沉积作用的另一表现是石灰华或钙华(
张捷, 1993 ;
Kleinteich et al ., 2017 ).钙华形成的物理化学过程实质是CaCO
3 的沉淀与CO
2 的脱气.苔藓植物在钙华形成过程中为CaCO
3 晶体提供了稳定的生长发育基础, 并且苔藓植物的形态结构控制着沉积物的形态变化(
Mili?a et al ., 2006 ), 且它们的发育形态随沉积环境而变化(
张朝晖等, 2004 ;
郭云等, 2018 ).如在洞穴弱光带一般形成洞穴石刺、向光钟乳石、洞口叠层石等, 这类形态有很强的方向性(
王福星等, 1998 ).苔藓植物在水生环境中也常常形成多样的沉积形态, 如凤尾藓(
Fissidens )钙华、石灰藓(
Hydrogonium )钙华形成的溪流瀑布沉积, 泽藓(
Philonotis )钙华、水灰藓(
Hygrohypnum )钙华形成的河流瀑布沉积, 地钱-真藓(
Marchantia-Bryum )钙华等形成的石灰华坝沉积, 以及净口藓(
Gymnostomum )钙华等形成的瀑布洞穴沉积(
张朝晖和陈家宽, 2007 ).同时, 也有研究发现, 水流环境也是影响苔藓钙华沉积的重要因素, 苔藓与钙华层之间的水流速度与钙华沉积速率呈正相关关系(
Wang et al ., 2014 ).钙华作为大陆古气候的潜在档案,
Yan等(2017) 利用碳氧稳定同位素比率(
δ 13 C和
δ 18 O), 研究钙华在沉积过程中碳、氧的循环规律, 分析认为, 钙华沉积与季节更替引起的气温、降水等的变化密切相关.目前对于苔藓植物沉积作用的研究主要集中在沉积形态以及成土量方面, 对于苔藓植物沉积速率, 对岩溶地区成土过程中的贡献, 以及岩溶沉积作用对生境的改造和环境对岩溶沉积作用的影响等方面的研究还未见报道. ...
碳酸盐岩风化成土作用的初步研究
1
1999
... 岩石中所含矿物种类及其晶体结构、产状、透水性等决定了岩石的溶解能力(
Voinot et al ., 2013 ;
黄奇波等, 2015 ), 例如, 一般情况下泥质含量高的碳酸盐岩溶蚀速度快, 岩石中酸不溶物含量越低, 越易被溶解(
王世杰等, 1999 ).同时, 岩石中酸不溶物含量对碳酸盐岩的成土速率具有重要影响, 酸不溶物含量越高岩石风化后可残留物越多, 成土物质越丰富(
陈洪松等, 2018 ).例如, 白云岩中酸不溶物含量高, 发育的土层较石灰岩地区土层厚(
陈洪松等, 2013 ).岩石中所含矿物的性质影响各元素的释放速率, Andrew等(2015)在25 ℃、pH = 2.6的开放试验系统与封闭试验系统内发现黑云母中的元素释放按照Fe > Mg > Al > Si的顺序进行, 且溶解速率符合 ${{r}_{i}}={{k}_{\text{H},i}}\times a_{{{\text{H}}^{\text{+}}}}^{xi}$,
k H 为表面速率常数((2.15-30.6) × 10
-10 mole s
biotite ·m
-2 ·s
-1 ),
a H+ 为H
+ 活性,
xi 为矿物与H
+ 的反应级数(0.31-0.58); 分析认为, 岩石中各矿物的溶解具有空间各向异性, 其边缘的反应效率强于基面, 而且各个元素的释放具有一定的规律性.苔藓植物种类的空间分布特征与岩性密切相关(
Gorozhankina & Konstantinov, 2001 ), 而且岩性和有机酸种类也会影响有机酸与岩石的反应速率(
Cama & Ganor, 2006 ;
Ghoorah et al ., 2014 ;
Mwangi et al ., 2018 ), 从而导致它们岩溶作用的差异性. ...
碳酸盐岩风化成土作用的初步研究
1
1999
... 岩石中所含矿物种类及其晶体结构、产状、透水性等决定了岩石的溶解能力(
Voinot et al ., 2013 ;
黄奇波等, 2015 ), 例如, 一般情况下泥质含量高的碳酸盐岩溶蚀速度快, 岩石中酸不溶物含量越低, 越易被溶解(
王世杰等, 1999 ).同时, 岩石中酸不溶物含量对碳酸盐岩的成土速率具有重要影响, 酸不溶物含量越高岩石风化后可残留物越多, 成土物质越丰富(
陈洪松等, 2018 ).例如, 白云岩中酸不溶物含量高, 发育的土层较石灰岩地区土层厚(
陈洪松等, 2013 ).岩石中所含矿物的性质影响各元素的释放速率, Andrew等(2015)在25 ℃、pH = 2.6的开放试验系统与封闭试验系统内发现黑云母中的元素释放按照Fe > Mg > Al > Si的顺序进行, 且溶解速率符合 ${{r}_{i}}={{k}_{\text{H},i}}\times a_{{{\text{H}}^{\text{+}}}}^{xi}$,
k H 为表面速率常数((2.15-30.6) × 10
-10 mole s
biotite ·m
-2 ·s
-1 ),
a H+ 为H
+ 活性,
xi 为矿物与H
+ 的反应级数(0.31-0.58); 分析认为, 岩石中各矿物的溶解具有空间各向异性, 其边缘的反应效率强于基面, 而且各个元素的释放具有一定的规律性.苔藓植物种类的空间分布特征与岩性密切相关(
Gorozhankina & Konstantinov, 2001 ), 而且岩性和有机酸种类也会影响有机酸与岩石的反应速率(
Cama & Ganor, 2006 ;
Ghoorah et al ., 2014 ;
Mwangi et al ., 2018 ), 从而导致它们岩溶作用的差异性. ...
Dissolution of metals from biosolid-treated soils by organic acid mixtures
1
2016
... 苔藓植物常以丛集型、交织型、匍匐型等着生于岩面(
图1 ), 假根根系不发达, 只起固着作用(
娄玉霞, 2013 ), 其特殊的叶表面结构和细胞特征(
娄红祥, 2012 ), 使苔藓具有耐高温、抗旱、蓄水能力强、保湿固土等功能(
Proctor & Pence, 2002 ;
Krol et al ., 2003 ;
Zhang et al ., 2017 ).
曹建华和袁道先(1999) 研究发现, 当岩石表面有苔藓植物时, 其蒸发失水和主动吸水时间比裸岩分别延长了48%和57%, 持水量提高了16.6倍, 分析认为苔藓植物可以延缓岩面水分蒸发时间, 增加岩石水化学作用时间, 但是苔藓植物的这种蓄水保湿能力存在明显的种间差异(
申家琛等, 2017 ;
张显强等, 2018a ).苔藓植物参与岩溶作用的另一大优势, 是其叶表面角质层不发育, 且阳离子交换位点多(
吴玉环等, 2001 ), 能够吸附多种阳离子, 这些阳离子在苔藓植物体内被转化为无机盐、有机盐以及其他形式的化合物储存于组织中.苔藓植物除了以生物化学方式对岩石进行溶蚀之外, 还可以通过物理机械作用破坏岩石.例如, 苔藓植物的假根可以伸入岩石裂隙, 对岩石有一定的破坏作用(
Jackson, 2015 ).再者, 当气候干旱时, 苔藓植物在卷缩过程中会拔起岩面的一些小颗粒, 从而加速了岩石被破坏和成土速度(
连宾等, 2008 ).此外, 苔藓植物常与藻类、地衣、微生物等共生形成生物结皮, 它们之间的相互关系以及因气候因素引起的干湿交替和冻融作用对岩石也具有一定的破坏性(
郑云普等, 2009 ).例如, 菌类的菌丝可以伸入岩石晶体结构中破坏岩石, 细菌与真菌形成的配合基能够络合三价铁离子, 细菌分泌的柠檬酸、乙酸以及地衣分泌的地衣酸(
连宾等, 2011 ), 可以溶解岩石中的矿物(
Won-Pyo et al ., 2016 ;
Wang et al ., 2017 ).因此在研究苔藓植物岩溶作用时, 应明确苔藓植物自身及其与其他生物形成的生物结皮两者之间的关系.目前, 有关岩面苔藓植物功能性状的研究主要集中在小范围内特定生境中的苔藓植物研究, 无法代表岩溶生态系统苔藓植物的整体情况.对于大尺度空间上, 因生境异质性高而造成相同物种在形态结构、光合特征及其钙(Ca)、碳(C)、氮(N)、磷(P) 等元素的代谢调节机制等方面的差异性的研究还未引起人们的广泛关注.岩溶生态系统的物质基础相对贫乏, 具有“缺氮少磷钙丰富”的特征.因此, 研究Ca、C、N、P等元素在苔藓植物体内的代谢机制对揭示岩溶生态系统苔藓植物和其他植物的生物多样性维持机制, 及其岩溶作用特征具有重要的科学 意义. ...
苔藓植物对环境变化的响应及适应性研究进展
1
2001
... 苔藓植物常以丛集型、交织型、匍匐型等着生于岩面(
图1 ), 假根根系不发达, 只起固着作用(
娄玉霞, 2013 ), 其特殊的叶表面结构和细胞特征(
娄红祥, 2012 ), 使苔藓具有耐高温、抗旱、蓄水能力强、保湿固土等功能(
Proctor & Pence, 2002 ;
Krol et al ., 2003 ;
Zhang et al ., 2017 ).
曹建华和袁道先(1999) 研究发现, 当岩石表面有苔藓植物时, 其蒸发失水和主动吸水时间比裸岩分别延长了48%和57%, 持水量提高了16.6倍, 分析认为苔藓植物可以延缓岩面水分蒸发时间, 增加岩石水化学作用时间, 但是苔藓植物的这种蓄水保湿能力存在明显的种间差异(
申家琛等, 2017 ;
张显强等, 2018a ).苔藓植物参与岩溶作用的另一大优势, 是其叶表面角质层不发育, 且阳离子交换位点多(
吴玉环等, 2001 ), 能够吸附多种阳离子, 这些阳离子在苔藓植物体内被转化为无机盐、有机盐以及其他形式的化合物储存于组织中.苔藓植物除了以生物化学方式对岩石进行溶蚀之外, 还可以通过物理机械作用破坏岩石.例如, 苔藓植物的假根可以伸入岩石裂隙, 对岩石有一定的破坏作用(
Jackson, 2015 ).再者, 当气候干旱时, 苔藓植物在卷缩过程中会拔起岩面的一些小颗粒, 从而加速了岩石被破坏和成土速度(
连宾等, 2008 ).此外, 苔藓植物常与藻类、地衣、微生物等共生形成生物结皮, 它们之间的相互关系以及因气候因素引起的干湿交替和冻融作用对岩石也具有一定的破坏性(
郑云普等, 2009 ).例如, 菌类的菌丝可以伸入岩石晶体结构中破坏岩石, 细菌与真菌形成的配合基能够络合三价铁离子, 细菌分泌的柠檬酸、乙酸以及地衣分泌的地衣酸(
连宾等, 2011 ), 可以溶解岩石中的矿物(
Won-Pyo et al ., 2016 ;
Wang et al ., 2017 ).因此在研究苔藓植物岩溶作用时, 应明确苔藓植物自身及其与其他生物形成的生物结皮两者之间的关系.目前, 有关岩面苔藓植物功能性状的研究主要集中在小范围内特定生境中的苔藓植物研究, 无法代表岩溶生态系统苔藓植物的整体情况.对于大尺度空间上, 因生境异质性高而造成相同物种在形态结构、光合特征及其钙(Ca)、碳(C)、氮(N)、磷(P) 等元素的代谢调节机制等方面的差异性的研究还未引起人们的广泛关注.岩溶生态系统的物质基础相对贫乏, 具有“缺氮少磷钙丰富”的特征.因此, 研究Ca、C、N、P等元素在苔藓植物体内的代谢机制对揭示岩溶生态系统苔藓植物和其他植物的生物多样性维持机制, 及其岩溶作用特征具有重要的科学 意义. ...
苔藓植物对环境变化的响应及适应性研究进展
1
2001
... 苔藓植物常以丛集型、交织型、匍匐型等着生于岩面(
图1 ), 假根根系不发达, 只起固着作用(
娄玉霞, 2013 ), 其特殊的叶表面结构和细胞特征(
娄红祥, 2012 ), 使苔藓具有耐高温、抗旱、蓄水能力强、保湿固土等功能(
Proctor & Pence, 2002 ;
Krol et al ., 2003 ;
Zhang et al ., 2017 ).
曹建华和袁道先(1999) 研究发现, 当岩石表面有苔藓植物时, 其蒸发失水和主动吸水时间比裸岩分别延长了48%和57%, 持水量提高了16.6倍, 分析认为苔藓植物可以延缓岩面水分蒸发时间, 增加岩石水化学作用时间, 但是苔藓植物的这种蓄水保湿能力存在明显的种间差异(
申家琛等, 2017 ;
张显强等, 2018a ).苔藓植物参与岩溶作用的另一大优势, 是其叶表面角质层不发育, 且阳离子交换位点多(
吴玉环等, 2001 ), 能够吸附多种阳离子, 这些阳离子在苔藓植物体内被转化为无机盐、有机盐以及其他形式的化合物储存于组织中.苔藓植物除了以生物化学方式对岩石进行溶蚀之外, 还可以通过物理机械作用破坏岩石.例如, 苔藓植物的假根可以伸入岩石裂隙, 对岩石有一定的破坏作用(
Jackson, 2015 ).再者, 当气候干旱时, 苔藓植物在卷缩过程中会拔起岩面的一些小颗粒, 从而加速了岩石被破坏和成土速度(
连宾等, 2008 ).此外, 苔藓植物常与藻类、地衣、微生物等共生形成生物结皮, 它们之间的相互关系以及因气候因素引起的干湿交替和冻融作用对岩石也具有一定的破坏性(
郑云普等, 2009 ).例如, 菌类的菌丝可以伸入岩石晶体结构中破坏岩石, 细菌与真菌形成的配合基能够络合三价铁离子, 细菌分泌的柠檬酸、乙酸以及地衣分泌的地衣酸(
连宾等, 2011 ), 可以溶解岩石中的矿物(
Won-Pyo et al ., 2016 ;
Wang et al ., 2017 ).因此在研究苔藓植物岩溶作用时, 应明确苔藓植物自身及其与其他生物形成的生物结皮两者之间的关系.目前, 有关岩面苔藓植物功能性状的研究主要集中在小范围内特定生境中的苔藓植物研究, 无法代表岩溶生态系统苔藓植物的整体情况.对于大尺度空间上, 因生境异质性高而造成相同物种在形态结构、光合特征及其钙(Ca)、碳(C)、氮(N)、磷(P) 等元素的代谢调节机制等方面的差异性的研究还未引起人们的广泛关注.岩溶生态系统的物质基础相对贫乏, 具有“缺氮少磷钙丰富”的特征.因此, 研究Ca、C、N、P等元素在苔藓植物体内的代谢机制对揭示岩溶生态系统苔藓植物和其他植物的生物多样性维持机制, 及其岩溶作用特征具有重要的科学 意义. ...
微生物碳酸酐酶在岩溶系统碳循环中的作用与应用研究进展
1
2015
... 此外, 苔藓植物体内的碳酸酐酶(CA)对岩溶作用也有促进作用(
Smith & Griffiths, 2000 ), 它主要催化CO
2 的水合反应: CO
2 + H
2 O $\rightleftharpoons $ HCO
3 - + H
+ (
吴雁雯和张金池, 2015 ).
李为等(2005) 研究发现无CA催化时上述可逆反应达到平衡的时间是有CA催化时的6 × 10
7 倍.生物体内均含有CA, 且CA活性因物种的不同而存在显著差异, 细菌、真菌、放线菌内均含有CA, 且微生物细胞内CA活性不同于细胞外CA, 细菌的CA对碳酸盐岩系统碳储存贡献在20%以上(
Li et al ., 2006 ), 真菌的CA对白云岩的溶蚀能力强于对石灰岩(
申泰铭等, 2014 ;
申泰铭, 2018 ).那么苔藓植物体内CA对岩溶作用的贡献是多少?各组织内的CA活性是否也存在这种差异?对此以及关于苔藓植物体内CA代谢的调节机制及原理仍不得而知.目前, 关于苔藓植物CA的研究, 主要集中在各类苔藓植物CA含量、活性及其对岩石溶蚀过程的促进作用等方面.例如,
刘天雷等(2017) 研究发现, 虽然苔藓植物都会分泌CA, 但CA分泌量、速率和活性存在种间差异.这种差异性可能是因生活型或者生长环境的不同而导致苔藓植物生理代谢与功能性状的差异引起的. ...
微生物碳酸酐酶在岩溶系统碳循环中的作用与应用研究进展
1
2015
... 此外, 苔藓植物体内的碳酸酐酶(CA)对岩溶作用也有促进作用(
Smith & Griffiths, 2000 ), 它主要催化CO
2 的水合反应: CO
2 + H
2 O $\rightleftharpoons $ HCO
3 - + H
+ (
吴雁雯和张金池, 2015 ).
李为等(2005) 研究发现无CA催化时上述可逆反应达到平衡的时间是有CA催化时的6 × 10
7 倍.生物体内均含有CA, 且CA活性因物种的不同而存在显著差异, 细菌、真菌、放线菌内均含有CA, 且微生物细胞内CA活性不同于细胞外CA, 细菌的CA对碳酸盐岩系统碳储存贡献在20%以上(
Li et al ., 2006 ), 真菌的CA对白云岩的溶蚀能力强于对石灰岩(
申泰铭等, 2014 ;
申泰铭, 2018 ).那么苔藓植物体内CA对岩溶作用的贡献是多少?各组织内的CA活性是否也存在这种差异?对此以及关于苔藓植物体内CA代谢的调节机制及原理仍不得而知.目前, 关于苔藓植物CA的研究, 主要集中在各类苔藓植物CA含量、活性及其对岩石溶蚀过程的促进作用等方面.例如,
刘天雷等(2017) 研究发现, 虽然苔藓植物都会分泌CA, 但CA分泌量、速率和活性存在种间差异.这种差异性可能是因生活型或者生长环境的不同而导致苔藓植物生理代谢与功能性状的差异引起的. ...
Secondary metabolites in bryophytes: An ecological aspect
1
2009
...
Xie和Lou (2009) 认为, 生物岩溶作用随pH值降低而增强, 如果单纯从H
+ + CaCO
3 $\rightleftharpoons $ CO
2 + H
2 O + Ca
2+ 角度分析pH对岩溶作用的影响, pH值越低溶蚀速率越快.然而岩石的最佳溶解pH由矿物自身属性决定(
Bray et al ., 2015 ), 同时有机酸的溶蚀能力也受pH影响(
Pachana et al ., 2012 ;
Chen et al ., 2016 ), 而且每类苔藓植物的最适生长pH也不尽相同.如泥炭藓属(
Sphagnum )植物适宜偏酸性环境, 细叶小羽藓(
Haplocladium microphyllum )配子体的最适生长pH值为7 (
陈文佳等, 2012 ).作为苔藓植物岩溶作用的重要参与者CA, 它的活性也受pH影响,
洪子茜(2018) 研究表明, 当pH在7-9条件下CA的活性最高.pH由外而内对苔藓植物岩溶作用的过程、速率、对象、各级参与者都产生不同程度的促进或抑制作用.目前我们对pH影响苔藓植物岩溶的规律以及调节机制还不得而知, 今后还需综合全面地进行深入研究. ...
苔藓植物DNA C-值测定及其变异特点分析
1
2018
... 因此, 湿度、温度、光照及其综合因子通过对苔藓植物生理代谢过程、基因型以及其他功能性状的影响(
Pharo & Zartman, 2006 ;
de la Rosa, 2016 ;
解梦, 2018 ), 而直接或间接地影响苔藓植物岩溶作用过程与效率.例如, 当温度在17.5-21.4 ℃, 湿度57.7%-72.6%, 光照2 700-5 900 lx条件下, 苔藓植物对碳酸盐岩的塑造作用最强(
申家琛等, 2018a ).但是目前对温湿度及其综合因子对苔藓植物岩溶作用的影响的关注度仍不够, 很多规律仍不清楚, 然而这些规律的发现是揭示苔藓植物岩溶机理的关键, 也是指导岩溶生态问题治理的重要科学依据. ...
苔藓植物DNA C-值测定及其变异特点分析
1
2018
... 因此, 湿度、温度、光照及其综合因子通过对苔藓植物生理代谢过程、基因型以及其他功能性状的影响(
Pharo & Zartman, 2006 ;
de la Rosa, 2016 ;
解梦, 2018 ), 而直接或间接地影响苔藓植物岩溶作用过程与效率.例如, 当温度在17.5-21.4 ℃, 湿度57.7%-72.6%, 光照2 700-5 900 lx条件下, 苔藓植物对碳酸盐岩的塑造作用最强(
申家琛等, 2018a ).但是目前对温湿度及其综合因子对苔藓植物岩溶作用的影响的关注度仍不够, 很多规律仍不清楚, 然而这些规律的发现是揭示苔藓植物岩溶机理的关键, 也是指导岩溶生态问题治理的重要科学依据. ...
2013
2013
Effect of in-stream physicochemical processes on the seasonal variations in δ
13 C and δ
18 O values in laminated travertine deposits in a mountain stream channel
1
2017
... 苔藓植物沉积作用的另一表现是石灰华或钙华(
张捷, 1993 ;
Kleinteich et al ., 2017 ).钙华形成的物理化学过程实质是CaCO
3 的沉淀与CO
2 的脱气.苔藓植物在钙华形成过程中为CaCO
3 晶体提供了稳定的生长发育基础, 并且苔藓植物的形态结构控制着沉积物的形态变化(
Mili?a et al ., 2006 ), 且它们的发育形态随沉积环境而变化(
张朝晖等, 2004 ;
郭云等, 2018 ).如在洞穴弱光带一般形成洞穴石刺、向光钟乳石、洞口叠层石等, 这类形态有很强的方向性(
王福星等, 1998 ).苔藓植物在水生环境中也常常形成多样的沉积形态, 如凤尾藓(
Fissidens )钙华、石灰藓(
Hydrogonium )钙华形成的溪流瀑布沉积, 泽藓(
Philonotis )钙华、水灰藓(
Hygrohypnum )钙华形成的河流瀑布沉积, 地钱-真藓(
Marchantia-Bryum )钙华等形成的石灰华坝沉积, 以及净口藓(
Gymnostomum )钙华等形成的瀑布洞穴沉积(
张朝晖和陈家宽, 2007 ).同时, 也有研究发现, 水流环境也是影响苔藓钙华沉积的重要因素, 苔藓与钙华层之间的水流速度与钙华沉积速率呈正相关关系(
Wang et al ., 2014 ).钙华作为大陆古气候的潜在档案,
Yan等(2017) 利用碳氧稳定同位素比率(
δ 13 C和
δ 18 O), 研究钙华在沉积过程中碳、氧的循环规律, 分析认为, 钙华沉积与季节更替引起的气温、降水等的变化密切相关.目前对于苔藓植物沉积作用的研究主要集中在沉积形态以及成土量方面, 对于苔藓植物沉积速率, 对岩溶地区成土过程中的贡献, 以及岩溶沉积作用对生境的改造和环境对岩溶沉积作用的影响等方面的研究还未见报道. ...
温度及CO
2 对方解石、白云石溶解度影响特征分析
1
2009
... 苔藓植物群落分布特征与生境条件密切相关(
Heino & Virtanen, 2006 ;
刘艳等, 2015 ;
Mod et al ., 2016 ;
李泽科等, 2018 ;
庞嘉鹏等, 2018 ), 温度和水分是影响苔藓植物生存和分布的主要决定性因素, 尤其是空气湿度(
娄红祥, 2012 ;
Guo & Zhao, 2018 ).
党成强等(2018) 研究表明, 南亚毛灰藓(
Homomallium simlaense )在干旱和高温条件下, 各项生理指标均与体内相对含水量呈正比.苔藓植物在干旱缺水的情况下会停止生理代谢进入休眠状态, 当水分充足时再进行生命活动.而且苔藓植物生物量与相对生长速率随光照增强而增加, 随光照时间的延长, 其长度、pH值降低能力逐渐提高(
St. Martin & Mallik, 2017 ).苔藓植物虽然能够忍受高温, 但是温度高于25 ℃时却很少有净光合产物(
Proctor, 2000 ;
Hanson et al ., 2014 ).同时, 温度也是影响酶活性的关键因素, 温度超过60 ℃时会严重影响CA的活性(
洪子茜, 2018 ).然而, CA参与苔藓植物岩溶作用的最佳酶促反应温度还不得而知.同时, 温度也影响有机酸的反应性、络合能力和岩石中矿物自身的溶解程度(
闫志为等, 2009 ;
Benbouzid et al ., 2012 ), 进而影响岩石的溶蚀强度(
陈传平等, 2008 ).而且有机酸与矿物反应生成有机酸盐的存在形态也随温度发生变化(
Peters et al ., 2018 ). ...
温度及CO
2 对方解石、白云石溶解度影响特征分析
1
2009
... 苔藓植物群落分布特征与生境条件密切相关(
Heino & Virtanen, 2006 ;
刘艳等, 2015 ;
Mod et al ., 2016 ;
李泽科等, 2018 ;
庞嘉鹏等, 2018 ), 温度和水分是影响苔藓植物生存和分布的主要决定性因素, 尤其是空气湿度(
娄红祥, 2012 ;
Guo & Zhao, 2018 ).
党成强等(2018) 研究表明, 南亚毛灰藓(
Homomallium simlaense )在干旱和高温条件下, 各项生理指标均与体内相对含水量呈正比.苔藓植物在干旱缺水的情况下会停止生理代谢进入休眠状态, 当水分充足时再进行生命活动.而且苔藓植物生物量与相对生长速率随光照增强而增加, 随光照时间的延长, 其长度、pH值降低能力逐渐提高(
St. Martin & Mallik, 2017 ).苔藓植物虽然能够忍受高温, 但是温度高于25 ℃时却很少有净光合产物(
Proctor, 2000 ;
Hanson et al ., 2014 ).同时, 温度也是影响酶活性的关键因素, 温度超过60 ℃时会严重影响CA的活性(
洪子茜, 2018 ).然而, CA参与苔藓植物岩溶作用的最佳酶促反应温度还不得而知.同时, 温度也影响有机酸的反应性、络合能力和岩石中矿物自身的溶解程度(
闫志为等, 2009 ;
Benbouzid et al ., 2012 ), 进而影响岩石的溶蚀强度(
陈传平等, 2008 ).而且有机酸与矿物反应生成有机酸盐的存在形态也随温度发生变化(
Peters et al ., 2018 ). ...
外源低分子量有机酸对土壤中钙素迁移特征的影响
1
2015
... 在岩石溶蚀过程中, 有些岩石因产状、矿物成分、气候、生物等因素利于溶蚀产物残留, 沉积物较多, 进而为沉积作用做贡献; 然而有些岩石可溶性矿物含量多, 则溶蚀残留物较少.例如, H
2 CO
3 与碳酸盐岩中的CaCO
3 反应生成Ca(HCO
3 )
2 随水溶失, 残留物较少.有机酸常常与岩石中钙等矿物结合生成有机酸钙等难溶化合物(
张大庚等, 2015 ), 赋存于残积物中, 进而影响土壤的矿物组分与含量.由于生物岩溶作用研究需要长时间尺度的监测, 研究难度较大, 所以对岩面因苔藓植物溶蚀作用而形成的溶蚀残积物中的物质种类、形态特征以及苔藓植物岩溶作用与生境之间关系的研究仍然很欠缺. ...
外源低分子量有机酸对土壤中钙素迁移特征的影响
1
2015
... 在岩石溶蚀过程中, 有些岩石因产状、矿物成分、气候、生物等因素利于溶蚀产物残留, 沉积物较多, 进而为沉积作用做贡献; 然而有些岩石可溶性矿物含量多, 则溶蚀残留物较少.例如, H
2 CO
3 与碳酸盐岩中的CaCO
3 反应生成Ca(HCO
3 )
2 随水溶失, 残留物较少.有机酸常常与岩石中钙等矿物结合生成有机酸钙等难溶化合物(
张大庚等, 2015 ), 赋存于残积物中, 进而影响土壤的矿物组分与含量.由于生物岩溶作用研究需要长时间尺度的监测, 研究难度较大, 所以对岩面因苔藓植物溶蚀作用而形成的溶蚀残积物中的物质种类、形态特征以及苔藓植物岩溶作用与生境之间关系的研究仍然很欠缺. ...
川西北眠山灰岩区岩溶堰塞湖形成中的生物作用
1
1993
... 苔藓植物沉积作用的另一表现是石灰华或钙华(
张捷, 1993 ;
Kleinteich et al ., 2017 ).钙华形成的物理化学过程实质是CaCO
3 的沉淀与CO
2 的脱气.苔藓植物在钙华形成过程中为CaCO
3 晶体提供了稳定的生长发育基础, 并且苔藓植物的形态结构控制着沉积物的形态变化(
Mili?a et al ., 2006 ), 且它们的发育形态随沉积环境而变化(
张朝晖等, 2004 ;
郭云等, 2018 ).如在洞穴弱光带一般形成洞穴石刺、向光钟乳石、洞口叠层石等, 这类形态有很强的方向性(
王福星等, 1998 ).苔藓植物在水生环境中也常常形成多样的沉积形态, 如凤尾藓(
Fissidens )钙华、石灰藓(
Hydrogonium )钙华形成的溪流瀑布沉积, 泽藓(
Philonotis )钙华、水灰藓(
Hygrohypnum )钙华形成的河流瀑布沉积, 地钱-真藓(
Marchantia-Bryum )钙华等形成的石灰华坝沉积, 以及净口藓(
Gymnostomum )钙华等形成的瀑布洞穴沉积(
张朝晖和陈家宽, 2007 ).同时, 也有研究发现, 水流环境也是影响苔藓钙华沉积的重要因素, 苔藓与钙华层之间的水流速度与钙华沉积速率呈正相关关系(
Wang et al ., 2014 ).钙华作为大陆古气候的潜在档案,
Yan等(2017) 利用碳氧稳定同位素比率(
δ 13 C和
δ 18 O), 研究钙华在沉积过程中碳、氧的循环规律, 分析认为, 钙华沉积与季节更替引起的气温、降水等的变化密切相关.目前对于苔藓植物沉积作用的研究主要集中在沉积形态以及成土量方面, 对于苔藓植物沉积速率, 对岩溶地区成土过程中的贡献, 以及岩溶沉积作用对生境的改造和环境对岩溶沉积作用的影响等方面的研究还未见报道. ...
川西北眠山灰岩区岩溶堰塞湖形成中的生物作用
1
1993
... 苔藓植物沉积作用的另一表现是石灰华或钙华(
张捷, 1993 ;
Kleinteich et al ., 2017 ).钙华形成的物理化学过程实质是CaCO
3 的沉淀与CO
2 的脱气.苔藓植物在钙华形成过程中为CaCO
3 晶体提供了稳定的生长发育基础, 并且苔藓植物的形态结构控制着沉积物的形态变化(
Mili?a et al ., 2006 ), 且它们的发育形态随沉积环境而变化(
张朝晖等, 2004 ;
郭云等, 2018 ).如在洞穴弱光带一般形成洞穴石刺、向光钟乳石、洞口叠层石等, 这类形态有很强的方向性(
王福星等, 1998 ).苔藓植物在水生环境中也常常形成多样的沉积形态, 如凤尾藓(
Fissidens )钙华、石灰藓(
Hydrogonium )钙华形成的溪流瀑布沉积, 泽藓(
Philonotis )钙华、水灰藓(
Hygrohypnum )钙华形成的河流瀑布沉积, 地钱-真藓(
Marchantia-Bryum )钙华等形成的石灰华坝沉积, 以及净口藓(
Gymnostomum )钙华等形成的瀑布洞穴沉积(
张朝晖和陈家宽, 2007 ).同时, 也有研究发现, 水流环境也是影响苔藓钙华沉积的重要因素, 苔藓与钙华层之间的水流速度与钙华沉积速率呈正相关关系(
Wang et al ., 2014 ).钙华作为大陆古气候的潜在档案,
Yan等(2017) 利用碳氧稳定同位素比率(
δ 13 C和
δ 18 O), 研究钙华在沉积过程中碳、氧的循环规律, 分析认为, 钙华沉积与季节更替引起的气温、降水等的变化密切相关.目前对于苔藓植物沉积作用的研究主要集中在沉积形态以及成土量方面, 对于苔藓植物沉积速率, 对岩溶地区成土过程中的贡献, 以及岩溶沉积作用对生境的改造和环境对岩溶沉积作用的影响等方面的研究还未见报道. ...
贵州喀斯特几种石生苔藓和土壤微生物对石灰岩的溶蚀作用
2
2017
... 苔藓植物常以丛集型、交织型、匍匐型等着生于岩面(
图1 ), 假根根系不发达, 只起固着作用(
娄玉霞, 2013 ), 其特殊的叶表面结构和细胞特征(
娄红祥, 2012 ), 使苔藓具有耐高温、抗旱、蓄水能力强、保湿固土等功能(
Proctor & Pence, 2002 ;
Krol et al ., 2003 ;
Zhang et al ., 2017 ).
曹建华和袁道先(1999) 研究发现, 当岩石表面有苔藓植物时, 其蒸发失水和主动吸水时间比裸岩分别延长了48%和57%, 持水量提高了16.6倍, 分析认为苔藓植物可以延缓岩面水分蒸发时间, 增加岩石水化学作用时间, 但是苔藓植物的这种蓄水保湿能力存在明显的种间差异(
申家琛等, 2017 ;
张显强等, 2018a ).苔藓植物参与岩溶作用的另一大优势, 是其叶表面角质层不发育, 且阳离子交换位点多(
吴玉环等, 2001 ), 能够吸附多种阳离子, 这些阳离子在苔藓植物体内被转化为无机盐、有机盐以及其他形式的化合物储存于组织中.苔藓植物除了以生物化学方式对岩石进行溶蚀之外, 还可以通过物理机械作用破坏岩石.例如, 苔藓植物的假根可以伸入岩石裂隙, 对岩石有一定的破坏作用(
Jackson, 2015 ).再者, 当气候干旱时, 苔藓植物在卷缩过程中会拔起岩面的一些小颗粒, 从而加速了岩石被破坏和成土速度(
连宾等, 2008 ).此外, 苔藓植物常与藻类、地衣、微生物等共生形成生物结皮, 它们之间的相互关系以及因气候因素引起的干湿交替和冻融作用对岩石也具有一定的破坏性(
郑云普等, 2009 ).例如, 菌类的菌丝可以伸入岩石晶体结构中破坏岩石, 细菌与真菌形成的配合基能够络合三价铁离子, 细菌分泌的柠檬酸、乙酸以及地衣分泌的地衣酸(
连宾等, 2011 ), 可以溶解岩石中的矿物(
Won-Pyo et al ., 2016 ;
Wang et al ., 2017 ).因此在研究苔藓植物岩溶作用时, 应明确苔藓植物自身及其与其他生物形成的生物结皮两者之间的关系.目前, 有关岩面苔藓植物功能性状的研究主要集中在小范围内特定生境中的苔藓植物研究, 无法代表岩溶生态系统苔藓植物的整体情况.对于大尺度空间上, 因生境异质性高而造成相同物种在形态结构、光合特征及其钙(Ca)、碳(C)、氮(N)、磷(P) 等元素的代谢调节机制等方面的差异性的研究还未引起人们的广泛关注.岩溶生态系统的物质基础相对贫乏, 具有“缺氮少磷钙丰富”的特征.因此, 研究Ca、C、N、P等元素在苔藓植物体内的代谢机制对揭示岩溶生态系统苔藓植物和其他植物的生物多样性维持机制, 及其岩溶作用特征具有重要的科学 意义. ...
... 苔藓植物对岩石的溶蚀作用原理与过程实质是岩石中矿物与苔藓植物之间的作用过程(
图2 ).苔藓植物因呼吸作用形成的H
2 CO
3 和代谢分泌物与岩石中的矿物发生酸化、碱化、氧化还原、螯合等反应, 随着反应过程不断推进, 岩石矿物成分发生变化, Ca、镁(Mg)、钠(Na)、钾(K)、铁(Fe)、铝(Al)、硅(Si)等从碳酸盐、硅酸盐、铝酸盐、硫化物、氧化物中溶出, 破坏原岩晶体结构, 达到溶蚀岩石, 促使风化的目的(
Street et al ., 2013 ;
Li et al ., 2015 ;
Lammers et al ., 2017 ).这种岩溶驱动力的大小因植物形态特征、岩石性质与生存环境改变而发生变化.有研究表明, 碳酸盐岩溶蚀过程中, 生物成因的CO
2 占溶蚀强度的49.26%, 有机酸占37.11% (
张楷燕, 2017 ).苔藓植物溶蚀作用过程释放的矿物离子和其他元素部分被苔藓植物拦截形成岩溶沉积物; 部分被苔藓植物吸收利用, 转化为其他物质存储于植物体内; 其余则随降雨冲刷而流失, 进入其他生态系统.关于苔藓植物呼吸作用和其分泌物引起的岩溶作用驱动力、效率及其与苔藓植物生活型、种间差异、生境特征之间的相互关系仍未引起人们的关注, 对苔藓植物溶蚀作用过程和机理、苔藓植物对岩石中矿物营养元素释放规律的影响, 以及这些矿物质在岩溶沉积过程中的形态转化过程及其与生境间的相互作用关系等的研究还未见报道, 这也正是揭示苔藓植物岩溶作用机理的关键点.岩石的溶解速率与有机酸的酸性和溶蚀能力密切相关(
刘胜亮等, 2017 ), 然而有机酸的理化特性取决于其官能团的属性(
赵志曼等, 2018 ).因此, 今后需结合有机化学、气相色谱质谱(GC-MS)、核磁共振(NMR)、液相色谱(HPLC)等技术手段深入研究苔藓植物的溶蚀过程与机理. ...
贵州喀斯特几种石生苔藓和土壤微生物对石灰岩的溶蚀作用
2
2017
... 苔藓植物常以丛集型、交织型、匍匐型等着生于岩面(
图1 ), 假根根系不发达, 只起固着作用(
娄玉霞, 2013 ), 其特殊的叶表面结构和细胞特征(
娄红祥, 2012 ), 使苔藓具有耐高温、抗旱、蓄水能力强、保湿固土等功能(
Proctor & Pence, 2002 ;
Krol et al ., 2003 ;
Zhang et al ., 2017 ).
曹建华和袁道先(1999) 研究发现, 当岩石表面有苔藓植物时, 其蒸发失水和主动吸水时间比裸岩分别延长了48%和57%, 持水量提高了16.6倍, 分析认为苔藓植物可以延缓岩面水分蒸发时间, 增加岩石水化学作用时间, 但是苔藓植物的这种蓄水保湿能力存在明显的种间差异(
申家琛等, 2017 ;
张显强等, 2018a ).苔藓植物参与岩溶作用的另一大优势, 是其叶表面角质层不发育, 且阳离子交换位点多(
吴玉环等, 2001 ), 能够吸附多种阳离子, 这些阳离子在苔藓植物体内被转化为无机盐、有机盐以及其他形式的化合物储存于组织中.苔藓植物除了以生物化学方式对岩石进行溶蚀之外, 还可以通过物理机械作用破坏岩石.例如, 苔藓植物的假根可以伸入岩石裂隙, 对岩石有一定的破坏作用(
Jackson, 2015 ).再者, 当气候干旱时, 苔藓植物在卷缩过程中会拔起岩面的一些小颗粒, 从而加速了岩石被破坏和成土速度(
连宾等, 2008 ).此外, 苔藓植物常与藻类、地衣、微生物等共生形成生物结皮, 它们之间的相互关系以及因气候因素引起的干湿交替和冻融作用对岩石也具有一定的破坏性(
郑云普等, 2009 ).例如, 菌类的菌丝可以伸入岩石晶体结构中破坏岩石, 细菌与真菌形成的配合基能够络合三价铁离子, 细菌分泌的柠檬酸、乙酸以及地衣分泌的地衣酸(
连宾等, 2011 ), 可以溶解岩石中的矿物(
Won-Pyo et al ., 2016 ;
Wang et al ., 2017 ).因此在研究苔藓植物岩溶作用时, 应明确苔藓植物自身及其与其他生物形成的生物结皮两者之间的关系.目前, 有关岩面苔藓植物功能性状的研究主要集中在小范围内特定生境中的苔藓植物研究, 无法代表岩溶生态系统苔藓植物的整体情况.对于大尺度空间上, 因生境异质性高而造成相同物种在形态结构、光合特征及其钙(Ca)、碳(C)、氮(N)、磷(P) 等元素的代谢调节机制等方面的差异性的研究还未引起人们的广泛关注.岩溶生态系统的物质基础相对贫乏, 具有“缺氮少磷钙丰富”的特征.因此, 研究Ca、C、N、P等元素在苔藓植物体内的代谢机制对揭示岩溶生态系统苔藓植物和其他植物的生物多样性维持机制, 及其岩溶作用特征具有重要的科学 意义. ...
... 苔藓植物对岩石的溶蚀作用原理与过程实质是岩石中矿物与苔藓植物之间的作用过程(
图2 ).苔藓植物因呼吸作用形成的H
2 CO
3 和代谢分泌物与岩石中的矿物发生酸化、碱化、氧化还原、螯合等反应, 随着反应过程不断推进, 岩石矿物成分发生变化, Ca、镁(Mg)、钠(Na)、钾(K)、铁(Fe)、铝(Al)、硅(Si)等从碳酸盐、硅酸盐、铝酸盐、硫化物、氧化物中溶出, 破坏原岩晶体结构, 达到溶蚀岩石, 促使风化的目的(
Street et al ., 2013 ;
Li et al ., 2015 ;
Lammers et al ., 2017 ).这种岩溶驱动力的大小因植物形态特征、岩石性质与生存环境改变而发生变化.有研究表明, 碳酸盐岩溶蚀过程中, 生物成因的CO
2 占溶蚀强度的49.26%, 有机酸占37.11% (
张楷燕, 2017 ).苔藓植物溶蚀作用过程释放的矿物离子和其他元素部分被苔藓植物拦截形成岩溶沉积物; 部分被苔藓植物吸收利用, 转化为其他物质存储于植物体内; 其余则随降雨冲刷而流失, 进入其他生态系统.关于苔藓植物呼吸作用和其分泌物引起的岩溶作用驱动力、效率及其与苔藓植物生活型、种间差异、生境特征之间的相互关系仍未引起人们的关注, 对苔藓植物溶蚀作用过程和机理、苔藓植物对岩石中矿物营养元素释放规律的影响, 以及这些矿物质在岩溶沉积过程中的形态转化过程及其与生境间的相互作用关系等的研究还未见报道, 这也正是揭示苔藓植物岩溶作用机理的关键点.岩石的溶解速率与有机酸的酸性和溶蚀能力密切相关(
刘胜亮等, 2017 ), 然而有机酸的理化特性取决于其官能团的属性(
赵志曼等, 2018 ).因此, 今后需结合有机化学、气相色谱质谱(GC-MS)、核磁共振(NMR)、液相色谱(HPLC)等技术手段深入研究苔藓植物的溶蚀过程与机理. ...
我国南方岩溶地区的土壤及其形成
1
1994
... 岩溶地区土壤的形成和发育是碳酸岩长期溶蚀、风化与生物富集过程的结果, 其一方面受岩石结构、性质影响, 另一方面也与植被作用密切相关(
张美良和邓自强, 1994 ).研究表明, 碳酸盐岩的成土速率与岩石溶蚀速率成正比(
曹建华等, 2004 ), 与酸不溶物含量密切相关(
曹沁智等, 2018 ).苔藓植物殖居于岩面, 假根系以及植物体的干湿交替冻融现象所产生的物理机械力作用于岩石, 产生岩石碎屑.同时, 有机物与矿物离子络合常常形成难溶物, 这些难溶物与苔藓及其生物结皮死亡残体黏结在一起, 不仅增加了土壤沉积量, 而且促进有机质积累, 增加土壤养分(
朱显谟, 1995 ;
刘天雷等, 2017 ).苔藓植物通过影响下垫面土壤湿度, 促进土壤对岩石的岩溶作用, 间接地影响沉积过程.同时, 苔藓植物的吸附拦截功能对防治岩石表面水土流失具有重要作用(
李军峰等, 2013 ;
贾少华等, 2014 ;
张显强等, 2018b ).苔藓植物的溶蚀作用为岩面土壤形成提供了原始物质, 关于苔藓对土壤沉积的贡献率与苔藓植物作用下岩面土壤的形成过程, 以及在此过程中岩石的物质、能量转化和土壤养分的变化等还未见报道. ...
我国南方岩溶地区的土壤及其形成
1
1994
... 岩溶地区土壤的形成和发育是碳酸岩长期溶蚀、风化与生物富集过程的结果, 其一方面受岩石结构、性质影响, 另一方面也与植被作用密切相关(
张美良和邓自强, 1994 ).研究表明, 碳酸盐岩的成土速率与岩石溶蚀速率成正比(
曹建华等, 2004 ), 与酸不溶物含量密切相关(
曹沁智等, 2018 ).苔藓植物殖居于岩面, 假根系以及植物体的干湿交替冻融现象所产生的物理机械力作用于岩石, 产生岩石碎屑.同时, 有机物与矿物离子络合常常形成难溶物, 这些难溶物与苔藓及其生物结皮死亡残体黏结在一起, 不仅增加了土壤沉积量, 而且促进有机质积累, 增加土壤养分(
朱显谟, 1995 ;
刘天雷等, 2017 ).苔藓植物通过影响下垫面土壤湿度, 促进土壤对岩石的岩溶作用, 间接地影响沉积过程.同时, 苔藓植物的吸附拦截功能对防治岩石表面水土流失具有重要作用(
李军峰等, 2013 ;
贾少华等, 2014 ;
张显强等, 2018b ).苔藓植物的溶蚀作用为岩面土壤形成提供了原始物质, 关于苔藓对土壤沉积的贡献率与苔藓植物作用下岩面土壤的形成过程, 以及在此过程中岩石的物质、能量转化和土壤养分的变化等还未见报道. ...
a). 贵州5种喀斯特石生藓类成土及保土生态功能研究
2018
a). 贵州5种喀斯特石生藓类成土及保土生态功能研究
2018
b). 贵州喀斯特地区5种石生藓类的持水性能及吸水特征比较
2018
b). 贵州喀斯特地区5种石生藓类的持水性能及吸水特征比较
2018
Responses of antioxidant defense system of epilithic mosses to drought stress in karst rock desertified areas
2017
黔中瀑布水生苔藓植物区系及其生物喀斯特沉积生态类型研究
1
2007
... 苔藓植物沉积作用的另一表现是石灰华或钙华(
张捷, 1993 ;
Kleinteich et al ., 2017 ).钙华形成的物理化学过程实质是CaCO
3 的沉淀与CO
2 的脱气.苔藓植物在钙华形成过程中为CaCO
3 晶体提供了稳定的生长发育基础, 并且苔藓植物的形态结构控制着沉积物的形态变化(
Mili?a et al ., 2006 ), 且它们的发育形态随沉积环境而变化(
张朝晖等, 2004 ;
郭云等, 2018 ).如在洞穴弱光带一般形成洞穴石刺、向光钟乳石、洞口叠层石等, 这类形态有很强的方向性(
王福星等, 1998 ).苔藓植物在水生环境中也常常形成多样的沉积形态, 如凤尾藓(
Fissidens )钙华、石灰藓(
Hydrogonium )钙华形成的溪流瀑布沉积, 泽藓(
Philonotis )钙华、水灰藓(
Hygrohypnum )钙华形成的河流瀑布沉积, 地钱-真藓(
Marchantia-Bryum )钙华等形成的石灰华坝沉积, 以及净口藓(
Gymnostomum )钙华等形成的瀑布洞穴沉积(
张朝晖和陈家宽, 2007 ).同时, 也有研究发现, 水流环境也是影响苔藓钙华沉积的重要因素, 苔藓与钙华层之间的水流速度与钙华沉积速率呈正相关关系(
Wang et al ., 2014 ).钙华作为大陆古气候的潜在档案,
Yan等(2017) 利用碳氧稳定同位素比率(
δ 13 C和
δ 18 O), 研究钙华在沉积过程中碳、氧的循环规律, 分析认为, 钙华沉积与季节更替引起的气温、降水等的变化密切相关.目前对于苔藓植物沉积作用的研究主要集中在沉积形态以及成土量方面, 对于苔藓植物沉积速率, 对岩溶地区成土过程中的贡献, 以及岩溶沉积作用对生境的改造和环境对岩溶沉积作用的影响等方面的研究还未见报道. ...
黔中瀑布水生苔藓植物区系及其生物喀斯特沉积生态类型研究
1
2007
... 苔藓植物沉积作用的另一表现是石灰华或钙华(
张捷, 1993 ;
Kleinteich et al ., 2017 ).钙华形成的物理化学过程实质是CaCO
3 的沉淀与CO
2 的脱气.苔藓植物在钙华形成过程中为CaCO
3 晶体提供了稳定的生长发育基础, 并且苔藓植物的形态结构控制着沉积物的形态变化(
Mili?a et al ., 2006 ), 且它们的发育形态随沉积环境而变化(
张朝晖等, 2004 ;
郭云等, 2018 ).如在洞穴弱光带一般形成洞穴石刺、向光钟乳石、洞口叠层石等, 这类形态有很强的方向性(
王福星等, 1998 ).苔藓植物在水生环境中也常常形成多样的沉积形态, 如凤尾藓(
Fissidens )钙华、石灰藓(
Hydrogonium )钙华形成的溪流瀑布沉积, 泽藓(
Philonotis )钙华、水灰藓(
Hygrohypnum )钙华形成的河流瀑布沉积, 地钱-真藓(
Marchantia-Bryum )钙华等形成的石灰华坝沉积, 以及净口藓(
Gymnostomum )钙华等形成的瀑布洞穴沉积(
张朝晖和陈家宽, 2007 ).同时, 也有研究发现, 水流环境也是影响苔藓钙华沉积的重要因素, 苔藓与钙华层之间的水流速度与钙华沉积速率呈正相关关系(
Wang et al ., 2014 ).钙华作为大陆古气候的潜在档案,
Yan等(2017) 利用碳氧稳定同位素比率(
δ 13 C和
δ 18 O), 研究钙华在沉积过程中碳、氧的循环规律, 分析认为, 钙华沉积与季节更替引起的气温、降水等的变化密切相关.目前对于苔藓植物沉积作用的研究主要集中在沉积形态以及成土量方面, 对于苔藓植物沉积速率, 对岩溶地区成土过程中的贡献, 以及岩溶沉积作用对生境的改造和环境对岩溶沉积作用的影响等方面的研究还未见报道. ...
中国昆明地区岩溶洞穴洞口带苔藓植物研究
1
2004
... 苔藓植物沉积作用的另一表现是石灰华或钙华(
张捷, 1993 ;
Kleinteich et al ., 2017 ).钙华形成的物理化学过程实质是CaCO
3 的沉淀与CO
2 的脱气.苔藓植物在钙华形成过程中为CaCO
3 晶体提供了稳定的生长发育基础, 并且苔藓植物的形态结构控制着沉积物的形态变化(
Mili?a et al ., 2006 ), 且它们的发育形态随沉积环境而变化(
张朝晖等, 2004 ;
郭云等, 2018 ).如在洞穴弱光带一般形成洞穴石刺、向光钟乳石、洞口叠层石等, 这类形态有很强的方向性(
王福星等, 1998 ).苔藓植物在水生环境中也常常形成多样的沉积形态, 如凤尾藓(
Fissidens )钙华、石灰藓(
Hydrogonium )钙华形成的溪流瀑布沉积, 泽藓(
Philonotis )钙华、水灰藓(
Hygrohypnum )钙华形成的河流瀑布沉积, 地钱-真藓(
Marchantia-Bryum )钙华等形成的石灰华坝沉积, 以及净口藓(
Gymnostomum )钙华等形成的瀑布洞穴沉积(
张朝晖和陈家宽, 2007 ).同时, 也有研究发现, 水流环境也是影响苔藓钙华沉积的重要因素, 苔藓与钙华层之间的水流速度与钙华沉积速率呈正相关关系(
Wang et al ., 2014 ).钙华作为大陆古气候的潜在档案,
Yan等(2017) 利用碳氧稳定同位素比率(
δ 13 C和
δ 18 O), 研究钙华在沉积过程中碳、氧的循环规律, 分析认为, 钙华沉积与季节更替引起的气温、降水等的变化密切相关.目前对于苔藓植物沉积作用的研究主要集中在沉积形态以及成土量方面, 对于苔藓植物沉积速率, 对岩溶地区成土过程中的贡献, 以及岩溶沉积作用对生境的改造和环境对岩溶沉积作用的影响等方面的研究还未见报道. ...
中国昆明地区岩溶洞穴洞口带苔藓植物研究
1
2004
... 苔藓植物沉积作用的另一表现是石灰华或钙华(
张捷, 1993 ;
Kleinteich et al ., 2017 ).钙华形成的物理化学过程实质是CaCO
3 的沉淀与CO
2 的脱气.苔藓植物在钙华形成过程中为CaCO
3 晶体提供了稳定的生长发育基础, 并且苔藓植物的形态结构控制着沉积物的形态变化(
Mili?a et al ., 2006 ), 且它们的发育形态随沉积环境而变化(
张朝晖等, 2004 ;
郭云等, 2018 ).如在洞穴弱光带一般形成洞穴石刺、向光钟乳石、洞口叠层石等, 这类形态有很强的方向性(
王福星等, 1998 ).苔藓植物在水生环境中也常常形成多样的沉积形态, 如凤尾藓(
Fissidens )钙华、石灰藓(
Hydrogonium )钙华形成的溪流瀑布沉积, 泽藓(
Philonotis )钙华、水灰藓(
Hygrohypnum )钙华形成的河流瀑布沉积, 地钱-真藓(
Marchantia-Bryum )钙华等形成的石灰华坝沉积, 以及净口藓(
Gymnostomum )钙华等形成的瀑布洞穴沉积(
张朝晖和陈家宽, 2007 ).同时, 也有研究发现, 水流环境也是影响苔藓钙华沉积的重要因素, 苔藓与钙华层之间的水流速度与钙华沉积速率呈正相关关系(
Wang et al ., 2014 ).钙华作为大陆古气候的潜在档案,
Yan等(2017) 利用碳氧稳定同位素比率(
δ 13 C和
δ 18 O), 研究钙华在沉积过程中碳、氧的循环规律, 分析认为, 钙华沉积与季节更替引起的气温、降水等的变化密切相关.目前对于苔藓植物沉积作用的研究主要集中在沉积形态以及成土量方面, 对于苔藓植物沉积速率, 对岩溶地区成土过程中的贡献, 以及岩溶沉积作用对生境的改造和环境对岩溶沉积作用的影响等方面的研究还未见报道. ...
含辅助官能团类有机酸对磷石膏晶体的影响
1
2018
... 苔藓植物对岩石的溶蚀作用原理与过程实质是岩石中矿物与苔藓植物之间的作用过程(
图2 ).苔藓植物因呼吸作用形成的H
2 CO
3 和代谢分泌物与岩石中的矿物发生酸化、碱化、氧化还原、螯合等反应, 随着反应过程不断推进, 岩石矿物成分发生变化, Ca、镁(Mg)、钠(Na)、钾(K)、铁(Fe)、铝(Al)、硅(Si)等从碳酸盐、硅酸盐、铝酸盐、硫化物、氧化物中溶出, 破坏原岩晶体结构, 达到溶蚀岩石, 促使风化的目的(
Street et al ., 2013 ;
Li et al ., 2015 ;
Lammers et al ., 2017 ).这种岩溶驱动力的大小因植物形态特征、岩石性质与生存环境改变而发生变化.有研究表明, 碳酸盐岩溶蚀过程中, 生物成因的CO
2 占溶蚀强度的49.26%, 有机酸占37.11% (
张楷燕, 2017 ).苔藓植物溶蚀作用过程释放的矿物离子和其他元素部分被苔藓植物拦截形成岩溶沉积物; 部分被苔藓植物吸收利用, 转化为其他物质存储于植物体内; 其余则随降雨冲刷而流失, 进入其他生态系统.关于苔藓植物呼吸作用和其分泌物引起的岩溶作用驱动力、效率及其与苔藓植物生活型、种间差异、生境特征之间的相互关系仍未引起人们的关注, 对苔藓植物溶蚀作用过程和机理、苔藓植物对岩石中矿物营养元素释放规律的影响, 以及这些矿物质在岩溶沉积过程中的形态转化过程及其与生境间的相互作用关系等的研究还未见报道, 这也正是揭示苔藓植物岩溶作用机理的关键点.岩石的溶解速率与有机酸的酸性和溶蚀能力密切相关(
刘胜亮等, 2017 ), 然而有机酸的理化特性取决于其官能团的属性(
赵志曼等, 2018 ).因此, 今后需结合有机化学、气相色谱质谱(GC-MS)、核磁共振(NMR)、液相色谱(HPLC)等技术手段深入研究苔藓植物的溶蚀过程与机理. ...
含辅助官能团类有机酸对磷石膏晶体的影响
1
2018
... 苔藓植物对岩石的溶蚀作用原理与过程实质是岩石中矿物与苔藓植物之间的作用过程(
图2 ).苔藓植物因呼吸作用形成的H
2 CO
3 和代谢分泌物与岩石中的矿物发生酸化、碱化、氧化还原、螯合等反应, 随着反应过程不断推进, 岩石矿物成分发生变化, Ca、镁(Mg)、钠(Na)、钾(K)、铁(Fe)、铝(Al)、硅(Si)等从碳酸盐、硅酸盐、铝酸盐、硫化物、氧化物中溶出, 破坏原岩晶体结构, 达到溶蚀岩石, 促使风化的目的(
Street et al ., 2013 ;
Li et al ., 2015 ;
Lammers et al ., 2017 ).这种岩溶驱动力的大小因植物形态特征、岩石性质与生存环境改变而发生变化.有研究表明, 碳酸盐岩溶蚀过程中, 生物成因的CO
2 占溶蚀强度的49.26%, 有机酸占37.11% (
张楷燕, 2017 ).苔藓植物溶蚀作用过程释放的矿物离子和其他元素部分被苔藓植物拦截形成岩溶沉积物; 部分被苔藓植物吸收利用, 转化为其他物质存储于植物体内; 其余则随降雨冲刷而流失, 进入其他生态系统.关于苔藓植物呼吸作用和其分泌物引起的岩溶作用驱动力、效率及其与苔藓植物生活型、种间差异、生境特征之间的相互关系仍未引起人们的关注, 对苔藓植物溶蚀作用过程和机理、苔藓植物对岩石中矿物营养元素释放规律的影响, 以及这些矿物质在岩溶沉积过程中的形态转化过程及其与生境间的相互作用关系等的研究还未见报道, 这也正是揭示苔藓植物岩溶作用机理的关键点.岩石的溶解速率与有机酸的酸性和溶蚀能力密切相关(
刘胜亮等, 2017 ), 然而有机酸的理化特性取决于其官能团的属性(
赵志曼等, 2018 ).因此, 今后需结合有机化学、气相色谱质谱(GC-MS)、核磁共振(NMR)、液相色谱(HPLC)等技术手段深入研究苔藓植物的溶蚀过程与机理. ...
荒漠生物结皮中藻类和苔藓植物研究进展
1
2009
... 苔藓植物常以丛集型、交织型、匍匐型等着生于岩面(
图1 ), 假根根系不发达, 只起固着作用(
娄玉霞, 2013 ), 其特殊的叶表面结构和细胞特征(
娄红祥, 2012 ), 使苔藓具有耐高温、抗旱、蓄水能力强、保湿固土等功能(
Proctor & Pence, 2002 ;
Krol et al ., 2003 ;
Zhang et al ., 2017 ).
曹建华和袁道先(1999) 研究发现, 当岩石表面有苔藓植物时, 其蒸发失水和主动吸水时间比裸岩分别延长了48%和57%, 持水量提高了16.6倍, 分析认为苔藓植物可以延缓岩面水分蒸发时间, 增加岩石水化学作用时间, 但是苔藓植物的这种蓄水保湿能力存在明显的种间差异(
申家琛等, 2017 ;
张显强等, 2018a ).苔藓植物参与岩溶作用的另一大优势, 是其叶表面角质层不发育, 且阳离子交换位点多(
吴玉环等, 2001 ), 能够吸附多种阳离子, 这些阳离子在苔藓植物体内被转化为无机盐、有机盐以及其他形式的化合物储存于组织中.苔藓植物除了以生物化学方式对岩石进行溶蚀之外, 还可以通过物理机械作用破坏岩石.例如, 苔藓植物的假根可以伸入岩石裂隙, 对岩石有一定的破坏作用(
Jackson, 2015 ).再者, 当气候干旱时, 苔藓植物在卷缩过程中会拔起岩面的一些小颗粒, 从而加速了岩石被破坏和成土速度(
连宾等, 2008 ).此外, 苔藓植物常与藻类、地衣、微生物等共生形成生物结皮, 它们之间的相互关系以及因气候因素引起的干湿交替和冻融作用对岩石也具有一定的破坏性(
郑云普等, 2009 ).例如, 菌类的菌丝可以伸入岩石晶体结构中破坏岩石, 细菌与真菌形成的配合基能够络合三价铁离子, 细菌分泌的柠檬酸、乙酸以及地衣分泌的地衣酸(
连宾等, 2011 ), 可以溶解岩石中的矿物(
Won-Pyo et al ., 2016 ;
Wang et al ., 2017 ).因此在研究苔藓植物岩溶作用时, 应明确苔藓植物自身及其与其他生物形成的生物结皮两者之间的关系.目前, 有关岩面苔藓植物功能性状的研究主要集中在小范围内特定生境中的苔藓植物研究, 无法代表岩溶生态系统苔藓植物的整体情况.对于大尺度空间上, 因生境异质性高而造成相同物种在形态结构、光合特征及其钙(Ca)、碳(C)、氮(N)、磷(P) 等元素的代谢调节机制等方面的差异性的研究还未引起人们的广泛关注.岩溶生态系统的物质基础相对贫乏, 具有“缺氮少磷钙丰富”的特征.因此, 研究Ca、C、N、P等元素在苔藓植物体内的代谢机制对揭示岩溶生态系统苔藓植物和其他植物的生物多样性维持机制, 及其岩溶作用特征具有重要的科学 意义. ...
荒漠生物结皮中藻类和苔藓植物研究进展
1
2009
... 苔藓植物常以丛集型、交织型、匍匐型等着生于岩面(
图1 ), 假根根系不发达, 只起固着作用(
娄玉霞, 2013 ), 其特殊的叶表面结构和细胞特征(
娄红祥, 2012 ), 使苔藓具有耐高温、抗旱、蓄水能力强、保湿固土等功能(
Proctor & Pence, 2002 ;
Krol et al ., 2003 ;
Zhang et al ., 2017 ).
曹建华和袁道先(1999) 研究发现, 当岩石表面有苔藓植物时, 其蒸发失水和主动吸水时间比裸岩分别延长了48%和57%, 持水量提高了16.6倍, 分析认为苔藓植物可以延缓岩面水分蒸发时间, 增加岩石水化学作用时间, 但是苔藓植物的这种蓄水保湿能力存在明显的种间差异(
申家琛等, 2017 ;
张显强等, 2018a ).苔藓植物参与岩溶作用的另一大优势, 是其叶表面角质层不发育, 且阳离子交换位点多(
吴玉环等, 2001 ), 能够吸附多种阳离子, 这些阳离子在苔藓植物体内被转化为无机盐、有机盐以及其他形式的化合物储存于组织中.苔藓植物除了以生物化学方式对岩石进行溶蚀之外, 还可以通过物理机械作用破坏岩石.例如, 苔藓植物的假根可以伸入岩石裂隙, 对岩石有一定的破坏作用(
Jackson, 2015 ).再者, 当气候干旱时, 苔藓植物在卷缩过程中会拔起岩面的一些小颗粒, 从而加速了岩石被破坏和成土速度(
连宾等, 2008 ).此外, 苔藓植物常与藻类、地衣、微生物等共生形成生物结皮, 它们之间的相互关系以及因气候因素引起的干湿交替和冻融作用对岩石也具有一定的破坏性(
郑云普等, 2009 ).例如, 菌类的菌丝可以伸入岩石晶体结构中破坏岩石, 细菌与真菌形成的配合基能够络合三价铁离子, 细菌分泌的柠檬酸、乙酸以及地衣分泌的地衣酸(
连宾等, 2011 ), 可以溶解岩石中的矿物(
Won-Pyo et al ., 2016 ;
Wang et al ., 2017 ).因此在研究苔藓植物岩溶作用时, 应明确苔藓植物自身及其与其他生物形成的生物结皮两者之间的关系.目前, 有关岩面苔藓植物功能性状的研究主要集中在小范围内特定生境中的苔藓植物研究, 无法代表岩溶生态系统苔藓植物的整体情况.对于大尺度空间上, 因生境异质性高而造成相同物种在形态结构、光合特征及其钙(Ca)、碳(C)、氮(N)、磷(P) 等元素的代谢调节机制等方面的差异性的研究还未引起人们的广泛关注.岩溶生态系统的物质基础相对贫乏, 具有“缺氮少磷钙丰富”的特征.因此, 研究Ca、C、N、P等元素在苔藓植物体内的代谢机制对揭示岩溶生态系统苔藓植物和其他植物的生物多样性维持机制, 及其岩溶作用特征具有重要的科学 意义. ...
论原始土壤的成土过程
1
1995
... 岩溶地区土壤的形成和发育是碳酸岩长期溶蚀、风化与生物富集过程的结果, 其一方面受岩石结构、性质影响, 另一方面也与植被作用密切相关(
张美良和邓自强, 1994 ).研究表明, 碳酸盐岩的成土速率与岩石溶蚀速率成正比(
曹建华等, 2004 ), 与酸不溶物含量密切相关(
曹沁智等, 2018 ).苔藓植物殖居于岩面, 假根系以及植物体的干湿交替冻融现象所产生的物理机械力作用于岩石, 产生岩石碎屑.同时, 有机物与矿物离子络合常常形成难溶物, 这些难溶物与苔藓及其生物结皮死亡残体黏结在一起, 不仅增加了土壤沉积量, 而且促进有机质积累, 增加土壤养分(
朱显谟, 1995 ;
刘天雷等, 2017 ).苔藓植物通过影响下垫面土壤湿度, 促进土壤对岩石的岩溶作用, 间接地影响沉积过程.同时, 苔藓植物的吸附拦截功能对防治岩石表面水土流失具有重要作用(
李军峰等, 2013 ;
贾少华等, 2014 ;
张显强等, 2018b ).苔藓植物的溶蚀作用为岩面土壤形成提供了原始物质, 关于苔藓对土壤沉积的贡献率与苔藓植物作用下岩面土壤的形成过程, 以及在此过程中岩石的物质、能量转化和土壤养分的变化等还未见报道. ...
论原始土壤的成土过程
1
1995
... 岩溶地区土壤的形成和发育是碳酸岩长期溶蚀、风化与生物富集过程的结果, 其一方面受岩石结构、性质影响, 另一方面也与植被作用密切相关(
张美良和邓自强, 1994 ).研究表明, 碳酸盐岩的成土速率与岩石溶蚀速率成正比(
曹建华等, 2004 ), 与酸不溶物含量密切相关(
曹沁智等, 2018 ).苔藓植物殖居于岩面, 假根系以及植物体的干湿交替冻融现象所产生的物理机械力作用于岩石, 产生岩石碎屑.同时, 有机物与矿物离子络合常常形成难溶物, 这些难溶物与苔藓及其生物结皮死亡残体黏结在一起, 不仅增加了土壤沉积量, 而且促进有机质积累, 增加土壤养分(
朱显谟, 1995 ;
刘天雷等, 2017 ).苔藓植物通过影响下垫面土壤湿度, 促进土壤对岩石的岩溶作用, 间接地影响沉积过程.同时, 苔藓植物的吸附拦截功能对防治岩石表面水土流失具有重要作用(
李军峰等, 2013 ;
贾少华等, 2014 ;
张显强等, 2018b ).苔藓植物的溶蚀作用为岩面土壤形成提供了原始物质, 关于苔藓对土壤沉积的贡献率与苔藓植物作用下岩面土壤的形成过程, 以及在此过程中岩石的物质、能量转化和土壤养分的变化等还未见报道. ...
 ,1,*, 冉景丞31.
,1,*, 冉景丞31. 
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT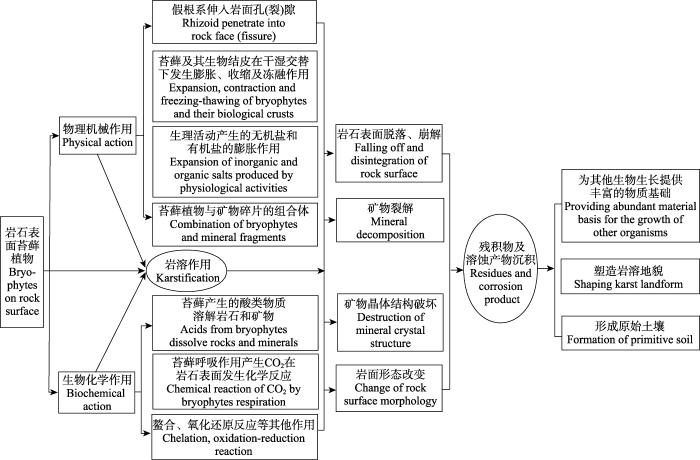 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT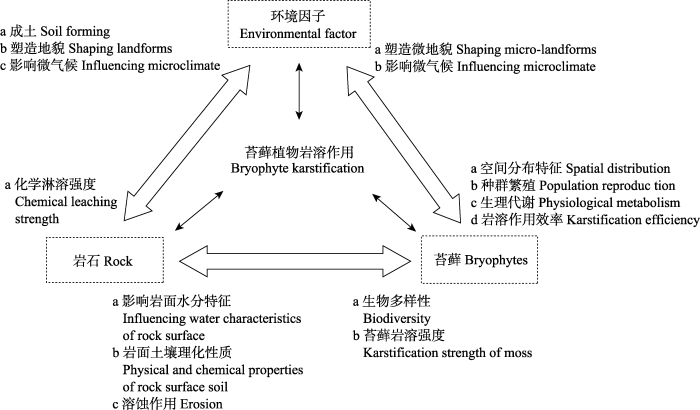 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}