Community assembly, diversity patterns and distributions of broad-leaved forests in North China
Jin-Shi XU1, Yong-Fu CHAI1, Xiao LIU1, Ming YUE,1,*, Yao-Xin GUO1, Mu-Yi KANG2,3, Quan-Ru LIU4, Cheng-Yang ZHENG5, Cheng-Jun JI5, Ming YAN6, Feng ZHANG7, Xian-Ming GAO8, Ren-Qing WANG9, Fu-Chen SHI10, Qin-Di ZHANG6, Mao WANG1,111Key Laboratory of Resource Biology and Biotechnology in Western China, Ministry of Education, Northwest University, Xi’an 710069, China 2State Key Laboratory of Earth Surface Processes and Resource Ecology, Beijing Normal University, Beijing 100875, China 3College of Resources Science & Technology, Faculty of Geographical Science, Beijing Normal University, Beijing 100875, China 4College of Life Sciences, Beijing Normal University, Beijing 100875, China 5Institute of Ecology, College of Urban and Environmental Sciences, Laboratory for Earth Surface Processes of the Ministry of Education, Peking University, Beijing 100871, China 6College of Life Sciences, Shanxi Normal University, Linfen, Shanxi 041000, China 7Institute of Loess Plateau, Shanxi University, Taiyuan 030006, China 8State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China 9School of Life Sciences, Shandong University, Jinan 250100, China 10College of Life Sciences, Nankai University, Tianjin 300071, China 11College of Grass Industry and Environmental Science, Xinjiang Agricultural University, ürümqi 830091, China
Supported by the National Basic Work of Science and Technology of China(2011FY110300) the China Postdoctoral Science Foundation(2018M643715) and the Natural Science Foundation of China(31700348)
Abstract Aims To understand the key processes driving the community assembly and diversity patterns in North China. Methods We investigated species composition of 87 plots from 29 sites. We applied phylogenetic approach, combined with community distribution information, to assess the community structure and diversity along environmental gradients. We then performed a variance partition to explore the relative importance of each environmental factor that influencing the patterns of community assembly and diversity process and a canonical correspondence analysis to analyze reason of community distributions. Important findings Similar communities showed similar habitat preferences, demonstrating that environments may shape species composition of the communities. The phylogenetic diversity showed a uni-modal pattern with the mean annual temperature (MAT), but increased with the mean annual precipitation (MAP), partly because of the strong disturbance in high-MAT regions. Temperature dominated the phylogenetic structure of the broad- leaved forests in North China. Environmental filtering dominate the community assembly processes in the areas with relatively low MAT. In addition, the effect of environment filtering increased with MAP. Keywords:community assembly;phylogenetic diversity;hydrothermal condition;species composition
PDF (1325KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 许金石, 柴永福, 刘晓, 岳明, 郭垚鑫, 康慕谊, 刘全儒, 郑成洋, 吉成均, 闫明, 张峰, 高贤明, 王仁卿, 石福臣, 张钦弟, 王茂. 华北区域环境梯度上阔叶林构建模式及分布成因. 植物生态学报, 2019, 43(9): 732-741. DOI: 10.17521/cjpe.2018.0183 XU Jin-Shi, CHAI Yong-Fu, LIU Xiao, YUE Ming, GUO Yao-Xin, KANG Mu-Yi, LIU Quan-Ru, ZHENG Cheng-Yang, JI Cheng-Jun, YAN Ming, ZHANG Feng, GAO Xian-Ming, WANG Ren-Qing, SHI Fu-Chen, ZHANG Qin-Di, WANG Mao. Community assembly, diversity patterns and distributions of broad-leaved forests in North China. Chinese Journal of Plant Ecology, 2019, 43(9): 732-741. DOI: 10.17521/cjpe.2018.0183
Table 2 表2 表2华北地区阔叶林环境因子对样方平均谱系距离(MPD)的方差分解结果 Table 2Result of variance decomposition of environment factors to mean phylogenetic distance (MPD) of broad-leaved forests in North China
环境因子 Factor
解释程度 Explanation (%)
坡度 Slope
3.200 3
坡向 Aspect
15.643 7
年平均气温 MAT
48.043 8
月平均气温极差 MMTD
16.433 6
年降水量 MAP
15.065 3
降水季节变化量 SVP
1.613 2
MAP, mean annual precipitation; MAT, mean annual temperature; MMTD, mean month temperature difference; SVP, seasonal variation of precipitation.
Fig. 2Pattern of the relationship between phylogenetic structure and mean annual temperature (MAT) of broad-leaved forests in North China. NRI, net relatedness index; NTI, nearest taxon index. The dotted line on 0 represented null expectation of phylogenetic structure.
Fig. 3Pattern of the relationship between phylogenetic structure and mean annual precipitation (MAP) of broad-leaved forests in North China. The dotted line on 0 represented null expectation of phylogenetic structure.
Fig. 4Pattern of the relationship between phylogenetic diversity (Faith’s PD) and mean annual temperature (MAT). Solid circles represent the value of Faithʼs PD. The solid curve represent regression curve of phylogenetic diversity.
Fig. 5Pattern of the relationship between phylogenetic diversity (Faith’s PD) and mean annual precipitation (MAP). Solid circles represent the value of Faithʼs PD. The solid line represent regression curve of phylogenetic diversity.
CaoK, RaoMD, YuJP, LiuXJ, MiXC, ChenJH ( 2013). The phylogenetic signal of functional traits and their effects on community structure in an evergreen broad-leaved forest Biodiversity Science, 21, 564-571. [本文引用: 1]
Cavender-BaresJ, KozakKH, FinePVA, KembelSW ( 2009). The merging of community ecology and phylogenetic biology Ecology Letters, 12, 693-715. [本文引用: 2]
ChaiYF, YueM, LiuX, GuoYX, WangM, XuJS, ZhangCG, ChenY, ZhangLX, ZhangRC ( 2016). Patterns of taxonomic, phylogenetic diversity during a long-term succession of forest on the Loess Plateau, China: Insights into assembly process Scientific Reports, 6, 27087. DOI: 10.1038/srep27087. [本文引用: 1]
ChaiYF, YueM ( 2016). Research advances in plant community assembly mechanisms Acta Ecologica Sinica, 36, 4557-4572. [本文引用: 1]
ChaseJM ( 2003). Community assembly: When should history matter? Oecologia, 136, 489-498. [本文引用: 2]
CheYD, LiuMX, LiLR, JiaoJ, XiaoW ( 2017). Exploring the community assembly of subalpine meadow communities based on functional traits and community phylogeny Chinese Journal of Plant Ecology, 41, 1157-1167. [本文引用: 1]
FangS, YuanZQ, LinF, YeJ, HaoZQ, WangXG ( 2014). Functional and phylogenetic structures of woody plants in broad-leaved Korean pine mixed forest in Changbai Mountains, Jilin, China Chinese Science Bulletin, 59, 2342-2348. [本文引用: 1]
FosterBL, DicksonTL, MurphyCA, KarelIS, SmithVH ( 2004). Propagule pools mediate community assembly and diversity—Ecosystem regulation along a grassland productivity gradient Journal of Ecology, 92, 435-449. [本文引用: 1]
GongGQ, HuangZL, HuangJX, YeWH, CaoHL, LianJY, LinGJ ( 2011). How individual species structure the community in Dinghushan 20 ha forest plot? Ecology and Environmental Sciences, 20, 991-995. [本文引用: 1]
GrimeJP ( 1998). Benefits of plant diversity to ecosystems: Immediate, filter and founder effects Journal of Ecology, 86, 902-910. [本文引用: 1]
HodkinsonID, CoulsonSJ, WebbNR ( 2003). Community assembly along proglacial chronosequences in the high Arctic: Vegetation and soil development in north-west Svalbard Journal of Ecology, 91, 651-663. [本文引用: 1]
HuangJX, ZhangJ, ShenY, LianJY, CaoHL, YeWH, WuLF, BinY ( 2014). Different relationships between temporal phylogenetic turnover and phylogenetic similarity and in two forests were detected by a new null model PLOS ONE, 9, e95703. DOI: 10.1371/journal.pone.0095703. [本文引用: 1]
HuangJX, ZhengFY, MiXC ( 2010). Influence of environmental factors on phylogenetic structure at multiple spatial scales in an evergreen broad-leaved forest of China Chinese Journal of Plant Ecology, 34, 309-315. [本文引用: 1]
HubbellSP ( 2001). The Unified Neutral Theory of Biodiversity and Biogeography. Princeton University Press, Princeton. 340-348. [本文引用: 1]
HulshofCM, ViolleC, SpasojevicMJ, McGillB, DamschenE, HarrisonS, EnquistBJ ( 2013). Intra-specific and inter-specific variation in specific leaf area reveal the importance of abiotic and biotic drivers of species diversity across elevation and latitude Journal of Vegetation Science, 24, 921-931. [本文引用: 1]
JiaP, DuGZ ( 2014). Measuring functional and phylogenetic diversity in community ecology Chinese Bulletin of Life Sciences, 26, 153-157. [本文引用: 1]
KellyCK, BowlerMG, PybusO, HarveyPH ( 2008). Phylogeny, niches, and relative abundance in natural communities Ecology, 89, 962-970. [本文引用: 1]
KraftNB, CornwellWK, WebbCO, AckerlyDD ( 2007). Trait evolution, community assembly, and the phylogenetic structure of ecological communities The American Naturalist, 170, 271-283. [本文引用: 2]
KylafisG, LoreauM ( 2011). Niche construction in the light of niche theory Ecology Letters, 14, 82-90. [本文引用: 1]
LessardJP, FordyceJA, GotelliNJ, SandersNJ ( 2009). Invasive ants alter the phylogenetic structure of ant communities Ecology, 90, 2664-2669. [本文引用: 1]
LuMM, HuangXC, CiXQ, YangGP, LiJ ( 2014). Phylogenetic community structure of subtropical forests along elevational gradients in Ailao Mountains of southwest China Biodiversity Science, 22, 438-448. [本文引用: 2]
MayfieldMM, LevineJM ( 2010). Opposing effects of competitive exclusion on the phylogenetic structure of communities Ecology Letters, 13, 1085-1093. [本文引用: 2]
MoriAS, ShionoT, KoideD, KitagawaR, OtaAT, MizumachiE ( 2013). Community assembly processes shape an altitudinal gradient of forest biodiversity Global Ecology and Biogeography, 22, 878-888. [本文引用: 2]
NiuHY, WangZF, LianJY, YeWH, ShenH ( 2011). New progress in community assembly: Community phylogenetic structure combining evolution and ecology Biodiversity Science, 19, 275-283. [本文引用: 1]
ParkDS, PotterD ( 2015). Why close relatives make bad neighbours: Phylogenetic conservatism in niche preferences and dispersal disproves Darwin’s naturalization hypothesis in the thistle tribe Molecular Ecology, 24, 3181-3193. [本文引用: 1]
PottierJ, DubuisA, PellissierL, MaioranoL, RossierL, RandinCF, VittozP, GuisanA ( 2013). The accuracy of plant assemblage prediction from species distribution models varies along environmental gradients Global Ecology and Biogeography, 22, 52-63. [本文引用: 1]
PurschkeO, SchmidBC, SykesMT, PoschlodP, MichalskiSG, DurkaW, KühnI, WinterM, PrenticeHC ( 2013). Contrasting changes in taxonomic, phylogenetic and functional diversity during a long-term succession: Insights into assembly processes Journal of Ecology, 101, 857-866. [本文引用: 1]
QianH, HaoZQ, ZhangJ ( 2014). Phylogenetic structure and phylogenetic diversity of angiosperm assemblages in forests along an elevational gradient in Changbaishan, China Journal of Plant Ecology, 7, 154-165. [本文引用: 1]
QiaoXJ, JabotF, TangZY, JiangMX, FangJY ( 2015). A latitudinal gradient in tree community assembly processes evidenced in Chinese forests Global Ecology and Biogeography, 24, 314-323. [本文引用: 2]
SwensonNG, EnquistBJ, ThompsonJ, ZimmermanJK ( 2007). The influence of spatial and size scale on phylogenetic relatedness in tropical forest communities Ecology, 88, 1770-1780. [本文引用: 1]
SwensonNG, EricksonDL, MiXC, BourgNA, Forero- Monta?aJ, GeXJ, HoweR, LakeJK, LiuXJ, MaKP, PeiNC, ThompsonJ, UriarteM, WolfA, WrightSJ, YeWH, ZhangJL, ZimmermanJK, KressWJ ( 2012). Phylogenetic and functional alpha and beta diversity in temperate and tropical tree communities Ecology, 93(Suppl.), S112-S125. [本文引用: 2]
TangZY, FangJY, ZhangL ( 2004). Patterns of woody plant species diversity along environmental gradients on Mt. Taibai, Qinling Mountains Biodiversity Science, 12, 115-122. [本文引用: 1]
TuckerCM, CadotteMW, CarvalhoSB, DaviesTJ, FerrierS, FritzSA, GrenyerR, HelmusMR, JinLS, MooersAO, PavoineS, PurschkeO, ReddingDW, RosauerDF, WinterM, MazelF ( 2017). A guide to phylogenetic metrics for conservation, community ecology and macroecology Biological Reviews, 92, 698-715. [本文引用: 1]
WebbCO ( 2000). Exploring the phylogenetic structure of ecological communities: An example for rain forest trees The American Naturalist, 156, 145-155. [本文引用: 1]
WebbCO, AckerlyDD, McPeekMA, DonoghueMJ ( 2002). Phylogenies and community ecology Annual Review of Ecology and Systematics, 33, 475-505. [本文引用: 1]
WebbCO, DonoghueMJ ( 2005). Phylomatic: Tree assembly for applied phylogenetics Molecular Ecology Notes, 5, 181-183. [本文引用: 1]
WehnckeEV, HubbellSP, FosterRB, DallingJW ( 2003). Seed dispersal patterns produced by white-faced monkeys: Implications for the dispersal limitation of neotropical tree species Journal of Ecology, 91, 677-685. [本文引用: 1]
WiensJJ, GrahamCH ( 2005). Niche conservatism: Integrating evolution, ecology, and conservation biology Annual Review of Ecology Evolution and Systematics, 36, 519-539. [本文引用: 1]
Wikstr?mN, SavolainenV, ChaseMW ( 2001). Evolution of the angiosperms: Calibrating the family tree The Proceedings of the Royal Society B, 268, 2211-2220. [本文引用: 1]
XuJS, ChaiYF, WangM, DangH, GuoYX, ChenY, ZhangCG, LiT, ZhangLX, YueM ( 2018). Shifts in plant community assembly processes across growth forms along a habitat severity gradient: A Test of the plant functional trait approach Frontiers in Plant Science, 9, 180. DOI: 10.3389/fpls.2018.00180. [本文引用: 3]
XuJS, ChenY, ZhangLX, ChaiYF, WangM, GuoYX, LiT, YueM ( 2017). Using phylogeny and functional traits for assessing community assembly along environmental gradients: A deterministic process driven by elevation Ecology and Evolution, 7, 5056-5069. [本文引用: 3]
YangJ, LuMM, CaoM, LiJ, LinLX ( 2014). Phylogenetic and functional alpha and beta diversity in mid-mountain humid evergreen broad-leaved forest Chinese Science Bulletin, 59, 2349-2358. [本文引用: 1]
YangYC, DaLJ ( 2006). A brief review of studies on differentiation of vegetation pattern along a topographic gradient in hilly regions Journal of Plant Ecology (Chinese Version), 30, 504-513. [本文引用: 1]
YuanX, MaKM, WangD ( 2011). Explaining the abundance-distribution relationship of plant species with niche breadth and position in the Yellow River Delta Acta Ecologica Sinica, 31, 1955-1961. [本文引用: 1]
ZhangH, GilbertB, ZhangXX, ZhouSR ( 2013). Community assembly along a successional gradient in sub-alpine meadows of the Qinghai-Tibetan Plateau, China Oikos, 122, 952-960. [本文引用: 1]
ZhaoMF, XueF, WangYH, WangGY, XingKX, KangMY, WangJL ( 2017). Phylogenetic structure and diversity of herbaceous communities in the conifer forests along an elevational gradient in Luya Mountain, Shanxi, China Chinese Journal of Plant Ecology, 41, 707-715. [本文引用: 1]
ZhouXG, LuWK, YeD, WenYG . ( 2014). Assembly mechanism of forest community based on phylogeny and functional traits Guangxi Sciences, 21, 525-533. [本文引用: 1]
... 群落内物种的多度反映了该物种在群落中的相对重要性, 代表了该物种占有资源的能力(Grime, 1998; 曹科等, 2013).基于物种组成数据, 可以探究相似物种组成的群落的分布模式.群落内的生物多样性则与多种环境因子有关(Qian et al., 2014; 车应弟等, 2017).通过多样性分析, 可以探究群落内物种形成、共存及时空变化规律(Swenson et al., 2012).因此, 研究环境因子与群落物种组成及多样性之间的关系, 有助于预测多样性的动态(Yang & Da, 2006; 周晓果等, 2014), 进而理解群落构建的过程. ...
古田山木本植物功能性状的系统发育信号及其对群落结构的影响 1 2013
... 群落内物种的多度反映了该物种在群落中的相对重要性, 代表了该物种占有资源的能力(Grime, 1998; 曹科等, 2013).基于物种组成数据, 可以探究相似物种组成的群落的分布模式.群落内的生物多样性则与多种环境因子有关(Qian et al., 2014; 车应弟等, 2017).通过多样性分析, 可以探究群落内物种形成、共存及时空变化规律(Swenson et al., 2012).因此, 研究环境因子与群落物种组成及多样性之间的关系, 有助于预测多样性的动态(Yang & Da, 2006; 周晓果等, 2014), 进而理解群落构建的过程. ...
The merging of community ecology and phylogenetic biology 2 2009
... 群落内的物种组成是进化过程与生态过程共同作用的结果(黄建雄等, 2010), 以群落内物种的亲缘关系即谱系为载体进行群落构建研究, 能够较好地从进化角度反映群落构建过程中的生态学机制(Park & Potter, 2015).基于系统发育的谱系多样性考虑了不同生物类群间的谱系关系及进化历史等信息, 能够较好地从进化角度反映群落的多样性模式(Cavender-Bares et al., 2009; 杨洁等, 2014), 并揭示多样性维持机制(即群落构建过程)(Purschke et al., 2013; 贾鹏和杜国祯, 2014).群落内物种可能因环境过滤或竞争排除而使谱系聚集或发散, 从而表现出特定的谱系结构(Webb, 2000; Swenson et al., 2012).例如, 环境过滤作用主导群落构建时, 筛选出的群落内物种亲缘关系可能较近, 从而适应当前生境(卢孟孟等, 2014), 其谱系结构表现为聚集; 竞争排除作用主导群落构建时, 亲缘关系较近的物种无法占用相似的生态位(Wiens & Graham, 2005), 从而排除部分近缘种, 其谱系结构表现为发散; 扩散限制作用主导群落构建时, 群落的形成与环境因子及生物间相互作用无关(Hubbell, 2001), 其谱系结构表现为随机(柴永福和岳明, 2016).先前的许多研究通过构建零模型来判断谱系结构的聚集与发散(牛红玉等, 2011), 从而证明环境过滤、竞争排除或扩散限制的存在, 阐明不同作用在群落构建过程中的相对重要性(Foster et al., 2004; Cavender-Bares et al., 2009; Mayfield & Levine, 2010; 姜晓燕等, 2016).基于此方法, 先前的研究认为, 环境因子造成的环境过滤往往主导了构建过程(Huang et al., 2014), 尤其在生境条件比较苛刻的地区, 如高海拔区域(Mori et al., 2013; Xu et al., 2017; 赵鸣飞等, 2017).然而, 海拔毕竟是一个间接化的生境因子, 其与温度、降水、紫外辐射强度、风速、土壤养分等诸多因子相关(Xu et al., 2018).探讨单一的水热梯度上群落构建的变化模式, 能够更清晰地阐明群落与水热因子的关系, 从而明确区域尺度上群落的构建模式.然而, 这方面的研究却鲜有报道. ...
... ; Cavender-Bares et al., 2009; Mayfield & Levine, 2010; 姜晓燕等, 2016).基于此方法, 先前的研究认为, 环境因子造成的环境过滤往往主导了构建过程(Huang et al., 2014), 尤其在生境条件比较苛刻的地区, 如高海拔区域(Mori et al., 2013; Xu et al., 2017; 赵鸣飞等, 2017).然而, 海拔毕竟是一个间接化的生境因子, 其与温度、降水、紫外辐射强度、风速、土壤养分等诸多因子相关(Xu et al., 2018).探讨单一的水热梯度上群落构建的变化模式, 能够更清晰地阐明群落与水热因子的关系, 从而明确区域尺度上群落的构建模式.然而, 这方面的研究却鲜有报道. ...
Patterns of taxonomic, phylogenetic diversity during a long-term succession of forest on the Loess Plateau, China: Insights into assembly process 1 2016
Propagule pools mediate community assembly and diversity—Ecosystem regulation along a grassland productivity gradient 1 2004
... 群落内的物种组成是进化过程与生态过程共同作用的结果(黄建雄等, 2010), 以群落内物种的亲缘关系即谱系为载体进行群落构建研究, 能够较好地从进化角度反映群落构建过程中的生态学机制(Park & Potter, 2015).基于系统发育的谱系多样性考虑了不同生物类群间的谱系关系及进化历史等信息, 能够较好地从进化角度反映群落的多样性模式(Cavender-Bares et al., 2009; 杨洁等, 2014), 并揭示多样性维持机制(即群落构建过程)(Purschke et al., 2013; 贾鹏和杜国祯, 2014).群落内物种可能因环境过滤或竞争排除而使谱系聚集或发散, 从而表现出特定的谱系结构(Webb, 2000; Swenson et al., 2012).例如, 环境过滤作用主导群落构建时, 筛选出的群落内物种亲缘关系可能较近, 从而适应当前生境(卢孟孟等, 2014), 其谱系结构表现为聚集; 竞争排除作用主导群落构建时, 亲缘关系较近的物种无法占用相似的生态位(Wiens & Graham, 2005), 从而排除部分近缘种, 其谱系结构表现为发散; 扩散限制作用主导群落构建时, 群落的形成与环境因子及生物间相互作用无关(Hubbell, 2001), 其谱系结构表现为随机(柴永福和岳明, 2016).先前的许多研究通过构建零模型来判断谱系结构的聚集与发散(牛红玉等, 2011), 从而证明环境过滤、竞争排除或扩散限制的存在, 阐明不同作用在群落构建过程中的相对重要性(Foster et al., 2004; Cavender-Bares et al., 2009; Mayfield & Levine, 2010; 姜晓燕等, 2016).基于此方法, 先前的研究认为, 环境因子造成的环境过滤往往主导了构建过程(Huang et al., 2014), 尤其在生境条件比较苛刻的地区, 如高海拔区域(Mori et al., 2013; Xu et al., 2017; 赵鸣飞等, 2017).然而, 海拔毕竟是一个间接化的生境因子, 其与温度、降水、紫外辐射强度、风速、土壤养分等诸多因子相关(Xu et al., 2018).探讨单一的水热梯度上群落构建的变化模式, 能够更清晰地阐明群落与水热因子的关系, 从而明确区域尺度上群落的构建模式.然而, 这方面的研究却鲜有报道. ...
鼎湖山20公顷森林样地单个物种对群落的构建 1 2011
... 群落构建即群落物种多样性的维持机制, 一直是群落生态学的研究热点(Chase, 2003).然而, 群落构建的形式尚存在争论(宫贵权等, 2011).目前, 基于确定性过程的生态位理论和基于随机过程的中性理论是最常用来检验群落构建模式的两个假说(Kelly et al., 2008; Kylafis & Loreau, 2011).由于群落构建过程中往往受到多种因子控制, 基于生态位理论的环境过滤、竞争排除(Webb et al., 2002)与基于中性理论的扩散限制(Wehncke et al., 2010)等作用可能同时影响着群落构建. ...
鼎湖山20公顷森林样地单个物种对群落的构建 1 2011
... 群落构建即群落物种多样性的维持机制, 一直是群落生态学的研究热点(Chase, 2003).然而, 群落构建的形式尚存在争论(宫贵权等, 2011).目前, 基于确定性过程的生态位理论和基于随机过程的中性理论是最常用来检验群落构建模式的两个假说(Kelly et al., 2008; Kylafis & Loreau, 2011).由于群落构建过程中往往受到多种因子控制, 基于生态位理论的环境过滤、竞争排除(Webb et al., 2002)与基于中性理论的扩散限制(Wehncke et al., 2010)等作用可能同时影响着群落构建. ...
Benefits of plant diversity to ecosystems: Immediate, filter and founder effects 1 1998
... 群落内物种的多度反映了该物种在群落中的相对重要性, 代表了该物种占有资源的能力(Grime, 1998; 曹科等, 2013).基于物种组成数据, 可以探究相似物种组成的群落的分布模式.群落内的生物多样性则与多种环境因子有关(Qian et al., 2014; 车应弟等, 2017).通过多样性分析, 可以探究群落内物种形成、共存及时空变化规律(Swenson et al., 2012).因此, 研究环境因子与群落物种组成及多样性之间的关系, 有助于预测多样性的动态(Yang & Da, 2006; 周晓果等, 2014), 进而理解群落构建的过程. ...
Community assembly along proglacial chronosequences in the high Arctic: Vegetation and soil development in north-west Svalbard 1 2003
... 单纯从水热梯度上进行群落构建研究的报道较少, 然而, 有大量的基于经纬度、海拔等地理因子梯度研究群落构建模式的报道.由于海拔与经纬度等信息中均包含了水热因素, 因此本研究可与这些研究做类比.例如, 大部分研究发现, 高海拔(意味着较低的温度, 由于凝冻, 也有较低的水分可用性)区域主要受到环境过滤的作用, 而低海拔(意味着较高的温度)区域则可以表现出中性或竞争排除的作用(Hulshof et al., 2013; Mori et al., 2013; Pottier et al., 2013; Xu et al., 2017); 而高纬度(意味着较低的温度、较少的降水)区域的构建模式应该是聚集的(Hodkinson et al., 2003).然而, 也有部分研究得出了不一致的结论, 例如, Zhang等(2013)发现, 亚高山草甸的群落构建模式与海拔等非生物因素无关, 而卢孟孟等(2014)发现, 哀牢山干热河谷的群落的谱系结构在高海拔区域趋于发散, 即在温度较低、水分可用性也较低的区域并未受到环境过滤的 作用. ...
Different relationships between temporal phylogenetic turnover and phylogenetic similarity and in two forests were detected by a new null model 1 2014
... 群落内的物种组成是进化过程与生态过程共同作用的结果(黄建雄等, 2010), 以群落内物种的亲缘关系即谱系为载体进行群落构建研究, 能够较好地从进化角度反映群落构建过程中的生态学机制(Park & Potter, 2015).基于系统发育的谱系多样性考虑了不同生物类群间的谱系关系及进化历史等信息, 能够较好地从进化角度反映群落的多样性模式(Cavender-Bares et al., 2009; 杨洁等, 2014), 并揭示多样性维持机制(即群落构建过程)(Purschke et al., 2013; 贾鹏和杜国祯, 2014).群落内物种可能因环境过滤或竞争排除而使谱系聚集或发散, 从而表现出特定的谱系结构(Webb, 2000; Swenson et al., 2012).例如, 环境过滤作用主导群落构建时, 筛选出的群落内物种亲缘关系可能较近, 从而适应当前生境(卢孟孟等, 2014), 其谱系结构表现为聚集; 竞争排除作用主导群落构建时, 亲缘关系较近的物种无法占用相似的生态位(Wiens & Graham, 2005), 从而排除部分近缘种, 其谱系结构表现为发散; 扩散限制作用主导群落构建时, 群落的形成与环境因子及生物间相互作用无关(Hubbell, 2001), 其谱系结构表现为随机(柴永福和岳明, 2016).先前的许多研究通过构建零模型来判断谱系结构的聚集与发散(牛红玉等, 2011), 从而证明环境过滤、竞争排除或扩散限制的存在, 阐明不同作用在群落构建过程中的相对重要性(Foster et al., 2004; Cavender-Bares et al., 2009; Mayfield & Levine, 2010; 姜晓燕等, 2016).基于此方法, 先前的研究认为, 环境因子造成的环境过滤往往主导了构建过程(Huang et al., 2014), 尤其在生境条件比较苛刻的地区, 如高海拔区域(Mori et al., 2013; Xu et al., 2017; 赵鸣飞等, 2017).然而, 海拔毕竟是一个间接化的生境因子, 其与温度、降水、紫外辐射强度、风速、土壤养分等诸多因子相关(Xu et al., 2018).探讨单一的水热梯度上群落构建的变化模式, 能够更清晰地阐明群落与水热因子的关系, 从而明确区域尺度上群落的构建模式.然而, 这方面的研究却鲜有报道. ...
不同尺度上环境因子对常绿阔叶林群落的谱系结构的影响 1 2010
... 群落内的物种组成是进化过程与生态过程共同作用的结果(黄建雄等, 2010), 以群落内物种的亲缘关系即谱系为载体进行群落构建研究, 能够较好地从进化角度反映群落构建过程中的生态学机制(Park & Potter, 2015).基于系统发育的谱系多样性考虑了不同生物类群间的谱系关系及进化历史等信息, 能够较好地从进化角度反映群落的多样性模式(Cavender-Bares et al., 2009; 杨洁等, 2014), 并揭示多样性维持机制(即群落构建过程)(Purschke et al., 2013; 贾鹏和杜国祯, 2014).群落内物种可能因环境过滤或竞争排除而使谱系聚集或发散, 从而表现出特定的谱系结构(Webb, 2000; Swenson et al., 2012).例如, 环境过滤作用主导群落构建时, 筛选出的群落内物种亲缘关系可能较近, 从而适应当前生境(卢孟孟等, 2014), 其谱系结构表现为聚集; 竞争排除作用主导群落构建时, 亲缘关系较近的物种无法占用相似的生态位(Wiens & Graham, 2005), 从而排除部分近缘种, 其谱系结构表现为发散; 扩散限制作用主导群落构建时, 群落的形成与环境因子及生物间相互作用无关(Hubbell, 2001), 其谱系结构表现为随机(柴永福和岳明, 2016).先前的许多研究通过构建零模型来判断谱系结构的聚集与发散(牛红玉等, 2011), 从而证明环境过滤、竞争排除或扩散限制的存在, 阐明不同作用在群落构建过程中的相对重要性(Foster et al., 2004; Cavender-Bares et al., 2009; Mayfield & Levine, 2010; 姜晓燕等, 2016).基于此方法, 先前的研究认为, 环境因子造成的环境过滤往往主导了构建过程(Huang et al., 2014), 尤其在生境条件比较苛刻的地区, 如高海拔区域(Mori et al., 2013; Xu et al., 2017; 赵鸣飞等, 2017).然而, 海拔毕竟是一个间接化的生境因子, 其与温度、降水、紫外辐射强度、风速、土壤养分等诸多因子相关(Xu et al., 2018).探讨单一的水热梯度上群落构建的变化模式, 能够更清晰地阐明群落与水热因子的关系, 从而明确区域尺度上群落的构建模式.然而, 这方面的研究却鲜有报道. ...
不同尺度上环境因子对常绿阔叶林群落的谱系结构的影响 1 2010
... 群落内的物种组成是进化过程与生态过程共同作用的结果(黄建雄等, 2010), 以群落内物种的亲缘关系即谱系为载体进行群落构建研究, 能够较好地从进化角度反映群落构建过程中的生态学机制(Park & Potter, 2015).基于系统发育的谱系多样性考虑了不同生物类群间的谱系关系及进化历史等信息, 能够较好地从进化角度反映群落的多样性模式(Cavender-Bares et al., 2009; 杨洁等, 2014), 并揭示多样性维持机制(即群落构建过程)(Purschke et al., 2013; 贾鹏和杜国祯, 2014).群落内物种可能因环境过滤或竞争排除而使谱系聚集或发散, 从而表现出特定的谱系结构(Webb, 2000; Swenson et al., 2012).例如, 环境过滤作用主导群落构建时, 筛选出的群落内物种亲缘关系可能较近, 从而适应当前生境(卢孟孟等, 2014), 其谱系结构表现为聚集; 竞争排除作用主导群落构建时, 亲缘关系较近的物种无法占用相似的生态位(Wiens & Graham, 2005), 从而排除部分近缘种, 其谱系结构表现为发散; 扩散限制作用主导群落构建时, 群落的形成与环境因子及生物间相互作用无关(Hubbell, 2001), 其谱系结构表现为随机(柴永福和岳明, 2016).先前的许多研究通过构建零模型来判断谱系结构的聚集与发散(牛红玉等, 2011), 从而证明环境过滤、竞争排除或扩散限制的存在, 阐明不同作用在群落构建过程中的相对重要性(Foster et al., 2004; Cavender-Bares et al., 2009; Mayfield & Levine, 2010; 姜晓燕等, 2016).基于此方法, 先前的研究认为, 环境因子造成的环境过滤往往主导了构建过程(Huang et al., 2014), 尤其在生境条件比较苛刻的地区, 如高海拔区域(Mori et al., 2013; Xu et al., 2017; 赵鸣飞等, 2017).然而, 海拔毕竟是一个间接化的生境因子, 其与温度、降水、紫外辐射强度、风速、土壤养分等诸多因子相关(Xu et al., 2018).探讨单一的水热梯度上群落构建的变化模式, 能够更清晰地阐明群落与水热因子的关系, 从而明确区域尺度上群落的构建模式.然而, 这方面的研究却鲜有报道. ...
1 2001
... 群落内的物种组成是进化过程与生态过程共同作用的结果(黄建雄等, 2010), 以群落内物种的亲缘关系即谱系为载体进行群落构建研究, 能够较好地从进化角度反映群落构建过程中的生态学机制(Park & Potter, 2015).基于系统发育的谱系多样性考虑了不同生物类群间的谱系关系及进化历史等信息, 能够较好地从进化角度反映群落的多样性模式(Cavender-Bares et al., 2009; 杨洁等, 2014), 并揭示多样性维持机制(即群落构建过程)(Purschke et al., 2013; 贾鹏和杜国祯, 2014).群落内物种可能因环境过滤或竞争排除而使谱系聚集或发散, 从而表现出特定的谱系结构(Webb, 2000; Swenson et al., 2012).例如, 环境过滤作用主导群落构建时, 筛选出的群落内物种亲缘关系可能较近, 从而适应当前生境(卢孟孟等, 2014), 其谱系结构表现为聚集; 竞争排除作用主导群落构建时, 亲缘关系较近的物种无法占用相似的生态位(Wiens & Graham, 2005), 从而排除部分近缘种, 其谱系结构表现为发散; 扩散限制作用主导群落构建时, 群落的形成与环境因子及生物间相互作用无关(Hubbell, 2001), 其谱系结构表现为随机(柴永福和岳明, 2016).先前的许多研究通过构建零模型来判断谱系结构的聚集与发散(牛红玉等, 2011), 从而证明环境过滤、竞争排除或扩散限制的存在, 阐明不同作用在群落构建过程中的相对重要性(Foster et al., 2004; Cavender-Bares et al., 2009; Mayfield & Levine, 2010; 姜晓燕等, 2016).基于此方法, 先前的研究认为, 环境因子造成的环境过滤往往主导了构建过程(Huang et al., 2014), 尤其在生境条件比较苛刻的地区, 如高海拔区域(Mori et al., 2013; Xu et al., 2017; 赵鸣飞等, 2017).然而, 海拔毕竟是一个间接化的生境因子, 其与温度、降水、紫外辐射强度、风速、土壤养分等诸多因子相关(Xu et al., 2018).探讨单一的水热梯度上群落构建的变化模式, 能够更清晰地阐明群落与水热因子的关系, 从而明确区域尺度上群落的构建模式.然而, 这方面的研究却鲜有报道. ...
Intra-specific and inter-specific variation in specific leaf area reveal the importance of abiotic and biotic drivers of species diversity across elevation and latitude 1 2013
... 单纯从水热梯度上进行群落构建研究的报道较少, 然而, 有大量的基于经纬度、海拔等地理因子梯度研究群落构建模式的报道.由于海拔与经纬度等信息中均包含了水热因素, 因此本研究可与这些研究做类比.例如, 大部分研究发现, 高海拔(意味着较低的温度, 由于凝冻, 也有较低的水分可用性)区域主要受到环境过滤的作用, 而低海拔(意味着较高的温度)区域则可以表现出中性或竞争排除的作用(Hulshof et al., 2013; Mori et al., 2013; Pottier et al., 2013; Xu et al., 2017); 而高纬度(意味着较低的温度、较少的降水)区域的构建模式应该是聚集的(Hodkinson et al., 2003).然而, 也有部分研究得出了不一致的结论, 例如, Zhang等(2013)发现, 亚高山草甸的群落构建模式与海拔等非生物因素无关, 而卢孟孟等(2014)发现, 哀牢山干热河谷的群落的谱系结构在高海拔区域趋于发散, 即在温度较低、水分可用性也较低的区域并未受到环境过滤的 作用. ...
生态学的多样性指数: 功能与系统发育 1 2014
... 群落内的物种组成是进化过程与生态过程共同作用的结果(黄建雄等, 2010), 以群落内物种的亲缘关系即谱系为载体进行群落构建研究, 能够较好地从进化角度反映群落构建过程中的生态学机制(Park & Potter, 2015).基于系统发育的谱系多样性考虑了不同生物类群间的谱系关系及进化历史等信息, 能够较好地从进化角度反映群落的多样性模式(Cavender-Bares et al., 2009; 杨洁等, 2014), 并揭示多样性维持机制(即群落构建过程)(Purschke et al., 2013; 贾鹏和杜国祯, 2014).群落内物种可能因环境过滤或竞争排除而使谱系聚集或发散, 从而表现出特定的谱系结构(Webb, 2000; Swenson et al., 2012).例如, 环境过滤作用主导群落构建时, 筛选出的群落内物种亲缘关系可能较近, 从而适应当前生境(卢孟孟等, 2014), 其谱系结构表现为聚集; 竞争排除作用主导群落构建时, 亲缘关系较近的物种无法占用相似的生态位(Wiens & Graham, 2005), 从而排除部分近缘种, 其谱系结构表现为发散; 扩散限制作用主导群落构建时, 群落的形成与环境因子及生物间相互作用无关(Hubbell, 2001), 其谱系结构表现为随机(柴永福和岳明, 2016).先前的许多研究通过构建零模型来判断谱系结构的聚集与发散(牛红玉等, 2011), 从而证明环境过滤、竞争排除或扩散限制的存在, 阐明不同作用在群落构建过程中的相对重要性(Foster et al., 2004; Cavender-Bares et al., 2009; Mayfield & Levine, 2010; 姜晓燕等, 2016).基于此方法, 先前的研究认为, 环境因子造成的环境过滤往往主导了构建过程(Huang et al., 2014), 尤其在生境条件比较苛刻的地区, 如高海拔区域(Mori et al., 2013; Xu et al., 2017; 赵鸣飞等, 2017).然而, 海拔毕竟是一个间接化的生境因子, 其与温度、降水、紫外辐射强度、风速、土壤养分等诸多因子相关(Xu et al., 2018).探讨单一的水热梯度上群落构建的变化模式, 能够更清晰地阐明群落与水热因子的关系, 从而明确区域尺度上群落的构建模式.然而, 这方面的研究却鲜有报道. ...
生态学的多样性指数: 功能与系统发育 1 2014
... 群落内的物种组成是进化过程与生态过程共同作用的结果(黄建雄等, 2010), 以群落内物种的亲缘关系即谱系为载体进行群落构建研究, 能够较好地从进化角度反映群落构建过程中的生态学机制(Park & Potter, 2015).基于系统发育的谱系多样性考虑了不同生物类群间的谱系关系及进化历史等信息, 能够较好地从进化角度反映群落的多样性模式(Cavender-Bares et al., 2009; 杨洁等, 2014), 并揭示多样性维持机制(即群落构建过程)(Purschke et al., 2013; 贾鹏和杜国祯, 2014).群落内物种可能因环境过滤或竞争排除而使谱系聚集或发散, 从而表现出特定的谱系结构(Webb, 2000; Swenson et al., 2012).例如, 环境过滤作用主导群落构建时, 筛选出的群落内物种亲缘关系可能较近, 从而适应当前生境(卢孟孟等, 2014), 其谱系结构表现为聚集; 竞争排除作用主导群落构建时, 亲缘关系较近的物种无法占用相似的生态位(Wiens & Graham, 2005), 从而排除部分近缘种, 其谱系结构表现为发散; 扩散限制作用主导群落构建时, 群落的形成与环境因子及生物间相互作用无关(Hubbell, 2001), 其谱系结构表现为随机(柴永福和岳明, 2016).先前的许多研究通过构建零模型来判断谱系结构的聚集与发散(牛红玉等, 2011), 从而证明环境过滤、竞争排除或扩散限制的存在, 阐明不同作用在群落构建过程中的相对重要性(Foster et al., 2004; Cavender-Bares et al., 2009; Mayfield & Levine, 2010; 姜晓燕等, 2016).基于此方法, 先前的研究认为, 环境因子造成的环境过滤往往主导了构建过程(Huang et al., 2014), 尤其在生境条件比较苛刻的地区, 如高海拔区域(Mori et al., 2013; Xu et al., 2017; 赵鸣飞等, 2017).然而, 海拔毕竟是一个间接化的生境因子, 其与温度、降水、紫外辐射强度、风速、土壤养分等诸多因子相关(Xu et al., 2018).探讨单一的水热梯度上群落构建的变化模式, 能够更清晰地阐明群落与水热因子的关系, 从而明确区域尺度上群落的构建模式.然而, 这方面的研究却鲜有报道. ...
山西霍山植物群落谱系结构的空间格局 1 2016
... 群落内的物种组成是进化过程与生态过程共同作用的结果(黄建雄等, 2010), 以群落内物种的亲缘关系即谱系为载体进行群落构建研究, 能够较好地从进化角度反映群落构建过程中的生态学机制(Park & Potter, 2015).基于系统发育的谱系多样性考虑了不同生物类群间的谱系关系及进化历史等信息, 能够较好地从进化角度反映群落的多样性模式(Cavender-Bares et al., 2009; 杨洁等, 2014), 并揭示多样性维持机制(即群落构建过程)(Purschke et al., 2013; 贾鹏和杜国祯, 2014).群落内物种可能因环境过滤或竞争排除而使谱系聚集或发散, 从而表现出特定的谱系结构(Webb, 2000; Swenson et al., 2012).例如, 环境过滤作用主导群落构建时, 筛选出的群落内物种亲缘关系可能较近, 从而适应当前生境(卢孟孟等, 2014), 其谱系结构表现为聚集; 竞争排除作用主导群落构建时, 亲缘关系较近的物种无法占用相似的生态位(Wiens & Graham, 2005), 从而排除部分近缘种, 其谱系结构表现为发散; 扩散限制作用主导群落构建时, 群落的形成与环境因子及生物间相互作用无关(Hubbell, 2001), 其谱系结构表现为随机(柴永福和岳明, 2016).先前的许多研究通过构建零模型来判断谱系结构的聚集与发散(牛红玉等, 2011), 从而证明环境过滤、竞争排除或扩散限制的存在, 阐明不同作用在群落构建过程中的相对重要性(Foster et al., 2004; Cavender-Bares et al., 2009; Mayfield & Levine, 2010; 姜晓燕等, 2016).基于此方法, 先前的研究认为, 环境因子造成的环境过滤往往主导了构建过程(Huang et al., 2014), 尤其在生境条件比较苛刻的地区, 如高海拔区域(Mori et al., 2013; Xu et al., 2017; 赵鸣飞等, 2017).然而, 海拔毕竟是一个间接化的生境因子, 其与温度、降水、紫外辐射强度、风速、土壤养分等诸多因子相关(Xu et al., 2018).探讨单一的水热梯度上群落构建的变化模式, 能够更清晰地阐明群落与水热因子的关系, 从而明确区域尺度上群落的构建模式.然而, 这方面的研究却鲜有报道. ...
山西霍山植物群落谱系结构的空间格局 1 2016
... 群落内的物种组成是进化过程与生态过程共同作用的结果(黄建雄等, 2010), 以群落内物种的亲缘关系即谱系为载体进行群落构建研究, 能够较好地从进化角度反映群落构建过程中的生态学机制(Park & Potter, 2015).基于系统发育的谱系多样性考虑了不同生物类群间的谱系关系及进化历史等信息, 能够较好地从进化角度反映群落的多样性模式(Cavender-Bares et al., 2009; 杨洁等, 2014), 并揭示多样性维持机制(即群落构建过程)(Purschke et al., 2013; 贾鹏和杜国祯, 2014).群落内物种可能因环境过滤或竞争排除而使谱系聚集或发散, 从而表现出特定的谱系结构(Webb, 2000; Swenson et al., 2012).例如, 环境过滤作用主导群落构建时, 筛选出的群落内物种亲缘关系可能较近, 从而适应当前生境(卢孟孟等, 2014), 其谱系结构表现为聚集; 竞争排除作用主导群落构建时, 亲缘关系较近的物种无法占用相似的生态位(Wiens & Graham, 2005), 从而排除部分近缘种, 其谱系结构表现为发散; 扩散限制作用主导群落构建时, 群落的形成与环境因子及生物间相互作用无关(Hubbell, 2001), 其谱系结构表现为随机(柴永福和岳明, 2016).先前的许多研究通过构建零模型来判断谱系结构的聚集与发散(牛红玉等, 2011), 从而证明环境过滤、竞争排除或扩散限制的存在, 阐明不同作用在群落构建过程中的相对重要性(Foster et al., 2004; Cavender-Bares et al., 2009; Mayfield & Levine, 2010; 姜晓燕等, 2016).基于此方法, 先前的研究认为, 环境因子造成的环境过滤往往主导了构建过程(Huang et al., 2014), 尤其在生境条件比较苛刻的地区, 如高海拔区域(Mori et al., 2013; Xu et al., 2017; 赵鸣飞等, 2017).然而, 海拔毕竟是一个间接化的生境因子, 其与温度、降水、紫外辐射强度、风速、土壤养分等诸多因子相关(Xu et al., 2018).探讨单一的水热梯度上群落构建的变化模式, 能够更清晰地阐明群落与水热因子的关系, 从而明确区域尺度上群落的构建模式.然而, 这方面的研究却鲜有报道. ...
Phylogeny, niches, and relative abundance in natural communities 1 2008
... 群落构建即群落物种多样性的维持机制, 一直是群落生态学的研究热点(Chase, 2003).然而, 群落构建的形式尚存在争论(宫贵权等, 2011).目前, 基于确定性过程的生态位理论和基于随机过程的中性理论是最常用来检验群落构建模式的两个假说(Kelly et al., 2008; Kylafis & Loreau, 2011).由于群落构建过程中往往受到多种因子控制, 基于生态位理论的环境过滤、竞争排除(Webb et al., 2002)与基于中性理论的扩散限制(Wehncke et al., 2010)等作用可能同时影响着群落构建. ...
Trait evolution, community assembly, and the phylogenetic structure of ecological communities 2 2007
Community assembly processes shape an altitudinal gradient of forest biodiversity 2 2013
... 群落内的物种组成是进化过程与生态过程共同作用的结果(黄建雄等, 2010), 以群落内物种的亲缘关系即谱系为载体进行群落构建研究, 能够较好地从进化角度反映群落构建过程中的生态学机制(Park & Potter, 2015).基于系统发育的谱系多样性考虑了不同生物类群间的谱系关系及进化历史等信息, 能够较好地从进化角度反映群落的多样性模式(Cavender-Bares et al., 2009; 杨洁等, 2014), 并揭示多样性维持机制(即群落构建过程)(Purschke et al., 2013; 贾鹏和杜国祯, 2014).群落内物种可能因环境过滤或竞争排除而使谱系聚集或发散, 从而表现出特定的谱系结构(Webb, 2000; Swenson et al., 2012).例如, 环境过滤作用主导群落构建时, 筛选出的群落内物种亲缘关系可能较近, 从而适应当前生境(卢孟孟等, 2014), 其谱系结构表现为聚集; 竞争排除作用主导群落构建时, 亲缘关系较近的物种无法占用相似的生态位(Wiens & Graham, 2005), 从而排除部分近缘种, 其谱系结构表现为发散; 扩散限制作用主导群落构建时, 群落的形成与环境因子及生物间相互作用无关(Hubbell, 2001), 其谱系结构表现为随机(柴永福和岳明, 2016).先前的许多研究通过构建零模型来判断谱系结构的聚集与发散(牛红玉等, 2011), 从而证明环境过滤、竞争排除或扩散限制的存在, 阐明不同作用在群落构建过程中的相对重要性(Foster et al., 2004; Cavender-Bares et al., 2009; Mayfield & Levine, 2010; 姜晓燕等, 2016).基于此方法, 先前的研究认为, 环境因子造成的环境过滤往往主导了构建过程(Huang et al., 2014), 尤其在生境条件比较苛刻的地区, 如高海拔区域(Mori et al., 2013; Xu et al., 2017; 赵鸣飞等, 2017).然而, 海拔毕竟是一个间接化的生境因子, 其与温度、降水、紫外辐射强度、风速、土壤养分等诸多因子相关(Xu et al., 2018).探讨单一的水热梯度上群落构建的变化模式, 能够更清晰地阐明群落与水热因子的关系, 从而明确区域尺度上群落的构建模式.然而, 这方面的研究却鲜有报道. ...
... 单纯从水热梯度上进行群落构建研究的报道较少, 然而, 有大量的基于经纬度、海拔等地理因子梯度研究群落构建模式的报道.由于海拔与经纬度等信息中均包含了水热因素, 因此本研究可与这些研究做类比.例如, 大部分研究发现, 高海拔(意味着较低的温度, 由于凝冻, 也有较低的水分可用性)区域主要受到环境过滤的作用, 而低海拔(意味着较高的温度)区域则可以表现出中性或竞争排除的作用(Hulshof et al., 2013; Mori et al., 2013; Pottier et al., 2013; Xu et al., 2017); 而高纬度(意味着较低的温度、较少的降水)区域的构建模式应该是聚集的(Hodkinson et al., 2003).然而, 也有部分研究得出了不一致的结论, 例如, Zhang等(2013)发现, 亚高山草甸的群落构建模式与海拔等非生物因素无关, 而卢孟孟等(2014)发现, 哀牢山干热河谷的群落的谱系结构在高海拔区域趋于发散, 即在温度较低、水分可用性也较低的区域并未受到环境过滤的 作用. ...
群落构建研究的新进展: 进化和生态相结合的群落谱系结构研究 1 2011
... 群落内的物种组成是进化过程与生态过程共同作用的结果(黄建雄等, 2010), 以群落内物种的亲缘关系即谱系为载体进行群落构建研究, 能够较好地从进化角度反映群落构建过程中的生态学机制(Park & Potter, 2015).基于系统发育的谱系多样性考虑了不同生物类群间的谱系关系及进化历史等信息, 能够较好地从进化角度反映群落的多样性模式(Cavender-Bares et al., 2009; 杨洁等, 2014), 并揭示多样性维持机制(即群落构建过程)(Purschke et al., 2013; 贾鹏和杜国祯, 2014).群落内物种可能因环境过滤或竞争排除而使谱系聚集或发散, 从而表现出特定的谱系结构(Webb, 2000; Swenson et al., 2012).例如, 环境过滤作用主导群落构建时, 筛选出的群落内物种亲缘关系可能较近, 从而适应当前生境(卢孟孟等, 2014), 其谱系结构表现为聚集; 竞争排除作用主导群落构建时, 亲缘关系较近的物种无法占用相似的生态位(Wiens & Graham, 2005), 从而排除部分近缘种, 其谱系结构表现为发散; 扩散限制作用主导群落构建时, 群落的形成与环境因子及生物间相互作用无关(Hubbell, 2001), 其谱系结构表现为随机(柴永福和岳明, 2016).先前的许多研究通过构建零模型来判断谱系结构的聚集与发散(牛红玉等, 2011), 从而证明环境过滤、竞争排除或扩散限制的存在, 阐明不同作用在群落构建过程中的相对重要性(Foster et al., 2004; Cavender-Bares et al., 2009; Mayfield & Levine, 2010; 姜晓燕等, 2016).基于此方法, 先前的研究认为, 环境因子造成的环境过滤往往主导了构建过程(Huang et al., 2014), 尤其在生境条件比较苛刻的地区, 如高海拔区域(Mori et al., 2013; Xu et al., 2017; 赵鸣飞等, 2017).然而, 海拔毕竟是一个间接化的生境因子, 其与温度、降水、紫外辐射强度、风速、土壤养分等诸多因子相关(Xu et al., 2018).探讨单一的水热梯度上群落构建的变化模式, 能够更清晰地阐明群落与水热因子的关系, 从而明确区域尺度上群落的构建模式.然而, 这方面的研究却鲜有报道. ...
群落构建研究的新进展: 进化和生态相结合的群落谱系结构研究 1 2011
... 群落内的物种组成是进化过程与生态过程共同作用的结果(黄建雄等, 2010), 以群落内物种的亲缘关系即谱系为载体进行群落构建研究, 能够较好地从进化角度反映群落构建过程中的生态学机制(Park & Potter, 2015).基于系统发育的谱系多样性考虑了不同生物类群间的谱系关系及进化历史等信息, 能够较好地从进化角度反映群落的多样性模式(Cavender-Bares et al., 2009; 杨洁等, 2014), 并揭示多样性维持机制(即群落构建过程)(Purschke et al., 2013; 贾鹏和杜国祯, 2014).群落内物种可能因环境过滤或竞争排除而使谱系聚集或发散, 从而表现出特定的谱系结构(Webb, 2000; Swenson et al., 2012).例如, 环境过滤作用主导群落构建时, 筛选出的群落内物种亲缘关系可能较近, 从而适应当前生境(卢孟孟等, 2014), 其谱系结构表现为聚集; 竞争排除作用主导群落构建时, 亲缘关系较近的物种无法占用相似的生态位(Wiens & Graham, 2005), 从而排除部分近缘种, 其谱系结构表现为发散; 扩散限制作用主导群落构建时, 群落的形成与环境因子及生物间相互作用无关(Hubbell, 2001), 其谱系结构表现为随机(柴永福和岳明, 2016).先前的许多研究通过构建零模型来判断谱系结构的聚集与发散(牛红玉等, 2011), 从而证明环境过滤、竞争排除或扩散限制的存在, 阐明不同作用在群落构建过程中的相对重要性(Foster et al., 2004; Cavender-Bares et al., 2009; Mayfield & Levine, 2010; 姜晓燕等, 2016).基于此方法, 先前的研究认为, 环境因子造成的环境过滤往往主导了构建过程(Huang et al., 2014), 尤其在生境条件比较苛刻的地区, 如高海拔区域(Mori et al., 2013; Xu et al., 2017; 赵鸣飞等, 2017).然而, 海拔毕竟是一个间接化的生境因子, 其与温度、降水、紫外辐射强度、风速、土壤养分等诸多因子相关(Xu et al., 2018).探讨单一的水热梯度上群落构建的变化模式, 能够更清晰地阐明群落与水热因子的关系, 从而明确区域尺度上群落的构建模式.然而, 这方面的研究却鲜有报道. ...
Why close relatives make bad neighbours: Phylogenetic conservatism in niche preferences and dispersal disproves Darwin’s naturalization hypothesis in the thistle tribe 1 2015
... 群落内的物种组成是进化过程与生态过程共同作用的结果(黄建雄等, 2010), 以群落内物种的亲缘关系即谱系为载体进行群落构建研究, 能够较好地从进化角度反映群落构建过程中的生态学机制(Park & Potter, 2015).基于系统发育的谱系多样性考虑了不同生物类群间的谱系关系及进化历史等信息, 能够较好地从进化角度反映群落的多样性模式(Cavender-Bares et al., 2009; 杨洁等, 2014), 并揭示多样性维持机制(即群落构建过程)(Purschke et al., 2013; 贾鹏和杜国祯, 2014).群落内物种可能因环境过滤或竞争排除而使谱系聚集或发散, 从而表现出特定的谱系结构(Webb, 2000; Swenson et al., 2012).例如, 环境过滤作用主导群落构建时, 筛选出的群落内物种亲缘关系可能较近, 从而适应当前生境(卢孟孟等, 2014), 其谱系结构表现为聚集; 竞争排除作用主导群落构建时, 亲缘关系较近的物种无法占用相似的生态位(Wiens & Graham, 2005), 从而排除部分近缘种, 其谱系结构表现为发散; 扩散限制作用主导群落构建时, 群落的形成与环境因子及生物间相互作用无关(Hubbell, 2001), 其谱系结构表现为随机(柴永福和岳明, 2016).先前的许多研究通过构建零模型来判断谱系结构的聚集与发散(牛红玉等, 2011), 从而证明环境过滤、竞争排除或扩散限制的存在, 阐明不同作用在群落构建过程中的相对重要性(Foster et al., 2004; Cavender-Bares et al., 2009; Mayfield & Levine, 2010; 姜晓燕等, 2016).基于此方法, 先前的研究认为, 环境因子造成的环境过滤往往主导了构建过程(Huang et al., 2014), 尤其在生境条件比较苛刻的地区, 如高海拔区域(Mori et al., 2013; Xu et al., 2017; 赵鸣飞等, 2017).然而, 海拔毕竟是一个间接化的生境因子, 其与温度、降水、紫外辐射强度、风速、土壤养分等诸多因子相关(Xu et al., 2018).探讨单一的水热梯度上群落构建的变化模式, 能够更清晰地阐明群落与水热因子的关系, 从而明确区域尺度上群落的构建模式.然而, 这方面的研究却鲜有报道. ...
The accuracy of plant assemblage prediction from species distribution models varies along environmental gradients 1 2013
... 单纯从水热梯度上进行群落构建研究的报道较少, 然而, 有大量的基于经纬度、海拔等地理因子梯度研究群落构建模式的报道.由于海拔与经纬度等信息中均包含了水热因素, 因此本研究可与这些研究做类比.例如, 大部分研究发现, 高海拔(意味着较低的温度, 由于凝冻, 也有较低的水分可用性)区域主要受到环境过滤的作用, 而低海拔(意味着较高的温度)区域则可以表现出中性或竞争排除的作用(Hulshof et al., 2013; Mori et al., 2013; Pottier et al., 2013; Xu et al., 2017); 而高纬度(意味着较低的温度、较少的降水)区域的构建模式应该是聚集的(Hodkinson et al., 2003).然而, 也有部分研究得出了不一致的结论, 例如, Zhang等(2013)发现, 亚高山草甸的群落构建模式与海拔等非生物因素无关, 而卢孟孟等(2014)发现, 哀牢山干热河谷的群落的谱系结构在高海拔区域趋于发散, 即在温度较低、水分可用性也较低的区域并未受到环境过滤的 作用. ...
Contrasting changes in taxonomic, phylogenetic and functional diversity during a long-term succession: Insights into assembly processes 1 2013
... 群落内的物种组成是进化过程与生态过程共同作用的结果(黄建雄等, 2010), 以群落内物种的亲缘关系即谱系为载体进行群落构建研究, 能够较好地从进化角度反映群落构建过程中的生态学机制(Park & Potter, 2015).基于系统发育的谱系多样性考虑了不同生物类群间的谱系关系及进化历史等信息, 能够较好地从进化角度反映群落的多样性模式(Cavender-Bares et al., 2009; 杨洁等, 2014), 并揭示多样性维持机制(即群落构建过程)(Purschke et al., 2013; 贾鹏和杜国祯, 2014).群落内物种可能因环境过滤或竞争排除而使谱系聚集或发散, 从而表现出特定的谱系结构(Webb, 2000; Swenson et al., 2012).例如, 环境过滤作用主导群落构建时, 筛选出的群落内物种亲缘关系可能较近, 从而适应当前生境(卢孟孟等, 2014), 其谱系结构表现为聚集; 竞争排除作用主导群落构建时, 亲缘关系较近的物种无法占用相似的生态位(Wiens & Graham, 2005), 从而排除部分近缘种, 其谱系结构表现为发散; 扩散限制作用主导群落构建时, 群落的形成与环境因子及生物间相互作用无关(Hubbell, 2001), 其谱系结构表现为随机(柴永福和岳明, 2016).先前的许多研究通过构建零模型来判断谱系结构的聚集与发散(牛红玉等, 2011), 从而证明环境过滤、竞争排除或扩散限制的存在, 阐明不同作用在群落构建过程中的相对重要性(Foster et al., 2004; Cavender-Bares et al., 2009; Mayfield & Levine, 2010; 姜晓燕等, 2016).基于此方法, 先前的研究认为, 环境因子造成的环境过滤往往主导了构建过程(Huang et al., 2014), 尤其在生境条件比较苛刻的地区, 如高海拔区域(Mori et al., 2013; Xu et al., 2017; 赵鸣飞等, 2017).然而, 海拔毕竟是一个间接化的生境因子, 其与温度、降水、紫外辐射强度、风速、土壤养分等诸多因子相关(Xu et al., 2018).探讨单一的水热梯度上群落构建的变化模式, 能够更清晰地阐明群落与水热因子的关系, 从而明确区域尺度上群落的构建模式.然而, 这方面的研究却鲜有报道. ...
Phylogenetic structure and phylogenetic diversity of angiosperm assemblages in forests along an elevational gradient in Changbaishan, China 1 2014
... 群落内物种的多度反映了该物种在群落中的相对重要性, 代表了该物种占有资源的能力(Grime, 1998; 曹科等, 2013).基于物种组成数据, 可以探究相似物种组成的群落的分布模式.群落内的生物多样性则与多种环境因子有关(Qian et al., 2014; 车应弟等, 2017).通过多样性分析, 可以探究群落内物种形成、共存及时空变化规律(Swenson et al., 2012).因此, 研究环境因子与群落物种组成及多样性之间的关系, 有助于预测多样性的动态(Yang & Da, 2006; 周晓果等, 2014), 进而理解群落构建的过程. ...
A latitudinal gradient in tree community assembly processes evidenced in Chinese forests 2 2015
... 热量与水分条件是驱动群落构建的最主要、最直接的因素(Xu et al., 2018).前人对于群落构建的研究发现, 海拔和纬度能够驱动群落构建(Qiao et al., 2015; Xu et al., 2017), 是因为海拔与纬度同热量条件呈明显的负相关关系(唐志尧等, 2004).在本研究区域内, 由于纬度跨度大, MAT也有很大差异.因此, 海拔和纬度同时影响了研究样地的MAT.本研究发现, 造成华北区域内阔叶林群落谱系关系变化的主要因子是MAT, 且还与MMTD有关(表2).对NRI、NTI及PD与MAT的显著拟合回归结果也证明了这一点(图2, 图4).另外, 谱系变化还与坡向有关(表2), 而坡向的差异正是导致群落热量差异的因素之一.因此, 热量是影响华北区域阔叶林群落构建的最主要因素.除此之外, 水分(如本研究中的MAP)也在较大程度上影响着群落构建的过程(表2). ...
Shifts in plant community assembly processes across growth forms along a habitat severity gradient: A Test of the plant functional trait approach 3 2018
... 群落内的物种组成是进化过程与生态过程共同作用的结果(黄建雄等, 2010), 以群落内物种的亲缘关系即谱系为载体进行群落构建研究, 能够较好地从进化角度反映群落构建过程中的生态学机制(Park & Potter, 2015).基于系统发育的谱系多样性考虑了不同生物类群间的谱系关系及进化历史等信息, 能够较好地从进化角度反映群落的多样性模式(Cavender-Bares et al., 2009; 杨洁等, 2014), 并揭示多样性维持机制(即群落构建过程)(Purschke et al., 2013; 贾鹏和杜国祯, 2014).群落内物种可能因环境过滤或竞争排除而使谱系聚集或发散, 从而表现出特定的谱系结构(Webb, 2000; Swenson et al., 2012).例如, 环境过滤作用主导群落构建时, 筛选出的群落内物种亲缘关系可能较近, 从而适应当前生境(卢孟孟等, 2014), 其谱系结构表现为聚集; 竞争排除作用主导群落构建时, 亲缘关系较近的物种无法占用相似的生态位(Wiens & Graham, 2005), 从而排除部分近缘种, 其谱系结构表现为发散; 扩散限制作用主导群落构建时, 群落的形成与环境因子及生物间相互作用无关(Hubbell, 2001), 其谱系结构表现为随机(柴永福和岳明, 2016).先前的许多研究通过构建零模型来判断谱系结构的聚集与发散(牛红玉等, 2011), 从而证明环境过滤、竞争排除或扩散限制的存在, 阐明不同作用在群落构建过程中的相对重要性(Foster et al., 2004; Cavender-Bares et al., 2009; Mayfield & Levine, 2010; 姜晓燕等, 2016).基于此方法, 先前的研究认为, 环境因子造成的环境过滤往往主导了构建过程(Huang et al., 2014), 尤其在生境条件比较苛刻的地区, 如高海拔区域(Mori et al., 2013; Xu et al., 2017; 赵鸣飞等, 2017).然而, 海拔毕竟是一个间接化的生境因子, 其与温度、降水、紫外辐射强度、风速、土壤养分等诸多因子相关(Xu et al., 2018).探讨单一的水热梯度上群落构建的变化模式, 能够更清晰地阐明群落与水热因子的关系, 从而明确区域尺度上群落的构建模式.然而, 这方面的研究却鲜有报道. ...
... 热量与水分条件是驱动群落构建的最主要、最直接的因素(Xu et al., 2018).前人对于群落构建的研究发现, 海拔和纬度能够驱动群落构建(Qiao et al., 2015; Xu et al., 2017), 是因为海拔与纬度同热量条件呈明显的负相关关系(唐志尧等, 2004).在本研究区域内, 由于纬度跨度大, MAT也有很大差异.因此, 海拔和纬度同时影响了研究样地的MAT.本研究发现, 造成华北区域内阔叶林群落谱系关系变化的主要因子是MAT, 且还与MMTD有关(表2).对NRI、NTI及PD与MAT的显著拟合回归结果也证明了这一点(图2, 图4).另外, 谱系变化还与坡向有关(表2), 而坡向的差异正是导致群落热量差异的因素之一.因此, 热量是影响华北区域阔叶林群落构建的最主要因素.除此之外, 水分(如本研究中的MAP)也在较大程度上影响着群落构建的过程(表2). ...
... 然而, 在MAP梯度上, NRI和NTI所表现出的变化模式不同, 这与前人得出的NRI与NTI表现模式不符(Swenson et al., 2007), 但也说明了二者的适用范围和敏感程度不一致, 有必要分别进行研究(Tucker et al., 2017).由于NRI的取值均未达到与零模型有显著差异的水平, NRI在MAP梯度上并未表现出明显的变化趋势; 而NTI则随MAP的增加表现出聚集的趋势, 表明受到了环境过滤的作用.一般而言, MAP较高的区域, 纬度更低, 更接近海岸, 且可能有较低的海拔.就本研究而言, MAP最高的几个样地位于河南、山东等省海拔较低的地区(表1), 而海拔较低的区域往往有更严重的人为扰动(Xu et al., 2018), 从而导致这些样地内的环境条件较差(Lessard et al., 2009), 使群落受到环境过滤的作用. ...
Using phylogeny and functional traits for assessing community assembly along environmental gradients: A deterministic process driven by elevation 3 2017
... 群落内的物种组成是进化过程与生态过程共同作用的结果(黄建雄等, 2010), 以群落内物种的亲缘关系即谱系为载体进行群落构建研究, 能够较好地从进化角度反映群落构建过程中的生态学机制(Park & Potter, 2015).基于系统发育的谱系多样性考虑了不同生物类群间的谱系关系及进化历史等信息, 能够较好地从进化角度反映群落的多样性模式(Cavender-Bares et al., 2009; 杨洁等, 2014), 并揭示多样性维持机制(即群落构建过程)(Purschke et al., 2013; 贾鹏和杜国祯, 2014).群落内物种可能因环境过滤或竞争排除而使谱系聚集或发散, 从而表现出特定的谱系结构(Webb, 2000; Swenson et al., 2012).例如, 环境过滤作用主导群落构建时, 筛选出的群落内物种亲缘关系可能较近, 从而适应当前生境(卢孟孟等, 2014), 其谱系结构表现为聚集; 竞争排除作用主导群落构建时, 亲缘关系较近的物种无法占用相似的生态位(Wiens & Graham, 2005), 从而排除部分近缘种, 其谱系结构表现为发散; 扩散限制作用主导群落构建时, 群落的形成与环境因子及生物间相互作用无关(Hubbell, 2001), 其谱系结构表现为随机(柴永福和岳明, 2016).先前的许多研究通过构建零模型来判断谱系结构的聚集与发散(牛红玉等, 2011), 从而证明环境过滤、竞争排除或扩散限制的存在, 阐明不同作用在群落构建过程中的相对重要性(Foster et al., 2004; Cavender-Bares et al., 2009; Mayfield & Levine, 2010; 姜晓燕等, 2016).基于此方法, 先前的研究认为, 环境因子造成的环境过滤往往主导了构建过程(Huang et al., 2014), 尤其在生境条件比较苛刻的地区, 如高海拔区域(Mori et al., 2013; Xu et al., 2017; 赵鸣飞等, 2017).然而, 海拔毕竟是一个间接化的生境因子, 其与温度、降水、紫外辐射强度、风速、土壤养分等诸多因子相关(Xu et al., 2018).探讨单一的水热梯度上群落构建的变化模式, 能够更清晰地阐明群落与水热因子的关系, 从而明确区域尺度上群落的构建模式.然而, 这方面的研究却鲜有报道. ...
... 热量与水分条件是驱动群落构建的最主要、最直接的因素(Xu et al., 2018).前人对于群落构建的研究发现, 海拔和纬度能够驱动群落构建(Qiao et al., 2015; Xu et al., 2017), 是因为海拔与纬度同热量条件呈明显的负相关关系(唐志尧等, 2004).在本研究区域内, 由于纬度跨度大, MAT也有很大差异.因此, 海拔和纬度同时影响了研究样地的MAT.本研究发现, 造成华北区域内阔叶林群落谱系关系变化的主要因子是MAT, 且还与MMTD有关(表2).对NRI、NTI及PD与MAT的显著拟合回归结果也证明了这一点(图2, 图4).另外, 谱系变化还与坡向有关(表2), 而坡向的差异正是导致群落热量差异的因素之一.因此, 热量是影响华北区域阔叶林群落构建的最主要因素.除此之外, 水分(如本研究中的MAP)也在较大程度上影响着群落构建的过程(表2). ...
... 单纯从水热梯度上进行群落构建研究的报道较少, 然而, 有大量的基于经纬度、海拔等地理因子梯度研究群落构建模式的报道.由于海拔与经纬度等信息中均包含了水热因素, 因此本研究可与这些研究做类比.例如, 大部分研究发现, 高海拔(意味着较低的温度, 由于凝冻, 也有较低的水分可用性)区域主要受到环境过滤的作用, 而低海拔(意味着较高的温度)区域则可以表现出中性或竞争排除的作用(Hulshof et al., 2013; Mori et al., 2013; Pottier et al., 2013; Xu et al., 2017); 而高纬度(意味着较低的温度、较少的降水)区域的构建模式应该是聚集的(Hodkinson et al., 2003).然而, 也有部分研究得出了不一致的结论, 例如, Zhang等(2013)发现, 亚高山草甸的群落构建模式与海拔等非生物因素无关, 而卢孟孟等(2014)发现, 哀牢山干热河谷的群落的谱系结构在高海拔区域趋于发散, 即在温度较低、水分可用性也较低的区域并未受到环境过滤的 作用. ...
中山湿性常绿阔叶林系统发育和功能性状的α及β多样性 1 2014
... 群落内的物种组成是进化过程与生态过程共同作用的结果(黄建雄等, 2010), 以群落内物种的亲缘关系即谱系为载体进行群落构建研究, 能够较好地从进化角度反映群落构建过程中的生态学机制(Park & Potter, 2015).基于系统发育的谱系多样性考虑了不同生物类群间的谱系关系及进化历史等信息, 能够较好地从进化角度反映群落的多样性模式(Cavender-Bares et al., 2009; 杨洁等, 2014), 并揭示多样性维持机制(即群落构建过程)(Purschke et al., 2013; 贾鹏和杜国祯, 2014).群落内物种可能因环境过滤或竞争排除而使谱系聚集或发散, 从而表现出特定的谱系结构(Webb, 2000; Swenson et al., 2012).例如, 环境过滤作用主导群落构建时, 筛选出的群落内物种亲缘关系可能较近, 从而适应当前生境(卢孟孟等, 2014), 其谱系结构表现为聚集; 竞争排除作用主导群落构建时, 亲缘关系较近的物种无法占用相似的生态位(Wiens & Graham, 2005), 从而排除部分近缘种, 其谱系结构表现为发散; 扩散限制作用主导群落构建时, 群落的形成与环境因子及生物间相互作用无关(Hubbell, 2001), 其谱系结构表现为随机(柴永福和岳明, 2016).先前的许多研究通过构建零模型来判断谱系结构的聚集与发散(牛红玉等, 2011), 从而证明环境过滤、竞争排除或扩散限制的存在, 阐明不同作用在群落构建过程中的相对重要性(Foster et al., 2004; Cavender-Bares et al., 2009; Mayfield & Levine, 2010; 姜晓燕等, 2016).基于此方法, 先前的研究认为, 环境因子造成的环境过滤往往主导了构建过程(Huang et al., 2014), 尤其在生境条件比较苛刻的地区, 如高海拔区域(Mori et al., 2013; Xu et al., 2017; 赵鸣飞等, 2017).然而, 海拔毕竟是一个间接化的生境因子, 其与温度、降水、紫外辐射强度、风速、土壤养分等诸多因子相关(Xu et al., 2018).探讨单一的水热梯度上群落构建的变化模式, 能够更清晰地阐明群落与水热因子的关系, 从而明确区域尺度上群落的构建模式.然而, 这方面的研究却鲜有报道. ...
中山湿性常绿阔叶林系统发育和功能性状的α及β多样性 1 2014
... 群落内的物种组成是进化过程与生态过程共同作用的结果(黄建雄等, 2010), 以群落内物种的亲缘关系即谱系为载体进行群落构建研究, 能够较好地从进化角度反映群落构建过程中的生态学机制(Park & Potter, 2015).基于系统发育的谱系多样性考虑了不同生物类群间的谱系关系及进化历史等信息, 能够较好地从进化角度反映群落的多样性模式(Cavender-Bares et al., 2009; 杨洁等, 2014), 并揭示多样性维持机制(即群落构建过程)(Purschke et al., 2013; 贾鹏和杜国祯, 2014).群落内物种可能因环境过滤或竞争排除而使谱系聚集或发散, 从而表现出特定的谱系结构(Webb, 2000; Swenson et al., 2012).例如, 环境过滤作用主导群落构建时, 筛选出的群落内物种亲缘关系可能较近, 从而适应当前生境(卢孟孟等, 2014), 其谱系结构表现为聚集; 竞争排除作用主导群落构建时, 亲缘关系较近的物种无法占用相似的生态位(Wiens & Graham, 2005), 从而排除部分近缘种, 其谱系结构表现为发散; 扩散限制作用主导群落构建时, 群落的形成与环境因子及生物间相互作用无关(Hubbell, 2001), 其谱系结构表现为随机(柴永福和岳明, 2016).先前的许多研究通过构建零模型来判断谱系结构的聚集与发散(牛红玉等, 2011), 从而证明环境过滤、竞争排除或扩散限制的存在, 阐明不同作用在群落构建过程中的相对重要性(Foster et al., 2004; Cavender-Bares et al., 2009; Mayfield & Levine, 2010; 姜晓燕等, 2016).基于此方法, 先前的研究认为, 环境因子造成的环境过滤往往主导了构建过程(Huang et al., 2014), 尤其在生境条件比较苛刻的地区, 如高海拔区域(Mori et al., 2013; Xu et al., 2017; 赵鸣飞等, 2017).然而, 海拔毕竟是一个间接化的生境因子, 其与温度、降水、紫外辐射强度、风速、土壤养分等诸多因子相关(Xu et al., 2018).探讨单一的水热梯度上群落构建的变化模式, 能够更清晰地阐明群落与水热因子的关系, 从而明确区域尺度上群落的构建模式.然而, 这方面的研究却鲜有报道. ...
丘陵地区地形梯度上植被格局的分异研究概述 1 2006
... 群落内物种的多度反映了该物种在群落中的相对重要性, 代表了该物种占有资源的能力(Grime, 1998; 曹科等, 2013).基于物种组成数据, 可以探究相似物种组成的群落的分布模式.群落内的生物多样性则与多种环境因子有关(Qian et al., 2014; 车应弟等, 2017).通过多样性分析, 可以探究群落内物种形成、共存及时空变化规律(Swenson et al., 2012).因此, 研究环境因子与群落物种组成及多样性之间的关系, 有助于预测多样性的动态(Yang & Da, 2006; 周晓果等, 2014), 进而理解群落构建的过程. ...
丘陵地区地形梯度上植被格局的分异研究概述 1 2006
... 群落内物种的多度反映了该物种在群落中的相对重要性, 代表了该物种占有资源的能力(Grime, 1998; 曹科等, 2013).基于物种组成数据, 可以探究相似物种组成的群落的分布模式.群落内的生物多样性则与多种环境因子有关(Qian et al., 2014; 车应弟等, 2017).通过多样性分析, 可以探究群落内物种形成、共存及时空变化规律(Swenson et al., 2012).因此, 研究环境因子与群落物种组成及多样性之间的关系, 有助于预测多样性的动态(Yang & Da, 2006; 周晓果等, 2014), 进而理解群落构建的过程. ...
 ,1,*, 郭垚鑫1, 康慕谊2,3, 刘全儒4, 郑成洋5, 吉成均5, 闫明6, 张峰7, 高贤明8, 王仁卿9, 石福臣10, 张钦弟6, 王茂1,11
,1,*, 郭垚鑫1, 康慕谊2,3, 刘全儒4, 郑成洋5, 吉成均5, 闫明6, 张峰7, 高贤明8, 王仁卿9, 石福臣10, 张钦弟6, 王茂1,11
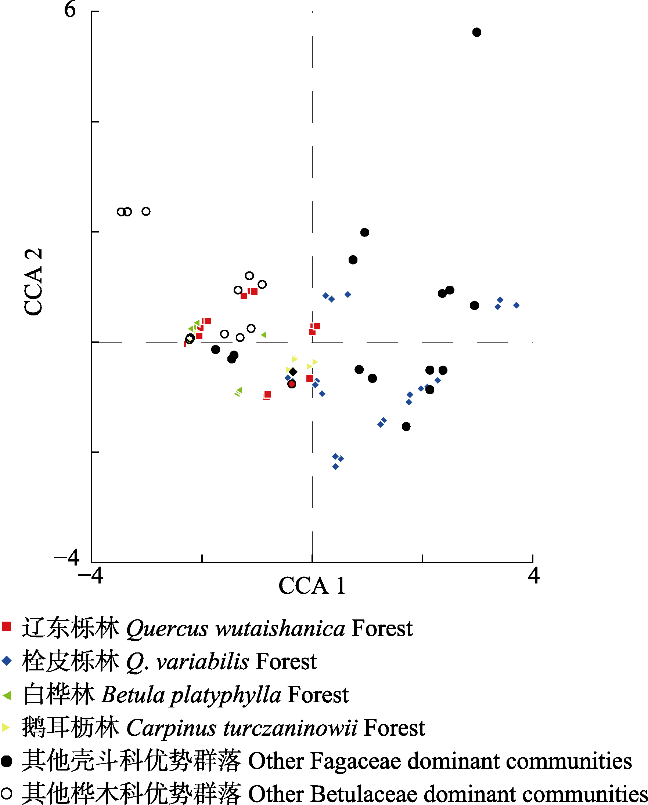 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT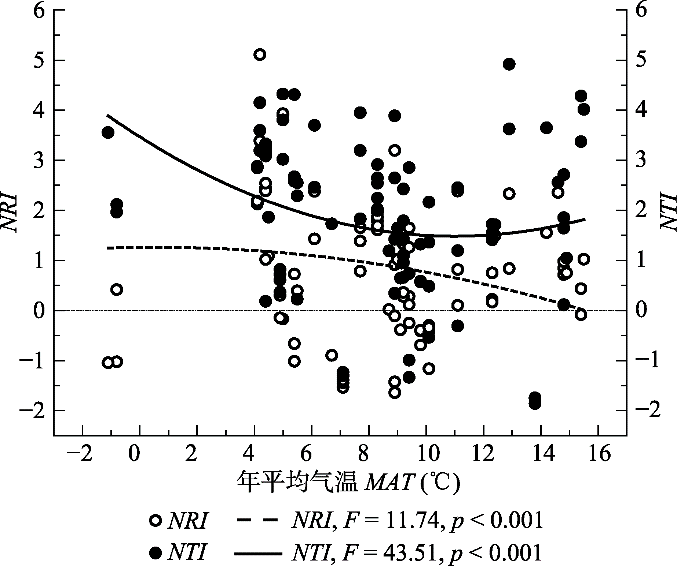 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT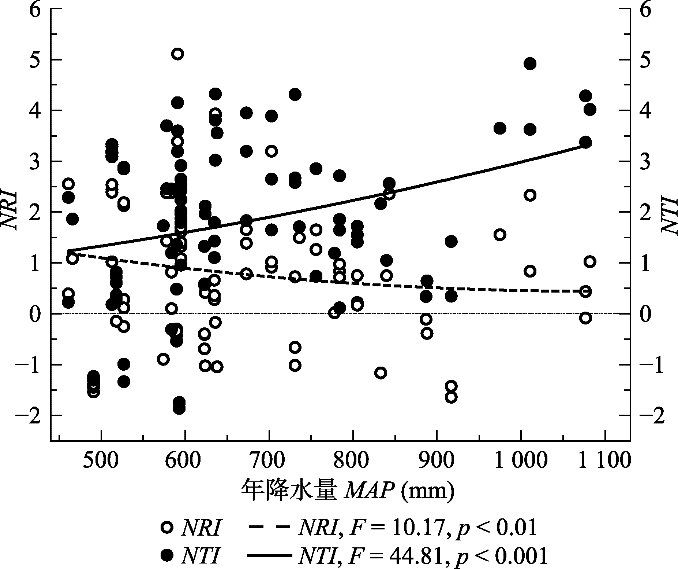 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT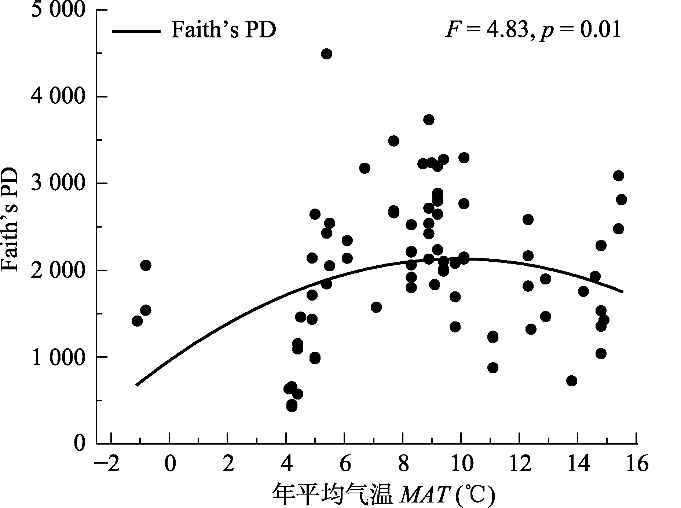 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT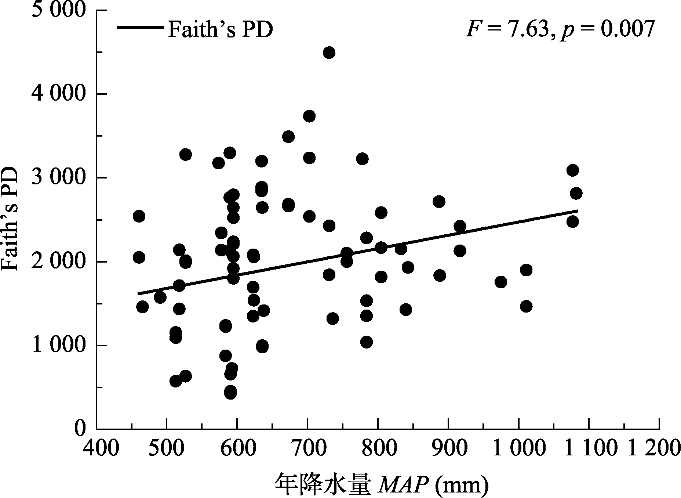 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}