Spatial distribution patterns and correlation of Tamarix chinensis population in coastal wetlands of Shandong, China
Pan WU,, Xi-Qiang PENG, Shu-Ren YANG, Ya-Nan GAO, Feng-Hua BAI, Shi-Jie YI, Ning DU,*, Wei-Hua GUO,*Institute of Ecology and Biodiversity, School of Life Sciences, Shandong University, Qingdao, Shandong 266237, China
Supported by the National Basic Work of Science and Technology of China(2011FY110300) and the National Natural Science Foundation of China(31470402) and the National Natural Science Foundation of China(31770361)
Abstract Aims Tamarix chinensis is a pioneering shrub species in temperate coastal saline wetlands, which plays an important role in plant community succession and preventing seawater intrusion in coastal wetlands. This study, which is focused on Tamarix chinensis population distribution characteristics and the correlations between Tamarix chinensis population distribution with individual diameters, can reveal the characteristics of population development and provide reference for species conservation and management of protected areas. This study may also provide basic information for scientific research on the succession and ecological management of coastal wetland vegetation ecosystems. Methods In the core area of Changyi National Marine Ecological Special Protection Zone, two sample tapes with a spacing of about 800 m are arranged along the parallel coastline. Each sample tape has three plots of 50 m × 50 m. A total of six sample plots are set up for each wood survey. The spatial distribution map of Tamarix chinensis population is drawn by Origin. The obtained data is divided into three different diameter grades according to their base diameter: diameter grade I (base diameter ≤ 4 cm) and diameter grade II (4 < base diameter ≤ 8 cm), diameter grade III (base diameter > 8 cm). The distribution pattern of Tamarix chinensis population and the relationship between different diameter grades are analyzed by the point pattern analysis method and Programita software. Important findings (1) A total of 374 individuals of Tamarix chinensis are investigated in six plots, including 14 in plot 1, 20 in plot 2, 36 in plot 3, 45 in plot 4, 221 in plot 5, and 38 in plot 6. (2) There is a large difference in population density of Tamarix chinensis between different plots, which indicates that Tamarix chinensis is not evenly distributed on the regional scale. (3) Tamarix chinensis populations show a clustered distribution on small scale (less than 5 m) while they appear as random distribution on large scale (greater than 15 m). The population of Tamarix chinensis exhibits a trend of transition from clustered distribution to random distribution with the increase of spatial scale. (4) The spatial association of Tamarix chinensis between any two diameter grades is positive on small scale and there is no significant spatial association between them on large scale. However, negative spatial association on diameter grade II and diameter grade III is detected at the scale of 15 m probably due to space competition. Keywords:Tamarix chinensis population;distribution pattern;spatial scale;diameter structure;spatial association
PDF (1531KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 吴盼, 彭希强, 杨树仁, 高亚男, 白丰桦, 衣世杰, 杜宁, 郭卫华. 山东省滨海湿地柽柳种群的空间分布格局及其关联性. 植物生态学报, 2019, 43(9): 817-824. DOI: 10.17521/cjpe.2018.0186 WU Pan, PENG Xi-Qiang, YANG Shu-Ren, GAO Ya-Nan, BAI Feng-Hua, YI Shi-Jie, DU Ning, GUO Wei-Hua. Spatial distribution patterns and correlation of Tamarix chinensis population in coastal wetlands of Shandong, China. Chinese Journal of Plant Ecology, 2019, 43(9): 817-824. DOI: 10.17521/cjpe.2018.0186
Fig. 2Scatterplot of spatial distribution of Tamarix chinensis in coastal wetlands of Shandong. A, Plot 1. B, Plot 2. C, Plot 3. D, Plot 4. E, Plot 5. F, Plot 6.
Table 1 表1 表1山东滨海湿地柽柳种群各样地基本信息及不同径级柽柳组成 Table 1Basic information and compositions of Tamarix chinensis at different diameter grades in each plot in coastal wetlands of Shandong
Fig. 3Point pattern analysis of Tamarix chinensis in coastal wetlands of Shandong. A, B, C, D, E and F represent plot 1, 2, 3, 4, 5 and 6, respectively. O(r), univariate O-ring function.
Fig. 4Spatial association of Tamarix chinensis at different diameter grades in coastal wetlands of Shandong. A, Diameter grade I and diameter grade II. B, Diameter grade I and diameter grade III. C, Diameter grade II and diameter grade III. O12(r), bivariate O-ring function.
柽柳种群不同径级的空间关联性从总体来讲, 径级I、II与III两两之间在小尺度上表现为正关联, 在大尺度上则以相互独立的空间关系为主。径级I与径级II及径级I与径级III在较小尺度上表现为正关联, 是由于植株对环境资源的需求具有相似性, 但基径小的植株个体相对幼小, 对水分、土壤养分等资源的竞争能力相对薄弱, 需要依附相对高大植株的庇护以提高存活几率(尤海舟等, 2010)。在本区域的前期研究中也发现, 柽柳的存在可以对其下生长的草本植物产生促进作用(Gao et al., 2015; Du et al., 2017)。在对黄河三角洲滨海湿地的研究中还发现, 柽柳的生长可以增加土壤有机质含量, 提高土壤氮、磷、钾的含量, 从而形成“沃岛” (封晓辉等, 2013; Rong et al., 2016)。
BaiC, YanM, BiRC, HeYH ( 2014). Spatial pattern analysis of dominant species in Exochorda giraldii community in Xingtang Temple of Taiyue Mountains, Shanxi, China Chinese Journal of Plant Ecology, 38, 1283-1295. [本文引用: 1]
BiXL, WenXH, YiHP, WuXQ, GaoM ( 2014). Succession in soil and vegetation caused by coastal embankment in southern Laizhou Bay, China—Flourish or degradation? Ocean & Coastal Management, 88, 1-7. [本文引用: 1]
Ca?adorI, TibérioS, CabralHN ( 2007). Species zonation in Corroios salt marsh in the Tagus estuary (Portugal) and its dynamics in the past fifty years Hydrobiologia, 587, 205-211. [本文引用: 1]
CuiBS, YangQC, ZhangKJ, ZhaoXS, YouZY ( 2010). Responses of saltcedar ( Tamarix chinensis) to water table depth and soil salinity in the Yellow River Delta, China Plant Ecology, 209, 279-290. [本文引用: 1]
DigglePJ ( 1983). Statistical Analysis of Spatial Point Patterns. Academic Press, New York. [本文引用: 1]
DuN, WuP, EllerF, ZhouDY, LiuJ, GanWH, YangRR, DaiM, ChenYD, WangRQ, GuoWH ( 2017). Facilitation or competition? The effects of the shrub species Tamarix chinensis on herbaceous communities are dependent on the successional stage in an impacted coastal wetland of north China Wetlands, 37, 899-911. [本文引用: 3]
FengXH, ZhangXM, LiuXJ, ChengRM, SunHR ( 2013). Growth dynamics of Tamarix chinensis plantations in heavy-saline coastal lands and related ecological effects Chinese Journal of Eco-Agriculture, 21, 1233-1240. [本文引用: 1]
GaoM, WangXX, HuiC, YiHP, ZhangCQ, WuXQ, BiXL, WangY, XiaoLX, WangD ( 2015). Assembly of plant communities in coastal wetlands-the role of saltcedar Tamarix chinensis during early succession Journal of Plant Ecology, 8, 539-548. [本文引用: 2]
GuoYL, WangB, XiangWS, DingT, LuSH, HuangYS, HuangFZ, LiDX, LiXK ( 2015). Spatial distribution of tree species in a tropical karst seasonal rainforest in Nonggang, Guangxi, southern China Biodiversity Science, 23, 183-191. [本文引用: 1]
HaoHM, HuangZ, LuR, JiaC, LiuY, LiuBR, WuGL ( 2017). Patches structure succession based on spatial point pattern features in semi-arid ecosystems of the water-wind erosion crisscross region Global Ecology and Conservation, 12, 158-165. [本文引用: 1]
JafariM, ChahoukiMAZ, TaviliA, AzarnivandH, AmiriGZ ( 2004). Effective environmental factors in the distribution of vegetation types in Poshtkouh rangelands of Yazd Province (Iran) Journal of Arid Environments, 56, 627-641. [本文引用: 1]
JiangZM, ChenYX, BaoY ( 2012). Population genetic structure of Tamarix chinensis in the Yellow River Delta, China Plant Systematics and Evolution, 298, 147-153. [本文引用: 1]
KangJP, MaYY, MaSQ, XueZW, YangLL, HanL, LiuWY ( 2019). Dynamic changes of spatial pattern and structure of the Tamarix ramosissima population at the desert-oasis ecotone of the Tarim Basin Acta Ecologica Sinica, 39, 265-276. [本文引用: 2]
LeiboldMA, McPeekMA ( 2006). Coexistence of the niche and neutral perspectives in community ecology Ecology, 87, 1399-1410. [本文引用: 1]
LiHD, ShenWS, FangY, YanSG, ZhangH, ZhaoW ( 2011). Point pattern analysis of several psammophyte populations in the riparian ecotone in the middle reaches of Yarlung Zangbo River of Tibet, China Chinese Journal of Plant Ecology, 35, 834-843. [本文引用: 2]
LiuJH, XiaJB, FangYM, LiT, LiuJT ( 2014). Effects of salt-drought stress on growth and physiobiochemical characteristics of Tamarix chinensis seedlings The Scientific World Journal, 765840. DOI: . URL [本文引用: 1]
McGillBJ ( 2003). A test of the unified neutral theory of biodiversity Nature, 422, 881-885. [本文引用: 1]
MeiJL, ZhuangFH, MaJM, QinYH, LiangSC, JiangY ( 2017). Spatial point pattern analysis of Alchornea trewioides population clonal growth in the karst area of Guilin Acta Ecologica Sinica, 37, 3164-3171. [本文引用: 2]
PerryGL, MillerBP, EnrightNJ, LamontBB ( 2014). Stochastic geometry best explains spatial associations among species pairs and plant functional types in species-rich shrublands Oikos, 123, 99-110. [本文引用: 1]
RipleyBD ( 1977). Modelling spatial patterns Journal of the Royal Statistical Society: Series B, 39, 172-192. [本文引用: 1]
RongQQ, LiuJT, CaiYP, LuZH, ZhaoZZ, YueWC, XiaJB ( 2016). “Fertile island” effects of Tamarix chinensis Lour. on soil N and P stoichiometry in the coastal wetland of Laizhou Bay, China Journal of Soils & Sediments, 16, 864-877. [本文引用: 2]
SongXJ, LiSN, GuoJ, YuYL, LiuZW, WeiW ( 2018). Effects of different salinity levels on the growth and physiological characteristics of roots of Tamarix chinensis cuttings. Acta Ecologica Sinica, 38, 606-614. [本文引用: 1]
SunLK, LiuWQ, LiuGX, ChenT, ZhangW, WuXK, ZhangGS, ZhangYH, LiL, ZhangBG, ZhangBL, WangB, YangRQ ( 2016). Temporal and spatial variations in the stable carbon isotope composition and carbon and nitrogen contents in current-season twigs of Tamarix chinensis Lour. and their relationships to environmental factors in the Laizhou Bay wetland in China Ecological Engineering, 90, 417-426. [本文引用: 1]
TangAK, LiuRH, XuLQ, WangJY, LiuYT ( 2011). Spatial heterogeneity of soil nutrients and distribution of plant community in Changyi Marine Ecological Special Protection Area Bulletin of Soil and Water Conservation, 31, 88-93. [本文引用: 2]
WangQD, SongJM, LiXG, YuanHM, LiN, CaoL ( 2015). Environmental radionuclides in a coastal wetland of the southern Laizhou Bay, China Marine Pollution Bulletin, 97, 506-511. [本文引用: 1]
WangXT, HouYL, LiuF, ChangY, WangW, LiangCZ, MiaoBL ( 2011). Point pattern analysis of dominant populations in a degraded community in Leymus chinensis + Stipa grandis steppe in Inner Mongolia, China Chinese Journal of Plant Ecology, 35, 1281-1289. [本文引用: 2]
WiegandT, MoloneyKA ( 2004). Rings, circles, and null- models for point pattern analysis in ecology Oikos, 104, 209-229. [本文引用: 1]
WuXQ, GaoM, WangD, WangY, LuQS, ZhangZD ( 2012). Framework and practice of integrated coastal zone management in Shandong Province, China Ocean & Coastal Management, 69, 58-67. [本文引用: 1]
XiaJB, ZhaoXM, LiuJH, ZhaoZG, LiuQ, ChenYP ( 2016). Environmental factors influencing the distribution of Tamarix chinense Lour. in the Laizhou Bay wetland of the Yellow River Delta Acta Ecologica Sinica, 36, 4801-4808. [本文引用: 1]
XieLP, WangM, WangBD, ShiXY, XinM, WeiQS, HeXP, GuoF ( 2017). Distribution pattern and influencing factors of vegetation carbon storage of Tamarix chinense in the coastal wetland of Laizhou Bay, China Chinese Journal of Applied Ecology, 28, 1103-1111. [本文引用: 3]
XuQ, LüJZ, MiaoYM, BiRC ( 2016). Spatial distribution patterns and association of major species in Elaeagnus mollis communities Chinese Bulletin of Botany, 51, 49-57. [本文引用: 2]
YangH, LiYL, ShenL, KangXG, YueG, WangY ( 2014). Spatial distribution patterns of seedling and sapling in a spruce-fir forest in the Changbai Mountains area in northeastern China Acta Ecologica Sinica, 34, 7311-7319. [本文引用: 1]
YouHZ, LiuXL, MiaoN, HeF, MaQY ( 2010). Individual association and scale effect of spatial pattern of Quercus aquifolioides populations along the elevation gradients Acta Ecologica Sinica, 30, 4004-4011. [本文引用: 1]
ZhangJT, MengDP ( 2004). Spatial pattern analysis of individuals in different age-classes of Larix principis??rupprechtii in Luya Mountain Reserve, Shanxi, China Acta Ecologica Sinica, 24, 35-40. [本文引用: 1]
ZhaoXS, CuiBS, SunT, LvJZ, LuF ( 2011). Analysis of spatial point pattern of Tamarix chinensis in different habitats Ecological Science, 30, 142-149. [本文引用: 1]
ZhuZ, ZhangLY, GaoLX, TangSQ, ZhaoY, YangJ ( 2016). Local habitat condition rather than geographic distance determines the genetic structure of Tamarix chinensis populations in Yellow River Delta, China Tree Genetics & Genomes, 12. DOI: 10.1007/S11295-016-0971-5. [本文引用: 1]
Responses of saltcedar ( Tamarix chinensis) to water table depth and soil salinity in the Yellow River Delta, China 1 2010
... 柽柳(Tamarix chinensis)是温带滨海盐碱湿地的先锋灌木物种, 主要分布在中国东部和东北部的河流三角洲和盐碱湿地等条件苛刻的生境中, 在防止沿海地区海水入侵中发挥着重要作用(Cui et al., 2010).昌邑国家海洋生态特别保护区地处渤海莱州湾南岸, 是全国唯一的以柽柳群落为主要保护和管理对象的国家级海洋特别保护区, 是研究柽柳个体、种群、群落等不同层次特征的理想区域, 也是我国“南红北柳”生态工程的重要组成部分(谢琳萍等, 2017).目前对于滨海盐碱湿地柽柳的研究多集中于个体层面的柽柳叶片生理生化特征(朱金方等, 2013; Liu et al., 2014), 根系生理特性(宋香静等, 2018), 种群层面的柽柳种群空间分布特征(Jiang et al., 2012; Zhu et al., 2016), 群落层面的柽柳群落的土壤环境特征(Rong et al., 2016; Sun et al., 2016)和柽柳对草本植物生物多样性的影响(Gao et al., 2015; Du et al., 2017)等方面, 但对于柽柳群落中不同径级柽柳的分布格局及不同径级之间关联性的研究鲜有报道.研究滨海湿地先锋物种柽柳种群的空间分布格局及其不同径级之间的关联性, 有助于深化对滨海湿地植物种群组成、结构与动态的认识, 揭示种群发展规律, 为保育柽柳提供理论依据, 并进一步为湿地生态系统的管理提供基础资料. ...
Facilitation or competition? The effects of the shrub species Tamarix chinensis on herbaceous communities are dependent on the successional stage in an impacted coastal wetland of north China 3 2017
... 柽柳(Tamarix chinensis)是温带滨海盐碱湿地的先锋灌木物种, 主要分布在中国东部和东北部的河流三角洲和盐碱湿地等条件苛刻的生境中, 在防止沿海地区海水入侵中发挥着重要作用(Cui et al., 2010).昌邑国家海洋生态特别保护区地处渤海莱州湾南岸, 是全国唯一的以柽柳群落为主要保护和管理对象的国家级海洋特别保护区, 是研究柽柳个体、种群、群落等不同层次特征的理想区域, 也是我国“南红北柳”生态工程的重要组成部分(谢琳萍等, 2017).目前对于滨海盐碱湿地柽柳的研究多集中于个体层面的柽柳叶片生理生化特征(朱金方等, 2013; Liu et al., 2014), 根系生理特性(宋香静等, 2018), 种群层面的柽柳种群空间分布特征(Jiang et al., 2012; Zhu et al., 2016), 群落层面的柽柳群落的土壤环境特征(Rong et al., 2016; Sun et al., 2016)和柽柳对草本植物生物多样性的影响(Gao et al., 2015; Du et al., 2017)等方面, 但对于柽柳群落中不同径级柽柳的分布格局及不同径级之间关联性的研究鲜有报道.研究滨海湿地先锋物种柽柳种群的空间分布格局及其不同径级之间的关联性, 有助于深化对滨海湿地植物种群组成、结构与动态的认识, 揭示种群发展规律, 为保育柽柳提供理论依据, 并进一步为湿地生态系统的管理提供基础资料. ...
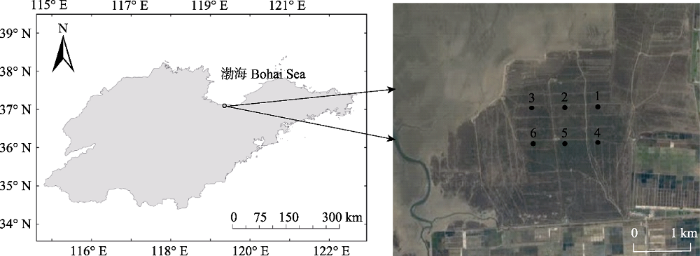
... 研究区位于山东省昌邑市北部堤河以东、海岸线以下的滩涂上(37.05°-37.12° N, 119.33°-119.38° E), 总面积为2 929.28 hm2, 其中天然柽柳林面积达2 070 hm2.保护区属典型的温带大陆性季风气候, 四季分明, 气候温和, 年平均气温11.9 ℃, 年降水量628.6 mm, 其中52%发生在7月和8月(Du et al., 2017).土壤以潮土和盐土为主, 地势南高北低, 地面坡降0.027%-0.031% (夏江宝等, 2016). ...
... 柽柳种群不同径级的空间关联性从总体来讲, 径级I、II与III两两之间在小尺度上表现为正关联, 在大尺度上则以相互独立的空间关系为主.径级I与径级II及径级I与径级III在较小尺度上表现为正关联, 是由于植株对环境资源的需求具有相似性, 但基径小的植株个体相对幼小, 对水分、土壤养分等资源的竞争能力相对薄弱, 需要依附相对高大植株的庇护以提高存活几率(尤海舟等, 2010).在本区域的前期研究中也发现, 柽柳的存在可以对其下生长的草本植物产生促进作用(Gao et al., 2015; Du et al., 2017).在对黄河三角洲滨海湿地的研究中还发现, 柽柳的生长可以增加土壤有机质含量, 提高土壤氮、磷、钾的含量, 从而形成“沃岛” (封晓辉等, 2013; Rong et al., 2016). ...
滨海重盐碱地人工栽植柽柳生长动态及生态效应 1 2013
... 柽柳种群不同径级的空间关联性从总体来讲, 径级I、II与III两两之间在小尺度上表现为正关联, 在大尺度上则以相互独立的空间关系为主.径级I与径级II及径级I与径级III在较小尺度上表现为正关联, 是由于植株对环境资源的需求具有相似性, 但基径小的植株个体相对幼小, 对水分、土壤养分等资源的竞争能力相对薄弱, 需要依附相对高大植株的庇护以提高存活几率(尤海舟等, 2010).在本区域的前期研究中也发现, 柽柳的存在可以对其下生长的草本植物产生促进作用(Gao et al., 2015; Du et al., 2017).在对黄河三角洲滨海湿地的研究中还发现, 柽柳的生长可以增加土壤有机质含量, 提高土壤氮、磷、钾的含量, 从而形成“沃岛” (封晓辉等, 2013; Rong et al., 2016). ...
滨海重盐碱地人工栽植柽柳生长动态及生态效应 1 2013
... 柽柳种群不同径级的空间关联性从总体来讲, 径级I、II与III两两之间在小尺度上表现为正关联, 在大尺度上则以相互独立的空间关系为主.径级I与径级II及径级I与径级III在较小尺度上表现为正关联, 是由于植株对环境资源的需求具有相似性, 但基径小的植株个体相对幼小, 对水分、土壤养分等资源的竞争能力相对薄弱, 需要依附相对高大植株的庇护以提高存活几率(尤海舟等, 2010).在本区域的前期研究中也发现, 柽柳的存在可以对其下生长的草本植物产生促进作用(Gao et al., 2015; Du et al., 2017).在对黄河三角洲滨海湿地的研究中还发现, 柽柳的生长可以增加土壤有机质含量, 提高土壤氮、磷、钾的含量, 从而形成“沃岛” (封晓辉等, 2013; Rong et al., 2016). ...
Assembly of plant communities in coastal wetlands-the role of saltcedar Tamarix chinensis during early succession 2 2015
... 柽柳(Tamarix chinensis)是温带滨海盐碱湿地的先锋灌木物种, 主要分布在中国东部和东北部的河流三角洲和盐碱湿地等条件苛刻的生境中, 在防止沿海地区海水入侵中发挥着重要作用(Cui et al., 2010).昌邑国家海洋生态特别保护区地处渤海莱州湾南岸, 是全国唯一的以柽柳群落为主要保护和管理对象的国家级海洋特别保护区, 是研究柽柳个体、种群、群落等不同层次特征的理想区域, 也是我国“南红北柳”生态工程的重要组成部分(谢琳萍等, 2017).目前对于滨海盐碱湿地柽柳的研究多集中于个体层面的柽柳叶片生理生化特征(朱金方等, 2013; Liu et al., 2014), 根系生理特性(宋香静等, 2018), 种群层面的柽柳种群空间分布特征(Jiang et al., 2012; Zhu et al., 2016), 群落层面的柽柳群落的土壤环境特征(Rong et al., 2016; Sun et al., 2016)和柽柳对草本植物生物多样性的影响(Gao et al., 2015; Du et al., 2017)等方面, 但对于柽柳群落中不同径级柽柳的分布格局及不同径级之间关联性的研究鲜有报道.研究滨海湿地先锋物种柽柳种群的空间分布格局及其不同径级之间的关联性, 有助于深化对滨海湿地植物种群组成、结构与动态的认识, 揭示种群发展规律, 为保育柽柳提供理论依据, 并进一步为湿地生态系统的管理提供基础资料. ...
... 柽柳种群不同径级的空间关联性从总体来讲, 径级I、II与III两两之间在小尺度上表现为正关联, 在大尺度上则以相互独立的空间关系为主.径级I与径级II及径级I与径级III在较小尺度上表现为正关联, 是由于植株对环境资源的需求具有相似性, 但基径小的植株个体相对幼小, 对水分、土壤养分等资源的竞争能力相对薄弱, 需要依附相对高大植株的庇护以提高存活几率(尤海舟等, 2010).在本区域的前期研究中也发现, 柽柳的存在可以对其下生长的草本植物产生促进作用(Gao et al., 2015; Du et al., 2017).在对黄河三角洲滨海湿地的研究中还发现, 柽柳的生长可以增加土壤有机质含量, 提高土壤氮、磷、钾的含量, 从而形成“沃岛” (封晓辉等, 2013; Rong et al., 2016). ...
Population genetic structure of Tamarix chinensis in the Yellow River Delta, China 1 2012
... 柽柳(Tamarix chinensis)是温带滨海盐碱湿地的先锋灌木物种, 主要分布在中国东部和东北部的河流三角洲和盐碱湿地等条件苛刻的生境中, 在防止沿海地区海水入侵中发挥着重要作用(Cui et al., 2010).昌邑国家海洋生态特别保护区地处渤海莱州湾南岸, 是全国唯一的以柽柳群落为主要保护和管理对象的国家级海洋特别保护区, 是研究柽柳个体、种群、群落等不同层次特征的理想区域, 也是我国“南红北柳”生态工程的重要组成部分(谢琳萍等, 2017).目前对于滨海盐碱湿地柽柳的研究多集中于个体层面的柽柳叶片生理生化特征(朱金方等, 2013; Liu et al., 2014), 根系生理特性(宋香静等, 2018), 种群层面的柽柳种群空间分布特征(Jiang et al., 2012; Zhu et al., 2016), 群落层面的柽柳群落的土壤环境特征(Rong et al., 2016; Sun et al., 2016)和柽柳对草本植物生物多样性的影响(Gao et al., 2015; Du et al., 2017)等方面, 但对于柽柳群落中不同径级柽柳的分布格局及不同径级之间关联性的研究鲜有报道.研究滨海湿地先锋物种柽柳种群的空间分布格局及其不同径级之间的关联性, 有助于深化对滨海湿地植物种群组成、结构与动态的认识, 揭示种群发展规律, 为保育柽柳提供理论依据, 并进一步为湿地生态系统的管理提供基础资料. ...
Effects of salt-drought stress on growth and physiobiochemical characteristics of Tamarix chinensis seedlings 1
... 柽柳(Tamarix chinensis)是温带滨海盐碱湿地的先锋灌木物种, 主要分布在中国东部和东北部的河流三角洲和盐碱湿地等条件苛刻的生境中, 在防止沿海地区海水入侵中发挥着重要作用(Cui et al., 2010).昌邑国家海洋生态特别保护区地处渤海莱州湾南岸, 是全国唯一的以柽柳群落为主要保护和管理对象的国家级海洋特别保护区, 是研究柽柳个体、种群、群落等不同层次特征的理想区域, 也是我国“南红北柳”生态工程的重要组成部分(谢琳萍等, 2017).目前对于滨海盐碱湿地柽柳的研究多集中于个体层面的柽柳叶片生理生化特征(朱金方等, 2013; Liu et al., 2014), 根系生理特性(宋香静等, 2018), 种群层面的柽柳种群空间分布特征(Jiang et al., 2012; Zhu et al., 2016), 群落层面的柽柳群落的土壤环境特征(Rong et al., 2016; Sun et al., 2016)和柽柳对草本植物生物多样性的影响(Gao et al., 2015; Du et al., 2017)等方面, 但对于柽柳群落中不同径级柽柳的分布格局及不同径级之间关联性的研究鲜有报道.研究滨海湿地先锋物种柽柳种群的空间分布格局及其不同径级之间的关联性, 有助于深化对滨海湿地植物种群组成、结构与动态的认识, 揭示种群发展规律, 为保育柽柳提供理论依据, 并进一步为湿地生态系统的管理提供基础资料. ...
A test of the unified neutral theory of biodiversity 1 2003
“Fertile island” effects of Tamarix chinensis Lour. on soil N and P stoichiometry in the coastal wetland of Laizhou Bay, China 2 2016
... 柽柳(Tamarix chinensis)是温带滨海盐碱湿地的先锋灌木物种, 主要分布在中国东部和东北部的河流三角洲和盐碱湿地等条件苛刻的生境中, 在防止沿海地区海水入侵中发挥着重要作用(Cui et al., 2010).昌邑国家海洋生态特别保护区地处渤海莱州湾南岸, 是全国唯一的以柽柳群落为主要保护和管理对象的国家级海洋特别保护区, 是研究柽柳个体、种群、群落等不同层次特征的理想区域, 也是我国“南红北柳”生态工程的重要组成部分(谢琳萍等, 2017).目前对于滨海盐碱湿地柽柳的研究多集中于个体层面的柽柳叶片生理生化特征(朱金方等, 2013; Liu et al., 2014), 根系生理特性(宋香静等, 2018), 种群层面的柽柳种群空间分布特征(Jiang et al., 2012; Zhu et al., 2016), 群落层面的柽柳群落的土壤环境特征(Rong et al., 2016; Sun et al., 2016)和柽柳对草本植物生物多样性的影响(Gao et al., 2015; Du et al., 2017)等方面, 但对于柽柳群落中不同径级柽柳的分布格局及不同径级之间关联性的研究鲜有报道.研究滨海湿地先锋物种柽柳种群的空间分布格局及其不同径级之间的关联性, 有助于深化对滨海湿地植物种群组成、结构与动态的认识, 揭示种群发展规律, 为保育柽柳提供理论依据, 并进一步为湿地生态系统的管理提供基础资料. ...
... 柽柳种群不同径级的空间关联性从总体来讲, 径级I、II与III两两之间在小尺度上表现为正关联, 在大尺度上则以相互独立的空间关系为主.径级I与径级II及径级I与径级III在较小尺度上表现为正关联, 是由于植株对环境资源的需求具有相似性, 但基径小的植株个体相对幼小, 对水分、土壤养分等资源的竞争能力相对薄弱, 需要依附相对高大植株的庇护以提高存活几率(尤海舟等, 2010).在本区域的前期研究中也发现, 柽柳的存在可以对其下生长的草本植物产生促进作用(Gao et al., 2015; Du et al., 2017).在对黄河三角洲滨海湿地的研究中还发现, 柽柳的生长可以增加土壤有机质含量, 提高土壤氮、磷、钾的含量, 从而形成“沃岛” (封晓辉等, 2013; Rong et al., 2016). ...
不同盐分水平对柽柳扦插苗根系生长及生理特性的影响 1 2018
... 柽柳(Tamarix chinensis)是温带滨海盐碱湿地的先锋灌木物种, 主要分布在中国东部和东北部的河流三角洲和盐碱湿地等条件苛刻的生境中, 在防止沿海地区海水入侵中发挥着重要作用(Cui et al., 2010).昌邑国家海洋生态特别保护区地处渤海莱州湾南岸, 是全国唯一的以柽柳群落为主要保护和管理对象的国家级海洋特别保护区, 是研究柽柳个体、种群、群落等不同层次特征的理想区域, 也是我国“南红北柳”生态工程的重要组成部分(谢琳萍等, 2017).目前对于滨海盐碱湿地柽柳的研究多集中于个体层面的柽柳叶片生理生化特征(朱金方等, 2013; Liu et al., 2014), 根系生理特性(宋香静等, 2018), 种群层面的柽柳种群空间分布特征(Jiang et al., 2012; Zhu et al., 2016), 群落层面的柽柳群落的土壤环境特征(Rong et al., 2016; Sun et al., 2016)和柽柳对草本植物生物多样性的影响(Gao et al., 2015; Du et al., 2017)等方面, 但对于柽柳群落中不同径级柽柳的分布格局及不同径级之间关联性的研究鲜有报道.研究滨海湿地先锋物种柽柳种群的空间分布格局及其不同径级之间的关联性, 有助于深化对滨海湿地植物种群组成、结构与动态的认识, 揭示种群发展规律, 为保育柽柳提供理论依据, 并进一步为湿地生态系统的管理提供基础资料. ...
不同盐分水平对柽柳扦插苗根系生长及生理特性的影响 1 2018
... 柽柳(Tamarix chinensis)是温带滨海盐碱湿地的先锋灌木物种, 主要分布在中国东部和东北部的河流三角洲和盐碱湿地等条件苛刻的生境中, 在防止沿海地区海水入侵中发挥着重要作用(Cui et al., 2010).昌邑国家海洋生态特别保护区地处渤海莱州湾南岸, 是全国唯一的以柽柳群落为主要保护和管理对象的国家级海洋特别保护区, 是研究柽柳个体、种群、群落等不同层次特征的理想区域, 也是我国“南红北柳”生态工程的重要组成部分(谢琳萍等, 2017).目前对于滨海盐碱湿地柽柳的研究多集中于个体层面的柽柳叶片生理生化特征(朱金方等, 2013; Liu et al., 2014), 根系生理特性(宋香静等, 2018), 种群层面的柽柳种群空间分布特征(Jiang et al., 2012; Zhu et al., 2016), 群落层面的柽柳群落的土壤环境特征(Rong et al., 2016; Sun et al., 2016)和柽柳对草本植物生物多样性的影响(Gao et al., 2015; Du et al., 2017)等方面, 但对于柽柳群落中不同径级柽柳的分布格局及不同径级之间关联性的研究鲜有报道.研究滨海湿地先锋物种柽柳种群的空间分布格局及其不同径级之间的关联性, 有助于深化对滨海湿地植物种群组成、结构与动态的认识, 揭示种群发展规律, 为保育柽柳提供理论依据, 并进一步为湿地生态系统的管理提供基础资料. ...
Temporal and spatial variations in the stable carbon isotope composition and carbon and nitrogen contents in current-season twigs of Tamarix chinensis Lour. and their relationships to environmental factors in the Laizhou Bay wetland in China 1 2016
... 柽柳(Tamarix chinensis)是温带滨海盐碱湿地的先锋灌木物种, 主要分布在中国东部和东北部的河流三角洲和盐碱湿地等条件苛刻的生境中, 在防止沿海地区海水入侵中发挥着重要作用(Cui et al., 2010).昌邑国家海洋生态特别保护区地处渤海莱州湾南岸, 是全国唯一的以柽柳群落为主要保护和管理对象的国家级海洋特别保护区, 是研究柽柳个体、种群、群落等不同层次特征的理想区域, 也是我国“南红北柳”生态工程的重要组成部分(谢琳萍等, 2017).目前对于滨海盐碱湿地柽柳的研究多集中于个体层面的柽柳叶片生理生化特征(朱金方等, 2013; Liu et al., 2014), 根系生理特性(宋香静等, 2018), 种群层面的柽柳种群空间分布特征(Jiang et al., 2012; Zhu et al., 2016), 群落层面的柽柳群落的土壤环境特征(Rong et al., 2016; Sun et al., 2016)和柽柳对草本植物生物多样性的影响(Gao et al., 2015; Du et al., 2017)等方面, 但对于柽柳群落中不同径级柽柳的分布格局及不同径级之间关联性的研究鲜有报道.研究滨海湿地先锋物种柽柳种群的空间分布格局及其不同径级之间的关联性, 有助于深化对滨海湿地植物种群组成、结构与动态的认识, 揭示种群发展规律, 为保育柽柳提供理论依据, 并进一步为湿地生态系统的管理提供基础资料. ...
昌邑海洋生态特别保护区土壤养分的空间异质性与植物群落的分布 2 2011
... 该保护区由中国国家海洋局于2007年成立, 被公认为是中国最大的天然柽柳灌丛湿地生态系统(Bi et al., 2014; Wang et al., 2015).植被以灌木和草本为主, 并呈平行于海岸线的带状分布(汤爱坤等, 2011).研究区内柽柳是唯一的灌木物种, 草本植物主要有猪毛蒿(Artemisia scoparia)、小蓬草(Conyza canadensis)、茵陈蒿(Artemisia capillaris)、狗尾草(Setaria viridis)和灰绿藜(Chenopodium glaucum)等, 藤本植物以鹅绒藤(Cynanchum chinense)为主. ...
... 研究区位于山东省昌邑市北部堤河以东、海岸线以下的滩涂上(37.05°-37.12° N, 119.33°-119.38° E), 总面积为2 929.28 hm2, 其中天然柽柳林面积达2 070 hm2.保护区属典型的温带大陆性季风气候, 四季分明, 气候温和, 年平均气温11.9 ℃, 年降水量628.6 mm, 其中52%发生在7月和8月(Du et al., 2017).土壤以潮土和盐土为主, 地势南高北低, 地面坡降0.027%-0.031% (夏江宝等, 2016). ...
黄河三角洲莱州湾湿地柽柳种群分布特征及其影响因素 1 2016
... 研究区位于山东省昌邑市北部堤河以东、海岸线以下的滩涂上(37.05°-37.12° N, 119.33°-119.38° E), 总面积为2 929.28 hm2, 其中天然柽柳林面积达2 070 hm2.保护区属典型的温带大陆性季风气候, 四季分明, 气候温和, 年平均气温11.9 ℃, 年降水量628.6 mm, 其中52%发生在7月和8月(Du et al., 2017).土壤以潮土和盐土为主, 地势南高北低, 地面坡降0.027%-0.031% (夏江宝等, 2016). ...
莱州湾滨海柽柳林湿地植被碳储量的分布特征及其影响因素 3 2017
... 柽柳(Tamarix chinensis)是温带滨海盐碱湿地的先锋灌木物种, 主要分布在中国东部和东北部的河流三角洲和盐碱湿地等条件苛刻的生境中, 在防止沿海地区海水入侵中发挥着重要作用(Cui et al., 2010).昌邑国家海洋生态特别保护区地处渤海莱州湾南岸, 是全国唯一的以柽柳群落为主要保护和管理对象的国家级海洋特别保护区, 是研究柽柳个体、种群、群落等不同层次特征的理想区域, 也是我国“南红北柳”生态工程的重要组成部分(谢琳萍等, 2017).目前对于滨海盐碱湿地柽柳的研究多集中于个体层面的柽柳叶片生理生化特征(朱金方等, 2013; Liu et al., 2014), 根系生理特性(宋香静等, 2018), 种群层面的柽柳种群空间分布特征(Jiang et al., 2012; Zhu et al., 2016), 群落层面的柽柳群落的土壤环境特征(Rong et al., 2016; Sun et al., 2016)和柽柳对草本植物生物多样性的影响(Gao et al., 2015; Du et al., 2017)等方面, 但对于柽柳群落中不同径级柽柳的分布格局及不同径级之间关联性的研究鲜有报道.研究滨海湿地先锋物种柽柳种群的空间分布格局及其不同径级之间的关联性, 有助于深化对滨海湿地植物种群组成、结构与动态的认识, 揭示种群发展规律, 为保育柽柳提供理论依据, 并进一步为湿地生态系统的管理提供基础资料. ...
... 柽柳(Tamarix chinensis)是温带滨海盐碱湿地的先锋灌木物种, 主要分布在中国东部和东北部的河流三角洲和盐碱湿地等条件苛刻的生境中, 在防止沿海地区海水入侵中发挥着重要作用(Cui et al., 2010).昌邑国家海洋生态特别保护区地处渤海莱州湾南岸, 是全国唯一的以柽柳群落为主要保护和管理对象的国家级海洋特别保护区, 是研究柽柳个体、种群、群落等不同层次特征的理想区域, 也是我国“南红北柳”生态工程的重要组成部分(谢琳萍等, 2017).目前对于滨海盐碱湿地柽柳的研究多集中于个体层面的柽柳叶片生理生化特征(朱金方等, 2013; Liu et al., 2014), 根系生理特性(宋香静等, 2018), 种群层面的柽柳种群空间分布特征(Jiang et al., 2012; Zhu et al., 2016), 群落层面的柽柳群落的土壤环境特征(Rong et al., 2016; Sun et al., 2016)和柽柳对草本植物生物多样性的影响(Gao et al., 2015; Du et al., 2017)等方面, 但对于柽柳群落中不同径级柽柳的分布格局及不同径级之间关联性的研究鲜有报道.研究滨海湿地先锋物种柽柳种群的空间分布格局及其不同径级之间的关联性, 有助于深化对滨海湿地植物种群组成、结构与动态的认识, 揭示种群发展规律, 为保育柽柳提供理论依据, 并进一步为湿地生态系统的管理提供基础资料. ...
... 柽柳(Tamarix chinensis)是温带滨海盐碱湿地的先锋灌木物种, 主要分布在中国东部和东北部的河流三角洲和盐碱湿地等条件苛刻的生境中, 在防止沿海地区海水入侵中发挥着重要作用(Cui et al., 2010).昌邑国家海洋生态特别保护区地处渤海莱州湾南岸, 是全国唯一的以柽柳群落为主要保护和管理对象的国家级海洋特别保护区, 是研究柽柳个体、种群、群落等不同层次特征的理想区域, 也是我国“南红北柳”生态工程的重要组成部分(谢琳萍等, 2017).目前对于滨海盐碱湿地柽柳的研究多集中于个体层面的柽柳叶片生理生化特征(朱金方等, 2013; Liu et al., 2014), 根系生理特性(宋香静等, 2018), 种群层面的柽柳种群空间分布特征(Jiang et al., 2012; Zhu et al., 2016), 群落层面的柽柳群落的土壤环境特征(Rong et al., 2016; Sun et al., 2016)和柽柳对草本植物生物多样性的影响(Gao et al., 2015; Du et al., 2017)等方面, 但对于柽柳群落中不同径级柽柳的分布格局及不同径级之间关联性的研究鲜有报道.研究滨海湿地先锋物种柽柳种群的空间分布格局及其不同径级之间的关联性, 有助于深化对滨海湿地植物种群组成、结构与动态的认识, 揭示种群发展规律, 为保育柽柳提供理论依据, 并进一步为湿地生态系统的管理提供基础资料. ...
盐旱交叉胁迫对柽柳幼苗渗透调节物质含量的影响 1 2013
... 柽柳(Tamarix chinensis)是温带滨海盐碱湿地的先锋灌木物种, 主要分布在中国东部和东北部的河流三角洲和盐碱湿地等条件苛刻的生境中, 在防止沿海地区海水入侵中发挥着重要作用(Cui et al., 2010).昌邑国家海洋生态特别保护区地处渤海莱州湾南岸, 是全国唯一的以柽柳群落为主要保护和管理对象的国家级海洋特别保护区, 是研究柽柳个体、种群、群落等不同层次特征的理想区域, 也是我国“南红北柳”生态工程的重要组成部分(谢琳萍等, 2017).目前对于滨海盐碱湿地柽柳的研究多集中于个体层面的柽柳叶片生理生化特征(朱金方等, 2013; Liu et al., 2014), 根系生理特性(宋香静等, 2018), 种群层面的柽柳种群空间分布特征(Jiang et al., 2012; Zhu et al., 2016), 群落层面的柽柳群落的土壤环境特征(Rong et al., 2016; Sun et al., 2016)和柽柳对草本植物生物多样性的影响(Gao et al., 2015; Du et al., 2017)等方面, 但对于柽柳群落中不同径级柽柳的分布格局及不同径级之间关联性的研究鲜有报道.研究滨海湿地先锋物种柽柳种群的空间分布格局及其不同径级之间的关联性, 有助于深化对滨海湿地植物种群组成、结构与动态的认识, 揭示种群发展规律, 为保育柽柳提供理论依据, 并进一步为湿地生态系统的管理提供基础资料. ...
Local habitat condition rather than geographic distance determines the genetic structure of Tamarix chinensis populations in Yellow River Delta, China 1 2016
... 柽柳(Tamarix chinensis)是温带滨海盐碱湿地的先锋灌木物种, 主要分布在中国东部和东北部的河流三角洲和盐碱湿地等条件苛刻的生境中, 在防止沿海地区海水入侵中发挥着重要作用(Cui et al., 2010).昌邑国家海洋生态特别保护区地处渤海莱州湾南岸, 是全国唯一的以柽柳群落为主要保护和管理对象的国家级海洋特别保护区, 是研究柽柳个体、种群、群落等不同层次特征的理想区域, 也是我国“南红北柳”生态工程的重要组成部分(谢琳萍等, 2017).目前对于滨海盐碱湿地柽柳的研究多集中于个体层面的柽柳叶片生理生化特征(朱金方等, 2013; Liu et al., 2014), 根系生理特性(宋香静等, 2018), 种群层面的柽柳种群空间分布特征(Jiang et al., 2012; Zhu et al., 2016), 群落层面的柽柳群落的土壤环境特征(Rong et al., 2016; Sun et al., 2016)和柽柳对草本植物生物多样性的影响(Gao et al., 2015; Du et al., 2017)等方面, 但对于柽柳群落中不同径级柽柳的分布格局及不同径级之间关联性的研究鲜有报道.研究滨海湿地先锋物种柽柳种群的空间分布格局及其不同径级之间的关联性, 有助于深化对滨海湿地植物种群组成、结构与动态的认识, 揭示种群发展规律, 为保育柽柳提供理论依据, 并进一步为湿地生态系统的管理提供基础资料. ...
 ,, 彭希强, 杨树仁, 高亚男, 白丰桦, 衣世杰, 杜宁
,, 彭希强, 杨树仁, 高亚男, 白丰桦, 衣世杰, 杜宁 ,*, 郭卫华
,*, 郭卫华
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT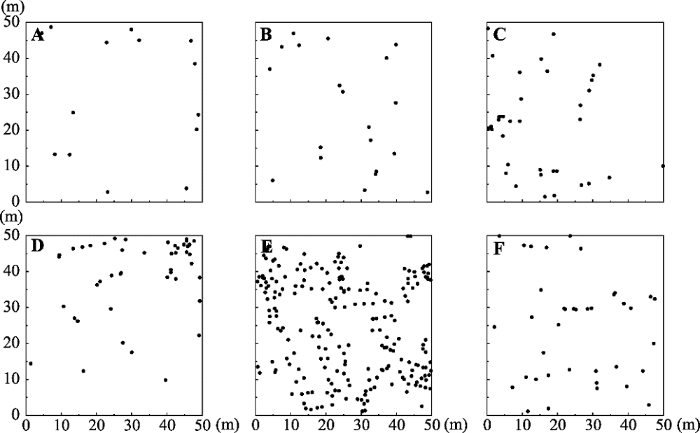 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT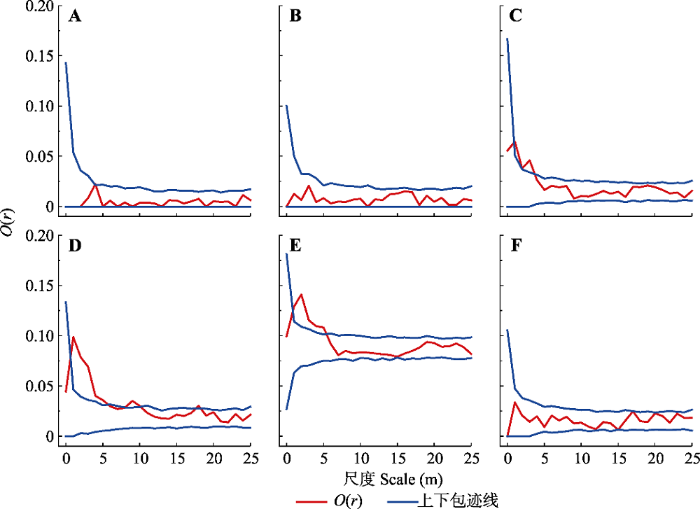 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT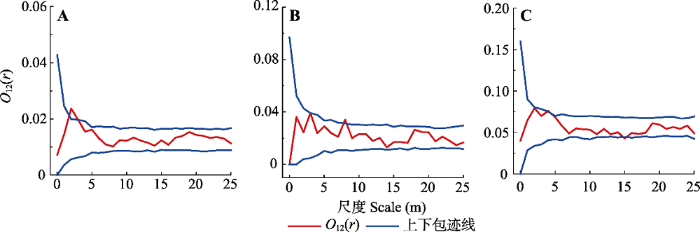 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}