方文静
1 , 蔡琼
1 , 朱江玲
1 , 吉成均
1 , 岳明
2 , 郭卫华
3 , 张峰
4 , 高贤明
5 , 唐志尧
1 , 方精云
, 1 , * 1 北京大学城市与环境学院, 北京大学生态研究中心, 地表过程分析与模拟教育部重点实验室, 北京 100871 2 西北大学西部资源生物与现代生物技术教育部重点实验室, 西安 710069 3 山东大学生命科学学院生态学与生物多样性研究所, 山东青岛 266237 4 山西大学黄土高原研究所, 太原 030006 5 中国科学院植物研究所植被与环境变化国家重点实验室, 北京 100093 Distribution, community structures and species diversity of larch forests in North China Wen-Jing FANG
1 , Qiong CAI
1 , Jiang-Ling ZHU
1 , Cheng-Jun JI
1 , Ming YUE
2 , Wei-Hua GUO
3 , Feng ZHANG
4 , Xian-Ming GAO
5 , Zhi-Yao TANG
1 , Jing-Yun FANG
, 1 , * 1 Institute of Ecology, College of Urban and Environmental Sciences, Laboratory for Earth Surface Processes of the Ministry of Education, Peking University, Beijing 100871, China 2 Key Laboratory of Resource Biology and Biotechnology in Western China, Ministry of Education, Northwest University, Xi’an 710069, China 3 Institute of Ecology and Biodiversity, School of Life Sciences, Shandong University, Qingdao, Shandong 266237, China 4 Institute of Loess Plateau, Shanxi University, Taiyuan 030006, China 5 State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China 通讯作者: * jyfang@urban.pku.edu.cn 编委: 江源
责任编辑: 李敏, 赵航(实习)
收稿日期: 2018-10-03
接受日期: 2018-12-24
网络出版日期: 2019-09-20
基金资助: 国家科技基础性工作专项 (2011FY110300 )国家科技基础性工作专项 (2015FY210200 )
Corresponding authors: * jyfang@urban.pku.edu.cn Received: 2018-10-03
Accepted: 2018-12-24
Online: 2019-09-20
Fund supported: Supported by the National Basic Work of Science and Technology of China (2011FY110300 )Supported by the National Basic Work of Science and Technology of China (2015FY210200 )
摘要 华北落叶松(
Larix principis-rupprechtii )林、日本落叶松(
L. kaempferi )林及太白红杉(
L. chinensis )林是华北地区常见的3种落叶松林类型, 其中日本落叶松林为人工林, 华北落叶松林既有天然分布又有人工种植, 太白红杉林则主要是天然林。该研究基于野外调查数据, 对这3种落叶松林的分布、物种组成、群落结构、物种多样性及其与环境间的关系进行了分析。研究发现, 3种落叶松林的分布受年平均气温的影响较大, 随着年平均气温的增加, 落叶松林的天然分布减少而人工种植的分布增加。3种森林中落叶松的林分径级及树高均为右偏分布, 说明3种落叶松林均处于相对稳定的演替阶段。3种落叶松林均拥有较高的物种丰富度且差异显著, 其中太白红杉林的物种丰富度最大(39.3 ± 17.9), 而华北落叶松林的物种丰富度最小(人工林27.2 ± 17.7, 天然林27.5 ± 13.8)。除最大树高与经度的关系不显著以外, 落叶松林的最大胸径和最大树高及物种丰富度均随经纬度的增加而显著降低, 随着年降水量的增加而显著增加。此外, 年平均气温对落叶松林的总物种丰富度影响不大, 但是对其群落结构影响显著。随着年平均气温的升高, 落叶松林的最大胸径显著降低而最大树高却显著增加。落叶松天然林和落叶松人工林物种多样性的地理分布格局及与气候因子间的关系与落叶松林总体的基本一致, 但群落结构的格局不尽相同: 随着经纬度的增加, 落叶松人工林的最大树高增加而天然林的最大树高减小; 落叶松天然林的最大胸径和最大树高分别随年平均气温的升高和年降水量的增加而减小, 而落叶松人工林的最大胸径和最大树高分别随年平均气温的升高和年降水量的增加而增大。
关键词: 落叶松林 ;
华北落叶松 ;
日本落叶松 ;
太白红杉 ;
群落结构 ;
物种组成 ;
物种多样性 Abstract Aims Larch forests are important for timber harvesting and water-soil conservation in North China. To explore the distribution, community structure and species diversity of larch forests is important for the vegetation conservation and sustainable utilization in North China.
Methods We collected species composition and local environment for 215 forest plots dominated by three common larch species, namely,
Larix principis-rupprechtii, L. kaempferi and
L. chinensis, in North China during 2000-2017. Among these types,
L. kaempferi forests are planted, while
L. chinensis forests are almost natural, and most of
L. principis-rupprechtii forests are natural. Based these data, we used the canonical correspondence analysis (CCA) to explore the relationship between species composition and environment. We also explored the pattern of community structure and species diversity of these three forests in relation to environmental factors.
Important findings Mean annual air temperature (
MAT ) was the most important factor for the distribution of these larch forests. The proportion of natural forest decreased, while that of planted forest increased, with
MAT . Diameter at breast height (
DBH ) and height distribution of three larch forests were right-skewed, indicating that all of these larch forests are at relatively stable successional stage. Species richness differ remarkably among different larch forests, which was highest in the
L. chinensis forests (39.3 ± 17.9), followed by the
L. kaempferi forests (37.4 ± 22.4), and lowest in the
L. principis-rupprechtii forests (planted forests 27.2 ± 17.7, natural forests 27.5 ± 13.8). Species richness, the maximum
DBH and the maximum height decreased with latitudes and longitudes. Species richness, the maximum
DBH and the maximum height increased with annual precipitation. However, species richness showed no significant trend, and the maximum height increased, while the maximum
DBH decreased, with
MAT . The patterns of species richness along geographical and climatic gradients were consistent between the planted, the natural and the overall (including both planted and natural) larch forests. However, the patterns of community structure differed remarkably among planted, natural and overall larch forests. The maximum height of planted forests increased, while that of natural forests decreased, with latitude and longitude. The maximum
DBH and height of natural forests decreased, while those of planted forests increased, with
MAT and annual precipitation, respectively.
Keywords: larch forest ;
Larix principis-rupprechtii ;
Larix kaempferi ;
Larix chinensis ;
community structure ;
species composition ;
species diversity PDF (8073KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 方文静, 蔡琼, 朱江玲, 吉成均, 岳明, 郭卫华, 张峰, 高贤明, 唐志尧, 方精云. 华北地区落叶松林的分布、群落结构和物种多样性. 植物生态学报, 2019, 43(9): 742-752. DOI:
10.17521/cjpe.2018.0244 FANG Wen-Jing, CAI Qiong, ZHU Jiang-Ling, JI Cheng-Jun, YUE Ming, GUO Wei-Hua, ZHANG Feng, GAO Xian-Ming, TANG Zhi-Yao, FANG Jing-Yun.
Distribution, community structures and species diversity of larch forests in North China .
Chinese Journal of Plant Ecology , 2019, 43(9): 742-752. DOI:
10.17521/cjpe.2018.0244 落叶松属(
Larix )植物是中国北方广泛分布的寒温性针叶林的建群种, 也是重要的用材树种(
吴征镒, 1980 ;
李楠, 1995 ;
刘增力等, 2002 )。在我国华北地区, 落叶松属植物是很多山地的林线物种, 如太白山的林线物种为太白红杉(
Larix chinensis )(
段仁燕等, 2010 ), 五台山、关帝山及小五台山的林线物种为华北落叶松(
L. principis-rupprechtii )(
Liu et al ., 2002 ), 这些落叶松对维护高海拔地区的生态环境有重要意义。目前中国华北地区主要分布有3种落叶松: 华北落叶松、日本落叶松(
L. kaempferi )及太白红杉(
郑万均, 1983 )。其中, 华北落叶松及太白红杉是中国特有种, 日本落叶松为华北地区广泛栽培种 (
郑万均和傅立国, 1978 )。这3种落叶松是华北地区落叶松林的优势种。其中, 日本落叶松林为人工林; 而华北落叶松林虽然在华北地区有天然分布, 但是由于其分布的海拔较低, 受人为干扰严重, 因而目前纯原始林较少, 大部分是受干扰的次生林或者栽培的人工林(
刘濂, 1996 ); 相较而言, 太白红杉林虽仅分布于我国秦岭山地, 但其分布海拔较高, 受人为干扰影响较小, 且其适应性极强, 在土层贫瘠气候寒冷的条件下也能适应, 为我国秦岭山区森林上限植被, 目前主要是原始林(
雷明德, 1999 ;
应俊生和陈梦玲, 2011 )。
落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要。前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面。而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面。而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道。
1 材料和方法 1.1 研究区域概况 研究区域涵盖了华北落叶松和太白红杉的天然分布区域, 包括了我国北方的大部分区域, 从东向西分别是山东、河北、北京、内蒙古、河南、山西、陕西、宁夏(
表1 ;
图1 )。三种落叶松林的森林样方如下: 日本落叶松林36个样方均为人工林; 太白红杉林29个样方均为天然林; 华北落叶松林150个样方中66个样方为人工林, 84个样方为天然林(
表1 )。
Table 1 表1 表1 华北地区3种落叶松林的调查区域
Table 1
Investigation area of three larch forests in North China 落叶松林类型 Larch forest type 起源类型 Origination 样方数量 No. of plots 主要调查地点 Main sites 华北落叶松林Larix principis-rupprechtii forest 人工林 Planted forest 66 山东: 泰山等 天然林 Natural forest 84 河北: 雾灵山、小五台山等 内蒙古: 大青山 山西: 太行山、中条山等 宁夏: 六盘山 日本落叶松林 L. kaempferi forest 人工林 Planted forest 36 山东: 崂山、昆嵛山等 河南: 玉皇山、龙峪湾林场 太白红杉林 L. chinensis forest 天然林 Natural forest 29 陕西: 太白山、朱雀森林公园等 总计 Total 215
新窗口打开 |
下载CSV 图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1华北地区3种落叶松林样方分布图。 Fig. 1Sampling sites of the three larch forests in North China. 1.2 调查方法 样方调查时间为2000-2017年, 面积20 m × 30 m。外业工作时, 记录样方内所有物种的物种名, 分乔木层、灌木层、草本层分别进行调查。对于乔木层, 对样方内胸径(
DBH ) ≥ 3 cm的所有乔木进行每木检尺, 测其
DBH 、树高; 对于灌木层, 将样方划成10 m × 10 m的小样方, 并从中选取2个进行调查, 记录平均高度、盖度、株数等; 对于草本层, 在每个10 m ×10 m小样方内选取1个1 m × 1 m的小样方进行调查, 记录多度、盖度、高度等(
方精云等, 2009 )。调查同时记录样地所在的经纬度、海拔及坡度和坡向等信息。
1.3 数据统计分析 计算乔木层物种的重要值。
(1) $相对频度=\frac{样方内该种的频度}{样方内全部种的频度和}\times100\%$ (2) 相对优势度 =样方内该种个体的胸高断面积/样方内全部种的胸高断面积总和$\times100\%$ (3) 重要值=(相对频度+相对优势度)/2 基于乔木层物种的胸高断面积, 对215个样地进行典型相关分析(CCA)排序。文中气候数据来源于WorldClim数据库(www.worldclim.org), 在ArcGIS 10.3支持下, 利用样方的经纬度信息获取所在样点的气候数据。本文使用的气候因子包括年平均气温(
MAT )和年降水量(
MAP )。地形因子包括坡度和坡向。选用物种丰富度(即物种数量)表征物种多样性。用多重比较的方法对比分析3种落叶松的林分径级结构及树高结构特征。本研究的数据统计均在R软件(3.4.3)中完成。
2 结果 2.1 华北地区落叶松林的地理分布及其与环境因子的关系 将华北地区3种落叶松林的物种矩阵结合地理因子及环境因子进行CCA排序, 其结果如
图2 所示。在地理因子与落叶松林物种矩阵的排序中, CCA第一轴反映了海拔和经纬度格局对落叶松林分布的交互影响, 落叶松天然林的分布与海拔正相关而与经纬度负相关, 大部分落叶松人工林的分布与海拔负相关而与经纬度正相关; 而CCA第二轴体现的主要是落叶松林分布的经纬度格局, 随着经纬度的增加, 落叶松林的分布由太白红杉林向日本落叶松林和华北落叶松林演变(
图2 A;
表2 )。在环境因子与物种矩阵的排序中, CCA第一轴主要体现了群落沿
MAT 的变化格局, 随着
MAT 的升高, 落叶松林的天然分布减少而人工种植分布增加, 落叶松林的类型也由太白红杉林向华北落叶松林和日本落叶松林演变; CCA第二轴主要体现了坡度分布规律, 华北落叶松林的分布与坡度大小存在正相关关系, 而日本落叶松林及太白红杉林的分布与坡度表现为负相关。综合而言, 华北地区落叶松人工林和天然林有显著的经纬度分布格局;
MAT 是华北地区落叶松人工林和天然林以及3种森林类型分布的最重要影响因素(
图2 ;
表2 )。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2华北地区落叶松林沿地理(A)及环境因子(B)的典范对应分析排序图。 Fig. 2Canonical correspondence analysis ordinations oflarch forests along geographical (A) and environmental variables (B) in North China. Environmental factors include Aspect, Slope, mean annual air temperature (MAT ) and mean annual precipitation (MAP ). LCF, L. chinensis forest; LKF, L. kaempferi forest; LPF, Larix principis-rupprechtii forest; NF, natural forest; PF, planted forest. Table 2 表2 表2 华北地区落叶松林物种矩阵与环境因子及地理因子典范对应分析排序得分
Table 2
Scores of the canonical correspondence analysis ordinations of larch forests in North China 地理因素 Geographical factor CCA 1 CCA 2 R 2 p 经度 Longitude 0.89 -0.46 0.44 <0.01 纬度 Latitude 0.84 -0.55 0.42 <0.01 海拔 Altitude -0.96 0.27 0.13 <0.01 气候及地形因素 CCA 1 CCA 2 R 2 p 年降水量 MAP 0.19 -0.98 0.08 <0.01 年平均气温 MAT 0.99 -0.11 0.53 <0.01 坡度 Slope -0.65 0.76 0.15 <0.01 坡向 Aspect -0.75 0.66 0.03 0.1
MAP, mean annual precipitation; MAT, mean annual air temperature.新窗口打开 |
下载CSV 2.2 华北地区落叶松林的常见物种组成 华北落叶松天然林乔木层物种主要为华北落叶松(重要值为74.6%)、油松(
Pinus tabuliformis, 6.0%)、红桦(
Betula albosinensis, 6.0%)及白桦(
B. platyphylla, 3.0%); 华北落叶松人工林乔木层物种主要为华北落叶松(88.7%)、油松(2.8%)、辽东桤木(
Alnus sibirica, 1.8%)及蒙古栎(
Quercus mongolica, 1.5%); 日本落叶松林乔木层物种主要为日本落叶松(70.2%)、水榆花楸(
Sorbus alnifolia, 5.3%)、白檀(
Symplocos paniculata, 4.2%)及三桠乌药(
Lindera obtusiloba, 2.8%); 太白红杉林乔木层物种主要为太白红杉(81.2%)、巴山冷杉(
Abies fargesii, 8.1%)、金背杜鹃(
Rhododendron clementinae subsp.
aureodorsale, 5.4%)及糙皮桦(
B. utilis, 2.2%)(
表3 )。
Table 3 表3 表3 华北地区3种落叶松林乔木层主要物种的重要值
Table 3
Important value of the main tree species in the canopy of three larch forests in North China 华北落叶松(人工林) 重要值 华北落叶松(天然林) 重要值 日本落叶松林(人工林) 重要值 太白红杉林(天然林) 重要值 华北落叶松Larix principis- Pinus tabuliformis Alnus sibirica Quercus mongolica Betula platyphylla B. albosinensis Pinus armandii 88.7 华北落叶松 L. principis-rupprechtii P. tabuliformis B. albosinensis B. platyphylla Picea meyeri P. wilsonii B. dahurica Populus davidiana P. koraiensis Juglans mandshurica Q. wutaishanica 74.6 日本落叶松 L. kaempferi Sorbus alnifolia Symplocos paniculata Lindera obtusiloba Q. acutissima Toxicodendron vernicifluum Chamaecyparis pisifera P. densiflora Juglans mandshurica Nyssa sinensis Q. wutaishanica Tilia tuan Fraxinus mandschurica P. armandii Robinia pseudoacacia 70.2 太白红杉 L. chinensis Abies fargesii Rhododendron clementinae subsp. aureodorsale B. utili Sorbus koehneana B. platyphylla 81.2
Plant names refer to Flora Republicae Popularis Sinicae (The Editorial Committee of Flora of China, 2004 ). IV, important value; LCF, L. chinensis forest; LKF, L. kaempferi forest; LPF, L. principis-rupprechtii forest; NF, natural forest; PF, planted forest. 表中植物名称参照《中国植物志》中文版(中国植物志编辑委员会, 2004 )。 新窗口打开 |
下载CSV 2.3 华北地区落叶松林的物种多样性 华北落叶松天然林乔、灌、草层的平均物种丰富度分别为3.1、5.8和11.4; 华北落叶松人工林乔、灌、草层的平均物种丰富度分别为2.5、4.7和20.4; 日本落叶松林分别为5.1、13.8和20.4; 太白红杉林分别为2.2、7.7和29.6; 所有天然林分别为2.9、6.2和21.8; 所有人工林分别为3.5、7.9和20.4 (
表4 )。3种落叶松林乔木层物种丰富度最大值分别是: 11、11和6; 灌木层: 29、38和22; 草本层: 55、56和73 (
表4 )。所有天然林和所有人工林的物种丰富度没有显著差异(
p > 0.05), 但是3种不同类型落叶松林乔、灌、草层的物种丰富度差异显著(
p < 0.05)(
图3 ;
表4 )。具体而言, 日本落叶松林乔木层物种丰富度显著大于华北落叶松林和太白红杉林(
p < 0.05), 华北落叶松林与太白红杉林的乔木层物种丰富度没有差异(
p > 0.05); 日本落叶松林灌木层物种丰富度显著大于华北落叶松林(
p < 0.05), 但与太白红杉林没有显著差异(
p > 0.05), 华北落叶松天然林灌木层物种丰富度与太白红杉林没有显著差异(
p > 0.05), 但是华北落叶松人工林的灌木层物种丰富度显著小于太白红杉林(
p < 0.05); 太白红杉林的草本层物种丰富度显著大于日本落叶松林及华北落叶松林(
p < 0.05), 日本落叶松林与华北落叶松林的草本层物种丰富度没有显著差异(
p > 0.05)(
图3 )。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3华北地区三种落叶松林乔木层(A)、灌木层(B)和草本层(C)的物种丰富度。不同小写字母代表多重比较的结果。其中LPF、LKF和LCF分别表示华北落叶松林、日本落叶松林和太白红杉林, NF和PF分别表示表示天然林和人工林。 Fig. 3Species richness of the tree (A), shrub (B) and herb (C) layers in the Larix principis-rupprechtii , L. kaempferi and L. chinensis forests in North China. Different lowercase letters mean the result of multiple comparisons. LCF, L. chinensis forest; LKF, L. kaempferi forest; LPF, L. principis-rupprechtii forest; NF, natural forest; PF, planted forest. Table 4 表4 表4 华北地区落叶松林的物种丰富度
Table 4
Species richness of larch forests in North China 落叶松林类型 乔木层 灌木层 草本层 总体 华北落叶松(人工林) LPF (PF) 2.5 ± 2.1b (1-9) 4.7 ± 4.1c (0-18) 20.4 ± 15.8b (0-55) 27.2 ± 17.7b (0-63) 华北落叶松(天然林) LPF (NF) 3.1 ± 2.1b (1-11) 5.8 ± 5.0bc (0-29) 11.4 ± 19.1b (0-50) 27.5 ± 13.8b (1-65) 日本落叶松林(人工林) LKF (PF) 5.1 ± 2.9a (1-11) 13.8 ± 9.1a (4-38) 20.4 ± 15.1b (4-56) 37.4 ± 22.4ab (14-81) 太白红杉林(天然林) LCF (NF) 2.2 ± 1.5b (1-6) 7.7 ± 3.6ab (0-22) 29.6 ± 15.8a (12-73) 39.3 ± 17.9a (17-98) 所有天然林 All natural forest 2.9 ± 2.0a (1-11) 6.2 ± 4.7a (0-29) 21.8 ± 13.4a (0-73) 30.5 ± 15.8a (1-98) 所有人工林 All planted forest 3.5 ± 2.7a (1-11) 7.9 ± 7.6a (0-38) 20.4 ± 15.5a (0-56) 30.8 ± 20a (1-81)
Values represent mean ± SD and (minimum - maximum) value. Different lowercase letters mean the result of multiple comparisons. LCF, L. chinensis forest; LKF, L. kaempferi forest; LPF, L. principis-rupprechtii forest; NF, natural forest; PF, planted forest. 数据表达为平均值±标准偏差及(最小值-最大值)。不同小写字母代表多重比较的结果。 新窗口打开 |
下载CSV 2.4 华北地区落叶松林的群落结构 三种落叶松林中优势种的胸径和树高结构如
表5 所示。日本落叶松的平均胸径最大, 太白红杉次之, 华北落叶松的平均胸径最小。日本落叶松的平均树高最高, 华北落叶松次之, 太白红杉最低。三种落叶松胸径、树高的偏度系数均大于0; 峰度系数均大于3。从森林起源来看, 落叶松天然林优势种的胸径大于人工林, 树高小于人工林, 无论天然林还是人工林均偏度系数大于0, 峰度系数大于3 (
表5 )。
Table 5 表5 表5 华北地区3种落叶松林中优势种的胸径、树高结构
Table 5
Diameter at breast height (DBH ) and height structure of dominant species in three larch forests in North China 胸径/树高DBH /Height 优势种 数量 均值±标准偏差SD 最小值-最大值 偏度系数 峰度系数 胸径 DBH (cm) 华北落叶松(人工林) LPF (PF) 4 251 15.1 ± 6.4d 3.0-36.2 0.2 2.2 华北落叶松(天然林) LPF (NF) 4 464 18.0 ± 7.5c 3.0-75.8 0.7 4.4 日本落叶松(人工林) LKF (PF) 1 463 20.7 ± 8.4a 3.2-56.0 0.8 3.7 太白红杉(天然林) LCF (NF) 1 278 19.5 ± 9.8b 3.0-80.0 0.8 4.5 所有天然林 All natural forest 5 742 18.3 ± 8.1a 3.0-80.0 0.8 4.8 所有人工林All planted forest 5 714 16.5 ± 7.4b 3.0-56.0 0.7 3.8 树高 Height (m) 华北落叶松(人工林) LPF (PF) 4 251 11.3 ± 4.2c 2.2-35.5 0.6 3.6 华北落叶松(天然林) LPF (NF) 4 464 12.3 ± 4.3b 2.0-30.0 0.3 3.2 日本落叶松(人工林) LKF (PF) 1 463 17.6 ± 8.2a 2.3-39.5 0.6 2.8 太白红杉(天然林) LCF (NF) 1 278 9.1 ± 4.8d 2.0-30.0 2.0 8.2 所有天然林 All natural forest 5 742 11.6 ± 4.6b 2.0-30.0 0.6 3.6 所有人工林 All planted forest 5 714 12.9 ± 6.1a 2.2-39.5 1.4 5.6
Different lowercase letters mean the result of multiple comparisons. LCF, L. chinensis forest; LKF, L. kaempferi forest; LPF, L. principis-rupprechtii forest; NF, natural forest; PF, planted forest. 不同小写字母代表多重比较的结果。 新窗口打开 |
下载CSV 2.5 华北地区落叶松林的群落结构和多样性与地理及气候因子的关系 地理和气候因子对华北地区落叶松林的群落结构和物种多样性影响显著(
p < 0.05)。就总体而言, 除最大树高与经度的关系不显著(
p > 0.05)以外, 落叶松林的最大胸径和最大树高及总体的物种丰富度随着经纬度的增加而显著降低(
p < 0.05)(
图4 ), 随着
MAP 的增加而显著增加(
p < 0.05)(
图4 D、4H、4L)。
图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4华北地区落叶松林的群落结构和物种丰富度与经纬度及年平均气温和年降水量之间的关系。 Fig. 4Changes in community structure and species richness with latitude, longitude, mean annual temperature (MAT ) and precipitation (MAP ) for the larch forests in North China. The structure measurements of forests include the maximum diameter at breast height (DBH max ) and height (Heightmax ). LCF, L. chinensis forest; LKF, L. kaempferi forest; LPF, L. principis-rupprechtii forest; NF, natural forest; PF, planted forest. 此外,
MAT 对华北地区落叶松林的总物种丰富度影响不大(
图4K ), 但是对其群落结构影响显著(
p < 0.05)。随着
MAT 的升高, 落叶松林的最大胸径显著降低而最大树高却显著增加(
p < 0.05)(
图4C 、4G)。从森林起源来看, 落叶松天然林的最大树高随着经纬度及
MAT 的增加而显著增加(
p < 0.05), 随
MAP 的增加而显著降低(
p < 0.05); 与此相反的, 其最大胸径却随着经纬度及
MAT 的增加而显著降低(
p < 0.05), 随
MAP 的增加而显著增加(
p < 0.05); 其物种丰富度与经度及
MAT 没有显著相关性(
p > 0.05), 随着纬度的增加而显著降低(
p < 0.05), 随着
MAP 的增加而显著增加(
p < 0.05)(
图4 )。落叶松人工林的群落结构和物种多样性与地理及气候因子间的关系与落叶松天然林及总体的规律基本一致。但是, 随经度的增加, 天然林的最大树高显著增加而人工林的最大树高显著降低(
p < 0.05)(
图4 I); 随
MAT 的增加, 落叶松天然林的最大胸径显著降低而人工林的最大胸径显著增加(
p < 0.05)(
图4C ); 随着
MAP 的增加, 落叶松天然林的最大树高显著降低而人工林的最大树高显著增加(
p < 0.05)(
图4 H)。
3 讨论 植物群落结构可以反映出群落的整体状态。3种落叶松林胸径和树高频度分布的偏度系数大于0且峰度系数大于3 (
表5 ), 即3种落叶松林的胸径和树高结构总体均呈右偏分布且其径级大小和高度大小相对集中, 表明3种落叶松林均处于相对稳定的演替阶段(
Tyrrell & Crow, 1994 ;
Aiba & Kitayama, 1999 ;
Fang et al ., 2012 )。
匡旭等(2014) 对中国东北长白山落叶松林的研究结果显示, 长白落叶松(
L. olgensis )有较多
DBH < 10 cm的个体, 群落处于生长状态。
赵文智(1997) 对河北坝上华北落叶松人工林的研究表明, 30年华北落叶松成熟林林分稳定, 有天然更新的潜力。我们的结果与以上研究结果一致。
物种丰富度是测度物种多样性的一个重要维度(
Gaston, 2000 )。华北地区, 3种落叶松林的物种丰富度总体比较丰富, 乔、灌、草层的物种丰富度有显著的差异(
图3 ;
表4 )。这与3种落叶松林的物种组成、气候条件以及乔木层郁闭度等因素有关(
Wang et al ., 2009 ;
Mölder et al ., 2014 )。具体而言, 太白红杉林为秦岭山地原始林(
雷明德, 1999 ), 华北落叶松林也多为人工林或次生林(
刘濂, 1996 ), 两种落叶松林优势种太白红杉及华北落叶松的重要值很高(
表3 ), 大部分甚至是纯林, 因而它们的乔木层的物种丰富度很低(
表4 ), 没有显著差异(
图3 A)。而日本落叶松林虽然是人工林, 但其分布海拔低于华北落叶松林和太白红杉林, 拥有更好的水热条件, 群落物种更为丰富; 通常与其他物种混交, 其乔木层优势种日本落叶松的重要值只有70.2% (
表3 ), 因而其乔木层的物种丰富度显著大于华北落叶松林和太白红杉林(
图3 A)。此外, 日本落叶松林与华北落叶松林均分布于中低海拔区域, 乔木层郁闭度大, 林下植物尤其是草本植物的生长受到影响, 其草本层丰富度相对较低, 没有显著差异(
图3 C)。而太白红杉林位于秦岭山地高海拔区域, 乔木层郁闭度相对较小, 有利于林下植物的生长, 其林下草本层的物种丰富度显著大于华北落叶松林和日本落叶松林(
图3 C)。但是, 也正是由于分布海拔高的影响, 其林下的生境条件较差, 所以对比华北地区纬度位置相差不大的日本落叶松林, 其灌木层物种丰富度没有显著优势(
图3 B)。
华北地区落叶松林的群落结构特征及物种多样性与其分布的地理位置和气候条件紧密相关。总体而言, 除最大树高与经度的关系不显著以外, 华北地区落叶松林最大胸径和最大树高及总体的物种丰富度均随着经纬度的增加而显著降低(
图4A 、4D、 4E、4F、4I、4J), 这与前人在全球地理格局的研究结果一致(
Gaston, 2000 ;
Moles et al ., 2009 ;
Tao et al ., 2016 )。此外, 诸多的研究表明, 水分条件是乔木胸径和高度及植物群落物种丰富度的重要影响因子(
Gaston, 2000 ;
Zhang et al ., 2013 ;
Tao et al ., 2016 )。在华北地区, 落叶松林最大胸径与最大树高及其总体的物种丰富度随着
MAP 的增加而增加(
图4D 、4H、4L), 与前人的研究结果相符(
Gaston, 2000 ;
Moles et al ., 2009 ;
Zhang et al ., 2013 )。而与经纬度和
MAP 对华北地区落叶松林最大胸径和最大树高的影响的一致性不同,
MAT 对华北地区落叶松林最大胸径与最大树高的影响正好相反(
图4C 、4G)。我们的结果显示: 华北地区落叶松林最大胸径随
MAT 的增加而 降低, 随
MAP 的增加而增加(
图4C 、4G), 与
Yan等 (2006) 对鼎湖山亚热带常绿阔叶林优势乔木胸径的研究及
Tao等(2016) 对全球树高的研究结果一致。落叶松属植物为阳性物种, 作为林线植被, 气候变暖对落叶松的生长及群落更新可能是有利的(
Devi et al ., 2008 )。在全球气候变暖的背景下, 华北地区的落叶松林更趋向于垂直生长。
从森林起源来看, 落叶松天然林和落叶松人工林的群落结构特征及物种多样性与地理及气候因子之间的关系, 与落叶松林总体的基本一致。其在地理因子上的差异主要体现在最大树高上, 落叶松人工林的最大树高随经纬度的增加而减少, 与前人在全球地理格局的研究结果一致(
Gaston, 2000 ;
Moles et al ., 2009 ;
Tao et al ., 2016 )。但是落叶松天然林的最大树高随经纬度的增加而增加, 与前人的研究结果(
Gaston, 2000 ;
Moles et al ., 2009 ;
Tao et al ., 2016 ) 相反, 其原因可能与海拔高度有关。华北地区低纬度和经度位置偏西的落叶松天然林主要是太白红杉林(
图1 ), 其分布海拔高, 树高相对低矮(
表5 ), 而高纬度位置及经度位置偏东的落叶松天然林为华北落叶松林, 其分布海拔较太白红杉林低(
郑万均和傅立国, 1978 ), 树高更高(
表5 ), 即华北地区落叶松林与地理因素的关系主要由落叶松人工林所主导。落叶松人工林和落叶松天然林在气候因子上的差异体现在最大胸径与
MAT 以及最大树高与
MAP 的关系上(
图4C 、4H)。落叶松天然林的最大胸径和最大树高分别随
MAT 和
MAP 的增大而减小, 而落叶松人工林的最大胸径和最大树高分别随
MAT 和
MAP 的增大而增大。落叶松林总体最大胸径与气候条件的关系与落叶松天然林一致, 最大树高与气候条件的关系与落叶松人工林一致, 而对比落叶松天然林和落叶松人工林优势种的胸径树高, 落叶松天然林优势种的胸径显著大于人工林, 而树高显著小于人工林(
表5 ), 即华北地区落叶松林与
MAT 的关系主要受落叶松天然林的影响, 而与
MAP 的关系主要受落叶松人工林的影响。
前人基于大尺度的研究表明, 气候因子尤其是环境能量因子对物种丰富度的影响显著(
Wang et al ., 2012 )。但我们的结果似乎并不支持这一结论。本研究中,
MAT 对华北地区落叶松林的物种丰富度没有显著影响(
图4K )。这可能与研究对象的差异有关。
Wang等(2012) 的研究是基于木本植物物种丰富度的结果, 而本研究中的研究结果是基于落叶松林木本植物及草本植物总的物种丰富度的结果。由
表4 可知, 华北地区落叶松林总体的物种丰富度中草本植物所占的比重最大, 因而其对环境条件的响应与木本植物的不同。
除地理及环境因子的影响外, 人为干扰也是影响森林群落结构和物种丰富度的重要影响因子。对落叶松天然林而言, 人为干扰可能主要体现在旅游方面。近年来, 由于生态旅游的兴起, 大量的游客涌入到自然保护区的核心区, 砍椽(搭设旅游接待帐篷用)、烧柴(取暖和生活用柴)以及生活垃圾污染等对山地环境产生重要影响, 诸如本文研究区域的秦岭太白山保护区就是近年来生态旅游的一大热点地区(
刘香妮, 2001 )。对落叶松人工林而言, 其初期的抚育间伐对落叶松幼龄林及中龄林的群落结构和物种丰富度影响甚大(
李国雷等, 2009 ;
李淑辉等, 2012 ;
郭琳等, 2014 )。而我们调查的落叶松林大部分为成熟林, 甚至部分已经向人工天然混交林状态发展, 群落结构及物种丰富度已经相对稳定。对比落叶松天然林和落叶松人工林的胸径树高结构发现, 落叶松天然林的胸径大于人工林, 而其树高则小于人工林(
表5 )。由于落叶松人工林主要营造于中低海拔地带, 而现存的落叶松天然林主要在高海拔区域。所以这除了可能与森林经营管理有关外, 落叶松天然林和人工林的分布海拔对其胸径树高结构的影响可能更大。对比落叶松天然林与人工林的物种丰富度并无显著差异(
表4 )。其原因可能是由于华北地区的落叶松人工林属于三北防护林工程的水土涵养林, 对于成过熟林乃至有天然化趋势的人工林, 人为的经营管理活动较少, 故而其群落结构和物种丰富度受到的影响可能较小(
表4 ,
表5 ), 而放牧等活动对该地区中低海拔落叶松人工林的影响或许 更大。
[1] Aiba S Kitayama K 1999 ). Structure, composition and species diversity in an altitude-substrate matrix of rain forest tree communities on Mount Kinabalu, BorneoPlant Ecology ,139 -157 . [本文引用: 1] [2] Chen DS Sun XM Zhang SG 2016 ). Biomass, carbon storage and nutrient characteristics in Larix kaempferi plantations at different stand agesChinese Journal of Applied Ecology, 3759 -3768 . [本文引用: 1] [ 陈东升 , 孙晓梅 , 张守攻 ( 2016 ). 不同年龄日本落叶松人工林生物量、碳储量及养分特征应用生态学报 , 27, 3759 -3768 .] [本文引用: 1] [3] Cheng WC 1983 ). Chinese Dendrology . Vol. 1. China Forestry Publishing House, Beijing. [本文引用: 1] [ 郑万均 ( 1983 ). 中国树木志(第1卷) . 中国林业出版社, 北京.] [本文引用: 1] [4] Cheng WC Fu LK 1978 ). GymnospermaeIn: Editorial Committee of Flora of China, Chinese Academy of Sciences. Flora of China. Science Press, Beijing. 169 -196 . [本文引用: 2] [ 郑万均 , 傅立国 ( 1978 ). 裸子植物门出自中国科学院中国植物志编辑委员会编著. 中国植物志 . 科学出版社, 北京. 169 -196 .] [本文引用: 2] [5] Devi N Hagedorn F Moiseev P Bugmann H Shiyatov S Mazepa V Rigling A 2008 ). Expanding forests and changing growth forms of Siberian larch at the Polar Urals treeline during the 20th centuryGlobal Change Biology ,1581 -1591 . [本文引用: 1] [6] Duan RY Wang XA Huang MY Wang ZG Wu GL 2010 ). Ecological characteristics of Larix chinensis population near timberline on Taibai Mountain in ChinaActa Ecologica Sinica, 519 -526 . [本文引用: 1] [ 段仁燕 , 王孝安 , 黄敏毅 , 王志高 , 吴甘霖 ( 2010 ). 太白山林线附近太白红杉种群的生态特征生态学报 , 30, 519 -526 .] [本文引用: 1] [7] Editorial Committee of Flora of China ( 2004 ). Flora of China . Science Press, Beijing. [本文引用: 2] [ 中国植物志编辑委员会 ( 2004 ). 中国植物志 . 科学出版社, 北京.] [本文引用: 2] [8] Fang JY Wang XP Liu YN Tang ZY White PS Sanders NJ 2012 ). Multi-scale patterns of forest structure and species composition in relation to climate in northeast ChinaEcography ,1072 -1082 . [本文引用: 1] [9] Fang JY Wang XP Shen ZH Tang ZY He JS Yu D Jiang Y Wang ZH Zheng CY Zhu JL Guo ZD 2009 ). Methods and protocols for plant community inventoryBiodiversity Science ,533 -548 . [本文引用: 1] [ 方精云 , 王襄平 , 沈泽昊 , 唐志尧 , 贺金生 , 于丹 , 江源 , 王志恒 , 郑成洋 , 朱江玲 , 郭兆迪 ( 2009 ). 植物群落清查的主要内容、方法和技术规范生物多样性 , 17, 533 -548 .] [本文引用: 1] [10] Gaston KJ 2000 ). Global patterns in biodiversityNature ,220 -227 . [本文引用: 6] [11] Guo H 2004 ). Studies on Spatial Pattern of Larix chinensis Populations in Qinling Mountain Master degree dissertation, Shaanxi Normal University, Xi’ an. 20 -31 . [本文引用: 1] [ 郭华 ( 2004 ). 秦岭太白红杉种群空间分布格局研究硕士学位论文, 陕西师范大学, 西安 . 20 -31 .] [本文引用: 1] [12] Guo L Zhu Q Zhang BG Yu J Lin L Zhou GX 2014 ). Research on tending thinning of Japanese larch forest in the Sichuan BasinJournal of Sichuan Forestry Science and Technology ,6 ), 64 -68 . [本文引用: 1] [ 郭琳 , 朱琴 , 张保刚 , 余军 , 林林 , 周桂香 ( 2014 ). 四川盆地日本落叶松林抚育间伐研究四川林业科技 , 35(6 ), 64 -68 .] [本文引用: 1] [13] Han YY 2006 ). Study on Effect of the Climatic Factors on the Increment of Larix leptolepis in Funiu Mountain in Western Henan Master degree dissertation, Henan Agricultural University, Zhengzhou . 20 -40 . [本文引用: 1] [ 韩艳英 ( 2006 ). 气象因子对豫西伏牛山区日本落叶松生长量影响研究硕士学位论文, 河南农业大学, 郑州 . 20 -40 .] [本文引用: 1] [14] Ji WJ Cheng XQ Han HR Kang FF Yang J Zhu J Zhao J Bai YC Ma JY 2016 ). The biomass and nutrient distribution in Larix principis-ruppechtii Magyr plantations at different forest age Chinese Journal of Applied and Environmental Biology, 277 -284 . [本文引用: 1] [ 纪文婧 , 程小琴 , 韩海荣 , 康峰峰 , 杨杰 , 朱江 , 赵敬 , 白英辰 , 马俊勇 ( 2016 ). 不同林龄华北落叶松人工林生物量及营养元素分布特征应用与环境生物学报 , 22, 277 -284 .] [本文引用: 1] [15] Kong LW Chen XW Lu SW Li SN Chen B Gao C Shi Y Yang XY 2014 ). Relationships among growth Larix principis-?rupprechtii , herbaceous plants diversity and landformBulletin of Soil and Water Conservation, 60 -66 . [本文引用: 1] [ 孔令伟 , 陈祥伟 , 鲁绍伟 , 李少宁 , 陈波 , 高琛 , 石媛 , 杨小燕 ( 2014 ). 华北落叶松林木生长、草本植物多样性及地形因子之间的关系水土保持通报 , 34, 60 -66 .] [本文引用: 1] [16] Kuang X Xing DL Zhang ZC Song HJ Wang YY Fang S Yuan ZQ Ye J Lin F Wang XG Hao ZQ 2014 ). Species composition and community structure of a spruce-fir forest and a larch forest on the northern slope of Changbai Mountains, Northeast ChinaChinese Journal of Applied Ecology ,2149 -2157 . [本文引用: 1] [ 匡旭 , 邢丁亮 , 张昭臣 , 宋厚娟 , 王芸芸 , 房帅 , 原作强 , 叶吉 , 蔺菲 , 王绪高 , 郝占庆 ( 2014 ). 长白山北坡云冷杉林和落叶松林物种组成与群落结构应用生态学报 , 25, 2149 -2157 .] [本文引用: 1] [17] Lei MD 1999 ). Vegetation of Shaanxi Province. [本文引用: 2] [ 雷明德 ( 1999 ). 陕西植被 . 科学出版社, 北京.] [本文引用: 2] [18] Li GL Liu Y Lü RH Yu HQ Li RS 2009 ). Responses of understory vegetation development to regulation of tree density in Larix principis-rupprechtii plantationsJournal of Beijing Forestry University, 1 ), 19 -24 . [本文引用: 1] [ 李国雷 , 刘勇 , 吕瑞恒 , 于海群 , 李瑞生 ( 2009 ). 华北落叶松人工林密度调控对林下植被发育的作用过程北京林业大学学报 , 31(1 ), 19 -24 .] [本文引用: 1] [19] Li N 1995 ). Studies on the geographic distribution, origin and dispersal of the Family Pinaceae LindlActa Phytotaxonomica Sinica ,105 -130 . [本文引用: 1] [ 李楠 ( 1995 ). 论松科植物的地理分布、起源和扩散植物分类学报 , 33, 105 -130 .] [本文引用: 1] [20] Li SH Liang F Guo JP 2012 ). Thinning effects on under-?crown vegetation of Larix principis-rupprechtii plantation in Guandishan MountainJournal of Shanxi Agricultural University (Natural Science Edition )256 -260 . [本文引用: 1] [ 李淑辉 , 梁芳 , 郭晋平 ( 2012 ). 关帝山林区华北落叶松人工林抚育间伐对林下植被的影响山西农业大学学报(自然科学版) , 32, 256 -260 .] [本文引用: 1] [21] Liu HY Tang ZY Dai JH Tang YX Cui HT 2002 ). Larch timberline and its development in North ChinaMountain Research and Development ,359 -367 . [本文引用: 1] [22] Liu L 1996 ). Vegetation of Hebei Province . [本文引用: 2] [ 刘濂 ( 1996 ). 河北植被 . 科学出版社, 北京.] [本文引用: 2] [23] Liu XN 2001 ). The investigation on the impact of human activity on Larix chinensis Shaanxi Forest Science and Technology ,3 ), 23 -24 . [本文引用: 1] [ 刘香妮 ( 2001 ). 人为活动对太白红杉林的影响调查陕西林业科技 , ( 3 ), 23 -24 .] [本文引用: 1] [24] Liu ZL Fang JY Piao SL 2002 ). Geographical distribution of species genera Abies, Picea and Larix in ChinaActa Geographica Sinica, 577 -586 . [本文引用: 1] [ 刘增力 , 方精云 , 朴世龙 ( 2002 ). 中国冷杉、云杉和落叶松属植物的地理分布地理学报 , 57, 577 -586 .] [本文引用: 1] [25] Luo YJ Zhang XQ Wang XK Zhu JH Zhang ZJ Sun GS Gao F 2009 ). Biomass and its distribution patterns of Larix principis-rupprechtii plantations in northern ChinaJournal of Beijing Forestry University, 1 ), 13 -18 . [本文引用: 1] [ 罗云建 , 张小全 , 王效科 , 朱建华 , 张治军 , 孙贵生 , 高峰 ( 2009 ). 华北落叶松人工林生物量及其分配模式北京林业大学学报 , 31(1 ), 13 -18 .] [本文引用: 1] [26] Ma FF Zhang CM Li YZ 2016 ). Density and distribution of organic carbon of Larix kaempferi plantation ecosystem in subtropicsJournal of Central South University of Forestry & Technology, 1 ), 94 -100 . [本文引用: 1] [ 马丰丰 , 张灿明 , 李有志 ( 2016 ). 亚热带日本落叶松人工林生态系统碳密度及其分配特征中南林业科技大学学报 , 36(1 ), 94 -100 .] [本文引用: 1] [27] Ma YP 2007 ). Study on Stand Structure of Larix kaempferi Forests and Its Increment Prediction—Taking Changlinggang Forest Farm as an Example PhD dissertation, Beijing Forestry University , Beijing. 71 -117 . [本文引用: 1] [ 马友平 ( 2007 ). 日本落叶松人工林林分结构与生长量预测研究——以长岭岗林场为例博士学位论文, 北京林业大学, 北京 . 71 -117 .] [本文引用: 1] [28] M?lder A Streit M Schmidt W 2014 ). When beech strikes back: How strict nature conservation reduces herb-layer diversity and productivity in Central European deciduous forestsForest Ecology and Management ,51 -61 . [本文引用: 1] [29] Moles AT Warton DI Warman L Swenson NG Laffan SW Zanne AE Pitman A Hemmings FA Leishman MR 2009 ). Global patterns in plant heightJournal of Ecology ,923 -932 . [本文引用: 4] [30] Su YM 1995 ). Study on biomass and productivity of Larix kaempferi plantationJournal of Sichuan Forestry Science and Technology, 3 ), 36 -42 . [本文引用: 1] [ 宿以明 ( 1995 ). 日本落叶松人工林生物量和生产力的研究四川林业科技 , 16(3 ), 36 -42 .] [本文引用: 1] [31] Tao SL Guo QH Li C Wang ZH Fang JY 2016 ). Global patterns and determinants of forest canopy heightEcology ,3265 -3270 . [本文引用: 5] [32] Tu YB 2008 ). Study on the Effect of Larix chinensis Forest Gap Characteristics on the Community Regeneration. Master degree dissertation, Shaanxi Normal University, Xi’an . 14 -35 . [本文引用: 1] [ 涂云博 ( 2008 ). 太白红杉林林窗特征及其对群落更新的影响研究硕士学位论文, 陕西师范大学, 西安 . 14 -35 .] [本文引用: 1] [33] Tyrrell LE Crow TR 1994 ). Structural characteristics of old-growth hemlock-hardwood forests in relation to ageEcology ,370 -386 . [本文引用: 1] [34] Wang XP Fang JY Sanders NJ White PS Tang ZY 2009 ). Relative importance of climate vs local factors in shaping the regional patterns of forest plant richness across northeast ChinaEcography ,133 -142 . [本文引用: 1] [35] Wang ZH Rahbek C Fang JY 2012 ). Effects of geographical extent on the determinants of woody plant diversityEcography ,1160 -1167 . [本文引用: 2] [36] Wu XJ Chang JG Yu JX An Y Guo JR 2010 ). Spatial distribution of Larix principis-rupprechtii-Picea spp. secondary forests along elevation gradients on north facing slope of Luya Mountain, ShanxiJournal of Northeast Forestry University, 11 ), 10 -14 . [本文引用: 1] [ 武秀娟 , 常建国 , 于吉祥 , 安雁 , 郭建荣 ( 2010 ). 芦芽山阴坡华北落叶松-云杉天然次生林群落特征的海拔梯度格局东北林业大学学报 , 38(11 ), 10 -14 .] [本文引用: 1] [37] Wu ZY 1980 ). Vegetation of China. [本文引用: 1] [ 吴征镒 ( 1980 ). 中国植被 . 科学出版社, 北京.] [本文引用: 1] [38] Xu LJ Peng H Chen CG 2005 a). Size-class and age-class structures of Larix chinensis forestsActa Botanica Boreali-?Occidentalia Sinica, 460 -465 . [本文引用: 1] [ 许林军 , 彭鸿 , 陈存根 ( 2005 a). 太白红杉林径级和龄级结构的研究西北植物学报 , 25, 460 -465 .] [本文引用: 1] [39] Xu LJ Peng H Chen CG Tang HL Yang YJ 2005 b). Quantitative analysis of the Larix chinensis forest’s distribution at Qinling Mountains and the character of the alpine timberline at Taibai MountainActa Botanica Boreali-? Occidentalia Sinica, 968 -972 . [本文引用: 1] [ 许林军 , 彭鸿 , 陈存根 , 唐红亮 , 杨亚娟 ( 2005 b). 秦岭太白红杉林分布及太白山高山林线特征的定量分析西北植物学报 , 25, 968 -972 .] [本文引用: 1] [40] Yan GQ Zhao GF Hu ZH 2001 ). The study on Larix chinensis community characteristics and species diversity in Mts QinlingActa Botanica Boreali-Occidentalia Sinica ,497 -506 . [本文引用: 1] [ 阎桂琴 , 赵桂仿 , 胡正海 ( 2001 ). 秦岭太白红杉群落特征及其物种多样性的研究西北植物学报 , 21, 497 -506 .] [本文引用: 1] [41] Yan HB Han YZ Yang XQ Wang LY Xiang XY 2010 ). Spatial distribution patterns and associations of tree species in typical natural secondary mountain forest communities of Northern ChinaActa Ecologica Sinica ,2311 -2321 . [本文引用: 1] [ 闫海冰 , 韩有志 , 杨秀清 , 王丽艳 , 项小英 ( 2010 ). 华北山地典型天然次生林群落的树种空间分布格局及其关联性生态学报 , 30, 2311 -2321 .] [本文引用: 1] [42] Yan JH Zhou GY Zhang DQ Tang XL Wang X 2006 ). Different patterns of changes in the dry season diameter at breast height of dominant and evergreen tree species in a mature subtropical forest in South ChinaJournal of Integrative Plant Biology ,906 -913 . [本文引用: 1] [43] Ying JS Chen ML 2011 ). Chinese Plant Geography [本文引用: 1] [ 应俊生 , 陈梦玲 ( 2011 ). 中国植物地理 . 上海科学技术出版社, 上海.] [本文引用: 1] [44] Zhang JL Zhou Y Zhou GS Xiao CW 2013 ). Structure and composition of natural gmelin larch ( Larix gmelinii var. gmelinii ) forests in response to spatial climatic changesPLOS ONE, e66668 . DOI: 10.1371/journal.?pone.?0066668 . [本文引用: 2] [45] Zhang QD Zhang JT Zhang B Cheng JJ Tian SG Suriguga ( 2010 ). Pattern of Larix principis-rupprechtii plantation and its environmental interpretation in Dongling MountainJournal of Wuhan Botanical Research, 577 -582 . [本文引用: 1] [ 张钦弟 , 张金屯 , 张斌 , 程佳佳 , 田世广 , 苏日古嘎 ( 2010 ). 东灵山华北落叶松人工林的分布格局及环境解释武汉植物学研究 , 28, 577 -582 .] [本文引用: 1] [46] Zhao TS Guang ZY Zhao YM Liu GW 1999 ). Study on biomass and productivity of Larix kaempferi plantationActa Agriculturae Universitatis Henanensis, 350 -353 . [本文引用: 1] [ 赵体顺 , 光增云 , 赵义民 , 刘国伟 ( 1999 ). 日本落叶松人工林生物量及生产力的研究河南农业大学学报 , 33, 350 -353 .] [本文引用: 1] [47] Zhao WZ 1997 ). Study on the artificial Larix principis- rupprechtii forests in Bashang, Hebei ProvinceJournal of Desert Research, 243 -249 . [本文引用: 1] [ 赵文智 ( 1997 ). 河北坝上半干旱/半湿润过渡带华北落叶松人工林研究中国沙漠 , 17, 243 -249 .] [本文引用: 1] Structure, composition and species diversity in an altitude-substrate matrix of rain forest tree communities on Mount Kinabalu, Borneo
1
1999
... 植物群落结构可以反映出群落的整体状态.3种落叶松林胸径和树高频度分布的偏度系数大于0且峰度系数大于3 (
表5 ), 即3种落叶松林的胸径和树高结构总体均呈右偏分布且其径级大小和高度大小相对集中, 表明3种落叶松林均处于相对稳定的演替阶段(
Tyrrell & Crow, 1994 ;
Aiba & Kitayama, 1999 ;
Fang et al ., 2012 ).
匡旭等(2014) 对中国东北长白山落叶松林的研究结果显示, 长白落叶松(
L. olgensis )有较多
DBH < 10 cm的个体, 群落处于生长状态.
赵文智(1997) 对河北坝上华北落叶松人工林的研究表明, 30年华北落叶松成熟林林分稳定, 有天然更新的潜力.我们的结果与以上研究结果一致. ...
不同年龄日本落叶松人工林生物量、碳储量及养分特征
1
2016
... 落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要.前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面.而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面.而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道. ...
不同年龄日本落叶松人工林生物量、碳储量及养分特征
1
2016
... 落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要.前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面.而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面.而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道. ...
1
1983
... 落叶松属(
Larix )植物是中国北方广泛分布的寒温性针叶林的建群种, 也是重要的用材树种(
吴征镒, 1980 ;
李楠, 1995 ;
刘增力等, 2002 ).在我国华北地区, 落叶松属植物是很多山地的林线物种, 如太白山的林线物种为太白红杉(
Larix chinensis )(
段仁燕等, 2010 ), 五台山、关帝山及小五台山的林线物种为华北落叶松(
L. principis-rupprechtii )(
Liu et al ., 2002 ), 这些落叶松对维护高海拔地区的生态环境有重要意义.目前中国华北地区主要分布有3种落叶松: 华北落叶松、日本落叶松(
L. kaempferi )及太白红杉(
郑万均, 1983 ).其中, 华北落叶松及太白红杉是中国特有种, 日本落叶松为华北地区广泛栽培种 (
郑万均和傅立国, 1978 ).这3种落叶松是华北地区落叶松林的优势种.其中, 日本落叶松林为人工林; 而华北落叶松林虽然在华北地区有天然分布, 但是由于其分布的海拔较低, 受人为干扰严重, 因而目前纯原始林较少, 大部分是受干扰的次生林或者栽培的人工林(
刘濂, 1996 ); 相较而言, 太白红杉林虽仅分布于我国秦岭山地, 但其分布海拔较高, 受人为干扰影响较小, 且其适应性极强, 在土层贫瘠气候寒冷的条件下也能适应, 为我国秦岭山区森林上限植被, 目前主要是原始林(
雷明德, 1999 ;
应俊生和陈梦玲, 2011 ). ...
1
1983
... 落叶松属(
Larix )植物是中国北方广泛分布的寒温性针叶林的建群种, 也是重要的用材树种(
吴征镒, 1980 ;
李楠, 1995 ;
刘增力等, 2002 ).在我国华北地区, 落叶松属植物是很多山地的林线物种, 如太白山的林线物种为太白红杉(
Larix chinensis )(
段仁燕等, 2010 ), 五台山、关帝山及小五台山的林线物种为华北落叶松(
L. principis-rupprechtii )(
Liu et al ., 2002 ), 这些落叶松对维护高海拔地区的生态环境有重要意义.目前中国华北地区主要分布有3种落叶松: 华北落叶松、日本落叶松(
L. kaempferi )及太白红杉(
郑万均, 1983 ).其中, 华北落叶松及太白红杉是中国特有种, 日本落叶松为华北地区广泛栽培种 (
郑万均和傅立国, 1978 ).这3种落叶松是华北地区落叶松林的优势种.其中, 日本落叶松林为人工林; 而华北落叶松林虽然在华北地区有天然分布, 但是由于其分布的海拔较低, 受人为干扰严重, 因而目前纯原始林较少, 大部分是受干扰的次生林或者栽培的人工林(
刘濂, 1996 ); 相较而言, 太白红杉林虽仅分布于我国秦岭山地, 但其分布海拔较高, 受人为干扰影响较小, 且其适应性极强, 在土层贫瘠气候寒冷的条件下也能适应, 为我国秦岭山区森林上限植被, 目前主要是原始林(
雷明德, 1999 ;
应俊生和陈梦玲, 2011 ). ...
裸子植物门
2
1978
... 落叶松属(
Larix )植物是中国北方广泛分布的寒温性针叶林的建群种, 也是重要的用材树种(
吴征镒, 1980 ;
李楠, 1995 ;
刘增力等, 2002 ).在我国华北地区, 落叶松属植物是很多山地的林线物种, 如太白山的林线物种为太白红杉(
Larix chinensis )(
段仁燕等, 2010 ), 五台山、关帝山及小五台山的林线物种为华北落叶松(
L. principis-rupprechtii )(
Liu et al ., 2002 ), 这些落叶松对维护高海拔地区的生态环境有重要意义.目前中国华北地区主要分布有3种落叶松: 华北落叶松、日本落叶松(
L. kaempferi )及太白红杉(
郑万均, 1983 ).其中, 华北落叶松及太白红杉是中国特有种, 日本落叶松为华北地区广泛栽培种 (
郑万均和傅立国, 1978 ).这3种落叶松是华北地区落叶松林的优势种.其中, 日本落叶松林为人工林; 而华北落叶松林虽然在华北地区有天然分布, 但是由于其分布的海拔较低, 受人为干扰严重, 因而目前纯原始林较少, 大部分是受干扰的次生林或者栽培的人工林(
刘濂, 1996 ); 相较而言, 太白红杉林虽仅分布于我国秦岭山地, 但其分布海拔较高, 受人为干扰影响较小, 且其适应性极强, 在土层贫瘠气候寒冷的条件下也能适应, 为我国秦岭山区森林上限植被, 目前主要是原始林(
雷明德, 1999 ;
应俊生和陈梦玲, 2011 ). ...
... 从森林起源来看, 落叶松天然林和落叶松人工林的群落结构特征及物种多样性与地理及气候因子之间的关系, 与落叶松林总体的基本一致.其在地理因子上的差异主要体现在最大树高上, 落叶松人工林的最大树高随经纬度的增加而减少, 与前人在全球地理格局的研究结果一致(
Gaston, 2000 ;
Moles et al ., 2009 ;
Tao et al ., 2016 ).但是落叶松天然林的最大树高随经纬度的增加而增加, 与前人的研究结果(
Gaston, 2000 ;
Moles et al ., 2009 ;
Tao et al ., 2016 ) 相反, 其原因可能与海拔高度有关.华北地区低纬度和经度位置偏西的落叶松天然林主要是太白红杉林(
图1 ), 其分布海拔高, 树高相对低矮(
表5 ), 而高纬度位置及经度位置偏东的落叶松天然林为华北落叶松林, 其分布海拔较太白红杉林低(
郑万均和傅立国, 1978 ), 树高更高(
表5 ), 即华北地区落叶松林与地理因素的关系主要由落叶松人工林所主导.落叶松人工林和落叶松天然林在气候因子上的差异体现在最大胸径与
MAT 以及最大树高与
MAP 的关系上(
图4C 、4H).落叶松天然林的最大胸径和最大树高分别随
MAT 和
MAP 的增大而减小, 而落叶松人工林的最大胸径和最大树高分别随
MAT 和
MAP 的增大而增大.落叶松林总体最大胸径与气候条件的关系与落叶松天然林一致, 最大树高与气候条件的关系与落叶松人工林一致, 而对比落叶松天然林和落叶松人工林优势种的胸径树高, 落叶松天然林优势种的胸径显著大于人工林, 而树高显著小于人工林(
表5 ), 即华北地区落叶松林与
MAT 的关系主要受落叶松天然林的影响, 而与
MAP 的关系主要受落叶松人工林的影响. ...
裸子植物门
2
1978
... 落叶松属(
Larix )植物是中国北方广泛分布的寒温性针叶林的建群种, 也是重要的用材树种(
吴征镒, 1980 ;
李楠, 1995 ;
刘增力等, 2002 ).在我国华北地区, 落叶松属植物是很多山地的林线物种, 如太白山的林线物种为太白红杉(
Larix chinensis )(
段仁燕等, 2010 ), 五台山、关帝山及小五台山的林线物种为华北落叶松(
L. principis-rupprechtii )(
Liu et al ., 2002 ), 这些落叶松对维护高海拔地区的生态环境有重要意义.目前中国华北地区主要分布有3种落叶松: 华北落叶松、日本落叶松(
L. kaempferi )及太白红杉(
郑万均, 1983 ).其中, 华北落叶松及太白红杉是中国特有种, 日本落叶松为华北地区广泛栽培种 (
郑万均和傅立国, 1978 ).这3种落叶松是华北地区落叶松林的优势种.其中, 日本落叶松林为人工林; 而华北落叶松林虽然在华北地区有天然分布, 但是由于其分布的海拔较低, 受人为干扰严重, 因而目前纯原始林较少, 大部分是受干扰的次生林或者栽培的人工林(
刘濂, 1996 ); 相较而言, 太白红杉林虽仅分布于我国秦岭山地, 但其分布海拔较高, 受人为干扰影响较小, 且其适应性极强, 在土层贫瘠气候寒冷的条件下也能适应, 为我国秦岭山区森林上限植被, 目前主要是原始林(
雷明德, 1999 ;
应俊生和陈梦玲, 2011 ). ...
... 从森林起源来看, 落叶松天然林和落叶松人工林的群落结构特征及物种多样性与地理及气候因子之间的关系, 与落叶松林总体的基本一致.其在地理因子上的差异主要体现在最大树高上, 落叶松人工林的最大树高随经纬度的增加而减少, 与前人在全球地理格局的研究结果一致(
Gaston, 2000 ;
Moles et al ., 2009 ;
Tao et al ., 2016 ).但是落叶松天然林的最大树高随经纬度的增加而增加, 与前人的研究结果(
Gaston, 2000 ;
Moles et al ., 2009 ;
Tao et al ., 2016 ) 相反, 其原因可能与海拔高度有关.华北地区低纬度和经度位置偏西的落叶松天然林主要是太白红杉林(
图1 ), 其分布海拔高, 树高相对低矮(
表5 ), 而高纬度位置及经度位置偏东的落叶松天然林为华北落叶松林, 其分布海拔较太白红杉林低(
郑万均和傅立国, 1978 ), 树高更高(
表5 ), 即华北地区落叶松林与地理因素的关系主要由落叶松人工林所主导.落叶松人工林和落叶松天然林在气候因子上的差异体现在最大胸径与
MAT 以及最大树高与
MAP 的关系上(
图4C 、4H).落叶松天然林的最大胸径和最大树高分别随
MAT 和
MAP 的增大而减小, 而落叶松人工林的最大胸径和最大树高分别随
MAT 和
MAP 的增大而增大.落叶松林总体最大胸径与气候条件的关系与落叶松天然林一致, 最大树高与气候条件的关系与落叶松人工林一致, 而对比落叶松天然林和落叶松人工林优势种的胸径树高, 落叶松天然林优势种的胸径显著大于人工林, 而树高显著小于人工林(
表5 ), 即华北地区落叶松林与
MAT 的关系主要受落叶松天然林的影响, 而与
MAP 的关系主要受落叶松人工林的影响. ...
Expanding forests and changing growth forms of Siberian larch at the Polar Urals treeline during the 20th century
1
2008
... 华北地区落叶松林的群落结构特征及物种多样性与其分布的地理位置和气候条件紧密相关.总体而言, 除最大树高与经度的关系不显著以外, 华北地区落叶松林最大胸径和最大树高及总体的物种丰富度均随着经纬度的增加而显著降低(
图4A 、4D、 4E、4F、4I、4J), 这与前人在全球地理格局的研究结果一致(
Gaston, 2000 ;
Moles et al ., 2009 ;
Tao et al ., 2016 ).此外, 诸多的研究表明, 水分条件是乔木胸径和高度及植物群落物种丰富度的重要影响因子(
Gaston, 2000 ;
Zhang et al ., 2013 ;
Tao et al ., 2016 ).在华北地区, 落叶松林最大胸径与最大树高及其总体的物种丰富度随着
MAP 的增加而增加(
图4D 、4H、4L), 与前人的研究结果相符(
Gaston, 2000 ;
Moles et al ., 2009 ;
Zhang et al ., 2013 ).而与经纬度和
MAP 对华北地区落叶松林最大胸径和最大树高的影响的一致性不同,
MAT 对华北地区落叶松林最大胸径与最大树高的影响正好相反(
图4C 、4G).我们的结果显示: 华北地区落叶松林最大胸径随
MAT 的增加而 降低, 随
MAP 的增加而增加(
图4C 、4G), 与
Yan等 (2006) 对鼎湖山亚热带常绿阔叶林优势乔木胸径的研究及
Tao等(2016) 对全球树高的研究结果一致.落叶松属植物为阳性物种, 作为林线植被, 气候变暖对落叶松的生长及群落更新可能是有利的(
Devi et al ., 2008 ).在全球气候变暖的背景下, 华北地区的落叶松林更趋向于垂直生长. ...
太白山林线附近太白红杉种群的生态特征
1
2010
... 落叶松属(
Larix )植物是中国北方广泛分布的寒温性针叶林的建群种, 也是重要的用材树种(
吴征镒, 1980 ;
李楠, 1995 ;
刘增力等, 2002 ).在我国华北地区, 落叶松属植物是很多山地的林线物种, 如太白山的林线物种为太白红杉(
Larix chinensis )(
段仁燕等, 2010 ), 五台山、关帝山及小五台山的林线物种为华北落叶松(
L. principis-rupprechtii )(
Liu et al ., 2002 ), 这些落叶松对维护高海拔地区的生态环境有重要意义.目前中国华北地区主要分布有3种落叶松: 华北落叶松、日本落叶松(
L. kaempferi )及太白红杉(
郑万均, 1983 ).其中, 华北落叶松及太白红杉是中国特有种, 日本落叶松为华北地区广泛栽培种 (
郑万均和傅立国, 1978 ).这3种落叶松是华北地区落叶松林的优势种.其中, 日本落叶松林为人工林; 而华北落叶松林虽然在华北地区有天然分布, 但是由于其分布的海拔较低, 受人为干扰严重, 因而目前纯原始林较少, 大部分是受干扰的次生林或者栽培的人工林(
刘濂, 1996 ); 相较而言, 太白红杉林虽仅分布于我国秦岭山地, 但其分布海拔较高, 受人为干扰影响较小, 且其适应性极强, 在土层贫瘠气候寒冷的条件下也能适应, 为我国秦岭山区森林上限植被, 目前主要是原始林(
雷明德, 1999 ;
应俊生和陈梦玲, 2011 ). ...
太白山林线附近太白红杉种群的生态特征
1
2010
... 落叶松属(
Larix )植物是中国北方广泛分布的寒温性针叶林的建群种, 也是重要的用材树种(
吴征镒, 1980 ;
李楠, 1995 ;
刘增力等, 2002 ).在我国华北地区, 落叶松属植物是很多山地的林线物种, 如太白山的林线物种为太白红杉(
Larix chinensis )(
段仁燕等, 2010 ), 五台山、关帝山及小五台山的林线物种为华北落叶松(
L. principis-rupprechtii )(
Liu et al ., 2002 ), 这些落叶松对维护高海拔地区的生态环境有重要意义.目前中国华北地区主要分布有3种落叶松: 华北落叶松、日本落叶松(
L. kaempferi )及太白红杉(
郑万均, 1983 ).其中, 华北落叶松及太白红杉是中国特有种, 日本落叶松为华北地区广泛栽培种 (
郑万均和傅立国, 1978 ).这3种落叶松是华北地区落叶松林的优势种.其中, 日本落叶松林为人工林; 而华北落叶松林虽然在华北地区有天然分布, 但是由于其分布的海拔较低, 受人为干扰严重, 因而目前纯原始林较少, 大部分是受干扰的次生林或者栽培的人工林(
刘濂, 1996 ); 相较而言, 太白红杉林虽仅分布于我国秦岭山地, 但其分布海拔较高, 受人为干扰影响较小, 且其适应性极强, 在土层贫瘠气候寒冷的条件下也能适应, 为我国秦岭山区森林上限植被, 目前主要是原始林(
雷明德, 1999 ;
应俊生和陈梦玲, 2011 ). ...
2
2004
... Plant names refer to
Flora Republicae Popularis Sinicae (
The Editorial Committee of Flora of China, 2004 ). IV, important value; LCF,
L. chinensis forest; LKF,
L. kaempferi forest; LPF,
L. principis-rupprechtii forest; NF, natural forest; PF, planted forest. ...
... 表中植物名称参照《中国植物志》中文版(
中国植物志编辑委员会, 2004 ). ...
2
2004
... Plant names refer to
Flora Republicae Popularis Sinicae (
The Editorial Committee of Flora of China, 2004 ). IV, important value; LCF,
L. chinensis forest; LKF,
L. kaempferi forest; LPF,
L. principis-rupprechtii forest; NF, natural forest; PF, planted forest. ...
... 表中植物名称参照《中国植物志》中文版(
中国植物志编辑委员会, 2004 ). ...
Multi-scale patterns of forest structure and species composition in relation to climate in northeast China
1
2012
... 植物群落结构可以反映出群落的整体状态.3种落叶松林胸径和树高频度分布的偏度系数大于0且峰度系数大于3 (
表5 ), 即3种落叶松林的胸径和树高结构总体均呈右偏分布且其径级大小和高度大小相对集中, 表明3种落叶松林均处于相对稳定的演替阶段(
Tyrrell & Crow, 1994 ;
Aiba & Kitayama, 1999 ;
Fang et al ., 2012 ).
匡旭等(2014) 对中国东北长白山落叶松林的研究结果显示, 长白落叶松(
L. olgensis )有较多
DBH < 10 cm的个体, 群落处于生长状态.
赵文智(1997) 对河北坝上华北落叶松人工林的研究表明, 30年华北落叶松成熟林林分稳定, 有天然更新的潜力.我们的结果与以上研究结果一致. ...
植物群落清查的主要内容、方法和技术规范
1
2009
... 样方调查时间为2000-2017年, 面积20 m × 30 m.外业工作时, 记录样方内所有物种的物种名, 分乔木层、灌木层、草本层分别进行调查.对于乔木层, 对样方内胸径(
DBH ) ≥ 3 cm的所有乔木进行每木检尺, 测其
DBH 、树高; 对于灌木层, 将样方划成10 m × 10 m的小样方, 并从中选取2个进行调查, 记录平均高度、盖度、株数等; 对于草本层, 在每个10 m ×10 m小样方内选取1个1 m × 1 m的小样方进行调查, 记录多度、盖度、高度等(
方精云等, 2009 ).调查同时记录样地所在的经纬度、海拔及坡度和坡向等信息. ...
植物群落清查的主要内容、方法和技术规范
1
2009
... 样方调查时间为2000-2017年, 面积20 m × 30 m.外业工作时, 记录样方内所有物种的物种名, 分乔木层、灌木层、草本层分别进行调查.对于乔木层, 对样方内胸径(
DBH ) ≥ 3 cm的所有乔木进行每木检尺, 测其
DBH 、树高; 对于灌木层, 将样方划成10 m × 10 m的小样方, 并从中选取2个进行调查, 记录平均高度、盖度、株数等; 对于草本层, 在每个10 m ×10 m小样方内选取1个1 m × 1 m的小样方进行调查, 记录多度、盖度、高度等(
方精云等, 2009 ).调查同时记录样地所在的经纬度、海拔及坡度和坡向等信息. ...
Global patterns in biodiversity
6
2000
... 物种丰富度是测度物种多样性的一个重要维度(
Gaston, 2000 ).华北地区, 3种落叶松林的物种丰富度总体比较丰富, 乔、灌、草层的物种丰富度有显著的差异(
图3 ;
表4 ).这与3种落叶松林的物种组成、气候条件以及乔木层郁闭度等因素有关(
Wang et al ., 2009 ;
Mölder et al ., 2014 ).具体而言, 太白红杉林为秦岭山地原始林(
雷明德, 1999 ), 华北落叶松林也多为人工林或次生林(
刘濂, 1996 ), 两种落叶松林优势种太白红杉及华北落叶松的重要值很高(
表3 ), 大部分甚至是纯林, 因而它们的乔木层的物种丰富度很低(
表4 ), 没有显著差异(
图3 A).而日本落叶松林虽然是人工林, 但其分布海拔低于华北落叶松林和太白红杉林, 拥有更好的水热条件, 群落物种更为丰富; 通常与其他物种混交, 其乔木层优势种日本落叶松的重要值只有70.2% (
表3 ), 因而其乔木层的物种丰富度显著大于华北落叶松林和太白红杉林(
图3 A).此外, 日本落叶松林与华北落叶松林均分布于中低海拔区域, 乔木层郁闭度大, 林下植物尤其是草本植物的生长受到影响, 其草本层丰富度相对较低, 没有显著差异(
图3 C).而太白红杉林位于秦岭山地高海拔区域, 乔木层郁闭度相对较小, 有利于林下植物的生长, 其林下草本层的物种丰富度显著大于华北落叶松林和日本落叶松林(
图3 C).但是, 也正是由于分布海拔高的影响, 其林下的生境条件较差, 所以对比华北地区纬度位置相差不大的日本落叶松林, 其灌木层物种丰富度没有显著优势(
图3 B). ...
... 华北地区落叶松林的群落结构特征及物种多样性与其分布的地理位置和气候条件紧密相关.总体而言, 除最大树高与经度的关系不显著以外, 华北地区落叶松林最大胸径和最大树高及总体的物种丰富度均随着经纬度的增加而显著降低(
图4A 、4D、 4E、4F、4I、4J), 这与前人在全球地理格局的研究结果一致(
Gaston, 2000 ;
Moles et al ., 2009 ;
Tao et al ., 2016 ).此外, 诸多的研究表明, 水分条件是乔木胸径和高度及植物群落物种丰富度的重要影响因子(
Gaston, 2000 ;
Zhang et al ., 2013 ;
Tao et al ., 2016 ).在华北地区, 落叶松林最大胸径与最大树高及其总体的物种丰富度随着
MAP 的增加而增加(
图4D 、4H、4L), 与前人的研究结果相符(
Gaston, 2000 ;
Moles et al ., 2009 ;
Zhang et al ., 2013 ).而与经纬度和
MAP 对华北地区落叶松林最大胸径和最大树高的影响的一致性不同,
MAT 对华北地区落叶松林最大胸径与最大树高的影响正好相反(
图4C 、4G).我们的结果显示: 华北地区落叶松林最大胸径随
MAT 的增加而 降低, 随
MAP 的增加而增加(
图4C 、4G), 与
Yan等 (2006) 对鼎湖山亚热带常绿阔叶林优势乔木胸径的研究及
Tao等(2016) 对全球树高的研究结果一致.落叶松属植物为阳性物种, 作为林线植被, 气候变暖对落叶松的生长及群落更新可能是有利的(
Devi et al ., 2008 ).在全球气候变暖的背景下, 华北地区的落叶松林更趋向于垂直生长. ...
... ).此外, 诸多的研究表明, 水分条件是乔木胸径和高度及植物群落物种丰富度的重要影响因子(
Gaston, 2000 ;
Zhang et al ., 2013 ;
Tao et al ., 2016 ).在华北地区, 落叶松林最大胸径与最大树高及其总体的物种丰富度随着
MAP 的增加而增加(
图4D 、4H、4L), 与前人的研究结果相符(
Gaston, 2000 ;
Moles et al ., 2009 ;
Zhang et al ., 2013 ).而与经纬度和
MAP 对华北地区落叶松林最大胸径和最大树高的影响的一致性不同,
MAT 对华北地区落叶松林最大胸径与最大树高的影响正好相反(
图4C 、4G).我们的结果显示: 华北地区落叶松林最大胸径随
MAT 的增加而 降低, 随
MAP 的增加而增加(
图4C 、4G), 与
Yan等 (2006) 对鼎湖山亚热带常绿阔叶林优势乔木胸径的研究及
Tao等(2016) 对全球树高的研究结果一致.落叶松属植物为阳性物种, 作为林线植被, 气候变暖对落叶松的生长及群落更新可能是有利的(
Devi et al ., 2008 ).在全球气候变暖的背景下, 华北地区的落叶松林更趋向于垂直生长. ...
... 、4H、4L), 与前人的研究结果相符(
Gaston, 2000 ;
Moles et al ., 2009 ;
Zhang et al ., 2013 ).而与经纬度和
MAP 对华北地区落叶松林最大胸径和最大树高的影响的一致性不同,
MAT 对华北地区落叶松林最大胸径与最大树高的影响正好相反(
图4C 、4G).我们的结果显示: 华北地区落叶松林最大胸径随
MAT 的增加而 降低, 随
MAP 的增加而增加(
图4C 、4G), 与
Yan等 (2006) 对鼎湖山亚热带常绿阔叶林优势乔木胸径的研究及
Tao等(2016) 对全球树高的研究结果一致.落叶松属植物为阳性物种, 作为林线植被, 气候变暖对落叶松的生长及群落更新可能是有利的(
Devi et al ., 2008 ).在全球气候变暖的背景下, 华北地区的落叶松林更趋向于垂直生长. ...
... 从森林起源来看, 落叶松天然林和落叶松人工林的群落结构特征及物种多样性与地理及气候因子之间的关系, 与落叶松林总体的基本一致.其在地理因子上的差异主要体现在最大树高上, 落叶松人工林的最大树高随经纬度的增加而减少, 与前人在全球地理格局的研究结果一致(
Gaston, 2000 ;
Moles et al ., 2009 ;
Tao et al ., 2016 ).但是落叶松天然林的最大树高随经纬度的增加而增加, 与前人的研究结果(
Gaston, 2000 ;
Moles et al ., 2009 ;
Tao et al ., 2016 ) 相反, 其原因可能与海拔高度有关.华北地区低纬度和经度位置偏西的落叶松天然林主要是太白红杉林(
图1 ), 其分布海拔高, 树高相对低矮(
表5 ), 而高纬度位置及经度位置偏东的落叶松天然林为华北落叶松林, 其分布海拔较太白红杉林低(
郑万均和傅立国, 1978 ), 树高更高(
表5 ), 即华北地区落叶松林与地理因素的关系主要由落叶松人工林所主导.落叶松人工林和落叶松天然林在气候因子上的差异体现在最大胸径与
MAT 以及最大树高与
MAP 的关系上(
图4C 、4H).落叶松天然林的最大胸径和最大树高分别随
MAT 和
MAP 的增大而减小, 而落叶松人工林的最大胸径和最大树高分别随
MAT 和
MAP 的增大而增大.落叶松林总体最大胸径与气候条件的关系与落叶松天然林一致, 最大树高与气候条件的关系与落叶松人工林一致, 而对比落叶松天然林和落叶松人工林优势种的胸径树高, 落叶松天然林优势种的胸径显著大于人工林, 而树高显著小于人工林(
表5 ), 即华北地区落叶松林与
MAT 的关系主要受落叶松天然林的影响, 而与
MAP 的关系主要受落叶松人工林的影响. ...
... ).但是落叶松天然林的最大树高随经纬度的增加而增加, 与前人的研究结果(
Gaston, 2000 ;
Moles et al ., 2009 ;
Tao et al ., 2016 ) 相反, 其原因可能与海拔高度有关.华北地区低纬度和经度位置偏西的落叶松天然林主要是太白红杉林(
图1 ), 其分布海拔高, 树高相对低矮(
表5 ), 而高纬度位置及经度位置偏东的落叶松天然林为华北落叶松林, 其分布海拔较太白红杉林低(
郑万均和傅立国, 1978 ), 树高更高(
表5 ), 即华北地区落叶松林与地理因素的关系主要由落叶松人工林所主导.落叶松人工林和落叶松天然林在气候因子上的差异体现在最大胸径与
MAT 以及最大树高与
MAP 的关系上(
图4C 、4H).落叶松天然林的最大胸径和最大树高分别随
MAT 和
MAP 的增大而减小, 而落叶松人工林的最大胸径和最大树高分别随
MAT 和
MAP 的增大而增大.落叶松林总体最大胸径与气候条件的关系与落叶松天然林一致, 最大树高与气候条件的关系与落叶松人工林一致, 而对比落叶松天然林和落叶松人工林优势种的胸径树高, 落叶松天然林优势种的胸径显著大于人工林, 而树高显著小于人工林(
表5 ), 即华北地区落叶松林与
MAT 的关系主要受落叶松天然林的影响, 而与
MAP 的关系主要受落叶松人工林的影响. ...
秦岭太白红杉种群空间分布格局研究
1
2004
... 落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要.前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面.而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面.而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道. ...
秦岭太白红杉种群空间分布格局研究
1
2004
... 落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要.前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面.而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面.而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道. ...
四川盆地日本落叶松林抚育间伐研究
1
2014
... 除地理及环境因子的影响外, 人为干扰也是影响森林群落结构和物种丰富度的重要影响因子.对落叶松天然林而言, 人为干扰可能主要体现在旅游方面.近年来, 由于生态旅游的兴起, 大量的游客涌入到自然保护区的核心区, 砍椽(搭设旅游接待帐篷用)、烧柴(取暖和生活用柴)以及生活垃圾污染等对山地环境产生重要影响, 诸如本文研究区域的秦岭太白山保护区就是近年来生态旅游的一大热点地区(
刘香妮, 2001 ).对落叶松人工林而言, 其初期的抚育间伐对落叶松幼龄林及中龄林的群落结构和物种丰富度影响甚大(
李国雷等, 2009 ;
李淑辉等, 2012 ;
郭琳等, 2014 ).而我们调查的落叶松林大部分为成熟林, 甚至部分已经向人工天然混交林状态发展, 群落结构及物种丰富度已经相对稳定.对比落叶松天然林和落叶松人工林的胸径树高结构发现, 落叶松天然林的胸径大于人工林, 而其树高则小于人工林(
表5 ).由于落叶松人工林主要营造于中低海拔地带, 而现存的落叶松天然林主要在高海拔区域.所以这除了可能与森林经营管理有关外, 落叶松天然林和人工林的分布海拔对其胸径树高结构的影响可能更大.对比落叶松天然林与人工林的物种丰富度并无显著差异(
表4 ).其原因可能是由于华北地区的落叶松人工林属于三北防护林工程的水土涵养林, 对于成过熟林乃至有天然化趋势的人工林, 人为的经营管理活动较少, 故而其群落结构和物种丰富度受到的影响可能较小(
表4 ,
表5 ), 而放牧等活动对该地区中低海拔落叶松人工林的影响或许 更大. ...
四川盆地日本落叶松林抚育间伐研究
1
2014
... 除地理及环境因子的影响外, 人为干扰也是影响森林群落结构和物种丰富度的重要影响因子.对落叶松天然林而言, 人为干扰可能主要体现在旅游方面.近年来, 由于生态旅游的兴起, 大量的游客涌入到自然保护区的核心区, 砍椽(搭设旅游接待帐篷用)、烧柴(取暖和生活用柴)以及生活垃圾污染等对山地环境产生重要影响, 诸如本文研究区域的秦岭太白山保护区就是近年来生态旅游的一大热点地区(
刘香妮, 2001 ).对落叶松人工林而言, 其初期的抚育间伐对落叶松幼龄林及中龄林的群落结构和物种丰富度影响甚大(
李国雷等, 2009 ;
李淑辉等, 2012 ;
郭琳等, 2014 ).而我们调查的落叶松林大部分为成熟林, 甚至部分已经向人工天然混交林状态发展, 群落结构及物种丰富度已经相对稳定.对比落叶松天然林和落叶松人工林的胸径树高结构发现, 落叶松天然林的胸径大于人工林, 而其树高则小于人工林(
表5 ).由于落叶松人工林主要营造于中低海拔地带, 而现存的落叶松天然林主要在高海拔区域.所以这除了可能与森林经营管理有关外, 落叶松天然林和人工林的分布海拔对其胸径树高结构的影响可能更大.对比落叶松天然林与人工林的物种丰富度并无显著差异(
表4 ).其原因可能是由于华北地区的落叶松人工林属于三北防护林工程的水土涵养林, 对于成过熟林乃至有天然化趋势的人工林, 人为的经营管理活动较少, 故而其群落结构和物种丰富度受到的影响可能较小(
表4 ,
表5 ), 而放牧等活动对该地区中低海拔落叶松人工林的影响或许 更大. ...
气象因子对豫西伏牛山区日本落叶松生长量影响研究
1
2006
... 落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要.前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面.而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面.而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道. ...
气象因子对豫西伏牛山区日本落叶松生长量影响研究
1
2006
... 落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要.前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面.而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面.而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道. ...
不同林龄华北落叶松人工林生物量及营养元素分布特征
1
2016
... 落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要.前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面.而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面.而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道. ...
不同林龄华北落叶松人工林生物量及营养元素分布特征
1
2016
... 落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要.前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面.而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面.而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道. ...
华北落叶松林木生长、草本植物多样性及地形因子之间的关系
1
2014
... 落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要.前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面.而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面.而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道. ...
华北落叶松林木生长、草本植物多样性及地形因子之间的关系
1
2014
... 落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要.前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面.而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面.而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道. ...
长白山北坡云冷杉林和落叶松林物种组成与群落结构
1
2014
... 植物群落结构可以反映出群落的整体状态.3种落叶松林胸径和树高频度分布的偏度系数大于0且峰度系数大于3 (
表5 ), 即3种落叶松林的胸径和树高结构总体均呈右偏分布且其径级大小和高度大小相对集中, 表明3种落叶松林均处于相对稳定的演替阶段(
Tyrrell & Crow, 1994 ;
Aiba & Kitayama, 1999 ;
Fang et al ., 2012 ).
匡旭等(2014) 对中国东北长白山落叶松林的研究结果显示, 长白落叶松(
L. olgensis )有较多
DBH < 10 cm的个体, 群落处于生长状态.
赵文智(1997) 对河北坝上华北落叶松人工林的研究表明, 30年华北落叶松成熟林林分稳定, 有天然更新的潜力.我们的结果与以上研究结果一致. ...
长白山北坡云冷杉林和落叶松林物种组成与群落结构
1
2014
... 植物群落结构可以反映出群落的整体状态.3种落叶松林胸径和树高频度分布的偏度系数大于0且峰度系数大于3 (
表5 ), 即3种落叶松林的胸径和树高结构总体均呈右偏分布且其径级大小和高度大小相对集中, 表明3种落叶松林均处于相对稳定的演替阶段(
Tyrrell & Crow, 1994 ;
Aiba & Kitayama, 1999 ;
Fang et al ., 2012 ).
匡旭等(2014) 对中国东北长白山落叶松林的研究结果显示, 长白落叶松(
L. olgensis )有较多
DBH < 10 cm的个体, 群落处于生长状态.
赵文智(1997) 对河北坝上华北落叶松人工林的研究表明, 30年华北落叶松成熟林林分稳定, 有天然更新的潜力.我们的结果与以上研究结果一致. ...
2
1999
... 落叶松属(
Larix )植物是中国北方广泛分布的寒温性针叶林的建群种, 也是重要的用材树种(
吴征镒, 1980 ;
李楠, 1995 ;
刘增力等, 2002 ).在我国华北地区, 落叶松属植物是很多山地的林线物种, 如太白山的林线物种为太白红杉(
Larix chinensis )(
段仁燕等, 2010 ), 五台山、关帝山及小五台山的林线物种为华北落叶松(
L. principis-rupprechtii )(
Liu et al ., 2002 ), 这些落叶松对维护高海拔地区的生态环境有重要意义.目前中国华北地区主要分布有3种落叶松: 华北落叶松、日本落叶松(
L. kaempferi )及太白红杉(
郑万均, 1983 ).其中, 华北落叶松及太白红杉是中国特有种, 日本落叶松为华北地区广泛栽培种 (
郑万均和傅立国, 1978 ).这3种落叶松是华北地区落叶松林的优势种.其中, 日本落叶松林为人工林; 而华北落叶松林虽然在华北地区有天然分布, 但是由于其分布的海拔较低, 受人为干扰严重, 因而目前纯原始林较少, 大部分是受干扰的次生林或者栽培的人工林(
刘濂, 1996 ); 相较而言, 太白红杉林虽仅分布于我国秦岭山地, 但其分布海拔较高, 受人为干扰影响较小, 且其适应性极强, 在土层贫瘠气候寒冷的条件下也能适应, 为我国秦岭山区森林上限植被, 目前主要是原始林(
雷明德, 1999 ;
应俊生和陈梦玲, 2011 ). ...
... 物种丰富度是测度物种多样性的一个重要维度(
Gaston, 2000 ).华北地区, 3种落叶松林的物种丰富度总体比较丰富, 乔、灌、草层的物种丰富度有显著的差异(
图3 ;
表4 ).这与3种落叶松林的物种组成、气候条件以及乔木层郁闭度等因素有关(
Wang et al ., 2009 ;
Mölder et al ., 2014 ).具体而言, 太白红杉林为秦岭山地原始林(
雷明德, 1999 ), 华北落叶松林也多为人工林或次生林(
刘濂, 1996 ), 两种落叶松林优势种太白红杉及华北落叶松的重要值很高(
表3 ), 大部分甚至是纯林, 因而它们的乔木层的物种丰富度很低(
表4 ), 没有显著差异(
图3 A).而日本落叶松林虽然是人工林, 但其分布海拔低于华北落叶松林和太白红杉林, 拥有更好的水热条件, 群落物种更为丰富; 通常与其他物种混交, 其乔木层优势种日本落叶松的重要值只有70.2% (
表3 ), 因而其乔木层的物种丰富度显著大于华北落叶松林和太白红杉林(
图3 A).此外, 日本落叶松林与华北落叶松林均分布于中低海拔区域, 乔木层郁闭度大, 林下植物尤其是草本植物的生长受到影响, 其草本层丰富度相对较低, 没有显著差异(
图3 C).而太白红杉林位于秦岭山地高海拔区域, 乔木层郁闭度相对较小, 有利于林下植物的生长, 其林下草本层的物种丰富度显著大于华北落叶松林和日本落叶松林(
图3 C).但是, 也正是由于分布海拔高的影响, 其林下的生境条件较差, 所以对比华北地区纬度位置相差不大的日本落叶松林, 其灌木层物种丰富度没有显著优势(
图3 B). ...
2
1999
... 落叶松属(
Larix )植物是中国北方广泛分布的寒温性针叶林的建群种, 也是重要的用材树种(
吴征镒, 1980 ;
李楠, 1995 ;
刘增力等, 2002 ).在我国华北地区, 落叶松属植物是很多山地的林线物种, 如太白山的林线物种为太白红杉(
Larix chinensis )(
段仁燕等, 2010 ), 五台山、关帝山及小五台山的林线物种为华北落叶松(
L. principis-rupprechtii )(
Liu et al ., 2002 ), 这些落叶松对维护高海拔地区的生态环境有重要意义.目前中国华北地区主要分布有3种落叶松: 华北落叶松、日本落叶松(
L. kaempferi )及太白红杉(
郑万均, 1983 ).其中, 华北落叶松及太白红杉是中国特有种, 日本落叶松为华北地区广泛栽培种 (
郑万均和傅立国, 1978 ).这3种落叶松是华北地区落叶松林的优势种.其中, 日本落叶松林为人工林; 而华北落叶松林虽然在华北地区有天然分布, 但是由于其分布的海拔较低, 受人为干扰严重, 因而目前纯原始林较少, 大部分是受干扰的次生林或者栽培的人工林(
刘濂, 1996 ); 相较而言, 太白红杉林虽仅分布于我国秦岭山地, 但其分布海拔较高, 受人为干扰影响较小, 且其适应性极强, 在土层贫瘠气候寒冷的条件下也能适应, 为我国秦岭山区森林上限植被, 目前主要是原始林(
雷明德, 1999 ;
应俊生和陈梦玲, 2011 ). ...
... 物种丰富度是测度物种多样性的一个重要维度(
Gaston, 2000 ).华北地区, 3种落叶松林的物种丰富度总体比较丰富, 乔、灌、草层的物种丰富度有显著的差异(
图3 ;
表4 ).这与3种落叶松林的物种组成、气候条件以及乔木层郁闭度等因素有关(
Wang et al ., 2009 ;
Mölder et al ., 2014 ).具体而言, 太白红杉林为秦岭山地原始林(
雷明德, 1999 ), 华北落叶松林也多为人工林或次生林(
刘濂, 1996 ), 两种落叶松林优势种太白红杉及华北落叶松的重要值很高(
表3 ), 大部分甚至是纯林, 因而它们的乔木层的物种丰富度很低(
表4 ), 没有显著差异(
图3 A).而日本落叶松林虽然是人工林, 但其分布海拔低于华北落叶松林和太白红杉林, 拥有更好的水热条件, 群落物种更为丰富; 通常与其他物种混交, 其乔木层优势种日本落叶松的重要值只有70.2% (
表3 ), 因而其乔木层的物种丰富度显著大于华北落叶松林和太白红杉林(
图3 A).此外, 日本落叶松林与华北落叶松林均分布于中低海拔区域, 乔木层郁闭度大, 林下植物尤其是草本植物的生长受到影响, 其草本层丰富度相对较低, 没有显著差异(
图3 C).而太白红杉林位于秦岭山地高海拔区域, 乔木层郁闭度相对较小, 有利于林下植物的生长, 其林下草本层的物种丰富度显著大于华北落叶松林和日本落叶松林(
图3 C).但是, 也正是由于分布海拔高的影响, 其林下的生境条件较差, 所以对比华北地区纬度位置相差不大的日本落叶松林, 其灌木层物种丰富度没有显著优势(
图3 B). ...
华北落叶松人工林密度调控对林下植被发育的作用过程
1
2009
... 除地理及环境因子的影响外, 人为干扰也是影响森林群落结构和物种丰富度的重要影响因子.对落叶松天然林而言, 人为干扰可能主要体现在旅游方面.近年来, 由于生态旅游的兴起, 大量的游客涌入到自然保护区的核心区, 砍椽(搭设旅游接待帐篷用)、烧柴(取暖和生活用柴)以及生活垃圾污染等对山地环境产生重要影响, 诸如本文研究区域的秦岭太白山保护区就是近年来生态旅游的一大热点地区(
刘香妮, 2001 ).对落叶松人工林而言, 其初期的抚育间伐对落叶松幼龄林及中龄林的群落结构和物种丰富度影响甚大(
李国雷等, 2009 ;
李淑辉等, 2012 ;
郭琳等, 2014 ).而我们调查的落叶松林大部分为成熟林, 甚至部分已经向人工天然混交林状态发展, 群落结构及物种丰富度已经相对稳定.对比落叶松天然林和落叶松人工林的胸径树高结构发现, 落叶松天然林的胸径大于人工林, 而其树高则小于人工林(
表5 ).由于落叶松人工林主要营造于中低海拔地带, 而现存的落叶松天然林主要在高海拔区域.所以这除了可能与森林经营管理有关外, 落叶松天然林和人工林的分布海拔对其胸径树高结构的影响可能更大.对比落叶松天然林与人工林的物种丰富度并无显著差异(
表4 ).其原因可能是由于华北地区的落叶松人工林属于三北防护林工程的水土涵养林, 对于成过熟林乃至有天然化趋势的人工林, 人为的经营管理活动较少, 故而其群落结构和物种丰富度受到的影响可能较小(
表4 ,
表5 ), 而放牧等活动对该地区中低海拔落叶松人工林的影响或许 更大. ...
华北落叶松人工林密度调控对林下植被发育的作用过程
1
2009
... 除地理及环境因子的影响外, 人为干扰也是影响森林群落结构和物种丰富度的重要影响因子.对落叶松天然林而言, 人为干扰可能主要体现在旅游方面.近年来, 由于生态旅游的兴起, 大量的游客涌入到自然保护区的核心区, 砍椽(搭设旅游接待帐篷用)、烧柴(取暖和生活用柴)以及生活垃圾污染等对山地环境产生重要影响, 诸如本文研究区域的秦岭太白山保护区就是近年来生态旅游的一大热点地区(
刘香妮, 2001 ).对落叶松人工林而言, 其初期的抚育间伐对落叶松幼龄林及中龄林的群落结构和物种丰富度影响甚大(
李国雷等, 2009 ;
李淑辉等, 2012 ;
郭琳等, 2014 ).而我们调查的落叶松林大部分为成熟林, 甚至部分已经向人工天然混交林状态发展, 群落结构及物种丰富度已经相对稳定.对比落叶松天然林和落叶松人工林的胸径树高结构发现, 落叶松天然林的胸径大于人工林, 而其树高则小于人工林(
表5 ).由于落叶松人工林主要营造于中低海拔地带, 而现存的落叶松天然林主要在高海拔区域.所以这除了可能与森林经营管理有关外, 落叶松天然林和人工林的分布海拔对其胸径树高结构的影响可能更大.对比落叶松天然林与人工林的物种丰富度并无显著差异(
表4 ).其原因可能是由于华北地区的落叶松人工林属于三北防护林工程的水土涵养林, 对于成过熟林乃至有天然化趋势的人工林, 人为的经营管理活动较少, 故而其群落结构和物种丰富度受到的影响可能较小(
表4 ,
表5 ), 而放牧等活动对该地区中低海拔落叶松人工林的影响或许 更大. ...
论松科植物的地理分布、起源和扩散
1
1995
... 落叶松属(
Larix )植物是中国北方广泛分布的寒温性针叶林的建群种, 也是重要的用材树种(
吴征镒, 1980 ;
李楠, 1995 ;
刘增力等, 2002 ).在我国华北地区, 落叶松属植物是很多山地的林线物种, 如太白山的林线物种为太白红杉(
Larix chinensis )(
段仁燕等, 2010 ), 五台山、关帝山及小五台山的林线物种为华北落叶松(
L. principis-rupprechtii )(
Liu et al ., 2002 ), 这些落叶松对维护高海拔地区的生态环境有重要意义.目前中国华北地区主要分布有3种落叶松: 华北落叶松、日本落叶松(
L. kaempferi )及太白红杉(
郑万均, 1983 ).其中, 华北落叶松及太白红杉是中国特有种, 日本落叶松为华北地区广泛栽培种 (
郑万均和傅立国, 1978 ).这3种落叶松是华北地区落叶松林的优势种.其中, 日本落叶松林为人工林; 而华北落叶松林虽然在华北地区有天然分布, 但是由于其分布的海拔较低, 受人为干扰严重, 因而目前纯原始林较少, 大部分是受干扰的次生林或者栽培的人工林(
刘濂, 1996 ); 相较而言, 太白红杉林虽仅分布于我国秦岭山地, 但其分布海拔较高, 受人为干扰影响较小, 且其适应性极强, 在土层贫瘠气候寒冷的条件下也能适应, 为我国秦岭山区森林上限植被, 目前主要是原始林(
雷明德, 1999 ;
应俊生和陈梦玲, 2011 ). ...
论松科植物的地理分布、起源和扩散
1
1995
... 落叶松属(
Larix )植物是中国北方广泛分布的寒温性针叶林的建群种, 也是重要的用材树种(
吴征镒, 1980 ;
李楠, 1995 ;
刘增力等, 2002 ).在我国华北地区, 落叶松属植物是很多山地的林线物种, 如太白山的林线物种为太白红杉(
Larix chinensis )(
段仁燕等, 2010 ), 五台山、关帝山及小五台山的林线物种为华北落叶松(
L. principis-rupprechtii )(
Liu et al ., 2002 ), 这些落叶松对维护高海拔地区的生态环境有重要意义.目前中国华北地区主要分布有3种落叶松: 华北落叶松、日本落叶松(
L. kaempferi )及太白红杉(
郑万均, 1983 ).其中, 华北落叶松及太白红杉是中国特有种, 日本落叶松为华北地区广泛栽培种 (
郑万均和傅立国, 1978 ).这3种落叶松是华北地区落叶松林的优势种.其中, 日本落叶松林为人工林; 而华北落叶松林虽然在华北地区有天然分布, 但是由于其分布的海拔较低, 受人为干扰严重, 因而目前纯原始林较少, 大部分是受干扰的次生林或者栽培的人工林(
刘濂, 1996 ); 相较而言, 太白红杉林虽仅分布于我国秦岭山地, 但其分布海拔较高, 受人为干扰影响较小, 且其适应性极强, 在土层贫瘠气候寒冷的条件下也能适应, 为我国秦岭山区森林上限植被, 目前主要是原始林(
雷明德, 1999 ;
应俊生和陈梦玲, 2011 ). ...
关帝山林区华北落叶松人工林抚育间伐对林下植被的影响
1
2012
... 除地理及环境因子的影响外, 人为干扰也是影响森林群落结构和物种丰富度的重要影响因子.对落叶松天然林而言, 人为干扰可能主要体现在旅游方面.近年来, 由于生态旅游的兴起, 大量的游客涌入到自然保护区的核心区, 砍椽(搭设旅游接待帐篷用)、烧柴(取暖和生活用柴)以及生活垃圾污染等对山地环境产生重要影响, 诸如本文研究区域的秦岭太白山保护区就是近年来生态旅游的一大热点地区(
刘香妮, 2001 ).对落叶松人工林而言, 其初期的抚育间伐对落叶松幼龄林及中龄林的群落结构和物种丰富度影响甚大(
李国雷等, 2009 ;
李淑辉等, 2012 ;
郭琳等, 2014 ).而我们调查的落叶松林大部分为成熟林, 甚至部分已经向人工天然混交林状态发展, 群落结构及物种丰富度已经相对稳定.对比落叶松天然林和落叶松人工林的胸径树高结构发现, 落叶松天然林的胸径大于人工林, 而其树高则小于人工林(
表5 ).由于落叶松人工林主要营造于中低海拔地带, 而现存的落叶松天然林主要在高海拔区域.所以这除了可能与森林经营管理有关外, 落叶松天然林和人工林的分布海拔对其胸径树高结构的影响可能更大.对比落叶松天然林与人工林的物种丰富度并无显著差异(
表4 ).其原因可能是由于华北地区的落叶松人工林属于三北防护林工程的水土涵养林, 对于成过熟林乃至有天然化趋势的人工林, 人为的经营管理活动较少, 故而其群落结构和物种丰富度受到的影响可能较小(
表4 ,
表5 ), 而放牧等活动对该地区中低海拔落叶松人工林的影响或许 更大. ...
关帝山林区华北落叶松人工林抚育间伐对林下植被的影响
1
2012
... 除地理及环境因子的影响外, 人为干扰也是影响森林群落结构和物种丰富度的重要影响因子.对落叶松天然林而言, 人为干扰可能主要体现在旅游方面.近年来, 由于生态旅游的兴起, 大量的游客涌入到自然保护区的核心区, 砍椽(搭设旅游接待帐篷用)、烧柴(取暖和生活用柴)以及生活垃圾污染等对山地环境产生重要影响, 诸如本文研究区域的秦岭太白山保护区就是近年来生态旅游的一大热点地区(
刘香妮, 2001 ).对落叶松人工林而言, 其初期的抚育间伐对落叶松幼龄林及中龄林的群落结构和物种丰富度影响甚大(
李国雷等, 2009 ;
李淑辉等, 2012 ;
郭琳等, 2014 ).而我们调查的落叶松林大部分为成熟林, 甚至部分已经向人工天然混交林状态发展, 群落结构及物种丰富度已经相对稳定.对比落叶松天然林和落叶松人工林的胸径树高结构发现, 落叶松天然林的胸径大于人工林, 而其树高则小于人工林(
表5 ).由于落叶松人工林主要营造于中低海拔地带, 而现存的落叶松天然林主要在高海拔区域.所以这除了可能与森林经营管理有关外, 落叶松天然林和人工林的分布海拔对其胸径树高结构的影响可能更大.对比落叶松天然林与人工林的物种丰富度并无显著差异(
表4 ).其原因可能是由于华北地区的落叶松人工林属于三北防护林工程的水土涵养林, 对于成过熟林乃至有天然化趋势的人工林, 人为的经营管理活动较少, 故而其群落结构和物种丰富度受到的影响可能较小(
表4 ,
表5 ), 而放牧等活动对该地区中低海拔落叶松人工林的影响或许 更大. ...
Larch timberline and its development in North China
1
2002
... 落叶松属(
Larix )植物是中国北方广泛分布的寒温性针叶林的建群种, 也是重要的用材树种(
吴征镒, 1980 ;
李楠, 1995 ;
刘增力等, 2002 ).在我国华北地区, 落叶松属植物是很多山地的林线物种, 如太白山的林线物种为太白红杉(
Larix chinensis )(
段仁燕等, 2010 ), 五台山、关帝山及小五台山的林线物种为华北落叶松(
L. principis-rupprechtii )(
Liu et al ., 2002 ), 这些落叶松对维护高海拔地区的生态环境有重要意义.目前中国华北地区主要分布有3种落叶松: 华北落叶松、日本落叶松(
L. kaempferi )及太白红杉(
郑万均, 1983 ).其中, 华北落叶松及太白红杉是中国特有种, 日本落叶松为华北地区广泛栽培种 (
郑万均和傅立国, 1978 ).这3种落叶松是华北地区落叶松林的优势种.其中, 日本落叶松林为人工林; 而华北落叶松林虽然在华北地区有天然分布, 但是由于其分布的海拔较低, 受人为干扰严重, 因而目前纯原始林较少, 大部分是受干扰的次生林或者栽培的人工林(
刘濂, 1996 ); 相较而言, 太白红杉林虽仅分布于我国秦岭山地, 但其分布海拔较高, 受人为干扰影响较小, 且其适应性极强, 在土层贫瘠气候寒冷的条件下也能适应, 为我国秦岭山区森林上限植被, 目前主要是原始林(
雷明德, 1999 ;
应俊生和陈梦玲, 2011 ). ...
2
1996
... 落叶松属(
Larix )植物是中国北方广泛分布的寒温性针叶林的建群种, 也是重要的用材树种(
吴征镒, 1980 ;
李楠, 1995 ;
刘增力等, 2002 ).在我国华北地区, 落叶松属植物是很多山地的林线物种, 如太白山的林线物种为太白红杉(
Larix chinensis )(
段仁燕等, 2010 ), 五台山、关帝山及小五台山的林线物种为华北落叶松(
L. principis-rupprechtii )(
Liu et al ., 2002 ), 这些落叶松对维护高海拔地区的生态环境有重要意义.目前中国华北地区主要分布有3种落叶松: 华北落叶松、日本落叶松(
L. kaempferi )及太白红杉(
郑万均, 1983 ).其中, 华北落叶松及太白红杉是中国特有种, 日本落叶松为华北地区广泛栽培种 (
郑万均和傅立国, 1978 ).这3种落叶松是华北地区落叶松林的优势种.其中, 日本落叶松林为人工林; 而华北落叶松林虽然在华北地区有天然分布, 但是由于其分布的海拔较低, 受人为干扰严重, 因而目前纯原始林较少, 大部分是受干扰的次生林或者栽培的人工林(
刘濂, 1996 ); 相较而言, 太白红杉林虽仅分布于我国秦岭山地, 但其分布海拔较高, 受人为干扰影响较小, 且其适应性极强, 在土层贫瘠气候寒冷的条件下也能适应, 为我国秦岭山区森林上限植被, 目前主要是原始林(
雷明德, 1999 ;
应俊生和陈梦玲, 2011 ). ...
... 物种丰富度是测度物种多样性的一个重要维度(
Gaston, 2000 ).华北地区, 3种落叶松林的物种丰富度总体比较丰富, 乔、灌、草层的物种丰富度有显著的差异(
图3 ;
表4 ).这与3种落叶松林的物种组成、气候条件以及乔木层郁闭度等因素有关(
Wang et al ., 2009 ;
Mölder et al ., 2014 ).具体而言, 太白红杉林为秦岭山地原始林(
雷明德, 1999 ), 华北落叶松林也多为人工林或次生林(
刘濂, 1996 ), 两种落叶松林优势种太白红杉及华北落叶松的重要值很高(
表3 ), 大部分甚至是纯林, 因而它们的乔木层的物种丰富度很低(
表4 ), 没有显著差异(
图3 A).而日本落叶松林虽然是人工林, 但其分布海拔低于华北落叶松林和太白红杉林, 拥有更好的水热条件, 群落物种更为丰富; 通常与其他物种混交, 其乔木层优势种日本落叶松的重要值只有70.2% (
表3 ), 因而其乔木层的物种丰富度显著大于华北落叶松林和太白红杉林(
图3 A).此外, 日本落叶松林与华北落叶松林均分布于中低海拔区域, 乔木层郁闭度大, 林下植物尤其是草本植物的生长受到影响, 其草本层丰富度相对较低, 没有显著差异(
图3 C).而太白红杉林位于秦岭山地高海拔区域, 乔木层郁闭度相对较小, 有利于林下植物的生长, 其林下草本层的物种丰富度显著大于华北落叶松林和日本落叶松林(
图3 C).但是, 也正是由于分布海拔高的影响, 其林下的生境条件较差, 所以对比华北地区纬度位置相差不大的日本落叶松林, 其灌木层物种丰富度没有显著优势(
图3 B). ...
2
1996
... 落叶松属(
Larix )植物是中国北方广泛分布的寒温性针叶林的建群种, 也是重要的用材树种(
吴征镒, 1980 ;
李楠, 1995 ;
刘增力等, 2002 ).在我国华北地区, 落叶松属植物是很多山地的林线物种, 如太白山的林线物种为太白红杉(
Larix chinensis )(
段仁燕等, 2010 ), 五台山、关帝山及小五台山的林线物种为华北落叶松(
L. principis-rupprechtii )(
Liu et al ., 2002 ), 这些落叶松对维护高海拔地区的生态环境有重要意义.目前中国华北地区主要分布有3种落叶松: 华北落叶松、日本落叶松(
L. kaempferi )及太白红杉(
郑万均, 1983 ).其中, 华北落叶松及太白红杉是中国特有种, 日本落叶松为华北地区广泛栽培种 (
郑万均和傅立国, 1978 ).这3种落叶松是华北地区落叶松林的优势种.其中, 日本落叶松林为人工林; 而华北落叶松林虽然在华北地区有天然分布, 但是由于其分布的海拔较低, 受人为干扰严重, 因而目前纯原始林较少, 大部分是受干扰的次生林或者栽培的人工林(
刘濂, 1996 ); 相较而言, 太白红杉林虽仅分布于我国秦岭山地, 但其分布海拔较高, 受人为干扰影响较小, 且其适应性极强, 在土层贫瘠气候寒冷的条件下也能适应, 为我国秦岭山区森林上限植被, 目前主要是原始林(
雷明德, 1999 ;
应俊生和陈梦玲, 2011 ). ...
... 物种丰富度是测度物种多样性的一个重要维度(
Gaston, 2000 ).华北地区, 3种落叶松林的物种丰富度总体比较丰富, 乔、灌、草层的物种丰富度有显著的差异(
图3 ;
表4 ).这与3种落叶松林的物种组成、气候条件以及乔木层郁闭度等因素有关(
Wang et al ., 2009 ;
Mölder et al ., 2014 ).具体而言, 太白红杉林为秦岭山地原始林(
雷明德, 1999 ), 华北落叶松林也多为人工林或次生林(
刘濂, 1996 ), 两种落叶松林优势种太白红杉及华北落叶松的重要值很高(
表3 ), 大部分甚至是纯林, 因而它们的乔木层的物种丰富度很低(
表4 ), 没有显著差异(
图3 A).而日本落叶松林虽然是人工林, 但其分布海拔低于华北落叶松林和太白红杉林, 拥有更好的水热条件, 群落物种更为丰富; 通常与其他物种混交, 其乔木层优势种日本落叶松的重要值只有70.2% (
表3 ), 因而其乔木层的物种丰富度显著大于华北落叶松林和太白红杉林(
图3 A).此外, 日本落叶松林与华北落叶松林均分布于中低海拔区域, 乔木层郁闭度大, 林下植物尤其是草本植物的生长受到影响, 其草本层丰富度相对较低, 没有显著差异(
图3 C).而太白红杉林位于秦岭山地高海拔区域, 乔木层郁闭度相对较小, 有利于林下植物的生长, 其林下草本层的物种丰富度显著大于华北落叶松林和日本落叶松林(
图3 C).但是, 也正是由于分布海拔高的影响, 其林下的生境条件较差, 所以对比华北地区纬度位置相差不大的日本落叶松林, 其灌木层物种丰富度没有显著优势(
图3 B). ...
人为活动对太白红杉林的影响调查
1
2001
... 除地理及环境因子的影响外, 人为干扰也是影响森林群落结构和物种丰富度的重要影响因子.对落叶松天然林而言, 人为干扰可能主要体现在旅游方面.近年来, 由于生态旅游的兴起, 大量的游客涌入到自然保护区的核心区, 砍椽(搭设旅游接待帐篷用)、烧柴(取暖和生活用柴)以及生活垃圾污染等对山地环境产生重要影响, 诸如本文研究区域的秦岭太白山保护区就是近年来生态旅游的一大热点地区(
刘香妮, 2001 ).对落叶松人工林而言, 其初期的抚育间伐对落叶松幼龄林及中龄林的群落结构和物种丰富度影响甚大(
李国雷等, 2009 ;
李淑辉等, 2012 ;
郭琳等, 2014 ).而我们调查的落叶松林大部分为成熟林, 甚至部分已经向人工天然混交林状态发展, 群落结构及物种丰富度已经相对稳定.对比落叶松天然林和落叶松人工林的胸径树高结构发现, 落叶松天然林的胸径大于人工林, 而其树高则小于人工林(
表5 ).由于落叶松人工林主要营造于中低海拔地带, 而现存的落叶松天然林主要在高海拔区域.所以这除了可能与森林经营管理有关外, 落叶松天然林和人工林的分布海拔对其胸径树高结构的影响可能更大.对比落叶松天然林与人工林的物种丰富度并无显著差异(
表4 ).其原因可能是由于华北地区的落叶松人工林属于三北防护林工程的水土涵养林, 对于成过熟林乃至有天然化趋势的人工林, 人为的经营管理活动较少, 故而其群落结构和物种丰富度受到的影响可能较小(
表4 ,
表5 ), 而放牧等活动对该地区中低海拔落叶松人工林的影响或许 更大. ...
人为活动对太白红杉林的影响调查
1
2001
... 除地理及环境因子的影响外, 人为干扰也是影响森林群落结构和物种丰富度的重要影响因子.对落叶松天然林而言, 人为干扰可能主要体现在旅游方面.近年来, 由于生态旅游的兴起, 大量的游客涌入到自然保护区的核心区, 砍椽(搭设旅游接待帐篷用)、烧柴(取暖和生活用柴)以及生活垃圾污染等对山地环境产生重要影响, 诸如本文研究区域的秦岭太白山保护区就是近年来生态旅游的一大热点地区(
刘香妮, 2001 ).对落叶松人工林而言, 其初期的抚育间伐对落叶松幼龄林及中龄林的群落结构和物种丰富度影响甚大(
李国雷等, 2009 ;
李淑辉等, 2012 ;
郭琳等, 2014 ).而我们调查的落叶松林大部分为成熟林, 甚至部分已经向人工天然混交林状态发展, 群落结构及物种丰富度已经相对稳定.对比落叶松天然林和落叶松人工林的胸径树高结构发现, 落叶松天然林的胸径大于人工林, 而其树高则小于人工林(
表5 ).由于落叶松人工林主要营造于中低海拔地带, 而现存的落叶松天然林主要在高海拔区域.所以这除了可能与森林经营管理有关外, 落叶松天然林和人工林的分布海拔对其胸径树高结构的影响可能更大.对比落叶松天然林与人工林的物种丰富度并无显著差异(
表4 ).其原因可能是由于华北地区的落叶松人工林属于三北防护林工程的水土涵养林, 对于成过熟林乃至有天然化趋势的人工林, 人为的经营管理活动较少, 故而其群落结构和物种丰富度受到的影响可能较小(
表4 ,
表5 ), 而放牧等活动对该地区中低海拔落叶松人工林的影响或许 更大. ...
中国冷杉、云杉和落叶松属植物的地理分布
1
2002
... 落叶松属(
Larix )植物是中国北方广泛分布的寒温性针叶林的建群种, 也是重要的用材树种(
吴征镒, 1980 ;
李楠, 1995 ;
刘增力等, 2002 ).在我国华北地区, 落叶松属植物是很多山地的林线物种, 如太白山的林线物种为太白红杉(
Larix chinensis )(
段仁燕等, 2010 ), 五台山、关帝山及小五台山的林线物种为华北落叶松(
L. principis-rupprechtii )(
Liu et al ., 2002 ), 这些落叶松对维护高海拔地区的生态环境有重要意义.目前中国华北地区主要分布有3种落叶松: 华北落叶松、日本落叶松(
L. kaempferi )及太白红杉(
郑万均, 1983 ).其中, 华北落叶松及太白红杉是中国特有种, 日本落叶松为华北地区广泛栽培种 (
郑万均和傅立国, 1978 ).这3种落叶松是华北地区落叶松林的优势种.其中, 日本落叶松林为人工林; 而华北落叶松林虽然在华北地区有天然分布, 但是由于其分布的海拔较低, 受人为干扰严重, 因而目前纯原始林较少, 大部分是受干扰的次生林或者栽培的人工林(
刘濂, 1996 ); 相较而言, 太白红杉林虽仅分布于我国秦岭山地, 但其分布海拔较高, 受人为干扰影响较小, 且其适应性极强, 在土层贫瘠气候寒冷的条件下也能适应, 为我国秦岭山区森林上限植被, 目前主要是原始林(
雷明德, 1999 ;
应俊生和陈梦玲, 2011 ). ...
中国冷杉、云杉和落叶松属植物的地理分布
1
2002
... 落叶松属(
Larix )植物是中国北方广泛分布的寒温性针叶林的建群种, 也是重要的用材树种(
吴征镒, 1980 ;
李楠, 1995 ;
刘增力等, 2002 ).在我国华北地区, 落叶松属植物是很多山地的林线物种, 如太白山的林线物种为太白红杉(
Larix chinensis )(
段仁燕等, 2010 ), 五台山、关帝山及小五台山的林线物种为华北落叶松(
L. principis-rupprechtii )(
Liu et al ., 2002 ), 这些落叶松对维护高海拔地区的生态环境有重要意义.目前中国华北地区主要分布有3种落叶松: 华北落叶松、日本落叶松(
L. kaempferi )及太白红杉(
郑万均, 1983 ).其中, 华北落叶松及太白红杉是中国特有种, 日本落叶松为华北地区广泛栽培种 (
郑万均和傅立国, 1978 ).这3种落叶松是华北地区落叶松林的优势种.其中, 日本落叶松林为人工林; 而华北落叶松林虽然在华北地区有天然分布, 但是由于其分布的海拔较低, 受人为干扰严重, 因而目前纯原始林较少, 大部分是受干扰的次生林或者栽培的人工林(
刘濂, 1996 ); 相较而言, 太白红杉林虽仅分布于我国秦岭山地, 但其分布海拔较高, 受人为干扰影响较小, 且其适应性极强, 在土层贫瘠气候寒冷的条件下也能适应, 为我国秦岭山区森林上限植被, 目前主要是原始林(
雷明德, 1999 ;
应俊生和陈梦玲, 2011 ). ...
华北落叶松人工林生物量及其分配模式
1
2009
... 落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要.前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面.而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面.而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道. ...
华北落叶松人工林生物量及其分配模式
1
2009
... 落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要.前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面.而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面.而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道. ...
亚热带日本落叶松人工林生态系统碳密度及其分配特征
1
2016
... 落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要.前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面.而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面.而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道. ...
亚热带日本落叶松人工林生态系统碳密度及其分配特征
1
2016
... 落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要.前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面.而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面.而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道. ...
日本落叶松人工林林分结构与生长量预测研究——以长岭岗林场为例
1
2007
... 落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要.前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面.而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面.而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道. ...
日本落叶松人工林林分结构与生长量预测研究——以长岭岗林场为例
1
2007
... 落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要.前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面.而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面.而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道. ...
When beech strikes back: How strict nature conservation reduces herb-layer diversity and productivity in Central European deciduous forests
1
2014
... 物种丰富度是测度物种多样性的一个重要维度(
Gaston, 2000 ).华北地区, 3种落叶松林的物种丰富度总体比较丰富, 乔、灌、草层的物种丰富度有显著的差异(
图3 ;
表4 ).这与3种落叶松林的物种组成、气候条件以及乔木层郁闭度等因素有关(
Wang et al ., 2009 ;
Mölder et al ., 2014 ).具体而言, 太白红杉林为秦岭山地原始林(
雷明德, 1999 ), 华北落叶松林也多为人工林或次生林(
刘濂, 1996 ), 两种落叶松林优势种太白红杉及华北落叶松的重要值很高(
表3 ), 大部分甚至是纯林, 因而它们的乔木层的物种丰富度很低(
表4 ), 没有显著差异(
图3 A).而日本落叶松林虽然是人工林, 但其分布海拔低于华北落叶松林和太白红杉林, 拥有更好的水热条件, 群落物种更为丰富; 通常与其他物种混交, 其乔木层优势种日本落叶松的重要值只有70.2% (
表3 ), 因而其乔木层的物种丰富度显著大于华北落叶松林和太白红杉林(
图3 A).此外, 日本落叶松林与华北落叶松林均分布于中低海拔区域, 乔木层郁闭度大, 林下植物尤其是草本植物的生长受到影响, 其草本层丰富度相对较低, 没有显著差异(
图3 C).而太白红杉林位于秦岭山地高海拔区域, 乔木层郁闭度相对较小, 有利于林下植物的生长, 其林下草本层的物种丰富度显著大于华北落叶松林和日本落叶松林(
图3 C).但是, 也正是由于分布海拔高的影响, 其林下的生境条件较差, 所以对比华北地区纬度位置相差不大的日本落叶松林, 其灌木层物种丰富度没有显著优势(
图3 B). ...
Global patterns in plant height
4
2009
... 华北地区落叶松林的群落结构特征及物种多样性与其分布的地理位置和气候条件紧密相关.总体而言, 除最大树高与经度的关系不显著以外, 华北地区落叶松林最大胸径和最大树高及总体的物种丰富度均随着经纬度的增加而显著降低(
图4A 、4D、 4E、4F、4I、4J), 这与前人在全球地理格局的研究结果一致(
Gaston, 2000 ;
Moles et al ., 2009 ;
Tao et al ., 2016 ).此外, 诸多的研究表明, 水分条件是乔木胸径和高度及植物群落物种丰富度的重要影响因子(
Gaston, 2000 ;
Zhang et al ., 2013 ;
Tao et al ., 2016 ).在华北地区, 落叶松林最大胸径与最大树高及其总体的物种丰富度随着
MAP 的增加而增加(
图4D 、4H、4L), 与前人的研究结果相符(
Gaston, 2000 ;
Moles et al ., 2009 ;
Zhang et al ., 2013 ).而与经纬度和
MAP 对华北地区落叶松林最大胸径和最大树高的影响的一致性不同,
MAT 对华北地区落叶松林最大胸径与最大树高的影响正好相反(
图4C 、4G).我们的结果显示: 华北地区落叶松林最大胸径随
MAT 的增加而 降低, 随
MAP 的增加而增加(
图4C 、4G), 与
Yan等 (2006) 对鼎湖山亚热带常绿阔叶林优势乔木胸径的研究及
Tao等(2016) 对全球树高的研究结果一致.落叶松属植物为阳性物种, 作为林线植被, 气候变暖对落叶松的生长及群落更新可能是有利的(
Devi et al ., 2008 ).在全球气候变暖的背景下, 华北地区的落叶松林更趋向于垂直生长. ...
... ;
Moles et al ., 2009 ;
Zhang et al ., 2013 ).而与经纬度和
MAP 对华北地区落叶松林最大胸径和最大树高的影响的一致性不同,
MAT 对华北地区落叶松林最大胸径与最大树高的影响正好相反(
图4C 、4G).我们的结果显示: 华北地区落叶松林最大胸径随
MAT 的增加而 降低, 随
MAP 的增加而增加(
图4C 、4G), 与
Yan等 (2006) 对鼎湖山亚热带常绿阔叶林优势乔木胸径的研究及
Tao等(2016) 对全球树高的研究结果一致.落叶松属植物为阳性物种, 作为林线植被, 气候变暖对落叶松的生长及群落更新可能是有利的(
Devi et al ., 2008 ).在全球气候变暖的背景下, 华北地区的落叶松林更趋向于垂直生长. ...
... 从森林起源来看, 落叶松天然林和落叶松人工林的群落结构特征及物种多样性与地理及气候因子之间的关系, 与落叶松林总体的基本一致.其在地理因子上的差异主要体现在最大树高上, 落叶松人工林的最大树高随经纬度的增加而减少, 与前人在全球地理格局的研究结果一致(
Gaston, 2000 ;
Moles et al ., 2009 ;
Tao et al ., 2016 ).但是落叶松天然林的最大树高随经纬度的增加而增加, 与前人的研究结果(
Gaston, 2000 ;
Moles et al ., 2009 ;
Tao et al ., 2016 ) 相反, 其原因可能与海拔高度有关.华北地区低纬度和经度位置偏西的落叶松天然林主要是太白红杉林(
图1 ), 其分布海拔高, 树高相对低矮(
表5 ), 而高纬度位置及经度位置偏东的落叶松天然林为华北落叶松林, 其分布海拔较太白红杉林低(
郑万均和傅立国, 1978 ), 树高更高(
表5 ), 即华北地区落叶松林与地理因素的关系主要由落叶松人工林所主导.落叶松人工林和落叶松天然林在气候因子上的差异体现在最大胸径与
MAT 以及最大树高与
MAP 的关系上(
图4C 、4H).落叶松天然林的最大胸径和最大树高分别随
MAT 和
MAP 的增大而减小, 而落叶松人工林的最大胸径和最大树高分别随
MAT 和
MAP 的增大而增大.落叶松林总体最大胸径与气候条件的关系与落叶松天然林一致, 最大树高与气候条件的关系与落叶松人工林一致, 而对比落叶松天然林和落叶松人工林优势种的胸径树高, 落叶松天然林优势种的胸径显著大于人工林, 而树高显著小于人工林(
表5 ), 即华北地区落叶松林与
MAT 的关系主要受落叶松天然林的影响, 而与
MAP 的关系主要受落叶松人工林的影响. ...
... ;
Moles et al ., 2009 ;
Tao et al ., 2016 ) 相反, 其原因可能与海拔高度有关.华北地区低纬度和经度位置偏西的落叶松天然林主要是太白红杉林(
图1 ), 其分布海拔高, 树高相对低矮(
表5 ), 而高纬度位置及经度位置偏东的落叶松天然林为华北落叶松林, 其分布海拔较太白红杉林低(
郑万均和傅立国, 1978 ), 树高更高(
表5 ), 即华北地区落叶松林与地理因素的关系主要由落叶松人工林所主导.落叶松人工林和落叶松天然林在气候因子上的差异体现在最大胸径与
MAT 以及最大树高与
MAP 的关系上(
图4C 、4H).落叶松天然林的最大胸径和最大树高分别随
MAT 和
MAP 的增大而减小, 而落叶松人工林的最大胸径和最大树高分别随
MAT 和
MAP 的增大而增大.落叶松林总体最大胸径与气候条件的关系与落叶松天然林一致, 最大树高与气候条件的关系与落叶松人工林一致, 而对比落叶松天然林和落叶松人工林优势种的胸径树高, 落叶松天然林优势种的胸径显著大于人工林, 而树高显著小于人工林(
表5 ), 即华北地区落叶松林与
MAT 的关系主要受落叶松天然林的影响, 而与
MAP 的关系主要受落叶松人工林的影响. ...
日本落叶松人工林生物量和生产力的研究
1
1995
... 落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要.前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面.而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面.而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道. ...
日本落叶松人工林生物量和生产力的研究
1
1995
... 落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要.前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面.而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面.而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道. ...
Global patterns and determinants of forest canopy height
5
2016
... 华北地区落叶松林的群落结构特征及物种多样性与其分布的地理位置和气候条件紧密相关.总体而言, 除最大树高与经度的关系不显著以外, 华北地区落叶松林最大胸径和最大树高及总体的物种丰富度均随着经纬度的增加而显著降低(
图4A 、4D、 4E、4F、4I、4J), 这与前人在全球地理格局的研究结果一致(
Gaston, 2000 ;
Moles et al ., 2009 ;
Tao et al ., 2016 ).此外, 诸多的研究表明, 水分条件是乔木胸径和高度及植物群落物种丰富度的重要影响因子(
Gaston, 2000 ;
Zhang et al ., 2013 ;
Tao et al ., 2016 ).在华北地区, 落叶松林最大胸径与最大树高及其总体的物种丰富度随着
MAP 的增加而增加(
图4D 、4H、4L), 与前人的研究结果相符(
Gaston, 2000 ;
Moles et al ., 2009 ;
Zhang et al ., 2013 ).而与经纬度和
MAP 对华北地区落叶松林最大胸径和最大树高的影响的一致性不同,
MAT 对华北地区落叶松林最大胸径与最大树高的影响正好相反(
图4C 、4G).我们的结果显示: 华北地区落叶松林最大胸径随
MAT 的增加而 降低, 随
MAP 的增加而增加(
图4C 、4G), 与
Yan等 (2006) 对鼎湖山亚热带常绿阔叶林优势乔木胸径的研究及
Tao等(2016) 对全球树高的研究结果一致.落叶松属植物为阳性物种, 作为林线植被, 气候变暖对落叶松的生长及群落更新可能是有利的(
Devi et al ., 2008 ).在全球气候变暖的背景下, 华北地区的落叶松林更趋向于垂直生长. ...
... ;
Tao et al ., 2016 ).在华北地区, 落叶松林最大胸径与最大树高及其总体的物种丰富度随着
MAP 的增加而增加(
图4D 、4H、4L), 与前人的研究结果相符(
Gaston, 2000 ;
Moles et al ., 2009 ;
Zhang et al ., 2013 ).而与经纬度和
MAP 对华北地区落叶松林最大胸径和最大树高的影响的一致性不同,
MAT 对华北地区落叶松林最大胸径与最大树高的影响正好相反(
图4C 、4G).我们的结果显示: 华北地区落叶松林最大胸径随
MAT 的增加而 降低, 随
MAP 的增加而增加(
图4C 、4G), 与
Yan等 (2006) 对鼎湖山亚热带常绿阔叶林优势乔木胸径的研究及
Tao等(2016) 对全球树高的研究结果一致.落叶松属植物为阳性物种, 作为林线植被, 气候变暖对落叶松的生长及群落更新可能是有利的(
Devi et al ., 2008 ).在全球气候变暖的背景下, 华北地区的落叶松林更趋向于垂直生长. ...
... 对鼎湖山亚热带常绿阔叶林优势乔木胸径的研究及
Tao等(2016) 对全球树高的研究结果一致.落叶松属植物为阳性物种, 作为林线植被, 气候变暖对落叶松的生长及群落更新可能是有利的(
Devi et al ., 2008 ).在全球气候变暖的背景下, 华北地区的落叶松林更趋向于垂直生长. ...
... 从森林起源来看, 落叶松天然林和落叶松人工林的群落结构特征及物种多样性与地理及气候因子之间的关系, 与落叶松林总体的基本一致.其在地理因子上的差异主要体现在最大树高上, 落叶松人工林的最大树高随经纬度的增加而减少, 与前人在全球地理格局的研究结果一致(
Gaston, 2000 ;
Moles et al ., 2009 ;
Tao et al ., 2016 ).但是落叶松天然林的最大树高随经纬度的增加而增加, 与前人的研究结果(
Gaston, 2000 ;
Moles et al ., 2009 ;
Tao et al ., 2016 ) 相反, 其原因可能与海拔高度有关.华北地区低纬度和经度位置偏西的落叶松天然林主要是太白红杉林(
图1 ), 其分布海拔高, 树高相对低矮(
表5 ), 而高纬度位置及经度位置偏东的落叶松天然林为华北落叶松林, 其分布海拔较太白红杉林低(
郑万均和傅立国, 1978 ), 树高更高(
表5 ), 即华北地区落叶松林与地理因素的关系主要由落叶松人工林所主导.落叶松人工林和落叶松天然林在气候因子上的差异体现在最大胸径与
MAT 以及最大树高与
MAP 的关系上(
图4C 、4H).落叶松天然林的最大胸径和最大树高分别随
MAT 和
MAP 的增大而减小, 而落叶松人工林的最大胸径和最大树高分别随
MAT 和
MAP 的增大而增大.落叶松林总体最大胸径与气候条件的关系与落叶松天然林一致, 最大树高与气候条件的关系与落叶松人工林一致, 而对比落叶松天然林和落叶松人工林优势种的胸径树高, 落叶松天然林优势种的胸径显著大于人工林, 而树高显著小于人工林(
表5 ), 即华北地区落叶松林与
MAT 的关系主要受落叶松天然林的影响, 而与
MAP 的关系主要受落叶松人工林的影响. ...
... ;
Tao et al ., 2016 ) 相反, 其原因可能与海拔高度有关.华北地区低纬度和经度位置偏西的落叶松天然林主要是太白红杉林(
图1 ), 其分布海拔高, 树高相对低矮(
表5 ), 而高纬度位置及经度位置偏东的落叶松天然林为华北落叶松林, 其分布海拔较太白红杉林低(
郑万均和傅立国, 1978 ), 树高更高(
表5 ), 即华北地区落叶松林与地理因素的关系主要由落叶松人工林所主导.落叶松人工林和落叶松天然林在气候因子上的差异体现在最大胸径与
MAT 以及最大树高与
MAP 的关系上(
图4C 、4H).落叶松天然林的最大胸径和最大树高分别随
MAT 和
MAP 的增大而减小, 而落叶松人工林的最大胸径和最大树高分别随
MAT 和
MAP 的增大而增大.落叶松林总体最大胸径与气候条件的关系与落叶松天然林一致, 最大树高与气候条件的关系与落叶松人工林一致, 而对比落叶松天然林和落叶松人工林优势种的胸径树高, 落叶松天然林优势种的胸径显著大于人工林, 而树高显著小于人工林(
表5 ), 即华北地区落叶松林与
MAT 的关系主要受落叶松天然林的影响, 而与
MAP 的关系主要受落叶松人工林的影响. ...
太白红杉林林窗特征及其对群落更新的影响研究
1
2008
... 落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要.前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面.而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面.而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道. ...
太白红杉林林窗特征及其对群落更新的影响研究
1
2008
... 落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要.前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面.而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面.而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道. ...
Structural characteristics of old-growth hemlock-hardwood forests in relation to age
1
1994
... 植物群落结构可以反映出群落的整体状态.3种落叶松林胸径和树高频度分布的偏度系数大于0且峰度系数大于3 (
表5 ), 即3种落叶松林的胸径和树高结构总体均呈右偏分布且其径级大小和高度大小相对集中, 表明3种落叶松林均处于相对稳定的演替阶段(
Tyrrell & Crow, 1994 ;
Aiba & Kitayama, 1999 ;
Fang et al ., 2012 ).
匡旭等(2014) 对中国东北长白山落叶松林的研究结果显示, 长白落叶松(
L. olgensis )有较多
DBH < 10 cm的个体, 群落处于生长状态.
赵文智(1997) 对河北坝上华北落叶松人工林的研究表明, 30年华北落叶松成熟林林分稳定, 有天然更新的潜力.我们的结果与以上研究结果一致. ...
Relative importance of climate vs local factors in shaping the regional patterns of forest plant richness across northeast China
1
2009
... 物种丰富度是测度物种多样性的一个重要维度(
Gaston, 2000 ).华北地区, 3种落叶松林的物种丰富度总体比较丰富, 乔、灌、草层的物种丰富度有显著的差异(
图3 ;
表4 ).这与3种落叶松林的物种组成、气候条件以及乔木层郁闭度等因素有关(
Wang et al ., 2009 ;
Mölder et al ., 2014 ).具体而言, 太白红杉林为秦岭山地原始林(
雷明德, 1999 ), 华北落叶松林也多为人工林或次生林(
刘濂, 1996 ), 两种落叶松林优势种太白红杉及华北落叶松的重要值很高(
表3 ), 大部分甚至是纯林, 因而它们的乔木层的物种丰富度很低(
表4 ), 没有显著差异(
图3 A).而日本落叶松林虽然是人工林, 但其分布海拔低于华北落叶松林和太白红杉林, 拥有更好的水热条件, 群落物种更为丰富; 通常与其他物种混交, 其乔木层优势种日本落叶松的重要值只有70.2% (
表3 ), 因而其乔木层的物种丰富度显著大于华北落叶松林和太白红杉林(
图3 A).此外, 日本落叶松林与华北落叶松林均分布于中低海拔区域, 乔木层郁闭度大, 林下植物尤其是草本植物的生长受到影响, 其草本层丰富度相对较低, 没有显著差异(
图3 C).而太白红杉林位于秦岭山地高海拔区域, 乔木层郁闭度相对较小, 有利于林下植物的生长, 其林下草本层的物种丰富度显著大于华北落叶松林和日本落叶松林(
图3 C).但是, 也正是由于分布海拔高的影响, 其林下的生境条件较差, 所以对比华北地区纬度位置相差不大的日本落叶松林, 其灌木层物种丰富度没有显著优势(
图3 B). ...
Effects of geographical extent on the determinants of woody plant diversity
2
2012
... 前人基于大尺度的研究表明, 气候因子尤其是环境能量因子对物种丰富度的影响显著(
Wang et al ., 2012 ).但我们的结果似乎并不支持这一结论.本研究中,
MAT 对华北地区落叶松林的物种丰富度没有显著影响(
图4K ).这可能与研究对象的差异有关.
Wang等(2012) 的研究是基于木本植物物种丰富度的结果, 而本研究中的研究结果是基于落叶松林木本植物及草本植物总的物种丰富度的结果.由
表4 可知, 华北地区落叶松林总体的物种丰富度中草本植物所占的比重最大, 因而其对环境条件的响应与木本植物的不同. ...
... ).这可能与研究对象的差异有关.
Wang等(2012) 的研究是基于木本植物物种丰富度的结果, 而本研究中的研究结果是基于落叶松林木本植物及草本植物总的物种丰富度的结果.由
表4 可知, 华北地区落叶松林总体的物种丰富度中草本植物所占的比重最大, 因而其对环境条件的响应与木本植物的不同. ...
芦芽山阴坡华北落叶松-云杉天然次生林群落特征的海拔梯度格局
1
2010
... 落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要.前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面.而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面.而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道. ...
芦芽山阴坡华北落叶松-云杉天然次生林群落特征的海拔梯度格局
1
2010
... 落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要.前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面.而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面.而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道. ...
1
1980
... 落叶松属(
Larix )植物是中国北方广泛分布的寒温性针叶林的建群种, 也是重要的用材树种(
吴征镒, 1980 ;
李楠, 1995 ;
刘增力等, 2002 ).在我国华北地区, 落叶松属植物是很多山地的林线物种, 如太白山的林线物种为太白红杉(
Larix chinensis )(
段仁燕等, 2010 ), 五台山、关帝山及小五台山的林线物种为华北落叶松(
L. principis-rupprechtii )(
Liu et al ., 2002 ), 这些落叶松对维护高海拔地区的生态环境有重要意义.目前中国华北地区主要分布有3种落叶松: 华北落叶松、日本落叶松(
L. kaempferi )及太白红杉(
郑万均, 1983 ).其中, 华北落叶松及太白红杉是中国特有种, 日本落叶松为华北地区广泛栽培种 (
郑万均和傅立国, 1978 ).这3种落叶松是华北地区落叶松林的优势种.其中, 日本落叶松林为人工林; 而华北落叶松林虽然在华北地区有天然分布, 但是由于其分布的海拔较低, 受人为干扰严重, 因而目前纯原始林较少, 大部分是受干扰的次生林或者栽培的人工林(
刘濂, 1996 ); 相较而言, 太白红杉林虽仅分布于我国秦岭山地, 但其分布海拔较高, 受人为干扰影响较小, 且其适应性极强, 在土层贫瘠气候寒冷的条件下也能适应, 为我国秦岭山区森林上限植被, 目前主要是原始林(
雷明德, 1999 ;
应俊生和陈梦玲, 2011 ). ...
1
1980
... 落叶松属(
Larix )植物是中国北方广泛分布的寒温性针叶林的建群种, 也是重要的用材树种(
吴征镒, 1980 ;
李楠, 1995 ;
刘增力等, 2002 ).在我国华北地区, 落叶松属植物是很多山地的林线物种, 如太白山的林线物种为太白红杉(
Larix chinensis )(
段仁燕等, 2010 ), 五台山、关帝山及小五台山的林线物种为华北落叶松(
L. principis-rupprechtii )(
Liu et al ., 2002 ), 这些落叶松对维护高海拔地区的生态环境有重要意义.目前中国华北地区主要分布有3种落叶松: 华北落叶松、日本落叶松(
L. kaempferi )及太白红杉(
郑万均, 1983 ).其中, 华北落叶松及太白红杉是中国特有种, 日本落叶松为华北地区广泛栽培种 (
郑万均和傅立国, 1978 ).这3种落叶松是华北地区落叶松林的优势种.其中, 日本落叶松林为人工林; 而华北落叶松林虽然在华北地区有天然分布, 但是由于其分布的海拔较低, 受人为干扰严重, 因而目前纯原始林较少, 大部分是受干扰的次生林或者栽培的人工林(
刘濂, 1996 ); 相较而言, 太白红杉林虽仅分布于我国秦岭山地, 但其分布海拔较高, 受人为干扰影响较小, 且其适应性极强, 在土层贫瘠气候寒冷的条件下也能适应, 为我国秦岭山区森林上限植被, 目前主要是原始林(
雷明德, 1999 ;
应俊生和陈梦玲, 2011 ). ...
太白红杉林径级和龄级结构的研究
1
2005
... 落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要.前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面.而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面.而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道. ...
太白红杉林径级和龄级结构的研究
1
2005
... 落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要.前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面.而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面.而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道. ...
秦岭太白红杉林分布及太白山高山林线特征的定量分析
1
2005
... 落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要.前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面.而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面.而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道. ...
秦岭太白红杉林分布及太白山高山林线特征的定量分析
1
2005
... 落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要.前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面.而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面.而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道. ...
秦岭太白红杉群落特征及其物种多样性的研究
1
2001
... 落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要.前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面.而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面.而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道. ...
秦岭太白红杉群落特征及其物种多样性的研究
1
2001
... 落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要.前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面.而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面.而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道. ...
华北山地典型天然次生林群落的树种空间分布格局及其关联性
1
2010
... 落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要.前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面.而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面.而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道. ...
华北山地典型天然次生林群落的树种空间分布格局及其关联性
1
2010
... 落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要.前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面.而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面.而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道. ...
Different patterns of changes in the dry season diameter at breast height of dominant and evergreen tree species in a mature subtropical forest in South China
1
2006
... 华北地区落叶松林的群落结构特征及物种多样性与其分布的地理位置和气候条件紧密相关.总体而言, 除最大树高与经度的关系不显著以外, 华北地区落叶松林最大胸径和最大树高及总体的物种丰富度均随着经纬度的增加而显著降低(
图4A 、4D、 4E、4F、4I、4J), 这与前人在全球地理格局的研究结果一致(
Gaston, 2000 ;
Moles et al ., 2009 ;
Tao et al ., 2016 ).此外, 诸多的研究表明, 水分条件是乔木胸径和高度及植物群落物种丰富度的重要影响因子(
Gaston, 2000 ;
Zhang et al ., 2013 ;
Tao et al ., 2016 ).在华北地区, 落叶松林最大胸径与最大树高及其总体的物种丰富度随着
MAP 的增加而增加(
图4D 、4H、4L), 与前人的研究结果相符(
Gaston, 2000 ;
Moles et al ., 2009 ;
Zhang et al ., 2013 ).而与经纬度和
MAP 对华北地区落叶松林最大胸径和最大树高的影响的一致性不同,
MAT 对华北地区落叶松林最大胸径与最大树高的影响正好相反(
图4C 、4G).我们的结果显示: 华北地区落叶松林最大胸径随
MAT 的增加而 降低, 随
MAP 的增加而增加(
图4C 、4G), 与
Yan等 (2006) 对鼎湖山亚热带常绿阔叶林优势乔木胸径的研究及
Tao等(2016) 对全球树高的研究结果一致.落叶松属植物为阳性物种, 作为林线植被, 气候变暖对落叶松的生长及群落更新可能是有利的(
Devi et al ., 2008 ).在全球气候变暖的背景下, 华北地区的落叶松林更趋向于垂直生长. ...
1
2011
... 落叶松属(
Larix )植物是中国北方广泛分布的寒温性针叶林的建群种, 也是重要的用材树种(
吴征镒, 1980 ;
李楠, 1995 ;
刘增力等, 2002 ).在我国华北地区, 落叶松属植物是很多山地的林线物种, 如太白山的林线物种为太白红杉(
Larix chinensis )(
段仁燕等, 2010 ), 五台山、关帝山及小五台山的林线物种为华北落叶松(
L. principis-rupprechtii )(
Liu et al ., 2002 ), 这些落叶松对维护高海拔地区的生态环境有重要意义.目前中国华北地区主要分布有3种落叶松: 华北落叶松、日本落叶松(
L. kaempferi )及太白红杉(
郑万均, 1983 ).其中, 华北落叶松及太白红杉是中国特有种, 日本落叶松为华北地区广泛栽培种 (
郑万均和傅立国, 1978 ).这3种落叶松是华北地区落叶松林的优势种.其中, 日本落叶松林为人工林; 而华北落叶松林虽然在华北地区有天然分布, 但是由于其分布的海拔较低, 受人为干扰严重, 因而目前纯原始林较少, 大部分是受干扰的次生林或者栽培的人工林(
刘濂, 1996 ); 相较而言, 太白红杉林虽仅分布于我国秦岭山地, 但其分布海拔较高, 受人为干扰影响较小, 且其适应性极强, 在土层贫瘠气候寒冷的条件下也能适应, 为我国秦岭山区森林上限植被, 目前主要是原始林(
雷明德, 1999 ;
应俊生和陈梦玲, 2011 ). ...
1
2011
... 落叶松属(
Larix )植物是中国北方广泛分布的寒温性针叶林的建群种, 也是重要的用材树种(
吴征镒, 1980 ;
李楠, 1995 ;
刘增力等, 2002 ).在我国华北地区, 落叶松属植物是很多山地的林线物种, 如太白山的林线物种为太白红杉(
Larix chinensis )(
段仁燕等, 2010 ), 五台山、关帝山及小五台山的林线物种为华北落叶松(
L. principis-rupprechtii )(
Liu et al ., 2002 ), 这些落叶松对维护高海拔地区的生态环境有重要意义.目前中国华北地区主要分布有3种落叶松: 华北落叶松、日本落叶松(
L. kaempferi )及太白红杉(
郑万均, 1983 ).其中, 华北落叶松及太白红杉是中国特有种, 日本落叶松为华北地区广泛栽培种 (
郑万均和傅立国, 1978 ).这3种落叶松是华北地区落叶松林的优势种.其中, 日本落叶松林为人工林; 而华北落叶松林虽然在华北地区有天然分布, 但是由于其分布的海拔较低, 受人为干扰严重, 因而目前纯原始林较少, 大部分是受干扰的次生林或者栽培的人工林(
刘濂, 1996 ); 相较而言, 太白红杉林虽仅分布于我国秦岭山地, 但其分布海拔较高, 受人为干扰影响较小, 且其适应性极强, 在土层贫瘠气候寒冷的条件下也能适应, 为我国秦岭山区森林上限植被, 目前主要是原始林(
雷明德, 1999 ;
应俊生和陈梦玲, 2011 ). ...
Structure and composition of natural gmelin larch (
Larix gmelinii var.
gmelinii ) forests in response to spatial climatic changes
2
2013
... 华北地区落叶松林的群落结构特征及物种多样性与其分布的地理位置和气候条件紧密相关.总体而言, 除最大树高与经度的关系不显著以外, 华北地区落叶松林最大胸径和最大树高及总体的物种丰富度均随着经纬度的增加而显著降低(
图4A 、4D、 4E、4F、4I、4J), 这与前人在全球地理格局的研究结果一致(
Gaston, 2000 ;
Moles et al ., 2009 ;
Tao et al ., 2016 ).此外, 诸多的研究表明, 水分条件是乔木胸径和高度及植物群落物种丰富度的重要影响因子(
Gaston, 2000 ;
Zhang et al ., 2013 ;
Tao et al ., 2016 ).在华北地区, 落叶松林最大胸径与最大树高及其总体的物种丰富度随着
MAP 的增加而增加(
图4D 、4H、4L), 与前人的研究结果相符(
Gaston, 2000 ;
Moles et al ., 2009 ;
Zhang et al ., 2013 ).而与经纬度和
MAP 对华北地区落叶松林最大胸径和最大树高的影响的一致性不同,
MAT 对华北地区落叶松林最大胸径与最大树高的影响正好相反(
图4C 、4G).我们的结果显示: 华北地区落叶松林最大胸径随
MAT 的增加而 降低, 随
MAP 的增加而增加(
图4C 、4G), 与
Yan等 (2006) 对鼎湖山亚热带常绿阔叶林优势乔木胸径的研究及
Tao等(2016) 对全球树高的研究结果一致.落叶松属植物为阳性物种, 作为林线植被, 气候变暖对落叶松的生长及群落更新可能是有利的(
Devi et al ., 2008 ).在全球气候变暖的背景下, 华北地区的落叶松林更趋向于垂直生长. ...
... ;
Zhang et al ., 2013 ).而与经纬度和
MAP 对华北地区落叶松林最大胸径和最大树高的影响的一致性不同,
MAT 对华北地区落叶松林最大胸径与最大树高的影响正好相反(
图4C 、4G).我们的结果显示: 华北地区落叶松林最大胸径随
MAT 的增加而 降低, 随
MAP 的增加而增加(
图4C 、4G), 与
Yan等 (2006) 对鼎湖山亚热带常绿阔叶林优势乔木胸径的研究及
Tao等(2016) 对全球树高的研究结果一致.落叶松属植物为阳性物种, 作为林线植被, 气候变暖对落叶松的生长及群落更新可能是有利的(
Devi et al ., 2008 ).在全球气候变暖的背景下, 华北地区的落叶松林更趋向于垂直生长. ...
东灵山华北落叶松人工林的分布格局及环境解释
1
2010
... 落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要.前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面.而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面.而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道. ...
东灵山华北落叶松人工林的分布格局及环境解释
1
2010
... 落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要.前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面.而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面.而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道. ...
日本落叶松人工林生物量及生产力的研究
1
1999
... 落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要.前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面.而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面.而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道. ...
日本落叶松人工林生物量及生产力的研究
1
1999
... 落叶松林作为我国华北地区重要的用材林及水土涵养林, 探讨其整体的分布规律及其群落结构和物种多样性的基础研究十分有必要.前人关于华北地区落叶松林的研究较为分散, 对于华北落叶松和日本落叶松林多集中在研究林分结构、林木生长和物种多样性(
韩艳英, 2006 ;
马友平, 2007 ;
武秀娟等; 2010 ;
孔令伟等, 2014 ), 生物量、生产力及碳密度(
宿以明, 1995 ;
赵体顺等, 1999 ;
罗云建等, 2009 ;
陈东升等, 2016 ;
纪文婧等, 2016 ;
马丰丰等, 2016 )以及空间分布格局(
闫海冰等, 2010 ;
张钦弟等, 2010 )等方面.而对太白红杉林的研究主要集中在群落分布及空间格局(
郭华, 2004 ;
许林军等, 2005b ), 群落结构特征及物种多样性(
阎桂琴等, 2001 ;
许林军等, 2005a )以及群落更新(
涂云博, 2008 )等方面.而立足于区域性的, 对中国华北地区落叶松林整体的研究鲜有报道. ...
河北坝上半干旱/半湿润过渡带华北落叶松人工林研究
1
1997
... 植物群落结构可以反映出群落的整体状态.3种落叶松林胸径和树高频度分布的偏度系数大于0且峰度系数大于3 (
表5 ), 即3种落叶松林的胸径和树高结构总体均呈右偏分布且其径级大小和高度大小相对集中, 表明3种落叶松林均处于相对稳定的演替阶段(
Tyrrell & Crow, 1994 ;
Aiba & Kitayama, 1999 ;
Fang et al ., 2012 ).
匡旭等(2014) 对中国东北长白山落叶松林的研究结果显示, 长白落叶松(
L. olgensis )有较多
DBH < 10 cm的个体, 群落处于生长状态.
赵文智(1997) 对河北坝上华北落叶松人工林的研究表明, 30年华北落叶松成熟林林分稳定, 有天然更新的潜力.我们的结果与以上研究结果一致. ...
河北坝上半干旱/半湿润过渡带华北落叶松人工林研究
1
1997
... 植物群落结构可以反映出群落的整体状态.3种落叶松林胸径和树高频度分布的偏度系数大于0且峰度系数大于3 (
表5 ), 即3种落叶松林的胸径和树高结构总体均呈右偏分布且其径级大小和高度大小相对集中, 表明3种落叶松林均处于相对稳定的演替阶段(
Tyrrell & Crow, 1994 ;
Aiba & Kitayama, 1999 ;
Fang et al ., 2012 ).
匡旭等(2014) 对中国东北长白山落叶松林的研究结果显示, 长白落叶松(
L. olgensis )有较多
DBH < 10 cm的个体, 群落处于生长状态.
赵文智(1997) 对河北坝上华北落叶松人工林的研究表明, 30年华北落叶松成熟林林分稳定, 有天然更新的潜力.我们的结果与以上研究结果一致. ...
 ,1,*
,1,*
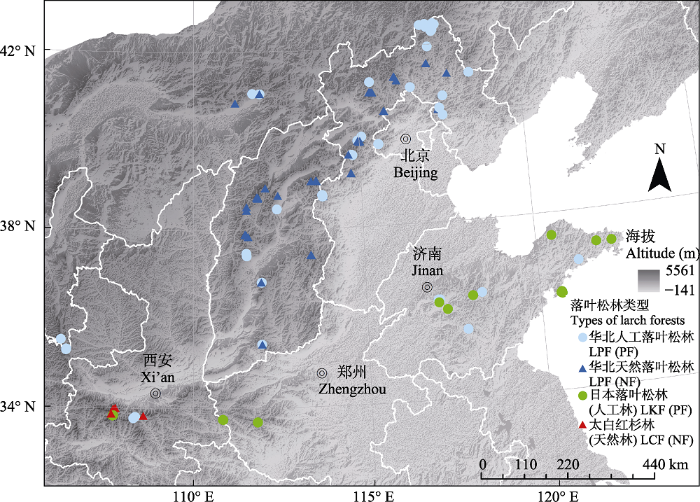 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT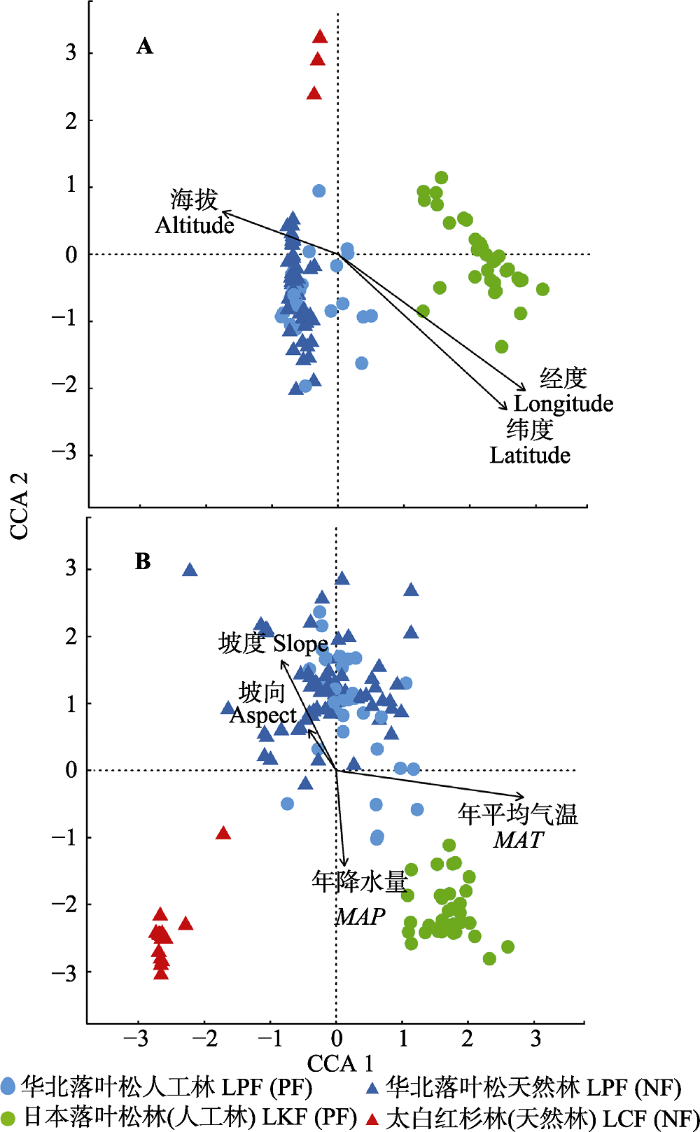 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT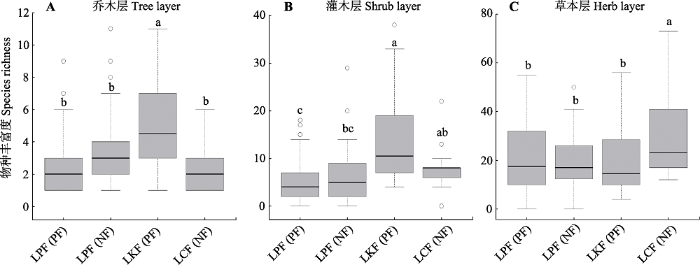 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT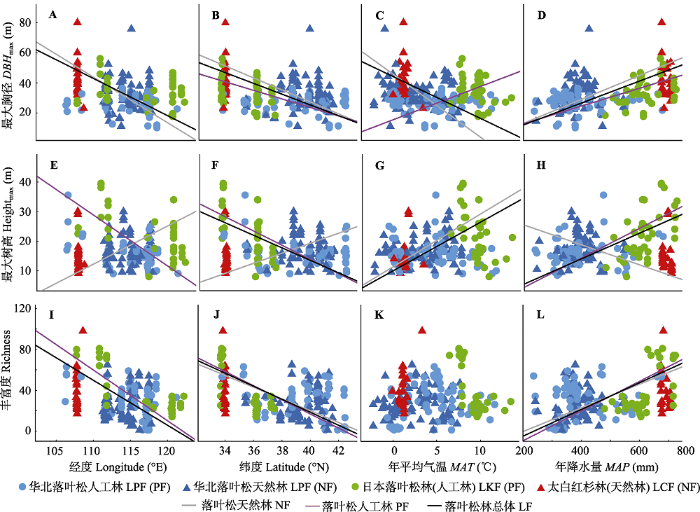 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}