Research advances in modelling plant species distribution in China
Xiao-Tong LIU,1, Quan YUAN1, Jian NI,,1,2,*1 College of Chemistry and Life Sciences, Zhejiang Normal University, Jinhua, Zhejiang 321004, China 2 Jinhua Mountain Observation and Research Station for Subtropical Forest Ecosystems, Jinhua, Zhejiang 321004, China
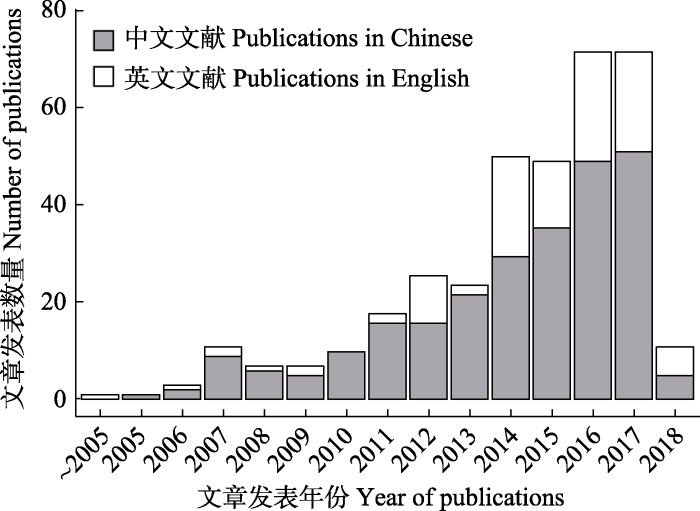
Abstract Species distribution models (SDMs) have been extensively used in simulations of geographical distribution of animal and plant species during the past 20 years. Taking the simulation of plant species distribution as an example, we used both the digitized and library databases including the China National Knowledge Infrastructure (CNKI), the VIP Chinese Journal Database (VIP) and the Web of Science (WoS) to compile available literatures published from 2000 to 2018. The number of publications, SDMs used, target plant species, data sources, and the purpose of studies about using various SDMs to simulate plant species distribution in China was statistically investigated. In total 366 publications were collected. Further analysis and synthesis showed that the application of SDMs in simulating Chinese plant species distribution has developed rapidly since 2011, especially during the past five years. SDMs have been used in studies of ecology, Chinese traditional medicine, agriculture, and forestry. The Maximum Entropy Model (MaxEnt) is the most widely used model among 33 commonly used SDMs. A half of the studies use climate data only, and another half of the studies use both climate, soil and topography data. The source of both environmental data and plant distribution data are diverse, derived from international and domestic databases. In these studies, researchers have simulated the distribution of 562 plant species, in which 52.7% are woody species and 41.8% are herbaceous species, including a large number of Chinese medicinal plants, fruit trees, garden plants, and crops. Studies aim mainly on two aspects, i.e. the impact of climate change on plant species distribution and their predicted pattern in the past, present, and future climate scenarios, and the assessment of the potential distribution of plant species and biodiversity trends (including the risk of invasive species). In future studies, more attention should be paid to both the basic science on the modelling of potential distribution of plant species and the impact from climate change, and the applied science on the prediction of suitable distribution area of plant species in order to popularize their plantation. More applications of SDMs in multiple disciplines and in multiple industries such as ecology, forestry, crop science and Chinese traditional medicine should be further developed. Joint simulations and inter-comparisons using multiple plant species, more SDMs and multiple data sources of environmental data, as well as the development of new and mechanism SDMs are encouraged. The extension of model applications in new research fields is also needed. Keywords:species distribution models;climate change;biodiversity conservation;potential distribution area;Maximum Entropy Model
Table 1 表1 表1发表植物物种分布模型研究的重要期刊(2000-01-2018-01) Table 1Top journals publishing articles of plant distribution modelling from China (from 2001-01 to 2018-01)
Table 2 表2 表2模拟中国植物种分布的模型及其应用次数 Table 2Statistics of publications modeling plant species in China using species distribution models
模型名称 Model
应用数量 Number of publications
模型名称 Model
应用数量 Number of publications
最大熵模型 Maximum Entropy Model (MaxEnt) 基于规则集的遗传算法 Genetic Algorithm for Rule-set Prediction (GARP) 生物气候模型 BIOCLIM 广义线性模型 Generalized Linear Model (GLM) 广义相加模型 Generalized Additive Model (GAM) 随机森林 Random Forest (RF) DOMAIN 推进式回归树 Generalized Boosted Regression Models/Boosted Regression Tree (GBM/BRT) 多元适应回归样条函数 Multivariate Adaptive Regression Splines (MARS) 人工神经网络 Artificial Neural Network (ANN) 柔性判别分析 Flexibled Discriminant Analysis (FDA) 支持向量机 Support Vector Machine (SVM) 分类树分析 Classification Tree Analysis (CTA) 分类回归树 Classification and Regression Tree (CART) 表面分布区分室模型 Surface Range Envelope (SRE) 复合型广义相加模型运算系 Mixed GAM Computation Vehicle (MGCV)
296 27
26 21 21 19 13 13
12
11 10 9 8 6 6 3
拟合神经网络 Fit Neural Networks (NNET) 循环分区回归树 Recursive Partitioning and Regression Trees (RPART) Logistic回归模型 Logistic Regression (LR) CLIMEX 作物生态需求 Crop Ecological Requirements (ECOCROP) 农业生态区模型 Agriculture Ecological Zone Model (AEZ) 决策树模型 Classification Tree Model (CT) 生态位因子分析模型 Ecological Niche Factor Analysis (ENFA) GREEN 生境适生性模型 Habitat Suitability Model (HSM) 线性判别分析 Linear Discriminant Analysis (LDA) 马氏距离 Mahalanobis Distance (MAHAL) 迭代决策树算法 Multiple Additive Regression Tree (MART) 空间明晰物种组合模型 Spatially Explicit Species Assemblage Model (SESAM) n维环境资源模型 n-Dimentional Environment and Resource Model 生态位模型 Niche model 随机预测模型 Random Predictive model
Fig. 2Number of species distribution models used in modelling China’s plant species distribution. The small plot is the number of models with MaxEnt excluded. See Table 2 for models.
AraújoMB, PetersonAT (2012). Uses and misuses of bioclimatic envelope modeling Ecology, 93, 1527-1539. [本文引用: 1]
BusbyJ ( 1991). BIOCLIM—A bioclimate analysis and prediction system Plant Protection Quarterly, 6, 8-9. [本文引用: 1]
ChenXM, LeiYC, ZhangXQ, JiaHY ( 2012). Effects of sample sizes on accuracy and stability of maximum entropy model in predicting species distribution Scientia Silvae Sinicae, 48(1), 53-59. DOI:10.11707/j.1001-7488.20120110Magsci [本文引用: 1] <p>以实际调查的4个物种的34个不同样本量(5,6,8,10,15,20,25,30,40,50,60,70,80,90,100,120,150,180,200,220,250,300,350,400,450,500,550,600,650,700,800,900,1 000,1 200)为例,模拟计算分析不同的样本量对MaxEnt物种分布模型的精度和稳定性的影响。结果表明: 总体上来看,样本量的大小对MaxEnt模型预测物种空间分布的精度影响不大,在样本量较小时,精度不稳定,随着样本量的增大(训练数据在样本量50左右,检验数据在样本量120左右),MaxEnt模型的预测精度越来越稳定。</p> [ 陈新美, 雷渊才, 张雄清, 贾宏炎 ( 2012). 样本量对MaxEnt模型预测物种分布精度和稳定性的影响 林业科学, 48(1), 53-59.] DOI:10.11707/j.1001-7488.20120110Magsci [本文引用: 1] <p>以实际调查的4个物种的34个不同样本量(5,6,8,10,15,20,25,30,40,50,60,70,80,90,100,120,150,180,200,220,250,300,350,400,450,500,550,600,650,700,800,900,1 000,1 200)为例,模拟计算分析不同的样本量对MaxEnt物种分布模型的精度和稳定性的影响。结果表明: 总体上来看,样本量的大小对MaxEnt模型预测物种空间分布的精度影响不大,在样本量较小时,精度不稳定,随着样本量的增大(训练数据在样本量50左右,检验数据在样本量120左右),MaxEnt模型的预测精度越来越稳定。</p>
CuiXQ, MaHP, HuangGL, HouM, XuM, ZhengGQ, CuiBX, ZhuoL, LiaoCZ ( 2016). Research on the land suitable for planting 6 major tree species in Qinghai Province Forest Resources Management, ( 4), 74-78. [本文引用: 1]
DaiG, YangJ, LuS, HuangC, JinJ, JiangP, YanP ( 2018). The potential impact of invasive woody oil plants on protected areas in China under future climate conditions Scientific Reports, 8, 1041. DOI: 10.1038/s41598-018-?19477-w. [本文引用: 1]
ElithJ, LeathwickJR ( 2009). Species distribution models: Ecological explanation and prediction across space and time Annual Review of Ecology, Evolution, and Systematics, 40, 677-697. [本文引用: 2]
FangJY, WangZH, TangZY (2011). Atlas of Woody Plants in China:Distribution and Climate. Higher Education Press, Beijing. [本文引用: 1]
GaoB, WeiHY, GuoYL, GuW ( 2015). Using GIS and MaxEnt to analyze the potential distribution of Abies chensiensis Chinese Journal of Ecology, 34, 843-852. [本文引用: 1]
GuisanA, ThuillerW ( 2005). Predicting species distribution: Offering more than simple habitat models Ecology Letters, 8, 993-1009. [本文引用: 1]
HarrisI, JonesPD, OsbornTJ, ListerDH ( 2014). Updated high-resolution grids of monthly climatic observations—?The CRU TS3.10 Dataset International Journal of Climatology, 34, 623-642. [本文引用: 1]
HeQ, ZhouG ( 2012). The climatic suitability for maize cultivation in China Chinese Science Bulletin, 57, 395-403. [本文引用: 1]
HijmansRJ, CameronSE, ParraJL, JonesPG, JarvisA ( 2005). Very high resolution interpolated climate surfaces for global land areas International Journal of Climatology, 25, 1965-1978. [本文引用: 1]
HuangMY, KongXQ, DuanRY, WuGL, ZhangZX ( 2016). The basic principle of virtual species and its application to evaluations of species distribution models Acta Ecologica Sinica, 36, 2460-2470. [本文引用: 1]
JiaX, MaFF, ZhouWM, ZhouL, YuDP, QinJ, DaiLM ( 2017). Impacts of climate change on the potential geographical distribution of broadleaved Korean pine (Pinus koraiensis) forests Acta Ecologica Sinica, 37, 464-473. [本文引用: 1]
JiangX, NiJ ( 2005). Species-climate relationships of 10 desert plant species and their estimated potential distribution range in the arid lands of northwestern China Acta Phytoecologica Sinica, 29, 98-107. [本文引用: 1]
LiGQ ( 2011). Evaluation the Ecological Niche Models and Predicting Species Potential Distribution Area PhD dissertation, Institute of Botany, Chinese Academy of Sciences,Beijing. [本文引用: 1]
LiY, YanHF, GeXJ ( 2012). Phylogeographic analysis and environmental niche modeling of widespread shrub Rhododendron simsii in China reveals multiple glacial refugia during the last glacial maximum Journal of Systematics and Evolution, 50, 362-373. [本文引用: 1]
LiuSJ, ZhouGS, FangSB, ZhangJH ( 2015). Effects of future climate change on climatic suitability of rubber plantation in China Chinese Journal of Applied Ecology, 26, 2083-2090. [本文引用: 2]
LuoM, WangH, LüZ ( 2017). Evaluating the performance of species distribution models Biomod2 and MaxEnt using the giant panda distribution data Chinese Journal of Applied Ecology, 28, 4001-4006. [本文引用: 1]
MaSM, ZhangML, ZhangHX, MengHH, ChenX ( 2010). Predicting potential geographical distributions and patterns of the relic plant Gymnocarpos przewalskii using Maximum Entropy and Genetic Algorithm for Rule-set Prediction Chinese Journal of Plant Ecology, 34, 1327-1335. [本文引用: 2]
MaoLH, LiY, LiuC, FangYM ( 2017). Predication of potential distribution of Haplocladium microphyllum in China based on MaxEnt model Chinese Journal of Ecology, 36, 54-60. [本文引用: 1]
NixH, McMahonJ, MackenzieD , (1977). Potential areas of production and the future of pigeon pea and other grain legumes in Australia. In: Wallis ES, Whiteman PC eds The Potential for Pigeon Pea in Australia: Proceedings of Pigeon Pea (Cajanus cajan (L.) Millsp.). University of Queensland,Queensland. 1-12. [本文引用: 1]
PengSZ, ZhaoCY, XuZL, AshiqMW ( 2016). Restoration and conservation potential of destroyed Qinghai spruce (Picea crassifolia) forests in the Qilian Mountains of northwest China Mitigation and Adaptation Strategies for Global Change, 21, 153-165. [本文引用: 1]
PengSZ, ZhaoCY, XuZL, WangC, LiuYY ( 2011). Potential distribution of Qinghai spruce and assessment of its growth status in the upper reaches of the Heihe River in the Qilian Mountains of China Chinese Journal of Plant Ecology, 35, 605-614. [本文引用: 1]
PhillipsSJ, AndersonRP, DudíkM, SchapireRE, BlairME ( 2017). Opening the black box: An open-source release of Maxent Ecography, 40, 887-893. [本文引用: 1]
PhillipsSJ, AndersonRP, SchapireRE ( 2006). Maximum entropy modeling of species geographic distributions Ecological Modelling, 190, 231-259. [本文引用: 1]
SvenningJC, Fl?jgaardC, MarskeKA, Nógues-BravoD, NormandS ( 2011). Applications of species distribution modeling to paleobiology Quaternary Science Reviews, 30, 2930-2947. [本文引用: 1]
SvenningJC, SkovF ( 2004). Limited filling of the potential range in European tree species Ecology Letters, 7, 565-573. [本文引用: 1]
WanJZ, WangCJ, HanSJ, YuJH ( 2014). Planning the priority protected areas of endangered orchid species in northeastern China Biodiversity and Conservation, 23, 1395-1409. [本文引用: 1]
WanJZ, WangCJ, YuFH ( 2017). Spatial conservation prioritization for dominant tree species of Chinese forest communities under climate change Climatic Change, 144, 303-316. [本文引用: 1]
WangC, LinHL, FengQS, JinCY, CaoAC, HeL ( 2017a). A new strategy for the prevention and control of Eupatorium adenophorum under climate change in China Sustainability, 9, 2037. DOI: 10.3390/su9112037. [本文引用: 1]
WangD, WeiHY, YangY, ShangZH, GuW ( 2017). MaxEnt and GIS for predicting the potential distribution of Bupleurum marginatum Journal of Chinese Medicinal Materials, 40, 301-305. [本文引用: 2]
WangLH, YangJX, XuXN ( 2015). Analysis of suitable bioclimatic characteristics of Pseudolarix amabilis by using MaxEnt model Scientia Silvae Sinicae, 51(1), 127-131. DOI:10.11707/j.1001-7488.20150115Magsci [本文引用: 2] [目的]利用MaxEnt模型分析金钱松适生的生物气候特征,为金钱松林业生产提供科学依据.[方法]从我国4个标本馆获取金钱松43个标本记录数据,并从WorldClim网站下载1950—2000年生物气候数据,用最大熵模型(MaxEnt)模拟金钱松的地理分布,检测其生物气候主导因子及其适宜值.[结果]金钱松最适宜(分布值为0.73~1.0)分布的地区是湖南东部、湖北东南部、江西的东北部、安徽的南部、浙江西北部和其东部沿海地区,其余具分布值0.37~0.73的区域是以上这些高分布值区域的向外连续延伸.刀切法检测表明:冷季均温、干季均温、暖季降雨量对金钱松的分布影响较大,其中冷季均温适宜值为1.15~8.1 ℃,最适值为4.62 ℃;干季均温适宜值为2.7~11.9 ℃,最适值为7.3 ℃;暖季降雨量适宜值为45.6~734.4 mm,最适值为546.9 mm.[结论]MaxEnt模拟可信度较高,精确反映出金钱松的地理分布范围,并阐明主导其分布的3个生物气候因子,揭示金钱松在地理分布上所需的定量生物气候条件. [ 王雷宏, 杨俊仙, 徐小牛 ( 2015). 基于MaxEnt分析金钱松适生的生物气候特征 林业科学, 51(1), 127-131.] DOI:10.11707/j.1001-7488.20150115Magsci [本文引用: 2] [目的]利用MaxEnt模型分析金钱松适生的生物气候特征,为金钱松林业生产提供科学依据.[方法]从我国4个标本馆获取金钱松43个标本记录数据,并从WorldClim网站下载1950—2000年生物气候数据,用最大熵模型(MaxEnt)模拟金钱松的地理分布,检测其生物气候主导因子及其适宜值.[结果]金钱松最适宜(分布值为0.73~1.0)分布的地区是湖南东部、湖北东南部、江西的东北部、安徽的南部、浙江西北部和其东部沿海地区,其余具分布值0.37~0.73的区域是以上这些高分布值区域的向外连续延伸.刀切法检测表明:冷季均温、干季均温、暖季降雨量对金钱松的分布影响较大,其中冷季均温适宜值为1.15~8.1 ℃,最适值为4.62 ℃;干季均温适宜值为2.7~11.9 ℃,最适值为7.3 ℃;暖季降雨量适宜值为45.6~734.4 mm,最适值为546.9 mm.[结论]MaxEnt模拟可信度较高,精确反映出金钱松的地理分布范围,并阐明主导其分布的3个生物气候因子,揭示金钱松在地理分布上所需的定量生物气候条件.
WangR, WanFH ( 2016). Predicting the potential invasive distribution and early-warning monitoring management of Solanum elaeagnifolium in China Chinese Journal of Ecology, 35, 1697-1703. [本文引用: 1]
WangSY, XuXT, ShresthaN, ZimmermannNE, TangZY, WangZH ( 2017b). Response of spatial vegetation distribution in China to climate changes since the Last Glacial Maximum (LGM) PLOS ONE, 12, e0175742. DOI: 10.1371/journal.pone.0175742. [本文引用: 1]
WangYH, JiangWM, ComesHP, HuFS, QiuYX, FuCX ( 2015). Molecular phylogeography and ecological niche modelling of a widespread herbaceous climber,Tetrastigma hemsleyanum(Vitaceae): Insights into Plio- Pleistocene range dynamics of evergreen forest in subtropical China New Phytologist, 206, 852-867.
XingDL, HaoZQ ( 2011). The principle of maximum entropy and its applications in ecology Biodiversity Science, 19, 295-302. [本文引用: 1]
XuZL, ZhaoCY, FengZD ( 2011). Species potential distribution models and evaluation based on dissimilarity index of variables of Qinghai spruce (Picea crassifolia) in Qilian mountains Journal of Lanzhou University (Nature Sciences), 47, 55-63. [本文引用: 1]
XuZL, ZhaoCY, FengZD ( 2012). Species distribution models to estimate the deforested area of Picea crassifolia in arid region recently protected: Qilian Mts. National Natural Reserve (China) Polish Journal of Ecology, 60, 515-524. [本文引用: 1]
XuZL, ZhaoCY, FengZD, PengHH, WangC (2009). The impact of climate change on potential distribution of species in semi-arid region: A case study of Qinghai spruce (Picea crassifolia) in Qilian Mountain, Gansu Province, China In: 2009 IEEE International Geoscience and Remote Sensing Symposium, Cape Town. 412-415. [本文引用: 1]
YanHF, ZhangCY, WangFY, HuCM, GeXJ, HaoG (2012). Population expanding with the phalanx model and lineages split by environmental heterogeneity: A case study of Primula obconica in subtropical China PLOS ONE, 7, e41315. DOI: 10.1371/journal.pone.0041315. [本文引用: 1]
YeYC, ZhouGS, YinXJ ( 2016). Changes in distribution and productivity of steppe vegetation in Inner Mongolia during 1961 to 2010: Analysis based on MaxEnt model and synthetic model Acta Ecologica Sinica, 36, 4718-4728. [本文引用: 1]
ZhangL, LiuSR, SunPS, WangTL, WangGY, ZhangXD, WangLL ( 2015). Consensus forecasting of species distributions: The effects of niche model performance and niche properties PLOS ONE, 10, e0120056. DOI: 10.1371/ ?journal.?pone.0120056.
ZhangMG, SlikJF, MaKP ( 2017). Priority areas for the conservation of perennial plants in China Biological Conservation, 210, 56-63. [本文引用: 1]
ZhangMG, ZhouZK, ChenWY, CannonCH, RaesN, SlikJF ( 2014). Major declines of woody plant species ranges under climate change in Yunnan, China Diversity and Distributions, 20, 405-415. [本文引用: 1]
ZhangMG, ZhouZK, ChenWY, SlikJF, CannonCH, RaesN (2012). Using species distribution modeling to improve conservation and land use planning of Yunnan, China Biological Conservation, 153, 257-264. [本文引用: 1]
ZhaoZF, WeiHY, GuoYL, GuW (2016). Potential distribution of Panax ginseng and its predicted responses to climate change Chinese Journal of Applied Ecology, 27, 3607-3615. [本文引用: 1]
ZhongGP ( 2008). Predicting the Potential Distribution of Invasive Alien Weeds in China Master degree dissertation, Southwest University, Chongqing. [本文引用: 1]
ZhouJ, LiQY, XiaoL, JiangJX, YiZL ( 2012). Potential distribution of Miscanthus sinensis and M. floridulus in China Chinese Journal of Plant Ecology, 36, 504-510. [本文引用: 1]
The potential impact of invasive woody oil plants on protected areas in China under future climate conditions 1 2018
... 全球变化一直是国内外科学界研究的重点领域, 我国物种分布模型过去在此方面的应用很多, 将来也应该继续更多关注全球变化的理论研究, 尤其是现状气候下的植物种潜在分布和适生区的模拟(周婧等, 2012; 王丹等, 2017), 以及未来气候变化情景下物种分布范围的变迁及其脆弱性和适应性(刘少军等, 2015; Dai et al., 2018), 当然也涉及过去气候变化对物种分布及其冰期避难所的研究(Li et al., 2012; Yan et al., 2012; Wang et al., 2015).物种分布模型能够直观地提供物种在不同气候时期的分布区大小, 通过对比可得出不同物种对于气候的响应模式(张路, 2015).古生态学研究也正在吸纳更多物种分布模型的应用, 这也是过去全球变化研究的需求(Svenning et al., 2011). ...
Species distribution models: Ecological explanation and prediction across space and time 2 2009
Phylogeographic analysis and environmental niche modeling of widespread shrub Rhododendron simsii in China reveals multiple glacial refugia during the last glacial maximum 1 2012
... 全球变化一直是国内外科学界研究的重点领域, 我国物种分布模型过去在此方面的应用很多, 将来也应该继续更多关注全球变化的理论研究, 尤其是现状气候下的植物种潜在分布和适生区的模拟(周婧等, 2012; 王丹等, 2017), 以及未来气候变化情景下物种分布范围的变迁及其脆弱性和适应性(刘少军等, 2015; Dai et al., 2018), 当然也涉及过去气候变化对物种分布及其冰期避难所的研究(Li et al., 2012; Yan et al., 2012; Wang et al., 2015).物种分布模型能够直观地提供物种在不同气候时期的分布区大小, 通过对比可得出不同物种对于气候的响应模式(张路, 2015).古生态学研究也正在吸纳更多物种分布模型的应用, 这也是过去全球变化研究的需求(Svenning et al., 2011). ...
... 全球变化一直是国内外科学界研究的重点领域, 我国物种分布模型过去在此方面的应用很多, 将来也应该继续更多关注全球变化的理论研究, 尤其是现状气候下的植物种潜在分布和适生区的模拟(周婧等, 2012; 王丹等, 2017), 以及未来气候变化情景下物种分布范围的变迁及其脆弱性和适应性(刘少军等, 2015; Dai et al., 2018), 当然也涉及过去气候变化对物种分布及其冰期避难所的研究(Li et al., 2012; Yan et al., 2012; Wang et al., 2015).物种分布模型能够直观地提供物种在不同气候时期的分布区大小, 通过对比可得出不同物种对于气候的响应模式(张路, 2015).古生态学研究也正在吸纳更多物种分布模型的应用, 这也是过去全球变化研究的需求(Svenning et al., 2011). ...
... 全球变化一直是国内外科学界研究的重点领域, 我国物种分布模型过去在此方面的应用很多, 将来也应该继续更多关注全球变化的理论研究, 尤其是现状气候下的植物种潜在分布和适生区的模拟(周婧等, 2012; 王丹等, 2017), 以及未来气候变化情景下物种分布范围的变迁及其脆弱性和适应性(刘少军等, 2015; Dai et al., 2018), 当然也涉及过去气候变化对物种分布及其冰期避难所的研究(Li et al., 2012; Yan et al., 2012; Wang et al., 2015).物种分布模型能够直观地提供物种在不同气候时期的分布区大小, 通过对比可得出不同物种对于气候的响应模式(张路, 2015).古生态学研究也正在吸纳更多物种分布模型的应用, 这也是过去全球变化研究的需求(Svenning et al., 2011). ...
Applications of species distribution modeling to paleobiology 1 2011
... 全球变化一直是国内外科学界研究的重点领域, 我国物种分布模型过去在此方面的应用很多, 将来也应该继续更多关注全球变化的理论研究, 尤其是现状气候下的植物种潜在分布和适生区的模拟(周婧等, 2012; 王丹等, 2017), 以及未来气候变化情景下物种分布范围的变迁及其脆弱性和适应性(刘少军等, 2015; Dai et al., 2018), 当然也涉及过去气候变化对物种分布及其冰期避难所的研究(Li et al., 2012; Yan et al., 2012; Wang et al., 2015).物种分布模型能够直观地提供物种在不同气候时期的分布区大小, 通过对比可得出不同物种对于气候的响应模式(张路, 2015).古生态学研究也正在吸纳更多物种分布模型的应用, 这也是过去全球变化研究的需求(Svenning et al., 2011). ...
Limited filling of the potential range in European tree species 1 2004
... 目前, 物种分布模型也被用于模拟植物群落和植被类型的地理分布(叶永昌等, 2016; Wan et al., 2017).由于气候变化对植被的影响是长期的, 利用物种分布模型对植被分布的模拟具有一定的时滞性(Svenning & Skov, 2004), 但不可否认它仍然是研究植被与气候关系的有效工具.如果能够突破其仅局限于模拟植物的主要功能类型, 而是如同动态植被模型那样, 更多地模拟各类植被类型, 那么物种分布模型在模拟植被地理分布研究中将会发挥更大的作用(Wang et al., 2017b).因此, 虽然现有的物种分布模型在生态学与生理学机理上较传统的植被模型有所欠缺(倪健, 2002), 但如同上文所言, 如果能够开发基于生态过程的而非基于统计学信息的机理性物种分布模型, 将它们同时应用于物种和生态系统的模拟, 应该是将来的一个重要发展方向. ...
Planning the priority protected areas of endangered orchid species in northeastern China 1 2014
... 生物多样性保护也是当前世界性环境问题, 物种分布模型在此可发挥其从理论到实践, 直至政策建议的一系列作用.模拟珍稀濒危物种的现代适应性及未来变化趋势, 评估生物多样性现状, 筛选物种迁地保护的生境, 从而基于模拟结果而推动一个地区自然保护政策的制订或调整, 比如建立新的自然保护区, 或扩展现有的自然保护区, 以更加有效地保护当地物种(马松梅等, 2010; Wan et al., 2014).珍稀濒危物种和经济植物的数据少, 无论是对于物种现有的地理分布还是对其潜在分布区的预测都会造成一定的阻碍.而物种分布模型可以利用较少的已知物种分布数据和现有的环境数据, 模拟物种可能的分布地区, 为珍稀濒危植物和经济植物的资源分布调查、就地或迁地保护、人工栽培提供理论依据. ...
Spatial conservation prioritization for dominant tree species of Chinese forest communities under climate change 1 2017
... 目前, 物种分布模型也被用于模拟植物群落和植被类型的地理分布(叶永昌等, 2016; Wan et al., 2017).由于气候变化对植被的影响是长期的, 利用物种分布模型对植被分布的模拟具有一定的时滞性(Svenning & Skov, 2004), 但不可否认它仍然是研究植被与气候关系的有效工具.如果能够突破其仅局限于模拟植物的主要功能类型, 而是如同动态植被模型那样, 更多地模拟各类植被类型, 那么物种分布模型在模拟植被地理分布研究中将会发挥更大的作用(Wang et al., 2017b).因此, 虽然现有的物种分布模型在生态学与生理学机理上较传统的植被模型有所欠缺(倪健, 2002), 但如同上文所言, 如果能够开发基于生态过程的而非基于统计学信息的机理性物种分布模型, 将它们同时应用于物种和生态系统的模拟, 应该是将来的一个重要发展方向. ...
A new strategy for the prevention and control of Eupatorium adenophorum under climate change in China 1 2017
... 在生物多样性研究中, 外来物种也是一个热门话题, 同样, 物种分布模型也被用来评估物种入侵的环境风险, 从而达到有效管理入侵物种的目标(王瑞和万方浩, 2016; Wang et al., 2017a).通过物种分布模型对外来入侵植物的预测, 探讨其扩散化趋势, 可以更好地协助控制外来物种对本地物种生存及自然环境的破坏, 为外来入侵植物的预防和治理提供决策依据. ...
... 全球变化一直是国内外科学界研究的重点领域, 我国物种分布模型过去在此方面的应用很多, 将来也应该继续更多关注全球变化的理论研究, 尤其是现状气候下的植物种潜在分布和适生区的模拟(周婧等, 2012; 王丹等, 2017), 以及未来气候变化情景下物种分布范围的变迁及其脆弱性和适应性(刘少军等, 2015; Dai et al., 2018), 当然也涉及过去气候变化对物种分布及其冰期避难所的研究(Li et al., 2012; Yan et al., 2012; Wang et al., 2015).物种分布模型能够直观地提供物种在不同气候时期的分布区大小, 通过对比可得出不同物种对于气候的响应模式(张路, 2015).古生态学研究也正在吸纳更多物种分布模型的应用, 这也是过去全球变化研究的需求(Svenning et al., 2011). ...
... 全球变化一直是国内外科学界研究的重点领域, 我国物种分布模型过去在此方面的应用很多, 将来也应该继续更多关注全球变化的理论研究, 尤其是现状气候下的植物种潜在分布和适生区的模拟(周婧等, 2012; 王丹等, 2017), 以及未来气候变化情景下物种分布范围的变迁及其脆弱性和适应性(刘少军等, 2015; Dai et al., 2018), 当然也涉及过去气候变化对物种分布及其冰期避难所的研究(Li et al., 2012; Yan et al., 2012; Wang et al., 2015).物种分布模型能够直观地提供物种在不同气候时期的分布区大小, 通过对比可得出不同物种对于气候的响应模式(张路, 2015).古生态学研究也正在吸纳更多物种分布模型的应用, 这也是过去全球变化研究的需求(Svenning et al., 2011). ...
... 全球变化一直是国内外科学界研究的重点领域, 我国物种分布模型过去在此方面的应用很多, 将来也应该继续更多关注全球变化的理论研究, 尤其是现状气候下的植物种潜在分布和适生区的模拟(周婧等, 2012; 王丹等, 2017), 以及未来气候变化情景下物种分布范围的变迁及其脆弱性和适应性(刘少军等, 2015; Dai et al., 2018), 当然也涉及过去气候变化对物种分布及其冰期避难所的研究(Li et al., 2012; Yan et al., 2012; Wang et al., 2015).物种分布模型能够直观地提供物种在不同气候时期的分布区大小, 通过对比可得出不同物种对于气候的响应模式(张路, 2015).古生态学研究也正在吸纳更多物种分布模型的应用, 这也是过去全球变化研究的需求(Svenning et al., 2011). ...
... 全球变化一直是国内外科学界研究的重点领域, 我国物种分布模型过去在此方面的应用很多, 将来也应该继续更多关注全球变化的理论研究, 尤其是现状气候下的植物种潜在分布和适生区的模拟(周婧等, 2012; 王丹等, 2017), 以及未来气候变化情景下物种分布范围的变迁及其脆弱性和适应性(刘少军等, 2015; Dai et al., 2018), 当然也涉及过去气候变化对物种分布及其冰期避难所的研究(Li et al., 2012; Yan et al., 2012; Wang et al., 2015).物种分布模型能够直观地提供物种在不同气候时期的分布区大小, 通过对比可得出不同物种对于气候的响应模式(张路, 2015).古生态学研究也正在吸纳更多物种分布模型的应用, 这也是过去全球变化研究的需求(Svenning et al., 2011). ...
入侵植物银毛龙葵在中国的适生区预测与早期监测预警 1 2016
... 在生物多样性研究中, 外来物种也是一个热门话题, 同样, 物种分布模型也被用来评估物种入侵的环境风险, 从而达到有效管理入侵物种的目标(王瑞和万方浩, 2016; Wang et al., 2017a).通过物种分布模型对外来入侵植物的预测, 探讨其扩散化趋势, 可以更好地协助控制外来物种对本地物种生存及自然环境的破坏, 为外来入侵植物的预防和治理提供决策依据. ...
入侵植物银毛龙葵在中国的适生区预测与早期监测预警 1 2016
... 在生物多样性研究中, 外来物种也是一个热门话题, 同样, 物种分布模型也被用来评估物种入侵的环境风险, 从而达到有效管理入侵物种的目标(王瑞和万方浩, 2016; Wang et al., 2017a).通过物种分布模型对外来入侵植物的预测, 探讨其扩散化趋势, 可以更好地协助控制外来物种对本地物种生存及自然环境的破坏, 为外来入侵植物的预防和治理提供决策依据. ...
Response of spatial vegetation distribution in China to climate changes since the Last Glacial Maximum (LGM) 1 2017
... 目前, 物种分布模型也被用于模拟植物群落和植被类型的地理分布(叶永昌等, 2016; Wan et al., 2017).由于气候变化对植被的影响是长期的, 利用物种分布模型对植被分布的模拟具有一定的时滞性(Svenning & Skov, 2004), 但不可否认它仍然是研究植被与气候关系的有效工具.如果能够突破其仅局限于模拟植物的主要功能类型, 而是如同动态植被模型那样, 更多地模拟各类植被类型, 那么物种分布模型在模拟植被地理分布研究中将会发挥更大的作用(Wang et al., 2017b).因此, 虽然现有的物种分布模型在生态学与生理学机理上较传统的植被模型有所欠缺(倪健, 2002), 但如同上文所言, 如果能够开发基于生态过程的而非基于统计学信息的机理性物种分布模型, 将它们同时应用于物种和生态系统的模拟, 应该是将来的一个重要发展方向. ...
Molecular phylogeography and ecological niche modelling of a widespread herbaceous climber,Tetrastigma hemsleyanum(Vitaceae): Insights into Plio- Pleistocene range dynamics of evergreen forest in subtropical China 2015
Species distribution models to estimate the deforested area of Picea crassifolia in arid region recently protected: Qilian Mts. National Natural Reserve (China) 1 2012
The impact of climate change on potential distribution of species in semi-arid region: A case study of Qinghai spruce (Picea crassifolia) in Qilian Mountain, Gansu Province, China 1 2009
Population expanding with the phalanx model and lineages split by environmental heterogeneity: A case study of Primula obconica in subtropical China 1 2012
... 全球变化一直是国内外科学界研究的重点领域, 我国物种分布模型过去在此方面的应用很多, 将来也应该继续更多关注全球变化的理论研究, 尤其是现状气候下的植物种潜在分布和适生区的模拟(周婧等, 2012; 王丹等, 2017), 以及未来气候变化情景下物种分布范围的变迁及其脆弱性和适应性(刘少军等, 2015; Dai et al., 2018), 当然也涉及过去气候变化对物种分布及其冰期避难所的研究(Li et al., 2012; Yan et al., 2012; Wang et al., 2015).物种分布模型能够直观地提供物种在不同气候时期的分布区大小, 通过对比可得出不同物种对于气候的响应模式(张路, 2015).古生态学研究也正在吸纳更多物种分布模型的应用, 这也是过去全球变化研究的需求(Svenning et al., 2011). ...
... 全球变化一直是国内外科学界研究的重点领域, 我国物种分布模型过去在此方面的应用很多, 将来也应该继续更多关注全球变化的理论研究, 尤其是现状气候下的植物种潜在分布和适生区的模拟(周婧等, 2012; 王丹等, 2017), 以及未来气候变化情景下物种分布范围的变迁及其脆弱性和适应性(刘少军等, 2015; Dai et al., 2018), 当然也涉及过去气候变化对物种分布及其冰期避难所的研究(Li et al., 2012; Yan et al., 2012; Wang et al., 2015).物种分布模型能够直观地提供物种在不同气候时期的分布区大小, 通过对比可得出不同物种对于气候的响应模式(张路, 2015).古生态学研究也正在吸纳更多物种分布模型的应用, 这也是过去全球变化研究的需求(Svenning et al., 2011). ...
... 全球变化一直是国内外科学界研究的重点领域, 我国物种分布模型过去在此方面的应用很多, 将来也应该继续更多关注全球变化的理论研究, 尤其是现状气候下的植物种潜在分布和适生区的模拟(周婧等, 2012; 王丹等, 2017), 以及未来气候变化情景下物种分布范围的变迁及其脆弱性和适应性(刘少军等, 2015; Dai et al., 2018), 当然也涉及过去气候变化对物种分布及其冰期避难所的研究(Li et al., 2012; Yan et al., 2012; Wang et al., 2015).物种分布模型能够直观地提供物种在不同气候时期的分布区大小, 通过对比可得出不同物种对于气候的响应模式(张路, 2015).古生态学研究也正在吸纳更多物种分布模型的应用, 这也是过去全球变化研究的需求(Svenning et al., 2011). ...
Consensus forecasting of species distributions: The effects of niche model performance and niche properties 2015
Priority areas for the conservation of perennial plants in China 1 2017
... 全球变化一直是国内外科学界研究的重点领域, 我国物种分布模型过去在此方面的应用很多, 将来也应该继续更多关注全球变化的理论研究, 尤其是现状气候下的植物种潜在分布和适生区的模拟(周婧等, 2012; 王丹等, 2017), 以及未来气候变化情景下物种分布范围的变迁及其脆弱性和适应性(刘少军等, 2015; Dai et al., 2018), 当然也涉及过去气候变化对物种分布及其冰期避难所的研究(Li et al., 2012; Yan et al., 2012; Wang et al., 2015).物种分布模型能够直观地提供物种在不同气候时期的分布区大小, 通过对比可得出不同物种对于气候的响应模式(张路, 2015).古生态学研究也正在吸纳更多物种分布模型的应用, 这也是过去全球变化研究的需求(Svenning et al., 2011). ...
芒和五节芒在中国的潜在分布 1 2012
... 全球变化一直是国内外科学界研究的重点领域, 我国物种分布模型过去在此方面的应用很多, 将来也应该继续更多关注全球变化的理论研究, 尤其是现状气候下的植物种潜在分布和适生区的模拟(周婧等, 2012; 王丹等, 2017), 以及未来气候变化情景下物种分布范围的变迁及其脆弱性和适应性(刘少军等, 2015; Dai et al., 2018), 当然也涉及过去气候变化对物种分布及其冰期避难所的研究(Li et al., 2012; Yan et al., 2012; Wang et al., 2015).物种分布模型能够直观地提供物种在不同气候时期的分布区大小, 通过对比可得出不同物种对于气候的响应模式(张路, 2015).古生态学研究也正在吸纳更多物种分布模型的应用, 这也是过去全球变化研究的需求(Svenning et al., 2011). ...
 ,1, 袁泉1, 倪健
,1, 袁泉1, 倪健 ,1,2,*
,1,2,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT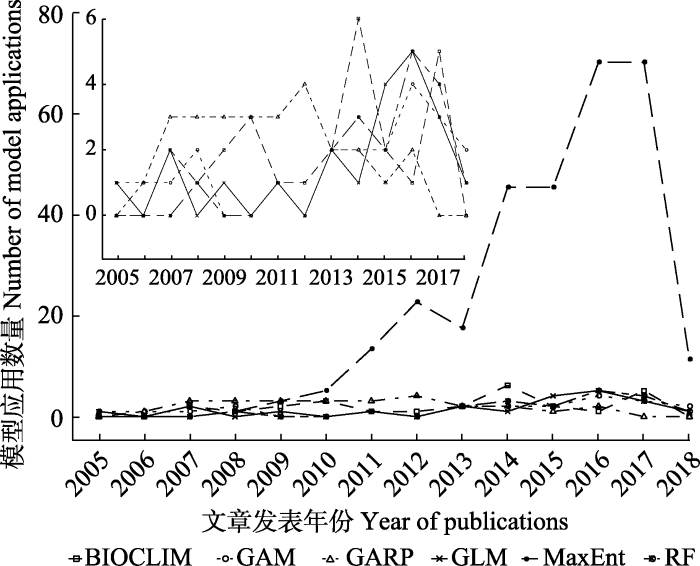 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}