Seasonal dynamics of soil microbial biomass carbon, nitrogen and phosphorus stoichiometry across global forest ecosystems
Pin LI,,1,*, TUERHANBAI Muledeer2, Di TIAN2, Zhao-Zhong FENG11 State Key Laboratory of Urban and Regional Ecology, Research Centre for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing 100085, China 2 College of Life Sciences, Capital Normal University, Beijing 100048, China
Supported by the National Natural Science Foundation of China(31870458) the Key Research Program of Frontier Sciences, Chinese Academy of Sciences(QYZDB-SSW-DQC019)
Abstract Aims Soil microorganisms in forest ecosystems play vital roles in regulating above- and belowground ecosystem processes and functions such as soil nutrient cycling, litter decomposition, net ecosystem productivity, and ecosystem succession. We aim to investigate broad-scale seasonal patterns of soil microbial biomass carbon (C), nitrogen (N) and phosphorus (P) stoichiometry. Methods By synthesizing 164 samples of soil microbial biomass C, N and P content derived from the published literature, we investigated global seasonal patterns of soil microbial C, N, P content and their ratios across three vegetation types of global forests. Important findings Soil microbial biomass C, N and P content in temperate and subtropical forests were lower in summer and higher in winter. Soil microbial biomass C, N and P content in tropical forests were lower than those in temperate and subtropical forests in four seasons. Soil microbial biomass C and N content in tropical forests were relatively the lowest in autumn, and soil microbial biomass P content was relatively constant in all seasons. The soil microbial biomass C:N of temperate forest was significantly higher than that of other two forest types in spring, and that of tropical forest was significantly higher than that of other two forest types in autumn. Soil microbial biomass N:P and C:P in temperate forests remained relatively constant in four seasons, while those in tropical forests were higher than those in other three seasons in summer. The soil microbial biomass C content, N content, N:P and C:P of broad-leaved trees were significantly higher than those of conifers in four seasons, while the soil microbial biomass P content of conifers was significantly higher than that of broad-leaved trees in four seasons. There was no significant difference in soil microbial biomass C:N between broad-leaved and coniferous trees in both spring and winter, but the soil microbial biomass C:N of coniferous trees was significantly higher than that of broad-leaved trees in summer and autumn. For the change of soil microbial biomass, season is not but forest type is the main significant factor, suggesting that the seasonal fluctuation of soil microbial biomass changes with the inherent periodic change of trees. Asynchronous nutrient uptake by plants and soil microorganisms is a trade-off mechanism between nutrient retention and ecological function maintenance. Keywords:microbial biomass carbon;microbial biomass nitrogen;microbial biomass phosphorus;forest soil;seasonal dynamics;stoichiometric ratio
PDF (1360KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 李品, 木勒德尔?吐尔汗拜, 田地, 冯兆忠. 全球森林土壤微生物生物量碳氮磷化学计量的季节动态. 植物生态学报, 2019, 43(6): 532-542. DOI: 10.17521/cjpe.2019.0075 LI Pin, Muledeer TUERHANBAI, TIAN Di, FENG Zhao-Zhong. Seasonal dynamics of soil microbial biomass carbon, nitrogen and phosphorus stoichiometry across global forest ecosystems. Chinese Journal of Plant Ecology, 2019, 43(6): 532-542. DOI: 10.17521/cjpe.2019.0075
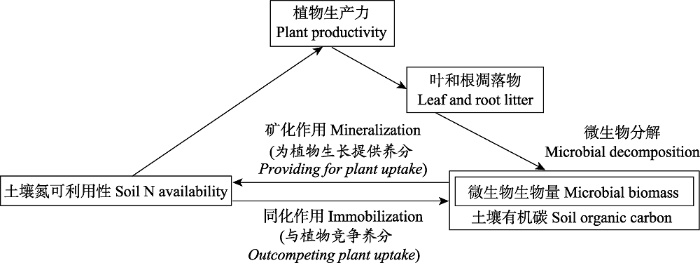
Fig. 1A conceptual model illustrating the links between plant and soil microbial activity in terrestrial ecosystems. On one hand, plants and microbes are mutualistic, since plant residues constitute the main substrate for microbial biomass that in turn produces inorganic nitrogen used by plants. On the other hand, their interaction is in competition for soil nutrients, N in particular. The competitive interaction is regulated by the relative distribution of roots and microbes in the soil, but it also depends on the pathway followed by the mineralized N. Redraw based on Zak et al. (2000).
Table 1 表1 表1土壤微生物生物量碳(C)、氮(N)、磷(P)含量和C:N、N:P、C:P在各个森林生态系统类型的统计结果 Table 1Soil microbial biomass carbon (C), nitrogen (N), phosphorus (P) content, C:N, N:P, C:P for each forest type across global forest ecosystems
森林类型 Forest type
微生物生物量 Microbial biomass (μg·g-1)
微生物生物量比值 Microbial biomass ratio
C
N
P
C:N
N:P
C:P
所有类型 Overall
629 ± 477 (80)
98 ± 81 (66)
32 ± 26 (34)
6.9a ± 2.5 (65)
5.4 ± 8.2 (33)
26.4 ± 25.0 (34)
温带针叶林 Temperate conifer forest
570ab ± 307 (14)
76b ± 49 (13)
76 ± 41 (5)
7.9a ± 2.4 (13)
1.3 ± 0.0 (4)
13.1 ± 9.9 (5)
温带阔叶林 Temperate broad-leaved forest
795ab ± 551 (12)
94b ± 57 (11)
54 (1)
7.4a ± 1.9 (10)
1.3 (1)
8.6 (1)
亚热带针叶林 Subtropical conifer forest
347b ± 125 (5)
35b ± 9 (3)
na
8.3a ± 2.6 (3)
na
na
亚热带阔叶林 Subtropical broad-leaved forest
971a ± 650 (17)
186a ± 94 (11)
28 ± 11 (7)
5.8a ± 1.7 (11)
9.1 ± 3.5 (7)
53.4 ± 22.2 (7)
热带森林 Tropical forest
454b ± 303 (32)
82b ± 78 (28)
22 ± 10 (21)
6.5a ± 2.7 (28)
5.2 ± 9.7 (21)
21.5 ± 22.8 (21)
Values are mean ± standard deviations of four seasons. Sample numbers are shown in parentheses. Different lowercase letters indicate significant differences among forest types at p < 0.05. na, not available. 数值代表平均值±标准偏差, 括号中是样本数; 不同的小写字母代表统计上有显著性差异(p < 0.05)。na, 无效。
Table 2 表2 表2各森林类型土壤微生物生物量碳(C)、氮(N)、磷(P)含量与环境因子的相关性 Table 2Correlation coefficients for the relationship between microbial biomass carbon (C), nitrogen (N), and phosphorus (P) content with environmental variables on different forest types
森林类型 Forest type
年平均气温 MAT
年降水量 MAP
pH
土壤有机碳 SOC
土壤总氮 TN
土壤总磷 TP
微生物生物量C Microbial biomass C
所有类型 Overall
-0.247***
0.037
-0.117*
0.671***
0.582***
0.461***
温带针叶林 Temperate conifer forest
-0.165
0.361**
-0.561***
0.858***
0.838***
-0.105
温带阔叶林 Temperate broad-leaved forest
-0.084
0.116
0.138
0.704***
0.696***
0.437***
亚热带针叶林 Subtropical conifer forest
0.087
0.231
-0.327*
0.303*
0.173
0.003
亚热带阔叶林 Subtropical broad-leaved forest
-0.208
0.249*
0.285*
0.614***
0.640***
0.505*
热带森林 Tropical
-0.172
0.289**
-0.350**
0.549***
0.609***
0.625***
微生物生物量N Microbial biomass N
所有类型 Overall
-0.226***
0.085
-0.047
0.535***
0.588***
0.346***
温带针叶林 Temperate conifer forest
-0.248
0.246
-0.285
0.638***
0.717***
-0.424
温带阔叶林 Temperate broad-leaved forest
0.039
0.059
0.355***
0.481***
0.597***
0.428*
亚热带针叶林 Subtropical conifer forest
0.180
0.517**
-0.498**
0.037
-0.007
-0.340
亚热带阔叶林 Subtropical broad-leaved forest
-0.419**
0.217
0.123
0.672***
0.676***
0.201
热带森林 Tropical forest
-0.262*
0.205
-0.339**
0.610***
0.691***
0.633***
微生物生物量P Microbial biomass P
所有类型 Overall
-0.555***
-0.323**
0.363***
0.215*
0.263*
0.474***
温带针叶林 Temperate conifer forest
0.891*
-0.147
0.351
0.943*
0.918*
-0.839
温带阔叶林 Temperate broad-leaved forest
0.434*
-0.434*
0.375
0.591**
0.776***
0.638**
亚热带针叶林 Subtropical conifer forest
0.154
0.287
0.442
0.806
0.867
-0.331
亚热带阔叶林 Subtropical broad-leaved forest
0.311
0.284
0.348
0.599*
0.643*
0.542
热带森林 Tropical forest
-0.082
0.024
-0.397*
0.245
0.303
0.347
***, p < 0.001; **, p < 0.01; *, p < 0.05. MAP, mean annual air precipitation; MAT, mean annual air temperature; SOC, soil organic carbon; TN, soil total nitrogen; TP, soil total phosphorus.
Fig. 2Seasonal dynamic changes in soil microbial biomass C, N, P content and their ratios in three climatic forest types including temperate forests, subtropical forests and tropical forests (mean ± SD). The asterisk designates statistical differences (p < 0.05).
Fig. 3Comparisons of soil microbial biomass and their ratios in three climatic forest types including temperate forests, subtropical forests and tropical forests (mean ± SD). Statistical differences are denoted by different lowercase letters (p < 0.05).
Fig. 4Box-Whisker plots showing the differences in soil microbial biomass between coniferous and broadleaf forests at four major seasons of spring, summer, autumn and winter (mean ± SD). Statistical differences are denoted by different lowercase letters (p < 0.05).
Table 3 表3 表3季节和森林类型双因子方差分析对土壤微生物生物量及其比值的交互影响 Table 3Two-way ANOVA analysis on the integrative effects of seasons and forest types on soil microbial biomass and their ratios
来源 Source
微生物生物量C含量 Microbial biomass C content
微生物生物量N含量 Microbial biomass N content
微生物生物量P含量 Microbial biomass P content
d.f.
MS
F
d.f.
MS
F
d.f.
MS
F
森林类型 Forest type
2
1 135 927.6
4.04*
2
90 638.4
13.41***
2
20 756.4
52.32***
季节 Season
3
172 792.3
0.62
3
3 484.8
0.52
3
1 142.8
2.88*
森林类型 × 季节 Forest type × Season
6
123 876.6
0.44
6
2 794.1
0.41
6
535.8
1.35
残差 Residual
264
281 003.8
216
6 761.1
105
396.7
来源 Source
微生物生物量C:N Microbial biomass C:N
微生物生物量N:P Microbial biomass N:P
微生物生物量C:P Microbial biomass C:P
d.f.
MS
F
d.f.
MS
F
d.f.
MS
F
森林类型 Forest type
2
93.1
7.76**
2
430.5
7.18**
2
11 677.1
6.20**
季节 Season
3
17.9
1.49
3
6.8
0.11
3
245.8
0.13
森林类型 × 季节 Forest type × Season
6
34.7
2.89*
6
20.3
0.34
6
649.4
0.35
残差 Residual
211
12.0
97
60.0
101
1 884.1
d.f., degree of freedom; MS, mean squares; F, variance ratio. ***, p < 0.001; **, p < 0.01; *, p < 0.05. Season: spring, summer, autumn and winter. Forest types: temperate, subtropical and tropical forest. d.f., 自由度; MS, 均方差; F, 方差比。 ***, p < 0.001; **, p < 0.01; *, p < 0.05。季节: 春季、夏季、秋季、冬季。森林类型: 温带森林, 亚热带森林和热带森林。
BarbhuiyaAR, ArunachalamA, PandeyHN, ArunachalamK, KhanML, NathPC (2004). Dynamics of soil microbial biomass C, N and P in disturbed and undisturbed stands of a tropical wet-evergreen forest European Journal of Soil Biology, 40, 113-121. [本文引用: 2]
BonanGB (2014). Connecting mathematical ecosystems, real-world ecosystems, and climate science New Phytologist, 202, 731-733. [本文引用: 1]
BrookesPC, OcioJA, WuJ (1990). The soil microbial biomass: Its measurement, properties and role in nitrogen carbon dynamics following substrate incorporation Soil Microorganisms, 35, 39-51. [本文引用: 3]
BrooksPD, WilliamsMW, SchmidtSK (1998). Inorganic N and microbial biomass dynamics before and during spring snowmelt Biogeochemistry, 43, 1-15. [本文引用: 1]
ClevelandCC, LiptzinD (2007). C:N:P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomass? Biogeochemistry, 85, 235-252. [本文引用: 1]
DeviNB, YadavaPS (2006). Seasonal dynamics in soil microbial biomass C, N and P in a mixed-oak forest ecosystem of Manipur, North-east India Applied Soil Ecology, 31, 220-227. [本文引用: 2]
EdwardsKA, McCullochJ, Peter KershawG, JefferiesRL (2006). Soil microbial and nutrient dynamics in a wet Arctic sedge meadow in late winter and early spring Soil Biology & Biochemistry, 38, 2843-2851. [本文引用: 2]
FangCM, SmithP, MoncrieffJB, SmithJU (2005). Similar response of labile and resistant soil organic matter pools to changes in temperature Nature, 433, 57-59. [本文引用: 1]
FauciMF, DickRP (1994). Soil microbial dynamics: Short- and long-term effects of inorganic and organic nitrogen Soil Science Society of America Journal, 58, 801-806. [本文引用: 1]
FengWT, ZouXM, SchaeferD (2009). Above- and belowground carbon inputs affect seasonal variations of soil microbial biomass in a subtropical monsoon forest of southwest China Soil Biology & Biochemistry, 41, 978-983. [本文引用: 1]
FiererN, StricklandMS, LiptzinD, BradfordMA, ClevelandCC (2009). Global patterns in belowground communities Ecology Letters, 12, 1238-1249. [本文引用: 1]
GroffmannPM, ZakDR, ChristensenS, MosierA, TiedjeJM (1993). Early spring nitrogen dynamics in a temperate forest landscape Ecology, 74, 1579-1585. [本文引用: 1]
HarteJ, KinzigAP (1993). Mutualism and competition between plants and decomposers: Implications for nutrient allocation in ecosystems The American Naturalist, 141, 829-846. [本文引用: 2]
HeZL, WuJ, O’DonnelAG, SyersJK (1997). Seasonal responses in microbial biomass carbon, phosphorus and sulphur in soils under pasture Biology and Fertility of Soils, 24, 421-428. [本文引用: 1]
KaiserC, FranklinO, DieckmannU, RichterA (2014). Microbial community dynamics alleviate stoichiometric constraints during litter decay Ecology Letters, 17, 680-690. [本文引用: 1]
KaiserC, FuchsluegerL, KorandaM, GorferM, StangeCF, KitzlerB, RascheF, StraussJ, SessitschA, Zechmeister-?BoltensternS, RichterA (2011). Plants control the seasonal dynamics of microbial N cycling in a beech forest soil by belowground C allocation Ecology, 92, 1036-1051. [本文引用: 2]
KayeJP, HartSC (1997). Competition for nitrogen between plants and soil microorganisms Trends in Ecology & Evolution, 12, 139-143. [本文引用: 2]
KorandaM, KaiserC, FuchsluegerL, KitzlerB, SessitschA, Zechmeister-BoltensternS, RichterA (2013). Seasonal variation in functional properties of microbial communities in beech forest soil Soil Biology & Biochemistry, 60, 95-104. [本文引用: 1]
LandgrafD, KloseS (2002). Mobile and readily available C and N fractions and their relationship to microbial biomass and selected enzyme activities in a sandy soil under different management systems Journal of Plant Nutrition and Soil Science, 165, 9-16. [本文引用: 1]
LiP, YangYH, HanWX, FangJY (2014). Global patterns of soil microbial nitrogen and phosphorus stoichiometry in forest ecosystems Global Ecology and Biogeography, 23, 979-987. [本文引用: 2]
LipsonDA, SchadtCW, SchmidtSK (2002). Changes in soil microbial community structure and function in an alpine dry meadow following spring snow melt Microbial Ecology, 43, 307-314. [本文引用: 1]
LipsonDA, SchmidtSK, MonsonRK (1999). Links between microbial population dynamics and nitrogen availability in an alpine ecosystem Ecology, 80, 1623-1631. [本文引用: 3]
LipsonDA, SchmidtSK, MonsonRK (2000). Carbon availability and temperature control the post-snowmelt decline in alpine soil microbial biomass Soil Biology & Biochemistry, 32, 441-448. [本文引用: 2]
LiuS, WangCK (2010). Spatio-temporal patterns of soil microbial biomass carbon and nitrogen in five temperate forest ecosystems Acta Ecologica Sinica, 30, 3135-3143. [本文引用: 2]
MullerRN, Herbert BormannF (1976). Role of Erythronium americanum Ker. in energy flow and nutrient dynamics of a northern hardwood forest ecosystem Science, 193, 1126-1128. [本文引用: 1]
NeillC, GignouxJ (2006). Soil organic matter decomposition driven by microbial growth: A simple model for a complex network of interactions Soil Biology & Biochemistry, 38, 803-811. [本文引用: 1]
NemergutDR, CostelloEK, MeyerAF, PescadorMY, WeintraubMN, SchmidtSK (2005). Structure and function of alpine and arctic soil microbial communities Research in Microbiology, 156, 775-784. [本文引用: 1]
SchimelJP, WeintraubMN (2003). The implications of exoenzyme activity on microbial carbon and nitrogen limitation in soil: A theoretical model Soil Biology & Biochemistry, 35, 549-563. [本文引用: 1]
SchmidtSK, LipsonDA (2004). Microbial growth under the snow: Implications for nutrient and allelochemical availability in temperate soils Plant and Soil, 259, 1-7. [本文引用: 1]
Scott-DentonLE, RosenstielTN, MonsonRK (2006). Differential controls by climate and substrate over the heterotrophic and rhizospheric components of soil respiration Global Change Biology, 12, 205-216. [本文引用: 2]
SinghJS, RaghubanshiAS, SinghRS, SrivastavaSC (1989). Microbial biomass acts as a source of plant nutrients in dry tropical forest and savanna Nature, 338, 499-500. [本文引用: 4]
TerrerC, ViccaS, StockerBD, HungateBA, PhillipsRP, ReichPB, FinziAC, PrenticeIC (2018). Ecosystem responses to elevated CO2 governed by plant-soil interactions and the cost of nitrogen acquisition New Phytologist, 217, 507-522. [本文引用: 3]
VanceED, BrookesPC, JenkinsonDS (1987). An extraction method for measuring soil microbial biomass C Soil Biology & Biochemistry, 19, 703-707. [本文引用: 1]
WangGB, RuanHH, TangYF, LuanYL, ChenYQ, TaoZF (2008). Seasonal fluctuation of soil microbial biomass carbon in secondary oak forest and Pinus taeda plantation in north subtropical area of China Chinese Journal of Applied Ecology, 19, 37-42. [本文引用: 2]
WangXQ, HanY, WangCK (2017). Soil microbial biomass and its seasonality in deciduous broadleaved forests with different stand ages in the Mao’ershan region, Northeast China Chinese Journal of Plant Ecology, 41, 597-609. [本文引用: 1]
YangK, ZhuJJ, ZhangJX, YanQL (2009). Seasonal dynamics of soil microbial biomass C and N in two larch plantation forests with different ages in Northeastern China Acta Ecologica Sinica, 29, 5500-5507. [本文引用: 1]
ZakDR, PregitzerKS, CurtisPS, TeeriJA, FogelR, RandlettDL (1993). Elevated atmospheric CO2 and feedback between carbon and nitrogen cycles Plant and Soil, 151, 105-117. [本文引用: 1]
ZakDR, PregitzerKS, KingJS, HolmesWE (2000). Elevated atmospheric CO2 fine roots and the response of soil microorganisms: A review and hypothesis New Phytologist, 147, 201-222. [本文引用: 4]
Dynamics of soil microbial biomass C, N and P in disturbed and undisturbed stands of a tropical wet-evergreen forest 2 2004
... 土壤微生物生物量的季节性动态变化是一个复杂过程, 其影响因子主要有土壤C含量和养分资源的限制(Landgraf & Klose, 2002; Koranda et al., 2013)、植物种类、土壤湿度以及土壤温度(杨凯等, 2009; López-Mondéjar et al., 2015)等.不同生态系统土壤微生物的季节变化呈现不同的格局(Devi & Yadava, 2006; Edwards et al., 2006).在温带森林生态系统, 土壤微生物生物量在植物生长季降低, 秋季逐渐升高, 呈现“W”形(Brookes et al., 1990; Lipson et al., 1999; 刘爽和王传宽, 2010); 在亚热带常绿阔叶林和热带季雨林, 土壤微生物生物量在夏季比冬季高(Devi & Yadava, 2006; Feng et al., 2009); 在北亚热带次生栎林和火炬松(Pinus taeda)人工林土壤微生物生物量在植物生长旺季维持在较低水平, 而在植物休眠季节维持在较高水平(王国兵等, 2008).在热带雨林, 土壤微生物生物量可能因为植物与土壤微生物之间剧烈的养分竞争而使土壤微生物量在树木生长旺盛时期较低(Barbhuiya et al., 2004).在湿润的北极苔原, 由于根系与土壤微生物之间的养分竞争, 使得土壤微生物生物量在春季冻融早期达到最大值, 随着温度回升而减少(Edwards et al., 2006).由此可见, 不同气候条件下的生态系统, 由于影响土壤微生物生物量的主导因子不同而使其季节变化格局产生差异. ...
... 温带森林和亚热带森林表现出的土壤微生物生物量“冬高夏低”格局可进一步理解为季节的变化使得积雪厚度、积雪存留时间和冬季土壤温度、自由水以及土壤微生物活力发生相应变化.土壤微生物生物量在冬季达到最大值(Brooks et al., 1998; Lipson et al., 1999, 2000, 2002; Schmidt & Lipson, 2004), 是由于在冬季, 真菌和细菌的生物量在积雪覆盖的土壤甚至比在夏季还要高(Brooks et al., 1996; Lipson et al., 2002).驱动雪下土壤微生物繁殖的碳和能量来源于植物枯枝落叶的输入, 温带森林、亚热带森林绝大多数枯枝落叶的凋落发生在秋季, 当地上生物量死亡或根系休眠的时候, 植物不会和土壤微生物竞争土壤养分, 且许多细根开始死亡并作为凋落物输入到土壤.这种枯枝落叶的数量可导致高浓度的纤维素和可溶性碳在冬季的释放(Lipson et al., 2000), 从而导致较高的土壤微生物生物量.另一方面, 冬季土壤经历了大量雪的积压, 导致根的破损, 而引起了蔗糖和其他可溶性糖向土壤的释放(Scott-Denton et al., 2006).土壤中来自于根的机械损伤而释放的蔗糖的浓度几乎是春天或夏天的8倍, 蔗糖的释放可作为降低细胞溶质冰点的一种方式, 防止在较浅的土层中根的冻伤(Scott-Denton et al., 2006).因此, 土壤中的可溶性碳在冬季迅速增加, 即发生在地下部分对光合产物的需求较低的时候.此外, 晚冬/早春期间, 由于土壤的冻融事件的频繁发生, 也可能会裂解植物和土壤微生物细胞而增加土壤碳的可获取性.而土壤微生物生物量在夏季达到最小值, 则可能是由于植物在夏季对土壤养分的大量需求限制了土壤微生物对养分的利用(Harte & Kinzig, 1993; Kaye & Hart, 1997; Barbhuiya et al., 2004), 土壤微生物与植物之间对养分的激烈竞争导致夏季土壤微生物量C、N、P含量较低. ...
Connecting mathematical ecosystems, real-world ecosystems, and climate science 1 2014
... 土壤微生物调节着土壤碳(C)的存储和氮(N)的固定与矿化, 其生物量的动态变化反映土壤养分及土壤碳的转化和循环, 其生物量的比值(C:N、N:P、C:P)可作为表征土壤营养限制的指标(Cleveland & Liptzin, 2007).土壤微生物生物量作为相对易分解和周转较快的营养库, 对C、N、磷(P)等化学元素的生物地球化学循环起到重要的驱动作用(Li et al., 2014); 同时, 也是许多碳循环模型的输入模块, 对预测气候变化对陆地生态系统的影响起到至关重要的作用(Bonan et al., 2014; Medlyn et al., 2015). ...
The soil microbial biomass: Its measurement, properties and role in nitrogen carbon dynamics following substrate incorporation 3 1990
... 土壤微生物生物量的季节性动态变化是一个复杂过程, 其影响因子主要有土壤C含量和养分资源的限制(Landgraf & Klose, 2002; Koranda et al., 2013)、植物种类、土壤湿度以及土壤温度(杨凯等, 2009; López-Mondéjar et al., 2015)等.不同生态系统土壤微生物的季节变化呈现不同的格局(Devi & Yadava, 2006; Edwards et al., 2006).在温带森林生态系统, 土壤微生物生物量在植物生长季降低, 秋季逐渐升高, 呈现“W”形(Brookes et al., 1990; Lipson et al., 1999; 刘爽和王传宽, 2010); 在亚热带常绿阔叶林和热带季雨林, 土壤微生物生物量在夏季比冬季高(Devi & Yadava, 2006; Feng et al., 2009); 在北亚热带次生栎林和火炬松(Pinus taeda)人工林土壤微生物生物量在植物生长旺季维持在较低水平, 而在植物休眠季节维持在较高水平(王国兵等, 2008).在热带雨林, 土壤微生物生物量可能因为植物与土壤微生物之间剧烈的养分竞争而使土壤微生物量在树木生长旺盛时期较低(Barbhuiya et al., 2004).在湿润的北极苔原, 由于根系与土壤微生物之间的养分竞争, 使得土壤微生物生物量在春季冻融早期达到最大值, 随着温度回升而减少(Edwards et al., 2006).由此可见, 不同气候条件下的生态系统, 由于影响土壤微生物生物量的主导因子不同而使其季节变化格局产生差异. ...
... 本研究发现土壤微生物生物量的季节波动与植物根系对营养的竞争以及林木的生长节律有关, 温带森林和亚热带森林的土壤微生物量的季节变化格局基本都呈现非生长季维持在较高水平, 而随着植物进入生长季而下降.这与其他一些温带森林的研究结果 (Brookes et al., 1990; Lipson et al., 1999; 王国兵等, 2008; 刘爽和王传宽, 2010; 王薪琪等, 2017)一致.可能的解释为: 秋季大量凋落物的输入为冬季土壤微生物提供了丰富的代谢底物.尽管冬季气温很低, 但处于积雪覆盖下的土壤温度基本能够维持土壤微生物生长和繁殖.另外, 由于冬季低温, 土壤微生物代谢较弱, 其维持呼吸所需要的能量较低, 在较低能量供给条件下就能维持较高的土壤微生物量(Brookes et al., 1990), 因此冬季土壤微生物量均较高.随着气温的升高进入春季, 早春植物(一些短命草本)和土壤微生物共同固持养分可能形成“春坝” (vernal dam)作用(Warren和邹晓明, 2003), 为植物根系和土壤微生物的快速生长提供养分, 从而使土壤微生物生物量呈现上升趋势.随着根系的恢复生长, 植物进入旺盛生长季(夏季), 土壤微生物与植物之间对养分的激烈竞争, 使土壤微生物生物量呈现下降趋势(Kaiser et al., 2011).由此可见, 在温带和亚热带森林生态系统中, 植物和土壤微生物对养分的不同步吸收是保留养分和维持生态系统生产力的一种机制(Muller & Herbert Bormann, 1976; Singh et al., 1989; Groffman et al., 1993; Zak et al., 1993; Kaiser et al., 2014). ...
... )一致.可能的解释为: 秋季大量凋落物的输入为冬季土壤微生物提供了丰富的代谢底物.尽管冬季气温很低, 但处于积雪覆盖下的土壤温度基本能够维持土壤微生物生长和繁殖.另外, 由于冬季低温, 土壤微生物代谢较弱, 其维持呼吸所需要的能量较低, 在较低能量供给条件下就能维持较高的土壤微生物量(Brookes et al., 1990), 因此冬季土壤微生物量均较高.随着气温的升高进入春季, 早春植物(一些短命草本)和土壤微生物共同固持养分可能形成“春坝” (vernal dam)作用(Warren和邹晓明, 2003), 为植物根系和土壤微生物的快速生长提供养分, 从而使土壤微生物生物量呈现上升趋势.随着根系的恢复生长, 植物进入旺盛生长季(夏季), 土壤微生物与植物之间对养分的激烈竞争, 使土壤微生物生物量呈现下降趋势(Kaiser et al., 2011).由此可见, 在温带和亚热带森林生态系统中, 植物和土壤微生物对养分的不同步吸收是保留养分和维持生态系统生产力的一种机制(Muller & Herbert Bormann, 1976; Singh et al., 1989; Groffman et al., 1993; Zak et al., 1993; Kaiser et al., 2014). ...
Microbial activity under alpine snowpacks, Niwot Ridge, Colorado 1 1996
... 温带森林和亚热带森林表现出的土壤微生物生物量“冬高夏低”格局可进一步理解为季节的变化使得积雪厚度、积雪存留时间和冬季土壤温度、自由水以及土壤微生物活力发生相应变化.土壤微生物生物量在冬季达到最大值(Brooks et al., 1998; Lipson et al., 1999, 2000, 2002; Schmidt & Lipson, 2004), 是由于在冬季, 真菌和细菌的生物量在积雪覆盖的土壤甚至比在夏季还要高(Brooks et al., 1996; Lipson et al., 2002).驱动雪下土壤微生物繁殖的碳和能量来源于植物枯枝落叶的输入, 温带森林、亚热带森林绝大多数枯枝落叶的凋落发生在秋季, 当地上生物量死亡或根系休眠的时候, 植物不会和土壤微生物竞争土壤养分, 且许多细根开始死亡并作为凋落物输入到土壤.这种枯枝落叶的数量可导致高浓度的纤维素和可溶性碳在冬季的释放(Lipson et al., 2000), 从而导致较高的土壤微生物生物量.另一方面, 冬季土壤经历了大量雪的积压, 导致根的破损, 而引起了蔗糖和其他可溶性糖向土壤的释放(Scott-Denton et al., 2006).土壤中来自于根的机械损伤而释放的蔗糖的浓度几乎是春天或夏天的8倍, 蔗糖的释放可作为降低细胞溶质冰点的一种方式, 防止在较浅的土层中根的冻伤(Scott-Denton et al., 2006).因此, 土壤中的可溶性碳在冬季迅速增加, 即发生在地下部分对光合产物的需求较低的时候.此外, 晚冬/早春期间, 由于土壤的冻融事件的频繁发生, 也可能会裂解植物和土壤微生物细胞而增加土壤碳的可获取性.而土壤微生物生物量在夏季达到最小值, 则可能是由于植物在夏季对土壤养分的大量需求限制了土壤微生物对养分的利用(Harte & Kinzig, 1993; Kaye & Hart, 1997; Barbhuiya et al., 2004), 土壤微生物与植物之间对养分的激烈竞争导致夏季土壤微生物量C、N、P含量较低. ...
Inorganic N and microbial biomass dynamics before and during spring snowmelt 1 1998
... 温带森林和亚热带森林表现出的土壤微生物生物量“冬高夏低”格局可进一步理解为季节的变化使得积雪厚度、积雪存留时间和冬季土壤温度、自由水以及土壤微生物活力发生相应变化.土壤微生物生物量在冬季达到最大值(Brooks et al., 1998; Lipson et al., 1999, 2000, 2002; Schmidt & Lipson, 2004), 是由于在冬季, 真菌和细菌的生物量在积雪覆盖的土壤甚至比在夏季还要高(Brooks et al., 1996; Lipson et al., 2002).驱动雪下土壤微生物繁殖的碳和能量来源于植物枯枝落叶的输入, 温带森林、亚热带森林绝大多数枯枝落叶的凋落发生在秋季, 当地上生物量死亡或根系休眠的时候, 植物不会和土壤微生物竞争土壤养分, 且许多细根开始死亡并作为凋落物输入到土壤.这种枯枝落叶的数量可导致高浓度的纤维素和可溶性碳在冬季的释放(Lipson et al., 2000), 从而导致较高的土壤微生物生物量.另一方面, 冬季土壤经历了大量雪的积压, 导致根的破损, 而引起了蔗糖和其他可溶性糖向土壤的释放(Scott-Denton et al., 2006).土壤中来自于根的机械损伤而释放的蔗糖的浓度几乎是春天或夏天的8倍, 蔗糖的释放可作为降低细胞溶质冰点的一种方式, 防止在较浅的土层中根的冻伤(Scott-Denton et al., 2006).因此, 土壤中的可溶性碳在冬季迅速增加, 即发生在地下部分对光合产物的需求较低的时候.此外, 晚冬/早春期间, 由于土壤的冻融事件的频繁发生, 也可能会裂解植物和土壤微生物细胞而增加土壤碳的可获取性.而土壤微生物生物量在夏季达到最小值, 则可能是由于植物在夏季对土壤养分的大量需求限制了土壤微生物对养分的利用(Harte & Kinzig, 1993; Kaye & Hart, 1997; Barbhuiya et al., 2004), 土壤微生物与植物之间对养分的激烈竞争导致夏季土壤微生物量C、N、P含量较低. ...
C:N:P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomass? 1 2007
... 土壤微生物调节着土壤碳(C)的存储和氮(N)的固定与矿化, 其生物量的动态变化反映土壤养分及土壤碳的转化和循环, 其生物量的比值(C:N、N:P、C:P)可作为表征土壤营养限制的指标(Cleveland & Liptzin, 2007).土壤微生物生物量作为相对易分解和周转较快的营养库, 对C、N、磷(P)等化学元素的生物地球化学循环起到重要的驱动作用(Li et al., 2014); 同时, 也是许多碳循环模型的输入模块, 对预测气候变化对陆地生态系统的影响起到至关重要的作用(Bonan et al., 2014; Medlyn et al., 2015). ...
Ecosystem scale trade-off in nitrogen acquisition pathways 1 2018
... 在陆地生态系统中, 植物是第一生产者, 土壤微生物是有机质的分解者.植物将光合产物以根系分泌物和植物残体(凋落物)的形式释放到土壤, 供给土壤微生物碳源(Zak et al., 2000); 而土壤微生物一方面通过自身分解和代谢将有机养分转化成无机养分, 以利于植物吸收利用(Schimel & Weintraub, 2003; Fang et al., 2005; Neill & Gignoux, 2006); 另一方面通过与根系的共生关系(菌根真菌)直接利用土壤里的小分子有机物质促进植物生长(Deng et al., 2018).这种植物-土壤微生物的相互作用维系着陆地生态系统的生态功能.尽管如此, 植物根系和土壤微生物之间存在着既相互依存又相互制约的关系(图1): 植物为土壤微生物的生长提供C源从而促进土壤微生物的活动, 而土壤微生物矿化有机养分提高植物对养分利用的有效性; 但是土壤微生物也同植物根系争夺营养元素, 它们对土壤中营养的利用属于竞争关系(Singh et al., 1989; Harte & Kinzig, 1993; Kaye & Hart, 1997; Terrer et al., 2018).这种植物和土壤微生物呈现的复杂的互利-竞争关系存在于季节交替过程中, 植物输入土壤中的凋落物在数量和质量上的改变, 土壤可利用营养在充足和缺乏之间变化, 加之植物在生长季和休眠季对养分需求的不同, 就有可能改变土壤微生物对N的需求(即矿化-同化途径), 限制植物对无机N的吸收(Zak et al., 2000; Terrer et al., 2018), 而从对养分的矿化转为对养分的同化, 增加自身的生物量, 从而在土壤微生物生物量上可能呈现一个季节波动特征. ...
Seasonal dynamics in soil microbial biomass C, N and P in a mixed-oak forest ecosystem of Manipur, North-east India 2 2006
... 土壤微生物生物量的季节性动态变化是一个复杂过程, 其影响因子主要有土壤C含量和养分资源的限制(Landgraf & Klose, 2002; Koranda et al., 2013)、植物种类、土壤湿度以及土壤温度(杨凯等, 2009; López-Mondéjar et al., 2015)等.不同生态系统土壤微生物的季节变化呈现不同的格局(Devi & Yadava, 2006; Edwards et al., 2006).在温带森林生态系统, 土壤微生物生物量在植物生长季降低, 秋季逐渐升高, 呈现“W”形(Brookes et al., 1990; Lipson et al., 1999; 刘爽和王传宽, 2010); 在亚热带常绿阔叶林和热带季雨林, 土壤微生物生物量在夏季比冬季高(Devi & Yadava, 2006; Feng et al., 2009); 在北亚热带次生栎林和火炬松(Pinus taeda)人工林土壤微生物生物量在植物生长旺季维持在较低水平, 而在植物休眠季节维持在较高水平(王国兵等, 2008).在热带雨林, 土壤微生物生物量可能因为植物与土壤微生物之间剧烈的养分竞争而使土壤微生物量在树木生长旺盛时期较低(Barbhuiya et al., 2004).在湿润的北极苔原, 由于根系与土壤微生物之间的养分竞争, 使得土壤微生物生物量在春季冻融早期达到最大值, 随着温度回升而减少(Edwards et al., 2006).由此可见, 不同气候条件下的生态系统, 由于影响土壤微生物生物量的主导因子不同而使其季节变化格局产生差异. ...
... ); 在亚热带常绿阔叶林和热带季雨林, 土壤微生物生物量在夏季比冬季高(Devi & Yadava, 2006; Feng et al., 2009); 在北亚热带次生栎林和火炬松(Pinus taeda)人工林土壤微生物生物量在植物生长旺季维持在较低水平, 而在植物休眠季节维持在较高水平(王国兵等, 2008).在热带雨林, 土壤微生物生物量可能因为植物与土壤微生物之间剧烈的养分竞争而使土壤微生物量在树木生长旺盛时期较低(Barbhuiya et al., 2004).在湿润的北极苔原, 由于根系与土壤微生物之间的养分竞争, 使得土壤微生物生物量在春季冻融早期达到最大值, 随着温度回升而减少(Edwards et al., 2006).由此可见, 不同气候条件下的生态系统, 由于影响土壤微生物生物量的主导因子不同而使其季节变化格局产生差异. ...
Soil microbial and nutrient dynamics in a wet Arctic sedge meadow in late winter and early spring 2 2006
... 土壤微生物生物量的季节性动态变化是一个复杂过程, 其影响因子主要有土壤C含量和养分资源的限制(Landgraf & Klose, 2002; Koranda et al., 2013)、植物种类、土壤湿度以及土壤温度(杨凯等, 2009; López-Mondéjar et al., 2015)等.不同生态系统土壤微生物的季节变化呈现不同的格局(Devi & Yadava, 2006; Edwards et al., 2006).在温带森林生态系统, 土壤微生物生物量在植物生长季降低, 秋季逐渐升高, 呈现“W”形(Brookes et al., 1990; Lipson et al., 1999; 刘爽和王传宽, 2010); 在亚热带常绿阔叶林和热带季雨林, 土壤微生物生物量在夏季比冬季高(Devi & Yadava, 2006; Feng et al., 2009); 在北亚热带次生栎林和火炬松(Pinus taeda)人工林土壤微生物生物量在植物生长旺季维持在较低水平, 而在植物休眠季节维持在较高水平(王国兵等, 2008).在热带雨林, 土壤微生物生物量可能因为植物与土壤微生物之间剧烈的养分竞争而使土壤微生物量在树木生长旺盛时期较低(Barbhuiya et al., 2004).在湿润的北极苔原, 由于根系与土壤微生物之间的养分竞争, 使得土壤微生物生物量在春季冻融早期达到最大值, 随着温度回升而减少(Edwards et al., 2006).由此可见, 不同气候条件下的生态系统, 由于影响土壤微生物生物量的主导因子不同而使其季节变化格局产生差异. ...
Competition for nitrogen between plants and soil microorganisms 2 1997
... 在陆地生态系统中, 植物是第一生产者, 土壤微生物是有机质的分解者.植物将光合产物以根系分泌物和植物残体(凋落物)的形式释放到土壤, 供给土壤微生物碳源(Zak et al., 2000); 而土壤微生物一方面通过自身分解和代谢将有机养分转化成无机养分, 以利于植物吸收利用(Schimel & Weintraub, 2003; Fang et al., 2005; Neill & Gignoux, 2006); 另一方面通过与根系的共生关系(菌根真菌)直接利用土壤里的小分子有机物质促进植物生长(Deng et al., 2018).这种植物-土壤微生物的相互作用维系着陆地生态系统的生态功能.尽管如此, 植物根系和土壤微生物之间存在着既相互依存又相互制约的关系(图1): 植物为土壤微生物的生长提供C源从而促进土壤微生物的活动, 而土壤微生物矿化有机养分提高植物对养分利用的有效性; 但是土壤微生物也同植物根系争夺营养元素, 它们对土壤中营养的利用属于竞争关系(Singh et al., 1989; Harte & Kinzig, 1993; Kaye & Hart, 1997; Terrer et al., 2018).这种植物和土壤微生物呈现的复杂的互利-竞争关系存在于季节交替过程中, 植物输入土壤中的凋落物在数量和质量上的改变, 土壤可利用营养在充足和缺乏之间变化, 加之植物在生长季和休眠季对养分需求的不同, 就有可能改变土壤微生物对N的需求(即矿化-同化途径), 限制植物对无机N的吸收(Zak et al., 2000; Terrer et al., 2018), 而从对养分的矿化转为对养分的同化, 增加自身的生物量, 从而在土壤微生物生物量上可能呈现一个季节波动特征. ...
... 温带森林和亚热带森林表现出的土壤微生物生物量“冬高夏低”格局可进一步理解为季节的变化使得积雪厚度、积雪存留时间和冬季土壤温度、自由水以及土壤微生物活力发生相应变化.土壤微生物生物量在冬季达到最大值(Brooks et al., 1998; Lipson et al., 1999, 2000, 2002; Schmidt & Lipson, 2004), 是由于在冬季, 真菌和细菌的生物量在积雪覆盖的土壤甚至比在夏季还要高(Brooks et al., 1996; Lipson et al., 2002).驱动雪下土壤微生物繁殖的碳和能量来源于植物枯枝落叶的输入, 温带森林、亚热带森林绝大多数枯枝落叶的凋落发生在秋季, 当地上生物量死亡或根系休眠的时候, 植物不会和土壤微生物竞争土壤养分, 且许多细根开始死亡并作为凋落物输入到土壤.这种枯枝落叶的数量可导致高浓度的纤维素和可溶性碳在冬季的释放(Lipson et al., 2000), 从而导致较高的土壤微生物生物量.另一方面, 冬季土壤经历了大量雪的积压, 导致根的破损, 而引起了蔗糖和其他可溶性糖向土壤的释放(Scott-Denton et al., 2006).土壤中来自于根的机械损伤而释放的蔗糖的浓度几乎是春天或夏天的8倍, 蔗糖的释放可作为降低细胞溶质冰点的一种方式, 防止在较浅的土层中根的冻伤(Scott-Denton et al., 2006).因此, 土壤中的可溶性碳在冬季迅速增加, 即发生在地下部分对光合产物的需求较低的时候.此外, 晚冬/早春期间, 由于土壤的冻融事件的频繁发生, 也可能会裂解植物和土壤微生物细胞而增加土壤碳的可获取性.而土壤微生物生物量在夏季达到最小值, 则可能是由于植物在夏季对土壤养分的大量需求限制了土壤微生物对养分的利用(Harte & Kinzig, 1993; Kaye & Hart, 1997; Barbhuiya et al., 2004), 土壤微生物与植物之间对养分的激烈竞争导致夏季土壤微生物量C、N、P含量较低. ...
Seasonal variation in functional properties of microbial communities in beech forest soil 1 2013
... 土壤微生物生物量的季节性动态变化是一个复杂过程, 其影响因子主要有土壤C含量和养分资源的限制(Landgraf & Klose, 2002; Koranda et al., 2013)、植物种类、土壤湿度以及土壤温度(杨凯等, 2009; López-Mondéjar et al., 2015)等.不同生态系统土壤微生物的季节变化呈现不同的格局(Devi & Yadava, 2006; Edwards et al., 2006).在温带森林生态系统, 土壤微生物生物量在植物生长季降低, 秋季逐渐升高, 呈现“W”形(Brookes et al., 1990; Lipson et al., 1999; 刘爽和王传宽, 2010); 在亚热带常绿阔叶林和热带季雨林, 土壤微生物生物量在夏季比冬季高(Devi & Yadava, 2006; Feng et al., 2009); 在北亚热带次生栎林和火炬松(Pinus taeda)人工林土壤微生物生物量在植物生长旺季维持在较低水平, 而在植物休眠季节维持在较高水平(王国兵等, 2008).在热带雨林, 土壤微生物生物量可能因为植物与土壤微生物之间剧烈的养分竞争而使土壤微生物量在树木生长旺盛时期较低(Barbhuiya et al., 2004).在湿润的北极苔原, 由于根系与土壤微生物之间的养分竞争, 使得土壤微生物生物量在春季冻融早期达到最大值, 随着温度回升而减少(Edwards et al., 2006).由此可见, 不同气候条件下的生态系统, 由于影响土壤微生物生物量的主导因子不同而使其季节变化格局产生差异. ...
Mobile and readily available C and N fractions and their relationship to microbial biomass and selected enzyme activities in a sandy soil under different management systems 1 2002
... 土壤微生物生物量的季节性动态变化是一个复杂过程, 其影响因子主要有土壤C含量和养分资源的限制(Landgraf & Klose, 2002; Koranda et al., 2013)、植物种类、土壤湿度以及土壤温度(杨凯等, 2009; López-Mondéjar et al., 2015)等.不同生态系统土壤微生物的季节变化呈现不同的格局(Devi & Yadava, 2006; Edwards et al., 2006).在温带森林生态系统, 土壤微生物生物量在植物生长季降低, 秋季逐渐升高, 呈现“W”形(Brookes et al., 1990; Lipson et al., 1999; 刘爽和王传宽, 2010); 在亚热带常绿阔叶林和热带季雨林, 土壤微生物生物量在夏季比冬季高(Devi & Yadava, 2006; Feng et al., 2009); 在北亚热带次生栎林和火炬松(Pinus taeda)人工林土壤微生物生物量在植物生长旺季维持在较低水平, 而在植物休眠季节维持在较高水平(王国兵等, 2008).在热带雨林, 土壤微生物生物量可能因为植物与土壤微生物之间剧烈的养分竞争而使土壤微生物量在树木生长旺盛时期较低(Barbhuiya et al., 2004).在湿润的北极苔原, 由于根系与土壤微生物之间的养分竞争, 使得土壤微生物生物量在春季冻融早期达到最大值, 随着温度回升而减少(Edwards et al., 2006).由此可见, 不同气候条件下的生态系统, 由于影响土壤微生物生物量的主导因子不同而使其季节变化格局产生差异. ...
Global patterns of soil microbial nitrogen and phosphorus stoichiometry in forest ecosystems 2 2014
... 土壤微生物调节着土壤碳(C)的存储和氮(N)的固定与矿化, 其生物量的动态变化反映土壤养分及土壤碳的转化和循环, 其生物量的比值(C:N、N:P、C:P)可作为表征土壤营养限制的指标(Cleveland & Liptzin, 2007).土壤微生物生物量作为相对易分解和周转较快的营养库, 对C、N、磷(P)等化学元素的生物地球化学循环起到重要的驱动作用(Li et al., 2014); 同时, 也是许多碳循环模型的输入模块, 对预测气候变化对陆地生态系统的影响起到至关重要的作用(Bonan et al., 2014; Medlyn et al., 2015). ...
An extraction method for measuring soil microbial biomass C 1 1987
... 为了避免文献选择的人为偏性, 收集的数据有3条筛选标准: (1)数据均来自森林对照组, 排除自然干扰(火灾、病虫害等)或人为干扰(如增温、施肥、间伐等)的数据.(2)所有数据的测定方法都是氯仿熏蒸浸提法(Vance et al., 1987), 该方法被认为是测定土壤微生物生物量最常用的方法, 并能够测得土壤中总微生物生物量(包括细菌和真菌)(Fierer et al., 2009).(3)选取样点土壤剖面表层(厚度0到20 cm)的数据. ...
北亚热带次生栎林与火炬松人工林土壤微生物生物量碳的季节动态 2 2008
... 土壤微生物生物量的季节性动态变化是一个复杂过程, 其影响因子主要有土壤C含量和养分资源的限制(Landgraf & Klose, 2002; Koranda et al., 2013)、植物种类、土壤湿度以及土壤温度(杨凯等, 2009; López-Mondéjar et al., 2015)等.不同生态系统土壤微生物的季节变化呈现不同的格局(Devi & Yadava, 2006; Edwards et al., 2006).在温带森林生态系统, 土壤微生物生物量在植物生长季降低, 秋季逐渐升高, 呈现“W”形(Brookes et al., 1990; Lipson et al., 1999; 刘爽和王传宽, 2010); 在亚热带常绿阔叶林和热带季雨林, 土壤微生物生物量在夏季比冬季高(Devi & Yadava, 2006; Feng et al., 2009); 在北亚热带次生栎林和火炬松(Pinus taeda)人工林土壤微生物生物量在植物生长旺季维持在较低水平, 而在植物休眠季节维持在较高水平(王国兵等, 2008).在热带雨林, 土壤微生物生物量可能因为植物与土壤微生物之间剧烈的养分竞争而使土壤微生物量在树木生长旺盛时期较低(Barbhuiya et al., 2004).在湿润的北极苔原, 由于根系与土壤微生物之间的养分竞争, 使得土壤微生物生物量在春季冻融早期达到最大值, 随着温度回升而减少(Edwards et al., 2006).由此可见, 不同气候条件下的生态系统, 由于影响土壤微生物生物量的主导因子不同而使其季节变化格局产生差异. ...
... 本研究发现土壤微生物生物量的季节波动与植物根系对营养的竞争以及林木的生长节律有关, 温带森林和亚热带森林的土壤微生物量的季节变化格局基本都呈现非生长季维持在较高水平, 而随着植物进入生长季而下降.这与其他一些温带森林的研究结果 (Brookes et al., 1990; Lipson et al., 1999; 王国兵等, 2008; 刘爽和王传宽, 2010; 王薪琪等, 2017)一致.可能的解释为: 秋季大量凋落物的输入为冬季土壤微生物提供了丰富的代谢底物.尽管冬季气温很低, 但处于积雪覆盖下的土壤温度基本能够维持土壤微生物生长和繁殖.另外, 由于冬季低温, 土壤微生物代谢较弱, 其维持呼吸所需要的能量较低, 在较低能量供给条件下就能维持较高的土壤微生物量(Brookes et al., 1990), 因此冬季土壤微生物量均较高.随着气温的升高进入春季, 早春植物(一些短命草本)和土壤微生物共同固持养分可能形成“春坝” (vernal dam)作用(Warren和邹晓明, 2003), 为植物根系和土壤微生物的快速生长提供养分, 从而使土壤微生物生物量呈现上升趋势.随着根系的恢复生长, 植物进入旺盛生长季(夏季), 土壤微生物与植物之间对养分的激烈竞争, 使土壤微生物生物量呈现下降趋势(Kaiser et al., 2011).由此可见, 在温带和亚热带森林生态系统中, 植物和土壤微生物对养分的不同步吸收是保留养分和维持生态系统生产力的一种机制(Muller & Herbert Bormann, 1976; Singh et al., 1989; Groffman et al., 1993; Zak et al., 1993; Kaiser et al., 2014). ...
北亚热带次生栎林与火炬松人工林土壤微生物生物量碳的季节动态 2 2008
... 土壤微生物生物量的季节性动态变化是一个复杂过程, 其影响因子主要有土壤C含量和养分资源的限制(Landgraf & Klose, 2002; Koranda et al., 2013)、植物种类、土壤湿度以及土壤温度(杨凯等, 2009; López-Mondéjar et al., 2015)等.不同生态系统土壤微生物的季节变化呈现不同的格局(Devi & Yadava, 2006; Edwards et al., 2006).在温带森林生态系统, 土壤微生物生物量在植物生长季降低, 秋季逐渐升高, 呈现“W”形(Brookes et al., 1990; Lipson et al., 1999; 刘爽和王传宽, 2010); 在亚热带常绿阔叶林和热带季雨林, 土壤微生物生物量在夏季比冬季高(Devi & Yadava, 2006; Feng et al., 2009); 在北亚热带次生栎林和火炬松(Pinus taeda)人工林土壤微生物生物量在植物生长旺季维持在较低水平, 而在植物休眠季节维持在较高水平(王国兵等, 2008).在热带雨林, 土壤微生物生物量可能因为植物与土壤微生物之间剧烈的养分竞争而使土壤微生物量在树木生长旺盛时期较低(Barbhuiya et al., 2004).在湿润的北极苔原, 由于根系与土壤微生物之间的养分竞争, 使得土壤微生物生物量在春季冻融早期达到最大值, 随着温度回升而减少(Edwards et al., 2006).由此可见, 不同气候条件下的生态系统, 由于影响土壤微生物生物量的主导因子不同而使其季节变化格局产生差异. ...
... 本研究发现土壤微生物生物量的季节波动与植物根系对营养的竞争以及林木的生长节律有关, 温带森林和亚热带森林的土壤微生物量的季节变化格局基本都呈现非生长季维持在较高水平, 而随着植物进入生长季而下降.这与其他一些温带森林的研究结果 (Brookes et al., 1990; Lipson et al., 1999; 王国兵等, 2008; 刘爽和王传宽, 2010; 王薪琪等, 2017)一致.可能的解释为: 秋季大量凋落物的输入为冬季土壤微生物提供了丰富的代谢底物.尽管冬季气温很低, 但处于积雪覆盖下的土壤温度基本能够维持土壤微生物生长和繁殖.另外, 由于冬季低温, 土壤微生物代谢较弱, 其维持呼吸所需要的能量较低, 在较低能量供给条件下就能维持较高的土壤微生物量(Brookes et al., 1990), 因此冬季土壤微生物量均较高.随着气温的升高进入春季, 早春植物(一些短命草本)和土壤微生物共同固持养分可能形成“春坝” (vernal dam)作用(Warren和邹晓明, 2003), 为植物根系和土壤微生物的快速生长提供养分, 从而使土壤微生物生物量呈现上升趋势.随着根系的恢复生长, 植物进入旺盛生长季(夏季), 土壤微生物与植物之间对养分的激烈竞争, 使土壤微生物生物量呈现下降趋势(Kaiser et al., 2011).由此可见, 在温带和亚热带森林生态系统中, 植物和土壤微生物对养分的不同步吸收是保留养分和维持生态系统生产力的一种机制(Muller & Herbert Bormann, 1976; Singh et al., 1989; Groffman et al., 1993; Zak et al., 1993; Kaiser et al., 2014). ...
帽儿山不同林龄落叶阔叶林土壤微生物生物量及其季节动态 1 2017
... 本研究发现土壤微生物生物量的季节波动与植物根系对营养的竞争以及林木的生长节律有关, 温带森林和亚热带森林的土壤微生物量的季节变化格局基本都呈现非生长季维持在较高水平, 而随着植物进入生长季而下降.这与其他一些温带森林的研究结果 (Brookes et al., 1990; Lipson et al., 1999; 王国兵等, 2008; 刘爽和王传宽, 2010; 王薪琪等, 2017)一致.可能的解释为: 秋季大量凋落物的输入为冬季土壤微生物提供了丰富的代谢底物.尽管冬季气温很低, 但处于积雪覆盖下的土壤温度基本能够维持土壤微生物生长和繁殖.另外, 由于冬季低温, 土壤微生物代谢较弱, 其维持呼吸所需要的能量较低, 在较低能量供给条件下就能维持较高的土壤微生物量(Brookes et al., 1990), 因此冬季土壤微生物量均较高.随着气温的升高进入春季, 早春植物(一些短命草本)和土壤微生物共同固持养分可能形成“春坝” (vernal dam)作用(Warren和邹晓明, 2003), 为植物根系和土壤微生物的快速生长提供养分, 从而使土壤微生物生物量呈现上升趋势.随着根系的恢复生长, 植物进入旺盛生长季(夏季), 土壤微生物与植物之间对养分的激烈竞争, 使土壤微生物生物量呈现下降趋势(Kaiser et al., 2011).由此可见, 在温带和亚热带森林生态系统中, 植物和土壤微生物对养分的不同步吸收是保留养分和维持生态系统生产力的一种机制(Muller & Herbert Bormann, 1976; Singh et al., 1989; Groffman et al., 1993; Zak et al., 1993; Kaiser et al., 2014). ...
帽儿山不同林龄落叶阔叶林土壤微生物生物量及其季节动态 1 2017
... 本研究发现土壤微生物生物量的季节波动与植物根系对营养的竞争以及林木的生长节律有关, 温带森林和亚热带森林的土壤微生物量的季节变化格局基本都呈现非生长季维持在较高水平, 而随着植物进入生长季而下降.这与其他一些温带森林的研究结果 (Brookes et al., 1990; Lipson et al., 1999; 王国兵等, 2008; 刘爽和王传宽, 2010; 王薪琪等, 2017)一致.可能的解释为: 秋季大量凋落物的输入为冬季土壤微生物提供了丰富的代谢底物.尽管冬季气温很低, 但处于积雪覆盖下的土壤温度基本能够维持土壤微生物生长和繁殖.另外, 由于冬季低温, 土壤微生物代谢较弱, 其维持呼吸所需要的能量较低, 在较低能量供给条件下就能维持较高的土壤微生物量(Brookes et al., 1990), 因此冬季土壤微生物量均较高.随着气温的升高进入春季, 早春植物(一些短命草本)和土壤微生物共同固持养分可能形成“春坝” (vernal dam)作用(Warren和邹晓明, 2003), 为植物根系和土壤微生物的快速生长提供养分, 从而使土壤微生物生物量呈现上升趋势.随着根系的恢复生长, 植物进入旺盛生长季(夏季), 土壤微生物与植物之间对养分的激烈竞争, 使土壤微生物生物量呈现下降趋势(Kaiser et al., 2011).由此可见, 在温带和亚热带森林生态系统中, 植物和土壤微生物对养分的不同步吸收是保留养分和维持生态系统生产力的一种机制(Muller & Herbert Bormann, 1976; Singh et al., 1989; Groffman et al., 1993; Zak et al., 1993; Kaiser et al., 2014). ...
温带阔叶林中氮的保留机制: “春坝”假设及研究实例 1 2003
... 本研究发现土壤微生物生物量的季节波动与植物根系对营养的竞争以及林木的生长节律有关, 温带森林和亚热带森林的土壤微生物量的季节变化格局基本都呈现非生长季维持在较高水平, 而随着植物进入生长季而下降.这与其他一些温带森林的研究结果 (Brookes et al., 1990; Lipson et al., 1999; 王国兵等, 2008; 刘爽和王传宽, 2010; 王薪琪等, 2017)一致.可能的解释为: 秋季大量凋落物的输入为冬季土壤微生物提供了丰富的代谢底物.尽管冬季气温很低, 但处于积雪覆盖下的土壤温度基本能够维持土壤微生物生长和繁殖.另外, 由于冬季低温, 土壤微生物代谢较弱, 其维持呼吸所需要的能量较低, 在较低能量供给条件下就能维持较高的土壤微生物量(Brookes et al., 1990), 因此冬季土壤微生物量均较高.随着气温的升高进入春季, 早春植物(一些短命草本)和土壤微生物共同固持养分可能形成“春坝” (vernal dam)作用(Warren和邹晓明, 2003), 为植物根系和土壤微生物的快速生长提供养分, 从而使土壤微生物生物量呈现上升趋势.随着根系的恢复生长, 植物进入旺盛生长季(夏季), 土壤微生物与植物之间对养分的激烈竞争, 使土壤微生物生物量呈现下降趋势(Kaiser et al., 2011).由此可见, 在温带和亚热带森林生态系统中, 植物和土壤微生物对养分的不同步吸收是保留养分和维持生态系统生产力的一种机制(Muller & Herbert Bormann, 1976; Singh et al., 1989; Groffman et al., 1993; Zak et al., 1993; Kaiser et al., 2014). ...
温带阔叶林中氮的保留机制: “春坝”假设及研究实例 1 2003
... 本研究发现土壤微生物生物量的季节波动与植物根系对营养的竞争以及林木的生长节律有关, 温带森林和亚热带森林的土壤微生物量的季节变化格局基本都呈现非生长季维持在较高水平, 而随着植物进入生长季而下降.这与其他一些温带森林的研究结果 (Brookes et al., 1990; Lipson et al., 1999; 王国兵等, 2008; 刘爽和王传宽, 2010; 王薪琪等, 2017)一致.可能的解释为: 秋季大量凋落物的输入为冬季土壤微生物提供了丰富的代谢底物.尽管冬季气温很低, 但处于积雪覆盖下的土壤温度基本能够维持土壤微生物生长和繁殖.另外, 由于冬季低温, 土壤微生物代谢较弱, 其维持呼吸所需要的能量较低, 在较低能量供给条件下就能维持较高的土壤微生物量(Brookes et al., 1990), 因此冬季土壤微生物量均较高.随着气温的升高进入春季, 早春植物(一些短命草本)和土壤微生物共同固持养分可能形成“春坝” (vernal dam)作用(Warren和邹晓明, 2003), 为植物根系和土壤微生物的快速生长提供养分, 从而使土壤微生物生物量呈现上升趋势.随着根系的恢复生长, 植物进入旺盛生长季(夏季), 土壤微生物与植物之间对养分的激烈竞争, 使土壤微生物生物量呈现下降趋势(Kaiser et al., 2011).由此可见, 在温带和亚热带森林生态系统中, 植物和土壤微生物对养分的不同步吸收是保留养分和维持生态系统生产力的一种机制(Muller & Herbert Bormann, 1976; Singh et al., 1989; Groffman et al., 1993; Zak et al., 1993; Kaiser et al., 2014). ...
不同林龄落叶松人工林土壤微生物生物量碳氮的季节变化 1 2009
... 土壤微生物生物量的季节性动态变化是一个复杂过程, 其影响因子主要有土壤C含量和养分资源的限制(Landgraf & Klose, 2002; Koranda et al., 2013)、植物种类、土壤湿度以及土壤温度(杨凯等, 2009; López-Mondéjar et al., 2015)等.不同生态系统土壤微生物的季节变化呈现不同的格局(Devi & Yadava, 2006; Edwards et al., 2006).在温带森林生态系统, 土壤微生物生物量在植物生长季降低, 秋季逐渐升高, 呈现“W”形(Brookes et al., 1990; Lipson et al., 1999; 刘爽和王传宽, 2010); 在亚热带常绿阔叶林和热带季雨林, 土壤微生物生物量在夏季比冬季高(Devi & Yadava, 2006; Feng et al., 2009); 在北亚热带次生栎林和火炬松(Pinus taeda)人工林土壤微生物生物量在植物生长旺季维持在较低水平, 而在植物休眠季节维持在较高水平(王国兵等, 2008).在热带雨林, 土壤微生物生物量可能因为植物与土壤微生物之间剧烈的养分竞争而使土壤微生物量在树木生长旺盛时期较低(Barbhuiya et al., 2004).在湿润的北极苔原, 由于根系与土壤微生物之间的养分竞争, 使得土壤微生物生物量在春季冻融早期达到最大值, 随着温度回升而减少(Edwards et al., 2006).由此可见, 不同气候条件下的生态系统, 由于影响土壤微生物生物量的主导因子不同而使其季节变化格局产生差异. ...
不同林龄落叶松人工林土壤微生物生物量碳氮的季节变化 1 2009
... 土壤微生物生物量的季节性动态变化是一个复杂过程, 其影响因子主要有土壤C含量和养分资源的限制(Landgraf & Klose, 2002; Koranda et al., 2013)、植物种类、土壤湿度以及土壤温度(杨凯等, 2009; López-Mondéjar et al., 2015)等.不同生态系统土壤微生物的季节变化呈现不同的格局(Devi & Yadava, 2006; Edwards et al., 2006).在温带森林生态系统, 土壤微生物生物量在植物生长季降低, 秋季逐渐升高, 呈现“W”形(Brookes et al., 1990; Lipson et al., 1999; 刘爽和王传宽, 2010); 在亚热带常绿阔叶林和热带季雨林, 土壤微生物生物量在夏季比冬季高(Devi & Yadava, 2006; Feng et al., 2009); 在北亚热带次生栎林和火炬松(Pinus taeda)人工林土壤微生物生物量在植物生长旺季维持在较低水平, 而在植物休眠季节维持在较高水平(王国兵等, 2008).在热带雨林, 土壤微生物生物量可能因为植物与土壤微生物之间剧烈的养分竞争而使土壤微生物量在树木生长旺盛时期较低(Barbhuiya et al., 2004).在湿润的北极苔原, 由于根系与土壤微生物之间的养分竞争, 使得土壤微生物生物量在春季冻融早期达到最大值, 随着温度回升而减少(Edwards et al., 2006).由此可见, 不同气候条件下的生态系统, 由于影响土壤微生物生物量的主导因子不同而使其季节变化格局产生差异. ...
Elevated atmospheric CO2 and feedback between carbon and nitrogen cycles 1 1993
... 本研究发现土壤微生物生物量的季节波动与植物根系对营养的竞争以及林木的生长节律有关, 温带森林和亚热带森林的土壤微生物量的季节变化格局基本都呈现非生长季维持在较高水平, 而随着植物进入生长季而下降.这与其他一些温带森林的研究结果 (Brookes et al., 1990; Lipson et al., 1999; 王国兵等, 2008; 刘爽和王传宽, 2010; 王薪琪等, 2017)一致.可能的解释为: 秋季大量凋落物的输入为冬季土壤微生物提供了丰富的代谢底物.尽管冬季气温很低, 但处于积雪覆盖下的土壤温度基本能够维持土壤微生物生长和繁殖.另外, 由于冬季低温, 土壤微生物代谢较弱, 其维持呼吸所需要的能量较低, 在较低能量供给条件下就能维持较高的土壤微生物量(Brookes et al., 1990), 因此冬季土壤微生物量均较高.随着气温的升高进入春季, 早春植物(一些短命草本)和土壤微生物共同固持养分可能形成“春坝” (vernal dam)作用(Warren和邹晓明, 2003), 为植物根系和土壤微生物的快速生长提供养分, 从而使土壤微生物生物量呈现上升趋势.随着根系的恢复生长, 植物进入旺盛生长季(夏季), 土壤微生物与植物之间对养分的激烈竞争, 使土壤微生物生物量呈现下降趋势(Kaiser et al., 2011).由此可见, 在温带和亚热带森林生态系统中, 植物和土壤微生物对养分的不同步吸收是保留养分和维持生态系统生产力的一种机制(Muller & Herbert Bormann, 1976; Singh et al., 1989; Groffman et al., 1993; Zak et al., 1993; Kaiser et al., 2014). ...
Elevated atmospheric CO2 fine roots and the response of soil microorganisms: A review and hypothesis 4 2000
... 在陆地生态系统中, 植物是第一生产者, 土壤微生物是有机质的分解者.植物将光合产物以根系分泌物和植物残体(凋落物)的形式释放到土壤, 供给土壤微生物碳源(Zak et al., 2000); 而土壤微生物一方面通过自身分解和代谢将有机养分转化成无机养分, 以利于植物吸收利用(Schimel & Weintraub, 2003; Fang et al., 2005; Neill & Gignoux, 2006); 另一方面通过与根系的共生关系(菌根真菌)直接利用土壤里的小分子有机物质促进植物生长(Deng et al., 2018).这种植物-土壤微生物的相互作用维系着陆地生态系统的生态功能.尽管如此, 植物根系和土壤微生物之间存在着既相互依存又相互制约的关系(图1): 植物为土壤微生物的生长提供C源从而促进土壤微生物的活动, 而土壤微生物矿化有机养分提高植物对养分利用的有效性; 但是土壤微生物也同植物根系争夺营养元素, 它们对土壤中营养的利用属于竞争关系(Singh et al., 1989; Harte & Kinzig, 1993; Kaye & Hart, 1997; Terrer et al., 2018).这种植物和土壤微生物呈现的复杂的互利-竞争关系存在于季节交替过程中, 植物输入土壤中的凋落物在数量和质量上的改变, 土壤可利用营养在充足和缺乏之间变化, 加之植物在生长季和休眠季对养分需求的不同, 就有可能改变土壤微生物对N的需求(即矿化-同化途径), 限制植物对无机N的吸收(Zak et al., 2000; Terrer et al., 2018), 而从对养分的矿化转为对养分的同化, 增加自身的生物量, 从而在土壤微生物生物量上可能呈现一个季节波动特征. ...
... ).这种植物和土壤微生物呈现的复杂的互利-竞争关系存在于季节交替过程中, 植物输入土壤中的凋落物在数量和质量上的改变, 土壤可利用营养在充足和缺乏之间变化, 加之植物在生长季和休眠季对养分需求的不同, 就有可能改变土壤微生物对N的需求(即矿化-同化途径), 限制植物对无机N的吸收(Zak et al., 2000; Terrer et al., 2018), 而从对养分的矿化转为对养分的同化, 增加自身的生物量, 从而在土壤微生物生物量上可能呈现一个季节波动特征. ...
... 陆地生态系统中植物和土壤微生物活动相互关联的概念模型.一方面植物凋落物为土壤微生物生长活动提供底物, 同时土壤微生物分解底物产生的无机氮供植物生长利用, 它们之间是互利共生的.另一方面, 当养分供应(特别是氮)缺乏时, 它们之间又转换为竞争关系.改绘自Zak等(2000). A conceptual model illustrating the links between plant and soil microbial activity in terrestrial ecosystems<i>. </i>On one hand, plants and microbes are mutualistic, since plant residues constitute the main substrate for microbial biomass that in turn produces inorganic nitrogen used by plants. On the other hand, their interaction is in competition for soil nutrients, N in particular. The competitive interaction is regulated by the relative distribution of roots and microbes in the soil, but it also depends on the pathway followed by the mineralized N. Redraw based on <xref ref-type="bibr" rid="b43">Zak <i>et al</i>. (2000)</xref>.Fig. 1 土壤微生物生物量的季节性动态变化是一个复杂过程, 其影响因子主要有土壤C含量和养分资源的限制(Landgraf & Klose, 2002; Koranda et al., 2013)、植物种类、土壤湿度以及土壤温度(杨凯等, 2009; López-Mondéjar et al., 2015)等.不同生态系统土壤微生物的季节变化呈现不同的格局(Devi & Yadava, 2006; Edwards et al., 2006).在温带森林生态系统, 土壤微生物生物量在植物生长季降低, 秋季逐渐升高, 呈现“W”形(Brookes et al., 1990; Lipson et al., 1999; 刘爽和王传宽, 2010); 在亚热带常绿阔叶林和热带季雨林, 土壤微生物生物量在夏季比冬季高(Devi & Yadava, 2006; Feng et al., 2009); 在北亚热带次生栎林和火炬松(Pinus taeda)人工林土壤微生物生物量在植物生长旺季维持在较低水平, 而在植物休眠季节维持在较高水平(王国兵等, 2008).在热带雨林, 土壤微生物生物量可能因为植物与土壤微生物之间剧烈的养分竞争而使土壤微生物量在树木生长旺盛时期较低(Barbhuiya et al., 2004).在湿润的北极苔原, 由于根系与土壤微生物之间的养分竞争, 使得土壤微生物生物量在春季冻融早期达到最大值, 随着温度回升而减少(Edwards et al., 2006).由此可见, 不同气候条件下的生态系统, 由于影响土壤微生物生物量的主导因子不同而使其季节变化格局产生差异. ...
... On one hand, plants and microbes are mutualistic, since plant residues constitute the main substrate for microbial biomass that in turn produces inorganic nitrogen used by plants. On the other hand, their interaction is in competition for soil nutrients, N in particular. The competitive interaction is regulated by the relative distribution of roots and microbes in the soil, but it also depends on the pathway followed by the mineralized N. Redraw based on Zak et al. (2000). Fig. 1 土壤微生物生物量的季节性动态变化是一个复杂过程, 其影响因子主要有土壤C含量和养分资源的限制(Landgraf & Klose, 2002; Koranda et al., 2013)、植物种类、土壤湿度以及土壤温度(杨凯等, 2009; López-Mondéjar et al., 2015)等.不同生态系统土壤微生物的季节变化呈现不同的格局(Devi & Yadava, 2006; Edwards et al., 2006).在温带森林生态系统, 土壤微生物生物量在植物生长季降低, 秋季逐渐升高, 呈现“W”形(Brookes et al., 1990; Lipson et al., 1999; 刘爽和王传宽, 2010); 在亚热带常绿阔叶林和热带季雨林, 土壤微生物生物量在夏季比冬季高(Devi & Yadava, 2006; Feng et al., 2009); 在北亚热带次生栎林和火炬松(Pinus taeda)人工林土壤微生物生物量在植物生长旺季维持在较低水平, 而在植物休眠季节维持在较高水平(王国兵等, 2008).在热带雨林, 土壤微生物生物量可能因为植物与土壤微生物之间剧烈的养分竞争而使土壤微生物量在树木生长旺盛时期较低(Barbhuiya et al., 2004).在湿润的北极苔原, 由于根系与土壤微生物之间的养分竞争, 使得土壤微生物生物量在春季冻融早期达到最大值, 随着温度回升而减少(Edwards et al., 2006).由此可见, 不同气候条件下的生态系统, 由于影响土壤微生物生物量的主导因子不同而使其季节变化格局产生差异. ...
 ,
, ,1,*, 木勒德尔?吐尔汗拜2, 田地2, 冯兆忠1
,1,*, 木勒德尔?吐尔汗拜2, 田地2, 冯兆忠1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT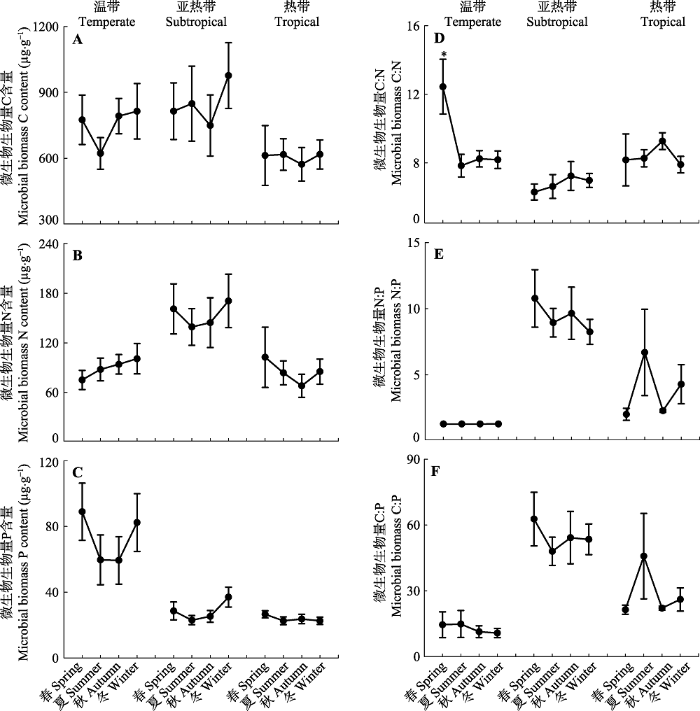 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT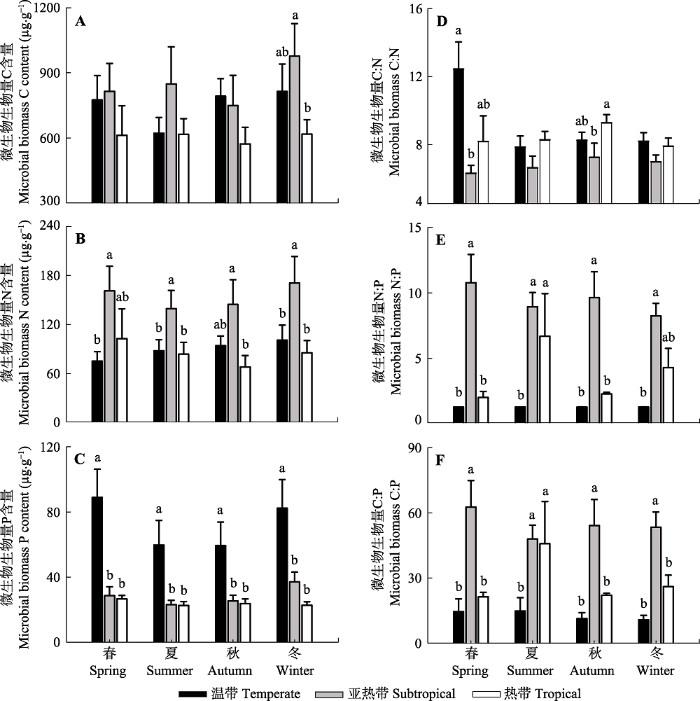 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT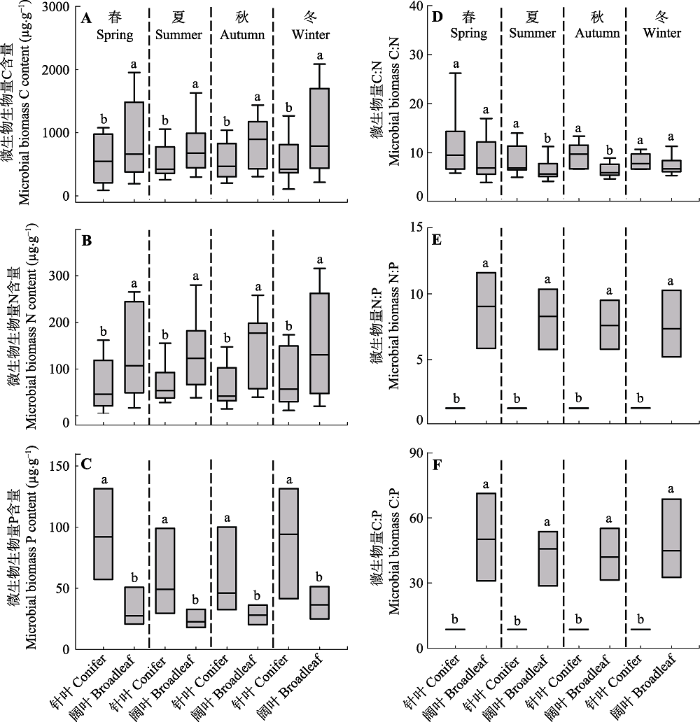 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}