Seasonal dynamics of non-structural carbohydrate content in branch of Quercus variabilis growing in east Qinling Mountain range
Yi-Ping ZHANG,1, Xu-Ying HAI1,2, Jun-Liang XU,1,*, Wen-Xia WU1, Peng-He CAO1,3, Wen-Jing AN41 College of Forestry, Henan University of Science and Technology, Luoyang, Henan 471023, China 2 College of Forestry, Northwest Agriculture and Forestry University, Yangling, Shaanxi 712100, China 3 College of Biology and the Environment, Nanjing Forestry University, Nanjing 210037, China 4 Beijing Jingxi Forestry Farm, Beijing 102300, China
Supported by the National Natural Science Foundation of China(41401063) Supported by the National Natural Science Foundation of China(41801026) the China Scholarship Council(201808410575) the China Scholarship Council(201908410061)
Abstract Aims Measure of non-structural carbohydrate (NSC) reserves indicates tree carbon surplus or shortage stored. Branches connect NSC sources (leaves) and NSC sinks (stemwood, root) of woody plants. Therefore, the seasonal dynamics of NSC concentration in branches will be of important implications for understanding and modeling plant carbon allocation. Methods We conducted a field survey monitoring branch NSC concentrations of Chinese cork oak (Quercus variabilis). We also synchronously observed the leaf phenology of the trees in uneven-aged secondary oak forests at its upper and lower distribution limits (650 m to 970 m) in east Qinling Mountain ranges. Sampling intervals were set semimonthly/monthly during the leaf unfolding period (March to May), and monthly/bimonthly during the tree’s full growing season (June to November) from May 2016 to May 2017. Important findings (1) The NSC measures in the tree branches had weak seasonal changes at both sites. However, the soluble sugar (SS) concentrations at the upper elevation site and the starch (S) concentrations at the lower site had significant seasonal changes. The relative stable NSC levels vs. larger seasonal oscillations of soluble sugar and starch may be explained by the mutual conversion between soluble sugar and starch in the tree branches. (2) Soluble sugar was the major contributor to the total NSC in oak branches, accounting for approximately 61% of it. Here the soluble sugars performed as quick C whereas starch acted mostly as reserved C for future use, it could be inferred that the Q. variabilis, a warm temperate deciduous tree species, developed this feature as its life strategy to survive in warm temperate climate. (3) Soil water availability was positively related to the NSC measures at the high elevation site, while vapor pressure deficit (VPD) was negatively related to the NSC at the low elevation site, indicating the oak may be more drought-susceptible to water stress at lower elevation. (4) The maximum and the minimum concentrations of NSC in tree branches were observed before bud break (late March, approximately 11%) and when full leaf expansion (late April, approximately 11%), respectively. These extremes of NSC could be partly explained by the simultaneous leaf phenological dynamics. Considering the fact of carbon supply for bud break and leaf development via branches rather than by photosynthesis, it was reasonable that the NSC concentration in branches of Q. variabilis reached its maximum before the bud break, and did not change significantly with elevation. Not surprisingly, the significant differences in branches NSC with elevation only occurred during bud break in spring, as a later phenology occurred at higher elevation. In conclusion, this study confirms that the leaf phenology of Q. variabilis directly affects the branches seasonal patterns of NSC, particularly in the spring. The relationship between the carbon budget of the oak branches and its aboveground phenology should be more emphasized for further comprehension on the NSC metabolism. Keywords:non-structural carbohydrate;Quercus variabilis;leaf phenology;altitude;vapor pressure deficit
PDF (1382KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 章异平, 海旭莹, 徐军亮, 吴文霞, 曹鹏鹤, 安文静. 秦岭东段栓皮栎枝条非结构性碳水化合物含量的季节动态. 植物生态学报, 2019, 43(6): 521-531. DOI: 10.17521/cjpe.2018.0325 ZHANG Yi-Ping, HAI Xu-Ying, XU Jun-Liang, WU Wen-Xia, CAO Peng-He, AN Wen-Jing. Seasonal dynamics of non-structural carbohydrate content in branch of Quercus variabilis growing in east Qinling Mountain range. Chinese Journal of Plant Ecology, 2019, 43(6): 521-531. DOI: 10.17521/cjpe.2018.0325
非结构性碳水化合物(NSC)是植物碳水化合物生产过剩时积累的可转化的暂时储存物, 是植物生长代谢过程中重要的能源物质, 在光合作用不足时起一定的调节和缓冲作用(Dietze et al., 2014)。NSC累积量的大小可以很好地反映植物体内碳供应与碳需求的平衡关系(Li et al., 2001; Fajardo et al., 2012), 因此, 研究植物NSC含量的动态变化规律是评价植物体内碳平衡的快速、有效的重要方法。
Table 1 表1 表1秦岭东段栓皮栎林2016和2017年高低海拔环境因子月值 Table 1Monthly measured environmental parameters at upper and lower altitudes in Quercus variabilis forests in the east Qinling Mountain range in 2016 and 2017
年 Year
月 Month
空气温度 Air temperature (℃)
土壤温度 Soil temperature (℃)
相对湿度 Relative humidity (%)
土壤体积含水量 SWC (%)
高海拔 Upper
低海拔 Lower
高海拔 Upper
低海拔 Lower
高海拔 Upper
低海拔 Lower
高海拔 Upper
低海拔 Lower
2016
3
8.2
8.9
5.6
7.0
47
51
16.6
15.4
4
14.7
15.1
10.8
13.2
62
69
17.9
15.9
5
16.5
16.8
13.5
15.6
65
71
14.8
11.9
6
22.0
22.2
18.1
20.4
72
77
17.8
14.2
7
23.5
24.0
20.4
23.2
84
86
16.7
13.1
8
22.6
23.3
20.9
23.6
87
90
16.5
13.2
9
18.6
19.5
18.1
20.1
77
80
12.0
11.0
10
11.4
12.6
13.6
14.9
89
90
13.8
12.5
11
6.1
6.7
7.6
8.8
74
79
20.6
17.4
12
2.0
3.3
2.6
6.0
74
76
21.6
NA
2017
1
-0.7
0.1
1.0
1.0
73
70
21.2
12.8
2
3.1
3.9
NA
NA
67
67
NA
NA
3
6.3
7.2
5.3
6.4
61
64
20.5
11.9
4
14.1
14.5
10.3
11.7
59
63
20.7
12.9
5
18.7
19.1
13.9
15.7
57
63
18.4
10.0
NA, data missed; SWC, soil volumetric water content. NA, 当月数据有缺失。
Table 2 表2 表2秦岭东段栓皮栎林样地基本情况 Table 2Physical (altitude, slope and aspect) and forest (canopy closure, average tree age, average tree height and average DBH) measures of sample plots in Quercus variabilis forest in the east Qinling Mountain range
Fig. 1Chronological changes of non-structural carbohydrate (NSC) and its components in the branches of Quercus variabilis at upper altitude in east Qinling Mountain (mean ± SD). Shaded area represent the non-growing season of Q. variabilis based on leaf phenology.
Fig. 2Chronological changes of non-structural carbohydrate (NSC) and its components in the branches of Quercus variabilis at lower altitude in east Qinling Mountain (mean ± SD). Shaded area represent the non-growing season of Q. variabilis based on leaf phenology.
新窗口打开|下载原图ZIP|生成PPT 图3秦岭东段不同海拔栓皮栎枝条非结构性碳水化合物(NSC)及其组分含量对比分析(平均值±标准偏差)。不同小写字母表示同一采样日不同海拔间NSC及其组分差异显著(p < 0.05, Paired-sample t test), 其中NSC用字母ab表示, 淀粉用字母cd表示, 可溶性糖用字母ef表示。
Fig. 3A comparison of non-structural carbohydrate (NSC) and its components in branches of Quercus variabilis at different altitudes in east Qinling Mountain (mean ± SD). Different lowercase letters indicate significant differences between two altitudes based on Paired-sample t test (p < 0 .05). Letters a and b refer to NSC, letters c and d refer to starch, and letters e and f refer to soluble sugars.
Table 3 表3 表3秦岭东段不同海拔环境因子与栓皮栎枝条非结构性碳水化合物(NSC)含量相关性分析 Table 3Pearson correlation between measured environmental variables and branch NSC contents of Quercus variabilis at upper and lower altitudes in east Qinling Mountain
样地 Plot
成分 Compound
空气温度 Ta (℃)
土壤温度 Ts (℃)
相对湿度 RH (%)
土壤体积含水量 SWC (%)
降水量 P (mm)
饱和水汽压差 VPD (kPa)
高海拔 Upper altitude
SS
-0.578
-0.589
-0.164
0.717*
-0.190
-0.278
S
-0.190
0.008
0.121
-0.537
0.049
-0.185
NSC
-0.590
-0.472
-0.055
0.235
-0.123
-0.349
SS/S
-0.178
-0.194
0.121
0.644*
-0.261
-0.297
低海拔 Lower altitude
SS
-0.591
-0.453
0.378
0.315
-0.051
-0.552
S
-0.343
-0.250
0.722*
0.302
-0.470
-0.788**
NSC
-0.564
-0.423
0.703*
0.382
-0.349
-0.845**
SS/S
0.296
0.174
-0.546
-0.246
0.821**
0.702**
Ta, air temperature; Ts, soil temperature; RH, relative humidity; SWC, soil volumetric water content; P, precipitation; VPD, vapor pressure deficit. NSC, non-structural carbohydrate; S, starch; SS, soluble sugar. Significant correlation (p < 0.05) is marked bold, ?* and ** correspond to significance levels at 0.05 and 0.01 (n = 10), respectively. NSC, 非结构性碳水化合物; S, 淀粉; SS, 可溶性糖。*和**代表置信度(双侧)为0.05和0.01时, 相关性显著, 并用加粗字体显示。
Fig. 4A comparison between measured and modeling values extracted from Gompertz fitting curve of leaf phenology of uppnr and lower altitude Quercus variabilis in east Qinling Mountain. ▲, measured value of leaf phenology at upper altitude; ■, measured value of leaf phenology at lower altitude; ┅, gompertz fitting curve of leaf phenology of upper altitude; ━, gompertz fitting curve of leaf phenology of lower altitude.
Table 4 表4 表4秦岭东段栓皮栎叶片物候主要阶段日期(拟合值) Table 4The dates of main leaf phenology phase of Quercus variabilis based on Gompertz fitting values in east Qinling Mountain
AugustoL, DaviesTJ, DelzonS, De SchrijverA (2014). The enigma of the rise of angiosperms: Can we untie the knot? Ecology Letters, 17, 1326-1338. [本文引用: 1]
BrüggemannN, GesslerA, KaylerZ, KeelSG, BadeckF, BarthelM, BoeckxP, BuchmannN, BrugnoliE, EsperschützJ, GavrichkovaO, GhashghaieJ, Gomez-CasanovasN, KeitelC, KnohlA, KuptzD, PalacioS, SalmonY, UchidaY, BahnM (2011). Carbon allocation and carbon isotope fluxes in the plant-soil-atmosphere continuum: A review Biogeosciences, 8, 3457-3489. [本文引用: 1]
CaoXX, DingSY, LiHM (2003). A study on vegetation in the state-owned tree farm of Quanbao Mountain in Luoning County, Henan Province Henan Science, 21, 183-186. [本文引用: 1]
CarboneMS, TrumboreSE (2007). Contribution of new photosynthetic assimilates to respiration by perennial grasses and shrubs: Residence times and allocation patterns New Phytologist, 176, 124-135. [本文引用: 1]
ChapinFS, SchulzeE, MooneyHA (1990). The ecology and economics of storage in plants Annual Review of Ecology and Systematics, 21, 423-447. [本文引用: 1]
ChenZC, WangL, DaiYX, WanXC, LiuSR (2017). Phenology- ?dependent variation in the non-structural carbohydrates of broadleaf evergreen species plays an important role in determining tolerance to defoliation (or herbivory) Scientific Reports, 7, 10125. DOI: 10.1038/s41598-017-09757-2. [本文引用: 1]
DengYP (2016). Nonstructural Carbohydrates and Nutrient Element Concentrations of Quercus variabilis of Different Latitude Master degree dissertation, Chinese Academy of Forestry Sciences, Beijing. [本文引用: 3]
EricssonA (1984). Effects of low temperature and light treatment, following winter cold storage, on starch accumulation in Scots pine seedlings Canadian Journal of Forest Research, 14, 114-118. [本文引用: 1]
FajardoA, PiperFI, PfundL, K?rnerC, HochG (2012). Variation of mobile carbon reserves in trees at the alpine treeline ecotone is under environmental control New Phytologist, 195, 794-802. [本文引用: 1]
HochG (2008). The carbon supply of Picea abies trees at a Swiss montane permafrost site Plant Ecology & Diversity, 1, 13-20. [本文引用: 1]
HochG, K?rnerC (2012). Global patterns of mobile carbon stores in trees at the high-elevation tree line Global Ecology and Biogeography, 21, 861-871. [本文引用: 1]
HochG, RichterA, K?rnerC (2003). Non-structural carbon compounds in temperate forest trees Plant, Cell & Environment, 26, 1067-1081. [本文引用: 3]
KeelSG, Sch?delC (2010). Expanding leaves of mature deciduous forest trees rapidly become autotrophic Tree Physiology, 30, 1253-1259. [本文引用: 1]
KitinP, FunadaR (2016). Earlywood vessels in ring-porous trees become functional for water transport after bud burst and before the maturation of the current-year leaves IAWA Journal, 37, 315-331. [本文引用: 1]
KleinT, HochG, YakirD, K?rnerC (2014). Drought stress, growth and nonstructural carbohydrate dynamics of pine trees in a semi-arid forest Tree Physiology, 34, 981-992. [本文引用: 1]
KleinT, VitasseY, HochG (2016). Coordination between growth, phenology and carbon storage in three coexisting deciduous tree species in a temperate forest Tree Physiology, 36, 847-855. [本文引用: 2]
Kontunen-SoppelaS, LankilaJ, L?hdesm?kiP, LaineK (2002). Response of protein and carbohydrate metabolism of Scots pine seedlings to low temperature Journal of Plant Physiology, 159, 175-180. [本文引用: 1]
KozlowskiSW, HattrupK (1992). A disagreement about within-group agreement: Disentangling issues of consistency versus consensus Journal of Applied Psychology, 77, 161-167. [本文引用: 1]
LiFD, LvDG, QinSJ, DuGD, MaHY, LiuGC (2014). Effects of grass coverage management on photosynthetic characteristics of apple leaves during leaf expanding Pratacultural Science, 31, 462-467. [本文引用: 1]
LiMH, HochG, K?rnerC (2001). Spatial variability of mobile carbohydrates within Pinus cembra trees at the alpine treeline Phyton-Annales Rei Botanicae, 41, 203-213. [本文引用: 1]
LiMH, XiaoWF, WangSG, ChengGW, CherubiniP, CaiXH, LiuXL, WangXD, ZhuWZ (2008). Mobile carbohydrates in Himalayan treeline trees I. Evidence for carbon gain limitation but not for growth limitation Tree Physiology, 28, 1287-1296. [本文引用: 1]
LiNN, HeNP, YuGR (2015). Non-structural carbohydrates in leaves of tree species from four typical forests in China Journal of Northwest Plants, 35, 1846-1854. [本文引用: 1]
Martínez-VilaltaJ, SalaA, AsensioD, GalianoL, HochG, PalacioS, PiperFI, LloretF (2016). Dynamics of non-structural carbohydrates in terrestrial plants: A global synthesis Ecological Monographs, 86, 495-516. [本文引用: 1]
MillardP, GreletGA (2010). Nitrogen storage and remobilization by trees: Ecophysiological relevance in a changing world Tree Physiology, 30, 1083-1095. [本文引用: 1]
PalacioS, CamareroJJ, MaestroM, AllaAQ, LahozE, Montserrat-MartíG (2018). Are storage and tree growth related? Seasonal nutrient and carbohydrate dynamics in evergreen and deciduous Mediterranean oaks Trees, 32, 777-790. [本文引用: 1]
PalacioS, MillaR, AlbuixechJ, Pérez-RontoméC, CamareroJJ, MaestroM, Montserrat-MartiG (2008). Seasonal variability of dry matter content and its relationship with shoot growth and nonstructural carbohydrates New Phytologist, 180, 133-142. [本文引用: 1]
Pérez-de-LisG, García-GonzálezI, RozasV, OlanoJM (2016). Feedbacks between earlywood anatomy and non-structural carbohydrates affect spring phenology and wood production in ring-porous oaks Biogeosciences, 13, 5499-5510. [本文引用: 1]
PiperFI (2011). Drought induces opposite changes in the concentration of non-structural carbohydrates of two evergreen Nothofagus species of differential drought resistance Annals of Forest Science, 68, 415-424. [本文引用: 1]
RichardsonAD, CarboneMS, KeenanTF, CzimczikCI, HollingerDY, MurakamiP, SchabergPG, XuXM (2013). Seasonal dynamics and age of stemwood nonstructural carbohydrates in temperate forest trees New Phytologist, 197, 850-861. [本文引用: 1]
SalaA, WoodruffDR, MeinzerFC (2012). Carbon dynamics in trees: Feast or famine? Tree Physiology, 32, 766-775. [本文引用: 1]
SavageJA, ClearwaterMJ, HainesDF, KleinT, MencucciniM, SevantoS, TurgeonR, ZhangC (2015). Allocation, stress tolerance and carbon transport in plants: How does phloem physiology affect plant ecology? Plant, Cell & Environment, 39, 709-725. [本文引用: 1]
Sch?delC, BlochlA, RichterA, HochG (2009). Short-term dynamics of nonstructural carbohydrates and hemicelluloses in young branches of temperate forest trees during bud break Tree Physiology, 29, 901-911. [本文引用: 2]
ShiP, K?rnerC, HochG (2006). End of season carbon supply status of woody species near the treeline in western China Basic and Applied Ecology, 7, 370-377. [本文引用: 1]
VerdaguerD, OjedaF (2002). Root starch storage and allocation patterns in seeder and resprouter seedlings of two Cape Erica ( Ericaceae) species American Journal of Botany, 89, 1189-1196. [本文引用: 1]
VoelkerSL, MeinzerFC, LachenbruchB, BrooksJR, GuyetteRP (2014). Drivers of radial growth and carbon isotope discrimination of bur oak ( Quercus macrocarpa Michx) across continental gradients in precipitation, vapour pressure deficit and irradiance Plant, Cell & Environment, 37, 766-779. [本文引用: 1]
WangB, JiangY, WangMC, DongMY, ZhangYP (2015). Variations of non-structural carbohydrate concentration of Picea meyeri at different elevations of Luya Mountain, China Chinese Journal of Plant Ecology, 39, 746-752. [本文引用: 2]
WangYR, ZhengCY, ZengFX (2016). Seasonal dynamic changes of non-structural carbohydrate in tissues of Picea mongolica in Baiyinaobao Acta Scientiarum Naturalium Universitatis Pekinensis, 52, 967-976. [本文引用: 2]
WongBL, BaggettKL, RyeAH (2003). Seasonal patterns of reserve and soluble carbohydrates in mature sugar maple (Acer saccharum) Canadian Journal of Botany, 81, 780-788. [本文引用: 1]
WürthMKR, Peláez-RiedlS, WrightSJ, K?rnerC (2005). Non-structural carbohydrate pools in a tropical forest Oecologia, 143, 11-24. [本文引用: 1]
ZhangHY (2013). Spatiotemporal Variability in Non-structural Carbohydrates of Stem and Branch of Twelve Tree Species in Chinese Temperate Forest. PhD dissertation, Northeast Forestry University, Haerbin. [本文引用: 2]
ZhangHY, WangCK, WangXC (2013). Comparison of concentrations of non-structural carbohydrates between new twigs and old branches for 12 temperate species Acta Ecologica Sinica, 33, 5675-5685. [本文引用: 1]
ZhangY, LiLY, ShuS, CuiGL (2013). Determination of soluble sugar and starch content in Pueraria lobata from different producing areas, varieties and harvest periods Journal of Chinese Medicinal Materials, 36, 1751-1754. [本文引用: 1]
ZhangYP, XuJL, SuW, ZhaoXP, XuXL (2019). Spring precipitation effects on formation of first row of earlywood vessels in Quercus variabilis at Qinling Mountain (China) Trees, 33, 457-468. [本文引用: 1]
ZhouYB, WuDD, YuDP, SuiCY (2009). Variations of nonstructural carbohydrate content in Betula ermanii at different elevations of Changbai Mountain, China Chinese Journal of Plant Ecology, 33, 118-124. [本文引用: 2]
ZhuWZ, CaoM, WangSG, XiaoWF, LiMH (2012). Seasonal dynamics of mobile carbon supply in Quercus aquifolioides at the upper elevational limit PLOS ONE, 7, e34213. DOI: 10.1371/journal.pone.0034213. [本文引用: 1]
ZrennerR, StittM (1991). Comparison of the effect of rapidly and gradually developing water-stress on carbohydrate metabolism in spinach leaves Plant, Cell & Environment, 14, 939-946. [本文引用: 1]
The enigma of the rise of angiosperms: Can we untie the knot? 1 2014
... 目前, 关于枝条NSC含量的研究仍存在以下不足: (1)较长的取样时间间隔(20-30天), 无法及时捕捉到春季展叶期枝条NSC含量的迅速变化(Hoch et al., 2003), 观测结果精度有待进一步提高.(2)仅观测萌芽前15-30天内枝条NSC含量, 缺乏对前一生长季末枝条NSC含量背景值的掌握.有研究表明, 前一年冬季树木NSC含量对春季叶片物候节律有直接影响(Pérez-de-Lis et al., 2016).因此掌握NSC含量背景值是有必要的, 这将有助于理解枝条NSC含量季节变化趋势及其与叶片物候的关系.(3)不少研究通过对比常绿针叶树和落叶阔叶树, 来分析叶片物候对枝条NSC含量的影响, 这一方法的合理性值得商榷(Millard & Grelet, 2010; Brüggemann et al., 2011).因为除叶片物候外, 不同树种自身的生理学、生物学特性亦存在较大差异, 将枝条NSC含量的变化笼统归因于叶片物候, 会直接降低结果的可信度(Augusto et al., 2014). ...
Carbon allocation and carbon isotope fluxes in the plant-soil-atmosphere continuum: A review 1 2011
... 目前, 关于枝条NSC含量的研究仍存在以下不足: (1)较长的取样时间间隔(20-30天), 无法及时捕捉到春季展叶期枝条NSC含量的迅速变化(Hoch et al., 2003), 观测结果精度有待进一步提高.(2)仅观测萌芽前15-30天内枝条NSC含量, 缺乏对前一生长季末枝条NSC含量背景值的掌握.有研究表明, 前一年冬季树木NSC含量对春季叶片物候节律有直接影响(Pérez-de-Lis et al., 2016).因此掌握NSC含量背景值是有必要的, 这将有助于理解枝条NSC含量季节变化趋势及其与叶片物候的关系.(3)不少研究通过对比常绿针叶树和落叶阔叶树, 来分析叶片物候对枝条NSC含量的影响, 这一方法的合理性值得商榷(Millard & Grelet, 2010; Brüggemann et al., 2011).因为除叶片物候外, 不同树种自身的生理学、生物学特性亦存在较大差异, 将枝条NSC含量的变化笼统归因于叶片物候, 会直接降低结果的可信度(Augusto et al., 2014). ...
河南省洛宁县全宝山国有林场植被的调查分析 1 2003
... 本研究的野外实验样地位于秦岭余脉熊耳山北坡, 豫西山区(河南省洛宁县西南部)的全宝山国有林场(34.10°-34.18° N, 111.38°-113.54° E).该研究区属于暖温带大陆性季风气候, 四季分明, 年平均气温10.5 ℃, 年降水量863.6 mm, 年蒸发量为1 562.8 mm, 全年无霜期186天.林场最高海拔2 100 m左右, 山地起伏较大, 坡度36°-50°, 土壤以棕壤为主.该区主要森林植被类型有壳斗科、槭树科、胡桃科等.栎属(Quercus)植物在低海拔(<1 000 m)地区常为优势种, 以次生林群落分布较多, 栓皮栎在此范围内常形成纯林(曹新向等, 2003). ...
河南省洛宁县全宝山国有林场植被的调查分析 1 2003
... 本研究的野外实验样地位于秦岭余脉熊耳山北坡, 豫西山区(河南省洛宁县西南部)的全宝山国有林场(34.10°-34.18° N, 111.38°-113.54° E).该研究区属于暖温带大陆性季风气候, 四季分明, 年平均气温10.5 ℃, 年降水量863.6 mm, 年蒸发量为1 562.8 mm, 全年无霜期186天.林场最高海拔2 100 m左右, 山地起伏较大, 坡度36°-50°, 土壤以棕壤为主.该区主要森林植被类型有壳斗科、槭树科、胡桃科等.栎属(Quercus)植物在低海拔(<1 000 m)地区常为优势种, 以次生林群落分布较多, 栓皮栎在此范围内常形成纯林(曹新向等, 2003). ...
Contribution of new photosynthetic assimilates to respiration by perennial grasses and shrubs: Residence times and allocation patterns 1 2007
... 枝条是碳供应器官(叶片)和碳需求器官(茎干、根)的连接者, 其NSC含量的季节变化与树木生长存在密切关系.因此, 枝条NSC含量的季节变化常常体现了树体的碳需求特征(Chapin et al., 1990; Kozlowski & Hattrup, 1992; Klein et al., 2016).研究表明, 枝条NSC含量的季节变化受生长型(乔木、灌木、藤本)(Carbone & Trumbore, 2007), 生活型(常绿、落叶)(Palacio et al., 2014)及环境因子(Martínez-Vilalta et al., 2016)等多因素的共同影响.已有研究发现, 在树体不同器官中, 叶片NSC含量存在明显的季节波动(Würth et al., 2005), 而茎干NSC含量相对稳定(张海燕, 2013), 但对枝条NSC含量的季节波动性结论存在较大分歧(Wong et al., 2003; 王逸然等, 2016).Hoch等(2003)研究指出, 虽然枝条NSC总含量(通常指淀粉和可溶性糖之和)的年内波动振幅在20% (Quercus petraea)到73% (Abies alba)之间, 但大多数温带落叶树种的枝条可溶性糖含量的季节变化并不显著, 仅在展叶期间有轻微的降低.因此, 枝条NSC总量和其组分的季节变化可能并不完全一致(Hoch, 2008; Klein et al., 2014).另一方面, 不少研究指出, 枝条NSC含量的季节变化与叶片物候有关, 因为枝条中存储的NSC常被用于春季树木发芽展叶(Palacio et al., 2008; Chen et al., 2017).张海燕等(2013)研究发现, 温带12个树种在展叶期间, 枝条NSC含量均明显下降, 与常绿树种相比, 落叶树种降幅更明显.因为春季展叶前, 落叶树种不具备进行光合作用的组织, 需要枝条提供春季萌芽所需的大量碳, 因此落叶树种枝条NSC含量常常在萌芽前下降, 在萌芽展叶后上升(Sch?del et al., 2009). ...
The ecology and economics of storage in plants 1 1990
... 枝条是碳供应器官(叶片)和碳需求器官(茎干、根)的连接者, 其NSC含量的季节变化与树木生长存在密切关系.因此, 枝条NSC含量的季节变化常常体现了树体的碳需求特征(Chapin et al., 1990; Kozlowski & Hattrup, 1992; Klein et al., 2016).研究表明, 枝条NSC含量的季节变化受生长型(乔木、灌木、藤本)(Carbone & Trumbore, 2007), 生活型(常绿、落叶)(Palacio et al., 2014)及环境因子(Martínez-Vilalta et al., 2016)等多因素的共同影响.已有研究发现, 在树体不同器官中, 叶片NSC含量存在明显的季节波动(Würth et al., 2005), 而茎干NSC含量相对稳定(张海燕, 2013), 但对枝条NSC含量的季节波动性结论存在较大分歧(Wong et al., 2003; 王逸然等, 2016).Hoch等(2003)研究指出, 虽然枝条NSC总含量(通常指淀粉和可溶性糖之和)的年内波动振幅在20% (Quercus petraea)到73% (Abies alba)之间, 但大多数温带落叶树种的枝条可溶性糖含量的季节变化并不显著, 仅在展叶期间有轻微的降低.因此, 枝条NSC总量和其组分的季节变化可能并不完全一致(Hoch, 2008; Klein et al., 2014).另一方面, 不少研究指出, 枝条NSC含量的季节变化与叶片物候有关, 因为枝条中存储的NSC常被用于春季树木发芽展叶(Palacio et al., 2008; Chen et al., 2017).张海燕等(2013)研究发现, 温带12个树种在展叶期间, 枝条NSC含量均明显下降, 与常绿树种相比, 落叶树种降幅更明显.因为春季展叶前, 落叶树种不具备进行光合作用的组织, 需要枝条提供春季萌芽所需的大量碳, 因此落叶树种枝条NSC含量常常在萌芽前下降, 在萌芽展叶后上升(Sch?del et al., 2009). ...
Phenology- ?dependent variation in the non-structural carbohydrates of broadleaf evergreen species plays an important role in determining tolerance to defoliation (or herbivory) 1 2017
... 枝条是碳供应器官(叶片)和碳需求器官(茎干、根)的连接者, 其NSC含量的季节变化与树木生长存在密切关系.因此, 枝条NSC含量的季节变化常常体现了树体的碳需求特征(Chapin et al., 1990; Kozlowski & Hattrup, 1992; Klein et al., 2016).研究表明, 枝条NSC含量的季节变化受生长型(乔木、灌木、藤本)(Carbone & Trumbore, 2007), 生活型(常绿、落叶)(Palacio et al., 2014)及环境因子(Martínez-Vilalta et al., 2016)等多因素的共同影响.已有研究发现, 在树体不同器官中, 叶片NSC含量存在明显的季节波动(Würth et al., 2005), 而茎干NSC含量相对稳定(张海燕, 2013), 但对枝条NSC含量的季节波动性结论存在较大分歧(Wong et al., 2003; 王逸然等, 2016).Hoch等(2003)研究指出, 虽然枝条NSC总含量(通常指淀粉和可溶性糖之和)的年内波动振幅在20% (Quercus petraea)到73% (Abies alba)之间, 但大多数温带落叶树种的枝条可溶性糖含量的季节变化并不显著, 仅在展叶期间有轻微的降低.因此, 枝条NSC总量和其组分的季节变化可能并不完全一致(Hoch, 2008; Klein et al., 2014).另一方面, 不少研究指出, 枝条NSC含量的季节变化与叶片物候有关, 因为枝条中存储的NSC常被用于春季树木发芽展叶(Palacio et al., 2008; Chen et al., 2017).张海燕等(2013)研究发现, 温带12个树种在展叶期间, 枝条NSC含量均明显下降, 与常绿树种相比, 落叶树种降幅更明显.因为春季展叶前, 落叶树种不具备进行光合作用的组织, 需要枝条提供春季萌芽所需的大量碳, 因此落叶树种枝条NSC含量常常在萌芽前下降, 在萌芽展叶后上升(Sch?del et al., 2009). ...
Earlywood vessels in ring-porous trees become functional for water transport after bud burst and before the maturation of the current-year leaves 1 2016
... 高低海拔枝条NSC含量在萌芽前后存在显著差异, 不同海拔叶片物候的时间差可能是造成这一现象的重要原因.通常认为, 枝条与芽的距离最近, 因此枝条很可能是春季芽萌发和叶生长所需的重要碳源(Palacio et al., 2018).有研究表明, 欧洲温带落叶树种如水青冈属(Fagus)在萌芽前枝条NSC含量会大幅度下降(Sch?del et al., 2009; Klein et al., 2016), 为萌芽展叶提供能量.本研究也发现, 栓皮栎高海拔枝条NSC含量分别在3月下旬(高海拔)和3月中旬(低海拔)明显降低.这一变化及其在高低海拔出现的时间差, 与叶片在高海拔4月初和低海拔3月底的展叶物候时间差是一致的(表4).这也解释了为何高低海拔枝条NSC含量的显著性差异仅出现在3月初和3月末(图3), 及高海拔枝条NSC含量仅在3月中旬高于低海拔.值得注意的是, Kitin和Funada (2016)研究指出, 环孔材栎类因其木质部形成层的启动时间早于萌芽, 因此木质部生长更依赖于树体(枝条、根部、茎干)存储的NSC.同为环孔材的栓皮栎, 其枝条NSC含量在萌芽前的下降, 是否也与木质部形成的生长启动有关, 亦或受萌芽和木质部生长的综合影响, 仍有待更进一步地全面研究. ...
Drought stress, growth and nonstructural carbohydrate dynamics of pine trees in a semi-arid forest 1 2014
... 枝条是碳供应器官(叶片)和碳需求器官(茎干、根)的连接者, 其NSC含量的季节变化与树木生长存在密切关系.因此, 枝条NSC含量的季节变化常常体现了树体的碳需求特征(Chapin et al., 1990; Kozlowski & Hattrup, 1992; Klein et al., 2016).研究表明, 枝条NSC含量的季节变化受生长型(乔木、灌木、藤本)(Carbone & Trumbore, 2007), 生活型(常绿、落叶)(Palacio et al., 2014)及环境因子(Martínez-Vilalta et al., 2016)等多因素的共同影响.已有研究发现, 在树体不同器官中, 叶片NSC含量存在明显的季节波动(Würth et al., 2005), 而茎干NSC含量相对稳定(张海燕, 2013), 但对枝条NSC含量的季节波动性结论存在较大分歧(Wong et al., 2003; 王逸然等, 2016).Hoch等(2003)研究指出, 虽然枝条NSC总含量(通常指淀粉和可溶性糖之和)的年内波动振幅在20% (Quercus petraea)到73% (Abies alba)之间, 但大多数温带落叶树种的枝条可溶性糖含量的季节变化并不显著, 仅在展叶期间有轻微的降低.因此, 枝条NSC总量和其组分的季节变化可能并不完全一致(Hoch, 2008; Klein et al., 2014).另一方面, 不少研究指出, 枝条NSC含量的季节变化与叶片物候有关, 因为枝条中存储的NSC常被用于春季树木发芽展叶(Palacio et al., 2008; Chen et al., 2017).张海燕等(2013)研究发现, 温带12个树种在展叶期间, 枝条NSC含量均明显下降, 与常绿树种相比, 落叶树种降幅更明显.因为春季展叶前, 落叶树种不具备进行光合作用的组织, 需要枝条提供春季萌芽所需的大量碳, 因此落叶树种枝条NSC含量常常在萌芽前下降, 在萌芽展叶后上升(Sch?del et al., 2009). ...
Coordination between growth, phenology and carbon storage in three coexisting deciduous tree species in a temperate forest 2 2016
... 枝条是碳供应器官(叶片)和碳需求器官(茎干、根)的连接者, 其NSC含量的季节变化与树木生长存在密切关系.因此, 枝条NSC含量的季节变化常常体现了树体的碳需求特征(Chapin et al., 1990; Kozlowski & Hattrup, 1992; Klein et al., 2016).研究表明, 枝条NSC含量的季节变化受生长型(乔木、灌木、藤本)(Carbone & Trumbore, 2007), 生活型(常绿、落叶)(Palacio et al., 2014)及环境因子(Martínez-Vilalta et al., 2016)等多因素的共同影响.已有研究发现, 在树体不同器官中, 叶片NSC含量存在明显的季节波动(Würth et al., 2005), 而茎干NSC含量相对稳定(张海燕, 2013), 但对枝条NSC含量的季节波动性结论存在较大分歧(Wong et al., 2003; 王逸然等, 2016).Hoch等(2003)研究指出, 虽然枝条NSC总含量(通常指淀粉和可溶性糖之和)的年内波动振幅在20% (Quercus petraea)到73% (Abies alba)之间, 但大多数温带落叶树种的枝条可溶性糖含量的季节变化并不显著, 仅在展叶期间有轻微的降低.因此, 枝条NSC总量和其组分的季节变化可能并不完全一致(Hoch, 2008; Klein et al., 2014).另一方面, 不少研究指出, 枝条NSC含量的季节变化与叶片物候有关, 因为枝条中存储的NSC常被用于春季树木发芽展叶(Palacio et al., 2008; Chen et al., 2017).张海燕等(2013)研究发现, 温带12个树种在展叶期间, 枝条NSC含量均明显下降, 与常绿树种相比, 落叶树种降幅更明显.因为春季展叶前, 落叶树种不具备进行光合作用的组织, 需要枝条提供春季萌芽所需的大量碳, 因此落叶树种枝条NSC含量常常在萌芽前下降, 在萌芽展叶后上升(Sch?del et al., 2009). ...
... 高低海拔枝条NSC含量在萌芽前后存在显著差异, 不同海拔叶片物候的时间差可能是造成这一现象的重要原因.通常认为, 枝条与芽的距离最近, 因此枝条很可能是春季芽萌发和叶生长所需的重要碳源(Palacio et al., 2018).有研究表明, 欧洲温带落叶树种如水青冈属(Fagus)在萌芽前枝条NSC含量会大幅度下降(Sch?del et al., 2009; Klein et al., 2016), 为萌芽展叶提供能量.本研究也发现, 栓皮栎高海拔枝条NSC含量分别在3月下旬(高海拔)和3月中旬(低海拔)明显降低.这一变化及其在高低海拔出现的时间差, 与叶片在高海拔4月初和低海拔3月底的展叶物候时间差是一致的(表4).这也解释了为何高低海拔枝条NSC含量的显著性差异仅出现在3月初和3月末(图3), 及高海拔枝条NSC含量仅在3月中旬高于低海拔.值得注意的是, Kitin和Funada (2016)研究指出, 环孔材栎类因其木质部形成层的启动时间早于萌芽, 因此木质部生长更依赖于树体(枝条、根部、茎干)存储的NSC.同为环孔材的栓皮栎, 其枝条NSC含量在萌芽前的下降, 是否也与木质部形成的生长启动有关, 亦或受萌芽和木质部生长的综合影响, 仍有待更进一步地全面研究. ...
Response of protein and carbohydrate metabolism of Scots pine seedlings to low temperature 1 2002
Dynamics of non-structural carbohydrates in terrestrial plants: A global synthesis 1 2016
... 枝条是碳供应器官(叶片)和碳需求器官(茎干、根)的连接者, 其NSC含量的季节变化与树木生长存在密切关系.因此, 枝条NSC含量的季节变化常常体现了树体的碳需求特征(Chapin et al., 1990; Kozlowski & Hattrup, 1992; Klein et al., 2016).研究表明, 枝条NSC含量的季节变化受生长型(乔木、灌木、藤本)(Carbone & Trumbore, 2007), 生活型(常绿、落叶)(Palacio et al., 2014)及环境因子(Martínez-Vilalta et al., 2016)等多因素的共同影响.已有研究发现, 在树体不同器官中, 叶片NSC含量存在明显的季节波动(Würth et al., 2005), 而茎干NSC含量相对稳定(张海燕, 2013), 但对枝条NSC含量的季节波动性结论存在较大分歧(Wong et al., 2003; 王逸然等, 2016).Hoch等(2003)研究指出, 虽然枝条NSC总含量(通常指淀粉和可溶性糖之和)的年内波动振幅在20% (Quercus petraea)到73% (Abies alba)之间, 但大多数温带落叶树种的枝条可溶性糖含量的季节变化并不显著, 仅在展叶期间有轻微的降低.因此, 枝条NSC总量和其组分的季节变化可能并不完全一致(Hoch, 2008; Klein et al., 2014).另一方面, 不少研究指出, 枝条NSC含量的季节变化与叶片物候有关, 因为枝条中存储的NSC常被用于春季树木发芽展叶(Palacio et al., 2008; Chen et al., 2017).张海燕等(2013)研究发现, 温带12个树种在展叶期间, 枝条NSC含量均明显下降, 与常绿树种相比, 落叶树种降幅更明显.因为春季展叶前, 落叶树种不具备进行光合作用的组织, 需要枝条提供春季萌芽所需的大量碳, 因此落叶树种枝条NSC含量常常在萌芽前下降, 在萌芽展叶后上升(Sch?del et al., 2009). ...
Nitrogen storage and remobilization by trees: Ecophysiological relevance in a changing world 1 2010
... 目前, 关于枝条NSC含量的研究仍存在以下不足: (1)较长的取样时间间隔(20-30天), 无法及时捕捉到春季展叶期枝条NSC含量的迅速变化(Hoch et al., 2003), 观测结果精度有待进一步提高.(2)仅观测萌芽前15-30天内枝条NSC含量, 缺乏对前一生长季末枝条NSC含量背景值的掌握.有研究表明, 前一年冬季树木NSC含量对春季叶片物候节律有直接影响(Pérez-de-Lis et al., 2016).因此掌握NSC含量背景值是有必要的, 这将有助于理解枝条NSC含量季节变化趋势及其与叶片物候的关系.(3)不少研究通过对比常绿针叶树和落叶阔叶树, 来分析叶片物候对枝条NSC含量的影响, 这一方法的合理性值得商榷(Millard & Grelet, 2010; Brüggemann et al., 2011).因为除叶片物候外, 不同树种自身的生理学、生物学特性亦存在较大差异, 将枝条NSC含量的变化笼统归因于叶片物候, 会直接降低结果的可信度(Augusto et al., 2014). ...
Are storage and tree growth related? Seasonal nutrient and carbohydrate dynamics in evergreen and deciduous Mediterranean oaks 1 2018
... 高低海拔枝条NSC含量在萌芽前后存在显著差异, 不同海拔叶片物候的时间差可能是造成这一现象的重要原因.通常认为, 枝条与芽的距离最近, 因此枝条很可能是春季芽萌发和叶生长所需的重要碳源(Palacio et al., 2018).有研究表明, 欧洲温带落叶树种如水青冈属(Fagus)在萌芽前枝条NSC含量会大幅度下降(Sch?del et al., 2009; Klein et al., 2016), 为萌芽展叶提供能量.本研究也发现, 栓皮栎高海拔枝条NSC含量分别在3月下旬(高海拔)和3月中旬(低海拔)明显降低.这一变化及其在高低海拔出现的时间差, 与叶片在高海拔4月初和低海拔3月底的展叶物候时间差是一致的(表4).这也解释了为何高低海拔枝条NSC含量的显著性差异仅出现在3月初和3月末(图3), 及高海拔枝条NSC含量仅在3月中旬高于低海拔.值得注意的是, Kitin和Funada (2016)研究指出, 环孔材栎类因其木质部形成层的启动时间早于萌芽, 因此木质部生长更依赖于树体(枝条、根部、茎干)存储的NSC.同为环孔材的栓皮栎, 其枝条NSC含量在萌芽前的下降, 是否也与木质部形成的生长启动有关, 亦或受萌芽和木质部生长的综合影响, 仍有待更进一步地全面研究. ...
Differential nitrogen cycling in semiarid sub-shrubs with contrasting leaf habit 1 2014
... 枝条是碳供应器官(叶片)和碳需求器官(茎干、根)的连接者, 其NSC含量的季节变化与树木生长存在密切关系.因此, 枝条NSC含量的季节变化常常体现了树体的碳需求特征(Chapin et al., 1990; Kozlowski & Hattrup, 1992; Klein et al., 2016).研究表明, 枝条NSC含量的季节变化受生长型(乔木、灌木、藤本)(Carbone & Trumbore, 2007), 生活型(常绿、落叶)(Palacio et al., 2014)及环境因子(Martínez-Vilalta et al., 2016)等多因素的共同影响.已有研究发现, 在树体不同器官中, 叶片NSC含量存在明显的季节波动(Würth et al., 2005), 而茎干NSC含量相对稳定(张海燕, 2013), 但对枝条NSC含量的季节波动性结论存在较大分歧(Wong et al., 2003; 王逸然等, 2016).Hoch等(2003)研究指出, 虽然枝条NSC总含量(通常指淀粉和可溶性糖之和)的年内波动振幅在20% (Quercus petraea)到73% (Abies alba)之间, 但大多数温带落叶树种的枝条可溶性糖含量的季节变化并不显著, 仅在展叶期间有轻微的降低.因此, 枝条NSC总量和其组分的季节变化可能并不完全一致(Hoch, 2008; Klein et al., 2014).另一方面, 不少研究指出, 枝条NSC含量的季节变化与叶片物候有关, 因为枝条中存储的NSC常被用于春季树木发芽展叶(Palacio et al., 2008; Chen et al., 2017).张海燕等(2013)研究发现, 温带12个树种在展叶期间, 枝条NSC含量均明显下降, 与常绿树种相比, 落叶树种降幅更明显.因为春季展叶前, 落叶树种不具备进行光合作用的组织, 需要枝条提供春季萌芽所需的大量碳, 因此落叶树种枝条NSC含量常常在萌芽前下降, 在萌芽展叶后上升(Sch?del et al., 2009). ...
Seasonal variability of dry matter content and its relationship with shoot growth and nonstructural carbohydrates 1 2008
... 枝条是碳供应器官(叶片)和碳需求器官(茎干、根)的连接者, 其NSC含量的季节变化与树木生长存在密切关系.因此, 枝条NSC含量的季节变化常常体现了树体的碳需求特征(Chapin et al., 1990; Kozlowski & Hattrup, 1992; Klein et al., 2016).研究表明, 枝条NSC含量的季节变化受生长型(乔木、灌木、藤本)(Carbone & Trumbore, 2007), 生活型(常绿、落叶)(Palacio et al., 2014)及环境因子(Martínez-Vilalta et al., 2016)等多因素的共同影响.已有研究发现, 在树体不同器官中, 叶片NSC含量存在明显的季节波动(Würth et al., 2005), 而茎干NSC含量相对稳定(张海燕, 2013), 但对枝条NSC含量的季节波动性结论存在较大分歧(Wong et al., 2003; 王逸然等, 2016).Hoch等(2003)研究指出, 虽然枝条NSC总含量(通常指淀粉和可溶性糖之和)的年内波动振幅在20% (Quercus petraea)到73% (Abies alba)之间, 但大多数温带落叶树种的枝条可溶性糖含量的季节变化并不显著, 仅在展叶期间有轻微的降低.因此, 枝条NSC总量和其组分的季节变化可能并不完全一致(Hoch, 2008; Klein et al., 2014).另一方面, 不少研究指出, 枝条NSC含量的季节变化与叶片物候有关, 因为枝条中存储的NSC常被用于春季树木发芽展叶(Palacio et al., 2008; Chen et al., 2017).张海燕等(2013)研究发现, 温带12个树种在展叶期间, 枝条NSC含量均明显下降, 与常绿树种相比, 落叶树种降幅更明显.因为春季展叶前, 落叶树种不具备进行光合作用的组织, 需要枝条提供春季萌芽所需的大量碳, 因此落叶树种枝条NSC含量常常在萌芽前下降, 在萌芽展叶后上升(Sch?del et al., 2009). ...
Feedbacks between earlywood anatomy and non-structural carbohydrates affect spring phenology and wood production in ring-porous oaks 1 2016
... 目前, 关于枝条NSC含量的研究仍存在以下不足: (1)较长的取样时间间隔(20-30天), 无法及时捕捉到春季展叶期枝条NSC含量的迅速变化(Hoch et al., 2003), 观测结果精度有待进一步提高.(2)仅观测萌芽前15-30天内枝条NSC含量, 缺乏对前一生长季末枝条NSC含量背景值的掌握.有研究表明, 前一年冬季树木NSC含量对春季叶片物候节律有直接影响(Pérez-de-Lis et al., 2016).因此掌握NSC含量背景值是有必要的, 这将有助于理解枝条NSC含量季节变化趋势及其与叶片物候的关系.(3)不少研究通过对比常绿针叶树和落叶阔叶树, 来分析叶片物候对枝条NSC含量的影响, 这一方法的合理性值得商榷(Millard & Grelet, 2010; Brüggemann et al., 2011).因为除叶片物候外, 不同树种自身的生理学、生物学特性亦存在较大差异, 将枝条NSC含量的变化笼统归因于叶片物候, 会直接降低结果的可信度(Augusto et al., 2014). ...
Drought induces opposite changes in the concentration of non-structural carbohydrates of two evergreen Nothofagus species of differential drought resistance 1 2011
Drivers of radial growth and carbon isotope discrimination of bur oak ( Quercus macrocarpa Michx) across continental gradients in precipitation, vapour pressure deficit and irradiance 1 2014
 ,1, 海旭莹1,2, 徐军亮
,1, 海旭莹1,2, 徐军亮 ,1,*, 吴文霞1, 曹鹏鹤1,3, 安文静4
,1,*, 吴文霞1, 曹鹏鹤1,3, 安文静4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT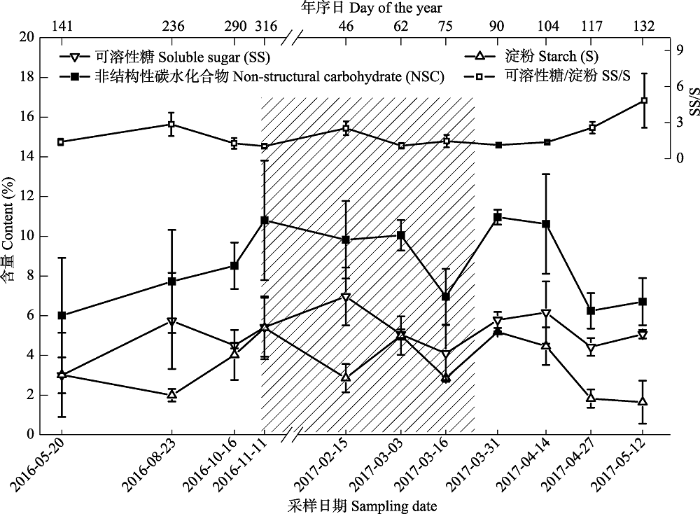 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT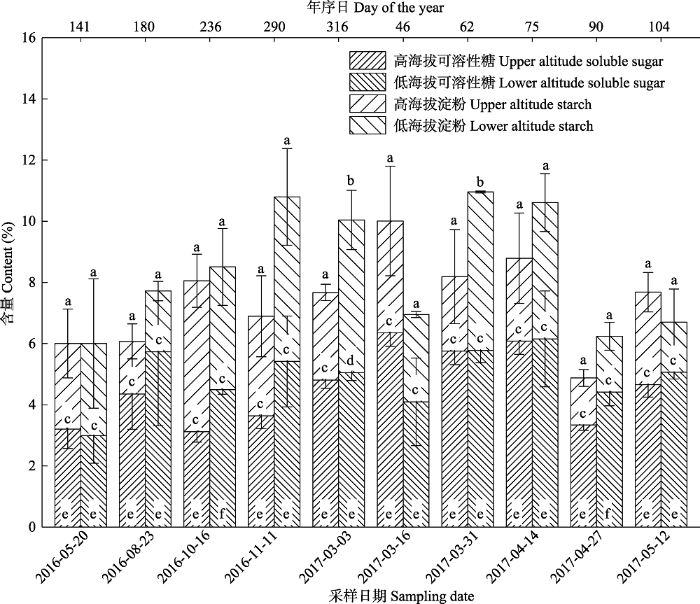 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT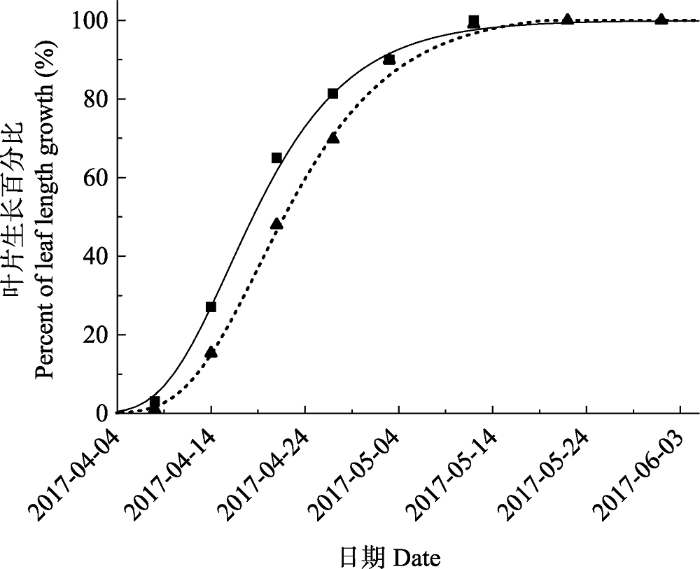 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}