 ,1,*
,1,*Patterns and affecting factors of nitrogen use efficiency of plant leaves and roots in Nei Mongol and Qinghai-Xizang Plateau grasslands
Yi-Wen FU1,2, Da-Shuan TIAN2, Jin-Song WANG2, Shu-Li NIU2,3, Ken-Tian ZHAO,1,*通讯作者: *zhaokt@sina.com
编委: 吕晓涛
责任编辑: 李敏, 赵航(实习)
收稿日期:2019-02-28接受日期:2019-07-10网络出版日期:2019-07-20
| 基金资助: |
Received:2019-02-28Accepted:2019-07-10Online:2019-07-20
| Fund supported: |

摘要
氮利用效率是植物的关键功能性状, 同时紧密关联生态系统功能, 但是目前对氮利用效率的区域格局及影响因素仍然不清楚。该研究分析了内蒙古和青藏高原草原82个调查地点、139种植物叶片和根系的氮利用效率及其与环境因素、植物功能群之间的关系, 实验结果显示: 1)草甸草原植物叶片的氮利用效率为53 g·g -1, 显著大于高寒草甸(46 g·g -1)、荒漠草原(41 g·g -1)和典型草原(39 g·g -1)。高寒草甸根系氮利用效率为108 g·g -1, 显著高于其他生态系统。2)叶片氮利用效率比根系对温度更加敏感, 但随着干旱指数的增加, 两者均表现出显著的降低趋势。3)杂类草叶片和根系氮利用效率低于莎草科和禾本科植物, 豆科植物叶片和根系氮利用效率分别比非豆科植物低48%和60%。4)植物氮利用效率与土壤氮含量之间没有显著关系。总体上, 内蒙古和青藏高原草原植物叶片和根系氮利用效率的空间格局存在差异, 主要影响因素为植物功能群和干旱指数。本研究系统揭示内蒙古和青藏高原草原植物氮利用效率的空间格局及关键驱动因子, 有助于在全球变化背景下了解我国草地生产力维持机制, 同时为草原生态系统管理提供科学依据。
关键词:
Abstract
Aims Nitrogen use efficiency (NUE) is a key functional trait in plants, which closely relates to ecosystem functions. However, it is still unclear about the regional patterns and affecting factors of plant NUE.
Methods This study quantified leaf and root NUE in 139 grassland plant species and explored their relationships with environmental factors and plant functional groups across 82 sampling sites in Nei Mongol and Qinghai-?Xizang Plateau.
Important findings 1) We found that leaf NUE (53 g·g -1) in meadow steppe was significantly greater than those in alpine meadow (46 g·g -1), desert steppe (41 g·g -1) and typical steppe (39 g·g -1). Root NUE (108 g·g -1) in alpine meadow was higher than those in other ecosystems. 2) Leaf NUE was more sensitive to temperature than root NUE, but with increasing drought index they all showed a significant decrease. 3) Leaf and root NUE in forbs were significantly lower than sedges and grasses. In addition, leaf and root NUE of legume were 48% and 60% lower than those of non-legume, respectively. 4) Plant NUE did not show any significant relationship with soil nitrogen content. Overall, there was significant difference between leaf and root NUE in their spatial patterns in the Nei Mongol and Qinghai-Xizang Plateau grasslands. The main impacting factors were plant functional group and drought index. The findings are helpful for better understanding the mechanisms underlying the variation of grassland productivity in China, and also provide more scientific basis for grassland management.
Keywords:
PDF (1454KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
符义稳, 田大栓, 汪金松, 牛书丽, 赵垦田. 内蒙古和青藏高原草原植物叶片与根系氮利用效率空间格局及影响因素. 植物生态学报, 2019, 43(7): 566-575. DOI: 10.17521/cjpe.2019.0044
FU Yi-Wen, TIAN Da-Shuan, WANG Jin-Song, NIU Shu-Li, ZHAO Ken-Tian.
氮是植物生长所必需的主要营养元素(Vermeer & Berendse, 1983), 而草原生态系统普遍受到氮限制(Vitousek & Howarth, 1991; Elser et al., 1996)。因而, 氮利用效率高的草原植物有利于适应氮限制生境, 同时对维持草原生态系统生产力具有重要作用(Li et al., 2010)。通常, 植物氮利用效率(NUE)定义为植物吸收单位氮所能生产的生物量(Birk & Vitousek, 1986; Berendse & Aerts, 1987; Hiremath & Ewel, 2001)。尽管前人对植物NUE已经进行了大量研究, 但大部分研究是基于植物叶片NUE (Field et al., 1983; Tian et al., 2017), 缺乏区域尺度上系统比较植物叶片与根系NUE空间格局差异及其主要影响因素的研究。
草原植物NUE主要受气候条件、土壤氮含量和植物功能群的共同影响。降水是限制草地生态系统生产力的主要因素, 尤其是干旱和半干旱草原(Le Houerou, 1984; Bai et al., 2008; Hu et al., 2010)。例如, McCulley等(2009)发现, 草原植物NUE随水分供应增加而增加, 主要由于水分添加缓解了植物水分胁迫, 促进植物生长进而提高NUE。此外, 高土壤含水量有利于把根际外土壤氮运输至根系表面, 便于根系吸收从而提高根系NUE (Cramer et al., 2009)。温度对植物NUE也具有重要调节作用。如在高寒草原生态系统中, 植物生长和温度变化密切相关(An et al., 2005; Reich, 2009; Niu et al., 2010; Norby et al., 2010)。高寒草原植物生命周期普遍较短, 温度升高会延长植物物候期和叶片寿命, 促进植物从老叶中回收养分, 从而促进植物生长和提高植物NUE (Chu et al., 2008; Yu et al., 2010)。
土壤氮含量是影响草原植物NUE的重要因素。植物主要通过根系从土壤中获取无机氮, 用于维持自身的生长发育(Xu et al., 2012)。然而, 土壤中90%以上的氮都是有机氮, 而大部分有机氮只有通过微生物矿化之后才能被根系吸收和利用(Burton et al., 2007; 王学霞等, 2018)。前人研究表明, 在水分适宜的条件下, 较高的土壤有效氮含量会促进草本植物NUE, 但是当有效氮含量超过一定阈值, NUE反而下降(Silver, 1994; Vázquez de Aldana & Berendse, 1997)。因此, 土壤氮含量与植物NUE之间的关系比较复杂。
草原植物NUE在不同功能群之间可能存在差异。相关研究表明, 不同植物功能群的氮利用策略不同, 如在氮吸收特点、氮在植物体内运输途径、分配状况以及氮再吸收等方面存在差异, 这些差异可能导致功能群之间NUE的不同(苏波等, 2000; Han et al., 2012)。例如, 豆科和非豆科植物NUE明显不同, 主要因为豆科植物根系与固氮微生物形成根瘤, 将大气中N2还原为氨, 进而为自身提供氮。与非豆科植物相比, 豆科植物无需最大限度地从土壤中获取氮和最大化自身NUE, 所以豆科植物NUE明显低于非豆科植物(Reed, 2017)。此外, 前人单点实验发现草原生态系统中莎草科与禾本科植物NUE没有差异, 但是它们与杂类草NUE有明显差异(Vázquez de Aldana & Berendse, 1997)。因此, 由于不同草原生态系统类型之间在气候条件、土壤氮含量以及植物功能群等方面的差异, 导致草原生态系统之间NUE可能也存在差异。
草原是全球陆地生态系统的重要组成部分。中国草原约占国土面积的40% (Wang & Wesche, 2016), 而青藏高原和内蒙古草原占中国草地面积的55% (Zhang et al., 2014; Briske et al., 2015)。因此, 本研究以内蒙古和青藏高原草原生态系统为研究对象, 旨在揭示植物NUE的区域格局, 并进一步解析草原植物NUE如何受气候、植物功能群、土壤氮含量和生态类型的影响。拟解决的具体科学问题如下: 1)内蒙古和青藏高原草原植物叶片和根系NUE的空间格局有何不同?2)影响草原植物NUE的主要因素是什么?
1 材料和方法
1.1 数据获取
本研究数据库来自于前人发表的数据文章(Geng et al., 2017)。迄今为止, 该数据库系统测定内蒙古和青藏高原草原植物叶片和根系的养分浓度及相关性状, 但是仍未解析和比较草原植物叶片和根系NUE的空间格局及影响因素。同时, 该数据库的优势还在于系统匹配地调查了草原植物叶片和根系性状, 并采用统一标准进行野外取样和室内样品处理和化学指标分析。数据库主要包括内蒙古和青藏高原草原生态系统, 植被类型具体包括荒漠草原、典型草原、草甸草原和高寒草甸。地理区域跨度90.8°-120.12° E, 30.31°-50.19° N, 海拔553-5 105 m, 共包含82个调查地点和139个植物种。样地年平均气温为-5.8-4.1 ℃, 年降水量148-604 mm, 年蒸发量300-608 mm。每个调查地点的年平均气温和年降水量是通过研究区680个气象监测站过去55年(1951-2005)记录数据, 用经度、纬度和海拔高度作为预测因素, 通过线性模型分析得到。水分蒸发散是通过水分平衡公式(Thornthwaite, 1948)计算得到。植物功能群主要包括禾草类(优势种为针茅属(Stipa))植物, 莎草类(优势种为嵩草属(Kobresia))植物, 以及杂类草(优势种为葱属(Allium)和委陵菜属(Potentilla))植物。植物叶片和根系样品在野外经过处理后, 在实验室内烘干至恒质量, 统一使用球磨仪(NM200, Retsch, Haan, Germany)粉碎。粉碎后的样品采用元素分析仪(2400 II CHN, Perkin-Elmer, Massachusetts, USA)测定氮含量。同时对每个地点0-10 cm土壤样品进行预处理, 新鲜土样过2 mm筛, 并将细根挑干净后分成两份。一份土样用KCl溶液浸提, 浸提液使用连续流动分析仪(SAN Plus, Skalar, Netherlands)测定土壤硝态氮和铵态氮含量; 另一份土样在室内风干之后, 使用球磨仪进行粉碎, 然后采用凯氏定氮仪(KjelFlex-360, BUCHI, Flawil, Switzerland)测定土壤全氮含量。相关数据的测定方法详见Geng等(2017)文中介绍。1.2 数据分析
植物NUE计算一般采用Chapin III和Moilanen (1991)提出的Chapin指数, 通过计算植物生物量与植物养分贮量之比来表征。因而, NUE等同于植物体养分浓度的倒数。NUE (g·g-1)的计算公式为1/E, E = Ap/m = m × Cp/m = Cp; 式中E为Chapin指数, m为植物生物量(kg·hm-2), Ap为植物养分贮量(kg·hm-2), Cp为植物体的养分含量(%)。统计分析首先采用单因素方差分析, 比较植物叶片和根系NUE受不同草地类型、干旱指数(r = E0/P, E0为年蒸发能力, P为年降水量; 根据草原干湿气候带标准, r < 1为湿润带, 1.0-1.6为半湿润带, 1.6-3.5为半干旱带)、植物功能群(豆科与非豆科植物; 杂类草、禾本科与莎草科)的影响。然后, 进一步利用线性回归, 分析植物叶片或根系NUE与年平均气温、年降水量、干旱指数、土壤全氮或有效氮含量之间的定量关系。上述统计分析均采用SPSS 22.0软件完成, 并使用SigmaPlot 12.5软件制图。最后, 基于SPSS Amos 21.0软件, 利用结构方程模型综合分析生物和非生物因素对植物叶片或根系NUE影响效应的方向和大小。由于生物因素植物功能群(杂类草、禾本科、莎草科)为分类变量, 我们参考前人研究(Zhang & Chen, 2015; Wang et al., 2019), 将杂类草、禾本科、莎草科功能群半定量为1、2、3代入模型分析, 并利用Microsoft Visio软件作图。2 结果和分析
基于对内蒙古和青藏高原调查数据的整合分析, 结果显示内蒙古和青藏高原草原植物叶片平均NUE为44 g·g-1, 显著低于根系NUE (100 g·g-1)(图1), 叶片NUE约为根系的44%。而在不同生态系统类型之间, 草甸草原(53 g·g-1)和高寒草甸(46 g·g-1)植物叶片NUE显著大于荒漠草原(41 g·g-1)和典型草原(39 g·g-1)。但是, 根系NUE在不同生态系统之间却表现出不同格局, 具体表现为高寒草甸根系NUE最高(108 g·g-1), 其他生态系统类型之间根系NUE没有显著差异。叶片和根系NUE对气候因子表现出不同的响应格局。例如, 随着年平均气温升高, 叶片NUE呈显著降低趋势, 斜率为-0.001 3。而根系NUE与年平均气温没有显著关系。沿着年降水量梯度, 叶片或根系NUE均没有显著变化趋势(图2)。但是, 随着干旱指数的增加, 叶片和根系NUE均表现出显著降低趋势(图3)。叶片NUE与干旱指数关系的斜率为-0.005 1, 低于根系NUE与干旱指数关系的斜率-0.012 8。图1
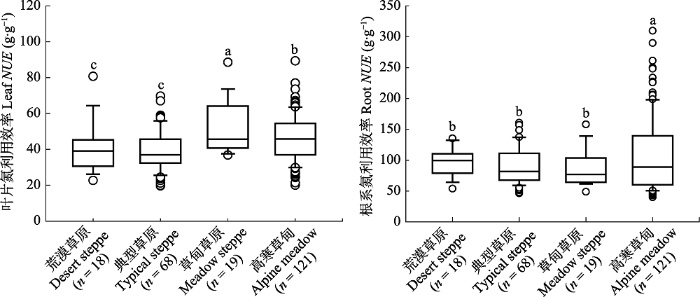 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1沿水分梯度不同草原生态系统类型植物叶片和根系氮利用效率。不同小写字母表示差异显著(p < 0.05), n代表样本量。
Fig. 1Differences of leaf and root nitrogen use efficiency (NUE) among different grassland ecosystems along a precipitation gradient. Different lowercase letters indicate a significant difference (p < 0.05), and n represents the sample size of observations.
图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2年平均气温和年降水量与草原植物叶片或根系氮利用效率之间的关系。n表示样本量。
Fig. 2Relationships of leaf or root nitrogen use efficiency (NUE) with mean annual air temperature and mean annual precipitation. n represents the sample size of observations.
图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3干旱指数与草原植物叶片或根系氮利用效率之间的关系。不同小写字母表示差异显著(p < 0.05), n表示样本量。
Fig. 3Relationships between drought index and leaf or root nitrogen use efficiency (NUE) in grassland plants. Different lowercase letters indicate a significant difference (p < 0.05), and n represents the sample size of observations.
在不同植物功能群之间, 草原植物叶片和根系NUE的响应规律一致, 均表现出莎草科植物NUE最高, 而杂类草NUE最低(图4)。例如, 莎草科、禾本科植物和杂类草的叶片NUE分别为50 g·g-1、49 g·g-1和42 g·g-1。莎草科、禾本科和杂类草植物根系NUE分别为181 g·g-1、114 g·g-1和85 g·g-1, 莎草科植物与杂类草根系NUE相差213%。非豆科植物叶片和根系NUE分别为46 g·g-1和104 g·g-1, 显著高于豆科植物48%和60%。但是, 土壤有效氮或全氮含量与草原植物NUE均没有显著关系(图5)。最后, 进一步基于结构方程模型综合分析, 我们发现植物功能群和干旱指数是影响草原植物叶片和根系NUE的主要因素。例如, 植物功能群影响叶片和根系NUE的路径系数分别为0.30和0.54, 干旱指数作用于叶片和根系NUE的路径系数分别为-0.19和-0.17, 而其他因素的效应不显著(图6)。
图4
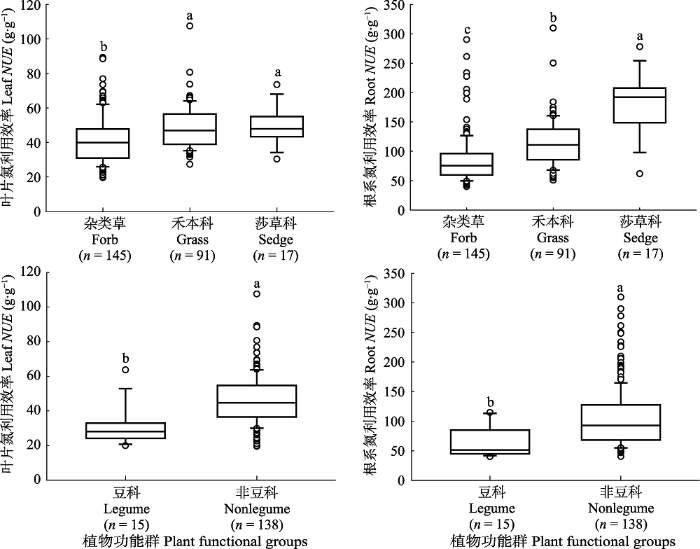 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4不同草原植物功能群的叶片和根系氮利用效率。不同小写字母表示差异显著(p < 0.05), n表示样本量。
Fig. 4Leaf and root nitrogen use efficiency (NUE) among different grassland plant functional groups. Different lowercase letters indicate a significant difference (p < 0.05), and n represents the sample size of observations.
图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5土壤全氮或有效氮含量与草原植物叶片或根系氮利用效率之间的关系。n表示样本量。
Fig. 5Relationships of grassland plant leaf or root nitrogen use efficiency (NUE) with soil total nitrogen or available nitrogen content. n represents the sample size of observations.
图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6生物(植物功能群)和非生物因素(年平均气温、年降水量和干旱指数)对植物叶片或根系氮利用效率(NUE)影响的直接和间接效应。图中线条粗细表示影响效应大小, 实线表示作用路径显著(p < 0.05), 虚线表示作用路径不显著(p > 0.05)。绿色表示负效应, 红色表示正效应。R2表示解释率。
Fig. 6Direct and indirect impacts of biotic (plant functional group (PFG)) and abiotic (mean annual air temperature (MAT), mean annual precipition (MAP) and drought index (r)) factors on leaf or root nitrogen use efficiency (NUE) in grassland plants. Line thickness indicates relative effect size. Solid lines denote a significant influence (p < 0.05), whereas dashed lines indicate no significant impact (p > 0.05). Green color represents a negative effect, while red color denotes a positive impact. R2 indicates how much variation can be explained.
3 讨 论
3.1 气候条件对植物叶片和根系NUE的影响
本研究结果发现植物叶片或根系NUE与年降水量之间没有显著关系, 这与前人的研究结果不一致。例如, McCulley等(2009)对北美中部草原植物叶片NUE的研究发现, 叶片NUE随年降水量增加而增加。同样, 吕晓涛等(2010)在内蒙古典型草原的增雨实验发现, 植物NUE随降雨量增加而增加。但是, 白春利(2013)在荒漠草原的研究发现, 短花针茅(Stipa breviflora)和无芒隐子草(Cleistogenes songorica)叶片NUE与降水量的关系是非线性的, 即植物NUE随降水量增加而增加, 但是当降水量超过一定阈值之后会导致植物NUE降低。尽管我们没有发现降水对草原植物NUE的显著影响, 但是结果发现高干旱指数会显著抑制植物NUE, 这可能是因为草原地区生态系统蒸发量大, 导致蒸发量主要决定土壤含水量, 而非降水量。我们的研究揭示湿润环境(低干旱指数)促进植物NUE, 可能包含几方面原因。第一, 高土壤含水量缓解植物水分限制(Yuan et al., 2006); 第二, 高土壤含水量有利于增强土壤氮运输至植物根际表面, 便于根系氮吸收(Steenvoorden, 1988); 第三, 高土壤水分倾向于延长氮在植物体内的滞留时间, 并提高植物从老叶中重吸收氮的效率, 进而增加植物NUE (Gong et al., 2011)。我们发现随着气温升高, 草原植物叶片NUE显著降低。虽然未见区域尺度上植物NUE与温度之间关系的研究报道, 但Yu等(2017)分析中国草地132个地点的植物叶片氮浓度发现, 叶片氮浓度随气温升高而降低。尽管植物氮浓度与氮利用效率之间存在着非线性(倒数函数)关系, 并非一一对应的线性关系, 但是Yu等(2017)的研究结果可能部分支持叶片NUE随气温升高而升高的观点, 本研究发现的随气温升高植物NUE呈降低趋势的结果与之不相符。这些结果之间的差异可能是由高温和低温草原地区不同植物功能群分布造成的。在低温的青藏高原地区, 主要优势物种是莎草科和禾本科植物。而我们的结果显示这两种植物功能群的NUE均比较高(图5)。此外, 我们发现草原植物NUE与土壤全氮或有效氮含量之间关系不显著, 这与以往研究发现草原植物NUE随土壤氮含量的增加而降低(Aerts & de Caluwe, 1994; de Aldana & Berendse, 1997; Yuan et al., 2006)的结果不一致。究其原因, 可能包括三方面。第一, 近年来随着氮沉降的不断累积, 草原生态系统氮限制程度减弱(杨晓霞等, 2014); 第二, 部分研究是基于室内氮添加实验, 而室内实验条件通常比较理想, 容易检测出氮添加效应。例如, Aerts和de Caluwe (1994)、de Aldana和Berendse (1997)分别在室内研究3种草原薹草植物和6种草原多年生草本植物NUE, 均发现氮添加降低植物NUE; 第三, 部分研究是基于较小空间尺度上的自然氮梯度实验。例如, Yuan等(2006)在内蒙古多伦县沿着差异明显的自然氮梯度, 人为选择20个地点分析两种针茅属植物NUE与土壤氮含量之间的关系, 发现植物NUE随土壤氮含量增加而降低。前人研究集中在局部区域尺度上, 年降水量相对较高(385 mm)且变异不大, 加之人为选择土壤氮梯度, 容易发现植物NUE与土壤氮之间的关系。但是, 本研究覆盖了内蒙古和青藏高原草原, 研究区域尺度和降水变化(148-604 mm)均比较大, 导致土壤水分成为内蒙古和青藏高原草原植物NUE的主要影响因素, 而非土壤氮含量。
3.2 草原植物功能群之间氮利用效率的差异
从草原植物不同功能群NUE来看, 莎草科和禾草科植物的NUE均高于杂类草, 这一结果部分地与前人研究结果一致。例如, He等(2008)对中国草地199个地点共计213个植物种叶片养分的研究得出, 杂类草叶片氮含量高于莎草科和禾本科植物, 可能指示出莎草科和禾草科植物NUE大于杂类草。具体原因可能如下: 第一, 不同植物功能群适应生境的策略不同(Xu et al., 2015)。莎草科和禾本科植物多以资源保守型物种为主, 倾向于高效利用资源以适应氮缺乏环境, 即提高根系和叶片吸收单位氮产生的生物量(Liu et al., 2018)。而杂类草是资源利用型植物, 倾向于在资源丰富的环境中增强竞争能力, 而适应贫瘠环境的能力较弱, 即养分利用效率低(Aerts & Chapin III, 1999; An et al., 2005)。第二, 莎草科和禾草科植物比叶面积均大于杂类草(杜占池等, 2001)。而前人研究发现, 植物比叶面积与光合能力正相关, 从而促进莎草科和禾草科植物的生长和NUE (Zuo et al., 2016)。同时, 我们还发现豆科植物NUE显著低于非豆科植物, 可能主要由豆科与非豆科植物在养分利用策略和环境适应性方面的差异所导致(彭昀月等, 2017)。第一, 从植物利用叶氮来看, 非豆科植物比豆科植物在光合系统内部具有更高效的氮分配机制, 如非豆科植物将更多叶氮分配到羧化系统中, 使非豆科植物具有更高效的氮利用机制(Poorter & Evans, 1998); 第二, 从植物环境适应性方面来看, 非豆科植物净光合速率和比叶面积均高于豆科植物, 而高净光合速率和比叶面积会促进植物生长和NUE (朱军涛等, 2010); 第三, 与植物自身生理特性有关。豆科植物与固氮微生物共生, 通过形成根瘤菌将大气中的氮气还原为氨, 进而为自身提供氮, 即豆科植物获取氮比非豆科植物更加容易。相比而言, 非豆科植物在相同氮养分条件下需通过提高自身NUE来满足生长过程所需要的氮养分(Shi et al., 2017)。
3.3 草原生态系统之间植物氮利用效率的差异
研究结果显示草甸草原植物叶片NUE显著高于荒漠草原、典型草原和高寒草甸, 而高寒草甸根系NUE显著高于其他生态系统。本研究中草甸草原年降水量为387 mm, 年平均气温-1.4 ℃, 干旱指数1.31, 优势种为禾本科植物。高寒草甸年降水量450 mm, 年均气温-2.4 ℃, 干旱指数0.87, 优势种为莎草科植物。进一步通过结构方程模型分析发现, 植物功能群对NUE影响最大, 干旱指数次之。这说明造成草原生态系统之间NUE的差异主要由植物功能群和气候条件共同决定。本研究中高寒草甸和草甸草原优势植物NUE均比较高, 并且它们的干旱指数低于其他生态系统。低干旱指数(高土壤含水量)有利于微生物对土壤有机质的分解和氨化, 从而提高土壤氮矿化速率(Patil et al., 2010)。同时, 高土壤含水量有利于将土壤氮运输至植物根际表面(Steenvoorden, 1988; Yuan et al., 2006), 进而增强植物氮吸收和利用。上述这些因素的共同作用促进草甸草原和高寒草甸植物NUE。4 结论
本研究首次从区域尺度上揭示内蒙古和青藏高原草原植物叶片和根系NUE空间格局的异同, 而传统研究主要关注植物叶片NUE。我们主要发现草原植物叶片NUE明显低于根系, 不同生态系统之间根系和叶片NUE的空间格局存在差异, 主要影响因素为植物功能群和干旱指数。沿着干旱指数梯度或不同植物功能群之间, 植物叶片和根系NUE均表现出类似格局。而植物NUE受土壤氮含量的影响较小。由于草原地区普遍受到氮限制, 并且NUE是反映植物适应氮限制环境的重要策略, 因此本研究有助于了解草原生态系统生产力的维持机制。同时, 在全球氮富集背景下, 植物NUE与土壤氮循环紧密关联(如氮淋溶等), 对我国环境保护具有积极的意义。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
DOI:10.2307/1940890URL [本文引用: 2]
DOI:10.1111/gcb.2005.11.issue-10URL [本文引用: 2]
[本文引用: 1]
[本文引用: 1]
DOI:10.1890/07-0992.1URLPMID:18724724 [本文引用: 1]

Understanding how the aboveground net primary production (ANPP) of arid and semiarid ecosystems of the world responds to variations in precipitation is crucial for assessing the impacts of climate change on terrestrial ecosystems. Rain-use efficiency (RUE) is an important measure for acquiring this understanding. However, little is known about the response pattern of RUE for the largest contiguous natural grassland region of the world, the Eurasian Steppe. Here we investigated the spatial and temporal patterns of ANPP and RUE and their key driving factors based on a long-term data set from 21 natural arid and semiarid ecosystem sites across the Inner Mongolia steppe region in northern China. Our results showed that, with increasing mean annual precipitation (MAP), (1) ANPP increased while the interannual variability of ANPP declined, (2) plant species richness increased and the relative abundance of key functional groups shifted predictably, and (3) RUE increased in space across different ecosystems but decreased with increasing annual precipitation within a given ecosystem. These results clearly indicate that the patterns of both ANPP and RUE are scale dependent, and the seemingly conflicting patterns of RUE in space vs. time suggest distinctive underlying mechanisms, involving interactions among precipitation, soil N, and biotic factors. Also, while our results supported the existence of a common maximum RUE, they also indicated that its value could be substantially increased by altering resource availability, such as adding nitrogen. Our findings have important implications for understanding and predicting ecological impacts of global climate change and for management practices in arid and semiarid ecosystems in the Inner Mongolia steppe region and beyond.
[本文引用: 1]
DOI:10.2307/1938504URL [本文引用: 1]
DOI:10.1016/j.jenvman.2014.07.036URLPMID:25687702 [本文引用: 1]
Semi-nomadic pastoralism was replaced by sedentary pastoralism in Inner Mongolia during the 1960's in response to changes in land use policy and increasing human population. Large increases in numbers of livestock and pastoralist households (11- and 9-fold, respectively) during the past 60?yrs have variously degraded the majority of grasslands in Inner Mongolia (78?M ha) and jeopardize the livelihoods of 24?M human inhabitants. A prevailing strategy for alleviating poverty and grassland degradation emphasizes intensification of livestock production systems to maintain both pastoral livelihoods and large livestock numbers. We consider this strategy unsustainable because maximization of livestock revenue incurs high supplemental feed costs, marginalizes net household income, and promotes larger flock sizes to create a positive feedback loop driving grassland degradation. We offer an alternative strategy that increases both livestock production efficiency and net pastoral income by marketing high quality animal products to an increasing affluent Chinese economy while simultaneously reducing livestock impacts on grasslands. We further caution that this strategy be designed and assessed within a social-ecological framework capable of coordinating market expansion for livestock products, sustainable livestock carrying capacities, modified pastoral perceptions of success, and incentives for ecosystem services to interrupt the positive feedback loop that exists between subsistence pastoralism and grassland degradation in Inner Mongolia.
DOI:10.1111/gcb.14962URLPMID:31838767 [本文引用: 1]
Numerous studies have demonstrated that fertilization with nutrients such as nitrogen, phosphorus, and potassium increase plant productivity in both natural and managed ecosystems, demonstrating that primary productivity is nutrient limited in most terrestrial ecosystems. In contrast, it has been demonstrated that heterotrophic microbial communities in soil are primarily limited by organic carbon or energy. While this concept of contrasting limitations, i.e., microbial carbon and plant nutrient limitation, is based on strong evidence that we review in this paper, it is often ignored in discussions of ecosystem response to global environment changes. The plant-centric perspective has equated plant-nutrient limitations with those of whole ecosystems, thereby ignoring the important role of the heterotrophs responsible for soil decomposition in driving ecosystem carbon storage. In order to truly integrate carbon and nutrient cycles in ecosystem science, we must account for the fact that while plant productivity may be nutrient- limited, the secondary productivity by heterotrophic communities is inherently carbon-limited. Ecosystem carbon cycling integrates the independent physiological responses of its individual components, as well as tightly coupled exchanges between autotrophs and heterotrophs. To the extent that the interacting autotrophic and heterotrophic processes are controlled by organisms that are limited by nutrient versus carbon accessibility, respectively, we propose that ecosystems by definition cannot be 'limited' by nutrients or carbon alone. Here, we outline how models aimed at predicting non-steady state ecosystem responses over time can benefit from dissecting ecosystems into the organismal components and their inherent limitations to better represent plant-microbe interactions in coupled carbon and nutrient models.
DOI:10.2307/2937210URL [本文引用: 1]
DOI:10.1111/j.1461-0248.2008.01228.xURLPMID:18684118 [本文引用: 1]
Theories based on competition for resources predict a monotonic negative relationship between population density and individual biomass in plant populations. They do not consider the role of facilitative interactions, which are known to be important in high stress environments. Using an individual-based 'zone-of-influence' model, we investigated the hypothesis that the balance between facilitative and competitive interactions determines biomass-density relationships. We tested model predictions with a field experiment on the clonal grass Elymus nutans in an alpine meadow. In the model, the relationship between mean individual biomass and density shifted from monotonic to humped as abiotic stress increased. The model results were supported by the field experiment, in which the greatest individual and population biomass were found at intermediate densities in a high-stress alpine habitat. Our results show that facilitation can affect biomass-density relationships.
DOI:10.1007/s00442-009-1364-3URLPMID:19449035 [本文引用: 1]
Transpiration is generally considered a wasteful but unavoidable consequence of photosynthesis, occurring because water is lost when stomata open for CO(2) uptake. Additionally, transpiration has been ascribed the functions of cooling leaves, driving root to shoot xylem transport and mass flow of nutrients through the soil to the rhizosphere. As a consequence of the link between nutrient mass flow and transpiration, nutrient availability, particularly that of NO(3)(-), partially regulates plant water flux. Nutrient regulation of transpiration may function through the concerted regulation of: (1) root hydraulic conductance through control of aquaporins by NO(3)(-), (2) shoot stomatal conductance (g(s)) through NO production, and (3) pH and phytohormone regulation of g(s). These mechanisms result in biphasic responses of water flux to NO(3)(-) availability. The consequent trade-off between water and nutrient flux has important implications for understanding plant distributions, for production of water use-efficient crops and for understanding the consequences of global-change-linked CO(2) suppression of transpiration for plant nutrient acquisition.
[本文引用: 1]
[本文引用: 1]
DOI:10.2307/1312897URL [本文引用: 1]
DOI:10.1007/BF00376856URLPMID:28310700 [本文引用: 1]
In five California evergreen trees and shrubs cooccurring in this study but most common in habitats of different moisture availability, leaf nitrogen was a major determinant of photosynthetic capacity. Within each species, stomatal conductance was highly correlated with photosynthetic capacity, resulting in little variation in the concentration of CO2 in the intercellular spaces. Among species, intercellular CO2 concentrations varied significantly. Under controlled conditions, the leaves that realized the highest photosynthesis per unit of leaf nitrogen tended to realize the lowest photosynthesis per unit of water transpired. The ratio of photosynthesis to transpiration, an instantaneous measure of intrinsic water-use efficiency, was highest in the species commonly found in the direst habitats and lowest in the species most common in the wettes habitats.
DOI:10.1002/ecy.1780URL [本文引用: 2]
DOI:10.1007/s11104-010-0525-9URL [本文引用: 1]
DOI:10.1111/j.1466-8238.2011.00677.xURL [本文引用: 1]
DOI:10.13287/j.1001-9332.201907.025URLPMID:31418219 [本文引用: 1]
Desert ecosystem has unique drought-enduring plants and stoichiometric characteristics. We collected leaf samples of 67 plant species from 63 desert sites in Xinjiang, and explored foliar carbon (C), nitrogen (N) and phosphorus (P) stoichiometry and the relationship between leaf nutrient stoichiometry and climatic factors. The results showed that the average content of leaf C, N and P in these plants were 394, 18.4 and 1.14 mg·g-1, respectively. The mean values of C:N, C:P and N:P were 28, 419 and 18, respectively. In general, shrubs had higher leaf N content than trees and herbs, while leaf P content was lower (higher) in shrubs than in herbaceous plants (trees). Plants with C3 photosynthesis pathway had higher leaf C, N, C:P and N:P than those with C4 pathway. With increasing mean annual precipitation, leaf C first decreased and then increased, while both leaf N and P showed the opposite trend. Leaf C:N and C:P first decreased and then increased, while leaf N:P changed insignificantly. With increasing mean annual temperature, leaf C first decreased and then increased, and leaf N and P decreased, while leaf C:P and N:P increased. Leaf C:N did not change significantly with mean annual temperature. Mean annual precipitation generally showed stronger control on the variation of leaf nutrient stoichiometry than MAT and plant functional types. These results could help predict responses of the biogeochemical cycling of C, N and P to the global climate changes and provide reference and basic data for biogeochemical modeling in the arid regions.
DOI:10.1007/s10021-001-0036-xURL [本文引用: 1]
DOI:10.1111/geb.2010.19.issue-6URL [本文引用: 1]
[本文引用: 1]
DOI:10.1139/X10-167URL [本文引用: 1]
DOI:10.1073/pnas.1700299114URLPMID:29666319 [本文引用: 1]
The structure and function of alpine grassland ecosystems, including their extensive soil carbon stocks, are largely shaped by temperature. The Tibetan Plateau in particular has experienced significant warming over the past 50 y, and this warming trend is projected to intensify in the future. Such climate change will likely alter plant species composition and net primary production (NPP). Here we combined 32 y of observations and monitoring with a manipulative experiment of temperature and precipitation to explore the effects of changing climate on plant community structure and ecosystem function. First, long-term climate warming from 1983 to 2014, which occurred without systematic changes in precipitation, led to higher grass abundance and lower sedge abundance, but did not affect aboveground NPP. Second, an experimental warming experiment conducted over 4 y had no effects on any aspect of NPP, whereas drought manipulation (reducing precipitation by 50%), shifted NPP allocation belowground without affecting total NPP. Third, both experimental warming and drought treatments, supported by a meta-analysis at nine sites across the plateau, increased grass abundance at the expense of biomass of sedges and forbs. This shift in functional group composition led to deeper root systems, which may have enabled plant communities to acquire more water and thus stabilize ecosystem primary production even with a changing climate. Overall, our study demonstrates that shifting plant species composition in response to climate change may have stabilized primary production in this high-elevation ecosystem, but it also caused a shift from aboveground to belowground productivity.
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s00442-008-1229-1URLPMID:19034525 [本文引用: 2]
Regional analyses and biogeochemical models predict that ecosystem N pools and N cycling rates must increase from the semi-arid shortgrass steppe to the sub-humid tallgrass prairie of the Central Great Plains, yet few field data exist to evaluate these predictions. In this paper, we measured rates of net N mineralization, N in above- and belowground primary production, total soil organic matter N pools, soil inorganic N pools and capture in resin bags, decomposition rates, foliar (15)N, and N use efficiency (NUE) across a precipitation gradient. We found that net N mineralization did not increase across the gradient, despite more N generally being found in plant production, suggesting higher N uptake, in the wetter areas. NUE of plants increased with precipitation, and delta(15)N foliar values and resin-captured N in soils decreased, all of which are consistent with the hypothesis that N cycling is tighter at the wet end of the gradient. Litter decomposition appeared to play a role in maintaining this regional N cycling trend: litter decomposed more slowly and released less N at the wet end of the gradient. These results suggest that immobilization of N within the plant-soil system increases from semi-arid shortgrass steppe to sub-humid tallgrass prairie. Despite the fact that N pools increase along a bio-climatic gradient from shortgrass steppe to mixed grass and tallgrass prairie, this element becomes relatively more limiting and is therefore more tightly conserved at the wettest end of the gradient. Similar to findings from forested systems, our results suggest that grassland N cycling becomes more open to N loss with increasing aridity.
DOI:10.1890/09-1634.1URLPMID:21141187 [本文引用: 1]
Modeling studies have shown that nitrogen (N) strongly regulates ecosystem responses and feedback to climate warming. However, it remains unclear what mechanisms underlie N regulation of ecosystem-climate interactions. To examine N regulation of ecosystem feedback to climate change, we have conducted a warming and clipping experiment since November 1999 in a tallgrass prairie of the Great Plains, USA. Infrared heaters were used to elevate soil temperature by an average of 1.96 degrees C at a depth of 2.5 cm from 2000 to 2008. Yearly biomass clipping mimicked hay or biofuel feedstock harvest. We measured carbon (C) and N concentrations, estimated their content and C:N ratio in plant, root, litter, and soil pools. Warming significantly stimulated C storage in aboveground plant, root, and litter pools by 17%, 38%, and 29%, respectively, averaged over the nine years (all P &lt; 0.05) but did not change soil C content or N content in any pool. Plant C:N ratio and nitrogen use efficiency increased in the warmed plots compared to the control plots, resulting primarily from increased dominance of C4 plants in the community. Clipping significantly decreased C and N storage in plant and litter pools (all P &lt; 0.05) but did not have interactive effects with warming on either C or N pools over the nine years. Our results suggest that increased ecosystem nitrogen use efficiency via a shift in species composition toward C4 dominance rather than plant N uptake is a key mechanism underlying warming stimulation of plant biomass growth.
DOI:10.1073/pnas.1006463107URLPMID:20974944 [本文引用: 1]
Stimulation of terrestrial plant production by rising CO(2) concentration is projected to reduce the airborne fraction of anthropogenic CO(2) emissions. Coupled climate-carbon cycle models are sensitive to this negative feedback on atmospheric CO(2), but model projections are uncertain because of the expectation that feedbacks through the nitrogen (N) cycle will reduce this so-called CO(2) fertilization effect. We assessed whether N limitation caused a reduced stimulation of net primary productivity (NPP) by elevated atmospheric CO(2) concentration over 11 y in a free-air CO(2) enrichment (FACE) experiment in a deciduous Liquidambar styraciflua (sweetgum) forest stand in Tennessee. During the first 6 y of the experiment, NPP was significantly enhanced in forest plots exposed to 550 ppm CO(2) compared with NPP in plots in current ambient CO(2), and this was a consistent and sustained response. However, the enhancement of NPP under elevated CO(2) declined from 24% in 2001-2003 to 9% in 2008. Global analyses that assume a sustained CO(2) fertilization effect are no longer supported by this FACE experiment. N budget analysis supports the premise that N availability was limiting to tree growth and declining over time--an expected consequence of stand development, which was exacerbated by elevated CO(2). Leaf- and stand-level observations provide mechanistic evidence that declining N availability constrained the tree response to elevated CO(2); these observations are consistent with stand-level model projections. This FACE experiment provides strong rationale and process understanding for incorporating N limitation and N feedback effects in ecosystem and global models used in climate change assessments.
DOI:10.1007/s11356-019-06538-4URLPMID:31838703 [本文引用: 1]
Valuation of environmental goods and services are frequently spatially heterogeneous, the significance of this heterogeneity for policy analysis is gradually recognized. For the valuation of environmental goods and services in the context of attaining a better environmental status across Heihe River basin (HRB), Northwest of China by 2020, a survey was conducted employing choice experiment approach across the entire river basin. In this research we estimate the impact of distance from inland rivers' origin (spatial attribute) on the inhabitants' willingness to pay for restoration of ecological attributes. A total of 11 ecological attributes were selected including 6 land related attributes and 5 water related attributes. The present study displays the outcomes of the research planned to examine differences in willingness to pay across different locations/distances. A total of five cities and 33 surround villages/townships were included to examine for location effect, while four ad hoc base distances split samples were recognized for distance effect i.e. ≤100?km, ≤200?km, ≤300 and?&gt;?300?km from rivers' origin. The results of mixed logit model recognized that people living at different locations value the attributes differently. For instance the people of Zhangye region are willing to pay RMB 95.66 annually for improvements in biodiversity while in Gaotai the individuals' willingness to pay (WTP) for the same attributes was RMB 45.68. Similarly, the respondents' living nearer to the origin of river were willing to pay relatively higher amount for the upgradation in the degraded ecosystem services than the remainders. The results of willingness to pay obtained by Krinsky Robb method confirmed that the attributes quality of agricultural products and water quality were the most preferred attributes with the willingness to pay RMB 91.09 and 122.89 respectively. The significant results of willingness to pay may serve as a reference for sustainable improvements and uplifting of the degraded ecological attributes.
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s004420050560URLPMID:28308535 [本文引用: 1]
Factors that contribute to interspecific variation in photosynthetic nitrogen-use efficiency (PNUE, the ratio of CO2 assimilation rate to leaf organic nitrogen content) were investigated, comparing ten dicotyledonous species that differ inherently in specific leaf area (SLA, leaf area:leaf dry mass). Plants were grown hydroponically in controlled environment cabinets at two irradiances (200 and 1000?μmol m-2 s-1). CO2 and irradiance response curves of photosynthesis were measured followed by analysis of the chlorophyll, Rubisco, nitrate and total nitrogen contents of the leaves. At both irradiances, SLA ranged more than twofold across species. High-SLA species had higher in situ rates of photosynthesis per unit leaf mass, but similar rates on an area basis. The organic N content per unit leaf area was lower for the high-SLA species and consequently PNUE at ambient light conditions (PNUEamb) was higher in those plants. Differences were somewhat smaller, but still present, when PNUE was determined at saturating irradiances (PNUEmax). An assessment was made of the relative importance of the various factors that underlay interspecific variation in PNUE. For plants grown under low irradiance, PNUEamb of high-SLA species was higher primarily due to their lower N content per unit leaf area. Low-SLA species clearly had an overinvestment in photosynthetic N under these conditions. In addition, high SLA-species allocated a larger fraction of organic nitrogen to thylakoids and Rubisco, which further increased PNUEamb. High-SLA species grown under high irradiance showed higher PNUEamb mainly due to a higher Rubisco specific activity. Other factors that contributed were again their lower contents of Norg per unit leaf area and a higher fraction of photosynthetic N in electron transport and Rubisco. For PNUEmax, differences between species in organic leaf nitrogen content per se were no longer important and higher PNUEmax of the high SLA species was due to a higher fraction of N in?photosynthetic compounds (for low-light plants) and a higher Rubisco specific activity (for high-light grown plants).
DOI:10.1111/nph.14390URLPMID:28000933 [本文引用: 1]
DOI:10.1126/science.1178820URLPMID:19965757 [本文引用: 1]
The interactive effects of rising atmospheric carbon dioxide (CO2) concentrations and elevated nitrogen (N) deposition on plant diversity are not well understood. This is of concern because both factors are important components of global environmental change and because each might suppress diversity, with their combined effects possibly additive or synergistic. In a long-term open-air experiment, grassland assemblages planted with 16 species were grown under all combinations of ambient and elevated CO2 and ambient and elevated N. Over 10 years, elevated N reduced species richness by 16% at ambient CO2 but by just 8% at elevated CO2. This resulted from multiple effects of CO2 and N on plant traits and soil resources that altered competitive interactions among species. Elevated CO2 thus ameliorated the negative effects of N enrichment on species richness.
DOI:10.1016/j.fcr.2017.08.022URL [本文引用: 1]
DOI:10.1007/BF00324222URLPMID:28313910 [本文引用: 1]
Data on soil nutrient availability for humid tropical forests are often reported, but are rarely integrated in an ecologically meaningful way with other measures of nutrient cycling. In this paper, estimates of soil nutrient availability and the inverse of litterfall nutrient concentrations (as an index of plant nutrient use) were compared, using data from 36 sites throughout the humid tropics, to determine if relationships exist between commonly used indices of nutrient cycling for plants and soils. Measures of both extractable and total soil P were significantly and positively correlated with the ratio of litterfall mass/P, particularly for montane tropical forests. Extractable soil P was also significantly correlated with litterfall mass for lowland humid tropical forests, explaining 58% of the variability in litterfall mass. A weak, albeit significant correlation was found between exchangeable soil Ca and litterfall mass/Ca, even though soil extraction techniques vary greatly. No significant relationship was found for total soil N, the most commonly measured soil N pool, and the inverse of litterfall N concentrations. The results suggest that our indices of soil P are related to litterfall processes, but that other measures, particularly total soil N, may not be as relevant to nutrient cycling by the vegetation.
DOI:10.1016/j.jenvman.2019.109817URLPMID:31783211 [本文引用: 2]
Mitigating the environmental impact of nonpoint source pollution from intensively managed urban and agricultural landscapes is of paramount concern to watershed managers. Golf course turfgrass systems, which receive significant fertilizer inputs, have been cited as significant sources of nutrient loading to groundwater and surface water, but a contemporary synthesis of golf course nutrient export rates is lacking. This review of nitrogen (N) and phosphorus (P) loss from golf courses and the factors affecting it aims to support watershed management efforts and decision making. We discuss previous literature reviews, examine seven golf course studies that quantify nutrient export from delineated drainage areas, and analyze the results of 40 turfgrass plot experiments. Studies were collected systematically and selected based on predetermined inclusion criteria. Combining evidence from both watershed- and plot-scale studies, typical inorganic N and P losses from golf courses via leaching and runoff are on the order of 2-12?kg?ha-1 yr-1 and 0.1-1.0?kg?ha-1 yr-1, respectively. Typical total N and P losses are around 2-20?kg?ha-1 yr-1 and 1.5-5?kg?ha-1 yr-1, respectively. However, the potential for large variation in export rates across 2-3 orders of magnitude must be emphasized. The body of turfgrass literature stresses the importance of best management practices (BMPs) related to applying fertilizer to match plant needs and reducing opportunities for its transport. Accounting for all sources of nutrients, especially soil P, in determining fertilizer application rates and avoiding excessive irrigation to prevent leaching of nutrients from the rootzone is particularly important. BMPs can also reduce nutrient leaching and runoff by controlling the movement of water across the landscape and promoting natural nutrient attenuation, such as with vegetative stream buffers.
[本文引用: 1]
[本文引用: 1]
DOI:10.2307/210739URL [本文引用: 1]
DOI:10.3390/su9112035URL [本文引用: 1]
DOI:10.1186/1471-2164-11-619URLPMID:21054883 [本文引用: 4]
As the only truly flying mammals, bats use their unique wing - consisting of four elongated digits (digits II-V) connected by membranes - to power their flight. In addition to the elongated digits II-V, the forelimb contains one shorter digit (digit I) that is morphologically similar to the hindlimb digits. Here, we capitalized on the morphological variation among the bat forelimb digits to investigate the molecular mechanisms underlying digit elongation and wing formation. Using next generation sequencing technology, we performed digital gene expression tag profiling (DGE-tag profiling) of developing digits in a pooled sample of two Myotis ricketti and validated our sequencing results using real-time quantitative PCR (RT-qPCR) of gene expression in the developing digits of two Hipposideros armiger.
DOI:10.1007/BF00043032URL [本文引用: 1]
[本文引用: 1]
DOI:10.1111/geb.v28.4URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s10531-015-1034-1URL [本文引用: 1]
DOI:10.1146/annurev-arplant-042811-105532URLPMID:22224450 [本文引用: 1]
Crop productivity relies heavily on nitrogen (N) fertilization. Production and application of N fertilizers consume huge amounts of energy, and excess is detrimental to the environment; therefore, increasing plant N use efficiency (NUE) is essential for the development of sustainable agriculture. Plant NUE is inherently complex, as each step-including N uptake, translocation, assimilation, and remobilization-is governed by multiple interacting genetic and environmental factors. The limiting factors in plant metabolism for maximizing NUE are different at high and low N supplies, indicating great potential for improving the NUE of current cultivars, which were bred in well-fertilized soil. Decreasing environmental losses and increasing the productivity of crop-acquired N requires the coordination of carbohydrate and N metabolism to give high yields. Increasing both the grain and N harvest index to drive N acquisition and utilization are important approaches for breeding future high-NUE cultivars.
DOI:10.1016/j.bse.2015.07.037URL [本文引用: 1]
DOI:10.3724/SP.J.1258.2014.00014URL [本文引用: 1]
DOI:10.3724/SP.J.1258.2014.00014URL [本文引用: 1]
DOI:10.1007/s11258-017-0719-9URL [本文引用: 2]
DOI:10.1073/pnas.1012490107URLPMID:21115833 [本文引用: 1]
Climate change has caused advances in spring phases of many plant species. Theoretically, however, strong warming in winter could slow the fulfillment of chilling requirements, which may delay spring phenology. This phenomenon should be particularly pronounced in regions that are experiencing rapid temperature increases and are characterized by highly temperature-responsive vegetation. To test this hypothesis, we used the Normalized Difference Vegetation Index ratio method to determine the beginning, end, and length of the growing season of meadow and steppe vegetation of the Tibetan Plateau in Western China between 1982 and 2006. We then correlated observed phenological dates with monthly temperatures for the entire period on record. For both vegetation types, spring phenology initially advanced, but started retreating in the mid-1990s in spite of continued warming. Together with an advancing end of the growing season for steppe vegetation, this led to a shortening of the growing period. Partial least-squares regression indicated that temperatures in both winter and spring had strong effects on spring phenology. Although warm springs led to an advance of the growing season, warm conditions in winter caused a delay of the spring phases. This delay appeared to be related to later fulfillment of chilling requirements. Because most plants from temperate and cold climates experience a period of dormancy in winter, it seems likely that similar effects occur in other environments. Continued warming may strengthen this effect and attenuate or even reverse the advancing trend in spring phenology that has dominated climate-change responses of plants thus far.
DOI:10.1007/s00442-006-0409-0URLPMID:16708228 [本文引用: 4]
The concept of nutrient use efficiency is central to understanding ecosystem functioning because it is the step in which plants can influence the return of nutrients to the soil pool and the quality of the litter. Theory suggests that nutrient efficiency increases unimodally with declining soil resources, but this has not been tested empirically for N and water in grassland ecosystems, where plant growth in these ecosystems is generally thought to be limited by soil N and moisture. In this paper, we tested the N uptake and the N use efficiency (NUE) of two Stipa species (S. grandis and S. krylovii) from 20 sites in the Inner Mongolia grassland by measuring the N content of net primary productivity (NPP). NUE is defined as the total net primary production per unit N absorbed. We further distinguished NUE from N response efficiency (NRE; production per unit N available). We found that NPP increased with soil N and water availability. Efficiency of whole-plant N use, uptake, and response increased monotonically with decreasing soil N and water, being higher on infertile (dry) habitats than on fertile (wet) habitats. We further considered NUE as the product of the N productivity (NP the rate of biomass increase per unit N in the plant) and the mean residence time (MRT; the ratio between the average N pool and the annual N uptake or loss). The NP and NUE of S. grandis growing usually in dry and N-poor habitats exceeded those of S. krylovii abundant in wet and N-rich habitats. NUE differed among sites, and was often affected by the evolutionary trade-off between NP and MRT, where plants and communities had adapted in a way to maximize either NP or MRT, but not both concurrently. Soil N availability and moisture influenced the community-level N uptake efficiency and ultimately the NRE, though the response to N was dependent on the plant community examined. These results show that soil N and water had exerted a great impact on the N efficiency in Stipa species. The intraspecific differences in N efficiency within both Stipa species along soil resource availability gradient may explain the differences in plant productivity on various soils, which will be conducive to our general understanding of the N cycling and vegetation dynamics in northern Chinese grasslands.
DOI:10.1111/1365-2745.12425URL [本文引用: 1]
DOI:10.1007/s11442-014-1087-1URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.ecolind.2016.01.012URL [本文引用: 1]
The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns
1
1999
... 从草原植物不同功能群NUE来看, 莎草科和禾草科植物的NUE均高于杂类草, 这一结果部分地与前人研究结果一致.例如,
Nitrogen use efficiency of Carex species in relation to nitrogen supply
2
1994
... 我们发现随着气温升高, 草原植物叶片NUE显著降低.虽然未见区域尺度上植物NUE与温度之间关系的研究报道, 但
... ); 第二, 部分研究是基于室内氮添加实验, 而室内实验条件通常比较理想, 容易检测出氮添加效应.例如,
Plant nitrogen concentration, use efficiency, and contents in a tallgrass prairie ecosystem under experimental warming
2
2005
... 草原植物NUE主要受气候条件、土壤氮含量和植物功能群的共同影响.降水是限制草地生态系统生产力的主要因素, 尤其是干旱和半干旱草原(
... 从草原植物不同功能群NUE来看, 莎草科和禾草科植物的NUE均高于杂类草, 这一结果部分地与前人研究结果一致.例如,
荒漠草原优势植物养分利用及化学计量特征研究
1
2013
... 本研究结果发现植物叶片或根系NUE与年降水量之间没有显著关系, 这与前人的研究结果不一致.例如,
荒漠草原优势植物养分利用及化学计量特征研究
1
2013
... 本研究结果发现植物叶片或根系NUE与年降水量之间没有显著关系, 这与前人的研究结果不一致.例如,
Primary production and rain use efficiency across a precipitation gradient on the Mongolia Plateau
1
2008
... 草原植物NUE主要受气候条件、土壤氮含量和植物功能群的共同影响.降水是限制草地生态系统生产力的主要因素, 尤其是干旱和半干旱草原(
Nitrogen-use-efficiency: A biologically meaningful definition?
1
1987
... 氮是植物生长所必需的主要营养元素(
Nitrogen availability and nitrogen use efficiency in loblolly pine stands
1
1986
... 氮是植物生长所必需的主要营养元素(
Strategies to alleviate poverty and grassland degradation in Inner Mongolia: Intensification vs production efficiency of livestock systems
1
2015
... 草原是全球陆地生态系统的重要组成部分.中国草原约占国土面积的40% (
Gross nitrogen transformations in adjacent native and plantation forests of subtropical Australia
1
2007
... 土壤氮含量是影响草原植物NUE的重要因素.植物主要通过根系从土壤中获取无机氮, 用于维持自身的生长发育(
Nutritional controls over nitrogen and phosphorus resorption from Alaskan birch leaves
1
1991
... 植物NUE计算一般采用
Balance between facilitation and resource competition determines biomass-density relationships in plant populations
1
2008
... 草原植物NUE主要受气候条件、土壤氮含量和植物功能群的共同影响.降水是限制草地生态系统生产力的主要因素, 尤其是干旱和半干旱草原(
The importance of nutritional regulation of plant water flux
1
2009
... 草原植物NUE主要受气候条件、土壤氮含量和植物功能群的共同影响.降水是限制草地生态系统生产力的主要因素, 尤其是干旱和半干旱草原(
内蒙古典型草原地区5类植物群落叶面积指数的比较研究
1
2001
... 从草原植物不同功能群NUE来看, 莎草科和禾草科植物的NUE均高于杂类草, 这一结果部分地与前人研究结果一致.例如,
内蒙古典型草原地区5类植物群落叶面积指数的比较研究
1
2001
... 从草原植物不同功能群NUE来看, 莎草科和禾草科植物的NUE均高于杂类草, 这一结果部分地与前人研究结果一致.例如,
Organism size, life history, and N:P stoichiometry: Toward a unified view of cellular and ecosystem processes
1
1996
... 氮是植物生长所必需的主要营养元素(
Compromises between water-use efficiency and nitrogen-use efficiency in five species of California evergreens
1
1983
... 氮是植物生长所必需的主要营养元素(
Linking above- and belowground traits to soil and climate variables: An integrated database on China’s grassland species
2
2017
... 本研究数据库来自于前人发表的数据文章(
... ))植物.植物叶片和根系样品在野外经过处理后, 在实验室内烘干至恒质量, 统一使用球磨仪(NM200, Retsch, Haan, Germany)粉碎.粉碎后的样品采用元素分析仪(2400 II CHN, Perkin-Elmer, Massachusetts, USA)测定氮含量.同时对每个地点0-10 cm土壤样品进行预处理, 新鲜土样过2 mm筛, 并将细根挑干净后分成两份.一份土样用KCl溶液浸提, 浸提液使用连续流动分析仪(SAN Plus, Skalar, Netherlands)测定土壤硝态氮和铵态氮含量; 另一份土样在室内风干之后, 使用球磨仪进行粉碎, 然后采用凯氏定氮仪(KjelFlex-360, BUCHI, Flawil, Switzerland)测定土壤全氮含量.相关数据的测定方法详见
Tradeoffs between nitrogen- and water-?use efficiency in dominant species of the semiarid steppe of Inner Mongolia
1
2011
... 本研究结果发现植物叶片或根系NUE与年降水量之间没有显著关系, 这与前人的研究结果不一致.例如,
Floral, climatic and soil pH controls on leaf ash content in China’s terrestrial plants
1
2012
... 草原植物NUE在不同功能群之间可能存在差异.相关研究表明, 不同植物功能群的氮利用策略不同, 如在氮吸收特点、氮在植物体内运输途径、分配状况以及氮再吸收等方面存在差异, 这些差异可能导致功能群之间NUE的不同(
Leaf nitrogen:phosphorus stoichiometry across Chinese grassland biomes
1
2008
... 从草原植物不同功能群NUE来看, 莎草科和禾草科植物的NUE均高于杂类草, 这一结果部分地与前人研究结果一致.例如,
Ecosystem nutrient use efficiency, productivity, and nutrient accrual in model tropical communities
1
2001
... 氮是植物生长所必需的主要营养元素(
Precipitation-use efficiency along a 4500-km grassland transect
1
2010
... 草原植物NUE主要受气候条件、土壤氮含量和植物功能群的共同影响.降水是限制草地生态系统生产力的主要因素, 尤其是干旱和半干旱草原(
Rain use efficiency: A unifying concept in arid-land ecology
1
1984
... 草原植物NUE主要受气候条件、土壤氮含量和植物功能群的共同影响.降水是限制草地生态系统生产力的主要因素, 尤其是干旱和半干旱草原(
Effects of nitrogen additions on nitrogen resorption and use efficiencies and foliar litterfall of six tree species in a mixed birch and poplar forest, northeastern China
1
2010
... 氮是植物生长所必需的主要营养元素(
Shifting plant species composition in response to climate change stabilizes grassland primary production
1
2018
... 从草原植物不同功能群NUE来看, 莎草科和禾草科植物的NUE均高于杂类草, 这一结果部分地与前人研究结果一致.例如,
生物多样性、水分和氮素添加对典型草原优势植物养分利用的影响
1
2010
... 本研究结果发现植物叶片或根系NUE与年降水量之间没有显著关系, 这与前人的研究结果不一致.例如,
生物多样性、水分和氮素添加对典型草原优势植物养分利用的影响
1
2010
... 本研究结果发现植物叶片或根系NUE与年降水量之间没有显著关系, 这与前人的研究结果不一致.例如,
Conservation of nitrogen increases with precipitation across a major grassland gradient in the Central Great Plains of North America
2
2009
... 草原植物NUE主要受气候条件、土壤氮含量和植物功能群的共同影响.降水是限制草地生态系统生产力的主要因素, 尤其是干旱和半干旱草原(
... 本研究结果发现植物叶片或根系NUE与年降水量之间没有显著关系, 这与前人的研究结果不一致.例如,
Nitrogen regulation of the climate-carbon feedback: Evidence from a long-term global change experiment
1
2010
... 草原植物NUE主要受气候条件、土壤氮含量和植物功能群的共同影响.降水是限制草地生态系统生产力的主要因素, 尤其是干旱和半干旱草原(
CO2 enhancement of forest productivity constrained by limited nitrogen availability
1
2010
... 草原植物NUE主要受气候条件、土壤氮含量和植物功能群的共同影响.降水是限制草地生态系统生产力的主要因素, 尤其是干旱和半干旱草原(
Effect of soil warming and rainfall patterns on soil N cycling in Northern Europe
1
2010
... 研究结果显示草甸草原植物叶片NUE显著高于荒漠草原、典型草原和高寒草甸, 而高寒草甸根系NUE显著高于其他生态系统.本研究中草甸草原年降水量为387 mm, 年平均气温-1.4 ℃, 干旱指数1.31, 优势种为禾本科植物.高寒草甸年降水量450 mm, 年均气温-2.4 ℃, 干旱指数0.87, 优势种为莎草科植物.进一步通过结构方程模型分析发现, 植物功能群对NUE影响最大, 干旱指数次之.这说明造成草原生态系统之间NUE的差异主要由植物功能群和气候条件共同决定.本研究中高寒草甸和草甸草原优势植物NUE均比较高, 并且它们的干旱指数低于其他生态系统.低干旱指数(高土壤含水量)有利于微生物对土壤有机质的分解和氨化, 从而提高土壤氮矿化速率(
豆科和非固氮植物氮磷利用效率的比较研究
1
2017
... 同时, 我们还发现豆科植物NUE显著低于非豆科植物, 可能主要由豆科与非豆科植物在养分利用策略和环境适应性方面的差异所导致(
豆科和非固氮植物氮磷利用效率的比较研究
1
2017
... 同时, 我们还发现豆科植物NUE显著低于非豆科植物, 可能主要由豆科与非豆科植物在养分利用策略和环境适应性方面的差异所导致(
Photosynthetic nitrogen-use efficiency of species that differ inherently in specific leaf area
1
1998
... 同时, 我们还发现豆科植物NUE显著低于非豆科植物, 可能主要由豆科与非豆科植物在养分利用策略和环境适应性方面的差异所导致(
Disentangling the complexities of how legumes and their symbionts regulate plant nitrogen access and storage
1
2017
... 草原植物NUE在不同功能群之间可能存在差异.相关研究表明, 不同植物功能群的氮利用策略不同, 如在氮吸收特点、氮在植物体内运输途径、分配状况以及氮再吸收等方面存在差异, 这些差异可能导致功能群之间NUE的不同(
Elevated CO2 reduces losses of plant diversity caused by nitrogen deposition
1
2009
... 草原植物NUE主要受气候条件、土壤氮含量和植物功能群的共同影响.降水是限制草地生态系统生产力的主要因素, 尤其是干旱和半干旱草原(
Fall nitrogen application increases seed yield, forage yield and nitrogen use efficiency more than spring nitrogen application in Leymus chinensis, a perennial grass
1
2017
... 同时, 我们还发现豆科植物NUE显著低于非豆科植物, 可能主要由豆科与非豆科植物在养分利用策略和环境适应性方面的差异所导致(
Is nutrient availability related to plant nutrient use in humid tropical forests?
1
1994
... 土壤氮含量是影响草原植物NUE的重要因素.植物主要通过根系从土壤中获取无机氮, 用于维持自身的生长发育(
How to reduce nitrogen losses in intensive-grassland management
2
1988
... 本研究结果发现植物叶片或根系NUE与年降水量之间没有显著关系, 这与前人的研究结果不一致.例如,
... 研究结果显示草甸草原植物叶片NUE显著高于荒漠草原、典型草原和高寒草甸, 而高寒草甸根系NUE显著高于其他生态系统.本研究中草甸草原年降水量为387 mm, 年平均气温-1.4 ℃, 干旱指数1.31, 优势种为禾本科植物.高寒草甸年降水量450 mm, 年均气温-2.4 ℃, 干旱指数0.87, 优势种为莎草科植物.进一步通过结构方程模型分析发现, 植物功能群对NUE影响最大, 干旱指数次之.这说明造成草原生态系统之间NUE的差异主要由植物功能群和气候条件共同决定.本研究中高寒草甸和草甸草原优势植物NUE均比较高, 并且它们的干旱指数低于其他生态系统.低干旱指数(高土壤含水量)有利于微生物对土壤有机质的分解和氨化, 从而提高土壤氮矿化速率(
植物的养分利用效率(NUE)及植物对养分胁迫环境的适应策略
1
2000
... 草原植物NUE在不同功能群之间可能存在差异.相关研究表明, 不同植物功能群的氮利用策略不同, 如在氮吸收特点、氮在植物体内运输途径、分配状况以及氮再吸收等方面存在差异, 这些差异可能导致功能群之间NUE的不同(
植物的养分利用效率(NUE)及植物对养分胁迫环境的适应策略
1
2000
... 草原植物NUE在不同功能群之间可能存在差异.相关研究表明, 不同植物功能群的氮利用策略不同, 如在氮吸收特点、氮在植物体内运输途径、分配状况以及氮再吸收等方面存在差异, 这些差异可能导致功能群之间NUE的不同(
An approach toward a rational classification of climate
1
1948
... 本研究数据库来自于前人发表的数据文章(
Effect of irrigation schemes on forage yield, water use efficiency, and nutrients in artificial grassland under arid conditions
1
2017
... 氮是植物生长所必需的主要营养元素(
Nitrogen-use efficiency in six perennial grasses from contrasting habitats
4
1997
... 土壤氮含量是影响草原植物NUE的重要因素.植物主要通过根系从土壤中获取无机氮, 用于维持自身的生长发育(
... 草原植物NUE在不同功能群之间可能存在差异.相关研究表明, 不同植物功能群的氮利用策略不同, 如在氮吸收特点、氮在植物体内运输途径、分配状况以及氮再吸收等方面存在差异, 这些差异可能导致功能群之间NUE的不同(
... 我们发现随着气温升高, 草原植物叶片NUE显著降低.虽然未见区域尺度上植物NUE与温度之间关系的研究报道, 但
... 、
The relationship between nutrient availability, shoot biomass and species richness in grassland and wetland communities
1
1983
... 氮是植物生长所必需的主要营养元素(
Nitrogen limitation on land and in the sea: How can it occur?
1
1991
... 氮是植物生长所必需的主要营养元素(
Vegetation type controls root turnover in global grasslands
1
2019
... 植物NUE计算一般采用
青藏高原退化高寒草地土壤氮矿化特征以及影响因素研究
1
2018
... 土壤氮含量是影响草原植物NUE的重要因素.植物主要通过根系从土壤中获取无机氮, 用于维持自身的生长发育(
青藏高原退化高寒草地土壤氮矿化特征以及影响因素研究
1
2018
... 土壤氮含量是影响草原植物NUE的重要因素.植物主要通过根系从土壤中获取无机氮, 用于维持自身的生长发育(
Vegetation and soil responses to livestock grazing in Central Asian grasslands: A review of Chinese literature
1
2016
... 草原是全球陆地生态系统的重要组成部分.中国草原约占国土面积的40% (
Plant nitrogen assimilation and use efficiency
1
2012
... 土壤氮含量是影响草原植物NUE的重要因素.植物主要通过根系从土壤中获取无机氮, 用于维持自身的生长发育(
The patterns of nitrogen and phosphorus stoichiometry across communities along altitudinal gradients in Qilian Mountains, China
1
2015
... 从草原植物不同功能群NUE来看, 莎草科和禾草科植物的NUE均高于杂类草, 这一结果部分地与前人研究结果一致.例如,
青藏高原高寒草甸植物群落生物量对氮、磷添加的响应
1
2014
... 我们发现随着气温升高, 草原植物叶片NUE显著降低.虽然未见区域尺度上植物NUE与温度之间关系的研究报道, 但
青藏高原高寒草甸植物群落生物量对氮、磷添加的响应
1
2014
... 我们发现随着气温升高, 草原植物叶片NUE显著降低.虽然未见区域尺度上植物NUE与温度之间关系的研究报道, 但
Relationships between below-ground biomass and foliar N:P stoichiometry along climatic and altitudinal gradients of the Chinese grassland transect
2
2017
... 我们发现随着气温升高, 草原植物叶片NUE显著降低.虽然未见区域尺度上植物NUE与温度之间关系的研究报道, 但
... 分析中国草地132个地点的植物叶片氮浓度发现, 叶片氮浓度随气温升高而降低.尽管植物氮浓度与氮利用效率之间存在着非线性(倒数函数)关系, 并非一一对应的线性关系, 但是
Winter and spring warming result in delayed spring phenology on the Tibetan Plateau
1
2010
... 草原植物NUE主要受气候条件、土壤氮含量和植物功能群的共同影响.降水是限制草地生态系统生产力的主要因素, 尤其是干旱和半干旱草原(
Nitrogen response efficiency increased monotonically with decreasing soil resource availability: A case study from a semiarid grassland in northern China
4
2006
... 本研究结果发现植物叶片或根系NUE与年降水量之间没有显著关系, 这与前人的研究结果不一致.例如,
... 我们发现随着气温升高, 草原植物叶片NUE显著降低.虽然未见区域尺度上植物NUE与温度之间关系的研究报道, 但
... 第三, 部分研究是基于较小空间尺度上的自然氮梯度实验.例如,
... 研究结果显示草甸草原植物叶片NUE显著高于荒漠草原、典型草原和高寒草甸, 而高寒草甸根系NUE显著高于其他生态系统.本研究中草甸草原年降水量为387 mm, 年平均气温-1.4 ℃, 干旱指数1.31, 优势种为禾本科植物.高寒草甸年降水量450 mm, 年均气温-2.4 ℃, 干旱指数0.87, 优势种为莎草科植物.进一步通过结构方程模型分析发现, 植物功能群对NUE影响最大, 干旱指数次之.这说明造成草原生态系统之间NUE的差异主要由植物功能群和气候条件共同决定.本研究中高寒草甸和草甸草原优势植物NUE均比较高, 并且它们的干旱指数低于其他生态系统.低干旱指数(高土壤含水量)有利于微生物对土壤有机质的分解和氨化, 从而提高土壤氮矿化速率(
Individual size inequality links forest diversity and above-ground biomass
1
2015
... 植物NUE计算一般采用
Spatial and temporal variability in the net primary production of alpine grassland on the Tibetan Plateau since 1982
1
2014
... 草原是全球陆地生态系统的重要组成部分.中国草原约占国土面积的40% (
塔克拉玛干沙漠南缘豆科与非豆科植物的氮分配
1
2010
... 同时, 我们还发现豆科植物NUE显著低于非豆科植物, 可能主要由豆科与非豆科植物在养分利用策略和环境适应性方面的差异所导致(
塔克拉玛干沙漠南缘豆科与非豆科植物的氮分配
1
2010
... 同时, 我们还发现豆科植物NUE显著低于非豆科植物, 可能主要由豆科与非豆科植物在养分利用策略和环境适应性方面的差异所导致(
Plant functional diversity mediates the effects of vegetation and soil properties on community-?level plant nitrogen use in the restoration of semiarid sandy grassland
1
2016
... 从草原植物不同功能群NUE来看, 莎草科和禾草科植物的NUE均高于杂类草, 这一结果部分地与前人研究结果一致.例如,

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}