王明明
1 , 2 , 刘新平
, 1 , 3 , * , 何玉惠
4 , 张铜会
1 , 3 , 魏静
5 , 车力木格
1 , 2 , 孙姗姗
1 , 2 1 中国科学院西北生态环境资源研究院奈曼沙漠化研究站, 兰州 7300002 中国科学院大学, 北京 1000493 中国科学院西北生态环境资源研究院乌拉特荒漠草原研究站, 兰州 7300004 中国科学院西北生态环境资源研究院皋兰生态与农业综合研究站, 兰州 7300005 北京大学深圳研究生院城市人居环境科学与技术重点实验室, 广东深圳 518000How enclosure influences restored plant community changes of different initial types in Horqin Sandy Land Ming-Ming WANG
1 , 2 , Xin-Ping LIU
, 1 , 3 , * , Yu-Hui HE
4 , Tong-Hui ZHANG
1 , 3 , Jing WEI
5 , Chelmge
1 , 2 , Shan-Shan SUN
1 , 2 1 Naiman Desertification Research Station, Northwest Institute of Eco-Environment and Resources, Chinese Academy of Sciences, Lanzhou 730000, China 2 University of Chinese Academy of Sciences, Beijing 100049, China 3 Urat Desert-grassland Research Station, Northwest Institute of Eco-Environment and Resources, Chinese Academy of Science, Lanzhou 730000, China 4 Gaolan Station of Agricultural and Ecological Experiment, Northwest Institute of Eco-Environment and Resources, Chinese Academy of Science, Lanzhou 730000, China 5 Key Laboratory for Urban Habitat Environmental Science and Technology, Peking University Shenzhen Graduate School, Shenzhen, Guangdong 518000, China 通讯作者: * (
liuxinping@lzb.ac.cn )
编委: 梁存柱
责任编辑: 李敏, 赵航(实习)
收稿日期: 2019-03-26
修回日期: 2019-07-23
网络出版日期: 2019-08-20
基金资助: 国家重点研发计划 (2017YFC0506706 )国家重点研发计划 (2016YFC0500907 )国家自然科学基金 (41801076 )内蒙古自治区科技重大专项 (Y749BJ1001 )中国科学院科技扶贫项目
Corresponding authors: * liuxinping@lzb.ac.cn Received: 2019-03-26
Revised: 2019-07-23
Online: 2019-08-20
Fund supported: National Key R&D Program of China (2017YFC0506706 )National Key R&D Program of China (2016YFC0500907 )National Natural Science Foundation of China (41801076 )Nei Mongol Autonomous Region Science and Technology Major Project (Y749BJ1001 )Science and Technology Poverty Alleviation Program of Chinese Academy of Sciences
摘要 封育是退化沙地植被恢复与生态重建的重要措施, 理解长期处于封育状态下不同类型沙地植物群落特征变化及其影响因素有利于沙地植被恢复和生态重建。该文基于对科尔沁沙地长期封育的流动沙丘(2005年封育)、固定沙丘(1985年封育)和沙质草地(1997年封育)连续多年(2005-2017年)的植物群落调查, 结合土壤种子库、土壤养分以及气象数据, 分析了植物群落特征变化及其对环境变化的响应。研究结果表明流动沙丘植被盖度显著增加, 群落生物量和物种多样性年际间波动变化, 但无明显趋势; 固定沙丘植物群落存在逆行演替趋势, 具体表现为群落生物量、灌木和半灌木以及豆科优势度显著下降, 而一年生和多年生杂类草优势度显著增加; 沙质草地群落物种丰富度和多年生禾草优势度存在降低趋势, 并且一年生杂类草优势度明显高于其他功能群, 群落存在退化现象。3类沙地土壤种子密度变化不显著, 而种子丰富度在流动沙丘显著增加, 在固定沙丘和沙质草地有下降趋势, 土壤养分仅有有效氮和有效磷含量增加。回归分析结果表明气温和降水是影响年内生物量积累的主要因素, 但对年际间群落生物量和物种丰富度变化影响不大。除趋势对应分析结果显示土壤种子库与植物群落之间存在很高的相似性, 典型相关分析结果表明沙质草地植物群落与土壤养分紧密相关, 而固定沙丘群落主要与土壤水分紧密相关。综合以上结果可知, 封育33年的固定沙丘群落和封育21年的沙质草地群落都存在退化现象, 而封育11年的流动沙丘群落正在缓慢恢复, 因此封育年限的设定对退化沙地植被恢复至关重要, 封育时间过长不仅不利于植物群落恢复, 反而会使群落发生逆行演替, 建议封育年限的设定应综合考虑植被退化程度、土壤养分状况、土壤种子库基础以及气候条件等因素的影响。
关键词: 沙地 ;
封育 ;
植被恢复 ;
群落特征 ;
影响因素 Abstract Aims Enclosure is one of the important measures for vegetation restoration of degraded sandy land. Understanding the plant community change of different initial types in long-term state of enclosing is vital for us to understand the vegetation restoration process or re-vegetation in sandy land. This paper aims to analyze the changes of plant communities and its comparative responses to long-term enclosure (2005-2017) of mobile dunes (enclosed in 2005), fixed dunes (enclosed in 1985) and sandy grassland (enclosed in 1997), in relation to soil seed bank, soil nutrient and precipitation and air temperature.
Methods The species composition, height, coverage and above-ground biomass were measured by quadrats in every year during 2005-2017. The soil organic carbon, soil nutrient and soil seed bank were measured by soil cores from the above quadrats in 2008 and 2017. Detrended correspondence analysis (DCA) and canonical correspondence analysis (CCA) were used to explore the relationship between plant community species composition and soil seed bank soil nutrients and soil moisture.
Important findings The results showed that the vegetation coverage and community species richness of mobile dune was significantly increased, but the community biomass had no obvious trend. The biomass of fixed dune community, shrubs, semi-shrubs and the perennial legume functional group dominances decreased significantly, but the dominance of the annual and perennial forbs increased significantly. The annual forbs were the dominant functional group, and community species richness and perennial grass dominances decreased significantly. The soil seed bank had no remarkable change in the three enclosed sandy land communities during 2008-2017, while the seed richness significantly increased in mobile dunes, and that of the fixed dunes and grassland presented downward trends. The soil available nitrogen and available phosphorus were increased significantly. Regression analysis showed the annual variation of plant community biomass was significantly affected by air temperature and precipitation, but there was a little effect on the inter-annual variation of plant community biomass and species richness. Detrended correspondence analysis (DCA) displayed that there was a high similarity between soil seed bank and plant community, and typical correlation analysis (CCA) results showed that grassland community species composition was closely related to soil nutrients. However, fixed dune community species composition was mainly related to soil moisture. In summary, the fixed dune plant community enclosed for 33 years and the sandy grassland plant community enclosed for 21 years present degraded trends, while the mobile dune plant community enclosed for 11 years is slowly recovering, thus, long-term enclosure is not always conducive to the restoration of degraded sandy land vegetation. We suggested that the influence of degradation degree of vegetation, soil nutrients and moisture, soil seed bank and precipitation should be synthetically considered when we set the duration of enclosure for restoration.
Keywords: sandy land ;
enclosure ;
vegetation restoration ;
community features ;
influencing factors PDF (4291KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 王明明, 刘新平, 何玉惠, 张铜会, 魏静, 车力木格, 孙姗姗. 科尔沁沙地封育恢复过程中植物群落特征变化及影响因素. 植物生态学报, 2019, 43(8): 672-684. DOI:
10.17521/cjpe.2019.0068 WANG Ming-Ming, LIU Xin-Ping, HE Yu-Hui, ZHANG Tong-Hui, WEI Jing, Chelmge , SUN Shan-Shan.
How enclosure influences restored plant community changes of different initial types in Horqin Sandy Land .
Chinese Journal of Plant Ecology , 2019, 43(8): 672-684. DOI:
10.17521/cjpe.2019.0068 封育恢复在一定时期内通过排除家畜采食、践踏等干扰使群落营养物质得以贮藏, 进而促进植被生产力恢复和群落自然更新, 由于其投资少、见效快, 已成为当前退化生态系统恢复与重建的重要措施, 并为世界各国所广泛采用(
Wairore et al ., 2016 )。封育为退化生态系统所带来的生态效应受许多因素的影响, 地区、封育时间以及封育方式都会使生态系统产生不同甚至相反的响应, 因此退化生态系统各要素对封育的响应机制、适宜的封育时间尺度以及封育方式等研究都受到国内外****的广泛关注(
闫玉春等, 2009 )。
科尔沁沙地作为中国北方最严重的一个沙漠化区域(
刘新民等, 1999 ), 封育禁牧被广泛用来恢复退化沙地植被, 同时也有许多研究对封育所产生的生态效应做了评述。如退化沙质草地封育后群落生物量响应最为敏感, 在较短时间内群落生物量迅速恢复, 同时凋落物量也显著增加(
苗仁辉等, 2013 ;
赵丽娅等, 2017 ;
吕朋等, 2018 )。随着封育时间的延长, 群落物种组成开始发生变化, 流动沙丘封育后沙蓬(
Agriophyllum squarrosum )作为先锋物种首先建立种群, 使沙丘流沙面积减小, 之后以差不嘎蒿(
Artemisia halodendron )为主的多年生灌木通过无性繁殖的方式开始在沙丘定植, 差不嘎蒿等灌木的出现逐渐营造出养分和水分相对富集的灌丛微生境, 进而为禾本科植物的种子萌发提供一个良好的环境(
Su et al ., 2005 )。随着植物群落的变化, 土壤种子库也随之改变(
赵丽娅等, 2006 ;
赵盼盼等, 2018 ), 并且随着流动沙丘的固定和生境的改善, 植物群落和土壤种子库之间的联系变得更加紧密(
Wang et al ., 2019 )。封育对土壤环境也有很大的改善, 对封育和放牧样地土壤性质进行比较, 发现封育样地土壤容重、紧实度及粗颗粒含量显著小于自由放牧样地(
单贵莲等, 2009 ), 而封育样地土壤有机质、全氮、有效氮和有效钾含量显著高于放牧样地(
张建鹏等, 2017 )。封育年限长短也是影响植物群落恢复的重要因素, 适当封育有利于植被恢复, 但过度封育反而会引起植被退化,
高凯等(2013) 在内蒙古草原的研究发现, 封育年限超过14年会使多年生禾草以及灌木和半灌木优势度显著降低, 而多年生杂类草优势度出现不同程度的升高。
尽管已经有许多研究对科尔沁沙地封育恢复的生态效应进行评价和研究, 但上述研究大多是通过对相同或相似的封育或放牧样地一次性调查取样来探讨封育对植物群落以及对土壤环境的影响, 而对不同类型沙地长期封育恢复过程中连续的植物群落特征变化及其影响因素并不清楚。本研究基于连续13年对科尔沁沙地3种长期封育沙地的植物群落的调查, 结合2008和2017年2期土壤种子库和养分监测数据以及多年气象数据, 分析3种沙地植物群落特征变化及其影响因素, 旨在揭示沙漠化土地封育恢复过程中植物群落变化的内在机制及其影响因子, 为退化土地植被恢复和生态重建提供科学指导。
1 材料和方法 1.1 研究区概况 研究区位于内蒙古自治区科尔沁沙地(42.92° N, 120.70° E), 属于半干旱陆地季风气候, 夏季高温, 冬季寒冷。年降水量360 mm, 75%的降水发生在6-9月。年平均气温约为6.4 ℃, 月平均气温在23.5 ℃ (7月)到-16.8 ℃ (1月)之间(
Zuo et al ., 2017 )。土壤类型主要有风沙土、草甸土、沙质栗钙土和沼泽土。地貌特征为固定沙丘、半固定沙丘、流动沙丘和面积不等的平缓沙质草地以及低洼地交错分布。根据奈曼沙漠化研究站2005-2018年间未发表的观测数据, 该区域地下水平均埋深12-15 m。植被斑块化分布和空间异质性较强, 植被盖度较小的流动沙丘上一年生先锋物种沙蓬为优势种, 半固定和固定沙丘上差不嘎蒿和小叶锦鸡儿(
Caragana microphylla )等灌木为优势种。盖度较高的沙质草地以菊科一二年生物种黄蒿(
Artemisia scoparia )以及多年的芦苇(
Phragmites australis )和白草(
Pennisetum flaccidum )为优势物种(
Zuo et al ., 2012 )。
1.2 实验设计与数据采集 自2005年起在科尔沁沙地选择长期封育的沙质草地(1997年封育, 面积34 500 m
2 )、固定沙丘(1985年封育, 面积20 000 m
2 )以及流动沙丘(2005年封育, 面积7 000 m
2 )为研究样地(
图1 ), 每个样地布设10个1 m × 1 m的样方, 每年8月下旬调查草本植物群落物种组成、高度、盖度、密度以及地上生物量, 地上生物量采用刈割法, 采样后烘干称质量; 另外设置5个5 m × 5 m的样方用于灌木和半灌木群落物种组成、高度、盖度、密度以及地上生物量调查, 其中灌木地上生物量测定采用标准枝法。每个样地布设3个土壤水分中子仪(CNC503B(DR), 北京超能科技公司), 用于土壤水分测定。土壤样品采集于2008和2017年的8月下旬, 在每个草本群落调查样方内用直径3 cm的土钻取0-20 cm深度的土壤样品, 每个样方3个重复, 用于测定土壤有机碳、全氮、有效氮、有效磷以及有效钾含量。土壤有机碳测定采用重铬酸钾容量法-外加热法, 土壤全氮测定采用电位法, 有效氮测定采用碱扩散法, 有效磷测定采用碳酸氢钠浸提-钼锑抗比色法, 土壤有效钾测定采用乙酸铵浸提-火焰光度法(
鲁如坤, 1999 )。土壤种子库样品采集于2008和2017年10月中旬, 每个沙地内草本植物群落调查样方旁设置1 m × 1 m的样方, 用土壤种子采样器在每个样方内采取深度为 5 cm, 面积为20 cm × 20 cm的原状土体装入布袋, 带回试验室采用室内萌发实验测定土壤种子库。多年气温和降水数据来源于奈曼沙漠化研究站自动气象观测站。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1科尔沁沙地地理位置及研究样地空间分布。 Fig. 1Geographical location of the Horqin Sandy Land (HSL) and spatial distribution of the sample plots. 1.3 数据统计分析 物种重要值
IV = (
A + B + C + D )/4 × 100%, 式中
A 为相对生物量,
B 为相对多度,
C 相对高度,
D 为相对盖度。群落物种丰富度(
R )、Simpson指数(
C )以及Shannon-Wiener指数(
H )分别计算如下:
(1) $R=S$ (2) $C=1-\underset{1}{\overset{S}{\mathop \sum }}\,P_{i}^{2}$ (3) $H=-\underset{1}{\overset{S}{\mathop \sum }}\,{{P}_{i}}\text{ln}{{P}_{i}}$ 式中,
S 为样方物种总数,
Pi 为第
i 物种密度占群落总密度的比例。用单因素方差分析来检验不同年份和沙地土壤水分、养分以及土壤有机质的差异性。群落物种丰富度、土壤种子丰富度以及土壤因子作为矩阵(
Plassmann et al ., 2009 ;
Yassir et al ., 2010 ), 应用除趋势对应分析(DCA)法来分析土壤种子库与地上群落之间的相似性, 应用典型相关分析(CCA)来分析植物群落和土壤因子之间的关系。所有统计分析均在R-3.2.2中完成, 其中单因素方差分析用‘agricolae’包实现, DCA和CCA分析以及群落
C 和
H 用‘vegan’包计算, 插图用‘ggplot2’包和Origin 2018绘制。
2 结果和分析 2.1 群落特征和物种组成变化 不同沙地2005-2017年间群落特征变化如
图2 所示。通过回归分析, 发现在13年间固定沙丘群落生物量存在显著下降趋势, 沙质草地和流动沙丘群落生物量变化趋势不明显, 但存在一定的年际波动。3种沙地草本层生物量均占到总生物量的60%以上, 并且其变化与群落生物量变化基本一致, 是引起群落生物量年际变化的主要部分。固定沙丘和沙质草地植被盖度有降低的趋势, 但未达到显著水平(
p < 0.05), 而流动沙丘植被盖度显著增加, 13年间增长超过一倍。群落物种丰富度在沙质草地显著下降, 在固定沙丘显著增加, 在流动沙丘有下降趋势但不显著。群落
C 和
H 在3种沙地均无显著变化趋势, 但年际间波动剧烈。为对比13年间3种沙地群落物种变化, 我们列出了2005和2017年群落物种优势度(
表1 ), 发现流动沙丘群落在2005年沙蓬占据优势地位, 到2017年沙蓬优势度下降, 蒺藜(
Tribulus terrestris )成为新的优势物种, 群落物种也由10种增长到14种; 固定沙丘2005年差不嘎蒿优势度最高, 2017年其优势度大幅下降, 群落物种减少4种; 沙质草地2005年多年生芦苇、花苜蓿(
Medicago ruthenica )以及白草等优势度较高, 而2017年多年生草本牻牛儿苗(
Erodium stephanianum )为优势种, 群落物种数也减少4种, 整体而言群落优势种年际间不断更替, 群落物种数也变化较大。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2科尔沁沙地不同类型封育沙地植物群落特征年际变化(平均值±标准偏差)。FD, 固定沙丘; MD, 流动沙丘; SG, 沙质草地。C, Simpson指数; H, Shannon-Wiener指数; k , 线性回归方程斜率; p , 显著性水平(p < 0.05变化趋势明显)。图中仅列出了置信水平较高的k, R 2 和p 值。 Fig. 2Interannual changes of plant community features in different enclosure types of sandy lands in the Horqin Sandy Land (mean ± SD ). FD, fixed dune; MD, mobile dune; SG, sandy grassland. C, Simpson index; H, Shannon-Wiener index; k , regression line slope; p , statistical significance (p < 0.05 indicates the change trend is significant). Only the k, R 2 and p values with higher confidence levels are listed in the figure. Table 1 表1 表1 2005和2017年科尔沁沙地不同类型封育沙地植物群落物种组成和物种优势度
Table 1
Species composition and dominance of plant community in different enclosure types of sandy lands in 2005 and 2017 物种 生活型 2005 2017 MD FD SG MD FD SG 沙蓬 Agriophyllum squarrosum AF 26.91 0 0 9.60 0 0 苦苣菜 Sonchus oleraceus AF 10.22 0 0 2.10 0 0 旋覆花 Inula japonica PF 9.26 0 0 8.75 0 0 狗尾草 Setaria viridis AG 8.65 2.81 1.52 7.45 3.72 6.72 蒺藜 Tribulus terrestris AF 5.24 1.00 0 15.77 0 0.92 地梢瓜 Cynanchum thesioides PF 5.48 0.98 0.84 1.47 2.18 1.88 大果虫实 Corispermum macrocarpum AF 5.32 1.49 0 3.30 2.12 0 差不嘎蒿 Artemisia halodendron SH 3.57 11.88 0 5.99 3.20 0 雾冰藜 Bassia dasyphylla AF 2.69 1.43 0 2.20 1.52 0 马唐 Digitaria sanguinalis AG 2.45 3.43 0.92 5.95 2.80 0 白草 Pennisetum flaccidum PG 0 1.92 4.27 0 0 10.55 猪毛菜 Salsola collina AF 0 0.85 0.85 2.52 7.50 2.17 花苜蓿 Medicago ruthenica PL 0 11.11 4.41 0 10.50 0 糙隐子草 Cleistogenes squarrosa PG 0 0 3.58 0 0 3.77 大籽蒿 Artemisia sieversiana AF 0 0 2.92 0 0 0 地锦 Euphorbia humifusa AF 0 13.27 0 0.94 1.99 0.85 二裂委陵菜 Potentilla bifurca AF 0 0 0.71 0 0 0 九顶草 Enneapogon desvauxii AG 0 0 0 0 0 1.90 兴安乌胡枝子 Lespedeza davurica SH/PL 0 1.39 2.74 0 2.24 2.83 虎尾草 Chloris virgata AG 0 0 1.19 0 0 0 画眉草 Eragrostis pilosa AG 0 3.41 0 1.72 9.99 0 黄蒿 Artemisia scoparia AF 0 0 22.09 0 6.53 6.11 灰绿藜 Chenopodium glaucum AF 0 2.46 0.69 0 0 0 鸡眼草 Kummerowia striata AL 0 0 1.54 0 0 0 尖头叶藜 Chenopodium acuminatum AF 0 0 0 3.54 4.01 4.94 芦苇 Phragmites australis PG 0 0 9.56 0 0 6.36 少花米口袋 Gueldenstaedtia verna PL 0 0 1.58 0 0 0 三芒草 Aristida adscensionis AG 0 3.07 4.37 0 0 0 砂蓝刺头 Echinops gmelinii AF 0 1.42 0 0 8.62 0 牻牛儿苗 Erodium stephanianum AF 0 0 1.67 0 0 12.76 中华苦荬菜 Ixeris chinensis AF 0 2.17 0 0 0 0 小叶锦鸡儿 Caragana microphylla SH/PL 0 2.46 0 0 0 0 独行菜 Lepidium apetalum AG 0 0 0 0 0 5.00 总计 Total - 10 18 18 14 14 13
FD, fixed dune; MD, mobile dune; SG, sandy grassland. AF, annual forbs; AG; annual grasses; AL, annual legumes; PF, perennial forbs; PG, perennial grasses; PL, perennial legumes; SH, shrubs or subshrubs. FD, 固定沙丘; MD, 流动沙丘; SG, 沙质草地。AF, 一年生杂类草; AG, 一年生禾草; AL, 一年生豆科植物; PF, 多年生杂类草; PG, 多年生禾草; PL, 多年生豆科植物; SH, 灌木或半灌木。 新窗口打开 |
下载CSV 2.2 群落生活型功能群重要值年际变化 群落生活型功能群重要值年际变化如
图3 所示。流动沙丘各功能群重要值在年际间波动变化, 但没有显著的趋势, 各功能群间也不存在显著相关关系, 但一年生杂类草重要值远大于其他功能群, 占据优势地位(
表2 )。固定沙丘也是一年生杂类草占据优势地位, 并且其重要值整体存在增加趋势, 多年生杂类草重要值也有增加趋势, 而灌木和半灌木以及多年生豆科植物重要值呈下降趋势, 多年生草本植物重要值呈波动变化。相关分析显示一年生杂类草重要值与灌木、半灌木以及多年生豆科植物重要值之间显著负相关。沙质草地也是一年生杂类草优势度最高, 并且其重要值年际间波动剧烈, 但不存在显著变化趋势。一年生禾草重要值与一年生杂类草重要值年际间消长变化, 并且二者也显著负相关, 表明这两类功能群之间存在补偿效应。多年生禾草重要值也与一年生杂类草显著负相关, 并且其重要值有显著下降趋势。多年生草本优势度变化与多年生禾草基本一致。灌木和半灌木、多年生杂类草重要值相对稳定, 与其他功能群相关性不显著。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3科尔沁沙地不同类型封育沙地植物群落功能群重要值年际变化。FD, 固定沙丘; MD, 流动沙丘; SG, 沙质草地。AF, 一年生杂类草; AG, 一年生禾草; PF, 多年生杂类草; PG, 多年生禾草; PH, 多年生草本植物, PL, 多年生豆科植物; SH, 灌木或半灌木。k , 回归线斜率。图中仅列出了达到显著性水平(p < 0.05)的检验参数值。 Fig. 3Interannual variation of the important value of plant community functional groups in different enclosure types of sandy lands in Horqin Sandy Land. FD, fixed dune; MD, mobile dune; SG, sandy grassland. AF, annual forbs; AG; annual grasses; PF, perennial forbs; PG, perennial grasses; PH, perennial grass layer; PL, perennial legumes; SH, shrubs or subshrubs. k , regression line slope. Only the test parameter values that reach the level of significance (p < 0.05) are listed in this figure. Table 2 表2 表2 科尔沁沙地不同类型封育沙地植物群落功能群重要值之间的Pearson相关系数
Table 2
Pearson’s correlation coefficient between important values of community functional groups in different types enclosed sandy land in Horqin Sandy Land 沙地 Sandy land 功能群 Functional group R p 沙地 Sandy land 功能群 Functional group R p MD AF vs AG -0.28 0.35 FD PL vs SH 0.33 0.28 MD AF vs SH -0.21 0.49 SG AF vs AG -0.72 <0.01 MD AG vs SH -0.11 0.72 SG AF vs PF -0.23 0.44 FD AF vs AG -0.28 0.36 SG AF vs PG -0.81 <0.01 FD AF vs PF -0.13 0.67 SG AF vs PL -0.17 0.58 FD AF vs PL -0.65 <0.05 SG AG vs PF 0.37 0.21 FD AF vs SH -0.71 <0.01 SG AG vs PG 0.22 0.47 FD AG vs PF -0.20 0.51 SG AG vs PL -0.14 0.64 FD AG vs PL -0.03 0.93 SG PF vs PG 0.14 0.64 FD AG vs SH -0.17 0.57 SG PF vs PL -0.04 0.90 FD PF vs PL -0.14 0.65 SG PG vs PL 0.53 0.14 FD PF vs SH -0.18 0.57 SG
FD, fixed dune; MD, mobile dune; SG, sandy grassland. AF, annual forbs; AG; annual grasses; PF, perennial forbs; PG, perennial grasses; PL, perennial legumes; SH, shrubs or subshrubs. FD, 固定沙丘; MD, 流动沙丘; SG, 沙质草地。AF, 一年生杂类草; AG, 一年生禾草; PF, 多年生杂类草; PG, 多年生禾草; PL, 多年生豆科植物; SH, 灌木或半灌木。 新窗口打开 |
下载CSV 2.3 土壤有机碳、养分和种子库变化 比较2008和2017年土壤养分测定结果(
表3 )发现, 10年间3种沙地仅有土壤有效氮和有效磷含量显著增加, 而土壤有机质碳、全氮以及有效钾含量在3种沙地均无显著变化。同期的土壤种子库比较发现种子密度在3种沙地间均无显著变化; 种子丰富度在流动沙丘有所提高, 而在固定沙丘和沙质草地有所下降, 这与植物群落物种变化趋势相同。不同沙地间土壤有机质在沙质草地的含量高于固定沙丘和流动沙丘, 但流动沙丘和固定沙丘之间无显著差异, 土壤全氮分布与有机质基本相同, 土壤有效氮含量随着沙地生境的逐渐改善而升高, 且在3种沙地间存在显著差异; 土壤有效磷含量在3种沙地间不存在显著差异, 有效钾在沙质草地的含量显著高于流动沙丘和固定沙丘。整体上随着沙地生境恢复, 土壤有效养分含量逐渐增加, 但积累速率缓慢, 土壤种子密度变化不大, 种子丰富度变化与群落变化基本一致。土壤水分含量在3类沙地间也显著不同, 其中沙质草地最高, 流动沙丘居中, 固定沙丘最低。
Table 3 表3 表3 科尔沁沙地不同类型封育沙地2008和2017年土壤养分、水分、有机质含量以及种子密度和丰富度比较(平均值±标准偏差)
Table 3
Multiple comparisons of soil nutrients, soil moisture, soil organic matter and soil seed density, seed richness in different enclosure types of sandy lands in the Horqin Sandy Land in 2008 and 2017 (mean ± SD ) 土壤因子 Soil factor MD FD SG 2008年 2017年 2008年 2017年 2008年 2017年 SOM (g·kg-1 ) 0.53 ± 0.08a 0.42 ± 0.05b 1.13 ± 0.39a 1.05 ± 0.36a 5.64 ± 2.18a 4.05 ± 0.51a TN (g·kg-1 ) 0.03 ± 0.05a 0.024 ± 0.01a 0.068 ± 0.02a 0.072 ± 0.06a 0.34 ± 0.11a 0.35 ± 0.07a AN (mg·kg-1 ) 5.35 ± 1.25b 8.80 ± 2.52a 6.35 ± 3.20b 11.28 + 3.60a 22.51 ± 2.98b 29.87 ± 4.89a AP (mg·kg-1 ) 4.14 ± 0.65b 9.88 ± 0.17a 3.88 ± 0.92b 9.87 ± 0.07a 3.22 ± 1.32b 9.43 ± 0.11a AK (mg·kg-1 ) 54.67 ± 7.08a 43.05 ± 4.60b 64.58 ± 9.80a 65.61 ± 11.30a 116.42 ± 23.30a 112.47 ± 20.20a SSD (粒·m-2 ) 492.0 ± 204.40a 670.0 ± 268.6a 2 669 ± 1 108a 1 860 ± 687a 3 344 ± 1 378a 4 072 ± 1 639a SSR 4.0 ± 1.20b 5 ± 1.30a 10 ± 0.50a 8 ± 1.33b 12 ± 2.10a 9 ± 1.24b SM (%) 2.43 ± 0.32b 2.00 ± 0.07c 3.72 ± 0.10a
FD, fixed dune; MD, mobile dune; SG, sandy grassland. AK, available potassium; AN, available nitrogen; AP, available phosphorus; SM, soil moisture; SOM, soil organic matter; SSD, soil seed density; SSR, soil seed richness; TN, total nitrogen. Lowercase letters indicate the difference in 95% confidence of soil environmental factors in three types of sandy land between 2008 and 2017. The last row shows the difference in soil moisture among the three types of sites. FD, 固定沙丘; MD, 流动沙丘; SG, 沙质草地。AK, 土壤有效钾; AN, 土壤有效氮; AP, 土壤有效磷; SM, 土壤水分; SOM, 土壤有机质; SSD, 土壤种子密度; SSR, 种子丰富度; TN, 土壤全氮。小写字母表示3种沙地2008年和2017年土壤环境因子之间95%置信度下的差异性。最后一行为3种样地间土壤体积含水量的差异。 新窗口打开 |
下载CSV 2.4 降水和气温对群落变化的影响 年降水量和年平均气温与3种沙地群落生物量和物种丰富度之间的关系如
表4 所示。观测期间在3种沙地群落生物量与年降水量和生长季(4-9月)降水量间均不存在显著的线性关系, 但随着沙地恢复, 二者之间的回归斜率逐渐增大。3种沙地物种丰富度也与降水不存在显著相关性。年平均气温与群落生物量和物种丰富度之间也不显著相关, 但与生物量之间的抛物线关系优于线性关系。进一步对2013- 2017年群落月生物量与累积降水量、生长季逐月积温之间做回归分析(
图4 ), 发现降水和生长季积温均与3种沙地群落月生物量之间存在显著正相关关系, 并且回归线斜率随着沙地的恢复而增加。气温、降水与月累积生物量之间的多变量回归结果也显示二者在沙质草地解释率达到68% (
R 2 = 0.68,
p < 0.01), 在固定沙丘解释率为36% (
R 2 = 0.36,
p < 0.01), 流动沙丘为60% (
R 2 = 0.60,
p < 0.01), 反映出不同生物量积累对水热条件的变化存在不同的响应, 其中沙质草地响应最为敏感。整体上年降水量和气温对群落生物量和物种丰富度年际变化贡献不大, 但却是影响年内生物量积累的主要因素。
Table 4 表4 表4 科尔沁沙地不同类型封育沙地植物群落生物量和物种丰富度与气候因子之间关系
Table 4
Relationships between plant community biomass and species richness with climatic factors in different habitats in different types enclosed sandy land in Horqin Sandy Land 气候因子 沙地 生物量 Biomass (g·m-2 ) 物种丰富度 Spices richness 方程 Equation R 2 p 方程 Equation R 2 p 年降水量 MD y = -0.1x + 17.080.3 0.98 y = -0.35lg x + 13.340.03 0.26 FD y = 0.09x + 55.960.22 0.45 y = 14.78lg x - 19.670.15 0.10 SG y = 0.21x + 75.110.45 0.13 y = 0.19lg x + 13.500.08 0.79 生长季降水量 MD y = 0.03x + 8.390.13 0.66 y = -0.35lg x + 13.350.03 0.27 FD y = 0.09x + 58.260.21 0.48 y = 14.53lg x - 18.320.15 0.11 SG y = 0.17x + 93.170.32 0.28 y = 0.19lg x + 13.50.08 0.79 年平均气温 MD y = 1.2x 2 - 22.64x + 117.040.24 0.23 y = -0.35x + 13.340.03 0.26 FD y = 2.42x 2 - 39.89x + 234.260.42 0.06 y = -0.09x + 16.000.09 0.84 SG y = 2.65x 2 - 44.46x + 306.240.19 0.35 y = 0.19x + 1.50.08 0.79
x , climatic factors; y , community features. FD, fixed dune; MD, mobile dune; SG, sandy grassland.x , 气候因子; y , 群落特征。FD, 固定沙丘; MD, 流动沙丘; SG, 沙质草地。新窗口打开 |
下载CSV 图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4科尔沁沙地不同类型封育沙地植物群落生物量与生长季降水累积量、积温之间的关系。FD, 固定沙丘; MD, 流动沙丘; SG, 沙质草地。 Fig. 4Relationships among plant community biomass, and growing season precipitation, growing season accumulated temperature in different enclosure types of sandy land in the Horqin Sandy Land. FD, fixed dune; MD, mobile dune; SG, sandy grassland. 2.5 群落与土壤因子之间的关系 对群落物种和土壤种子库物种做DCA分析, 土壤因子与群落物种、土壤种子库之间做CCA分析, 得到二维排序图(
图5 )。DCA分析图中群落物种分布和土壤种子库物种分布存在很好的一致性(
图5 A), 说明土壤种子库与地上群落之间密切相关。群落物种与土壤种子的CCA图(
图5 B)中, 不同沙地群落物种分布在不同的象限, 其中沙质草地物种集中分布在第一轴上下, 并且第一轴与土壤有机质、全氮、有效氮以及有效钾含量之间的夹角很小, 反映出沙质草地群落物种主要受土壤养分的影响。固定沙丘群落物种集中分布在第三象限, 且与第二轴距离较远, 但与土壤水分与相关性较强, 表明固定沙丘群落在一定程度上受土壤水分限制, 实际监测也发现固定沙丘土壤水分含量最低。流动沙丘群落物种集中分布在第二象限, 且与各土壤因子之间的连线都较远。土壤种子库物种与土壤因子之间的CCA分析结果(
图5 C)与植物群落和土壤因子的CCA分析结果基本一致。
图5 新窗口打开 |
下载原图ZIP |
生成PPT 图5科尔沁沙地不同类型封育沙地植物群落物种、土壤种子库、土壤养分之间的除趋势对应分析(DCA)和典型相关分析(CCA)。A, 群落物种和土壤种子库的DCA分析。B, 群落物种和土壤因子的CCA分析。C, 土壤种子库物种和土壤因子的CCA分析。FD, 固定沙丘; MD, 流动沙丘; SG, 沙质草地。AK, 土壤有效钾; AN, 土壤有效氮; AP, 土壤有效磷; SM, 土壤水分; SOM, 土壤有机质; TN, 土壤全氮。 Fig. 5Detrended correspondence analysis (DCA) and canonical correspondence analysis (CCA) among plant community species, soil seed bank and soil nutrients. A, the DCA analysis of community species and soil seed bank. B, the CCA analysis of community species and soil nutrients. C, the CCA analysis of soil seed bank species and soil nutrients. FD, fixed dune; MD, mobile dune; SG, sandy grassland. AK, available potassium; AN, available nitrogen; AP, available phosphorus; SM, soil moisture; SOM, soil organic matter; TN, total nitrogen. Veg, vegtation. 3 讨论 3.1 群落变化特征 退化沙地恢复的速度和演替方向一方面取决于退化程度和干扰的消除程度, 另一方面取决于土壤、气候等环境条件(
赵哈林等, 2003 )。本研究中流动沙丘13年封育期间植被盖度和物种丰富度显著增加, 但群落生物量没有显著变化, 整体上群落在逐渐恢复, 但速度缓慢, 群落生产力有限。许多研究发现干扰退化草地在短期封育下可以快速恢复(
赵哈林等, 2009 ;
苗仁辉等, 2013 ;
吕朋等, 2018 ), 其根本原因是现存群落具有拓殖能力和退化群落资源过剩(
王炜等, 1996 ), 但对于流动沙丘, 群落资源处于极其匮乏的状态, 现存群落拓殖能力有限, 土壤种子库基础也相对薄弱, 因此植被恢复受到很大限制。另外植被恢复也受时间积累效应的影响, 物种要进入某一环境空间并建立种群, 需要有足够的时间去完成其繁殖体从种源地到目标空间的传播、散布和在目标空间的定居、繁殖过程, 这一过程进行的速度直接影响物种多样性恢复的进度(
张继义等, 2004 ), 本研究中流动沙丘封育仅13年, 植物建群时间有限, 因此时间积累可能相对不足, 植被恢复程度有限。有研究认为在降水条件较好的半干旱沙区, 采取封沙育草措施后10-15年的时间, 植被能够自然恢复(
张华等, 2003 ), 然而本研究的结果显示, 流动沙丘恢复到灌木、半灌木或多年生草本优势度较高的固定沙丘群落需要更长的时间, 因此封育年限的设定应综合考虑植被退化程度、气候条件以及土壤环境等因素。
固定沙丘群落恢复演替过程中一年生杂类草群落属于演替的早期阶段, 多年生植物占优势的群落处于演替的中期阶段, 随着沙丘群落演替的推进, 一年生杂类草优势度会逐渐下降, 多年生草本优势度会逐渐增加, 相应的豆科物种优势度也会逐渐上升(
赵哈林等, 2003 )。而在本研究13年的观测期间, 固定沙丘群落一年生和多年生杂类草优势度有显著增加的趋势, 而灌木和半灌木以及多年生豆科植物优势度有显著下降的趋势, 这与正常的固定沙丘群落恢复演替的方向相反, 这说明封育33年的固定沙丘群落存在逆行演替的现象。另外, 一年生杂类草重要值与灌木和半灌木以及多年生豆科植物重要值显著负相关, 但与其他功能群重要值的相关性不显著, 群落生物量和物种丰富度也存在下降趋势, 反映出一年生杂类草优势度的增长抑制了其他功能群的生长, 这也表明过度封育不利于固定沙丘植被 恢复。
退化沙质草地封育恢复是群落生物量和物种多样性显著增加, 多年生植物优势度提高, 群落结构逐渐趋于稳定, 群落生态功能不断增强的过程(
张继义等, 2004 )。在本研究中封育21年的沙质草地群落生物量不存在显著的变化趋势, 但群落物种丰富度显著下降, 多年生禾草功能群重要值也不断降低, 这也与正常退化草地封育恢复过程中的群落演替相反, 反映出固定沙丘群落在观测期间存在退化趋势。根据生态系统可持续性原理(
戎郁萍等, 2004 ), 退化草地的封育时间不应是无限期的, 我们的研究结果也显示封育时间过长不仅不利于群落恢复, 反而会导致群落发生逆行演替, 因此封育一段时间后应适当实施刈割、放牧等干扰措施, 刺激牧草分蘖, 促进植物再生, 改善沙质草地质量(
程积民等, 1995 ), 也有研究发现轻度放牧是内蒙古草原最有利的草地利用方式(
Gao et al ., 2015 )。
3.2 群落变化与气候因子之间的关系 大多数陆地生态系统天然植物群落都是特定气候条件下的产物(
赵哈林等, 2003 ), 气候条件的变化, 特别是气温和降水的变化会引起植物群落特征的变化(
Liu & Lei, 2015 ;
Liu et al ., 2016 )。
赵哈林等(2006) 认为气候是我国北方植被恢复的外在驱动力, 决定着演替的方向和终点。然而我们13年的观测结果显示群落生物量和物种丰富度与降水和气温之间不存在显著关系, 但气温和降水与生物量的线性回归斜率随着沙地的恢复而逐渐增大, 斜率的大小反映出植物群落对降水年际变化的响应敏感性(
Sala et al ., 2012 ), 在一定程度上也反映出群落对降水的利用效率(
Bai et al ., 2008 )。随着沙地生境恢复, 群落对年际降水波动的响应敏感性逐渐增加, 降水利用效率也相应地提升, 这种变化可能与资源的可利用性密切相关。许多研究发现土壤氮等资源的可利用性越高, 植被对降水变化的响应越敏感(
Wilcox et al ., 2016 ,
2017 ;
Knapp et al ., 2017 ), 在科尔沁沙地大尺度的研究也发现土壤养分梯度是影响植物群落分布最主要的因素(
左小安等, 2007 )。本研究中沙质草地土壤环境条件最优, 固定沙丘次之, 流动沙丘最差, 因此这种资源分布的差异导致不同沙地群落对降水变化的响应不同。群落组成也会影响群落对降水变化的响应。通过对生活型功能群重要值的年变化分析发现沙质草地群落不同功能群之间存在补偿效应, 这种补偿效应的存在可以有效地减缓降水等气候因子变化带来的影响。
Bai等(2004) 在内蒙古草原的研究也发现群落功能群之间存在补偿效应, 这种效应的存在更有利于群落稳定性的维持。这些因素的综合作用可能导致了群落生物量年变化对降水波动响应微弱。然而需要注意的是许多关于降水与群落生产力之间关系的结论都是在长时间尺度或者大空间尺度统计得来的(
Knapp et al ., 2017 ;
Wilcox et al ., 2017 ), 而本研究仅是在时间跨度为13年的小区域中开展的, 因此还需要更长时间的观测资料来研究群落对降水变化的响应。年内生物量积累与积温和累积降水量显著相关, 甚至在沙质草地二者对生物量年内积累的解释率达到68%, 因此积温和累积降水量是年内生物量积累的重要驱动因素, 这与植被生长过程中的水热需求规律一致。
3.3 群落物种与土壤环境之间的关系 气候是植被恢复的外在驱动力, 决定群落演替的方向和终点, 而植被的自我修复是内在驱动力, 决定着植物恢复的起始速度和演替速度(
赵哈林等, 2006 )。植被自我修复的基础是土壤种子库,
Plassmann等(2009) 研究发现相比于当前群落, 土壤种子库更能反映演替初期的植被组成, 但在我们的研究中发现种子丰富度变化与群落物种丰富度变化相似, DCA分析也发现地上植物群落物种和土壤种子库之间有很高的相似性, 这反映出科尔沁沙地植被恢复和土壤种子库变化存在较强的关联性。
Wang等(2019) 在科尔沁沙地的研究也表明植被恢复过程中地上群落和土壤种子库之间存在紧密联系, 并且随生境恢复二者之间的联系更加紧密。
退化沙地土壤环境的恢复速率直接影响植被的演替进程, 土壤环境的改善是植被对土壤作用的结果, 但又反馈于植被的演替(
赵哈林等, 2006 )。我们的监测结果显示3种封育沙地在10年内仅有土壤有效氮和有效磷含量增加, 并且增加幅度不大, 土壤养分缓慢的积累速率对植被恢复有很大的限制作用, 尤其是在养分含量极低的流动沙丘这种限制作用可能会更大, 因此封育13年的流动沙丘群落生物量没有显著增加。另外, 土壤养分的空间异质性直接影响植被分布模式(
Zuo et al ., 2014 )。本研究3种沙地土壤环境存在显著差异, 群落物种与土壤因子之间的CCA分析也显示不同沙地群落物种受不同因素的影响, 其中沙质草地植物群落物种与土壤养分之间相关性很强, 固定沙丘群落物种和土壤水分联系紧密, 而流动沙丘群落物种与各土壤因子并不相关。3类沙地中沙质草地土壤养分和水分条件最优, 其相应的群落生物量和物种密度也最高, 但沙质草地多为草本植物, 密布的根系层对降水存在滞留作用(
牛存洋等, 2015 ), 另外草本植物相比于灌木和半灌木耗水相对较少, 并且较高的植被盖度产生的地表遮阴效果也能有效减弱土壤蒸发(
常昌明等, 2016 ), 这些因素的综合作用使其土壤水分含量最高, 进而使水分对群落的限制作用可能弱于土壤养分。而在固定沙丘土壤水分含量最低, 这是因为在固定沙丘优势物种主要为根系较深、耗水强烈的差不嘎蒿、小叶锦鸡儿等灌木, 使得土壤水分消耗强烈, 导致群落主要受土壤水分的限制。流动沙丘由于表面干沙层的存在抑制了土壤水分的蒸发(
段利民, 2011 ), 其土壤水分含量反而高于固定沙丘, 但其土壤养分和种子库非常贫瘠, 因此二者可能共同限制了群落的恢复。
4 结论 我们对3种典型封育沙地连续13年的植物群落调查发现, 封育13年的流动沙丘群落生物量没有显著增加, 但物种丰富度和植被盖度增加了一倍; 封育33年的固定沙丘群落杂类草优势度显著上升, 灌木和半灌木以及豆科植物优势度显著下降, 群落存在逆行演替趋势, 另外群落生物量和物种丰富度也存在降低趋势; 封育21年的沙质草地群落多年生禾草优势度和物种丰富度存在显著下降趋势, 表明群落存在退化现象, 但一年生禾草和一年生杂类草功能群之间存在补偿效应, 使得群落生产力和物种多样性相对稳定。封育是退化沙地植被恢复的有效措施, 但过度的封育不利于植被的持续恢复, 因此适当封育后应进行刈割、放牧等干扰以保持群落持续恢复。土壤环境的改善是群落持续恢复的基础, 我们的研究发现3种封育沙地10年间土壤养分积累速率缓慢, 土壤种子丰富度与群落物种变化一致, 在固定沙丘和沙质草地甚至有退化现象, 并且不同沙地土壤资源分布的差异导致群落对气温和降水变化存在不同的响应, 因此封育年限和策略的设定应全面考虑沙地植被退化程度、土壤养分状况、土壤种子库基础以及气候条件等因素的综合影响。
[1] Bai Y Han X Wu J Chen Z Li L 2004 ). Ecosystem stability and compensatory effects in the Inner Mongolia grasslandNature, 431, 181 -184 . [本文引用: 1] [2] Bai YF Wu JG Xing Q Pan QM Huang JH Yang DL Han XG 2008 ). Primary production and rain use efficiency across a precipitation gradient on the Mongolia plateauEcology, 89, 2140 -2153 . [本文引用: 1] [3] Chang CM Niu JM Wang H Zhang Q Dong JJ Kang SR Han F Na RS 2016 ). Dynamic change of soil moisture and its response to rainfall in a Stipa klemenzii steppeArid Zone Research, 260 -265 . [本文引用: 1] [ 常昌明 , 牛建明 , 王海 , 张庆 , 董建军 , 康萨如拉 , 韩芳 , 那日苏 (2016 ). 小针茅荒漠草原土壤水分动态及其对降雨的响应干旱区研究 , 33, 260 -265 .] [本文引用: 1] [4] Cheng JM Zou HY Akio H 1995 ). The rational utilization of grassland and successional course of grassland vegetation in the Loess PlateauActa Prataculturae Sinica, 4(4 ), 17 -22 . [本文引用: 1] [ 程积民 , 邹后远 , 本江昭夫 (1995 ). 黄土高原草地合理利用与草地植被演替过程的实验研究草业学报 , 4(4 ), 17 -22 .] [本文引用: 1] [5] Duan LM 2011 ). Dynamic Interrelation of Hydrology, Soil and Vegetation in the Horqin Sandy Land with Sand-Meadow Land Features. PhD dissertation, Inner Mongolia Agricultural University, Hohhot. [本文引用: 1] [ 段利民 (2011 ). 科尔沁沙地沙丘-草甸相间地区水文-土壤-植被动态响应关系研究博士学位论文, 内蒙古农业大学, 呼和浩特 .] [本文引用: 1] [6] Gao K Zhu TX Han GD 2013 ). Impact of enclosure duration on plant functional and species diversity in Inner Mongolian grasslandActa Prataculturae Sinica, 22(6 ), 39 -45 . [本文引用: 1] [ 高凯 , 朱铁霞 , 韩国栋 (2013 ). 围封年限对内蒙古羊草-针茅典型草原植物功能群及其多样性的影响草业学报 , 22(6 ), 39 -45 .] [本文引用: 1] [7] Gao TM Zhang RQ Yue ZW 2015 ). Influence of enclosure on vegetation productivity and biodiversity on Xilamuren Grassland, Inner Mogolia2015 4th International Conference on Energy and Environmental Protection : 654 -657 . . Cited: 2019-03-20 URL [本文引用: 1] [8] Knapp AK Ciais P Smith MD 2017 ). Reconciling inconsistencies in precipitation-productivity relationships: Implications for climate changeNew Phytologist, 214, 41 -47 . [本文引用: 2] [9] Liu R Cieraad E Li Y Ma J 2016 ). Precipitation pattern determines the inter-annual variation of herbaceous layer and carbon fluxes in a phreatophyte-dominated desert ecosystemEcosystems, 19, 601 -614 . [本文引用: 1] [10] Liu XM Zhao HL Zhao AF 1999 ). Wind-Sand Environment and Vegetation in Horqin Sandy Land . Science Press, Beijing. 191 -221 . [本文引用: 1] [ 刘新民 , 赵哈林 , 赵爱芬 (1999 ). 科尔沁沙地风沙环境与植被 . 科学出版社, 北京. 191 -221 .] [本文引用: 1] [11] Liu YL Lei HM 2015 ). Responses of natural vegetation dynamics to climate drivers in China from 1982 to 2011Remote Sensing, 7, 10243 -10268 . [本文引用: 1] [12] Lu RK 1999 ). Analytical Method of Soil Agricultural Chemistry . China Agriculture Science and Technique Press, Beijing. [本文引用: 1] [ 鲁如坤 (1999 ). 土壤农业化学分析方法 . 中国农业科技出版社, 北京.] [本文引用: 1] [13] Lü P Zuo XA Yue XY Zhang J Zhao SL Cheng QP 2018 ). Temporal changes of vegetation characteristics during the long-term grazing exclusion in Horqin Sandy LandChinese Journal of Ecology, 37, 2880 -2888 . [本文引用: 2] [ 吕朋 , 左小安 , 岳喜元 , 张晶 , 赵生龙 , 程清平 (2018 ). 科尔沁沙地封育过程中植被特征的动态变化生态学杂志 , 37, 2880 -2888 .] [本文引用: 2] [14] Miao RH Jiang DM Wang YC 2013 ). Change and mechanism of vegetation in the fenced sandy grassland in Horqin Sandy LandArid Zone Research, 30, 264 -270 . [本文引用: 2] [ 苗仁辉 , 蒋德明 , 王永翠 (2013 ). 科尔沁沙质草地封育过程中的植被变化及其机制干旱区研究 , 30, 264 -270 .] [本文引用: 2] [15] Niu CY Alamusa , Liu Y Guo YH Tian YH Wang JL Zhang W 2015 ). The characteristics of sand-fixation plantations roots and soil moisture Horqin Sandy LandJournal of Arid Land Resources and Environment, 29(10 ), 106 -111 . [本文引用: 1] [ 牛存洋 , 阿拉木萨 , 刘亚 , 郭宇航 , 田英华 , 王甲立 , 张巍 (2015 ). 科尔沁沙地固沙植物根系与土壤水分特征研究干旱区资源与环境 , 29(10 ), 106 -111 .] [本文引用: 1] [16] Plassmann K Brown N Jones MLM Edwards-Jones G 2009 ). Can soil seed banks contribute to the restoration of dune slacks under conservation management?Applied Vegetation Science, 12, 199 -210 . [本文引用: 2] [17] Rong YP Zhao ML Han GD 2004 ). Principle and Technology of Grassland Resource Sustainable Utilization . Chemical Industry Press, Beijing. [本文引用: 1] [ 戎郁萍 , 赵萌莉 , 韩国栋 (2004 ). 草地资源可持续利用原理与技术 . 化学工业出版社, 北京.] [本文引用: 1] [18] Sala OE Gherardi LA Reichmann L Jobbágy E Peters D 2012 ). Legacies of precipitation fluctuations on primary production: Theory and data synthesisPhilosophical Transactions of the Royal Society B: Biological Sciences, 367, 3135 -3144 . [本文引用: 1] [19] Shan GL Xu Z Ning F Jiao Y 2009 ). Influence of seasonal on exclosure on plant and soil characteristics in typical steppeActa Prataculturae Sinica, 18, 3 -10 . [本文引用: 1] [ 单贵莲 , 徐柱 , 宁发 , 焦燕 (2009 ). 围封年限对典型草原植被与土壤特征的影响草业学报 , 18, 3 -10 .] [本文引用: 1] [20] Su YZ Zhang TH Li YL Wang F 2005 ). Changes in soil properties after establishment of Artemisia halodendron and Caragana microphylla on shifting sand dunes in semiarid Horqin Sandy Land, Northern ChinaEnvironmental Management 272 -281 . [本文引用: 1] [21] Wairore JN Mureithi SM Wasonga OV Nyberg G 2016 ). Benefits derived from rehabilitating a degraded semi-arid rangeland in private enclosures in West Pokot County, KenyaLand Degradation & Development, 27, 532 -541 . [本文引用: 1] [22] Wang W Liu ZL Hao DY Liang CZ 1996 ). Research on the restoring succession of the degenerated grassland in Inner Mongolia Ⅰ. Basic characteristics and driving force for restoration of the degenerated grasslandActa Phytoecologica Sinica, 20, 449 -459 . [本文引用: 1] [ 王炜 , 刘钟龄 , 郝敦元 , 梁存柱 (1996 ). 内蒙古草原退化群落恢复演替的研究——I. 退化草原的基本特征与恢复演替动力植物生态学报 , 20, 449 -459 .] [本文引用: 1] [23] Wang Y Chu L Daryanto S Lü L Ala MS Wang L 2019 ). Sand dune stabilization changes the vegetation characteristics and soil seed bank and their correlations with environmental factorsScience of the Total Environment, 648, 500 -507 . [本文引用: 2] [24] Wilcox KR Blair JM Smith MD Knapp AK 2016 ). Does ecosystem sensitivity to precipitation at the site-level conform to regional-scale predictions?Ecology, 97, 561 -568 . [本文引用: 1] [25] Wilcox KR Shi Z Gherardi LA Lemoine NP Koerner SE Hoover DL Bork E Byrne KM Cahill Jr J Collins SL Evans S Gilgen AK Holub P Jiang L Knapp AK LeCain D Liang J Garcia-Palacios P Pe?uelas J Pockman WT Smith MD Sun S White SR Yahdjian L Zhu K Luo Y 2017 ). Asymmetric responses of primary productivity to precipitation extremes: A synthesis of grassland precipitation manipulation experimentsGlobal Change Biology, 23, 4376 -4385 . [本文引用: 2] [26] Yan YC Tang HP Xin XP Wang X 2009 ). Advances in research on the effects of exclosure on grasslands,Acta Ecologica Sinica, 29, 5039 -5046 . [本文引用: 1] [ 闫玉春 , 唐海萍 , 辛晓平 , 王旭 (2009 ). 围封对草地的影响研究进展生态学报 , 29, 5039 -5046 .] [本文引用: 1] [27] Yassir I van der Kamp J Buurman P 2010 ). Secondary succession after fire in Imperata grasslands of East Kalimantan, IndonesiaAgriculture, Ecosystems & Environment 172 -182 . [本文引用: 1] [28] Zhang H Fu QK Li FR Shirato Y 2003 ). Features of soil-plant system changes in different restorative stages of degraded sandy grasslandsBulletin of Soil and Water Conservation, 23(6 ), 1 -6 . [本文引用: 1] [ 张华 , 伏乾科 , 李锋瑞 , Shirato Y 2003 ). 退化沙质草地自然恢复过程中土壤-植物系统的变化特征水土保持通报 , 23(6 ), 1 -6 .] [本文引用: 1] [29] Zhang JP Li YQ Zhao XY Zhang TH She QN Liu M Wei SL 2017 ). Effects of exclosure on soil physicochemical properties and carbon sequestration potential recovery of desertified grasslandJournal of Desert Research, 37, 491 -499 . [本文引用: 1] [ 张建鹏 , 李玉强 , 赵学勇 , 张铜会 , 佘倩楠 , 刘敏 , 魏水莲 (2017 ). 围封对沙漠化草地土壤理化性质和固碳潜力恢复的影响中国沙漠 , 37, 491 -499 .] [本文引用: 1] [30] Zhang JY Zhao HL Zhang TH Zhao XY 2004 ). Dynamics of species diversity of communities in restoration processes in Horqin Sandy LandActa Phytoecologica Sinica, 28, 86 -92 . [本文引用: 2] [ 张继义 , 赵哈林 , 张铜会 , 赵学勇 (2004 ). 科尔沁沙地植被恢复系列上群落演替与物种多样性的恢复动态植物生态学报 , 28, 86 -92 .] [本文引用: 2] [31] Zhao HL Okuro T Li YL Zuo XA Huang G Zhou RL 2009 ). Changes of plant community in grazing and restoration processes in Horqin Sandy Land, Inner MongoliaJournal of Desert Research, 29, 229 -235 . [本文引用: 1] [ 赵哈林 , 大黑俊哉 , 李玉霖 , 左小安 , 黄刚 , 周瑞莲 (2009 ). 科尔沁沙质草地植物群落的放牧退化及其自然恢复过程中国沙漠 , 29, 229 -235 .] [本文引用: 1] [32] Zhao HL Su YZ Zhou RL 2006 ). Restoration mechanism of degraded vegetation in sandy areas of northern ChinaJournal of Desert Research, 26, 323 -328 . [本文引用: 3] [ 赵哈林 , 苏永中 , 周瑞莲 (2006 ). 我国北方沙区退化植被的恢复机理中国沙漠 , 26, 323 -328 .] [本文引用: 3] [33] Zhao HL Zhao XY Zhang TH Wu W 2003 ). Desertification Process and Restoration Mechanism in Horqin Sandy Land . China Ocean Press, Beijing. [本文引用: 3] [ 赵哈林 , 赵学勇 , 张铜会 , 吴薇 (2003 ). 科尔沁沙地沙漠化过程及其恢复机理 . 海洋出版社, 北京.] [本文引用: 3] [34] Zhao LY Li ZH Zhao JH Zhao HL Zhao XY 2006 ). Comparison on the difference in soil seed bank between grazed and enclosed grasslands in Horqin Sandy LandJournal of Plant Ecology (Chinese Version )617 -623 . [本文引用: 1] [ 赵丽娅 , 李兆华 , 赵锦慧 , 赵哈林 , 赵学勇 (2006 ). 科尔沁沙质草地放牧和围封条件下的土壤种子库植物生态学报 , 30, 617 -623 .] [本文引用: 1] [35] Zhao LY Zhang XY Xiong BQ Zhang J 2017 ). Influence of fencing and grazing on the soil and standing vegetation changes in Horqin sandy grasslandEcology and Environmental Sciences, 26, 971 -977 . [本文引用: 1] [ 赵丽娅 , 张晓雨 , 熊炳桥 , 张劲 (2017 ). 围封和放牧对科尔沁沙质草地植被和土壤的影响生态环境学报 , 26, 971 -977 .] [本文引用: 1] [36] Zhao PP Li GQ Shao WS Jin CQ 2018 ). Influence of herbivore exclusion on the soil seed bank and the aboveground vegetation characteristics of Agropyron mongolicum dominant desert steppe grasslandActa Prataculturae Sinica, 1 ), 42 -52 . [本文引用: 1] [ 赵盼盼 , 李国旗 , 邵文山 , 靳长青 (2018 ). 围封对荒漠草原区沙芦草群落土壤种子库及地上植被的影响草业学报 , 27(1 ), 42 -52 .] [本文引用: 1] [37] Zuo XA Wang SK Zhao XY Lian J 2014 ). Scale dependence of plant species richness and vegetation-environment relationship along a gradient of dune stabilization in Horqin Sandy Land, Northern ChinaJournal of Arid Land, 6, 334 -342 . [本文引用: 1] [38] Zuo XA Yue XY Lv P Yu Q Chen M Zhang J Luo YQ Wang SK Zhang J 2017 ). Contrasting effects of plant inter- and intraspecific variation on community trait responses to restoration of a sandy grassland ecosystemEcology and Evolution, 7, 1125 -1134 . [本文引用: 1] [39] Zuo XA Zhao XY Zhao HL Li YQ Guo YR Zhao YP 2007 ). Changes of species diversity and productivity in relation to soil properties in sandy grassland in Horqin Sandy LandEnvironmental Science, 28, 945 -951 . [本文引用: 1] [ 左小安 , 赵学勇 , 赵哈林 , 李玉强 , 郭轶瑞 , 赵玉萍 (2007 ). 科尔沁沙质草地群落物种多样性、生产力与土壤特性的关系环境科学 , 28, 945 -951 .] [本文引用: 1] [40] Zuo XA Zhao XY Zhao HL Zhang TH Li YL Wang SK Li WJ Powers R 2012 ). Scale dependent effects of environmental factors on vegetation pattern and composition in Horqin Sandy Land, Northern ChinaGeoderma, 1 -9 . [本文引用: 1] Ecosystem stability and compensatory effects in the Inner Mongolia grassland
1
2004
... 大多数陆地生态系统天然植物群落都是特定气候条件下的产物(
赵哈林等, 2003 ), 气候条件的变化, 特别是气温和降水的变化会引起植物群落特征的变化(
Liu & Lei, 2015 ;
Liu et al ., 2016 ).
赵哈林等(2006) 认为气候是我国北方植被恢复的外在驱动力, 决定着演替的方向和终点.然而我们13年的观测结果显示群落生物量和物种丰富度与降水和气温之间不存在显著关系, 但气温和降水与生物量的线性回归斜率随着沙地的恢复而逐渐增大, 斜率的大小反映出植物群落对降水年际变化的响应敏感性(
Sala et al ., 2012 ), 在一定程度上也反映出群落对降水的利用效率(
Bai et al ., 2008 ).随着沙地生境恢复, 群落对年际降水波动的响应敏感性逐渐增加, 降水利用效率也相应地提升, 这种变化可能与资源的可利用性密切相关.许多研究发现土壤氮等资源的可利用性越高, 植被对降水变化的响应越敏感(
Wilcox et al ., 2016 ,
2017 ;
Knapp et al ., 2017 ), 在科尔沁沙地大尺度的研究也发现土壤养分梯度是影响植物群落分布最主要的因素(
左小安等, 2007 ).本研究中沙质草地土壤环境条件最优, 固定沙丘次之, 流动沙丘最差, 因此这种资源分布的差异导致不同沙地群落对降水变化的响应不同.群落组成也会影响群落对降水变化的响应.通过对生活型功能群重要值的年变化分析发现沙质草地群落不同功能群之间存在补偿效应, 这种补偿效应的存在可以有效地减缓降水等气候因子变化带来的影响.
Bai等(2004) 在内蒙古草原的研究也发现群落功能群之间存在补偿效应, 这种效应的存在更有利于群落稳定性的维持.这些因素的综合作用可能导致了群落生物量年变化对降水波动响应微弱.然而需要注意的是许多关于降水与群落生产力之间关系的结论都是在长时间尺度或者大空间尺度统计得来的(
Knapp et al ., 2017 ;
Wilcox et al ., 2017 ), 而本研究仅是在时间跨度为13年的小区域中开展的, 因此还需要更长时间的观测资料来研究群落对降水变化的响应.年内生物量积累与积温和累积降水量显著相关, 甚至在沙质草地二者对生物量年内积累的解释率达到68%, 因此积温和累积降水量是年内生物量积累的重要驱动因素, 这与植被生长过程中的水热需求规律一致. ...
Primary production and rain use efficiency across a precipitation gradient on the Mongolia plateau
1
2008
... 大多数陆地生态系统天然植物群落都是特定气候条件下的产物(
赵哈林等, 2003 ), 气候条件的变化, 特别是气温和降水的变化会引起植物群落特征的变化(
Liu & Lei, 2015 ;
Liu et al ., 2016 ).
赵哈林等(2006) 认为气候是我国北方植被恢复的外在驱动力, 决定着演替的方向和终点.然而我们13年的观测结果显示群落生物量和物种丰富度与降水和气温之间不存在显著关系, 但气温和降水与生物量的线性回归斜率随着沙地的恢复而逐渐增大, 斜率的大小反映出植物群落对降水年际变化的响应敏感性(
Sala et al ., 2012 ), 在一定程度上也反映出群落对降水的利用效率(
Bai et al ., 2008 ).随着沙地生境恢复, 群落对年际降水波动的响应敏感性逐渐增加, 降水利用效率也相应地提升, 这种变化可能与资源的可利用性密切相关.许多研究发现土壤氮等资源的可利用性越高, 植被对降水变化的响应越敏感(
Wilcox et al ., 2016 ,
2017 ;
Knapp et al ., 2017 ), 在科尔沁沙地大尺度的研究也发现土壤养分梯度是影响植物群落分布最主要的因素(
左小安等, 2007 ).本研究中沙质草地土壤环境条件最优, 固定沙丘次之, 流动沙丘最差, 因此这种资源分布的差异导致不同沙地群落对降水变化的响应不同.群落组成也会影响群落对降水变化的响应.通过对生活型功能群重要值的年变化分析发现沙质草地群落不同功能群之间存在补偿效应, 这种补偿效应的存在可以有效地减缓降水等气候因子变化带来的影响.
Bai等(2004) 在内蒙古草原的研究也发现群落功能群之间存在补偿效应, 这种效应的存在更有利于群落稳定性的维持.这些因素的综合作用可能导致了群落生物量年变化对降水波动响应微弱.然而需要注意的是许多关于降水与群落生产力之间关系的结论都是在长时间尺度或者大空间尺度统计得来的(
Knapp et al ., 2017 ;
Wilcox et al ., 2017 ), 而本研究仅是在时间跨度为13年的小区域中开展的, 因此还需要更长时间的观测资料来研究群落对降水变化的响应.年内生物量积累与积温和累积降水量显著相关, 甚至在沙质草地二者对生物量年内积累的解释率达到68%, 因此积温和累积降水量是年内生物量积累的重要驱动因素, 这与植被生长过程中的水热需求规律一致. ...
小针茅荒漠草原土壤水分动态及其对降雨的响应
1
2016
... 退化沙地土壤环境的恢复速率直接影响植被的演替进程, 土壤环境的改善是植被对土壤作用的结果, 但又反馈于植被的演替(
赵哈林等, 2006 ).我们的监测结果显示3种封育沙地在10年内仅有土壤有效氮和有效磷含量增加, 并且增加幅度不大, 土壤养分缓慢的积累速率对植被恢复有很大的限制作用, 尤其是在养分含量极低的流动沙丘这种限制作用可能会更大, 因此封育13年的流动沙丘群落生物量没有显著增加.另外, 土壤养分的空间异质性直接影响植被分布模式(
Zuo et al ., 2014 ).本研究3种沙地土壤环境存在显著差异, 群落物种与土壤因子之间的CCA分析也显示不同沙地群落物种受不同因素的影响, 其中沙质草地植物群落物种与土壤养分之间相关性很强, 固定沙丘群落物种和土壤水分联系紧密, 而流动沙丘群落物种与各土壤因子并不相关.3类沙地中沙质草地土壤养分和水分条件最优, 其相应的群落生物量和物种密度也最高, 但沙质草地多为草本植物, 密布的根系层对降水存在滞留作用(
牛存洋等, 2015 ), 另外草本植物相比于灌木和半灌木耗水相对较少, 并且较高的植被盖度产生的地表遮阴效果也能有效减弱土壤蒸发(
常昌明等, 2016 ), 这些因素的综合作用使其土壤水分含量最高, 进而使水分对群落的限制作用可能弱于土壤养分.而在固定沙丘土壤水分含量最低, 这是因为在固定沙丘优势物种主要为根系较深、耗水强烈的差不嘎蒿、小叶锦鸡儿等灌木, 使得土壤水分消耗强烈, 导致群落主要受土壤水分的限制.流动沙丘由于表面干沙层的存在抑制了土壤水分的蒸发(
段利民, 2011 ), 其土壤水分含量反而高于固定沙丘, 但其土壤养分和种子库非常贫瘠, 因此二者可能共同限制了群落的恢复. ...
小针茅荒漠草原土壤水分动态及其对降雨的响应
1
2016
... 退化沙地土壤环境的恢复速率直接影响植被的演替进程, 土壤环境的改善是植被对土壤作用的结果, 但又反馈于植被的演替(
赵哈林等, 2006 ).我们的监测结果显示3种封育沙地在10年内仅有土壤有效氮和有效磷含量增加, 并且增加幅度不大, 土壤养分缓慢的积累速率对植被恢复有很大的限制作用, 尤其是在养分含量极低的流动沙丘这种限制作用可能会更大, 因此封育13年的流动沙丘群落生物量没有显著增加.另外, 土壤养分的空间异质性直接影响植被分布模式(
Zuo et al ., 2014 ).本研究3种沙地土壤环境存在显著差异, 群落物种与土壤因子之间的CCA分析也显示不同沙地群落物种受不同因素的影响, 其中沙质草地植物群落物种与土壤养分之间相关性很强, 固定沙丘群落物种和土壤水分联系紧密, 而流动沙丘群落物种与各土壤因子并不相关.3类沙地中沙质草地土壤养分和水分条件最优, 其相应的群落生物量和物种密度也最高, 但沙质草地多为草本植物, 密布的根系层对降水存在滞留作用(
牛存洋等, 2015 ), 另外草本植物相比于灌木和半灌木耗水相对较少, 并且较高的植被盖度产生的地表遮阴效果也能有效减弱土壤蒸发(
常昌明等, 2016 ), 这些因素的综合作用使其土壤水分含量最高, 进而使水分对群落的限制作用可能弱于土壤养分.而在固定沙丘土壤水分含量最低, 这是因为在固定沙丘优势物种主要为根系较深、耗水强烈的差不嘎蒿、小叶锦鸡儿等灌木, 使得土壤水分消耗强烈, 导致群落主要受土壤水分的限制.流动沙丘由于表面干沙层的存在抑制了土壤水分的蒸发(
段利民, 2011 ), 其土壤水分含量反而高于固定沙丘, 但其土壤养分和种子库非常贫瘠, 因此二者可能共同限制了群落的恢复. ...
黄土高原草地合理利用与草地植被演替过程的实验研究
1
1995
... 退化沙质草地封育恢复是群落生物量和物种多样性显著增加, 多年生植物优势度提高, 群落结构逐渐趋于稳定, 群落生态功能不断增强的过程(
张继义等, 2004 ).在本研究中封育21年的沙质草地群落生物量不存在显著的变化趋势, 但群落物种丰富度显著下降, 多年生禾草功能群重要值也不断降低, 这也与正常退化草地封育恢复过程中的群落演替相反, 反映出固定沙丘群落在观测期间存在退化趋势.根据生态系统可持续性原理(
戎郁萍等, 2004 ), 退化草地的封育时间不应是无限期的, 我们的研究结果也显示封育时间过长不仅不利于群落恢复, 反而会导致群落发生逆行演替, 因此封育一段时间后应适当实施刈割、放牧等干扰措施, 刺激牧草分蘖, 促进植物再生, 改善沙质草地质量(
程积民等, 1995 ), 也有研究发现轻度放牧是内蒙古草原最有利的草地利用方式(
Gao et al ., 2015 ). ...
黄土高原草地合理利用与草地植被演替过程的实验研究
1
1995
... 退化沙质草地封育恢复是群落生物量和物种多样性显著增加, 多年生植物优势度提高, 群落结构逐渐趋于稳定, 群落生态功能不断增强的过程(
张继义等, 2004 ).在本研究中封育21年的沙质草地群落生物量不存在显著的变化趋势, 但群落物种丰富度显著下降, 多年生禾草功能群重要值也不断降低, 这也与正常退化草地封育恢复过程中的群落演替相反, 反映出固定沙丘群落在观测期间存在退化趋势.根据生态系统可持续性原理(
戎郁萍等, 2004 ), 退化草地的封育时间不应是无限期的, 我们的研究结果也显示封育时间过长不仅不利于群落恢复, 反而会导致群落发生逆行演替, 因此封育一段时间后应适当实施刈割、放牧等干扰措施, 刺激牧草分蘖, 促进植物再生, 改善沙质草地质量(
程积民等, 1995 ), 也有研究发现轻度放牧是内蒙古草原最有利的草地利用方式(
Gao et al ., 2015 ). ...
科尔沁沙地沙丘-草甸相间地区水文-土壤-植被动态响应关系研究
1
2011
... 退化沙地土壤环境的恢复速率直接影响植被的演替进程, 土壤环境的改善是植被对土壤作用的结果, 但又反馈于植被的演替(
赵哈林等, 2006 ).我们的监测结果显示3种封育沙地在10年内仅有土壤有效氮和有效磷含量增加, 并且增加幅度不大, 土壤养分缓慢的积累速率对植被恢复有很大的限制作用, 尤其是在养分含量极低的流动沙丘这种限制作用可能会更大, 因此封育13年的流动沙丘群落生物量没有显著增加.另外, 土壤养分的空间异质性直接影响植被分布模式(
Zuo et al ., 2014 ).本研究3种沙地土壤环境存在显著差异, 群落物种与土壤因子之间的CCA分析也显示不同沙地群落物种受不同因素的影响, 其中沙质草地植物群落物种与土壤养分之间相关性很强, 固定沙丘群落物种和土壤水分联系紧密, 而流动沙丘群落物种与各土壤因子并不相关.3类沙地中沙质草地土壤养分和水分条件最优, 其相应的群落生物量和物种密度也最高, 但沙质草地多为草本植物, 密布的根系层对降水存在滞留作用(
牛存洋等, 2015 ), 另外草本植物相比于灌木和半灌木耗水相对较少, 并且较高的植被盖度产生的地表遮阴效果也能有效减弱土壤蒸发(
常昌明等, 2016 ), 这些因素的综合作用使其土壤水分含量最高, 进而使水分对群落的限制作用可能弱于土壤养分.而在固定沙丘土壤水分含量最低, 这是因为在固定沙丘优势物种主要为根系较深、耗水强烈的差不嘎蒿、小叶锦鸡儿等灌木, 使得土壤水分消耗强烈, 导致群落主要受土壤水分的限制.流动沙丘由于表面干沙层的存在抑制了土壤水分的蒸发(
段利民, 2011 ), 其土壤水分含量反而高于固定沙丘, 但其土壤养分和种子库非常贫瘠, 因此二者可能共同限制了群落的恢复. ...
科尔沁沙地沙丘-草甸相间地区水文-土壤-植被动态响应关系研究
1
2011
... 退化沙地土壤环境的恢复速率直接影响植被的演替进程, 土壤环境的改善是植被对土壤作用的结果, 但又反馈于植被的演替(
赵哈林等, 2006 ).我们的监测结果显示3种封育沙地在10年内仅有土壤有效氮和有效磷含量增加, 并且增加幅度不大, 土壤养分缓慢的积累速率对植被恢复有很大的限制作用, 尤其是在养分含量极低的流动沙丘这种限制作用可能会更大, 因此封育13年的流动沙丘群落生物量没有显著增加.另外, 土壤养分的空间异质性直接影响植被分布模式(
Zuo et al ., 2014 ).本研究3种沙地土壤环境存在显著差异, 群落物种与土壤因子之间的CCA分析也显示不同沙地群落物种受不同因素的影响, 其中沙质草地植物群落物种与土壤养分之间相关性很强, 固定沙丘群落物种和土壤水分联系紧密, 而流动沙丘群落物种与各土壤因子并不相关.3类沙地中沙质草地土壤养分和水分条件最优, 其相应的群落生物量和物种密度也最高, 但沙质草地多为草本植物, 密布的根系层对降水存在滞留作用(
牛存洋等, 2015 ), 另外草本植物相比于灌木和半灌木耗水相对较少, 并且较高的植被盖度产生的地表遮阴效果也能有效减弱土壤蒸发(
常昌明等, 2016 ), 这些因素的综合作用使其土壤水分含量最高, 进而使水分对群落的限制作用可能弱于土壤养分.而在固定沙丘土壤水分含量最低, 这是因为在固定沙丘优势物种主要为根系较深、耗水强烈的差不嘎蒿、小叶锦鸡儿等灌木, 使得土壤水分消耗强烈, 导致群落主要受土壤水分的限制.流动沙丘由于表面干沙层的存在抑制了土壤水分的蒸发(
段利民, 2011 ), 其土壤水分含量反而高于固定沙丘, 但其土壤养分和种子库非常贫瘠, 因此二者可能共同限制了群落的恢复. ...
围封年限对内蒙古羊草-针茅典型草原植物功能群及其多样性的影响
1
2013
... 科尔沁沙地作为中国北方最严重的一个沙漠化区域(
刘新民等, 1999 ), 封育禁牧被广泛用来恢复退化沙地植被, 同时也有许多研究对封育所产生的生态效应做了评述.如退化沙质草地封育后群落生物量响应最为敏感, 在较短时间内群落生物量迅速恢复, 同时凋落物量也显著增加(
苗仁辉等, 2013 ;
赵丽娅等, 2017 ;
吕朋等, 2018 ).随着封育时间的延长, 群落物种组成开始发生变化, 流动沙丘封育后沙蓬(
Agriophyllum squarrosum )作为先锋物种首先建立种群, 使沙丘流沙面积减小, 之后以差不嘎蒿(
Artemisia halodendron )为主的多年生灌木通过无性繁殖的方式开始在沙丘定植, 差不嘎蒿等灌木的出现逐渐营造出养分和水分相对富集的灌丛微生境, 进而为禾本科植物的种子萌发提供一个良好的环境(
Su et al ., 2005 ).随着植物群落的变化, 土壤种子库也随之改变(
赵丽娅等, 2006 ;
赵盼盼等, 2018 ), 并且随着流动沙丘的固定和生境的改善, 植物群落和土壤种子库之间的联系变得更加紧密(
Wang et al ., 2019 ).封育对土壤环境也有很大的改善, 对封育和放牧样地土壤性质进行比较, 发现封育样地土壤容重、紧实度及粗颗粒含量显著小于自由放牧样地(
单贵莲等, 2009 ), 而封育样地土壤有机质、全氮、有效氮和有效钾含量显著高于放牧样地(
张建鹏等, 2017 ).封育年限长短也是影响植物群落恢复的重要因素, 适当封育有利于植被恢复, 但过度封育反而会引起植被退化,
高凯等(2013) 在内蒙古草原的研究发现, 封育年限超过14年会使多年生禾草以及灌木和半灌木优势度显著降低, 而多年生杂类草优势度出现不同程度的升高. ...
围封年限对内蒙古羊草-针茅典型草原植物功能群及其多样性的影响
1
2013
... 科尔沁沙地作为中国北方最严重的一个沙漠化区域(
刘新民等, 1999 ), 封育禁牧被广泛用来恢复退化沙地植被, 同时也有许多研究对封育所产生的生态效应做了评述.如退化沙质草地封育后群落生物量响应最为敏感, 在较短时间内群落生物量迅速恢复, 同时凋落物量也显著增加(
苗仁辉等, 2013 ;
赵丽娅等, 2017 ;
吕朋等, 2018 ).随着封育时间的延长, 群落物种组成开始发生变化, 流动沙丘封育后沙蓬(
Agriophyllum squarrosum )作为先锋物种首先建立种群, 使沙丘流沙面积减小, 之后以差不嘎蒿(
Artemisia halodendron )为主的多年生灌木通过无性繁殖的方式开始在沙丘定植, 差不嘎蒿等灌木的出现逐渐营造出养分和水分相对富集的灌丛微生境, 进而为禾本科植物的种子萌发提供一个良好的环境(
Su et al ., 2005 ).随着植物群落的变化, 土壤种子库也随之改变(
赵丽娅等, 2006 ;
赵盼盼等, 2018 ), 并且随着流动沙丘的固定和生境的改善, 植物群落和土壤种子库之间的联系变得更加紧密(
Wang et al ., 2019 ).封育对土壤环境也有很大的改善, 对封育和放牧样地土壤性质进行比较, 发现封育样地土壤容重、紧实度及粗颗粒含量显著小于自由放牧样地(
单贵莲等, 2009 ), 而封育样地土壤有机质、全氮、有效氮和有效钾含量显著高于放牧样地(
张建鹏等, 2017 ).封育年限长短也是影响植物群落恢复的重要因素, 适当封育有利于植被恢复, 但过度封育反而会引起植被退化,
高凯等(2013) 在内蒙古草原的研究发现, 封育年限超过14年会使多年生禾草以及灌木和半灌木优势度显著降低, 而多年生杂类草优势度出现不同程度的升高. ...
Influence of enclosure on vegetation productivity and biodiversity on Xilamuren Grassland, Inner Mogolia
1
2015
... 退化沙质草地封育恢复是群落生物量和物种多样性显著增加, 多年生植物优势度提高, 群落结构逐渐趋于稳定, 群落生态功能不断增强的过程(
张继义等, 2004 ).在本研究中封育21年的沙质草地群落生物量不存在显著的变化趋势, 但群落物种丰富度显著下降, 多年生禾草功能群重要值也不断降低, 这也与正常退化草地封育恢复过程中的群落演替相反, 反映出固定沙丘群落在观测期间存在退化趋势.根据生态系统可持续性原理(
戎郁萍等, 2004 ), 退化草地的封育时间不应是无限期的, 我们的研究结果也显示封育时间过长不仅不利于群落恢复, 反而会导致群落发生逆行演替, 因此封育一段时间后应适当实施刈割、放牧等干扰措施, 刺激牧草分蘖, 促进植物再生, 改善沙质草地质量(
程积民等, 1995 ), 也有研究发现轻度放牧是内蒙古草原最有利的草地利用方式(
Gao et al ., 2015 ). ...
Reconciling inconsistencies in precipitation-productivity relationships: Implications for climate change
2
2017
... 大多数陆地生态系统天然植物群落都是特定气候条件下的产物(
赵哈林等, 2003 ), 气候条件的变化, 特别是气温和降水的变化会引起植物群落特征的变化(
Liu & Lei, 2015 ;
Liu et al ., 2016 ).
赵哈林等(2006) 认为气候是我国北方植被恢复的外在驱动力, 决定着演替的方向和终点.然而我们13年的观测结果显示群落生物量和物种丰富度与降水和气温之间不存在显著关系, 但气温和降水与生物量的线性回归斜率随着沙地的恢复而逐渐增大, 斜率的大小反映出植物群落对降水年际变化的响应敏感性(
Sala et al ., 2012 ), 在一定程度上也反映出群落对降水的利用效率(
Bai et al ., 2008 ).随着沙地生境恢复, 群落对年际降水波动的响应敏感性逐渐增加, 降水利用效率也相应地提升, 这种变化可能与资源的可利用性密切相关.许多研究发现土壤氮等资源的可利用性越高, 植被对降水变化的响应越敏感(
Wilcox et al ., 2016 ,
2017 ;
Knapp et al ., 2017 ), 在科尔沁沙地大尺度的研究也发现土壤养分梯度是影响植物群落分布最主要的因素(
左小安等, 2007 ).本研究中沙质草地土壤环境条件最优, 固定沙丘次之, 流动沙丘最差, 因此这种资源分布的差异导致不同沙地群落对降水变化的响应不同.群落组成也会影响群落对降水变化的响应.通过对生活型功能群重要值的年变化分析发现沙质草地群落不同功能群之间存在补偿效应, 这种补偿效应的存在可以有效地减缓降水等气候因子变化带来的影响.
Bai等(2004) 在内蒙古草原的研究也发现群落功能群之间存在补偿效应, 这种效应的存在更有利于群落稳定性的维持.这些因素的综合作用可能导致了群落生物量年变化对降水波动响应微弱.然而需要注意的是许多关于降水与群落生产力之间关系的结论都是在长时间尺度或者大空间尺度统计得来的(
Knapp et al ., 2017 ;
Wilcox et al ., 2017 ), 而本研究仅是在时间跨度为13年的小区域中开展的, 因此还需要更长时间的观测资料来研究群落对降水变化的响应.年内生物量积累与积温和累积降水量显著相关, 甚至在沙质草地二者对生物量年内积累的解释率达到68%, 因此积温和累积降水量是年内生物量积累的重要驱动因素, 这与植被生长过程中的水热需求规律一致. ...
... 在内蒙古草原的研究也发现群落功能群之间存在补偿效应, 这种效应的存在更有利于群落稳定性的维持.这些因素的综合作用可能导致了群落生物量年变化对降水波动响应微弱.然而需要注意的是许多关于降水与群落生产力之间关系的结论都是在长时间尺度或者大空间尺度统计得来的(
Knapp et al ., 2017 ;
Wilcox et al ., 2017 ), 而本研究仅是在时间跨度为13年的小区域中开展的, 因此还需要更长时间的观测资料来研究群落对降水变化的响应.年内生物量积累与积温和累积降水量显著相关, 甚至在沙质草地二者对生物量年内积累的解释率达到68%, 因此积温和累积降水量是年内生物量积累的重要驱动因素, 这与植被生长过程中的水热需求规律一致. ...
Precipitation pattern determines the inter-annual variation of herbaceous layer and carbon fluxes in a phreatophyte-dominated desert ecosystem
1
2016
... 大多数陆地生态系统天然植物群落都是特定气候条件下的产物(
赵哈林等, 2003 ), 气候条件的变化, 特别是气温和降水的变化会引起植物群落特征的变化(
Liu & Lei, 2015 ;
Liu et al ., 2016 ).
赵哈林等(2006) 认为气候是我国北方植被恢复的外在驱动力, 决定着演替的方向和终点.然而我们13年的观测结果显示群落生物量和物种丰富度与降水和气温之间不存在显著关系, 但气温和降水与生物量的线性回归斜率随着沙地的恢复而逐渐增大, 斜率的大小反映出植物群落对降水年际变化的响应敏感性(
Sala et al ., 2012 ), 在一定程度上也反映出群落对降水的利用效率(
Bai et al ., 2008 ).随着沙地生境恢复, 群落对年际降水波动的响应敏感性逐渐增加, 降水利用效率也相应地提升, 这种变化可能与资源的可利用性密切相关.许多研究发现土壤氮等资源的可利用性越高, 植被对降水变化的响应越敏感(
Wilcox et al ., 2016 ,
2017 ;
Knapp et al ., 2017 ), 在科尔沁沙地大尺度的研究也发现土壤养分梯度是影响植物群落分布最主要的因素(
左小安等, 2007 ).本研究中沙质草地土壤环境条件最优, 固定沙丘次之, 流动沙丘最差, 因此这种资源分布的差异导致不同沙地群落对降水变化的响应不同.群落组成也会影响群落对降水变化的响应.通过对生活型功能群重要值的年变化分析发现沙质草地群落不同功能群之间存在补偿效应, 这种补偿效应的存在可以有效地减缓降水等气候因子变化带来的影响.
Bai等(2004) 在内蒙古草原的研究也发现群落功能群之间存在补偿效应, 这种效应的存在更有利于群落稳定性的维持.这些因素的综合作用可能导致了群落生物量年变化对降水波动响应微弱.然而需要注意的是许多关于降水与群落生产力之间关系的结论都是在长时间尺度或者大空间尺度统计得来的(
Knapp et al ., 2017 ;
Wilcox et al ., 2017 ), 而本研究仅是在时间跨度为13年的小区域中开展的, 因此还需要更长时间的观测资料来研究群落对降水变化的响应.年内生物量积累与积温和累积降水量显著相关, 甚至在沙质草地二者对生物量年内积累的解释率达到68%, 因此积温和累积降水量是年内生物量积累的重要驱动因素, 这与植被生长过程中的水热需求规律一致. ...
1
1999
... 科尔沁沙地作为中国北方最严重的一个沙漠化区域(
刘新民等, 1999 ), 封育禁牧被广泛用来恢复退化沙地植被, 同时也有许多研究对封育所产生的生态效应做了评述.如退化沙质草地封育后群落生物量响应最为敏感, 在较短时间内群落生物量迅速恢复, 同时凋落物量也显著增加(
苗仁辉等, 2013 ;
赵丽娅等, 2017 ;
吕朋等, 2018 ).随着封育时间的延长, 群落物种组成开始发生变化, 流动沙丘封育后沙蓬(
Agriophyllum squarrosum )作为先锋物种首先建立种群, 使沙丘流沙面积减小, 之后以差不嘎蒿(
Artemisia halodendron )为主的多年生灌木通过无性繁殖的方式开始在沙丘定植, 差不嘎蒿等灌木的出现逐渐营造出养分和水分相对富集的灌丛微生境, 进而为禾本科植物的种子萌发提供一个良好的环境(
Su et al ., 2005 ).随着植物群落的变化, 土壤种子库也随之改变(
赵丽娅等, 2006 ;
赵盼盼等, 2018 ), 并且随着流动沙丘的固定和生境的改善, 植物群落和土壤种子库之间的联系变得更加紧密(
Wang et al ., 2019 ).封育对土壤环境也有很大的改善, 对封育和放牧样地土壤性质进行比较, 发现封育样地土壤容重、紧实度及粗颗粒含量显著小于自由放牧样地(
单贵莲等, 2009 ), 而封育样地土壤有机质、全氮、有效氮和有效钾含量显著高于放牧样地(
张建鹏等, 2017 ).封育年限长短也是影响植物群落恢复的重要因素, 适当封育有利于植被恢复, 但过度封育反而会引起植被退化,
高凯等(2013) 在内蒙古草原的研究发现, 封育年限超过14年会使多年生禾草以及灌木和半灌木优势度显著降低, 而多年生杂类草优势度出现不同程度的升高. ...
1
1999
... 科尔沁沙地作为中国北方最严重的一个沙漠化区域(
刘新民等, 1999 ), 封育禁牧被广泛用来恢复退化沙地植被, 同时也有许多研究对封育所产生的生态效应做了评述.如退化沙质草地封育后群落生物量响应最为敏感, 在较短时间内群落生物量迅速恢复, 同时凋落物量也显著增加(
苗仁辉等, 2013 ;
赵丽娅等, 2017 ;
吕朋等, 2018 ).随着封育时间的延长, 群落物种组成开始发生变化, 流动沙丘封育后沙蓬(
Agriophyllum squarrosum )作为先锋物种首先建立种群, 使沙丘流沙面积减小, 之后以差不嘎蒿(
Artemisia halodendron )为主的多年生灌木通过无性繁殖的方式开始在沙丘定植, 差不嘎蒿等灌木的出现逐渐营造出养分和水分相对富集的灌丛微生境, 进而为禾本科植物的种子萌发提供一个良好的环境(
Su et al ., 2005 ).随着植物群落的变化, 土壤种子库也随之改变(
赵丽娅等, 2006 ;
赵盼盼等, 2018 ), 并且随着流动沙丘的固定和生境的改善, 植物群落和土壤种子库之间的联系变得更加紧密(
Wang et al ., 2019 ).封育对土壤环境也有很大的改善, 对封育和放牧样地土壤性质进行比较, 发现封育样地土壤容重、紧实度及粗颗粒含量显著小于自由放牧样地(
单贵莲等, 2009 ), 而封育样地土壤有机质、全氮、有效氮和有效钾含量显著高于放牧样地(
张建鹏等, 2017 ).封育年限长短也是影响植物群落恢复的重要因素, 适当封育有利于植被恢复, 但过度封育反而会引起植被退化,
高凯等(2013) 在内蒙古草原的研究发现, 封育年限超过14年会使多年生禾草以及灌木和半灌木优势度显著降低, 而多年生杂类草优势度出现不同程度的升高. ...
Responses of natural vegetation dynamics to climate drivers in China from 1982 to 2011
1
2015
... 大多数陆地生态系统天然植物群落都是特定气候条件下的产物(
赵哈林等, 2003 ), 气候条件的变化, 特别是气温和降水的变化会引起植物群落特征的变化(
Liu & Lei, 2015 ;
Liu et al ., 2016 ).
赵哈林等(2006) 认为气候是我国北方植被恢复的外在驱动力, 决定着演替的方向和终点.然而我们13年的观测结果显示群落生物量和物种丰富度与降水和气温之间不存在显著关系, 但气温和降水与生物量的线性回归斜率随着沙地的恢复而逐渐增大, 斜率的大小反映出植物群落对降水年际变化的响应敏感性(
Sala et al ., 2012 ), 在一定程度上也反映出群落对降水的利用效率(
Bai et al ., 2008 ).随着沙地生境恢复, 群落对年际降水波动的响应敏感性逐渐增加, 降水利用效率也相应地提升, 这种变化可能与资源的可利用性密切相关.许多研究发现土壤氮等资源的可利用性越高, 植被对降水变化的响应越敏感(
Wilcox et al ., 2016 ,
2017 ;
Knapp et al ., 2017 ), 在科尔沁沙地大尺度的研究也发现土壤养分梯度是影响植物群落分布最主要的因素(
左小安等, 2007 ).本研究中沙质草地土壤环境条件最优, 固定沙丘次之, 流动沙丘最差, 因此这种资源分布的差异导致不同沙地群落对降水变化的响应不同.群落组成也会影响群落对降水变化的响应.通过对生活型功能群重要值的年变化分析发现沙质草地群落不同功能群之间存在补偿效应, 这种补偿效应的存在可以有效地减缓降水等气候因子变化带来的影响.
Bai等(2004) 在内蒙古草原的研究也发现群落功能群之间存在补偿效应, 这种效应的存在更有利于群落稳定性的维持.这些因素的综合作用可能导致了群落生物量年变化对降水波动响应微弱.然而需要注意的是许多关于降水与群落生产力之间关系的结论都是在长时间尺度或者大空间尺度统计得来的(
Knapp et al ., 2017 ;
Wilcox et al ., 2017 ), 而本研究仅是在时间跨度为13年的小区域中开展的, 因此还需要更长时间的观测资料来研究群落对降水变化的响应.年内生物量积累与积温和累积降水量显著相关, 甚至在沙质草地二者对生物量年内积累的解释率达到68%, 因此积温和累积降水量是年内生物量积累的重要驱动因素, 这与植被生长过程中的水热需求规律一致. ...
1
1999
... 自2005年起在科尔沁沙地选择长期封育的沙质草地(1997年封育, 面积34 500 m
2 )、固定沙丘(1985年封育, 面积20 000 m
2 )以及流动沙丘(2005年封育, 面积7 000 m
2 )为研究样地(
图1 ), 每个样地布设10个1 m × 1 m的样方, 每年8月下旬调查草本植物群落物种组成、高度、盖度、密度以及地上生物量, 地上生物量采用刈割法, 采样后烘干称质量; 另外设置5个5 m × 5 m的样方用于灌木和半灌木群落物种组成、高度、盖度、密度以及地上生物量调查, 其中灌木地上生物量测定采用标准枝法.每个样地布设3个土壤水分中子仪(CNC503B(DR), 北京超能科技公司), 用于土壤水分测定.土壤样品采集于2008和2017年的8月下旬, 在每个草本群落调查样方内用直径3 cm的土钻取0-20 cm深度的土壤样品, 每个样方3个重复, 用于测定土壤有机碳、全氮、有效氮、有效磷以及有效钾含量.土壤有机碳测定采用重铬酸钾容量法-外加热法, 土壤全氮测定采用电位法, 有效氮测定采用碱扩散法, 有效磷测定采用碳酸氢钠浸提-钼锑抗比色法, 土壤有效钾测定采用乙酸铵浸提-火焰光度法(
鲁如坤, 1999 ).土壤种子库样品采集于2008和2017年10月中旬, 每个沙地内草本植物群落调查样方旁设置1 m × 1 m的样方, 用土壤种子采样器在每个样方内采取深度为 5 cm, 面积为20 cm × 20 cm的原状土体装入布袋, 带回试验室采用室内萌发实验测定土壤种子库.多年气温和降水数据来源于奈曼沙漠化研究站自动气象观测站. ...
1
1999
... 自2005年起在科尔沁沙地选择长期封育的沙质草地(1997年封育, 面积34 500 m
2 )、固定沙丘(1985年封育, 面积20 000 m
2 )以及流动沙丘(2005年封育, 面积7 000 m
2 )为研究样地(
图1 ), 每个样地布设10个1 m × 1 m的样方, 每年8月下旬调查草本植物群落物种组成、高度、盖度、密度以及地上生物量, 地上生物量采用刈割法, 采样后烘干称质量; 另外设置5个5 m × 5 m的样方用于灌木和半灌木群落物种组成、高度、盖度、密度以及地上生物量调查, 其中灌木地上生物量测定采用标准枝法.每个样地布设3个土壤水分中子仪(CNC503B(DR), 北京超能科技公司), 用于土壤水分测定.土壤样品采集于2008和2017年的8月下旬, 在每个草本群落调查样方内用直径3 cm的土钻取0-20 cm深度的土壤样品, 每个样方3个重复, 用于测定土壤有机碳、全氮、有效氮、有效磷以及有效钾含量.土壤有机碳测定采用重铬酸钾容量法-外加热法, 土壤全氮测定采用电位法, 有效氮测定采用碱扩散法, 有效磷测定采用碳酸氢钠浸提-钼锑抗比色法, 土壤有效钾测定采用乙酸铵浸提-火焰光度法(
鲁如坤, 1999 ).土壤种子库样品采集于2008和2017年10月中旬, 每个沙地内草本植物群落调查样方旁设置1 m × 1 m的样方, 用土壤种子采样器在每个样方内采取深度为 5 cm, 面积为20 cm × 20 cm的原状土体装入布袋, 带回试验室采用室内萌发实验测定土壤种子库.多年气温和降水数据来源于奈曼沙漠化研究站自动气象观测站. ...
科尔沁沙地封育过程中植被特征的动态变化
2
2018
... 科尔沁沙地作为中国北方最严重的一个沙漠化区域(
刘新民等, 1999 ), 封育禁牧被广泛用来恢复退化沙地植被, 同时也有许多研究对封育所产生的生态效应做了评述.如退化沙质草地封育后群落生物量响应最为敏感, 在较短时间内群落生物量迅速恢复, 同时凋落物量也显著增加(
苗仁辉等, 2013 ;
赵丽娅等, 2017 ;
吕朋等, 2018 ).随着封育时间的延长, 群落物种组成开始发生变化, 流动沙丘封育后沙蓬(
Agriophyllum squarrosum )作为先锋物种首先建立种群, 使沙丘流沙面积减小, 之后以差不嘎蒿(
Artemisia halodendron )为主的多年生灌木通过无性繁殖的方式开始在沙丘定植, 差不嘎蒿等灌木的出现逐渐营造出养分和水分相对富集的灌丛微生境, 进而为禾本科植物的种子萌发提供一个良好的环境(
Su et al ., 2005 ).随着植物群落的变化, 土壤种子库也随之改变(
赵丽娅等, 2006 ;
赵盼盼等, 2018 ), 并且随着流动沙丘的固定和生境的改善, 植物群落和土壤种子库之间的联系变得更加紧密(
Wang et al ., 2019 ).封育对土壤环境也有很大的改善, 对封育和放牧样地土壤性质进行比较, 发现封育样地土壤容重、紧实度及粗颗粒含量显著小于自由放牧样地(
单贵莲等, 2009 ), 而封育样地土壤有机质、全氮、有效氮和有效钾含量显著高于放牧样地(
张建鹏等, 2017 ).封育年限长短也是影响植物群落恢复的重要因素, 适当封育有利于植被恢复, 但过度封育反而会引起植被退化,
高凯等(2013) 在内蒙古草原的研究发现, 封育年限超过14年会使多年生禾草以及灌木和半灌木优势度显著降低, 而多年生杂类草优势度出现不同程度的升高. ...
... 退化沙地恢复的速度和演替方向一方面取决于退化程度和干扰的消除程度, 另一方面取决于土壤、气候等环境条件(
赵哈林等, 2003 ).本研究中流动沙丘13年封育期间植被盖度和物种丰富度显著增加, 但群落生物量没有显著变化, 整体上群落在逐渐恢复, 但速度缓慢, 群落生产力有限.许多研究发现干扰退化草地在短期封育下可以快速恢复(
赵哈林等, 2009 ;
苗仁辉等, 2013 ;
吕朋等, 2018 ), 其根本原因是现存群落具有拓殖能力和退化群落资源过剩(
王炜等, 1996 ), 但对于流动沙丘, 群落资源处于极其匮乏的状态, 现存群落拓殖能力有限, 土壤种子库基础也相对薄弱, 因此植被恢复受到很大限制.另外植被恢复也受时间积累效应的影响, 物种要进入某一环境空间并建立种群, 需要有足够的时间去完成其繁殖体从种源地到目标空间的传播、散布和在目标空间的定居、繁殖过程, 这一过程进行的速度直接影响物种多样性恢复的进度(
张继义等, 2004 ), 本研究中流动沙丘封育仅13年, 植物建群时间有限, 因此时间积累可能相对不足, 植被恢复程度有限.有研究认为在降水条件较好的半干旱沙区, 采取封沙育草措施后10-15年的时间, 植被能够自然恢复(
张华等, 2003 ), 然而本研究的结果显示, 流动沙丘恢复到灌木、半灌木或多年生草本优势度较高的固定沙丘群落需要更长的时间, 因此封育年限的设定应综合考虑植被退化程度、气候条件以及土壤环境等因素. ...
科尔沁沙地封育过程中植被特征的动态变化
2
2018
... 科尔沁沙地作为中国北方最严重的一个沙漠化区域(
刘新民等, 1999 ), 封育禁牧被广泛用来恢复退化沙地植被, 同时也有许多研究对封育所产生的生态效应做了评述.如退化沙质草地封育后群落生物量响应最为敏感, 在较短时间内群落生物量迅速恢复, 同时凋落物量也显著增加(
苗仁辉等, 2013 ;
赵丽娅等, 2017 ;
吕朋等, 2018 ).随着封育时间的延长, 群落物种组成开始发生变化, 流动沙丘封育后沙蓬(
Agriophyllum squarrosum )作为先锋物种首先建立种群, 使沙丘流沙面积减小, 之后以差不嘎蒿(
Artemisia halodendron )为主的多年生灌木通过无性繁殖的方式开始在沙丘定植, 差不嘎蒿等灌木的出现逐渐营造出养分和水分相对富集的灌丛微生境, 进而为禾本科植物的种子萌发提供一个良好的环境(
Su et al ., 2005 ).随着植物群落的变化, 土壤种子库也随之改变(
赵丽娅等, 2006 ;
赵盼盼等, 2018 ), 并且随着流动沙丘的固定和生境的改善, 植物群落和土壤种子库之间的联系变得更加紧密(
Wang et al ., 2019 ).封育对土壤环境也有很大的改善, 对封育和放牧样地土壤性质进行比较, 发现封育样地土壤容重、紧实度及粗颗粒含量显著小于自由放牧样地(
单贵莲等, 2009 ), 而封育样地土壤有机质、全氮、有效氮和有效钾含量显著高于放牧样地(
张建鹏等, 2017 ).封育年限长短也是影响植物群落恢复的重要因素, 适当封育有利于植被恢复, 但过度封育反而会引起植被退化,
高凯等(2013) 在内蒙古草原的研究发现, 封育年限超过14年会使多年生禾草以及灌木和半灌木优势度显著降低, 而多年生杂类草优势度出现不同程度的升高. ...
... 退化沙地恢复的速度和演替方向一方面取决于退化程度和干扰的消除程度, 另一方面取决于土壤、气候等环境条件(
赵哈林等, 2003 ).本研究中流动沙丘13年封育期间植被盖度和物种丰富度显著增加, 但群落生物量没有显著变化, 整体上群落在逐渐恢复, 但速度缓慢, 群落生产力有限.许多研究发现干扰退化草地在短期封育下可以快速恢复(
赵哈林等, 2009 ;
苗仁辉等, 2013 ;
吕朋等, 2018 ), 其根本原因是现存群落具有拓殖能力和退化群落资源过剩(
王炜等, 1996 ), 但对于流动沙丘, 群落资源处于极其匮乏的状态, 现存群落拓殖能力有限, 土壤种子库基础也相对薄弱, 因此植被恢复受到很大限制.另外植被恢复也受时间积累效应的影响, 物种要进入某一环境空间并建立种群, 需要有足够的时间去完成其繁殖体从种源地到目标空间的传播、散布和在目标空间的定居、繁殖过程, 这一过程进行的速度直接影响物种多样性恢复的进度(
张继义等, 2004 ), 本研究中流动沙丘封育仅13年, 植物建群时间有限, 因此时间积累可能相对不足, 植被恢复程度有限.有研究认为在降水条件较好的半干旱沙区, 采取封沙育草措施后10-15年的时间, 植被能够自然恢复(
张华等, 2003 ), 然而本研究的结果显示, 流动沙丘恢复到灌木、半灌木或多年生草本优势度较高的固定沙丘群落需要更长的时间, 因此封育年限的设定应综合考虑植被退化程度、气候条件以及土壤环境等因素. ...
科尔沁沙质草地封育过程中的植被变化及其机制
2
2013
... 科尔沁沙地作为中国北方最严重的一个沙漠化区域(
刘新民等, 1999 ), 封育禁牧被广泛用来恢复退化沙地植被, 同时也有许多研究对封育所产生的生态效应做了评述.如退化沙质草地封育后群落生物量响应最为敏感, 在较短时间内群落生物量迅速恢复, 同时凋落物量也显著增加(
苗仁辉等, 2013 ;
赵丽娅等, 2017 ;
吕朋等, 2018 ).随着封育时间的延长, 群落物种组成开始发生变化, 流动沙丘封育后沙蓬(
Agriophyllum squarrosum )作为先锋物种首先建立种群, 使沙丘流沙面积减小, 之后以差不嘎蒿(
Artemisia halodendron )为主的多年生灌木通过无性繁殖的方式开始在沙丘定植, 差不嘎蒿等灌木的出现逐渐营造出养分和水分相对富集的灌丛微生境, 进而为禾本科植物的种子萌发提供一个良好的环境(
Su et al ., 2005 ).随着植物群落的变化, 土壤种子库也随之改变(
赵丽娅等, 2006 ;
赵盼盼等, 2018 ), 并且随着流动沙丘的固定和生境的改善, 植物群落和土壤种子库之间的联系变得更加紧密(
Wang et al ., 2019 ).封育对土壤环境也有很大的改善, 对封育和放牧样地土壤性质进行比较, 发现封育样地土壤容重、紧实度及粗颗粒含量显著小于自由放牧样地(
单贵莲等, 2009 ), 而封育样地土壤有机质、全氮、有效氮和有效钾含量显著高于放牧样地(
张建鹏等, 2017 ).封育年限长短也是影响植物群落恢复的重要因素, 适当封育有利于植被恢复, 但过度封育反而会引起植被退化,
高凯等(2013) 在内蒙古草原的研究发现, 封育年限超过14年会使多年生禾草以及灌木和半灌木优势度显著降低, 而多年生杂类草优势度出现不同程度的升高. ...
... 退化沙地恢复的速度和演替方向一方面取决于退化程度和干扰的消除程度, 另一方面取决于土壤、气候等环境条件(
赵哈林等, 2003 ).本研究中流动沙丘13年封育期间植被盖度和物种丰富度显著增加, 但群落生物量没有显著变化, 整体上群落在逐渐恢复, 但速度缓慢, 群落生产力有限.许多研究发现干扰退化草地在短期封育下可以快速恢复(
赵哈林等, 2009 ;
苗仁辉等, 2013 ;
吕朋等, 2018 ), 其根本原因是现存群落具有拓殖能力和退化群落资源过剩(
王炜等, 1996 ), 但对于流动沙丘, 群落资源处于极其匮乏的状态, 现存群落拓殖能力有限, 土壤种子库基础也相对薄弱, 因此植被恢复受到很大限制.另外植被恢复也受时间积累效应的影响, 物种要进入某一环境空间并建立种群, 需要有足够的时间去完成其繁殖体从种源地到目标空间的传播、散布和在目标空间的定居、繁殖过程, 这一过程进行的速度直接影响物种多样性恢复的进度(
张继义等, 2004 ), 本研究中流动沙丘封育仅13年, 植物建群时间有限, 因此时间积累可能相对不足, 植被恢复程度有限.有研究认为在降水条件较好的半干旱沙区, 采取封沙育草措施后10-15年的时间, 植被能够自然恢复(
张华等, 2003 ), 然而本研究的结果显示, 流动沙丘恢复到灌木、半灌木或多年生草本优势度较高的固定沙丘群落需要更长的时间, 因此封育年限的设定应综合考虑植被退化程度、气候条件以及土壤环境等因素. ...
科尔沁沙质草地封育过程中的植被变化及其机制
2
2013
... 科尔沁沙地作为中国北方最严重的一个沙漠化区域(
刘新民等, 1999 ), 封育禁牧被广泛用来恢复退化沙地植被, 同时也有许多研究对封育所产生的生态效应做了评述.如退化沙质草地封育后群落生物量响应最为敏感, 在较短时间内群落生物量迅速恢复, 同时凋落物量也显著增加(
苗仁辉等, 2013 ;
赵丽娅等, 2017 ;
吕朋等, 2018 ).随着封育时间的延长, 群落物种组成开始发生变化, 流动沙丘封育后沙蓬(
Agriophyllum squarrosum )作为先锋物种首先建立种群, 使沙丘流沙面积减小, 之后以差不嘎蒿(
Artemisia halodendron )为主的多年生灌木通过无性繁殖的方式开始在沙丘定植, 差不嘎蒿等灌木的出现逐渐营造出养分和水分相对富集的灌丛微生境, 进而为禾本科植物的种子萌发提供一个良好的环境(
Su et al ., 2005 ).随着植物群落的变化, 土壤种子库也随之改变(
赵丽娅等, 2006 ;
赵盼盼等, 2018 ), 并且随着流动沙丘的固定和生境的改善, 植物群落和土壤种子库之间的联系变得更加紧密(
Wang et al ., 2019 ).封育对土壤环境也有很大的改善, 对封育和放牧样地土壤性质进行比较, 发现封育样地土壤容重、紧实度及粗颗粒含量显著小于自由放牧样地(
单贵莲等, 2009 ), 而封育样地土壤有机质、全氮、有效氮和有效钾含量显著高于放牧样地(
张建鹏等, 2017 ).封育年限长短也是影响植物群落恢复的重要因素, 适当封育有利于植被恢复, 但过度封育反而会引起植被退化,
高凯等(2013) 在内蒙古草原的研究发现, 封育年限超过14年会使多年生禾草以及灌木和半灌木优势度显著降低, 而多年生杂类草优势度出现不同程度的升高. ...
... 退化沙地恢复的速度和演替方向一方面取决于退化程度和干扰的消除程度, 另一方面取决于土壤、气候等环境条件(
赵哈林等, 2003 ).本研究中流动沙丘13年封育期间植被盖度和物种丰富度显著增加, 但群落生物量没有显著变化, 整体上群落在逐渐恢复, 但速度缓慢, 群落生产力有限.许多研究发现干扰退化草地在短期封育下可以快速恢复(
赵哈林等, 2009 ;
苗仁辉等, 2013 ;
吕朋等, 2018 ), 其根本原因是现存群落具有拓殖能力和退化群落资源过剩(
王炜等, 1996 ), 但对于流动沙丘, 群落资源处于极其匮乏的状态, 现存群落拓殖能力有限, 土壤种子库基础也相对薄弱, 因此植被恢复受到很大限制.另外植被恢复也受时间积累效应的影响, 物种要进入某一环境空间并建立种群, 需要有足够的时间去完成其繁殖体从种源地到目标空间的传播、散布和在目标空间的定居、繁殖过程, 这一过程进行的速度直接影响物种多样性恢复的进度(
张继义等, 2004 ), 本研究中流动沙丘封育仅13年, 植物建群时间有限, 因此时间积累可能相对不足, 植被恢复程度有限.有研究认为在降水条件较好的半干旱沙区, 采取封沙育草措施后10-15年的时间, 植被能够自然恢复(
张华等, 2003 ), 然而本研究的结果显示, 流动沙丘恢复到灌木、半灌木或多年生草本优势度较高的固定沙丘群落需要更长的时间, 因此封育年限的设定应综合考虑植被退化程度、气候条件以及土壤环境等因素. ...
科尔沁沙地固沙植物根系与土壤水分特征研究
1
2015
... 退化沙地土壤环境的恢复速率直接影响植被的演替进程, 土壤环境的改善是植被对土壤作用的结果, 但又反馈于植被的演替(
赵哈林等, 2006 ).我们的监测结果显示3种封育沙地在10年内仅有土壤有效氮和有效磷含量增加, 并且增加幅度不大, 土壤养分缓慢的积累速率对植被恢复有很大的限制作用, 尤其是在养分含量极低的流动沙丘这种限制作用可能会更大, 因此封育13年的流动沙丘群落生物量没有显著增加.另外, 土壤养分的空间异质性直接影响植被分布模式(
Zuo et al ., 2014 ).本研究3种沙地土壤环境存在显著差异, 群落物种与土壤因子之间的CCA分析也显示不同沙地群落物种受不同因素的影响, 其中沙质草地植物群落物种与土壤养分之间相关性很强, 固定沙丘群落物种和土壤水分联系紧密, 而流动沙丘群落物种与各土壤因子并不相关.3类沙地中沙质草地土壤养分和水分条件最优, 其相应的群落生物量和物种密度也最高, 但沙质草地多为草本植物, 密布的根系层对降水存在滞留作用(
牛存洋等, 2015 ), 另外草本植物相比于灌木和半灌木耗水相对较少, 并且较高的植被盖度产生的地表遮阴效果也能有效减弱土壤蒸发(
常昌明等, 2016 ), 这些因素的综合作用使其土壤水分含量最高, 进而使水分对群落的限制作用可能弱于土壤养分.而在固定沙丘土壤水分含量最低, 这是因为在固定沙丘优势物种主要为根系较深、耗水强烈的差不嘎蒿、小叶锦鸡儿等灌木, 使得土壤水分消耗强烈, 导致群落主要受土壤水分的限制.流动沙丘由于表面干沙层的存在抑制了土壤水分的蒸发(
段利民, 2011 ), 其土壤水分含量反而高于固定沙丘, 但其土壤养分和种子库非常贫瘠, 因此二者可能共同限制了群落的恢复. ...
科尔沁沙地固沙植物根系与土壤水分特征研究
1
2015
... 退化沙地土壤环境的恢复速率直接影响植被的演替进程, 土壤环境的改善是植被对土壤作用的结果, 但又反馈于植被的演替(
赵哈林等, 2006 ).我们的监测结果显示3种封育沙地在10年内仅有土壤有效氮和有效磷含量增加, 并且增加幅度不大, 土壤养分缓慢的积累速率对植被恢复有很大的限制作用, 尤其是在养分含量极低的流动沙丘这种限制作用可能会更大, 因此封育13年的流动沙丘群落生物量没有显著增加.另外, 土壤养分的空间异质性直接影响植被分布模式(
Zuo et al ., 2014 ).本研究3种沙地土壤环境存在显著差异, 群落物种与土壤因子之间的CCA分析也显示不同沙地群落物种受不同因素的影响, 其中沙质草地植物群落物种与土壤养分之间相关性很强, 固定沙丘群落物种和土壤水分联系紧密, 而流动沙丘群落物种与各土壤因子并不相关.3类沙地中沙质草地土壤养分和水分条件最优, 其相应的群落生物量和物种密度也最高, 但沙质草地多为草本植物, 密布的根系层对降水存在滞留作用(
牛存洋等, 2015 ), 另外草本植物相比于灌木和半灌木耗水相对较少, 并且较高的植被盖度产生的地表遮阴效果也能有效减弱土壤蒸发(
常昌明等, 2016 ), 这些因素的综合作用使其土壤水分含量最高, 进而使水分对群落的限制作用可能弱于土壤养分.而在固定沙丘土壤水分含量最低, 这是因为在固定沙丘优势物种主要为根系较深、耗水强烈的差不嘎蒿、小叶锦鸡儿等灌木, 使得土壤水分消耗强烈, 导致群落主要受土壤水分的限制.流动沙丘由于表面干沙层的存在抑制了土壤水分的蒸发(
段利民, 2011 ), 其土壤水分含量反而高于固定沙丘, 但其土壤养分和种子库非常贫瘠, 因此二者可能共同限制了群落的恢复. ...
Can soil seed banks contribute to the restoration of dune slacks under conservation management?
2
2009
... 式中,
S 为样方物种总数,
Pi 为第
i 物种密度占群落总密度的比例.用单因素方差分析来检验不同年份和沙地土壤水分、养分以及土壤有机质的差异性.群落物种丰富度、土壤种子丰富度以及土壤因子作为矩阵(
Plassmann et al ., 2009 ;
Yassir et al ., 2010 ), 应用除趋势对应分析(DCA)法来分析土壤种子库与地上群落之间的相似性, 应用典型相关分析(CCA)来分析植物群落和土壤因子之间的关系.所有统计分析均在R-3.2.2中完成, 其中单因素方差分析用‘agricolae’包实现, DCA和CCA分析以及群落
C 和
H 用‘vegan’包计算, 插图用‘ggplot2’包和Origin 2018绘制. ...
... 气候是植被恢复的外在驱动力, 决定群落演替的方向和终点, 而植被的自我修复是内在驱动力, 决定着植物恢复的起始速度和演替速度(
赵哈林等, 2006 ).植被自我修复的基础是土壤种子库,
Plassmann等(2009) 研究发现相比于当前群落, 土壤种子库更能反映演替初期的植被组成, 但在我们的研究中发现种子丰富度变化与群落物种丰富度变化相似, DCA分析也发现地上植物群落物种和土壤种子库之间有很高的相似性, 这反映出科尔沁沙地植被恢复和土壤种子库变化存在较强的关联性.
Wang等(2019) 在科尔沁沙地的研究也表明植被恢复过程中地上群落和土壤种子库之间存在紧密联系, 并且随生境恢复二者之间的联系更加紧密. ...
1
2004
... 退化沙质草地封育恢复是群落生物量和物种多样性显著增加, 多年生植物优势度提高, 群落结构逐渐趋于稳定, 群落生态功能不断增强的过程(
张继义等, 2004 ).在本研究中封育21年的沙质草地群落生物量不存在显著的变化趋势, 但群落物种丰富度显著下降, 多年生禾草功能群重要值也不断降低, 这也与正常退化草地封育恢复过程中的群落演替相反, 反映出固定沙丘群落在观测期间存在退化趋势.根据生态系统可持续性原理(
戎郁萍等, 2004 ), 退化草地的封育时间不应是无限期的, 我们的研究结果也显示封育时间过长不仅不利于群落恢复, 反而会导致群落发生逆行演替, 因此封育一段时间后应适当实施刈割、放牧等干扰措施, 刺激牧草分蘖, 促进植物再生, 改善沙质草地质量(
程积民等, 1995 ), 也有研究发现轻度放牧是内蒙古草原最有利的草地利用方式(
Gao et al ., 2015 ). ...
1
2004
... 退化沙质草地封育恢复是群落生物量和物种多样性显著增加, 多年生植物优势度提高, 群落结构逐渐趋于稳定, 群落生态功能不断增强的过程(
张继义等, 2004 ).在本研究中封育21年的沙质草地群落生物量不存在显著的变化趋势, 但群落物种丰富度显著下降, 多年生禾草功能群重要值也不断降低, 这也与正常退化草地封育恢复过程中的群落演替相反, 反映出固定沙丘群落在观测期间存在退化趋势.根据生态系统可持续性原理(
戎郁萍等, 2004 ), 退化草地的封育时间不应是无限期的, 我们的研究结果也显示封育时间过长不仅不利于群落恢复, 反而会导致群落发生逆行演替, 因此封育一段时间后应适当实施刈割、放牧等干扰措施, 刺激牧草分蘖, 促进植物再生, 改善沙质草地质量(
程积民等, 1995 ), 也有研究发现轻度放牧是内蒙古草原最有利的草地利用方式(
Gao et al ., 2015 ). ...
Legacies of precipitation fluctuations on primary production: Theory and data synthesis
1
2012
... 大多数陆地生态系统天然植物群落都是特定气候条件下的产物(
赵哈林等, 2003 ), 气候条件的变化, 特别是气温和降水的变化会引起植物群落特征的变化(
Liu & Lei, 2015 ;
Liu et al ., 2016 ).
赵哈林等(2006) 认为气候是我国北方植被恢复的外在驱动力, 决定着演替的方向和终点.然而我们13年的观测结果显示群落生物量和物种丰富度与降水和气温之间不存在显著关系, 但气温和降水与生物量的线性回归斜率随着沙地的恢复而逐渐增大, 斜率的大小反映出植物群落对降水年际变化的响应敏感性(
Sala et al ., 2012 ), 在一定程度上也反映出群落对降水的利用效率(
Bai et al ., 2008 ).随着沙地生境恢复, 群落对年际降水波动的响应敏感性逐渐增加, 降水利用效率也相应地提升, 这种变化可能与资源的可利用性密切相关.许多研究发现土壤氮等资源的可利用性越高, 植被对降水变化的响应越敏感(
Wilcox et al ., 2016 ,
2017 ;
Knapp et al ., 2017 ), 在科尔沁沙地大尺度的研究也发现土壤养分梯度是影响植物群落分布最主要的因素(
左小安等, 2007 ).本研究中沙质草地土壤环境条件最优, 固定沙丘次之, 流动沙丘最差, 因此这种资源分布的差异导致不同沙地群落对降水变化的响应不同.群落组成也会影响群落对降水变化的响应.通过对生活型功能群重要值的年变化分析发现沙质草地群落不同功能群之间存在补偿效应, 这种补偿效应的存在可以有效地减缓降水等气候因子变化带来的影响.
Bai等(2004) 在内蒙古草原的研究也发现群落功能群之间存在补偿效应, 这种效应的存在更有利于群落稳定性的维持.这些因素的综合作用可能导致了群落生物量年变化对降水波动响应微弱.然而需要注意的是许多关于降水与群落生产力之间关系的结论都是在长时间尺度或者大空间尺度统计得来的(
Knapp et al ., 2017 ;
Wilcox et al ., 2017 ), 而本研究仅是在时间跨度为13年的小区域中开展的, 因此还需要更长时间的观测资料来研究群落对降水变化的响应.年内生物量积累与积温和累积降水量显著相关, 甚至在沙质草地二者对生物量年内积累的解释率达到68%, 因此积温和累积降水量是年内生物量积累的重要驱动因素, 这与植被生长过程中的水热需求规律一致. ...
围封年限对典型草原植被与土壤特征的影响
1
2009
... 科尔沁沙地作为中国北方最严重的一个沙漠化区域(
刘新民等, 1999 ), 封育禁牧被广泛用来恢复退化沙地植被, 同时也有许多研究对封育所产生的生态效应做了评述.如退化沙质草地封育后群落生物量响应最为敏感, 在较短时间内群落生物量迅速恢复, 同时凋落物量也显著增加(
苗仁辉等, 2013 ;
赵丽娅等, 2017 ;
吕朋等, 2018 ).随着封育时间的延长, 群落物种组成开始发生变化, 流动沙丘封育后沙蓬(
Agriophyllum squarrosum )作为先锋物种首先建立种群, 使沙丘流沙面积减小, 之后以差不嘎蒿(
Artemisia halodendron )为主的多年生灌木通过无性繁殖的方式开始在沙丘定植, 差不嘎蒿等灌木的出现逐渐营造出养分和水分相对富集的灌丛微生境, 进而为禾本科植物的种子萌发提供一个良好的环境(
Su et al ., 2005 ).随着植物群落的变化, 土壤种子库也随之改变(
赵丽娅等, 2006 ;
赵盼盼等, 2018 ), 并且随着流动沙丘的固定和生境的改善, 植物群落和土壤种子库之间的联系变得更加紧密(
Wang et al ., 2019 ).封育对土壤环境也有很大的改善, 对封育和放牧样地土壤性质进行比较, 发现封育样地土壤容重、紧实度及粗颗粒含量显著小于自由放牧样地(
单贵莲等, 2009 ), 而封育样地土壤有机质、全氮、有效氮和有效钾含量显著高于放牧样地(
张建鹏等, 2017 ).封育年限长短也是影响植物群落恢复的重要因素, 适当封育有利于植被恢复, 但过度封育反而会引起植被退化,
高凯等(2013) 在内蒙古草原的研究发现, 封育年限超过14年会使多年生禾草以及灌木和半灌木优势度显著降低, 而多年生杂类草优势度出现不同程度的升高. ...
围封年限对典型草原植被与土壤特征的影响
1
2009
... 科尔沁沙地作为中国北方最严重的一个沙漠化区域(
刘新民等, 1999 ), 封育禁牧被广泛用来恢复退化沙地植被, 同时也有许多研究对封育所产生的生态效应做了评述.如退化沙质草地封育后群落生物量响应最为敏感, 在较短时间内群落生物量迅速恢复, 同时凋落物量也显著增加(
苗仁辉等, 2013 ;
赵丽娅等, 2017 ;
吕朋等, 2018 ).随着封育时间的延长, 群落物种组成开始发生变化, 流动沙丘封育后沙蓬(
Agriophyllum squarrosum )作为先锋物种首先建立种群, 使沙丘流沙面积减小, 之后以差不嘎蒿(
Artemisia halodendron )为主的多年生灌木通过无性繁殖的方式开始在沙丘定植, 差不嘎蒿等灌木的出现逐渐营造出养分和水分相对富集的灌丛微生境, 进而为禾本科植物的种子萌发提供一个良好的环境(
Su et al ., 2005 ).随着植物群落的变化, 土壤种子库也随之改变(
赵丽娅等, 2006 ;
赵盼盼等, 2018 ), 并且随着流动沙丘的固定和生境的改善, 植物群落和土壤种子库之间的联系变得更加紧密(
Wang et al ., 2019 ).封育对土壤环境也有很大的改善, 对封育和放牧样地土壤性质进行比较, 发现封育样地土壤容重、紧实度及粗颗粒含量显著小于自由放牧样地(
单贵莲等, 2009 ), 而封育样地土壤有机质、全氮、有效氮和有效钾含量显著高于放牧样地(
张建鹏等, 2017 ).封育年限长短也是影响植物群落恢复的重要因素, 适当封育有利于植被恢复, 但过度封育反而会引起植被退化,
高凯等(2013) 在内蒙古草原的研究发现, 封育年限超过14年会使多年生禾草以及灌木和半灌木优势度显著降低, 而多年生杂类草优势度出现不同程度的升高. ...
Changes in soil properties after establishment of
Artemisia halodendron and
Caragana microphylla on shifting sand dunes in semiarid Horqin Sandy Land, Northern China
1
2005
... 科尔沁沙地作为中国北方最严重的一个沙漠化区域(
刘新民等, 1999 ), 封育禁牧被广泛用来恢复退化沙地植被, 同时也有许多研究对封育所产生的生态效应做了评述.如退化沙质草地封育后群落生物量响应最为敏感, 在较短时间内群落生物量迅速恢复, 同时凋落物量也显著增加(
苗仁辉等, 2013 ;
赵丽娅等, 2017 ;
吕朋等, 2018 ).随着封育时间的延长, 群落物种组成开始发生变化, 流动沙丘封育后沙蓬(
Agriophyllum squarrosum )作为先锋物种首先建立种群, 使沙丘流沙面积减小, 之后以差不嘎蒿(
Artemisia halodendron )为主的多年生灌木通过无性繁殖的方式开始在沙丘定植, 差不嘎蒿等灌木的出现逐渐营造出养分和水分相对富集的灌丛微生境, 进而为禾本科植物的种子萌发提供一个良好的环境(
Su et al ., 2005 ).随着植物群落的变化, 土壤种子库也随之改变(
赵丽娅等, 2006 ;
赵盼盼等, 2018 ), 并且随着流动沙丘的固定和生境的改善, 植物群落和土壤种子库之间的联系变得更加紧密(
Wang et al ., 2019 ).封育对土壤环境也有很大的改善, 对封育和放牧样地土壤性质进行比较, 发现封育样地土壤容重、紧实度及粗颗粒含量显著小于自由放牧样地(
单贵莲等, 2009 ), 而封育样地土壤有机质、全氮、有效氮和有效钾含量显著高于放牧样地(
张建鹏等, 2017 ).封育年限长短也是影响植物群落恢复的重要因素, 适当封育有利于植被恢复, 但过度封育反而会引起植被退化,
高凯等(2013) 在内蒙古草原的研究发现, 封育年限超过14年会使多年生禾草以及灌木和半灌木优势度显著降低, 而多年生杂类草优势度出现不同程度的升高. ...
Benefits derived from rehabilitating a degraded semi-arid rangeland in private enclosures in West Pokot County, Kenya
1
2016
... 封育恢复在一定时期内通过排除家畜采食、践踏等干扰使群落营养物质得以贮藏, 进而促进植被生产力恢复和群落自然更新, 由于其投资少、见效快, 已成为当前退化生态系统恢复与重建的重要措施, 并为世界各国所广泛采用(
Wairore et al ., 2016 ).封育为退化生态系统所带来的生态效应受许多因素的影响, 地区、封育时间以及封育方式都会使生态系统产生不同甚至相反的响应, 因此退化生态系统各要素对封育的响应机制、适宜的封育时间尺度以及封育方式等研究都受到国内外****的广泛关注(
闫玉春等, 2009 ). ...
内蒙古草原退化群落恢复演替的研究——I. 退化草原的基本特征与恢复演替动力
1
1996
... 退化沙地恢复的速度和演替方向一方面取决于退化程度和干扰的消除程度, 另一方面取决于土壤、气候等环境条件(
赵哈林等, 2003 ).本研究中流动沙丘13年封育期间植被盖度和物种丰富度显著增加, 但群落生物量没有显著变化, 整体上群落在逐渐恢复, 但速度缓慢, 群落生产力有限.许多研究发现干扰退化草地在短期封育下可以快速恢复(
赵哈林等, 2009 ;
苗仁辉等, 2013 ;
吕朋等, 2018 ), 其根本原因是现存群落具有拓殖能力和退化群落资源过剩(
王炜等, 1996 ), 但对于流动沙丘, 群落资源处于极其匮乏的状态, 现存群落拓殖能力有限, 土壤种子库基础也相对薄弱, 因此植被恢复受到很大限制.另外植被恢复也受时间积累效应的影响, 物种要进入某一环境空间并建立种群, 需要有足够的时间去完成其繁殖体从种源地到目标空间的传播、散布和在目标空间的定居、繁殖过程, 这一过程进行的速度直接影响物种多样性恢复的进度(
张继义等, 2004 ), 本研究中流动沙丘封育仅13年, 植物建群时间有限, 因此时间积累可能相对不足, 植被恢复程度有限.有研究认为在降水条件较好的半干旱沙区, 采取封沙育草措施后10-15年的时间, 植被能够自然恢复(
张华等, 2003 ), 然而本研究的结果显示, 流动沙丘恢复到灌木、半灌木或多年生草本优势度较高的固定沙丘群落需要更长的时间, 因此封育年限的设定应综合考虑植被退化程度、气候条件以及土壤环境等因素. ...
内蒙古草原退化群落恢复演替的研究——I. 退化草原的基本特征与恢复演替动力
1
1996
... 退化沙地恢复的速度和演替方向一方面取决于退化程度和干扰的消除程度, 另一方面取决于土壤、气候等环境条件(
赵哈林等, 2003 ).本研究中流动沙丘13年封育期间植被盖度和物种丰富度显著增加, 但群落生物量没有显著变化, 整体上群落在逐渐恢复, 但速度缓慢, 群落生产力有限.许多研究发现干扰退化草地在短期封育下可以快速恢复(
赵哈林等, 2009 ;
苗仁辉等, 2013 ;
吕朋等, 2018 ), 其根本原因是现存群落具有拓殖能力和退化群落资源过剩(
王炜等, 1996 ), 但对于流动沙丘, 群落资源处于极其匮乏的状态, 现存群落拓殖能力有限, 土壤种子库基础也相对薄弱, 因此植被恢复受到很大限制.另外植被恢复也受时间积累效应的影响, 物种要进入某一环境空间并建立种群, 需要有足够的时间去完成其繁殖体从种源地到目标空间的传播、散布和在目标空间的定居、繁殖过程, 这一过程进行的速度直接影响物种多样性恢复的进度(
张继义等, 2004 ), 本研究中流动沙丘封育仅13年, 植物建群时间有限, 因此时间积累可能相对不足, 植被恢复程度有限.有研究认为在降水条件较好的半干旱沙区, 采取封沙育草措施后10-15年的时间, 植被能够自然恢复(
张华等, 2003 ), 然而本研究的结果显示, 流动沙丘恢复到灌木、半灌木或多年生草本优势度较高的固定沙丘群落需要更长的时间, 因此封育年限的设定应综合考虑植被退化程度、气候条件以及土壤环境等因素. ...
Sand dune stabilization changes the vegetation characteristics and soil seed bank and their correlations with environmental factors
2
2019
... 科尔沁沙地作为中国北方最严重的一个沙漠化区域(
刘新民等, 1999 ), 封育禁牧被广泛用来恢复退化沙地植被, 同时也有许多研究对封育所产生的生态效应做了评述.如退化沙质草地封育后群落生物量响应最为敏感, 在较短时间内群落生物量迅速恢复, 同时凋落物量也显著增加(
苗仁辉等, 2013 ;
赵丽娅等, 2017 ;
吕朋等, 2018 ).随着封育时间的延长, 群落物种组成开始发生变化, 流动沙丘封育后沙蓬(
Agriophyllum squarrosum )作为先锋物种首先建立种群, 使沙丘流沙面积减小, 之后以差不嘎蒿(
Artemisia halodendron )为主的多年生灌木通过无性繁殖的方式开始在沙丘定植, 差不嘎蒿等灌木的出现逐渐营造出养分和水分相对富集的灌丛微生境, 进而为禾本科植物的种子萌发提供一个良好的环境(
Su et al ., 2005 ).随着植物群落的变化, 土壤种子库也随之改变(
赵丽娅等, 2006 ;
赵盼盼等, 2018 ), 并且随着流动沙丘的固定和生境的改善, 植物群落和土壤种子库之间的联系变得更加紧密(
Wang et al ., 2019 ).封育对土壤环境也有很大的改善, 对封育和放牧样地土壤性质进行比较, 发现封育样地土壤容重、紧实度及粗颗粒含量显著小于自由放牧样地(
单贵莲等, 2009 ), 而封育样地土壤有机质、全氮、有效氮和有效钾含量显著高于放牧样地(
张建鹏等, 2017 ).封育年限长短也是影响植物群落恢复的重要因素, 适当封育有利于植被恢复, 但过度封育反而会引起植被退化,
高凯等(2013) 在内蒙古草原的研究发现, 封育年限超过14年会使多年生禾草以及灌木和半灌木优势度显著降低, 而多年生杂类草优势度出现不同程度的升高. ...
... 气候是植被恢复的外在驱动力, 决定群落演替的方向和终点, 而植被的自我修复是内在驱动力, 决定着植物恢复的起始速度和演替速度(
赵哈林等, 2006 ).植被自我修复的基础是土壤种子库,
Plassmann等(2009) 研究发现相比于当前群落, 土壤种子库更能反映演替初期的植被组成, 但在我们的研究中发现种子丰富度变化与群落物种丰富度变化相似, DCA分析也发现地上植物群落物种和土壤种子库之间有很高的相似性, 这反映出科尔沁沙地植被恢复和土壤种子库变化存在较强的关联性.
Wang等(2019) 在科尔沁沙地的研究也表明植被恢复过程中地上群落和土壤种子库之间存在紧密联系, 并且随生境恢复二者之间的联系更加紧密. ...
Does ecosystem sensitivity to precipitation at the site-level conform to regional-scale predictions?
1
2016
... 大多数陆地生态系统天然植物群落都是特定气候条件下的产物(
赵哈林等, 2003 ), 气候条件的变化, 特别是气温和降水的变化会引起植物群落特征的变化(
Liu & Lei, 2015 ;
Liu et al ., 2016 ).
赵哈林等(2006) 认为气候是我国北方植被恢复的外在驱动力, 决定着演替的方向和终点.然而我们13年的观测结果显示群落生物量和物种丰富度与降水和气温之间不存在显著关系, 但气温和降水与生物量的线性回归斜率随着沙地的恢复而逐渐增大, 斜率的大小反映出植物群落对降水年际变化的响应敏感性(
Sala et al ., 2012 ), 在一定程度上也反映出群落对降水的利用效率(
Bai et al ., 2008 ).随着沙地生境恢复, 群落对年际降水波动的响应敏感性逐渐增加, 降水利用效率也相应地提升, 这种变化可能与资源的可利用性密切相关.许多研究发现土壤氮等资源的可利用性越高, 植被对降水变化的响应越敏感(
Wilcox et al ., 2016 ,
2017 ;
Knapp et al ., 2017 ), 在科尔沁沙地大尺度的研究也发现土壤养分梯度是影响植物群落分布最主要的因素(
左小安等, 2007 ).本研究中沙质草地土壤环境条件最优, 固定沙丘次之, 流动沙丘最差, 因此这种资源分布的差异导致不同沙地群落对降水变化的响应不同.群落组成也会影响群落对降水变化的响应.通过对生活型功能群重要值的年变化分析发现沙质草地群落不同功能群之间存在补偿效应, 这种补偿效应的存在可以有效地减缓降水等气候因子变化带来的影响.
Bai等(2004) 在内蒙古草原的研究也发现群落功能群之间存在补偿效应, 这种效应的存在更有利于群落稳定性的维持.这些因素的综合作用可能导致了群落生物量年变化对降水波动响应微弱.然而需要注意的是许多关于降水与群落生产力之间关系的结论都是在长时间尺度或者大空间尺度统计得来的(
Knapp et al ., 2017 ;
Wilcox et al ., 2017 ), 而本研究仅是在时间跨度为13年的小区域中开展的, 因此还需要更长时间的观测资料来研究群落对降水变化的响应.年内生物量积累与积温和累积降水量显著相关, 甚至在沙质草地二者对生物量年内积累的解释率达到68%, 因此积温和累积降水量是年内生物量积累的重要驱动因素, 这与植被生长过程中的水热需求规律一致. ...
Asymmetric responses of primary productivity to precipitation extremes: A synthesis of grassland precipitation manipulation experiments
2
2017
... 大多数陆地生态系统天然植物群落都是特定气候条件下的产物(
赵哈林等, 2003 ), 气候条件的变化, 特别是气温和降水的变化会引起植物群落特征的变化(
Liu & Lei, 2015 ;
Liu et al ., 2016 ).
赵哈林等(2006) 认为气候是我国北方植被恢复的外在驱动力, 决定着演替的方向和终点.然而我们13年的观测结果显示群落生物量和物种丰富度与降水和气温之间不存在显著关系, 但气温和降水与生物量的线性回归斜率随着沙地的恢复而逐渐增大, 斜率的大小反映出植物群落对降水年际变化的响应敏感性(
Sala et al ., 2012 ), 在一定程度上也反映出群落对降水的利用效率(
Bai et al ., 2008 ).随着沙地生境恢复, 群落对年际降水波动的响应敏感性逐渐增加, 降水利用效率也相应地提升, 这种变化可能与资源的可利用性密切相关.许多研究发现土壤氮等资源的可利用性越高, 植被对降水变化的响应越敏感(
Wilcox et al ., 2016 ,
2017 ;
Knapp et al ., 2017 ), 在科尔沁沙地大尺度的研究也发现土壤养分梯度是影响植物群落分布最主要的因素(
左小安等, 2007 ).本研究中沙质草地土壤环境条件最优, 固定沙丘次之, 流动沙丘最差, 因此这种资源分布的差异导致不同沙地群落对降水变化的响应不同.群落组成也会影响群落对降水变化的响应.通过对生活型功能群重要值的年变化分析发现沙质草地群落不同功能群之间存在补偿效应, 这种补偿效应的存在可以有效地减缓降水等气候因子变化带来的影响.
Bai等(2004) 在内蒙古草原的研究也发现群落功能群之间存在补偿效应, 这种效应的存在更有利于群落稳定性的维持.这些因素的综合作用可能导致了群落生物量年变化对降水波动响应微弱.然而需要注意的是许多关于降水与群落生产力之间关系的结论都是在长时间尺度或者大空间尺度统计得来的(
Knapp et al ., 2017 ;
Wilcox et al ., 2017 ), 而本研究仅是在时间跨度为13年的小区域中开展的, 因此还需要更长时间的观测资料来研究群落对降水变化的响应.年内生物量积累与积温和累积降水量显著相关, 甚至在沙质草地二者对生物量年内积累的解释率达到68%, 因此积温和累积降水量是年内生物量积累的重要驱动因素, 这与植被生长过程中的水热需求规律一致. ...
... ;
Wilcox et al ., 2017 ), 而本研究仅是在时间跨度为13年的小区域中开展的, 因此还需要更长时间的观测资料来研究群落对降水变化的响应.年内生物量积累与积温和累积降水量显著相关, 甚至在沙质草地二者对生物量年内积累的解释率达到68%, 因此积温和累积降水量是年内生物量积累的重要驱动因素, 这与植被生长过程中的水热需求规律一致. ...
围封对草地的影响研究进展
1
2009
... 封育恢复在一定时期内通过排除家畜采食、践踏等干扰使群落营养物质得以贮藏, 进而促进植被生产力恢复和群落自然更新, 由于其投资少、见效快, 已成为当前退化生态系统恢复与重建的重要措施, 并为世界各国所广泛采用(
Wairore et al ., 2016 ).封育为退化生态系统所带来的生态效应受许多因素的影响, 地区、封育时间以及封育方式都会使生态系统产生不同甚至相反的响应, 因此退化生态系统各要素对封育的响应机制、适宜的封育时间尺度以及封育方式等研究都受到国内外****的广泛关注(
闫玉春等, 2009 ). ...
围封对草地的影响研究进展
1
2009
... 封育恢复在一定时期内通过排除家畜采食、践踏等干扰使群落营养物质得以贮藏, 进而促进植被生产力恢复和群落自然更新, 由于其投资少、见效快, 已成为当前退化生态系统恢复与重建的重要措施, 并为世界各国所广泛采用(
Wairore et al ., 2016 ).封育为退化生态系统所带来的生态效应受许多因素的影响, 地区、封育时间以及封育方式都会使生态系统产生不同甚至相反的响应, 因此退化生态系统各要素对封育的响应机制、适宜的封育时间尺度以及封育方式等研究都受到国内外****的广泛关注(
闫玉春等, 2009 ). ...
Secondary succession after fire in
Imperata grasslands of East Kalimantan, Indonesia
1
2010
... 式中,
S 为样方物种总数,
Pi 为第
i 物种密度占群落总密度的比例.用单因素方差分析来检验不同年份和沙地土壤水分、养分以及土壤有机质的差异性.群落物种丰富度、土壤种子丰富度以及土壤因子作为矩阵(
Plassmann et al ., 2009 ;
Yassir et al ., 2010 ), 应用除趋势对应分析(DCA)法来分析土壤种子库与地上群落之间的相似性, 应用典型相关分析(CCA)来分析植物群落和土壤因子之间的关系.所有统计分析均在R-3.2.2中完成, 其中单因素方差分析用‘agricolae’包实现, DCA和CCA分析以及群落
C 和
H 用‘vegan’包计算, 插图用‘ggplot2’包和Origin 2018绘制. ...
退化沙质草地自然恢复过程中土壤-植物系统的变化特征
1
2003
... 退化沙地恢复的速度和演替方向一方面取决于退化程度和干扰的消除程度, 另一方面取决于土壤、气候等环境条件(
赵哈林等, 2003 ).本研究中流动沙丘13年封育期间植被盖度和物种丰富度显著增加, 但群落生物量没有显著变化, 整体上群落在逐渐恢复, 但速度缓慢, 群落生产力有限.许多研究发现干扰退化草地在短期封育下可以快速恢复(
赵哈林等, 2009 ;
苗仁辉等, 2013 ;
吕朋等, 2018 ), 其根本原因是现存群落具有拓殖能力和退化群落资源过剩(
王炜等, 1996 ), 但对于流动沙丘, 群落资源处于极其匮乏的状态, 现存群落拓殖能力有限, 土壤种子库基础也相对薄弱, 因此植被恢复受到很大限制.另外植被恢复也受时间积累效应的影响, 物种要进入某一环境空间并建立种群, 需要有足够的时间去完成其繁殖体从种源地到目标空间的传播、散布和在目标空间的定居、繁殖过程, 这一过程进行的速度直接影响物种多样性恢复的进度(
张继义等, 2004 ), 本研究中流动沙丘封育仅13年, 植物建群时间有限, 因此时间积累可能相对不足, 植被恢复程度有限.有研究认为在降水条件较好的半干旱沙区, 采取封沙育草措施后10-15年的时间, 植被能够自然恢复(
张华等, 2003 ), 然而本研究的结果显示, 流动沙丘恢复到灌木、半灌木或多年生草本优势度较高的固定沙丘群落需要更长的时间, 因此封育年限的设定应综合考虑植被退化程度、气候条件以及土壤环境等因素. ...
退化沙质草地自然恢复过程中土壤-植物系统的变化特征
1
2003
... 退化沙地恢复的速度和演替方向一方面取决于退化程度和干扰的消除程度, 另一方面取决于土壤、气候等环境条件(
赵哈林等, 2003 ).本研究中流动沙丘13年封育期间植被盖度和物种丰富度显著增加, 但群落生物量没有显著变化, 整体上群落在逐渐恢复, 但速度缓慢, 群落生产力有限.许多研究发现干扰退化草地在短期封育下可以快速恢复(
赵哈林等, 2009 ;
苗仁辉等, 2013 ;
吕朋等, 2018 ), 其根本原因是现存群落具有拓殖能力和退化群落资源过剩(
王炜等, 1996 ), 但对于流动沙丘, 群落资源处于极其匮乏的状态, 现存群落拓殖能力有限, 土壤种子库基础也相对薄弱, 因此植被恢复受到很大限制.另外植被恢复也受时间积累效应的影响, 物种要进入某一环境空间并建立种群, 需要有足够的时间去完成其繁殖体从种源地到目标空间的传播、散布和在目标空间的定居、繁殖过程, 这一过程进行的速度直接影响物种多样性恢复的进度(
张继义等, 2004 ), 本研究中流动沙丘封育仅13年, 植物建群时间有限, 因此时间积累可能相对不足, 植被恢复程度有限.有研究认为在降水条件较好的半干旱沙区, 采取封沙育草措施后10-15年的时间, 植被能够自然恢复(
张华等, 2003 ), 然而本研究的结果显示, 流动沙丘恢复到灌木、半灌木或多年生草本优势度较高的固定沙丘群落需要更长的时间, 因此封育年限的设定应综合考虑植被退化程度、气候条件以及土壤环境等因素. ...
围封对沙漠化草地土壤理化性质和固碳潜力恢复的影响
1
2017
... 科尔沁沙地作为中国北方最严重的一个沙漠化区域(
刘新民等, 1999 ), 封育禁牧被广泛用来恢复退化沙地植被, 同时也有许多研究对封育所产生的生态效应做了评述.如退化沙质草地封育后群落生物量响应最为敏感, 在较短时间内群落生物量迅速恢复, 同时凋落物量也显著增加(
苗仁辉等, 2013 ;
赵丽娅等, 2017 ;
吕朋等, 2018 ).随着封育时间的延长, 群落物种组成开始发生变化, 流动沙丘封育后沙蓬(
Agriophyllum squarrosum )作为先锋物种首先建立种群, 使沙丘流沙面积减小, 之后以差不嘎蒿(
Artemisia halodendron )为主的多年生灌木通过无性繁殖的方式开始在沙丘定植, 差不嘎蒿等灌木的出现逐渐营造出养分和水分相对富集的灌丛微生境, 进而为禾本科植物的种子萌发提供一个良好的环境(
Su et al ., 2005 ).随着植物群落的变化, 土壤种子库也随之改变(
赵丽娅等, 2006 ;
赵盼盼等, 2018 ), 并且随着流动沙丘的固定和生境的改善, 植物群落和土壤种子库之间的联系变得更加紧密(
Wang et al ., 2019 ).封育对土壤环境也有很大的改善, 对封育和放牧样地土壤性质进行比较, 发现封育样地土壤容重、紧实度及粗颗粒含量显著小于自由放牧样地(
单贵莲等, 2009 ), 而封育样地土壤有机质、全氮、有效氮和有效钾含量显著高于放牧样地(
张建鹏等, 2017 ).封育年限长短也是影响植物群落恢复的重要因素, 适当封育有利于植被恢复, 但过度封育反而会引起植被退化,
高凯等(2013) 在内蒙古草原的研究发现, 封育年限超过14年会使多年生禾草以及灌木和半灌木优势度显著降低, 而多年生杂类草优势度出现不同程度的升高. ...
围封对沙漠化草地土壤理化性质和固碳潜力恢复的影响
1
2017
... 科尔沁沙地作为中国北方最严重的一个沙漠化区域(
刘新民等, 1999 ), 封育禁牧被广泛用来恢复退化沙地植被, 同时也有许多研究对封育所产生的生态效应做了评述.如退化沙质草地封育后群落生物量响应最为敏感, 在较短时间内群落生物量迅速恢复, 同时凋落物量也显著增加(
苗仁辉等, 2013 ;
赵丽娅等, 2017 ;
吕朋等, 2018 ).随着封育时间的延长, 群落物种组成开始发生变化, 流动沙丘封育后沙蓬(
Agriophyllum squarrosum )作为先锋物种首先建立种群, 使沙丘流沙面积减小, 之后以差不嘎蒿(
Artemisia halodendron )为主的多年生灌木通过无性繁殖的方式开始在沙丘定植, 差不嘎蒿等灌木的出现逐渐营造出养分和水分相对富集的灌丛微生境, 进而为禾本科植物的种子萌发提供一个良好的环境(
Su et al ., 2005 ).随着植物群落的变化, 土壤种子库也随之改变(
赵丽娅等, 2006 ;
赵盼盼等, 2018 ), 并且随着流动沙丘的固定和生境的改善, 植物群落和土壤种子库之间的联系变得更加紧密(
Wang et al ., 2019 ).封育对土壤环境也有很大的改善, 对封育和放牧样地土壤性质进行比较, 发现封育样地土壤容重、紧实度及粗颗粒含量显著小于自由放牧样地(
单贵莲等, 2009 ), 而封育样地土壤有机质、全氮、有效氮和有效钾含量显著高于放牧样地(
张建鹏等, 2017 ).封育年限长短也是影响植物群落恢复的重要因素, 适当封育有利于植被恢复, 但过度封育反而会引起植被退化,
高凯等(2013) 在内蒙古草原的研究发现, 封育年限超过14年会使多年生禾草以及灌木和半灌木优势度显著降低, 而多年生杂类草优势度出现不同程度的升高. ...
科尔沁沙地植被恢复系列上群落演替与物种多样性的恢复动态
2
2004
... 退化沙地恢复的速度和演替方向一方面取决于退化程度和干扰的消除程度, 另一方面取决于土壤、气候等环境条件(
赵哈林等, 2003 ).本研究中流动沙丘13年封育期间植被盖度和物种丰富度显著增加, 但群落生物量没有显著变化, 整体上群落在逐渐恢复, 但速度缓慢, 群落生产力有限.许多研究发现干扰退化草地在短期封育下可以快速恢复(
赵哈林等, 2009 ;
苗仁辉等, 2013 ;
吕朋等, 2018 ), 其根本原因是现存群落具有拓殖能力和退化群落资源过剩(
王炜等, 1996 ), 但对于流动沙丘, 群落资源处于极其匮乏的状态, 现存群落拓殖能力有限, 土壤种子库基础也相对薄弱, 因此植被恢复受到很大限制.另外植被恢复也受时间积累效应的影响, 物种要进入某一环境空间并建立种群, 需要有足够的时间去完成其繁殖体从种源地到目标空间的传播、散布和在目标空间的定居、繁殖过程, 这一过程进行的速度直接影响物种多样性恢复的进度(
张继义等, 2004 ), 本研究中流动沙丘封育仅13年, 植物建群时间有限, 因此时间积累可能相对不足, 植被恢复程度有限.有研究认为在降水条件较好的半干旱沙区, 采取封沙育草措施后10-15年的时间, 植被能够自然恢复(
张华等, 2003 ), 然而本研究的结果显示, 流动沙丘恢复到灌木、半灌木或多年生草本优势度较高的固定沙丘群落需要更长的时间, 因此封育年限的设定应综合考虑植被退化程度、气候条件以及土壤环境等因素. ...
... 退化沙质草地封育恢复是群落生物量和物种多样性显著增加, 多年生植物优势度提高, 群落结构逐渐趋于稳定, 群落生态功能不断增强的过程(
张继义等, 2004 ).在本研究中封育21年的沙质草地群落生物量不存在显著的变化趋势, 但群落物种丰富度显著下降, 多年生禾草功能群重要值也不断降低, 这也与正常退化草地封育恢复过程中的群落演替相反, 反映出固定沙丘群落在观测期间存在退化趋势.根据生态系统可持续性原理(
戎郁萍等, 2004 ), 退化草地的封育时间不应是无限期的, 我们的研究结果也显示封育时间过长不仅不利于群落恢复, 反而会导致群落发生逆行演替, 因此封育一段时间后应适当实施刈割、放牧等干扰措施, 刺激牧草分蘖, 促进植物再生, 改善沙质草地质量(
程积民等, 1995 ), 也有研究发现轻度放牧是内蒙古草原最有利的草地利用方式(
Gao et al ., 2015 ). ...
科尔沁沙地植被恢复系列上群落演替与物种多样性的恢复动态
2
2004
... 退化沙地恢复的速度和演替方向一方面取决于退化程度和干扰的消除程度, 另一方面取决于土壤、气候等环境条件(
赵哈林等, 2003 ).本研究中流动沙丘13年封育期间植被盖度和物种丰富度显著增加, 但群落生物量没有显著变化, 整体上群落在逐渐恢复, 但速度缓慢, 群落生产力有限.许多研究发现干扰退化草地在短期封育下可以快速恢复(
赵哈林等, 2009 ;
苗仁辉等, 2013 ;
吕朋等, 2018 ), 其根本原因是现存群落具有拓殖能力和退化群落资源过剩(
王炜等, 1996 ), 但对于流动沙丘, 群落资源处于极其匮乏的状态, 现存群落拓殖能力有限, 土壤种子库基础也相对薄弱, 因此植被恢复受到很大限制.另外植被恢复也受时间积累效应的影响, 物种要进入某一环境空间并建立种群, 需要有足够的时间去完成其繁殖体从种源地到目标空间的传播、散布和在目标空间的定居、繁殖过程, 这一过程进行的速度直接影响物种多样性恢复的进度(
张继义等, 2004 ), 本研究中流动沙丘封育仅13年, 植物建群时间有限, 因此时间积累可能相对不足, 植被恢复程度有限.有研究认为在降水条件较好的半干旱沙区, 采取封沙育草措施后10-15年的时间, 植被能够自然恢复(
张华等, 2003 ), 然而本研究的结果显示, 流动沙丘恢复到灌木、半灌木或多年生草本优势度较高的固定沙丘群落需要更长的时间, 因此封育年限的设定应综合考虑植被退化程度、气候条件以及土壤环境等因素. ...
... 退化沙质草地封育恢复是群落生物量和物种多样性显著增加, 多年生植物优势度提高, 群落结构逐渐趋于稳定, 群落生态功能不断增强的过程(
张继义等, 2004 ).在本研究中封育21年的沙质草地群落生物量不存在显著的变化趋势, 但群落物种丰富度显著下降, 多年生禾草功能群重要值也不断降低, 这也与正常退化草地封育恢复过程中的群落演替相反, 反映出固定沙丘群落在观测期间存在退化趋势.根据生态系统可持续性原理(
戎郁萍等, 2004 ), 退化草地的封育时间不应是无限期的, 我们的研究结果也显示封育时间过长不仅不利于群落恢复, 反而会导致群落发生逆行演替, 因此封育一段时间后应适当实施刈割、放牧等干扰措施, 刺激牧草分蘖, 促进植物再生, 改善沙质草地质量(
程积民等, 1995 ), 也有研究发现轻度放牧是内蒙古草原最有利的草地利用方式(
Gao et al ., 2015 ). ...
科尔沁沙质草地植物群落的放牧退化及其自然恢复过程
1
2009
... 退化沙地恢复的速度和演替方向一方面取决于退化程度和干扰的消除程度, 另一方面取决于土壤、气候等环境条件(
赵哈林等, 2003 ).本研究中流动沙丘13年封育期间植被盖度和物种丰富度显著增加, 但群落生物量没有显著变化, 整体上群落在逐渐恢复, 但速度缓慢, 群落生产力有限.许多研究发现干扰退化草地在短期封育下可以快速恢复(
赵哈林等, 2009 ;
苗仁辉等, 2013 ;
吕朋等, 2018 ), 其根本原因是现存群落具有拓殖能力和退化群落资源过剩(
王炜等, 1996 ), 但对于流动沙丘, 群落资源处于极其匮乏的状态, 现存群落拓殖能力有限, 土壤种子库基础也相对薄弱, 因此植被恢复受到很大限制.另外植被恢复也受时间积累效应的影响, 物种要进入某一环境空间并建立种群, 需要有足够的时间去完成其繁殖体从种源地到目标空间的传播、散布和在目标空间的定居、繁殖过程, 这一过程进行的速度直接影响物种多样性恢复的进度(
张继义等, 2004 ), 本研究中流动沙丘封育仅13年, 植物建群时间有限, 因此时间积累可能相对不足, 植被恢复程度有限.有研究认为在降水条件较好的半干旱沙区, 采取封沙育草措施后10-15年的时间, 植被能够自然恢复(
张华等, 2003 ), 然而本研究的结果显示, 流动沙丘恢复到灌木、半灌木或多年生草本优势度较高的固定沙丘群落需要更长的时间, 因此封育年限的设定应综合考虑植被退化程度、气候条件以及土壤环境等因素. ...
科尔沁沙质草地植物群落的放牧退化及其自然恢复过程
1
2009
... 退化沙地恢复的速度和演替方向一方面取决于退化程度和干扰的消除程度, 另一方面取决于土壤、气候等环境条件(
赵哈林等, 2003 ).本研究中流动沙丘13年封育期间植被盖度和物种丰富度显著增加, 但群落生物量没有显著变化, 整体上群落在逐渐恢复, 但速度缓慢, 群落生产力有限.许多研究发现干扰退化草地在短期封育下可以快速恢复(
赵哈林等, 2009 ;
苗仁辉等, 2013 ;
吕朋等, 2018 ), 其根本原因是现存群落具有拓殖能力和退化群落资源过剩(
王炜等, 1996 ), 但对于流动沙丘, 群落资源处于极其匮乏的状态, 现存群落拓殖能力有限, 土壤种子库基础也相对薄弱, 因此植被恢复受到很大限制.另外植被恢复也受时间积累效应的影响, 物种要进入某一环境空间并建立种群, 需要有足够的时间去完成其繁殖体从种源地到目标空间的传播、散布和在目标空间的定居、繁殖过程, 这一过程进行的速度直接影响物种多样性恢复的进度(
张继义等, 2004 ), 本研究中流动沙丘封育仅13年, 植物建群时间有限, 因此时间积累可能相对不足, 植被恢复程度有限.有研究认为在降水条件较好的半干旱沙区, 采取封沙育草措施后10-15年的时间, 植被能够自然恢复(
张华等, 2003 ), 然而本研究的结果显示, 流动沙丘恢复到灌木、半灌木或多年生草本优势度较高的固定沙丘群落需要更长的时间, 因此封育年限的设定应综合考虑植被退化程度、气候条件以及土壤环境等因素. ...
我国北方沙区退化植被的恢复机理
3
2006
... 大多数陆地生态系统天然植物群落都是特定气候条件下的产物(
赵哈林等, 2003 ), 气候条件的变化, 特别是气温和降水的变化会引起植物群落特征的变化(
Liu & Lei, 2015 ;
Liu et al ., 2016 ).
赵哈林等(2006) 认为气候是我国北方植被恢复的外在驱动力, 决定着演替的方向和终点.然而我们13年的观测结果显示群落生物量和物种丰富度与降水和气温之间不存在显著关系, 但气温和降水与生物量的线性回归斜率随着沙地的恢复而逐渐增大, 斜率的大小反映出植物群落对降水年际变化的响应敏感性(
Sala et al ., 2012 ), 在一定程度上也反映出群落对降水的利用效率(
Bai et al ., 2008 ).随着沙地生境恢复, 群落对年际降水波动的响应敏感性逐渐增加, 降水利用效率也相应地提升, 这种变化可能与资源的可利用性密切相关.许多研究发现土壤氮等资源的可利用性越高, 植被对降水变化的响应越敏感(
Wilcox et al ., 2016 ,
2017 ;
Knapp et al ., 2017 ), 在科尔沁沙地大尺度的研究也发现土壤养分梯度是影响植物群落分布最主要的因素(
左小安等, 2007 ).本研究中沙质草地土壤环境条件最优, 固定沙丘次之, 流动沙丘最差, 因此这种资源分布的差异导致不同沙地群落对降水变化的响应不同.群落组成也会影响群落对降水变化的响应.通过对生活型功能群重要值的年变化分析发现沙质草地群落不同功能群之间存在补偿效应, 这种补偿效应的存在可以有效地减缓降水等气候因子变化带来的影响.
Bai等(2004) 在内蒙古草原的研究也发现群落功能群之间存在补偿效应, 这种效应的存在更有利于群落稳定性的维持.这些因素的综合作用可能导致了群落生物量年变化对降水波动响应微弱.然而需要注意的是许多关于降水与群落生产力之间关系的结论都是在长时间尺度或者大空间尺度统计得来的(
Knapp et al ., 2017 ;
Wilcox et al ., 2017 ), 而本研究仅是在时间跨度为13年的小区域中开展的, 因此还需要更长时间的观测资料来研究群落对降水变化的响应.年内生物量积累与积温和累积降水量显著相关, 甚至在沙质草地二者对生物量年内积累的解释率达到68%, 因此积温和累积降水量是年内生物量积累的重要驱动因素, 这与植被生长过程中的水热需求规律一致. ...
... 气候是植被恢复的外在驱动力, 决定群落演替的方向和终点, 而植被的自我修复是内在驱动力, 决定着植物恢复的起始速度和演替速度(
赵哈林等, 2006 ).植被自我修复的基础是土壤种子库,
Plassmann等(2009) 研究发现相比于当前群落, 土壤种子库更能反映演替初期的植被组成, 但在我们的研究中发现种子丰富度变化与群落物种丰富度变化相似, DCA分析也发现地上植物群落物种和土壤种子库之间有很高的相似性, 这反映出科尔沁沙地植被恢复和土壤种子库变化存在较强的关联性.
Wang等(2019) 在科尔沁沙地的研究也表明植被恢复过程中地上群落和土壤种子库之间存在紧密联系, 并且随生境恢复二者之间的联系更加紧密. ...
... 退化沙地土壤环境的恢复速率直接影响植被的演替进程, 土壤环境的改善是植被对土壤作用的结果, 但又反馈于植被的演替(
赵哈林等, 2006 ).我们的监测结果显示3种封育沙地在10年内仅有土壤有效氮和有效磷含量增加, 并且增加幅度不大, 土壤养分缓慢的积累速率对植被恢复有很大的限制作用, 尤其是在养分含量极低的流动沙丘这种限制作用可能会更大, 因此封育13年的流动沙丘群落生物量没有显著增加.另外, 土壤养分的空间异质性直接影响植被分布模式(
Zuo et al ., 2014 ).本研究3种沙地土壤环境存在显著差异, 群落物种与土壤因子之间的CCA分析也显示不同沙地群落物种受不同因素的影响, 其中沙质草地植物群落物种与土壤养分之间相关性很强, 固定沙丘群落物种和土壤水分联系紧密, 而流动沙丘群落物种与各土壤因子并不相关.3类沙地中沙质草地土壤养分和水分条件最优, 其相应的群落生物量和物种密度也最高, 但沙质草地多为草本植物, 密布的根系层对降水存在滞留作用(
牛存洋等, 2015 ), 另外草本植物相比于灌木和半灌木耗水相对较少, 并且较高的植被盖度产生的地表遮阴效果也能有效减弱土壤蒸发(
常昌明等, 2016 ), 这些因素的综合作用使其土壤水分含量最高, 进而使水分对群落的限制作用可能弱于土壤养分.而在固定沙丘土壤水分含量最低, 这是因为在固定沙丘优势物种主要为根系较深、耗水强烈的差不嘎蒿、小叶锦鸡儿等灌木, 使得土壤水分消耗强烈, 导致群落主要受土壤水分的限制.流动沙丘由于表面干沙层的存在抑制了土壤水分的蒸发(
段利民, 2011 ), 其土壤水分含量反而高于固定沙丘, 但其土壤养分和种子库非常贫瘠, 因此二者可能共同限制了群落的恢复. ...
我国北方沙区退化植被的恢复机理
3
2006
... 大多数陆地生态系统天然植物群落都是特定气候条件下的产物(
赵哈林等, 2003 ), 气候条件的变化, 特别是气温和降水的变化会引起植物群落特征的变化(
Liu & Lei, 2015 ;
Liu et al ., 2016 ).
赵哈林等(2006) 认为气候是我国北方植被恢复的外在驱动力, 决定着演替的方向和终点.然而我们13年的观测结果显示群落生物量和物种丰富度与降水和气温之间不存在显著关系, 但气温和降水与生物量的线性回归斜率随着沙地的恢复而逐渐增大, 斜率的大小反映出植物群落对降水年际变化的响应敏感性(
Sala et al ., 2012 ), 在一定程度上也反映出群落对降水的利用效率(
Bai et al ., 2008 ).随着沙地生境恢复, 群落对年际降水波动的响应敏感性逐渐增加, 降水利用效率也相应地提升, 这种变化可能与资源的可利用性密切相关.许多研究发现土壤氮等资源的可利用性越高, 植被对降水变化的响应越敏感(
Wilcox et al ., 2016 ,
2017 ;
Knapp et al ., 2017 ), 在科尔沁沙地大尺度的研究也发现土壤养分梯度是影响植物群落分布最主要的因素(
左小安等, 2007 ).本研究中沙质草地土壤环境条件最优, 固定沙丘次之, 流动沙丘最差, 因此这种资源分布的差异导致不同沙地群落对降水变化的响应不同.群落组成也会影响群落对降水变化的响应.通过对生活型功能群重要值的年变化分析发现沙质草地群落不同功能群之间存在补偿效应, 这种补偿效应的存在可以有效地减缓降水等气候因子变化带来的影响.
Bai等(2004) 在内蒙古草原的研究也发现群落功能群之间存在补偿效应, 这种效应的存在更有利于群落稳定性的维持.这些因素的综合作用可能导致了群落生物量年变化对降水波动响应微弱.然而需要注意的是许多关于降水与群落生产力之间关系的结论都是在长时间尺度或者大空间尺度统计得来的(
Knapp et al ., 2017 ;
Wilcox et al ., 2017 ), 而本研究仅是在时间跨度为13年的小区域中开展的, 因此还需要更长时间的观测资料来研究群落对降水变化的响应.年内生物量积累与积温和累积降水量显著相关, 甚至在沙质草地二者对生物量年内积累的解释率达到68%, 因此积温和累积降水量是年内生物量积累的重要驱动因素, 这与植被生长过程中的水热需求规律一致. ...
... 气候是植被恢复的外在驱动力, 决定群落演替的方向和终点, 而植被的自我修复是内在驱动力, 决定着植物恢复的起始速度和演替速度(
赵哈林等, 2006 ).植被自我修复的基础是土壤种子库,
Plassmann等(2009) 研究发现相比于当前群落, 土壤种子库更能反映演替初期的植被组成, 但在我们的研究中发现种子丰富度变化与群落物种丰富度变化相似, DCA分析也发现地上植物群落物种和土壤种子库之间有很高的相似性, 这反映出科尔沁沙地植被恢复和土壤种子库变化存在较强的关联性.
Wang等(2019) 在科尔沁沙地的研究也表明植被恢复过程中地上群落和土壤种子库之间存在紧密联系, 并且随生境恢复二者之间的联系更加紧密. ...
... 退化沙地土壤环境的恢复速率直接影响植被的演替进程, 土壤环境的改善是植被对土壤作用的结果, 但又反馈于植被的演替(
赵哈林等, 2006 ).我们的监测结果显示3种封育沙地在10年内仅有土壤有效氮和有效磷含量增加, 并且增加幅度不大, 土壤养分缓慢的积累速率对植被恢复有很大的限制作用, 尤其是在养分含量极低的流动沙丘这种限制作用可能会更大, 因此封育13年的流动沙丘群落生物量没有显著增加.另外, 土壤养分的空间异质性直接影响植被分布模式(
Zuo et al ., 2014 ).本研究3种沙地土壤环境存在显著差异, 群落物种与土壤因子之间的CCA分析也显示不同沙地群落物种受不同因素的影响, 其中沙质草地植物群落物种与土壤养分之间相关性很强, 固定沙丘群落物种和土壤水分联系紧密, 而流动沙丘群落物种与各土壤因子并不相关.3类沙地中沙质草地土壤养分和水分条件最优, 其相应的群落生物量和物种密度也最高, 但沙质草地多为草本植物, 密布的根系层对降水存在滞留作用(
牛存洋等, 2015 ), 另外草本植物相比于灌木和半灌木耗水相对较少, 并且较高的植被盖度产生的地表遮阴效果也能有效减弱土壤蒸发(
常昌明等, 2016 ), 这些因素的综合作用使其土壤水分含量最高, 进而使水分对群落的限制作用可能弱于土壤养分.而在固定沙丘土壤水分含量最低, 这是因为在固定沙丘优势物种主要为根系较深、耗水强烈的差不嘎蒿、小叶锦鸡儿等灌木, 使得土壤水分消耗强烈, 导致群落主要受土壤水分的限制.流动沙丘由于表面干沙层的存在抑制了土壤水分的蒸发(
段利民, 2011 ), 其土壤水分含量反而高于固定沙丘, 但其土壤养分和种子库非常贫瘠, 因此二者可能共同限制了群落的恢复. ...
3
2003
... 退化沙地恢复的速度和演替方向一方面取决于退化程度和干扰的消除程度, 另一方面取决于土壤、气候等环境条件(
赵哈林等, 2003 ).本研究中流动沙丘13年封育期间植被盖度和物种丰富度显著增加, 但群落生物量没有显著变化, 整体上群落在逐渐恢复, 但速度缓慢, 群落生产力有限.许多研究发现干扰退化草地在短期封育下可以快速恢复(
赵哈林等, 2009 ;
苗仁辉等, 2013 ;
吕朋等, 2018 ), 其根本原因是现存群落具有拓殖能力和退化群落资源过剩(
王炜等, 1996 ), 但对于流动沙丘, 群落资源处于极其匮乏的状态, 现存群落拓殖能力有限, 土壤种子库基础也相对薄弱, 因此植被恢复受到很大限制.另外植被恢复也受时间积累效应的影响, 物种要进入某一环境空间并建立种群, 需要有足够的时间去完成其繁殖体从种源地到目标空间的传播、散布和在目标空间的定居、繁殖过程, 这一过程进行的速度直接影响物种多样性恢复的进度(
张继义等, 2004 ), 本研究中流动沙丘封育仅13年, 植物建群时间有限, 因此时间积累可能相对不足, 植被恢复程度有限.有研究认为在降水条件较好的半干旱沙区, 采取封沙育草措施后10-15年的时间, 植被能够自然恢复(
张华等, 2003 ), 然而本研究的结果显示, 流动沙丘恢复到灌木、半灌木或多年生草本优势度较高的固定沙丘群落需要更长的时间, 因此封育年限的设定应综合考虑植被退化程度、气候条件以及土壤环境等因素. ...
... 固定沙丘群落恢复演替过程中一年生杂类草群落属于演替的早期阶段, 多年生植物占优势的群落处于演替的中期阶段, 随着沙丘群落演替的推进, 一年生杂类草优势度会逐渐下降, 多年生草本优势度会逐渐增加, 相应的豆科物种优势度也会逐渐上升(
赵哈林等, 2003 ).而在本研究13年的观测期间, 固定沙丘群落一年生和多年生杂类草优势度有显著增加的趋势, 而灌木和半灌木以及多年生豆科植物优势度有显著下降的趋势, 这与正常的固定沙丘群落恢复演替的方向相反, 这说明封育33年的固定沙丘群落存在逆行演替的现象.另外, 一年生杂类草重要值与灌木和半灌木以及多年生豆科植物重要值显著负相关, 但与其他功能群重要值的相关性不显著, 群落生物量和物种丰富度也存在下降趋势, 反映出一年生杂类草优势度的增长抑制了其他功能群的生长, 这也表明过度封育不利于固定沙丘植被 恢复. ...
... 大多数陆地生态系统天然植物群落都是特定气候条件下的产物(
赵哈林等, 2003 ), 气候条件的变化, 特别是气温和降水的变化会引起植物群落特征的变化(
Liu & Lei, 2015 ;
Liu et al ., 2016 ).
赵哈林等(2006) 认为气候是我国北方植被恢复的外在驱动力, 决定着演替的方向和终点.然而我们13年的观测结果显示群落生物量和物种丰富度与降水和气温之间不存在显著关系, 但气温和降水与生物量的线性回归斜率随着沙地的恢复而逐渐增大, 斜率的大小反映出植物群落对降水年际变化的响应敏感性(
Sala et al ., 2012 ), 在一定程度上也反映出群落对降水的利用效率(
Bai et al ., 2008 ).随着沙地生境恢复, 群落对年际降水波动的响应敏感性逐渐增加, 降水利用效率也相应地提升, 这种变化可能与资源的可利用性密切相关.许多研究发现土壤氮等资源的可利用性越高, 植被对降水变化的响应越敏感(
Wilcox et al ., 2016 ,
2017 ;
Knapp et al ., 2017 ), 在科尔沁沙地大尺度的研究也发现土壤养分梯度是影响植物群落分布最主要的因素(
左小安等, 2007 ).本研究中沙质草地土壤环境条件最优, 固定沙丘次之, 流动沙丘最差, 因此这种资源分布的差异导致不同沙地群落对降水变化的响应不同.群落组成也会影响群落对降水变化的响应.通过对生活型功能群重要值的年变化分析发现沙质草地群落不同功能群之间存在补偿效应, 这种补偿效应的存在可以有效地减缓降水等气候因子变化带来的影响.
Bai等(2004) 在内蒙古草原的研究也发现群落功能群之间存在补偿效应, 这种效应的存在更有利于群落稳定性的维持.这些因素的综合作用可能导致了群落生物量年变化对降水波动响应微弱.然而需要注意的是许多关于降水与群落生产力之间关系的结论都是在长时间尺度或者大空间尺度统计得来的(
Knapp et al ., 2017 ;
Wilcox et al ., 2017 ), 而本研究仅是在时间跨度为13年的小区域中开展的, 因此还需要更长时间的观测资料来研究群落对降水变化的响应.年内生物量积累与积温和累积降水量显著相关, 甚至在沙质草地二者对生物量年内积累的解释率达到68%, 因此积温和累积降水量是年内生物量积累的重要驱动因素, 这与植被生长过程中的水热需求规律一致. ...
3
2003
... 退化沙地恢复的速度和演替方向一方面取决于退化程度和干扰的消除程度, 另一方面取决于土壤、气候等环境条件(
赵哈林等, 2003 ).本研究中流动沙丘13年封育期间植被盖度和物种丰富度显著增加, 但群落生物量没有显著变化, 整体上群落在逐渐恢复, 但速度缓慢, 群落生产力有限.许多研究发现干扰退化草地在短期封育下可以快速恢复(
赵哈林等, 2009 ;
苗仁辉等, 2013 ;
吕朋等, 2018 ), 其根本原因是现存群落具有拓殖能力和退化群落资源过剩(
王炜等, 1996 ), 但对于流动沙丘, 群落资源处于极其匮乏的状态, 现存群落拓殖能力有限, 土壤种子库基础也相对薄弱, 因此植被恢复受到很大限制.另外植被恢复也受时间积累效应的影响, 物种要进入某一环境空间并建立种群, 需要有足够的时间去完成其繁殖体从种源地到目标空间的传播、散布和在目标空间的定居、繁殖过程, 这一过程进行的速度直接影响物种多样性恢复的进度(
张继义等, 2004 ), 本研究中流动沙丘封育仅13年, 植物建群时间有限, 因此时间积累可能相对不足, 植被恢复程度有限.有研究认为在降水条件较好的半干旱沙区, 采取封沙育草措施后10-15年的时间, 植被能够自然恢复(
张华等, 2003 ), 然而本研究的结果显示, 流动沙丘恢复到灌木、半灌木或多年生草本优势度较高的固定沙丘群落需要更长的时间, 因此封育年限的设定应综合考虑植被退化程度、气候条件以及土壤环境等因素. ...
... 固定沙丘群落恢复演替过程中一年生杂类草群落属于演替的早期阶段, 多年生植物占优势的群落处于演替的中期阶段, 随着沙丘群落演替的推进, 一年生杂类草优势度会逐渐下降, 多年生草本优势度会逐渐增加, 相应的豆科物种优势度也会逐渐上升(
赵哈林等, 2003 ).而在本研究13年的观测期间, 固定沙丘群落一年生和多年生杂类草优势度有显著增加的趋势, 而灌木和半灌木以及多年生豆科植物优势度有显著下降的趋势, 这与正常的固定沙丘群落恢复演替的方向相反, 这说明封育33年的固定沙丘群落存在逆行演替的现象.另外, 一年生杂类草重要值与灌木和半灌木以及多年生豆科植物重要值显著负相关, 但与其他功能群重要值的相关性不显著, 群落生物量和物种丰富度也存在下降趋势, 反映出一年生杂类草优势度的增长抑制了其他功能群的生长, 这也表明过度封育不利于固定沙丘植被 恢复. ...
... 大多数陆地生态系统天然植物群落都是特定气候条件下的产物(
赵哈林等, 2003 ), 气候条件的变化, 特别是气温和降水的变化会引起植物群落特征的变化(
Liu & Lei, 2015 ;
Liu et al ., 2016 ).
赵哈林等(2006) 认为气候是我国北方植被恢复的外在驱动力, 决定着演替的方向和终点.然而我们13年的观测结果显示群落生物量和物种丰富度与降水和气温之间不存在显著关系, 但气温和降水与生物量的线性回归斜率随着沙地的恢复而逐渐增大, 斜率的大小反映出植物群落对降水年际变化的响应敏感性(
Sala et al ., 2012 ), 在一定程度上也反映出群落对降水的利用效率(
Bai et al ., 2008 ).随着沙地生境恢复, 群落对年际降水波动的响应敏感性逐渐增加, 降水利用效率也相应地提升, 这种变化可能与资源的可利用性密切相关.许多研究发现土壤氮等资源的可利用性越高, 植被对降水变化的响应越敏感(
Wilcox et al ., 2016 ,
2017 ;
Knapp et al ., 2017 ), 在科尔沁沙地大尺度的研究也发现土壤养分梯度是影响植物群落分布最主要的因素(
左小安等, 2007 ).本研究中沙质草地土壤环境条件最优, 固定沙丘次之, 流动沙丘最差, 因此这种资源分布的差异导致不同沙地群落对降水变化的响应不同.群落组成也会影响群落对降水变化的响应.通过对生活型功能群重要值的年变化分析发现沙质草地群落不同功能群之间存在补偿效应, 这种补偿效应的存在可以有效地减缓降水等气候因子变化带来的影响.
Bai等(2004) 在内蒙古草原的研究也发现群落功能群之间存在补偿效应, 这种效应的存在更有利于群落稳定性的维持.这些因素的综合作用可能导致了群落生物量年变化对降水波动响应微弱.然而需要注意的是许多关于降水与群落生产力之间关系的结论都是在长时间尺度或者大空间尺度统计得来的(
Knapp et al ., 2017 ;
Wilcox et al ., 2017 ), 而本研究仅是在时间跨度为13年的小区域中开展的, 因此还需要更长时间的观测资料来研究群落对降水变化的响应.年内生物量积累与积温和累积降水量显著相关, 甚至在沙质草地二者对生物量年内积累的解释率达到68%, 因此积温和累积降水量是年内生物量积累的重要驱动因素, 这与植被生长过程中的水热需求规律一致. ...
科尔沁沙质草地放牧和围封条件下的土壤种子库
1
2006
... 科尔沁沙地作为中国北方最严重的一个沙漠化区域(
刘新民等, 1999 ), 封育禁牧被广泛用来恢复退化沙地植被, 同时也有许多研究对封育所产生的生态效应做了评述.如退化沙质草地封育后群落生物量响应最为敏感, 在较短时间内群落生物量迅速恢复, 同时凋落物量也显著增加(
苗仁辉等, 2013 ;
赵丽娅等, 2017 ;
吕朋等, 2018 ).随着封育时间的延长, 群落物种组成开始发生变化, 流动沙丘封育后沙蓬(
Agriophyllum squarrosum )作为先锋物种首先建立种群, 使沙丘流沙面积减小, 之后以差不嘎蒿(
Artemisia halodendron )为主的多年生灌木通过无性繁殖的方式开始在沙丘定植, 差不嘎蒿等灌木的出现逐渐营造出养分和水分相对富集的灌丛微生境, 进而为禾本科植物的种子萌发提供一个良好的环境(
Su et al ., 2005 ).随着植物群落的变化, 土壤种子库也随之改变(
赵丽娅等, 2006 ;
赵盼盼等, 2018 ), 并且随着流动沙丘的固定和生境的改善, 植物群落和土壤种子库之间的联系变得更加紧密(
Wang et al ., 2019 ).封育对土壤环境也有很大的改善, 对封育和放牧样地土壤性质进行比较, 发现封育样地土壤容重、紧实度及粗颗粒含量显著小于自由放牧样地(
单贵莲等, 2009 ), 而封育样地土壤有机质、全氮、有效氮和有效钾含量显著高于放牧样地(
张建鹏等, 2017 ).封育年限长短也是影响植物群落恢复的重要因素, 适当封育有利于植被恢复, 但过度封育反而会引起植被退化,
高凯等(2013) 在内蒙古草原的研究发现, 封育年限超过14年会使多年生禾草以及灌木和半灌木优势度显著降低, 而多年生杂类草优势度出现不同程度的升高. ...
科尔沁沙质草地放牧和围封条件下的土壤种子库
1
2006
... 科尔沁沙地作为中国北方最严重的一个沙漠化区域(
刘新民等, 1999 ), 封育禁牧被广泛用来恢复退化沙地植被, 同时也有许多研究对封育所产生的生态效应做了评述.如退化沙质草地封育后群落生物量响应最为敏感, 在较短时间内群落生物量迅速恢复, 同时凋落物量也显著增加(
苗仁辉等, 2013 ;
赵丽娅等, 2017 ;
吕朋等, 2018 ).随着封育时间的延长, 群落物种组成开始发生变化, 流动沙丘封育后沙蓬(
Agriophyllum squarrosum )作为先锋物种首先建立种群, 使沙丘流沙面积减小, 之后以差不嘎蒿(
Artemisia halodendron )为主的多年生灌木通过无性繁殖的方式开始在沙丘定植, 差不嘎蒿等灌木的出现逐渐营造出养分和水分相对富集的灌丛微生境, 进而为禾本科植物的种子萌发提供一个良好的环境(
Su et al ., 2005 ).随着植物群落的变化, 土壤种子库也随之改变(
赵丽娅等, 2006 ;
赵盼盼等, 2018 ), 并且随着流动沙丘的固定和生境的改善, 植物群落和土壤种子库之间的联系变得更加紧密(
Wang et al ., 2019 ).封育对土壤环境也有很大的改善, 对封育和放牧样地土壤性质进行比较, 发现封育样地土壤容重、紧实度及粗颗粒含量显著小于自由放牧样地(
单贵莲等, 2009 ), 而封育样地土壤有机质、全氮、有效氮和有效钾含量显著高于放牧样地(
张建鹏等, 2017 ).封育年限长短也是影响植物群落恢复的重要因素, 适当封育有利于植被恢复, 但过度封育反而会引起植被退化,
高凯等(2013) 在内蒙古草原的研究发现, 封育年限超过14年会使多年生禾草以及灌木和半灌木优势度显著降低, 而多年生杂类草优势度出现不同程度的升高. ...
围封和放牧对科尔沁沙质草地植被和土壤的影响
1
2017
... 科尔沁沙地作为中国北方最严重的一个沙漠化区域(
刘新民等, 1999 ), 封育禁牧被广泛用来恢复退化沙地植被, 同时也有许多研究对封育所产生的生态效应做了评述.如退化沙质草地封育后群落生物量响应最为敏感, 在较短时间内群落生物量迅速恢复, 同时凋落物量也显著增加(
苗仁辉等, 2013 ;
赵丽娅等, 2017 ;
吕朋等, 2018 ).随着封育时间的延长, 群落物种组成开始发生变化, 流动沙丘封育后沙蓬(
Agriophyllum squarrosum )作为先锋物种首先建立种群, 使沙丘流沙面积减小, 之后以差不嘎蒿(
Artemisia halodendron )为主的多年生灌木通过无性繁殖的方式开始在沙丘定植, 差不嘎蒿等灌木的出现逐渐营造出养分和水分相对富集的灌丛微生境, 进而为禾本科植物的种子萌发提供一个良好的环境(
Su et al ., 2005 ).随着植物群落的变化, 土壤种子库也随之改变(
赵丽娅等, 2006 ;
赵盼盼等, 2018 ), 并且随着流动沙丘的固定和生境的改善, 植物群落和土壤种子库之间的联系变得更加紧密(
Wang et al ., 2019 ).封育对土壤环境也有很大的改善, 对封育和放牧样地土壤性质进行比较, 发现封育样地土壤容重、紧实度及粗颗粒含量显著小于自由放牧样地(
单贵莲等, 2009 ), 而封育样地土壤有机质、全氮、有效氮和有效钾含量显著高于放牧样地(
张建鹏等, 2017 ).封育年限长短也是影响植物群落恢复的重要因素, 适当封育有利于植被恢复, 但过度封育反而会引起植被退化,
高凯等(2013) 在内蒙古草原的研究发现, 封育年限超过14年会使多年生禾草以及灌木和半灌木优势度显著降低, 而多年生杂类草优势度出现不同程度的升高. ...
围封和放牧对科尔沁沙质草地植被和土壤的影响
1
2017
... 科尔沁沙地作为中国北方最严重的一个沙漠化区域(
刘新民等, 1999 ), 封育禁牧被广泛用来恢复退化沙地植被, 同时也有许多研究对封育所产生的生态效应做了评述.如退化沙质草地封育后群落生物量响应最为敏感, 在较短时间内群落生物量迅速恢复, 同时凋落物量也显著增加(
苗仁辉等, 2013 ;
赵丽娅等, 2017 ;
吕朋等, 2018 ).随着封育时间的延长, 群落物种组成开始发生变化, 流动沙丘封育后沙蓬(
Agriophyllum squarrosum )作为先锋物种首先建立种群, 使沙丘流沙面积减小, 之后以差不嘎蒿(
Artemisia halodendron )为主的多年生灌木通过无性繁殖的方式开始在沙丘定植, 差不嘎蒿等灌木的出现逐渐营造出养分和水分相对富集的灌丛微生境, 进而为禾本科植物的种子萌发提供一个良好的环境(
Su et al ., 2005 ).随着植物群落的变化, 土壤种子库也随之改变(
赵丽娅等, 2006 ;
赵盼盼等, 2018 ), 并且随着流动沙丘的固定和生境的改善, 植物群落和土壤种子库之间的联系变得更加紧密(
Wang et al ., 2019 ).封育对土壤环境也有很大的改善, 对封育和放牧样地土壤性质进行比较, 发现封育样地土壤容重、紧实度及粗颗粒含量显著小于自由放牧样地(
单贵莲等, 2009 ), 而封育样地土壤有机质、全氮、有效氮和有效钾含量显著高于放牧样地(
张建鹏等, 2017 ).封育年限长短也是影响植物群落恢复的重要因素, 适当封育有利于植被恢复, 但过度封育反而会引起植被退化,
高凯等(2013) 在内蒙古草原的研究发现, 封育年限超过14年会使多年生禾草以及灌木和半灌木优势度显著降低, 而多年生杂类草优势度出现不同程度的升高. ...
围封对荒漠草原区沙芦草群落土壤种子库及地上植被的影响
1
2018
... 科尔沁沙地作为中国北方最严重的一个沙漠化区域(
刘新民等, 1999 ), 封育禁牧被广泛用来恢复退化沙地植被, 同时也有许多研究对封育所产生的生态效应做了评述.如退化沙质草地封育后群落生物量响应最为敏感, 在较短时间内群落生物量迅速恢复, 同时凋落物量也显著增加(
苗仁辉等, 2013 ;
赵丽娅等, 2017 ;
吕朋等, 2018 ).随着封育时间的延长, 群落物种组成开始发生变化, 流动沙丘封育后沙蓬(
Agriophyllum squarrosum )作为先锋物种首先建立种群, 使沙丘流沙面积减小, 之后以差不嘎蒿(
Artemisia halodendron )为主的多年生灌木通过无性繁殖的方式开始在沙丘定植, 差不嘎蒿等灌木的出现逐渐营造出养分和水分相对富集的灌丛微生境, 进而为禾本科植物的种子萌发提供一个良好的环境(
Su et al ., 2005 ).随着植物群落的变化, 土壤种子库也随之改变(
赵丽娅等, 2006 ;
赵盼盼等, 2018 ), 并且随着流动沙丘的固定和生境的改善, 植物群落和土壤种子库之间的联系变得更加紧密(
Wang et al ., 2019 ).封育对土壤环境也有很大的改善, 对封育和放牧样地土壤性质进行比较, 发现封育样地土壤容重、紧实度及粗颗粒含量显著小于自由放牧样地(
单贵莲等, 2009 ), 而封育样地土壤有机质、全氮、有效氮和有效钾含量显著高于放牧样地(
张建鹏等, 2017 ).封育年限长短也是影响植物群落恢复的重要因素, 适当封育有利于植被恢复, 但过度封育反而会引起植被退化,
高凯等(2013) 在内蒙古草原的研究发现, 封育年限超过14年会使多年生禾草以及灌木和半灌木优势度显著降低, 而多年生杂类草优势度出现不同程度的升高. ...
围封对荒漠草原区沙芦草群落土壤种子库及地上植被的影响
1
2018
... 科尔沁沙地作为中国北方最严重的一个沙漠化区域(
刘新民等, 1999 ), 封育禁牧被广泛用来恢复退化沙地植被, 同时也有许多研究对封育所产生的生态效应做了评述.如退化沙质草地封育后群落生物量响应最为敏感, 在较短时间内群落生物量迅速恢复, 同时凋落物量也显著增加(
苗仁辉等, 2013 ;
赵丽娅等, 2017 ;
吕朋等, 2018 ).随着封育时间的延长, 群落物种组成开始发生变化, 流动沙丘封育后沙蓬(
Agriophyllum squarrosum )作为先锋物种首先建立种群, 使沙丘流沙面积减小, 之后以差不嘎蒿(
Artemisia halodendron )为主的多年生灌木通过无性繁殖的方式开始在沙丘定植, 差不嘎蒿等灌木的出现逐渐营造出养分和水分相对富集的灌丛微生境, 进而为禾本科植物的种子萌发提供一个良好的环境(
Su et al ., 2005 ).随着植物群落的变化, 土壤种子库也随之改变(
赵丽娅等, 2006 ;
赵盼盼等, 2018 ), 并且随着流动沙丘的固定和生境的改善, 植物群落和土壤种子库之间的联系变得更加紧密(
Wang et al ., 2019 ).封育对土壤环境也有很大的改善, 对封育和放牧样地土壤性质进行比较, 发现封育样地土壤容重、紧实度及粗颗粒含量显著小于自由放牧样地(
单贵莲等, 2009 ), 而封育样地土壤有机质、全氮、有效氮和有效钾含量显著高于放牧样地(
张建鹏等, 2017 ).封育年限长短也是影响植物群落恢复的重要因素, 适当封育有利于植被恢复, 但过度封育反而会引起植被退化,
高凯等(2013) 在内蒙古草原的研究发现, 封育年限超过14年会使多年生禾草以及灌木和半灌木优势度显著降低, 而多年生杂类草优势度出现不同程度的升高. ...
Scale dependence of plant species richness and vegetation-environment relationship along a gradient of dune stabilization in Horqin Sandy Land, Northern China
1
2014
... 退化沙地土壤环境的恢复速率直接影响植被的演替进程, 土壤环境的改善是植被对土壤作用的结果, 但又反馈于植被的演替(
赵哈林等, 2006 ).我们的监测结果显示3种封育沙地在10年内仅有土壤有效氮和有效磷含量增加, 并且增加幅度不大, 土壤养分缓慢的积累速率对植被恢复有很大的限制作用, 尤其是在养分含量极低的流动沙丘这种限制作用可能会更大, 因此封育13年的流动沙丘群落生物量没有显著增加.另外, 土壤养分的空间异质性直接影响植被分布模式(
Zuo et al ., 2014 ).本研究3种沙地土壤环境存在显著差异, 群落物种与土壤因子之间的CCA分析也显示不同沙地群落物种受不同因素的影响, 其中沙质草地植物群落物种与土壤养分之间相关性很强, 固定沙丘群落物种和土壤水分联系紧密, 而流动沙丘群落物种与各土壤因子并不相关.3类沙地中沙质草地土壤养分和水分条件最优, 其相应的群落生物量和物种密度也最高, 但沙质草地多为草本植物, 密布的根系层对降水存在滞留作用(
牛存洋等, 2015 ), 另外草本植物相比于灌木和半灌木耗水相对较少, 并且较高的植被盖度产生的地表遮阴效果也能有效减弱土壤蒸发(
常昌明等, 2016 ), 这些因素的综合作用使其土壤水分含量最高, 进而使水分对群落的限制作用可能弱于土壤养分.而在固定沙丘土壤水分含量最低, 这是因为在固定沙丘优势物种主要为根系较深、耗水强烈的差不嘎蒿、小叶锦鸡儿等灌木, 使得土壤水分消耗强烈, 导致群落主要受土壤水分的限制.流动沙丘由于表面干沙层的存在抑制了土壤水分的蒸发(
段利民, 2011 ), 其土壤水分含量反而高于固定沙丘, 但其土壤养分和种子库非常贫瘠, 因此二者可能共同限制了群落的恢复. ...
Contrasting effects of plant inter- and intraspecific variation on community trait responses to restoration of a sandy grassland ecosystem
1
2017
... 研究区位于内蒙古自治区科尔沁沙地(42.92° N, 120.70° E), 属于半干旱陆地季风气候, 夏季高温, 冬季寒冷.年降水量360 mm, 75%的降水发生在6-9月.年平均气温约为6.4 ℃, 月平均气温在23.5 ℃ (7月)到-16.8 ℃ (1月)之间(
Zuo et al ., 2017 ).土壤类型主要有风沙土、草甸土、沙质栗钙土和沼泽土.地貌特征为固定沙丘、半固定沙丘、流动沙丘和面积不等的平缓沙质草地以及低洼地交错分布.根据奈曼沙漠化研究站2005-2018年间未发表的观测数据, 该区域地下水平均埋深12-15 m.植被斑块化分布和空间异质性较强, 植被盖度较小的流动沙丘上一年生先锋物种沙蓬为优势种, 半固定和固定沙丘上差不嘎蒿和小叶锦鸡儿(
Caragana microphylla )等灌木为优势种.盖度较高的沙质草地以菊科一二年生物种黄蒿(
Artemisia scoparia )以及多年的芦苇(
Phragmites australis )和白草(
Pennisetum flaccidum )为优势物种(
Zuo et al ., 2012 ). ...
科尔沁沙质草地群落物种多样性、生产力与土壤特性的关系
1
2007
... 大多数陆地生态系统天然植物群落都是特定气候条件下的产物(
赵哈林等, 2003 ), 气候条件的变化, 特别是气温和降水的变化会引起植物群落特征的变化(
Liu & Lei, 2015 ;
Liu et al ., 2016 ).
赵哈林等(2006) 认为气候是我国北方植被恢复的外在驱动力, 决定着演替的方向和终点.然而我们13年的观测结果显示群落生物量和物种丰富度与降水和气温之间不存在显著关系, 但气温和降水与生物量的线性回归斜率随着沙地的恢复而逐渐增大, 斜率的大小反映出植物群落对降水年际变化的响应敏感性(
Sala et al ., 2012 ), 在一定程度上也反映出群落对降水的利用效率(
Bai et al ., 2008 ).随着沙地生境恢复, 群落对年际降水波动的响应敏感性逐渐增加, 降水利用效率也相应地提升, 这种变化可能与资源的可利用性密切相关.许多研究发现土壤氮等资源的可利用性越高, 植被对降水变化的响应越敏感(
Wilcox et al ., 2016 ,
2017 ;
Knapp et al ., 2017 ), 在科尔沁沙地大尺度的研究也发现土壤养分梯度是影响植物群落分布最主要的因素(
左小安等, 2007 ).本研究中沙质草地土壤环境条件最优, 固定沙丘次之, 流动沙丘最差, 因此这种资源分布的差异导致不同沙地群落对降水变化的响应不同.群落组成也会影响群落对降水变化的响应.通过对生活型功能群重要值的年变化分析发现沙质草地群落不同功能群之间存在补偿效应, 这种补偿效应的存在可以有效地减缓降水等气候因子变化带来的影响.
Bai等(2004) 在内蒙古草原的研究也发现群落功能群之间存在补偿效应, 这种效应的存在更有利于群落稳定性的维持.这些因素的综合作用可能导致了群落生物量年变化对降水波动响应微弱.然而需要注意的是许多关于降水与群落生产力之间关系的结论都是在长时间尺度或者大空间尺度统计得来的(
Knapp et al ., 2017 ;
Wilcox et al ., 2017 ), 而本研究仅是在时间跨度为13年的小区域中开展的, 因此还需要更长时间的观测资料来研究群落对降水变化的响应.年内生物量积累与积温和累积降水量显著相关, 甚至在沙质草地二者对生物量年内积累的解释率达到68%, 因此积温和累积降水量是年内生物量积累的重要驱动因素, 这与植被生长过程中的水热需求规律一致. ...
科尔沁沙质草地群落物种多样性、生产力与土壤特性的关系
1
2007
... 大多数陆地生态系统天然植物群落都是特定气候条件下的产物(
赵哈林等, 2003 ), 气候条件的变化, 特别是气温和降水的变化会引起植物群落特征的变化(
Liu & Lei, 2015 ;
Liu et al ., 2016 ).
赵哈林等(2006) 认为气候是我国北方植被恢复的外在驱动力, 决定着演替的方向和终点.然而我们13年的观测结果显示群落生物量和物种丰富度与降水和气温之间不存在显著关系, 但气温和降水与生物量的线性回归斜率随着沙地的恢复而逐渐增大, 斜率的大小反映出植物群落对降水年际变化的响应敏感性(
Sala et al ., 2012 ), 在一定程度上也反映出群落对降水的利用效率(
Bai et al ., 2008 ).随着沙地生境恢复, 群落对年际降水波动的响应敏感性逐渐增加, 降水利用效率也相应地提升, 这种变化可能与资源的可利用性密切相关.许多研究发现土壤氮等资源的可利用性越高, 植被对降水变化的响应越敏感(
Wilcox et al ., 2016 ,
2017 ;
Knapp et al ., 2017 ), 在科尔沁沙地大尺度的研究也发现土壤养分梯度是影响植物群落分布最主要的因素(
左小安等, 2007 ).本研究中沙质草地土壤环境条件最优, 固定沙丘次之, 流动沙丘最差, 因此这种资源分布的差异导致不同沙地群落对降水变化的响应不同.群落组成也会影响群落对降水变化的响应.通过对生活型功能群重要值的年变化分析发现沙质草地群落不同功能群之间存在补偿效应, 这种补偿效应的存在可以有效地减缓降水等气候因子变化带来的影响.
Bai等(2004) 在内蒙古草原的研究也发现群落功能群之间存在补偿效应, 这种效应的存在更有利于群落稳定性的维持.这些因素的综合作用可能导致了群落生物量年变化对降水波动响应微弱.然而需要注意的是许多关于降水与群落生产力之间关系的结论都是在长时间尺度或者大空间尺度统计得来的(
Knapp et al ., 2017 ;
Wilcox et al ., 2017 ), 而本研究仅是在时间跨度为13年的小区域中开展的, 因此还需要更长时间的观测资料来研究群落对降水变化的响应.年内生物量积累与积温和累积降水量显著相关, 甚至在沙质草地二者对生物量年内积累的解释率达到68%, 因此积温和累积降水量是年内生物量积累的重要驱动因素, 这与植被生长过程中的水热需求规律一致. ...
Scale dependent effects of environmental factors on vegetation pattern and composition in Horqin Sandy Land, Northern China
1
2012
... 研究区位于内蒙古自治区科尔沁沙地(42.92° N, 120.70° E), 属于半干旱陆地季风气候, 夏季高温, 冬季寒冷.年降水量360 mm, 75%的降水发生在6-9月.年平均气温约为6.4 ℃, 月平均气温在23.5 ℃ (7月)到-16.8 ℃ (1月)之间(
Zuo et al ., 2017 ).土壤类型主要有风沙土、草甸土、沙质栗钙土和沼泽土.地貌特征为固定沙丘、半固定沙丘、流动沙丘和面积不等的平缓沙质草地以及低洼地交错分布.根据奈曼沙漠化研究站2005-2018年间未发表的观测数据, 该区域地下水平均埋深12-15 m.植被斑块化分布和空间异质性较强, 植被盖度较小的流动沙丘上一年生先锋物种沙蓬为优势种, 半固定和固定沙丘上差不嘎蒿和小叶锦鸡儿(
Caragana microphylla )等灌木为优势种.盖度较高的沙质草地以菊科一二年生物种黄蒿(
Artemisia scoparia )以及多年的芦苇(
Phragmites australis )和白草(
Pennisetum flaccidum )为优势物种(
Zuo et al ., 2012 ). ...
 ,1,3,*, 何玉惠4, 张铜会1,3, 魏静5, 车力木格1,2, 孙姗姗1,2
,1,3,*, 何玉惠4, 张铜会1,3, 魏静5, 车力木格1,2, 孙姗姗1,2
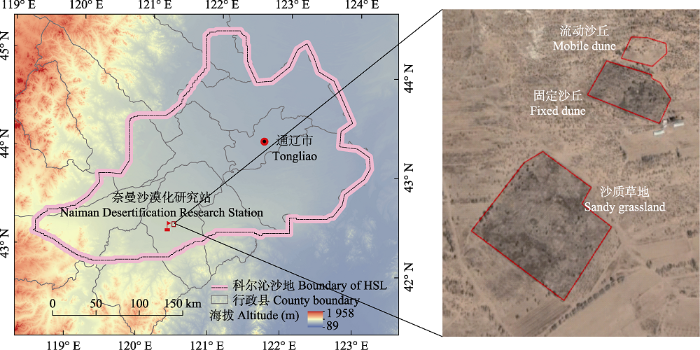 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT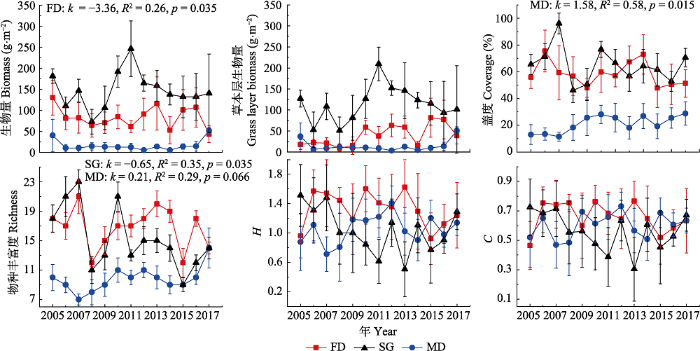 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT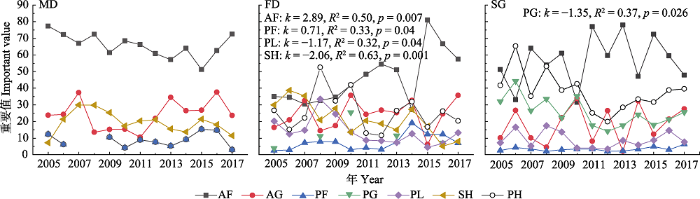 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT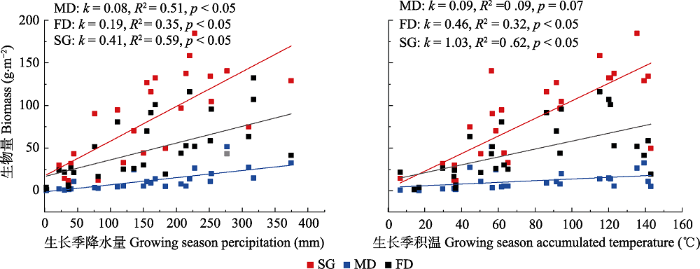 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT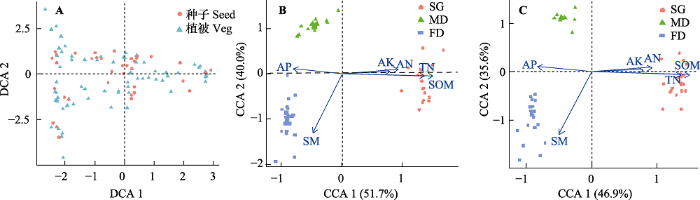 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}