Origin and distribution of neutral sugars in soils
Cheng-Zhu LIU1,2, Juan JIA1,2, Guo-Hua DAI1, Tian MA1,2, Xiao-Juan FENG,,1,2,*1 State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China 2 University of Chinese Academy of Sciences, Beijing 100049, China
Supported by the Chinese National Key Development Program for Basic Research(2015CB954201) the National Natural Science Foundation of China(41773067) the National Natural Science Foundation of China(41422304) the International Partnership Program of Chinese Academy of Sciences(151111KYSB20160014)
Abstract Carbohydrates are important components of soil organic matter, which can be decomposed to different types of monosaccharides. Neutral monosaccharides in the soil are also called neutral sugars, including xylose, ribose, arabinose, glucose, galactose, mannose, fucose and rhamnose. Among them, plant-derived sugars mainly include pentoses, such as xylose and arabinose, while microbial-derived sugars mainly consist of hexoses including galactose, mannose, fucose and rhamnose. Generally, the ratios of hexoses to pentoses are used to evaluate the contribution of microbial- versus plant-derived sugars. Neutral sugars are the main carbon and energy resources for soil microorganisms and play a vital role in aggregates formation. In this study, we review studies about neutral sugars in soils over the past 30 years and compare different methods for neutral sugar analysis. Furthermore, we compare the distribution patterns and turnover of soil neutral sugars across diverse land-use regimes, different soil density and particle size fractions and their influencing factors. The lowest neutral sugar content is found in arable soils compared with other four land-use types (coniferous forests, deciduous forests, shrublands and grasslands) in terms of absolute and relative contents. No significant difference is observed for the (galactose + mannose)/?(arabinose + xylose)(GM/AX) ratios across the five land-use regimes. Nevertheless, the ratio of (rhamnose + fucose)/(arabinose + xylose)(RF/AX) indicates that microbially derived neutral sugars are more abundant in the soils of grasslands than coniferous forests or farmlands. The heavy fraction is characterized by an enrichment of microbial neutral sugars but a lower content of total neutral sugars compared to the light fraction. Concerning the distribution of neutral sugars across different soil size fractions (or aggregates), the microbial-derived neutral sugars are more abundant in the clay fraction (or microaggregates). As for the factors affecting neutral sugar content and distribution, many studies have focused on the human disturbances like agriculture and grazing, while the influence of environmental factors such as temperature, precipitation is poorly investigated. Keywords:soil;neutral sugars;origin;distribution;influencing factor
产率高; 不会破坏单糖结构; 具有挥发性, 可通过旋转蒸发去除; 水解的多糖主要为半纤维素 High yields; Not destructive to monosaccharides; TFA is volatile and can be easily removed by evaporation; Hydrolysis products are mainly released from hemicellulose
虽然不断有研究对各类衍生化方法进行改进, 但是衍生化仍旧是一个繁琐耗时的过程, 相比GC-MS检测方法, HPLC和HPAEC-PDA检测法较为简单, 在定量前无需衍生化。但这两种检测方法都需要特殊的仪器或检测器, 并且检测敏感度、准确度和精度较低, 因此目前没有广泛应用(Zhang et al., 2013; Wang et al., 2017)。
Fig. 1Content and distribution of neutral sugars across different land-use regimes in the top soils (Nierop et al., 2001; Spielvogel et al., 2007; Eder et al., 2010; Rumpel et al., 2010; Zhao et al., 2014; Conti et al., 2016; Cui et al., 2016; Wang et al., 2016; Creme et al., 2017; Llorente et al., 2017; Evgrafova et al., 2018; Zhu et al., 2018). A, Neutral sugar absolute content. B, Neutral sugar relative content. C, GM/AX ((galactose + mannose)/(arabinose + xylose)). D, RF/AX ((rhamnose + fucose)/(arabinose + xylose)). The upper and lower end of boxes denote the 0.25 and 0.75 percentiles, respectively. The solid bar in the box mark the median of each dataset. The circles indicate outliers of each dataset. Different lowercase letters indicate differences in various land-use regimes (p < 0.05). n = 8, 25, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the A, C). n = 8, 22, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the B). n = 3, 19, 4, 8, 27 (from deciduous, coniferous, shrub, grassland to crops in the D). OC, soil organic carbon.
AbdelrahmanHM, OlkDC, DinnesD, VentrellaD, MianoT, CocozzaC ( 2016). Occurrence and abundance of carbohydrates and amino compounds in sequentially extracted labile soil organic matter fractions Journal of Soils and Sediments, 16, 2375-2384. [本文引用: 1]
AmelungW, CheshireMV, GuggenbergerG ( 1996). Determination of neutral and acidic sugars in soil by capillary gas-liquid chromatography after trifluoroacetic acid hydrolysis Soil Biology & Biochemistry, 28, 1631-1639. [本文引用: 11]
AndrewsMA ( 1989). Capillary gas-chromatographic analysis of monosaccharides: Improvements and comparisons using trifluoroacetylation and trimethylsilylation of sugar O-benzyl- and O-methyl-oximes Carbohydrate Research, 194, 1-19. [本文引用: 1]
AngersDA, MehuysGR ( 1989). Effects of cropping on carbohydrate content and water-stable aggregation of a clay soil Canadian Journal of Soil Science, 69, 373-380. [本文引用: 1]
AngersDA, NadeauP, MehuysGR ( 1988). Determination of carbohydrate-composition of soil hydrolysates by high- performance liquid-chromatography Journal of Chromatography, 454, 444-449. [本文引用: 2]
Barron-GaffordG, MartensD, GrieveK, BielK, KudeyarovV, MclainJET, LipsonD, MurthyR ( 2005). Growth of eastern cottonwoods (Populus deltoides) in elevated CO2 stimulates stand-level respiration and rhizodeposition of carbohydrates, accelerates soil nutrient depletion, yet stimulates above- and belowground biomass production Global Change Biology, 11, 1220-1233. [本文引用: 1]
BaslerA, DippoldM, HelfrichM, DyckmansJ ( 2015a). Microbial carbon recycling: An underestimated process controlling soil carbon dynamics―Part 1: A long-term laboratory incubation experiment Biogeosciences, 12, 5929-5940. [本文引用: 2]
BaslerA, DippoldM, HelfrichM, DyckmansJ ( 2015b). Microbial carbon recycling: An underestimated process controlling soil carbon dynamics―Part 2: A C3-C4 vegetation change field labelling experiment Biogeosciences, 12, 6291-6299. [本文引用: 1]
BaslerA, DyckmansJ ( 2013). Compound-specific δ13C analysis of monosaccharides from soil extracts by high- performance liquid chromatography/isotope ratio mass spectrometry Rapid Communications in Mass Spectrometry , 27, 2546-2550. [本文引用: 1]
BiernathC, FischerH, KuzyakovY ( 2008). Root uptake of N-containing and N-free low molecular weight organic substances by maize: A 14C/ 15N tracer study Soil Biology & Biochemistry, 40, 2237-2245. [本文引用: 1]
BischoffN, MikuttaR, ShibistovaO, DohrmannR, HerdtleD, GerhardL, FritzscheF, PuzanovA, SilantevaM, GrebennikovaA, GuggenbergerG ( 2018). Organic matter dynamics along a salinity gradient in Siberian steppe soils Biogeosciences, 15, 13-29. [本文引用: 2]
BlackGE, FoxA ( 1996). Recent progress in the analysis of sugar monomers from complex matrices using chromatography in conjunction with mass spectrometry or stand- alone tandem mass spectrometry Journal of Chromatography A, 720, 51-60. [本文引用: 2]
BlagodatskayaE, KuzyakovY ( 2013). Active microorganisms in soil: Critical review of estimation criteria and approaches Soil Biology & Biochemistry, 67, 192-211. [本文引用: 1]
BockM, GlaserB, MillarN ( 2007). Sequestration and turnover of plant- and microbially derived sugars in a temperate grassland soil during 7 years exposed to elevated atmospheric pCO2 Global Change Biology, 13, 478-490. [本文引用: 1]
BrugginkC, MaurerR, HerrmannH, CavalliS, HoeflerF ( 2005). Analysis of carbohydrates by anion exchange chromatography and mass spectrometry Journal of Chromatography A, 1085, 104-109. [本文引用: 2]
CheshireMV (1979). Nature and Origin of Carbohydrates in Soils. Academic Press, London. [本文引用: 2]
CheshireMV, RussellJD, FraserAR, BracewellJM, RobertsonsGW, Benzing-PurdieLM, RatcliffeCI, RipmeesterJA, GoodmanBA ( 1992). Nature of soil carbohydrate and its association with soil humic substances Journal of Soil Science, 43, 359-373. [本文引用: 1]
ContiG, KowaljowE, BaptistF, RumpelC, CuchiettiA, Perez HarguindeguyN, DiazS ( 2016). Altered soil carbon dynamics under different land-use regimes in subtropical seasonally-dry forests of central Argentina Plant and Soil, 403, 375-387. [本文引用: 5]
CremeA, ChabbiA, GastalF, RumpelC ( 2017). Biogeochemical nature of grassland soil organic matter under plant communities with two nitrogen sources Plant and Soil, 415, 189-201. [本文引用: 2]
CuiLF, LiangC, DuncanDS, BaoXL, XieHT, HeHB, WickingsK, ZhangXD, ChenFS ( 2016). Impacts of vegetation type and climatic zone on neutral sugar distribution in natural forest soils Geoderma, 282, 139-146. [本文引用: 4]
DerrienD, MarolC, BalabaneM, BalesdentJ (2006). The turnover of carbohydrate carbon in a cultivated soil estimated by 13C natural abundances European Journal of Soil Science, 57, 547-557. [本文引用: 2]
DerrienD, MarolC, BalesdentJ (2007). Microbial biosyntheses of individual neutral sugars among sets of substrates and soils Geoderma, 139, 190-198. [本文引用: 2]
EderE, SpielvogelS, KoelblA, AlbertG, K?gel-KnabnerI ( 2010). Analysis of hydrolysable neutral sugars in mineral soils: Improvement of alditol acetylation for gas chromatographic separation and measurement Organic Geochemistry, 41, 580-585. [本文引用: 5]
EvgrafovaA, De La HayeTR, HaaseI, ShibistovaO, GuggenbergerG, TananaevN, SauheitlL, SpielvogelS ( 2018). Small-scale spatial patterns of soil organic carbon and nitrogen stocks in permafrost-affected soils of northern Siberia Geoderma, 329, 91-107. [本文引用: 2]
FarhadiA, KeshavarzianA, FieldsJZ, SheikhM, BananA ( 2006). Resolution of common dietary sugars from probe sugars for test of intestinal permeability using capillary column gas chromatography Journal of Chromatography B, 836, 63-68. [本文引用: 1]
FellerC, BeareMH ( 1997). Physical control of soil organic matter dynamics in the tropics Geoderma, 79, 69-116. [本文引用: 1]
FischerH, IngwersenJ, KuzyakovY ( 2010). Microbial uptake of low-molecular-weight organic substances out-competes sorption in soil European Journal of Soil Science, 61, 504-513. [本文引用: 1]
GentschN, MikuttaR, AlvesRJE, BartaJ, ?apekP, GittelA, HugeliusG, KuhryP, LashchinskiyN, PalmtagJ ( 2015a). Storage and transformation of organic matter fractions in cryoturbated permafrost soils across the Siberian Arctic Biogeosciences Discussions, 12, 2697-2743. [本文引用: 1]
GentschN, MikuttaR, ShibistovaO, WildB, SchneckerJ, RichterA, UrichT, GittelA, ?antr??kováH, BártaJ ( 2015b). Properties and bioavailability of particulate and mineral-associated organic matter in Arctic permafrost soils European Journal of Soil Science, 66, 722-734. [本文引用: 1]
GleixnerG, PoirierN, BolR, BalesdentJ ( 2002). Molecular dynamics of organic matter in a cultivated soil Organic Geochemistry, 33, 357-366. [本文引用: 2]
GuanZH, LiXG, WangL, MouXM, KuzyakovY ( 2018). Conversion of Tibetan grasslands to croplands decreases accumulation of microbially synthesized compounds in soil Soil Biology & Biochemistry, 123, 10-20. [本文引用: 2]
GuggenbergerG, ChristensenBT, ZechW ( 1994). Land-use effects on the composition of organic matter in particle-?size separates of soil: I. Lignin and carbohydrate signature European Journal of Soil Science, 45, 449-458. [本文引用: 2]
GuninaA, KuzyakovY (2015). Sugars in soil and sweets for microorganisms: Review of origin, content, composition and fate Soil Biology & Biochemistry, 90, 87-100. [本文引用: 10]
HamadaR, OnoA ( 1984). Determination of carbohydrates in hydrolysates of volcanic ash soil by liquid chromatography with fluorescence spectroscopy Soil Science & Plant Nutrition, 30, 145-150. [本文引用: 1]
HuS, ColemanDC, BeareMH, HendrixPF ( 1995a). Soil carbohydrates in aggrading and degrading agroecosystems: Influences of fungi and aggregates Agriculture Ecosystems & Environment, 54, 77-88. [本文引用: 1]
HuS, ColemanDC, HendrixPF, BeareMH ( 1995b). Biotic manipulation effects on soil carbohydrates and microbial biomass in a cultivated soil Soil Biology & Biochemistry, 27, 1127-1135. [本文引用: 1]
JolivetC, AngersDA, ChantignyMH, AndreuxF, ArrouaysD ( 2006). Carbohydrate dynamics in particle-size fractions of sandy spodosols following forest conversion to maize cropping Soil Biology & Biochemistry, 38, 2834-2842. [本文引用: 2]
KleberM, EusterhuesK, KeiluweitM, MikuttaC, MikuttaR, NicoPS ( 2015). Chapter One―Mineral-organic associations: Formation, properties, and relevance in soil environments Advances in Agronomy, 130, 1-140. [本文引用: 1]
K?gel-KnabnerI ( 2002). The macromolecular organic composition of plant and microbial residues as inputs to soil organic matter Soil Biology & Biochemistry, 34, 139-162. [本文引用: 8]
K?gel-KnabnerI, AmelungW ( 2014). Dynamics, chemistry, and preservation of organic matter in soils Treatise on Geochemistry, 13, 157-215. [本文引用: 1]
KuzyakovY ( 2010). Priming effects: Interactions between living and dead organic matter Soil Biology & Biochemistry, 42, 1363-1371. [本文引用: 1]
KuzyakovY, DomanskiG ( 2000). Carbon input by plants into the soil. Review Journal of Plant Nutrition and Soil Science, 163, 421-431. [本文引用: 1]
KuzyakovY, JonesDL ( 2006). Glucose uptake by maize roots and its transformation in the rhizosphere Soil Biology & Biochemistry, 38, 851-860. [本文引用: 1]
Larré-LarrouyMC, BlanchartE, AlbrechtA, FellerC ( 2004). Carbon and monosaccharides of a tropical vertisol under pasture and market-gardening: Distribution in secondary organomineral separates Geoderma, 119, 163-178. [本文引用: 5]
Larré-LarrouyMC, FellerC ( 1997). Determination of carbohydrates in two ferrallitic soils: Analysis by capillary gas chromatography after derivatization by silylation Soil Biology & Biochemistry, 29, 1585-1589. [本文引用: 1]
LlorenteM, GlaserB, TurrionMB ( 2017). Effect of land use change on contents and distribution of monosacharides within density fractions of calcareous soil Soil Biology & Biochemistry, 107, 260-268. [本文引用: 6]
MartensDA, FrankenbergerWT ( 1993). Soil saccharide extraction and detection Plant and Soil, 149, 145-147. [本文引用: 1]
MartensDA, ReedyTE, LewisDT ( 2004). Soil organic carbon content and composition of 130-year crop, pasture and forest land-use managements Global Change Biology, 10, 65-78. [本文引用: 3]
MastrolonardoG, RumpelC, ForteC, DoerrSH, CertiniG ( 2015). Abundance and composition of free and aggregate- occluded carbohydrates and lignin in two forest soils as affected by wildfires of different severity Geoderma, 245, 40-51. [本文引用: 1]
MiltnerA, BombachP, Schmidt-BrückenB, K?stnerM ( 2012). SOM genesis: Microbial biomass as a significant source Biogeochemistry, 111, 41-55. [本文引用: 1]
MoersMEC, JonesDM, EakinPA, FallickAE, GriffithsH, LarterSR ( 1993). Carbohydrate diagenesis in hypersaline environments: Application of GC-IRMS to the stable isotope analysis of derivatized saccharides from surficial and buried sediments Organic Geochemistry, 20, 927-933. [本文引用: 1]
MurataT, TanakaH, YasueS, HamadaR, SakagamiK, KurokawaY ( 1999). Seasonal variations in soil microbial biomass content and soil neutral sugar composition in grassland in the Japanese Temperate Zone Applied Soil Ecology, 11, 253-259. [本文引用: 1]
NacroHB, Larré-larrouyMC, FellerC, AbbadieL ( 2005). Hydrolysable carbohydrate in tropical soils under adjacent forest and savanna vegetation in Lamto, C?te d’Ivoire Soil Research, 43, 705-711. [本文引用: 1]
NavarreteIA, TsutsukiK ( 2008). Land-use impact on soil carbon, nitrogen, neutral sugar composition and related chemical properties in a degraded Ultisol in Leyte, Philippines Soil Science & Plant Nutrition, 54, 321-331. [本文引用: 1]
NieropKGJ, Van LagenB, BuurmanP ( 2001). Composition of plant tissues and soil organic matter in the first stages of a vegetation succession Geoderma, 100, 1-24. [本文引用: 4]
OadesJM ( 1984). Soil organic matter and structural stability: Mechanisms and implications for management Plant and Soil, 76, 319-337. [本文引用: 3]
OadesJM, KirkmanMA, WagnerGH ( 1970). The use of gas-liquid chromatography for the determination of sugars extracted from soils by sulfuric acid Soil Science Society of America Journal, 34, 230-235. [本文引用: 1]
PhilbenM, HolmquistJ, MacdonaldG, DuanD, KaiserK, BennerR ( 2015). Temperature, oxygen, and vegetation controls on decomposition in a James Bay peatland Global Biogeochemical Cycles, 29, 729-743. [本文引用: 1]
Phuong-ThiN, RumpelC, Thu-ThuyD, JouquetP ( 2012). The effect of earthworms on carbon storage and soil organic matter composition in tropical soil amended with compost and vermicompost Soil Biology & Biochemistry, 50, 214-220. [本文引用: 2]
PrietzelJ, DechampsN, SpielvogelS ( 2012). Analysis of non-cellulosic polysaccharides helps to reveal the history of thick organic surface layers on calcareous Alpine soils Plant and Soil, 365, 93-114. [本文引用: 1]
PugetP, AngersDA, ChenuC ( 1999). Nature of carbohydrates associated with water-stable aggregates of two cultivated soils Soil Biology & Biochemistry, 31, 55-63. [本文引用: 1]
Ruiz-MatuteAI, Hernandez-HernandezO, Rodriguez-SanchezS, SanzML, Martinez-CastroI ( 2011). Derivatization of carbohydrates for GC and GC-MS analyses Journal of Chromatography B-Analytical Technologies in the Biomedical and Life Sciences, 879, 1226-1240. [本文引用: 3]
RumpelC, DignacMF ( 2006). Gas chromatographic analysis of monosaccharides in a forest soil profile: Analysis by gas chromatography after trifluoroacetic acid hydrolysis and reduction-acetylation Soil Biology & Biochemistry, 38, 1478-1481. [本文引用: 5]
RumpelC, EusterhuesK, K?gel-KnabnerI ( 2010). Non- cellulosic neutral sugar contribution to mineral associated organic matter in top- and subsoil horizons of two acid forest soils Soil Biology & Biochemistry, 42, 379-382. [本文引用: 4]
SixJ, ConantRT, PaulEA, PaustianK ( 2002). Stabilization mechanisms of soil organic matter: Implications for C-saturation of soils Plant and Soil, 241, 155-176. [本文引用: 1]
SollinsP, HomannP, CaldwellBA ( 1996). Stabilization and destabilization of soil organic matter: Mechanisms and controls Geoderma, 74, 65-105. [本文引用: 1]
SowdenFJ, IvarsonKC ( 1962). Methods for the analysis of carbohydrate material in soil: 2 Soil Science, 94, 340-344. [本文引用: 1]
SpielvogelS, PrietzelJ, K?gel-KnabnerI ( 2007). Changes of lignin phenols and neutral sugars in different soil types of a high-elevation forest ecosystem 25 years after forest dieback Soil Biology & Biochemistry, 39, 655-668. [本文引用: 5]
SteffensM, K?lblA, K?gel-KnabnerI ( 2009). Alteration of soil organic matter pools and aggregation in semi-arid steppe topsoils as driven by organic matter input European Journal of Soil Science, 60, 198-212. [本文引用: 1]
TakeuchiM, TakasakiS, InoueN, KobataA ( 1987). Sensitive method for carbohydrate-composition analysis of glycoproteins by high-performance liquid- chromatography Journal of Chromatography, 400, 207-213. [本文引用: 1]
TanakaH, HamadaR, KondohA, SakagamiK (1990). Determination of component sugars in soil organic-matter by HPLC Zentralblatt für Mikrobiologie, 145, 621-628. [本文引用: 3]
ThompsonTL, ZaadyE, HuanchengP, WilsonTB, MartensDA ( 2006). Soil C and N pools in patchy shrublands of the Negev and Chihuahuan Deserts Soil Biology & Biochemistry, 38, 1943-1955. [本文引用: 1]
TianQX, ZhangB, HeHB, ZhangXD, ChenWX ( 2013). Distribution pattern of neutral sugar in forest soils along an altitude gradient in Changbai Mountains, Northeast China Chinese Journal of Applied Ecology, 24, 1777-1783. [本文引用: 1]
TrouveC, DisnarJR, MariottiA, GuilletB ( 1996). Changes in the amount and distribution of neutral monosaccharides of savanna soils after plantation of Pinus and Eucalyptus in the Congo European Journal of Soil Science, 47, 51-59. [本文引用: 1]
UzakiM, IshiwatariR ( 1983). Determination of cellulose and non-cellulose carbohydrates in recent sediments by gas chromatography Journal of Chromatography A, 260, 487-492. [本文引用: 2]
Von Lu?tzowM, K?gel-KnabnerI, EkschmittK, MatznerE, GuggenbergerG, MarschnerB, FlessaH ( 2006). Stabilization of organic matter in temperate soils: Mechanisms and their relevance under different soil conditions―A review European Journal of Soil Science, 57, 426-445. [本文引用: 1]
WangX, ZhangL, WuJ, XuW, WangX, LuX ( 2017). Improvement of simultaneous determination of neutral monosaccharides and uronic acids by gas chromatography Food Chemistry, 220, 198-207. [本文引用: 3]
WangYP, LiXG, FuT, WangL, TurnerNC, SiddiqueKHM, LiFM ( 2016). Multi-site assessment of the effects of plastic- film mulch on the soil organic carbon balance in semiarid areas of China Agricultural and Forest Meteorology, 228, 42-51. [本文引用: 2]
WeilJ, BrewerM, HendricksonR, SarikayaA, LadischMR ( 1998). Continuous pH monitoring during pretreatment of yellow poplar wood sawdust by pressure cooking in water Applied Biochemistry and Biotechnology, 70-72, 99-111. [本文引用: 1]
XieH, LiJ, ZhuP, PengC, WangJ, HeH, ZhangX ( 2014). Long-term manure amendments enhance neutral sugar accumulation in bulk soil and particulate organic matter in a Mollisol Soil Biology & Biochemistry, 78, 45-53. [本文引用: 1]
ZhangB, DuJF, XieHT, LiWF, WangLF, ZhangXD ( 2010). Effects of long-term fertilization on features of neutral sugars in particulate organic matter Chinese Journal of Soil Science, 41, 617-621. [本文引用: 1]
ZhangS, LiC, ZhouG, CheG, YouJ, SuoY ( 2013). Determination of the carbohydrates from Notopterygium forbesii Boiss by HPLC with fluorescence detection Carbohydrate Polymers, 97, 794-799. [本文引用: 1]
ZhangW, HeH, ZhangX ( 2007). Determination of neutral sugars in soil by capillary gas chromatography after derivatization to aldononitrile acetates Soil Biology & Biochemistry, 39, 2665-2669. [本文引用: 5]
ZhaoNN, GuggenbergerG, ShibistovaO, ThaoDT, ShiWJ, LiXG ( 2014). Aspect-vegetation complex effects on biochemical characteristics and decomposability of soil organic carbon on the eastern Qinghai-Tibetan Plateau Plant and Soil, 384, 289-301. [本文引用: 5]
ZhuX, LiangC, MastersMD, KantolaIB, DeluciaEH ( 2018). The impacts of four potential bioenergy crops on soil carbon dynamics as shown by biomarker analyses and DRIFT spectroscopy Global Change Biology Bioenergy, 10, 489-500. [本文引用: 2]
Occurrence and abundance of carbohydrates and amino compounds in sequentially extracted labile soil organic matter fractions 1 2016
... 关于影响中性糖的人为因素的研究主要集中在放牧、耕作和土地利用方式改变等方面.放牧的影响存在较多争议, 如有研究发现放牧通过影响植物生物量输入使草地生态系统碳水化合物含量减少(Thompson et al., 2006), 但也有研究发现放牧对中性糖含量没有影响(Steffens et al., 2009).Abdelrahman等(2016)指出施肥和种植的农作物种类对碳水化合物含量均无显著影响, 但也有研究表明施用混合肥的土壤中性糖含量显著高于施用蚯蚓堆肥的土壤, 可能是施用混合肥土壤中微生物分解作用更小的原因(Phuong-Thi et al., 2012).对中国东北地区的研究发现, 长期单施有机肥以及有机肥和化肥配合施用可以提高作物产量, 有利于颗粒有机质中植物来源的中性糖的积累, 而单施化肥对土壤中性糖含量无显著影响(张彬等, 2010). ...
Determination of neutral and acidic sugars in soil by capillary gas-liquid chromatography after trifluoroacetic acid hydrolysis 11 1996
... 碳水化合物, 即糖类, 可分为单糖、低聚糖和多糖(Cheshire et al., 1992).碳水化合物占维管束植物干质量的75%, 细菌干质量的40% (Moers et al., 1993), 土壤中来源于植物和微生物的碳水化合物占土壤有机质的5%-25%, 是土壤有机质的重要组分(Angers et al., 1988; Murata et al., 1999).碳水化合物是土壤微生物的主要能量来源和碳源(Amelung et al., 1996), 几乎所有的低聚糖和单糖都可溶于水, 属于土壤中易降解的有机质, 易被微生物利用, 并用于自身生长代谢(Gunina & Kuzyakov, 2015).碳水化合物还可驱动微生物由休眠状态转变为活跃状态(Blagodatskaya & Kuzyakov, 2013), 是激发效应产生的有效基质(Kuzyakov, 2010).此外, 碳水化合物是团聚体形成的重要胶结物质(Feller & Beare, 1997), 例如, 阿拉伯糖和木糖是植物黏胶糖的重要来源, 对团聚体的形成起着促进作用(Oades, 1984).Larré-Larrouy等(2004)研究发现, 耕作后土壤团聚体稳定性的改变与团聚体中木糖的减少有关.Angers和Mehuys (1989)提出碳水化合物含量和团聚体的平均质量直径正相关, 表明碳水化合物含量的变化对水稳定性团聚体的改变有着一定的影响.此外, Puget等(1999)发现在更稳定的黏-粉团聚体中微生物来源的碳水化合物较丰富, 并推测团聚体的稳定性受胞外的微生物多糖调控.由此可见, 碳水化合物在团聚体稳定性方面扮演着重要角色.综上所述, 研究土壤碳水化合物对于评估土壤有机质的组成、来源、质量与动态变化具有重要意义(Guggenberger et al., 1994; Trouve et al., 1996; Nacro et al., 2005; Navarrete & Tsutsuki, 2008). ...
... 土壤中性糖的提取主要包括水解、纯化和检测三个步骤.水解可以破坏糖类与土壤基质之间的氢键、共价键(Martens & Frankenberger, 1993), 以及多糖分子结构内部的键合作用(如糖苷键)(K?gel- Knabner, 2002), 将中性单糖从半纤维素中释放出来.土壤中性单糖的水解一般用硫酸(Sowden & Ivarson, 1962; Oades et al., 1970; Cheshire, 1979)、盐酸(Uzaki & Ishiwatari, 1983)或三氟乙酸(Amelung et al., 1996)实现(表1).由于硫酸不易被除去, 而三氟乙酸挥发性强, 可通过旋转蒸发去除, 且提取效率也较硫酸高, 因此三氟乙酸水解得到了广泛的应用(Amelung et al., 1996; Rumpel & Dignac, 2006; Zhang et al., 2007; Eder et al., 2010).目前最常用的水解方法是利用4 mol·L-1的三氟乙酸在105 ℃水解4 h, 既不会破坏单糖的结构, 也能获得最高的提取效率(Amelung et al., 1996).由于盐酸的水解效率较三氟乙酸和硫酸低, 该方法基本不再使用(Amelung et al., 1996).上述水解产物的纯化通常使用旋转蒸发法去除三氟乙酸, 使用螯合剂防止铁的氢氧化物(或氧化物)干扰, 或调节pH值将铁的氢氧化物(或氧化物)以沉淀形式去除(Zhang et al., 2007; Eder et al., 2010), 也有研究使用吸附剂和阳离子交换树脂去除杂质(Amelung et al., 1996; Martens et al., 2004). ...
... ).由于硫酸不易被除去, 而三氟乙酸挥发性强, 可通过旋转蒸发去除, 且提取效率也较硫酸高, 因此三氟乙酸水解得到了广泛的应用(Amelung et al., 1996; Rumpel & Dignac, 2006; Zhang et al., 2007; Eder et al., 2010).目前最常用的水解方法是利用4 mol·L-1的三氟乙酸在105 ℃水解4 h, 既不会破坏单糖的结构, 也能获得最高的提取效率(Amelung et al., 1996).由于盐酸的水解效率较三氟乙酸和硫酸低, 该方法基本不再使用(Amelung et al., 1996).上述水解产物的纯化通常使用旋转蒸发法去除三氟乙酸, 使用螯合剂防止铁的氢氧化物(或氧化物)干扰, 或调节pH值将铁的氢氧化物(或氧化物)以沉淀形式去除(Zhang et al., 2007; Eder et al., 2010), 也有研究使用吸附剂和阳离子交换树脂去除杂质(Amelung et al., 1996; Martens et al., 2004). ...
... 的三氟乙酸在105 ℃水解4 h, 既不会破坏单糖的结构, 也能获得最高的提取效率(Amelung et al., 1996).由于盐酸的水解效率较三氟乙酸和硫酸低, 该方法基本不再使用(Amelung et al., 1996).上述水解产物的纯化通常使用旋转蒸发法去除三氟乙酸, 使用螯合剂防止铁的氢氧化物(或氧化物)干扰, 或调节pH值将铁的氢氧化物(或氧化物)以沉淀形式去除(Zhang et al., 2007; Eder et al., 2010), 也有研究使用吸附剂和阳离子交换树脂去除杂质(Amelung et al., 1996; Martens et al., 2004). ...
... ).由于盐酸的水解效率较三氟乙酸和硫酸低, 该方法基本不再使用(Amelung et al., 1996).上述水解产物的纯化通常使用旋转蒸发法去除三氟乙酸, 使用螯合剂防止铁的氢氧化物(或氧化物)干扰, 或调节pH值将铁的氢氧化物(或氧化物)以沉淀形式去除(Zhang et al., 2007; Eder et al., 2010), 也有研究使用吸附剂和阳离子交换树脂去除杂质(Amelung et al., 1996; Martens et al., 2004). ...
... ), 也有研究使用吸附剂和阳离子交换树脂去除杂质(Amelung et al., 1996; Martens et al., 2004). ...
... Comparison of extraction and detection methods of neutral sugars in soils Table 1
步骤 Procedure
方法 Method
优点 Advantage
缺点 Drawback
参考文献 Reference
提取 Extraction
硫酸 H2SO4
硫酸不易被除去 H2SO4 can not be removed easily
Tanaka et al., 1990
盐酸 HCl
会水解一部分纤维素; 产率较低 The hydrolysis products include a few cellulosic neutral sugars; low yields
Uzaki & Ishiwatari, 1983
三氟乙酸 TFA
产率高; 不会破坏单糖结构; 具有挥发性, 可通过旋转蒸发去除; 水解的多糖主要为半纤维素 High yields; Not destructive to monosaccharides; TFA is volatile and can be easily removed by evaporation; Hydrolysis products are mainly released from hemicellulose
Amelung et al., 1996
检测 Detection
GC-MS
精度、准确度、敏感性和效率较高 High accuracy, precision, sensibility and efficiency
需要衍生化 Derivatization is required
Amelung et al., 1996; Wang et al., 2017
HPLC
无需衍生化; 纯化过程简单 No need for derivatization; Simple purification procedures
精度、准确度、敏感性较低 Low accuracy, precision, sensibility and efficiency
Hamada & Ono, 1984; Angers et al., 1988;Tanaka et al., 1990;
HPAEC-PAD
无需衍生化; 应用范围广, 可同时分析糖醛酸和中性糖 No need for derivatization; Wide application and simultaneous analysis of uronic acid and neutral sugars
精度、准确度、敏感性较低 Low accuracy, precision, sensibility and efficiency
Bruggink et al., 2005; Zhang et al., 2012
TFA, trifluoroacetic acid; GC-MS, gas chromatography-mass spectrometry; HPLC, high-performance liquid chromatography; HPAEC-PAD, high-performance anion-exchange chromatography with pulsed amperometric detection. ...
... Amelung et al., 1996; Wang et al., 2017
HPLC
无需衍生化; 纯化过程简单 No need for derivatization; Simple purification procedures
精度、准确度、敏感性较低 Low accuracy, precision, sensibility and efficiency
Hamada & Ono, 1984; Angers et al., 1988;Tanaka et al., 1990;
HPAEC-PAD
无需衍生化; 应用范围广, 可同时分析糖醛酸和中性糖 No need for derivatization; Wide application and simultaneous analysis of uronic acid and neutral sugars
精度、准确度、敏感性较低 Low accuracy, precision, sensibility and efficiency
Bruggink et al., 2005; Zhang et al., 2012
TFA, trifluoroacetic acid; GC-MS, gas chromatography-mass spectrometry; HPLC, high-performance liquid chromatography; HPAEC-PAD, high-performance anion-exchange chromatography with pulsed amperometric detection. ...
... 中性糖的检测分析可使用气相色谱-质谱联用仪(GC-MS)(Rumpel & Dignac, 2006; Zhang et al., 2007; Eder et al., 2010), 液相色谱(HPLC)(Takeuchi et al., 1987; Tanaka et al., 1990; Amelung et al., 1996; Basler & Dyckmans, 2013)和高效阴离子交换色谱-脉冲安培检测法(HPAEC-PAD)(Bruggink et al., 2005; Zhang et al., 2012).由于中性糖不易挥发, 因此在使用GC-MS检测前需衍生化, 衍生化是影响土壤中性糖定量的关键环节.下面将介绍几种常用的中性糖衍生化方法. ...
... 此外, 糖腈乙酰酯衍生化也是目前使用较多的方法之一(Xie et al., 2014; Cui et al., 2016), 该衍生化方法最开始用于土壤中氨基糖含量的测定(Amelung et al., 1996), 随后Zhang等(2007)将其应用于土壤中性糖的测定.糖腈乙酰酯衍生化方法的优点是重复性较好, 获得的衍生化产物也较稳定(Zhang et al., 2007). ...
... 糖肟三甲基硅醚衍生化(trimethylsilyl oximes, TMSO)或三甲基硅烷基肟衍生化(trimethylsilyl alkyl oximes)以及三甲基硅烷基衍生化(trimethylsilyl, TMS)也被用于醛糖和酮糖的测定(Andrews, 1989; Amelung et al., 1996; Larré-Larrouy & Feller, 1997; Nierop et al., 2001; Farhadi et al., 2006), 此类衍生化步骤较简单, 但会产生多峰, 也易受到水的影响, 同时衍生化产物也不稳定(Ruiz-Matute et al., 2011). ...
Capillary gas-chromatographic analysis of monosaccharides: Improvements and comparisons using trifluoroacetylation and trimethylsilylation of sugar O-benzyl- and O-methyl-oximes 1 1989
... 糖肟三甲基硅醚衍生化(trimethylsilyl oximes, TMSO)或三甲基硅烷基肟衍生化(trimethylsilyl alkyl oximes)以及三甲基硅烷基衍生化(trimethylsilyl, TMS)也被用于醛糖和酮糖的测定(Andrews, 1989; Amelung et al., 1996; Larré-Larrouy & Feller, 1997; Nierop et al., 2001; Farhadi et al., 2006), 此类衍生化步骤较简单, 但会产生多峰, 也易受到水的影响, 同时衍生化产物也不稳定(Ruiz-Matute et al., 2011). ...
Effects of cropping on carbohydrate content and water-stable aggregation of a clay soil 1 1989
... 碳水化合物, 即糖类, 可分为单糖、低聚糖和多糖(Cheshire et al., 1992).碳水化合物占维管束植物干质量的75%, 细菌干质量的40% (Moers et al., 1993), 土壤中来源于植物和微生物的碳水化合物占土壤有机质的5%-25%, 是土壤有机质的重要组分(Angers et al., 1988; Murata et al., 1999).碳水化合物是土壤微生物的主要能量来源和碳源(Amelung et al., 1996), 几乎所有的低聚糖和单糖都可溶于水, 属于土壤中易降解的有机质, 易被微生物利用, 并用于自身生长代谢(Gunina & Kuzyakov, 2015).碳水化合物还可驱动微生物由休眠状态转变为活跃状态(Blagodatskaya & Kuzyakov, 2013), 是激发效应产生的有效基质(Kuzyakov, 2010).此外, 碳水化合物是团聚体形成的重要胶结物质(Feller & Beare, 1997), 例如, 阿拉伯糖和木糖是植物黏胶糖的重要来源, 对团聚体的形成起着促进作用(Oades, 1984).Larré-Larrouy等(2004)研究发现, 耕作后土壤团聚体稳定性的改变与团聚体中木糖的减少有关.Angers和Mehuys (1989)提出碳水化合物含量和团聚体的平均质量直径正相关, 表明碳水化合物含量的变化对水稳定性团聚体的改变有着一定的影响.此外, Puget等(1999)发现在更稳定的黏-粉团聚体中微生物来源的碳水化合物较丰富, 并推测团聚体的稳定性受胞外的微生物多糖调控.由此可见, 碳水化合物在团聚体稳定性方面扮演着重要角色.综上所述, 研究土壤碳水化合物对于评估土壤有机质的组成、来源、质量与动态变化具有重要意义(Guggenberger et al., 1994; Trouve et al., 1996; Nacro et al., 2005; Navarrete & Tsutsuki, 2008). ...
Determination of carbohydrate-composition of soil hydrolysates by high- performance liquid-chromatography 2 1988
... 碳水化合物, 即糖类, 可分为单糖、低聚糖和多糖(Cheshire et al., 1992).碳水化合物占维管束植物干质量的75%, 细菌干质量的40% (Moers et al., 1993), 土壤中来源于植物和微生物的碳水化合物占土壤有机质的5%-25%, 是土壤有机质的重要组分(Angers et al., 1988; Murata et al., 1999).碳水化合物是土壤微生物的主要能量来源和碳源(Amelung et al., 1996), 几乎所有的低聚糖和单糖都可溶于水, 属于土壤中易降解的有机质, 易被微生物利用, 并用于自身生长代谢(Gunina & Kuzyakov, 2015).碳水化合物还可驱动微生物由休眠状态转变为活跃状态(Blagodatskaya & Kuzyakov, 2013), 是激发效应产生的有效基质(Kuzyakov, 2010).此外, 碳水化合物是团聚体形成的重要胶结物质(Feller & Beare, 1997), 例如, 阿拉伯糖和木糖是植物黏胶糖的重要来源, 对团聚体的形成起着促进作用(Oades, 1984).Larré-Larrouy等(2004)研究发现, 耕作后土壤团聚体稳定性的改变与团聚体中木糖的减少有关.Angers和Mehuys (1989)提出碳水化合物含量和团聚体的平均质量直径正相关, 表明碳水化合物含量的变化对水稳定性团聚体的改变有着一定的影响.此外, Puget等(1999)发现在更稳定的黏-粉团聚体中微生物来源的碳水化合物较丰富, 并推测团聚体的稳定性受胞外的微生物多糖调控.由此可见, 碳水化合物在团聚体稳定性方面扮演着重要角色.综上所述, 研究土壤碳水化合物对于评估土壤有机质的组成、来源、质量与动态变化具有重要意义(Guggenberger et al., 1994; Trouve et al., 1996; Nacro et al., 2005; Navarrete & Tsutsuki, 2008). ...
... Comparison of extraction and detection methods of neutral sugars in soils Table 1
步骤 Procedure
方法 Method
优点 Advantage
缺点 Drawback
参考文献 Reference
提取 Extraction
硫酸 H2SO4
硫酸不易被除去 H2SO4 can not be removed easily
Tanaka et al., 1990
盐酸 HCl
会水解一部分纤维素; 产率较低 The hydrolysis products include a few cellulosic neutral sugars; low yields
Uzaki & Ishiwatari, 1983
三氟乙酸 TFA
产率高; 不会破坏单糖结构; 具有挥发性, 可通过旋转蒸发去除; 水解的多糖主要为半纤维素 High yields; Not destructive to monosaccharides; TFA is volatile and can be easily removed by evaporation; Hydrolysis products are mainly released from hemicellulose
Amelung et al., 1996
检测 Detection
GC-MS
精度、准确度、敏感性和效率较高 High accuracy, precision, sensibility and efficiency
需要衍生化 Derivatization is required
Amelung et al., 1996; Wang et al., 2017
HPLC
无需衍生化; 纯化过程简单 No need for derivatization; Simple purification procedures
精度、准确度、敏感性较低 Low accuracy, precision, sensibility and efficiency
Hamada & Ono, 1984; Angers et al., 1988;Tanaka et al., 1990;
HPAEC-PAD
无需衍生化; 应用范围广, 可同时分析糖醛酸和中性糖 No need for derivatization; Wide application and simultaneous analysis of uronic acid and neutral sugars
精度、准确度、敏感性较低 Low accuracy, precision, sensibility and efficiency
Bruggink et al., 2005; Zhang et al., 2012
TFA, trifluoroacetic acid; GC-MS, gas chromatography-mass spectrometry; HPLC, high-performance liquid chromatography; HPAEC-PAD, high-performance anion-exchange chromatography with pulsed amperometric detection. ...
Growth of eastern cottonwoods (Populus deltoides) in elevated CO2 stimulates stand-level respiration and rhizodeposition of carbohydrates, accelerates soil nutrient depletion, yet stimulates above- and belowground biomass production 1 2005
... 影响中性糖含量的因素可以分为自然因素和人为因素.自然因素有环境因素(CO2、O2、海拔、温度、降水等), 生物扰动(土壤动物和微生物), 以及自然灾害(如火灾和病虫害).Barron-Gafford等(2005)在半封闭式的生态系统(生物圈Ⅱ号)中的研究表明, 在高CO2浓度处理下, 土壤碳水化合物含量增加, 主要是由植物的地下生物量增加引起的.另外, 有研究使用开放式CO2富集系统(FACE)发现高CO2处理7年后, 由于植物生物量增加, 与对照组相比, 温带草地土壤中性糖含量增加了28% (Bock et al., 2007).与CO2不同, 当泥炭土在O2中的暴露时间延长, 土壤中性糖降解速率加快, 含量降低(Philben et al., 2015).田秋香等(2013)通过对中国长白山不同海拔梯度森林土壤中性糖分布的研究发现, 不同海拔间的温度差异是造成土壤中性糖含量差异的主要原因, 在高海拔的低温环境中微生物的分解作用减弱, 再加上微生物对外源碳的利用效率提高, 被微生物转化后的外源碳以微生物残体的形式固存在土壤中.综上所述, 高海拔有利于中性糖的积累.有关生物扰动的研究发现蚯蚓并不会影响土壤中性糖含量(Phuong-Thi et al., 2012), 而土壤微生物对于土壤中性糖的降解影响很大, 当微生物多样性较高时, 单糖如阿拉伯糖和木糖的降解速率增加, 含量减少(Baumann et al., 2013).此外, 自然灾害如火灾对土壤中性糖的影响并没有一致的规律, 不同程度的火灾对土壤不同组分的中性糖含量和组成的影响都不同(Mastrolonardo et al., 2015). ...
Microbial carbon recycling: An underestimated process controlling soil carbon dynamics―Part 1: A long-term laboratory incubation experiment 2 2015
... 有研究提出有机质的化学特性决定了其在土壤中的降解速率, 即难降解的化合物会被选择性地保存在土壤中(Stevenson, 1994).传统观点认为, 中性糖等碳水化合物属于易降解化合物, 在土壤中的周转应该较快.但越来越多的研究表明土壤中碳水化合物的周转行为与其易降解的化学特性并不相符, 这可以从土壤中碳水化合物的平均年龄较高, 与原土中总有机碳年龄相似得到印证(Gleixner et al., 2002; Derrien et al., 2006; Derrien et al., 2007).其背后的机制也备受关注, 一方面是物理保护作用或化学难降解性(Sollins et al., 1996; Six et al., 2002; Von Lu?tzow et al., 2006), 另一方面是微生物对有机质的再利用(Gleixner et al., 2002).为明确土壤碳水化合物的周转特征及其驱动机制, 现有的研究常利用以下2种方法: 1) C3-C4植物轮作: 如Derrien等(2006)在长期种植小麦的土壤中种植玉米23年, 通过13C自然丰度的变化来评估土壤中性糖的周转发现, 在>200 μm粒级的土壤中, 碳水化合物的年龄(5年)比总土壤有机碳(7年)年轻, 在50-200 μm和0-50 μm粒级的土壤中, 碳水化合物的平均年龄分别为19和101年, 与相应粒级总的土壤有机碳年龄(32和91年)相似.即, 中性糖在>200 μm粒级中的周转较快(与50-200 μm和0-50 μm粒级相比).Basler等(2015a)利用该方法发现中性糖在土壤重组分中周转比在轻组分中慢: 矿物组分(>2 g·cm-3)以及原土中的中性糖碳的平均滞留时间和总碳相似, 但是闭蓄态组分(≤2 g·cm-3)中性糖碳的平均滞留时间比总碳短; 此外, 微生物来源的中性糖的周转也比植物来源周转慢(在未分组分的原土中), Basler等(2015a)认为有机质的再循环是控制土壤有机质动态的重要因子.2)添加13C标记的葡萄糖的培养实验: Basler等(2015b)通过添加13C标记的葡萄糖进行了为期3年的培养实验, 发现周转较快的中性糖碳库平均滞留时间为25-60天, 与Derrien等(2007)为期1年的短期培养实验结果相符(17天); 而较稳定的中性糖碳库(微生物来源的中性糖)的平均滞留时间长达365天, 进一步说明微生物对土壤中性糖的再循环利用过程是影响中性糖周转的重要环节. ...
Microbial carbon recycling: An underestimated process controlling soil carbon dynamics―Part 2: A C3-C4 vegetation change field labelling experiment 1 2015
... 有研究提出有机质的化学特性决定了其在土壤中的降解速率, 即难降解的化合物会被选择性地保存在土壤中(Stevenson, 1994).传统观点认为, 中性糖等碳水化合物属于易降解化合物, 在土壤中的周转应该较快.但越来越多的研究表明土壤中碳水化合物的周转行为与其易降解的化学特性并不相符, 这可以从土壤中碳水化合物的平均年龄较高, 与原土中总有机碳年龄相似得到印证(Gleixner et al., 2002; Derrien et al., 2006; Derrien et al., 2007).其背后的机制也备受关注, 一方面是物理保护作用或化学难降解性(Sollins et al., 1996; Six et al., 2002; Von Lu?tzow et al., 2006), 另一方面是微生物对有机质的再利用(Gleixner et al., 2002).为明确土壤碳水化合物的周转特征及其驱动机制, 现有的研究常利用以下2种方法: 1) C3-C4植物轮作: 如Derrien等(2006)在长期种植小麦的土壤中种植玉米23年, 通过13C自然丰度的变化来评估土壤中性糖的周转发现, 在>200 μm粒级的土壤中, 碳水化合物的年龄(5年)比总土壤有机碳(7年)年轻, 在50-200 μm和0-50 μm粒级的土壤中, 碳水化合物的平均年龄分别为19和101年, 与相应粒级总的土壤有机碳年龄(32和91年)相似.即, 中性糖在>200 μm粒级中的周转较快(与50-200 μm和0-50 μm粒级相比).Basler等(2015a)利用该方法发现中性糖在土壤重组分中周转比在轻组分中慢: 矿物组分(>2 g·cm-3)以及原土中的中性糖碳的平均滞留时间和总碳相似, 但是闭蓄态组分(≤2 g·cm-3)中性糖碳的平均滞留时间比总碳短; 此外, 微生物来源的中性糖的周转也比植物来源周转慢(在未分组分的原土中), Basler等(2015a)认为有机质的再循环是控制土壤有机质动态的重要因子.2)添加13C标记的葡萄糖的培养实验: Basler等(2015b)通过添加13C标记的葡萄糖进行了为期3年的培养实验, 发现周转较快的中性糖碳库平均滞留时间为25-60天, 与Derrien等(2007)为期1年的短期培养实验结果相符(17天); 而较稳定的中性糖碳库(微生物来源的中性糖)的平均滞留时间长达365天, 进一步说明微生物对土壤中性糖的再循环利用过程是影响中性糖周转的重要环节. ...
Compound-specific δ13C analysis of monosaccharides from soil extracts by high- performance liquid chromatography/isotope ratio mass spectrometry 1 2013
... 中性糖的检测分析可使用气相色谱-质谱联用仪(GC-MS)(Rumpel & Dignac, 2006; Zhang et al., 2007; Eder et al., 2010), 液相色谱(HPLC)(Takeuchi et al., 1987; Tanaka et al., 1990; Amelung et al., 1996; Basler & Dyckmans, 2013)和高效阴离子交换色谱-脉冲安培检测法(HPAEC-PAD)(Bruggink et al., 2005; Zhang et al., 2012).由于中性糖不易挥发, 因此在使用GC-MS检测前需衍生化, 衍生化是影响土壤中性糖定量的关键环节.下面将介绍几种常用的中性糖衍生化方法. ...
Soil microbial diversity affects soil organic matter decomposition in a silty grassland soil 1 2013
... 影响中性糖含量的因素可以分为自然因素和人为因素.自然因素有环境因素(CO2、O2、海拔、温度、降水等), 生物扰动(土壤动物和微生物), 以及自然灾害(如火灾和病虫害).Barron-Gafford等(2005)在半封闭式的生态系统(生物圈Ⅱ号)中的研究表明, 在高CO2浓度处理下, 土壤碳水化合物含量增加, 主要是由植物的地下生物量增加引起的.另外, 有研究使用开放式CO2富集系统(FACE)发现高CO2处理7年后, 由于植物生物量增加, 与对照组相比, 温带草地土壤中性糖含量增加了28% (Bock et al., 2007).与CO2不同, 当泥炭土在O2中的暴露时间延长, 土壤中性糖降解速率加快, 含量降低(Philben et al., 2015).田秋香等(2013)通过对中国长白山不同海拔梯度森林土壤中性糖分布的研究发现, 不同海拔间的温度差异是造成土壤中性糖含量差异的主要原因, 在高海拔的低温环境中微生物的分解作用减弱, 再加上微生物对外源碳的利用效率提高, 被微生物转化后的外源碳以微生物残体的形式固存在土壤中.综上所述, 高海拔有利于中性糖的积累.有关生物扰动的研究发现蚯蚓并不会影响土壤中性糖含量(Phuong-Thi et al., 2012), 而土壤微生物对于土壤中性糖的降解影响很大, 当微生物多样性较高时, 单糖如阿拉伯糖和木糖的降解速率增加, 含量减少(Baumann et al., 2013).此外, 自然灾害如火灾对土壤中性糖的影响并没有一致的规律, 不同程度的火灾对土壤不同组分的中性糖含量和组成的影响都不同(Mastrolonardo et al., 2015). ...
Root uptake of N-containing and N-free low molecular weight organic substances by maize: A 14C/ 15N tracer study 1 2008
... 按密度分组, 中性糖在轻质组分中的相对含量(129.0-410.0 mg·g-1)较重质组分中(7.5-322.0 mg·g-1)高.Llorente等(2017)在含碳酸钙的土壤中也发现中性糖含量在游离态组分中最高, 闭蓄态组分次之, 重质组分中最低; 由于微生物更易进入, 因此游离态组分中含有较多的甘露糖.另外, Bischoff等(2018)在西伯利亚高原不同盐分梯度的土壤中也发现, 无论是在表层还是底层, 中性糖含量在轻质组分中最高.但是, 与全土相比, 与矿质相结合的重质组分中中性糖含量更丰富, 该组分中以微生物来源的六碳糖最为丰富; 此外, 该研究中放射性碳的数据显示年龄较老的土壤样品中微生物来源的中性糖含量较高, 说明中性糖在矿质土中的稳定储存是通过连续的微生物转化或矿物的物理保护实现的(Rumpel et al., 2010).Dao等(2018)在西伯利亚东北部冻土中的研究也发现微生物来源的中性糖在重质组分土壤中含量较丰富, 从较低的C:N和较高的碳稳定同位素比率(δ13C)可以看出这与微生物来源的中性糖的选择性积累有关(Gentsch et al., 2015a, 2015b).此外, 微生物残体(Miltner et al., 2012)及微生物分泌的胞外多聚物中富含的多糖(Kleber et al., 2015), 也可能通过矿物的吸附作用在重质组分土壤中积累. ...
Recent progress in the analysis of sugar monomers from complex matrices using chromatography in conjunction with mass spectrometry or stand- alone tandem mass spectrometry 2 1996
... 糖醇乙酸酯衍生化是目前使用最广泛的一种中性糖衍生化方法, 其原理是将醛糖的羰基还原, 最终形成糖醇乙酸酯(Black & Fox, 1996; Ruiz-Matute et al., 2011).该衍生化的优点是: 得到的每种糖醇产物只产生一个单独的色谱峰, 易于检测, 且衍生化产物较稳定, 可稳定储存20天(Wang et al., 2017), 因此被广泛用于中性糖的定量分析(Rumpel & Dignac, 2006; Spielvogel et al., 2007; Rumpel et al., 2010).然而, 该衍生化方法耗时, 步骤繁琐(Black & Fox, 1996; Rumpel & Dignac, 2006; Ruiz-Matute et al., 2011), 一直有研究在对该方法进行不断改进. ...
Active microorganisms in soil: Critical review of estimation criteria and approaches 1 2013
... 碳水化合物, 即糖类, 可分为单糖、低聚糖和多糖(Cheshire et al., 1992).碳水化合物占维管束植物干质量的75%, 细菌干质量的40% (Moers et al., 1993), 土壤中来源于植物和微生物的碳水化合物占土壤有机质的5%-25%, 是土壤有机质的重要组分(Angers et al., 1988; Murata et al., 1999).碳水化合物是土壤微生物的主要能量来源和碳源(Amelung et al., 1996), 几乎所有的低聚糖和单糖都可溶于水, 属于土壤中易降解的有机质, 易被微生物利用, 并用于自身生长代谢(Gunina & Kuzyakov, 2015).碳水化合物还可驱动微生物由休眠状态转变为活跃状态(Blagodatskaya & Kuzyakov, 2013), 是激发效应产生的有效基质(Kuzyakov, 2010).此外, 碳水化合物是团聚体形成的重要胶结物质(Feller & Beare, 1997), 例如, 阿拉伯糖和木糖是植物黏胶糖的重要来源, 对团聚体的形成起着促进作用(Oades, 1984).Larré-Larrouy等(2004)研究发现, 耕作后土壤团聚体稳定性的改变与团聚体中木糖的减少有关.Angers和Mehuys (1989)提出碳水化合物含量和团聚体的平均质量直径正相关, 表明碳水化合物含量的变化对水稳定性团聚体的改变有着一定的影响.此外, Puget等(1999)发现在更稳定的黏-粉团聚体中微生物来源的碳水化合物较丰富, 并推测团聚体的稳定性受胞外的微生物多糖调控.由此可见, 碳水化合物在团聚体稳定性方面扮演着重要角色.综上所述, 研究土壤碳水化合物对于评估土壤有机质的组成、来源、质量与动态变化具有重要意义(Guggenberger et al., 1994; Trouve et al., 1996; Nacro et al., 2005; Navarrete & Tsutsuki, 2008). ...
Sequestration and turnover of plant- and microbially derived sugars in a temperate grassland soil during 7 years exposed to elevated atmospheric pCO2 1 2007
... 影响中性糖含量的因素可以分为自然因素和人为因素.自然因素有环境因素(CO2、O2、海拔、温度、降水等), 生物扰动(土壤动物和微生物), 以及自然灾害(如火灾和病虫害).Barron-Gafford等(2005)在半封闭式的生态系统(生物圈Ⅱ号)中的研究表明, 在高CO2浓度处理下, 土壤碳水化合物含量增加, 主要是由植物的地下生物量增加引起的.另外, 有研究使用开放式CO2富集系统(FACE)发现高CO2处理7年后, 由于植物生物量增加, 与对照组相比, 温带草地土壤中性糖含量增加了28% (Bock et al., 2007).与CO2不同, 当泥炭土在O2中的暴露时间延长, 土壤中性糖降解速率加快, 含量降低(Philben et al., 2015).田秋香等(2013)通过对中国长白山不同海拔梯度森林土壤中性糖分布的研究发现, 不同海拔间的温度差异是造成土壤中性糖含量差异的主要原因, 在高海拔的低温环境中微生物的分解作用减弱, 再加上微生物对外源碳的利用效率提高, 被微生物转化后的外源碳以微生物残体的形式固存在土壤中.综上所述, 高海拔有利于中性糖的积累.有关生物扰动的研究发现蚯蚓并不会影响土壤中性糖含量(Phuong-Thi et al., 2012), 而土壤微生物对于土壤中性糖的降解影响很大, 当微生物多样性较高时, 单糖如阿拉伯糖和木糖的降解速率增加, 含量减少(Baumann et al., 2013).此外, 自然灾害如火灾对土壤中性糖的影响并没有一致的规律, 不同程度的火灾对土壤不同组分的中性糖含量和组成的影响都不同(Mastrolonardo et al., 2015). ...
Analysis of carbohydrates by anion exchange chromatography and mass spectrometry 2 2005
... Comparison of extraction and detection methods of neutral sugars in soils Table 1
步骤 Procedure
方法 Method
优点 Advantage
缺点 Drawback
参考文献 Reference
提取 Extraction
硫酸 H2SO4
硫酸不易被除去 H2SO4 can not be removed easily
Tanaka et al., 1990
盐酸 HCl
会水解一部分纤维素; 产率较低 The hydrolysis products include a few cellulosic neutral sugars; low yields
Uzaki & Ishiwatari, 1983
三氟乙酸 TFA
产率高; 不会破坏单糖结构; 具有挥发性, 可通过旋转蒸发去除; 水解的多糖主要为半纤维素 High yields; Not destructive to monosaccharides; TFA is volatile and can be easily removed by evaporation; Hydrolysis products are mainly released from hemicellulose
Amelung et al., 1996
检测 Detection
GC-MS
精度、准确度、敏感性和效率较高 High accuracy, precision, sensibility and efficiency
需要衍生化 Derivatization is required
Amelung et al., 1996; Wang et al., 2017
HPLC
无需衍生化; 纯化过程简单 No need for derivatization; Simple purification procedures
精度、准确度、敏感性较低 Low accuracy, precision, sensibility and efficiency
Hamada & Ono, 1984; Angers et al., 1988;Tanaka et al., 1990;
HPAEC-PAD
无需衍生化; 应用范围广, 可同时分析糖醛酸和中性糖 No need for derivatization; Wide application and simultaneous analysis of uronic acid and neutral sugars
精度、准确度、敏感性较低 Low accuracy, precision, sensibility and efficiency
Bruggink et al., 2005; Zhang et al., 2012
TFA, trifluoroacetic acid; GC-MS, gas chromatography-mass spectrometry; HPLC, high-performance liquid chromatography; HPAEC-PAD, high-performance anion-exchange chromatography with pulsed amperometric detection. ...
... 中性糖的检测分析可使用气相色谱-质谱联用仪(GC-MS)(Rumpel & Dignac, 2006; Zhang et al., 2007; Eder et al., 2010), 液相色谱(HPLC)(Takeuchi et al., 1987; Tanaka et al., 1990; Amelung et al., 1996; Basler & Dyckmans, 2013)和高效阴离子交换色谱-脉冲安培检测法(HPAEC-PAD)(Bruggink et al., 2005; Zhang et al., 2012).由于中性糖不易挥发, 因此在使用GC-MS检测前需衍生化, 衍生化是影响土壤中性糖定量的关键环节.下面将介绍几种常用的中性糖衍生化方法. ...
Content and distribution of neutral sugars across different land-use regimes in the top soils (<xref ref-type="bibr" rid="b58">Nierop<i> et al</i>., 2001</xref>; <xref ref-type="bibr" rid="b71">Spielvogel<i> et al</i>., 2007</xref>; <xref ref-type="bibr" rid="b25">Eder<i> et al</i>., 2010</xref>; <xref ref-type="bibr" rid="b67">Rumpel<i> et al</i>., 2010</xref>; <xref ref-type="bibr" rid="b90">Zhao<i> et al</i>., 2014</xref>; <xref ref-type="bibr" rid="b19">Conti<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b21">Cui<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b82">Wang<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b20">Creme<i> et al</i>., 2017</xref>; <xref ref-type="bibr" rid="b49">Llorente<i> et al</i>., 2017</xref>; <xref ref-type="bibr" rid="b26">Evgrafova <i>et al</i>., 2018</xref>; <xref ref-type="bibr" rid="b91">Zhu <i>et al</i>., 2018</xref>). A, Neutral sugar absolute content. B, Neutral sugar relative content. C, GM/AX ((galactose + mannose)/(arabinose + xylose)). D, RF/AX ((rhamnose + fucose)/(arabinose + xylose)). The upper and lower end of boxes denote the 0.25 and 0.75 percentiles, respectively. The solid bar in the box mark the median of each dataset. The circles indicate outliers of each dataset. Different lowercase letters indicate differences in various land-use regimes (<i>p</i> < 0.05). <i>n</i> = 8, 25, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the A, C). <i>n</i> = 8, 22, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the B). <i>n</i> = 3, 19, 4, 8, 27 (from deciduous, coniferous, shrub, grassland to crops in the D). OC, soil organic carbon.Fig. 1 为考察不同土地利用类型中的中性糖对土壤有机碳(OC)的潜在贡献, 我们利用土壤有机碳含量对中性糖含量进行了归一化.结果显示: 阔叶林(84.5-253.9 mg·g-1)、针叶林(55.2-291.0 mg·g-1)、灌丛(139.0-208.9 mg·g-1)和草地(161.0- 371.0 mg·g-1)土壤中性糖的相对含量无显著差异(p > 0.05; 图1B), 除灌丛外, 其他类型的中性糖相对含量均显著高于耕地(65.0-370.7 mg·g-1)(p < 0.05; 图1B).这与上述中性糖的绝对含量结果一致, 说明耕种会通过减少植物残体的输入从而使土壤中性糖含量减少.也有研究发现虽然土地利用类型由森林转变为灌丛, 但主要的维管束植物种类未改变, 凋落物质量没有变化导致两种土地利用类型土壤中性糖含量无显著差异(Conti et al., 2016).而Zhao等(2014)在青藏高原的研究表明, 草地土壤中性糖的相对含量显著高于灌丛, 可能是草本植物凋落物的半纤维素含量比灌丛植物凋落物中的含量高导致的. ...
... ; Conti et al., 2016; Cui et al., 2016; Wang et al., 2016; Creme et al., 2017; Llorente et al., 2017; Evgrafova et al., 2018; Zhu et al., 2018). A, Neutral sugar absolute content. B, Neutral sugar relative content. C, GM/AX ((galactose + mannose)/(arabinose + xylose)). D, RF/AX ((rhamnose + fucose)/(arabinose + xylose)). The upper and lower end of boxes denote the 0.25 and 0.75 percentiles, respectively. The solid bar in the box mark the median of each dataset. The circles indicate outliers of each dataset. Different lowercase letters indicate differences in various land-use regimes (p < 0.05). n = 8, 25, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the A, C). n = 8, 22, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the B). n = 3, 19, 4, 8, 27 (from deciduous, coniferous, shrub, grassland to crops in the D). OC, soil organic carbon. Fig. 1 为考察不同土地利用类型中的中性糖对土壤有机碳(OC)的潜在贡献, 我们利用土壤有机碳含量对中性糖含量进行了归一化.结果显示: 阔叶林(84.5-253.9 mg·g-1)、针叶林(55.2-291.0 mg·g-1)、灌丛(139.0-208.9 mg·g-1)和草地(161.0- 371.0 mg·g-1)土壤中性糖的相对含量无显著差异(p > 0.05; 图1B), 除灌丛外, 其他类型的中性糖相对含量均显著高于耕地(65.0-370.7 mg·g-1)(p < 0.05; 图1B).这与上述中性糖的绝对含量结果一致, 说明耕种会通过减少植物残体的输入从而使土壤中性糖含量减少.也有研究发现虽然土地利用类型由森林转变为灌丛, 但主要的维管束植物种类未改变, 凋落物质量没有变化导致两种土地利用类型土壤中性糖含量无显著差异(Conti et al., 2016).而Zhao等(2014)在青藏高原的研究表明, 草地土壤中性糖的相对含量显著高于灌丛, 可能是草本植物凋落物的半纤维素含量比灌丛植物凋落物中的含量高导致的. ...
Content and distribution of neutral sugars across different land-use regimes in the top soils (<xref ref-type="bibr" rid="b58">Nierop<i> et al</i>., 2001</xref>; <xref ref-type="bibr" rid="b71">Spielvogel<i> et al</i>., 2007</xref>; <xref ref-type="bibr" rid="b25">Eder<i> et al</i>., 2010</xref>; <xref ref-type="bibr" rid="b67">Rumpel<i> et al</i>., 2010</xref>; <xref ref-type="bibr" rid="b90">Zhao<i> et al</i>., 2014</xref>; <xref ref-type="bibr" rid="b19">Conti<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b21">Cui<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b82">Wang<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b20">Creme<i> et al</i>., 2017</xref>; <xref ref-type="bibr" rid="b49">Llorente<i> et al</i>., 2017</xref>; <xref ref-type="bibr" rid="b26">Evgrafova <i>et al</i>., 2018</xref>; <xref ref-type="bibr" rid="b91">Zhu <i>et al</i>., 2018</xref>). A, Neutral sugar absolute content. B, Neutral sugar relative content. C, GM/AX ((galactose + mannose)/(arabinose + xylose)). D, RF/AX ((rhamnose + fucose)/(arabinose + xylose)). The upper and lower end of boxes denote the 0.25 and 0.75 percentiles, respectively. The solid bar in the box mark the median of each dataset. The circles indicate outliers of each dataset. Different lowercase letters indicate differences in various land-use regimes (<i>p</i> < 0.05). <i>n</i> = 8, 25, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the A, C). <i>n</i> = 8, 22, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the B). <i>n</i> = 3, 19, 4, 8, 27 (from deciduous, coniferous, shrub, grassland to crops in the D). OC, soil organic carbon.Fig. 1 为考察不同土地利用类型中的中性糖对土壤有机碳(OC)的潜在贡献, 我们利用土壤有机碳含量对中性糖含量进行了归一化.结果显示: 阔叶林(84.5-253.9 mg·g-1)、针叶林(55.2-291.0 mg·g-1)、灌丛(139.0-208.9 mg·g-1)和草地(161.0- 371.0 mg·g-1)土壤中性糖的相对含量无显著差异(p > 0.05; 图1B), 除灌丛外, 其他类型的中性糖相对含量均显著高于耕地(65.0-370.7 mg·g-1)(p < 0.05; 图1B).这与上述中性糖的绝对含量结果一致, 说明耕种会通过减少植物残体的输入从而使土壤中性糖含量减少.也有研究发现虽然土地利用类型由森林转变为灌丛, 但主要的维管束植物种类未改变, 凋落物质量没有变化导致两种土地利用类型土壤中性糖含量无显著差异(Conti et al., 2016).而Zhao等(2014)在青藏高原的研究表明, 草地土壤中性糖的相对含量显著高于灌丛, 可能是草本植物凋落物的半纤维素含量比灌丛植物凋落物中的含量高导致的. ...
... ; Creme et al., 2017; Llorente et al., 2017; Evgrafova et al., 2018; Zhu et al., 2018). A, Neutral sugar absolute content. B, Neutral sugar relative content. C, GM/AX ((galactose + mannose)/(arabinose + xylose)). D, RF/AX ((rhamnose + fucose)/(arabinose + xylose)). The upper and lower end of boxes denote the 0.25 and 0.75 percentiles, respectively. The solid bar in the box mark the median of each dataset. The circles indicate outliers of each dataset. Different lowercase letters indicate differences in various land-use regimes (p < 0.05). n = 8, 25, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the A, C). n = 8, 22, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the B). n = 3, 19, 4, 8, 27 (from deciduous, coniferous, shrub, grassland to crops in the D). OC, soil organic carbon. Fig. 1 为考察不同土地利用类型中的中性糖对土壤有机碳(OC)的潜在贡献, 我们利用土壤有机碳含量对中性糖含量进行了归一化.结果显示: 阔叶林(84.5-253.9 mg·g-1)、针叶林(55.2-291.0 mg·g-1)、灌丛(139.0-208.9 mg·g-1)和草地(161.0- 371.0 mg·g-1)土壤中性糖的相对含量无显著差异(p > 0.05; 图1B), 除灌丛外, 其他类型的中性糖相对含量均显著高于耕地(65.0-370.7 mg·g-1)(p < 0.05; 图1B).这与上述中性糖的绝对含量结果一致, 说明耕种会通过减少植物残体的输入从而使土壤中性糖含量减少.也有研究发现虽然土地利用类型由森林转变为灌丛, 但主要的维管束植物种类未改变, 凋落物质量没有变化导致两种土地利用类型土壤中性糖含量无显著差异(Conti et al., 2016).而Zhao等(2014)在青藏高原的研究表明, 草地土壤中性糖的相对含量显著高于灌丛, 可能是草本植物凋落物的半纤维素含量比灌丛植物凋落物中的含量高导致的. ...
Impacts of vegetation type and climatic zone on neutral sugar distribution in natural forest soils 4 2016
... 此外, 糖腈乙酰酯衍生化也是目前使用较多的方法之一(Xie et al., 2014; Cui et al., 2016), 该衍生化方法最开始用于土壤中氨基糖含量的测定(Amelung et al., 1996), 随后Zhang等(2007)将其应用于土壤中性糖的测定.糖腈乙酰酯衍生化方法的优点是重复性较好, 获得的衍生化产物也较稳定(Zhang et al., 2007). ...
... 本研究整合了5种土地利用类型的土壤中性糖数据, 发现阔叶林(7.0-82.8 mg·g-1)、针叶林(1.1-90.1 mg·g-1)、灌丛(2.5-80.6 mg·g-1)和草地(1.1-15.1 mg·g-1)土壤中性糖的绝对含量无显著差异(p > 0.05; 图1A), 除草地外, 其他类型均显著高于耕地土壤(0.6-6.3 mg·g-1)(p < 0.05; 图1A).不同地区(西班牙西北部、美国中西部、法国西南部)、不同土壤类型(碳酸钙土壤、沙质淋溶土)中的研究均表明阔叶林和针叶林土壤中性糖(或碳水化合物, 包括纤维素和半纤维中性糖, 下同)的绝对含量显著高于耕地土壤, 并认为可能是人为收获引起的地上输入量减少, 以及耕作扰动导致土壤有机层中的有机质向矿质层迁移造成的(Martens et al., 2004; Jolivet et al., 2006; Llorente et al., 2017).但是有研究报道中性糖(或碳水化合物)在草地土壤中更丰富(与耕地相比)(Larré-Larrouy et al., 2004; Martens et al., 2004; Guan et al., 2018).如, Guan等(2018)发现青藏高原的自然草地经过长期(>50年)耕种后, 植物地下生物量降低使植物碳向土壤中的输入减少, 加上耕作的扰动作用强烈影响了微生物的群落结构和丰度, 导致与草地相比, 耕地土壤中性糖含量降低了57%.Larré-Larrouy等(2004)也认为农作物的根系不发达, 来源于根系的碳水化合物较少是导致此现象的主要原因.由于本研究的数据基于土壤类型、气候条件等各异的研究, 中性糖含量在各研究之间存在差异, 加之数据量的限制导致了与上述研究的结果不一致.
Content and distribution of neutral sugars across different land-use regimes in the top soils (<xref ref-type="bibr" rid="b58">Nierop<i> et al</i>., 2001</xref>; <xref ref-type="bibr" rid="b71">Spielvogel<i> et al</i>., 2007</xref>; <xref ref-type="bibr" rid="b25">Eder<i> et al</i>., 2010</xref>; <xref ref-type="bibr" rid="b67">Rumpel<i> et al</i>., 2010</xref>; <xref ref-type="bibr" rid="b90">Zhao<i> et al</i>., 2014</xref>; <xref ref-type="bibr" rid="b19">Conti<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b21">Cui<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b82">Wang<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b20">Creme<i> et al</i>., 2017</xref>; <xref ref-type="bibr" rid="b49">Llorente<i> et al</i>., 2017</xref>; <xref ref-type="bibr" rid="b26">Evgrafova <i>et al</i>., 2018</xref>; <xref ref-type="bibr" rid="b91">Zhu <i>et al</i>., 2018</xref>). A, Neutral sugar absolute content. B, Neutral sugar relative content. C, GM/AX ((galactose + mannose)/(arabinose + xylose)). D, RF/AX ((rhamnose + fucose)/(arabinose + xylose)). The upper and lower end of boxes denote the 0.25 and 0.75 percentiles, respectively. The solid bar in the box mark the median of each dataset. The circles indicate outliers of each dataset. Different lowercase letters indicate differences in various land-use regimes (<i>p</i> < 0.05). <i>n</i> = 8, 25, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the A, C). <i>n</i> = 8, 22, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the B). <i>n</i> = 3, 19, 4, 8, 27 (from deciduous, coniferous, shrub, grassland to crops in the D). OC, soil organic carbon.Fig. 1 为考察不同土地利用类型中的中性糖对土壤有机碳(OC)的潜在贡献, 我们利用土壤有机碳含量对中性糖含量进行了归一化.结果显示: 阔叶林(84.5-253.9 mg·g-1)、针叶林(55.2-291.0 mg·g-1)、灌丛(139.0-208.9 mg·g-1)和草地(161.0- 371.0 mg·g-1)土壤中性糖的相对含量无显著差异(p > 0.05; 图1B), 除灌丛外, 其他类型的中性糖相对含量均显著高于耕地(65.0-370.7 mg·g-1)(p < 0.05; 图1B).这与上述中性糖的绝对含量结果一致, 说明耕种会通过减少植物残体的输入从而使土壤中性糖含量减少.也有研究发现虽然土地利用类型由森林转变为灌丛, 但主要的维管束植物种类未改变, 凋落物质量没有变化导致两种土地利用类型土壤中性糖含量无显著差异(Conti et al., 2016).而Zhao等(2014)在青藏高原的研究表明, 草地土壤中性糖的相对含量显著高于灌丛, 可能是草本植物凋落物的半纤维素含量比灌丛植物凋落物中的含量高导致的. ...
... ; Cui et al., 2016; Wang et al., 2016; Creme et al., 2017; Llorente et al., 2017; Evgrafova et al., 2018; Zhu et al., 2018). A, Neutral sugar absolute content. B, Neutral sugar relative content. C, GM/AX ((galactose + mannose)/(arabinose + xylose)). D, RF/AX ((rhamnose + fucose)/(arabinose + xylose)). The upper and lower end of boxes denote the 0.25 and 0.75 percentiles, respectively. The solid bar in the box mark the median of each dataset. The circles indicate outliers of each dataset. Different lowercase letters indicate differences in various land-use regimes (p < 0.05). n = 8, 25, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the A, C). n = 8, 22, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the B). n = 3, 19, 4, 8, 27 (from deciduous, coniferous, shrub, grassland to crops in the D). OC, soil organic carbon. Fig. 1 为考察不同土地利用类型中的中性糖对土壤有机碳(OC)的潜在贡献, 我们利用土壤有机碳含量对中性糖含量进行了归一化.结果显示: 阔叶林(84.5-253.9 mg·g-1)、针叶林(55.2-291.0 mg·g-1)、灌丛(139.0-208.9 mg·g-1)和草地(161.0- 371.0 mg·g-1)土壤中性糖的相对含量无显著差异(p > 0.05; 图1B), 除灌丛外, 其他类型的中性糖相对含量均显著高于耕地(65.0-370.7 mg·g-1)(p < 0.05; 图1B).这与上述中性糖的绝对含量结果一致, 说明耕种会通过减少植物残体的输入从而使土壤中性糖含量减少.也有研究发现虽然土地利用类型由森林转变为灌丛, 但主要的维管束植物种类未改变, 凋落物质量没有变化导致两种土地利用类型土壤中性糖含量无显著差异(Conti et al., 2016).而Zhao等(2014)在青藏高原的研究表明, 草地土壤中性糖的相对含量显著高于灌丛, 可能是草本植物凋落物的半纤维素含量比灌丛植物凋落物中的含量高导致的. ...
Analysis of hydrolysable neutral sugars in mineral soils: Improvement of alditol acetylation for gas chromatographic separation and measurement 5 2010
... 土壤中性糖的提取主要包括水解、纯化和检测三个步骤.水解可以破坏糖类与土壤基质之间的氢键、共价键(Martens & Frankenberger, 1993), 以及多糖分子结构内部的键合作用(如糖苷键)(K?gel- Knabner, 2002), 将中性单糖从半纤维素中释放出来.土壤中性单糖的水解一般用硫酸(Sowden & Ivarson, 1962; Oades et al., 1970; Cheshire, 1979)、盐酸(Uzaki & Ishiwatari, 1983)或三氟乙酸(Amelung et al., 1996)实现(表1).由于硫酸不易被除去, 而三氟乙酸挥发性强, 可通过旋转蒸发去除, 且提取效率也较硫酸高, 因此三氟乙酸水解得到了广泛的应用(Amelung et al., 1996; Rumpel & Dignac, 2006; Zhang et al., 2007; Eder et al., 2010).目前最常用的水解方法是利用4 mol·L-1的三氟乙酸在105 ℃水解4 h, 既不会破坏单糖的结构, 也能获得最高的提取效率(Amelung et al., 1996).由于盐酸的水解效率较三氟乙酸和硫酸低, 该方法基本不再使用(Amelung et al., 1996).上述水解产物的纯化通常使用旋转蒸发法去除三氟乙酸, 使用螯合剂防止铁的氢氧化物(或氧化物)干扰, 或调节pH值将铁的氢氧化物(或氧化物)以沉淀形式去除(Zhang et al., 2007; Eder et al., 2010), 也有研究使用吸附剂和阳离子交换树脂去除杂质(Amelung et al., 1996; Martens et al., 2004). ...
... ; Eder et al., 2010), 也有研究使用吸附剂和阳离子交换树脂去除杂质(Amelung et al., 1996; Martens et al., 2004). ...
... 中性糖的检测分析可使用气相色谱-质谱联用仪(GC-MS)(Rumpel & Dignac, 2006; Zhang et al., 2007; Eder et al., 2010), 液相色谱(HPLC)(Takeuchi et al., 1987; Tanaka et al., 1990; Amelung et al., 1996; Basler & Dyckmans, 2013)和高效阴离子交换色谱-脉冲安培检测法(HPAEC-PAD)(Bruggink et al., 2005; Zhang et al., 2012).由于中性糖不易挥发, 因此在使用GC-MS检测前需衍生化, 衍生化是影响土壤中性糖定量的关键环节.下面将介绍几种常用的中性糖衍生化方法. ...
... 本研究整合了5种土地利用类型的土壤中性糖数据, 发现阔叶林(7.0-82.8 mg·g-1)、针叶林(1.1-90.1 mg·g-1)、灌丛(2.5-80.6 mg·g-1)和草地(1.1-15.1 mg·g-1)土壤中性糖的绝对含量无显著差异(p > 0.05; 图1A), 除草地外, 其他类型均显著高于耕地土壤(0.6-6.3 mg·g-1)(p < 0.05; 图1A).不同地区(西班牙西北部、美国中西部、法国西南部)、不同土壤类型(碳酸钙土壤、沙质淋溶土)中的研究均表明阔叶林和针叶林土壤中性糖(或碳水化合物, 包括纤维素和半纤维中性糖, 下同)的绝对含量显著高于耕地土壤, 并认为可能是人为收获引起的地上输入量减少, 以及耕作扰动导致土壤有机层中的有机质向矿质层迁移造成的(Martens et al., 2004; Jolivet et al., 2006; Llorente et al., 2017).但是有研究报道中性糖(或碳水化合物)在草地土壤中更丰富(与耕地相比)(Larré-Larrouy et al., 2004; Martens et al., 2004; Guan et al., 2018).如, Guan等(2018)发现青藏高原的自然草地经过长期(>50年)耕种后, 植物地下生物量降低使植物碳向土壤中的输入减少, 加上耕作的扰动作用强烈影响了微生物的群落结构和丰度, 导致与草地相比, 耕地土壤中性糖含量降低了57%.Larré-Larrouy等(2004)也认为农作物的根系不发达, 来源于根系的碳水化合物较少是导致此现象的主要原因.由于本研究的数据基于土壤类型、气候条件等各异的研究, 中性糖含量在各研究之间存在差异, 加之数据量的限制导致了与上述研究的结果不一致.
Content and distribution of neutral sugars across different land-use regimes in the top soils (<xref ref-type="bibr" rid="b58">Nierop<i> et al</i>., 2001</xref>; <xref ref-type="bibr" rid="b71">Spielvogel<i> et al</i>., 2007</xref>; <xref ref-type="bibr" rid="b25">Eder<i> et al</i>., 2010</xref>; <xref ref-type="bibr" rid="b67">Rumpel<i> et al</i>., 2010</xref>; <xref ref-type="bibr" rid="b90">Zhao<i> et al</i>., 2014</xref>; <xref ref-type="bibr" rid="b19">Conti<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b21">Cui<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b82">Wang<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b20">Creme<i> et al</i>., 2017</xref>; <xref ref-type="bibr" rid="b49">Llorente<i> et al</i>., 2017</xref>; <xref ref-type="bibr" rid="b26">Evgrafova <i>et al</i>., 2018</xref>; <xref ref-type="bibr" rid="b91">Zhu <i>et al</i>., 2018</xref>). A, Neutral sugar absolute content. B, Neutral sugar relative content. C, GM/AX ((galactose + mannose)/(arabinose + xylose)). D, RF/AX ((rhamnose + fucose)/(arabinose + xylose)). The upper and lower end of boxes denote the 0.25 and 0.75 percentiles, respectively. The solid bar in the box mark the median of each dataset. The circles indicate outliers of each dataset. Different lowercase letters indicate differences in various land-use regimes (<i>p</i> < 0.05). <i>n</i> = 8, 25, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the A, C). <i>n</i> = 8, 22, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the B). <i>n</i> = 3, 19, 4, 8, 27 (from deciduous, coniferous, shrub, grassland to crops in the D). OC, soil organic carbon.Fig. 1 为考察不同土地利用类型中的中性糖对土壤有机碳(OC)的潜在贡献, 我们利用土壤有机碳含量对中性糖含量进行了归一化.结果显示: 阔叶林(84.5-253.9 mg·g-1)、针叶林(55.2-291.0 mg·g-1)、灌丛(139.0-208.9 mg·g-1)和草地(161.0- 371.0 mg·g-1)土壤中性糖的相对含量无显著差异(p > 0.05; 图1B), 除灌丛外, 其他类型的中性糖相对含量均显著高于耕地(65.0-370.7 mg·g-1)(p < 0.05; 图1B).这与上述中性糖的绝对含量结果一致, 说明耕种会通过减少植物残体的输入从而使土壤中性糖含量减少.也有研究发现虽然土地利用类型由森林转变为灌丛, 但主要的维管束植物种类未改变, 凋落物质量没有变化导致两种土地利用类型土壤中性糖含量无显著差异(Conti et al., 2016).而Zhao等(2014)在青藏高原的研究表明, 草地土壤中性糖的相对含量显著高于灌丛, 可能是草本植物凋落物的半纤维素含量比灌丛植物凋落物中的含量高导致的. ...
... ; Eder et al., 2010; Rumpel et al., 2010; Zhao et al., 2014; Conti et al., 2016; Cui et al., 2016; Wang et al., 2016; Creme et al., 2017; Llorente et al., 2017; Evgrafova et al., 2018; Zhu et al., 2018). A, Neutral sugar absolute content. B, Neutral sugar relative content. C, GM/AX ((galactose + mannose)/(arabinose + xylose)). D, RF/AX ((rhamnose + fucose)/(arabinose + xylose)). The upper and lower end of boxes denote the 0.25 and 0.75 percentiles, respectively. The solid bar in the box mark the median of each dataset. The circles indicate outliers of each dataset. Different lowercase letters indicate differences in various land-use regimes (p < 0.05). n = 8, 25, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the A, C). n = 8, 22, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the B). n = 3, 19, 4, 8, 27 (from deciduous, coniferous, shrub, grassland to crops in the D). OC, soil organic carbon. Fig. 1 为考察不同土地利用类型中的中性糖对土壤有机碳(OC)的潜在贡献, 我们利用土壤有机碳含量对中性糖含量进行了归一化.结果显示: 阔叶林(84.5-253.9 mg·g-1)、针叶林(55.2-291.0 mg·g-1)、灌丛(139.0-208.9 mg·g-1)和草地(161.0- 371.0 mg·g-1)土壤中性糖的相对含量无显著差异(p > 0.05; 图1B), 除灌丛外, 其他类型的中性糖相对含量均显著高于耕地(65.0-370.7 mg·g-1)(p < 0.05; 图1B).这与上述中性糖的绝对含量结果一致, 说明耕种会通过减少植物残体的输入从而使土壤中性糖含量减少.也有研究发现虽然土地利用类型由森林转变为灌丛, 但主要的维管束植物种类未改变, 凋落物质量没有变化导致两种土地利用类型土壤中性糖含量无显著差异(Conti et al., 2016).而Zhao等(2014)在青藏高原的研究表明, 草地土壤中性糖的相对含量显著高于灌丛, 可能是草本植物凋落物的半纤维素含量比灌丛植物凋落物中的含量高导致的. ...
Small-scale spatial patterns of soil organic carbon and nitrogen stocks in permafrost-affected soils of northern Siberia 2 2018
... 本研究整合了5种土地利用类型的土壤中性糖数据, 发现阔叶林(7.0-82.8 mg·g-1)、针叶林(1.1-90.1 mg·g-1)、灌丛(2.5-80.6 mg·g-1)和草地(1.1-15.1 mg·g-1)土壤中性糖的绝对含量无显著差异(p > 0.05; 图1A), 除草地外, 其他类型均显著高于耕地土壤(0.6-6.3 mg·g-1)(p < 0.05; 图1A).不同地区(西班牙西北部、美国中西部、法国西南部)、不同土壤类型(碳酸钙土壤、沙质淋溶土)中的研究均表明阔叶林和针叶林土壤中性糖(或碳水化合物, 包括纤维素和半纤维中性糖, 下同)的绝对含量显著高于耕地土壤, 并认为可能是人为收获引起的地上输入量减少, 以及耕作扰动导致土壤有机层中的有机质向矿质层迁移造成的(Martens et al., 2004; Jolivet et al., 2006; Llorente et al., 2017).但是有研究报道中性糖(或碳水化合物)在草地土壤中更丰富(与耕地相比)(Larré-Larrouy et al., 2004; Martens et al., 2004; Guan et al., 2018).如, Guan等(2018)发现青藏高原的自然草地经过长期(>50年)耕种后, 植物地下生物量降低使植物碳向土壤中的输入减少, 加上耕作的扰动作用强烈影响了微生物的群落结构和丰度, 导致与草地相比, 耕地土壤中性糖含量降低了57%.Larré-Larrouy等(2004)也认为农作物的根系不发达, 来源于根系的碳水化合物较少是导致此现象的主要原因.由于本研究的数据基于土壤类型、气候条件等各异的研究, 中性糖含量在各研究之间存在差异, 加之数据量的限制导致了与上述研究的结果不一致.
Content and distribution of neutral sugars across different land-use regimes in the top soils (<xref ref-type="bibr" rid="b58">Nierop<i> et al</i>., 2001</xref>; <xref ref-type="bibr" rid="b71">Spielvogel<i> et al</i>., 2007</xref>; <xref ref-type="bibr" rid="b25">Eder<i> et al</i>., 2010</xref>; <xref ref-type="bibr" rid="b67">Rumpel<i> et al</i>., 2010</xref>; <xref ref-type="bibr" rid="b90">Zhao<i> et al</i>., 2014</xref>; <xref ref-type="bibr" rid="b19">Conti<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b21">Cui<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b82">Wang<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b20">Creme<i> et al</i>., 2017</xref>; <xref ref-type="bibr" rid="b49">Llorente<i> et al</i>., 2017</xref>; <xref ref-type="bibr" rid="b26">Evgrafova <i>et al</i>., 2018</xref>; <xref ref-type="bibr" rid="b91">Zhu <i>et al</i>., 2018</xref>). A, Neutral sugar absolute content. B, Neutral sugar relative content. C, GM/AX ((galactose + mannose)/(arabinose + xylose)). D, RF/AX ((rhamnose + fucose)/(arabinose + xylose)). The upper and lower end of boxes denote the 0.25 and 0.75 percentiles, respectively. The solid bar in the box mark the median of each dataset. The circles indicate outliers of each dataset. Different lowercase letters indicate differences in various land-use regimes (<i>p</i> < 0.05). <i>n</i> = 8, 25, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the A, C). <i>n</i> = 8, 22, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the B). <i>n</i> = 3, 19, 4, 8, 27 (from deciduous, coniferous, shrub, grassland to crops in the D). OC, soil organic carbon.Fig. 1 为考察不同土地利用类型中的中性糖对土壤有机碳(OC)的潜在贡献, 我们利用土壤有机碳含量对中性糖含量进行了归一化.结果显示: 阔叶林(84.5-253.9 mg·g-1)、针叶林(55.2-291.0 mg·g-1)、灌丛(139.0-208.9 mg·g-1)和草地(161.0- 371.0 mg·g-1)土壤中性糖的相对含量无显著差异(p > 0.05; 图1B), 除灌丛外, 其他类型的中性糖相对含量均显著高于耕地(65.0-370.7 mg·g-1)(p < 0.05; 图1B).这与上述中性糖的绝对含量结果一致, 说明耕种会通过减少植物残体的输入从而使土壤中性糖含量减少.也有研究发现虽然土地利用类型由森林转变为灌丛, 但主要的维管束植物种类未改变, 凋落物质量没有变化导致两种土地利用类型土壤中性糖含量无显著差异(Conti et al., 2016).而Zhao等(2014)在青藏高原的研究表明, 草地土壤中性糖的相对含量显著高于灌丛, 可能是草本植物凋落物的半纤维素含量比灌丛植物凋落物中的含量高导致的. ...
... ; Evgrafova et al., 2018; Zhu et al., 2018). A, Neutral sugar absolute content. B, Neutral sugar relative content. C, GM/AX ((galactose + mannose)/(arabinose + xylose)). D, RF/AX ((rhamnose + fucose)/(arabinose + xylose)). The upper and lower end of boxes denote the 0.25 and 0.75 percentiles, respectively. The solid bar in the box mark the median of each dataset. The circles indicate outliers of each dataset. Different lowercase letters indicate differences in various land-use regimes (p < 0.05). n = 8, 25, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the A, C). n = 8, 22, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the B). n = 3, 19, 4, 8, 27 (from deciduous, coniferous, shrub, grassland to crops in the D). OC, soil organic carbon. Fig. 1 为考察不同土地利用类型中的中性糖对土壤有机碳(OC)的潜在贡献, 我们利用土壤有机碳含量对中性糖含量进行了归一化.结果显示: 阔叶林(84.5-253.9 mg·g-1)、针叶林(55.2-291.0 mg·g-1)、灌丛(139.0-208.9 mg·g-1)和草地(161.0- 371.0 mg·g-1)土壤中性糖的相对含量无显著差异(p > 0.05; 图1B), 除灌丛外, 其他类型的中性糖相对含量均显著高于耕地(65.0-370.7 mg·g-1)(p < 0.05; 图1B).这与上述中性糖的绝对含量结果一致, 说明耕种会通过减少植物残体的输入从而使土壤中性糖含量减少.也有研究发现虽然土地利用类型由森林转变为灌丛, 但主要的维管束植物种类未改变, 凋落物质量没有变化导致两种土地利用类型土壤中性糖含量无显著差异(Conti et al., 2016).而Zhao等(2014)在青藏高原的研究表明, 草地土壤中性糖的相对含量显著高于灌丛, 可能是草本植物凋落物的半纤维素含量比灌丛植物凋落物中的含量高导致的. ...
Resolution of common dietary sugars from probe sugars for test of intestinal permeability using capillary column gas chromatography 1 2006
... 糖肟三甲基硅醚衍生化(trimethylsilyl oximes, TMSO)或三甲基硅烷基肟衍生化(trimethylsilyl alkyl oximes)以及三甲基硅烷基衍生化(trimethylsilyl, TMS)也被用于醛糖和酮糖的测定(Andrews, 1989; Amelung et al., 1996; Larré-Larrouy & Feller, 1997; Nierop et al., 2001; Farhadi et al., 2006), 此类衍生化步骤较简单, 但会产生多峰, 也易受到水的影响, 同时衍生化产物也不稳定(Ruiz-Matute et al., 2011). ...
Physical control of soil organic matter dynamics in the tropics 1 1997
... 碳水化合物, 即糖类, 可分为单糖、低聚糖和多糖(Cheshire et al., 1992).碳水化合物占维管束植物干质量的75%, 细菌干质量的40% (Moers et al., 1993), 土壤中来源于植物和微生物的碳水化合物占土壤有机质的5%-25%, 是土壤有机质的重要组分(Angers et al., 1988; Murata et al., 1999).碳水化合物是土壤微生物的主要能量来源和碳源(Amelung et al., 1996), 几乎所有的低聚糖和单糖都可溶于水, 属于土壤中易降解的有机质, 易被微生物利用, 并用于自身生长代谢(Gunina & Kuzyakov, 2015).碳水化合物还可驱动微生物由休眠状态转变为活跃状态(Blagodatskaya & Kuzyakov, 2013), 是激发效应产生的有效基质(Kuzyakov, 2010).此外, 碳水化合物是团聚体形成的重要胶结物质(Feller & Beare, 1997), 例如, 阿拉伯糖和木糖是植物黏胶糖的重要来源, 对团聚体的形成起着促进作用(Oades, 1984).Larré-Larrouy等(2004)研究发现, 耕作后土壤团聚体稳定性的改变与团聚体中木糖的减少有关.Angers和Mehuys (1989)提出碳水化合物含量和团聚体的平均质量直径正相关, 表明碳水化合物含量的变化对水稳定性团聚体的改变有着一定的影响.此外, Puget等(1999)发现在更稳定的黏-粉团聚体中微生物来源的碳水化合物较丰富, 并推测团聚体的稳定性受胞外的微生物多糖调控.由此可见, 碳水化合物在团聚体稳定性方面扮演着重要角色.综上所述, 研究土壤碳水化合物对于评估土壤有机质的组成、来源、质量与动态变化具有重要意义(Guggenberger et al., 1994; Trouve et al., 1996; Nacro et al., 2005; Navarrete & Tsutsuki, 2008). ...
Microbial uptake of low-molecular-weight organic substances out-competes sorption in soil 1 2010
Determination of carbohydrates in hydrolysates of volcanic ash soil by liquid chromatography with fluorescence spectroscopy 1 1984
... Comparison of extraction and detection methods of neutral sugars in soils Table 1
步骤 Procedure
方法 Method
优点 Advantage
缺点 Drawback
参考文献 Reference
提取 Extraction
硫酸 H2SO4
硫酸不易被除去 H2SO4 can not be removed easily
Tanaka et al., 1990
盐酸 HCl
会水解一部分纤维素; 产率较低 The hydrolysis products include a few cellulosic neutral sugars; low yields
Uzaki & Ishiwatari, 1983
三氟乙酸 TFA
产率高; 不会破坏单糖结构; 具有挥发性, 可通过旋转蒸发去除; 水解的多糖主要为半纤维素 High yields; Not destructive to monosaccharides; TFA is volatile and can be easily removed by evaporation; Hydrolysis products are mainly released from hemicellulose
Amelung et al., 1996
检测 Detection
GC-MS
精度、准确度、敏感性和效率较高 High accuracy, precision, sensibility and efficiency
需要衍生化 Derivatization is required
Amelung et al., 1996; Wang et al., 2017
HPLC
无需衍生化; 纯化过程简单 No need for derivatization; Simple purification procedures
精度、准确度、敏感性较低 Low accuracy, precision, sensibility and efficiency
Hamada & Ono, 1984; Angers et al., 1988;Tanaka et al., 1990;
HPAEC-PAD
无需衍生化; 应用范围广, 可同时分析糖醛酸和中性糖 No need for derivatization; Wide application and simultaneous analysis of uronic acid and neutral sugars
精度、准确度、敏感性较低 Low accuracy, precision, sensibility and efficiency
Bruggink et al., 2005; Zhang et al., 2012
TFA, trifluoroacetic acid; GC-MS, gas chromatography-mass spectrometry; HPLC, high-performance liquid chromatography; HPAEC-PAD, high-performance anion-exchange chromatography with pulsed amperometric detection. ...
Soil carbohydrates in aggrading and degrading agroecosystems: Influences of fungi and aggregates 1 1995
... 本研究整合了5种土地利用类型的土壤中性糖数据, 发现阔叶林(7.0-82.8 mg·g-1)、针叶林(1.1-90.1 mg·g-1)、灌丛(2.5-80.6 mg·g-1)和草地(1.1-15.1 mg·g-1)土壤中性糖的绝对含量无显著差异(p > 0.05; 图1A), 除草地外, 其他类型均显著高于耕地土壤(0.6-6.3 mg·g-1)(p < 0.05; 图1A).不同地区(西班牙西北部、美国中西部、法国西南部)、不同土壤类型(碳酸钙土壤、沙质淋溶土)中的研究均表明阔叶林和针叶林土壤中性糖(或碳水化合物, 包括纤维素和半纤维中性糖, 下同)的绝对含量显著高于耕地土壤, 并认为可能是人为收获引起的地上输入量减少, 以及耕作扰动导致土壤有机层中的有机质向矿质层迁移造成的(Martens et al., 2004; Jolivet et al., 2006; Llorente et al., 2017).但是有研究报道中性糖(或碳水化合物)在草地土壤中更丰富(与耕地相比)(Larré-Larrouy et al., 2004; Martens et al., 2004; Guan et al., 2018).如, Guan等(2018)发现青藏高原的自然草地经过长期(>50年)耕种后, 植物地下生物量降低使植物碳向土壤中的输入减少, 加上耕作的扰动作用强烈影响了微生物的群落结构和丰度, 导致与草地相比, 耕地土壤中性糖含量降低了57%.Larré-Larrouy等(2004)也认为农作物的根系不发达, 来源于根系的碳水化合物较少是导致此现象的主要原因.由于本研究的数据基于土壤类型、气候条件等各异的研究, 中性糖含量在各研究之间存在差异, 加之数据量的限制导致了与上述研究的结果不一致. ...
... ; Llorente et al., 2017; Evgrafova et al., 2018; Zhu et al., 2018).A, 中性糖绝对含量.B, 中性糖相对含量.C, GM/AX ((半乳糖+甘露糖)/(阿拉伯糖+木糖)).D, RF/AX ((鼠李糖+岩藻糖)/(阿拉伯糖+木糖)).箱式图上方和下方的线段分别表示上四分位数和下四分位数,箱式图内部的横线表示数据的中位数,箱式图中在最上方或最下方的圆圈表示样本数据中的极端值.不同小写字母表示不同土地利用类型间差异显著(p < 0.05).A, C, 阔叶林n = 8, 针叶林n = 25, 灌丛n = 4, 草地n = 8, 耕地n = 15.B, 阔叶林n = 8, 针叶林n = 22, 灌丛n = 4, 草地n = 8, 耕地n = 15.D, 阔叶林n = 3, 针叶林n = 19, 灌丛n = 4, 草地n = 8, 耕地n = 27.OC, 土壤有机碳. Content and distribution of neutral sugars across different land-use regimes in the top soils (<xref ref-type="bibr" rid="b58">Nierop<i> et al</i>., 2001</xref>; <xref ref-type="bibr" rid="b71">Spielvogel<i> et al</i>., 2007</xref>; <xref ref-type="bibr" rid="b25">Eder<i> et al</i>., 2010</xref>; <xref ref-type="bibr" rid="b67">Rumpel<i> et al</i>., 2010</xref>; <xref ref-type="bibr" rid="b90">Zhao<i> et al</i>., 2014</xref>; <xref ref-type="bibr" rid="b19">Conti<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b21">Cui<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b82">Wang<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b20">Creme<i> et al</i>., 2017</xref>; <xref ref-type="bibr" rid="b49">Llorente<i> et al</i>., 2017</xref>; <xref ref-type="bibr" rid="b26">Evgrafova <i>et al</i>., 2018</xref>; <xref ref-type="bibr" rid="b91">Zhu <i>et al</i>., 2018</xref>). A, Neutral sugar absolute content. B, Neutral sugar relative content. C, GM/AX ((galactose + mannose)/(arabinose + xylose)). D, RF/AX ((rhamnose + fucose)/(arabinose + xylose)). The upper and lower end of boxes denote the 0.25 and 0.75 percentiles, respectively. The solid bar in the box mark the median of each dataset. The circles indicate outliers of each dataset. Different lowercase letters indicate differences in various land-use regimes (<i>p</i> < 0.05). <i>n</i> = 8, 25, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the A, C). <i>n</i> = 8, 22, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the B). <i>n</i> = 3, 19, 4, 8, 27 (from deciduous, coniferous, shrub, grassland to crops in the D). OC, soil organic carbon.Fig. 1 为考察不同土地利用类型中的中性糖对土壤有机碳(OC)的潜在贡献, 我们利用土壤有机碳含量对中性糖含量进行了归一化.结果显示: 阔叶林(84.5-253.9 mg·g-1)、针叶林(55.2-291.0 mg·g-1)、灌丛(139.0-208.9 mg·g-1)和草地(161.0- 371.0 mg·g-1)土壤中性糖的相对含量无显著差异(p > 0.05; 图1B), 除灌丛外, 其他类型的中性糖相对含量均显著高于耕地(65.0-370.7 mg·g-1)(p < 0.05; 图1B).这与上述中性糖的绝对含量结果一致, 说明耕种会通过减少植物残体的输入从而使土壤中性糖含量减少.也有研究发现虽然土地利用类型由森林转变为灌丛, 但主要的维管束植物种类未改变, 凋落物质量没有变化导致两种土地利用类型土壤中性糖含量无显著差异(Conti et al., 2016).而Zhao等(2014)在青藏高原的研究表明, 草地土壤中性糖的相对含量显著高于灌丛, 可能是草本植物凋落物的半纤维素含量比灌丛植物凋落物中的含量高导致的. ...
... ; Llorente et al., 2017; Evgrafova et al., 2018; Zhu et al., 2018). A, Neutral sugar absolute content. B, Neutral sugar relative content. C, GM/AX ((galactose + mannose)/(arabinose + xylose)). D, RF/AX ((rhamnose + fucose)/(arabinose + xylose)). The upper and lower end of boxes denote the 0.25 and 0.75 percentiles, respectively. The solid bar in the box mark the median of each dataset. The circles indicate outliers of each dataset. Different lowercase letters indicate differences in various land-use regimes (p < 0.05). n = 8, 25, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the A, C). n = 8, 22, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the B). n = 3, 19, 4, 8, 27 (from deciduous, coniferous, shrub, grassland to crops in the D). OC, soil organic carbon. Fig. 1 为考察不同土地利用类型中的中性糖对土壤有机碳(OC)的潜在贡献, 我们利用土壤有机碳含量对中性糖含量进行了归一化.结果显示: 阔叶林(84.5-253.9 mg·g-1)、针叶林(55.2-291.0 mg·g-1)、灌丛(139.0-208.9 mg·g-1)和草地(161.0- 371.0 mg·g-1)土壤中性糖的相对含量无显著差异(p > 0.05; 图1B), 除灌丛外, 其他类型的中性糖相对含量均显著高于耕地(65.0-370.7 mg·g-1)(p < 0.05; 图1B).这与上述中性糖的绝对含量结果一致, 说明耕种会通过减少植物残体的输入从而使土壤中性糖含量减少.也有研究发现虽然土地利用类型由森林转变为灌丛, 但主要的维管束植物种类未改变, 凋落物质量没有变化导致两种土地利用类型土壤中性糖含量无显著差异(Conti et al., 2016).而Zhao等(2014)在青藏高原的研究表明, 草地土壤中性糖的相对含量显著高于灌丛, 可能是草本植物凋落物的半纤维素含量比灌丛植物凋落物中的含量高导致的. ...
... 按密度分组, 中性糖在轻质组分中的相对含量(129.0-410.0 mg·g-1)较重质组分中(7.5-322.0 mg·g-1)高.Llorente等(2017)在含碳酸钙的土壤中也发现中性糖含量在游离态组分中最高, 闭蓄态组分次之, 重质组分中最低; 由于微生物更易进入, 因此游离态组分中含有较多的甘露糖.另外, Bischoff等(2018)在西伯利亚高原不同盐分梯度的土壤中也发现, 无论是在表层还是底层, 中性糖含量在轻质组分中最高.但是, 与全土相比, 与矿质相结合的重质组分中中性糖含量更丰富, 该组分中以微生物来源的六碳糖最为丰富; 此外, 该研究中放射性碳的数据显示年龄较老的土壤样品中微生物来源的中性糖含量较高, 说明中性糖在矿质土中的稳定储存是通过连续的微生物转化或矿物的物理保护实现的(Rumpel et al., 2010).Dao等(2018)在西伯利亚东北部冻土中的研究也发现微生物来源的中性糖在重质组分土壤中含量较丰富, 从较低的C:N和较高的碳稳定同位素比率(δ13C)可以看出这与微生物来源的中性糖的选择性积累有关(Gentsch et al., 2015a, 2015b).此外, 微生物残体(Miltner et al., 2012)及微生物分泌的胞外多聚物中富含的多糖(Kleber et al., 2015), 也可能通过矿物的吸附作用在重质组分土壤中积累. ...
Soil saccharide extraction and detection 1 1993
... 土壤中性糖的提取主要包括水解、纯化和检测三个步骤.水解可以破坏糖类与土壤基质之间的氢键、共价键(Martens & Frankenberger, 1993), 以及多糖分子结构内部的键合作用(如糖苷键)(K?gel- Knabner, 2002), 将中性单糖从半纤维素中释放出来.土壤中性单糖的水解一般用硫酸(Sowden & Ivarson, 1962; Oades et al., 1970; Cheshire, 1979)、盐酸(Uzaki & Ishiwatari, 1983)或三氟乙酸(Amelung et al., 1996)实现(表1).由于硫酸不易被除去, 而三氟乙酸挥发性强, 可通过旋转蒸发去除, 且提取效率也较硫酸高, 因此三氟乙酸水解得到了广泛的应用(Amelung et al., 1996; Rumpel & Dignac, 2006; Zhang et al., 2007; Eder et al., 2010).目前最常用的水解方法是利用4 mol·L-1的三氟乙酸在105 ℃水解4 h, 既不会破坏单糖的结构, 也能获得最高的提取效率(Amelung et al., 1996).由于盐酸的水解效率较三氟乙酸和硫酸低, 该方法基本不再使用(Amelung et al., 1996).上述水解产物的纯化通常使用旋转蒸发法去除三氟乙酸, 使用螯合剂防止铁的氢氧化物(或氧化物)干扰, 或调节pH值将铁的氢氧化物(或氧化物)以沉淀形式去除(Zhang et al., 2007; Eder et al., 2010), 也有研究使用吸附剂和阳离子交换树脂去除杂质(Amelung et al., 1996; Martens et al., 2004). ...
Soil organic carbon content and composition of 130-year crop, pasture and forest land-use managements 3 2004
... 土壤中性糖的提取主要包括水解、纯化和检测三个步骤.水解可以破坏糖类与土壤基质之间的氢键、共价键(Martens & Frankenberger, 1993), 以及多糖分子结构内部的键合作用(如糖苷键)(K?gel- Knabner, 2002), 将中性单糖从半纤维素中释放出来.土壤中性单糖的水解一般用硫酸(Sowden & Ivarson, 1962; Oades et al., 1970; Cheshire, 1979)、盐酸(Uzaki & Ishiwatari, 1983)或三氟乙酸(Amelung et al., 1996)实现(表1).由于硫酸不易被除去, 而三氟乙酸挥发性强, 可通过旋转蒸发去除, 且提取效率也较硫酸高, 因此三氟乙酸水解得到了广泛的应用(Amelung et al., 1996; Rumpel & Dignac, 2006; Zhang et al., 2007; Eder et al., 2010).目前最常用的水解方法是利用4 mol·L-1的三氟乙酸在105 ℃水解4 h, 既不会破坏单糖的结构, 也能获得最高的提取效率(Amelung et al., 1996).由于盐酸的水解效率较三氟乙酸和硫酸低, 该方法基本不再使用(Amelung et al., 1996).上述水解产物的纯化通常使用旋转蒸发法去除三氟乙酸, 使用螯合剂防止铁的氢氧化物(或氧化物)干扰, 或调节pH值将铁的氢氧化物(或氧化物)以沉淀形式去除(Zhang et al., 2007; Eder et al., 2010), 也有研究使用吸附剂和阳离子交换树脂去除杂质(Amelung et al., 1996; Martens et al., 2004). ...
... 本研究整合了5种土地利用类型的土壤中性糖数据, 发现阔叶林(7.0-82.8 mg·g-1)、针叶林(1.1-90.1 mg·g-1)、灌丛(2.5-80.6 mg·g-1)和草地(1.1-15.1 mg·g-1)土壤中性糖的绝对含量无显著差异(p > 0.05; 图1A), 除草地外, 其他类型均显著高于耕地土壤(0.6-6.3 mg·g-1)(p < 0.05; 图1A).不同地区(西班牙西北部、美国中西部、法国西南部)、不同土壤类型(碳酸钙土壤、沙质淋溶土)中的研究均表明阔叶林和针叶林土壤中性糖(或碳水化合物, 包括纤维素和半纤维中性糖, 下同)的绝对含量显著高于耕地土壤, 并认为可能是人为收获引起的地上输入量减少, 以及耕作扰动导致土壤有机层中的有机质向矿质层迁移造成的(Martens et al., 2004; Jolivet et al., 2006; Llorente et al., 2017).但是有研究报道中性糖(或碳水化合物)在草地土壤中更丰富(与耕地相比)(Larré-Larrouy et al., 2004; Martens et al., 2004; Guan et al., 2018).如, Guan等(2018)发现青藏高原的自然草地经过长期(>50年)耕种后, 植物地下生物量降低使植物碳向土壤中的输入减少, 加上耕作的扰动作用强烈影响了微生物的群落结构和丰度, 导致与草地相比, 耕地土壤中性糖含量降低了57%.Larré-Larrouy等(2004)也认为农作物的根系不发达, 来源于根系的碳水化合物较少是导致此现象的主要原因.由于本研究的数据基于土壤类型、气候条件等各异的研究, 中性糖含量在各研究之间存在差异, 加之数据量的限制导致了与上述研究的结果不一致. ...
... ; Martens et al., 2004; Guan et al., 2018).如, Guan等(2018)发现青藏高原的自然草地经过长期(>50年)耕种后, 植物地下生物量降低使植物碳向土壤中的输入减少, 加上耕作的扰动作用强烈影响了微生物的群落结构和丰度, 导致与草地相比, 耕地土壤中性糖含量降低了57%.Larré-Larrouy等(2004)也认为农作物的根系不发达, 来源于根系的碳水化合物较少是导致此现象的主要原因.由于本研究的数据基于土壤类型、气候条件等各异的研究, 中性糖含量在各研究之间存在差异, 加之数据量的限制导致了与上述研究的结果不一致. ...
Abundance and composition of free and aggregate- occluded carbohydrates and lignin in two forest soils as affected by wildfires of different severity 1 2015
... 影响中性糖含量的因素可以分为自然因素和人为因素.自然因素有环境因素(CO2、O2、海拔、温度、降水等), 生物扰动(土壤动物和微生物), 以及自然灾害(如火灾和病虫害).Barron-Gafford等(2005)在半封闭式的生态系统(生物圈Ⅱ号)中的研究表明, 在高CO2浓度处理下, 土壤碳水化合物含量增加, 主要是由植物的地下生物量增加引起的.另外, 有研究使用开放式CO2富集系统(FACE)发现高CO2处理7年后, 由于植物生物量增加, 与对照组相比, 温带草地土壤中性糖含量增加了28% (Bock et al., 2007).与CO2不同, 当泥炭土在O2中的暴露时间延长, 土壤中性糖降解速率加快, 含量降低(Philben et al., 2015).田秋香等(2013)通过对中国长白山不同海拔梯度森林土壤中性糖分布的研究发现, 不同海拔间的温度差异是造成土壤中性糖含量差异的主要原因, 在高海拔的低温环境中微生物的分解作用减弱, 再加上微生物对外源碳的利用效率提高, 被微生物转化后的外源碳以微生物残体的形式固存在土壤中.综上所述, 高海拔有利于中性糖的积累.有关生物扰动的研究发现蚯蚓并不会影响土壤中性糖含量(Phuong-Thi et al., 2012), 而土壤微生物对于土壤中性糖的降解影响很大, 当微生物多样性较高时, 单糖如阿拉伯糖和木糖的降解速率增加, 含量减少(Baumann et al., 2013).此外, 自然灾害如火灾对土壤中性糖的影响并没有一致的规律, 不同程度的火灾对土壤不同组分的中性糖含量和组成的影响都不同(Mastrolonardo et al., 2015). ...
SOM genesis: Microbial biomass as a significant source 1 2012
... 按密度分组, 中性糖在轻质组分中的相对含量(129.0-410.0 mg·g-1)较重质组分中(7.5-322.0 mg·g-1)高.Llorente等(2017)在含碳酸钙的土壤中也发现中性糖含量在游离态组分中最高, 闭蓄态组分次之, 重质组分中最低; 由于微生物更易进入, 因此游离态组分中含有较多的甘露糖.另外, Bischoff等(2018)在西伯利亚高原不同盐分梯度的土壤中也发现, 无论是在表层还是底层, 中性糖含量在轻质组分中最高.但是, 与全土相比, 与矿质相结合的重质组分中中性糖含量更丰富, 该组分中以微生物来源的六碳糖最为丰富; 此外, 该研究中放射性碳的数据显示年龄较老的土壤样品中微生物来源的中性糖含量较高, 说明中性糖在矿质土中的稳定储存是通过连续的微生物转化或矿物的物理保护实现的(Rumpel et al., 2010).Dao等(2018)在西伯利亚东北部冻土中的研究也发现微生物来源的中性糖在重质组分土壤中含量较丰富, 从较低的C:N和较高的碳稳定同位素比率(δ13C)可以看出这与微生物来源的中性糖的选择性积累有关(Gentsch et al., 2015a, 2015b).此外, 微生物残体(Miltner et al., 2012)及微生物分泌的胞外多聚物中富含的多糖(Kleber et al., 2015), 也可能通过矿物的吸附作用在重质组分土壤中积累. ...
Carbohydrate diagenesis in hypersaline environments: Application of GC-IRMS to the stable isotope analysis of derivatized saccharides from surficial and buried sediments 1 1993
... 碳水化合物, 即糖类, 可分为单糖、低聚糖和多糖(Cheshire et al., 1992).碳水化合物占维管束植物干质量的75%, 细菌干质量的40% (Moers et al., 1993), 土壤中来源于植物和微生物的碳水化合物占土壤有机质的5%-25%, 是土壤有机质的重要组分(Angers et al., 1988; Murata et al., 1999).碳水化合物是土壤微生物的主要能量来源和碳源(Amelung et al., 1996), 几乎所有的低聚糖和单糖都可溶于水, 属于土壤中易降解的有机质, 易被微生物利用, 并用于自身生长代谢(Gunina & Kuzyakov, 2015).碳水化合物还可驱动微生物由休眠状态转变为活跃状态(Blagodatskaya & Kuzyakov, 2013), 是激发效应产生的有效基质(Kuzyakov, 2010).此外, 碳水化合物是团聚体形成的重要胶结物质(Feller & Beare, 1997), 例如, 阿拉伯糖和木糖是植物黏胶糖的重要来源, 对团聚体的形成起着促进作用(Oades, 1984).Larré-Larrouy等(2004)研究发现, 耕作后土壤团聚体稳定性的改变与团聚体中木糖的减少有关.Angers和Mehuys (1989)提出碳水化合物含量和团聚体的平均质量直径正相关, 表明碳水化合物含量的变化对水稳定性团聚体的改变有着一定的影响.此外, Puget等(1999)发现在更稳定的黏-粉团聚体中微生物来源的碳水化合物较丰富, 并推测团聚体的稳定性受胞外的微生物多糖调控.由此可见, 碳水化合物在团聚体稳定性方面扮演着重要角色.综上所述, 研究土壤碳水化合物对于评估土壤有机质的组成、来源、质量与动态变化具有重要意义(Guggenberger et al., 1994; Trouve et al., 1996; Nacro et al., 2005; Navarrete & Tsutsuki, 2008). ...
Seasonal variations in soil microbial biomass content and soil neutral sugar composition in grassland in the Japanese Temperate Zone 1 1999
... 碳水化合物, 即糖类, 可分为单糖、低聚糖和多糖(Cheshire et al., 1992).碳水化合物占维管束植物干质量的75%, 细菌干质量的40% (Moers et al., 1993), 土壤中来源于植物和微生物的碳水化合物占土壤有机质的5%-25%, 是土壤有机质的重要组分(Angers et al., 1988; Murata et al., 1999).碳水化合物是土壤微生物的主要能量来源和碳源(Amelung et al., 1996), 几乎所有的低聚糖和单糖都可溶于水, 属于土壤中易降解的有机质, 易被微生物利用, 并用于自身生长代谢(Gunina & Kuzyakov, 2015).碳水化合物还可驱动微生物由休眠状态转变为活跃状态(Blagodatskaya & Kuzyakov, 2013), 是激发效应产生的有效基质(Kuzyakov, 2010).此外, 碳水化合物是团聚体形成的重要胶结物质(Feller & Beare, 1997), 例如, 阿拉伯糖和木糖是植物黏胶糖的重要来源, 对团聚体的形成起着促进作用(Oades, 1984).Larré-Larrouy等(2004)研究发现, 耕作后土壤团聚体稳定性的改变与团聚体中木糖的减少有关.Angers和Mehuys (1989)提出碳水化合物含量和团聚体的平均质量直径正相关, 表明碳水化合物含量的变化对水稳定性团聚体的改变有着一定的影响.此外, Puget等(1999)发现在更稳定的黏-粉团聚体中微生物来源的碳水化合物较丰富, 并推测团聚体的稳定性受胞外的微生物多糖调控.由此可见, 碳水化合物在团聚体稳定性方面扮演着重要角色.综上所述, 研究土壤碳水化合物对于评估土壤有机质的组成、来源、质量与动态变化具有重要意义(Guggenberger et al., 1994; Trouve et al., 1996; Nacro et al., 2005; Navarrete & Tsutsuki, 2008). ...
Hydrolysable carbohydrate in tropical soils under adjacent forest and savanna vegetation in Lamto, C?te d’Ivoire 1 2005
... 碳水化合物, 即糖类, 可分为单糖、低聚糖和多糖(Cheshire et al., 1992).碳水化合物占维管束植物干质量的75%, 细菌干质量的40% (Moers et al., 1993), 土壤中来源于植物和微生物的碳水化合物占土壤有机质的5%-25%, 是土壤有机质的重要组分(Angers et al., 1988; Murata et al., 1999).碳水化合物是土壤微生物的主要能量来源和碳源(Amelung et al., 1996), 几乎所有的低聚糖和单糖都可溶于水, 属于土壤中易降解的有机质, 易被微生物利用, 并用于自身生长代谢(Gunina & Kuzyakov, 2015).碳水化合物还可驱动微生物由休眠状态转变为活跃状态(Blagodatskaya & Kuzyakov, 2013), 是激发效应产生的有效基质(Kuzyakov, 2010).此外, 碳水化合物是团聚体形成的重要胶结物质(Feller & Beare, 1997), 例如, 阿拉伯糖和木糖是植物黏胶糖的重要来源, 对团聚体的形成起着促进作用(Oades, 1984).Larré-Larrouy等(2004)研究发现, 耕作后土壤团聚体稳定性的改变与团聚体中木糖的减少有关.Angers和Mehuys (1989)提出碳水化合物含量和团聚体的平均质量直径正相关, 表明碳水化合物含量的变化对水稳定性团聚体的改变有着一定的影响.此外, Puget等(1999)发现在更稳定的黏-粉团聚体中微生物来源的碳水化合物较丰富, 并推测团聚体的稳定性受胞外的微生物多糖调控.由此可见, 碳水化合物在团聚体稳定性方面扮演着重要角色.综上所述, 研究土壤碳水化合物对于评估土壤有机质的组成、来源、质量与动态变化具有重要意义(Guggenberger et al., 1994; Trouve et al., 1996; Nacro et al., 2005; Navarrete & Tsutsuki, 2008). ...
Land-use impact on soil carbon, nitrogen, neutral sugar composition and related chemical properties in a degraded Ultisol in Leyte, Philippines 1 2008
... 碳水化合物, 即糖类, 可分为单糖、低聚糖和多糖(Cheshire et al., 1992).碳水化合物占维管束植物干质量的75%, 细菌干质量的40% (Moers et al., 1993), 土壤中来源于植物和微生物的碳水化合物占土壤有机质的5%-25%, 是土壤有机质的重要组分(Angers et al., 1988; Murata et al., 1999).碳水化合物是土壤微生物的主要能量来源和碳源(Amelung et al., 1996), 几乎所有的低聚糖和单糖都可溶于水, 属于土壤中易降解的有机质, 易被微生物利用, 并用于自身生长代谢(Gunina & Kuzyakov, 2015).碳水化合物还可驱动微生物由休眠状态转变为活跃状态(Blagodatskaya & Kuzyakov, 2013), 是激发效应产生的有效基质(Kuzyakov, 2010).此外, 碳水化合物是团聚体形成的重要胶结物质(Feller & Beare, 1997), 例如, 阿拉伯糖和木糖是植物黏胶糖的重要来源, 对团聚体的形成起着促进作用(Oades, 1984).Larré-Larrouy等(2004)研究发现, 耕作后土壤团聚体稳定性的改变与团聚体中木糖的减少有关.Angers和Mehuys (1989)提出碳水化合物含量和团聚体的平均质量直径正相关, 表明碳水化合物含量的变化对水稳定性团聚体的改变有着一定的影响.此外, Puget等(1999)发现在更稳定的黏-粉团聚体中微生物来源的碳水化合物较丰富, 并推测团聚体的稳定性受胞外的微生物多糖调控.由此可见, 碳水化合物在团聚体稳定性方面扮演着重要角色.综上所述, 研究土壤碳水化合物对于评估土壤有机质的组成、来源、质量与动态变化具有重要意义(Guggenberger et al., 1994; Trouve et al., 1996; Nacro et al., 2005; Navarrete & Tsutsuki, 2008). ...
Composition of plant tissues and soil organic matter in the first stages of a vegetation succession 4 2001
Content and distribution of neutral sugars across different land-use regimes in the top soils (<xref ref-type="bibr" rid="b58">Nierop<i> et al</i>., 2001</xref>; <xref ref-type="bibr" rid="b71">Spielvogel<i> et al</i>., 2007</xref>; <xref ref-type="bibr" rid="b25">Eder<i> et al</i>., 2010</xref>; <xref ref-type="bibr" rid="b67">Rumpel<i> et al</i>., 2010</xref>; <xref ref-type="bibr" rid="b90">Zhao<i> et al</i>., 2014</xref>; <xref ref-type="bibr" rid="b19">Conti<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b21">Cui<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b82">Wang<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b20">Creme<i> et al</i>., 2017</xref>; <xref ref-type="bibr" rid="b49">Llorente<i> et al</i>., 2017</xref>; <xref ref-type="bibr" rid="b26">Evgrafova <i>et al</i>., 2018</xref>; <xref ref-type="bibr" rid="b91">Zhu <i>et al</i>., 2018</xref>). A, Neutral sugar absolute content. B, Neutral sugar relative content. C, GM/AX ((galactose + mannose)/(arabinose + xylose)). D, RF/AX ((rhamnose + fucose)/(arabinose + xylose)). The upper and lower end of boxes denote the 0.25 and 0.75 percentiles, respectively. The solid bar in the box mark the median of each dataset. The circles indicate outliers of each dataset. Different lowercase letters indicate differences in various land-use regimes (<i>p</i> < 0.05). <i>n</i> = 8, 25, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the A, C). <i>n</i> = 8, 22, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the B). <i>n</i> = 3, 19, 4, 8, 27 (from deciduous, coniferous, shrub, grassland to crops in the D). OC, soil organic carbon.Fig. 1 为考察不同土地利用类型中的中性糖对土壤有机碳(OC)的潜在贡献, 我们利用土壤有机碳含量对中性糖含量进行了归一化.结果显示: 阔叶林(84.5-253.9 mg·g-1)、针叶林(55.2-291.0 mg·g-1)、灌丛(139.0-208.9 mg·g-1)和草地(161.0- 371.0 mg·g-1)土壤中性糖的相对含量无显著差异(p > 0.05; 图1B), 除灌丛外, 其他类型的中性糖相对含量均显著高于耕地(65.0-370.7 mg·g-1)(p < 0.05; 图1B).这与上述中性糖的绝对含量结果一致, 说明耕种会通过减少植物残体的输入从而使土壤中性糖含量减少.也有研究发现虽然土地利用类型由森林转变为灌丛, 但主要的维管束植物种类未改变, 凋落物质量没有变化导致两种土地利用类型土壤中性糖含量无显著差异(Conti et al., 2016).而Zhao等(2014)在青藏高原的研究表明, 草地土壤中性糖的相对含量显著高于灌丛, 可能是草本植物凋落物的半纤维素含量比灌丛植物凋落物中的含量高导致的. ...
... Content and distribution of neutral sugars across different land-use regimes in the top soils (Nierop et al., 2001; Spielvogel et al., 2007; Eder et al., 2010; Rumpel et al., 2010; Zhao et al., 2014; Conti et al., 2016; Cui et al., 2016; Wang et al., 2016; Creme et al., 2017; Llorente et al., 2017; Evgrafova et al., 2018; Zhu et al., 2018). A, Neutral sugar absolute content. B, Neutral sugar relative content. C, GM/AX ((galactose + mannose)/(arabinose + xylose)). D, RF/AX ((rhamnose + fucose)/(arabinose + xylose)). The upper and lower end of boxes denote the 0.25 and 0.75 percentiles, respectively. The solid bar in the box mark the median of each dataset. The circles indicate outliers of each dataset. Different lowercase letters indicate differences in various land-use regimes (p < 0.05). n = 8, 25, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the A, C). n = 8, 22, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the B). n = 3, 19, 4, 8, 27 (from deciduous, coniferous, shrub, grassland to crops in the D). OC, soil organic carbon. Fig. 1 为考察不同土地利用类型中的中性糖对土壤有机碳(OC)的潜在贡献, 我们利用土壤有机碳含量对中性糖含量进行了归一化.结果显示: 阔叶林(84.5-253.9 mg·g-1)、针叶林(55.2-291.0 mg·g-1)、灌丛(139.0-208.9 mg·g-1)和草地(161.0- 371.0 mg·g-1)土壤中性糖的相对含量无显著差异(p > 0.05; 图1B), 除灌丛外, 其他类型的中性糖相对含量均显著高于耕地(65.0-370.7 mg·g-1)(p < 0.05; 图1B).这与上述中性糖的绝对含量结果一致, 说明耕种会通过减少植物残体的输入从而使土壤中性糖含量减少.也有研究发现虽然土地利用类型由森林转变为灌丛, 但主要的维管束植物种类未改变, 凋落物质量没有变化导致两种土地利用类型土壤中性糖含量无显著差异(Conti et al., 2016).而Zhao等(2014)在青藏高原的研究表明, 草地土壤中性糖的相对含量显著高于灌丛, 可能是草本植物凋落物的半纤维素含量比灌丛植物凋落物中的含量高导致的. ...
Soil organic matter and structural stability: Mechanisms and implications for management 3 1984
... 碳水化合物, 即糖类, 可分为单糖、低聚糖和多糖(Cheshire et al., 1992).碳水化合物占维管束植物干质量的75%, 细菌干质量的40% (Moers et al., 1993), 土壤中来源于植物和微生物的碳水化合物占土壤有机质的5%-25%, 是土壤有机质的重要组分(Angers et al., 1988; Murata et al., 1999).碳水化合物是土壤微生物的主要能量来源和碳源(Amelung et al., 1996), 几乎所有的低聚糖和单糖都可溶于水, 属于土壤中易降解的有机质, 易被微生物利用, 并用于自身生长代谢(Gunina & Kuzyakov, 2015).碳水化合物还可驱动微生物由休眠状态转变为活跃状态(Blagodatskaya & Kuzyakov, 2013), 是激发效应产生的有效基质(Kuzyakov, 2010).此外, 碳水化合物是团聚体形成的重要胶结物质(Feller & Beare, 1997), 例如, 阿拉伯糖和木糖是植物黏胶糖的重要来源, 对团聚体的形成起着促进作用(Oades, 1984).Larré-Larrouy等(2004)研究发现, 耕作后土壤团聚体稳定性的改变与团聚体中木糖的减少有关.Angers和Mehuys (1989)提出碳水化合物含量和团聚体的平均质量直径正相关, 表明碳水化合物含量的变化对水稳定性团聚体的改变有着一定的影响.此外, Puget等(1999)发现在更稳定的黏-粉团聚体中微生物来源的碳水化合物较丰富, 并推测团聚体的稳定性受胞外的微生物多糖调控.由此可见, 碳水化合物在团聚体稳定性方面扮演着重要角色.综上所述, 研究土壤碳水化合物对于评估土壤有机质的组成、来源、质量与动态变化具有重要意义(Guggenberger et al., 1994; Trouve et al., 1996; Nacro et al., 2005; Navarrete & Tsutsuki, 2008). ...
Nature of carbohydrates associated with water-stable aggregates of two cultivated soils 1 1999
... 碳水化合物, 即糖类, 可分为单糖、低聚糖和多糖(Cheshire et al., 1992).碳水化合物占维管束植物干质量的75%, 细菌干质量的40% (Moers et al., 1993), 土壤中来源于植物和微生物的碳水化合物占土壤有机质的5%-25%, 是土壤有机质的重要组分(Angers et al., 1988; Murata et al., 1999).碳水化合物是土壤微生物的主要能量来源和碳源(Amelung et al., 1996), 几乎所有的低聚糖和单糖都可溶于水, 属于土壤中易降解的有机质, 易被微生物利用, 并用于自身生长代谢(Gunina & Kuzyakov, 2015).碳水化合物还可驱动微生物由休眠状态转变为活跃状态(Blagodatskaya & Kuzyakov, 2013), 是激发效应产生的有效基质(Kuzyakov, 2010).此外, 碳水化合物是团聚体形成的重要胶结物质(Feller & Beare, 1997), 例如, 阿拉伯糖和木糖是植物黏胶糖的重要来源, 对团聚体的形成起着促进作用(Oades, 1984).Larré-Larrouy等(2004)研究发现, 耕作后土壤团聚体稳定性的改变与团聚体中木糖的减少有关.Angers和Mehuys (1989)提出碳水化合物含量和团聚体的平均质量直径正相关, 表明碳水化合物含量的变化对水稳定性团聚体的改变有着一定的影响.此外, Puget等(1999)发现在更稳定的黏-粉团聚体中微生物来源的碳水化合物较丰富, 并推测团聚体的稳定性受胞外的微生物多糖调控.由此可见, 碳水化合物在团聚体稳定性方面扮演着重要角色.综上所述, 研究土壤碳水化合物对于评估土壤有机质的组成、来源、质量与动态变化具有重要意义(Guggenberger et al., 1994; Trouve et al., 1996; Nacro et al., 2005; Navarrete & Tsutsuki, 2008). ...
Derivatization of carbohydrates for GC and GC-MS analyses 3 2011
... 糖醇乙酸酯衍生化是目前使用最广泛的一种中性糖衍生化方法, 其原理是将醛糖的羰基还原, 最终形成糖醇乙酸酯(Black & Fox, 1996; Ruiz-Matute et al., 2011).该衍生化的优点是: 得到的每种糖醇产物只产生一个单独的色谱峰, 易于检测, 且衍生化产物较稳定, 可稳定储存20天(Wang et al., 2017), 因此被广泛用于中性糖的定量分析(Rumpel & Dignac, 2006; Spielvogel et al., 2007; Rumpel et al., 2010).然而, 该衍生化方法耗时, 步骤繁琐(Black & Fox, 1996; Rumpel & Dignac, 2006; Ruiz-Matute et al., 2011), 一直有研究在对该方法进行不断改进. ...
... ; Ruiz-Matute et al., 2011), 一直有研究在对该方法进行不断改进. ...
... 糖肟三甲基硅醚衍生化(trimethylsilyl oximes, TMSO)或三甲基硅烷基肟衍生化(trimethylsilyl alkyl oximes)以及三甲基硅烷基衍生化(trimethylsilyl, TMS)也被用于醛糖和酮糖的测定(Andrews, 1989; Amelung et al., 1996; Larré-Larrouy & Feller, 1997; Nierop et al., 2001; Farhadi et al., 2006), 此类衍生化步骤较简单, 但会产生多峰, 也易受到水的影响, 同时衍生化产物也不稳定(Ruiz-Matute et al., 2011). ...
Gas chromatographic analysis of monosaccharides in a forest soil profile: Analysis by gas chromatography after trifluoroacetic acid hydrolysis and reduction-acetylation 5 2006
... 土壤中性糖的提取主要包括水解、纯化和检测三个步骤.水解可以破坏糖类与土壤基质之间的氢键、共价键(Martens & Frankenberger, 1993), 以及多糖分子结构内部的键合作用(如糖苷键)(K?gel- Knabner, 2002), 将中性单糖从半纤维素中释放出来.土壤中性单糖的水解一般用硫酸(Sowden & Ivarson, 1962; Oades et al., 1970; Cheshire, 1979)、盐酸(Uzaki & Ishiwatari, 1983)或三氟乙酸(Amelung et al., 1996)实现(表1).由于硫酸不易被除去, 而三氟乙酸挥发性强, 可通过旋转蒸发去除, 且提取效率也较硫酸高, 因此三氟乙酸水解得到了广泛的应用(Amelung et al., 1996; Rumpel & Dignac, 2006; Zhang et al., 2007; Eder et al., 2010).目前最常用的水解方法是利用4 mol·L-1的三氟乙酸在105 ℃水解4 h, 既不会破坏单糖的结构, 也能获得最高的提取效率(Amelung et al., 1996).由于盐酸的水解效率较三氟乙酸和硫酸低, 该方法基本不再使用(Amelung et al., 1996).上述水解产物的纯化通常使用旋转蒸发法去除三氟乙酸, 使用螯合剂防止铁的氢氧化物(或氧化物)干扰, 或调节pH值将铁的氢氧化物(或氧化物)以沉淀形式去除(Zhang et al., 2007; Eder et al., 2010), 也有研究使用吸附剂和阳离子交换树脂去除杂质(Amelung et al., 1996; Martens et al., 2004). ...
... 中性糖的检测分析可使用气相色谱-质谱联用仪(GC-MS)(Rumpel & Dignac, 2006; Zhang et al., 2007; Eder et al., 2010), 液相色谱(HPLC)(Takeuchi et al., 1987; Tanaka et al., 1990; Amelung et al., 1996; Basler & Dyckmans, 2013)和高效阴离子交换色谱-脉冲安培检测法(HPAEC-PAD)(Bruggink et al., 2005; Zhang et al., 2012).由于中性糖不易挥发, 因此在使用GC-MS检测前需衍生化, 衍生化是影响土壤中性糖定量的关键环节.下面将介绍几种常用的中性糖衍生化方法. ...
... 糖醇乙酸酯衍生化是目前使用最广泛的一种中性糖衍生化方法, 其原理是将醛糖的羰基还原, 最终形成糖醇乙酸酯(Black & Fox, 1996; Ruiz-Matute et al., 2011).该衍生化的优点是: 得到的每种糖醇产物只产生一个单独的色谱峰, 易于检测, 且衍生化产物较稳定, 可稳定储存20天(Wang et al., 2017), 因此被广泛用于中性糖的定量分析(Rumpel & Dignac, 2006; Spielvogel et al., 2007; Rumpel et al., 2010).然而, 该衍生化方法耗时, 步骤繁琐(Black & Fox, 1996; Rumpel & Dignac, 2006; Ruiz-Matute et al., 2011), 一直有研究在对该方法进行不断改进. ...
Content and distribution of neutral sugars across different land-use regimes in the top soils (<xref ref-type="bibr" rid="b58">Nierop<i> et al</i>., 2001</xref>; <xref ref-type="bibr" rid="b71">Spielvogel<i> et al</i>., 2007</xref>; <xref ref-type="bibr" rid="b25">Eder<i> et al</i>., 2010</xref>; <xref ref-type="bibr" rid="b67">Rumpel<i> et al</i>., 2010</xref>; <xref ref-type="bibr" rid="b90">Zhao<i> et al</i>., 2014</xref>; <xref ref-type="bibr" rid="b19">Conti<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b21">Cui<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b82">Wang<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b20">Creme<i> et al</i>., 2017</xref>; <xref ref-type="bibr" rid="b49">Llorente<i> et al</i>., 2017</xref>; <xref ref-type="bibr" rid="b26">Evgrafova <i>et al</i>., 2018</xref>; <xref ref-type="bibr" rid="b91">Zhu <i>et al</i>., 2018</xref>). A, Neutral sugar absolute content. B, Neutral sugar relative content. C, GM/AX ((galactose + mannose)/(arabinose + xylose)). D, RF/AX ((rhamnose + fucose)/(arabinose + xylose)). The upper and lower end of boxes denote the 0.25 and 0.75 percentiles, respectively. The solid bar in the box mark the median of each dataset. The circles indicate outliers of each dataset. Different lowercase letters indicate differences in various land-use regimes (<i>p</i> < 0.05). <i>n</i> = 8, 25, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the A, C). <i>n</i> = 8, 22, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the B). <i>n</i> = 3, 19, 4, 8, 27 (from deciduous, coniferous, shrub, grassland to crops in the D). OC, soil organic carbon.Fig. 1 为考察不同土地利用类型中的中性糖对土壤有机碳(OC)的潜在贡献, 我们利用土壤有机碳含量对中性糖含量进行了归一化.结果显示: 阔叶林(84.5-253.9 mg·g-1)、针叶林(55.2-291.0 mg·g-1)、灌丛(139.0-208.9 mg·g-1)和草地(161.0- 371.0 mg·g-1)土壤中性糖的相对含量无显著差异(p > 0.05; 图1B), 除灌丛外, 其他类型的中性糖相对含量均显著高于耕地(65.0-370.7 mg·g-1)(p < 0.05; 图1B).这与上述中性糖的绝对含量结果一致, 说明耕种会通过减少植物残体的输入从而使土壤中性糖含量减少.也有研究发现虽然土地利用类型由森林转变为灌丛, 但主要的维管束植物种类未改变, 凋落物质量没有变化导致两种土地利用类型土壤中性糖含量无显著差异(Conti et al., 2016).而Zhao等(2014)在青藏高原的研究表明, 草地土壤中性糖的相对含量显著高于灌丛, 可能是草本植物凋落物的半纤维素含量比灌丛植物凋落物中的含量高导致的. ...
... ; Rumpel et al., 2010; Zhao et al., 2014; Conti et al., 2016; Cui et al., 2016; Wang et al., 2016; Creme et al., 2017; Llorente et al., 2017; Evgrafova et al., 2018; Zhu et al., 2018). A, Neutral sugar absolute content. B, Neutral sugar relative content. C, GM/AX ((galactose + mannose)/(arabinose + xylose)). D, RF/AX ((rhamnose + fucose)/(arabinose + xylose)). The upper and lower end of boxes denote the 0.25 and 0.75 percentiles, respectively. The solid bar in the box mark the median of each dataset. The circles indicate outliers of each dataset. Different lowercase letters indicate differences in various land-use regimes (p < 0.05). n = 8, 25, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the A, C). n = 8, 22, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the B). n = 3, 19, 4, 8, 27 (from deciduous, coniferous, shrub, grassland to crops in the D). OC, soil organic carbon. Fig. 1 为考察不同土地利用类型中的中性糖对土壤有机碳(OC)的潜在贡献, 我们利用土壤有机碳含量对中性糖含量进行了归一化.结果显示: 阔叶林(84.5-253.9 mg·g-1)、针叶林(55.2-291.0 mg·g-1)、灌丛(139.0-208.9 mg·g-1)和草地(161.0- 371.0 mg·g-1)土壤中性糖的相对含量无显著差异(p > 0.05; 图1B), 除灌丛外, 其他类型的中性糖相对含量均显著高于耕地(65.0-370.7 mg·g-1)(p < 0.05; 图1B).这与上述中性糖的绝对含量结果一致, 说明耕种会通过减少植物残体的输入从而使土壤中性糖含量减少.也有研究发现虽然土地利用类型由森林转变为灌丛, 但主要的维管束植物种类未改变, 凋落物质量没有变化导致两种土地利用类型土壤中性糖含量无显著差异(Conti et al., 2016).而Zhao等(2014)在青藏高原的研究表明, 草地土壤中性糖的相对含量显著高于灌丛, 可能是草本植物凋落物的半纤维素含量比灌丛植物凋落物中的含量高导致的. ...
... 按密度分组, 中性糖在轻质组分中的相对含量(129.0-410.0 mg·g-1)较重质组分中(7.5-322.0 mg·g-1)高.Llorente等(2017)在含碳酸钙的土壤中也发现中性糖含量在游离态组分中最高, 闭蓄态组分次之, 重质组分中最低; 由于微生物更易进入, 因此游离态组分中含有较多的甘露糖.另外, Bischoff等(2018)在西伯利亚高原不同盐分梯度的土壤中也发现, 无论是在表层还是底层, 中性糖含量在轻质组分中最高.但是, 与全土相比, 与矿质相结合的重质组分中中性糖含量更丰富, 该组分中以微生物来源的六碳糖最为丰富; 此外, 该研究中放射性碳的数据显示年龄较老的土壤样品中微生物来源的中性糖含量较高, 说明中性糖在矿质土中的稳定储存是通过连续的微生物转化或矿物的物理保护实现的(Rumpel et al., 2010).Dao等(2018)在西伯利亚东北部冻土中的研究也发现微生物来源的中性糖在重质组分土壤中含量较丰富, 从较低的C:N和较高的碳稳定同位素比率(δ13C)可以看出这与微生物来源的中性糖的选择性积累有关(Gentsch et al., 2015a, 2015b).此外, 微生物残体(Miltner et al., 2012)及微生物分泌的胞外多聚物中富含的多糖(Kleber et al., 2015), 也可能通过矿物的吸附作用在重质组分土壤中积累. ...
Stabilization mechanisms of soil organic matter: Implications for C-saturation of soils 1 2002
... 有研究提出有机质的化学特性决定了其在土壤中的降解速率, 即难降解的化合物会被选择性地保存在土壤中(Stevenson, 1994).传统观点认为, 中性糖等碳水化合物属于易降解化合物, 在土壤中的周转应该较快.但越来越多的研究表明土壤中碳水化合物的周转行为与其易降解的化学特性并不相符, 这可以从土壤中碳水化合物的平均年龄较高, 与原土中总有机碳年龄相似得到印证(Gleixner et al., 2002; Derrien et al., 2006; Derrien et al., 2007).其背后的机制也备受关注, 一方面是物理保护作用或化学难降解性(Sollins et al., 1996; Six et al., 2002; Von Lu?tzow et al., 2006), 另一方面是微生物对有机质的再利用(Gleixner et al., 2002).为明确土壤碳水化合物的周转特征及其驱动机制, 现有的研究常利用以下2种方法: 1) C3-C4植物轮作: 如Derrien等(2006)在长期种植小麦的土壤中种植玉米23年, 通过13C自然丰度的变化来评估土壤中性糖的周转发现, 在>200 μm粒级的土壤中, 碳水化合物的年龄(5年)比总土壤有机碳(7年)年轻, 在50-200 μm和0-50 μm粒级的土壤中, 碳水化合物的平均年龄分别为19和101年, 与相应粒级总的土壤有机碳年龄(32和91年)相似.即, 中性糖在>200 μm粒级中的周转较快(与50-200 μm和0-50 μm粒级相比).Basler等(2015a)利用该方法发现中性糖在土壤重组分中周转比在轻组分中慢: 矿物组分(>2 g·cm-3)以及原土中的中性糖碳的平均滞留时间和总碳相似, 但是闭蓄态组分(≤2 g·cm-3)中性糖碳的平均滞留时间比总碳短; 此外, 微生物来源的中性糖的周转也比植物来源周转慢(在未分组分的原土中), Basler等(2015a)认为有机质的再循环是控制土壤有机质动态的重要因子.2)添加13C标记的葡萄糖的培养实验: Basler等(2015b)通过添加13C标记的葡萄糖进行了为期3年的培养实验, 发现周转较快的中性糖碳库平均滞留时间为25-60天, 与Derrien等(2007)为期1年的短期培养实验结果相符(17天); 而较稳定的中性糖碳库(微生物来源的中性糖)的平均滞留时间长达365天, 进一步说明微生物对土壤中性糖的再循环利用过程是影响中性糖周转的重要环节. ...
Stabilization and destabilization of soil organic matter: Mechanisms and controls 1 1996
... 有研究提出有机质的化学特性决定了其在土壤中的降解速率, 即难降解的化合物会被选择性地保存在土壤中(Stevenson, 1994).传统观点认为, 中性糖等碳水化合物属于易降解化合物, 在土壤中的周转应该较快.但越来越多的研究表明土壤中碳水化合物的周转行为与其易降解的化学特性并不相符, 这可以从土壤中碳水化合物的平均年龄较高, 与原土中总有机碳年龄相似得到印证(Gleixner et al., 2002; Derrien et al., 2006; Derrien et al., 2007).其背后的机制也备受关注, 一方面是物理保护作用或化学难降解性(Sollins et al., 1996; Six et al., 2002; Von Lu?tzow et al., 2006), 另一方面是微生物对有机质的再利用(Gleixner et al., 2002).为明确土壤碳水化合物的周转特征及其驱动机制, 现有的研究常利用以下2种方法: 1) C3-C4植物轮作: 如Derrien等(2006)在长期种植小麦的土壤中种植玉米23年, 通过13C自然丰度的变化来评估土壤中性糖的周转发现, 在>200 μm粒级的土壤中, 碳水化合物的年龄(5年)比总土壤有机碳(7年)年轻, 在50-200 μm和0-50 μm粒级的土壤中, 碳水化合物的平均年龄分别为19和101年, 与相应粒级总的土壤有机碳年龄(32和91年)相似.即, 中性糖在>200 μm粒级中的周转较快(与50-200 μm和0-50 μm粒级相比).Basler等(2015a)利用该方法发现中性糖在土壤重组分中周转比在轻组分中慢: 矿物组分(>2 g·cm-3)以及原土中的中性糖碳的平均滞留时间和总碳相似, 但是闭蓄态组分(≤2 g·cm-3)中性糖碳的平均滞留时间比总碳短; 此外, 微生物来源的中性糖的周转也比植物来源周转慢(在未分组分的原土中), Basler等(2015a)认为有机质的再循环是控制土壤有机质动态的重要因子.2)添加13C标记的葡萄糖的培养实验: Basler等(2015b)通过添加13C标记的葡萄糖进行了为期3年的培养实验, 发现周转较快的中性糖碳库平均滞留时间为25-60天, 与Derrien等(2007)为期1年的短期培养实验结果相符(17天); 而较稳定的中性糖碳库(微生物来源的中性糖)的平均滞留时间长达365天, 进一步说明微生物对土壤中性糖的再循环利用过程是影响中性糖周转的重要环节. ...
Methods for the analysis of carbohydrate material in soil: 2 1 1962
... 土壤中性糖的提取主要包括水解、纯化和检测三个步骤.水解可以破坏糖类与土壤基质之间的氢键、共价键(Martens & Frankenberger, 1993), 以及多糖分子结构内部的键合作用(如糖苷键)(K?gel- Knabner, 2002), 将中性单糖从半纤维素中释放出来.土壤中性单糖的水解一般用硫酸(Sowden & Ivarson, 1962; Oades et al., 1970; Cheshire, 1979)、盐酸(Uzaki & Ishiwatari, 1983)或三氟乙酸(Amelung et al., 1996)实现(表1).由于硫酸不易被除去, 而三氟乙酸挥发性强, 可通过旋转蒸发去除, 且提取效率也较硫酸高, 因此三氟乙酸水解得到了广泛的应用(Amelung et al., 1996; Rumpel & Dignac, 2006; Zhang et al., 2007; Eder et al., 2010).目前最常用的水解方法是利用4 mol·L-1的三氟乙酸在105 ℃水解4 h, 既不会破坏单糖的结构, 也能获得最高的提取效率(Amelung et al., 1996).由于盐酸的水解效率较三氟乙酸和硫酸低, 该方法基本不再使用(Amelung et al., 1996).上述水解产物的纯化通常使用旋转蒸发法去除三氟乙酸, 使用螯合剂防止铁的氢氧化物(或氧化物)干扰, 或调节pH值将铁的氢氧化物(或氧化物)以沉淀形式去除(Zhang et al., 2007; Eder et al., 2010), 也有研究使用吸附剂和阳离子交换树脂去除杂质(Amelung et al., 1996; Martens et al., 2004). ...
Changes of lignin phenols and neutral sugars in different soil types of a high-elevation forest ecosystem 25 years after forest dieback 5 2007
... 糖醇乙酸酯衍生化是目前使用最广泛的一种中性糖衍生化方法, 其原理是将醛糖的羰基还原, 最终形成糖醇乙酸酯(Black & Fox, 1996; Ruiz-Matute et al., 2011).该衍生化的优点是: 得到的每种糖醇产物只产生一个单独的色谱峰, 易于检测, 且衍生化产物较稳定, 可稳定储存20天(Wang et al., 2017), 因此被广泛用于中性糖的定量分析(Rumpel & Dignac, 2006; Spielvogel et al., 2007; Rumpel et al., 2010).然而, 该衍生化方法耗时, 步骤繁琐(Black & Fox, 1996; Rumpel & Dignac, 2006; Ruiz-Matute et al., 2011), 一直有研究在对该方法进行不断改进. ...
... 本研究整合了5种土地利用类型的土壤中性糖数据, 发现阔叶林(7.0-82.8 mg·g-1)、针叶林(1.1-90.1 mg·g-1)、灌丛(2.5-80.6 mg·g-1)和草地(1.1-15.1 mg·g-1)土壤中性糖的绝对含量无显著差异(p > 0.05; 图1A), 除草地外, 其他类型均显著高于耕地土壤(0.6-6.3 mg·g-1)(p < 0.05; 图1A).不同地区(西班牙西北部、美国中西部、法国西南部)、不同土壤类型(碳酸钙土壤、沙质淋溶土)中的研究均表明阔叶林和针叶林土壤中性糖(或碳水化合物, 包括纤维素和半纤维中性糖, 下同)的绝对含量显著高于耕地土壤, 并认为可能是人为收获引起的地上输入量减少, 以及耕作扰动导致土壤有机层中的有机质向矿质层迁移造成的(Martens et al., 2004; Jolivet et al., 2006; Llorente et al., 2017).但是有研究报道中性糖(或碳水化合物)在草地土壤中更丰富(与耕地相比)(Larré-Larrouy et al., 2004; Martens et al., 2004; Guan et al., 2018).如, Guan等(2018)发现青藏高原的自然草地经过长期(>50年)耕种后, 植物地下生物量降低使植物碳向土壤中的输入减少, 加上耕作的扰动作用强烈影响了微生物的群落结构和丰度, 导致与草地相比, 耕地土壤中性糖含量降低了57%.Larré-Larrouy等(2004)也认为农作物的根系不发达, 来源于根系的碳水化合物较少是导致此现象的主要原因.由于本研究的数据基于土壤类型、气候条件等各异的研究, 中性糖含量在各研究之间存在差异, 加之数据量的限制导致了与上述研究的结果不一致.
Content and distribution of neutral sugars across different land-use regimes in the top soils (<xref ref-type="bibr" rid="b58">Nierop<i> et al</i>., 2001</xref>; <xref ref-type="bibr" rid="b71">Spielvogel<i> et al</i>., 2007</xref>; <xref ref-type="bibr" rid="b25">Eder<i> et al</i>., 2010</xref>; <xref ref-type="bibr" rid="b67">Rumpel<i> et al</i>., 2010</xref>; <xref ref-type="bibr" rid="b90">Zhao<i> et al</i>., 2014</xref>; <xref ref-type="bibr" rid="b19">Conti<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b21">Cui<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b82">Wang<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b20">Creme<i> et al</i>., 2017</xref>; <xref ref-type="bibr" rid="b49">Llorente<i> et al</i>., 2017</xref>; <xref ref-type="bibr" rid="b26">Evgrafova <i>et al</i>., 2018</xref>; <xref ref-type="bibr" rid="b91">Zhu <i>et al</i>., 2018</xref>). A, Neutral sugar absolute content. B, Neutral sugar relative content. C, GM/AX ((galactose + mannose)/(arabinose + xylose)). D, RF/AX ((rhamnose + fucose)/(arabinose + xylose)). The upper and lower end of boxes denote the 0.25 and 0.75 percentiles, respectively. The solid bar in the box mark the median of each dataset. The circles indicate outliers of each dataset. Different lowercase letters indicate differences in various land-use regimes (<i>p</i> < 0.05). <i>n</i> = 8, 25, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the A, C). <i>n</i> = 8, 22, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the B). <i>n</i> = 3, 19, 4, 8, 27 (from deciduous, coniferous, shrub, grassland to crops in the D). OC, soil organic carbon.Fig. 1 为考察不同土地利用类型中的中性糖对土壤有机碳(OC)的潜在贡献, 我们利用土壤有机碳含量对中性糖含量进行了归一化.结果显示: 阔叶林(84.5-253.9 mg·g-1)、针叶林(55.2-291.0 mg·g-1)、灌丛(139.0-208.9 mg·g-1)和草地(161.0- 371.0 mg·g-1)土壤中性糖的相对含量无显著差异(p > 0.05; 图1B), 除灌丛外, 其他类型的中性糖相对含量均显著高于耕地(65.0-370.7 mg·g-1)(p < 0.05; 图1B).这与上述中性糖的绝对含量结果一致, 说明耕种会通过减少植物残体的输入从而使土壤中性糖含量减少.也有研究发现虽然土地利用类型由森林转变为灌丛, 但主要的维管束植物种类未改变, 凋落物质量没有变化导致两种土地利用类型土壤中性糖含量无显著差异(Conti et al., 2016).而Zhao等(2014)在青藏高原的研究表明, 草地土壤中性糖的相对含量显著高于灌丛, 可能是草本植物凋落物的半纤维素含量比灌丛植物凋落物中的含量高导致的. ...
... ; Spielvogel et al., 2007; Eder et al., 2010; Rumpel et al., 2010; Zhao et al., 2014; Conti et al., 2016; Cui et al., 2016; Wang et al., 2016; Creme et al., 2017; Llorente et al., 2017; Evgrafova et al., 2018; Zhu et al., 2018). A, Neutral sugar absolute content. B, Neutral sugar relative content. C, GM/AX ((galactose + mannose)/(arabinose + xylose)). D, RF/AX ((rhamnose + fucose)/(arabinose + xylose)). The upper and lower end of boxes denote the 0.25 and 0.75 percentiles, respectively. The solid bar in the box mark the median of each dataset. The circles indicate outliers of each dataset. Different lowercase letters indicate differences in various land-use regimes (p < 0.05). n = 8, 25, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the A, C). n = 8, 22, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the B). n = 3, 19, 4, 8, 27 (from deciduous, coniferous, shrub, grassland to crops in the D). OC, soil organic carbon. Fig. 1 为考察不同土地利用类型中的中性糖对土壤有机碳(OC)的潜在贡献, 我们利用土壤有机碳含量对中性糖含量进行了归一化.结果显示: 阔叶林(84.5-253.9 mg·g-1)、针叶林(55.2-291.0 mg·g-1)、灌丛(139.0-208.9 mg·g-1)和草地(161.0- 371.0 mg·g-1)土壤中性糖的相对含量无显著差异(p > 0.05; 图1B), 除灌丛外, 其他类型的中性糖相对含量均显著高于耕地(65.0-370.7 mg·g-1)(p < 0.05; 图1B).这与上述中性糖的绝对含量结果一致, 说明耕种会通过减少植物残体的输入从而使土壤中性糖含量减少.也有研究发现虽然土地利用类型由森林转变为灌丛, 但主要的维管束植物种类未改变, 凋落物质量没有变化导致两种土地利用类型土壤中性糖含量无显著差异(Conti et al., 2016).而Zhao等(2014)在青藏高原的研究表明, 草地土壤中性糖的相对含量显著高于灌丛, 可能是草本植物凋落物的半纤维素含量比灌丛植物凋落物中的含量高导致的. ...
... 有研究提出有机质的化学特性决定了其在土壤中的降解速率, 即难降解的化合物会被选择性地保存在土壤中(Stevenson, 1994).传统观点认为, 中性糖等碳水化合物属于易降解化合物, 在土壤中的周转应该较快.但越来越多的研究表明土壤中碳水化合物的周转行为与其易降解的化学特性并不相符, 这可以从土壤中碳水化合物的平均年龄较高, 与原土中总有机碳年龄相似得到印证(Gleixner et al., 2002; Derrien et al., 2006; Derrien et al., 2007).其背后的机制也备受关注, 一方面是物理保护作用或化学难降解性(Sollins et al., 1996; Six et al., 2002; Von Lu?tzow et al., 2006), 另一方面是微生物对有机质的再利用(Gleixner et al., 2002).为明确土壤碳水化合物的周转特征及其驱动机制, 现有的研究常利用以下2种方法: 1) C3-C4植物轮作: 如Derrien等(2006)在长期种植小麦的土壤中种植玉米23年, 通过13C自然丰度的变化来评估土壤中性糖的周转发现, 在>200 μm粒级的土壤中, 碳水化合物的年龄(5年)比总土壤有机碳(7年)年轻, 在50-200 μm和0-50 μm粒级的土壤中, 碳水化合物的平均年龄分别为19和101年, 与相应粒级总的土壤有机碳年龄(32和91年)相似.即, 中性糖在>200 μm粒级中的周转较快(与50-200 μm和0-50 μm粒级相比).Basler等(2015a)利用该方法发现中性糖在土壤重组分中周转比在轻组分中慢: 矿物组分(>2 g·cm-3)以及原土中的中性糖碳的平均滞留时间和总碳相似, 但是闭蓄态组分(≤2 g·cm-3)中性糖碳的平均滞留时间比总碳短; 此外, 微生物来源的中性糖的周转也比植物来源周转慢(在未分组分的原土中), Basler等(2015a)认为有机质的再循环是控制土壤有机质动态的重要因子.2)添加13C标记的葡萄糖的培养实验: Basler等(2015b)通过添加13C标记的葡萄糖进行了为期3年的培养实验, 发现周转较快的中性糖碳库平均滞留时间为25-60天, 与Derrien等(2007)为期1年的短期培养实验结果相符(17天); 而较稳定的中性糖碳库(微生物来源的中性糖)的平均滞留时间长达365天, 进一步说明微生物对土壤中性糖的再循环利用过程是影响中性糖周转的重要环节. ...
Sensitive method for carbohydrate-composition analysis of glycoproteins by high-performance liquid- chromatography 1 1987
... 中性糖的检测分析可使用气相色谱-质谱联用仪(GC-MS)(Rumpel & Dignac, 2006; Zhang et al., 2007; Eder et al., 2010), 液相色谱(HPLC)(Takeuchi et al., 1987; Tanaka et al., 1990; Amelung et al., 1996; Basler & Dyckmans, 2013)和高效阴离子交换色谱-脉冲安培检测法(HPAEC-PAD)(Bruggink et al., 2005; Zhang et al., 2012).由于中性糖不易挥发, 因此在使用GC-MS检测前需衍生化, 衍生化是影响土壤中性糖定量的关键环节.下面将介绍几种常用的中性糖衍生化方法. ...
Determination of component sugars in soil organic-matter by HPLC 3 1990
... Comparison of extraction and detection methods of neutral sugars in soils Table 1
步骤 Procedure
方法 Method
优点 Advantage
缺点 Drawback
参考文献 Reference
提取 Extraction
硫酸 H2SO4
硫酸不易被除去 H2SO4 can not be removed easily
Tanaka et al., 1990
盐酸 HCl
会水解一部分纤维素; 产率较低 The hydrolysis products include a few cellulosic neutral sugars; low yields
Uzaki & Ishiwatari, 1983
三氟乙酸 TFA
产率高; 不会破坏单糖结构; 具有挥发性, 可通过旋转蒸发去除; 水解的多糖主要为半纤维素 High yields; Not destructive to monosaccharides; TFA is volatile and can be easily removed by evaporation; Hydrolysis products are mainly released from hemicellulose
Amelung et al., 1996
检测 Detection
GC-MS
精度、准确度、敏感性和效率较高 High accuracy, precision, sensibility and efficiency
需要衍生化 Derivatization is required
Amelung et al., 1996; Wang et al., 2017
HPLC
无需衍生化; 纯化过程简单 No need for derivatization; Simple purification procedures
精度、准确度、敏感性较低 Low accuracy, precision, sensibility and efficiency
Hamada & Ono, 1984; Angers et al., 1988;Tanaka et al., 1990;
HPAEC-PAD
无需衍生化; 应用范围广, 可同时分析糖醛酸和中性糖 No need for derivatization; Wide application and simultaneous analysis of uronic acid and neutral sugars
精度、准确度、敏感性较低 Low accuracy, precision, sensibility and efficiency
Bruggink et al., 2005; Zhang et al., 2012
TFA, trifluoroacetic acid; GC-MS, gas chromatography-mass spectrometry; HPLC, high-performance liquid chromatography; HPAEC-PAD, high-performance anion-exchange chromatography with pulsed amperometric detection. ...
... ;Tanaka et al., 1990;
HPAEC-PAD
无需衍生化; 应用范围广, 可同时分析糖醛酸和中性糖 No need for derivatization; Wide application and simultaneous analysis of uronic acid and neutral sugars
精度、准确度、敏感性较低 Low accuracy, precision, sensibility and efficiency
Bruggink et al., 2005; Zhang et al., 2012
TFA, trifluoroacetic acid; GC-MS, gas chromatography-mass spectrometry; HPLC, high-performance liquid chromatography; HPAEC-PAD, high-performance anion-exchange chromatography with pulsed amperometric detection. ...
... 中性糖的检测分析可使用气相色谱-质谱联用仪(GC-MS)(Rumpel & Dignac, 2006; Zhang et al., 2007; Eder et al., 2010), 液相色谱(HPLC)(Takeuchi et al., 1987; Tanaka et al., 1990; Amelung et al., 1996; Basler & Dyckmans, 2013)和高效阴离子交换色谱-脉冲安培检测法(HPAEC-PAD)(Bruggink et al., 2005; Zhang et al., 2012).由于中性糖不易挥发, 因此在使用GC-MS检测前需衍生化, 衍生化是影响土壤中性糖定量的关键环节.下面将介绍几种常用的中性糖衍生化方法. ...
Soil C and N pools in patchy shrublands of the Negev and Chihuahuan Deserts 1 2006
... 关于影响中性糖的人为因素的研究主要集中在放牧、耕作和土地利用方式改变等方面.放牧的影响存在较多争议, 如有研究发现放牧通过影响植物生物量输入使草地生态系统碳水化合物含量减少(Thompson et al., 2006), 但也有研究发现放牧对中性糖含量没有影响(Steffens et al., 2009).Abdelrahman等(2016)指出施肥和种植的农作物种类对碳水化合物含量均无显著影响, 但也有研究表明施用混合肥的土壤中性糖含量显著高于施用蚯蚓堆肥的土壤, 可能是施用混合肥土壤中微生物分解作用更小的原因(Phuong-Thi et al., 2012).对中国东北地区的研究发现, 长期单施有机肥以及有机肥和化肥配合施用可以提高作物产量, 有利于颗粒有机质中植物来源的中性糖的积累, 而单施化肥对土壤中性糖含量无显著影响(张彬等, 2010). ...
长白山不同海拔梯度森林土壤中性糖分布特征 1 2013
... 影响中性糖含量的因素可以分为自然因素和人为因素.自然因素有环境因素(CO2、O2、海拔、温度、降水等), 生物扰动(土壤动物和微生物), 以及自然灾害(如火灾和病虫害).Barron-Gafford等(2005)在半封闭式的生态系统(生物圈Ⅱ号)中的研究表明, 在高CO2浓度处理下, 土壤碳水化合物含量增加, 主要是由植物的地下生物量增加引起的.另外, 有研究使用开放式CO2富集系统(FACE)发现高CO2处理7年后, 由于植物生物量增加, 与对照组相比, 温带草地土壤中性糖含量增加了28% (Bock et al., 2007).与CO2不同, 当泥炭土在O2中的暴露时间延长, 土壤中性糖降解速率加快, 含量降低(Philben et al., 2015).田秋香等(2013)通过对中国长白山不同海拔梯度森林土壤中性糖分布的研究发现, 不同海拔间的温度差异是造成土壤中性糖含量差异的主要原因, 在高海拔的低温环境中微生物的分解作用减弱, 再加上微生物对外源碳的利用效率提高, 被微生物转化后的外源碳以微生物残体的形式固存在土壤中.综上所述, 高海拔有利于中性糖的积累.有关生物扰动的研究发现蚯蚓并不会影响土壤中性糖含量(Phuong-Thi et al., 2012), 而土壤微生物对于土壤中性糖的降解影响很大, 当微生物多样性较高时, 单糖如阿拉伯糖和木糖的降解速率增加, 含量减少(Baumann et al., 2013).此外, 自然灾害如火灾对土壤中性糖的影响并没有一致的规律, 不同程度的火灾对土壤不同组分的中性糖含量和组成的影响都不同(Mastrolonardo et al., 2015). ...
长白山不同海拔梯度森林土壤中性糖分布特征 1 2013
... 影响中性糖含量的因素可以分为自然因素和人为因素.自然因素有环境因素(CO2、O2、海拔、温度、降水等), 生物扰动(土壤动物和微生物), 以及自然灾害(如火灾和病虫害).Barron-Gafford等(2005)在半封闭式的生态系统(生物圈Ⅱ号)中的研究表明, 在高CO2浓度处理下, 土壤碳水化合物含量增加, 主要是由植物的地下生物量增加引起的.另外, 有研究使用开放式CO2富集系统(FACE)发现高CO2处理7年后, 由于植物生物量增加, 与对照组相比, 温带草地土壤中性糖含量增加了28% (Bock et al., 2007).与CO2不同, 当泥炭土在O2中的暴露时间延长, 土壤中性糖降解速率加快, 含量降低(Philben et al., 2015).田秋香等(2013)通过对中国长白山不同海拔梯度森林土壤中性糖分布的研究发现, 不同海拔间的温度差异是造成土壤中性糖含量差异的主要原因, 在高海拔的低温环境中微生物的分解作用减弱, 再加上微生物对外源碳的利用效率提高, 被微生物转化后的外源碳以微生物残体的形式固存在土壤中.综上所述, 高海拔有利于中性糖的积累.有关生物扰动的研究发现蚯蚓并不会影响土壤中性糖含量(Phuong-Thi et al., 2012), 而土壤微生物对于土壤中性糖的降解影响很大, 当微生物多样性较高时, 单糖如阿拉伯糖和木糖的降解速率增加, 含量减少(Baumann et al., 2013).此外, 自然灾害如火灾对土壤中性糖的影响并没有一致的规律, 不同程度的火灾对土壤不同组分的中性糖含量和组成的影响都不同(Mastrolonardo et al., 2015). ...
Changes in the amount and distribution of neutral monosaccharides of savanna soils after plantation of Pinus and Eucalyptus in the Congo 1 1996
... 碳水化合物, 即糖类, 可分为单糖、低聚糖和多糖(Cheshire et al., 1992).碳水化合物占维管束植物干质量的75%, 细菌干质量的40% (Moers et al., 1993), 土壤中来源于植物和微生物的碳水化合物占土壤有机质的5%-25%, 是土壤有机质的重要组分(Angers et al., 1988; Murata et al., 1999).碳水化合物是土壤微生物的主要能量来源和碳源(Amelung et al., 1996), 几乎所有的低聚糖和单糖都可溶于水, 属于土壤中易降解的有机质, 易被微生物利用, 并用于自身生长代谢(Gunina & Kuzyakov, 2015).碳水化合物还可驱动微生物由休眠状态转变为活跃状态(Blagodatskaya & Kuzyakov, 2013), 是激发效应产生的有效基质(Kuzyakov, 2010).此外, 碳水化合物是团聚体形成的重要胶结物质(Feller & Beare, 1997), 例如, 阿拉伯糖和木糖是植物黏胶糖的重要来源, 对团聚体的形成起着促进作用(Oades, 1984).Larré-Larrouy等(2004)研究发现, 耕作后土壤团聚体稳定性的改变与团聚体中木糖的减少有关.Angers和Mehuys (1989)提出碳水化合物含量和团聚体的平均质量直径正相关, 表明碳水化合物含量的变化对水稳定性团聚体的改变有着一定的影响.此外, Puget等(1999)发现在更稳定的黏-粉团聚体中微生物来源的碳水化合物较丰富, 并推测团聚体的稳定性受胞外的微生物多糖调控.由此可见, 碳水化合物在团聚体稳定性方面扮演着重要角色.综上所述, 研究土壤碳水化合物对于评估土壤有机质的组成、来源、质量与动态变化具有重要意义(Guggenberger et al., 1994; Trouve et al., 1996; Nacro et al., 2005; Navarrete & Tsutsuki, 2008). ...
Determination of cellulose and non-cellulose carbohydrates in recent sediments by gas chromatography 2 1983
... 土壤中性糖的提取主要包括水解、纯化和检测三个步骤.水解可以破坏糖类与土壤基质之间的氢键、共价键(Martens & Frankenberger, 1993), 以及多糖分子结构内部的键合作用(如糖苷键)(K?gel- Knabner, 2002), 将中性单糖从半纤维素中释放出来.土壤中性单糖的水解一般用硫酸(Sowden & Ivarson, 1962; Oades et al., 1970; Cheshire, 1979)、盐酸(Uzaki & Ishiwatari, 1983)或三氟乙酸(Amelung et al., 1996)实现(表1).由于硫酸不易被除去, 而三氟乙酸挥发性强, 可通过旋转蒸发去除, 且提取效率也较硫酸高, 因此三氟乙酸水解得到了广泛的应用(Amelung et al., 1996; Rumpel & Dignac, 2006; Zhang et al., 2007; Eder et al., 2010).目前最常用的水解方法是利用4 mol·L-1的三氟乙酸在105 ℃水解4 h, 既不会破坏单糖的结构, 也能获得最高的提取效率(Amelung et al., 1996).由于盐酸的水解效率较三氟乙酸和硫酸低, 该方法基本不再使用(Amelung et al., 1996).上述水解产物的纯化通常使用旋转蒸发法去除三氟乙酸, 使用螯合剂防止铁的氢氧化物(或氧化物)干扰, 或调节pH值将铁的氢氧化物(或氧化物)以沉淀形式去除(Zhang et al., 2007; Eder et al., 2010), 也有研究使用吸附剂和阳离子交换树脂去除杂质(Amelung et al., 1996; Martens et al., 2004). ...
... Comparison of extraction and detection methods of neutral sugars in soils Table 1
步骤 Procedure
方法 Method
优点 Advantage
缺点 Drawback
参考文献 Reference
提取 Extraction
硫酸 H2SO4
硫酸不易被除去 H2SO4 can not be removed easily
Tanaka et al., 1990
盐酸 HCl
会水解一部分纤维素; 产率较低 The hydrolysis products include a few cellulosic neutral sugars; low yields
Uzaki & Ishiwatari, 1983
三氟乙酸 TFA
产率高; 不会破坏单糖结构; 具有挥发性, 可通过旋转蒸发去除; 水解的多糖主要为半纤维素 High yields; Not destructive to monosaccharides; TFA is volatile and can be easily removed by evaporation; Hydrolysis products are mainly released from hemicellulose
Amelung et al., 1996
检测 Detection
GC-MS
精度、准确度、敏感性和效率较高 High accuracy, precision, sensibility and efficiency
需要衍生化 Derivatization is required
Amelung et al., 1996; Wang et al., 2017
HPLC
无需衍生化; 纯化过程简单 No need for derivatization; Simple purification procedures
精度、准确度、敏感性较低 Low accuracy, precision, sensibility and efficiency
Hamada & Ono, 1984; Angers et al., 1988;Tanaka et al., 1990;
HPAEC-PAD
无需衍生化; 应用范围广, 可同时分析糖醛酸和中性糖 No need for derivatization; Wide application and simultaneous analysis of uronic acid and neutral sugars
精度、准确度、敏感性较低 Low accuracy, precision, sensibility and efficiency
Bruggink et al., 2005; Zhang et al., 2012
TFA, trifluoroacetic acid; GC-MS, gas chromatography-mass spectrometry; HPLC, high-performance liquid chromatography; HPAEC-PAD, high-performance anion-exchange chromatography with pulsed amperometric detection. ...
Stabilization of organic matter in temperate soils: Mechanisms and their relevance under different soil conditions―A review 1 2006
... 有研究提出有机质的化学特性决定了其在土壤中的降解速率, 即难降解的化合物会被选择性地保存在土壤中(Stevenson, 1994).传统观点认为, 中性糖等碳水化合物属于易降解化合物, 在土壤中的周转应该较快.但越来越多的研究表明土壤中碳水化合物的周转行为与其易降解的化学特性并不相符, 这可以从土壤中碳水化合物的平均年龄较高, 与原土中总有机碳年龄相似得到印证(Gleixner et al., 2002; Derrien et al., 2006; Derrien et al., 2007).其背后的机制也备受关注, 一方面是物理保护作用或化学难降解性(Sollins et al., 1996; Six et al., 2002; Von Lu?tzow et al., 2006), 另一方面是微生物对有机质的再利用(Gleixner et al., 2002).为明确土壤碳水化合物的周转特征及其驱动机制, 现有的研究常利用以下2种方法: 1) C3-C4植物轮作: 如Derrien等(2006)在长期种植小麦的土壤中种植玉米23年, 通过13C自然丰度的变化来评估土壤中性糖的周转发现, 在>200 μm粒级的土壤中, 碳水化合物的年龄(5年)比总土壤有机碳(7年)年轻, 在50-200 μm和0-50 μm粒级的土壤中, 碳水化合物的平均年龄分别为19和101年, 与相应粒级总的土壤有机碳年龄(32和91年)相似.即, 中性糖在>200 μm粒级中的周转较快(与50-200 μm和0-50 μm粒级相比).Basler等(2015a)利用该方法发现中性糖在土壤重组分中周转比在轻组分中慢: 矿物组分(>2 g·cm-3)以及原土中的中性糖碳的平均滞留时间和总碳相似, 但是闭蓄态组分(≤2 g·cm-3)中性糖碳的平均滞留时间比总碳短; 此外, 微生物来源的中性糖的周转也比植物来源周转慢(在未分组分的原土中), Basler等(2015a)认为有机质的再循环是控制土壤有机质动态的重要因子.2)添加13C标记的葡萄糖的培养实验: Basler等(2015b)通过添加13C标记的葡萄糖进行了为期3年的培养实验, 发现周转较快的中性糖碳库平均滞留时间为25-60天, 与Derrien等(2007)为期1年的短期培养实验结果相符(17天); 而较稳定的中性糖碳库(微生物来源的中性糖)的平均滞留时间长达365天, 进一步说明微生物对土壤中性糖的再循环利用过程是影响中性糖周转的重要环节. ...
Improvement of simultaneous determination of neutral monosaccharides and uronic acids by gas chromatography 3 2017
... Comparison of extraction and detection methods of neutral sugars in soils Table 1
步骤 Procedure
方法 Method
优点 Advantage
缺点 Drawback
参考文献 Reference
提取 Extraction
硫酸 H2SO4
硫酸不易被除去 H2SO4 can not be removed easily
Tanaka et al., 1990
盐酸 HCl
会水解一部分纤维素; 产率较低 The hydrolysis products include a few cellulosic neutral sugars; low yields
Uzaki & Ishiwatari, 1983
三氟乙酸 TFA
产率高; 不会破坏单糖结构; 具有挥发性, 可通过旋转蒸发去除; 水解的多糖主要为半纤维素 High yields; Not destructive to monosaccharides; TFA is volatile and can be easily removed by evaporation; Hydrolysis products are mainly released from hemicellulose
Amelung et al., 1996
检测 Detection
GC-MS
精度、准确度、敏感性和效率较高 High accuracy, precision, sensibility and efficiency
需要衍生化 Derivatization is required
Amelung et al., 1996; Wang et al., 2017
HPLC
无需衍生化; 纯化过程简单 No need for derivatization; Simple purification procedures
精度、准确度、敏感性较低 Low accuracy, precision, sensibility and efficiency
Hamada & Ono, 1984; Angers et al., 1988;Tanaka et al., 1990;
HPAEC-PAD
无需衍生化; 应用范围广, 可同时分析糖醛酸和中性糖 No need for derivatization; Wide application and simultaneous analysis of uronic acid and neutral sugars
精度、准确度、敏感性较低 Low accuracy, precision, sensibility and efficiency
Bruggink et al., 2005; Zhang et al., 2012
TFA, trifluoroacetic acid; GC-MS, gas chromatography-mass spectrometry; HPLC, high-performance liquid chromatography; HPAEC-PAD, high-performance anion-exchange chromatography with pulsed amperometric detection. ...
... 糖醇乙酸酯衍生化是目前使用最广泛的一种中性糖衍生化方法, 其原理是将醛糖的羰基还原, 最终形成糖醇乙酸酯(Black & Fox, 1996; Ruiz-Matute et al., 2011).该衍生化的优点是: 得到的每种糖醇产物只产生一个单独的色谱峰, 易于检测, 且衍生化产物较稳定, 可稳定储存20天(Wang et al., 2017), 因此被广泛用于中性糖的定量分析(Rumpel & Dignac, 2006; Spielvogel et al., 2007; Rumpel et al., 2010).然而, 该衍生化方法耗时, 步骤繁琐(Black & Fox, 1996; Rumpel & Dignac, 2006; Ruiz-Matute et al., 2011), 一直有研究在对该方法进行不断改进. ...
... 虽然不断有研究对各类衍生化方法进行改进, 但是衍生化仍旧是一个繁琐耗时的过程, 相比GC-MS检测方法, HPLC和HPAEC-PDA检测法较为简单, 在定量前无需衍生化.但这两种检测方法都需要特殊的仪器或检测器, 并且检测敏感度、准确度和精度较低, 因此目前没有广泛应用(Zhang et al., 2013; Wang et al., 2017). ...
Multi-site assessment of the effects of plastic- film mulch on the soil organic carbon balance in semiarid areas of China 2 2016
... 本研究整合了5种土地利用类型的土壤中性糖数据, 发现阔叶林(7.0-82.8 mg·g-1)、针叶林(1.1-90.1 mg·g-1)、灌丛(2.5-80.6 mg·g-1)和草地(1.1-15.1 mg·g-1)土壤中性糖的绝对含量无显著差异(p > 0.05; 图1A), 除草地外, 其他类型均显著高于耕地土壤(0.6-6.3 mg·g-1)(p < 0.05; 图1A).不同地区(西班牙西北部、美国中西部、法国西南部)、不同土壤类型(碳酸钙土壤、沙质淋溶土)中的研究均表明阔叶林和针叶林土壤中性糖(或碳水化合物, 包括纤维素和半纤维中性糖, 下同)的绝对含量显著高于耕地土壤, 并认为可能是人为收获引起的地上输入量减少, 以及耕作扰动导致土壤有机层中的有机质向矿质层迁移造成的(Martens et al., 2004; Jolivet et al., 2006; Llorente et al., 2017).但是有研究报道中性糖(或碳水化合物)在草地土壤中更丰富(与耕地相比)(Larré-Larrouy et al., 2004; Martens et al., 2004; Guan et al., 2018).如, Guan等(2018)发现青藏高原的自然草地经过长期(>50年)耕种后, 植物地下生物量降低使植物碳向土壤中的输入减少, 加上耕作的扰动作用强烈影响了微生物的群落结构和丰度, 导致与草地相比, 耕地土壤中性糖含量降低了57%.Larré-Larrouy等(2004)也认为农作物的根系不发达, 来源于根系的碳水化合物较少是导致此现象的主要原因.由于本研究的数据基于土壤类型、气候条件等各异的研究, 中性糖含量在各研究之间存在差异, 加之数据量的限制导致了与上述研究的结果不一致.
Content and distribution of neutral sugars across different land-use regimes in the top soils (<xref ref-type="bibr" rid="b58">Nierop<i> et al</i>., 2001</xref>; <xref ref-type="bibr" rid="b71">Spielvogel<i> et al</i>., 2007</xref>; <xref ref-type="bibr" rid="b25">Eder<i> et al</i>., 2010</xref>; <xref ref-type="bibr" rid="b67">Rumpel<i> et al</i>., 2010</xref>; <xref ref-type="bibr" rid="b90">Zhao<i> et al</i>., 2014</xref>; <xref ref-type="bibr" rid="b19">Conti<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b21">Cui<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b82">Wang<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b20">Creme<i> et al</i>., 2017</xref>; <xref ref-type="bibr" rid="b49">Llorente<i> et al</i>., 2017</xref>; <xref ref-type="bibr" rid="b26">Evgrafova <i>et al</i>., 2018</xref>; <xref ref-type="bibr" rid="b91">Zhu <i>et al</i>., 2018</xref>). A, Neutral sugar absolute content. B, Neutral sugar relative content. C, GM/AX ((galactose + mannose)/(arabinose + xylose)). D, RF/AX ((rhamnose + fucose)/(arabinose + xylose)). The upper and lower end of boxes denote the 0.25 and 0.75 percentiles, respectively. The solid bar in the box mark the median of each dataset. The circles indicate outliers of each dataset. Different lowercase letters indicate differences in various land-use regimes (<i>p</i> < 0.05). <i>n</i> = 8, 25, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the A, C). <i>n</i> = 8, 22, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the B). <i>n</i> = 3, 19, 4, 8, 27 (from deciduous, coniferous, shrub, grassland to crops in the D). OC, soil organic carbon.Fig. 1 为考察不同土地利用类型中的中性糖对土壤有机碳(OC)的潜在贡献, 我们利用土壤有机碳含量对中性糖含量进行了归一化.结果显示: 阔叶林(84.5-253.9 mg·g-1)、针叶林(55.2-291.0 mg·g-1)、灌丛(139.0-208.9 mg·g-1)和草地(161.0- 371.0 mg·g-1)土壤中性糖的相对含量无显著差异(p > 0.05; 图1B), 除灌丛外, 其他类型的中性糖相对含量均显著高于耕地(65.0-370.7 mg·g-1)(p < 0.05; 图1B).这与上述中性糖的绝对含量结果一致, 说明耕种会通过减少植物残体的输入从而使土壤中性糖含量减少.也有研究发现虽然土地利用类型由森林转变为灌丛, 但主要的维管束植物种类未改变, 凋落物质量没有变化导致两种土地利用类型土壤中性糖含量无显著差异(Conti et al., 2016).而Zhao等(2014)在青藏高原的研究表明, 草地土壤中性糖的相对含量显著高于灌丛, 可能是草本植物凋落物的半纤维素含量比灌丛植物凋落物中的含量高导致的. ...
... ; Wang et al., 2016; Creme et al., 2017; Llorente et al., 2017; Evgrafova et al., 2018; Zhu et al., 2018). A, Neutral sugar absolute content. B, Neutral sugar relative content. C, GM/AX ((galactose + mannose)/(arabinose + xylose)). D, RF/AX ((rhamnose + fucose)/(arabinose + xylose)). The upper and lower end of boxes denote the 0.25 and 0.75 percentiles, respectively. The solid bar in the box mark the median of each dataset. The circles indicate outliers of each dataset. Different lowercase letters indicate differences in various land-use regimes (p < 0.05). n = 8, 25, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the A, C). n = 8, 22, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the B). n = 3, 19, 4, 8, 27 (from deciduous, coniferous, shrub, grassland to crops in the D). OC, soil organic carbon. Fig. 1 为考察不同土地利用类型中的中性糖对土壤有机碳(OC)的潜在贡献, 我们利用土壤有机碳含量对中性糖含量进行了归一化.结果显示: 阔叶林(84.5-253.9 mg·g-1)、针叶林(55.2-291.0 mg·g-1)、灌丛(139.0-208.9 mg·g-1)和草地(161.0- 371.0 mg·g-1)土壤中性糖的相对含量无显著差异(p > 0.05; 图1B), 除灌丛外, 其他类型的中性糖相对含量均显著高于耕地(65.0-370.7 mg·g-1)(p < 0.05; 图1B).这与上述中性糖的绝对含量结果一致, 说明耕种会通过减少植物残体的输入从而使土壤中性糖含量减少.也有研究发现虽然土地利用类型由森林转变为灌丛, 但主要的维管束植物种类未改变, 凋落物质量没有变化导致两种土地利用类型土壤中性糖含量无显著差异(Conti et al., 2016).而Zhao等(2014)在青藏高原的研究表明, 草地土壤中性糖的相对含量显著高于灌丛, 可能是草本植物凋落物的半纤维素含量比灌丛植物凋落物中的含量高导致的. ...
Continuous pH monitoring during pretreatment of yellow poplar wood sawdust by pressure cooking in water 1 1998
Long-term manure amendments enhance neutral sugar accumulation in bulk soil and particulate organic matter in a Mollisol 1 2014
... 此外, 糖腈乙酰酯衍生化也是目前使用较多的方法之一(Xie et al., 2014; Cui et al., 2016), 该衍生化方法最开始用于土壤中氨基糖含量的测定(Amelung et al., 1996), 随后Zhang等(2007)将其应用于土壤中性糖的测定.糖腈乙酰酯衍生化方法的优点是重复性较好, 获得的衍生化产物也较稳定(Zhang et al., 2007). ...
长期施肥对颗粒有机质中中性糖特性的影响 1 2010
... 关于影响中性糖的人为因素的研究主要集中在放牧、耕作和土地利用方式改变等方面.放牧的影响存在较多争议, 如有研究发现放牧通过影响植物生物量输入使草地生态系统碳水化合物含量减少(Thompson et al., 2006), 但也有研究发现放牧对中性糖含量没有影响(Steffens et al., 2009).Abdelrahman等(2016)指出施肥和种植的农作物种类对碳水化合物含量均无显著影响, 但也有研究表明施用混合肥的土壤中性糖含量显著高于施用蚯蚓堆肥的土壤, 可能是施用混合肥土壤中微生物分解作用更小的原因(Phuong-Thi et al., 2012).对中国东北地区的研究发现, 长期单施有机肥以及有机肥和化肥配合施用可以提高作物产量, 有利于颗粒有机质中植物来源的中性糖的积累, 而单施化肥对土壤中性糖含量无显著影响(张彬等, 2010). ...
长期施肥对颗粒有机质中中性糖特性的影响 1 2010
... 关于影响中性糖的人为因素的研究主要集中在放牧、耕作和土地利用方式改变等方面.放牧的影响存在较多争议, 如有研究发现放牧通过影响植物生物量输入使草地生态系统碳水化合物含量减少(Thompson et al., 2006), 但也有研究发现放牧对中性糖含量没有影响(Steffens et al., 2009).Abdelrahman等(2016)指出施肥和种植的农作物种类对碳水化合物含量均无显著影响, 但也有研究表明施用混合肥的土壤中性糖含量显著高于施用蚯蚓堆肥的土壤, 可能是施用混合肥土壤中微生物分解作用更小的原因(Phuong-Thi et al., 2012).对中国东北地区的研究发现, 长期单施有机肥以及有机肥和化肥配合施用可以提高作物产量, 有利于颗粒有机质中植物来源的中性糖的积累, 而单施化肥对土壤中性糖含量无显著影响(张彬等, 2010). ...
Determination of the carbohydrates from Notopterygium forbesii Boiss by HPLC with fluorescence detection 1 2013
... 虽然不断有研究对各类衍生化方法进行改进, 但是衍生化仍旧是一个繁琐耗时的过程, 相比GC-MS检测方法, HPLC和HPAEC-PDA检测法较为简单, 在定量前无需衍生化.但这两种检测方法都需要特殊的仪器或检测器, 并且检测敏感度、准确度和精度较低, 因此目前没有广泛应用(Zhang et al., 2013; Wang et al., 2017). ...
Determination of neutral sugars in soil by capillary gas chromatography after derivatization to aldononitrile acetates 5 2007
... 土壤中性糖的提取主要包括水解、纯化和检测三个步骤.水解可以破坏糖类与土壤基质之间的氢键、共价键(Martens & Frankenberger, 1993), 以及多糖分子结构内部的键合作用(如糖苷键)(K?gel- Knabner, 2002), 将中性单糖从半纤维素中释放出来.土壤中性单糖的水解一般用硫酸(Sowden & Ivarson, 1962; Oades et al., 1970; Cheshire, 1979)、盐酸(Uzaki & Ishiwatari, 1983)或三氟乙酸(Amelung et al., 1996)实现(表1).由于硫酸不易被除去, 而三氟乙酸挥发性强, 可通过旋转蒸发去除, 且提取效率也较硫酸高, 因此三氟乙酸水解得到了广泛的应用(Amelung et al., 1996; Rumpel & Dignac, 2006; Zhang et al., 2007; Eder et al., 2010).目前最常用的水解方法是利用4 mol·L-1的三氟乙酸在105 ℃水解4 h, 既不会破坏单糖的结构, 也能获得最高的提取效率(Amelung et al., 1996).由于盐酸的水解效率较三氟乙酸和硫酸低, 该方法基本不再使用(Amelung et al., 1996).上述水解产物的纯化通常使用旋转蒸发法去除三氟乙酸, 使用螯合剂防止铁的氢氧化物(或氧化物)干扰, 或调节pH值将铁的氢氧化物(或氧化物)以沉淀形式去除(Zhang et al., 2007; Eder et al., 2010), 也有研究使用吸附剂和阳离子交换树脂去除杂质(Amelung et al., 1996; Martens et al., 2004). ...
... ).上述水解产物的纯化通常使用旋转蒸发法去除三氟乙酸, 使用螯合剂防止铁的氢氧化物(或氧化物)干扰, 或调节pH值将铁的氢氧化物(或氧化物)以沉淀形式去除(Zhang et al., 2007; Eder et al., 2010), 也有研究使用吸附剂和阳离子交换树脂去除杂质(Amelung et al., 1996; Martens et al., 2004). ...
... 中性糖的检测分析可使用气相色谱-质谱联用仪(GC-MS)(Rumpel & Dignac, 2006; Zhang et al., 2007; Eder et al., 2010), 液相色谱(HPLC)(Takeuchi et al., 1987; Tanaka et al., 1990; Amelung et al., 1996; Basler & Dyckmans, 2013)和高效阴离子交换色谱-脉冲安培检测法(HPAEC-PAD)(Bruggink et al., 2005; Zhang et al., 2012).由于中性糖不易挥发, 因此在使用GC-MS检测前需衍生化, 衍生化是影响土壤中性糖定量的关键环节.下面将介绍几种常用的中性糖衍生化方法. ...
... 此外, 糖腈乙酰酯衍生化也是目前使用较多的方法之一(Xie et al., 2014; Cui et al., 2016), 该衍生化方法最开始用于土壤中氨基糖含量的测定(Amelung et al., 1996), 随后Zhang等(2007)将其应用于土壤中性糖的测定.糖腈乙酰酯衍生化方法的优点是重复性较好, 获得的衍生化产物也较稳定(Zhang et al., 2007). ...
... 将其应用于土壤中性糖的测定.糖腈乙酰酯衍生化方法的优点是重复性较好, 获得的衍生化产物也较稳定(Zhang et al., 2007). ...
Complete monosaccharide analysis by high-performance anion-exchange chromatography with pulsed amperometric detection 2 2012
... Comparison of extraction and detection methods of neutral sugars in soils Table 1
步骤 Procedure
方法 Method
优点 Advantage
缺点 Drawback
参考文献 Reference
提取 Extraction
硫酸 H2SO4
硫酸不易被除去 H2SO4 can not be removed easily
Tanaka et al., 1990
盐酸 HCl
会水解一部分纤维素; 产率较低 The hydrolysis products include a few cellulosic neutral sugars; low yields
Uzaki & Ishiwatari, 1983
三氟乙酸 TFA
产率高; 不会破坏单糖结构; 具有挥发性, 可通过旋转蒸发去除; 水解的多糖主要为半纤维素 High yields; Not destructive to monosaccharides; TFA is volatile and can be easily removed by evaporation; Hydrolysis products are mainly released from hemicellulose
Amelung et al., 1996
检测 Detection
GC-MS
精度、准确度、敏感性和效率较高 High accuracy, precision, sensibility and efficiency
需要衍生化 Derivatization is required
Amelung et al., 1996; Wang et al., 2017
HPLC
无需衍生化; 纯化过程简单 No need for derivatization; Simple purification procedures
精度、准确度、敏感性较低 Low accuracy, precision, sensibility and efficiency
Hamada & Ono, 1984; Angers et al., 1988;Tanaka et al., 1990;
HPAEC-PAD
无需衍生化; 应用范围广, 可同时分析糖醛酸和中性糖 No need for derivatization; Wide application and simultaneous analysis of uronic acid and neutral sugars
精度、准确度、敏感性较低 Low accuracy, precision, sensibility and efficiency
Bruggink et al., 2005; Zhang et al., 2012
TFA, trifluoroacetic acid; GC-MS, gas chromatography-mass spectrometry; HPLC, high-performance liquid chromatography; HPAEC-PAD, high-performance anion-exchange chromatography with pulsed amperometric detection. ...
... 中性糖的检测分析可使用气相色谱-质谱联用仪(GC-MS)(Rumpel & Dignac, 2006; Zhang et al., 2007; Eder et al., 2010), 液相色谱(HPLC)(Takeuchi et al., 1987; Tanaka et al., 1990; Amelung et al., 1996; Basler & Dyckmans, 2013)和高效阴离子交换色谱-脉冲安培检测法(HPAEC-PAD)(Bruggink et al., 2005; Zhang et al., 2012).由于中性糖不易挥发, 因此在使用GC-MS检测前需衍生化, 衍生化是影响土壤中性糖定量的关键环节.下面将介绍几种常用的中性糖衍生化方法. ...
Aspect-vegetation complex effects on biochemical characteristics and decomposability of soil organic carbon on the eastern Qinghai-Tibetan Plateau 5 2014
... 本研究整合了5种土地利用类型的土壤中性糖数据, 发现阔叶林(7.0-82.8 mg·g-1)、针叶林(1.1-90.1 mg·g-1)、灌丛(2.5-80.6 mg·g-1)和草地(1.1-15.1 mg·g-1)土壤中性糖的绝对含量无显著差异(p > 0.05; 图1A), 除草地外, 其他类型均显著高于耕地土壤(0.6-6.3 mg·g-1)(p < 0.05; 图1A).不同地区(西班牙西北部、美国中西部、法国西南部)、不同土壤类型(碳酸钙土壤、沙质淋溶土)中的研究均表明阔叶林和针叶林土壤中性糖(或碳水化合物, 包括纤维素和半纤维中性糖, 下同)的绝对含量显著高于耕地土壤, 并认为可能是人为收获引起的地上输入量减少, 以及耕作扰动导致土壤有机层中的有机质向矿质层迁移造成的(Martens et al., 2004; Jolivet et al., 2006; Llorente et al., 2017).但是有研究报道中性糖(或碳水化合物)在草地土壤中更丰富(与耕地相比)(Larré-Larrouy et al., 2004; Martens et al., 2004; Guan et al., 2018).如, Guan等(2018)发现青藏高原的自然草地经过长期(>50年)耕种后, 植物地下生物量降低使植物碳向土壤中的输入减少, 加上耕作的扰动作用强烈影响了微生物的群落结构和丰度, 导致与草地相比, 耕地土壤中性糖含量降低了57%.Larré-Larrouy等(2004)也认为农作物的根系不发达, 来源于根系的碳水化合物较少是导致此现象的主要原因.由于本研究的数据基于土壤类型、气候条件等各异的研究, 中性糖含量在各研究之间存在差异, 加之数据量的限制导致了与上述研究的结果不一致.
Content and distribution of neutral sugars across different land-use regimes in the top soils (<xref ref-type="bibr" rid="b58">Nierop<i> et al</i>., 2001</xref>; <xref ref-type="bibr" rid="b71">Spielvogel<i> et al</i>., 2007</xref>; <xref ref-type="bibr" rid="b25">Eder<i> et al</i>., 2010</xref>; <xref ref-type="bibr" rid="b67">Rumpel<i> et al</i>., 2010</xref>; <xref ref-type="bibr" rid="b90">Zhao<i> et al</i>., 2014</xref>; <xref ref-type="bibr" rid="b19">Conti<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b21">Cui<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b82">Wang<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b20">Creme<i> et al</i>., 2017</xref>; <xref ref-type="bibr" rid="b49">Llorente<i> et al</i>., 2017</xref>; <xref ref-type="bibr" rid="b26">Evgrafova <i>et al</i>., 2018</xref>; <xref ref-type="bibr" rid="b91">Zhu <i>et al</i>., 2018</xref>). A, Neutral sugar absolute content. B, Neutral sugar relative content. C, GM/AX ((galactose + mannose)/(arabinose + xylose)). D, RF/AX ((rhamnose + fucose)/(arabinose + xylose)). The upper and lower end of boxes denote the 0.25 and 0.75 percentiles, respectively. The solid bar in the box mark the median of each dataset. The circles indicate outliers of each dataset. Different lowercase letters indicate differences in various land-use regimes (<i>p</i> < 0.05). <i>n</i> = 8, 25, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the A, C). <i>n</i> = 8, 22, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the B). <i>n</i> = 3, 19, 4, 8, 27 (from deciduous, coniferous, shrub, grassland to crops in the D). OC, soil organic carbon.Fig. 1 为考察不同土地利用类型中的中性糖对土壤有机碳(OC)的潜在贡献, 我们利用土壤有机碳含量对中性糖含量进行了归一化.结果显示: 阔叶林(84.5-253.9 mg·g-1)、针叶林(55.2-291.0 mg·g-1)、灌丛(139.0-208.9 mg·g-1)和草地(161.0- 371.0 mg·g-1)土壤中性糖的相对含量无显著差异(p > 0.05; 图1B), 除灌丛外, 其他类型的中性糖相对含量均显著高于耕地(65.0-370.7 mg·g-1)(p < 0.05; 图1B).这与上述中性糖的绝对含量结果一致, 说明耕种会通过减少植物残体的输入从而使土壤中性糖含量减少.也有研究发现虽然土地利用类型由森林转变为灌丛, 但主要的维管束植物种类未改变, 凋落物质量没有变化导致两种土地利用类型土壤中性糖含量无显著差异(Conti et al., 2016).而Zhao等(2014)在青藏高原的研究表明, 草地土壤中性糖的相对含量显著高于灌丛, 可能是草本植物凋落物的半纤维素含量比灌丛植物凋落物中的含量高导致的. ...
... ; Zhao et al., 2014; Conti et al., 2016; Cui et al., 2016; Wang et al., 2016; Creme et al., 2017; Llorente et al., 2017; Evgrafova et al., 2018; Zhu et al., 2018). A, Neutral sugar absolute content. B, Neutral sugar relative content. C, GM/AX ((galactose + mannose)/(arabinose + xylose)). D, RF/AX ((rhamnose + fucose)/(arabinose + xylose)). The upper and lower end of boxes denote the 0.25 and 0.75 percentiles, respectively. The solid bar in the box mark the median of each dataset. The circles indicate outliers of each dataset. Different lowercase letters indicate differences in various land-use regimes (p < 0.05). n = 8, 25, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the A, C). n = 8, 22, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the B). n = 3, 19, 4, 8, 27 (from deciduous, coniferous, shrub, grassland to crops in the D). OC, soil organic carbon. Fig. 1 为考察不同土地利用类型中的中性糖对土壤有机碳(OC)的潜在贡献, 我们利用土壤有机碳含量对中性糖含量进行了归一化.结果显示: 阔叶林(84.5-253.9 mg·g-1)、针叶林(55.2-291.0 mg·g-1)、灌丛(139.0-208.9 mg·g-1)和草地(161.0- 371.0 mg·g-1)土壤中性糖的相对含量无显著差异(p > 0.05; 图1B), 除灌丛外, 其他类型的中性糖相对含量均显著高于耕地(65.0-370.7 mg·g-1)(p < 0.05; 图1B).这与上述中性糖的绝对含量结果一致, 说明耕种会通过减少植物残体的输入从而使土壤中性糖含量减少.也有研究发现虽然土地利用类型由森林转变为灌丛, 但主要的维管束植物种类未改变, 凋落物质量没有变化导致两种土地利用类型土壤中性糖含量无显著差异(Conti et al., 2016).而Zhao等(2014)在青藏高原的研究表明, 草地土壤中性糖的相对含量显著高于灌丛, 可能是草本植物凋落物的半纤维素含量比灌丛植物凋落物中的含量高导致的. ...
Content and distribution of neutral sugars across different land-use regimes in the top soils (<xref ref-type="bibr" rid="b58">Nierop<i> et al</i>., 2001</xref>; <xref ref-type="bibr" rid="b71">Spielvogel<i> et al</i>., 2007</xref>; <xref ref-type="bibr" rid="b25">Eder<i> et al</i>., 2010</xref>; <xref ref-type="bibr" rid="b67">Rumpel<i> et al</i>., 2010</xref>; <xref ref-type="bibr" rid="b90">Zhao<i> et al</i>., 2014</xref>; <xref ref-type="bibr" rid="b19">Conti<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b21">Cui<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b82">Wang<i> et al</i>., 2016</xref>; <xref ref-type="bibr" rid="b20">Creme<i> et al</i>., 2017</xref>; <xref ref-type="bibr" rid="b49">Llorente<i> et al</i>., 2017</xref>; <xref ref-type="bibr" rid="b26">Evgrafova <i>et al</i>., 2018</xref>; <xref ref-type="bibr" rid="b91">Zhu <i>et al</i>., 2018</xref>). A, Neutral sugar absolute content. B, Neutral sugar relative content. C, GM/AX ((galactose + mannose)/(arabinose + xylose)). D, RF/AX ((rhamnose + fucose)/(arabinose + xylose)). The upper and lower end of boxes denote the 0.25 and 0.75 percentiles, respectively. The solid bar in the box mark the median of each dataset. The circles indicate outliers of each dataset. Different lowercase letters indicate differences in various land-use regimes (<i>p</i> < 0.05). <i>n</i> = 8, 25, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the A, C). <i>n</i> = 8, 22, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the B). <i>n</i> = 3, 19, 4, 8, 27 (from deciduous, coniferous, shrub, grassland to crops in the D). OC, soil organic carbon.Fig. 1 为考察不同土地利用类型中的中性糖对土壤有机碳(OC)的潜在贡献, 我们利用土壤有机碳含量对中性糖含量进行了归一化.结果显示: 阔叶林(84.5-253.9 mg·g-1)、针叶林(55.2-291.0 mg·g-1)、灌丛(139.0-208.9 mg·g-1)和草地(161.0- 371.0 mg·g-1)土壤中性糖的相对含量无显著差异(p > 0.05; 图1B), 除灌丛外, 其他类型的中性糖相对含量均显著高于耕地(65.0-370.7 mg·g-1)(p < 0.05; 图1B).这与上述中性糖的绝对含量结果一致, 说明耕种会通过减少植物残体的输入从而使土壤中性糖含量减少.也有研究发现虽然土地利用类型由森林转变为灌丛, 但主要的维管束植物种类未改变, 凋落物质量没有变化导致两种土地利用类型土壤中性糖含量无显著差异(Conti et al., 2016).而Zhao等(2014)在青藏高原的研究表明, 草地土壤中性糖的相对含量显著高于灌丛, 可能是草本植物凋落物的半纤维素含量比灌丛植物凋落物中的含量高导致的. ...
... ; Zhu et al., 2018). A, Neutral sugar absolute content. B, Neutral sugar relative content. C, GM/AX ((galactose + mannose)/(arabinose + xylose)). D, RF/AX ((rhamnose + fucose)/(arabinose + xylose)). The upper and lower end of boxes denote the 0.25 and 0.75 percentiles, respectively. The solid bar in the box mark the median of each dataset. The circles indicate outliers of each dataset. Different lowercase letters indicate differences in various land-use regimes (p < 0.05). n = 8, 25, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the A, C). n = 8, 22, 4, 8, 15 (from deciduous, coniferous, shrub, grassland to crops in the B). n = 3, 19, 4, 8, 27 (from deciduous, coniferous, shrub, grassland to crops in the D). OC, soil organic carbon. Fig. 1 为考察不同土地利用类型中的中性糖对土壤有机碳(OC)的潜在贡献, 我们利用土壤有机碳含量对中性糖含量进行了归一化.结果显示: 阔叶林(84.5-253.9 mg·g-1)、针叶林(55.2-291.0 mg·g-1)、灌丛(139.0-208.9 mg·g-1)和草地(161.0- 371.0 mg·g-1)土壤中性糖的相对含量无显著差异(p > 0.05; 图1B), 除灌丛外, 其他类型的中性糖相对含量均显著高于耕地(65.0-370.7 mg·g-1)(p < 0.05; 图1B).这与上述中性糖的绝对含量结果一致, 说明耕种会通过减少植物残体的输入从而使土壤中性糖含量减少.也有研究发现虽然土地利用类型由森林转变为灌丛, 但主要的维管束植物种类未改变, 凋落物质量没有变化导致两种土地利用类型土壤中性糖含量无显著差异(Conti et al., 2016).而Zhao等(2014)在青藏高原的研究表明, 草地土壤中性糖的相对含量显著高于灌丛, 可能是草本植物凋落物的半纤维素含量比灌丛植物凋落物中的含量高导致的. ...
 ,
, ,1,2,*
,1,2,*
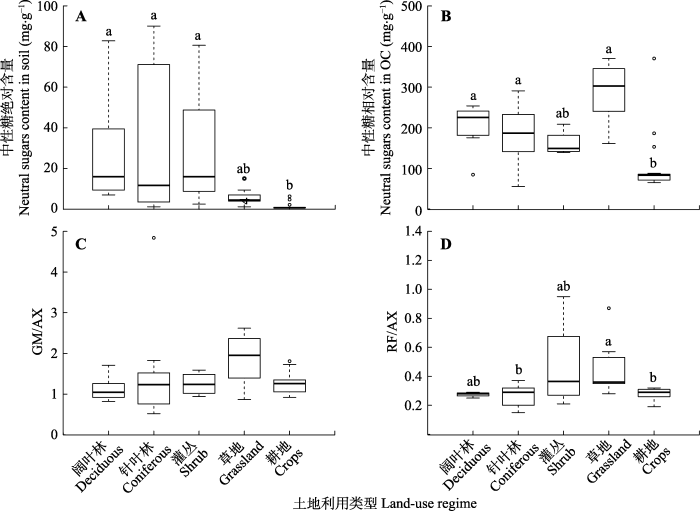 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}