Modeling phosphorus effects on the carbon cycle in terrestrial ecosystems
Mei HUANG,1,*, Na WANG1,2, Zhao-Sheng WANG1, He GONG11 Key Laboratory of Ecosystem Network Observation and Modeling, Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101, China 2 University of Chinese Academy of Sciences, Beijing 100049, China
Supported by the National Natural Science Foundation of China(41671101) the Strategic Priority Research Program for the Chinese Academy of Sciences(XDA23100202)
摘要 全球气候变暖已大大改变了陆地植物碳吸收能力, 提高了全球植被净初级生产力。随着气候变暖的加剧, 磷对植物生长的限制作用逐渐显现且不断增强, 磷影响陆地生态系统碳循环的机理和模型研究已成为研究热点。该文系统分析了磷影响陆地生态系统碳循环的相关机理以及模型对相关过程的定量化表达方法。综合对比分析了国际上的Carnegie- Ames-Stanford Approach-CNP (CASA-CNP)、Community Land Model-CNP (CLM-CNP)和Jena Scheme for Biosphere-Atmosphere Coupling in Hamburg-CNP (JSBACH-CNP)等碳、氮、磷耦合模型中磷影响植物光合作用与同化物分配过程、植物对磷的吸收过程、土壤中磷的转化过程以及生态系统磷输入与输出等过程的相关数学表达方法, 指出了模型算法的局限与不确定性以及未来模型发展与改进的方向。同时综合对比分析了CASA-CNP、CLM-CNP、JSBACH-CNP模型的基本特征, 总结了磷循环模型的建模方法, 为未来开展磷影响陆地生态系统碳循环的模型模拟研究提供了借鉴方法与参考思路。 关键词:磷限制;陆地生态系统;碳循环;过程模型;气候变化
Abstract Climate warming has significantly alerted the terrestrial carbon dynamics, resulting in enhanced vegetation productivity, especially in the northern hemisphere. However, most of the prior modeling studies have neglected the effects of nutrient availability, such as the phosphorus limitation, on carbon processes, which potentially leads to an overestimation of the capacity of terrestrial ecosystems to sequester additional carbon. Here, we reviewed recent progress in phosphorus limitation and its interactions with carbon dynamics in the context of climate change, with a focus on the process-based modeling approach. We comparatively analyzed quantitative representations of phosphorus-associated biological processes in some models (i.e., Carnegie-Ames-Stanford Approach (CASA), Community Land Model (CLM), and Jena Scheme for Biosphere-Atmosphere Coupling in Hamburg (JSBACH)), such as photosynthesis and distribution of assimilates, phosphorus uptake by plants, the transformation of phosphorus pools in soil, phosphorus inputs and outputs, etc. We also discussed the key characteristics of these models and summarized the mathematical representations of the terrestrial phosphorus cycle. In addition, we identified and discussed the limitations, uncertainties and future needs in process-based modeling in terms of nutrient and carbon dynamics. Our study highlighted the importance of including phosphorus limitation in regional carbon estimation and provided deep insights related to biogeochemical modeling at broad scales. Keywords:phosphorus limitation;terrestrial ecosystem;carbon cycle;process-based modeling;climate change
近年来, 随着全球变暖的加剧以及农田生态系统中磷肥的施入, 全球范围内植被净初级生产力普遍提高, 磷对植物生长的限制性作用逐渐加强(Goll et al., 2012; Sun et al., 2017)。因此, 能模拟磷循环的生物地球化学模型对于评估与预测全球碳循环至关重要。研究表明, 生物地球化学模型如果不考虑磷限制因素则可能高估植被的碳吸收(Zhang et al., 2014; Reed et al., 2015)。虽然早在2000年初开始出现模拟磷影响碳循环的模型(Wang et al., 2007), 但是目前国际上只有少数生物地球化学模型能够模拟磷循环。因此, 系统总结磷影响陆地生态系统碳循环过程及其模型表达方法, 一方面能提高我们对该领域研究态势和趋势的认识, 另一方面对我国生物地球化学模型的发展具有指导意义。
Carnegie-Ames-Stanford Approach-CNP (CASA- CNP)、Jena Scheme for Biosphere-Atmosphere Coupling in Hamburg-CNP (JSBACH-CNP)、Community Land Model-CNP (CLM-CNP)模型是目前国际上主流的能模拟碳、氮、磷循环的生物地球化学模型。Wang等(2010)基于CASA-CNP模型模拟了全球陆地生态系统中碳、氮、磷的空间分布, 发现磷的限制使得热带地区植被净初级生产力减少了20%。Goll等(2012)基于JSBACH-CNP模型模拟评估了磷对CO2浓度升高引起的陆地生态系统碳吸收的影响, 发现磷限制使得全球碳吸收减少了16%。Yang等(2014)基于CLM-CNP模型模拟得出热带与温带森林1970-1999年间的磷吸收速率为0.18-0.71 mg·m-2·a-1。上述过程模型在模拟评估磷对于陆地生态系统碳循环的影响方面取得了较大的进展。本文在分析磷参与生态系统碳循环的过程与机理的基础上, 系统总结了JSBACH-CNP、CASA-CNP、CLM- CNP模型中磷在生态系统中循环转化的定量表达方法, 提出了模型的构建原则和框架, 以期为未来发展与改进与磷相关的生物地球化学模型提供参考。
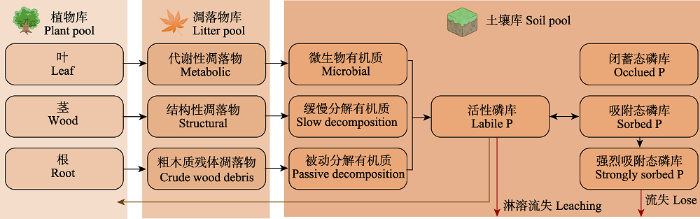
1 磷在土壤中的循环过程机理与模型表达
1.1 磷输入生态系统过程的机理与模型表达
大量的实验研究表明磷主要通过风化、沉降、地表径流等方式输入陆地生态系统, 其中, 风化磷是由生物和有机磷酸盐通过存储转化而产生的磷的释放, 土壤的风化基面是磷酸盐暂时的存储库, 岩性、土壤发育程度、径流、气温、降水、植被类型等是影响磷风化通量大小的主要因子(Goll et al., 2014; Hartmann et al., 2014)。虽然在某些地区沉降是磷输入生态系统的重要方式, 但通过沉降方式输入生态系统的磷可利用性相对较小(Pe?uelas et al., 2014), 影响磷沉降通量大小的因子相对复杂, 因此对其机理的研究还有较大的不确定性。有研究表明以沉降方式输入生态系统的磷通量与以风化方式输入的磷通量的量级大致相当(Wang et al., 2015)。径流是磷输入生态系统的途径之一, 但由于其输入数量相对较小, 对其机理的研究结果也具有很大的不确定性。
Table 1 表1 表1CASA-CNP、CLM-CNP和JSBACH-CNP模型的主要构架对比分析 Table 1Comparison of phosphorus processes in CASA-CNP, CLM-CNP, and JSBACH-CNP
模型 Model
磷库数量 Number of phosphate pools
时间步长 Time step
碳磷比参数 C:P ratio parameters
矿化过程模拟 Simulation of the mineralization process
适用范围 Scope of application
CASA-CNP
12
1 d
不同植被类型、植物不同器官具有不同的碳磷比 C:P ratios vary among different organs in various vegetation types
只量化了生物化学矿化过程 Only consider the biochemical mineralization process
温带和热带森林生态系统 温带和热带草原生态系统 Temperate and tropical forest and grassland ecosystems
CLM-CNP
15
30 min
不同植被类型、植物不同器官具有不同的碳磷比 C:P ratios vary among different organs in various vegetation types
模拟了生物矿化与生物化学矿化两个过程 Consider both biomineralization and biochemical mineralization
热带森林生态系统 热带草原生态系统 Tropical forest and grassland ecosystems
JSBACH-CNP
8
1 d
不同植被类型具有不同碳磷比, 但植物不同器官的碳磷比相同 C:P ratios are the same for organs but vary among various vegetation types
模拟了生物矿化与生物化学矿化两个过程 Consider both biomineralization and biochemical mineralization
温带和热带森林生态系统 温带和热带草原生态系统 Temperate and tropical forest and grassland ecosystems
CASA-CNP, Carnegie-Ames-Stanford Approach-CNP; JSBACH-CNP, Jena Scheme for Biosphere-Atmosphere Coupling in Hamburg-CNP; CLM-CNP, Community Land Model-CNP.
AchatDL, BakkerMR, MorelC (2009). Process-based assessment of phosphorus availability in a low phosphorus sorbing forest soil using isotopic dilution methods Soil Science Society of America Journal, 73, 2131-2142. [本文引用: 1]
AertsR, Chapin IIIFS (2000). The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns Advances in Ecological Research, 30, 1-67. [本文引用: 1]
AtkinOK, BloomfieldKJ, ReichPB, TjoelkerMG, AsnerGP, BonalD, B?nischG, BradfordMG, CernusakLA, CosioEG (2015). Global variability in leaf respiration in relation to climate, plant functional types and leaf traits New Phytologist, 206, 614-636. [本文引用: 2]
BattiniF, Gr?nlundM, AgnolucciM, GiovannettiM, JakobsenI (2017). Facilitation of phosphorus uptake in maize plants by mycorrhizosphere bacteria Scientific Reports, 7, 4686. DOI: 10.1038/s41598-017-04959-0. [本文引用: 1]
BenderL, StiebelingB, NeumannKH (1986). Investigations on photosynthesis and assimilate translocation in Daucus carota L. as influenced by a varied phosphorus supply and changes in the endogenous hormonal system following GA3 treatments Journal of Plant Nutrition and Soil Science, 149, 533-540. [本文引用: 1]
BowmanRA, ColeCV (1978). An exploratory method for fractionation of organic phosphorus from grassland soils Soil Science, 125, 95-101. [本文引用: 1]
CampbellLB, RaczGJ (1975). Organic and inorganic P content, movement and mineralization of P in soil beneath a feedlot Canadian Journal of Soil Science, 55, 457-466. [本文引用: 1]
DavidsonEA, JanssensIA (2006). Temperature sensitivity of soil carbon decomposition and feedbacks to climate change Nature, 440, 165-173. [本文引用: 1]
de GraaffMA, van GroenigenKJ, SixJ, HungateB, van KesselC (2006). Interactions between plant growth and soil nutrient cycling under elevated CO2: A meta-analysis Global Change Biology, 12, 2077-2091. [本文引用: 1]
DeepikaS, KothamasiD (2015). Soil moisture—A regulator of arbuscular mycorrhizal fungal community assembly and symbiotic phosphorus uptake Mycorrhiza, 25, 67-75. [本文引用: 1]
DominguesTF, MeirP, FeldpauschTR, SaizG, VeenendaalEM, SchrodtF, BirdM, DjagbleteyG, HienF, CompaoreH, DialloA, GraceJ, LloydJ (2010). Co-limitation of photosynthetic capacity by nitrogen and phosphorus in West Africa woodlands Plant, Cell & Environment, 33, 959-980. [本文引用: 1]
DongXB, HaoMD, GuoSA, ShiXJ, MaT, LiuGS (2014). The effects of nitrogen fertilizers and phosphate fertilizer rates on the yield, nutrient uptake and quality of Leymus chinensis Acta Agrestia Sinica, 22, 1232-1238. [本文引用: 1]
EllsworthD, CrousKY, LambersH, CookeJ (2015). Phosphorus recycling in photorespiration maintains high photosynthetic capacity in woody species Plant, Cell & Environment, 38, 1142-1156. [本文引用: 2]
Fernández-MartínezM, ViccaS, JanssensIA, SardansJ, LuyssaertS, CampioliM, Chapin IIIFS, CiaisP, MalhiY, ObersteinerM, PapaleD, PiaoSL, ReichsteinM, RodàF, Pe?uelasJ (2014). Nutrient availability as the key regulator of global forest carbon balance Nature Climate Change, 4, 471-476. [本文引用: 1]
F?llmiKB (1996). The phosphorus cycle, phosphogenesis and marine phosphate-rich deposits Earth-Science Reviews, 40, 55-124. [本文引用: 1]
FredeenAL, Madhusudana RaoI, TerryN (2018). Influence of phosphorus nutrition on growth and carbon partitioning in Glycine max Plant Physiology, 89, 225-230. [本文引用: 1]
GhannoumO (2008). C4 photosynthesis and water stress Annals of Botany, 103, 635-644. [本文引用: 1]
GollDS, BrovkinV, ParidaBR, ReickCH, KattgeJ, ReichPB, van BodegomPM, Niinemetsü (2012). Nutrient limitation reduces land carbon uptake in simulations with a model of combined carbon, nitrogen and phosphorus cycling Biogeosciences, 9, 3547-3569. [本文引用: 2]
GollDS, MoosdorfN, HartmannJ, BrovkinV (2014). Climate driven changes in chemical weathering and associated phosphorus release since 1850: Implications for the land carbon balance Geophysical Research Letters, 41, 3553-3558. [本文引用: 1]
HartmannJ, MoosdorfN, LauerwaldR, HindererM, WestAJ (2014). Global chemical weathering and associated P-release—The role of lithology, temperature and soil properties Chemical Geology, 363, 145-163. [本文引用: 1]
JiangJ, GuoS, ZhangY, LiuQ, WangR, WangZ, LiN, LiR (2015). Changes in temperature sensitivity of soil respiration in the phases of a three-year crop rotation system Soil & Tillage Research, 150, 139-146. [本文引用: 2]
JohnsonAH, FrizanoJ, VannDR (2003). Biogeochemical implications of labile phosphorus in forest soils determined by the Hedley fractionation procedure Oecologia, 135, 487-499. [本文引用: 1]
JungkA (2001). Root hairs and the acquisition of plant nutrients from soil Journal of Plant Nutrition and Soil Science, 164, 121-129. [本文引用: 1]
LiL, HuangM, GuFX, ZhangL (2013). The modeling algorithms for the effects of nitrogen on terrestrial vegetation carbon cycle process Journal of Natural Resources, 28, 2012-2022. [本文引用: 1]
LiuC, WangY, WangN, WangGX (2012). Advances research in plant nitrogen, phosphorus and stoichiometry in terrestrial ecosystems: A review Chinese Journal of Plant Ecology, 36, 1205-1216. [本文引用: 1]
LloydJ, BirdMI, VeenendaalEM, KruijtB (2001). Global Biogeochemical Cycles in the Climate System. Academic Press, London. 96-144. [本文引用: 2]
LuSY, LiuXX, LiK, GaoSH, JiaJL, YangGY (2016). Phosphorus removal by ecological planting tank treating rainwater runoff Chinese Journal of Environmental Engineering, 10, 3434-3438. [本文引用: 1]
[ 卢少勇, 刘学欣, 李珂, 高硕晗, 贾建丽, 杨光亚 (2016). 模拟生态种植槽去除雨水径流中的磷 环境工程学报, 10, 3434-3438.], Kubilay N, Losno R, Luo C, Maenhaut U McGee KA, Okin GS, Siefert RL Tsukuda. [本文引用: 1]
MahowaldN, JickellsTD, BakerAR, ArtaxoP, Benitez-NelsonCR, BergamettiG, BondTC, ChenY, CohenDD, HerutB, KubilayN, LosnoR, LuoC, MaenhautW, McGeeKA, OkinGS, SiefertRL, TsukudS (2008). Global distribution of atmospheric phosphorus sources, concentrations and deposition rates, and anthropogenic impacts Global Biogeochemical Cycles, 22, GB4026. DOI: 10.1029/2008GB003240. [本文引用: 3]
McgroddyME, DaufresneT, HedinLO (2004). Scaling of C:N:P stoichiometry in forests worldwide: Implications of terrestrial Redfield-type ratios Ecology, 85, 2390-2401. [本文引用: 2]
MeirP, GraceJ, MirandaAC (2001). Leaf respiration in two tropical rainforests: Constraints on physiology by phosphorus, nitrogen and temperature Functional Ecology, 15, 378-387. [本文引用: 1]
MonteithJL, MossCJ (1977). Climate and the efficiency of crop production in Britain Philosophical Transactions of the Royal Society B: Biological Sciences, 281, 277-294. [本文引用: 1]
NewmanEI (1995). Phosphorus inputs to terrestrial ecosystems Journal of Ecology, 83, 713-726. [本文引用: 1]
NiuYF, ChaiRS, JinGL, WangH, TangCX, ZhangYS (2013). Responses of root architecture development to low phosphorus availability: A review Annals of Botany, 112, 391-408. [本文引用: 2]
NorbyRJ, GuL, HaworthIC, JensenAM, TurnerBL, WalkerAP, WarrenJM, WestonDJ, XuC, WinterK (2017). Informing models through empirical relationships between foliar phosphorus, nitrogen and photosynthesis across diverse woody species in tropical forests of Panama New Phytologist, 215, 1425-1437. [本文引用: 1]
NorbyRJ, de KauweMG, DominguesTF, DuursmaRA, EllsworthDS, GollDS, LapolaDM, LuusKA, MackenzieAR, MedlynBE, PavlickR, RammingA, SmithB, ThomasR, ThonickeK, WalkerAP, YangX, ZaehleS (2015). Model-data synthesis for the next generation of forest free-air CO2 enrichment (FACE) experiments New Phytologist, 209, 17-28. [本文引用: 1]
NorbyRJ, WarrenJM, IversenCM, MedlynBE, McMurtrieRE (2010). CO2 enhancement of forest productivity constrained by limited nitrogen availability Proceedings of the National Academy of Sciences of the United States of America, 107, 19368-19373. [本文引用: 1]
PierrouU (1976). The global phosphorus cycle Ecological Bulletins, 48(22), 75-88. [本文引用: 1]
PlaxtonWC, PodestáFE (2006). The functional organization and control of plant respiration Critical Reviews in Plant Sciences, 25, 159-198. [本文引用: 1]
PoteDH, DanielTC (1996). Relating extractable soil phosphorus to phosphorus losses in runoff Soil Science Society of America Journal, 60, 855-859. [本文引用: 1]
ReedSC, YangX, ThorntonPE (2015). Incorporating phosphorus cycling into global modeling efforts: A worthwhile, tractable endeavor New Phytologist, 208, 324-329. [本文引用: 1]
ReichPB, OleksynJ (2004). Global patterns of plant leaf N and P in relation to temperature and latitude Proceedings of the National Academy of Sciences of the United States of America, 101, 11001-11006. [本文引用: 1]
ScottJT, CondronLM (2005). Short term effects of radiata pine and selected pasture species on soil organic phosphorus mineralisation Plant and Soil, 266, 153-163. [本文引用: 1]
SlotM, Rey-SánchezC, WinterK, KitajimaK (2014). Trait-based scaling of temperature-dependent foliar respiration in a species-rich tropical forest canopy Functional Ecology, 28, 1074-1086. [本文引用: 1]
SmilV (2000). Phosphorus in the environment: Natural flows and human interferences Annual Review of Energy and the Environment, 25, 53-88. [本文引用: 1]
SpohnM, Zavi?i?A, NassalP, BergkemperF, SchulzS, MarhanS, SchloterM, KandelerE, PolleA (2018). Temporal variations of phosphorus uptake by soil microbial biomass and young beech trees in two forest soils with contrasting phosphorus stocks Soil Biology & Biochemistry, 117, 191-202. [本文引用: 1]
StittM, HurryV (2002). A plant for all seasons: Alterations in photosynthetic carbon metabolism during cold acclimation in Arabidopsis Current Opinion in Plant Biology, 5, 199-206. [本文引用: 1]
ThomasDS, MontaguKD, ConroyJP (2006). Leaf inorganic phosphorus as a potential indicator of phosphorus status, photosynthesis and growth of Eucalyptus grandis seedlings Forest Ecology and Management, 223, 267-274. [本文引用: 1]
TurnerBL, MahieuN, CondronLM (2003). Phosphorus-31 nuclear magnetic resonance spectral assignments of phosphorus compounds in soil NaOH-EDTA extracts Soil Science Society of America Journal, 67, 497-510. [本文引用: 1]
UshioM, FujikiY, HidakaA, KitayamaK (2015). Linkage of root physiology and morphology as an adaptation to soil phosphorus impoverishment in tropical montane forests Functional Ecology, 29, 1235-1245. [本文引用: 1]
van WijkMT, WilliamsM, GoughL, HobbieSE, ShaverGR (2003). Luxury consumption of soil nutrients: A possible competitive strategy in above-ground and below-ground biomass allocation and root morphology for slow-growing arctic vegetation? Journal of Ecology, 91, 664-676. [本文引用: 1]
WanS, HuiD, WallaceL, LuoY (2005). Direct and indirect effects of experimental warming on ecosystem carbon processes in a tallgrass prairie Global Biogeochemical Cycles, 19, GB2014. DOI: 10.1029/2004GB002315. [本文引用: 1]
WangF, SongMH, HuangM, ZhangJS (2014). The spatial distribution of soil nutrients and the controlling factors of temperate forest and steppe in northeastern China Ecology and Environmental Sciences, 23, 1280-1285. [本文引用: 1]
WangR, BalkanskiY, BoucherO, CiaisP, Pe?uelasJ, TaoS (2015). Significant contribution of combustion-related emissions to the atmospheric phosphorus budget Nature Geoscience, 8, 48-54. [本文引用: 1]
WangR, SunQQ, WangY, LiuQF, DuLL, ZhaoM, GaoX, HuYX, GuoSL (2017). Temperature sensitivity of soil respiration: Synthetic effects of nitrogen and phosphorus fertilization on Chinese Loess Plateau Science of the Total Environment, 574, 1665-1673. [本文引用: 1]
WangYP, HoultonBZ, FieldCB (2007). A model of biogeochemical cycles of carbon, nitrogen, and phosphorus including symbiotic nitrogen fixation and phosphatase production Global Biogeochemical Cycles, 21, GB1018. DOI: 10.1029/2006GB002797. [本文引用: 1]
WangYP, LawRM, PakB (2010). A global model of carbon, nitrogen and phosphorus cycles for the terrestrial biosphere Biogeosciences, 7, 2261-2282. [本文引用: 4]
YanXF, LiuYY, GuoQW, HuangDW (2016). Nitrogen and phosphorus elimination by composited aggregate Bahia grass-planting concrete Chinese Journal of Environmental Engineering, 10, 1171-1176. [本文引用: 1]
YangX, ThorntonPE, RicciutoDM, PostWM (2014). The role of phosphorus dynamics in tropical forests—A modeling study using CLM-CNP Biogeosciences, 11, 1667-1681. [本文引用: 2]
ZhangDS, ZhangCC, TangXY, LiHG, ZhangFS, RengelZ, WhalleyWR, DaviesWJ, ShenJB (2016). Increased soil phosphorus availability induced by faba bean root exudation stimulates root growth and phosphorus uptake in neighbouring maize New Phytologist, 209, 823-831. [本文引用: 1]
ZhangQ, WangYP, MatearRJ, PitmanAJ, DaiYJ (2014). Nitrogen and phosphorous limitations significantly reduce future allowable CO2 emissions Geophysical Research Letters, 41, 632-637. [本文引用: 1]
ZhangYM, ZhouGS (2012). Primary simulation on the response of leaf maximum carboxylation rate to multiple environmental factors Chinese Science Bulletin, 57, 1112-1118. [本文引用: 1]
ZhouZH, WangCK (2016). Changes of the relationships between soil and microbes in carbon, nitrogen and phosphorus stoichiometry during ecosystem succession Chinese Journal of Plant Ecology, 40, 1257-1266. [本文引用: 1]
Facilitation of phosphorus uptake in maize plants by mycorrhizosphere bacteria 1 2017
... 根是植物体从土壤中吸收磷的重要器官, 植物根对磷的获取能力影响着植物体生长发育的全过程(Ushio et al., 2015; Battini et al., 2017), 而影响植物根部对磷吸收的主要因素分为两类, 一类为植物体生理特征, 如根系的形态(廖红等, 2001)、根的结构(严小龙等, 2000)、根毛(Jungk, 2001)等均影响根部吸收速率(Zhang et al., 2016); 另一类为根部土壤环境, 如土壤中磷酸酶(酸性磷酸酶APA)的含量、微生物的丰富度(Spohn et al., 2018)、菌群、水分(Deepika & Kothamasi, 2015)等.同时, 不同的物种与不同的气候环境下根部的磷吸收速率也存在差异.总体而言, 植物可以通过根系的生长、形成根毛及菌根等方式提高土壤磷的利用效率. ...
Investigations on photosynthesis and assimilate translocation in Daucus carota L. as influenced by a varied phosphorus supply and changes in the endogenous hormonal system following GA3 treatments 1 1986
Nutrient limitation reduces land carbon uptake in simulations with a model of combined carbon, nitrogen and phosphorus cycling 2 2012
... 近年来, 随着全球变暖的加剧以及农田生态系统中磷肥的施入, 全球范围内植被净初级生产力普遍提高, 磷对植物生长的限制性作用逐渐加强(Goll et al., 2012; Sun et al., 2017).因此, 能模拟磷循环的生物地球化学模型对于评估与预测全球碳循环至关重要.研究表明, 生物地球化学模型如果不考虑磷限制因素则可能高估植被的碳吸收(Zhang et al., 2014; Reed et al., 2015).虽然早在2000年初开始出现模拟磷影响碳循环的模型(Wang et al., 2007), 但是目前国际上只有少数生物地球化学模型能够模拟磷循环.因此, 系统总结磷影响陆地生态系统碳循环过程及其模型表达方法, 一方面能提高我们对该领域研究态势和趋势的认识, 另一方面对我国生物地球化学模型的发展具有指导意义. ...
... Carnegie-Ames-Stanford Approach-CNP (CASA- CNP)、Jena Scheme for Biosphere-Atmosphere Coupling in Hamburg-CNP (JSBACH-CNP)、Community Land Model-CNP (CLM-CNP)模型是目前国际上主流的能模拟碳、氮、磷循环的生物地球化学模型.Wang等(2010)基于CASA-CNP模型模拟了全球陆地生态系统中碳、氮、磷的空间分布, 发现磷的限制使得热带地区植被净初级生产力减少了20%.Goll等(2012)基于JSBACH-CNP模型模拟评估了磷对CO2浓度升高引起的陆地生态系统碳吸收的影响, 发现磷限制使得全球碳吸收减少了16%.Yang等(2014)基于CLM-CNP模型模拟得出热带与温带森林1970-1999年间的磷吸收速率为0.18-0.71 mg·m-2·a-1.上述过程模型在模拟评估磷对于陆地生态系统碳循环的影响方面取得了较大的进展.本文在分析磷参与生态系统碳循环的过程与机理的基础上, 系统总结了JSBACH-CNP、CASA-CNP、CLM- CNP模型中磷在生态系统中循环转化的定量表达方法, 提出了模型的构建原则和框架, 以期为未来发展与改进与磷相关的生物地球化学模型提供参考. ...
Climate driven changes in chemical weathering and associated phosphorus release since 1850: Implications for the land carbon balance 1 2014
... 大量的实验研究表明磷主要通过风化、沉降、地表径流等方式输入陆地生态系统, 其中, 风化磷是由生物和有机磷酸盐通过存储转化而产生的磷的释放, 土壤的风化基面是磷酸盐暂时的存储库, 岩性、土壤发育程度、径流、气温、降水、植被类型等是影响磷风化通量大小的主要因子(Goll et al., 2014; Hartmann et al., 2014).虽然在某些地区沉降是磷输入生态系统的重要方式, 但通过沉降方式输入生态系统的磷可利用性相对较小(Pe?uelas et al., 2014), 影响磷沉降通量大小的因子相对复杂, 因此对其机理的研究还有较大的不确定性.有研究表明以沉降方式输入生态系统的磷通量与以风化方式输入的磷通量的量级大致相当(Wang et al., 2015).径流是磷输入生态系统的途径之一, 但由于其输入数量相对较小, 对其机理的研究结果也具有很大的不确定性. ...
Global chemical weathering and associated P-release—The role of lithology, temperature and soil properties 1 2014
... 大量的实验研究表明磷主要通过风化、沉降、地表径流等方式输入陆地生态系统, 其中, 风化磷是由生物和有机磷酸盐通过存储转化而产生的磷的释放, 土壤的风化基面是磷酸盐暂时的存储库, 岩性、土壤发育程度、径流、气温、降水、植被类型等是影响磷风化通量大小的主要因子(Goll et al., 2014; Hartmann et al., 2014).虽然在某些地区沉降是磷输入生态系统的重要方式, 但通过沉降方式输入生态系统的磷可利用性相对较小(Pe?uelas et al., 2014), 影响磷沉降通量大小的因子相对复杂, 因此对其机理的研究还有较大的不确定性.有研究表明以沉降方式输入生态系统的磷通量与以风化方式输入的磷通量的量级大致相当(Wang et al., 2015).径流是磷输入生态系统的途径之一, 但由于其输入数量相对较小, 对其机理的研究结果也具有很大的不确定性. ...
Changes in temperature sensitivity of soil respiration in the phases of a three-year crop rotation system 2 2015
Informing models through empirical relationships between foliar phosphorus, nitrogen and photosynthesis across diverse woody species in tropical forests of Panama 1 2017
... 土壤与叶片中的磷均影响植物光合作用过程.实验表明, 土壤中以无机磷为主导的多种形态的磷, 不仅会直接影响光合作用的能力和速率, 而且也会间接地影响着植物叶片中磷的浓度(Ellsworth et al., 2015).一方面, 土壤中的无机磷可以被植物体吸收, 其不仅充当光合作用的基底物, 而且是形成光合作用的生物分子的重要元素(Reich & Oleksy, 2004).另一方面, 土壤中的无机磷影响着二磷酸核酮糖羧化酶的形成, 从而影响着光合作用的速率(Norby et al., 2015, 2017).同样, 植物叶片中磷的浓度与植物叶片光合速率的相关性十分显著, 如果叶片中磷供应不足会影响磷酸甘油酸的还原和再生(Fredeen et al., 2018), 降低叶绿体间质的pH值以及参与卡尔文循环的各种酶的活性, 最终影响光合作用的强度、羧化速率等(Lloyd et al., 2001; Wang et al., 2017). ...
Model-data synthesis for the next generation of forest free-air CO2 enrichment (FACE) experiments 1 2015
... 土壤与叶片中的磷均影响植物光合作用过程.实验表明, 土壤中以无机磷为主导的多种形态的磷, 不仅会直接影响光合作用的能力和速率, 而且也会间接地影响着植物叶片中磷的浓度(Ellsworth et al., 2015).一方面, 土壤中的无机磷可以被植物体吸收, 其不仅充当光合作用的基底物, 而且是形成光合作用的生物分子的重要元素(Reich & Oleksy, 2004).另一方面, 土壤中的无机磷影响着二磷酸核酮糖羧化酶的形成, 从而影响着光合作用的速率(Norby et al., 2015, 2017).同样, 植物叶片中磷的浓度与植物叶片光合速率的相关性十分显著, 如果叶片中磷供应不足会影响磷酸甘油酸的还原和再生(Fredeen et al., 2018), 降低叶绿体间质的pH值以及参与卡尔文循环的各种酶的活性, 最终影响光合作用的强度、羧化速率等(Lloyd et al., 2001; Wang et al., 2017). ...
CO2 enhancement of forest productivity constrained by limited nitrogen availability 1 2010
... 磷(P)是植物体内有机物的组成元素, 广泛参与植物有机体的代谢过程(刘超等, 2012), 与氮(N)一起通过影响植物的光合作用、呼吸作用和土壤有机质分解等一系列过程实现与碳氮循环过程的耦合(李雷等, 2013; 卢少勇等, 2016).在地史时期, 生物圈的活动形成了磷在全球范围内的大循环, 磷的循环与转化在重建气候、环境、生态以及全球变化的长期反馈机制中有着重要的作用(F?llmi, 1996).一方面, 地球生命从起源开始就依赖磷, 磷是所有生命形式的基本元素, 参与生物体的遗传物质与能量物质的转化, 是构成许多生命大分子(核酸、磷脂、二磷酸腺苷(ADP)、三磷酸腺苷(ATP)等)的关键元素, 在整个生态系统内能量的贮存、迁移、转化过程中发挥着重要的作用(Pierrou, 1976; 赵琼和曾德慧, 2005; Achat et al., 2009; Wang et al., 2010).另一方面, 众多研究表明磷的有效性强烈地影响着陆地生态系统的碳循环(de Graaff et al., 2006; Norby et al., 2010; Fernández-Martínez et al., 2014): 光合作用产生的糖类如果不经过磷酸化, 碳的固定则是无效的; 磷也会加强光合作用过程中碳水化合物的合成与运输; 在植物与土壤中, 磷与碳、氮的化学计量关系不仅会影响着植物体内碳的积累与分配(周正虎和王传宽, 2016), 还决定着全球陆地生态系统功能以及碳的源与汇(Aerts & Chapin III, 2000; van Wijk et al., 2003; Wan et al., 2005). ...
Biogenic volatile emissions from the soil 1 2014
... 大量的实验研究表明磷主要通过风化、沉降、地表径流等方式输入陆地生态系统, 其中, 风化磷是由生物和有机磷酸盐通过存储转化而产生的磷的释放, 土壤的风化基面是磷酸盐暂时的存储库, 岩性、土壤发育程度、径流、气温、降水、植被类型等是影响磷风化通量大小的主要因子(Goll et al., 2014; Hartmann et al., 2014).虽然在某些地区沉降是磷输入生态系统的重要方式, 但通过沉降方式输入生态系统的磷可利用性相对较小(Pe?uelas et al., 2014), 影响磷沉降通量大小的因子相对复杂, 因此对其机理的研究还有较大的不确定性.有研究表明以沉降方式输入生态系统的磷通量与以风化方式输入的磷通量的量级大致相当(Wang et al., 2015).径流是磷输入生态系统的途径之一, 但由于其输入数量相对较小, 对其机理的研究结果也具有很大的不确定性. ...
The global phosphorus cycle 1 1976
... 磷(P)是植物体内有机物的组成元素, 广泛参与植物有机体的代谢过程(刘超等, 2012), 与氮(N)一起通过影响植物的光合作用、呼吸作用和土壤有机质分解等一系列过程实现与碳氮循环过程的耦合(李雷等, 2013; 卢少勇等, 2016).在地史时期, 生物圈的活动形成了磷在全球范围内的大循环, 磷的循环与转化在重建气候、环境、生态以及全球变化的长期反馈机制中有着重要的作用(F?llmi, 1996).一方面, 地球生命从起源开始就依赖磷, 磷是所有生命形式的基本元素, 参与生物体的遗传物质与能量物质的转化, 是构成许多生命大分子(核酸、磷脂、二磷酸腺苷(ADP)、三磷酸腺苷(ATP)等)的关键元素, 在整个生态系统内能量的贮存、迁移、转化过程中发挥着重要的作用(Pierrou, 1976; 赵琼和曾德慧, 2005; Achat et al., 2009; Wang et al., 2010).另一方面, 众多研究表明磷的有效性强烈地影响着陆地生态系统的碳循环(de Graaff et al., 2006; Norby et al., 2010; Fernández-Martínez et al., 2014): 光合作用产生的糖类如果不经过磷酸化, 碳的固定则是无效的; 磷也会加强光合作用过程中碳水化合物的合成与运输; 在植物与土壤中, 磷与碳、氮的化学计量关系不仅会影响着植物体内碳的积累与分配(周正虎和王传宽, 2016), 还决定着全球陆地生态系统功能以及碳的源与汇(Aerts & Chapin III, 2000; van Wijk et al., 2003; Wan et al., 2005). ...
The functional organization and control of plant respiration 1 2006
Relating extractable soil phosphorus to phosphorus losses in runoff 1 1996
... 磷主要通过地表径流沿着土壤剖面, 通过淋溶、侵蚀、吸附以及向闭蓄态磷库转化等方式输出生态系统(Pote & Daniel, 1996; McGroddy et al., 2004; Mahowald et al., 2008).影响磷输出生态系统过程的机理相对复杂, 土壤的磷含量、植被覆盖状况、土壤类型、土壤微生物以及气候条件等都是影响磷流失的因子.活性磷库的大小以及径流量是影响磷以淋溶方式输出生态系统的主要因子(Mahowald et al., 2008), 同时气候与环境因素对以淋溶、侵蚀等方式流失的磷具有影响(McGroddy et al., 2004), 但是气候与环境因子量化难度较高. ...
Incorporating phosphorus cycling into global modeling efforts: A worthwhile, tractable endeavor 1 2015
... 近年来, 随着全球变暖的加剧以及农田生态系统中磷肥的施入, 全球范围内植被净初级生产力普遍提高, 磷对植物生长的限制性作用逐渐加强(Goll et al., 2012; Sun et al., 2017).因此, 能模拟磷循环的生物地球化学模型对于评估与预测全球碳循环至关重要.研究表明, 生物地球化学模型如果不考虑磷限制因素则可能高估植被的碳吸收(Zhang et al., 2014; Reed et al., 2015).虽然早在2000年初开始出现模拟磷影响碳循环的模型(Wang et al., 2007), 但是目前国际上只有少数生物地球化学模型能够模拟磷循环.因此, 系统总结磷影响陆地生态系统碳循环过程及其模型表达方法, 一方面能提高我们对该领域研究态势和趋势的认识, 另一方面对我国生物地球化学模型的发展具有指导意义. ...
Global patterns of plant leaf N and P in relation to temperature and latitude 1 2004
... 土壤与叶片中的磷均影响植物光合作用过程.实验表明, 土壤中以无机磷为主导的多种形态的磷, 不仅会直接影响光合作用的能力和速率, 而且也会间接地影响着植物叶片中磷的浓度(Ellsworth et al., 2015).一方面, 土壤中的无机磷可以被植物体吸收, 其不仅充当光合作用的基底物, 而且是形成光合作用的生物分子的重要元素(Reich & Oleksy, 2004).另一方面, 土壤中的无机磷影响着二磷酸核酮糖羧化酶的形成, 从而影响着光合作用的速率(Norby et al., 2015, 2017).同样, 植物叶片中磷的浓度与植物叶片光合速率的相关性十分显著, 如果叶片中磷供应不足会影响磷酸甘油酸的还原和再生(Fredeen et al., 2018), 降低叶绿体间质的pH值以及参与卡尔文循环的各种酶的活性, 最终影响光合作用的强度、羧化速率等(Lloyd et al., 2001; Wang et al., 2017). ...
Short term effects of radiata pine and selected pasture species on soil organic phosphorus mineralisation 1 2005
... 土壤中的有机磷只有经过矿化后才能被植物吸收利用, 有机磷的矿化过程可以源源不断地为植物提供新的磷来源, 有机磷的矿化过程包括生物矿化过程和生物化学矿化过程两部分, 其中生物化学矿化过程是有机磷矿化的主导过程.土壤中矿化的磷通量大小不仅取决于环境因子, 还取决于生物因子(Niu et al., 2013).土壤中有机磷的矿化速率与有机碳的含量也有一定的关系, 很多研究发现有机碳的含量与有机磷的矿化速率呈显著的正相关关系(Scott & Condron, 2005; Thomas et al., 2006).另外, 土壤的温度、湿度、孔隙度等理化性质影响着有机磷矿化通量的大小, 如Campbell和Racz (1975)研究发现在含水量较高的土壤中有机磷的矿化较强, 同时也有研究表明土壤温度升高可以加快土壤中有机磷的矿化速率(Turner et al., 2003).土壤中的酶是有机磷矿化过程的重要催化剂, 如磷酸酶是有机磷矿化的首要催化剂, 可以促进土壤中有机物释放无机磷, 其活性的高低影响着土壤中有机磷的分解转化(Wang et al., 2010). ...
Trait-based scaling of temperature-dependent foliar respiration in a species-rich tropical forest canopy 1 2014
... 磷影响植物吸收全过程.叶片中磷浓度与植物呼吸之间具有较强的相关性, 在热带地区这种相关性尤为明显.研究发现, 热带森林中叶片磷浓度显著影响着植物的呼吸作用(Meir et al., 2001; Atkin et al., 2015), Domingues等(2010)在热带地区观测到较低的叶片磷浓度明显影响叶片暗呼吸(Slot et al., 2014).另外, Atkin等(2015)发现磷的可利用性还影响着叶片暗呼吸与氮之间的关系. ...
Phosphorus in the environment: Natural flows and human interferences 1 2000
Diagnosing phosphorus limitations in natural terrestrial ecosystems in carbon cycle models 1 2017
... 近年来, 随着全球变暖的加剧以及农田生态系统中磷肥的施入, 全球范围内植被净初级生产力普遍提高, 磷对植物生长的限制性作用逐渐加强(Goll et al., 2012; Sun et al., 2017).因此, 能模拟磷循环的生物地球化学模型对于评估与预测全球碳循环至关重要.研究表明, 生物地球化学模型如果不考虑磷限制因素则可能高估植被的碳吸收(Zhang et al., 2014; Reed et al., 2015).虽然早在2000年初开始出现模拟磷影响碳循环的模型(Wang et al., 2007), 但是目前国际上只有少数生物地球化学模型能够模拟磷循环.因此, 系统总结磷影响陆地生态系统碳循环过程及其模型表达方法, 一方面能提高我们对该领域研究态势和趋势的认识, 另一方面对我国生物地球化学模型的发展具有指导意义. ...
Leaf inorganic phosphorus as a potential indicator of phosphorus status, photosynthesis and growth of Eucalyptus grandis seedlings 1 2006
... 土壤中的有机磷只有经过矿化后才能被植物吸收利用, 有机磷的矿化过程可以源源不断地为植物提供新的磷来源, 有机磷的矿化过程包括生物矿化过程和生物化学矿化过程两部分, 其中生物化学矿化过程是有机磷矿化的主导过程.土壤中矿化的磷通量大小不仅取决于环境因子, 还取决于生物因子(Niu et al., 2013).土壤中有机磷的矿化速率与有机碳的含量也有一定的关系, 很多研究发现有机碳的含量与有机磷的矿化速率呈显著的正相关关系(Scott & Condron, 2005; Thomas et al., 2006).另外, 土壤的温度、湿度、孔隙度等理化性质影响着有机磷矿化通量的大小, 如Campbell和Racz (1975)研究发现在含水量较高的土壤中有机磷的矿化较强, 同时也有研究表明土壤温度升高可以加快土壤中有机磷的矿化速率(Turner et al., 2003).土壤中的酶是有机磷矿化过程的重要催化剂, 如磷酸酶是有机磷矿化的首要催化剂, 可以促进土壤中有机物释放无机磷, 其活性的高低影响着土壤中有机磷的分解转化(Wang et al., 2010). ...
Phosphorus-31 nuclear magnetic resonance spectral assignments of phosphorus compounds in soil NaOH-EDTA extracts 1 2003
... 土壤中的有机磷只有经过矿化后才能被植物吸收利用, 有机磷的矿化过程可以源源不断地为植物提供新的磷来源, 有机磷的矿化过程包括生物矿化过程和生物化学矿化过程两部分, 其中生物化学矿化过程是有机磷矿化的主导过程.土壤中矿化的磷通量大小不仅取决于环境因子, 还取决于生物因子(Niu et al., 2013).土壤中有机磷的矿化速率与有机碳的含量也有一定的关系, 很多研究发现有机碳的含量与有机磷的矿化速率呈显著的正相关关系(Scott & Condron, 2005; Thomas et al., 2006).另外, 土壤的温度、湿度、孔隙度等理化性质影响着有机磷矿化通量的大小, 如Campbell和Racz (1975)研究发现在含水量较高的土壤中有机磷的矿化较强, 同时也有研究表明土壤温度升高可以加快土壤中有机磷的矿化速率(Turner et al., 2003).土壤中的酶是有机磷矿化过程的重要催化剂, 如磷酸酶是有机磷矿化的首要催化剂, 可以促进土壤中有机物释放无机磷, 其活性的高低影响着土壤中有机磷的分解转化(Wang et al., 2010). ...
Linkage of root physiology and morphology as an adaptation to soil phosphorus impoverishment in tropical montane forests 1 2015
... 根是植物体从土壤中吸收磷的重要器官, 植物根对磷的获取能力影响着植物体生长发育的全过程(Ushio et al., 2015; Battini et al., 2017), 而影响植物根部对磷吸收的主要因素分为两类, 一类为植物体生理特征, 如根系的形态(廖红等, 2001)、根的结构(严小龙等, 2000)、根毛(Jungk, 2001)等均影响根部吸收速率(Zhang et al., 2016); 另一类为根部土壤环境, 如土壤中磷酸酶(酸性磷酸酶APA)的含量、微生物的丰富度(Spohn et al., 2018)、菌群、水分(Deepika & Kothamasi, 2015)等.同时, 不同的物种与不同的气候环境下根部的磷吸收速率也存在差异.总体而言, 植物可以通过根系的生长、形成根毛及菌根等方式提高土壤磷的利用效率. ...
Luxury consumption of soil nutrients: A possible competitive strategy in above-ground and below-ground biomass allocation and root morphology for slow-growing arctic vegetation? 1 2003
... 磷(P)是植物体内有机物的组成元素, 广泛参与植物有机体的代谢过程(刘超等, 2012), 与氮(N)一起通过影响植物的光合作用、呼吸作用和土壤有机质分解等一系列过程实现与碳氮循环过程的耦合(李雷等, 2013; 卢少勇等, 2016).在地史时期, 生物圈的活动形成了磷在全球范围内的大循环, 磷的循环与转化在重建气候、环境、生态以及全球变化的长期反馈机制中有着重要的作用(F?llmi, 1996).一方面, 地球生命从起源开始就依赖磷, 磷是所有生命形式的基本元素, 参与生物体的遗传物质与能量物质的转化, 是构成许多生命大分子(核酸、磷脂、二磷酸腺苷(ADP)、三磷酸腺苷(ATP)等)的关键元素, 在整个生态系统内能量的贮存、迁移、转化过程中发挥着重要的作用(Pierrou, 1976; 赵琼和曾德慧, 2005; Achat et al., 2009; Wang et al., 2010).另一方面, 众多研究表明磷的有效性强烈地影响着陆地生态系统的碳循环(de Graaff et al., 2006; Norby et al., 2010; Fernández-Martínez et al., 2014): 光合作用产生的糖类如果不经过磷酸化, 碳的固定则是无效的; 磷也会加强光合作用过程中碳水化合物的合成与运输; 在植物与土壤中, 磷与碳、氮的化学计量关系不仅会影响着植物体内碳的积累与分配(周正虎和王传宽, 2016), 还决定着全球陆地生态系统功能以及碳的源与汇(Aerts & Chapin III, 2000; van Wijk et al., 2003; Wan et al., 2005). ...
Direct and indirect effects of experimental warming on ecosystem carbon processes in a tallgrass prairie 1 2005
... 磷(P)是植物体内有机物的组成元素, 广泛参与植物有机体的代谢过程(刘超等, 2012), 与氮(N)一起通过影响植物的光合作用、呼吸作用和土壤有机质分解等一系列过程实现与碳氮循环过程的耦合(李雷等, 2013; 卢少勇等, 2016).在地史时期, 生物圈的活动形成了磷在全球范围内的大循环, 磷的循环与转化在重建气候、环境、生态以及全球变化的长期反馈机制中有着重要的作用(F?llmi, 1996).一方面, 地球生命从起源开始就依赖磷, 磷是所有生命形式的基本元素, 参与生物体的遗传物质与能量物质的转化, 是构成许多生命大分子(核酸、磷脂、二磷酸腺苷(ADP)、三磷酸腺苷(ATP)等)的关键元素, 在整个生态系统内能量的贮存、迁移、转化过程中发挥着重要的作用(Pierrou, 1976; 赵琼和曾德慧, 2005; Achat et al., 2009; Wang et al., 2010).另一方面, 众多研究表明磷的有效性强烈地影响着陆地生态系统的碳循环(de Graaff et al., 2006; Norby et al., 2010; Fernández-Martínez et al., 2014): 光合作用产生的糖类如果不经过磷酸化, 碳的固定则是无效的; 磷也会加强光合作用过程中碳水化合物的合成与运输; 在植物与土壤中, 磷与碳、氮的化学计量关系不仅会影响着植物体内碳的积累与分配(周正虎和王传宽, 2016), 还决定着全球陆地生态系统功能以及碳的源与汇(Aerts & Chapin III, 2000; van Wijk et al., 2003; Wan et al., 2005). ...
Significant contribution of combustion-related emissions to the atmospheric phosphorus budget 1 2015
... 大量的实验研究表明磷主要通过风化、沉降、地表径流等方式输入陆地生态系统, 其中, 风化磷是由生物和有机磷酸盐通过存储转化而产生的磷的释放, 土壤的风化基面是磷酸盐暂时的存储库, 岩性、土壤发育程度、径流、气温、降水、植被类型等是影响磷风化通量大小的主要因子(Goll et al., 2014; Hartmann et al., 2014).虽然在某些地区沉降是磷输入生态系统的重要方式, 但通过沉降方式输入生态系统的磷可利用性相对较小(Pe?uelas et al., 2014), 影响磷沉降通量大小的因子相对复杂, 因此对其机理的研究还有较大的不确定性.有研究表明以沉降方式输入生态系统的磷通量与以风化方式输入的磷通量的量级大致相当(Wang et al., 2015).径流是磷输入生态系统的途径之一, 但由于其输入数量相对较小, 对其机理的研究结果也具有很大的不确定性. ...
Temperature sensitivity of soil respiration: Synthetic effects of nitrogen and phosphorus fertilization on Chinese Loess Plateau 1 2017
... 土壤与叶片中的磷均影响植物光合作用过程.实验表明, 土壤中以无机磷为主导的多种形态的磷, 不仅会直接影响光合作用的能力和速率, 而且也会间接地影响着植物叶片中磷的浓度(Ellsworth et al., 2015).一方面, 土壤中的无机磷可以被植物体吸收, 其不仅充当光合作用的基底物, 而且是形成光合作用的生物分子的重要元素(Reich & Oleksy, 2004).另一方面, 土壤中的无机磷影响着二磷酸核酮糖羧化酶的形成, 从而影响着光合作用的速率(Norby et al., 2015, 2017).同样, 植物叶片中磷的浓度与植物叶片光合速率的相关性十分显著, 如果叶片中磷供应不足会影响磷酸甘油酸的还原和再生(Fredeen et al., 2018), 降低叶绿体间质的pH值以及参与卡尔文循环的各种酶的活性, 最终影响光合作用的强度、羧化速率等(Lloyd et al., 2001; Wang et al., 2017). ...
A model of biogeochemical cycles of carbon, nitrogen, and phosphorus including symbiotic nitrogen fixation and phosphatase production 1 2007
... 近年来, 随着全球变暖的加剧以及农田生态系统中磷肥的施入, 全球范围内植被净初级生产力普遍提高, 磷对植物生长的限制性作用逐渐加强(Goll et al., 2012; Sun et al., 2017).因此, 能模拟磷循环的生物地球化学模型对于评估与预测全球碳循环至关重要.研究表明, 生物地球化学模型如果不考虑磷限制因素则可能高估植被的碳吸收(Zhang et al., 2014; Reed et al., 2015).虽然早在2000年初开始出现模拟磷影响碳循环的模型(Wang et al., 2007), 但是目前国际上只有少数生物地球化学模型能够模拟磷循环.因此, 系统总结磷影响陆地生态系统碳循环过程及其模型表达方法, 一方面能提高我们对该领域研究态势和趋势的认识, 另一方面对我国生物地球化学模型的发展具有指导意义. ...
A global model of carbon, nitrogen and phosphorus cycles for the terrestrial biosphere 4 2010
... 磷(P)是植物体内有机物的组成元素, 广泛参与植物有机体的代谢过程(刘超等, 2012), 与氮(N)一起通过影响植物的光合作用、呼吸作用和土壤有机质分解等一系列过程实现与碳氮循环过程的耦合(李雷等, 2013; 卢少勇等, 2016).在地史时期, 生物圈的活动形成了磷在全球范围内的大循环, 磷的循环与转化在重建气候、环境、生态以及全球变化的长期反馈机制中有着重要的作用(F?llmi, 1996).一方面, 地球生命从起源开始就依赖磷, 磷是所有生命形式的基本元素, 参与生物体的遗传物质与能量物质的转化, 是构成许多生命大分子(核酸、磷脂、二磷酸腺苷(ADP)、三磷酸腺苷(ATP)等)的关键元素, 在整个生态系统内能量的贮存、迁移、转化过程中发挥着重要的作用(Pierrou, 1976; 赵琼和曾德慧, 2005; Achat et al., 2009; Wang et al., 2010).另一方面, 众多研究表明磷的有效性强烈地影响着陆地生态系统的碳循环(de Graaff et al., 2006; Norby et al., 2010; Fernández-Martínez et al., 2014): 光合作用产生的糖类如果不经过磷酸化, 碳的固定则是无效的; 磷也会加强光合作用过程中碳水化合物的合成与运输; 在植物与土壤中, 磷与碳、氮的化学计量关系不仅会影响着植物体内碳的积累与分配(周正虎和王传宽, 2016), 还决定着全球陆地生态系统功能以及碳的源与汇(Aerts & Chapin III, 2000; van Wijk et al., 2003; Wan et al., 2005). ...
... Carnegie-Ames-Stanford Approach-CNP (CASA- CNP)、Jena Scheme for Biosphere-Atmosphere Coupling in Hamburg-CNP (JSBACH-CNP)、Community Land Model-CNP (CLM-CNP)模型是目前国际上主流的能模拟碳、氮、磷循环的生物地球化学模型.Wang等(2010)基于CASA-CNP模型模拟了全球陆地生态系统中碳、氮、磷的空间分布, 发现磷的限制使得热带地区植被净初级生产力减少了20%.Goll等(2012)基于JSBACH-CNP模型模拟评估了磷对CO2浓度升高引起的陆地生态系统碳吸收的影响, 发现磷限制使得全球碳吸收减少了16%.Yang等(2014)基于CLM-CNP模型模拟得出热带与温带森林1970-1999年间的磷吸收速率为0.18-0.71 mg·m-2·a-1.上述过程模型在模拟评估磷对于陆地生态系统碳循环的影响方面取得了较大的进展.本文在分析磷参与生态系统碳循环的过程与机理的基础上, 系统总结了JSBACH-CNP、CASA-CNP、CLM- CNP模型中磷在生态系统中循环转化的定量表达方法, 提出了模型的构建原则和框架, 以期为未来发展与改进与磷相关的生物地球化学模型提供参考. ...
... 根是植物体从土壤中吸收磷的重要器官, 植物根对磷的获取能力影响着植物体生长发育的全过程(Ushio et al., 2015; Battini et al., 2017), 而影响植物根部对磷吸收的主要因素分为两类, 一类为植物体生理特征, 如根系的形态(廖红等, 2001)、根的结构(严小龙等, 2000)、根毛(Jungk, 2001)等均影响根部吸收速率(Zhang et al., 2016); 另一类为根部土壤环境, 如土壤中磷酸酶(酸性磷酸酶APA)的含量、微生物的丰富度(Spohn et al., 2018)、菌群、水分(Deepika & Kothamasi, 2015)等.同时, 不同的物种与不同的气候环境下根部的磷吸收速率也存在差异.总体而言, 植物可以通过根系的生长、形成根毛及菌根等方式提高土壤磷的利用效率. ...
植物根构型特性与磷吸收效率 1 2000
... 根是植物体从土壤中吸收磷的重要器官, 植物根对磷的获取能力影响着植物体生长发育的全过程(Ushio et al., 2015; Battini et al., 2017), 而影响植物根部对磷吸收的主要因素分为两类, 一类为植物体生理特征, 如根系的形态(廖红等, 2001)、根的结构(严小龙等, 2000)、根毛(Jungk, 2001)等均影响根部吸收速率(Zhang et al., 2016); 另一类为根部土壤环境, 如土壤中磷酸酶(酸性磷酸酶APA)的含量、微生物的丰富度(Spohn et al., 2018)、菌群、水分(Deepika & Kothamasi, 2015)等.同时, 不同的物种与不同的气候环境下根部的磷吸收速率也存在差异.总体而言, 植物可以通过根系的生长、形成根毛及菌根等方式提高土壤磷的利用效率. ...
The role of phosphorus dynamics in tropical forests—A modeling study using CLM-CNP 2 2014
... Carnegie-Ames-Stanford Approach-CNP (CASA- CNP)、Jena Scheme for Biosphere-Atmosphere Coupling in Hamburg-CNP (JSBACH-CNP)、Community Land Model-CNP (CLM-CNP)模型是目前国际上主流的能模拟碳、氮、磷循环的生物地球化学模型.Wang等(2010)基于CASA-CNP模型模拟了全球陆地生态系统中碳、氮、磷的空间分布, 发现磷的限制使得热带地区植被净初级生产力减少了20%.Goll等(2012)基于JSBACH-CNP模型模拟评估了磷对CO2浓度升高引起的陆地生态系统碳吸收的影响, 发现磷限制使得全球碳吸收减少了16%.Yang等(2014)基于CLM-CNP模型模拟得出热带与温带森林1970-1999年间的磷吸收速率为0.18-0.71 mg·m-2·a-1.上述过程模型在模拟评估磷对于陆地生态系统碳循环的影响方面取得了较大的进展.本文在分析磷参与生态系统碳循环的过程与机理的基础上, 系统总结了JSBACH-CNP、CASA-CNP、CLM- CNP模型中磷在生态系统中循环转化的定量表达方法, 提出了模型的构建原则和框架, 以期为未来发展与改进与磷相关的生物地球化学模型提供参考. ...
... 植物体根部吸收磷后, 按照一定的比例输送、分配给根、茎、叶器官.模型对植物体磷的分配的定量描述主要依据根、茎、叶中的碳、氮、磷的化学计量关系来计算(Yang et al., 2014).植物体各器官获得的磷通量(Fu,i)的通用表达式为: ...
Increased soil phosphorus availability induced by faba bean root exudation stimulates root growth and phosphorus uptake in neighbouring maize 1 2016
... 根是植物体从土壤中吸收磷的重要器官, 植物根对磷的获取能力影响着植物体生长发育的全过程(Ushio et al., 2015; Battini et al., 2017), 而影响植物根部对磷吸收的主要因素分为两类, 一类为植物体生理特征, 如根系的形态(廖红等, 2001)、根的结构(严小龙等, 2000)、根毛(Jungk, 2001)等均影响根部吸收速率(Zhang et al., 2016); 另一类为根部土壤环境, 如土壤中磷酸酶(酸性磷酸酶APA)的含量、微生物的丰富度(Spohn et al., 2018)、菌群、水分(Deepika & Kothamasi, 2015)等.同时, 不同的物种与不同的气候环境下根部的磷吸收速率也存在差异.总体而言, 植物可以通过根系的生长、形成根毛及菌根等方式提高土壤磷的利用效率. ...
Nitrogen and phosphorous limitations significantly reduce future allowable CO2 emissions 1 2014
... 近年来, 随着全球变暖的加剧以及农田生态系统中磷肥的施入, 全球范围内植被净初级生产力普遍提高, 磷对植物生长的限制性作用逐渐加强(Goll et al., 2012; Sun et al., 2017).因此, 能模拟磷循环的生物地球化学模型对于评估与预测全球碳循环至关重要.研究表明, 生物地球化学模型如果不考虑磷限制因素则可能高估植被的碳吸收(Zhang et al., 2014; Reed et al., 2015).虽然早在2000年初开始出现模拟磷影响碳循环的模型(Wang et al., 2007), 但是目前国际上只有少数生物地球化学模型能够模拟磷循环.因此, 系统总结磷影响陆地生态系统碳循环过程及其模型表达方法, 一方面能提高我们对该领域研究态势和趋势的认识, 另一方面对我国生物地球化学模型的发展具有指导意义. ...
 ,1,*, 王娜1,2, 王昭生1, 巩贺1
,1,*, 王娜1,2, 王昭生1, 巩贺1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}