Distribution and species diversity patterns of Vitex negundo var. heterophylla shrublands in North China
Li-Li TANG1, Tong YANG1, Hong-Yan LIU2, Mu-Yi KANG3,4, Ren-Qing WANG5, Feng ZHANG6, Xian-Ming GAO7, Ming YUE8, Mei ZHANG1, Pu-Fan ZHENG1, Fu-Chen SHI,1,*1College of Life Sciences, Nankai University, Tianjin 300071, China 2Institute of Ecology, College of Urban and Environmental Sciences, Laboratory for Earth Surface Processes of the Ministry of Education, Peking University, Beijing 100871, China 3State Key Laboratory of Earth Surface Processes and Resource Ecology, Beijing Normal University, Beijing 100875, China 4College of Resources Science & Technology, Faculty of Geographical Science, Beijing Normal University, Beijing 100875, China 5School of Life Sciences, Shandong University, Jinan 250100, China 6Institute of Loess Plateau, Shanxi University, Taiyuan 030006, China 7State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academic of Science, Beijing 100093, China 8Key Laboratory of Resource Biology and Biotechnology in Western China, Ministry of Education, Northwest University, Xi’an 710069, China
Supported by the National Basic work of Science and Technology of China(2011FY110300) the Special Fund for the Industrial System Construction of Modern Agriculture of Tianjin, China(ITTFPRS2018001) and the Program of Science and Technology Commission Foundation of Tianjin, China(18ZXZYNC00120)
Abstract Aims To explore the factors controlling the spatial distribution, as well as the species diversity, of the Vitex negundo var. heterophylla shrublands in North China. Methods We investigated the species composition and environments for 370 shrubland plots of Vitex negundo var. heterophylla shrublands in North China. We calculated biodiversity indices (Shannon-Wiener index, Simpson index and Pielou’s evenness index) and then applied multivariate regression trees (MRT), analysis of variance (ANOVA), Pearson correlation analysis to disentangle distribution of the Vitex negundo var. heterophylla community and patterns of species diversity along both horizontal and elevational gradients. Important findings The results showed that Vitex negundo var. heterophylla shrublands distributed in a large area stretching for more than 10° of both latitude and longitude, together with an elevational range of more than 1 000 m (117 m to 1 248 m). The Shannon-Wiener index, Simpson index and Pielou’s evenness index were higher in the herbaceous layer than those in the shrub layers. The species diversity of shrub layer increased with latitude but decreased with elevation. In contrast, that of the herbaceous layer decreased with latitude but increased with elevation. The Shannon-Wiener indexes of both layers were significantly correlated with environmental factors. The results of Pearson correlation analysis indicated that the Shannon-Wiener indexes of shrub layer had a significantly negative relationship with its temperature and slope position, and a significantly positive relationship with its precipitation. However, that of herb layer showed opposite trends. Keywords:Vitex negundo var. heterophylla shrubland;species diversity index;spatial pattern;plant community
PDF (2811KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 唐丽丽, 杨彤, 刘鸿雁, 康慕谊, 王仁卿, 张峰, 高贤明, 岳明, 张梅, 郑璞帆, 石福臣. 华北地区荆条灌丛分布及物种多样性空间分异 规律. 植物生态学报, 2019, 43(9): 825-833. DOI: 10.17521/cjpe.2018.0162 TANG Li-Li, YANG Tong, LIU Hong-Yan, KANG Mu-Yi, WANG Ren-Qing, ZHANG Feng, GAO Xian-Ming, YUE Ming, ZHANG Mei, ZHENG Pu-Fan, SHI Fu-Chen. Distribution and species diversity patterns of Vitex negundo var. heterophylla shrublands in North China. Chinese Journal of Plant Ecology, 2019, 43(9): 825-833. DOI: 10.17521/cjpe.2018.0162
Table 2 表2 表2华北不同纬度带荆条灌丛物种多样性指数 Table 2Biodiversity indices of Vitex negundo var. heterophylla shrublands in different latitudinal zones in North China
Table 3 表3 表3华北不同垂直带荆条灌丛物种多样性指数 Table 3Biodiversity indices of Vitex negundo var. heterophylla shrublands in different elevational zones in North China
Table 4 表4 表4华北地区荆条灌丛不同分布区环境因子单因素方差分析(平均值±标准误差) Table 4ANOVA of environmental variables of the Vitex negundo var. heterophylla shrublands in different latitudinal and elevational zones in North China (mean ± SE)
群丛 Association
BIO3
BIO5
BIO15
BIO18
1
28.08 ± 0.05c
30.01 ± 0.07c
129.44 ± 0.47a
378.50 ± 2.58ab
2
28.50 ± 0.37b
32.24 ± 0.29a
114.50 ± 1.28b
351.18 ± 2.60b
3
29.11 ± 0.16b
31.15 ± 0.16b
113.09 ± 0.79b
345.25 ± 7.25b
4
30.58 ± 0.24a
28.29 ± 0.30d
110.32 ± 1.54b
293.21 ± 6.73c
5
28.90 ± 0.14b
30.41 ± 0.14bc
95.83 ± 1.49c
402.91 ± 6.41a
Different lowercase letters indicate significant differences among different formation (p < 0.05). BIO3, isothermality; BIO5, max temperature of warmest month; BIO15, precipitation seasonality (coefficient of variation); BIO18, precipitation of warmest quarter. 同一列数据不同小写字母表示差异显著(p < 0.05)。BIO3, 等温性; BIO5, 最热月份最高气温; BIO15, 降水量季节变异系数; BIO18, 最热季度降水量。
Table 5 表5 表5华北地区荆条灌丛物种多样性与环境因子相关性检验 Table 5Pearson correlation between species diversity and environmental factors of the Vitex negundo var. heterophylla shrublands in North China
环境因子 Variables
灌木层物种多样性 Biodiversity indices of the shrub layer
草本层物种多样性 Biodiversity indices of the herbaceous layer
相关系数 Correlation coefficient
相关系数 Correlation coefficient
BIO3
-0.41**
0.14**
BIO5
0.07
-0.13*
BIO15
0.69**
-0.51**
BIO18
-0.02
0.27**
ELE
-0.37**
0.22**
ASP
-0.06
0.10
SLO
0.34**
-0.08
SLOP
-0.17**
0.28**
**, p < 0.01; *, p < 0.05. BIO3, isothermality; BIO5, max temperature of warmest month; BIO15, precipitation seasonality (coefficient of variation); BIO18, precipitation of warmest quarter; ASP, aspect; Ele, elevation; SLO, slope; SLOP, slope position. **, p < 0.01; *, p < 0.05。BIO3, 等温性; BIO5, 最热月份最高气温; BIO15, 降水量季节变异系数; BIO18, 最热季度降水量; ASP, 坡向; Ele, 海拔; SLO, 坡度; SLOP, 坡位。
AllenAP, BrownJH, GilloolyJF ( 2002). Global biodiversity, biochemical kinetics, and the energetic-equivalence rule Science, 297, 1545-1548. [本文引用: 1]
BaoYF, GaoJR, HeMY, ZhangJR ( 2008). Study on age structure of Vitex negundo community in Miyun Reservoir watershed Forest Resources Management, ( 6), 67-70. [本文引用: 1]
BlanckYL, GowdaJ, M?rtenssonLM, SandbergJ, FranssonAM ( 2011). Plant species richness in a natural Argentinian matorral shrub-land correlates negatively with levels of plant phosphorus Plant and Soil, 345, 11-21. [本文引用: 1]
BobbinkR, HicksK, GallowayJ, SprangerT, AlkemadeR, AshmoreM, BustamanteM, CinderbyS, DavidsonE, DentenerF, EmmettB, ErismanJW, FennM, GilliamF, NordinA, PardoL, de VriesW ( 2010). Global assessment of nitrogen deposition effects on terrestrial plant diversity: A synthesis Ecological Applications, 20, 30-59. [本文引用: 1]
ChenY, WangHL, HanJW, WeiBL, JiaHR, YeYZ, YuanZL ( 2014). Numerical classification, ordination and species diversity along elevation gradients of the forest community in Xiaoqinling Acta Ecologica Sinica, 34, 2068-2075. [本文引用: 2]
ChengJ, LiuYH, ZhangJE ( 2016). The distribution and growth trait of Vitex negundo shrub in the loess region. Journal of Soil and Water Conservation, 30, 348-352. [本文引用: 2]
DaiXB ( 1989). The seasonal dynamics of the biomass of Vitex shrubland in mountainous area of Huairou County in Beijing Acta Botanica Sinica, 31, 307-315. [本文引用: 1]
GaoML, GongZN, ZhaoWJ, GaoY, HuD ( 2014). The study of Vitex negundo shrubs canopy biomass inversion in Beijing Jundu mountainous area based on vegetation indices Acta Ecologica Sinica, 34, 1178-1188. [本文引用: 1]
JinH, ZhaoY, ZhaoW, YinH, QinLW, CuiKF, LiXJ, HuangXT ( 2015). Elevational changes in species diversity and similarity analysis of a Rhododendron chrysanthum community on Changbai Mountain Acta Ecologica Sinica, 35, 125-133. [本文引用: 1]
le BrocqueAF, BuckneyRT ( 2003). Species richness-environment relationships within coastal sclerophyll and mesophyll vegetation in Ku-ring-gai Chase National Park, New South Wales, Australia Austral Ecology, 28, 404-412. [本文引用: 1]
LiSQ, YangBS, WuDM ( 2008). Community succession analysis of naturally colonized plants on coal gob piles in Shanxi mining areas, China Water Air & Soil Pollution, 193, 211-228. [本文引用: 1]
LiXH ( 2011). The Research of Distribution Pattern of Vitex negundo Shrub in the Watershed of Miyun Reservoir Master degree dissertation, Beijing Forestry University, Beijing. [本文引用: 1]
LiXR, TanHJ, HeMZ, WangXP, LiXJ ( 2009). Patterns of shrub species richness and abundance in relation to environmental factors on the Alxa Plateau: Prerequisites for conserving shrub diversity in extreme arid desert regions Science in China Series D-Earth Sciences, 52, 669-680. [本文引用: 1]
LiuYY, ZhouGZ ( 2011). Study on drought resistance of three shrub tree species Northern Horticulture, ( 8), 88-90. [本文引用: 1]
ModHK, HeikkinenRK, le RouxPC, WiszMS, LuotoM ( 2016). Impact of biotic interactions on biodiversity varies across a landscape Journal of Biogeography, 43, 2412-2423. [本文引用: 1]
NieEB ( 2006). Study on Quantitative Ecology of Vitex negundo L. var. heterophylla (Franch) Communities in Canyong Destrict of Taihang Mountains, in the southern Shanxi, China Master degree dissertation, Shanxi University, Taiyuan. [本文引用: 1]
NieEB, WangYQ, ZhangJT ( 2009a). Interspecific relationships of Vitex negundo var. heterophylla community of Taihang Mountain in Shanxi Province Pratacultural Science, 26(10), 24-29. [本文引用: 1]
NeiEB, WangYQ, ZhangJT ( 2009b). Study on the diversity of Vitex negundo var. heterophylla community in Taihang Mountain of Shanxi Province Pratacultural Science, 26(6), 6-10. [本文引用: 1]
NiklasKJ, MidgleyJJ, RandRH ( 2003). Size-dependent species richness: Trends within plant communities and across latitude Ecology Letters, 6, 631-636. [本文引用: 1]
NorthM, OakleyB, FiegenerR, GrayA, BarbourM ( 2005). Influence of light and soil moisture on Sierran mixed- conifer understory communities Plant Ecology, 177, 13-24. [本文引用: 1]
OlivierB, ValeriaDC, AntoineG ( 2017). ecospat: Spatial ecology miscellaneous methods. R package version 2.2.0
RosenzweigML ( 1995). Species Diversity in Space and Time. Cambridge University Press, Cambridge, UK. [本文引用: 1]
RuWM ( 1993). The community structure of Vitex negundo var. heterophylla ( Franch) Rehd scrub and its biomass in mountainous area of Lingchuan County Shanxi Province Journal of Shanxi Normal University (Natural Science Edition), ( S2), 55-58. [本文引用: 2]
ShenZH, LiuZL, FangJY ( 2004). Altitudinal changes in species diversity and community structure of Abies fabri communities at Hailuo Valley of Mt. Gongga, Sichuan Biodiversity Science, 12, 237-244. [本文引用: 1]
SwensonNG, WeiserMD, MaoLF, NormandS, RodríguezMá, LinLX, CaoM, SvenningJC ( 2016). Constancy in functional space across a species richness anomaly The American Naturalist, 187, 83-92. [本文引用: 1]
TherneauTM, AtkinsonB ( 2014). mvpart: Multivariate partitioning. R package version 1.6-1
WangXB, HanLB, LiuCX ( 2007). Experiments on germination of seeds of Vitex negundo var. Heterophylla—A superior shrub for soil and water conservation Liaoning Forestry Science and Technology, ( 4), 30-32. [本文引用: 2]
WangYQ, NieEB ( 2009). Quantitative analysis of the Vitex negundo var. heterophylla communities in Taihang Mountain, Shanxi Province Pratacultural Science, 26(11), 32-36. [本文引用: 1]
WenY, YangWB, YanSX, WangJY, YanDR ( 2006). Conparison of resistance of 8 kinds of local tree species Journal of Arid Land Resources and Environment, 20(4), 204-208. [本文引用: 1]
WilligMR, KaufmanDM, StevensRD ( 2003). Latitudinal gradients of biodiversity: Pattern, process, scale, and synthesis Annual Review of Ecology Evolution and Systematics, 34, 273-309. [本文引用: 1]
WuYJ, MiaoYM, BiRC ( 2015). Herbaceous plant species diversity and regeneration in shrub gaps in Huoshan Mountain of Shanxi Province Plant Diversity and Resources, 37, 203-208. [本文引用: 1]
XuM, MaL, JiaY, LiuM ( 2017). Integrating the effects of latitude and altitude on the spatial differentiation of plant community diversity in a mountainous ecosystem in China PLOS ONE, 12, e0174231. DOI: 10.1371/journal. pone.0174231. [本文引用: 1]
XuYJ, ChenYN, LiWH, FuAH, MaXD, GuiDW, ChenYP ( 2010). Distribution pattern and environmental interpretation of plant species diversity in the mountainous region of Ili River Valley, Xinjiang, China Chinese Journal of Plant Ecology, 34, 1142-1154. [本文引用: 2]
YangY, HanJ, LiuY, ZhongYC, ShiS, SinaC, XuY, YingL, ZhangW, ShenZ ( 2016). A comparison of the altitudinal patterns in plant species diversity within the dry valleys of the Three Parallel Rivers region, northwestern Yunnan Biodiversity Science, 24, 440-452. [本文引用: 3]
ZhangCX ( 2007). Study on Bioecological Characteristics of Soil and Water Conservation Shurb—Vitex negundo Master degree dissertation, Beijing Forestry University, Beijing. [本文引用: 2]
Global biodiversity, biochemical kinetics, and the energetic-equivalence rule 1 2002
... 物种多样性受诸多因素影响, 在大尺度下, 纬度不同所带来的气候差异是多样性变化的最主要原因(Rosenzweig, 1995; Allen et al., 2002; Xu et al., 2017), 多个研究表明, 随着纬度的降低, 物种多样性增加(Niklas et al., 2003; Willig et al., 2003).小尺度下, 海拔、土壤的营养成分、土壤含水量、物种本身的一些功能属性等都是物种多样性变化的原因, 研究表明随着海拔的增高物种多样性降低, 土壤因子和植物功能属性则对多样性有不同程度的影响(North et al., 2005; Bobbink et al., 2010; 徐远杰等, 2010; Swenson et al., 2016; Yang et al., 2016). ...
Plant species richness in a natural Argentinian matorral shrub-land correlates negatively with levels of plant phosphorus 1 2011
... 在水平分布带上, 随着纬度的降低, 灌木层物种丰富度(H)、分布的均匀性(D)降低, 草本层物种丰富度、分布的均匀性增加, 但物种种类的均匀度(J)在中纬度地区更高, 不呈纬度性变化.在垂直分布带上, 随着海拔的升高, 灌木层物种丰富度降低, 草本层物种丰富度有所升高, 这与大多数研究结果一致, 高海拔地区草本物种更加丰富(Le Brocque & Buckney, 2003; Li et al., 2009; Blanck et al., 2011; Mod et al., 2016; Yang et al., 2016). ...
Global assessment of nitrogen deposition effects on terrestrial plant diversity: A synthesis 1 2010
... 物种多样性受诸多因素影响, 在大尺度下, 纬度不同所带来的气候差异是多样性变化的最主要原因(Rosenzweig, 1995; Allen et al., 2002; Xu et al., 2017), 多个研究表明, 随着纬度的降低, 物种多样性增加(Niklas et al., 2003; Willig et al., 2003).小尺度下, 海拔、土壤的营养成分、土壤含水量、物种本身的一些功能属性等都是物种多样性变化的原因, 研究表明随着海拔的增高物种多样性降低, 土壤因子和植物功能属性则对多样性有不同程度的影响(North et al., 2005; Bobbink et al., 2010; 徐远杰等, 2010; Swenson et al., 2016; Yang et al., 2016). ...
Species richness-environment relationships within coastal sclerophyll and mesophyll vegetation in Ku-ring-gai Chase National Park, New South Wales, Australia 1 2003
... 在水平分布带上, 随着纬度的降低, 灌木层物种丰富度(H)、分布的均匀性(D)降低, 草本层物种丰富度、分布的均匀性增加, 但物种种类的均匀度(J)在中纬度地区更高, 不呈纬度性变化.在垂直分布带上, 随着海拔的升高, 灌木层物种丰富度降低, 草本层物种丰富度有所升高, 这与大多数研究结果一致, 高海拔地区草本物种更加丰富(Le Brocque & Buckney, 2003; Li et al., 2009; Blanck et al., 2011; Mod et al., 2016; Yang et al., 2016). ...
Community succession analysis of naturally colonized plants on coal gob piles in Shanxi mining areas, China 1 2008
Patterns of shrub species richness and abundance in relation to environmental factors on the Alxa Plateau: Prerequisites for conserving shrub diversity in extreme arid desert regions 1 2009
... 在水平分布带上, 随着纬度的降低, 灌木层物种丰富度(H)、分布的均匀性(D)降低, 草本层物种丰富度、分布的均匀性增加, 但物种种类的均匀度(J)在中纬度地区更高, 不呈纬度性变化.在垂直分布带上, 随着海拔的升高, 灌木层物种丰富度降低, 草本层物种丰富度有所升高, 这与大多数研究结果一致, 高海拔地区草本物种更加丰富(Le Brocque & Buckney, 2003; Li et al., 2009; Blanck et al., 2011; Mod et al., 2016; Yang et al., 2016). ...
Impact of biotic interactions on biodiversity varies across a landscape 1 2016
... 在水平分布带上, 随着纬度的降低, 灌木层物种丰富度(H)、分布的均匀性(D)降低, 草本层物种丰富度、分布的均匀性增加, 但物种种类的均匀度(J)在中纬度地区更高, 不呈纬度性变化.在垂直分布带上, 随着海拔的升高, 灌木层物种丰富度降低, 草本层物种丰富度有所升高, 这与大多数研究结果一致, 高海拔地区草本物种更加丰富(Le Brocque & Buckney, 2003; Li et al., 2009; Blanck et al., 2011; Mod et al., 2016; Yang et al., 2016). ...
Size-dependent species richness: Trends within plant communities and across latitude 1 2003
... 物种多样性受诸多因素影响, 在大尺度下, 纬度不同所带来的气候差异是多样性变化的最主要原因(Rosenzweig, 1995; Allen et al., 2002; Xu et al., 2017), 多个研究表明, 随着纬度的降低, 物种多样性增加(Niklas et al., 2003; Willig et al., 2003).小尺度下, 海拔、土壤的营养成分、土壤含水量、物种本身的一些功能属性等都是物种多样性变化的原因, 研究表明随着海拔的增高物种多样性降低, 土壤因子和植物功能属性则对多样性有不同程度的影响(North et al., 2005; Bobbink et al., 2010; 徐远杰等, 2010; Swenson et al., 2016; Yang et al., 2016). ...
Influence of light and soil moisture on Sierran mixed- conifer understory communities 1 2005
... 物种多样性受诸多因素影响, 在大尺度下, 纬度不同所带来的气候差异是多样性变化的最主要原因(Rosenzweig, 1995; Allen et al., 2002; Xu et al., 2017), 多个研究表明, 随着纬度的降低, 物种多样性增加(Niklas et al., 2003; Willig et al., 2003).小尺度下, 海拔、土壤的营养成分、土壤含水量、物种本身的一些功能属性等都是物种多样性变化的原因, 研究表明随着海拔的增高物种多样性降低, 土壤因子和植物功能属性则对多样性有不同程度的影响(North et al., 2005; Bobbink et al., 2010; 徐远杰等, 2010; Swenson et al., 2016; Yang et al., 2016). ...
ecospat: Spatial ecology miscellaneous methods. R package version 2.2.0 1 2017
... 物种多样性受诸多因素影响, 在大尺度下, 纬度不同所带来的气候差异是多样性变化的最主要原因(Rosenzweig, 1995; Allen et al., 2002; Xu et al., 2017), 多个研究表明, 随着纬度的降低, 物种多样性增加(Niklas et al., 2003; Willig et al., 2003).小尺度下, 海拔、土壤的营养成分、土壤含水量、物种本身的一些功能属性等都是物种多样性变化的原因, 研究表明随着海拔的增高物种多样性降低, 土壤因子和植物功能属性则对多样性有不同程度的影响(North et al., 2005; Bobbink et al., 2010; 徐远杰等, 2010; Swenson et al., 2016; Yang et al., 2016). ...
Constancy in functional space across a species richness anomaly 1 2016
... 物种多样性受诸多因素影响, 在大尺度下, 纬度不同所带来的气候差异是多样性变化的最主要原因(Rosenzweig, 1995; Allen et al., 2002; Xu et al., 2017), 多个研究表明, 随着纬度的降低, 物种多样性增加(Niklas et al., 2003; Willig et al., 2003).小尺度下, 海拔、土壤的营养成分、土壤含水量、物种本身的一些功能属性等都是物种多样性变化的原因, 研究表明随着海拔的增高物种多样性降低, 土壤因子和植物功能属性则对多样性有不同程度的影响(North et al., 2005; Bobbink et al., 2010; 徐远杰等, 2010; Swenson et al., 2016; Yang et al., 2016). ...
mvpart: Multivariate partitioning. R package version 1.6-1 1 2014
Latitudinal gradients of biodiversity: Pattern, process, scale, and synthesis 1 2003
... 物种多样性受诸多因素影响, 在大尺度下, 纬度不同所带来的气候差异是多样性变化的最主要原因(Rosenzweig, 1995; Allen et al., 2002; Xu et al., 2017), 多个研究表明, 随着纬度的降低, 物种多样性增加(Niklas et al., 2003; Willig et al., 2003).小尺度下, 海拔、土壤的营养成分、土壤含水量、物种本身的一些功能属性等都是物种多样性变化的原因, 研究表明随着海拔的增高物种多样性降低, 土壤因子和植物功能属性则对多样性有不同程度的影响(North et al., 2005; Bobbink et al., 2010; 徐远杰等, 2010; Swenson et al., 2016; Yang et al., 2016). ...
Integrating the effects of latitude and altitude on the spatial differentiation of plant community diversity in a mountainous ecosystem in China 1 2017
... 物种多样性受诸多因素影响, 在大尺度下, 纬度不同所带来的气候差异是多样性变化的最主要原因(Rosenzweig, 1995; Allen et al., 2002; Xu et al., 2017), 多个研究表明, 随着纬度的降低, 物种多样性增加(Niklas et al., 2003; Willig et al., 2003).小尺度下, 海拔、土壤的营养成分、土壤含水量、物种本身的一些功能属性等都是物种多样性变化的原因, 研究表明随着海拔的增高物种多样性降低, 土壤因子和植物功能属性则对多样性有不同程度的影响(North et al., 2005; Bobbink et al., 2010; 徐远杰等, 2010; Swenson et al., 2016; Yang et al., 2016). ...
... 物种多样性受诸多因素影响, 在大尺度下, 纬度不同所带来的气候差异是多样性变化的最主要原因(Rosenzweig, 1995; Allen et al., 2002; Xu et al., 2017), 多个研究表明, 随着纬度的降低, 物种多样性增加(Niklas et al., 2003; Willig et al., 2003).小尺度下, 海拔、土壤的营养成分、土壤含水量、物种本身的一些功能属性等都是物种多样性变化的原因, 研究表明随着海拔的增高物种多样性降低, 土壤因子和植物功能属性则对多样性有不同程度的影响(North et al., 2005; Bobbink et al., 2010; 徐远杰等, 2010; Swenson et al., 2016; Yang et al., 2016). ...
... 物种多样性受诸多因素影响, 在大尺度下, 纬度不同所带来的气候差异是多样性变化的最主要原因(Rosenzweig, 1995; Allen et al., 2002; Xu et al., 2017), 多个研究表明, 随着纬度的降低, 物种多样性增加(Niklas et al., 2003; Willig et al., 2003).小尺度下, 海拔、土壤的营养成分、土壤含水量、物种本身的一些功能属性等都是物种多样性变化的原因, 研究表明随着海拔的增高物种多样性降低, 土壤因子和植物功能属性则对多样性有不同程度的影响(North et al., 2005; Bobbink et al., 2010; 徐远杰等, 2010; Swenson et al., 2016; Yang et al., 2016). ...
A comparison of the altitudinal patterns in plant species diversity within the dry valleys of the Three Parallel Rivers region, northwestern Yunnan 3 2016
... 在水平分布带上, 随着纬度的降低, 灌木层物种丰富度(H)、分布的均匀性(D)降低, 草本层物种丰富度、分布的均匀性增加, 但物种种类的均匀度(J)在中纬度地区更高, 不呈纬度性变化.在垂直分布带上, 随着海拔的升高, 灌木层物种丰富度降低, 草本层物种丰富度有所升高, 这与大多数研究结果一致, 高海拔地区草本物种更加丰富(Le Brocque & Buckney, 2003; Li et al., 2009; Blanck et al., 2011; Mod et al., 2016; Yang et al., 2016). ...
... 物种多样性受诸多因素影响, 在大尺度下, 纬度不同所带来的气候差异是多样性变化的最主要原因(Rosenzweig, 1995; Allen et al., 2002; Xu et al., 2017), 多个研究表明, 随着纬度的降低, 物种多样性增加(Niklas et al., 2003; Willig et al., 2003).小尺度下, 海拔、土壤的营养成分、土壤含水量、物种本身的一些功能属性等都是物种多样性变化的原因, 研究表明随着海拔的增高物种多样性降低, 土壤因子和植物功能属性则对多样性有不同程度的影响(North et al., 2005; Bobbink et al., 2010; 徐远杰等, 2010; Swenson et al., 2016; Yang et al., 2016). ...
 ,1,*
,1,*
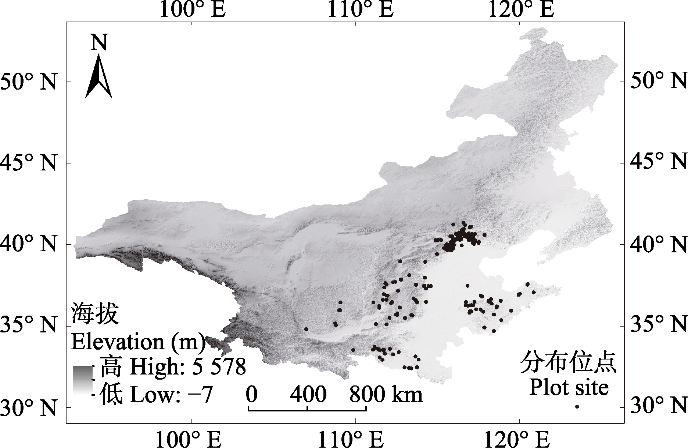 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT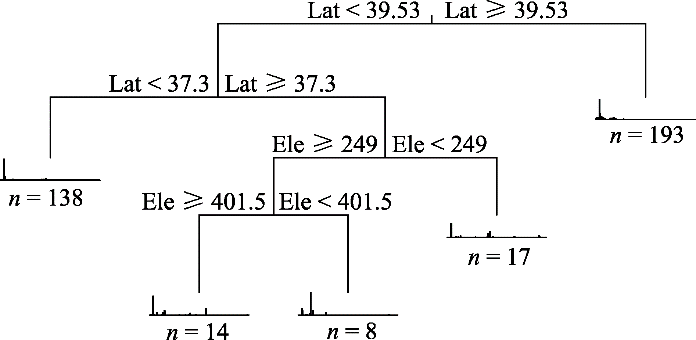 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT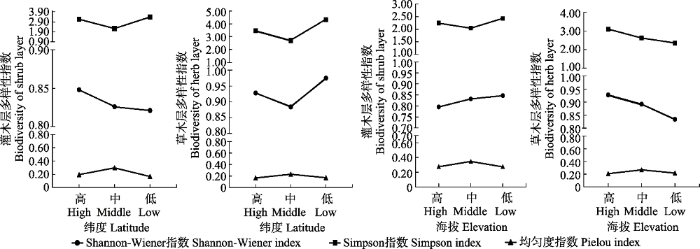 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}