Effects of topography on leaf functional traits across plant life forms in Xingdou Mountain, Hubei, China
Jin WANG1, Jiang ZHU,1,*, Xun-Ru AI1, Lan YAO1, Xiao HUANG1, Man-Ling WU1, Qiang ZHU1, Jian- Feng HONG21. School of Forestry and Horticulture, Hubei Minzu University, Enshi, Hubei 445000, China 2. Bureau of Xingdoushan National Nature Reserve, Enshi, Hubei 445000, China
Abstract Aims Exploring the effects of topography on leaf functional traits across plant life forms is helpful to understand species diversity maintenance in forests. The objective of this study is to test how leaf traits among trees, shrubs and woody lianas respond to topographic variability distinctly. Methods Five leaf functional traits (leaf area, leaf thickness, leaf dry mass, leaf dry matter content and specific leaf area) were measured for 224 woody plant species with contrasting life forms (trees, shrubs and woody lianas) across 50 plots of evergreen and deciduous broadleaved mixed forests in Xingdou Mountain, Hubei Province, China. The variation in leaf functional traits of trees, shrubs and woody lianas were analyzed by one-way ANOVA. We tested the significance of the effect of topographic variability on leaf functional traits of woody plants by Partial Mantel test at community level and species level, respectively. Important findings The coefficient of variation of leaf functional traits in different life forms ranged from 23.42% to 110.45%, and there were significant differences in leaf functional traits among different life forms. At the community level, there was a significant positive correlation between altitude and tree leaf dry mass, shrub leaf area and woody liana leaf thickness. Slope only had significant influences on shrub and woody liana leaf area, and slope aspect had significant effects on shrubs leaf thickness. There was a significant positive correlation between leaf dry mass and specific leaf area. At the species level, the effects of slope and slope aspect on plant leaf functional traits were more significant than those of altitude, and the sensitivity in the responses of functional traits to topographic variation was not consistent across different species. After controlling for the effects of spatial structure, the effects of topographic factors on functional traits of plant leaves were reduced. The results showed that the responses of leaf functional traits to topographic variation were different in different life forms, which might be one of the major mechanisms maintaining plant diversity in the evergreen and deciduous broadleaved mixed forests of Xingdou Mountain. Keywords:evergreen and deciduous broadleaved mixed forest;variation of traits;spatial structure;Partial Mantel test
PDF (1391KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 王进, 朱江, 艾训儒, 姚兰, 黄小, 吴漫玲, 朱强, 洪建峰. 湖北星斗山地形变化对不同生活型植物叶功能性状的影响. 植物生态学报, 2019, 43(5): 447-457. DOI: 10.17521/cjpe.2018.0228 WANG Jin, ZHU Jiang, AI Xun-Ru, YAO Lan, HUANG Xiao, WU Man-Ling, ZHU Qiang, HONG Jian- Feng. Effects of topography on leaf functional traits across plant life forms in Xingdou Mountain, Hubei, China. Chinese Journal of Plant Ecology, 2019, 43(5): 447-457. DOI: 10.17521/cjpe.2018.0228
Fig. 1Basic topography status of 50 plots in Xingdou Moutain. The altitude and slope were presented as the actual observation data, and the aspects were sinusoidally transformed.
Fig. 2Species diversity of plants from different life forms in 50 plots of Xingdou Mountain. In the box, “□” is the average, “—” is the median. “○” is the abnormal value.
新窗口打开|下载原图ZIP|生成PPT 图3群落水平3种生活型植物叶功能性状变异。图中百分数据为变异系数(CV); 箱体中“█”为平均值、“—”为中位数; “○”为异常值。**, p < 0.01; *, p < 0.05; ns, p > 0.05。
Fig. 3Variation of leaf functional traits across three life forms of plants at community level. The figures on top of the boxed are the coefficients of variation (CV) of that trait across all plots. In the box, “█” is the average, “—” is the median. “○” is the abnormal value. LA, leaf area; LT, leaf thickness; LDM, leaf dry mass; LDMC, leaf dry matter content; SLA, specific leaf area. **, p < 0.01; *, p < 0.05; ns, p > 0.05.
Fig. 4Variation in leaf functional traits across dominant species (species level). LA, leaf area; LT, leaf thickness; LDM, leaf dry mass; LDMC, leaf dry matter content; SLA, specific leaf area. The figures on top of the boxes are the coefficients of variation of each trait for a species across all sampled plots. In the box, “█” is the average, “—” is the median. “○” is the abnormal value. 1-6 represent species in the abscissa: 1, Carpinus fargesiana; 2, Enkianthus serrulatus; 3, Eurya alata; 4, Stranvaesia davidiana; 5, Dalbergia dyeriana; 6, Actinidia chinensis.
Table 1 表1 表1群落水平上不同生活型植物叶功能性状与地形因子的偏曼特尔检验 Table 1Partial Mantel tests for the relationships between leaf functional traits of different life forms and topographic factors at community level
生活型 Life form
性状 Trait
空间结构 Spatial structure
未控制空间结构影响 Uncontrolled spatial structure effect
控制空间结构影响 Controlling the influence of spatial structure
海拔 Altitude
坡度 Slope
坡向 Slope aspect
海拔 Altitude
坡度 Slope
坡向 Slope aspect
乔木 Tree
LA (cm2)
0.26*
0.27*
0.07
0.15*
0.10
0.00
0.09
LT (mm)
-0.02
-0.02
-0.02
0.00
-0.01
-0.02
0.01
LDM (g)
0.15
0.20*
0.02
0.12
0.15*
-0.02
0.09
LDMC (g·g-1)
0.13*
0.08
-0.00
0.05
-0.09
-0.04
0.02
SLA (cm2·g-1)
0.06
0.07
0.01
0.09
0.04
-0.01
0.08
灌木 Shrub
LA (cm2)
0.18*
0.23**
0.05
0.13*
0.15*
-0.00
0.09
LT (mm)
0.39**
0.36**
0.02
0.19**
0.03
-0.09
0.11*
LDM (g)
0.32**
0.31**
0.19*
0.34**
0.07
0.12
0.29**
LDMC (g·g-1)
0.15
0.11
0.12
0.02
-0.04
0.08
-0.01
SLA (cm2·g-1)
0.30**
0.31**
0.23**
0.19**
0.10
0.16*
0.13*
木质藤本Woody liana
LA (cm2)
-0.02
-0.03
-0.07
0.01
-0.02
-0.07
0.02
LT (mm)
0.23
0.39*
0.21
0.11
0.40**
0.14
0.05
LDM (g)
-0.15
-0.16*
-0.04
-0.05
-0.05
0.01
-0.02
LDMC (g·g-1)
-0.07
-0.08
0.04
-0.09
-0.05
0.06
-0.07
SLA (cm2·g-1)
0.16
0.20
-0.06
0.02
0.12
-0.12*
-0.02
LA, leaf area; LT, leaf thickness; LDM, leaf dry mass; LDMC, leaf dry matter content; SLA, specific leaf area. The Partial Mantel coefficients and their significance were obtained by 999 times of randomization . **, p < 0.01; *, p < 0.05. LA, 叶面积; LT, 叶厚度; LDM, 叶干质量; LDMC, 叶干物质含量; SLA, 比叶面积。根据控制不同因子和叶功能性状之间进行999次置换得出Partial Mantel系数及其显著性。**, p < 0.01; *, p < 0.05。
Table 2 表2 表2物种水平上不同生活型植物叶功能性状与地形因子的偏曼特尔检验 Table 2Partial Mantel tests for the relationships between leaf functional traits of different life forms and topographic factors at species level across
物种 Species
性状 Trait
空间结构 Spatial structure
未控制空间结构影响 Uncontrolled spatial structure effect
控制空间结构影响 Controlling the influence of spatial structure
海拔 Altitude
坡度 Slope
坡向 Slope aspect
海拔 Altitude
坡度 Slope
坡向 Slope aspect
川陕鹅耳枥 Carpinus fargesiana
LA (cm2)
0.27*
0.34*
0.16
0.20
0.24*
0.07
0.12
LT (mm)
0.40**
0.43**
0.09
0.29**
0.15
-0.07
0.20*
LDM (g)
0.37**
0.38**
0.19*
0.16*
0.12
0.06
0.07
LDMC (g·g-1)
0.21*
0.12
-0.05
0.17*
-0.16
-0.14*
0.11
SLA (cm2·g-1)
0.35*
0.38*
0.21*
0.15
0.16
0.09
0.06
齿缘吊钟花 Enkianthus serrulatus
LA (cm2)
0.18*
0.17*
-0.01
-0.03
0.03
-0.02
-0.09
LT (mm)
-0.10
-0.10
0.04
-0.06
-0.03
0.04
-0.03
LDM (g)
0.29*
0.30*
0.03
0.05
0.11
0.02
-0.04
LDMC (g·g-1)
0.17
0.06
0.02
0.04
-0.17*
0.02
-0.01
SLA (cm2·g-1)
0.06
0.08
-0.01
-0.06
0.07
-0.01
-0.08
翅柃 Eurya alata
LA (cm2)
0.38**
0.40**
0.12
0.04
0.17*
0.02
-0.06
LT (mm)
0.02
0.05
-0.02
0.07
0.08
-0.03
0.07
LDM (g)
0.15*
0.18*
0.01
0.07
0.12
-0.03
0.04
LDMC (g·g-1)
0.22**
0.20*
-0.02
0.02
0.01
-0.08
-0.04
SLA (cm2·g-1)
0.11
0.07
0.02
0.20**
-0.07
-0.01
0.18**
红果树 Stranvaesia davidiana
LA (cm2)
0.15
0.24*
0.16*
0.03
0.21*
0.13
-0.02
LT (mm)
-0.03
-0.08
-0.05
0.05
-0.10
-0.04
0.07
LDM (g)
0.30*
0.37*
0.00
0.39**
0.23*
-0.08
0.32**
LDMC (g·g-1)
0.09
0.16
0.03
0.11
0.15
0.01
0.09
SLA (cm2·g-1)
0.29*
0.36*
0.02
0.32**
0.23*
-0.05
0.25**
大金刚藤 Dalbergia dyeriana
LA (cm2)
0.21
0.38**
-0.14
0.62**
0.33*
-0.14*
0.60*
LT (mm)
0.37*
0.07
-0.14
0.05
-0.13
-0.15
-0.12
LDM (g)
-0.05
0.00
-0.08
-0.13
0.03
-0.08
-0.12
LDMC (g·g-1)
0.27
-0.15
-0.05
-0.09
-0.32**
-0.05
-0.23*
SLA (cm2·g-1)
0.01
0.27*
0.01
0.12
0.29*
0.01
0.12
中华猕猴桃 Actinidia chinensis
LA (cm2)
0.43**
0.50**
0.06
-0.01
0.29*
-0.25**
-0.10
LT (mm)
-0.08
-0.08
-0.07
0.01
-0.02
-0.03
0.02
LDM (g)
0.21
0.11
0.07
-0.01
-0.17
-0.06
-0.05
LDMC (g·g-1)
-0.20*
-0.17*
-0.17**
-0.16
0.03
-0.07
-0.13
SLA (cm2·g-1)
0.12
0.28*
-0.12
0.11
0.40**
-0.23**
0.09
LA, leaf area; LT, leaf thickness; LDM, leaf dry mass; LDMC, leaf dry matter content; SLA, specific leaf area. The Partial Mantel coefficients and their significance were obtained by 999 times of randomization. **, p < 0.01; *, p < 0.05. LA, 叶面积; LT, 叶厚度; LDM, 叶干质量; LDMC, 叶干物质含量; SLA, 比叶面积。根据控制不同因子和叶功能性状之间进行999次置换得出Partial Mantel系数及其显著性。**, p < 0.01; *, p < 0.05。
AemW, FulePZ, CovingtonWW, MooreMM (2003). Diversity in ponderosa pine forest structure following ecological restoration treatments Forest Science, 49, 885-900. [本文引用: 1]
CantónY, BarrioGD, Solé-BenetA, LázaroR (2004). Topographic controls on the spatial distribution of ground cover in the Tabernas badlands of SE Spain Catena, 55, 341-365. [本文引用: 1]
ChenC, LiuDH, WuJJ, KangMY, ZhangJT, LiuQR, LiangY (2015). Leaf traits of Quercus wutaishanica and their relationship with topographic factors in Mount Dongling Chinese Journal of Ecology, 34, 2131-2139. [本文引用: 1]
ConditR (1998). Tropical Forest Census Plots: Methods and Results from Barro Colorado Island, Panama and A Comparison with Other Plots. Springer-Verlag, Berlin Heidelberg, New York. [本文引用: 1]
CornelissenJHC, CeraboliniB, CastrodíezP, VillarsalvadorP, MontserratmartíG, PuyravaudJP, MaestroM, WergerMJA, AertsR (2003a). Functional traits of woody plants: Correspondence of species rankings between field adults and laboratory-grown seedlings? Journal of Vegetation Science, 14, 311-322. [本文引用: 1]
CornelissenJHC, LavorelS, GarnierE, BuchmannN, GurvichDE, ReichPB, SteegeHT, MorganHD, HeijdenMGAV (2003b). A handbook of protocols for standardised and easy measurement of plant functional traits worldwide Australian Journal of Botany, 51, 335-380. [本文引用: 2]
CornwellWK, AckerlyDD (2009). Community assembly and shifts in plant trait distributions across an environmental gradient in coastal California Ecological Monographs, 79, 109-126.
CornwellWK, SchwilkLDW, AckerlyDD (2006). A trait- based test for habitat filtering: Convex hull volume Ecology, 87, 1465-1471. [本文引用: 1]
DaiN, HirabukiY, MochidaY (2010). Influence of micro- landforms on forest structure, tree death and recruitment in a Japanese temperate mixed forest Ecological Research, 18, 533-547. [本文引用: 1]
DangJJ, ZhaoCZ, LiY, HouZJ, DongXG (2014). Variations with slope in stem and leaf traits of Melica przewalskyi in alpine grassland Chinese Journal of Plant Ecology, 38, 1307-1314. [本文引用: 1]
D??azS, CabidoM (2001). Vive la difference: Plant functional diversity matters to ecosystem processes Trends in Ecology & Evolution, 16, 646-655. [本文引用: 1]
DingJ, WuQ, YanH, ZhangSR (2011). Effects of topographic variations and soil characteristics on plant functional traits in a subtropical ever green broad-leaved forest Biodiversity Science, 19, 158-167. [本文引用: 2]
GaoXM, ChenLZ (1998). The revision of plant life form system and an analysis of the life-form spectrum of forest plants in the warm temperate zone of China Acta Botanica Sinica, 40, 553-559. [本文引用: 1]
HaraM, HirataK, FujiharaM, OonoK (1996). Vegetation structure in relation to micro-landform in an evergreen broad-leaved forest on Amami Ohshima Island, south-west Japan Ecological Research, 11, 325-337. [本文引用: 1]
HarrodRJ, McraeBH, HartlWE (1999). Historical stand reconstruction in ponderosa pine forests to guide silvicultural prescriptions Forest Ecology and Management, 114, 433-446. [本文引用: 1]
HeD, YanER (2018). Size-dependent variations in individual traits and trait scaling relationships within a shade-tolerant evergreen tree species American Journal of Botany, 105, 1165-1174. [本文引用: 1]
HomyackJA, HarrisonDJ, KrohnWB (2004). Structural differences between precommercially thinned and unthinned conifer stands Forest Ecology and Management, 194, 131-143. [本文引用: 1]
HuangHX, YangXD, SunBW, ZhangZH, YanER (2013). Variability and association of leaf traits between current- year and former-year leaves in evergreen trees in Tiantong, Zhejiang, China Chinese Journal of Plant Ecology, 37, 912-921. [本文引用: 1]
HuangYT, YaoL, AiXR, LüSA, DingY (2015). Quantitative classification of the subtropical evergreen-deciduous broadleaved mixed forest and the deciduous and evergreen species composition structure across two national nature reserves in the southwest of Hubei, China Chinese Journal of Plant Ecology, 39, 990-1002. [本文引用: 1]
KayamaM, SasaK, KoikeT (2002). Needle life span, photosynthetic rate and nutrient concentration of Picea glehnii, P. jezoensis and P. abies planted on serpentine soil in northern Japan Tree Physiology, 22, 707-716. [本文引用: 1]
KenzoT, InoueY, YoshimuraM, YamashitaM, TanakaodaA, IchieT (2015). Height-related changes in leaf photosynthetic traits in diverse Bornean tropical rain forest trees Oecologia, 177, 191-202. [本文引用: 1]
LiHW, WangXA, GuoH (2012). Leaf functional traits of different forest communities in Ziwuling Mountains of Loess Plateau Chinese Journal of Ecology, 31, 544-550.
LiJX, XuWT, XiongGM, WangY, ZhaoCM, LuZJ, LiYL, XieZQ (2017). Leaf nitrogen and phosphorus concentration and the empirical regulations in dominant woody plants of shrublands across southern China Chinese Journal of Plant Ecology, 41, 31-42. [本文引用: 1]
LiY, ZhaoCZ, DongXG, HouZJ, MaXL, ZhangQ (2013a). Twig and leaf trait differences in Stellera chamaejasme with slope in alpine grassland Chinese Journal of Plant Ecology, 37, 709-717. [本文引用: 2]
LiY, ZhaoCZ, DongXG, HouZJ, MaXL, ZhangQ (2013b). Responses of Stellera chamaejasme twig and leaf traits to slope aspect in alpine grassland of Northwest China Chinese Journal of Ecology, 32, 3145-3151. [本文引用: 2]
LiuYP, LiuGF, BaiyilaDF, ChengWY, ChenZJ, JiangLL (2017). Effects of topographic factors on leaf traits of dominant species in different forest communities in Daqinggou Nature Reserves Scientia Silvae Sinicae, 53(3), 154-162.
MarkAF, DickinsonKJM, AllenJ, SmithR, WestCJ (2010). Vegetation patterns, plant distribution and life forms across the alpine zone in southern Tierra del Fuego, Argentina Austral Ecology, 26, 423-440. [本文引用: 1]
PoorterL, WrightSJ, PazH, AckerlyDD, ConditR, Ibarra-?manríquezG, HarmsKE, LiconaJC, Martínez-ramosM, MazerSJ (2008). Are functional traits good predictors of demographic rates? Evidence from five neotropical forests Ecology, 89, 1908-1920. [本文引用: 1]
PutzFE (1984). How trees avoid and shed liana Biotropica, 1, 19-23.
ReichPB (2003). The evolution of plant functional variation: Traits, spectra, and strategies International Journal of Plant Sciences, 164, S143-S164. [本文引用: 1]
SakaiA, OhsawaM (1994). Topographical pattern of the forest vegetation on a river basin in a warm-temperate hilly region, central Japan Ecological Research, 9, 269-280. [本文引用: 1]
SchnitzerSA, BongersF (2002). The ecology of lianas and their role in forests Trends in Ecology & Evolution, 5, 223-230.
ShenZH (2002). A multi-scale study on the vegetation-?environment relationship of a mountain forest transect Acta Ecologica Sinica, 22, 461-470. [本文引用: 1]
ToledoM, PoorterL, ClarosMP, AlarcónA, BalcázarJ, Lea?oC, LiconaJC, LlanqueO, VroomansV, ZuidemaP, BongersF (2011). Climate is a stronger driver of tree and forest growth rates than soil and disturbance Journal of Ecology, 99, 254-264. [本文引用: 1]
WaltzAEM, FuléPZ, CovingtonWW, MooreMM (2003). Diversity in ponderosa pine forest structure following ecological restoration treatments Forest Science, 49, 885-900. [本文引用: 1]
WangCS, WangSP (2015). A review of research on responses of leaf traits to climate change Chinese Journal of Plant Ecology, 39, 206-216. [本文引用: 1]
WangHY, ChenH (2013). Plant functional groups based on vegetative and reproductive traits in a subtropical forest community Journal of Forest Research, 18, 482-490. [本文引用: 1]
WestobyM, WrightIJ (2006). Land-plant ecology on the basis of functional traits Trends in Ecology & Evolution, 21, 261-268. [本文引用: 1]
WuZY (1980). Chinese Vegetation Science Press, Beijing. 279-306. [本文引用: 1]
ZhangQP (2011). Differentiation Patterns of Leaf Traits of Main Trees Along Altitudinal and Micro-Topographical Gradients on Tianmushan Mountain Master degree dissertation, East China Normal University, Shanghai. [本文引用: 1]
ZhongQL, LiuLB, XuX, YangY, GuoYM, XuHY, CaiXL, NiJ (2018). Variations of plant functional traits and adaptive strategy of woody species in a karst forest of central Guizhou Province, southwestern China Chinese Journal of Plant Ecology, 42, 562-572. [本文引用: 1]
ZhuJD, MengTT, NiJ, SuHX, XieZQ, ZhangSR, ZhengYR, XiaoCW (2011). Within-leaf allometric relationships of mature forests in different bioclimatic zones vary with plant functional types Chinese Journal of Plant Ecology, 35, 687-698. [本文引用: 1]
Needle life span, photosynthetic rate and nutrient concentration of Picea glehnii, P. jezoensis and P. abies planted on serpentine soil in northern Japan 1 2002
 ,1,*, 艾训儒1, 姚兰1, 黄小1, 吴漫玲1, 朱强1, 洪建峰21.
,1,*, 艾训儒1, 姚兰1, 黄小1, 吴漫玲1, 朱强1, 洪建峰21. 
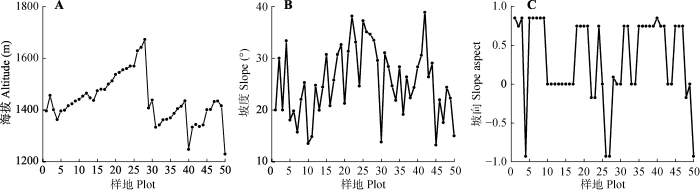 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT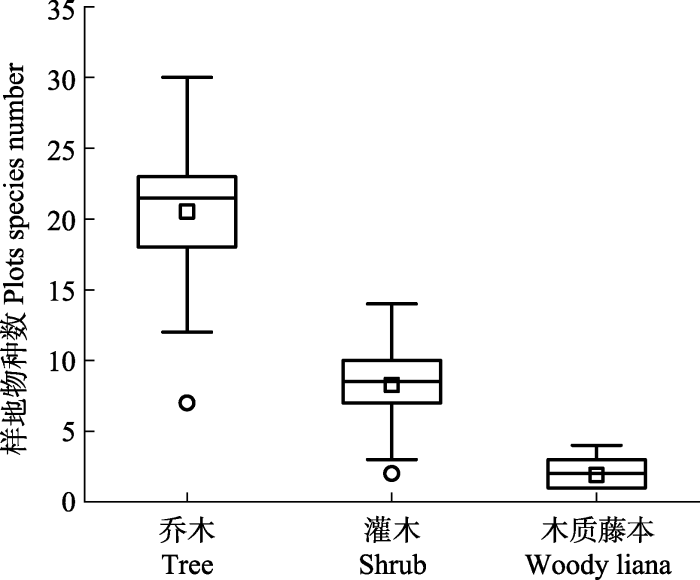 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT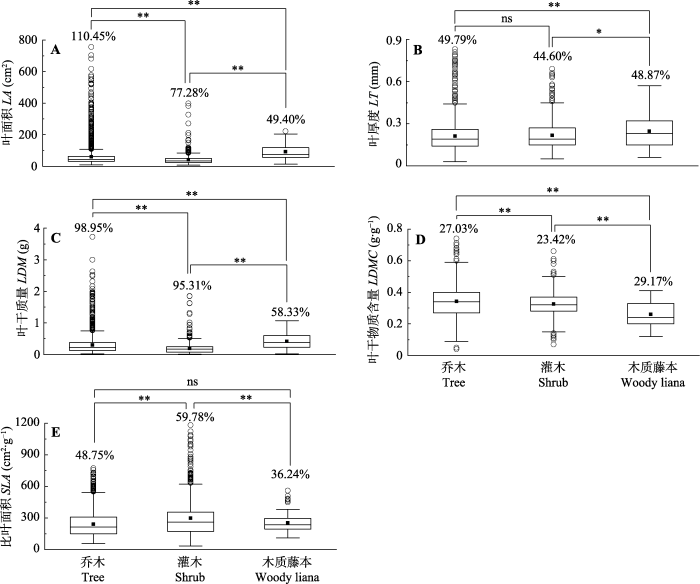 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT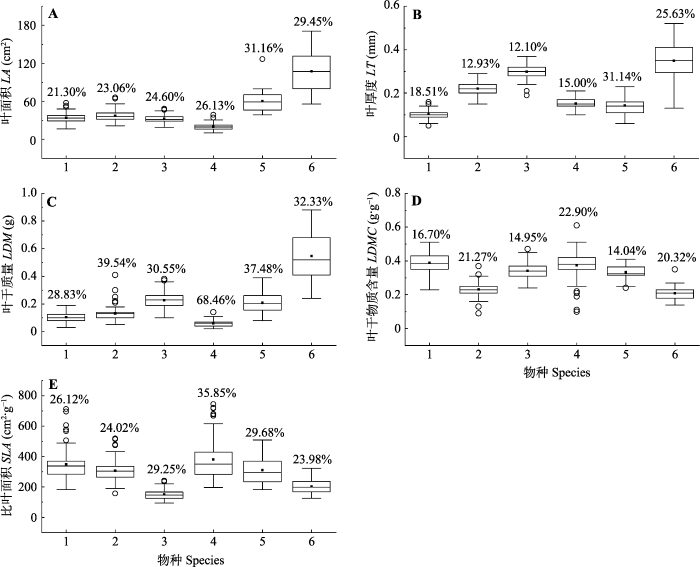 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}