艾沙江?阿不都沙拉木
1 , 2 , 迪丽娜尔?阿布拉
1 , 张凯
1 , 买热也木古?吐尔逊
1 , 卡迪尔?阿布都热西提
, 1 , 2 , * , 李玲
, 1 , 2 , * 1.
喀什大学生命与地理科学学院, 新疆喀什 844006 2.
喀什大学新疆维吾尔自治区叶尔羌绿洲生态与生物资源自治区高校重点实验室, 新疆喀什 844006 Fruit set and seed germination traits of Zygophyllum kaschgaricum ABDUSALAM Aysajan
1 , 2 , ABULA Dilinaer
1 , Kai ZHANG
1 , TUERXUN Maireyemugu
1 , ABDULRASHID Kadir
, 1 , 2 , * , Ling LI
, 1 , 2 , * 1.
College of Life and Geographic Sciences, Kashi University, Kashi, Xinjiang 844006, China; 2.
The Key Laboratory of Ecology and Biological Resources in Yarkand oasis at Colleges & Universities Under the Department of Education of Xinjiang Uygur Autonomous Region, Kashi University, Kashi, Xinjiang 844006, China; 通讯作者: * Abdulrashid K:
kadir_ks@126.com ;Li L:
liling502@126.com 编委: 王雷
责任编辑: 李敏, 实习编辑: 赵航
收稿日期: 2018-11-17
接受日期: 2019-04-17
网络出版日期: 2019-05-20
基金资助: 国家自然科学基金 (31860121 )国家自然科学基金 (31400279 )新疆维吾尔自治区高层次人才培养计划项目 (QN2016BS0597 )新疆维吾尔自治区高校计划重点项目 (XJEDU2016I042 )
Editorial board: WANG Lei Editor: LI Min, ZHAO Hang Received: 2018-11-17
Accepted: 2019-04-17
Online: 2019-05-20
Fund supported: Supported by the National Natural Science Foundation of China (31860121 )Supported by the National Natural Science Foundation of China (31400279 )The High-level Personnel Training Program of the Xinjiang Uygur Autonomous Region (QN2016BS0597 )The Key Scientific Research Program of the Higher Education Institution of XinJiang (XJEDU2016I042 )
摘要 喀什霸王(
Zygophyllum kaschgaricum )是生长于中国新疆南部荒漠环境的稀有种及二级保护植物。当前, 该物种在自然种群中呈分散式及片段化分布, 且种群密度低, 种群老龄化较严重。因此, 为了了解该物种种子萌发特性及其对荒漠环境的响应, 该文采用室内控制实验方法, 对该物种的自然坐果率、结籽率、种子吸水特性、种子休眠和萌发特性及对干旱胁迫的响应进行了比较研究。结果表明: 喀什霸王在自然种群中的坐果率及结籽率较低, 且种子的败育率较高。不同干藏时间种子的吸水速率间存在显著差异; 随着干藏时间的延长, 种子的吸水率逐步增强。刚成熟的种子在不同温度及光周期下均可萌发; 其中高温(10/20 ℃, 20/30 ℃)及黑暗条件下的萌发率比低温(10/5 ℃, 5/2 ℃)及光照条件下的萌发率高。不同干藏时间的种子在不同浓度赤霉素(GA
3 )下的萌发率均较高; 但低温储藏时间对该物种种子的打破休眠及萌发率没有促进作用。以上结果说明该物种存在非深度生理休眠; 而干藏时间、高温且黑暗及高浓度(50 mmoloL
-1 ) GA
3 是打破休眠及促进种子萌发的最合适条件。高温条件下的干旱胁迫对喀什霸王种子萌发具有抑制作用; 春季和秋季降水量决定种子的萌发率。总之, 喀什霸王种子在物候上表现出的春秋季萌动及非深度生理休眠以提高幼苗存活力及保障种群稳定性, 是一种对新疆南部干旱及高温胁迫荒漠环境的适应策略。
关键词: 种子萌发 ;
干藏时间 ;
吸水特性 ;
非深度生理休眠 ;
暖温带荒漠环境 Abstract Aims Zygophyllum kaschgaricum is a rare and secondary protection plant, which grows in the desert environment in southern Xinjiang, China. At present, this species not only distributes decentralized and fragmentally, but also is low population density and severe ageing of populations in the natural populations. To better understand how this species is adapted to its desert habitat, we studied seed set and its germination characteristics and addressed the following questions. 1) What are the seed set characteristics and water absorption of seeds in different dry storage duration in natural population? 2) What are the seed dormancy and drought stress responses of this species in desert environment?
Methods For the seed set and seed germination characters, we compared the natural seed set of each fruit and water absorption characteristics of each seed in the different dry storage duration, seed dormancy and germination and their responses to drought stress, and the adaptive strategies of this species in southern Xinjiang (Taklimakan desert) natural populations were analyzed.
Important findings The natural fruit and seed set rate of this species is low and seed abortion rate is high in natural populations. There is significant difference in the water absorption capacity of seeds subjected to different durations of dry storage time (p < 0.001), and water absorption capacity of seeds increases with the extension of dry storage time. The seed germination rate increased with the extension of dry storage time under different temperature conditions. The newly matured seeds germinate under different temperature conditions and light cycle (dark 12 h and light 12 h); The germination rate at high temperature (10/20 ℃, 20/30 ℃) and dark conditions is higher than that at low temperature (10/5 ℃, 5/2 ℃) and light conditions. Seed germination rate at different light and temperature conditions under different GA
3 concentration was high, but storage period at low temperature does not promote breaking of dormancy and germination in Z. kaschgaricum seeds. The results suggest that the species has shallow physiological dormancy. Storage at dry environment, high temperature and dark mode conditions, and high concentration (50 mmoloL
-1 ) of GA
3 were the most suitable factors to break dormancy and thus promote seed germination of Z. kaschgaricum. Drought stress under the high temperature inhibited seed germination. The rainfall during spring and autumn is the most important factor to control the seed germination rate. However, shallow physiological dormancy and seed germination at spring and autumn characteristics can improve for the seedling viability and population expansion of this Z. kaschgaricum in the desert, which may be an adaptive strategy to drought and high temperature stress of desert environment in Taklimakan (southern Xinjiang), China.
Keywords: Key words seed germination; ;
dry storage duration; ;
water absorption characteristics; ;
shallow physiological dormancy; ;
warm temperate desert environment; PDF (1635KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 艾沙江?阿不都沙拉木, 迪丽娜尔?阿布拉, 张凯, 买热也木古?吐尔逊, 卡迪尔?阿布都热西提, 李玲. 喀什霸王的结实和种子萌发特性. 植物生态学报, 2019, 43(5): 437-446. DOI:
10.17521/cjpe.2018.0293 Aysajan ABDUSALAM, Dilinaer ABULA, ZHANG Kai, Maireyemugu TUERXUN, Kadir ABDULRASHID, LI Ling.
Fruit set and seed germination traits of Zygophyllum kaschgaricum .
Chinese Journal of Plant Ecology , 2019, 43(5): 437-446. DOI:
10.17521/cjpe.2018.0293 生物多样性保护是有关生态系统安全和稳定性维护的关键科学问题(
Western, 2001 ;
Ren et al ., 2012 ;
覃海宁等, 2017 )。在全球变暖、自然灾害频繁发生、荒漠化严重及种群片段化等内外因素的影响下(
Parmesan & Yohe, 2003 ;
Leadley et al ., 2010 ), 生存在极端环境中的物种受到了不同程度的灭绝威胁(
Western, 2001 ;
张文辉等, 2004 ;
覃海宁等, 2017 )。我国植物多样性丰富, 目前有4 000-5 000种植物正处于不同程度的危机(稀有、濒危、极危)状态中(
Ren et al ., 2012 )。这些植物是生物多样性保护的重要组成部分, 常具有地理分布狭窄、种群规模小、个体数量少以及生境严酷且脆弱等特点(
张文辉等, 2004 ;
Brown et al ., 2015 ;
覃海宁等, 2017 ); 此外, 坐果率、结籽率、种子萌发率和幼苗存活力低等生活史特征也是导致其濒危或灭绝的主要原因(
Neaves et al ., 2015 )。因此, 对该类群的生活史及其对生存环境响应的研究, 在保护稀有濒危植物方面具有重要的科学及实际意义(
Western, 2001 ;
覃海宁等, 2017 )。
与生活史特征相关的种子休眠及幼苗形成过程是种子植物类群后代产生及种群更新的关键阶段(
Baskin & Baskin, 2014 ;
Lu et al ., 2014 ;
孙清琳等, 2019 )。不同植物类群在自身生存及对自然环境的长期适应过程中, 表现出特殊的休眠和萌发特性来保证物种和种群的稳定性(
Nordborg & Bergelson, 1999 ;
Nurulla et al ., 2015 ;
Xia et al ., 2018 )。休眠程度和萌发时间往往受外部因素(光照、温度、水分)及种子自身因素(胚的成熟程度、种皮厚度、种子生活力、调控基因的表达及营养物质)的影响(
Venable et al ., 1987 ;
Casas et al ., 2012 ;
刘波等, 2015 ;
Soltani et al ., 2017 )。根据以上因素的组合程度不同, 种子休眠类型分为物理休眠(PY)、生理休眠(PD)、综合休眠(PY + PD)、形态休眠(MD)和形态生理休眠(MPD) 5种类型(
Galloway & Etterson, 2009 ;
Albrecht & McCarthy, 2011 ;
Baskin & Baskin, 2014 ;
Lu et al ., 2014 ;
Pluntz et al ., 2018 ), 其中生理休眠是自然界中普遍存在且较复杂的休眠类型(
Baskin & Baskin, 2014 )。
生理休眠的植物在极端环境生存及适应过程中表现出较高的生活周期多样性(
Gutterman, 1993 ;
Baskin & Baskin, 2014 ;
Nurulla et al ., 2015 ), 其中非深度生理休眠植物的幼苗存活力往往受到不同环境胁迫(干旱、盐、高温、紫外线)的影响(
Albrecht & McCarthy, 2011 ;
Baskin & Baskin, 2014 )。在干旱环境中普遍存在的干旱及盐胁迫能影响植物种子萌发物候及生活史特点(
Tobe et al ., 2001 ;
Ahmad et al ., 2009 ;
宋乃平等, 2018 )。国内有关研究人员在盐及干旱(缺水)胁迫对寒温带干旱、半干旱荒漠分布植物的生活史特征及其对极端环境的响应(
黄振英等, 2001 ;
Sun et al ., 2009 ;
Lu et al ., 2014 ,
2015 ;
Nurulla et al ., 2015 )方面开展了研究; 但对在我国暖温带干旱荒漠生长的濒危植物种子的生物学特性及其对当地干旱(缺水)环境的响应研究鲜有报道。为此, 本研究对该区域稀有植物喀什霸王(
Zygophyllum kaschgaricum )的结籽率和种子萌发特性进行了研究。
喀什霸王是主要分布于我国塔克拉玛干沙漠西南部的多年生小灌木; 是生长于海拔400-2 100 m荒漠环境的稀有种及新疆二级保护植物(
尹林克, 2006 )。该物种在自然种群中呈分散式的片段化分布, 种群老龄化严重; 此外, 自然环境的年降水量少而温度较高。因此, 我们认为该区域的极端环境(高温及干旱胁迫)及该物种的休眠特点是抑制其种子萌发及种群更新的主要因素。为了证实该推测, 我们对该物种的自然坐果率、结籽率、种子休眠及萌发特性进行了室内比较研究, 为进一步探讨我国塔克拉玛干沙漠西南部荒漠植物在暖温带荒漠环境中的生活史特征及其适应机制研究提供理论资料。
1 材料和方法 1.1 研究材料与研究地点 喀什霸王为隶属于蒺藜科霸王属的小灌木, 主要分布于我国新疆南部(塔克拉玛干沙漠西南部)的荒漠中。该物种平均株高100 cm左右, 最高可达约150 cm, 分枝多, 株丛寿命20年以上。该物种主要以种子繁殖为主, 果实是由三个心皮组成的蒴果, 果翅由果皮发育而来, 每室有1个胚珠(
尹林克, 2006 )。
本研究的果实采集于新疆阿图什市格达良荒漠化草原的自然种群(地理坐标为39.83° N, 76.61° E,海拔1 200 m); 已采集的果实装入纸袋后保存在室内(温度为23-26 ℃, 相对湿度为30-35 %)备用。研究地点位于塔克拉玛干沙漠西南部, 属于大陆性暖温带荒漠气候。根据阿图什气象局提供的数据(2015-2017年), 当地年平均气温14 ℃, 7月气温最高为28 ℃, 最低气温出现在1、4和9月, 平均气温为20 ℃, 平均年降水量为118.6 mm, 8月降水量最高为35.2 mm, 1和12月降水量最低为1 mm, 9月降水量平均为10 mm (
图1 )。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1喀什霸王自然种群的每月累计降水量和月平均气温(2015-2017年)(平均值±标准误差)。 Fig. 1Monthly precipitation and monthly mean air temperatures in natural population of Zygophyllum kaschgaricum in (2015-2017)(mean ± SE ). 1.2 研究方法 1.2.1 自然结籽率及种子吸水特性的测定
1.2.1.1 自然结籽率 为了判断该物种的自然结籽率, 随机选择刚采回来的200个果实, 统计每个果实的结籽率及完整种子数。用便携式SE 2000型电子游标卡尺(桂林迪吉特电子有限公司, 广西桂林, 中国; 精度为0.02 mm)及FA1004型电子天平(华丰电子仪器厂, 浙江宁波, 中国; 精度为0.000 1 g), 测量每粒种子长宽度及10粒种子的质量; 各重复5次。
1.2.1.2 吸水特性 为了判断种子在休眠及非休眠状态时吸水特点的差异, 自2017年6月开始, 从刚采回来(0个月)及室温条件下干藏6个月的饱满种子中各选取50粒, 每个时间段的种子均分5个组(每个组10粒); 用电子天平测每组种子的质量, 然后将每组种子置于2层滤纸的培养皿(直径为9 cm)中, 加适量蒸馏水, 隔1 h称量一次(直到种子质量不再增加为止); 最后比较不同干藏时间种子的吸水速率。按照
Baskin和Baskin (2004) 的公式:
W r = ((
W f -
W i )/
W i ) × 100, 计算种子吸水速率(
W r ), 其中
W i 及
W f 是吸水前和吸水1 h后的质量。
1.2.2 室内干藏时间对种子萌发的影响
为了判断种子生理后熟对种子萌发的影响, 将从自然环境采集, 后在室内温度条件下干藏0、3、6及9个月的种子放于直径9 cm的培养皿里(每个培养皿25粒种子, 重复3次), 然后置于5/2 ℃、15/5 ℃、20/10 ℃及30/20 ℃ 4个不同温度、光照/黑暗(培养皿用不透光的黑色袋子包裹)各12 h条件下的变温光照培养箱中培养28天。每天统计一次各培养皿中种子的萌发率, 并加入2 mL蒸馏水以保持湿度一致。本实验的4个温度代表该物种自然生长种群(格达良荒漠环境)不同月的日平均、最高和最低气温: 30/15 ℃ (5月、7月、8月)、20/10 ℃ (4月和10月)、15/5 ℃ (3月和11月)及5/2 ℃ (12月和2月)。实验结束后参考
Baskin和Baskin (2004) 的TTC (0.1%的氯化三苯四氮唑溶液)处理方法, 对各组未萌发的种子活力进行检测。TTC处理后的种胚在20 ℃全黑暗条件下放置24 h, 有活力的种子将被染成红或粉红色。
1.2.3 低温储藏对种子萌发的影响
为了确定低温储藏对种子萌发的影响, 将刚从野外采回来的种子晾干后装到塑料袋(25 cm × 20 cm)中, 并在4 ℃的冷藏箱储藏0、3及6个月后, 将种子置于5/2 ℃、15/5 ℃、20/10 ℃及30/20 ℃ 4个温度、12 h光照/12 h黑暗条件下培养28天。每24 h统计一次萌发率。
1.2.4 赤霉素处理对种子萌发的影响
为了判断赤霉素(GA
3 )浓度对该物种休眠及非休眠状态种子萌发的影响, 刚成熟(0个月)及室温条件下干藏6个月的种子均分别置于0 (蒸馏水)、1、10及50 mmoloL
-1 浓度的GA
3 溶液(置于直径为9 cm的培养皿)中, 在5/2 ℃、15/5 ℃、20/10 ℃、30/20 ℃温度及不同光周期(每天光照/黑暗各12 h或全黑暗两种)的变温培养箱里培养28天, 其中向全黑暗处理的培养皿里加5 mL的不同浓度GA
3 溶液后用封口膜密封培养皿并用不透光的黑色袋子包裹, 以防止水分蒸发引起水势变动影响处理。为了保持不同光照及温度处理的培养皿中GA
3 溶液浓度一致, 每天对每个培养皿加0.5 mL不同浓度的GA
3 溶液, 每天统计一次萌发率。
1.2.5 室内干旱胁迫(PEG)对种子萌发的影响
为了判断干旱胁迫对喀什霸王种子萌发的影响, 将刚成熟(0个月)及室温度条件下干藏6个月的种子置于5%、10%、15%、20%的聚乙二醇(PEG 6000)溶液培养皿中, 在5/2 ℃、15/5 ℃、20/10 ℃及30/20 ℃ 4个温度和不同光周期(每天光照/黑暗各12 h)条件下培养28天。为了维持培养皿小环境溶液的浓度用封口膜密封培养皿, 以防止水分蒸发引起水势变动。每天统计一次萌发率。
1.3 数据处理 本文的所有数据在SPSS 16.0 (SPSS, Chicago, USA)统计软件中进行分析。首先对所有数据在分析前均进行正态分布和方差齐性检验, 对非正态分布或方差不齐的数据进行对数或平方根函数转换; 当所有数据符合方差齐性后, 对结籽率、吸水特性和种子萌发率进行单因素方差分析、多因素方差分析或独立样本
t 检验。Tukey’s HSD用于检验处理间多重比较的差异显著性(
α = 0.05)。统计数据以平均值±标准误差(mean ±
SE )表示。采用SigmaPlot 10.0软件进行绘图。
2 结果和分析 2.1 自然坐果率、结籽率及种子吸水特性 喀什霸王果实的自然坐果率及结籽率分别为(74 ± 3.25)%和(46.97 ± 2.56)%, 其中每个果实的平均结籽数为1.93粒。其种子为肾形, 种皮呈褐色或黄褐色, 种子长度为(0.846 ± 0.024) cm, 宽度为(0.219 ± 0.004) cm。不同干藏处理种子的百粒质量间存在显著差异(
t = 29.544,
p < 0.001), 其中刚成熟种子的百粒质量(1.218 ± 0.044) g大于干藏6个月种子的百粒质量(0.866 ± 0.068) g。种子的吸水率受到干藏时间(
F = 210.111,
p < 0.001)、吸水时间(
F = 16.900,
p < 0.001)的影响(
图2 ), 但未受到二者间相互作用(
F = 1.804,
p = 0.115)的影响; 其中干藏6个月种子的吸水率比刚成熟种子的吸水率强, 随着干藏时间的延长, 种子对水分的需求增高(
图2 )。两种干藏处理种子吸水7 h后质量增加百分比为(66.13 ± 5.23)%及(97.27 ± 3.20)%, 此时吸水达到饱和(
图2 )。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2不同干藏时间喀什霸王种子的吸水曲线(平均值±标准误差)。 Fig. 2Water uptake curves of Zygophyllum kaschgaricum seeds for different duration of dry storage at room temperatures (mean ± SE ). 2.2 室内干藏时间对种子萌发的影响 喀什霸王种子的萌发率随着干藏时间的延长而逐渐增高。种子萌发率显著受干藏时间(
p < 0.001)、温度(
p < 0.001)、光照(
p < 0.001)、干藏时间和温度交互作用(
p < 0.001)、温度和光照交互作用(
p < 0.001)以及干藏时间、光照和温度交互作用(
p < 0.001)的影响(
图3 ;
表1 ), 但未受到干藏时间和光照交互作用(
p = 0.170)的影响; 其中不同温度处理种子的萌发率随着干藏时间的延长而增高; 说明干藏时间是打破该物种种子休眠及提高种子萌发率的一个主要因素。此外, 黑暗条件下的种子萌发率高于光照条件下的萌发率(
图3 ), 而不同干藏时间处理种子在高温(30/20 ℃及20/10 ℃)条件下的萌发率高于低温(5/2 ℃及15/5 ℃)条件下的萌发率; 其中干藏9个月种子在不同光照(黑暗及光照)及高温条件下的萌发率高于80%, 尤其是黑暗30/20 ℃处理种子的最终萌发率达到95%; 虽然低温处理种子的萌发率随着干藏时间的延长而提高, 但干藏9个月的最终萌发率往往低于60% (
图3 ); 说明高温及黑暗能提高种子萌发率。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3室内干藏不同时间段(0、3、6及9个月)的喀什霸王种子在不同温度光照(A)及全黑暗(B)条件下的萌发率(平均值±标准误差)。不同小写字母表示同一温度不同干藏时间间存在显著差异(p < 0.05)。 Fig. 3Effects of dry storage period (0、3、6 and 9 months) at room temperature on the germination rate of Zygophyllum kaschgaricum seeds in different temperature under light (A) and darkness (B) (mean ± SE ). Bars with different lowercase letters indicate significant differences between different dry storage periods within a temperature range treatment (p < 0.05). Table 1 表1 表1 光照条件、温度和干藏时间及三者之间交互作用对喀什霸王种子萌发率的影响
Table 1
Three-way ANoVA of effects of light condition, temperature and different dry storage period and their interactions on seed germination percentages of Zygophyllum kaschgaricum 处理 自由度 平方和 均方 F p 温度 3 31 596.125 10 532.042 1 040.202 <0.001 光照 Light (B) 1 1 584.375 1 584.375 156.481 <0.001 干藏时间 3 17 573.458 857.819 578.550 <0.001 A × B 3 339.125 113.042 11.165 <0.001 A × C 9 1 966.375 218.486 21.578 <0.001 B × C 3 52.458 17.486 1.727 0.307 A × B × C 9 902.042 100.227 9.898 <0.001 合计 Total 96 329 652
新窗口打开 |
下载CSV 2.3 低温储藏对种子萌发的影响 喀什霸王种子的萌发率受温度(
F = 765.741,
p < 0.001)、光照(
F = 94.127,
p < 0.001)、低温储藏时间(
F = 46.127,
p < 0.001)及温度和光照(
F = 16.906,
p < 0.001)、温度和低温储藏时间(
F = 22.970,
p < 0.001)及三者交互作用的影响(
F = 2.315,
p < 0.05), 但种子萌发率未受到光照和低温储藏时间交互作用(
F = 0.910,
p = 0.409)的影响。随着低温储藏时间的延长, 同一温度梯度光照和黑暗条件下种子的萌发率未发生变化(
图4 ), 但低温储藏处理的种子在高温条件下的萌发率较高。说明低温储藏时间对该物种种子的打破休眠及萌发率未有促进作用。
图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4不同低温储藏时间(0, 3及6个月)处理的喀什霸王种子在不同温度光照(A)及全黑暗(B)条件下的萌发率(平均值±标准误差)。不同小写字母表示同一温度不同低温储藏时间间存在显著差异(p < 0.05)。 Fig. 4Effects of storage periods (0, 3 and 6 months) at low temperature on the germination rate of Zygophyllum kaschgaricum seeds in different temperature under light (A) and darkness (B) (mean ± SE ). Bars with different lowercase letters indicate significant differences between different storage periods within a temperature range treatment (p < 0.05). 2.4 赤霉素处理对种子萌发的影响 刚成熟及干藏6个月种子的萌发率受GA
3 溶液浓度的影响(
图5 ); 其中不同干藏时间种子的萌发率随着GA
3 浓度的提高而逐渐提高。GA
3 存在时光照(
F = 1.715,
p = 0.167)、干藏和光照(
F = 0.336,
p = 0.800)、温度和光照(
F = 0.685,
p = 0.721)及干藏时间、温度和光照(
F = 1.350,
p = 0.218)的交互作用对种子萌发率没有显著的影响(
图5 ;
表2 ); 尤其是在10 mmoloL
-1 及50 mmoloL
-1 的GA
3 浓度下高温处理种子的(30/20 ℃及20/10 ℃)萌发率高于90%; 而50 mmoloL
-1 的GA
3 浓度下低温处理种子(5/2 ℃及15/5 ℃)的萌发率也达到80%左右。说明GA
3 浓度能打破喀什霸王种子休眠; 其中50 mmoloL
-1 的GA
3 浓度是打破种子休眠的最合适浓度。
图5 新窗口打开 |
下载原图ZIP |
生成PPT 图5喀什霸王刚成熟(A, B)及干藏6个月(C, D)处理的种子在不同GA3 浓度及不同温度光照(A, C)及全黑暗(B, D)条件下的萌发率(平均值±标准误差)。不同小字母表示同一温度不同GA3 浓度间存在显著差异(p < 0.05)。 Fig. 5Germination rate of Zygophyllum kaschgaricum seeds from post-harvest (A, B) and dry storage for 6 months (C, D) in different GA3 concentrations and temperature under light (A, C) and darkness (B, D) (mean ± SE ). Bars with different lowercase letters indicate significant differences between GA3 concentrations within a temperature range treatment (p < 0.05). Table 2 表2 表2 GA
3 浓度、光照条件、温度和干藏时间及四者之间交互作用对喀什霸王种子萌发的影响
Table 2
Four-way ANoVA of effects of light condition, temperature, GA3 concentration and different dry storage period and their interactions on seed germination rate of Zygophyllum kaschgaricum 处理Treatment 自由度 平方和 均方 F p 干藏时间 1 6 007.688 6 007.688 283.06 <0.001 温度 3 37 412.38 12 470.79 587.58 <0.001 光照Light (C) 1 2 338.02 2 338.02 110.16 <0.001 赤霉素 GA3 (D) 3 38 984.708 12 994.903 612.275 <0.001 A × B 3 987.85 329.29 15.52 <0.001 A × C 1 70.08 70.08 3.30 <0.072 A × D 3 2 607.85 869.29 40.96 <0.001 B × C 3 312.60 104.20 4.91 <0.001 B × D 9 5 908.83 656.54 30.93 <0.001 C × D 3 109.19 36.40 1.72 0.167 A × B × C 3 174.29 58.09 2.74 <0.05 A × B × D 9 1 051.60 116.85 5.51 <0.001 A × C × D 3 21.38 7.13 0.34 0.800 B × C × D 9 130.85 14.54 0.69 0.721 A × B × C × D 9 257.92 28.66 1.35 0.218 合计 Total 192 1 047 186
新窗口打开 |
下载CSV 2.5 室内干旱胁迫(PEG)对种子萌发的影响 刚成熟及干藏6个月种子的萌发率受PEG溶液浓度的影响。干旱胁迫程度(
F = 254.30,
p < 0.001)、光照(
F = 239.66,
p < 0.001)、温度(
F = 479.74,
p < 0.001)、干藏时间(
F = 593.52,
p < 0.001)以及四因素间的不同交互作用对种子萌发有一定的负面影响(
图6 ); 但干藏时间、温度及干旱胁迫程度(
F = 1.46,
p = 0.171)以及四因素共同的交互作用(
F = 0.986,
p = 0.455)对种子萌发没有影响。随着PEG浓度的提高, 喀什霸王种子的萌发率逐渐降低, 其中PEG对高温处理种子萌发率的影响比低温处理大(
图6 )。此外, PEG浓度对刚成熟和干藏6个月的种子萌发率的影响间存在显著差异(
p < 0.001); 其中PEG对干藏6个月种子萌发率的负面影响(
p < 0.001)比刚成熟种子大; 20%浓度的PEG对20/30 ℃及10/20 ℃条件下处理种子萌发率的负面影响最显著。说明干旱胁迫是抑制喀什霸王种子萌发的主要因素(
图6 )。
图6 新窗口打开 |
下载原图ZIP |
生成PPT 图6喀什霸王刚成熟(A, B)及室内干藏6月(C, D)处理的种子在不同浓度PEG溶液及不同温度光照(A, C)及全黑暗(B, D)条件下的萌发率(平均值±标准误差)。不同小写字母表示同一温度不同PEG浓度间存在显著差异(p < 0.05)。 Fig. 6Germination rate of Zygophyllum kaschgaricum seeds from post-harvest (A, B) and dry storage for 6 months (C, D) in different PEG concentrations and temperature under light (A, C) and darkness (B, D) (mean ± SE ). Bars with different lowercase letters indicate significant differences between PEG concentrations within a temperature range treatment (p < 0.05). 3 讨论 3.1 果实结籽率及种子休眠及萌发特性 在严酷的极端荒漠环境中, 植物的坐果率和结籽率、种子休眠程度及幼苗存活率能决定植物生活史特征及种群的更新水平。其中果实坐果率及结籽率与植物交配方式及传粉效率有关, 而休眠程度、种子萌发率及幼苗存活率与自然环境的温度和水分等外部因素有关(
黄振英等, 2001 ;
Baskin & Baskin, 2004 ;
Ahmad et al ., 2009 ;
Pluntz et al ., 2018 ;
Xia et al ., 2018 )。本研究中, 喀什霸王果实的自然坐果率及结籽率较低, 因为在其开花的4月中旬, 塔克拉玛干沙漠西南部荒漠环境的昼夜温度差异及风速较大, 早春活动的昆虫种类少; 此外, 该物种存在花内雌性先熟现象(未发表数据)。以上因素导致的传粉限制可能是降低自然坐果率及结籽率(46.97 ± 2.56)%的主要因素, 此推测需要进一步验证。
生理休眠植物中有非深度生理休眠的新鲜种子在任何温度及光照下均不能萌发或只能在很窄的温度或光照范围内萌发(
Sun et al ., 2009 ;
Baskin & Baskin, 2014 ;
刘波等, 2015 ;
Soltani et al ., 2017 )。喀什霸王刚采回来的种子在不同温度、光照或黑暗条件下均能萌发, 且彼此间存在显著差异(
p < 0.01,
图3 ); 其中高温黑暗(20/10 ℃及30/20 ℃)处理种子的萌发率高于低温(5/2 ℃和15/2 ℃)黑暗处理种子的萌发率。随着贮藏时间的延长, 贮藏种子的萌发率逐渐提高, 而休眠逐渐解除, 各温变周期下的种子萌发率逐渐提高(
图3 ); 但干藏9个月的种子在低温条件下萌发率低于60%。虽然该物种种子在各温度和不同光周期下均萌发, 但高温和黑暗是最佳的萌发条件。此外, 刚成熟及干藏6个月的种子在不同浓度的GA
3 处理后, 同一浓度GA
3 在各温变周期(光照/黑暗)下的不同干藏处理种子的萌发率间未存在显著差异(
p > 0.05); 但不同浓度GA
3 处理种子的萌发率间存在显著差异(
p < 0.05); 其中50 mmoloL
-1 浓度的萌发率最高, 是打破种子休眠的最合适浓度(
图3 )。从以上结果可以判断喀什霸王种子的休眠类型属于非深度生理休眠类型, 这与我国北部寒温带分布的石生霸王(
Zygophyllum rosovii )和石炭酸灌木(
Larrea divaricate )的生理休眠表现相似(
Venable et al ., 1987 ;
Mandák, 2001 ;
Baskin & Baskin, 2014 )。喀什霸王种子的非深度生理休眠特征可能与该物种长期适应南疆秋天和春天较短的光日照时间及高温过程中进化出的种子萌发对策有关。
3.2 干旱胁迫对种子萌发的影响 种子的不同季节萌发能促进植物物候的多样性(
Baskin & Baskin, 1988 ;
刘波等, 2015 ); 春秋季萌发是种子植物在不可预测环境的两头下注策略和种子生理后熟之间的权衡(
Lu et al ., 2014 )。在本研究中, 室内不同浓度PEG处理种子在低温条件下的萌发率比高温高(
图5 )。此外, 随着干藏时间的延长, 种子对水分的需求提高(
图1 )。塔克拉玛干沙漠西南部自然环境的平均月降水量低于10 mm, 该物种果实成熟的6-9月的平均气温在25 ℃以上, 气候特点与以色列的Negev荒漠(
Gutterman, 1993 ,
2000 )相似。因此, 该区域春季的降水量、气温及光照强度等气候特点能限制有非深度生理休眠种子的萌发及幼苗存活率。在降水时间分布不均匀的塔克拉玛干沙漠西南部荒漠环境中(
图1 ), 该物种种子的萌发对策主要是对降水做出响应(
Tobe et al ., 2001 ;
Ahmad et al ., 2009 ), 其中种子的春季萌发及幼苗形成过程被认为是一种逃避冬天死亡的有利方式, 而秋天萌发是种子对低温的适应及竞争优势(
Gutterman, 1993 )。喀什霸王干藏6个月的种子在水分充足时才萌发, 说明该物种在塔克拉玛干沙漠西南部的极端环境的长期生存过程中, 随时面临着自然环境难以预测的生存风险, 通过种子的非深度生理休眠可获得更多的生存机会, 保持物种亲本性状。
3.3 休眠特性对种群更新的影响 种子萌发作为植物生活周期的主要阶段, 可影响植物雌性(种子)适合度和种群维持及扩大能力(
Baskin & Baskin, 2014 ;
Copete et al ., 2015 )。被子植物在种子物候上所表现出的异质性(
Kalisz & Wardle, 1994 ;
Nordborg & Bergelson, 1999 )作为一种可避免种群灭绝的、谨慎的萌发对策(
Gutterman, 2000 ), 有利于在荒漠极端环境中生长的植物种群的维持和更新(
姚红和谭敦炎, 2005 ;
Nurulla et al ., 2015 )。在本研究中, 非深度生理休眠的喀什霸王种子在春季和秋季均萌发, 虽然该物种在秋季存在一定程度的休眠。在自然环境中, 春季和秋季的温度较低而降水量较多, 因此, 该物种种子通过春秋季萌动的繁殖策略及非深度生理休眠来维持种群的动态平衡及保证种群的更新; 但4-6月气温的上升及降水量的减少导致的干旱胁迫可降低喀什霸王种子的萌发率或幼苗存活能力(
Tobe et al ., 2001 ), 说明新疆南部荒漠环境中降水量是影响非深度生理休眠植物喀什霸王种群更新能力的主要因素(
Lu et al ., 2014 ;
Nurulla et al ., 2015 )。总之, 种子物候上表现出的春秋季萌动繁殖策略及非深度生理休眠是该物种在干旱胁迫及高温环境中为保持种群稳定性而演化出的一种适应策略。在今后的研究中, 喀什霸王幼苗存活力及种群更新与环境因子的关系等方面还需要进一步的研究。
[1] Ahmad S Ahmad R Ashraf MY Ashraf M Waraich EA 2009 ). Sunflower (Helianthus annuus L.) response to drought stress at germination and seedling growth stagesPakistan Journal of Botany, 647 -654 . [本文引用: 3] [2] Albrecht MA McCarthy BC 2011 ). Variation in dormancy and germination in three co-occurring perennial forest herbsPlant Ecology, 212, 1465 -1477 . [本文引用: 2] [3] Baskin CC Baskin JM 1988 ). Germination ecophysiology of herbaceous plant species in a temperate regionAmerican Journal of Botany, 75, 286 -305 . [本文引用: 1] [4] Baskin CC Baskin JM 2014 ). Seed: Ecology, Biogeography, and Evolution of Dormancy and Germination . Academic Press, San Diego, USA. 215 -375 . [5] Baskin JM Baskin CC 2004 ). A classification system for seed dormancySeed Science Research, 14, 1 -16 . [本文引用: 3] [6] Brown AR owena SF Petersa J 2015 ). Climate change and pollution speed declines in zebra fish populationsProceedings of the National Academy of the Sciences of the United States of America, 112, 1237 -1246 . [本文引用: 1] [7] Casas RR Kovach K Dittmar E 2012 ). Seed after-ripening and dormancy determine adult life history independently of germination timingNew Phytologist, 194, 868 -879 . [本文引用: 1] [8] Copete MA Herranz JM Ferrandis P 2015 ). Annual dormancy cycles in buried seeds of shrub species: Germination ecology of Sideritis serrata(Labiatae)Plant Biology, 798 -807 . [本文引用: 1] [9] Galloway LF Etterson JR 2009 ). Plasticity to canopy shade in a monocarpic herb: Within- and between-generation effectsNew Phytologist, 182, 1003 -1012 . [本文引用: 1] [10] Gutterman Y 1993 ). Seed Germination in Desert Plants Springer-Verlag, Berlin Heidelberg. [本文引用: 3] [11] Gutterman Y 2000 ). Environmental factors and survival strategies of annual plant species in the Negev Desert, IsraelPlant Species Biology, 15, 113 -125 . [本文引用: 2] [12] Huang ZY Gutterman Y Hu ZH Zhang XS 2001 ). Seed germination in Artemisia sphaerocephala II. The influencing of environmental factorsActa Phytoecologica Sinica, 240 -246 . [本文引用: 2] [ 黄振英 , Gutterman Y 胡正海 , 张新时 (2001 ). 白沙蒿种子萌发特性的研究II. 环境因素的影响植物生态学报 , 25, 240 -246 .] [本文引用: 2] [13] Kalisz S Wardle GM 1994 ). Life history variation in Campanula americana(Campanulaceae): Population differentiationAmerican Journal of Botany, 521 -527 . [本文引用: 1] [14] Leadley P Pereira HM Alkemade R 2010 ). Biodiversity Scenarios: Projections of 21st Century Change in Biodiversity and Associated Ecosystem Services Secretariat of the Convention on Biological Diversity, Montreal . 121 -133 . [本文引用: 1] [15] Liu B Lü XG Jiang M Zhang WG Wu HT 2015 ). Effects of light and water depth on seed germination of Phragmites australis in the wetlands of Songnen PlainChinese Journal of Plant Ecology, 616 -620 . [本文引用: 3] [ 刘波 , 吕宪国 , 姜明 , 张文广 , 武海涛 (2015 ). 光照、水深交互作用对松嫩湿地芦苇种子萌发的影响植物生态学报 , 39, 616 -620 .] [本文引用: 3] [16] Lu JJ Tan DY Baskin JM Baskin CC 2014 ). Germination season and watering regime, but not seed morph, affect life history traits in a cold desert diaspore-heteromorphic annualPLoS oNE, e102018 . DOI:10.1371/journal.pone.0102018. [本文引用: 13] [17] Lu JJ Zhou YM Tan DY Baskin CC Baskin JM 2015 ). Seed dormancy in six cold desert Brassicaceae species with indehiscent fruitsSeed Science Research, 25, 276 -285 . [本文引用: 1] [18] Mandák B 2001 ). Germination requirements of invasive and non-invasive Atriplex species: A comparative studyFlora, 45 -54 . [本文引用: 1] [19] Neaves LE Eales J Whitlock R 2015 ). The fitness consequences of inbreeding in natural populations and their implications for species conservation—A systematic mapEnvironmental Evidence, 4, 2 -17 . [本文引用: 1] [20] Nordborg M Bergelson J 1999 ). The effect of seed and rosette cold treatment on germination and flowering time in some Arabidopsis thaliana(Brassicaceae) ecotypesAmerican Journal of Botany, 470 -475 . [本文引用: 2] [21] Nurulla M Baskin CC Lu JJ Tan DY Baskin JM 2015 ). Intermediate morphophysiological dormancy allows for life-cycle diversity in the annual weed,Turgenia latifolia(Apiaceae)Australian Journal of Botany, 630 -637 . [本文引用: 5] [22] Parmesan C Yohe G 2003 ). A globally coherent fingerprint of climate change impacts across natural systemsNatur e, 37 -42 . [本文引用: 1] [23] Pluntz M Coz SL Peyrard N Pradel R Choquet R Cheptou Po 2018 ). A general method for estimating seed dormancy and colonisation in annual plants from the observation of existing floraEcology Letter s, 1311 -1318 . [本文引用: 2] [24] Qin HN Zhao LN Yu SX Liu HY Liu B Xia NH Peng H Li ZY Zhang ZX He XJ Yin LK Lin YL Liu QR Hou YT Liu Y Liu QX Cao W Li JQ Chen SL Jin XH Gao TG Chen WL Ma HY Geng YY Jin XF Chang CY Jiang H Cai L Zang CX Wu JY Ye JF Lai YJ Liu B Lin QW Xue NX 2017 ). Evaluating the endangerment status of China’s angiosperms through the red list assessmentBiodiversity Science, 25, 745 -757 . [本文引用: 4] [ 覃海宁 , 赵莉娜 , 于胜祥 , 刘慧圆 , 刘博 , 夏念和 , 彭华 , 李振宇 , 张志翔 , 何兴金 , 尹林克 , 林余霖 , 刘全儒 , 侯元同 , 刘演 , 刘启新 , 曹伟 , 李建强 , 陈世龙 , 金效华 , 高天刚 , 陈文俐 , 马海英 , 耿玉英 , 金孝锋 , 常朝阳 , 蒋宏 , 蔡蕾 , 臧春鑫 , 武建勇 , 叶建飞 , 赖阳均 , 刘冰 , 林秦文 , 薛纳新 (2017 ). 中国被子植物濒危等级的评估生物多样性 , 25, 745 -757 .] [本文引用: 4] [25] Ren H Zhang QM Lu HF Liu HX Guo QF Wang J Jian SG Bao Ho 2012 ). Wild plant species with extremely small populations require conservation and reintroduction in ChinaAMBIo , 41, 913 -917 . [本文引用: 2] [26] Soltani E Gruber S oveisi M Salehi N Alahdadi I Javid MG 2017 ). Water stress, temperature regimes and light control induction, and loss of secondary dormancy in Brassica napus L. seedsSeed Science Research, 217 -230 . [本文引用: 2] [27] Song NP Wang X Chen L Xue Y Chen J Sui JM Wang L Yang XG 2018 ). Co-existence mechanisms of plant species within “soil islands” habitat of desert steppeBiodiversity Science, 26, 667 -677 . [本文引用: 1] [ 宋乃平 , 王兴 , 陈林 , 薛毅 , 陈娟 , 随金明 , 王磊 , 杨新国 (2018 ). 荒漠草原“土岛”生境群落物种共存机制生物多样性 , 26, 667 -677 .] [本文引用: 1] [28] Sun HZ Lu JJ Tan DY Baskin JM Baskin CC 2009 ). Dormancy and germination characteristics of the trimorphic achenes of Garhadiolus papposus( Asteraceae), an annual ephemeral from the Junggar Desert, ChinaSouth African Journal of Botany, 537 -545 . [本文引用: 2] [29] Sun QL Zhang RH Yi SG Yang L Li QY Zhou JH Lai LM Jiang LH Zheng YR 2019 ). Responses of germination and seedling emergence of Stipa bungeana to key environmental factorsActa Ecologica Sinica, 2034 -2042 . [本文引用: 1] [ 孙清琳 , 张瑞红 , 易三桂 , 杨柳 , 李巧燕 , 周继华 , 来利明 , 姜联合 , 郑元润 (2019 ). 长芒草(Stipa bungeana )种子萌发与出苗对关键环境因子的响应生态学报 , 39, 2034 -2042 .] [本文引用: 1] [30] Tobe K Zhang L Yu QG 2001 ). Characteristics of seed germination in five non-halophytic Chinese desert shrub speciesJournal of Arid Environments, 47, 191 -201 . [本文引用: 3] [31] Venable DL Búrquez A Corral G Morales E Espinosa F 1987 ). The ecology of seed heteromorphism in Heterosperma pinnatum in central MexicoEcology 65 -76 . [本文引用: 2] [32] Western D 2001 ). Human-modified ecosystems and future evolutionProceedings of the National Academy of Sciences of United States of America, 98, 5458 -5465 . [本文引用: 3] [33] Xia Q Maharajah P Cueff G Rajjouc L Prodhommed D Gibond Y Baillya C Corbineaua F Meimouna P El-Maarouf-Bouteau H 2018 ). Integrating proteomics and enzymatic profiling to decipher seed metabolism affected by temperature in seed dormancy and germinationPlant Science, 269, 118 -125 . [本文引用: 2] [34] Yao H Tan DY 2005 ). Size-dependent reproductive output and life-history strategies in four ephemeral species of Trigonella Acta Phytoecologica Sinica, 954 -960 . [本文引用: 1] [ 姚红 , 谭敦炎 (2005 ). 胡卢巴属4种短命植物个体大小依赖的繁殖输出与生活史对策植物生态学报 , 29, 954 -960 .] [本文引用: 1] [35] Yin LK 2006 ). Rare, Endangered and Endemic Higher Plants in Xinjiang . Xinjiang Science and Technology Press, ürü mqi. 35 -45 . [ 尹林克 (2006 ). 新疆珍稀濒危特有高等植物 . 新疆科学技术出版社, 乌鲁木齐. 35 -45 .] [36] Zhang WH Xu XB Zhou JY 2004 ). Distribution and bio- ecological characteristics of Abies chensiensis , an endangered plantChinese Biodiversity, 419 -426 . [本文引用: 4] [ 张文辉 , 许晓波 , 周建云 (2004 ). 濒危植物秦岭冷杉地理分布和生物生态学特性研究生物多样性 , 12, 419 -426 .] [本文引用: 4] Sunflower (
Helianthus annuus L.) response to drought stress at germination and seedling growth stages
3
2009
... 生理休眠的植物在极端环境生存及适应过程中表现出较高的生活周期多样性(
Gutterman, 1993 ;
Baskin & Baskin, 2014 ;
Nurulla et al ., 2015 ), 其中非深度生理休眠植物的幼苗存活力往往受到不同环境胁迫(干旱、盐、高温、紫外线)的影响(
Albrecht & McCarthy, 2011 ;
Baskin & Baskin, 2014 ).在干旱环境中普遍存在的干旱及盐胁迫能影响植物种子萌发物候及生活史特点(
Tobe et al ., 2001 ;
Ahmad et al ., 2009 ;
宋乃平等, 2018 ).国内有关研究人员在盐及干旱(缺水)胁迫对寒温带干旱、半干旱荒漠分布植物的生活史特征及其对极端环境的响应(
黄振英等, 2001 ;
Sun et al ., 2009 ;
Lu et al ., 2014 ,
2015 ;
Nurulla et al ., 2015 )方面开展了研究; 但对在我国暖温带干旱荒漠生长的濒危植物种子的生物学特性及其对当地干旱(缺水)环境的响应研究鲜有报道.为此, 本研究对该区域稀有植物喀什霸王(
Zygophyllum kaschgaricum )的结籽率和种子萌发特性进行了研究. ...
... 在严酷的极端荒漠环境中, 植物的坐果率和结籽率、种子休眠程度及幼苗存活率能决定植物生活史特征及种群的更新水平.其中果实坐果率及结籽率与植物交配方式及传粉效率有关, 而休眠程度、种子萌发率及幼苗存活率与自然环境的温度和水分等外部因素有关(
黄振英等, 2001 ;
Baskin & Baskin, 2004 ;
Ahmad et al ., 2009 ;
Pluntz et al ., 2018 ;
Xia et al ., 2018 ).本研究中, 喀什霸王果实的自然坐果率及结籽率较低, 因为在其开花的4月中旬, 塔克拉玛干沙漠西南部荒漠环境的昼夜温度差异及风速较大, 早春活动的昆虫种类少; 此外, 该物种存在花内雌性先熟现象(未发表数据).以上因素导致的传粉限制可能是降低自然坐果率及结籽率(46.97 ± 2.56)%的主要因素, 此推测需要进一步验证. ...
... 种子的不同季节萌发能促进植物物候的多样性(
Baskin & Baskin, 1988 ;
刘波等, 2015 ); 春秋季萌发是种子植物在不可预测环境的两头下注策略和种子生理后熟之间的权衡(
Lu et al ., 2014 ).在本研究中, 室内不同浓度PEG处理种子在低温条件下的萌发率比高温高(
图5 ).此外, 随着干藏时间的延长, 种子对水分的需求提高(
图1 ).塔克拉玛干沙漠西南部自然环境的平均月降水量低于10 mm, 该物种果实成熟的6-9月的平均气温在25 ℃以上, 气候特点与以色列的Negev荒漠(
Gutterman, 1993 ,
2000 )相似.因此, 该区域春季的降水量、气温及光照强度等气候特点能限制有非深度生理休眠种子的萌发及幼苗存活率.在降水时间分布不均匀的塔克拉玛干沙漠西南部荒漠环境中(
图1 ), 该物种种子的萌发对策主要是对降水做出响应(
Tobe et al ., 2001 ;
Ahmad et al ., 2009 ), 其中种子的春季萌发及幼苗形成过程被认为是一种逃避冬天死亡的有利方式, 而秋天萌发是种子对低温的适应及竞争优势(
Gutterman, 1993 ).喀什霸王干藏6个月的种子在水分充足时才萌发, 说明该物种在塔克拉玛干沙漠西南部的极端环境的长期生存过程中, 随时面临着自然环境难以预测的生存风险, 通过种子的非深度生理休眠可获得更多的生存机会, 保持物种亲本性状. ...
Variation in dormancy and germination in three co-occurring perennial forest herbs
2
2011
... 与生活史特征相关的种子休眠及幼苗形成过程是种子植物类群后代产生及种群更新的关键阶段(
Baskin & Baskin, 2014 ;
Lu et al ., 2014 ;
孙清琳等, 2019 ).不同植物类群在自身生存及对自然环境的长期适应过程中, 表现出特殊的休眠和萌发特性来保证物种和种群的稳定性(
Nordborg & Bergelson, 1999 ;
Nurulla et al ., 2015 ;
Xia et al ., 2018 ).休眠程度和萌发时间往往受外部因素(光照、温度、水分)及种子自身因素(胚的成熟程度、种皮厚度、种子生活力、调控基因的表达及营养物质)的影响(
Venable et al ., 1987 ;
Casas et al ., 2012 ;
刘波等, 2015 ;
Soltani et al ., 2017 ).根据以上因素的组合程度不同, 种子休眠类型分为物理休眠(PY)、生理休眠(PD)、综合休眠(PY + PD)、形态休眠(MD)和形态生理休眠(MPD) 5种类型(
Galloway & Etterson, 2009 ;
Albrecht & McCarthy, 2011 ;
Baskin & Baskin, 2014 ;
Lu et al ., 2014 ;
Pluntz et al ., 2018 ), 其中生理休眠是自然界中普遍存在且较复杂的休眠类型(
Baskin & Baskin, 2014 ). ...
... 生理休眠的植物在极端环境生存及适应过程中表现出较高的生活周期多样性(
Gutterman, 1993 ;
Baskin & Baskin, 2014 ;
Nurulla et al ., 2015 ), 其中非深度生理休眠植物的幼苗存活力往往受到不同环境胁迫(干旱、盐、高温、紫外线)的影响(
Albrecht & McCarthy, 2011 ;
Baskin & Baskin, 2014 ).在干旱环境中普遍存在的干旱及盐胁迫能影响植物种子萌发物候及生活史特点(
Tobe et al ., 2001 ;
Ahmad et al ., 2009 ;
宋乃平等, 2018 ).国内有关研究人员在盐及干旱(缺水)胁迫对寒温带干旱、半干旱荒漠分布植物的生活史特征及其对极端环境的响应(
黄振英等, 2001 ;
Sun et al ., 2009 ;
Lu et al ., 2014 ,
2015 ;
Nurulla et al ., 2015 )方面开展了研究; 但对在我国暖温带干旱荒漠生长的濒危植物种子的生物学特性及其对当地干旱(缺水)环境的响应研究鲜有报道.为此, 本研究对该区域稀有植物喀什霸王(
Zygophyllum kaschgaricum )的结籽率和种子萌发特性进行了研究. ...
Germination ecophysiology of herbaceous plant species in a temperate region
1
1988
... 种子的不同季节萌发能促进植物物候的多样性(
Baskin & Baskin, 1988 ;
刘波等, 2015 ); 春秋季萌发是种子植物在不可预测环境的两头下注策略和种子生理后熟之间的权衡(
Lu et al ., 2014 ).在本研究中, 室内不同浓度PEG处理种子在低温条件下的萌发率比高温高(
图5 ).此外, 随着干藏时间的延长, 种子对水分的需求提高(
图1 ).塔克拉玛干沙漠西南部自然环境的平均月降水量低于10 mm, 该物种果实成熟的6-9月的平均气温在25 ℃以上, 气候特点与以色列的Negev荒漠(
Gutterman, 1993 ,
2000 )相似.因此, 该区域春季的降水量、气温及光照强度等气候特点能限制有非深度生理休眠种子的萌发及幼苗存活率.在降水时间分布不均匀的塔克拉玛干沙漠西南部荒漠环境中(
图1 ), 该物种种子的萌发对策主要是对降水做出响应(
Tobe et al ., 2001 ;
Ahmad et al ., 2009 ), 其中种子的春季萌发及幼苗形成过程被认为是一种逃避冬天死亡的有利方式, 而秋天萌发是种子对低温的适应及竞争优势(
Gutterman, 1993 ).喀什霸王干藏6个月的种子在水分充足时才萌发, 说明该物种在塔克拉玛干沙漠西南部的极端环境的长期生存过程中, 随时面临着自然环境难以预测的生存风险, 通过种子的非深度生理休眠可获得更多的生存机会, 保持物种亲本性状. ...
2014
A classification system for seed dormancy
3
2004
... 1.2.1.2 吸水特性 为了判断种子在休眠及非休眠状态时吸水特点的差异, 自2017年6月开始, 从刚采回来(0个月)及室温条件下干藏6个月的饱满种子中各选取50粒, 每个时间段的种子均分5个组(每个组10粒); 用电子天平测每组种子的质量, 然后将每组种子置于2层滤纸的培养皿(直径为9 cm)中, 加适量蒸馏水, 隔1 h称量一次(直到种子质量不再增加为止); 最后比较不同干藏时间种子的吸水速率.按照
Baskin和Baskin (2004) 的公式:
W r = ((
W f -
W i )/
W i ) × 100, 计算种子吸水速率(
W r ), 其中
W i 及
W f 是吸水前和吸水1 h后的质量. ...
... 为了判断种子生理后熟对种子萌发的影响, 将从自然环境采集, 后在室内温度条件下干藏0、3、6及9个月的种子放于直径9 cm的培养皿里(每个培养皿25粒种子, 重复3次), 然后置于5/2 ℃、15/5 ℃、20/10 ℃及30/20 ℃ 4个不同温度、光照/黑暗(培养皿用不透光的黑色袋子包裹)各12 h条件下的变温光照培养箱中培养28天.每天统计一次各培养皿中种子的萌发率, 并加入2 mL蒸馏水以保持湿度一致.本实验的4个温度代表该物种自然生长种群(格达良荒漠环境)不同月的日平均、最高和最低气温: 30/15 ℃ (5月、7月、8月)、20/10 ℃ (4月和10月)、15/5 ℃ (3月和11月)及5/2 ℃ (12月和2月).实验结束后参考
Baskin和Baskin (2004) 的TTC (0.1%的氯化三苯四氮唑溶液)处理方法, 对各组未萌发的种子活力进行检测.TTC处理后的种胚在20 ℃全黑暗条件下放置24 h, 有活力的种子将被染成红或粉红色. ...
... 在严酷的极端荒漠环境中, 植物的坐果率和结籽率、种子休眠程度及幼苗存活率能决定植物生活史特征及种群的更新水平.其中果实坐果率及结籽率与植物交配方式及传粉效率有关, 而休眠程度、种子萌发率及幼苗存活率与自然环境的温度和水分等外部因素有关(
黄振英等, 2001 ;
Baskin & Baskin, 2004 ;
Ahmad et al ., 2009 ;
Pluntz et al ., 2018 ;
Xia et al ., 2018 ).本研究中, 喀什霸王果实的自然坐果率及结籽率较低, 因为在其开花的4月中旬, 塔克拉玛干沙漠西南部荒漠环境的昼夜温度差异及风速较大, 早春活动的昆虫种类少; 此外, 该物种存在花内雌性先熟现象(未发表数据).以上因素导致的传粉限制可能是降低自然坐果率及结籽率(46.97 ± 2.56)%的主要因素, 此推测需要进一步验证. ...
Climate change and pollution speed declines in zebra fish populations
1
2015
... 生物多样性保护是有关生态系统安全和稳定性维护的关键科学问题(
Western, 2001 ;
Ren et al ., 2012 ;
覃海宁等, 2017 ).在全球变暖、自然灾害频繁发生、荒漠化严重及种群片段化等内外因素的影响下(
Parmesan & Yohe, 2003 ;
Leadley et al ., 2010 ), 生存在极端环境中的物种受到了不同程度的灭绝威胁(
Western, 2001 ;
张文辉等, 2004 ;
覃海宁等, 2017 ).我国植物多样性丰富, 目前有4 000-5 000种植物正处于不同程度的危机(稀有、濒危、极危)状态中(
Ren et al ., 2012 ).这些植物是生物多样性保护的重要组成部分, 常具有地理分布狭窄、种群规模小、个体数量少以及生境严酷且脆弱等特点(
张文辉等, 2004 ;
Brown et al ., 2015 ;
覃海宁等, 2017 ); 此外, 坐果率、结籽率、种子萌发率和幼苗存活力低等生活史特征也是导致其濒危或灭绝的主要原因(
Neaves et al ., 2015 ).因此, 对该类群的生活史及其对生存环境响应的研究, 在保护稀有濒危植物方面具有重要的科学及实际意义(
Western, 2001 ;
覃海宁等, 2017 ). ...
Seed after-ripening and dormancy determine adult life history independently of germination timing
1
2012
... 与生活史特征相关的种子休眠及幼苗形成过程是种子植物类群后代产生及种群更新的关键阶段(
Baskin & Baskin, 2014 ;
Lu et al ., 2014 ;
孙清琳等, 2019 ).不同植物类群在自身生存及对自然环境的长期适应过程中, 表现出特殊的休眠和萌发特性来保证物种和种群的稳定性(
Nordborg & Bergelson, 1999 ;
Nurulla et al ., 2015 ;
Xia et al ., 2018 ).休眠程度和萌发时间往往受外部因素(光照、温度、水分)及种子自身因素(胚的成熟程度、种皮厚度、种子生活力、调控基因的表达及营养物质)的影响(
Venable et al ., 1987 ;
Casas et al ., 2012 ;
刘波等, 2015 ;
Soltani et al ., 2017 ).根据以上因素的组合程度不同, 种子休眠类型分为物理休眠(PY)、生理休眠(PD)、综合休眠(PY + PD)、形态休眠(MD)和形态生理休眠(MPD) 5种类型(
Galloway & Etterson, 2009 ;
Albrecht & McCarthy, 2011 ;
Baskin & Baskin, 2014 ;
Lu et al ., 2014 ;
Pluntz et al ., 2018 ), 其中生理休眠是自然界中普遍存在且较复杂的休眠类型(
Baskin & Baskin, 2014 ). ...
Annual dormancy cycles in buried seeds of shrub species: Germination ecology of Sideritis serrata(Labiatae)
1
2015
... 种子萌发作为植物生活周期的主要阶段, 可影响植物雌性(种子)适合度和种群维持及扩大能力(
Baskin & Baskin, 2014 ;
Copete et al ., 2015 ).被子植物在种子物候上所表现出的异质性(
Kalisz & Wardle, 1994 ;
Nordborg & Bergelson, 1999 )作为一种可避免种群灭绝的、谨慎的萌发对策(
Gutterman, 2000 ), 有利于在荒漠极端环境中生长的植物种群的维持和更新(
姚红和谭敦炎, 2005 ;
Nurulla et al ., 2015 ).在本研究中, 非深度生理休眠的喀什霸王种子在春季和秋季均萌发, 虽然该物种在秋季存在一定程度的休眠.在自然环境中, 春季和秋季的温度较低而降水量较多, 因此, 该物种种子通过春秋季萌动的繁殖策略及非深度生理休眠来维持种群的动态平衡及保证种群的更新; 但4-6月气温的上升及降水量的减少导致的干旱胁迫可降低喀什霸王种子的萌发率或幼苗存活能力(
Tobe et al ., 2001 ), 说明新疆南部荒漠环境中降水量是影响非深度生理休眠植物喀什霸王种群更新能力的主要因素(
Lu et al ., 2014 ;
Nurulla et al ., 2015 ).总之, 种子物候上表现出的春秋季萌动繁殖策略及非深度生理休眠是该物种在干旱胁迫及高温环境中为保持种群稳定性而演化出的一种适应策略.在今后的研究中, 喀什霸王幼苗存活力及种群更新与环境因子的关系等方面还需要进一步的研究. ...
Plasticity to canopy shade in a monocarpic herb: Within- and between-generation effects
1
2009
... 与生活史特征相关的种子休眠及幼苗形成过程是种子植物类群后代产生及种群更新的关键阶段(
Baskin & Baskin, 2014 ;
Lu et al ., 2014 ;
孙清琳等, 2019 ).不同植物类群在自身生存及对自然环境的长期适应过程中, 表现出特殊的休眠和萌发特性来保证物种和种群的稳定性(
Nordborg & Bergelson, 1999 ;
Nurulla et al ., 2015 ;
Xia et al ., 2018 ).休眠程度和萌发时间往往受外部因素(光照、温度、水分)及种子自身因素(胚的成熟程度、种皮厚度、种子生活力、调控基因的表达及营养物质)的影响(
Venable et al ., 1987 ;
Casas et al ., 2012 ;
刘波等, 2015 ;
Soltani et al ., 2017 ).根据以上因素的组合程度不同, 种子休眠类型分为物理休眠(PY)、生理休眠(PD)、综合休眠(PY + PD)、形态休眠(MD)和形态生理休眠(MPD) 5种类型(
Galloway & Etterson, 2009 ;
Albrecht & McCarthy, 2011 ;
Baskin & Baskin, 2014 ;
Lu et al ., 2014 ;
Pluntz et al ., 2018 ), 其中生理休眠是自然界中普遍存在且较复杂的休眠类型(
Baskin & Baskin, 2014 ). ...
Seed Germination in Desert Plants 3
1993
... 生理休眠的植物在极端环境生存及适应过程中表现出较高的生活周期多样性(
Gutterman, 1993 ;
Baskin & Baskin, 2014 ;
Nurulla et al ., 2015 ), 其中非深度生理休眠植物的幼苗存活力往往受到不同环境胁迫(干旱、盐、高温、紫外线)的影响(
Albrecht & McCarthy, 2011 ;
Baskin & Baskin, 2014 ).在干旱环境中普遍存在的干旱及盐胁迫能影响植物种子萌发物候及生活史特点(
Tobe et al ., 2001 ;
Ahmad et al ., 2009 ;
宋乃平等, 2018 ).国内有关研究人员在盐及干旱(缺水)胁迫对寒温带干旱、半干旱荒漠分布植物的生活史特征及其对极端环境的响应(
黄振英等, 2001 ;
Sun et al ., 2009 ;
Lu et al ., 2014 ,
2015 ;
Nurulla et al ., 2015 )方面开展了研究; 但对在我国暖温带干旱荒漠生长的濒危植物种子的生物学特性及其对当地干旱(缺水)环境的响应研究鲜有报道.为此, 本研究对该区域稀有植物喀什霸王(
Zygophyllum kaschgaricum )的结籽率和种子萌发特性进行了研究. ...
... 种子的不同季节萌发能促进植物物候的多样性(
Baskin & Baskin, 1988 ;
刘波等, 2015 ); 春秋季萌发是种子植物在不可预测环境的两头下注策略和种子生理后熟之间的权衡(
Lu et al ., 2014 ).在本研究中, 室内不同浓度PEG处理种子在低温条件下的萌发率比高温高(
图5 ).此外, 随着干藏时间的延长, 种子对水分的需求提高(
图1 ).塔克拉玛干沙漠西南部自然环境的平均月降水量低于10 mm, 该物种果实成熟的6-9月的平均气温在25 ℃以上, 气候特点与以色列的Negev荒漠(
Gutterman, 1993 ,
2000 )相似.因此, 该区域春季的降水量、气温及光照强度等气候特点能限制有非深度生理休眠种子的萌发及幼苗存活率.在降水时间分布不均匀的塔克拉玛干沙漠西南部荒漠环境中(
图1 ), 该物种种子的萌发对策主要是对降水做出响应(
Tobe et al ., 2001 ;
Ahmad et al ., 2009 ), 其中种子的春季萌发及幼苗形成过程被认为是一种逃避冬天死亡的有利方式, 而秋天萌发是种子对低温的适应及竞争优势(
Gutterman, 1993 ).喀什霸王干藏6个月的种子在水分充足时才萌发, 说明该物种在塔克拉玛干沙漠西南部的极端环境的长期生存过程中, 随时面临着自然环境难以预测的生存风险, 通过种子的非深度生理休眠可获得更多的生存机会, 保持物种亲本性状. ...
... ), 其中种子的春季萌发及幼苗形成过程被认为是一种逃避冬天死亡的有利方式, 而秋天萌发是种子对低温的适应及竞争优势(
Gutterman, 1993 ).喀什霸王干藏6个月的种子在水分充足时才萌发, 说明该物种在塔克拉玛干沙漠西南部的极端环境的长期生存过程中, 随时面临着自然环境难以预测的生存风险, 通过种子的非深度生理休眠可获得更多的生存机会, 保持物种亲本性状. ...
Environmental factors and survival strategies of annual plant species in the Negev Desert, Israel
2
2000
... 种子的不同季节萌发能促进植物物候的多样性(
Baskin & Baskin, 1988 ;
刘波等, 2015 ); 春秋季萌发是种子植物在不可预测环境的两头下注策略和种子生理后熟之间的权衡(
Lu et al ., 2014 ).在本研究中, 室内不同浓度PEG处理种子在低温条件下的萌发率比高温高(
图5 ).此外, 随着干藏时间的延长, 种子对水分的需求提高(
图1 ).塔克拉玛干沙漠西南部自然环境的平均月降水量低于10 mm, 该物种果实成熟的6-9月的平均气温在25 ℃以上, 气候特点与以色列的Negev荒漠(
Gutterman, 1993 ,
2000 )相似.因此, 该区域春季的降水量、气温及光照强度等气候特点能限制有非深度生理休眠种子的萌发及幼苗存活率.在降水时间分布不均匀的塔克拉玛干沙漠西南部荒漠环境中(
图1 ), 该物种种子的萌发对策主要是对降水做出响应(
Tobe et al ., 2001 ;
Ahmad et al ., 2009 ), 其中种子的春季萌发及幼苗形成过程被认为是一种逃避冬天死亡的有利方式, 而秋天萌发是种子对低温的适应及竞争优势(
Gutterman, 1993 ).喀什霸王干藏6个月的种子在水分充足时才萌发, 说明该物种在塔克拉玛干沙漠西南部的极端环境的长期生存过程中, 随时面临着自然环境难以预测的生存风险, 通过种子的非深度生理休眠可获得更多的生存机会, 保持物种亲本性状. ...
... 种子萌发作为植物生活周期的主要阶段, 可影响植物雌性(种子)适合度和种群维持及扩大能力(
Baskin & Baskin, 2014 ;
Copete et al ., 2015 ).被子植物在种子物候上所表现出的异质性(
Kalisz & Wardle, 1994 ;
Nordborg & Bergelson, 1999 )作为一种可避免种群灭绝的、谨慎的萌发对策(
Gutterman, 2000 ), 有利于在荒漠极端环境中生长的植物种群的维持和更新(
姚红和谭敦炎, 2005 ;
Nurulla et al ., 2015 ).在本研究中, 非深度生理休眠的喀什霸王种子在春季和秋季均萌发, 虽然该物种在秋季存在一定程度的休眠.在自然环境中, 春季和秋季的温度较低而降水量较多, 因此, 该物种种子通过春秋季萌动的繁殖策略及非深度生理休眠来维持种群的动态平衡及保证种群的更新; 但4-6月气温的上升及降水量的减少导致的干旱胁迫可降低喀什霸王种子的萌发率或幼苗存活能力(
Tobe et al ., 2001 ), 说明新疆南部荒漠环境中降水量是影响非深度生理休眠植物喀什霸王种群更新能力的主要因素(
Lu et al ., 2014 ;
Nurulla et al ., 2015 ).总之, 种子物候上表现出的春秋季萌动繁殖策略及非深度生理休眠是该物种在干旱胁迫及高温环境中为保持种群稳定性而演化出的一种适应策略.在今后的研究中, 喀什霸王幼苗存活力及种群更新与环境因子的关系等方面还需要进一步的研究. ...
白沙蒿种子萌发特性的研究II. 环境因素的影响
2
2001
... 生理休眠的植物在极端环境生存及适应过程中表现出较高的生活周期多样性(
Gutterman, 1993 ;
Baskin & Baskin, 2014 ;
Nurulla et al ., 2015 ), 其中非深度生理休眠植物的幼苗存活力往往受到不同环境胁迫(干旱、盐、高温、紫外线)的影响(
Albrecht & McCarthy, 2011 ;
Baskin & Baskin, 2014 ).在干旱环境中普遍存在的干旱及盐胁迫能影响植物种子萌发物候及生活史特点(
Tobe et al ., 2001 ;
Ahmad et al ., 2009 ;
宋乃平等, 2018 ).国内有关研究人员在盐及干旱(缺水)胁迫对寒温带干旱、半干旱荒漠分布植物的生活史特征及其对极端环境的响应(
黄振英等, 2001 ;
Sun et al ., 2009 ;
Lu et al ., 2014 ,
2015 ;
Nurulla et al ., 2015 )方面开展了研究; 但对在我国暖温带干旱荒漠生长的濒危植物种子的生物学特性及其对当地干旱(缺水)环境的响应研究鲜有报道.为此, 本研究对该区域稀有植物喀什霸王(
Zygophyllum kaschgaricum )的结籽率和种子萌发特性进行了研究. ...
... 在严酷的极端荒漠环境中, 植物的坐果率和结籽率、种子休眠程度及幼苗存活率能决定植物生活史特征及种群的更新水平.其中果实坐果率及结籽率与植物交配方式及传粉效率有关, 而休眠程度、种子萌发率及幼苗存活率与自然环境的温度和水分等外部因素有关(
黄振英等, 2001 ;
Baskin & Baskin, 2004 ;
Ahmad et al ., 2009 ;
Pluntz et al ., 2018 ;
Xia et al ., 2018 ).本研究中, 喀什霸王果实的自然坐果率及结籽率较低, 因为在其开花的4月中旬, 塔克拉玛干沙漠西南部荒漠环境的昼夜温度差异及风速较大, 早春活动的昆虫种类少; 此外, 该物种存在花内雌性先熟现象(未发表数据).以上因素导致的传粉限制可能是降低自然坐果率及结籽率(46.97 ± 2.56)%的主要因素, 此推测需要进一步验证. ...
白沙蒿种子萌发特性的研究II. 环境因素的影响
2
2001
... 生理休眠的植物在极端环境生存及适应过程中表现出较高的生活周期多样性(
Gutterman, 1993 ;
Baskin & Baskin, 2014 ;
Nurulla et al ., 2015 ), 其中非深度生理休眠植物的幼苗存活力往往受到不同环境胁迫(干旱、盐、高温、紫外线)的影响(
Albrecht & McCarthy, 2011 ;
Baskin & Baskin, 2014 ).在干旱环境中普遍存在的干旱及盐胁迫能影响植物种子萌发物候及生活史特点(
Tobe et al ., 2001 ;
Ahmad et al ., 2009 ;
宋乃平等, 2018 ).国内有关研究人员在盐及干旱(缺水)胁迫对寒温带干旱、半干旱荒漠分布植物的生活史特征及其对极端环境的响应(
黄振英等, 2001 ;
Sun et al ., 2009 ;
Lu et al ., 2014 ,
2015 ;
Nurulla et al ., 2015 )方面开展了研究; 但对在我国暖温带干旱荒漠生长的濒危植物种子的生物学特性及其对当地干旱(缺水)环境的响应研究鲜有报道.为此, 本研究对该区域稀有植物喀什霸王(
Zygophyllum kaschgaricum )的结籽率和种子萌发特性进行了研究. ...
... 在严酷的极端荒漠环境中, 植物的坐果率和结籽率、种子休眠程度及幼苗存活率能决定植物生活史特征及种群的更新水平.其中果实坐果率及结籽率与植物交配方式及传粉效率有关, 而休眠程度、种子萌发率及幼苗存活率与自然环境的温度和水分等外部因素有关(
黄振英等, 2001 ;
Baskin & Baskin, 2004 ;
Ahmad et al ., 2009 ;
Pluntz et al ., 2018 ;
Xia et al ., 2018 ).本研究中, 喀什霸王果实的自然坐果率及结籽率较低, 因为在其开花的4月中旬, 塔克拉玛干沙漠西南部荒漠环境的昼夜温度差异及风速较大, 早春活动的昆虫种类少; 此外, 该物种存在花内雌性先熟现象(未发表数据).以上因素导致的传粉限制可能是降低自然坐果率及结籽率(46.97 ± 2.56)%的主要因素, 此推测需要进一步验证. ...
Life history variation in Campanula americana(Campanulaceae): Population differentiation
1
1994
... 种子萌发作为植物生活周期的主要阶段, 可影响植物雌性(种子)适合度和种群维持及扩大能力(
Baskin & Baskin, 2014 ;
Copete et al ., 2015 ).被子植物在种子物候上所表现出的异质性(
Kalisz & Wardle, 1994 ;
Nordborg & Bergelson, 1999 )作为一种可避免种群灭绝的、谨慎的萌发对策(
Gutterman, 2000 ), 有利于在荒漠极端环境中生长的植物种群的维持和更新(
姚红和谭敦炎, 2005 ;
Nurulla et al ., 2015 ).在本研究中, 非深度生理休眠的喀什霸王种子在春季和秋季均萌发, 虽然该物种在秋季存在一定程度的休眠.在自然环境中, 春季和秋季的温度较低而降水量较多, 因此, 该物种种子通过春秋季萌动的繁殖策略及非深度生理休眠来维持种群的动态平衡及保证种群的更新; 但4-6月气温的上升及降水量的减少导致的干旱胁迫可降低喀什霸王种子的萌发率或幼苗存活能力(
Tobe et al ., 2001 ), 说明新疆南部荒漠环境中降水量是影响非深度生理休眠植物喀什霸王种群更新能力的主要因素(
Lu et al ., 2014 ;
Nurulla et al ., 2015 ).总之, 种子物候上表现出的春秋季萌动繁殖策略及非深度生理休眠是该物种在干旱胁迫及高温环境中为保持种群稳定性而演化出的一种适应策略.在今后的研究中, 喀什霸王幼苗存活力及种群更新与环境因子的关系等方面还需要进一步的研究. ...
Biodiversity Scenarios: Projections of 21st Century Change in Biodiversity and Associated Ecosystem Services 1
2010
... 生物多样性保护是有关生态系统安全和稳定性维护的关键科学问题(
Western, 2001 ;
Ren et al ., 2012 ;
覃海宁等, 2017 ).在全球变暖、自然灾害频繁发生、荒漠化严重及种群片段化等内外因素的影响下(
Parmesan & Yohe, 2003 ;
Leadley et al ., 2010 ), 生存在极端环境中的物种受到了不同程度的灭绝威胁(
Western, 2001 ;
张文辉等, 2004 ;
覃海宁等, 2017 ).我国植物多样性丰富, 目前有4 000-5 000种植物正处于不同程度的危机(稀有、濒危、极危)状态中(
Ren et al ., 2012 ).这些植物是生物多样性保护的重要组成部分, 常具有地理分布狭窄、种群规模小、个体数量少以及生境严酷且脆弱等特点(
张文辉等, 2004 ;
Brown et al ., 2015 ;
覃海宁等, 2017 ); 此外, 坐果率、结籽率、种子萌发率和幼苗存活力低等生活史特征也是导致其濒危或灭绝的主要原因(
Neaves et al ., 2015 ).因此, 对该类群的生活史及其对生存环境响应的研究, 在保护稀有濒危植物方面具有重要的科学及实际意义(
Western, 2001 ;
覃海宁等, 2017 ). ...
光照、水深交互作用对松嫩湿地芦苇种子萌发的影响
3
2015
... 与生活史特征相关的种子休眠及幼苗形成过程是种子植物类群后代产生及种群更新的关键阶段(
Baskin & Baskin, 2014 ;
Lu et al ., 2014 ;
孙清琳等, 2019 ).不同植物类群在自身生存及对自然环境的长期适应过程中, 表现出特殊的休眠和萌发特性来保证物种和种群的稳定性(
Nordborg & Bergelson, 1999 ;
Nurulla et al ., 2015 ;
Xia et al ., 2018 ).休眠程度和萌发时间往往受外部因素(光照、温度、水分)及种子自身因素(胚的成熟程度、种皮厚度、种子生活力、调控基因的表达及营养物质)的影响(
Venable et al ., 1987 ;
Casas et al ., 2012 ;
刘波等, 2015 ;
Soltani et al ., 2017 ).根据以上因素的组合程度不同, 种子休眠类型分为物理休眠(PY)、生理休眠(PD)、综合休眠(PY + PD)、形态休眠(MD)和形态生理休眠(MPD) 5种类型(
Galloway & Etterson, 2009 ;
Albrecht & McCarthy, 2011 ;
Baskin & Baskin, 2014 ;
Lu et al ., 2014 ;
Pluntz et al ., 2018 ), 其中生理休眠是自然界中普遍存在且较复杂的休眠类型(
Baskin & Baskin, 2014 ). ...
... 生理休眠植物中有非深度生理休眠的新鲜种子在任何温度及光照下均不能萌发或只能在很窄的温度或光照范围内萌发(
Sun et al ., 2009 ;
Baskin & Baskin, 2014 ;
刘波等, 2015 ;
Soltani et al ., 2017 ).喀什霸王刚采回来的种子在不同温度、光照或黑暗条件下均能萌发, 且彼此间存在显著差异(
p < 0.01,
图3 ); 其中高温黑暗(20/10 ℃及30/20 ℃)处理种子的萌发率高于低温(5/2 ℃和15/2 ℃)黑暗处理种子的萌发率.随着贮藏时间的延长, 贮藏种子的萌发率逐渐提高, 而休眠逐渐解除, 各温变周期下的种子萌发率逐渐提高(
图3 ); 但干藏9个月的种子在低温条件下萌发率低于60%.虽然该物种种子在各温度和不同光周期下均萌发, 但高温和黑暗是最佳的萌发条件.此外, 刚成熟及干藏6个月的种子在不同浓度的GA
3 处理后, 同一浓度GA
3 在各温变周期(光照/黑暗)下的不同干藏处理种子的萌发率间未存在显著差异(
p > 0.05); 但不同浓度GA
3 处理种子的萌发率间存在显著差异(
p < 0.05); 其中50 mmoloL
-1 浓度的萌发率最高, 是打破种子休眠的最合适浓度(
图3 ).从以上结果可以判断喀什霸王种子的休眠类型属于非深度生理休眠类型, 这与我国北部寒温带分布的石生霸王(
Zygophyllum rosovii )和石炭酸灌木(
Larrea divaricate )的生理休眠表现相似(
Venable et al ., 1987 ;
Mandák, 2001 ;
Baskin & Baskin, 2014 ).喀什霸王种子的非深度生理休眠特征可能与该物种长期适应南疆秋天和春天较短的光日照时间及高温过程中进化出的种子萌发对策有关. ...
... 种子的不同季节萌发能促进植物物候的多样性(
Baskin & Baskin, 1988 ;
刘波等, 2015 ); 春秋季萌发是种子植物在不可预测环境的两头下注策略和种子生理后熟之间的权衡(
Lu et al ., 2014 ).在本研究中, 室内不同浓度PEG处理种子在低温条件下的萌发率比高温高(
图5 ).此外, 随着干藏时间的延长, 种子对水分的需求提高(
图1 ).塔克拉玛干沙漠西南部自然环境的平均月降水量低于10 mm, 该物种果实成熟的6-9月的平均气温在25 ℃以上, 气候特点与以色列的Negev荒漠(
Gutterman, 1993 ,
2000 )相似.因此, 该区域春季的降水量、气温及光照强度等气候特点能限制有非深度生理休眠种子的萌发及幼苗存活率.在降水时间分布不均匀的塔克拉玛干沙漠西南部荒漠环境中(
图1 ), 该物种种子的萌发对策主要是对降水做出响应(
Tobe et al ., 2001 ;
Ahmad et al ., 2009 ), 其中种子的春季萌发及幼苗形成过程被认为是一种逃避冬天死亡的有利方式, 而秋天萌发是种子对低温的适应及竞争优势(
Gutterman, 1993 ).喀什霸王干藏6个月的种子在水分充足时才萌发, 说明该物种在塔克拉玛干沙漠西南部的极端环境的长期生存过程中, 随时面临着自然环境难以预测的生存风险, 通过种子的非深度生理休眠可获得更多的生存机会, 保持物种亲本性状. ...
光照、水深交互作用对松嫩湿地芦苇种子萌发的影响
3
2015
... 与生活史特征相关的种子休眠及幼苗形成过程是种子植物类群后代产生及种群更新的关键阶段(
Baskin & Baskin, 2014 ;
Lu et al ., 2014 ;
孙清琳等, 2019 ).不同植物类群在自身生存及对自然环境的长期适应过程中, 表现出特殊的休眠和萌发特性来保证物种和种群的稳定性(
Nordborg & Bergelson, 1999 ;
Nurulla et al ., 2015 ;
Xia et al ., 2018 ).休眠程度和萌发时间往往受外部因素(光照、温度、水分)及种子自身因素(胚的成熟程度、种皮厚度、种子生活力、调控基因的表达及营养物质)的影响(
Venable et al ., 1987 ;
Casas et al ., 2012 ;
刘波等, 2015 ;
Soltani et al ., 2017 ).根据以上因素的组合程度不同, 种子休眠类型分为物理休眠(PY)、生理休眠(PD)、综合休眠(PY + PD)、形态休眠(MD)和形态生理休眠(MPD) 5种类型(
Galloway & Etterson, 2009 ;
Albrecht & McCarthy, 2011 ;
Baskin & Baskin, 2014 ;
Lu et al ., 2014 ;
Pluntz et al ., 2018 ), 其中生理休眠是自然界中普遍存在且较复杂的休眠类型(
Baskin & Baskin, 2014 ). ...
... 生理休眠植物中有非深度生理休眠的新鲜种子在任何温度及光照下均不能萌发或只能在很窄的温度或光照范围内萌发(
Sun et al ., 2009 ;
Baskin & Baskin, 2014 ;
刘波等, 2015 ;
Soltani et al ., 2017 ).喀什霸王刚采回来的种子在不同温度、光照或黑暗条件下均能萌发, 且彼此间存在显著差异(
p < 0.01,
图3 ); 其中高温黑暗(20/10 ℃及30/20 ℃)处理种子的萌发率高于低温(5/2 ℃和15/2 ℃)黑暗处理种子的萌发率.随着贮藏时间的延长, 贮藏种子的萌发率逐渐提高, 而休眠逐渐解除, 各温变周期下的种子萌发率逐渐提高(
图3 ); 但干藏9个月的种子在低温条件下萌发率低于60%.虽然该物种种子在各温度和不同光周期下均萌发, 但高温和黑暗是最佳的萌发条件.此外, 刚成熟及干藏6个月的种子在不同浓度的GA
3 处理后, 同一浓度GA
3 在各温变周期(光照/黑暗)下的不同干藏处理种子的萌发率间未存在显著差异(
p > 0.05); 但不同浓度GA
3 处理种子的萌发率间存在显著差异(
p < 0.05); 其中50 mmoloL
-1 浓度的萌发率最高, 是打破种子休眠的最合适浓度(
图3 ).从以上结果可以判断喀什霸王种子的休眠类型属于非深度生理休眠类型, 这与我国北部寒温带分布的石生霸王(
Zygophyllum rosovii )和石炭酸灌木(
Larrea divaricate )的生理休眠表现相似(
Venable et al ., 1987 ;
Mandák, 2001 ;
Baskin & Baskin, 2014 ).喀什霸王种子的非深度生理休眠特征可能与该物种长期适应南疆秋天和春天较短的光日照时间及高温过程中进化出的种子萌发对策有关. ...
... 种子的不同季节萌发能促进植物物候的多样性(
Baskin & Baskin, 1988 ;
刘波等, 2015 ); 春秋季萌发是种子植物在不可预测环境的两头下注策略和种子生理后熟之间的权衡(
Lu et al ., 2014 ).在本研究中, 室内不同浓度PEG处理种子在低温条件下的萌发率比高温高(
图5 ).此外, 随着干藏时间的延长, 种子对水分的需求提高(
图1 ).塔克拉玛干沙漠西南部自然环境的平均月降水量低于10 mm, 该物种果实成熟的6-9月的平均气温在25 ℃以上, 气候特点与以色列的Negev荒漠(
Gutterman, 1993 ,
2000 )相似.因此, 该区域春季的降水量、气温及光照强度等气候特点能限制有非深度生理休眠种子的萌发及幼苗存活率.在降水时间分布不均匀的塔克拉玛干沙漠西南部荒漠环境中(
图1 ), 该物种种子的萌发对策主要是对降水做出响应(
Tobe et al ., 2001 ;
Ahmad et al ., 2009 ), 其中种子的春季萌发及幼苗形成过程被认为是一种逃避冬天死亡的有利方式, 而秋天萌发是种子对低温的适应及竞争优势(
Gutterman, 1993 ).喀什霸王干藏6个月的种子在水分充足时才萌发, 说明该物种在塔克拉玛干沙漠西南部的极端环境的长期生存过程中, 随时面临着自然环境难以预测的生存风险, 通过种子的非深度生理休眠可获得更多的生存机会, 保持物种亲本性状. ...
Germination season and watering regime, but not seed morph, affect life history traits in a cold desert diaspore-heteromorphic annual
13
2014
... 与生活史特征相关的种子休眠及幼苗形成过程是种子植物类群后代产生及种群更新的关键阶段(
Baskin & Baskin, 2014 ;
Lu et al ., 2014 ;
孙清琳等, 2019 ).不同植物类群在自身生存及对自然环境的长期适应过程中, 表现出特殊的休眠和萌发特性来保证物种和种群的稳定性(
Nordborg & Bergelson, 1999 ;
Nurulla et al ., 2015 ;
Xia et al ., 2018 ).休眠程度和萌发时间往往受外部因素(光照、温度、水分)及种子自身因素(胚的成熟程度、种皮厚度、种子生活力、调控基因的表达及营养物质)的影响(
Venable et al ., 1987 ;
Casas et al ., 2012 ;
刘波等, 2015 ;
Soltani et al ., 2017 ).根据以上因素的组合程度不同, 种子休眠类型分为物理休眠(PY)、生理休眠(PD)、综合休眠(PY + PD)、形态休眠(MD)和形态生理休眠(MPD) 5种类型(
Galloway & Etterson, 2009 ;
Albrecht & McCarthy, 2011 ;
Baskin & Baskin, 2014 ;
Lu et al ., 2014 ;
Pluntz et al ., 2018 ), 其中生理休眠是自然界中普遍存在且较复杂的休眠类型(
Baskin & Baskin, 2014 ). ...
... ;
Lu et al ., 2014 ;
孙清琳等, 2019 ).不同植物类群在自身生存及对自然环境的长期适应过程中, 表现出特殊的休眠和萌发特性来保证物种和种群的稳定性(
Nordborg & Bergelson, 1999 ;
Nurulla et al ., 2015 ;
Xia et al ., 2018 ).休眠程度和萌发时间往往受外部因素(光照、温度、水分)及种子自身因素(胚的成熟程度、种皮厚度、种子生活力、调控基因的表达及营养物质)的影响(
Venable et al ., 1987 ;
Casas et al ., 2012 ;
刘波等, 2015 ;
Soltani et al ., 2017 ).根据以上因素的组合程度不同, 种子休眠类型分为物理休眠(PY)、生理休眠(PD)、综合休眠(PY + PD)、形态休眠(MD)和形态生理休眠(MPD) 5种类型(
Galloway & Etterson, 2009 ;
Albrecht & McCarthy, 2011 ;
Baskin & Baskin, 2014 ;
Lu et al ., 2014 ;
Pluntz et al ., 2018 ), 其中生理休眠是自然界中普遍存在且较复杂的休眠类型(
Baskin & Baskin, 2014 ). ...
... ;
Baskin & Baskin, 2014 ;
Lu et al ., 2014 ;
Pluntz et al ., 2018 ), 其中生理休眠是自然界中普遍存在且较复杂的休眠类型(
Baskin & Baskin, 2014 ). ...
... ;
Lu et al ., 2014 ;
Pluntz et al ., 2018 ), 其中生理休眠是自然界中普遍存在且较复杂的休眠类型(
Baskin & Baskin, 2014 ). ...
... ), 其中生理休眠是自然界中普遍存在且较复杂的休眠类型(
Baskin & Baskin, 2014 ). ...
... 生理休眠的植物在极端环境生存及适应过程中表现出较高的生活周期多样性(
Gutterman, 1993 ;
Baskin & Baskin, 2014 ;
Nurulla et al ., 2015 ), 其中非深度生理休眠植物的幼苗存活力往往受到不同环境胁迫(干旱、盐、高温、紫外线)的影响(
Albrecht & McCarthy, 2011 ;
Baskin & Baskin, 2014 ).在干旱环境中普遍存在的干旱及盐胁迫能影响植物种子萌发物候及生活史特点(
Tobe et al ., 2001 ;
Ahmad et al ., 2009 ;
宋乃平等, 2018 ).国内有关研究人员在盐及干旱(缺水)胁迫对寒温带干旱、半干旱荒漠分布植物的生活史特征及其对极端环境的响应(
黄振英等, 2001 ;
Sun et al ., 2009 ;
Lu et al ., 2014 ,
2015 ;
Nurulla et al ., 2015 )方面开展了研究; 但对在我国暖温带干旱荒漠生长的濒危植物种子的生物学特性及其对当地干旱(缺水)环境的响应研究鲜有报道.为此, 本研究对该区域稀有植物喀什霸王(
Zygophyllum kaschgaricum )的结籽率和种子萌发特性进行了研究. ...
... ;
Baskin & Baskin, 2014 ).在干旱环境中普遍存在的干旱及盐胁迫能影响植物种子萌发物候及生活史特点(
Tobe et al ., 2001 ;
Ahmad et al ., 2009 ;
宋乃平等, 2018 ).国内有关研究人员在盐及干旱(缺水)胁迫对寒温带干旱、半干旱荒漠分布植物的生活史特征及其对极端环境的响应(
黄振英等, 2001 ;
Sun et al ., 2009 ;
Lu et al ., 2014 ,
2015 ;
Nurulla et al ., 2015 )方面开展了研究; 但对在我国暖温带干旱荒漠生长的濒危植物种子的生物学特性及其对当地干旱(缺水)环境的响应研究鲜有报道.为此, 本研究对该区域稀有植物喀什霸王(
Zygophyllum kaschgaricum )的结籽率和种子萌发特性进行了研究. ...
... ;
Lu et al ., 2014 ,
2015 ;
Nurulla et al ., 2015 )方面开展了研究; 但对在我国暖温带干旱荒漠生长的濒危植物种子的生物学特性及其对当地干旱(缺水)环境的响应研究鲜有报道.为此, 本研究对该区域稀有植物喀什霸王(
Zygophyllum kaschgaricum )的结籽率和种子萌发特性进行了研究. ...
... 生理休眠植物中有非深度生理休眠的新鲜种子在任何温度及光照下均不能萌发或只能在很窄的温度或光照范围内萌发(
Sun et al ., 2009 ;
Baskin & Baskin, 2014 ;
刘波等, 2015 ;
Soltani et al ., 2017 ).喀什霸王刚采回来的种子在不同温度、光照或黑暗条件下均能萌发, 且彼此间存在显著差异(
p < 0.01,
图3 ); 其中高温黑暗(20/10 ℃及30/20 ℃)处理种子的萌发率高于低温(5/2 ℃和15/2 ℃)黑暗处理种子的萌发率.随着贮藏时间的延长, 贮藏种子的萌发率逐渐提高, 而休眠逐渐解除, 各温变周期下的种子萌发率逐渐提高(
图3 ); 但干藏9个月的种子在低温条件下萌发率低于60%.虽然该物种种子在各温度和不同光周期下均萌发, 但高温和黑暗是最佳的萌发条件.此外, 刚成熟及干藏6个月的种子在不同浓度的GA
3 处理后, 同一浓度GA
3 在各温变周期(光照/黑暗)下的不同干藏处理种子的萌发率间未存在显著差异(
p > 0.05); 但不同浓度GA
3 处理种子的萌发率间存在显著差异(
p < 0.05); 其中50 mmoloL
-1 浓度的萌发率最高, 是打破种子休眠的最合适浓度(
图3 ).从以上结果可以判断喀什霸王种子的休眠类型属于非深度生理休眠类型, 这与我国北部寒温带分布的石生霸王(
Zygophyllum rosovii )和石炭酸灌木(
Larrea divaricate )的生理休眠表现相似(
Venable et al ., 1987 ;
Mandák, 2001 ;
Baskin & Baskin, 2014 ).喀什霸王种子的非深度生理休眠特征可能与该物种长期适应南疆秋天和春天较短的光日照时间及高温过程中进化出的种子萌发对策有关. ...
... ;
Baskin & Baskin, 2014 ).喀什霸王种子的非深度生理休眠特征可能与该物种长期适应南疆秋天和春天较短的光日照时间及高温过程中进化出的种子萌发对策有关. ...
... 种子的不同季节萌发能促进植物物候的多样性(
Baskin & Baskin, 1988 ;
刘波等, 2015 ); 春秋季萌发是种子植物在不可预测环境的两头下注策略和种子生理后熟之间的权衡(
Lu et al ., 2014 ).在本研究中, 室内不同浓度PEG处理种子在低温条件下的萌发率比高温高(
图5 ).此外, 随着干藏时间的延长, 种子对水分的需求提高(
图1 ).塔克拉玛干沙漠西南部自然环境的平均月降水量低于10 mm, 该物种果实成熟的6-9月的平均气温在25 ℃以上, 气候特点与以色列的Negev荒漠(
Gutterman, 1993 ,
2000 )相似.因此, 该区域春季的降水量、气温及光照强度等气候特点能限制有非深度生理休眠种子的萌发及幼苗存活率.在降水时间分布不均匀的塔克拉玛干沙漠西南部荒漠环境中(
图1 ), 该物种种子的萌发对策主要是对降水做出响应(
Tobe et al ., 2001 ;
Ahmad et al ., 2009 ), 其中种子的春季萌发及幼苗形成过程被认为是一种逃避冬天死亡的有利方式, 而秋天萌发是种子对低温的适应及竞争优势(
Gutterman, 1993 ).喀什霸王干藏6个月的种子在水分充足时才萌发, 说明该物种在塔克拉玛干沙漠西南部的极端环境的长期生存过程中, 随时面临着自然环境难以预测的生存风险, 通过种子的非深度生理休眠可获得更多的生存机会, 保持物种亲本性状. ...
... 种子萌发作为植物生活周期的主要阶段, 可影响植物雌性(种子)适合度和种群维持及扩大能力(
Baskin & Baskin, 2014 ;
Copete et al ., 2015 ).被子植物在种子物候上所表现出的异质性(
Kalisz & Wardle, 1994 ;
Nordborg & Bergelson, 1999 )作为一种可避免种群灭绝的、谨慎的萌发对策(
Gutterman, 2000 ), 有利于在荒漠极端环境中生长的植物种群的维持和更新(
姚红和谭敦炎, 2005 ;
Nurulla et al ., 2015 ).在本研究中, 非深度生理休眠的喀什霸王种子在春季和秋季均萌发, 虽然该物种在秋季存在一定程度的休眠.在自然环境中, 春季和秋季的温度较低而降水量较多, 因此, 该物种种子通过春秋季萌动的繁殖策略及非深度生理休眠来维持种群的动态平衡及保证种群的更新; 但4-6月气温的上升及降水量的减少导致的干旱胁迫可降低喀什霸王种子的萌发率或幼苗存活能力(
Tobe et al ., 2001 ), 说明新疆南部荒漠环境中降水量是影响非深度生理休眠植物喀什霸王种群更新能力的主要因素(
Lu et al ., 2014 ;
Nurulla et al ., 2015 ).总之, 种子物候上表现出的春秋季萌动繁殖策略及非深度生理休眠是该物种在干旱胁迫及高温环境中为保持种群稳定性而演化出的一种适应策略.在今后的研究中, 喀什霸王幼苗存活力及种群更新与环境因子的关系等方面还需要进一步的研究. ...
... ), 说明新疆南部荒漠环境中降水量是影响非深度生理休眠植物喀什霸王种群更新能力的主要因素(
Lu et al ., 2014 ;
Nurulla et al ., 2015 ).总之, 种子物候上表现出的春秋季萌动繁殖策略及非深度生理休眠是该物种在干旱胁迫及高温环境中为保持种群稳定性而演化出的一种适应策略.在今后的研究中, 喀什霸王幼苗存活力及种群更新与环境因子的关系等方面还需要进一步的研究. ...
Seed dormancy in six cold desert Brassicaceae species with indehiscent fruits
1
2015
... 生理休眠的植物在极端环境生存及适应过程中表现出较高的生活周期多样性(
Gutterman, 1993 ;
Baskin & Baskin, 2014 ;
Nurulla et al ., 2015 ), 其中非深度生理休眠植物的幼苗存活力往往受到不同环境胁迫(干旱、盐、高温、紫外线)的影响(
Albrecht & McCarthy, 2011 ;
Baskin & Baskin, 2014 ).在干旱环境中普遍存在的干旱及盐胁迫能影响植物种子萌发物候及生活史特点(
Tobe et al ., 2001 ;
Ahmad et al ., 2009 ;
宋乃平等, 2018 ).国内有关研究人员在盐及干旱(缺水)胁迫对寒温带干旱、半干旱荒漠分布植物的生活史特征及其对极端环境的响应(
黄振英等, 2001 ;
Sun et al ., 2009 ;
Lu et al ., 2014 ,
2015 ;
Nurulla et al ., 2015 )方面开展了研究; 但对在我国暖温带干旱荒漠生长的濒危植物种子的生物学特性及其对当地干旱(缺水)环境的响应研究鲜有报道.为此, 本研究对该区域稀有植物喀什霸王(
Zygophyllum kaschgaricum )的结籽率和种子萌发特性进行了研究. ...
Germination requirements of invasive and non-invasive
Atriplex species: A comparative study
1
2001
... 生理休眠植物中有非深度生理休眠的新鲜种子在任何温度及光照下均不能萌发或只能在很窄的温度或光照范围内萌发(
Sun et al ., 2009 ;
Baskin & Baskin, 2014 ;
刘波等, 2015 ;
Soltani et al ., 2017 ).喀什霸王刚采回来的种子在不同温度、光照或黑暗条件下均能萌发, 且彼此间存在显著差异(
p < 0.01,
图3 ); 其中高温黑暗(20/10 ℃及30/20 ℃)处理种子的萌发率高于低温(5/2 ℃和15/2 ℃)黑暗处理种子的萌发率.随着贮藏时间的延长, 贮藏种子的萌发率逐渐提高, 而休眠逐渐解除, 各温变周期下的种子萌发率逐渐提高(
图3 ); 但干藏9个月的种子在低温条件下萌发率低于60%.虽然该物种种子在各温度和不同光周期下均萌发, 但高温和黑暗是最佳的萌发条件.此外, 刚成熟及干藏6个月的种子在不同浓度的GA
3 处理后, 同一浓度GA
3 在各温变周期(光照/黑暗)下的不同干藏处理种子的萌发率间未存在显著差异(
p > 0.05); 但不同浓度GA
3 处理种子的萌发率间存在显著差异(
p < 0.05); 其中50 mmoloL
-1 浓度的萌发率最高, 是打破种子休眠的最合适浓度(
图3 ).从以上结果可以判断喀什霸王种子的休眠类型属于非深度生理休眠类型, 这与我国北部寒温带分布的石生霸王(
Zygophyllum rosovii )和石炭酸灌木(
Larrea divaricate )的生理休眠表现相似(
Venable et al ., 1987 ;
Mandák, 2001 ;
Baskin & Baskin, 2014 ).喀什霸王种子的非深度生理休眠特征可能与该物种长期适应南疆秋天和春天较短的光日照时间及高温过程中进化出的种子萌发对策有关. ...
The fitness consequences of inbreeding in natural populations and their implications for species conservation—A systematic map
1
2015
... 生物多样性保护是有关生态系统安全和稳定性维护的关键科学问题(
Western, 2001 ;
Ren et al ., 2012 ;
覃海宁等, 2017 ).在全球变暖、自然灾害频繁发生、荒漠化严重及种群片段化等内外因素的影响下(
Parmesan & Yohe, 2003 ;
Leadley et al ., 2010 ), 生存在极端环境中的物种受到了不同程度的灭绝威胁(
Western, 2001 ;
张文辉等, 2004 ;
覃海宁等, 2017 ).我国植物多样性丰富, 目前有4 000-5 000种植物正处于不同程度的危机(稀有、濒危、极危)状态中(
Ren et al ., 2012 ).这些植物是生物多样性保护的重要组成部分, 常具有地理分布狭窄、种群规模小、个体数量少以及生境严酷且脆弱等特点(
张文辉等, 2004 ;
Brown et al ., 2015 ;
覃海宁等, 2017 ); 此外, 坐果率、结籽率、种子萌发率和幼苗存活力低等生活史特征也是导致其濒危或灭绝的主要原因(
Neaves et al ., 2015 ).因此, 对该类群的生活史及其对生存环境响应的研究, 在保护稀有濒危植物方面具有重要的科学及实际意义(
Western, 2001 ;
覃海宁等, 2017 ). ...
The effect of seed and rosette cold treatment on germination and flowering time in some Arabidopsis thaliana(Brassicaceae) ecotypes
2
1999
... 与生活史特征相关的种子休眠及幼苗形成过程是种子植物类群后代产生及种群更新的关键阶段(
Baskin & Baskin, 2014 ;
Lu et al ., 2014 ;
孙清琳等, 2019 ).不同植物类群在自身生存及对自然环境的长期适应过程中, 表现出特殊的休眠和萌发特性来保证物种和种群的稳定性(
Nordborg & Bergelson, 1999 ;
Nurulla et al ., 2015 ;
Xia et al ., 2018 ).休眠程度和萌发时间往往受外部因素(光照、温度、水分)及种子自身因素(胚的成熟程度、种皮厚度、种子生活力、调控基因的表达及营养物质)的影响(
Venable et al ., 1987 ;
Casas et al ., 2012 ;
刘波等, 2015 ;
Soltani et al ., 2017 ).根据以上因素的组合程度不同, 种子休眠类型分为物理休眠(PY)、生理休眠(PD)、综合休眠(PY + PD)、形态休眠(MD)和形态生理休眠(MPD) 5种类型(
Galloway & Etterson, 2009 ;
Albrecht & McCarthy, 2011 ;
Baskin & Baskin, 2014 ;
Lu et al ., 2014 ;
Pluntz et al ., 2018 ), 其中生理休眠是自然界中普遍存在且较复杂的休眠类型(
Baskin & Baskin, 2014 ). ...
... 种子萌发作为植物生活周期的主要阶段, 可影响植物雌性(种子)适合度和种群维持及扩大能力(
Baskin & Baskin, 2014 ;
Copete et al ., 2015 ).被子植物在种子物候上所表现出的异质性(
Kalisz & Wardle, 1994 ;
Nordborg & Bergelson, 1999 )作为一种可避免种群灭绝的、谨慎的萌发对策(
Gutterman, 2000 ), 有利于在荒漠极端环境中生长的植物种群的维持和更新(
姚红和谭敦炎, 2005 ;
Nurulla et al ., 2015 ).在本研究中, 非深度生理休眠的喀什霸王种子在春季和秋季均萌发, 虽然该物种在秋季存在一定程度的休眠.在自然环境中, 春季和秋季的温度较低而降水量较多, 因此, 该物种种子通过春秋季萌动的繁殖策略及非深度生理休眠来维持种群的动态平衡及保证种群的更新; 但4-6月气温的上升及降水量的减少导致的干旱胁迫可降低喀什霸王种子的萌发率或幼苗存活能力(
Tobe et al ., 2001 ), 说明新疆南部荒漠环境中降水量是影响非深度生理休眠植物喀什霸王种群更新能力的主要因素(
Lu et al ., 2014 ;
Nurulla et al ., 2015 ).总之, 种子物候上表现出的春秋季萌动繁殖策略及非深度生理休眠是该物种在干旱胁迫及高温环境中为保持种群稳定性而演化出的一种适应策略.在今后的研究中, 喀什霸王幼苗存活力及种群更新与环境因子的关系等方面还需要进一步的研究. ...
Intermediate morphophysiological dormancy allows for life-cycle diversity in the annual weed,Turgenia latifolia(Apiaceae)
5
2015
... 与生活史特征相关的种子休眠及幼苗形成过程是种子植物类群后代产生及种群更新的关键阶段(
Baskin & Baskin, 2014 ;
Lu et al ., 2014 ;
孙清琳等, 2019 ).不同植物类群在自身生存及对自然环境的长期适应过程中, 表现出特殊的休眠和萌发特性来保证物种和种群的稳定性(
Nordborg & Bergelson, 1999 ;
Nurulla et al ., 2015 ;
Xia et al ., 2018 ).休眠程度和萌发时间往往受外部因素(光照、温度、水分)及种子自身因素(胚的成熟程度、种皮厚度、种子生活力、调控基因的表达及营养物质)的影响(
Venable et al ., 1987 ;
Casas et al ., 2012 ;
刘波等, 2015 ;
Soltani et al ., 2017 ).根据以上因素的组合程度不同, 种子休眠类型分为物理休眠(PY)、生理休眠(PD)、综合休眠(PY + PD)、形态休眠(MD)和形态生理休眠(MPD) 5种类型(
Galloway & Etterson, 2009 ;
Albrecht & McCarthy, 2011 ;
Baskin & Baskin, 2014 ;
Lu et al ., 2014 ;
Pluntz et al ., 2018 ), 其中生理休眠是自然界中普遍存在且较复杂的休眠类型(
Baskin & Baskin, 2014 ). ...
... 生理休眠的植物在极端环境生存及适应过程中表现出较高的生活周期多样性(
Gutterman, 1993 ;
Baskin & Baskin, 2014 ;
Nurulla et al ., 2015 ), 其中非深度生理休眠植物的幼苗存活力往往受到不同环境胁迫(干旱、盐、高温、紫外线)的影响(
Albrecht & McCarthy, 2011 ;
Baskin & Baskin, 2014 ).在干旱环境中普遍存在的干旱及盐胁迫能影响植物种子萌发物候及生活史特点(
Tobe et al ., 2001 ;
Ahmad et al ., 2009 ;
宋乃平等, 2018 ).国内有关研究人员在盐及干旱(缺水)胁迫对寒温带干旱、半干旱荒漠分布植物的生活史特征及其对极端环境的响应(
黄振英等, 2001 ;
Sun et al ., 2009 ;
Lu et al ., 2014 ,
2015 ;
Nurulla et al ., 2015 )方面开展了研究; 但对在我国暖温带干旱荒漠生长的濒危植物种子的生物学特性及其对当地干旱(缺水)环境的响应研究鲜有报道.为此, 本研究对该区域稀有植物喀什霸王(
Zygophyllum kaschgaricum )的结籽率和种子萌发特性进行了研究. ...
... ;
Nurulla et al ., 2015 )方面开展了研究; 但对在我国暖温带干旱荒漠生长的濒危植物种子的生物学特性及其对当地干旱(缺水)环境的响应研究鲜有报道.为此, 本研究对该区域稀有植物喀什霸王(
Zygophyllum kaschgaricum )的结籽率和种子萌发特性进行了研究. ...
... 种子萌发作为植物生活周期的主要阶段, 可影响植物雌性(种子)适合度和种群维持及扩大能力(
Baskin & Baskin, 2014 ;
Copete et al ., 2015 ).被子植物在种子物候上所表现出的异质性(
Kalisz & Wardle, 1994 ;
Nordborg & Bergelson, 1999 )作为一种可避免种群灭绝的、谨慎的萌发对策(
Gutterman, 2000 ), 有利于在荒漠极端环境中生长的植物种群的维持和更新(
姚红和谭敦炎, 2005 ;
Nurulla et al ., 2015 ).在本研究中, 非深度生理休眠的喀什霸王种子在春季和秋季均萌发, 虽然该物种在秋季存在一定程度的休眠.在自然环境中, 春季和秋季的温度较低而降水量较多, 因此, 该物种种子通过春秋季萌动的繁殖策略及非深度生理休眠来维持种群的动态平衡及保证种群的更新; 但4-6月气温的上升及降水量的减少导致的干旱胁迫可降低喀什霸王种子的萌发率或幼苗存活能力(
Tobe et al ., 2001 ), 说明新疆南部荒漠环境中降水量是影响非深度生理休眠植物喀什霸王种群更新能力的主要因素(
Lu et al ., 2014 ;
Nurulla et al ., 2015 ).总之, 种子物候上表现出的春秋季萌动繁殖策略及非深度生理休眠是该物种在干旱胁迫及高温环境中为保持种群稳定性而演化出的一种适应策略.在今后的研究中, 喀什霸王幼苗存活力及种群更新与环境因子的关系等方面还需要进一步的研究. ...
... ;
Nurulla et al ., 2015 ).总之, 种子物候上表现出的春秋季萌动繁殖策略及非深度生理休眠是该物种在干旱胁迫及高温环境中为保持种群稳定性而演化出的一种适应策略.在今后的研究中, 喀什霸王幼苗存活力及种群更新与环境因子的关系等方面还需要进一步的研究. ...
A globally coherent fingerprint of climate change impacts across natural systems
1
2003
... 生物多样性保护是有关生态系统安全和稳定性维护的关键科学问题(
Western, 2001 ;
Ren et al ., 2012 ;
覃海宁等, 2017 ).在全球变暖、自然灾害频繁发生、荒漠化严重及种群片段化等内外因素的影响下(
Parmesan & Yohe, 2003 ;
Leadley et al ., 2010 ), 生存在极端环境中的物种受到了不同程度的灭绝威胁(
Western, 2001 ;
张文辉等, 2004 ;
覃海宁等, 2017 ).我国植物多样性丰富, 目前有4 000-5 000种植物正处于不同程度的危机(稀有、濒危、极危)状态中(
Ren et al ., 2012 ).这些植物是生物多样性保护的重要组成部分, 常具有地理分布狭窄、种群规模小、个体数量少以及生境严酷且脆弱等特点(
张文辉等, 2004 ;
Brown et al ., 2015 ;
覃海宁等, 2017 ); 此外, 坐果率、结籽率、种子萌发率和幼苗存活力低等生活史特征也是导致其濒危或灭绝的主要原因(
Neaves et al ., 2015 ).因此, 对该类群的生活史及其对生存环境响应的研究, 在保护稀有濒危植物方面具有重要的科学及实际意义(
Western, 2001 ;
覃海宁等, 2017 ). ...
A general method for estimating seed dormancy and colonisation in annual plants from the observation of existing flora
2
2018
... 与生活史特征相关的种子休眠及幼苗形成过程是种子植物类群后代产生及种群更新的关键阶段(
Baskin & Baskin, 2014 ;
Lu et al ., 2014 ;
孙清琳等, 2019 ).不同植物类群在自身生存及对自然环境的长期适应过程中, 表现出特殊的休眠和萌发特性来保证物种和种群的稳定性(
Nordborg & Bergelson, 1999 ;
Nurulla et al ., 2015 ;
Xia et al ., 2018 ).休眠程度和萌发时间往往受外部因素(光照、温度、水分)及种子自身因素(胚的成熟程度、种皮厚度、种子生活力、调控基因的表达及营养物质)的影响(
Venable et al ., 1987 ;
Casas et al ., 2012 ;
刘波等, 2015 ;
Soltani et al ., 2017 ).根据以上因素的组合程度不同, 种子休眠类型分为物理休眠(PY)、生理休眠(PD)、综合休眠(PY + PD)、形态休眠(MD)和形态生理休眠(MPD) 5种类型(
Galloway & Etterson, 2009 ;
Albrecht & McCarthy, 2011 ;
Baskin & Baskin, 2014 ;
Lu et al ., 2014 ;
Pluntz et al ., 2018 ), 其中生理休眠是自然界中普遍存在且较复杂的休眠类型(
Baskin & Baskin, 2014 ). ...
... 在严酷的极端荒漠环境中, 植物的坐果率和结籽率、种子休眠程度及幼苗存活率能决定植物生活史特征及种群的更新水平.其中果实坐果率及结籽率与植物交配方式及传粉效率有关, 而休眠程度、种子萌发率及幼苗存活率与自然环境的温度和水分等外部因素有关(
黄振英等, 2001 ;
Baskin & Baskin, 2004 ;
Ahmad et al ., 2009 ;
Pluntz et al ., 2018 ;
Xia et al ., 2018 ).本研究中, 喀什霸王果实的自然坐果率及结籽率较低, 因为在其开花的4月中旬, 塔克拉玛干沙漠西南部荒漠环境的昼夜温度差异及风速较大, 早春活动的昆虫种类少; 此外, 该物种存在花内雌性先熟现象(未发表数据).以上因素导致的传粉限制可能是降低自然坐果率及结籽率(46.97 ± 2.56)%的主要因素, 此推测需要进一步验证. ...
中国被子植物濒危等级的评估
4
2017
... 生物多样性保护是有关生态系统安全和稳定性维护的关键科学问题(
Western, 2001 ;
Ren et al ., 2012 ;
覃海宁等, 2017 ).在全球变暖、自然灾害频繁发生、荒漠化严重及种群片段化等内外因素的影响下(
Parmesan & Yohe, 2003 ;
Leadley et al ., 2010 ), 生存在极端环境中的物种受到了不同程度的灭绝威胁(
Western, 2001 ;
张文辉等, 2004 ;
覃海宁等, 2017 ).我国植物多样性丰富, 目前有4 000-5 000种植物正处于不同程度的危机(稀有、濒危、极危)状态中(
Ren et al ., 2012 ).这些植物是生物多样性保护的重要组成部分, 常具有地理分布狭窄、种群规模小、个体数量少以及生境严酷且脆弱等特点(
张文辉等, 2004 ;
Brown et al ., 2015 ;
覃海宁等, 2017 ); 此外, 坐果率、结籽率、种子萌发率和幼苗存活力低等生活史特征也是导致其濒危或灭绝的主要原因(
Neaves et al ., 2015 ).因此, 对该类群的生活史及其对生存环境响应的研究, 在保护稀有濒危植物方面具有重要的科学及实际意义(
Western, 2001 ;
覃海宁等, 2017 ). ...
... ;
覃海宁等, 2017 ).我国植物多样性丰富, 目前有4 000-5 000种植物正处于不同程度的危机(稀有、濒危、极危)状态中(
Ren et al ., 2012 ).这些植物是生物多样性保护的重要组成部分, 常具有地理分布狭窄、种群规模小、个体数量少以及生境严酷且脆弱等特点(
张文辉等, 2004 ;
Brown et al ., 2015 ;
覃海宁等, 2017 ); 此外, 坐果率、结籽率、种子萌发率和幼苗存活力低等生活史特征也是导致其濒危或灭绝的主要原因(
Neaves et al ., 2015 ).因此, 对该类群的生活史及其对生存环境响应的研究, 在保护稀有濒危植物方面具有重要的科学及实际意义(
Western, 2001 ;
覃海宁等, 2017 ). ...
... ;
覃海宁等, 2017 ); 此外, 坐果率、结籽率、种子萌发率和幼苗存活力低等生活史特征也是导致其濒危或灭绝的主要原因(
Neaves et al ., 2015 ).因此, 对该类群的生活史及其对生存环境响应的研究, 在保护稀有濒危植物方面具有重要的科学及实际意义(
Western, 2001 ;
覃海宁等, 2017 ). ...
... ;
覃海宁等, 2017 ). ...
中国被子植物濒危等级的评估
4
2017
... 生物多样性保护是有关生态系统安全和稳定性维护的关键科学问题(
Western, 2001 ;
Ren et al ., 2012 ;
覃海宁等, 2017 ).在全球变暖、自然灾害频繁发生、荒漠化严重及种群片段化等内外因素的影响下(
Parmesan & Yohe, 2003 ;
Leadley et al ., 2010 ), 生存在极端环境中的物种受到了不同程度的灭绝威胁(
Western, 2001 ;
张文辉等, 2004 ;
覃海宁等, 2017 ).我国植物多样性丰富, 目前有4 000-5 000种植物正处于不同程度的危机(稀有、濒危、极危)状态中(
Ren et al ., 2012 ).这些植物是生物多样性保护的重要组成部分, 常具有地理分布狭窄、种群规模小、个体数量少以及生境严酷且脆弱等特点(
张文辉等, 2004 ;
Brown et al ., 2015 ;
覃海宁等, 2017 ); 此外, 坐果率、结籽率、种子萌发率和幼苗存活力低等生活史特征也是导致其濒危或灭绝的主要原因(
Neaves et al ., 2015 ).因此, 对该类群的生活史及其对生存环境响应的研究, 在保护稀有濒危植物方面具有重要的科学及实际意义(
Western, 2001 ;
覃海宁等, 2017 ). ...
... ;
覃海宁等, 2017 ).我国植物多样性丰富, 目前有4 000-5 000种植物正处于不同程度的危机(稀有、濒危、极危)状态中(
Ren et al ., 2012 ).这些植物是生物多样性保护的重要组成部分, 常具有地理分布狭窄、种群规模小、个体数量少以及生境严酷且脆弱等特点(
张文辉等, 2004 ;
Brown et al ., 2015 ;
覃海宁等, 2017 ); 此外, 坐果率、结籽率、种子萌发率和幼苗存活力低等生活史特征也是导致其濒危或灭绝的主要原因(
Neaves et al ., 2015 ).因此, 对该类群的生活史及其对生存环境响应的研究, 在保护稀有濒危植物方面具有重要的科学及实际意义(
Western, 2001 ;
覃海宁等, 2017 ). ...
... ;
覃海宁等, 2017 ); 此外, 坐果率、结籽率、种子萌发率和幼苗存活力低等生活史特征也是导致其濒危或灭绝的主要原因(
Neaves et al ., 2015 ).因此, 对该类群的生活史及其对生存环境响应的研究, 在保护稀有濒危植物方面具有重要的科学及实际意义(
Western, 2001 ;
覃海宁等, 2017 ). ...
... ;
覃海宁等, 2017 ). ...
Wild plant species with extremely small populations require conservation and reintroduction in China
2
2012
... 生物多样性保护是有关生态系统安全和稳定性维护的关键科学问题(
Western, 2001 ;
Ren et al ., 2012 ;
覃海宁等, 2017 ).在全球变暖、自然灾害频繁发生、荒漠化严重及种群片段化等内外因素的影响下(
Parmesan & Yohe, 2003 ;
Leadley et al ., 2010 ), 生存在极端环境中的物种受到了不同程度的灭绝威胁(
Western, 2001 ;
张文辉等, 2004 ;
覃海宁等, 2017 ).我国植物多样性丰富, 目前有4 000-5 000种植物正处于不同程度的危机(稀有、濒危、极危)状态中(
Ren et al ., 2012 ).这些植物是生物多样性保护的重要组成部分, 常具有地理分布狭窄、种群规模小、个体数量少以及生境严酷且脆弱等特点(
张文辉等, 2004 ;
Brown et al ., 2015 ;
覃海宁等, 2017 ); 此外, 坐果率、结籽率、种子萌发率和幼苗存活力低等生活史特征也是导致其濒危或灭绝的主要原因(
Neaves et al ., 2015 ).因此, 对该类群的生活史及其对生存环境响应的研究, 在保护稀有濒危植物方面具有重要的科学及实际意义(
Western, 2001 ;
覃海宁等, 2017 ). ...
... ).我国植物多样性丰富, 目前有4 000-5 000种植物正处于不同程度的危机(稀有、濒危、极危)状态中(
Ren et al ., 2012 ).这些植物是生物多样性保护的重要组成部分, 常具有地理分布狭窄、种群规模小、个体数量少以及生境严酷且脆弱等特点(
张文辉等, 2004 ;
Brown et al ., 2015 ;
覃海宁等, 2017 ); 此外, 坐果率、结籽率、种子萌发率和幼苗存活力低等生活史特征也是导致其濒危或灭绝的主要原因(
Neaves et al ., 2015 ).因此, 对该类群的生活史及其对生存环境响应的研究, 在保护稀有濒危植物方面具有重要的科学及实际意义(
Western, 2001 ;
覃海宁等, 2017 ). ...
Water stress, temperature regimes and light control induction, and loss of secondary dormancy in
Brassica napus L. seeds
2
2017
... 与生活史特征相关的种子休眠及幼苗形成过程是种子植物类群后代产生及种群更新的关键阶段(
Baskin & Baskin, 2014 ;
Lu et al ., 2014 ;
孙清琳等, 2019 ).不同植物类群在自身生存及对自然环境的长期适应过程中, 表现出特殊的休眠和萌发特性来保证物种和种群的稳定性(
Nordborg & Bergelson, 1999 ;
Nurulla et al ., 2015 ;
Xia et al ., 2018 ).休眠程度和萌发时间往往受外部因素(光照、温度、水分)及种子自身因素(胚的成熟程度、种皮厚度、种子生活力、调控基因的表达及营养物质)的影响(
Venable et al ., 1987 ;
Casas et al ., 2012 ;
刘波等, 2015 ;
Soltani et al ., 2017 ).根据以上因素的组合程度不同, 种子休眠类型分为物理休眠(PY)、生理休眠(PD)、综合休眠(PY + PD)、形态休眠(MD)和形态生理休眠(MPD) 5种类型(
Galloway & Etterson, 2009 ;
Albrecht & McCarthy, 2011 ;
Baskin & Baskin, 2014 ;
Lu et al ., 2014 ;
Pluntz et al ., 2018 ), 其中生理休眠是自然界中普遍存在且较复杂的休眠类型(
Baskin & Baskin, 2014 ). ...
... 生理休眠植物中有非深度生理休眠的新鲜种子在任何温度及光照下均不能萌发或只能在很窄的温度或光照范围内萌发(
Sun et al ., 2009 ;
Baskin & Baskin, 2014 ;
刘波等, 2015 ;
Soltani et al ., 2017 ).喀什霸王刚采回来的种子在不同温度、光照或黑暗条件下均能萌发, 且彼此间存在显著差异(
p < 0.01,
图3 ); 其中高温黑暗(20/10 ℃及30/20 ℃)处理种子的萌发率高于低温(5/2 ℃和15/2 ℃)黑暗处理种子的萌发率.随着贮藏时间的延长, 贮藏种子的萌发率逐渐提高, 而休眠逐渐解除, 各温变周期下的种子萌发率逐渐提高(
图3 ); 但干藏9个月的种子在低温条件下萌发率低于60%.虽然该物种种子在各温度和不同光周期下均萌发, 但高温和黑暗是最佳的萌发条件.此外, 刚成熟及干藏6个月的种子在不同浓度的GA
3 处理后, 同一浓度GA
3 在各温变周期(光照/黑暗)下的不同干藏处理种子的萌发率间未存在显著差异(
p > 0.05); 但不同浓度GA
3 处理种子的萌发率间存在显著差异(
p < 0.05); 其中50 mmoloL
-1 浓度的萌发率最高, 是打破种子休眠的最合适浓度(
图3 ).从以上结果可以判断喀什霸王种子的休眠类型属于非深度生理休眠类型, 这与我国北部寒温带分布的石生霸王(
Zygophyllum rosovii )和石炭酸灌木(
Larrea divaricate )的生理休眠表现相似(
Venable et al ., 1987 ;
Mandák, 2001 ;
Baskin & Baskin, 2014 ).喀什霸王种子的非深度生理休眠特征可能与该物种长期适应南疆秋天和春天较短的光日照时间及高温过程中进化出的种子萌发对策有关. ...
荒漠草原“土岛”生境群落物种共存机制
1
2018
... 生理休眠的植物在极端环境生存及适应过程中表现出较高的生活周期多样性(
Gutterman, 1993 ;
Baskin & Baskin, 2014 ;
Nurulla et al ., 2015 ), 其中非深度生理休眠植物的幼苗存活力往往受到不同环境胁迫(干旱、盐、高温、紫外线)的影响(
Albrecht & McCarthy, 2011 ;
Baskin & Baskin, 2014 ).在干旱环境中普遍存在的干旱及盐胁迫能影响植物种子萌发物候及生活史特点(
Tobe et al ., 2001 ;
Ahmad et al ., 2009 ;
宋乃平等, 2018 ).国内有关研究人员在盐及干旱(缺水)胁迫对寒温带干旱、半干旱荒漠分布植物的生活史特征及其对极端环境的响应(
黄振英等, 2001 ;
Sun et al ., 2009 ;
Lu et al ., 2014 ,
2015 ;
Nurulla et al ., 2015 )方面开展了研究; 但对在我国暖温带干旱荒漠生长的濒危植物种子的生物学特性及其对当地干旱(缺水)环境的响应研究鲜有报道.为此, 本研究对该区域稀有植物喀什霸王(
Zygophyllum kaschgaricum )的结籽率和种子萌发特性进行了研究. ...
荒漠草原“土岛”生境群落物种共存机制
1
2018
... 生理休眠的植物在极端环境生存及适应过程中表现出较高的生活周期多样性(
Gutterman, 1993 ;
Baskin & Baskin, 2014 ;
Nurulla et al ., 2015 ), 其中非深度生理休眠植物的幼苗存活力往往受到不同环境胁迫(干旱、盐、高温、紫外线)的影响(
Albrecht & McCarthy, 2011 ;
Baskin & Baskin, 2014 ).在干旱环境中普遍存在的干旱及盐胁迫能影响植物种子萌发物候及生活史特点(
Tobe et al ., 2001 ;
Ahmad et al ., 2009 ;
宋乃平等, 2018 ).国内有关研究人员在盐及干旱(缺水)胁迫对寒温带干旱、半干旱荒漠分布植物的生活史特征及其对极端环境的响应(
黄振英等, 2001 ;
Sun et al ., 2009 ;
Lu et al ., 2014 ,
2015 ;
Nurulla et al ., 2015 )方面开展了研究; 但对在我国暖温带干旱荒漠生长的濒危植物种子的生物学特性及其对当地干旱(缺水)环境的响应研究鲜有报道.为此, 本研究对该区域稀有植物喀什霸王(
Zygophyllum kaschgaricum )的结籽率和种子萌发特性进行了研究. ...
Dormancy and germination characteristics of the trimorphic achenes of Garhadiolus papposus( Asteraceae), an annual ephemeral from the Junggar Desert, China
2
2009
... 生理休眠的植物在极端环境生存及适应过程中表现出较高的生活周期多样性(
Gutterman, 1993 ;
Baskin & Baskin, 2014 ;
Nurulla et al ., 2015 ), 其中非深度生理休眠植物的幼苗存活力往往受到不同环境胁迫(干旱、盐、高温、紫外线)的影响(
Albrecht & McCarthy, 2011 ;
Baskin & Baskin, 2014 ).在干旱环境中普遍存在的干旱及盐胁迫能影响植物种子萌发物候及生活史特点(
Tobe et al ., 2001 ;
Ahmad et al ., 2009 ;
宋乃平等, 2018 ).国内有关研究人员在盐及干旱(缺水)胁迫对寒温带干旱、半干旱荒漠分布植物的生活史特征及其对极端环境的响应(
黄振英等, 2001 ;
Sun et al ., 2009 ;
Lu et al ., 2014 ,
2015 ;
Nurulla et al ., 2015 )方面开展了研究; 但对在我国暖温带干旱荒漠生长的濒危植物种子的生物学特性及其对当地干旱(缺水)环境的响应研究鲜有报道.为此, 本研究对该区域稀有植物喀什霸王(
Zygophyllum kaschgaricum )的结籽率和种子萌发特性进行了研究. ...
... 生理休眠植物中有非深度生理休眠的新鲜种子在任何温度及光照下均不能萌发或只能在很窄的温度或光照范围内萌发(
Sun et al ., 2009 ;
Baskin & Baskin, 2014 ;
刘波等, 2015 ;
Soltani et al ., 2017 ).喀什霸王刚采回来的种子在不同温度、光照或黑暗条件下均能萌发, 且彼此间存在显著差异(
p < 0.01,
图3 ); 其中高温黑暗(20/10 ℃及30/20 ℃)处理种子的萌发率高于低温(5/2 ℃和15/2 ℃)黑暗处理种子的萌发率.随着贮藏时间的延长, 贮藏种子的萌发率逐渐提高, 而休眠逐渐解除, 各温变周期下的种子萌发率逐渐提高(
图3 ); 但干藏9个月的种子在低温条件下萌发率低于60%.虽然该物种种子在各温度和不同光周期下均萌发, 但高温和黑暗是最佳的萌发条件.此外, 刚成熟及干藏6个月的种子在不同浓度的GA
3 处理后, 同一浓度GA
3 在各温变周期(光照/黑暗)下的不同干藏处理种子的萌发率间未存在显著差异(
p > 0.05); 但不同浓度GA
3 处理种子的萌发率间存在显著差异(
p < 0.05); 其中50 mmoloL
-1 浓度的萌发率最高, 是打破种子休眠的最合适浓度(
图3 ).从以上结果可以判断喀什霸王种子的休眠类型属于非深度生理休眠类型, 这与我国北部寒温带分布的石生霸王(
Zygophyllum rosovii )和石炭酸灌木(
Larrea divaricate )的生理休眠表现相似(
Venable et al ., 1987 ;
Mandák, 2001 ;
Baskin & Baskin, 2014 ).喀什霸王种子的非深度生理休眠特征可能与该物种长期适应南疆秋天和春天较短的光日照时间及高温过程中进化出的种子萌发对策有关. ...
长芒草(
Stipa bungeana )种子萌发与出苗对关键环境因子的响应
1
2019
... 与生活史特征相关的种子休眠及幼苗形成过程是种子植物类群后代产生及种群更新的关键阶段(
Baskin & Baskin, 2014 ;
Lu et al ., 2014 ;
孙清琳等, 2019 ).不同植物类群在自身生存及对自然环境的长期适应过程中, 表现出特殊的休眠和萌发特性来保证物种和种群的稳定性(
Nordborg & Bergelson, 1999 ;
Nurulla et al ., 2015 ;
Xia et al ., 2018 ).休眠程度和萌发时间往往受外部因素(光照、温度、水分)及种子自身因素(胚的成熟程度、种皮厚度、种子生活力、调控基因的表达及营养物质)的影响(
Venable et al ., 1987 ;
Casas et al ., 2012 ;
刘波等, 2015 ;
Soltani et al ., 2017 ).根据以上因素的组合程度不同, 种子休眠类型分为物理休眠(PY)、生理休眠(PD)、综合休眠(PY + PD)、形态休眠(MD)和形态生理休眠(MPD) 5种类型(
Galloway & Etterson, 2009 ;
Albrecht & McCarthy, 2011 ;
Baskin & Baskin, 2014 ;
Lu et al ., 2014 ;
Pluntz et al ., 2018 ), 其中生理休眠是自然界中普遍存在且较复杂的休眠类型(
Baskin & Baskin, 2014 ). ...
长芒草(
Stipa bungeana )种子萌发与出苗对关键环境因子的响应
1
2019
... 与生活史特征相关的种子休眠及幼苗形成过程是种子植物类群后代产生及种群更新的关键阶段(
Baskin & Baskin, 2014 ;
Lu et al ., 2014 ;
孙清琳等, 2019 ).不同植物类群在自身生存及对自然环境的长期适应过程中, 表现出特殊的休眠和萌发特性来保证物种和种群的稳定性(
Nordborg & Bergelson, 1999 ;
Nurulla et al ., 2015 ;
Xia et al ., 2018 ).休眠程度和萌发时间往往受外部因素(光照、温度、水分)及种子自身因素(胚的成熟程度、种皮厚度、种子生活力、调控基因的表达及营养物质)的影响(
Venable et al ., 1987 ;
Casas et al ., 2012 ;
刘波等, 2015 ;
Soltani et al ., 2017 ).根据以上因素的组合程度不同, 种子休眠类型分为物理休眠(PY)、生理休眠(PD)、综合休眠(PY + PD)、形态休眠(MD)和形态生理休眠(MPD) 5种类型(
Galloway & Etterson, 2009 ;
Albrecht & McCarthy, 2011 ;
Baskin & Baskin, 2014 ;
Lu et al ., 2014 ;
Pluntz et al ., 2018 ), 其中生理休眠是自然界中普遍存在且较复杂的休眠类型(
Baskin & Baskin, 2014 ). ...
Characteristics of seed germination in five non-halophytic Chinese desert shrub species
3
2001
... 生理休眠的植物在极端环境生存及适应过程中表现出较高的生活周期多样性(
Gutterman, 1993 ;
Baskin & Baskin, 2014 ;
Nurulla et al ., 2015 ), 其中非深度生理休眠植物的幼苗存活力往往受到不同环境胁迫(干旱、盐、高温、紫外线)的影响(
Albrecht & McCarthy, 2011 ;
Baskin & Baskin, 2014 ).在干旱环境中普遍存在的干旱及盐胁迫能影响植物种子萌发物候及生活史特点(
Tobe et al ., 2001 ;
Ahmad et al ., 2009 ;
宋乃平等, 2018 ).国内有关研究人员在盐及干旱(缺水)胁迫对寒温带干旱、半干旱荒漠分布植物的生活史特征及其对极端环境的响应(
黄振英等, 2001 ;
Sun et al ., 2009 ;
Lu et al ., 2014 ,
2015 ;
Nurulla et al ., 2015 )方面开展了研究; 但对在我国暖温带干旱荒漠生长的濒危植物种子的生物学特性及其对当地干旱(缺水)环境的响应研究鲜有报道.为此, 本研究对该区域稀有植物喀什霸王(
Zygophyllum kaschgaricum )的结籽率和种子萌发特性进行了研究. ...
... 种子的不同季节萌发能促进植物物候的多样性(
Baskin & Baskin, 1988 ;
刘波等, 2015 ); 春秋季萌发是种子植物在不可预测环境的两头下注策略和种子生理后熟之间的权衡(
Lu et al ., 2014 ).在本研究中, 室内不同浓度PEG处理种子在低温条件下的萌发率比高温高(
图5 ).此外, 随着干藏时间的延长, 种子对水分的需求提高(
图1 ).塔克拉玛干沙漠西南部自然环境的平均月降水量低于10 mm, 该物种果实成熟的6-9月的平均气温在25 ℃以上, 气候特点与以色列的Negev荒漠(
Gutterman, 1993 ,
2000 )相似.因此, 该区域春季的降水量、气温及光照强度等气候特点能限制有非深度生理休眠种子的萌发及幼苗存活率.在降水时间分布不均匀的塔克拉玛干沙漠西南部荒漠环境中(
图1 ), 该物种种子的萌发对策主要是对降水做出响应(
Tobe et al ., 2001 ;
Ahmad et al ., 2009 ), 其中种子的春季萌发及幼苗形成过程被认为是一种逃避冬天死亡的有利方式, 而秋天萌发是种子对低温的适应及竞争优势(
Gutterman, 1993 ).喀什霸王干藏6个月的种子在水分充足时才萌发, 说明该物种在塔克拉玛干沙漠西南部的极端环境的长期生存过程中, 随时面临着自然环境难以预测的生存风险, 通过种子的非深度生理休眠可获得更多的生存机会, 保持物种亲本性状. ...
... 种子萌发作为植物生活周期的主要阶段, 可影响植物雌性(种子)适合度和种群维持及扩大能力(
Baskin & Baskin, 2014 ;
Copete et al ., 2015 ).被子植物在种子物候上所表现出的异质性(
Kalisz & Wardle, 1994 ;
Nordborg & Bergelson, 1999 )作为一种可避免种群灭绝的、谨慎的萌发对策(
Gutterman, 2000 ), 有利于在荒漠极端环境中生长的植物种群的维持和更新(
姚红和谭敦炎, 2005 ;
Nurulla et al ., 2015 ).在本研究中, 非深度生理休眠的喀什霸王种子在春季和秋季均萌发, 虽然该物种在秋季存在一定程度的休眠.在自然环境中, 春季和秋季的温度较低而降水量较多, 因此, 该物种种子通过春秋季萌动的繁殖策略及非深度生理休眠来维持种群的动态平衡及保证种群的更新; 但4-6月气温的上升及降水量的减少导致的干旱胁迫可降低喀什霸王种子的萌发率或幼苗存活能力(
Tobe et al ., 2001 ), 说明新疆南部荒漠环境中降水量是影响非深度生理休眠植物喀什霸王种群更新能力的主要因素(
Lu et al ., 2014 ;
Nurulla et al ., 2015 ).总之, 种子物候上表现出的春秋季萌动繁殖策略及非深度生理休眠是该物种在干旱胁迫及高温环境中为保持种群稳定性而演化出的一种适应策略.在今后的研究中, 喀什霸王幼苗存活力及种群更新与环境因子的关系等方面还需要进一步的研究. ...
The ecology of seed heteromorphism in
Heterosperma pinnatum in central Mexico
2
1987
... 与生活史特征相关的种子休眠及幼苗形成过程是种子植物类群后代产生及种群更新的关键阶段(
Baskin & Baskin, 2014 ;
Lu et al ., 2014 ;
孙清琳等, 2019 ).不同植物类群在自身生存及对自然环境的长期适应过程中, 表现出特殊的休眠和萌发特性来保证物种和种群的稳定性(
Nordborg & Bergelson, 1999 ;
Nurulla et al ., 2015 ;
Xia et al ., 2018 ).休眠程度和萌发时间往往受外部因素(光照、温度、水分)及种子自身因素(胚的成熟程度、种皮厚度、种子生活力、调控基因的表达及营养物质)的影响(
Venable et al ., 1987 ;
Casas et al ., 2012 ;
刘波等, 2015 ;
Soltani et al ., 2017 ).根据以上因素的组合程度不同, 种子休眠类型分为物理休眠(PY)、生理休眠(PD)、综合休眠(PY + PD)、形态休眠(MD)和形态生理休眠(MPD) 5种类型(
Galloway & Etterson, 2009 ;
Albrecht & McCarthy, 2011 ;
Baskin & Baskin, 2014 ;
Lu et al ., 2014 ;
Pluntz et al ., 2018 ), 其中生理休眠是自然界中普遍存在且较复杂的休眠类型(
Baskin & Baskin, 2014 ). ...
... 生理休眠植物中有非深度生理休眠的新鲜种子在任何温度及光照下均不能萌发或只能在很窄的温度或光照范围内萌发(
Sun et al ., 2009 ;
Baskin & Baskin, 2014 ;
刘波等, 2015 ;
Soltani et al ., 2017 ).喀什霸王刚采回来的种子在不同温度、光照或黑暗条件下均能萌发, 且彼此间存在显著差异(
p < 0.01,
图3 ); 其中高温黑暗(20/10 ℃及30/20 ℃)处理种子的萌发率高于低温(5/2 ℃和15/2 ℃)黑暗处理种子的萌发率.随着贮藏时间的延长, 贮藏种子的萌发率逐渐提高, 而休眠逐渐解除, 各温变周期下的种子萌发率逐渐提高(
图3 ); 但干藏9个月的种子在低温条件下萌发率低于60%.虽然该物种种子在各温度和不同光周期下均萌发, 但高温和黑暗是最佳的萌发条件.此外, 刚成熟及干藏6个月的种子在不同浓度的GA
3 处理后, 同一浓度GA
3 在各温变周期(光照/黑暗)下的不同干藏处理种子的萌发率间未存在显著差异(
p > 0.05); 但不同浓度GA
3 处理种子的萌发率间存在显著差异(
p < 0.05); 其中50 mmoloL
-1 浓度的萌发率最高, 是打破种子休眠的最合适浓度(
图3 ).从以上结果可以判断喀什霸王种子的休眠类型属于非深度生理休眠类型, 这与我国北部寒温带分布的石生霸王(
Zygophyllum rosovii )和石炭酸灌木(
Larrea divaricate )的生理休眠表现相似(
Venable et al ., 1987 ;
Mandák, 2001 ;
Baskin & Baskin, 2014 ).喀什霸王种子的非深度生理休眠特征可能与该物种长期适应南疆秋天和春天较短的光日照时间及高温过程中进化出的种子萌发对策有关. ...
Human-modified ecosystems and future evolution
3
2001
... 生物多样性保护是有关生态系统安全和稳定性维护的关键科学问题(
Western, 2001 ;
Ren et al ., 2012 ;
覃海宁等, 2017 ).在全球变暖、自然灾害频繁发生、荒漠化严重及种群片段化等内外因素的影响下(
Parmesan & Yohe, 2003 ;
Leadley et al ., 2010 ), 生存在极端环境中的物种受到了不同程度的灭绝威胁(
Western, 2001 ;
张文辉等, 2004 ;
覃海宁等, 2017 ).我国植物多样性丰富, 目前有4 000-5 000种植物正处于不同程度的危机(稀有、濒危、极危)状态中(
Ren et al ., 2012 ).这些植物是生物多样性保护的重要组成部分, 常具有地理分布狭窄、种群规模小、个体数量少以及生境严酷且脆弱等特点(
张文辉等, 2004 ;
Brown et al ., 2015 ;
覃海宁等, 2017 ); 此外, 坐果率、结籽率、种子萌发率和幼苗存活力低等生活史特征也是导致其濒危或灭绝的主要原因(
Neaves et al ., 2015 ).因此, 对该类群的生活史及其对生存环境响应的研究, 在保护稀有濒危植物方面具有重要的科学及实际意义(
Western, 2001 ;
覃海宁等, 2017 ). ...
... ), 生存在极端环境中的物种受到了不同程度的灭绝威胁(
Western, 2001 ;
张文辉等, 2004 ;
覃海宁等, 2017 ).我国植物多样性丰富, 目前有4 000-5 000种植物正处于不同程度的危机(稀有、濒危、极危)状态中(
Ren et al ., 2012 ).这些植物是生物多样性保护的重要组成部分, 常具有地理分布狭窄、种群规模小、个体数量少以及生境严酷且脆弱等特点(
张文辉等, 2004 ;
Brown et al ., 2015 ;
覃海宁等, 2017 ); 此外, 坐果率、结籽率、种子萌发率和幼苗存活力低等生活史特征也是导致其濒危或灭绝的主要原因(
Neaves et al ., 2015 ).因此, 对该类群的生活史及其对生存环境响应的研究, 在保护稀有濒危植物方面具有重要的科学及实际意义(
Western, 2001 ;
覃海宁等, 2017 ). ...
... ).因此, 对该类群的生活史及其对生存环境响应的研究, 在保护稀有濒危植物方面具有重要的科学及实际意义(
Western, 2001 ;
覃海宁等, 2017 ). ...
Integrating proteomics and enzymatic profiling to decipher seed metabolism affected by temperature in seed dormancy and germination
2
2018
... 与生活史特征相关的种子休眠及幼苗形成过程是种子植物类群后代产生及种群更新的关键阶段(
Baskin & Baskin, 2014 ;
Lu et al ., 2014 ;
孙清琳等, 2019 ).不同植物类群在自身生存及对自然环境的长期适应过程中, 表现出特殊的休眠和萌发特性来保证物种和种群的稳定性(
Nordborg & Bergelson, 1999 ;
Nurulla et al ., 2015 ;
Xia et al ., 2018 ).休眠程度和萌发时间往往受外部因素(光照、温度、水分)及种子自身因素(胚的成熟程度、种皮厚度、种子生活力、调控基因的表达及营养物质)的影响(
Venable et al ., 1987 ;
Casas et al ., 2012 ;
刘波等, 2015 ;
Soltani et al ., 2017 ).根据以上因素的组合程度不同, 种子休眠类型分为物理休眠(PY)、生理休眠(PD)、综合休眠(PY + PD)、形态休眠(MD)和形态生理休眠(MPD) 5种类型(
Galloway & Etterson, 2009 ;
Albrecht & McCarthy, 2011 ;
Baskin & Baskin, 2014 ;
Lu et al ., 2014 ;
Pluntz et al ., 2018 ), 其中生理休眠是自然界中普遍存在且较复杂的休眠类型(
Baskin & Baskin, 2014 ). ...
... 在严酷的极端荒漠环境中, 植物的坐果率和结籽率、种子休眠程度及幼苗存活率能决定植物生活史特征及种群的更新水平.其中果实坐果率及结籽率与植物交配方式及传粉效率有关, 而休眠程度、种子萌发率及幼苗存活率与自然环境的温度和水分等外部因素有关(
黄振英等, 2001 ;
Baskin & Baskin, 2004 ;
Ahmad et al ., 2009 ;
Pluntz et al ., 2018 ;
Xia et al ., 2018 ).本研究中, 喀什霸王果实的自然坐果率及结籽率较低, 因为在其开花的4月中旬, 塔克拉玛干沙漠西南部荒漠环境的昼夜温度差异及风速较大, 早春活动的昆虫种类少; 此外, 该物种存在花内雌性先熟现象(未发表数据).以上因素导致的传粉限制可能是降低自然坐果率及结籽率(46.97 ± 2.56)%的主要因素, 此推测需要进一步验证. ...
胡卢巴属4种短命植物个体大小依赖的繁殖输出与生活史对策
1
2005
... 种子萌发作为植物生活周期的主要阶段, 可影响植物雌性(种子)适合度和种群维持及扩大能力(
Baskin & Baskin, 2014 ;
Copete et al ., 2015 ).被子植物在种子物候上所表现出的异质性(
Kalisz & Wardle, 1994 ;
Nordborg & Bergelson, 1999 )作为一种可避免种群灭绝的、谨慎的萌发对策(
Gutterman, 2000 ), 有利于在荒漠极端环境中生长的植物种群的维持和更新(
姚红和谭敦炎, 2005 ;
Nurulla et al ., 2015 ).在本研究中, 非深度生理休眠的喀什霸王种子在春季和秋季均萌发, 虽然该物种在秋季存在一定程度的休眠.在自然环境中, 春季和秋季的温度较低而降水量较多, 因此, 该物种种子通过春秋季萌动的繁殖策略及非深度生理休眠来维持种群的动态平衡及保证种群的更新; 但4-6月气温的上升及降水量的减少导致的干旱胁迫可降低喀什霸王种子的萌发率或幼苗存活能力(
Tobe et al ., 2001 ), 说明新疆南部荒漠环境中降水量是影响非深度生理休眠植物喀什霸王种群更新能力的主要因素(
Lu et al ., 2014 ;
Nurulla et al ., 2015 ).总之, 种子物候上表现出的春秋季萌动繁殖策略及非深度生理休眠是该物种在干旱胁迫及高温环境中为保持种群稳定性而演化出的一种适应策略.在今后的研究中, 喀什霸王幼苗存活力及种群更新与环境因子的关系等方面还需要进一步的研究. ...
胡卢巴属4种短命植物个体大小依赖的繁殖输出与生活史对策
1
2005
... 种子萌发作为植物生活周期的主要阶段, 可影响植物雌性(种子)适合度和种群维持及扩大能力(
Baskin & Baskin, 2014 ;
Copete et al ., 2015 ).被子植物在种子物候上所表现出的异质性(
Kalisz & Wardle, 1994 ;
Nordborg & Bergelson, 1999 )作为一种可避免种群灭绝的、谨慎的萌发对策(
Gutterman, 2000 ), 有利于在荒漠极端环境中生长的植物种群的维持和更新(
姚红和谭敦炎, 2005 ;
Nurulla et al ., 2015 ).在本研究中, 非深度生理休眠的喀什霸王种子在春季和秋季均萌发, 虽然该物种在秋季存在一定程度的休眠.在自然环境中, 春季和秋季的温度较低而降水量较多, 因此, 该物种种子通过春秋季萌动的繁殖策略及非深度生理休眠来维持种群的动态平衡及保证种群的更新; 但4-6月气温的上升及降水量的减少导致的干旱胁迫可降低喀什霸王种子的萌发率或幼苗存活能力(
Tobe et al ., 2001 ), 说明新疆南部荒漠环境中降水量是影响非深度生理休眠植物喀什霸王种群更新能力的主要因素(
Lu et al ., 2014 ;
Nurulla et al ., 2015 ).总之, 种子物候上表现出的春秋季萌动繁殖策略及非深度生理休眠是该物种在干旱胁迫及高温环境中为保持种群稳定性而演化出的一种适应策略.在今后的研究中, 喀什霸王幼苗存活力及种群更新与环境因子的关系等方面还需要进一步的研究. ...
2006
2006
濒危植物秦岭冷杉地理分布和生物生态学特性研究
4
2004
... 生物多样性保护是有关生态系统安全和稳定性维护的关键科学问题(
Western, 2001 ;
Ren et al ., 2012 ;
覃海宁等, 2017 ).在全球变暖、自然灾害频繁发生、荒漠化严重及种群片段化等内外因素的影响下(
Parmesan & Yohe, 2003 ;
Leadley et al ., 2010 ), 生存在极端环境中的物种受到了不同程度的灭绝威胁(
Western, 2001 ;
张文辉等, 2004 ;
覃海宁等, 2017 ).我国植物多样性丰富, 目前有4 000-5 000种植物正处于不同程度的危机(稀有、濒危、极危)状态中(
Ren et al ., 2012 ).这些植物是生物多样性保护的重要组成部分, 常具有地理分布狭窄、种群规模小、个体数量少以及生境严酷且脆弱等特点(
张文辉等, 2004 ;
Brown et al ., 2015 ;
覃海宁等, 2017 ); 此外, 坐果率、结籽率、种子萌发率和幼苗存活力低等生活史特征也是导致其濒危或灭绝的主要原因(
Neaves et al ., 2015 ).因此, 对该类群的生活史及其对生存环境响应的研究, 在保护稀有濒危植物方面具有重要的科学及实际意义(
Western, 2001 ;
覃海宁等, 2017 ). ...
... ).这些植物是生物多样性保护的重要组成部分, 常具有地理分布狭窄、种群规模小、个体数量少以及生境严酷且脆弱等特点(
张文辉等, 2004 ;
Brown et al ., 2015 ;
覃海宁等, 2017 ); 此外, 坐果率、结籽率、种子萌发率和幼苗存活力低等生活史特征也是导致其濒危或灭绝的主要原因(
Neaves et al ., 2015 ).因此, 对该类群的生活史及其对生存环境响应的研究, 在保护稀有濒危植物方面具有重要的科学及实际意义(
Western, 2001 ;
覃海宁等, 2017 ). ...
... 喀什霸王是主要分布于我国塔克拉玛干沙漠西南部的多年生小灌木; 是生长于海拔400-2 100 m荒漠环境的稀有种及新疆二级保护植物(
尹林克, 2006 ).该物种在自然种群中呈分散式的片段化分布, 种群老龄化严重; 此外, 自然环境的年降水量少而温度较高.因此, 我们认为该区域的极端环境(高温及干旱胁迫)及该物种的休眠特点是抑制其种子萌发及种群更新的主要因素.为了证实该推测, 我们对该物种的自然坐果率、结籽率、种子休眠及萌发特性进行了室内比较研究, 为进一步探讨我国塔克拉玛干沙漠西南部荒漠植物在暖温带荒漠环境中的生活史特征及其适应机制研究提供理论资料. ...
... 喀什霸王为隶属于蒺藜科霸王属的小灌木, 主要分布于我国新疆南部(塔克拉玛干沙漠西南部)的荒漠中.该物种平均株高100 cm左右, 最高可达约150 cm, 分枝多, 株丛寿命20年以上.该物种主要以种子繁殖为主, 果实是由三个心皮组成的蒴果, 果翅由果皮发育而来, 每室有1个胚珠(
尹林克, 2006 ). ...
濒危植物秦岭冷杉地理分布和生物生态学特性研究
4
2004
... 生物多样性保护是有关生态系统安全和稳定性维护的关键科学问题(
Western, 2001 ;
Ren et al ., 2012 ;
覃海宁等, 2017 ).在全球变暖、自然灾害频繁发生、荒漠化严重及种群片段化等内外因素的影响下(
Parmesan & Yohe, 2003 ;
Leadley et al ., 2010 ), 生存在极端环境中的物种受到了不同程度的灭绝威胁(
Western, 2001 ;
张文辉等, 2004 ;
覃海宁等, 2017 ).我国植物多样性丰富, 目前有4 000-5 000种植物正处于不同程度的危机(稀有、濒危、极危)状态中(
Ren et al ., 2012 ).这些植物是生物多样性保护的重要组成部分, 常具有地理分布狭窄、种群规模小、个体数量少以及生境严酷且脆弱等特点(
张文辉等, 2004 ;
Brown et al ., 2015 ;
覃海宁等, 2017 ); 此外, 坐果率、结籽率、种子萌发率和幼苗存活力低等生活史特征也是导致其濒危或灭绝的主要原因(
Neaves et al ., 2015 ).因此, 对该类群的生活史及其对生存环境响应的研究, 在保护稀有濒危植物方面具有重要的科学及实际意义(
Western, 2001 ;
覃海宁等, 2017 ). ...
... ).这些植物是生物多样性保护的重要组成部分, 常具有地理分布狭窄、种群规模小、个体数量少以及生境严酷且脆弱等特点(
张文辉等, 2004 ;
Brown et al ., 2015 ;
覃海宁等, 2017 ); 此外, 坐果率、结籽率、种子萌发率和幼苗存活力低等生活史特征也是导致其濒危或灭绝的主要原因(
Neaves et al ., 2015 ).因此, 对该类群的生活史及其对生存环境响应的研究, 在保护稀有濒危植物方面具有重要的科学及实际意义(
Western, 2001 ;
覃海宁等, 2017 ). ...
... 喀什霸王是主要分布于我国塔克拉玛干沙漠西南部的多年生小灌木; 是生长于海拔400-2 100 m荒漠环境的稀有种及新疆二级保护植物(
尹林克, 2006 ).该物种在自然种群中呈分散式的片段化分布, 种群老龄化严重; 此外, 自然环境的年降水量少而温度较高.因此, 我们认为该区域的极端环境(高温及干旱胁迫)及该物种的休眠特点是抑制其种子萌发及种群更新的主要因素.为了证实该推测, 我们对该物种的自然坐果率、结籽率、种子休眠及萌发特性进行了室内比较研究, 为进一步探讨我国塔克拉玛干沙漠西南部荒漠植物在暖温带荒漠环境中的生活史特征及其适应机制研究提供理论资料. ...
... 喀什霸王为隶属于蒺藜科霸王属的小灌木, 主要分布于我国新疆南部(塔克拉玛干沙漠西南部)的荒漠中.该物种平均株高100 cm左右, 最高可达约150 cm, 分枝多, 株丛寿命20年以上.该物种主要以种子繁殖为主, 果实是由三个心皮组成的蒴果, 果翅由果皮发育而来, 每室有1个胚珠(
尹林克, 2006 ). ...
 ,1,2,*, 李玲
,1,2,*, 李玲
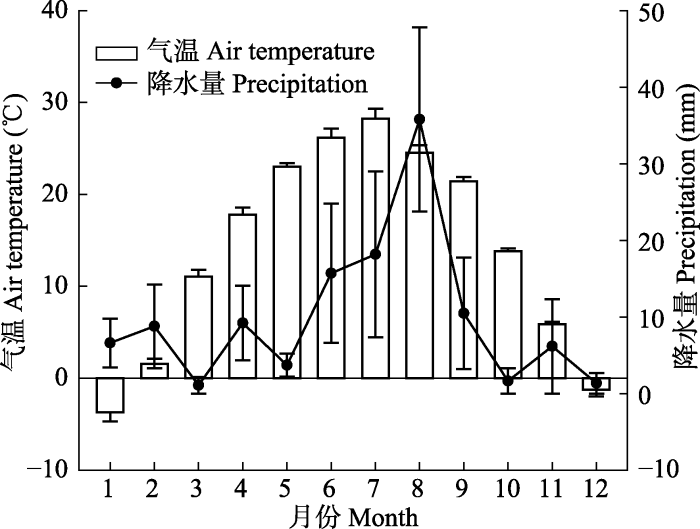 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT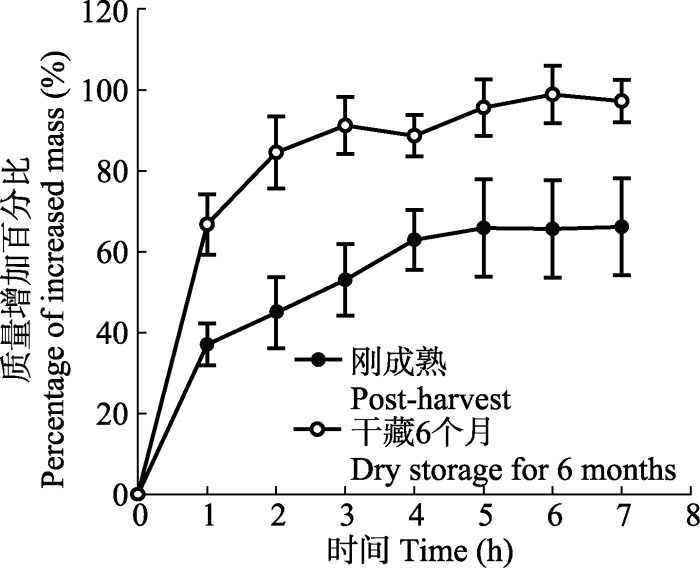 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT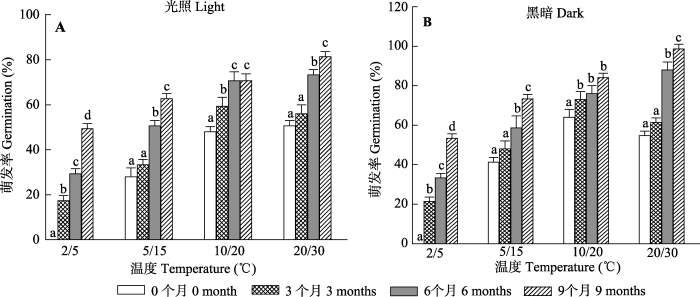 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT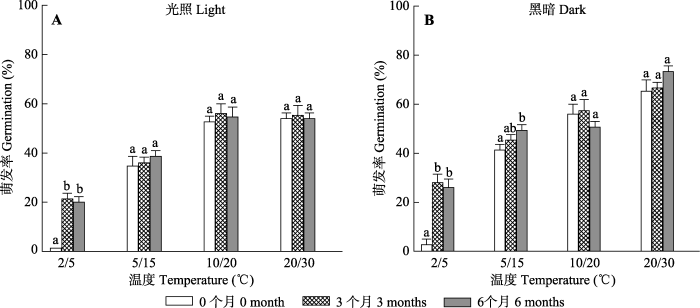 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT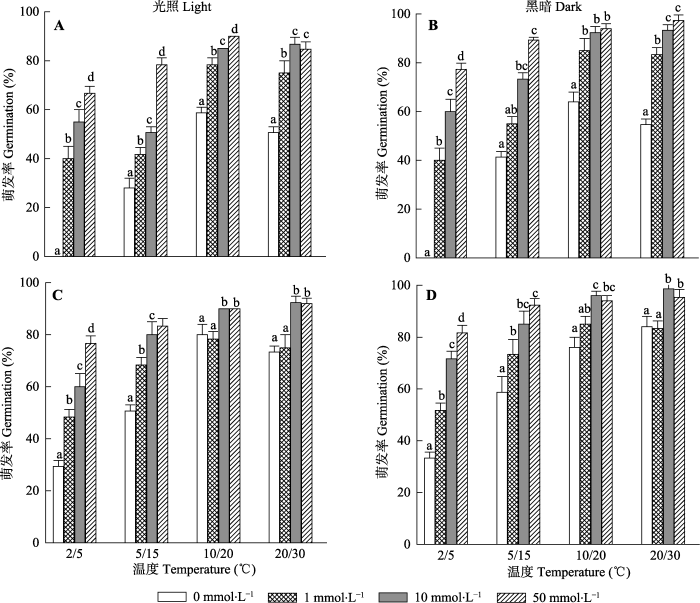 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT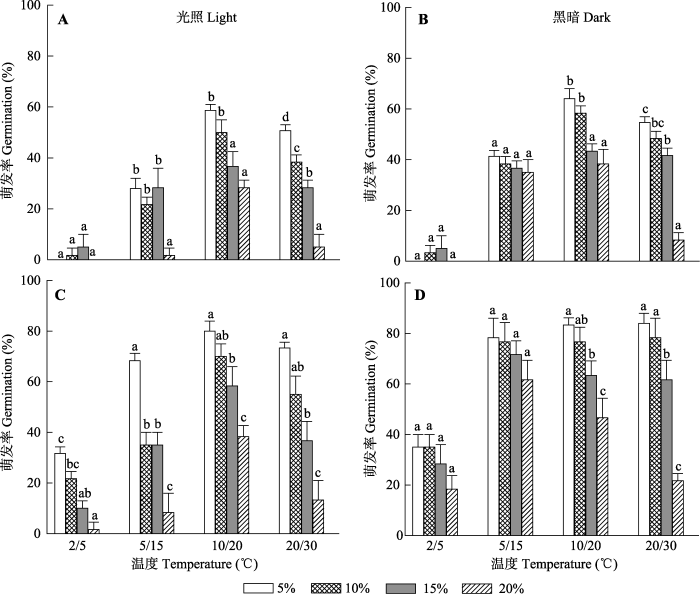 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}