Advances in the study of population genetic diversity at plant species’ margins
Xin-Xin ZHANG,, Xi WANG, Ying HU, Wei ZHOU, Xiao-Yang CHEN, Xin-Sheng HU,,*Guangdong Key Laboratory for Innovative Development and Utilization of Forest Plant Germplasm, College of Forestry and Landscape Architecture, South China Agricultural University, Guangzhou 510642, China
Abstract Marginal populations are those at the geographical edge of a species’ distribution. Appropriate evaluation of genetic diversity in marginal populations is of crucial significance for understanding the impacts of climate changes on species expansion or contraction in the post Quaternary glaciations, conservation of genetic resources and exploitation, and peripatric speciation. Here, we discuss the evolutionary mechanisms for maintaining genetic diversity in marginal populations, analyze the role of plant mating system in shaping a plant species’ range and the genetic diversity in marginal populations, assess the difference or similarity in genetic diversity between central and marginal populations and the underlying ecological and evolutionary processes, and discuss the species genetic diversity correlation (SGDC) and the theory underlying such correlations. We proposed that future research includes the use of genome-wide sequences or transcriptome data to study the adaptive differential between leading- and rear-edge populations or between central and marginal populations and the molecular mechanisms of the interactions between the genetic diversity in marginal populations and the species diversity in the resident community of a focal species. This may help to understand the adaptability of marginal populations to local habitats and the ecological and evolutionary processes for SGDC at species’ edges. Keywords:marginal population;central population;genetic diversity;mating system;species diversity
PDF (1059KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 张新新, 王茜, 胡颖, 周玮, 陈晓阳, 胡新生. 植物边缘种群遗传多样性研究进展. 植物生态学报, 2019, 43(5): 383-395. DOI: 10.17521/cjpe.2018.0252 ZHANG Xin-Xin, WANG Xi, HU Ying, ZHOU Wei, CHEN Xiao-Yang, HU Xin-Sheng. Advances in the study of population genetic diversity at plant species’ margins. Chinese Journal of Plant Ecology, 2019, 43(5): 383-395. DOI: 10.17521/cjpe.2018.0252
Table 1 表1 表1物种分布理论模型 Table 1Theoretical models of a species’ distribution
模型 Model
主要观点 Main point
参考文献 Reference
随机生态位模型或断棒模型 Stochastic niche model or broken stick model
每一物种生态位超体积的占有和分布不受其他物种影响。 The niche occupation and its size distribution of each species are random and independent of the niche sizes of other species.
物种占有的生态位是随机分布的并受大量因素综合影响, 并不优待某些种。 The niche size of a species is random and determined by the joint effects of a large number of factors, and no selective advantage is present among species.
第一位优势种优先占领生态位空间大部, 第二位占领其余下的大部, 以此类推, 末位只占留下的极少空间。 The first dominant species occupies the largest niche space, followed by the species that occupies the second largest niche in the remaining space, and so on. The last species occupies the minimum niche.
群落内个体总数固定, 某一物种多度的增加必然伴随其他物种的减少; 所有个体出生率、死亡率相同。 The community size is fixed, and a decrease of one species’ abundance is equally compensated by other species. All individuals in the community have the same birth and death rates.
自然选择-基因流机制 Mechanism of natural selection-gene flow
基因由中心向边缘种群的迁移与边缘种群的自然选择作用持衡。 Effects of gene flow from the central to marginal population are in balance with the effects of natural selection in the marginal population.
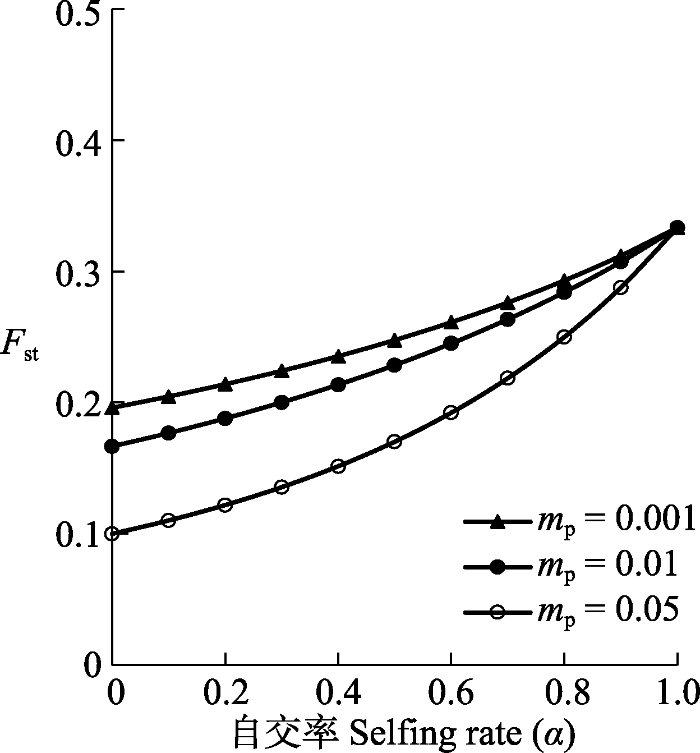
Fig. 1Effects of selfing on population genetic structure. The population differentiation coefficient Fst was calculated according to the equation $\frac{1}{F_{st}}=1+4N_{e}(1-\frac{1}{2}\alpha)(m_{s}+\frac{1-\alpha}{2}m_{p})$, and the parameters used were seed flow ms = 0.02, effective population size Ne = 50, and pollen flow mp were 0.001, 0.01 and 0.05.
Table 3 表3 表3边缘与中心种群遗传多样性比较及可能的生态或进化过程 Table 3Comparison of genetic diversity between central and marginal populations of various plant species and the potential ecological or evolutionary processes responsible for the observed differences
植物种 Species
边缘种群与中心种群遗传多样性 Genetic diversity in marginal vs. central populations
生态或进化机制 Ecological or evolutionary mechanisms
参考文献 Reference
樱桃 Cerasus pseudocerasus
边缘种群低于中心种群 marginal populations < central populations
AvolioML, SmithMD (2013). Correlations between genetic and species diversity: Effects of resource quantity and heterogeneity Journal of Vegetation Science, 24, 1185-1194. [本文引用: 1]
BakerHG (1955). Self-compatibility and establishment after “long-distance” dispersal Evolution, 9, 347-349. [本文引用: 1]
BakerHG (1967). Support for Baker’s Law—As a rule Evolution, 21, 853-856. [本文引用: 1]
BakkerEG, StahlEA, ToomajianC, NordborgM, KreitmanM, BergelsonJ (2006). Distribution of genetic variation within and among local populations of Arabidopsis thaliana over its species range Molecular Ecology, 15, 1405-1418. [本文引用: 1]
BarrettSCH (1995). Mating-system evolution in flowering plants: Micro- and macroevolutionary approaches Acta Botanica Neerlandica, 44, 385-402. [本文引用: 2]
BarrettSCH (2014). Evolution of Mating Systems: Outcrossing Versus Selfing. Princeton University Press, Princeton. 356-362. [本文引用: 1]
BartonN (2001). Adaptation at the edge of a species’ range. In: Silvertown J, Antonovics J eds. Integrating Ecology and Evolution in a Spatial Context Blackwell Science,London. 365-392. [本文引用: 4]
BlumMJ, BagleyMJ, WaltersDM, DanielFB, ChaloudDJ, CadeBS (2012). Genetic diversity and species diversity of stream fishes covary across a land-use gradient Oecologia, 168, 83-95. [本文引用: 1]
BuschJW (2005). The evolution of self-compatibility in geographically marginal populations of Leavenworthia alabamica(Brassicaceae) American Journal of Botany, 92, 1503-1512. [本文引用: 2]
CallahanCM, RoweCA, RyelRJ, ShawJD, MadritchMD, MockKE (2013). Continental-scale assessment of genetic diversity and population structure in quaking aspen (Populus tremuloides) Journal of Biogeography, 40, 1780-1791. [本文引用: 1]
CaseTJ, TaperML (2000). Interspecific competition, environmental gradients, gene flow, and the coevolution of species’ borders The American Naturalist, 155, 583-605. [本文引用: 1]
CharlesworthD (2006). Evolution of plant breeding systems Current Biology, 16, 726-735. [本文引用: 1]
ChenDM, KangHZ, LiuCJ (2011). An overview on the potential quaternary glacial refugia of plants in China mainland Bulletin of Botanical Research, 31, 623-632.
ChhatreVE, RajoraOP (2014). Genetic divergence and signatures of natural selection in marginal populations of a keystone, long-lived conifer, eastern white pine (Pinus strobus) from northern Ontario PLOS ONE, 9, e97291. DOI: 10.1371/journal.pone.0097291. [本文引用: 1]
ChuCJ, MaestreFT, XiaoS, WeinerJ, WangYS, DuanZH, WangG (2008). Balance between facilitation and resource competition determines biomass-density relationships in plant populations Ecology Letters, 11, 1189-1197. [本文引用: 1]
CoyneJA, OrrHA (2004). Speciation Sinauer Associates, Sunderland, USA. [本文引用: 1]
DartSR, SamisKE, AustenE, EcketCG (2012). Broad geographic covariation between floral traits and the mating system in Camissoniopsis cheiranthifolia(Onagraceae): Multiple stable mixed mating systems across the species’ range . Annals of Botany, 109, 599-611. [本文引用: 1]
DarwinC (1859). On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life John Murray, London. [本文引用: 1]
de LafontaineG, DucoussoA, LefevreS, MagnanouE, PetitRJ (2013). Stronger spatial genetic structure in recolonized areas than in refugia in the European beech Molecular Ecology, 22, 4397-4412. [本文引用: 1]
DurkaW (1999). Genetic diversity in peripheral and subcentral populations of Corrigiola litoralis L.( lllecebraceae) Heredity, 83, 476-484. [本文引用: 1]
EckertCG, KaliszS, GeberMA, SargentR, ElleE, CheptouPO, GoodwillieC, JohnstonMO, KellyJK, MoellerDA, PorcherE, ReeRH, Vallejo-MarinM, WinnAA (2010). Plant mating systems in a changing world Trends in Ecology & Evolution, 25, 35-43. [本文引用: 1]
EckertCG, SamisKE, LougheedSC (2008). Genetic variation across species’ geographical ranges: The central-marginal hypothesis and beyond Molecular Ecology, 17, 1170-1188. [本文引用: 1]
FengFJ, HanSJ, WangHM (2006). Genetic diversity and genetic differentiation of natural Pinus koraiensis population Journal of Forestry Research, 17, 21-24. [本文引用: 2]
FisherRA (1930). The Genetical Theory of Natural Selection. Oxford University Press, Oxford. [本文引用: 1]
GaoLZ, GaoCW (2016). Lowered diversity and increased inbreeding depression within peripheral populations of wild rice Oryza rufipogon PLOS ONE, 11, e0150468. DOI: 10.1371/journal.pone.0150468. [本文引用: 1]
GastonK (2003). The Structure and Dynamics of Geographic Ranges. Oxford University Press, Oxford. 100-101. [本文引用: 2]
GriffinPC, WilliY (2014). Evolutionary shifts to self- fertilisation restricted to geographic margins in North American Arabidopsis lyrata Ecology Letters, 17, 484-490. [本文引用: 2]
GrossenbacherD, Briscoe RunquistRD, GoldbergEE, BrandvainY (2015). Geographic range size is predicted by plant mating system Ecology Letters, 18, 706-713. [本文引用: 1]
GrossenbacherD, Briscoe RunquistRD, GoldbergEE, BrandvainY (2016). No association between plant mating system and geographic range overlap American Journal of Botany, 103, 110-117. [本文引用: 1]
GuggerPF, González-RodríguezA, Rodríguez-CorreaH, SugitaS, Cavender-BaresJ (2011). Southward Pleistocene migration of Douglas-fir into Mexico: Phylogeography, ecological niche modeling, and conservation of “rear edge” populations New Phytologist, 189, 1185-1199. [本文引用: 2]
GuoQ (2012). Incorporating latitudinal and central-marginal trends in assessing genetic variation across species ranges Molecular Ecology, 21, 5396-5403. [本文引用: 1]
HaldaneJBS (1956). The relation between density regulation and natural selection Proceedings of the Royal Society of London: Series B, Biological Sciences, 145, 306-308. [本文引用: 4]
HampeA, PetitRJ (2005). Conserving biodiversity under climate change: The rear edge matters Ecology Letters, 8, 461-467. [本文引用: 4]
HaoL, ZhangL, ZhangGS, WangY, HanSL, BaiYR (2017). Genetic diversity and population genetic structure of Salix psammophila Acta Botanica Boreali-Occidentalia Sinica, 37, 1507-1516. [本文引用: 2]
HavrdovaA, DoudaJ, KrakK, VitP, HadincovaV, ZakravskyP, MandakB (2015). Higher genetic diversity in recolonized areas than in refugia of Alnus glutinosa triggered by continent-wide lineage admixture Molecular Ecology, 24, 4759-4777. [本文引用: 1]
HeT, LamontBB, KraussSL, EnrightNJ, MillerBP (2008). Covariation between intraspecific genetic diversity and species diversity within a plant functional group Journal of Ecology, 96, 956-961. [本文引用: 1]
HiraoAS, WatanabeM, TsuyuzakiS, ShimonoA, LiX, MasuzawaT, WadaN (2017). Genetic diversity within populations of an arctic-alpine species declines with decreasing latitude across the Northern Hemisphere Journal of Biogeography, 44, 2740-2751. [本文引用: 2]
HirschH, WagnerV, DanihelkaJ, RuprechtE, Sánchez-GómezP, SeifertM, HensenI (2015). High genetic diversity declines towards the geographic range periphery of Adonis vernalis, a Eurasian dry grassland plant Plant Biology, 17, 1233-1241. [本文引用: 1]
HuXS (2011). Mating system and the critical migration rate for swamping selection Genetics Research, 93, 233-254. [本文引用: 2]
HuXS, EnnosRA (1999). Scoring the mating systems of natural populations of three Larix taxa in China: L. gmelinii(Rupr.) Rupr., L. olgensis Henry and L. principis-?rupprechtii Mayr Scientia Silvae Sinicae, 35(1), 21-31. [本文引用: 1]
HuXS, HeFL (2005). Background selection and population differentiation Journal of Theoretical Biology, 235, 207-219. [本文引用: 1]
HuXS, HeFL, HubbellSP (2006). Neutral theory in macroecology and population genetics Oikos, 113, 548-556. [本文引用: 2]
HuXS, YehFC, WangZQ (2011). Structural genomics: Correlation blocks, population structure, and genome architecture Current Genomics, 12, 55-70.
HuXS, ZengW, LiBL (2003). Impacts of one-way gene flow on genetic variance components in a natural population Silvae Genetica, 52, 18-24. [本文引用: 1]
HuXS, ZhangXX, ZhouW, HuY, WangX, ChenXY (2019). Mating system shifts a species’ range Evolution, 73, 158-174. [本文引用: 1]
HubbellSP (2001). The Unified Neutral Theory of Biodiversity and Biogeography. Princeton University Press, Princeton. 1772. [本文引用: 2]
JohnsonMTJ, LajeunesseMJ, AgrawalAA (2006). Additive and interactive effects of plant genotypic diversity on arthropod communities and plant fitness Ecology Letters, 9, 24-34. [本文引用: 1]
JohnstonMO, PorcherE, CheptouPO, EckertCG, ElleE, GeberMA, KaliszS, KellyJK, MoellerDA, Vallejo-MarinM, WinnAA (2009). Correlations among fertility components can maintain mixed mating in plants The American Naturalist, 173, 1-11. [本文引用: 1]
JumpAS, WoodwardEI, BurkeT (2003). Cirsium species show disparity in patterns of genetic variation at their range-edge, despite similar patterns of reproduction and isolation New Phytologist, 160, 359-370. [本文引用: 1]
KaweckiTJ (2008). Adaptation to marginal habitats Annual Review of Ecology, Evolution, and Systematics, 39, 321-342. [本文引用: 1]
KimminsJP (2004). Forestry Ecology: A Foundation for Sustainable Forest Management and Environmental Ethics in Forestry. 3rd edn Pearson Education, Upper Saddle River, New Jersey. [本文引用: 2]
KirkpatrickM, BartonNH (1997). Evolution of a species’ range The American Naturalist, 150, 1-23. [本文引用: 2]
KirkpatrickM, RavignéV (2002). Speciation by natural and sexual selection: Models and experiments The American Naturalist, 159, 22-35. [本文引用: 1]
KropfM (2012). Genetic variation, biogeographical history, and conservation of Anthyllis montana L. ssp. jacquinii(Kern.) Hayek (Fabaceae) at its northern distribution limit International Journal of Plant Sciences, 173, 789-801.
LaikreL, AllendorfFW, AronerLC, AronerLC, BakerCS, GregovichDP, HansenMM, JacksonJA, KendallKC, McKelveyK, NeelMC, OlivieriI, RymanN, SchwartzMK, BullRS, StetzJB, TallmonDA, TaylorBL, VojtaCD, WallerDM, WaplesRS (2010). Neglect of genetic diversity in implementation of the convention on biological diversity Conservation Biology, 24, 86-88. [本文引用: 1]
LamyT, JarneP, LarocheF, PointierJP, HuthG, SegardA, DavidP (2013). Variation in habitat connectivity generates positive correlations between species and genetic diversity in a metacommunity Molecular Ecology, 2, 4445-4456. [本文引用: 1]
LarocheF, JarneP, LamyT, DavidP, MassolF (2015). A neutral theory for interpreting correlations between species and genetic diversity in communities The American Naturalist, 185, 59-69. [本文引用: 1]
LázaronogalA, MatesanzS, GarcíafernándezA, TravesetA, ValladaresF (2017). Population size, center-periphery, and seed dispersers’ effects on the genetic diversity and population structure of the Mediterranean relict shrub Cneorum tricoccon Ecology and Evolution, 7, 7231-7242. [本文引用: 1]
LepaisO, MullerSD, SamiaBS-L, BenslamaM, RhaziL, Belouahem-AbedD, Daoud-BouttourA, GammarAM, Ghrabi-gammarZ, BaclesCFE (2013). High genetic diversity and distinctiveness of rear-edge climate relicts maintained by ancient tetraploidisation for Alnus glutinosa PLOS ONE, 8, e75029. DOI: 10.1371/journal.pone.0075029. [本文引用: 1]
LevinDA (2012). Mating system shifts on the trailing edge Annals of Botany, 109, 613-620. [本文引用: 1]
LiuJ, JiangJM, ZouJ, XuJL, ShenH, DiaoSF (2013). Genetic diversity of central and marginal populations of Toona ciliata var. pubescens, an endangered tree species endemic to China Chinese Journal of Plant Ecology, 37, 52-60. [本文引用: 2]
MacArthurRH (1957). On the relative abundance of bird species Proceedings of the National Academy of Sciences of the United States of America, 43, 283-295. [本文引用: 1]
MandakB, BimovaKA, PlackovaI, MahelkaV, ChrtekJ (2005). Loss of genetic variation in geographically marginal populations of Atriplex tatatica(Chenopodiaceae) Annals of Botany, 96, 901-902. [本文引用: 1]
MatosPG, ClarissePS, Bodanese-ZanettiniMH, ChristianL, FernandaB (2015). Limited pollen flow and high selfing rates toward geographic range limit in an Atlantic forest bromeliad Flora, 211, 1-10. [本文引用: 1]
MichalskiSG, DurkaW (2007). High selfing and high inbreeding depression in peripheral populations of Juncus atratus Molecular Ecology, 16, 4715-4727. [本文引用: 1]
MoellerDA, Briscoe RunquistRD, MoeAM, GeberMA, GoodwillieC, CheptouP, EckertCG, ElleE, JohnstonMO, KaliszS, ReeRH, SargentRD, Vallejo-MarinM, WinnAA (2017). Global biogeography of mating system variation in seed plants Ecology Letters, 20, 375-384. [本文引用: 1]
MunozF, ViolleC, CheptouPO (2016). CSR ecological strategies and plant mating systems: Outcrossing increases with competitiveness but stress-tolerance is related to mixed mating Oikos, 125, 1296-1303. [本文引用: 1]
OngeKRST, KallmanT, SlotteT, LascouxM, PalmeAE (2011). Contrasting demographic history and population structure in Capsella rubella and Capsella grandiflora, two closely related species with different mating systems Molecular Ecology, 20, 3306-3320. [本文引用: 1]
OttoSP, MarksJC (1996). Mating systems and the evolutionary transition between haploidy and diploidy Biological Journal of the Linnean Society, 57, 197-218. [本文引用: 1]
PannelJR (2015). Evolution of the mating system in colonizing plants Molecular Ecology, 24, 2018-2037. [本文引用: 1]
PellissierL, EidesenPB, EhrichD, DescombesP, SchonswetterP, TribschA, WestergaardKB, AlvarezN, GuisanA, ZimmermannNE, NormandS, VittozP, LuotoM, DamgaardC, BrochmannC, WiszMS, AlsosIG (2016). Past climate-driven range shifts and population genetic diversity in arctic plants Journal of Biogeography, 43, 461-470. [本文引用: 1]
PerssonH, WideB, AnderssonS, SvenssonL (2004). Allozyme diversity and genetic structure of marginal and central populations of Corylus avellana L.(Betulaceae) in Europe Plant Systematics and Evolution, 244, 157-179. [本文引用: 1]
PetanidouT, GodfreeRC, SongDS, KantsaA, DupontYL, WaserNM (2011). Self-compatibility and plant invasiveness: Comparing species in native and invasive ranges Perspectives in Plant Ecology, Evolution and Systematics, 14, 3-12. [本文引用: 2]
PettengillJB, Briscoe RunquistRD, MoellerDA (2016). Mating system divergence affects the distribution of sequence diversity within and among populations of recently diverged subspecies of Clarkia xantiana(Onagraceae) American Journal of Botany, 103, 99-109. [本文引用: 1]
PirononS, PapugaG, VillellasJ, AngertAL, GarciaMB, ThompsonJD (2017). Geographic variation in genetic and demographic performance: New insights from an old biogeographical paradigm Biological Reviews, 92, 1877-1909. [本文引用: 1]
PolechovaJ, BartonNH (2015). Limits to adaptation along environmental gradients Proceedings of the National Academy of Sciences of the United States of America, 112, 6401-6406. [本文引用: 1]
PrestonFW (1948). The commonness and rarity of species Ecology, 29, 254-283. [本文引用: 1]
ProvanJ (2013). The effects of past, present and future climate change on range-wide genetic diversity in northern North Atlantic marine species Frontiers of Biogeography, 5, 60-66. [本文引用: 1]
ProvanJ, MaggsCA (2012). Unique genetic variation at a species’ rear edge is under threat from global climate change Proceedings: Biological Sciences, 279, 39-47. [本文引用: 1]
Prus-GlowackiW, UrbaniakL, BujasE, CurtuAL (2012). Genetic variation of isolated and peripheral populations of Pinus sylvestris(L.) from glacial refugia Flora, 207, 150-158. [本文引用: 2]
Pu?ca?M, TaberletP, CholerP (2008). No positive correlation between species and genetic diversity in European alpine grasslands dominated by Carex curvula Diversity and Distributions, 14, 852-861. [本文引用: 1]
RazanajatovoM, MaurelN, DawsonW, EsslF, KreftH, PerglJ, PysekP, WeigeltP, WinterM, KleunenMV (2016). Plants capable of selfing are more likely to become naturalized Nature Communications, 30, 1511-1520. [本文引用: 1]
RestouxG, SilvaDE, SagnardF, TorreF, KleinE, FadyB (2008). Life at the margin: The mating system of Mediterranean conifers Web Ecology, 8, 94-102. [本文引用: 2]
SagarinRD, GainesSD, GaylordB (2006). Moving beyond assumptions to understand abundance distributions across the ranges of species Trends in Ecology & Evolution, 21, 524-530. [本文引用: 1]
ScalfiM, PiottiA, RossiM, PiovaniP (2009). Genetic variability of Italian southern Scots pine (Pinus sylvestris L.) populations: The rear edge of the range European Journal of Forest Research, 128, 377-386. [本文引用: 1]
SextonJP, MclntyrePJ, AngertAL, RiceKJ (2009). Evolution and ecology of species range limits Annual Review of Ecology, Evolution, and Systematics, 40, 415-436. [本文引用: 2]
SextonJP, StraussSY, RiceKJ (2011). Gene flow increases fitness at the warm edge of a species’ range Proceedings of the National Academy of Sciences of the United States of America, 108, 11704-11709. [本文引用: 1]
StebbinsGL (1957). Self fertilization and population variability in the higher plants The American Naturalist, 91, 337-354. [本文引用: 1]
TaberletP, ZimmermannNE, EnglischT, TribschA, HoldereggerR, AlvarezN, NiklfeldH, ColdeaG, MirekZ, MoilanenA, AhlmerW, MarsanPA, BonaE, BovioM, CholerP, Cie?lakE, ColliL, CristeaV, DalmasJP, FrajmanB, GarraudL, GaudeulM, GiellyL, GutermannW, JoganN, KagaloAA, KorbeckaG, KüpferP, LequetteB, LetzDR, ManelS, MansionG, MarholdK, MartiniF, NegriniR, Ni?oF, PaunO, PellecchiaM, PericoG, Pi?ko?-MirkowaH, ProsserF, Pu?ca?M, RonikierM, ScheuererM, SchneeweissGM, Sch?nswetterP, Schratt-EhrendorferL, SchüpferF, SelvaggiA, SteinmannK, Thiel-EgenterC, van LooM, WinklerM, WohlgemuthT, WraberT, GugerliF, VellendM (2012). Genetic diversity in widespread species is not congruent with species richness in alpine plant communities Ecology Letters, 15, 1439-1448. [本文引用: 1]
TrotterMV, SpencerHG (2013). Models of frequency dependent selection with mutation from parental alleles Genetics, 195, 231-242. [本文引用: 1]
VekemansX, PouxC, GoubetPM, CastricV (2014). The evolution of selfing from outcrossing ancestors in Brassicaceae: What have we learned from variation at the S-locus? Journal of Evolutionary Biology, 27, 1372-1385. [本文引用: 1]
VellendM (2005). Species diversity and genetic diversity: Parallel processes and correlated patterns The American Naturalist, 166, 199-215. [本文引用: 1]
VolisS, OrmanbekovaD, ShulginaI (2016a). Role of selection and gene flow in population differentiation at the edge vs. interior of the species range differing in climatic conditions Molecular Ecology, 25, 1449-1464. [本文引用: 1]
VolisS, OrmanbekovaD, YermekbayevK, SongM, ShulginaI (2016b). The conservation value of peripheral populations and a relationship between quantitative trait and molecular variation Evolutionary Biology, 43, 26-36. [本文引用: 1]
WagnerV, DurkaW, HensenI (2011). Increased genetic differentiation but no reduced genetic diversity in peripheral vs. central populations of a steppe grass American Journal of Botany, 98, 1173-1179. [本文引用: 1]
WagnerV, TreiberJ, DanihelkaJ, ReprechtE, WescheK, HensenI (2012). Declining genetic diversity and increasing genetic isolation toward the range periphery of Stipa pennata, a Eurasian feather grass International Journal of Plant Sciences, 173, 802-811. [本文引用: 1]
WeiXZ, BaoDC, MengHJ, JiangMX (2017). Pattern and drivers of species-genetic diversity correlation in natural forest tree communities across a biodiversity hotspot Journal of Plant Ecology, 11, 761-770. [本文引用: 2]
WeiXZ, SorkVL, MengHJ, JiangMX (2016). Genetic evidence for central-marginal hypothesis in a Cenozoic relict tree species across its distribution in China Journal of Biogeography, 43, 2173-2185. [本文引用: 2]
WhittakerRH (1972). Evolution and measurement of species diversity Taxon, 21, 213-351. [本文引用: 1]
WhittakerRJ, TriantisKA, LadleRJ (2010). A general dynamic theory of oceanic island biogeography: Extending the MacArthur-Wilson theory to accommodate the rise and fall of volcanic islands. In: Losos JB, Ricklefs RE, MacArthur RH eds. The Theory of Island Biogeography Revisited. Princeton University Press, Princeton. 88-115. [本文引用: 1]
WilliY, MaeaettaenenK (2010). Evolutionary dynamics of mating system shifts in Arabidopsis lyrata Journal of Evolutionary Biology, 23, 2123-2131. [本文引用: 1]
WrightS (1969). Evolution and the Genetics of Populations. University Chicago Press, Chicago. 1191-1192. [本文引用: 1]
WuJB, GaoYB, BaoXY, GaoH, JiaMQ, LiJ, ZhaoNX (2010). Genetic diversity of Stipa grandis P. Smirn populations across the species’ range in the Inner Mongolia Plateau of China Biochemical Systematics and Ecology, 38, 471-477. [本文引用: 1]
WuXP, ShenYF, WangHQ (2016). Analysis of genetic diversity and population genetic structure of Medicago archiducis-nolai and Medicago ruthenica populations based on cpDNA trnL-trnF sequences Pratacultural Science, 33, 1136-1146. [本文引用: 2]
XuWM, LiuL, HeTH, CaoM, ShaLQ, HuYH, LiQM, LiJ (2016). Soil properties drive a negative correlation between species diversity and genetic diversity in a tropical seasonal rainforest Scientific Reports, 6, 20652. DOI: 10.1038/srep20652. [本文引用: 1]
YangAH, DickCW, YaoX, HuangH (2016). Impacts of biogeographic history and marginal population genetics on species range limits: A case study of Liriodendron chinense Scientific Reports, 6, 25632. DOI: 10.1038/srep25632. [本文引用: 2]
ZhangDY, JiangXH (2001). Mating system evolution, resource allocation, and genetic diversity in plants Acta Phytoecologia Sinica, 25, 130-143. [本文引用: 1]
... Comparison of genetic diversity between central and marginal populations of various plant species and the potential ecological or evolutionary processes responsible for the observed differences Table 3
植物种 Species
边缘种群与中心种群遗传多样性 Genetic diversity in marginal vs. central populations
生态或进化机制 Ecological or evolutionary mechanisms
参考文献 Reference
樱桃 Cerasus pseudocerasus
边缘种群低于中心种群 marginal populations < central populations
奠基者效应、瓶颈效应 Founder effect, bottleneck effect
Chen et al., 2012
北沙柳 Salix psammophila
边缘种群低于中心种群 marginal populations < central populations
奠基者效应 Founder effect
Hao et al., 2017
花苜蓿、青海苜蓿 Medicago ruthenica, M. archiducis-nicolai
边缘种群低于中心种群 marginal populations < central populations
奠基者效应 Founder effect
Wu et al., 2016
毛红椿 Toona ciliata var. pubescens
边缘种群高于中心种群 marginal populations > central populations
生境破碎化 Habitat fragmentation
Liu et al., 2013
红松 Pinus koraiensis
边缘种群低于中心种群 marginal populations < central populations
奠基者效应、瓶颈效应 Founder effect, bottleneck effect
Feng et al., 2006
领春木 Euptelea pleiospermum
边缘种群低于中心种群 marginal populations < central populations
冰期后扩张、不对称基因流 Postglacial expansion and asymmetric gene flow
... Comparison of genetic diversity between central and marginal populations of various plant species and the potential ecological or evolutionary processes responsible for the observed differences Table 3
植物种 Species
边缘种群与中心种群遗传多样性 Genetic diversity in marginal vs. central populations
生态或进化机制 Ecological or evolutionary mechanisms
参考文献 Reference
樱桃 Cerasus pseudocerasus
边缘种群低于中心种群 marginal populations < central populations
奠基者效应、瓶颈效应 Founder effect, bottleneck effect
Chen et al., 2012
北沙柳 Salix psammophila
边缘种群低于中心种群 marginal populations < central populations
奠基者效应 Founder effect
Hao et al., 2017
花苜蓿、青海苜蓿 Medicago ruthenica, M. archiducis-nicolai
边缘种群低于中心种群 marginal populations < central populations
奠基者效应 Founder effect
Wu et al., 2016
毛红椿 Toona ciliata var. pubescens
边缘种群高于中心种群 marginal populations > central populations
生境破碎化 Habitat fragmentation
Liu et al., 2013
红松 Pinus koraiensis
边缘种群低于中心种群 marginal populations < central populations
奠基者效应、瓶颈效应 Founder effect, bottleneck effect
Feng et al., 2006
领春木 Euptelea pleiospermum
边缘种群低于中心种群 marginal populations < central populations
冰期后扩张、不对称基因流 Postglacial expansion and asymmetric gene flow
Genetic divergence and signatures of natural selection in marginal populations of a keystone, long-lived conifer, eastern white pine (Pinus strobus) from northern Ontario 1 2014
... 当物种呈离散分布或离散与连续混合分布时, 边缘种群处于相对隔离状态, 如物种分布先扩张后收缩, 容易产生与主要种群完全分离的小种群, 隔离的边缘种群可逐渐形成新物种(Coyne & Orr, 2004).在无迁移时, 其他基本进化动力都可参与维持隔离种群的遗传多样性.当边缘种群很小时, 如冰期后仍处于隔离状态的原避难所种群, 漂变效应起主导作用, 导致等位基因丢失, 杂合子缺失, 遗传多样性降低.Corylus avellana边缘种群比中心种群的遗传多样性低, 这是由冰期后物种边缘扩张时的瓶颈效应或遗传漂变所致(Persson et al., 2004);Stipa pennata的边缘种群遗传多样性低也是由遗传漂变效应引起的(Wagner et al., 2012).类似的报道还有对鹅掌楸(Liriodendron chinense)?(Yang et al., 2016)、欧亚大陆的Adonis vernalis (Hirsch et al., 2015)、我国的野生稻(Oryza rufipogon)?(Gao & Gao, 2016)等的研究, 这些研究揭示了以遗传漂变为主的进化过程对边缘种群遗传多样性的影响. ...
Intraspecific diversity and dominant genotypes resist plant invasions 1 2008
... 此外, 当单个物种边缘种群发生在一个或多个不同的生物群落组成的过渡地带时, 所在群落的物种多样性和生态学特征不同于中心区域的群落(Kimmins, 2004).已有报道证明单个物种遗传多样性对生态系统功能有直接或间接的影响.例如, Kotowska等(2010)发现当一年生植物拟南芥(Arabidopsis thaliana)的基因型多样性较高时, 拟南芥及其天敌粉纹夜蛾(Trichoplusia ni)均会有较高的生物量和存活率.Johnson等(2006)研究发现遗传多样化的植物群落能支持更加多样的动物群落, 且对干扰的耐受性也会更强(Crutsinger et al., 2008).虽然物种遗传多样性与群落物种多样性的关系已有许多报道(Jump et al., 2003; He et al., 2008; Laikre et al., 2010; Wei et al., 2017), 但边缘种群遗传多样性与所在群落物种多样性的相互作用很少有报道, 因此, 分析边缘种群遗传多样性有助于深入理解群落过渡带或非过渡带生物多样性的维持机制. ...
Broad geographic covariation between floral traits and the mating system in Camissoniopsis cheiranthifolia(Onagraceae): Multiple stable mixed mating systems across the species’ range 1 2012
... Contrasts in mating systems between central and marginal populations of a range of plant species Table 2
分类群 Taxonomic group
中心/亚中心种群 Central/subcentral population
边缘种群 Marginal population
参考文献 Reference
Leavenworthia alabamica
自交不亲和 Self-incompatibility
自交亲和/自我受精 Self-compatible/self-fertilization
Busch, 2005
Juncus atratus
低近交率 Low inbreeding rates
高近交率, 异交率为5.6% High inbreeding, outcrossing rate = 5.6%
... Comparison of genetic diversity between central and marginal populations of various plant species and the potential ecological or evolutionary processes responsible for the observed differences Table 3
植物种 Species
边缘种群与中心种群遗传多样性 Genetic diversity in marginal vs. central populations
生态或进化机制 Ecological or evolutionary mechanisms
参考文献 Reference
樱桃 Cerasus pseudocerasus
边缘种群低于中心种群 marginal populations < central populations
奠基者效应、瓶颈效应 Founder effect, bottleneck effect
Chen et al., 2012
北沙柳 Salix psammophila
边缘种群低于中心种群 marginal populations < central populations
奠基者效应 Founder effect
Hao et al., 2017
花苜蓿、青海苜蓿 Medicago ruthenica, M. archiducis-nicolai
边缘种群低于中心种群 marginal populations < central populations
奠基者效应 Founder effect
Wu et al., 2016
毛红椿 Toona ciliata var. pubescens
边缘种群高于中心种群 marginal populations > central populations
生境破碎化 Habitat fragmentation
Liu et al., 2013
红松 Pinus koraiensis
边缘种群低于中心种群 marginal populations < central populations
奠基者效应、瓶颈效应 Founder effect, bottleneck effect
Feng et al., 2006
领春木 Euptelea pleiospermum
边缘种群低于中心种群 marginal populations < central populations
冰期后扩张、不对称基因流 Postglacial expansion and asymmetric gene flow
Southward Pleistocene migration of Douglas-fir into Mexico: Phylogeography, ecological niche modeling, and conservation of “rear edge” populations 2 2011
... Theoretical models of a species’ distribution Table 1
模型 Model
主要观点 Main point
参考文献 Reference
随机生态位模型或断棒模型 Stochastic niche model or broken stick model
每一物种生态位超体积的占有和分布不受其他物种影响. The niche occupation and its size distribution of each species are random and independent of the niche sizes of other species.
MacArthur, 1957; Whittaker et al., 2010
对数正态分布模型 Lognormal distribution model
物种占有的生态位是随机分布的并受大量因素综合影响, 并不优待某些种. The niche size of a species is random and determined by the joint effects of a large number of factors, and no selective advantage is present among species.
Preston, 1948
生态位优先占领模型 Niche pre-emption model
第一位优势种优先占领生态位空间大部, 第二位占领其余下的大部, 以此类推, 末位只占留下的极少空间. The first dominant species occupies the largest niche space, followed by the species that occupies the second largest niche in the remaining space, and so on. The last species occupies the minimum niche.
Whittaker, 1972
群落中性理论 Neutral community theory
群落内个体总数固定, 某一物种多度的增加必然伴随其他物种的减少; 所有个体出生率、死亡率相同. The community size is fixed, and a decrease of one species’ abundance is equally compensated by other species. All individuals in the community have the same birth and death rates.
Hubbell, 2001
自然选择-基因流机制 Mechanism of natural selection-gene flow
基因由中心向边缘种群的迁移与边缘种群的自然选择作用持衡. Effects of gene flow from the central to marginal population are in balance with the effects of natural selection in the marginal population.
... Comparison of genetic diversity between central and marginal populations of various plant species and the potential ecological or evolutionary processes responsible for the observed differences Table 3
植物种 Species
边缘种群与中心种群遗传多样性 Genetic diversity in marginal vs. central populations
生态或进化机制 Ecological or evolutionary mechanisms
参考文献 Reference
樱桃 Cerasus pseudocerasus
边缘种群低于中心种群 marginal populations < central populations
奠基者效应、瓶颈效应 Founder effect, bottleneck effect
Chen et al., 2012
北沙柳 Salix psammophila
边缘种群低于中心种群 marginal populations < central populations
奠基者效应 Founder effect
Hao et al., 2017
花苜蓿、青海苜蓿 Medicago ruthenica, M. archiducis-nicolai
边缘种群低于中心种群 marginal populations < central populations
奠基者效应 Founder effect
Wu et al., 2016
毛红椿 Toona ciliata var. pubescens
边缘种群高于中心种群 marginal populations > central populations
生境破碎化 Habitat fragmentation
Liu et al., 2013
红松 Pinus koraiensis
边缘种群低于中心种群 marginal populations < central populations
奠基者效应、瓶颈效应 Founder effect, bottleneck effect
Feng et al., 2006
领春木 Euptelea pleiospermum
边缘种群低于中心种群 marginal populations < central populations
冰期后扩张、不对称基因流 Postglacial expansion and asymmetric gene flow
... Comparison of genetic diversity between central and marginal populations of various plant species and the potential ecological or evolutionary processes responsible for the observed differences Table 3
植物种 Species
边缘种群与中心种群遗传多样性 Genetic diversity in marginal vs. central populations
生态或进化机制 Ecological or evolutionary mechanisms
参考文献 Reference
樱桃 Cerasus pseudocerasus
边缘种群低于中心种群 marginal populations < central populations
奠基者效应、瓶颈效应 Founder effect, bottleneck effect
Chen et al., 2012
北沙柳 Salix psammophila
边缘种群低于中心种群 marginal populations < central populations
奠基者效应 Founder effect
Hao et al., 2017
花苜蓿、青海苜蓿 Medicago ruthenica, M. archiducis-nicolai
边缘种群低于中心种群 marginal populations < central populations
奠基者效应 Founder effect
Wu et al., 2016
毛红椿 Toona ciliata var. pubescens
边缘种群高于中心种群 marginal populations > central populations
生境破碎化 Habitat fragmentation
Liu et al., 2013
红松 Pinus koraiensis
边缘种群低于中心种群 marginal populations < central populations
奠基者效应、瓶颈效应 Founder effect, bottleneck effect
Feng et al., 2006
领春木 Euptelea pleiospermum
边缘种群低于中心种群 marginal populations < central populations
冰期后扩张、不对称基因流 Postglacial expansion and asymmetric gene flow
... 由于前缘、后缘种群以及中心种群的形成或维持机制不同, 在基因组遗传结构变异上也会表现出不同的遗传多样性分布模式, 例如杂合子频率和Fst异常值在位点间的分布或二者的相关性, 以及在染色体上的分布(Hu & He, 2005; Hu et al., 2011), 中性与适应性位点在染色体上的分布等.今后的一个重要研究方向就是利用高通量基因组测序分析边缘与中心种群在适应性位点上的差异, 或者利用转录组数据筛选出种群间的差异表达基因, 寻找与种群边缘适应性相关的基因分布, 揭示导致边缘种群遗传多样性差异的生态和进化过程(Provan, 2013). ...
Scoring the mating systems of natural populations of three Larix taxa in China: L. gmelinii(Rupr.) Rupr., L. olgensis Henry and L. principis-?rupprechtii Mayr 1 1999
... Theoretical models of a species’ distribution Table 1
模型 Model
主要观点 Main point
参考文献 Reference
随机生态位模型或断棒模型 Stochastic niche model or broken stick model
每一物种生态位超体积的占有和分布不受其他物种影响. The niche occupation and its size distribution of each species are random and independent of the niche sizes of other species.
MacArthur, 1957; Whittaker et al., 2010
对数正态分布模型 Lognormal distribution model
物种占有的生态位是随机分布的并受大量因素综合影响, 并不优待某些种. The niche size of a species is random and determined by the joint effects of a large number of factors, and no selective advantage is present among species.
Preston, 1948
生态位优先占领模型 Niche pre-emption model
第一位优势种优先占领生态位空间大部, 第二位占领其余下的大部, 以此类推, 末位只占留下的极少空间. The first dominant species occupies the largest niche space, followed by the species that occupies the second largest niche in the remaining space, and so on. The last species occupies the minimum niche.
Whittaker, 1972
群落中性理论 Neutral community theory
群落内个体总数固定, 某一物种多度的增加必然伴随其他物种的减少; 所有个体出生率、死亡率相同. The community size is fixed, and a decrease of one species’ abundance is equally compensated by other species. All individuals in the community have the same birth and death rates.
Hubbell, 2001
自然选择-基因流机制 Mechanism of natural selection-gene flow
基因由中心向边缘种群的迁移与边缘种群的自然选择作用持衡. Effects of gene flow from the central to marginal population are in balance with the effects of natural selection in the marginal population.
... 此外, 当单个物种边缘种群发生在一个或多个不同的生物群落组成的过渡地带时, 所在群落的物种多样性和生态学特征不同于中心区域的群落(Kimmins, 2004).已有报道证明单个物种遗传多样性对生态系统功能有直接或间接的影响.例如, Kotowska等(2010)发现当一年生植物拟南芥(Arabidopsis thaliana)的基因型多样性较高时, 拟南芥及其天敌粉纹夜蛾(Trichoplusia ni)均会有较高的生物量和存活率.Johnson等(2006)研究发现遗传多样化的植物群落能支持更加多样的动物群落, 且对干扰的耐受性也会更强(Crutsinger et al., 2008).虽然物种遗传多样性与群落物种多样性的关系已有许多报道(Jump et al., 2003; He et al., 2008; Laikre et al., 2010; Wei et al., 2017), 但边缘种群遗传多样性与所在群落物种多样性的相互作用很少有报道, 因此, 分析边缘种群遗传多样性有助于深入理解群落过渡带或非过渡带生物多样性的维持机制. ...
Genetic variation, biogeographical history, and conservation of Anthyllis montana L. ssp. jacquinii(Kern.) Hayek (Fabaceae) at its northern distribution limit 2012
Neglect of genetic diversity in implementation of the convention on biological diversity 1 2010
... 此外, 当单个物种边缘种群发生在一个或多个不同的生物群落组成的过渡地带时, 所在群落的物种多样性和生态学特征不同于中心区域的群落(Kimmins, 2004).已有报道证明单个物种遗传多样性对生态系统功能有直接或间接的影响.例如, Kotowska等(2010)发现当一年生植物拟南芥(Arabidopsis thaliana)的基因型多样性较高时, 拟南芥及其天敌粉纹夜蛾(Trichoplusia ni)均会有较高的生物量和存活率.Johnson等(2006)研究发现遗传多样化的植物群落能支持更加多样的动物群落, 且对干扰的耐受性也会更强(Crutsinger et al., 2008).虽然物种遗传多样性与群落物种多样性的关系已有许多报道(Jump et al., 2003; He et al., 2008; Laikre et al., 2010; Wei et al., 2017), 但边缘种群遗传多样性与所在群落物种多样性的相互作用很少有报道, 因此, 分析边缘种群遗传多样性有助于深入理解群落过渡带或非过渡带生物多样性的维持机制. ...
Variation in habitat connectivity generates positive correlations between species and genetic diversity in a metacommunity 1 2013
... 理论上群落物种多样性与种群遗传多样性是两个不同层次但又相互关联的概念(Hu et al., 2006).在群落中性理论假设下(Hubbell, 2001), 若有效群落数量相对稳定, 群落漂变会导致群落内种群遗传漂变, 群落内物种之间的多度应呈负相关关系, 也可导致所谓的单个物种遗传多样性与群落物种多样性相关(species-gene diversity correlation (SGDC); Vellend, 2005).Laroche等(2015)证明当突变率低时, 群落间个体迁移变异可产生正的SGDC; 当突变率高时, 突变、迁移及竞争联合作用可产生负的SGDC, 因此, 中性过程并不总是产生负的SGDC.实际研究发现单个物种遗传多样性与群落物种多样性既有正相关关系(Blum et al., 2012; Lamy et al., 2013), 又有负相关关系(Pu?ca? et al., 2008), 或不相关(Taberlet et al., 2012; Avolio & Smith, 2013), 反映了不同的生态和进化过程作用于两个不同层次系统的结果. ...
A neutral theory for interpreting correlations between species and genetic diversity in communities 1 2015
... 理论上群落物种多样性与种群遗传多样性是两个不同层次但又相互关联的概念(Hu et al., 2006).在群落中性理论假设下(Hubbell, 2001), 若有效群落数量相对稳定, 群落漂变会导致群落内种群遗传漂变, 群落内物种之间的多度应呈负相关关系, 也可导致所谓的单个物种遗传多样性与群落物种多样性相关(species-gene diversity correlation (SGDC); Vellend, 2005).Laroche等(2015)证明当突变率低时, 群落间个体迁移变异可产生正的SGDC; 当突变率高时, 突变、迁移及竞争联合作用可产生负的SGDC, 因此, 中性过程并不总是产生负的SGDC.实际研究发现单个物种遗传多样性与群落物种多样性既有正相关关系(Blum et al., 2012; Lamy et al., 2013), 又有负相关关系(Pu?ca? et al., 2008), 或不相关(Taberlet et al., 2012; Avolio & Smith, 2013), 反映了不同的生态和进化过程作用于两个不同层次系统的结果. ...
Population size, center-periphery, and seed dispersers’ effects on the genetic diversity and population structure of the Mediterranean relict shrub Cneorum tricoccon 1 2017
... Comparison of genetic diversity between central and marginal populations of various plant species and the potential ecological or evolutionary processes responsible for the observed differences Table 3
植物种 Species
边缘种群与中心种群遗传多样性 Genetic diversity in marginal vs. central populations
生态或进化机制 Ecological or evolutionary mechanisms
参考文献 Reference
樱桃 Cerasus pseudocerasus
边缘种群低于中心种群 marginal populations < central populations
奠基者效应、瓶颈效应 Founder effect, bottleneck effect
Chen et al., 2012
北沙柳 Salix psammophila
边缘种群低于中心种群 marginal populations < central populations
奠基者效应 Founder effect
Hao et al., 2017
花苜蓿、青海苜蓿 Medicago ruthenica, M. archiducis-nicolai
边缘种群低于中心种群 marginal populations < central populations
奠基者效应 Founder effect
Wu et al., 2016
毛红椿 Toona ciliata var. pubescens
边缘种群高于中心种群 marginal populations > central populations
生境破碎化 Habitat fragmentation
Liu et al., 2013
红松 Pinus koraiensis
边缘种群低于中心种群 marginal populations < central populations
奠基者效应、瓶颈效应 Founder effect, bottleneck effect
Feng et al., 2006
领春木 Euptelea pleiospermum
边缘种群低于中心种群 marginal populations < central populations
冰期后扩张、不对称基因流 Postglacial expansion and asymmetric gene flow
... Comparison of genetic diversity between central and marginal populations of various plant species and the potential ecological or evolutionary processes responsible for the observed differences Table 3
植物种 Species
边缘种群与中心种群遗传多样性 Genetic diversity in marginal vs. central populations
生态或进化机制 Ecological or evolutionary mechanisms
参考文献 Reference
樱桃 Cerasus pseudocerasus
边缘种群低于中心种群 marginal populations < central populations
奠基者效应、瓶颈效应 Founder effect, bottleneck effect
Chen et al., 2012
北沙柳 Salix psammophila
边缘种群低于中心种群 marginal populations < central populations
奠基者效应 Founder effect
Hao et al., 2017
花苜蓿、青海苜蓿 Medicago ruthenica, M. archiducis-nicolai
边缘种群低于中心种群 marginal populations < central populations
奠基者效应 Founder effect
Wu et al., 2016
毛红椿 Toona ciliata var. pubescens
边缘种群高于中心种群 marginal populations > central populations
生境破碎化 Habitat fragmentation
Liu et al., 2013
红松 Pinus koraiensis
边缘种群低于中心种群 marginal populations < central populations
奠基者效应、瓶颈效应 Founder effect, bottleneck effect
Feng et al., 2006
领春木 Euptelea pleiospermum
边缘种群低于中心种群 marginal populations < central populations
冰期后扩张、不对称基因流 Postglacial expansion and asymmetric gene flow
... Theoretical models of a species’ distribution Table 1
模型 Model
主要观点 Main point
参考文献 Reference
随机生态位模型或断棒模型 Stochastic niche model or broken stick model
每一物种生态位超体积的占有和分布不受其他物种影响. The niche occupation and its size distribution of each species are random and independent of the niche sizes of other species.
MacArthur, 1957; Whittaker et al., 2010
对数正态分布模型 Lognormal distribution model
物种占有的生态位是随机分布的并受大量因素综合影响, 并不优待某些种. The niche size of a species is random and determined by the joint effects of a large number of factors, and no selective advantage is present among species.
Preston, 1948
生态位优先占领模型 Niche pre-emption model
第一位优势种优先占领生态位空间大部, 第二位占领其余下的大部, 以此类推, 末位只占留下的极少空间. The first dominant species occupies the largest niche space, followed by the species that occupies the second largest niche in the remaining space, and so on. The last species occupies the minimum niche.
Whittaker, 1972
群落中性理论 Neutral community theory
群落内个体总数固定, 某一物种多度的增加必然伴随其他物种的减少; 所有个体出生率、死亡率相同. The community size is fixed, and a decrease of one species’ abundance is equally compensated by other species. All individuals in the community have the same birth and death rates.
Hubbell, 2001
自然选择-基因流机制 Mechanism of natural selection-gene flow
基因由中心向边缘种群的迁移与边缘种群的自然选择作用持衡. Effects of gene flow from the central to marginal population are in balance with the effects of natural selection in the marginal population.
Contrasting demographic history and population structure in Capsella rubella and Capsella grandiflora, two closely related species with different mating systems 1 2011
... 自交降低种群内的遗传多样性, 但增加了种群间的遗传分化(张大勇和姜新华, 2001; Bakker et al., 2006; Onge et al., 2011; Pettengill et al., 2016), 包括增加边缘与中心种群以及边缘种群间的遗传分化(Hampe & Petit, 2005; Griffin & Willi, 2014).在迁移和漂变联合作用下, 自交率(α)与种群遗传分化系数(Fst)的理论关系可以从Wright (1969)、Caballero和Hill (1992)及Hu (2011)的研究结果中获得近似表达, 即 ...
Mating systems and the evolutionary transition between haploidy and diploidy 1 1996
Mating system divergence affects the distribution of sequence diversity within and among populations of recently diverged subspecies of Clarkia xantiana(Onagraceae) 1 2016
... 自交降低种群内的遗传多样性, 但增加了种群间的遗传分化(张大勇和姜新华, 2001; Bakker et al., 2006; Onge et al., 2011; Pettengill et al., 2016), 包括增加边缘与中心种群以及边缘种群间的遗传分化(Hampe & Petit, 2005; Griffin & Willi, 2014).在迁移和漂变联合作用下, 自交率(α)与种群遗传分化系数(Fst)的理论关系可以从Wright (1969)、Caballero和Hill (1992)及Hu (2011)的研究结果中获得近似表达, 即 ...
Geographic variation in genetic and demographic performance: New insights from an old biogeographical paradigm 1 2017
... 中心-边缘模型在理论和实践上也受到了挑战(Gaston, 2003; Sagarin et al., 2006; Sexton et al., 2009).理论上, Guo (2012)提出将物种纬度线性变异趋势与中心-边缘模式综合生成偏正态驼峰型分布, 用于评价物种全分布范围内的遗传变异趋势(Hirao et al., 2017).同样, 该模型预测的边缘种群遗传多样性格局也受到挑战, Eckert等(2008)分析了134个研究中115个物种的结果, 发现64.2%研究结果显示边缘种群遗传多样性下降, 70.2%的研究显示边缘种群间遗传分化增强, 但多数研究中中心与边缘种群遗传多样性差异不显著.近期, Pironon等(2017)分析了248个研究, 认为81%的研究显示物种出现在中心区域的频率要比在边缘区域高, 但只有51%的研究显示物种的密度变异模式支持中心-边缘模型, 47%的研究结果显示遗传多样性从中心到边缘逐渐减小. ...
Limits to adaptation along environmental gradients 1 2015
... Theoretical models of a species’ distribution Table 1
模型 Model
主要观点 Main point
参考文献 Reference
随机生态位模型或断棒模型 Stochastic niche model or broken stick model
每一物种生态位超体积的占有和分布不受其他物种影响. The niche occupation and its size distribution of each species are random and independent of the niche sizes of other species.
MacArthur, 1957; Whittaker et al., 2010
对数正态分布模型 Lognormal distribution model
物种占有的生态位是随机分布的并受大量因素综合影响, 并不优待某些种. The niche size of a species is random and determined by the joint effects of a large number of factors, and no selective advantage is present among species.
Preston, 1948
生态位优先占领模型 Niche pre-emption model
第一位优势种优先占领生态位空间大部, 第二位占领其余下的大部, 以此类推, 末位只占留下的极少空间. The first dominant species occupies the largest niche space, followed by the species that occupies the second largest niche in the remaining space, and so on. The last species occupies the minimum niche.
Whittaker, 1972
群落中性理论 Neutral community theory
群落内个体总数固定, 某一物种多度的增加必然伴随其他物种的减少; 所有个体出生率、死亡率相同. The community size is fixed, and a decrease of one species’ abundance is equally compensated by other species. All individuals in the community have the same birth and death rates.
Hubbell, 2001
自然选择-基因流机制 Mechanism of natural selection-gene flow
基因由中心向边缘种群的迁移与边缘种群的自然选择作用持衡. Effects of gene flow from the central to marginal population are in balance with the effects of natural selection in the marginal population.
... Comparison of genetic diversity between central and marginal populations of various plant species and the potential ecological or evolutionary processes responsible for the observed differences Table 3
植物种 Species
边缘种群与中心种群遗传多样性 Genetic diversity in marginal vs. central populations
生态或进化机制 Ecological or evolutionary mechanisms
参考文献 Reference
樱桃 Cerasus pseudocerasus
边缘种群低于中心种群 marginal populations < central populations
奠基者效应、瓶颈效应 Founder effect, bottleneck effect
Chen et al., 2012
北沙柳 Salix psammophila
边缘种群低于中心种群 marginal populations < central populations
奠基者效应 Founder effect
Hao et al., 2017
花苜蓿、青海苜蓿 Medicago ruthenica, M. archiducis-nicolai
边缘种群低于中心种群 marginal populations < central populations
奠基者效应 Founder effect
Wu et al., 2016
毛红椿 Toona ciliata var. pubescens
边缘种群高于中心种群 marginal populations > central populations
生境破碎化 Habitat fragmentation
Liu et al., 2013
红松 Pinus koraiensis
边缘种群低于中心种群 marginal populations < central populations
奠基者效应、瓶颈效应 Founder effect, bottleneck effect
Feng et al., 2006
领春木 Euptelea pleiospermum
边缘种群低于中心种群 marginal populations < central populations
冰期后扩张、不对称基因流 Postglacial expansion and asymmetric gene flow
Evolution and measurement of species diversity 1 1972
... Theoretical models of a species’ distribution Table 1
模型 Model
主要观点 Main point
参考文献 Reference
随机生态位模型或断棒模型 Stochastic niche model or broken stick model
每一物种生态位超体积的占有和分布不受其他物种影响. The niche occupation and its size distribution of each species are random and independent of the niche sizes of other species.
MacArthur, 1957; Whittaker et al., 2010
对数正态分布模型 Lognormal distribution model
物种占有的生态位是随机分布的并受大量因素综合影响, 并不优待某些种. The niche size of a species is random and determined by the joint effects of a large number of factors, and no selective advantage is present among species.
Preston, 1948
生态位优先占领模型 Niche pre-emption model
第一位优势种优先占领生态位空间大部, 第二位占领其余下的大部, 以此类推, 末位只占留下的极少空间. The first dominant species occupies the largest niche space, followed by the species that occupies the second largest niche in the remaining space, and so on. The last species occupies the minimum niche.
Whittaker, 1972
群落中性理论 Neutral community theory
群落内个体总数固定, 某一物种多度的增加必然伴随其他物种的减少; 所有个体出生率、死亡率相同. The community size is fixed, and a decrease of one species’ abundance is equally compensated by other species. All individuals in the community have the same birth and death rates.
Hubbell, 2001
自然选择-基因流机制 Mechanism of natural selection-gene flow
基因由中心向边缘种群的迁移与边缘种群的自然选择作用持衡. Effects of gene flow from the central to marginal population are in balance with the effects of natural selection in the marginal population.
... Theoretical models of a species’ distribution Table 1
模型 Model
主要观点 Main point
参考文献 Reference
随机生态位模型或断棒模型 Stochastic niche model or broken stick model
每一物种生态位超体积的占有和分布不受其他物种影响. The niche occupation and its size distribution of each species are random and independent of the niche sizes of other species.
MacArthur, 1957; Whittaker et al., 2010
对数正态分布模型 Lognormal distribution model
物种占有的生态位是随机分布的并受大量因素综合影响, 并不优待某些种. The niche size of a species is random and determined by the joint effects of a large number of factors, and no selective advantage is present among species.
Preston, 1948
生态位优先占领模型 Niche pre-emption model
第一位优势种优先占领生态位空间大部, 第二位占领其余下的大部, 以此类推, 末位只占留下的极少空间. The first dominant species occupies the largest niche space, followed by the species that occupies the second largest niche in the remaining space, and so on. The last species occupies the minimum niche.
Whittaker, 1972
群落中性理论 Neutral community theory
群落内个体总数固定, 某一物种多度的增加必然伴随其他物种的减少; 所有个体出生率、死亡率相同. The community size is fixed, and a decrease of one species’ abundance is equally compensated by other species. All individuals in the community have the same birth and death rates.
Hubbell, 2001
自然选择-基因流机制 Mechanism of natural selection-gene flow
基因由中心向边缘种群的迁移与边缘种群的自然选择作用持衡. Effects of gene flow from the central to marginal population are in balance with the effects of natural selection in the marginal population.
... Comparison of genetic diversity between central and marginal populations of various plant species and the potential ecological or evolutionary processes responsible for the observed differences Table 3
植物种 Species
边缘种群与中心种群遗传多样性 Genetic diversity in marginal vs. central populations
生态或进化机制 Ecological or evolutionary mechanisms
参考文献 Reference
樱桃 Cerasus pseudocerasus
边缘种群低于中心种群 marginal populations < central populations
奠基者效应、瓶颈效应 Founder effect, bottleneck effect
Chen et al., 2012
北沙柳 Salix psammophila
边缘种群低于中心种群 marginal populations < central populations
奠基者效应 Founder effect
Hao et al., 2017
花苜蓿、青海苜蓿 Medicago ruthenica, M. archiducis-nicolai
边缘种群低于中心种群 marginal populations < central populations
奠基者效应 Founder effect
Wu et al., 2016
毛红椿 Toona ciliata var. pubescens
边缘种群高于中心种群 marginal populations > central populations
生境破碎化 Habitat fragmentation
Liu et al., 2013
红松 Pinus koraiensis
边缘种群低于中心种群 marginal populations < central populations
奠基者效应、瓶颈效应 Founder effect, bottleneck effect
Feng et al., 2006
领春木 Euptelea pleiospermum
边缘种群低于中心种群 marginal populations < central populations
冰期后扩张、不对称基因流 Postglacial expansion and asymmetric gene flow
... Comparison of genetic diversity between central and marginal populations of various plant species and the potential ecological or evolutionary processes responsible for the observed differences Table 3
植物种 Species
边缘种群与中心种群遗传多样性 Genetic diversity in marginal vs. central populations
生态或进化机制 Ecological or evolutionary mechanisms
参考文献 Reference
樱桃 Cerasus pseudocerasus
边缘种群低于中心种群 marginal populations < central populations
奠基者效应、瓶颈效应 Founder effect, bottleneck effect
Chen et al., 2012
北沙柳 Salix psammophila
边缘种群低于中心种群 marginal populations < central populations
奠基者效应 Founder effect
Hao et al., 2017
花苜蓿、青海苜蓿 Medicago ruthenica, M. archiducis-nicolai
边缘种群低于中心种群 marginal populations < central populations
奠基者效应 Founder effect
Wu et al., 2016
毛红椿 Toona ciliata var. pubescens
边缘种群高于中心种群 marginal populations > central populations
生境破碎化 Habitat fragmentation
Liu et al., 2013
红松 Pinus koraiensis
边缘种群低于中心种群 marginal populations < central populations
奠基者效应、瓶颈效应 Founder effect, bottleneck effect
Feng et al., 2006
领春木 Euptelea pleiospermum
边缘种群低于中心种群 marginal populations < central populations
冰期后扩张、不对称基因流 Postglacial expansion and asymmetric gene flow
 ,, 王茜, 胡颖, 周玮, 陈晓阳, 胡新生
,, 王茜, 胡颖, 周玮, 陈晓阳, 胡新生 ,*广东省植物种质资源与利用重点实验室, 华南农业大学林学与风景园林学院, 广州 510642
,*广东省植物种质资源与利用重点实验室, 华南农业大学林学与风景园林学院, 广州 510642
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}