Effects of water and nutrient additions on functional traits and aboveground biomass of Leymus chinensis
Dan-Dan ZHAO1,2, Hong-Yuan MA,1,*, Yang LI1,2, Ji-Ping WEI1, Zhi-Chun WANG11 Northeast Institute of Geography and Agroecology, Chinese Academy of Sciences, Changchun 130102, China 2 University of Chinese Academy of Science, Beijing 100049, China
Supported by the National Natural Science Foundation of China(41771058) the National Key R&D Program of China(2016YFC0501200) the National Basic Research Program of China(2015CB150802) the National Key Basic Survey of Resources(2015FY110500) the Category A Strategic Leading Science and Technology Project of Chinese Academy of Sciences(XDA2306040303)
Abstract Aims The research on the response of plant functional traits to environmental change, such as precipitation change and nutrient additions is very important to understand how plant species adapt to variable environments. Methods We conducted a pot experiment with a gradient of water treatments (increase precipitation by 50%, HW; decrease precipitation by 50%, LW; take 498 mm precipitation as control, MW) and nutrient additions (without nutrient addition, CK; nitrogen (N) addition, NA; phosphorus (P) addition, PA; nitrogen and phosphorus additions, N+P). We investigated 11 plant functional traits and aboveground biomass of Leymus chinensis. Important findings The effects of moisture on plant height, tillers, stem biomass, leaf biomass, leaf area, leaf mass, net photosynthetic rate, transpiration rate, water use efficiency were significant. The effects of fertilizers on tillers, stem biomass, net photosynthetic rate, transpiration rate, water use efficiency were significant. And the combination of fertilizers and moisture had a significant influence on tillers, stem biomass, transpiration rate and water use efficiency (two-way ANOVA). The pattern of functional traits in response to precipitation differed between plants with varied fertilizer additions. Tillers and leaf area were increased in treatments with HW under N and N+P additions, but not changed under CK and P addition. Stem biomass increased along the precipitation gradients under CK, N addition and P addition, but did not change under N+P additions. Specific leaf area with HW was significantly higher than that of MW under N addition, but not changed under other nutrient addition. Short-term nutrient additions significantly affected photosynthetic physiological traits of L. chinensis, but it had no significant effect on morphological traits under the same precipitation. The aboveground biomass of L. chinensis increased with the increase of precipitation, and reached the highest level of 522.55 g·m -2 with HW treatment under N addition. In conclusion, our results indicate that the functional traits in L. chinensis respond to precipitation addition and the patterns of responses differ under different nutrient additions, reflecting the adaptation to changes in water and nutrient availability. Keywords:plant functional traits;morphological traits;physiological traits;aboveground biomass
PDF (1297KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 赵丹丹, 马红媛, 李阳, 魏继平, 王志春. 水分和养分添加对羊草功能性状和地上生物量的影响. 植物生态学报, 2019, 43(6): 501-511. DOI: 10.17521/cjpe.2019.0041 ZHAO Dan-Dan, MA Hong-Yuan, LI Yang, WEI Ji-Ping, WANG Zhi-Chun. Effects of water and nutrient additions on functional traits and aboveground biomass of Leymus chinensis. Chinese Journal of Plant Ecology, 2019, 43(6): 501-511. DOI: 10.17521/cjpe.2019.0041
盆栽试验开始于2016年6月中旬, 在中国科学院东北地理与农业生态研究所(125.38° E, 43.98° N, 海拔190 m)进行, 该区域气候属于温带季风气候, 1953-2012年的平均年降水量为498.0 mm, 年降水量极大值为754.0 mm (1956年), 比平均水平高出约50%, 年降水量极小值为244.1 mm (1982年)(李晶等, 2015), 比平均水平低50%。土壤类型为黑土, 有机质平均含量为2.83%、全N含量为1 368.56 mg·kg-1、全P含量为669.18 mg·kg-1。试验在遮雨棚内进行, 试验盆直径30 cm, 高27 cm, 盆内装入试验地附近的0-20 cm土层的过筛土, 每盆移栽10株大小均一的羊草。晴天时收起遮雨棚, 下雨时进行遮挡以防止盆栽接收自然降水。定期浇水, 剔除杂草, 保证羊草植株生长。
Table 1 表1 表1水处理、施肥处理及其交互作用对羊草功能性状影响的双因素方差分析 Table 1Results of two-way ANOVAs for the effects of water treatments, fertilization treatment and their interactions on functional traits of Leymus chinensis
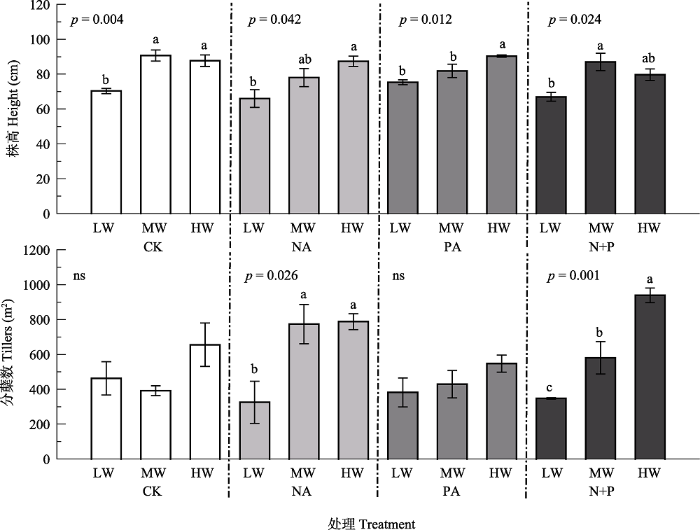
Fig. 1Effects of fertilization and water on height and tillers of Leymus chinensis (mean ± SE). CK, control; NA, N addition; PA, P addition; N+P, N and P additions; LW, MW, HW represent low, moderate and high precipitation levels. Different lowercase letters indicate significant difference (p < 0.05) among different water treatments at the same fertilizer addition, and ns indicates non-significant differences (p > 0.05) among water treatments.
Fig. 2Effects of fertilization and water on biomass allocation of Leymus chinensis (mean ± SE). CK, control; NA, N addition; PA, P addition; N+P, N and P additions; LW, MW, HW represent low, moderate and high precipitation levels. Different lowercase letters indicate significant difference (p < 0.05) among different water treatments at the same fertilizer addition, and ns indicates non-significant differences (p > 0.05) among water treatments.
Fig. 3Effects of fertilization and water on morphological traits of Leymus chinensis (mean ± SE). LA, leaf area; SLA, specific leaf area. CK, control; NA, N addition; PA, P addition; N+P, N and P additions; LW, MW, HW represent low, moderate and high precipitation levels. Different lowercase letters indicate significant difference (p < 0.05) among different water treatments at the same fertilizer addition, and ns indicates non-significant differences (p > 0.05) among water treatments.
Fig. 4Effects of fertilization and water on photosynthetic traits of Leymus chinensis (mean ± SE). Pn, net photosynthesis rate; WUE, water use efficiency. CK, control; NA, N addition; PA, P addition; N+P, N and P additions; LW, MW, HW represent low, moderate and high precipitation levels. Different lowercase letters indicate significant difference (p < 0.05) among different water treatments at the same fertilizer addition, and ns indicates non-significant differences (p > 0.05) among water treatments.
Fig. 5Effects of fertilization and water on above-ground biomass of Leymus chinensis (mean ± SE). CK, control; NA, N addition; PA, P addition; N+P, N and P additions; LW, MW, HW represent low, moderate and high precipitation levels. Different lowercase letters indicate significant difference (p < 0.05) among different water treatments at the same fertilizer addition, and ns indicates non-significant differences (p > 0.05) among water treatments.
BaiX, ChengJH, ZhengSX, ZhanSX, BaiYF (2014). Ecophysiological responses of Leymus chinensis to nitrogen and phosphorus additions in a typical steppe Chinese Journal of Plant Ecology, 38, 103-115. [本文引用: 1]
BaiYF, WuJG, ClarkCM, NaeemS, PanQM, HuangJH, ZhangLX, HanXG (2010). Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: Evidence from Inner Mongolia Grasslands Global Change Biology, 16, 358-372. [本文引用: 1]
BalachowskiJA, VolaireFA (2018). Implications of plant functional traits and drought survival strategies for ecological restoration Journal of Applied Ecology, 55, 631-640. [本文引用: 1]
BarkerDH, VanierC, NamburgE, CharletTN, NielsenKM, NewinghamBA, SmithSD (2005). Enhanced monsoon precipitation and nitrogen deposition affect leaf traits and photosynthesis differently in spring and summer in the desert shrub Larrea tridentate New Phytologist, 169, 799-808. [本文引用: 1]
BeheraSK, PandaRK (2009). Integrated management of irrigation water and fertilizers for wheat crop using field experiments and simulation modeling Agricultural Water Management, 96, 1532-1540. [本文引用: 1]
BobbinkR, HicksK, GallowayJ, SprangerT, AlkemadeR, AshmoreM, BustamanteM, CinderbyS, DavidsonE, DentenerF, EmmettB, ErismanJW, FennM, GilliamF, NordinA, PardoL, de VriesW (2010). Global assessment of nitrogen deposition effects on terrestrial plant diversity: A synthesis Ecological Applications, 20, 30-59. [本文引用: 2]
BuckleyH, YoungCA, CharltonND, HendricksWQ, HaleyB, NagabhyruP, RudgersJA (2019). Leaf endophytes mediate fertilizer effects on plant yield and traits in northern oat grass (Trisetum spicatum) Plant and Soil, 434, 425-440. [本文引用: 1]
ChenSP, BaiYF, ZhangLX, HanXG (2005). Comparing physiological responses of two dominant grass species to nitrogen addition in Xilin River Basin of China Environmental and Experimental Botany, 53, 65-75. [本文引用: 1]
DíazS, LavorelS, McintyreS, FalczukV, CasanovesF, MilchunasDG, SkarpeC, RuschG, SternbergM, Noy-MeirI, LandsbergJ, ZhangW, ClarkH, CampbellBD (2007). Plant trait responses to grazing—A global synthesis Global Change Biology, 13, 313-341. [本文引用: 1]
DjamanK, MelVC, DiopL, SowA, EI-NamakyR, MannehB, SatioK, FutakuchiK, IrmakS (2018). Effects of alternate wetting and drying irrigation regime and nitrogen fertilizer on yield and nitrogen use efficiency of irrigated rice in the Sahel Water, 10, 711. DOI: 10.3390/w10060711. [本文引用: 1]
FauconMP, HoubenD, LambersH (2017). Plant functional traits: Soil and ecosystem services Trends in Plant Science, 22, 385-394. [本文引用: 1]
FonsecaCR, OvertonJM, CollinsB, WestobyM (2000). Shifts in trait-combinations along rainfall and phosphorus gradients Journal of Ecology, 88, 964-977. [本文引用: 1]
ForrestelEJ, DonoghueMJ, EdwardsEJ, JetzW, du ToitJCO, SmithMD (2017). Different clades and traits yield similar grassland functional responses Proceedings of the National Academy of Sciences of the United States of America, 114, 705-710. [本文引用: 1]
GaoR, YangX, LiuG, HuangZ, WalckJL (2015). Effects of rainfall pattern on the growth and fecundity of a dominant dune annual in a semi-arid ecosystem Plant and Soil, 389, 335-347. [本文引用: 1]
GüsewellS (2004). N:P ratios in terrestrial plants: Variation and functional significance New Phytologist, 164, 243-266. [本文引用: 1]
HuZM, YuGR, FanJW, ZhongHP, WangSQ, LiSG (2010). Precipitation-use efficiency along a 4500-km grassland transect Global Ecology and Biogeography, 19, 842-851. [本文引用: 1]
HuangJY, XuP, YuHL, YuanZY, LiLH (2012). Responses of biomass, nutrient allocation of Leymus chinensis along N, P and water gradients Pratacultural Science, 29, 1589-1595. [本文引用: 1]
HuangJY, YuanZY, LiLH (2009). Changes in [N], [P] and specific leaf area of green leaves of Leymus chinensis along nitrogen, phosphorus and water gradients Chinese Journal of Plant Ecology, 33, 442-448. [本文引用: 1]
JanR, AgaFA, BaharFA, SinghT, LoneR (2018). Effect of nitrogen and silicon on growth and yield attributes of transplanted rice (Oryza sativa L.) under Kashmir conditions Journal of Pharmacognosy and Phytochemistry, 7, 328-332. [本文引用: 1]
LavorelS, GrigulisK, LamarqueP (2011). Using plant functional traits to understand the landscape distribution of multiple ecosystem services Journal of Ecology, 99, 135-147. [本文引用: 1]
LeBauerDS, TresederKK (2008). Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed Ecology, 89, 371-379. [本文引用: 1]
LiDJ, MoJM, FangYT, CaiXA, XueJH, XuGL (2004). Effects of simulated nitrogen deposition on growth and photosynthesis of Schima superba, Castanopsis chinensis and Cryptocarya concinna seedlings Acta Ecologica Sinica,2004,24, 876-882. [本文引用: 1]
LiJ, LiuZS, XueXQ, YuXG, ZhangB, YueYT (2015). Study of rainfall characteristics and pattern in Changchun city China Water & Wastewater, 31, 100-104. [本文引用: 1]
LiKF, ZhangFC, QiYL, XingYY, LiZJ (2010). Effects of water-fertilizer spatial coupling in root zone on winter wheat growth and yield Chinese Journal of Applied Ecology, 21, 3154-3160. [本文引用: 1]
LiSX, LiSQ, GaoYJ, WangXQ, HeHX, DuJJ (1994). The mechanism and effects of N fertilization in increasing water use efficiency Agricultural Research in the Arid Areas, 12, 38-46. [本文引用: 1]
LiXF (2011). Effects of Water and Fertilizer Cooperation on the Growth Nutrient Absorption and Root Activity of Summer Maize Master degree dissertation, Hebei Agricultural University, Baoding, Hebei. [本文引用: 1]
LiYH (1993). Grazing dynamics of the species diversity in Aneurolepidium chinense steppe and Stipa grandis steppe Acta Botanica Sinica, 35, 877-884. [本文引用: 1]
LüXT, ReedS, YuQ, HeNP, WangZW, HanXG (2013). Convergent responses of nitrogen and phosphorus resorption to nitrogen inputs in a semiarid grassland Global Change Biology, 19, 2775-2784. [本文引用: 1]
LüXT, ReedSC, YuQ, HanXG (2016). Nutrient resorption helps drive intra-specific coupling of foliar nitrogen and phosphorus under nutrient-enriched conditions Plant and Soil, 398, 111-120. [本文引用: 1]
MaHY, YangHY, LiangZW, OoiMKJ (2015). Effects of 10-year management regimes on the soil seed bank in saline- ??alkaline grassland PLOS ONE, 10, e0122319. DOI: 10.1371/ ?journal.?pone.0122319. [本文引用: 1]
MengFC, ZhangJH, YaoFM (2014). Interactive effects of elevated CO2 concentration and increasing precipitation on yield and growth development in maize Chinese Journal of Plant Ecology, 38, 1064-1073. [本文引用: 1]
MokanyK, AshJ, RoxburghS (2010). Functional identity is more important than diversity in influencing ecosystem processes in a temperate native grassland Journal of Ecology, 96, 884-893. [本文引用: 1]
MoorH, HylanderK, NorbergJ (2015). Predicting climate change effects on wetland ecosystem services using species distribution modeling and plant functional traits Ambio, 44, 113-126. [本文引用: 1]
OsborneCP, Charles-DominiqueT, StevensN, BondWJ, MidgleyG, LehmannCER (2018). Human impacts in African savannas are mediated by plant functional traits New Phytologist, 220, 10-24. [本文引用: 1]
PoorterL, RozendaalDMA (2008). Leaf size and leaf display of thirty-eight tropical tree species Oecologia, 158, 35-46. [本文引用: 1]
RaoLE, AllenEB (2010). Combined effects of precipitation and nitrogen deposition on native and invasive winter annual production in California deserts Oecologia, 162, 1035-1046. [本文引用: 1]
ReichPB, OleksynJ (2004). Global patterns of plant leaf N and P in relation to temperature and latitude Proceedings of the National Academy of Sciences of the United States of America, 101, 11001-11006. [本文引用: 1]
RosbakhS, R?mermannC, PoschlodP (2015). Specific leaf area correlates with temperature: New evidence of trait variation at the population, species and community levels Alpine Botany, 125, 79-86. [本文引用: 1]
ShipleyB (2006). Net assimilation rate, specific leaf area and leaf mass ratio: Which is most closely correlated with relative growth rate? A meta-analysis Functional Ecology, 20, 565-574. [本文引用: 1]
SongYT, LiQ, WangP, ZhouDW, Wuyunna (2016). Response of Leymus chinensis functional traits and aboveground biomass to nitrogen addition in Songnen grassland in northeast China Pratacultural Science, 33, 1383-1390. [本文引用: 1]
SwensonNG, EnquistBJ (2007). Ecological and evolutionary determinants of a key plant functional trait: Wood density and its community-wide variation across latitude and elevation American Journal of Botany, 94, 451-459. [本文引用: 1]
ViolleC, NavasML, VileD, KazakouE, FortunerlC, HummelI, GarnierE (2007). Let the concept of trait be functional! Oikos, 116, 882-892. [本文引用: 1]
WanHW, YangY, BaiSQ, XuYH, BaiYF (2008). Variations in leaf functional traits of six species along a nitrogen addition gradient in Leymus chinensis steppe in Inner Mongolia Journal of Plant Ecology (Chinese Version), 32, 611-621. [本文引用: 2]
WangS (2008). Effect of N Rates on Growth of Summer Maize and Drought Threshold under Different Drought Levels Master degree dissertation, Huazhong Agricultural University, Wuhan. [本文引用: 1]
XuXX, ChenGL, MuXM (1995). The coordinative effect of moisture and fertilizer upon spring wheat yields Agricultural Research in the Arid Areas, 13, 34-38. [本文引用: 1]
XuZZ, ZhouGS (2005). Effects of soil moisture on growth characteristics of Leymus chinensis seedlings under different temperature conditions Chinese Journal of Ecology, 24, 256-260. [本文引用: 1]
XuZZ, ZhouGS (2006). Nitrogen metabolism and photosynthesis in Leymus chinensis in response to long-term soil drought Journal of Plant Growth Regulation, 25, 252-266. [本文引用: 1]
YangX, LiJ, ZhaoT, MoL, ZhangJ, RenH, ZhaoN, GaoY (2019). Variation and heritability of morphological and physiological traits among Leymus chinensis genotypes under different environmental conditions Journal of Arid Land, 11, 66-74. [本文引用: 1]
YangY, FangJ, MaW, WangW (2008). Relationship between variability in aboveground net primary production and precipitation in global grasslands Geophysical Research Letters, 35, L23710. DOI: 10.1029/2008GL035408. [本文引用: 1]
YiJ, LiQF, GuAL, MenZH, HeWX (2001). Advances on biology characteristics the rhizomatous grasses Journal of Arid Land Resources and Environment, 15(S1), 1-16. [本文引用: 1]
YueX, ZuoX, YuQ, XuC, LvP, ZhangJ, KnappAK, SmithMD (2019). Response of plant functional traits of Leymus chinensis to extreme drought in Inner Mongolia grasslands Plant Ecology, 220, 141-149. [本文引用: 1]
ZengXP, ZhaoP, CaiXA, SunGC, PengSL (2004). Physioecological characteristics of Woonyoungia septentrionalis seedlings under various soil water conditions Chinese Journal of Ecology, 23(2), 26-31. [本文引用: 1]
ZhangGT, WangKX, WangLX, WangY (2007). Couple effect of the soil water and fertility in sustainable agriculture in Liaoning Journal of Anhui Agricultural Sciences, 35, 7531-7555. [本文引用: 1]
ZhangHX, GaoYZ, TasisaBY, BaskinJM, BaskinCC, LüXT, ZhouDW (2019). Divergent responses to water and nitrogen addition of three perennial bunchgrass species from variously degraded typical steppe in Inner Mongolia Science of the Total Environment, 647, 1344-1350. [本文引用: 1]
ZhangZN, WuGL, WangD, DengL, HaoHM, YangZ, ShangguanZP (2014). Plant community structure and soil moisture in the semi-arid natural grassland of the Loess Plateau Acta Prataculturae Sinica, 23, 313-319. [本文引用: 1]
ZhaoXF, XuHL, ZhangP, TuWX, ZhangQQ (2014). Effects of nutrient and water additions on plant community structure and species diversity in desert grasslands Chinese Journal of Plant Ecology, 38, 167-177. [本文引用: 1]
ZhouP, GengY, MaWH, HeJS (2010). Linkages of functional traits among plant organs in the dominant species of the Inner Mongolia grassland, China Chinese Journal of Plant Ecology, 34, 7-16. [本文引用: 1]
ZhuJT, LiXY, ZhangXM, ZengFJ, YangSG (2010). Leaf functional traits of Ceratoides latens in northern slope of Kunlun Mountain and its regional difference with the altitude Journal of Desert Research, 30, 1325-1330. [本文引用: 1]
Implications of plant functional traits and drought survival strategies for ecological restoration 1 2018
... 功能性状作为解释变量, 对生态系统功能和服务的变化有显著的指示作用(Garnier et al., 2004; Mokany et al., 2010; Moor et al., 2015; Osborne et al., 2018), 能够显示植物对环境异质性的适应能力(Reich & Oleksyn, 2004; Faucon et al., 2017).植被生长过程中, 功能性状会受到降水量和施肥处理不同程度的影响.降水量增加使植株高度增高, 叶面积增大, 净光合速率升高(曾小平等, 2004; 许振柱和周广胜, 2005; 孟凡超等, 2014; Yue et al., 2019), 本研究结果与之一致, 主要原因是水分充足与否直接影响到羊草根系的发育, 根系发育的良好程度又直接影响到植物对土壤养分的吸收及养分在植物体内的循环, 进而对植物功能性状产生影响(李秀芳, 2011; Balachowski & Volaire, 2018).已有研究表明, 植物的光合能力与相对生长速率紧密相关(Shipley, 2006; Morgan et al., 2011), 叶面积反映了叶片对光的截获能力(Poorter & Rozendaal, 2008).因此, 降水量增加与氮磷共施条件下较高的光合速率和较大的叶面积可能是较高的叶生物量和茎生物量的主要原因.Fonseca等(2000)的研究表明澳大利亚东南部多年生植物的SLA随降水量的增加而增加; 驼绒藜(Ceratoides latens)的SLA与降水量呈正相关关系(朱军涛等, 2010); 植物叶质量也会随降水量的增加而减小(Wright et al., 2004; Rosbakh et al., 2015; Buckley et al., 2019).而本研究中, 降水量对羊草SLA没有显著影响(p > 0.05), 可能是由于羊草对短期水分处理响应不明显(Gao et al., 2015; Forrestel et al., 2017). ...
Enhanced monsoon precipitation and nitrogen deposition affect leaf traits and photosynthesis differently in spring and summer in the desert shrub Larrea tridentate 1 2005
... 大部分陆地生态系统都受到N和P限制, 适当添加养分可以提高植物生产力(LeBauer & Treseder, 2008; Djaman et al., 2018).有****指出氮供应充分可以提高植物的光合能力, 进而促进植物的生长, 提高植物生物量(Chen et al., 2005; Jan et al., 2018).在干旱半干旱地区, 植物生长不仅受到养分的限制,较低的降水量导致水分也成为该地区的主要限制因子(Rao & Allen, 2010).本研究结果表明, 降水量增加显著提高了羊草地上生物量.在减少降水量的情况下, 氮添加、氮磷共同添加对羊草地上生物量的促进效应明显低于正常和增加降水量.这是因为植物对养分的吸收和运输往往依赖于水分, 养分要溶解于水中才能被植物吸收, 同时植物地上部分的蒸腾作用拉力成为养分运输的动力(Barker et al., 2005).另外土壤中氮的矿化和硝化作用与水分也有密切的联系(王双, 2008).因此, 干旱可能使溶解于水中的养分降低, 同时使植物的蒸腾作用降低, 导致减少降水时养分添加对植物的促进作用弱于其他处理. ...
Integrated management of irrigation water and fertilizers for wheat crop using field experiments and simulation modeling 1 2009
... 短期的养分添加主要影响羊草的生理性状, 如叶片光合速率、蒸腾速率和水分利用效率均显著高于不施肥处理, 这些性状主要与光合作用相关.这说明养分添加缓解了植物对养分的竞争, 使物种对资源的竞争从地下部分转向地上部分, 即增加植物对光资源的竞争(Bobbink et al., 2010).然而, 养分添加对羊草形态性状的影响作用相对较小, 这与万宏伟等(2008)的研究结果不一致, 可能是因为本研究时间较短, 植物性状随着资源水平的改变, 首先是调整生理性状使其快速适应, 而形态性状相对滞后, 需要较长的时间才能发生显著变化(宋彦涛等, 2016; Yang et al., 2019). ...
Leaf endophytes mediate fertilizer effects on plant yield and traits in northern oat grass (Trisetum spicatum) 1 2019
... 功能性状作为解释变量, 对生态系统功能和服务的变化有显著的指示作用(Garnier et al., 2004; Mokany et al., 2010; Moor et al., 2015; Osborne et al., 2018), 能够显示植物对环境异质性的适应能力(Reich & Oleksyn, 2004; Faucon et al., 2017).植被生长过程中, 功能性状会受到降水量和施肥处理不同程度的影响.降水量增加使植株高度增高, 叶面积增大, 净光合速率升高(曾小平等, 2004; 许振柱和周广胜, 2005; 孟凡超等, 2014; Yue et al., 2019), 本研究结果与之一致, 主要原因是水分充足与否直接影响到羊草根系的发育, 根系发育的良好程度又直接影响到植物对土壤养分的吸收及养分在植物体内的循环, 进而对植物功能性状产生影响(李秀芳, 2011; Balachowski & Volaire, 2018).已有研究表明, 植物的光合能力与相对生长速率紧密相关(Shipley, 2006; Morgan et al., 2011), 叶面积反映了叶片对光的截获能力(Poorter & Rozendaal, 2008).因此, 降水量增加与氮磷共施条件下较高的光合速率和较大的叶面积可能是较高的叶生物量和茎生物量的主要原因.Fonseca等(2000)的研究表明澳大利亚东南部多年生植物的SLA随降水量的增加而增加; 驼绒藜(Ceratoides latens)的SLA与降水量呈正相关关系(朱军涛等, 2010); 植物叶质量也会随降水量的增加而减小(Wright et al., 2004; Rosbakh et al., 2015; Buckley et al., 2019).而本研究中, 降水量对羊草SLA没有显著影响(p > 0.05), 可能是由于羊草对短期水分处理响应不明显(Gao et al., 2015; Forrestel et al., 2017). ...
Comparing physiological responses of two dominant grass species to nitrogen addition in Xilin River Basin of China 1 2005
... 大部分陆地生态系统都受到N和P限制, 适当添加养分可以提高植物生产力(LeBauer & Treseder, 2008; Djaman et al., 2018).有****指出氮供应充分可以提高植物的光合能力, 进而促进植物的生长, 提高植物生物量(Chen et al., 2005; Jan et al., 2018).在干旱半干旱地区, 植物生长不仅受到养分的限制,较低的降水量导致水分也成为该地区的主要限制因子(Rao & Allen, 2010).本研究结果表明, 降水量增加显著提高了羊草地上生物量.在减少降水量的情况下, 氮添加、氮磷共同添加对羊草地上生物量的促进效应明显低于正常和增加降水量.这是因为植物对养分的吸收和运输往往依赖于水分, 养分要溶解于水中才能被植物吸收, 同时植物地上部分的蒸腾作用拉力成为养分运输的动力(Barker et al., 2005).另外土壤中氮的矿化和硝化作用与水分也有密切的联系(王双, 2008).因此, 干旱可能使溶解于水中的养分降低, 同时使植物的蒸腾作用降低, 导致减少降水时养分添加对植物的促进作用弱于其他处理. ...
Plant trait responses to grazing—A global synthesis 1 2007
... 植物功能性状是指植物具有的与其定植、存活、生长和死亡紧密相关的一系列核心植物属性, 且这些属性能够反映植物个体对环境的响应和适应, 将环境变化同植物群落的结构和功能联系起来(Díaz et al., 2007; Swenson & Enquist, 2007; Violle et al., 2007; Lavorel et al., 2011; 刘晓娟和马克平, 2015).全球变化和植物功能性状之间的关系是当前植物生态学研究的热点问题. ...
Effects of alternate wetting and drying irrigation regime and nitrogen fertilizer on yield and nitrogen use efficiency of irrigated rice in the Sahel 1 2018
... 大部分陆地生态系统都受到N和P限制, 适当添加养分可以提高植物生产力(LeBauer & Treseder, 2008; Djaman et al., 2018).有****指出氮供应充分可以提高植物的光合能力, 进而促进植物的生长, 提高植物生物量(Chen et al., 2005; Jan et al., 2018).在干旱半干旱地区, 植物生长不仅受到养分的限制,较低的降水量导致水分也成为该地区的主要限制因子(Rao & Allen, 2010).本研究结果表明, 降水量增加显著提高了羊草地上生物量.在减少降水量的情况下, 氮添加、氮磷共同添加对羊草地上生物量的促进效应明显低于正常和增加降水量.这是因为植物对养分的吸收和运输往往依赖于水分, 养分要溶解于水中才能被植物吸收, 同时植物地上部分的蒸腾作用拉力成为养分运输的动力(Barker et al., 2005).另外土壤中氮的矿化和硝化作用与水分也有密切的联系(王双, 2008).因此, 干旱可能使溶解于水中的养分降低, 同时使植物的蒸腾作用降低, 导致减少降水时养分添加对植物的促进作用弱于其他处理. ...
Plant functional traits: Soil and ecosystem services 1 2017
... 功能性状作为解释变量, 对生态系统功能和服务的变化有显著的指示作用(Garnier et al., 2004; Mokany et al., 2010; Moor et al., 2015; Osborne et al., 2018), 能够显示植物对环境异质性的适应能力(Reich & Oleksyn, 2004; Faucon et al., 2017).植被生长过程中, 功能性状会受到降水量和施肥处理不同程度的影响.降水量增加使植株高度增高, 叶面积增大, 净光合速率升高(曾小平等, 2004; 许振柱和周广胜, 2005; 孟凡超等, 2014; Yue et al., 2019), 本研究结果与之一致, 主要原因是水分充足与否直接影响到羊草根系的发育, 根系发育的良好程度又直接影响到植物对土壤养分的吸收及养分在植物体内的循环, 进而对植物功能性状产生影响(李秀芳, 2011; Balachowski & Volaire, 2018).已有研究表明, 植物的光合能力与相对生长速率紧密相关(Shipley, 2006; Morgan et al., 2011), 叶面积反映了叶片对光的截获能力(Poorter & Rozendaal, 2008).因此, 降水量增加与氮磷共施条件下较高的光合速率和较大的叶面积可能是较高的叶生物量和茎生物量的主要原因.Fonseca等(2000)的研究表明澳大利亚东南部多年生植物的SLA随降水量的增加而增加; 驼绒藜(Ceratoides latens)的SLA与降水量呈正相关关系(朱军涛等, 2010); 植物叶质量也会随降水量的增加而减小(Wright et al., 2004; Rosbakh et al., 2015; Buckley et al., 2019).而本研究中, 降水量对羊草SLA没有显著影响(p > 0.05), 可能是由于羊草对短期水分处理响应不明显(Gao et al., 2015; Forrestel et al., 2017). ...
Shifts in trait-combinations along rainfall and phosphorus gradients 1 2000
... 功能性状作为解释变量, 对生态系统功能和服务的变化有显著的指示作用(Garnier et al., 2004; Mokany et al., 2010; Moor et al., 2015; Osborne et al., 2018), 能够显示植物对环境异质性的适应能力(Reich & Oleksyn, 2004; Faucon et al., 2017).植被生长过程中, 功能性状会受到降水量和施肥处理不同程度的影响.降水量增加使植株高度增高, 叶面积增大, 净光合速率升高(曾小平等, 2004; 许振柱和周广胜, 2005; 孟凡超等, 2014; Yue et al., 2019), 本研究结果与之一致, 主要原因是水分充足与否直接影响到羊草根系的发育, 根系发育的良好程度又直接影响到植物对土壤养分的吸收及养分在植物体内的循环, 进而对植物功能性状产生影响(李秀芳, 2011; Balachowski & Volaire, 2018).已有研究表明, 植物的光合能力与相对生长速率紧密相关(Shipley, 2006; Morgan et al., 2011), 叶面积反映了叶片对光的截获能力(Poorter & Rozendaal, 2008).因此, 降水量增加与氮磷共施条件下较高的光合速率和较大的叶面积可能是较高的叶生物量和茎生物量的主要原因.Fonseca等(2000)的研究表明澳大利亚东南部多年生植物的SLA随降水量的增加而增加; 驼绒藜(Ceratoides latens)的SLA与降水量呈正相关关系(朱军涛等, 2010); 植物叶质量也会随降水量的增加而减小(Wright et al., 2004; Rosbakh et al., 2015; Buckley et al., 2019).而本研究中, 降水量对羊草SLA没有显著影响(p > 0.05), 可能是由于羊草对短期水分处理响应不明显(Gao et al., 2015; Forrestel et al., 2017). ...
Different clades and traits yield similar grassland functional responses 1 2017
... 功能性状作为解释变量, 对生态系统功能和服务的变化有显著的指示作用(Garnier et al., 2004; Mokany et al., 2010; Moor et al., 2015; Osborne et al., 2018), 能够显示植物对环境异质性的适应能力(Reich & Oleksyn, 2004; Faucon et al., 2017).植被生长过程中, 功能性状会受到降水量和施肥处理不同程度的影响.降水量增加使植株高度增高, 叶面积增大, 净光合速率升高(曾小平等, 2004; 许振柱和周广胜, 2005; 孟凡超等, 2014; Yue et al., 2019), 本研究结果与之一致, 主要原因是水分充足与否直接影响到羊草根系的发育, 根系发育的良好程度又直接影响到植物对土壤养分的吸收及养分在植物体内的循环, 进而对植物功能性状产生影响(李秀芳, 2011; Balachowski & Volaire, 2018).已有研究表明, 植物的光合能力与相对生长速率紧密相关(Shipley, 2006; Morgan et al., 2011), 叶面积反映了叶片对光的截获能力(Poorter & Rozendaal, 2008).因此, 降水量增加与氮磷共施条件下较高的光合速率和较大的叶面积可能是较高的叶生物量和茎生物量的主要原因.Fonseca等(2000)的研究表明澳大利亚东南部多年生植物的SLA随降水量的增加而增加; 驼绒藜(Ceratoides latens)的SLA与降水量呈正相关关系(朱军涛等, 2010); 植物叶质量也会随降水量的增加而减小(Wright et al., 2004; Rosbakh et al., 2015; Buckley et al., 2019).而本研究中, 降水量对羊草SLA没有显著影响(p > 0.05), 可能是由于羊草对短期水分处理响应不明显(Gao et al., 2015; Forrestel et al., 2017). ...
Effects of rainfall pattern on the growth and fecundity of a dominant dune annual in a semi-arid ecosystem 1 2015
... 功能性状作为解释变量, 对生态系统功能和服务的变化有显著的指示作用(Garnier et al., 2004; Mokany et al., 2010; Moor et al., 2015; Osborne et al., 2018), 能够显示植物对环境异质性的适应能力(Reich & Oleksyn, 2004; Faucon et al., 2017).植被生长过程中, 功能性状会受到降水量和施肥处理不同程度的影响.降水量增加使植株高度增高, 叶面积增大, 净光合速率升高(曾小平等, 2004; 许振柱和周广胜, 2005; 孟凡超等, 2014; Yue et al., 2019), 本研究结果与之一致, 主要原因是水分充足与否直接影响到羊草根系的发育, 根系发育的良好程度又直接影响到植物对土壤养分的吸收及养分在植物体内的循环, 进而对植物功能性状产生影响(李秀芳, 2011; Balachowski & Volaire, 2018).已有研究表明, 植物的光合能力与相对生长速率紧密相关(Shipley, 2006; Morgan et al., 2011), 叶面积反映了叶片对光的截获能力(Poorter & Rozendaal, 2008).因此, 降水量增加与氮磷共施条件下较高的光合速率和较大的叶面积可能是较高的叶生物量和茎生物量的主要原因.Fonseca等(2000)的研究表明澳大利亚东南部多年生植物的SLA随降水量的增加而增加; 驼绒藜(Ceratoides latens)的SLA与降水量呈正相关关系(朱军涛等, 2010); 植物叶质量也会随降水量的增加而减小(Wright et al., 2004; Rosbakh et al., 2015; Buckley et al., 2019).而本研究中, 降水量对羊草SLA没有显著影响(p > 0.05), 可能是由于羊草对短期水分处理响应不明显(Gao et al., 2015; Forrestel et al., 2017). ...
... 盆栽试验开始于2016年6月中旬, 在中国科学院东北地理与农业生态研究所(125.38° E, 43.98° N, 海拔190 m)进行, 该区域气候属于温带季风气候, 1953-2012年的平均年降水量为498.0 mm, 年降水量极大值为754.0 mm (1956年), 比平均水平高出约50%, 年降水量极小值为244.1 mm (1982年)(李晶等, 2015), 比平均水平低50%.土壤类型为黑土, 有机质平均含量为2.83%、全N含量为1 368.56 mg·kg-1、全P含量为669.18 mg·kg-1.试验在遮雨棚内进行, 试验盆直径30 cm, 高27 cm, 盆内装入试验地附近的0-20 cm土层的过筛土, 每盆移栽10株大小均一的羊草.晴天时收起遮雨棚, 下雨时进行遮挡以防止盆栽接收自然降水.定期浇水, 剔除杂草, 保证羊草植株生长. ...
长春市降雨特征及雨型分析研究 1 2015
... 盆栽试验开始于2016年6月中旬, 在中国科学院东北地理与农业生态研究所(125.38° E, 43.98° N, 海拔190 m)进行, 该区域气候属于温带季风气候, 1953-2012年的平均年降水量为498.0 mm, 年降水量极大值为754.0 mm (1956年), 比平均水平高出约50%, 年降水量极小值为244.1 mm (1982年)(李晶等, 2015), 比平均水平低50%.土壤类型为黑土, 有机质平均含量为2.83%、全N含量为1 368.56 mg·kg-1、全P含量为669.18 mg·kg-1.试验在遮雨棚内进行, 试验盆直径30 cm, 高27 cm, 盆内装入试验地附近的0-20 cm土层的过筛土, 每盆移栽10株大小均一的羊草.晴天时收起遮雨棚, 下雨时进行遮挡以防止盆栽接收自然降水.定期浇水, 剔除杂草, 保证羊草植株生长. ...
根区水肥空间耦合对冬小麦生长及产量的影响 1 2010
... 水分和养分是植物生产力水平提高的关键因素(李生秀等, 1994; 徐学选等, 1995; 汪德水, 1999; 李开峰等, 2010), 二者相互作用、互相影响, 从而对植物的生长发育以及产量产生水肥耦合效应(张广涛等, 2007).水分变化在植物的生长发育和生理生态过程中扮演着重要角色, 当降水量增加时, 植物能够通过调节其叶片干物质含量、比叶面积来适应环境(赵新风等, 2014; Kunstler et al., 2016); 随着降水量的增加, 植物的叶质量也会减小(Wright et al., 2004).一般情况下, 植物地上生物量随降水量的增加而增加(Hu et al., 2010; 张志南等, 2014).虽然近年来降水量与地上生物量之间的正相关关系已经逐渐得到广泛证实, 但其关系形态在不同研究中还存在一定差异(Yang et al., 2008; Zhang et al., 2019).土壤水分显著影响土壤养分有效性乃至陆地生态系统养分循环进程, 合理控水对于提高生产力具有重要意义. ...
根区水肥空间耦合对冬小麦生长及产量的影响 1 2010
... 水分和养分是植物生产力水平提高的关键因素(李生秀等, 1994; 徐学选等, 1995; 汪德水, 1999; 李开峰等, 2010), 二者相互作用、互相影响, 从而对植物的生长发育以及产量产生水肥耦合效应(张广涛等, 2007).水分变化在植物的生长发育和生理生态过程中扮演着重要角色, 当降水量增加时, 植物能够通过调节其叶片干物质含量、比叶面积来适应环境(赵新风等, 2014; Kunstler et al., 2016); 随着降水量的增加, 植物的叶质量也会减小(Wright et al., 2004).一般情况下, 植物地上生物量随降水量的增加而增加(Hu et al., 2010; 张志南等, 2014).虽然近年来降水量与地上生物量之间的正相关关系已经逐渐得到广泛证实, 但其关系形态在不同研究中还存在一定差异(Yang et al., 2008; Zhang et al., 2019).土壤水分显著影响土壤养分有效性乃至陆地生态系统养分循环进程, 合理控水对于提高生产力具有重要意义. ...
施用氮肥对提高旱地作物利用土壤水分的作用机理和效果 1 1994
... 水分和养分是植物生产力水平提高的关键因素(李生秀等, 1994; 徐学选等, 1995; 汪德水, 1999; 李开峰等, 2010), 二者相互作用、互相影响, 从而对植物的生长发育以及产量产生水肥耦合效应(张广涛等, 2007).水分变化在植物的生长发育和生理生态过程中扮演着重要角色, 当降水量增加时, 植物能够通过调节其叶片干物质含量、比叶面积来适应环境(赵新风等, 2014; Kunstler et al., 2016); 随着降水量的增加, 植物的叶质量也会减小(Wright et al., 2004).一般情况下, 植物地上生物量随降水量的增加而增加(Hu et al., 2010; 张志南等, 2014).虽然近年来降水量与地上生物量之间的正相关关系已经逐渐得到广泛证实, 但其关系形态在不同研究中还存在一定差异(Yang et al., 2008; Zhang et al., 2019).土壤水分显著影响土壤养分有效性乃至陆地生态系统养分循环进程, 合理控水对于提高生产力具有重要意义. ...
施用氮肥对提高旱地作物利用土壤水分的作用机理和效果 1 1994
... 水分和养分是植物生产力水平提高的关键因素(李生秀等, 1994; 徐学选等, 1995; 汪德水, 1999; 李开峰等, 2010), 二者相互作用、互相影响, 从而对植物的生长发育以及产量产生水肥耦合效应(张广涛等, 2007).水分变化在植物的生长发育和生理生态过程中扮演着重要角色, 当降水量增加时, 植物能够通过调节其叶片干物质含量、比叶面积来适应环境(赵新风等, 2014; Kunstler et al., 2016); 随着降水量的增加, 植物的叶质量也会减小(Wright et al., 2004).一般情况下, 植物地上生物量随降水量的增加而增加(Hu et al., 2010; 张志南等, 2014).虽然近年来降水量与地上生物量之间的正相关关系已经逐渐得到广泛证实, 但其关系形态在不同研究中还存在一定差异(Yang et al., 2008; Zhang et al., 2019).土壤水分显著影响土壤养分有效性乃至陆地生态系统养分循环进程, 合理控水对于提高生产力具有重要意义. ...
水肥互作对夏玉米生长与养分吸收及根系活性的影响 1 2011
... 功能性状作为解释变量, 对生态系统功能和服务的变化有显著的指示作用(Garnier et al., 2004; Mokany et al., 2010; Moor et al., 2015; Osborne et al., 2018), 能够显示植物对环境异质性的适应能力(Reich & Oleksyn, 2004; Faucon et al., 2017).植被生长过程中, 功能性状会受到降水量和施肥处理不同程度的影响.降水量增加使植株高度增高, 叶面积增大, 净光合速率升高(曾小平等, 2004; 许振柱和周广胜, 2005; 孟凡超等, 2014; Yue et al., 2019), 本研究结果与之一致, 主要原因是水分充足与否直接影响到羊草根系的发育, 根系发育的良好程度又直接影响到植物对土壤养分的吸收及养分在植物体内的循环, 进而对植物功能性状产生影响(李秀芳, 2011; Balachowski & Volaire, 2018).已有研究表明, 植物的光合能力与相对生长速率紧密相关(Shipley, 2006; Morgan et al., 2011), 叶面积反映了叶片对光的截获能力(Poorter & Rozendaal, 2008).因此, 降水量增加与氮磷共施条件下较高的光合速率和较大的叶面积可能是较高的叶生物量和茎生物量的主要原因.Fonseca等(2000)的研究表明澳大利亚东南部多年生植物的SLA随降水量的增加而增加; 驼绒藜(Ceratoides latens)的SLA与降水量呈正相关关系(朱军涛等, 2010); 植物叶质量也会随降水量的增加而减小(Wright et al., 2004; Rosbakh et al., 2015; Buckley et al., 2019).而本研究中, 降水量对羊草SLA没有显著影响(p > 0.05), 可能是由于羊草对短期水分处理响应不明显(Gao et al., 2015; Forrestel et al., 2017). ...
水肥互作对夏玉米生长与养分吸收及根系活性的影响 1 2011
... 功能性状作为解释变量, 对生态系统功能和服务的变化有显著的指示作用(Garnier et al., 2004; Mokany et al., 2010; Moor et al., 2015; Osborne et al., 2018), 能够显示植物对环境异质性的适应能力(Reich & Oleksyn, 2004; Faucon et al., 2017).植被生长过程中, 功能性状会受到降水量和施肥处理不同程度的影响.降水量增加使植株高度增高, 叶面积增大, 净光合速率升高(曾小平等, 2004; 许振柱和周广胜, 2005; 孟凡超等, 2014; Yue et al., 2019), 本研究结果与之一致, 主要原因是水分充足与否直接影响到羊草根系的发育, 根系发育的良好程度又直接影响到植物对土壤养分的吸收及养分在植物体内的循环, 进而对植物功能性状产生影响(李秀芳, 2011; Balachowski & Volaire, 2018).已有研究表明, 植物的光合能力与相对生长速率紧密相关(Shipley, 2006; Morgan et al., 2011), 叶面积反映了叶片对光的截获能力(Poorter & Rozendaal, 2008).因此, 降水量增加与氮磷共施条件下较高的光合速率和较大的叶面积可能是较高的叶生物量和茎生物量的主要原因.Fonseca等(2000)的研究表明澳大利亚东南部多年生植物的SLA随降水量的增加而增加; 驼绒藜(Ceratoides latens)的SLA与降水量呈正相关关系(朱军涛等, 2010); 植物叶质量也会随降水量的增加而减小(Wright et al., 2004; Rosbakh et al., 2015; Buckley et al., 2019).而本研究中, 降水量对羊草SLA没有显著影响(p > 0.05), 可能是由于羊草对短期水分处理响应不明显(Gao et al., 2015; Forrestel et al., 2017). ...
... 功能性状作为解释变量, 对生态系统功能和服务的变化有显著的指示作用(Garnier et al., 2004; Mokany et al., 2010; Moor et al., 2015; Osborne et al., 2018), 能够显示植物对环境异质性的适应能力(Reich & Oleksyn, 2004; Faucon et al., 2017).植被生长过程中, 功能性状会受到降水量和施肥处理不同程度的影响.降水量增加使植株高度增高, 叶面积增大, 净光合速率升高(曾小平等, 2004; 许振柱和周广胜, 2005; 孟凡超等, 2014; Yue et al., 2019), 本研究结果与之一致, 主要原因是水分充足与否直接影响到羊草根系的发育, 根系发育的良好程度又直接影响到植物对土壤养分的吸收及养分在植物体内的循环, 进而对植物功能性状产生影响(李秀芳, 2011; Balachowski & Volaire, 2018).已有研究表明, 植物的光合能力与相对生长速率紧密相关(Shipley, 2006; Morgan et al., 2011), 叶面积反映了叶片对光的截获能力(Poorter & Rozendaal, 2008).因此, 降水量增加与氮磷共施条件下较高的光合速率和较大的叶面积可能是较高的叶生物量和茎生物量的主要原因.Fonseca等(2000)的研究表明澳大利亚东南部多年生植物的SLA随降水量的增加而增加; 驼绒藜(Ceratoides latens)的SLA与降水量呈正相关关系(朱军涛等, 2010); 植物叶质量也会随降水量的增加而减小(Wright et al., 2004; Rosbakh et al., 2015; Buckley et al., 2019).而本研究中, 降水量对羊草SLA没有显著影响(p > 0.05), 可能是由于羊草对短期水分处理响应不明显(Gao et al., 2015; Forrestel et al., 2017). ...
CO2浓度升高和降水增加协同作用对玉米产量及生长发育的影响 1 2014
... 功能性状作为解释变量, 对生态系统功能和服务的变化有显著的指示作用(Garnier et al., 2004; Mokany et al., 2010; Moor et al., 2015; Osborne et al., 2018), 能够显示植物对环境异质性的适应能力(Reich & Oleksyn, 2004; Faucon et al., 2017).植被生长过程中, 功能性状会受到降水量和施肥处理不同程度的影响.降水量增加使植株高度增高, 叶面积增大, 净光合速率升高(曾小平等, 2004; 许振柱和周广胜, 2005; 孟凡超等, 2014; Yue et al., 2019), 本研究结果与之一致, 主要原因是水分充足与否直接影响到羊草根系的发育, 根系发育的良好程度又直接影响到植物对土壤养分的吸收及养分在植物体内的循环, 进而对植物功能性状产生影响(李秀芳, 2011; Balachowski & Volaire, 2018).已有研究表明, 植物的光合能力与相对生长速率紧密相关(Shipley, 2006; Morgan et al., 2011), 叶面积反映了叶片对光的截获能力(Poorter & Rozendaal, 2008).因此, 降水量增加与氮磷共施条件下较高的光合速率和较大的叶面积可能是较高的叶生物量和茎生物量的主要原因.Fonseca等(2000)的研究表明澳大利亚东南部多年生植物的SLA随降水量的增加而增加; 驼绒藜(Ceratoides latens)的SLA与降水量呈正相关关系(朱军涛等, 2010); 植物叶质量也会随降水量的增加而减小(Wright et al., 2004; Rosbakh et al., 2015; Buckley et al., 2019).而本研究中, 降水量对羊草SLA没有显著影响(p > 0.05), 可能是由于羊草对短期水分处理响应不明显(Gao et al., 2015; Forrestel et al., 2017). ...
Functional identity is more important than diversity in influencing ecosystem processes in a temperate native grassland 1 2010
... 功能性状作为解释变量, 对生态系统功能和服务的变化有显著的指示作用(Garnier et al., 2004; Mokany et al., 2010; Moor et al., 2015; Osborne et al., 2018), 能够显示植物对环境异质性的适应能力(Reich & Oleksyn, 2004; Faucon et al., 2017).植被生长过程中, 功能性状会受到降水量和施肥处理不同程度的影响.降水量增加使植株高度增高, 叶面积增大, 净光合速率升高(曾小平等, 2004; 许振柱和周广胜, 2005; 孟凡超等, 2014; Yue et al., 2019), 本研究结果与之一致, 主要原因是水分充足与否直接影响到羊草根系的发育, 根系发育的良好程度又直接影响到植物对土壤养分的吸收及养分在植物体内的循环, 进而对植物功能性状产生影响(李秀芳, 2011; Balachowski & Volaire, 2018).已有研究表明, 植物的光合能力与相对生长速率紧密相关(Shipley, 2006; Morgan et al., 2011), 叶面积反映了叶片对光的截获能力(Poorter & Rozendaal, 2008).因此, 降水量增加与氮磷共施条件下较高的光合速率和较大的叶面积可能是较高的叶生物量和茎生物量的主要原因.Fonseca等(2000)的研究表明澳大利亚东南部多年生植物的SLA随降水量的增加而增加; 驼绒藜(Ceratoides latens)的SLA与降水量呈正相关关系(朱军涛等, 2010); 植物叶质量也会随降水量的增加而减小(Wright et al., 2004; Rosbakh et al., 2015; Buckley et al., 2019).而本研究中, 降水量对羊草SLA没有显著影响(p > 0.05), 可能是由于羊草对短期水分处理响应不明显(Gao et al., 2015; Forrestel et al., 2017). ...
Predicting climate change effects on wetland ecosystem services using species distribution modeling and plant functional traits 1 2015
... 功能性状作为解释变量, 对生态系统功能和服务的变化有显著的指示作用(Garnier et al., 2004; Mokany et al., 2010; Moor et al., 2015; Osborne et al., 2018), 能够显示植物对环境异质性的适应能力(Reich & Oleksyn, 2004; Faucon et al., 2017).植被生长过程中, 功能性状会受到降水量和施肥处理不同程度的影响.降水量增加使植株高度增高, 叶面积增大, 净光合速率升高(曾小平等, 2004; 许振柱和周广胜, 2005; 孟凡超等, 2014; Yue et al., 2019), 本研究结果与之一致, 主要原因是水分充足与否直接影响到羊草根系的发育, 根系发育的良好程度又直接影响到植物对土壤养分的吸收及养分在植物体内的循环, 进而对植物功能性状产生影响(李秀芳, 2011; Balachowski & Volaire, 2018).已有研究表明, 植物的光合能力与相对生长速率紧密相关(Shipley, 2006; Morgan et al., 2011), 叶面积反映了叶片对光的截获能力(Poorter & Rozendaal, 2008).因此, 降水量增加与氮磷共施条件下较高的光合速率和较大的叶面积可能是较高的叶生物量和茎生物量的主要原因.Fonseca等(2000)的研究表明澳大利亚东南部多年生植物的SLA随降水量的增加而增加; 驼绒藜(Ceratoides latens)的SLA与降水量呈正相关关系(朱军涛等, 2010); 植物叶质量也会随降水量的增加而减小(Wright et al., 2004; Rosbakh et al., 2015; Buckley et al., 2019).而本研究中, 降水量对羊草SLA没有显著影响(p > 0.05), 可能是由于羊草对短期水分处理响应不明显(Gao et al., 2015; Forrestel et al., 2017). ...
C4 grasses prosper as carbon dioxide eliminates desiccation in warmed semi-arid grassland 1 2011
... 功能性状作为解释变量, 对生态系统功能和服务的变化有显著的指示作用(Garnier et al., 2004; Mokany et al., 2010; Moor et al., 2015; Osborne et al., 2018), 能够显示植物对环境异质性的适应能力(Reich & Oleksyn, 2004; Faucon et al., 2017).植被生长过程中, 功能性状会受到降水量和施肥处理不同程度的影响.降水量增加使植株高度增高, 叶面积增大, 净光合速率升高(曾小平等, 2004; 许振柱和周广胜, 2005; 孟凡超等, 2014; Yue et al., 2019), 本研究结果与之一致, 主要原因是水分充足与否直接影响到羊草根系的发育, 根系发育的良好程度又直接影响到植物对土壤养分的吸收及养分在植物体内的循环, 进而对植物功能性状产生影响(李秀芳, 2011; Balachowski & Volaire, 2018).已有研究表明, 植物的光合能力与相对生长速率紧密相关(Shipley, 2006; Morgan et al., 2011), 叶面积反映了叶片对光的截获能力(Poorter & Rozendaal, 2008).因此, 降水量增加与氮磷共施条件下较高的光合速率和较大的叶面积可能是较高的叶生物量和茎生物量的主要原因.Fonseca等(2000)的研究表明澳大利亚东南部多年生植物的SLA随降水量的增加而增加; 驼绒藜(Ceratoides latens)的SLA与降水量呈正相关关系(朱军涛等, 2010); 植物叶质量也会随降水量的增加而减小(Wright et al., 2004; Rosbakh et al., 2015; Buckley et al., 2019).而本研究中, 降水量对羊草SLA没有显著影响(p > 0.05), 可能是由于羊草对短期水分处理响应不明显(Gao et al., 2015; Forrestel et al., 2017). ...
Human impacts in African savannas are mediated by plant functional traits 1 2018
... 功能性状作为解释变量, 对生态系统功能和服务的变化有显著的指示作用(Garnier et al., 2004; Mokany et al., 2010; Moor et al., 2015; Osborne et al., 2018), 能够显示植物对环境异质性的适应能力(Reich & Oleksyn, 2004; Faucon et al., 2017).植被生长过程中, 功能性状会受到降水量和施肥处理不同程度的影响.降水量增加使植株高度增高, 叶面积增大, 净光合速率升高(曾小平等, 2004; 许振柱和周广胜, 2005; 孟凡超等, 2014; Yue et al., 2019), 本研究结果与之一致, 主要原因是水分充足与否直接影响到羊草根系的发育, 根系发育的良好程度又直接影响到植物对土壤养分的吸收及养分在植物体内的循环, 进而对植物功能性状产生影响(李秀芳, 2011; Balachowski & Volaire, 2018).已有研究表明, 植物的光合能力与相对生长速率紧密相关(Shipley, 2006; Morgan et al., 2011), 叶面积反映了叶片对光的截获能力(Poorter & Rozendaal, 2008).因此, 降水量增加与氮磷共施条件下较高的光合速率和较大的叶面积可能是较高的叶生物量和茎生物量的主要原因.Fonseca等(2000)的研究表明澳大利亚东南部多年生植物的SLA随降水量的增加而增加; 驼绒藜(Ceratoides latens)的SLA与降水量呈正相关关系(朱军涛等, 2010); 植物叶质量也会随降水量的增加而减小(Wright et al., 2004; Rosbakh et al., 2015; Buckley et al., 2019).而本研究中, 降水量对羊草SLA没有显著影响(p > 0.05), 可能是由于羊草对短期水分处理响应不明显(Gao et al., 2015; Forrestel et al., 2017). ...
Leaf size and leaf display of thirty-eight tropical tree species 1 2008
... 功能性状作为解释变量, 对生态系统功能和服务的变化有显著的指示作用(Garnier et al., 2004; Mokany et al., 2010; Moor et al., 2015; Osborne et al., 2018), 能够显示植物对环境异质性的适应能力(Reich & Oleksyn, 2004; Faucon et al., 2017).植被生长过程中, 功能性状会受到降水量和施肥处理不同程度的影响.降水量增加使植株高度增高, 叶面积增大, 净光合速率升高(曾小平等, 2004; 许振柱和周广胜, 2005; 孟凡超等, 2014; Yue et al., 2019), 本研究结果与之一致, 主要原因是水分充足与否直接影响到羊草根系的发育, 根系发育的良好程度又直接影响到植物对土壤养分的吸收及养分在植物体内的循环, 进而对植物功能性状产生影响(李秀芳, 2011; Balachowski & Volaire, 2018).已有研究表明, 植物的光合能力与相对生长速率紧密相关(Shipley, 2006; Morgan et al., 2011), 叶面积反映了叶片对光的截获能力(Poorter & Rozendaal, 2008).因此, 降水量增加与氮磷共施条件下较高的光合速率和较大的叶面积可能是较高的叶生物量和茎生物量的主要原因.Fonseca等(2000)的研究表明澳大利亚东南部多年生植物的SLA随降水量的增加而增加; 驼绒藜(Ceratoides latens)的SLA与降水量呈正相关关系(朱军涛等, 2010); 植物叶质量也会随降水量的增加而减小(Wright et al., 2004; Rosbakh et al., 2015; Buckley et al., 2019).而本研究中, 降水量对羊草SLA没有显著影响(p > 0.05), 可能是由于羊草对短期水分处理响应不明显(Gao et al., 2015; Forrestel et al., 2017). ...
Combined effects of precipitation and nitrogen deposition on native and invasive winter annual production in California deserts 1 2010
... 大部分陆地生态系统都受到N和P限制, 适当添加养分可以提高植物生产力(LeBauer & Treseder, 2008; Djaman et al., 2018).有****指出氮供应充分可以提高植物的光合能力, 进而促进植物的生长, 提高植物生物量(Chen et al., 2005; Jan et al., 2018).在干旱半干旱地区, 植物生长不仅受到养分的限制,较低的降水量导致水分也成为该地区的主要限制因子(Rao & Allen, 2010).本研究结果表明, 降水量增加显著提高了羊草地上生物量.在减少降水量的情况下, 氮添加、氮磷共同添加对羊草地上生物量的促进效应明显低于正常和增加降水量.这是因为植物对养分的吸收和运输往往依赖于水分, 养分要溶解于水中才能被植物吸收, 同时植物地上部分的蒸腾作用拉力成为养分运输的动力(Barker et al., 2005).另外土壤中氮的矿化和硝化作用与水分也有密切的联系(王双, 2008).因此, 干旱可能使溶解于水中的养分降低, 同时使植物的蒸腾作用降低, 导致减少降水时养分添加对植物的促进作用弱于其他处理. ...
Global patterns of plant leaf N and P in relation to temperature and latitude 1 2004
... 功能性状作为解释变量, 对生态系统功能和服务的变化有显著的指示作用(Garnier et al., 2004; Mokany et al., 2010; Moor et al., 2015; Osborne et al., 2018), 能够显示植物对环境异质性的适应能力(Reich & Oleksyn, 2004; Faucon et al., 2017).植被生长过程中, 功能性状会受到降水量和施肥处理不同程度的影响.降水量增加使植株高度增高, 叶面积增大, 净光合速率升高(曾小平等, 2004; 许振柱和周广胜, 2005; 孟凡超等, 2014; Yue et al., 2019), 本研究结果与之一致, 主要原因是水分充足与否直接影响到羊草根系的发育, 根系发育的良好程度又直接影响到植物对土壤养分的吸收及养分在植物体内的循环, 进而对植物功能性状产生影响(李秀芳, 2011; Balachowski & Volaire, 2018).已有研究表明, 植物的光合能力与相对生长速率紧密相关(Shipley, 2006; Morgan et al., 2011), 叶面积反映了叶片对光的截获能力(Poorter & Rozendaal, 2008).因此, 降水量增加与氮磷共施条件下较高的光合速率和较大的叶面积可能是较高的叶生物量和茎生物量的主要原因.Fonseca等(2000)的研究表明澳大利亚东南部多年生植物的SLA随降水量的增加而增加; 驼绒藜(Ceratoides latens)的SLA与降水量呈正相关关系(朱军涛等, 2010); 植物叶质量也会随降水量的增加而减小(Wright et al., 2004; Rosbakh et al., 2015; Buckley et al., 2019).而本研究中, 降水量对羊草SLA没有显著影响(p > 0.05), 可能是由于羊草对短期水分处理响应不明显(Gao et al., 2015; Forrestel et al., 2017). ...
Specific leaf area correlates with temperature: New evidence of trait variation at the population, species and community levels 1 2015
... 功能性状作为解释变量, 对生态系统功能和服务的变化有显著的指示作用(Garnier et al., 2004; Mokany et al., 2010; Moor et al., 2015; Osborne et al., 2018), 能够显示植物对环境异质性的适应能力(Reich & Oleksyn, 2004; Faucon et al., 2017).植被生长过程中, 功能性状会受到降水量和施肥处理不同程度的影响.降水量增加使植株高度增高, 叶面积增大, 净光合速率升高(曾小平等, 2004; 许振柱和周广胜, 2005; 孟凡超等, 2014; Yue et al., 2019), 本研究结果与之一致, 主要原因是水分充足与否直接影响到羊草根系的发育, 根系发育的良好程度又直接影响到植物对土壤养分的吸收及养分在植物体内的循环, 进而对植物功能性状产生影响(李秀芳, 2011; Balachowski & Volaire, 2018).已有研究表明, 植物的光合能力与相对生长速率紧密相关(Shipley, 2006; Morgan et al., 2011), 叶面积反映了叶片对光的截获能力(Poorter & Rozendaal, 2008).因此, 降水量增加与氮磷共施条件下较高的光合速率和较大的叶面积可能是较高的叶生物量和茎生物量的主要原因.Fonseca等(2000)的研究表明澳大利亚东南部多年生植物的SLA随降水量的增加而增加; 驼绒藜(Ceratoides latens)的SLA与降水量呈正相关关系(朱军涛等, 2010); 植物叶质量也会随降水量的增加而减小(Wright et al., 2004; Rosbakh et al., 2015; Buckley et al., 2019).而本研究中, 降水量对羊草SLA没有显著影响(p > 0.05), 可能是由于羊草对短期水分处理响应不明显(Gao et al., 2015; Forrestel et al., 2017). ...
Net assimilation rate, specific leaf area and leaf mass ratio: Which is most closely correlated with relative growth rate? A meta-analysis 1 2006
... 功能性状作为解释变量, 对生态系统功能和服务的变化有显著的指示作用(Garnier et al., 2004; Mokany et al., 2010; Moor et al., 2015; Osborne et al., 2018), 能够显示植物对环境异质性的适应能力(Reich & Oleksyn, 2004; Faucon et al., 2017).植被生长过程中, 功能性状会受到降水量和施肥处理不同程度的影响.降水量增加使植株高度增高, 叶面积增大, 净光合速率升高(曾小平等, 2004; 许振柱和周广胜, 2005; 孟凡超等, 2014; Yue et al., 2019), 本研究结果与之一致, 主要原因是水分充足与否直接影响到羊草根系的发育, 根系发育的良好程度又直接影响到植物对土壤养分的吸收及养分在植物体内的循环, 进而对植物功能性状产生影响(李秀芳, 2011; Balachowski & Volaire, 2018).已有研究表明, 植物的光合能力与相对生长速率紧密相关(Shipley, 2006; Morgan et al., 2011), 叶面积反映了叶片对光的截获能力(Poorter & Rozendaal, 2008).因此, 降水量增加与氮磷共施条件下较高的光合速率和较大的叶面积可能是较高的叶生物量和茎生物量的主要原因.Fonseca等(2000)的研究表明澳大利亚东南部多年生植物的SLA随降水量的增加而增加; 驼绒藜(Ceratoides latens)的SLA与降水量呈正相关关系(朱军涛等, 2010); 植物叶质量也会随降水量的增加而减小(Wright et al., 2004; Rosbakh et al., 2015; Buckley et al., 2019).而本研究中, 降水量对羊草SLA没有显著影响(p > 0.05), 可能是由于羊草对短期水分处理响应不明显(Gao et al., 2015; Forrestel et al., 2017). ...
羊草功能性状和地上生物量对氮素添加的响应 1 2016
... 短期的养分添加主要影响羊草的生理性状, 如叶片光合速率、蒸腾速率和水分利用效率均显著高于不施肥处理, 这些性状主要与光合作用相关.这说明养分添加缓解了植物对养分的竞争, 使物种对资源的竞争从地下部分转向地上部分, 即增加植物对光资源的竞争(Bobbink et al., 2010).然而, 养分添加对羊草形态性状的影响作用相对较小, 这与万宏伟等(2008)的研究结果不一致, 可能是因为本研究时间较短, 植物性状随着资源水平的改变, 首先是调整生理性状使其快速适应, 而形态性状相对滞后, 需要较长的时间才能发生显著变化(宋彦涛等, 2016; Yang et al., 2019). ...
羊草功能性状和地上生物量对氮素添加的响应 1 2016
... 短期的养分添加主要影响羊草的生理性状, 如叶片光合速率、蒸腾速率和水分利用效率均显著高于不施肥处理, 这些性状主要与光合作用相关.这说明养分添加缓解了植物对养分的竞争, 使物种对资源的竞争从地下部分转向地上部分, 即增加植物对光资源的竞争(Bobbink et al., 2010).然而, 养分添加对羊草形态性状的影响作用相对较小, 这与万宏伟等(2008)的研究结果不一致, 可能是因为本研究时间较短, 植物性状随着资源水平的改变, 首先是调整生理性状使其快速适应, 而形态性状相对滞后, 需要较长的时间才能发生显著变化(宋彦涛等, 2016; Yang et al., 2019). ...
Ecological and evolutionary determinants of a key plant functional trait: Wood density and its community-wide variation across latitude and elevation 1 2007
... 植物功能性状是指植物具有的与其定植、存活、生长和死亡紧密相关的一系列核心植物属性, 且这些属性能够反映植物个体对环境的响应和适应, 将环境变化同植物群落的结构和功能联系起来(Díaz et al., 2007; Swenson & Enquist, 2007; Violle et al., 2007; Lavorel et al., 2011; 刘晓娟和马克平, 2015).全球变化和植物功能性状之间的关系是当前植物生态学研究的热点问题. ...
Let the concept of trait be functional! 1 2007
... 植物功能性状是指植物具有的与其定植、存活、生长和死亡紧密相关的一系列核心植物属性, 且这些属性能够反映植物个体对环境的响应和适应, 将环境变化同植物群落的结构和功能联系起来(Díaz et al., 2007; Swenson & Enquist, 2007; Violle et al., 2007; Lavorel et al., 2011; 刘晓娟和马克平, 2015).全球变化和植物功能性状之间的关系是当前植物生态学研究的热点问题. ...
Response of plant functional traits of Leymus chinensis to extreme drought in Inner Mongolia grasslands 1 2019
... 功能性状作为解释变量, 对生态系统功能和服务的变化有显著的指示作用(Garnier et al., 2004; Mokany et al., 2010; Moor et al., 2015; Osborne et al., 2018), 能够显示植物对环境异质性的适应能力(Reich & Oleksyn, 2004; Faucon et al., 2017).植被生长过程中, 功能性状会受到降水量和施肥处理不同程度的影响.降水量增加使植株高度增高, 叶面积增大, 净光合速率升高(曾小平等, 2004; 许振柱和周广胜, 2005; 孟凡超等, 2014; Yue et al., 2019), 本研究结果与之一致, 主要原因是水分充足与否直接影响到羊草根系的发育, 根系发育的良好程度又直接影响到植物对土壤养分的吸收及养分在植物体内的循环, 进而对植物功能性状产生影响(李秀芳, 2011; Balachowski & Volaire, 2018).已有研究表明, 植物的光合能力与相对生长速率紧密相关(Shipley, 2006; Morgan et al., 2011), 叶面积反映了叶片对光的截获能力(Poorter & Rozendaal, 2008).因此, 降水量增加与氮磷共施条件下较高的光合速率和较大的叶面积可能是较高的叶生物量和茎生物量的主要原因.Fonseca等(2000)的研究表明澳大利亚东南部多年生植物的SLA随降水量的增加而增加; 驼绒藜(Ceratoides latens)的SLA与降水量呈正相关关系(朱军涛等, 2010); 植物叶质量也会随降水量的增加而减小(Wright et al., 2004; Rosbakh et al., 2015; Buckley et al., 2019).而本研究中, 降水量对羊草SLA没有显著影响(p > 0.05), 可能是由于羊草对短期水分处理响应不明显(Gao et al., 2015; Forrestel et al., 2017). ...
不同土壤水分条件下焕镛木幼苗的生理生态特性 1 2004
... 功能性状作为解释变量, 对生态系统功能和服务的变化有显著的指示作用(Garnier et al., 2004; Mokany et al., 2010; Moor et al., 2015; Osborne et al., 2018), 能够显示植物对环境异质性的适应能力(Reich & Oleksyn, 2004; Faucon et al., 2017).植被生长过程中, 功能性状会受到降水量和施肥处理不同程度的影响.降水量增加使植株高度增高, 叶面积增大, 净光合速率升高(曾小平等, 2004; 许振柱和周广胜, 2005; 孟凡超等, 2014; Yue et al., 2019), 本研究结果与之一致, 主要原因是水分充足与否直接影响到羊草根系的发育, 根系发育的良好程度又直接影响到植物对土壤养分的吸收及养分在植物体内的循环, 进而对植物功能性状产生影响(李秀芳, 2011; Balachowski & Volaire, 2018).已有研究表明, 植物的光合能力与相对生长速率紧密相关(Shipley, 2006; Morgan et al., 2011), 叶面积反映了叶片对光的截获能力(Poorter & Rozendaal, 2008).因此, 降水量增加与氮磷共施条件下较高的光合速率和较大的叶面积可能是较高的叶生物量和茎生物量的主要原因.Fonseca等(2000)的研究表明澳大利亚东南部多年生植物的SLA随降水量的增加而增加; 驼绒藜(Ceratoides latens)的SLA与降水量呈正相关关系(朱军涛等, 2010); 植物叶质量也会随降水量的增加而减小(Wright et al., 2004; Rosbakh et al., 2015; Buckley et al., 2019).而本研究中, 降水量对羊草SLA没有显著影响(p > 0.05), 可能是由于羊草对短期水分处理响应不明显(Gao et al., 2015; Forrestel et al., 2017). ...
不同土壤水分条件下焕镛木幼苗的生理生态特性 1 2004
... 功能性状作为解释变量, 对生态系统功能和服务的变化有显著的指示作用(Garnier et al., 2004; Mokany et al., 2010; Moor et al., 2015; Osborne et al., 2018), 能够显示植物对环境异质性的适应能力(Reich & Oleksyn, 2004; Faucon et al., 2017).植被生长过程中, 功能性状会受到降水量和施肥处理不同程度的影响.降水量增加使植株高度增高, 叶面积增大, 净光合速率升高(曾小平等, 2004; 许振柱和周广胜, 2005; 孟凡超等, 2014; Yue et al., 2019), 本研究结果与之一致, 主要原因是水分充足与否直接影响到羊草根系的发育, 根系发育的良好程度又直接影响到植物对土壤养分的吸收及养分在植物体内的循环, 进而对植物功能性状产生影响(李秀芳, 2011; Balachowski & Volaire, 2018).已有研究表明, 植物的光合能力与相对生长速率紧密相关(Shipley, 2006; Morgan et al., 2011), 叶面积反映了叶片对光的截获能力(Poorter & Rozendaal, 2008).因此, 降水量增加与氮磷共施条件下较高的光合速率和较大的叶面积可能是较高的叶生物量和茎生物量的主要原因.Fonseca等(2000)的研究表明澳大利亚东南部多年生植物的SLA随降水量的增加而增加; 驼绒藜(Ceratoides latens)的SLA与降水量呈正相关关系(朱军涛等, 2010); 植物叶质量也会随降水量的增加而减小(Wright et al., 2004; Rosbakh et al., 2015; Buckley et al., 2019).而本研究中, 降水量对羊草SLA没有显著影响(p > 0.05), 可能是由于羊草对短期水分处理响应不明显(Gao et al., 2015; Forrestel et al., 2017). ...
水肥耦合技术在辽宁地区农业可持续发展中的应用分析 1 2007
... 水分和养分是植物生产力水平提高的关键因素(李生秀等, 1994; 徐学选等, 1995; 汪德水, 1999; 李开峰等, 2010), 二者相互作用、互相影响, 从而对植物的生长发育以及产量产生水肥耦合效应(张广涛等, 2007).水分变化在植物的生长发育和生理生态过程中扮演着重要角色, 当降水量增加时, 植物能够通过调节其叶片干物质含量、比叶面积来适应环境(赵新风等, 2014; Kunstler et al., 2016); 随着降水量的增加, 植物的叶质量也会减小(Wright et al., 2004).一般情况下, 植物地上生物量随降水量的增加而增加(Hu et al., 2010; 张志南等, 2014).虽然近年来降水量与地上生物量之间的正相关关系已经逐渐得到广泛证实, 但其关系形态在不同研究中还存在一定差异(Yang et al., 2008; Zhang et al., 2019).土壤水分显著影响土壤养分有效性乃至陆地生态系统养分循环进程, 合理控水对于提高生产力具有重要意义. ...
水肥耦合技术在辽宁地区农业可持续发展中的应用分析 1 2007
... 水分和养分是植物生产力水平提高的关键因素(李生秀等, 1994; 徐学选等, 1995; 汪德水, 1999; 李开峰等, 2010), 二者相互作用、互相影响, 从而对植物的生长发育以及产量产生水肥耦合效应(张广涛等, 2007).水分变化在植物的生长发育和生理生态过程中扮演着重要角色, 当降水量增加时, 植物能够通过调节其叶片干物质含量、比叶面积来适应环境(赵新风等, 2014; Kunstler et al., 2016); 随着降水量的增加, 植物的叶质量也会减小(Wright et al., 2004).一般情况下, 植物地上生物量随降水量的增加而增加(Hu et al., 2010; 张志南等, 2014).虽然近年来降水量与地上生物量之间的正相关关系已经逐渐得到广泛证实, 但其关系形态在不同研究中还存在一定差异(Yang et al., 2008; Zhang et al., 2019).土壤水分显著影响土壤养分有效性乃至陆地生态系统养分循环进程, 合理控水对于提高生产力具有重要意义. ...
Divergent responses to water and nitrogen addition of three perennial bunchgrass species from variously degraded typical steppe in Inner Mongolia 1 2019
... 水分和养分是植物生产力水平提高的关键因素(李生秀等, 1994; 徐学选等, 1995; 汪德水, 1999; 李开峰等, 2010), 二者相互作用、互相影响, 从而对植物的生长发育以及产量产生水肥耦合效应(张广涛等, 2007).水分变化在植物的生长发育和生理生态过程中扮演着重要角色, 当降水量增加时, 植物能够通过调节其叶片干物质含量、比叶面积来适应环境(赵新风等, 2014; Kunstler et al., 2016); 随着降水量的增加, 植物的叶质量也会减小(Wright et al., 2004).一般情况下, 植物地上生物量随降水量的增加而增加(Hu et al., 2010; 张志南等, 2014).虽然近年来降水量与地上生物量之间的正相关关系已经逐渐得到广泛证实, 但其关系形态在不同研究中还存在一定差异(Yang et al., 2008; Zhang et al., 2019).土壤水分显著影响土壤养分有效性乃至陆地生态系统养分循环进程, 合理控水对于提高生产力具有重要意义. ...
黄土高原半干旱区天然草地群落结构与土壤水分关系 1 2014
... 水分和养分是植物生产力水平提高的关键因素(李生秀等, 1994; 徐学选等, 1995; 汪德水, 1999; 李开峰等, 2010), 二者相互作用、互相影响, 从而对植物的生长发育以及产量产生水肥耦合效应(张广涛等, 2007).水分变化在植物的生长发育和生理生态过程中扮演着重要角色, 当降水量增加时, 植物能够通过调节其叶片干物质含量、比叶面积来适应环境(赵新风等, 2014; Kunstler et al., 2016); 随着降水量的增加, 植物的叶质量也会减小(Wright et al., 2004).一般情况下, 植物地上生物量随降水量的增加而增加(Hu et al., 2010; 张志南等, 2014).虽然近年来降水量与地上生物量之间的正相关关系已经逐渐得到广泛证实, 但其关系形态在不同研究中还存在一定差异(Yang et al., 2008; Zhang et al., 2019).土壤水分显著影响土壤养分有效性乃至陆地生态系统养分循环进程, 合理控水对于提高生产力具有重要意义. ...
黄土高原半干旱区天然草地群落结构与土壤水分关系 1 2014
... 水分和养分是植物生产力水平提高的关键因素(李生秀等, 1994; 徐学选等, 1995; 汪德水, 1999; 李开峰等, 2010), 二者相互作用、互相影响, 从而对植物的生长发育以及产量产生水肥耦合效应(张广涛等, 2007).水分变化在植物的生长发育和生理生态过程中扮演着重要角色, 当降水量增加时, 植物能够通过调节其叶片干物质含量、比叶面积来适应环境(赵新风等, 2014; Kunstler et al., 2016); 随着降水量的增加, 植物的叶质量也会减小(Wright et al., 2004).一般情况下, 植物地上生物量随降水量的增加而增加(Hu et al., 2010; 张志南等, 2014).虽然近年来降水量与地上生物量之间的正相关关系已经逐渐得到广泛证实, 但其关系形态在不同研究中还存在一定差异(Yang et al., 2008; Zhang et al., 2019).土壤水分显著影响土壤养分有效性乃至陆地生态系统养分循环进程, 合理控水对于提高生产力具有重要意义. ...
养分和水分添加对荒漠草地植物群落结构和物种多样性的影响 1 2014
... 水分和养分是植物生产力水平提高的关键因素(李生秀等, 1994; 徐学选等, 1995; 汪德水, 1999; 李开峰等, 2010), 二者相互作用、互相影响, 从而对植物的生长发育以及产量产生水肥耦合效应(张广涛等, 2007).水分变化在植物的生长发育和生理生态过程中扮演着重要角色, 当降水量增加时, 植物能够通过调节其叶片干物质含量、比叶面积来适应环境(赵新风等, 2014; Kunstler et al., 2016); 随着降水量的增加, 植物的叶质量也会减小(Wright et al., 2004).一般情况下, 植物地上生物量随降水量的增加而增加(Hu et al., 2010; 张志南等, 2014).虽然近年来降水量与地上生物量之间的正相关关系已经逐渐得到广泛证实, 但其关系形态在不同研究中还存在一定差异(Yang et al., 2008; Zhang et al., 2019).土壤水分显著影响土壤养分有效性乃至陆地生态系统养分循环进程, 合理控水对于提高生产力具有重要意义. ...
养分和水分添加对荒漠草地植物群落结构和物种多样性的影响 1 2014
... 水分和养分是植物生产力水平提高的关键因素(李生秀等, 1994; 徐学选等, 1995; 汪德水, 1999; 李开峰等, 2010), 二者相互作用、互相影响, 从而对植物的生长发育以及产量产生水肥耦合效应(张广涛等, 2007).水分变化在植物的生长发育和生理生态过程中扮演着重要角色, 当降水量增加时, 植物能够通过调节其叶片干物质含量、比叶面积来适应环境(赵新风等, 2014; Kunstler et al., 2016); 随着降水量的增加, 植物的叶质量也会减小(Wright et al., 2004).一般情况下, 植物地上生物量随降水量的增加而增加(Hu et al., 2010; 张志南等, 2014).虽然近年来降水量与地上生物量之间的正相关关系已经逐渐得到广泛证实, 但其关系形态在不同研究中还存在一定差异(Yang et al., 2008; Zhang et al., 2019).土壤水分显著影响土壤养分有效性乃至陆地生态系统养分循环进程, 合理控水对于提高生产力具有重要意义. ...
 ,1,*, 李阳1,2, 魏继平1, 王志春1
,1,*, 李阳1,2, 魏继平1, 王志春1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT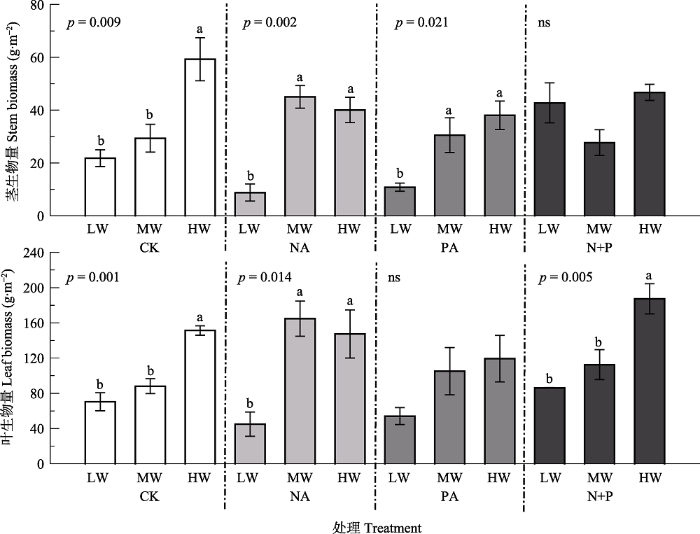 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT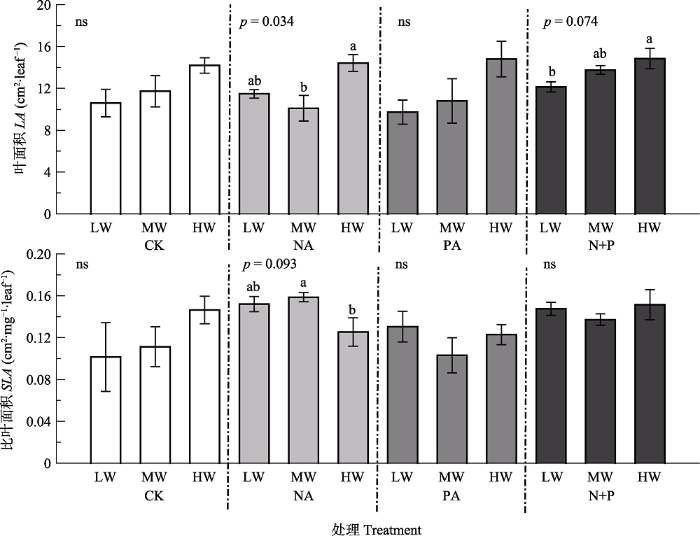 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT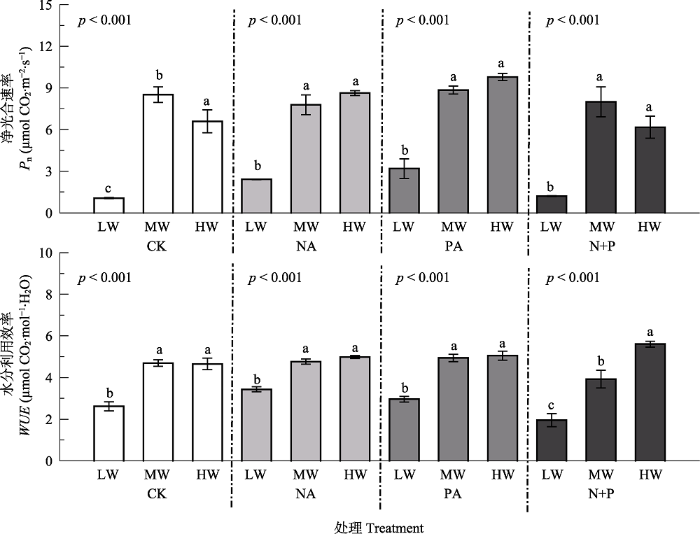 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT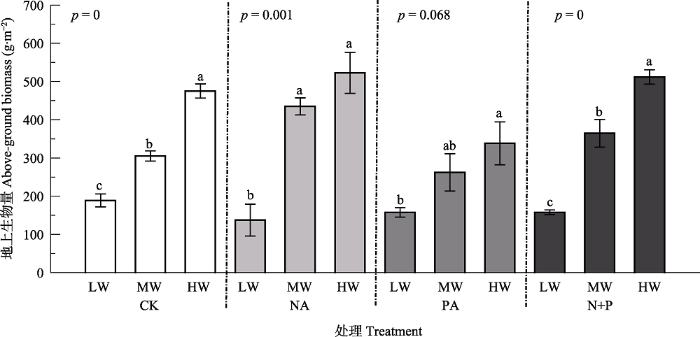 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}