 ,
, ,*, 刘明国,**, 董胜君, 吴月亮, 张皓凯沈阳农业大学林学院, 沈阳 110161
,*, 刘明国,**, 董胜君, 吴月亮, 张皓凯沈阳农业大学林学院, 沈阳 110161Diversity and geographical variations of germplasm resources of Armeniaca mandshurica
Hao XU,,*, Ming-Guo LIU,**, Sheng-Jun DONG, Yue-Liang WU, Hao-Kai ZHANGCollege of Forestry, Shenyang Agricultural University, Shenyang 110161, China通讯作者: * E-mail:2017220539@stu.syau.edu.cn**liumingguo916@163.com
编委: 何维明
责任编辑: 李敏, 赵航(实习)
收稿日期:2019-03-18接受日期:2019-05-15网络出版日期:2019-07-20
| 基金资助: |
Received:2019-03-18Accepted:2019-05-15Online:2019-07-20
| Fund supported: |

摘要
东北杏(Armeniaca mandshurica)是集观赏、经济、用材于一体的重要树种, 长期以来主要处于野生和半野生状态, 鲜有相关研究报道。该研究对东北杏主要分布区内种质资源状况开展了调查, 选择来自辽宁、吉林与黑龙江的47份典型种质进行了22个定量描述性状和7个定性描述性状的观测, 旨在为该树种的种质资源收集、评价与保护提供重要参考。采用变异系数等指标分析定量描述性状多样性, 使用频率分布等指标分析定性描述性状多样性。通过趋势面方法分析定量描述性状的地理变化规律。利用99对SSR引物对47份东北杏种质进行PCR扩增, 应用遗传相似系数分析东北杏种质的遗传多样性。基于表型性状和SSR分子标记采用聚类分析方法分别对种质进行分类。东北杏不同种质间呈现出较高的表型多样性, 其中19个定量描述性状指标的变异系数在9.40%-55.98%之间, 变异系数最大的为小枝长度, 变异系数最小的为种仁宽; 7个定性描述性状的Shannon-Wiener指数在0.58-1.22之间。由于调查区域的地理位置与主要气候因子间存在着显著的相关性, 东北杏种质的定量描述性状与其地理位置有着密切的关系。其中, 小枝长度呈由东向西逐渐增大的梯度变化, 小枝粗度、种子质量呈由北向南逐渐增大的梯度变化, 果柄长呈由东北向西南逐渐增大的梯度变化; 小枝长度、果柄长与海拔呈正相关关系, 小枝粗度与海拔呈负相关关系, 种子质量与海拔相关性很小。基于26个表型性状进行系统聚类, 将47份东北杏种质分为4类, 分类结果主要体现了东北杏种质特征的差别, 同时也在一定程度上体现了种质产地的效应; 基于遗传相似系数进行聚类分析, 将47份东北杏种质也分为4类, 分类结果体现了种质产地效应; 卡方检验表明, 两种聚类结果相关性不显著, 外在环境是影响东北杏表型变异的重要因素。
关键词:
Abstract
Aims Armeniaca mandshurica is an important species which serves the need of ornamental, wood and other economical uses. This species has been in wild or semi-wild state for a long time and few studies about this species have been conducted. This paper aimed to provide an important reference for the collection, evaluation and protection of germplasm resources of A. mandshurica.
Methods The status of the germplasm resources in the main distribution area of A. mandshurica was investigated. In total, 47 typical sample trees from Liaoning, Jilin and Heilongjiang Provinces were selected. For each tree, 22 quantitative traits and 7 qualitative traits were measured. The diversity of quantitative traits was represented by indicators such as coefficient of variation, and the diversity of qualitative traits was represented by indicators such as frequency distribution. Trend surface analysis was used to explore the geographical variation in the quantitative traits. A total of 115 pairs of SSR primers were used for PCR amplification for 47 A. mandshurica germplasms, and the genetic diversity of A. mandshurica germplasms was analyzed using genetic similarity coefficient. Germplasms were classified by cluster analysis based on phenotypic traits and SSR markers separately.
Important findings High phenotypic diversity was found among different germplasms in A. mandshurica. The coefficients of variation (CV) of the 19 quantitative traits ranged from 9.40% to 55.98%. Among the 19 traits, twig length had the highest CV and kernel width had the lowest CV. The Shannon-Wiener index of 7 qualitative traits ranged from 0.58 to 1.22. Due to the significant correlation between geographical locations and main climatic factors within the study area, the quantitative traits of A. mandshurica germplasms were closely related to their geographical locations. Among them, twig length increased from east to west, twig thickness and seed mass increased from north to south, and fruit handle length increased from northeast to southwest. Twig length and fruit handle length were positively correlated with altitude, twig thickness was negatively correlated with altitude, and seed mass was not correlated with altitude. Clustering analysis based on 26 phenotypic traits showed that the 47 A. mandshurica germplasms could be divided into 4 categories, which mainly reflected the difference of the germplasm characteristics in A. mandshurica and to some extent also reflected the difference of germplasm productions. Clustering analysis based on genetic similarity coefficients showed that the 47 A. mandshurica germplasms were also divided into 4 categories, which reflected the difference of germplasm productions. Chi-square test showed that the correlation between the two clustering results was not significant, and that the external environment was the key factor affecting phenotypic variations in A. mandshurica.
Keywords:
PDF (1878KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
徐豪, 刘明国, 董胜君, 吴月亮, 张皓凯. 东北杏种质资源多样性及其地理变化. 植物生态学报, 2019, 43(7): 585-600. DOI: 10.17521/cjpe.2019.0060
XU Hao, LIU Ming-Guo, DONG Sheng-Jun, WU Yue-Liang, ZHANG Hao-Kai.
东北杏(Armeniaca mandshurica)为蔷薇科杏属植物, 主要分布于我国黑龙江、吉林和辽宁的东部地区, 在朝鲜的中北部和俄罗斯南乌苏里地区也有分布。东北杏用途很广, 是集观赏、经济、用材于一身的重要树种(庞振伟等, 2001; 王利兵, 2010)。目前该树种尚未得到充分开发, 主要处于野生和半野生状态, 除了少量园林绿化栽培外, 其他利用方式鲜见报道。同时由于缺乏保护措施, 其资源破坏严重, 数量日渐减少。
种质资源是林木改良的物质基础, 同时也是遗传学、分类学、生物学等理论研究的重要材料, 进行种质资源调查、评价是对其合理保护、利用的前提。目前, 林木种质资源调查、评价研究主要集中于主要经济林树种(Wu et al., 2007; 刘小利等, 2015; 刘胤等, 2016)和用材林树种(Liu et al., 1997; Wan & Zhang, 2013; 郭琪等, 2019)。山杏是野生的蔷薇科杏属植物统称, 相关研究在西伯利亚杏(Armeniaca sibirica)上比较活跃, 已从形态标记(李明等, 2011b; 尹健等, 2015)、蛋白质和酶标记(刘明国等, 2006; 李明等, 2011a)、孢粉学(刘明国等, 2015)及SSR分子标记(Wang et al., 2014; 金玲等, 2018)等方面开展了种质资源评价和分类研究。但作为山杏重要组成树种的东北杏, 未见相关研究报道。
遗传多样性是指物种种内基因组成的多样性, 不同的遗传多样性研究及检测方法存在差异, 可从形态学、细胞学及DNA分子等水平上进行, 其中表型测定是最简单和最直接的方法(李文英和顾万春, 2005)。表型性状是由基因型和环境共同作用的结果, 表型多样性是环境多样性和遗传多样性的综合体现, 通过对表型变异程度和地理变化规律的研究, 不仅可以揭示物种适应和进化的方式, 而且有助于了解种群遗传变异的大小, 这也是遗传育种工作的基础(苏世平等, 2013; 杨晓霞等, 2016)。近年来, 对野杏(Armeniaca vulgaris var. ansu)(曹倩等, 2015)、枫香树(Liquidambar formosana)(何庆海等, 2018)、尾叶樱桃(Cerasus dielsiana)(朱弘等, 2018)等植物的表型变异特征和Azadirachta indica (Kundu & Tigerstedt, 1997)、无患子(Sapindus mukorossi)(邵文豪等, 2013)、乌饭树(Vaccinium bracteatum)(刘仁林等, 2016)等树种的表型地理变异已有较多研究报道。
本研究以辽宁、吉林和黑龙江的47份东北杏种质为试验材料, 对其22个定量描述性状和7个定性描述性状进行了观测, 并进行了SSR分子标记试验, 在此基础上分析了种质资源多样性及其地理变化, 以了解性状地理分布格局, 揭示变异特点、变异程度及其与地理环境的关系, 旨在为收集、评价与保护东北杏种质资源提供重要参考, 同时为该树种的良种选育工作提供科学依据。
1 材料和方法
1.1 研究区域自然概况
本研究调查区域包括辽宁凤城市、本溪县、桓仁县、新宾县、清原县、沈阳市, 吉林抚松县、磐石市、桦甸市、敦化市、吉林市, 黑龙江宁安市、尚志市、鸡西市和阿城区等地。调查区域的年平均气温为3.4-8.5 ℃, 7月最高气温平均为25.3-29.0 ℃, 1月最低气温平均为-25.4- -14.9 ℃, 年降水量为530.2-1 077.8 mm。地貌类型为中低山地、低山丘陵和平原, 土壤类型主要有棕壤、暗棕壤、白浆土、草甸土等, 植被属长白植物区系。1.2 试验材料
在2016年6-8月, 开展了对上述15个地区的东北杏种质资源调查, 共选择47份东北杏典型种质(表1)进行表型性状观测。Table 1
表1
表1供试东北杏种质的地理位置
Table 1
| 种质编号 Germplasm No. | 地点 Site | 海拔 Altitude (m) | 经纬度 Longitude (E) and Latitude (N) | 种质编号 Germplasm No. | 地点 Site | 海拔 Altitude (m) | 经纬度 Longitude (E) and Latitude (N) |
|---|---|---|---|---|---|---|---|
| 701 | 凤城 Fengcheng | 185.6 | 123.95°, 40.42° | 747 | 桦甸 Huadian | 268.5 | 127.04°, 42.98° |
| 702 | 凤城 Fengcheng | 187.6 | 123.95°, 40.42° | 748 | 敦化 Dunhua | 495.0 | 128.22°, 43.38° |
| 703 | 凤城 Fengcheng | 185.4 | 123.95°, 40.42° | 749 | 敦化 Dunhua | 509.6 | 128.22°, 43.38° |
| 704 | 本溪 Benxi | 406.0 | 123.96°, 40.88° | 750 | 敦化 Dunhua | 517.3 | 128.22°, 43.38° |
| 705 | 本溪 Benxi | 405.8 | 123.96°, 40.88° | 751 | 敦化 Dunhua | 598.8 | 128.54°, 43.41° |
| 706 | 本溪 Benxi | 406.3 | 123.96°, 40.88° | 752 | 敦化 Dunhua | 599.1 | 128.54°, 43.41° |
| 707 | 桓仁 Huanren | 277.6 | 125.37°, 41.25° | 753 | 敦化 Dunhua | 595.3 | 128.54°, 43.41° |
| 708 | 桓仁 Huanren | 272.6 | 125.37°, 41.25° | 755 | 吉林 Jilin | 301.4 | 126.80°, 43.97° |
| 709 | 桓仁 Huanren | 283.3 | 125.37°, 41.25° | 756 | 吉林 Jilin | 302.2 | 126.80°, 43.97° |
| 710 | 新宾 Xinbin | 387.1 | 125.10°, 41.77° | 757 | 吉林 Jilin | 303.7 | 126.80°, 43.97° |
| 711 | 新宾 Xinbin | 416.5 | 125.10°, 41.77° | 758 | 吉林 Jilin | 200.0 | 126.55°, 43.85° |
| 712 | 新宾 Xinbin | 419.1 | 125.10°, 41.77° | 772 | 宁安 Ningan | 263.8 | 129.52°, 44.42° |
| 713 | 清原 Qingyuan | 315.7 | 124.81°, 42.34° | 773 | 宁安 Ningan | 262.2 | 129.52°, 44.42° |
| 714 | 清原 Qingyuan | 318.7 | 124.81°, 42.34° | 774 | 宁安 Ningan | 268.6 | 129.52°, 44.42° |
| 715 | 清原 Qingyuan | 315.7 | 124.81°, 42.34° | 775 | 宁安 Ningan | 279.6 | 129.52°, 44.42° |
| 724 | 沈阳 Shenyang | 60.0 | 123.57°, 41.82° | 776 | 尚志 Shangzhi | 327.6 | 127.55°, 45.27° |
| 729 | 沈阳 Shenyang | 58.0 | 123.55°, 41.82° | 777 | 鸡西 Jixi | 303.6 | 130.52°, 45.21° |
| 730 | 沈阳 Shenyang | 65.0 | 123.55°, 41.82° | 778 | 鸡西 Jixi | 282.6 | 130.54°, 45.18° |
| 741 | 抚松 Fusong | 461.9 | 127.20°, 42.34° | 779 | 鸡西 Jixi | 372.4 | 130.69°, 45.02° |
| 742 | 抚松 Fusong | 460.9 | 127.20°, 42.34° | 783 | 阿城 Acheng | 243.0 | 127.05°, 45.44° |
| 743 | 抚松 Fusong | 449.7 | 127.20°, 42.34° | 784 | 阿城 Acheng | 262.0 | 127.05°, 45.43° |
| 744 | 磐石 Panshi | 399.2 | 126.06°, 42.86° | 785 | 阿城 Acheng | 262.1 | 127.05°, 45.43° |
| 745 | 磐石 Panshi | 402.5 | 126.06°, 42.86° | 786 | 阿城 Acheng | 216.4 | 127.06°, 45.44° |
| 746 | 磐石 Panshi | 401.3 | 126.06°, 42.86° |
新窗口打开|下载CSV
1.3 表型性状观测内容与方法
参照《中国果树志: 杏卷》(张加延和张钊, 2003)和《山杏种质资源调查及评价技术规程》(吴月亮等, 2015), 对7个定性描述性状指标、22个定量描述性状指标进行观测(表2)。Table 2
表2
表2东北杏种质表型性状及其描述
Table 2
| 序号 No. | 性状 Traits | 表型性状描述 Description of phenotypic traits |
|---|---|---|
| 1 | 小枝色泽 Twig color | (1)绿色 Green (2)黄褐色 Tawny (3)灰褐色 Grayish-brown |
| 2 | 叶背被毛 Leaf back fuzzed | (1)有 Hairy (2)无 Glabrous |
| 3 | 叶表被毛 Leaf surface fuzzed | (1)有 Hairy (2)无 Glabrous |
| 4 | 叶基形状 Leaf base shape | (1)宽楔形 Wide wedge (2)圆形 Round (3)窄楔形 Narrow wedge |
| 5 | 叶缘形状 Leaf edge shape | (1)锐重锯齿 Sharp double sawtooth (2)钝重锯齿 Blunt double sawtooth |
| 6 | 腺体数量 Number of glands | (1) 1个 One (2) 2个 Two (3)无 Zero |
| 7 | 果实形状 Fruit shape | (1)球形 Sphericity (2)扁球形 Flat sphericity (3)扁长球形 Prolate sphericity (4)扁宽球形 Flat and broad sphericity (5)长球形 Long sphericity |
| 8 | 树高 Tree height | 树木从根颈到树梢之间的距离 The distance between the treetop and the root neck of tree |
| 9 | 胸径 Diameter at breast height | 树干1.3 m高度处的直径 Diameter of the trunk at 1.3 m height |
| 10 | 冠幅 Crown diameter | 树冠南北和东西方向宽度的平均值 The average value of the north-south and east-west width of tree crown |
| 11 | 主枝基角 Main branch base angle | 主枝与中心干之间的分枝角度 Angle between the main branch and the center stem of tree |
| 12 | 小枝长度 Twig length | 一年生枝平均长度 Average length of annual branches |
| 13 | 小枝粗度 Twig width | 一年生枝平均粗度 Average width of annual branches |
| 14 | 叶长 Leaf length | 从叶基切线至叶尖顶部的长度 Length from the tangent of leaf base to the top of leaf tip |
| 15 | 叶宽 Leaf width | 叶片最宽处的长度 The length of the blade at its widest point |
| 16 | 叶柄长 Petiole length | 叶柄的长度 Length of petiole |
| 17 | 单果质量 Single fruit mass | 单个成熟果实的质量 The mass of single ripe fruit |
| 18 | 果长 Fruit length | 成熟果实从顶部到底部的最大距离 Maximum distance from top to bottom of ripe fruit |
| 19 | 果宽 Fruit width | 成熟果实两条缝合线之间的最大距离 Maximum distance between two sutures of ripe fruit |
| 20 | 果厚 Fruit thickness | 成熟果实腹面观时的最大距离 Maximum distance from the ventral view of ripe fruit |
| 21 | 果柄长 Fruit handle length | 果柄的长度 Length of fruit handle |
| 22 | 种子质量 Seed mass | 单个成熟种子的质量 The mass of single ripe seed |
| 23 | 种子长 Seed length | 成熟种子从顶部到底部的最大距离 Maximum distance from top to bottom of ripe seed |
| 24 | 种子宽 Seed width | 成熟种子脊背到缝翅的最大距离 Maximum distance from ridge to sewed wing of ripe seed |
| 25 | 种子厚 Seed thickness | 成熟种子腹面观时的最大距离 Maximum distance from the ventral view of ripe seed |
| 26 | 种仁质量 Kernel mass | 单个成熟种仁的质量 The mass of single mature kernel |
| 27 | 种仁长 Kernel length | 成熟种仁从顶部到底部的最大距离 Maximum distance from top to bottom of ripe kernel |
| 28 | 种仁宽 Kernel width | 成熟种仁两条缝合线之间的最大距离 Maximum distance between two sutures of ripe kernel |
| 29 | 种仁厚 Kernel thickness | 成熟种仁腹面观时的最大距离 Maximum distance from the ventral view of ripe kernel |
新窗口打开|下载CSV
观测指标包括树体、小枝、叶片、果、种子、种仁6个方面。树体性状: 测量树高、胸径、冠幅、主枝基角; 小枝性状: 选择种质东、南、西、北4个方向的外围中、上部当年生枝条, 测量其长度、粗度, 记载其色泽; 叶片性状: 选择种质东、南、西、北4个方向的当年生枝条各1个, 从梢端数第5-7片成熟叶, 测量其叶长、叶宽、叶柄长, 记载叶表、叶背被毛情况, 叶基形状, 叶缘形状和腺体数量; 果实性状: 果实成熟后, 在树体东、南、西、北4个方向的小枝上各选取10-20个健康、无病虫害的果实, 分别测量果长、果宽、果厚、单果质量、果柄长, 记载果实形状; 种子性状: 测量种子长、种子宽、种子厚、种子质量; 种仁性状: 测量种仁长、种仁宽、种仁厚、种仁质量。
1.4 SSR分子标记试验
在2016年6月中旬, 采集47份供试东北杏种质向阳面一年生枝条上的叶片, 放入液氮罐中速冻, 带回实验室, 置于-80 ℃超低温冰箱中备用。试验步骤包括DNA提取、SSR引物筛选、PCR反应体系及扩增程序、聚丙烯酰胺凝胶电泳检测及数据的读取, 具体参照金玲等(2018)的方法。
1.5 数据统计分析
使用SPSS 20.0软件对所得表型性状数据进行统计与分析, 分别计算47份供试东北杏种质定量描述性状的平均值($\bar{X}$)、标准偏差(SD)、极小值(Min)、极大值(Max)、变异系数(CV = SD/$\bar{X}$)和K-S值, 并对定性描述性状进行频率统计、计算Shannon- Wiener指数(H′), H′的计算参照王永康等(2014)的方法。通过Pearson系数分析法研究定量描述性状与地理位置之间的相关关系, 采用R 3.3.2软件对定量描述性状进行多项式线性回归分析, 使用Surfer软件分析定量描述性状的地理变化规律。基于26个表型性状, 运用R 3.3.2对47份东北杏种质进行主成分分析。基于主成分得分值, 利用平方欧氏距离, 采用离差平方和法(Ward)进行系统聚类。使用Cervus 3.0软件计算SSR标记各位点的多态性信息含量(PIC = $1-\sum{{{P}_{j}}^{2}}$, 其中Pj2为第j个基因的频率); 按照Nei和Li (1979)的方法计算种质间的遗传相似系数(GS): GS = 2Nij/(Ni + Nj), 式中Ni和Nj为i和j种质的谱带数, Nij为i和j种质共有的谱带数。基于遗传相似系数, 利用NTSYS 2.1软件, 采用非加权组平均法(UPGMA)对47份东北杏种质进行聚类分析。
2 结果和分析
2.1 东北杏种质资源表型性状多样性
2.1.1 定性描述性状频率分布及多样性指数对东北杏种质的7个定性描述性状进行频率统计及变异分析(表3), 由表可知, 小枝色泽以绿色为主(80.85%); 叶背、叶表均以被毛为主, 分布频率分别为74.47%和68.09%; 叶基形状以宽楔形为主(74.47%); 叶缘可分为锐重锯齿和钝重锯齿, 两种类型分布频率相差不大, 分别为51.06%和48.94%; 叶柄腺体数量分为1个(34.04%)、2个(40.43%)和无(25.53%)三类, 其中叶柄具有2个腺体的种质较多; 果实以扁球形为主(53.19%)。
Table 3
表3
表3东北杏种质定性描述性状频率分布及多样性指数
Table 3
| 序号 No. | 性状 Traits | 频率分布 Frequency distribution (%) | Shannon-Wiener 多样性指数 Shannon-Wiener diversity index | ||||
|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | |||
| 1 | 小枝色泽 Twig color | 80.85 | 14.89 | 4.26 | 0.59 | ||
| 2 | 叶背被毛 Leaf back fuzzed | 74.47 | 25.53 | 0.57 | |||
| 3 | 叶表被毛 Leaf surface fuzzed | 68.09 | 31.91 | 0.63 | |||
| 4 | 叶基形状 Leaf base shape | 74.47 | 23.40 | 2.13 | 0.64 | ||
| 5 | 叶缘形状 Leaf edge shape | 51.06 | 48.94 | 0.69 | |||
| 6 | 腺体数量 Number of glands | 34.04 | 40.43 | 25.53 | 1.08 | ||
| 7 | 果实形状 Fruit shape | 23.41 | 53.19 | 14.89 | 6.38 | 2.13 | 1.22 |
新窗口打开|下载CSV
从Shannon-Wiener多样性指数来看, 不同定性描述性状多样性之间差异明显, 范围在0.57-1.22之间。其中, 果实形状、腺体数量多样性指数较大, 分别为1.22和1.08 (表3)。
2.1.2 定量描述性状的分布特征及变异程度
表4为东北杏19个定量描述性状的K-S正态性分布检验和变异程度分析结果, 树高、胸径和冠幅3个定量描述性状受树龄影响较大, 未进行相应的统计分析。由表4可知, 除了果柄长度外, 其余18个定量描述性状的p值均大于0.05, 表明这些性状呈正态 分布。
Table 4
Table 4Basic statistics and K-S test of the quantitative traits of Armeniaca mandshurica germplasms.
| 性状 Traits | 极小值 Min | 极大值 Max | 平均值 $\bar{X}$ | 标准偏差 SD | 变异系数 CV | K-S值 K-S value | 显著性 p |
|---|---|---|---|---|---|---|---|
| 主枝基角 MBBA (°) | 18.00 | 51.00 | 33.60 | 7.04 | 0.2096 | 0.833 | 0.491 |
| 小枝长度 TL (cm) | 10.23 | 120.00 | 59.82 | 33.49 | 0.5598 | 0.622 | 0.835 |
| 小枝粗度 TW (mm) | 1.19 | 3.18 | 2.02 | 0.58 | 0.2868 | 0.897 | 0.397 |
| 叶长 LL (mm) | 76.59 | 122.53 | 98.08 | 11.20 | 0.1142 | 0.822 | 0.509 |
| 叶宽 LW (mm) | 44.96 | 78.46 | 62.36 | 7.34 | 0.1177 | 0.289 | 1.000 |
| 叶柄长 PL (mm) | 18.48 | 45.29 | 30.25 | 6.19 | 0.2048 | 0.745 | 0.636 |
| 单果质量 SFM (g) | 1.83 | 8.70 | 4.44 | 1.55 | 0.3477 | 0.971 | 0.302 |
| 果长 FL (mm) | 15.17 | 24.93 | 20.38 | 2.31 | 0.1133 | 0.447 | 0.988 |
| 果宽 FW (mm) | 12.92 | 26.16 | 19.92 | 2.72 | 0.1365 | 1.031 | 0.238 |
| 果厚 FT (mm) | 10.65 | 24.06 | 17.49 | 2.89 | 0.1651 | 0.816 | 0.518 |
| 种子质量 SM (g) | 0.58 | 1.45 | 1.01 | 0.24 | 0.2352 | 0.736 | 0.650 |
| 种子长 SL (mm) | 13.11 | 24.01 | 16.51 | 1.95 | 0.1181 | 1.052 | 0.218 |
| 种子宽 SW (mm) | 11.57 | 20.87 | 14.46 | 1.57 | 0.1085 | 0.914 | 0.374 |
| 种子厚 ST (mm) | 8.66 | 15.72 | 10.34 | 1.16 | 0.1121 | 0.981 | 0.291 |
| 种仁质量 KM (g) | 0.14 | 0.40 | 0.29 | 0.06 | 0.1936 | 0.572 | 0.898 |
| 种仁长 KL (mm) | 8.65 | 13.59 | 11.27 | 1.06 | 0.0944 | 0.793 | 0.556 |
| 种仁宽 KW (mm) | 6.68 | 11.11 | 8.59 | 0.81 | 0.0940 | 0.886 | 0.412 |
| 种仁厚 KT (mm) | 4.21 | 7.69 | 5.71 | 0.82 | 0.1439 | 0.520 | 0.950 |
| 果柄长 FHL (mm) | 3.24 | 14.52 | 7.04 | 2.87 | 0.4071 | 1.503 | 0.022 |
新窗口打开|下载CSV
由表4可以看出, 19个定量描述性状中, 小枝长度、果柄长和单果质量的变异系数较大, 分别为55.98%、40.71%和34.77%, 种仁质量、叶柄长、主枝基角、种子质量、小枝粗度的变异系数次之, 在19.36%-28.68%之间; 其他11个指标变异系数较小, 在9.40%-16.51%之间。由此可见, 东北杏定量描述性状指标变异较大, 体现出较高的遗传多样性。
2.2 东北杏种质资源定量描述性状的地理变化
2.2.1 调查区域地理位置与气候因子的相关性15个调查区域的经度、纬度及海拔与气候因子(气候指标为1981-2010年的平均值, 数据来自中国气象数据网data.cma.cn)的相关性分析(表5)总结如下: 经度与年均气温、7月平均最高气温、1月平均最低气温及平均年降水量均呈负相关关系, 其中与年平均气温、7月平均最低气温及平均年降水量的相关性达极显著水平(p < 0.01), 与1月平均最高气温的相关性达显著水平(p < 0.05); 纬度与年平均气温、1月平均最低气温及平均年降水量均呈极显著负相关关系(p < 0.01); 海拔与年均气温、7月平均最高气温、1月平均最低气温及平均年降水量均呈负相关关系, 其中与年平均气温、1月平均最高气温的相关性达显著水平(p < 0.05), 与7月平均最高气温的相关性达极显著水平(p < 0.01)。
Table 5
表5
表5调查区域地理位置与气候因子的相关性
Table 5
| 指标 Index | 年均气温 AAT (℃) | 7月平均最高气温 JAMT1 (℃) | 1月平均最低气温 JAMT2 (℃) | 年降水量 AAP (mm) | ||||
|---|---|---|---|---|---|---|---|---|
| 相关系数 R | 显著性 p | 相关系数 R | 显著性 p | 相关系数 R | 显著性 p | 相关系数 R | 显著性 p | |
| 经度 Longitude | -0.810** | <0.001 | -0.649** | 0.009 | -0.589* | 0.021 | -0.693** | 0.004 |
| 纬度 Latitude | -0.785** | 0.001 | -0.322 | 0.242 | -0.656** | 0.008 | -0.864** | <0.001 |
| 海拔 Altitude | -0.633* | 0.011 | -0.778** | 0.001 | -0.529* | 0.043 | -0.132 | 0.639 |
新窗口打开|下载CSV
2.2.2 定量描述性状与地理位置的相关性
对47份供试东北杏种质地理分布的纬度、经度及海拔与定量描述性状指标(树高、胸径和冠幅3个指标受树龄影响较大, 故未参与相关分析)进行相关性分析(表6), 结果表明: 19个定量描述性状中,主枝基角与纬度呈显著正相关关系(p < 0.05); 小枝长度与经度、纬度显著负相关(p < 0.05), 而与海拔显著正相关(p < 0.05); 小枝粗度与经度、纬度的相关性均为显著负相关(p < 0.05); 种子质量与经度呈显著负相关关系(p < 0.05), 与纬度极显著负相关(p < 0.01); 果柄长与经度、纬度均呈极显著负相关关系(p < 0.01); 叶宽与经度显著负相关(p < 0.05)。其他指标与经度、纬度及海拔没有表现出显著的相关性(p > 0.05)。
Table 6
表6
表6东北杏种质定量描述性状与地理位置的相关性
Table 6
| 性状 Traits | 经度 Longitude (E) | p | 纬度 Latitude (N) | p | 海拔 Altitude (m) | p |
|---|---|---|---|---|---|---|
| 主枝基角 Main branch base angle | 0.277 | 0.060 | 0.351* | 0.016 | 0.043 | 0.774 |
| 小枝长度 Twig length | -0.307* | 0.036 | -0.370* | 0.011 | 0.289* | 0.049 |
| 小枝粗度 Twig width | -0.330* | 0.023 | -0.341* | 0.019 | -0.274 | 0.062 |
| 叶长 Leaf length | 0.121 | 0.417 | 0.081 | 0.589 | 0.100 | 0.502 |
| 叶宽 Leaf width | -0.305* | 0.037 | -0.226 | 0.127 | -0.109 | 0.466 |
| 叶柄长 Petiole length | -0.088 | 0.555 | -0.138 | 0.354 | 0.176 | 0.236 |
| 单果质量 Single fruit mass | -0.181 | 0.222 | 0.130 | 0.383 | -0.214 | 0.149 |
| 果长 Fruit length | 0.053 | 0.721 | 0.264 | 0.073 | -0.111 | 0.459 |
| 果宽 Fruit width | -0.243 | 0.100 | 0.036 | 0.810 | -0.238 | 0.106 |
| 果厚 Fruit thickness | -0.167 | 0.261 | 0.144 | 0.336 | -0.254 | 0.085 |
| 种子质量 Seed mass | -0.348* | 0.016 | -0.447** | 0.002 | -0.013 | 0.929 |
| 种子长 Seed length | 0.163 | 0.273 | 0.080 | 0.594 | 0.200 | 0.177 |
| 种子宽 Seed width | -0.195 | 0.190 | -0.188 | 0.205 | 0.084 | 0.576 |
| 种子厚 Seed thickness | -0.106 | 0.479 | -0.073 | 0.625 | 0.167 | 0.261 |
| 种仁质量 Kernel mass | -0.080 | 0.592 | 0.037 | 0.805 | -0.163 | 0.273 |
| 种仁长 Kernel length | 0.043 | 0.776 | -0.073 | 0.625 | 0.067 | 0.656 |
| 种仁宽 Kernel width | -0.146 | 0.327 | -0.099 | 0.510 | -0.178 | 0.232 |
| 种仁厚 Kernel thickness | -0.019 | 0.900 | 0.102 | 0.495 | 0.053 | 0.721 |
| 果柄长 Fruit handle length | -0.434** | 0.002 | -0.539** | <0.001 | 0.234 | 0.114 |
新窗口打开|下载CSV
2.2.3 定量描述性状与种质产地经度、纬度及海拔的多项式回归分析
上述分析表明, 主枝基角、小枝长度、小枝粗度、叶宽、种子质量和果柄长与种质所处地理位置显著相关, 采用R软件分别将这些指标与经度、纬度及海拔进行多项式线性回归分析(表7)。由表7可知, 小枝长度、小枝粗度、种子质量、果柄长的拟合系数较高且均达到了显著水平(p < 0.05), 可以揭示其地理变化规律; 而主枝基角和果长的拟合度不够, 不能揭示其地理变化规律。通过表中p值大小可知: 海拔对小枝长度和果柄长的影响达显著水平(p < 0.05); 纬度、经度和海拔对小枝粗度、种子质量的影响均未达显著水平(p > 0.05)。
Table 7
表7
表7东北杏种质定量描述性状与地理位置多项式回归分析
Table 7
| 性状 Traits | 多项式变量 Polynomial variable | 分项系数 Coefficient | 标准误差 Standard Error | 系数显著性 p of coefficient | 方程显著性 p of equation | 拟合系数 C |
|---|---|---|---|---|---|---|
| 主枝基角 Main branch base angle | 常数项 Constant term | -24.139 | 81.028 | 0.767 | 0.123 | 0.124 |
| 纬度 Latitude | 1.746 | 1.186 | 0.148 | |||
| 经度 Longitude | -0.143 | 0.950 | 0.881 | |||
| 海拔 Altitude | 0.002 | 0.009 | 0.808 | |||
| 小枝长度 Twig length | 常数项 Constant term | 949.078 | 349.745 | 0.010 | 0.003 | 0.278 |
| 纬度 Latitude | -1.059 | 5.121 | 0.837 | |||
| 经度 Longitude | -6.967 | 4.099 | 0.096 | |||
| 海拔 Altitude | 0.115 | 0.040 | 0.006 | |||
| 小枝粗度 Twig width | 常数项 Constant term | 5.605 | 6.419 | 0.387 | 0.030 | 0.186 |
| 纬度 Latitude | -0.151 | 0.094 | 0.115 | |||
| 经度 Longitude | 0.026 | 0.075 | 0.728 | |||
| 海拔 Altitude | -0.001 | 0.001 | 0.082 | |||
| 叶宽 Leaf width | 常数项 Constant term | 210.474 | 85.886 | 0.018 | 0.232 | 0.094 |
| 纬度 Latitude | 0.261 | 1.257 | 0.836 | |||
| 经度 Longitude | -1.261 | 1.007 | 0.217 | |||
| 海拔 Altitude | 0.001 | 0.010 | 0.925 | |||
| 种子质量 Seed mass | 常数项 Constant term | 3.809 | 2.608 | 0.151 | 0.021 | 0.200 |
| 纬度 Latitude | -0.071 | 0.038 | 0.070 | |||
| 经度 Longitude | 0.002 | 0.031 | 0.950 | |||
| 海拔 Altitude | 1.853 × 10-6 | <0.001 | 0.995 | |||
| 果柄长 Fruit handle length | 常数项 Constant term | 87.172 | 27.568 | 0.003 | <0.001 | 0.388 |
| 纬度 Latitude | -0.528 | 0.404 | 0.198 | |||
| 经度 Longitude | -0.475 | 0.323 | 0.149 | |||
| 海拔 Altitude | 0.008 | 0.003 | 0.012 |
新窗口打开|下载CSV
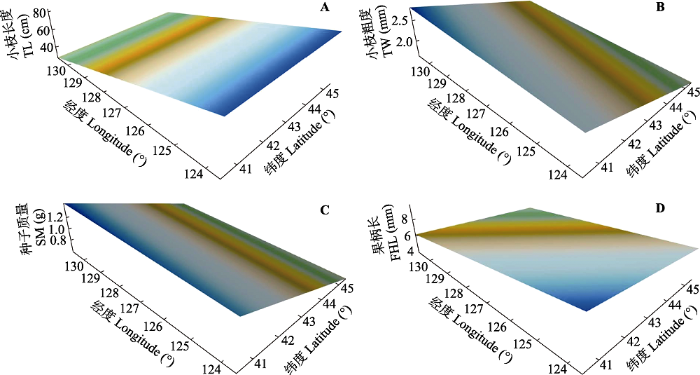
将供试种质所处的海拔平均值代入小枝长度、小枝粗度、种子质量和果柄长的回归方程中(廖柏勇等, 2016), 采用Surfer 12.0软件分别做经、纬度趋势面图(图1)。由图可知: 小枝长度呈由东向西逐渐增加的梯度变化, 小枝粗度呈由北到南逐渐增加的梯度变化, 种子质量呈由北到南逐渐增加的梯度变化, 果柄长呈由东北到西南逐渐增加的梯度变化。
图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1东北杏种质定量描述性状与经纬度趋势面图。A, 小枝长度。B, 小枝粗度。C, 种子质量。D, 果柄长。
Fig. 1Trend surface analyses between the quantitative traits and the geographical coordinates (i.e. latitude and longitude) of Armeniaca mandshurica germplasms. A, Trend surface map between twig length (TL), latitude and longitude. B, Trend surface map between twig width (TW), latitude and longitude. C, Trend surface map between seed mass (SM), latitude and longitude. D, Trend surface map between fruit handle length (FHL), latitude and longitude.
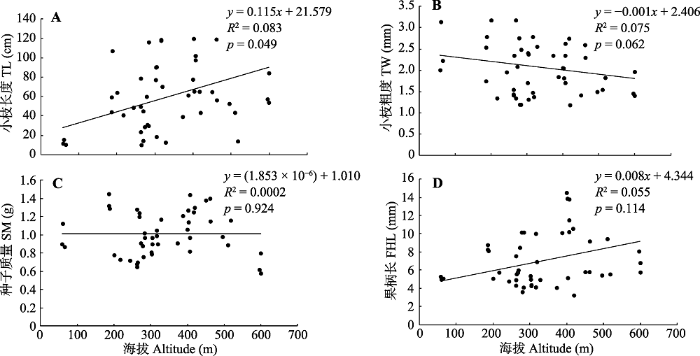
再将平均经度和平均纬度代入回归方程, 采用Excel 2010分别做海拔趋势曲线图(图2)。由图2可知: 小枝长度、果柄长与海拔呈正相关关系, 随海拔升高而增加; 小枝粗度与海拔呈负相关关系, 随海拔升高而减小; 种子质量与海拔相关性很小。
图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2东北杏种质定量描述性状与海拔趋势曲线图。A, 小枝长度。B, 小枝粗度。C, 种子质量。D, 果柄长。
Fig. 2Relationships between quantitative traits and altitude of Armeniaca mandshurica germplasms. A, Trend curve between twig length (TL) and altitude. B, Trend curve between twig width (TW) and altitude. C, Trend curve between seed mass (SM) and altitude. D, Trend curve between fruit handle length (FHL) and altitude.
2.3 基于表型性状的东北杏种质分类
本文根据19个定量描述性状和7个定性描述性状对47份东北杏种质进行主成分分析, 按照特征值大于1进行抽取(因树高、胸径和冠幅3个指标受树龄影响较大, 故未参与主成分分析)。表8列出了各个指标对应的方差分量(载荷值的平方)及各主成分对应的特征根和贡献率。Table 8
表8
表8东北杏种质表型性状的主成分分析
Table 8
| 性状 Traits | 主成分 Principal component | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| PC1 | PC2 | PC3 | PC4 | PC5 | PC6 | PC7 | PC8 | PC9 | PC10 | |
| 主枝基角 Main branch base angle | 0.005 | 0.009 | 0.282 | 0.008 | 0.097 | 0.007 | 0.091 | 0.239 | 0.009 | 0.015 |
| 小枝长度 Twig length | 0.001 | 0.002 | 0.450 | 0.000 | 0.174 | 0.027 | 0.019 | 0.076 | 0.052 | 0.001 |
| 小枝粗度 Twig width | 0.008 | 0.004 | 0.004 | 0.290 | 0.144 | 0.064 | 0.120 | 0.003 | 0.004 | 0.005 |
| 叶长 Leaf length | 0.000 | 0.049 | 0.337 | 0.282 | 0.012 | 0.019 | 0.041 | 0.004 | 0.061 | 0.006 |
| 叶宽 Leaf width | 0.000 | 0.011 | 0.108 | 0.120 | 0.074 | 0.019 | 0.093 | 0.003 | 0.407 | 0.000 |
| 叶柄长 Petiole length | 0.006 | 0.216 | 0.112 | 0.068 | 0.008 | 0.164 | 0.002 | 0.007 | 0.021 | 0.056 |
| 单果质量 Single fruit mass | 0.591 | 0.139 | 0.001 | 0.004 | 0.009 | 0.009 | 0.007 | 0.001 | 0.003 | 0.019 |
| 果长 Fruit length | 0.659 | 0.006 | 0.053 | 0.023 | 0.000 | 0.141 | 0.003 | 0.008 | 0.011 | 0.002 |
| 果宽 Fruit width | 0.648 | 0.129 | 0.022 | 0.006 | 0.039 | 0.000 | 0.026 | 0.004 | 0.000 | 0.006 |
| 果厚 Fruit thickness | 0.665 | 0.184 | 0.020 | 0.011 | 0.028 | 0.001 | 0.001 | 0.032 | 0.001 | 0.000 |
| 种子质量 Seed mass | 0.455 | 0.192 | 0.074 | 0.036 | 0.006 | 0.002 | 0.008 | 0.001 | 0.006 | 0.003 |
| 种子长 Seed length | 0.337 | 0.283 | 0.010 | 0.047 | 0.058 | 0.097 | 0.000 | 0.075 | 0.001 | 0.031 |
| 种子宽 Seed width | 0.476 | 0.145 | 0.009 | 0.014 | 0.006 | 0.094 | 0.050 | 0.039 | 0.020 | 0.001 |
| 种子厚 Seed thickness | 0.449 | 0.096 | 0.002 | 0.012 | 0.049 | 0.172 | 0.004 | 0.046 | 0.001 | 0.020 |
| 种仁质量 Kernel mass | 0.461 | 0.005 | 0.104 | 0.084 | 0.040 | 0.014 | 0.026 | 0.029 | 0.008 | 0.000 |
| 种仁长 Kernel length | 0.334 | 0.178 | 0.032 | 0.016 | 0.033 | 0.180 | 0.001 | 0.015 | 0.000 | 0.035 |
| 种仁宽 Kernel width | 0.319 | 0.046 | 0.058 | 0.153 | 0.105 | 0.064 | 0.008 | 0.038 | 0.006 | 0.020 |
| 种仁厚 Kernel thickness | 0.066 | 0.023 | 0.186 | 0.047 | 0.085 | 0.130 | 0.116 | 0.000 | 0.023 | 0.072 |
| 果柄长 Fruit handle length | 0.016 | 0.033 | 0.112 | 0.052 | 0.377 | 0.043 | 0.037 | 0.014 | 0.016 | 0.015 |
| 小枝色泽 Twig color | 0.227 | 0.205 | 0.090 | 0.023 | 0.134 | 0.139 | 0.149 | 0.159 | 0.016 | 0.168 |
| 叶背被毛 Leaf back fuzzed | 0.014 | 0.056 | 0.016 | 0.116 | 0.069 | 0.028 | 0.186 | 0.006 | 0.099 | 0.146 |
| 叶表被毛 Leaf surface fuzzed | 0.000 | 0.212 | 0.155 | 0.177 | 0.041 | 0.051 | 0.007 | 0.047 | 0.054 | 0.000 |
| 叶基形状 Leaf base shape | 0.030 | 0.273 | 0.017 | 0.205 | 0.238 | 0.250 | 0.304 | 0.156 | 0.069 | 0.044 |
| 叶缘形状 Leaf edge shape | 0.001 | 0.262 | 0.214 | 0.122 | 0.002 | 0.001 | 0.019 | 0.088 | 0.049 | 0.010 |
| 腺体数量 Number of glands | 0.068 | 0.230 | 0.037 | 0.263 | 0.098 | 0.021 | 0.098 | 0.129 | 0.137 | 0.055 |
| 果实形状 Fruit shape | 0.210 | 0.388 | 0.141 | 0.163 | 0.317 | 0.345 | 0.424 | 0.151 | 0.205 | 0.398 |
| 特征根 Eigenvalue | 6.047 | 3.376 | 2.644 | 2.345 | 2.244 | 2.083 | 1.841 | 1.369 | 1.280 | 1.127 |
| 贡献率(%) Contribution rate | 18.896 | 10.549 | 8.261 | 7.329 | 7.013 | 6.510 | 5.754 | 4.279 | 4.001 | 3.521 |
| 累积贡献率(%) Cumulative contribution rate | 18.896 | 29.445 | 37.706 | 45.035 | 52.048 | 58.558 | 64.312 | 68.591 | 72.592 | 76.113 |
新窗口打开|下载CSV
由表8可知, 前10个主成分累计贡献率为76.113%。第一主成分中单果质量、果长、果宽、果厚、种子质量、种子长、种子宽、种子厚、种仁质量、种仁长、种仁宽的方差分量较大, 可看作是反映果、核、仁大小的综合指标。第二主成分中叶基形状、叶缘形状的方差分量较大, 可作为叶片形态特征的综合指标。第三主成分中主枝基角、小枝长度、叶长具有较大的方差分量, 即第三主成分主要代表了树体特征。第四主成分中小枝粗度、腺体数量具有较大的方差分量。第五主成分中果柄长的方差分量较大。第七主成分中果实形状具有较大的方差分量。第九主成分中叶宽的方差分量较大。其余主成分指示作用不明显。
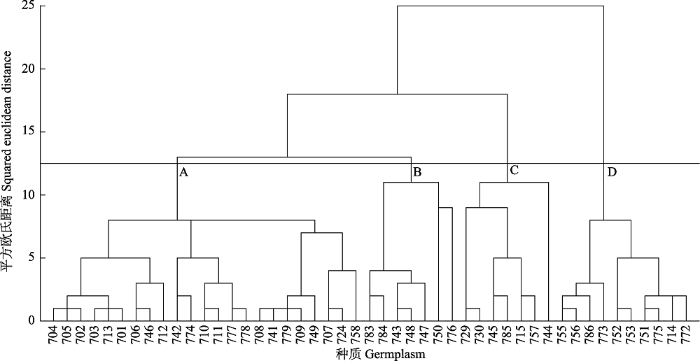
基于47份东北杏种质的主成分得分值, 运用Ward法进行系统聚类, 结果如图3所示。
图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3基于表型性状的东北杏种质聚类结果。
Fig. 3Clustering result of Armeniaca mandshurica germplasms based on phenotypic traits.
在横坐标12.5处划线, 将种质划分为四类(分别用A、B、C、D表示)。表9为各类主要定量描述性状。
Table 9
表9
表9东北杏种质各类别主要定量描述性状
Table 9
| 类别 Categories | 单果质量 SFM (g) | 种子质量 SM (g) | 种仁质量 SM (g) | 小枝长度 TL (cm) | 小枝粗度 TW (mm) | 主枝基角 MBBA (°) | 叶长 LL (mm) | 叶宽 LW (mm) | 叶柄长 PL (mm) | 果柄长 FHL (mm) |
|---|---|---|---|---|---|---|---|---|---|---|
| A | 4.35 (0.89) | 1.11 (0.21) | 0.30 (0.05) | 67.80 (34.75) | 2.25 (0.58) | 30.61 (6.37) | 97.92 (9.43) | 63.25 (7.02) | 31.58 (6.75) | 8.02 (3.15) |
| B | 5.27 (1.84) | 1.04 (0.26) | 0.28 (0.42) | 36.80 (22.20) | 1.98 (0.66) | 39.86 (6.47) | 101.79 (13.00) | 63.80 (4.65) | 31.09 (6.20) | 5.23 (0.56) |
| C | 6.01 (1.95) | 1.03 (0.15) | 0.32 (0.40) | 57.00 (42.44) | 1.76 (0.36) | 35.57 (7.91) | 97.22 (12.64) | 60.10 (7.68) | 27.01 (5.61) | 7.43 (6.38) |
| D | 2.99 (0.81) | 0.73 (0.10) | 0.24 (0.06) | 59.55 (25.85) | 1.68 (0.43) | 34.70 (5.19) | 96.45 (13.82) | 60.91 (9.54) | 28.87 (4.77) | 5.78 (1.24) |
| 群体平均值 Average value of population | 4.65 (1.30) | 0.98 (0.17) | 0.29 (0.03) | 55.29 (13.15) | 1.92 (0.26) | 35.18 (3.79) | 98.35 (2.37) | 62.02 (1.79) | 29.64 (2.11) | 6.61 (1.32) |
新窗口打开|下载CSV
A类包括23份种质, 该类总体特征为主枝基角小, 小枝长且粗; 果柄长, 果大小中等, 种子大, 种仁较大; 叶柄长, 叶片大小中等。叶基均为宽楔形,大部分种质小枝为绿色(701和708号种质除外)。种质产地以调查区域的南部为主, 其中14份种质位于辽宁东部地区(704、705、702、703、713、701、706、712、710、711、708、709、707、724), 5份种质位于吉林东部地区(746、742、741、749、758), 4份种质位于黑龙江东部地区(774、777、778、779)。
B类包括7份种质, 该类总体特征为主枝基角大, 小枝短、较粗; 叶柄长, 叶片大; 果柄短, 果较大, 种子、种仁大小中等。叶背均被毛, 大部分种质叶缘为锐重锯齿(783号种质除外)。种质产地为调查区域的中部和北部, 其中4份种质位于吉林东部地区(743、748、747、750), 3份种质位于黑龙江东部地区(783、784、776)。
C类包括7份种质, 该类总体特征为主枝基角中等, 小枝细、长度中等; 叶柄短, 叶片小; 果柄较长, 果大, 种仁大, 种子大小中等; 多数种质叶缘为钝重锯齿且叶柄具2个腺体(785号种质除外)。种质产地以调查区域的中部和南部为主, 其中3份种质位于辽宁东部地区(729、730、715), 3份种质位于吉林东部地区(745、757、744), 1份种质位于黑龙江东部地区(785)。
D类包括10份种质, 该类总体特征为主枝基角中等, 小枝细、较长; 叶柄长度中等, 叶片小; 果柄短, 果、种子、种仁小。所有种质小枝均为绿色。种质产地以调查区域的中部和北部为主, 其中3份种质位于黑龙江东部地区(786、773、775), 5份种质位于吉林东部地区(755、756、752、753、751), 1份种质位于辽宁东部地区(714)。
上述分类结果主要体现了东北杏种质特征的差异, 在一定程度上也体现了种质产地的效应。
2.4 表型聚类和SSR分子标记聚类的相关分析
根据沈阳农业大学山杏课题组2014年进行的山杏简化基因组测序结果, 由北京赛百盛基因技术有限公司合成600对SSR引物, 从中筛选出99对扩增条带清晰、多态性高的引物, 对47份东北杏种质的基因组DNA进行扩增, 平均每个位点的等位基因数为8.879个, 多态性信息含量为0.718 (各位点信息详见附录Ⅰ)。基于99个SSR位点数据, 分析47份东北杏种质间的遗传相似性, 结果表明, 遗传相似系数在0.726-0.934之间, 平均为0.797, 遗传差异较小, 亲缘关系较近。以遗传相似系数为基础, 采用UPGMA法进行聚类分析, 在相似系数0.83处可以分为4大类(图4)。图4
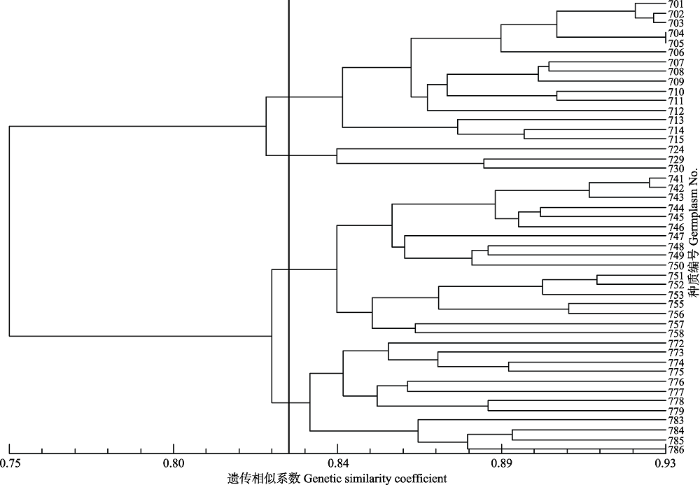 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4基于SSR分子标记的东北杏种质聚类结果。种质编号同
Fig. 4Clustering result of Armeniaca mandshurica germplasms based on SSR molecular markers. Germplasm No. see
第1大类包括15份种质(701、702、703、704、705、706、707、708、709、710、711、712、713、714、715), 均来自于辽宁东部地区, 即调查区域的东南部; 第2大类包括3份种质(724、729、730), 均来自于辽宁沈阳市, 即调查区域的西南部; 第3大类包括17份种质(741、742、743、744、745、746、747、748、749、750、751、752、753、755、756、757、758), 均来自于吉林东部地区, 即调查区域的中部; 第4大类包括12份种质(772、773、774、775、776、777、778、779、783、784、785、786), 均来自于黑龙江东部地区, 即调查区域的北部。聚类结果反映了种质产地的效应。
为了探讨表型聚类与SSR分子标记聚类结果的关系, 进行卡方检验(表10), 结果表明, 两种聚类结果的相关性未达到显著水平(p > 0.05)。
Table 10
表10
表10东北杏种质基于表型性状和SSR分子标记聚类结果间的相关性
Table 10
| 表型性状聚类 Clustering based on phenotypic traits | SSR分子标记聚类 Clustering based on SSR molecular markers | χ2 | p | |||
|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | |||
| 1 | 13 | 1 | 5 | 4 | 3.250a | 0.777 |
| 2 | 0 | 0 | 4 | 3 | ||
| 3 | 1 | 2 | 3 | 1 | ||
| 4 | 1 | 0 | 5 | 3 | ||
新窗口打开|下载CSV
3 讨论
3.1 东北杏种质表型性状呈现出遗传多样性
遗传参数中变异系数可在一定程度上反映性状的离散程度, 变异系数越大, 离散程度越高, 多样性程度越高(柳江群等, 2017), 一般情况下变异系数大于10%, 说明该性状在不同种质间的差异较大, 多样性丰富(白史且等, 2002); Shannon-Wiener多样性指数是用来描述不同性状出现的紊乱和不确定性,不确定性越高, 多样性也就越高(孙儒泳, 2002)。本研究基于47份东北杏种质的26个表型性状进行统计分析, 19个定量描述性状指标的变异系数在9.40%- 55.98%之间, 7个定性描述性状指标的Shannon- Wiener多样性指数在0.57-1.22之间, 表现出较高的遗传多样性。其中小枝长度、果柄长和单果质量的变异系数较大, 分别为55.98%、40.71%和34.77%; 种仁质量、叶柄长、主枝基角、种子质量、小枝粗度的变异系数次之, 在19.36%-28.68%之间。雷鸣雷等(2012)研究认为, 宁夏彭阳县野杏变异系数较大的指标为嫩枝长、雌蕊发育程度、果质量、节间长度、出核率、仁质量、核质量, 其大小分别为39.8%、38.4%、30.2%、30.1%、26.2%、26.2%和23.9%, 本研究结果与之基本一致。仲维平等(2015)研究表明, 内蒙古扎兰屯地区山杏(Armeniaca sibirica)的嫩枝长、节间长度、叶柄长、完全花比例、嫩枝粗、叶宽、叶长/叶柄长、核质量的变异系数均超过20%, 本研究结果在小枝性状、叶柄性状及种子大小方面与之一致。与上述文献不同的是, 本研究中东北杏在果柄长、主枝基角上也呈现出较大的变异, 这是由树种生物学特性和研究区域的自然条件特点所决定的。对19个主要定量描述性状进行K-S正态性检验的结果表明, 除果柄长外的18个定量描述性状均符合正态分布, 说明该树种受人为干扰较少, 主要处于野生、半野生状态, 与赵海娟等(2013)对普通杏(Armeniaca vulgaris)的研究结果类似。3.2 东北杏种质定量描述性状的地理变化
相关性分析结果(表5)表明, 调查区域的地理位置与主要气候因子之间存在着显著的负相关性, 地理位置梯度的变化反映了气候条件的变化。因此, 随着经、纬度的升高, 气温降低, 降水减少, 东北杏种质小枝长度、小枝粗度、种子质量和果柄长呈现出显著或极显著变小的趋势。在其地理变化中, 小枝长度表现为由东到西逐渐增大的梯度变化, 经度是主要影响因子; 小枝粗度、种子质量表现为由北到南逐渐增大的梯度变化, 纬度是主要影响因子; 果柄长表现为由东北到西南逐渐增大的梯度变化, 经、纬度均是主要影响因子。五角枫(Acer mono)果柄长、任豆(Zenia insignis)种子质量的地理变化规律与东北杏一致(姬志峰, 2013; 林玮等, 2016)。本研究发现, 叶宽与经度显著负相关, 这是因为随着经度的升高, 降水减少, 叶片变小不仅可以削弱自身的边界层阻力, 增加叶片的保水性, 而且可以减缓叶温升高的速率, 从而提高植物对有限水分的利用效率(McDonald et al., 2003; 任红剑等, 2018); 而主枝基角与纬度显著正相关, 这是因为随着纬度的增加, 气温降低, 年日照时间变短, 光谱中短波成分增加, 为了适应这些环境胁迫, 植株通过增大主枝基角, 更好地接受太阳光, 提高树冠内的温度, 有利于植株的光合作用, 促进生长(苏渤海等, 2008; 李军如和宋涛, 2013)。随着海拔的升高, 各种气象要素也发生相应变化, 在一定海拔高度范围内, 海拔升高, 温度降低, 大气压减小, 空气所能容纳的水汽含量降低, 进而导致大气相对湿度的增加(郑征等, 2007)。本研究表明, 小枝粗度与海拔呈负相关关系, 这与经纬度的作用机理一致。但小枝长度、果柄长与海拔均呈正相关关系, 其中小枝长度与海拔的相关性达显著水平, 这与李曼等(2017)的研究结果相似。这是因为在高海拔处, 水分、温度等胁迫因子加剧, 植物小枝和果柄不仅需要承担运输水分和养分的功能, 还需要增加对茎的投入, 以支撑风、雪等因素对茎产生的压力。恒定胁迫模型理论认为, 令植物茎横截面外缘弯曲所需要做的功(即弯曲矩) M主要是由作用于茎上的力(F)和茎的长度(L)决定, 即M = F × L。当植物小枝和果柄面临大风、降雪等压力时, 通过增加其长度, 进而增大茎基部所能承受的最大弯曲胁迫, 有利于保持茎的稳定性。其次, 通过增加小枝的长度, 以便将更多叶片伸展到冠层以外, 减少叶片之间的互相遮蔽, 有利于植物更有效地获取光能, 提高光合效率(Niklas, 1999; 李俊慧等, 2017)。
地理位置对东北杏种质表型性状影响较大, 所以在进行良种选育时, 应将野外优选种质定植于相同条件下, 采取同样的经营措施, 以排除自然条件、林龄及经营措施所造成的影响, 在此基础上进行形态观测, 筛选出优良单株。
3.3 东北杏种质的分类
通过主成分分析和聚类分析, 基于表型性状和分子标记均可将47份东北杏种质分为4类, 但从聚类结果分析来看, 2种方法的分类存在较大差异, 在柚(Citrus maxima)(彭瑜等, 2008)、油橄榄(Olea europaea)(秦倩等, 2016)、牡丹(Paeonia spp.)(翟立娟, 2019)等植物的研究中也存在形态学聚类和分子标记聚类结果不一致的情况。本研究调查区域包括辽宁、吉林和黑龙江的15个地区, 基本覆盖了东北杏的主要分布区, 调查区域内年平均气温为3.4-8.5 ℃, 降水量为530.2- 1 077.8 mm, 由于地理隔离和长期的自然选择, 不同产地的种质形成了相应的遗传特性(金玲等, 2018), 同时东北杏为虫媒传粉植物, 同风媒传粉植物花粉相比, 其花粉的传播距离受到限制, 导致种群间的基因流水平相对较低, 增加了种群间分化的可能性(刁松锋等, 2014), 因此基于SSR分子标记的东北杏种质聚类结果与种质产地高度吻合。基于表型性状的聚类结果中既存在地理位置较近的种质聚为一类的情况, 也存在地理位置较远的种质聚为一类的情况, 表型性状表现为空间上的不连续变异, 彭兴民等(2012)对Azadirachta indica和高张莹等(2017)对核桃楸(Juglans mandshurica)的聚类分析结果也体现出这一规律。本研究中基于分子标记和表型性状的两种聚类结果的相关性不显著, SSR分子标记的东北杏种质遗传差异较小, 表型性状多样性较高, 这是因为除不同种质产地的气候差异外, 海拔、地形、土壤等方面的环境异质性对表型性状多样性的形成也起到了重要作用(曾杰等, 2005)。
附录? 东北杏种质99个SSR位点的多态性信息
Supplement ? Polymorphism information of 99 SSR loci in Armeniaca mandshurica germplasms
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.17521/cjpe.2017.0229URL [本文引用: 1]
DOI:10.17521/cjpe.2017.0229URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.17521/cjpe.2016.0376URL [本文引用: 1]
DOI:10.17521/cjpe.2016.0376URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s11356-019-06538-4URLPMID:31838703 [本文引用: 1]

Valuation of environmental goods and services are frequently spatially heterogeneous, the significance of this heterogeneity for policy analysis is gradually recognized. For the valuation of environmental goods and services in the context of attaining a better environmental status across Heihe River basin (HRB), Northwest of China by 2020, a survey was conducted employing choice experiment approach across the entire river basin. In this research we estimate the impact of distance from inland rivers' origin (spatial attribute) on the inhabitants' willingness to pay for restoration of ecological attributes. A total of 11 ecological attributes were selected including 6 land related attributes and 5 water related attributes. The present study displays the outcomes of the research planned to examine differences in willingness to pay across different locations/distances. A total of five cities and 33 surround villages/townships were included to examine for location effect, while four ad hoc base distances split samples were recognized for distance effect i.e. ≤100?km, ≤200?km, ≤300 and?&gt;?300?km from rivers' origin. The results of mixed logit model recognized that people living at different locations value the attributes differently. For instance the people of Zhangye region are willing to pay RMB 95.66 annually for improvements in biodiversity while in Gaotai the individuals' willingness to pay (WTP) for the same attributes was RMB 45.68. Similarly, the respondents' living nearer to the origin of river were willing to pay relatively higher amount for the upgradation in the degraded ecosystem services than the remainders. The results of willingness to pay obtained by Krinsky Robb method confirmed that the attributes quality of agricultural products and water quality were the most preferred attributes with the willingness to pay RMB 91.09 and 122.89 respectively. The significant results of willingness to pay may serve as a reference for sustainable improvements and uplifting of the degraded ecological attributes.
DOI:10.1007/s11356-019-06538-4URLPMID:31838703 [本文引用: 1]
Valuation of environmental goods and services are frequently spatially heterogeneous, the significance of this heterogeneity for policy analysis is gradually recognized. For the valuation of environmental goods and services in the context of attaining a better environmental status across Heihe River basin (HRB), Northwest of China by 2020, a survey was conducted employing choice experiment approach across the entire river basin. In this research we estimate the impact of distance from inland rivers' origin (spatial attribute) on the inhabitants' willingness to pay for restoration of ecological attributes. A total of 11 ecological attributes were selected including 6 land related attributes and 5 water related attributes. The present study displays the outcomes of the research planned to examine differences in willingness to pay across different locations/distances. A total of five cities and 33 surround villages/townships were included to examine for location effect, while four ad hoc base distances split samples were recognized for distance effect i.e. ≤100?km, ≤200?km, ≤300 and?&gt;?300?km from rivers' origin. The results of mixed logit model recognized that people living at different locations value the attributes differently. For instance the people of Zhangye region are willing to pay RMB 95.66 annually for improvements in biodiversity while in Gaotai the individuals' willingness to pay (WTP) for the same attributes was RMB 45.68. Similarly, the respondents' living nearer to the origin of river were willing to pay relatively higher amount for the upgradation in the degraded ecosystem services than the remainders. The results of willingness to pay obtained by Krinsky Robb method confirmed that the attributes quality of agricultural products and water quality were the most preferred attributes with the willingness to pay RMB 91.09 and 122.89 respectively. The significant results of willingness to pay may serve as a reference for sustainable improvements and uplifting of the degraded ecological attributes.
[本文引用: 1]
[本文引用: 1]
DOI:10.13287/j.1001-9332.201702.039URLPMID:29749162 [本文引用: 1]
To analyze the tradeoff relationship between twigs and leaves, the traits of Pinus hwang-shanensis including leaf area, leaf number, twig length and twig diameter were investigated in Wuyi Mountains along an altitudinal gradient. The results indicated that leaf number, twig length, twig diameter, leafing intensity and twig stem cross-sectional area of P. hwangshanensis increased gra-dually with the increasing altitude, while individual leaf area decreased gradually. Leafing intensity of P. hwangshanensis at different altitudes had significant negative relationships with leaf area. The cross-sectional area of P. hwangshanensis had significant positive relationship with total leaf area. Twig length and twig diameter of P. hwangshanensis correlated negatively with leafing intensity, but positively with leaf area, leaf number and total leaf area. To enhance the competitiveness and resource utilization efficiency, P. hwangshanensis at low altitude tended to have relatively few large leaves on short twigs, and those at high altitude tended to have a large number of small leaves on long twigs. Such tradeoff between twigs and leaves reflected the strategy of resource utilization and the balance of biomass allocation mechanism of P. hwangshanensis responding to the altitudinal change.
DOI:10.13287/j.1001-9332.201702.039URLPMID:29749162 [本文引用: 1]
To analyze the tradeoff relationship between twigs and leaves, the traits of Pinus hwang-shanensis including leaf area, leaf number, twig length and twig diameter were investigated in Wuyi Mountains along an altitudinal gradient. The results indicated that leaf number, twig length, twig diameter, leafing intensity and twig stem cross-sectional area of P. hwangshanensis increased gra-dually with the increasing altitude, while individual leaf area decreased gradually. Leafing intensity of P. hwangshanensis at different altitudes had significant negative relationships with leaf area. The cross-sectional area of P. hwangshanensis had significant positive relationship with total leaf area. Twig length and twig diameter of P. hwangshanensis correlated negatively with leafing intensity, but positively with leaf area, leaf number and total leaf area. To enhance the competitiveness and resource utilization efficiency, P. hwangshanensis at low altitude tended to have relatively few large leaves on short twigs, and those at high altitude tended to have a large number of small leaves on long twigs. Such tradeoff between twigs and leaves reflected the strategy of resource utilization and the balance of biomass allocation mechanism of P. hwangshanensis responding to the altitudinal change.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1002/jcp.29399URLPMID:31838757 [本文引用: 1]
In Italy, in the eastern area of the Campania region, the illegal dumping and burning of waste have been documented, which could potentially affect the local population's health. In particular, toxic waste exposure has been suggested to associate with increased cancer development/mortality in these areas, although a causal link has not yet been established. In this pilot study, we evaluated blood levels of toxic heavy metals and persistent organic pollutants (POPs) in 95 patients with different cancer types residing in this area and in 27 healthy individuals. While we did not find any significant correlation between the blood levels of POPs and the provenance of the patients, we did observe high blood concentrations of heavy metals in some municipalities, including Giugliano, where many illegal waste disposal sites have previously been documented. Our results showed that patients with different cancer types from Giugliano had higher blood levels of heavy metals than healthy controls. Despite the obvious limitations of this exploratory study, our preliminary observations encourage further research assessing the possible association between exposure to hazardous waste, increased blood metals, and increased risk of cancer.
DOI:10.17521/cjpe.2017.0104URL [本文引用: 1]
DOI:10.17521/cjpe.2017.0104URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1046/j.1365-2435.2003.00698.xURL [本文引用: 1]
DOI:10.1073/pnas.76.10.5269URLPMID:291943 [本文引用: 1]
A mathematical model for the evolutionary change of restriction sites in mitochondrial DNA is developed. Formulas based on this model are presented for estimating the number of nucleotide substitutions between two populations or species. To express the degree of polymorphism in a population at the nucleotide level, a measure called &quot;nucleotide diversity&quot; is proposed.
DOI:10.1046/j.1469-8137.1999.00441.xURL [本文引用: 1]
DOI:10.1111/gcb.14962URLPMID:31838767 [本文引用: 1]
Numerous studies have demonstrated that fertilization with nutrients such as nitrogen, phosphorus, and potassium increase plant productivity in both natural and managed ecosystems, demonstrating that primary productivity is nutrient limited in most terrestrial ecosystems. In contrast, it has been demonstrated that heterotrophic microbial communities in soil are primarily limited by organic carbon or energy. While this concept of contrasting limitations, i.e., microbial carbon and plant nutrient limitation, is based on strong evidence that we review in this paper, it is often ignored in discussions of ecosystem response to global environment changes. The plant-centric perspective has equated plant-nutrient limitations with those of whole ecosystems, thereby ignoring the important role of the heterotrophs responsible for soil decomposition in driving ecosystem carbon storage. In order to truly integrate carbon and nutrient cycles in ecosystem science, we must account for the fact that while plant productivity may be nutrient- limited, the secondary productivity by heterotrophic communities is inherently carbon-limited. Ecosystem carbon cycling integrates the independent physiological responses of its individual components, as well as tightly coupled exchanges between autotrophs and heterotrophs. To the extent that the interacting autotrophic and heterotrophic processes are controlled by organisms that are limited by nutrient versus carbon accessibility, respectively, we propose that ecosystems by definition cannot be 'limited' by nutrients or carbon alone. Here, we outline how models aimed at predicting non-steady state ecosystem responses over time can benefit from dissecting ecosystems into the organismal components and their inherent limitations to better represent plant-microbe interactions in coupled carbon and nutrient models.
DOI:10.1111/gcb.14962URLPMID:31838767 [本文引用: 1]
Numerous studies have demonstrated that fertilization with nutrients such as nitrogen, phosphorus, and potassium increase plant productivity in both natural and managed ecosystems, demonstrating that primary productivity is nutrient limited in most terrestrial ecosystems. In contrast, it has been demonstrated that heterotrophic microbial communities in soil are primarily limited by organic carbon or energy. While this concept of contrasting limitations, i.e., microbial carbon and plant nutrient limitation, is based on strong evidence that we review in this paper, it is often ignored in discussions of ecosystem response to global environment changes. The plant-centric perspective has equated plant-nutrient limitations with those of whole ecosystems, thereby ignoring the important role of the heterotrophs responsible for soil decomposition in driving ecosystem carbon storage. In order to truly integrate carbon and nutrient cycles in ecosystem science, we must account for the fact that while plant productivity may be nutrient- limited, the secondary productivity by heterotrophic communities is inherently carbon-limited. Ecosystem carbon cycling integrates the independent physiological responses of its individual components, as well as tightly coupled exchanges between autotrophs and heterotrophs. To the extent that the interacting autotrophic and heterotrophic processes are controlled by organisms that are limited by nutrient versus carbon accessibility, respectively, we propose that ecosystems by definition cannot be 'limited' by nutrients or carbon alone. Here, we outline how models aimed at predicting non-steady state ecosystem responses over time can benefit from dissecting ecosystems into the organismal components and their inherent limitations to better represent plant-microbe interactions in coupled carbon and nutrient models.
DOI:10.3724/SP.J.1258.2012.00560URL [本文引用: 1]
DOI:10.3724/SP.J.1258.2012.00560URL [本文引用: 1]
DOI:10.1007/s11356-019-06538-4URLPMID:31838703 [本文引用: 1]
Valuation of environmental goods and services are frequently spatially heterogeneous, the significance of this heterogeneity for policy analysis is gradually recognized. For the valuation of environmental goods and services in the context of attaining a better environmental status across Heihe River basin (HRB), Northwest of China by 2020, a survey was conducted employing choice experiment approach across the entire river basin. In this research we estimate the impact of distance from inland rivers' origin (spatial attribute) on the inhabitants' willingness to pay for restoration of ecological attributes. A total of 11 ecological attributes were selected including 6 land related attributes and 5 water related attributes. The present study displays the outcomes of the research planned to examine differences in willingness to pay across different locations/distances. A total of five cities and 33 surround villages/townships were included to examine for location effect, while four ad hoc base distances split samples were recognized for distance effect i.e. ≤100?km, ≤200?km, ≤300 and?&gt;?300?km from rivers' origin. The results of mixed logit model recognized that people living at different locations value the attributes differently. For instance the people of Zhangye region are willing to pay RMB 95.66 annually for improvements in biodiversity while in Gaotai the individuals' willingness to pay (WTP) for the same attributes was RMB 45.68. Similarly, the respondents' living nearer to the origin of river were willing to pay relatively higher amount for the upgradation in the degraded ecosystem services than the remainders. The results of willingness to pay obtained by Krinsky Robb method confirmed that the attributes quality of agricultural products and water quality were the most preferred attributes with the willingness to pay RMB 91.09 and 122.89 respectively. The significant results of willingness to pay may serve as a reference for sustainable improvements and uplifting of the degraded ecological attributes.
DOI:10.1007/s11356-019-06538-4URLPMID:31838703 [本文引用: 1]
Valuation of environmental goods and services are frequently spatially heterogeneous, the significance of this heterogeneity for policy analysis is gradually recognized. For the valuation of environmental goods and services in the context of attaining a better environmental status across Heihe River basin (HRB), Northwest of China by 2020, a survey was conducted employing choice experiment approach across the entire river basin. In this research we estimate the impact of distance from inland rivers' origin (spatial attribute) on the inhabitants' willingness to pay for restoration of ecological attributes. A total of 11 ecological attributes were selected including 6 land related attributes and 5 water related attributes. The present study displays the outcomes of the research planned to examine differences in willingness to pay across different locations/distances. A total of five cities and 33 surround villages/townships were included to examine for location effect, while four ad hoc base distances split samples were recognized for distance effect i.e. ≤100?km, ≤200?km, ≤300 and?&gt;?300?km from rivers' origin. The results of mixed logit model recognized that people living at different locations value the attributes differently. For instance the people of Zhangye region are willing to pay RMB 95.66 annually for improvements in biodiversity while in Gaotai the individuals' willingness to pay (WTP) for the same attributes was RMB 45.68. Similarly, the respondents' living nearer to the origin of river were willing to pay relatively higher amount for the upgradation in the degraded ecosystem services than the remainders. The results of willingness to pay obtained by Krinsky Robb method confirmed that the attributes quality of agricultural products and water quality were the most preferred attributes with the willingness to pay RMB 91.09 and 122.89 respectively. The significant results of willingness to pay may serve as a reference for sustainable improvements and uplifting of the degraded ecological attributes.
[本文引用: 1]
[本文引用: 1]
DOI:10.1088/0952-4746/33/1/113URLPMID:23296360 [本文引用: 1]
Living organisms are exposed to a mixture of environmental stressors, and the resultant effects are referred to as multiple stressor effects. In the present work, we studied the multiple stressor effect in embryos of the zebrafish (Danio rerio) from simultaneous exposure to ionising radiation (alpha particles) and cadmium through quantification of apoptotic signals at 24 h postfertilisation (hpf) revealed by vital dye acridine orange staining. For each set of experiments, 32-40 dechorionated embryos were deployed, which were divided into four groups each having 8-10 embryos. The four groups of embryos were referred to as (1) the control group (C), which received no further treatments after dechorionation; (2) the Cd-dosed and irradiated group (CdIr), which was exposed to 100 μM Cd from 5 to 24 hpf, and also received about 4.4 mGy from alpha particles at 5 hpf; (3) the irradiated group (Ir), which received about 4.4 mGy from alpha particles at 5 hpf; and (4) the Cd-dosed group (Cd), which was exposed to 100 μM Cd from 5 to 24 hpf. In general, the CdIr, Ir and Cd groups had more apoptotic signals than the C group. Within the 12 sets of experimental results, two showed significant synergistic effects, one showed a weakly synergistic effect and nine showed additive effects. The multiple stressor effect of 100 μM Cd with ~4.4 mGy alpha-particle radiation resulted in an additive or synergistic effect, but no antagonistic effect. The failure to identify significant synergistic effects for some sets of data, and thus their subsequent classification as additive effects, might be a result of the relatively small magnitude of the synergistic effects. The results showed that the radiation risk could be perturbed by another environmental stressor such as a heavy metal, and as such a realistic human radiation risk assessment should in general take into account the multiple stressor effects.
DOI:10.1088/0952-4746/33/1/113URLPMID:23296360 [本文引用: 1]
Living organisms are exposed to a mixture of environmental stressors, and the resultant effects are referred to as multiple stressor effects. In the present work, we studied the multiple stressor effect in embryos of the zebrafish (Danio rerio) from simultaneous exposure to ionising radiation (alpha particles) and cadmium through quantification of apoptotic signals at 24 h postfertilisation (hpf) revealed by vital dye acridine orange staining. For each set of experiments, 32-40 dechorionated embryos were deployed, which were divided into four groups each having 8-10 embryos. The four groups of embryos were referred to as (1) the control group (C), which received no further treatments after dechorionation; (2) the Cd-dosed and irradiated group (CdIr), which was exposed to 100 μM Cd from 5 to 24 hpf, and also received about 4.4 mGy from alpha particles at 5 hpf; (3) the irradiated group (Ir), which received about 4.4 mGy from alpha particles at 5 hpf; and (4) the Cd-dosed group (Cd), which was exposed to 100 μM Cd from 5 to 24 hpf. In general, the CdIr, Ir and Cd groups had more apoptotic signals than the C group. Within the 12 sets of experimental results, two showed significant synergistic effects, one showed a weakly synergistic effect and nine showed additive effects. The multiple stressor effect of 100 μM Cd with ~4.4 mGy alpha-particle radiation resulted in an additive or synergistic effect, but no antagonistic effect. The failure to identify significant synergistic effects for some sets of data, and thus their subsequent classification as additive effects, might be a result of the relatively small magnitude of the synergistic effects. The results showed that the radiation risk could be perturbed by another environmental stressor such as a heavy metal, and as such a realistic human radiation risk assessment should in general take into account the multiple stressor effects.
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s11356-019-06538-4URLPMID:31838703 [本文引用: 1]
Valuation of environmental goods and services are frequently spatially heterogeneous, the significance of this heterogeneity for policy analysis is gradually recognized. For the valuation of environmental goods and services in the context of attaining a better environmental status across Heihe River basin (HRB), Northwest of China by 2020, a survey was conducted employing choice experiment approach across the entire river basin. In this research we estimate the impact of distance from inland rivers' origin (spatial attribute) on the inhabitants' willingness to pay for restoration of ecological attributes. A total of 11 ecological attributes were selected including 6 land related attributes and 5 water related attributes. The present study displays the outcomes of the research planned to examine differences in willingness to pay across different locations/distances. A total of five cities and 33 surround villages/townships were included to examine for location effect, while four ad hoc base distances split samples were recognized for distance effect i.e. ≤100?km, ≤200?km, ≤300 and?&gt;?300?km from rivers' origin. The results of mixed logit model recognized that people living at different locations value the attributes differently. For instance the people of Zhangye region are willing to pay RMB 95.66 annually for improvements in biodiversity while in Gaotai the individuals' willingness to pay (WTP) for the same attributes was RMB 45.68. Similarly, the respondents' living nearer to the origin of river were willing to pay relatively higher amount for the upgradation in the degraded ecosystem services than the remainders. The results of willingness to pay obtained by Krinsky Robb method confirmed that the attributes quality of agricultural products and water quality were the most preferred attributes with the willingness to pay RMB 91.09 and 122.89 respectively. The significant results of willingness to pay may serve as a reference for sustainable improvements and uplifting of the degraded ecological attributes.
DOI:10.1007/s11356-019-06538-4URLPMID:31838703 [本文引用: 1]
Valuation of environmental goods and services are frequently spatially heterogeneous, the significance of this heterogeneity for policy analysis is gradually recognized. For the valuation of environmental goods and services in the context of attaining a better environmental status across Heihe River basin (HRB), Northwest of China by 2020, a survey was conducted employing choice experiment approach across the entire river basin. In this research we estimate the impact of distance from inland rivers' origin (spatial attribute) on the inhabitants' willingness to pay for restoration of ecological attributes. A total of 11 ecological attributes were selected including 6 land related attributes and 5 water related attributes. The present study displays the outcomes of the research planned to examine differences in willingness to pay across different locations/distances. A total of five cities and 33 surround villages/townships were included to examine for location effect, while four ad hoc base distances split samples were recognized for distance effect i.e. ≤100?km, ≤200?km, ≤300 and?&gt;?300?km from rivers' origin. The results of mixed logit model recognized that people living at different locations value the attributes differently. For instance the people of Zhangye region are willing to pay RMB 95.66 annually for improvements in biodiversity while in Gaotai the individuals' willingness to pay (WTP) for the same attributes was RMB 45.68. Similarly, the respondents' living nearer to the origin of river were willing to pay relatively higher amount for the upgradation in the degraded ecosystem services than the remainders. The results of willingness to pay obtained by Krinsky Robb method confirmed that the attributes quality of agricultural products and water quality were the most preferred attributes with the willingness to pay RMB 91.09 and 122.89 respectively. The significant results of willingness to pay may serve as a reference for sustainable improvements and uplifting of the degraded ecological attributes.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.5558/tfc2013-013URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
DOI:10.1371/journal.pone.0087381URLPMID:24516551
Siberian apricot (Prunus sibirica L.), an ecologically and economically important tree species with a high degree of tolerance to a variety of extreme environmental conditions, is widely distributed across the mountains of northeastern and northern China, eastern and southeastern regions of Mongolia, Eastern Siberia, and the Maritime Territory of Russia. However, few studies have examined the genetic diversity and population structure of this species. Using 31 nuclear microsatellites, we investigated the level of genetic diversity and population structure of Siberian apricot sampled from 22 populations across China. The number of alleles per locus ranged from 5 to 33, with an average of 19.323 alleles. The observed heterozygosity and expected heterozygosity ranged from 0.037 to 0.874 and 0.040 to 0.924 with average values of 0.639 and 0.774, respectively. A STRUCTURE-based analysis clustered all of the populations into four genetic clusters. Significant genetic differentiation was observed between all population pairs. A hierarchical analysis of molecular variance attributed about 94% of the variation to within populations. No significant difference was detected between the wild and semi-wild groups, indicating that recent cultivation practices have had little impact on the genetic diversity of Siberian apricot. The Mantel test showed that the genetic distance among the populations was not significantly correlated with geographic distance (r?=?0.4651, p?=?0.9940). Our study represents the most comprehensive investigation of the genetic diversity and population structure of Siberian apricot in China to date, and it provides valuable information for the collection of genetic resources for the breeding of Siberian apricot and related species.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.17521/cjpe.2018.0196URL [本文引用: 1]
DOI:10.17521/cjpe.2018.0196URL [本文引用: 1]
假俭草种群变异与生态特性的研究
1
2002
... 遗传参数中变异系数可在一定程度上反映性状的离散程度, 变异系数越大, 离散程度越高, 多样性程度越高(
假俭草种群变异与生态特性的研究
1
2002
... 遗传参数中变异系数可在一定程度上反映性状的离散程度, 变异系数越大, 离散程度越高, 多样性程度越高(
基于ISSR分子标记分析新疆野杏遗传多样性
1
2015
... 遗传多样性是指物种种内基因组成的多样性, 不同的遗传多样性研究及检测方法存在差异, 可从形态学、细胞学及DNA分子等水平上进行, 其中表型测定是最简单和最直接的方法(
基于ISSR分子标记分析新疆野杏遗传多样性
1
2015
... 遗传多样性是指物种种内基因组成的多样性, 不同的遗传多样性研究及检测方法存在差异, 可从形态学、细胞学及DNA分子等水平上进行, 其中表型测定是最简单和最直接的方法(
基于种实性状的无患子天然群体表型多样性研究
1
2014
... 本研究调查区域包括辽宁、吉林和黑龙江的15个地区, 基本覆盖了东北杏的主要分布区, 调查区域内年平均气温为3.4-8.5 ℃, 降水量为530.2- 1 077.8 mm, 由于地理隔离和长期的自然选择, 不同产地的种质形成了相应的遗传特性(
基于种实性状的无患子天然群体表型多样性研究
1
2014
... 本研究调查区域包括辽宁、吉林和黑龙江的15个地区, 基本覆盖了东北杏的主要分布区, 调查区域内年平均气温为3.4-8.5 ℃, 降水量为530.2- 1 077.8 mm, 由于地理隔离和长期的自然选择, 不同产地的种质形成了相应的遗传特性(
核桃楸种群果核形态及地理变异
1
2017
... 本研究调查区域包括辽宁、吉林和黑龙江的15个地区, 基本覆盖了东北杏的主要分布区, 调查区域内年平均气温为3.4-8.5 ℃, 降水量为530.2- 1 077.8 mm, 由于地理隔离和长期的自然选择, 不同产地的种质形成了相应的遗传特性(
核桃楸种群果核形态及地理变异
1
2017
... 本研究调查区域包括辽宁、吉林和黑龙江的15个地区, 基本覆盖了东北杏的主要分布区, 调查区域内年平均气温为3.4-8.5 ℃, 降水量为530.2- 1 077.8 mm, 由于地理隔离和长期的自然选择, 不同产地的种质形成了相应的遗传特性(
山西刺槐种质资源的叶片表型多样性分析
1
2019
... 种质资源是林木改良的物质基础, 同时也是遗传学、分类学、生物学等理论研究的重要材料, 进行种质资源调查、评价是对其合理保护、利用的前提.目前, 林木种质资源调查、评价研究主要集中于主要经济林树种(
山西刺槐种质资源的叶片表型多样性分析
1
2019
... 种质资源是林木改良的物质基础, 同时也是遗传学、分类学、生物学等理论研究的重要材料, 进行种质资源调查、评价是对其合理保护、利用的前提.目前, 林木种质资源调查、评价研究主要集中于主要经济林树种(
枫香树种群种子与果实表型性状变异分析
1
2018
... 遗传多样性是指物种种内基因组成的多样性, 不同的遗传多样性研究及检测方法存在差异, 可从形态学、细胞学及DNA分子等水平上进行, 其中表型测定是最简单和最直接的方法(
枫香树种群种子与果实表型性状变异分析
1
2018
... 遗传多样性是指物种种内基因组成的多样性, 不同的遗传多样性研究及检测方法存在差异, 可从形态学、细胞学及DNA分子等水平上进行, 其中表型测定是最简单和最直接的方法(
山西五角枫天然种群表型多样性研究
1
2013
... 相关性分析结果(
山西五角枫天然种群表型多样性研究
1
2013
... 相关性分析结果(
97个山杏无性系的遗传多样性及SSR指纹图谱
3
2018
... 种质资源是林木改良的物质基础, 同时也是遗传学、分类学、生物学等理论研究的重要材料, 进行种质资源调查、评价是对其合理保护、利用的前提.目前, 林木种质资源调查、评价研究主要集中于主要经济林树种(
... 试验步骤包括DNA提取、SSR引物筛选、PCR反应体系及扩增程序、聚丙烯酰胺凝胶电泳检测及数据的读取, 具体参照
... 本研究调查区域包括辽宁、吉林和黑龙江的15个地区, 基本覆盖了东北杏的主要分布区, 调查区域内年平均气温为3.4-8.5 ℃, 降水量为530.2- 1 077.8 mm, 由于地理隔离和长期的自然选择, 不同产地的种质形成了相应的遗传特性(
97个山杏无性系的遗传多样性及SSR指纹图谱
3
2018
... 种质资源是林木改良的物质基础, 同时也是遗传学、分类学、生物学等理论研究的重要材料, 进行种质资源调查、评价是对其合理保护、利用的前提.目前, 林木种质资源调查、评价研究主要集中于主要经济林树种(
... 试验步骤包括DNA提取、SSR引物筛选、PCR反应体系及扩增程序、聚丙烯酰胺凝胶电泳检测及数据的读取, 具体参照
... 本研究调查区域包括辽宁、吉林和黑龙江的15个地区, 基本覆盖了东北杏的主要分布区, 调查区域内年平均气温为3.4-8.5 ℃, 降水量为530.2- 1 077.8 mm, 由于地理隔离和长期的自然选择, 不同产地的种质形成了相应的遗传特性(
Geographical variation in seed and seedling traits of neem ( Azadirachta indica A. Juss.) among ten populations studied in growth chamber
1
1997
... 遗传多样性是指物种种内基因组成的多样性, 不同的遗传多样性研究及检测方法存在差异, 可从形态学、细胞学及DNA分子等水平上进行, 其中表型测定是最简单和最直接的方法(
宁夏彭阳县山杏群体数量性状变异及其与经济性状的相关性
1
2012
... 遗传参数中变异系数可在一定程度上反映性状的离散程度, 变异系数越大, 离散程度越高, 多样性程度越高(
宁夏彭阳县山杏群体数量性状变异及其与经济性状的相关性
1
2012
... 遗传参数中变异系数可在一定程度上反映性状的离散程度, 变异系数越大, 离散程度越高, 多样性程度越高(
常绿和落叶阔叶物种当年生小枝茎长度和茎纤细率对展叶效率的影响
1
2017
... 随着海拔的升高, 各种气象要素也发生相应变化, 在一定海拔高度范围内, 海拔升高, 温度降低, 大气压减小, 空气所能容纳的水汽含量降低, 进而导致大气相对湿度的增加(
常绿和落叶阔叶物种当年生小枝茎长度和茎纤细率对展叶效率的影响
1
2017
... 随着海拔的升高, 各种气象要素也发生相应变化, 在一定海拔高度范围内, 海拔升高, 温度降低, 大气压减小, 空气所能容纳的水汽含量降低, 进而导致大气相对湿度的增加(
不同主枝开张角度下轮台白杏树冠微域气候及生长结果差异分析
1
2013
... 相关性分析结果(
不同主枝开张角度下轮台白杏树冠微域气候及生长结果差异分析
1
2013
... 相关性分析结果(
黄土高原不同县域山杏种质遗传多样性研究
1
2011
... 种质资源是林木改良的物质基础, 同时也是遗传学、分类学、生物学等理论研究的重要材料, 进行种质资源调查、评价是对其合理保护、利用的前提.目前, 林木种质资源调查、评价研究主要集中于主要经济林树种(
黄土高原不同县域山杏种质遗传多样性研究
1
2011
... 种质资源是林木改良的物质基础, 同时也是遗传学、分类学、生物学等理论研究的重要材料, 进行种质资源调查、评价是对其合理保护、利用的前提.目前, 林木种质资源调查、评价研究主要集中于主要经济林树种(
黄土高原山杏种质资源分类研究
1
2011
... 种质资源是林木改良的物质基础, 同时也是遗传学、分类学、生物学等理论研究的重要材料, 进行种质资源调查、评价是对其合理保护、利用的前提.目前, 林木种质资源调查、评价研究主要集中于主要经济林树种(
黄土高原山杏种质资源分类研究
1
2011
... 种质资源是林木改良的物质基础, 同时也是遗传学、分类学、生物学等理论研究的重要材料, 进行种质资源调查、评价是对其合理保护、利用的前提.目前, 林木种质资源调查、评价研究主要集中于主要经济林树种(
武夷山不同海拔黄山松枝叶大小关系
1
2017
... 随着海拔的升高, 各种气象要素也发生相应变化, 在一定海拔高度范围内, 海拔升高, 温度降低, 大气压减小, 空气所能容纳的水汽含量降低, 进而导致大气相对湿度的增加(
武夷山不同海拔黄山松枝叶大小关系
1
2017
... 随着海拔的升高, 各种气象要素也发生相应变化, 在一定海拔高度范围内, 海拔升高, 温度降低, 大气压减小, 空气所能容纳的水汽含量降低, 进而导致大气相对湿度的增加(
蒙古栎天然群体表型多样性研究
1
2005
... 遗传多样性是指物种种内基因组成的多样性, 不同的遗传多样性研究及检测方法存在差异, 可从形态学、细胞学及DNA分子等水平上进行, 其中表型测定是最简单和最直接的方法(
蒙古栎天然群体表型多样性研究
1
2005
... 遗传多样性是指物种种内基因组成的多样性, 不同的遗传多样性研究及检测方法存在差异, 可从形态学、细胞学及DNA分子等水平上进行, 其中表型测定是最简单和最直接的方法(
苦楝种源果核和种子性状地理变异的趋势面分析
1
2016
... 将供试种质所处的海拔平均值代入小枝长度、小枝粗度、种子质量和果柄长的回归方程中(
苦楝种源果核和种子性状地理变异的趋势面分析
1
2016
... 将供试种质所处的海拔平均值代入小枝长度、小枝粗度、种子质量和果柄长的回归方程中(
任豆种源种子性状地理变异研究
1
2016
... 相关性分析结果(
任豆种源种子性状地理变异研究
1
2016
... 相关性分析结果(
Study on the provenance of Fraxinus mandshurica
1
1997
... 种质资源是林木改良的物质基础, 同时也是遗传学、分类学、生物学等理论研究的重要材料, 进行种质资源调查、评价是对其合理保护、利用的前提.目前, 林木种质资源调查、评价研究主要集中于主要经济林树种(
长柄扁桃天然种群表型变异
1
2017
... 遗传参数中变异系数可在一定程度上反映性状的离散程度, 变异系数越大, 离散程度越高, 多样性程度越高(
长柄扁桃天然种群表型变异
1
2017
... 遗传参数中变异系数可在一定程度上反映性状的离散程度, 变异系数越大, 离散程度越高, 多样性程度越高(
山杏花粉形态特征与花粉萌发的关系
1
2015
... 种质资源是林木改良的物质基础, 同时也是遗传学、分类学、生物学等理论研究的重要材料, 进行种质资源调查、评价是对其合理保护、利用的前提.目前, 林木种质资源调查、评价研究主要集中于主要经济林树种(
山杏花粉形态特征与花粉萌发的关系
1
2015
... 种质资源是林木改良的物质基础, 同时也是遗传学、分类学、生物学等理论研究的重要材料, 进行种质资源调查、评价是对其合理保护、利用的前提.目前, 林木种质资源调查、评价研究主要集中于主要经济林树种(
山杏种内POD同工酶及种子可溶性蛋白分析
1
2006
... 种质资源是林木改良的物质基础, 同时也是遗传学、分类学、生物学等理论研究的重要材料, 进行种质资源调查、评价是对其合理保护、利用的前提.目前, 林木种质资源调查、评价研究主要集中于主要经济林树种(
山杏种内POD同工酶及种子可溶性蛋白分析
1
2006
... 种质资源是林木改良的物质基础, 同时也是遗传学、分类学、生物学等理论研究的重要材料, 进行种质资源调查、评价是对其合理保护、利用的前提.目前, 林木种质资源调查、评价研究主要集中于主要经济林树种(
乌饭树果实大小的地理变异研究
1
2016
... 遗传多样性是指物种种内基因组成的多样性, 不同的遗传多样性研究及检测方法存在差异, 可从形态学、细胞学及DNA分子等水平上进行, 其中表型测定是最简单和最直接的方法(
乌饭树果实大小的地理变异研究
1
2016
... 遗传多样性是指物种种内基因组成的多样性, 不同的遗传多样性研究及检测方法存在差异, 可从形态学、细胞学及DNA分子等水平上进行, 其中表型测定是最简单和最直接的方法(
青海高原核桃种质资源调查及坚果表型多样性分析
1
2015
... 种质资源是林木改良的物质基础, 同时也是遗传学、分类学、生物学等理论研究的重要材料, 进行种质资源调查、评价是对其合理保护、利用的前提.目前, 林木种质资源调查、评价研究主要集中于主要经济林树种(
青海高原核桃种质资源调查及坚果表型多样性分析
1
2015
... 种质资源是林木改良的物质基础, 同时也是遗传学、分类学、生物学等理论研究的重要材料, 进行种质资源调查、评价是对其合理保护、利用的前提.目前, 林木种质资源调查、评价研究主要集中于主要经济林树种(
中国樱桃地方种质资源表型性状遗传多样性分析
1
2016
... 种质资源是林木改良的物质基础, 同时也是遗传学、分类学、生物学等理论研究的重要材料, 进行种质资源调查、评价是对其合理保护、利用的前提.目前, 林木种质资源调查、评价研究主要集中于主要经济林树种(
中国樱桃地方种质资源表型性状遗传多样性分析
1
2016
... 种质资源是林木改良的物质基础, 同时也是遗传学、分类学、生物学等理论研究的重要材料, 进行种质资源调查、评价是对其合理保护、利用的前提.目前, 林木种质资源调查、评价研究主要集中于主要经济林树种(
Leaf-size divergence along rainfall and soil-nutrient gradients: Is the method of size reduction common among clades?
1
2003
... 相关性分析结果(
Mathematical model for studying genetic variation in terms of restriction endonucleases
1
1979
... 使用Cervus 3.0软件计算SSR标记各位点的多态性信息含量(PIC = $1-\sum{{{P}_{j}}^{2}}$, 其中Pj2为第j个基因的频率); 按照
A mechanical perspective on foliage leaf form and function
1
1999
... 随着海拔的升高, 各种气象要素也发生相应变化, 在一定海拔高度范围内, 海拔升高, 温度降低, 大气压减小, 空气所能容纳的水汽含量降低, 进而导致大气相对湿度的增加(
东北杏栽培技术与效益分析
1
2001
... 东北杏(Armeniaca mandshurica)为蔷薇科杏属植物, 主要分布于我国黑龙江、吉林和辽宁的东部地区, 在朝鲜的中北部和俄罗斯南乌苏里地区也有分布.东北杏用途很广, 是集观赏、经济、用材于一身的重要树种(
东北杏栽培技术与效益分析
1
2001
... 东北杏(Armeniaca mandshurica)为蔷薇科杏属植物, 主要分布于我国黑龙江、吉林和辽宁的东部地区, 在朝鲜的中北部和俄罗斯南乌苏里地区也有分布.东北杏用途很广, 是集观赏、经济、用材于一身的重要树种(
云南引种印楝实生种群的表型变异
1
2012
... 本研究调查区域包括辽宁、吉林和黑龙江的15个地区, 基本覆盖了东北杏的主要分布区, 调查区域内年平均气温为3.4-8.5 ℃, 降水量为530.2- 1 077.8 mm, 由于地理隔离和长期的自然选择, 不同产地的种质形成了相应的遗传特性(
云南引种印楝实生种群的表型变异
1
2012
... 本研究调查区域包括辽宁、吉林和黑龙江的15个地区, 基本覆盖了东北杏的主要分布区, 调查区域内年平均气温为3.4-8.5 ℃, 降水量为530.2- 1 077.8 mm, 由于地理隔离和长期的自然选择, 不同产地的种质形成了相应的遗传特性(
利用叶片形态学性状和ISSR标记检测柚类的遗传多样性
1
2008
... 通过主成分分析和聚类分析, 基于表型性状和分子标记均可将47份东北杏种质分为4类, 但从聚类结果分析来看, 2种方法的分类存在较大差异, 在柚(Citrus maxima)(
利用叶片形态学性状和ISSR标记检测柚类的遗传多样性
1
2008
... 通过主成分分析和聚类分析, 基于表型性状和分子标记均可将47份东北杏种质分为4类, 但从聚类结果分析来看, 2种方法的分类存在较大差异, 在柚(Citrus maxima)(
油橄榄品种表型和SSR标记的多样性及聚类分析
1
2016
... 通过主成分分析和聚类分析, 基于表型性状和分子标记均可将47份东北杏种质分为4类, 但从聚类结果分析来看, 2种方法的分类存在较大差异, 在柚(Citrus maxima)(
油橄榄品种表型和SSR标记的多样性及聚类分析
1
2016
... 通过主成分分析和聚类分析, 基于表型性状和分子标记均可将47份东北杏种质分为4类, 但从聚类结果分析来看, 2种方法的分类存在较大差异, 在柚(Citrus maxima)(
元宝枫叶片形态特征的地理变异
1
2018
... 相关性分析结果(
元宝枫叶片形态特征的地理变异
1
2018
... 相关性分析结果(
无患子种实形态及经济性状的地理变异
1
2013
... 遗传多样性是指物种种内基因组成的多样性, 不同的遗传多样性研究及检测方法存在差异, 可从形态学、细胞学及DNA分子等水平上进行, 其中表型测定是最简单和最直接的方法(
无患子种实形态及经济性状的地理变异
1
2013
... 遗传多样性是指物种种内基因组成的多样性, 不同的遗传多样性研究及检测方法存在差异, 可从形态学、细胞学及DNA分子等水平上进行, 其中表型测定是最简单和最直接的方法(
红富士苹果改形过程中不同树形光照分布及其对产量品质的影响
1
2008
... 相关性分析结果(
红富士苹果改形过程中不同树形光照分布及其对产量品质的影响
1
2008
... 相关性分析结果(
河西走廊不同红砂天然群体种子表型性状相关性研究
1
2013
... 遗传多样性是指物种种内基因组成的多样性, 不同的遗传多样性研究及检测方法存在差异, 可从形态学、细胞学及DNA分子等水平上进行, 其中表型测定是最简单和最直接的方法(
河西走廊不同红砂天然群体种子表型性状相关性研究
1
2013
... 遗传多样性是指物种种内基因组成的多样性, 不同的遗传多样性研究及检测方法存在差异, 可从形态学、细胞学及DNA分子等水平上进行, 其中表型测定是最简单和最直接的方法(
1
2002
... 遗传参数中变异系数可在一定程度上反映性状的离散程度, 变异系数越大, 离散程度越高, 多样性程度越高(
1
2002
... 遗传参数中变异系数可在一定程度上反映性状的离散程度, 变异系数越大, 离散程度越高, 多样性程度越高(
An overview of Populus genetic resources in southwest China
1
2013
... 种质资源是林木改良的物质基础, 同时也是遗传学、分类学、生物学等理论研究的重要材料, 进行种质资源调查、评价是对其合理保护、利用的前提.目前, 林木种质资源调查、评价研究主要集中于主要经济林树种(
我国3种杏的地理分布及其植物学性状
1
2010
... 东北杏(Armeniaca mandshurica)为蔷薇科杏属植物, 主要分布于我国黑龙江、吉林和辽宁的东部地区, 在朝鲜的中北部和俄罗斯南乌苏里地区也有分布.东北杏用途很广, 是集观赏、经济、用材于一身的重要树种(
我国3种杏的地理分布及其植物学性状
1
2010
... 东北杏(Armeniaca mandshurica)为蔷薇科杏属植物, 主要分布于我国黑龙江、吉林和辽宁的东部地区, 在朝鲜的中北部和俄罗斯南乌苏里地区也有分布.东北杏用途很广, 是集观赏、经济、用材于一身的重要树种(
枣种质资源的表型遗传多样性
2
2014
... 种质资源是林木改良的物质基础, 同时也是遗传学、分类学、生物学等理论研究的重要材料, 进行种质资源调查、评价是对其合理保护、利用的前提.目前, 林木种质资源调查、评价研究主要集中于主要经济林树种(
... 使用SPSS 20.0软件对所得表型性状数据进行统计与分析, 分别计算47份供试东北杏种质定量描述性状的平均值($\bar{X}$)、标准偏差(SD)、极小值(Min)、极大值(Max)、变异系数(CV = SD/$\bar{X}$)和K-S值, 并对定性描述性状进行频率统计、计算Shannon- Wiener指数(H′), H′的计算参照
枣种质资源的表型遗传多样性
2
2014
... 种质资源是林木改良的物质基础, 同时也是遗传学、分类学、生物学等理论研究的重要材料, 进行种质资源调查、评价是对其合理保护、利用的前提.目前, 林木种质资源调查、评价研究主要集中于主要经济林树种(
... 使用SPSS 20.0软件对所得表型性状数据进行统计与分析, 分别计算47份供试东北杏种质定量描述性状的平均值($\bar{X}$)、标准偏差(SD)、极小值(Min)、极大值(Max)、变异系数(CV = SD/$\bar{X}$)和K-S值, 并对定性描述性状进行频率统计、计算Shannon- Wiener指数(H′), H′的计算参照
High-level genetic diversity and complex population structure of siberian apricot ( Prunus sibirica L.) in China as revealed by nuclear SSR markers
2014
DB21/T 2462-2015 山杏种质资源调查及评价技术规程
1
2015
... 参照《中国果树志: 杏卷》(
DB21/T 2462-2015 山杏种质资源调查及评价技术规程
1
2015
... 参照《中国果树志: 杏卷》(
The advancement of research on litchi and longan germplasm resources in China
1
2007
... 种质资源是林木改良的物质基础, 同时也是遗传学、分类学、生物学等理论研究的重要材料, 进行种质资源调查、评价是对其合理保护、利用的前提.目前, 林木种质资源调查、评价研究主要集中于主要经济林树种(
暴马丁香不同种源种子和幼苗的表型性状变异及其与地理-气候因子的相关性
1
2016
... 遗传多样性是指物种种内基因组成的多样性, 不同的遗传多样性研究及检测方法存在差异, 可从形态学、细胞学及DNA分子等水平上进行, 其中表型测定是最简单和最直接的方法(
暴马丁香不同种源种子和幼苗的表型性状变异及其与地理-气候因子的相关性
1
2016
... 遗传多样性是指物种种内基因组成的多样性, 不同的遗传多样性研究及检测方法存在差异, 可从形态学、细胞学及DNA分子等水平上进行, 其中表型测定是最简单和最直接的方法(
内蒙古扎兰屯地区西伯利亚杏种质资源的数量分类
1
2015
... 种质资源是林木改良的物质基础, 同时也是遗传学、分类学、生物学等理论研究的重要材料, 进行种质资源调查、评价是对其合理保护、利用的前提.目前, 林木种质资源调查、评价研究主要集中于主要经济林树种(
内蒙古扎兰屯地区西伯利亚杏种质资源的数量分类
1
2015
... 种质资源是林木改良的物质基础, 同时也是遗传学、分类学、生物学等理论研究的重要材料, 进行种质资源调查、评价是对其合理保护、利用的前提.目前, 林木种质资源调查、评价研究主要集中于主要经济林树种(
广西西南桦天然居群的表型变异
1
2005
... 本研究调查区域包括辽宁、吉林和黑龙江的15个地区, 基本覆盖了东北杏的主要分布区, 调查区域内年平均气温为3.4-8.5 ℃, 降水量为530.2- 1 077.8 mm, 由于地理隔离和长期的自然选择, 不同产地的种质形成了相应的遗传特性(
广西西南桦天然居群的表型变异
1
2005
... 本研究调查区域包括辽宁、吉林和黑龙江的15个地区, 基本覆盖了东北杏的主要分布区, 调查区域内年平均气温为3.4-8.5 ℃, 降水量为530.2- 1 077.8 mm, 由于地理隔离和长期的自然选择, 不同产地的种质形成了相应的遗传特性(
基于表型性状和CDDP分子标记延安万花山牡丹遗传多样性分析
1
2019
... 通过主成分分析和聚类分析, 基于表型性状和分子标记均可将47份东北杏种质分为4类, 但从聚类结果分析来看, 2种方法的分类存在较大差异, 在柚(Citrus maxima)(
基于表型性状和CDDP分子标记延安万花山牡丹遗传多样性分析
1
2019
... 通过主成分分析和聚类分析, 基于表型性状和分子标记均可将47份东北杏种质分为4类, 但从聚类结果分析来看, 2种方法的分类存在较大差异, 在柚(Citrus maxima)(
1
2003
... 参照《中国果树志: 杏卷》(
1
2003
... 参照《中国果树志: 杏卷》(
普通杏(Armeniaca vulgaris)种质资源果实主要数量性状变异及概率分级
1
2013
... 遗传参数中变异系数可在一定程度上反映性状的离散程度, 变异系数越大, 离散程度越高, 多样性程度越高(
普通杏(Armeniaca vulgaris)种质资源果实主要数量性状变异及概率分级
1
2013
... 遗传参数中变异系数可在一定程度上反映性状的离散程度, 变异系数越大, 离散程度越高, 多样性程度越高(
西双版纳海拔变化对水湿状况的影响
1
2007
... 随着海拔的升高, 各种气象要素也发生相应变化, 在一定海拔高度范围内, 海拔升高, 温度降低, 大气压减小, 空气所能容纳的水汽含量降低, 进而导致大气相对湿度的增加(
西双版纳海拔变化对水湿状况的影响
1
2007
... 随着海拔的升高, 各种气象要素也发生相应变化, 在一定海拔高度范围内, 海拔升高, 温度降低, 大气压减小, 空气所能容纳的水汽含量降低, 进而导致大气相对湿度的增加(
内蒙古扎兰屯地区山杏群体性状研究
1
2015
... 遗传参数中变异系数可在一定程度上反映性状的离散程度, 变异系数越大, 离散程度越高, 多样性程度越高(
内蒙古扎兰屯地区山杏群体性状研究
1
2015
... 遗传参数中变异系数可在一定程度上反映性状的离散程度, 变异系数越大, 离散程度越高, 多样性程度越高(
尾叶樱桃天然种群叶表型性状变异研究
1
2018
... 遗传多样性是指物种种内基因组成的多样性, 不同的遗传多样性研究及检测方法存在差异, 可从形态学、细胞学及DNA分子等水平上进行, 其中表型测定是最简单和最直接的方法(
尾叶樱桃天然种群叶表型性状变异研究
1
2018
... 遗传多样性是指物种种内基因组成的多样性, 不同的遗传多样性研究及检测方法存在差异, 可从形态学、细胞学及DNA分子等水平上进行, 其中表型测定是最简单和最直接的方法(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}