Abstract Embolism induced by the freeze-thaw commonly occurs in the woody plant in mid- and high-latitude regions. In cold environments, the resistance capacity to freeze-thaw embolism of woody plants is critical for their growth and successful survival throughout winter, which directly determines their distribution in the earth. During freeze-thaw cycles, the freeze-thaw embolism of vessels is induced by the bubbles that are from dissolved frozen water because of the low solubility of ice. The resistance capacity to freeze-thaw embolism varies from plants with different anatomical structures. Plants can also reduce potential damages of freeze-thaw embolism through adjusting the xylem positive pressure by refilling embolism and metabolism, and/or increasing the sap solute contents to avoid the stress. Compared to embolism induced by drought, however, few studies have been conducted on embolism by freeze-thaw, and the underlying mechanisms of plant responses and regulation are poorly understood. In this paper, we first reviewed the process of formation and repair of freeze-thaw embolism of xylem, including the strategies of escaping freezing, tolerance, and repairing freeze-thaw cavitation. Then we summarized physiological characteristics of plant resistance to low-temperature stress, influencing factors and evaluating criteria, based on the multiple trade-offs between low-temperature resistance, drought resistance, and hydraulic efficiency. Finally, we proposed five priorities for future study in this field: (1) the threshold of freezing temperature for different plants; (2) the vulnerability segmentation of frost-induced embolism; (3) the relationships between freeze-thaw cavitation repair and metabolism exhaustion; (4) the potential trade-offs between low-temperature resistance, drought resistance and hydraulic efficiency; and (5) integration of the traits of freeze-thaw embolism resistance into the economic spectrum of plants. Keywords:freeze-thaw embolism;embolism repair;hydraulic efficiency;trade-off;hydraulic characteristic
Fig. 2A conceptual diagram of multiple trade-offs among low-temperature resistance, drought resistance and hydraulic efficiency of the major vegetation types in the world. High (low) hydraulic efficiency is associated with weak (strong) low-temperature resistance, but the capacity of positive refilling embolism may influence the trade-off between hydraulic efficiency and low-temperature resistance, and low-temperature also affects the plant’s frost fatigue. High (low) hydraulic efficiency is associated with weak (strong) drought resistance, but many species have both low hydraulic efficiency and drought resistance rather than high hydraulic efficiency and drought resistance. Drought stress may increase low-temperature resistance (asynchronous), and vice versa; concomitant drought and low-temperature stresses aggravate the loss of hydraulic conductance, but there are no related impacts of drought and low-temperature stresses on the hydraulic system.
AméglioT, BodetC, LacointeA, CochardH (2002). Winter embolism, mechanisms of xylem hydraulic conductivity recovery and springtime growth patterns in walnut and peach trees Tree Physiology, 22, 1211-1220. [本文引用: 2]
AméglioT, CochardH, EwersFW (2001). Stem diameter variations and cold hardiness in walnut trees Journal of Experimental Botany, 52, 2135-2142. [本文引用: 2]
AméglioT, DecourteixM, AlvesG, ValentinV, SakrS, JulienJL, PetelG, GuilliotA, LacointeA (2004). Temperature effects on xylem sap osmolarity in walnut trees: Evidence for a vitalistic model of winter embolism repair Tree Physiology, 24, 785-793. [本文引用: 1]
AriasNS, BucciSJ, ScholzFG, GoldsteinG (2015). Freezing avoidance by supercooling in Olea europaea cultivars: The role of apoplastic water, solute content and cell wall rigidity Plant, Cell & Environment, 38, 2061-2070. [本文引用: 4]
AriasNS, ScholzFG, GoldsteinG, BucciSJ (2017). The cost of avoiding freezing in stems: Trade-off between xylem resistance to cavitation and supercooling capacity in woody plants Tree Physiology, 37, 1251-1262. [本文引用: 4]
BachofenC, WohlgemuthT, GhazoulJ, MoserB (2016). Cold temperature extremes during spring do not limit the range shift of Mediterranean pines into regions with intermittent frost Functional Ecology, 30, 856-865. [本文引用: 2]
BallMC, CannyMJ, HuangCX, HeadyRD (2004). Structural changes in acclimated and unacclimated leaves during freezing and thawing Functional Plant Biology, 31, 29-40. [本文引用: 2]
BlackmanCJ, BrodribbTJ, JordanGJ (2010). Leaf hydraulic vulnerability is related to conduit dimensions and drought resistance across a diverse range of woody angiosperms New Phytologist, 188, 1113-1123. [本文引用: 1]
BowlingDR, LoganBA, HufkensK, AubrechtDM, RichardsonAD, BurnsSP, AndereggWRL, BlankenPD, EirikssonDP (2018). Limitations to winter and spring photosynthesis of a Rocky Mountain subalpine forest Agricultural and Forest Meteorology, 252, 241-255. [本文引用: 3]
BrodersenCR, McElroneAJ (2013). Maintenance of xylem network transport capacity: A review of embolism repair in vascular plants Frontiers in Plant Science, 4, 108. DOI: 10.3389/fpls.2013.00108. [本文引用: 2]
BrodribbTJ, FeildTS, JordanGJ (2007). Leaf maximum photosynthetic rate and venation are linked by hydraulics Plant Physiology, 144, 1890-1898. [本文引用: 1]
BuchnerO, NeunerG (2011). Winter frost resistance of Pinus cembra measured in situ at the alpine timberline as affected by temperature conditions Tree Physiology, 31, 1217-1227. [本文引用: 1]
CaoKF, YangSJ, ZhangYJ, BrodribbTJ (2012). The maximum height of grasses is determined by roots Ecology Letters, 15, 666-672. [本文引用: 1]
CarlesS, LamhamediMS, StoweDC, BernierPY, VeilleuxL, MargolisHA (2011). Relationships between frost hardiness, root growth potential, and photosynthesis of nursery- grown white spruce seedlings Annals of Forest Science, 68, 1303-1313. [本文引用: 1]
Charra-VaskouK, BadelE, CharrierG, PonomarenkoA, BonhommeM, FoucatL, MayrS, AméglioT (2016). Cavitation and water fluxes driven by ice water potential in Juglans regia during freeze-thaw cycles Journal of Experimental Botany, 67, 739-750. [本文引用: 3]
CharrierG, Charra-VaskouK, KasugaJ, CochardH, MayrS, AméglioT (2014). Freeze-thaw stress: Effects of temperature on hydraulic conductivity and ultrasonic activity in ten woody angiosperms Plant Physiology, 164, 992-998. [本文引用: 1]
CharrierG, CochardH, AméglioT (2013). Evaluation of the impact of frost resistances on potential altitudinal limit of trees Tree Physiology, 33, 891-902. [本文引用: 3]
CharrierG, NgaoJ, SaudreauM, AméglioT (2015). Effects of environmental factors and management practices on microclimate, winter physiology, and frost resistance in trees Frontiers in Plant Science, 6, 259. DOI: 10.3389/fpls.?2015.??00259. [本文引用: 1]
CharrierG, NolfM, LeitingerG, Charra-VaskouK, LossoA, TappeinerU, AméglioT, MayrS (2017). Monitoring of freezing dynamics in trees: A simple phase shift causes complexity Plant Physiology, 173, 2196-2207. [本文引用: 5]
ChoatB, BrodieTW, CobbAR, ZwienieckiMA, HolbrookNM (2006). Direct measurements of intervessel pit membrane hydraulic resistance in two angiosperm tree species American Journal of Botany, 93, 993-1000. [本文引用: 1]
ChoatB, BrodribbTJ, BrodersenCR, DuursmaRA, LópezR, MedlynBE (2018). Triggers of tree mortality under drought Nature, 558, 531-539. [本文引用: 2]
ChoatB, JansenS, BrodribbTJ, CochardH, DelzonS, BhaskarR, BucciSJ, FeildTS, GleasonSM, HackeUG, JacobsenAL, LensF, MaheraliH, Martínez-VilaltaJ, MayrS, MencucciniM, MitchellPJ, NardiniA, PittermannJ, PrattRB, SperryJS, WestobyM, WrightIJ, ZanneAE (2012). Global convergence in the vulnerability of forests to drought Nature, 491, 752-755. [本文引用: 1]
ChoatB, MedekDE, StuartSA, Pasquet-KokJ, EgertonJJG, SalariH, SackL, BallMC (2011). Xylem traits mediate a trade-off between resistance to freeze-thaw-induced embolism and photosynthetic capacity in overwintering evergreens New Phytologist, 191, 996-1005. [本文引用: 5]
Christensen-DalsgaardKK, TyreeMT (2013). Does freezing and dynamic flexing of frozen branches impact the cavitation resistance of Malus domestica and the Populus clone walker? Oecologia, 173, 665-674. [本文引用: 2]
Christensen-DalsgaardKK, TyreeMT (2014). Frost fatigue and spring recovery of xylem vessels in three diffuse-porous trees in situ Plant, Cell & Environment, 37, 1074-1085. [本文引用: 2]
ClimentJ, Costa e SilvaF, ChambelMR, PardosM, AlmeidaMH (2009). Freezing injury in primary and secondary needles of Mediterranean pine species of contrasting ecological niches Annals of Forest Science, 66, 407. DOI: 10.1051/forest/2009016. [本文引用: 1]
CochardH, TyreeMT (1990). Xylem dysfunction in Quercus: Vessel sizes, tyloses, cavitation and seasonal changes in embolism Tree Physiology, 6, 393-407. [本文引用: 1]
DavisSD, SperryJS, HackeUG (1999). The relationship between xylem conduit diameter and cavitation caused by freezing American Journal of Botany, 86, 1367-1372. [本文引用: 1]
de SwaefT, HanssensJ, CornelisA, SteppeK (2013). Non-destructive estimation of root pressure using sap flow, stem diameter measurements and mechanistic modelling Annals of Botany, 111, 271-282. [本文引用: 1]
DomecJC (2011). Let’s not forget the critical role of surface tension in xylem water relations Tree Physiology, 31, 359-360. [本文引用: 1]
FengF, DingF, TyreeMT (2015). Investigations concerning cavitation and frost fatigue in clonal 84K poplar using high-resolution cavitron measurements Plant Physiology, 168, 144-155. [本文引用: 3]
Fernández-PérezL, Villar-SalvadorP, Martínez-VillataJ, TocaA, ZavalaMA (2018). Distribution of pines in the Iberian Peninsula agrees with species differences in foliage frost tolerance, not with vulnerability to freezing-induced xylem embolism Tree Physiology, 38, 507-516. [本文引用: 6]
FeildTS, BrodribbT (2001). Stem water transport and freeze-thaw xylem embolism in conifers and angiosperms in a Tasmanian treeline heath Oecologia, 127, 314-320. [本文引用: 8]
GleasonSM, BlackmanCJ, GleasonST, McCullohKA, OcheltreeTW, WestobyM (2018). Vessel scaling in evergreen angiosperm leaves conforms with Murray’s law and area-filling assumptions: Implications for plant size, leaf size and cold tolerance New Phytologist, 218, 1360-1370. [本文引用: 1]
GleasonSM, WestobyM, JansenS, ChoatB, HackeUG, PrattRB, BhaskarR, BrodribbTJ, BucciSJ, CaoKF, CochardH, DelzonS, DomecJC, FanZX, FeildTS, JacobsenAL, JohnsonDM, LensF, MaheraliH, Martínez-VilaltaJ, MayrS, McCullohKA, MencucciniM, MitchellPJ, MorrisH, NardiniA, PittermannJ, PlavcováL, SchreiberSG, SperryJS, WrightIJ, ZanneAE (2016). Weak tradeoff between xylem safety and xylem-specific hydraulic efficiency across the world’s woody plant species New Phytologist, 209, 123-136. [本文引用: 1]
GrandaE, ScoffoniC, Rubio-CasalAE, SackL, ValladaresF (2014). Leaf and stem physiological responses to summer and winter extremes of woody species across temperate ecosystems Oikos, 123, 1281-1290. [本文引用: 3]
HackeUG, LachenbruchB, PittermannJ, MayrS, DomecJC, SchultePJ (2015). The hydraulic architecture of conifers In: Hacke U ed. Functional and Ecological Xylem Anatomy. Springer, Cham, Switzerland. 39-75. [本文引用: 5]
HackeUG, SperryJS (2001). Functional and ecological xylem anatomy Perspectives in Plant Ecology, Evolution and Systematics, 4, 97-115. [本文引用: 3]
HackeUG, SpicerR, SchreiberSG, PlavcováL (2017). An ecophysiological and developmental perspective on variation in vessel diameter Plant, Cell & Environment, 40, 831-845. [本文引用: 5]
HajekP, LeuschnerC, HertelD, DelzonS, SchuldtB (2014). Trade-offs between xylem hydraulic properties, wood anatomy and yield in Populus Tree Physiology, 34, 744-756. [本文引用: 1]
HaoGY, HoffmannWA, ScholzFG, BucciSJ, MeinzerFC, FrancoAC, CaoKF, GoldsteinG (2008). Stem and leaf hydraulics of congeneric tree species from adjacent tropical savanna and forest ecosystems Oecologia, 155, 405-415. [本文引用: 1]
HaoGY, WheelerJK, HolbrookNM, GoldsteinG (2013). Investigating xylem embolism formation, refilling and water storage in tree trunks using frequency domain reflectometry Journal of Experimental Botany, 64, 2321-2332. [本文引用: 1]
HoermillerII, RuschhauptM, HeyerAG (2018). Mechanisms of frost resistance in Arabidopsis thaliana. Planta, 248, 827-835. [本文引用: 2]
HoffmanL, DaCostaM, EbdonJS, ZhaoJZ (2012). Effects of drought preconditioning on freezing tolerance of perennial ryegrass Environmental and Experimental Botany, 79, 11-20. [本文引用: 1]
HoffmannWA, MarchinRM, AbitP, LauOL (2011). Hydraulic failure and tree dieback are associated with high wood density in a temperate forest under extreme drought Global Change Biology, 17, 2731-2742. [本文引用: 1]
H?ltt?T, Dominguez CarrascoMDR, SalmonY, AaltoJ, VanhataloA, B?ckJ, LintunenA (2018). Water relations in silver birch during springtime: How is sap pressurised? Plant Biology, 20, 834-847. [本文引用: 1]
Jiménez-CastilloM, LuskCH (2013). Vascular performance of woody plants in a temperate rain forest: Lianas suffer higher levels of freeze-thaw embolism than associated trees Functional Ecology, 27, 403-412. [本文引用: 1]
JinY, WangCK, ZhouZH, LiZM (2016). Co-ordinated performance of leaf hydraulics and economics in 10 Chinese temperate tree species Functional Plant Biology, 43, 1082-1090. [本文引用: 2]
JinY, WangCK (2016). Leaf hydraulic traits and their trade-offs for nine Chinese temperate tree species with different wood properties Chinese Journal of Plant Ecology, 40, 702-710. [本文引用: 3]
JinY, WangCK, ZhouZH (2016). Mechanisms of xylem embolism repair in woody plants: Research progress and questions Chinese Journal of Plant Ecology, 40, 834-846. [本文引用: 1]
JohnsonDM, McCullohKA, WoodruffDR, MeinzerFC (2012). Hydraulic safety margins and embolism reversal in stems and leaves: Why are conifers and angiosperms so different? Plant Science, 195, 48-53. [本文引用: 1]
KongRS, HenryHAL (2016). Prior exposure to freezing stress enhances the survival and recovery of Poa pratensis exposed to severe drought American Journal of Botany, 103, 1890-1896. [本文引用: 1]
KreylingJ, SchmidS, AasG (2015). Cold tolerance of tree species is related to the climate of their native ranges Journal of Biogeography, 42, 156-166. [本文引用: 2]
KreylingJ, WiesenbergGLB, ThielD, WohlfartC, HuberG, WalterJ, JentschA, KonnertM, BeierkuhnleinC (2012). Cold hardiness of Pinus nigra Arnold as influenced by geographic origin, warming, and extreme summer drought Environmental and Experimental Botany, 78, 99-108. [本文引用: 1]
LemoineD, GranierA, CochardH (1999). Mechanism of freeze-induced embolism in Fagus sylvatica L Trees, 13, 206-210. [本文引用: 1]
LensF, SperryJS, ChristmanMA, ChoatB, RabaeyD, JansenS (2011). Testing hypotheses that link wood anatomy to cavitation resistance and hydraulic conductivity in the genus Acer New Phytologist, 190, 709-723. [本文引用: 1]
LensF, TixierA, CochardH, SperryJS, JansenS, HerbetteS (2013). Embolism resistance as a key mechanism to understand adaptive plant strategies Current Opinion in Plant Biology, 16, 287-292. [本文引用: 2]
LindforsL, H?ltt?T, LintunenA, Porcar-CastellA, NikinmaaE, JuurolaE (2015). Dynamics of leaf gas exchange, chlorophyll fluorescence and stem diameter changes during freezing and thawing of Scots pine seedlings Tree Physiology, 35, 1314-1324. [本文引用: 1]
LintunenA, H?ltt?T, KulmalaM (2013). Anatomical regulation of ice nucleation and cavitation helps trees to survive freezing and drought stress Scientific Reports, 3, 2031. DOI: 10.1038/srep02031. [本文引用: 1]
LintunenA, LindforsL, KolariP, JuurolaE, NikinmaaE, H?ltt?T (2014). Bursts of CO2 released during freezing offer a new perspective on avoidance of winter embolism in trees Annals of Botany, 114, 1711-1718. [本文引用: 1]
LintunenA, LindforsL, NikinmaaE, H?ltt?T (2016). Xylem diameter changes during osmotic stress, desiccation and freezing in Pinus sylvestris and Populus tremula Tree Physiology, 37, 491-500. [本文引用: 5]
LintunenA, MayrS, SalmonY, CochardH, H?ltt?T (2018). Drivers of apoplastic freezing in gymnosperm and angiosperm branches Ecology and Evolution, 8, 333-343. [本文引用: 5]
LossoA, NardiniA, D?monB, MayrS (2018). Xylem sap chemistry: Seasonal changes in timberline conifers Pinus cembra, Picea abies, and Larix decidua Biologia Plantarum, 62, 157-165. [本文引用: 1]
Martínez-VilaltaJ, SalaAN, AsensioD, GalianoL, HochG, PalacioS, PiperFI, LloretF (2016). Dynamics of non-structural carbohydrates in terrestrial plants: A global synthesis Ecological Monographs, 86, 495-516. [本文引用: 1]
MayrS, AméglioT (2016). Freezing stress in tree xylem Progress in Botany, 77, 381-414. [本文引用: 3]
MayrS, CochardH, AméglioT, KikutaSB (2007). Embolism formation during freezing in the wood of Picea abies Plant Physiology, 143, 60-67. [本文引用: 2]
MayrS, HackeU, SchmidP, SchwienbacherF, GruberA (2006). Frost drought in conifers at the alpine timberline: Xylem dysfunction and adaptations Ecology, 87, 3175-3185. [本文引用: 1]
MayrS, SchmidP, LaurJ, RosnerS, Charra-VaskouK, D?monB, HackeUG (2014). Uptake of water via branches helps timberline conifers refill embolized xylem in late winter Plant Physiology, 164, 1731-1740. [本文引用: 1]
MayrS, SchwienbacherF, BauerH (2003). Winter at the alpine timberline. Why does embolism occur in Norway spruce but not in stone pine? Plant Physiology, 131, 780-792. [本文引用: 1]
MayrS, SperryJS (2010). Freeze-thaw-induced embolism in Pinus contorta: Centrifuge experiments validate the ‘thaw-?expansion hypothesis’ but conflict with ultrasonic emission data New Phytologist, 185, 1016-1024. [本文引用: 1]
MayrS, WolfschwengerM, BauerH (2002). Winter-drought induced embolism in Norway spruce (Picea abies) at the Alpine timberline . Physiologia Plantarum, 115, 74-80. [本文引用: 1]
McCullyME, CannyMJ, HuangCX (2004). The management of extracellular ice by petioles of frost-resistant herbaceous plants Annals of Botany, 94, 665-674. [本文引用: 1]
McDowellN, PockmanWT, AllenCD, BreshearsDD, CobbN, KolbT, PlautJ, SperryJ, WestA, WilliamsDG, YepezEA (2008). Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytologist, 178, 719-739. [本文引用: 1]
MedeirosJS, DanielsonSC (2018). Renewed interest in whole-plant physiology sheds light on the complexity of plant stress response architecture Tree Physiology, 38, 503-506. [本文引用: 1]
MedeirosJS, PockmanWT (2011). Drought increases freezing tolerance of both leaves and xylem of Larrea tridentate. Plant, Cell & Environments, 34, 43-51. [本文引用: 1]
NardiniA, Lo GulloMA, SalleoS (2011). Refilling embolized xylem conduits: Is it a matter of phloem unloading? Plant Science, 180, 604-611. [本文引用: 1]
NardiniA, LuglioJ (2014). Leaf hydraulic capacity and drought vulnerability: Possible trade-offs and correlations with climate across three major biomes Functional Ecology, 28, 810-818. [本文引用: 2]
NardiniA, PedàG, La RoccaN (2012). Trade-offs between leaf hydraulic capacity and drought vulnerability: Morpho-?anatomical bases, carbon costs and ecological consequences New Phytologist, 196, 788-798. [本文引用: 1]
NiuCY, MeinzerFC, HaoGY (2017). Divergence in strategies for coping with winter embolism among co-occurring temperate tree species: The role of positive xylem pressure, wood type and tree stature Functional Ecology, 31, 1550-1560. [本文引用: 6]
PittermannJ, SperryJ (2003). Tracheid diameter is the key trait determining the extent of freezing-induced embolism in conifers Tree Physiology, 23, 907-914. [本文引用: 3]
PittermannJ, SperryJS (2006). Analysis of freeze-thaw embolism in conifers. The interaction between cavitation pressure and tracheid size Plant Physiology, 140, 374-382. [本文引用: 4]
PivovaroffAL, SackL, SantiagoLS (2014). Coordination of stem and leaf hydraulic conductance in southern California shrubs: A test of the hydraulic segmentation hypothesis New Phytologist, 203, 842-850. [本文引用: 1]
PloyetR, SolerM, CarochaV, LadouceN, AlvesA, RodriguesJ, HarvengtL, MarqueC, TeulièresC, Grima-PettenatiJ, MounetF (2018). Long cold exposure induces transcriptional and biochemical remodelling of xylem secondary cell wall in Eucalyptus Tree Physiology, 38, 409-422. [本文引用: 1]
PrendinAL, MayrS, BeikircherB, von ArxG, PetitG (2018). Xylem anatomical adjustments prioritize hydraulic efficiency over safety as Norway spruce trees grow taller Tree Physiology, 38, 1088-1097. [本文引用: 1]
PrestonJC, SandveSR (2013). Adaptation to seasonality and the winter freeze Frontiers in Plant Science, 4, 167. DOI: 10.3389/fpls.2013.00167. [本文引用: 2]
RasmussenDH, APDH (1972). Effect of solute on ice-solution interfacial free energy; calculation from measured homogeneous nucleation temperatures In: Jellinek HHG ed. Water Structure at the Water-polymer Interface. Springer, Boston. 126-145. [本文引用: 1]
ReichPB (2014). The world-wide “fast-slow” plant economics spectrum: A traits manifesto Journal of Ecology, 102, 275-301. [本文引用: 2]
ReichPB, SendallKM, StefanskiA, WeiXR, RichRL, MontgomeryRA (2016). Boreal and temperate trees show strong acclimation of respiration to warming Nature, 531, 633-636. [本文引用: 1]
RoratoDG, AraujoMM, TabaldiLA, TurchettoF, GriebelerAM, BerghettiáLP, BarbosaFM (2018). Tolerance and resilience of forest species to frost in restoration planting in southern Brazil Restoration Ecology, 26, 537-542. [本文引用: 1]
SalleoS, Lo GulloMA, TrifilòP, NardiniA (2004). New evidence for a role of vessel-associated cells and phloem in the rapid xylem refilling of cavitated stems of Laurus nobilis L Plant, Cell & Environment, 27, 1065-1076. [本文引用: 2]
SantiagoLS, De GuzmanME, BaralotoC, VogenbergJE, BrodieM, HéraultB, FortunelC, BonalD (2018). Coordination and trade-offs among hydraulic safety, efficiency and drought avoidance traits in Amazonian rainforest canopy tree species New Phytologist, 218, 1015-1024. [本文引用: 1]
SchreiberSG, HamannA, HackeUG, ThomasBR (2013). Sixteen years of winter stress: An assessment of cold hardiness, growth performance and survival of hybrid poplar clones at a boreal planting site Plant, Cell & Environment, 36, 419-428. [本文引用: 1]
SevantoS, SuniT, PumpanenJ, Gr?nholmT, KolariP, NikinmaaE, HariP, VesalaT (2006). Wintertime photosynthesis and water uptake in a boreal forest Tree Physiology, 26, 749-757. [本文引用: 1]
Sierra-AlmeidaA, Reyes-BahamondeC, CavieresLA (2016). Drought increases the freezing resistance of high-elevation plants of the Central Chilean Andes Oecologia, 181, 1011-1023. [本文引用: 1]
SperlingO, SecchiF, GodfreyJ, ZwienieckiMA (2017). Acclimation of Pistacia integerrima trees to frost in semi-arid environments depends on autumn’s drought Planta, 245, 671-679. [本文引用: 1]
SperryJS, HackeUG, PittermannJ (2006). Size and function in conifer tracheids and angiosperm vessels American Journal of Botany, 93, 1490-1500. [本文引用: 1]
SperryJS, LoveDM (2015). What plant hydraulics can tell us about responses to climate-change droughts New Phytologist, 207, 14-27. [本文引用: 1]
SperryJS, SullivanJEM (1992). Xylem embolism in response to freeze-thaw cycles and water stress in ring-porous, diffuse-porous, and conifer species Plant Physiology, 100, 605-613. [本文引用: 2]
StillerV, SperryJS (2002). Cavitation fatigue and its reversal in sunflower ( Helianthus annuus L.) Journal of Experimental Botany, 53, 1155-1161. [本文引用: 1]
TanedaH, TatenoM (2005). Hydraulic conductivity, photosynthesis and leaf water balance in six evergreen woody species from fall to winter Tree Physiology, 25, 299-306. [本文引用: 1]
TocaA, OlietJA, Villar-SalvadorP, MarotoJ, JacobsDF (2018). Species ecology determines the role of nitrogen nutrition in the frost tolerance of pine seedlings Tree Physiology, 38, 96-108. [本文引用: 1]
TyreeMT, EwersFW (1991). The hydraulic architecture of trees and other woody plants New Phytologist, 119, 345-360. [本文引用: 2]
VenturasMD, SperryJS, HackeUG (2017). Plant xylem hydraulics: What we understand, current research, and future challenges Journal of Integrative Plant Biology, 59, 356-389. [本文引用: 2]
VerhoevenA, García-PlazaolaJI, Fernández-MarínB (2018). Shared mechanisms of photoprotection in photosynthetic organisms tolerant to desiccation or to low temperature Environmental and Experimental Botany, 154, 66-79. [本文引用: 2]
VitasseY, LenzA, K?rnerC (2014). The interaction between freezing tolerance and phenology in temperate deciduous trees Frontiers in Plant Science, 5, 541. DOI: 10.3389/?fpls.2014.00541. [本文引用: 1]
WalterJ, JentschA, BeierkuhnleinC, KreylingJ (2013). Ecological stress memory and cross stress tolerance in plants in the face of climate extremes Environmental and Experimental Botany, 94, 3-8. [本文引用: 1]
WangAY, HanSJ, ZhangJH, WangM, YinXH, FangLD, YangD, HaoGY (2018). The interaction between nonstructural carbohydrate reserves and xylem hydraulics in Korean pine trees across an altitudinal gradient Tree Physiology, 38, 1792-1804. [本文引用: 2]
WesthoffM, SchneiderH, ZimmermannD, MimietzS, StinzingA, WegnerLH, KaiserW, KrohneG, ShirleyS, JakobP, BambergE, BentrupFW, ZimmermannU (2008). The mechanisms of refilling of xylem conduits and bleeding of tall birch during spring Plant Biology, 10, 604-623. [本文引用: 1]
WillickIR, GustaLV, FowlerDB, TaninoKK (2019). Ice segregation in the crown of winter cereals: Evidence for extraorgan and extratissue freezing Plant, Cell & Environment, 42, 701-716. [本文引用: 2]
WillickIR, TakahashiD, FowlerDB, UemuraM, TaninoKK (2018). Tissue-specific changes in apoplastic proteins and cell wall structure during cold acclimation of winter wheat crowns Journal of Experimental Botany, 69, 1221-1234. [本文引用: 3]
WillsonCJ, JacksonRB (2006). Xylem cavitation caused by drought and freezing stress in four co-occurring Juniperus species Physiologia Plantarum, 127, 374-382. [本文引用: 1]
YinXH, HaoGY (2018). Divergence between ring- and diffuse-?porous wood types in broadleaf trees of Changbai Mountains results in substantial differences in hydraulic traits Chinese Journal of Applied Ecology, 29, 352-360. [本文引用: 7]
YinXH, SterckF, HaoGY (2018). Divergent hydraulic strategies to cope with freezing in co-occurring temperate tree species with special reference to root and stem pressure generation New Phytologist, 219, 530-541. [本文引用: 1]
ZanneAE, TankDC, CornwellWK, EastmanJM, SmithSA, FitzJohnRG, McGlinnDJ, O’MearaBC, MolesAT, ReichPB, RoyerDL, SoltisDE, StevensPF, WestobyM, WrightIJ, AarssenL, BertinRI, CalaminusA, GovaertsR, HemmingsF, LeishmanMR, OleksynJ, SoltisPS, SwensonNG, WarmanL, BeaulieuJM (2014). Three keys to the radiation of angiosperms into freezing environments Nature, 506, 89-92. [本文引用: 2]
ZhangHY, WangCK, WangXC (2013). Comparison of concentrations of non-structural carbohydrates between new twigs and old branches for 12 temperate species Acta Ecologica Sinica, 33, 5675-5685. [本文引用: 1]
ZhangW, FengF, TyreeMT (2018). Seasonality of cavitation and frost fatigue in Acer mono Maxim Plant, Cell & Environment, 41, 1278-1286. [本文引用: 5]
ZhangYJ, BucciSJ, AriasNS, ScholzFG, HaoGY, CaoKF, GoldsteinG (2016). Freezing resistance in Patagonian woody shrubs: The role of cell wall elasticity and stem vessel size Tree Physiology, 36, 1007-1018. [本文引用: 3]
ZhuSD, LiRH, HePC, SiddiqZ, CaoKF, YeQ (2019). Large branch and leaf hydraulic safety margins in subtropical evergreen broadleaved forest Tree Physiology, 39, 1405-1415. [本文引用: 1]
ZimmermannMH (1983). Xylem Structure and the Ascent of Sap Spring, Berlin. [本文引用: 1]
Winter embolism, mechanisms of xylem hydraulic conductivity recovery and springtime growth patterns in walnut and peach trees 2 2002
... 栓塞是降低木质部内水分运输效率的关键因子(Choat et al., 2012; 金鹰等, 2016), 主要包括干旱栓塞和冻融栓塞(Améglio et al., 2002; Willson & Jackson, 2006; Venturas et al., 2017).两种栓塞形成的主要机理均为空气进入导管或管胞, 从而阻断木质部水柱, 形成栓塞(Sperry & Sullivan, 1992), 而不同之处在于引发空气进入的过程(Hacke & Sperry, 2001).根据内聚力-张力理论, 木质部导管内水分持续运输动力主要来自叶片蒸腾失水引起的拉力, 因此导管中的水分始终处于亚稳态(Sperry & Love, 2015).当蒸腾加剧和/或土壤水分供应不足而使木质部导管内负压超过一定阈值时, 就会造成其水柱断裂, 致使周围组织中的气体进入导管, 从而形成干旱栓塞(Choat et al., 2018).而冻融栓塞主要发生在木质部经历多次冻融循环的过程中(Sperry & Sullivan, 1992; Feild & Brodribb, 2001; Charra-?Vaskou et al., 2016).当木质部水分冰冻时, 水中原有溶解气体因在冰中的溶解度很低而逸散到导管中(Lintunen et al., 2014).而当冰冻融化时, 导管中的留存气体或者重新溶进水中(气泡压力大于大气压时, Hacke & Sperry, 2001), 或者由于冰冻压力的释放(冰融化后体积减小)等使气体在导管中扩大(Lemoine et al., 1999; Mayr et al., 2002; Pittermann & Sperry, 2006), 多次冻融交替之后, 导管中气体不断增加和扩张, 最终引发冻融栓塞, 该现象也被称为“融化-扩张假说” (Mayr & Sperry, 2010; Mayr & Améglio, 2016).干旱栓塞和冻融栓塞均会阻碍植物水分从土壤到叶片的运输, 从而降低叶片的光合速率(Brodribb et al., 2007; Reich, 2014; Jin et al., 2016), 导致植物生产力降低甚至死亡(McDowell et al., 2008; Hoffmann et al., 2011; McDowell, 2011; Choat et al., 2018). ...
... 修复是植物应对冻融栓塞的又一重要策略(Preston & Sandve, 2013; Wang et al., 2018).有研究表明, 采用正压(主要包括根压和茎压)恢复水力功能的植物可能具有更强的抗冷冻疲劳能力(即木质部冰冻后对水力功能丢失更敏感的现象)(Niu et al., 2017; Yin et al., 2018), 这是植物对冻融栓塞的一种主动修复.Hao等(2013)发现, 早春纸桦(Betula papyrifera)树干水分含量明显高于生长季, 表明根压在修复冻融栓塞导管的过程中具有重要作用.Améglio等(2002)报道, 冬季胡桃(Juglans regia) PLC可高达到100%, 但是由于冬季和春季茎压修复, 木质部冻融栓塞程度有一定的降低.另外, 近期研究表明, 能产生木质部正压的植物, 其抗冻融栓塞能力可能不受导管直径的影响(Niu et al., 2017).根压和茎压主要来源于糖类水解成的蔗糖被运输到栓塞导管中而产生渗透浓度梯度(Nardini et al., 2011; Cao et al., 2012), 从而使水分流向栓塞导管, 即渗透调节假说(金鹰等, 2016).两者的不同之处在于: 根压主要在无蒸腾流情况下产生作用(Swaef et al., 2013), 且修复栓塞作用较小(Salleo et al., 2004), 特别是对高大植物上层木质部栓塞的修复作用很小(Westhoff et al., 2008).而茎压产生的渗透梯度可能来自韧皮部的糖类(韧皮部卸载假说, Salleo et al., 2004; Hölttä et al., 2018), 因此在多数情况下均有茎压存在, 这也被称为“新的再冲水” (novel refilling)假说(Brodersen & McElrone, 2013; 金鹰等, 2016).Améglio等(2004)发现, 胡桃在深冬时茎中淀粉含量最低, 在发芽前茎中再次合成淀粉, 这不仅表明胡桃在冬季低温下进行淀粉和蔗糖的转换, 而且蔗糖累积是其冻融栓塞修复中的关键.张海燕等(2013)也发现, 12个温带树种老枝的淀粉含量变化具有相同趋势. ...
Stem diameter variations and cold hardiness in walnut trees 2 2001
... 冻的树干(Charrier et al., 2017)无法及时补充水分丢失(Bowling et al., 2018), 从而使植物叶片和小枝的木质部含水量降低.另外, 含水量变化还可能会引起质外体组织冰冻时的体积发生变化(Lintunen et al., 2016).低温胁迫使木质组织首先脱水收缩, 再经历液态水冰冻时的体积膨大(+9%)(Améglio et al., 2001).在冰冻前, 植物主动降低质外体含水量, 增加溶质浓度, 此时脱水引起的体积减小能够降低过冷却点(Lintunen et al., 2013; Willick et al., 2019).因此, 冰冻前含水量越低(体积越小), 植物对低温胁迫的忍耐力越强(Lintunen et al., 2018).当冰冻发生时, 由于水分由液态转为固态的体积增加, 对周围细胞壁产生机械压力, 植物需构建更强健的细胞壁(Zhang et al., 2016; Ployet et al., 2018)和改变细胞壁上蛋白组分(Willick et al., 2018)来抵御低温胁迫, 因此, 质外体较高的水分含量可能对植物产生更严重的低温损伤.研究表明, 木质部通过寒冷驯化 后其直径少量收缩(Améglio et al., 2001; Charra- Vaskou et al., 2016; Lintunen et al., 2016).例如, 欧洲赤松(Pinus sylvestris)和欧洲山杨(Populus tremula)木质部直径经寒冷驯化后分别收缩了0.5%和1.0%, 小于渗透胁迫和干旱胁迫引起的收缩量(Lintunen et al., 2016).此外, 植物通过寒冷驯化, 可以降低质外体的含水量, 并将水分转移到冰储存位点, 同时不同冰区之间形成隔离带(Willick et al., 2019), 从而尽可能减少低温损伤. ...
... )来抵御低温胁迫, 因此, 质外体较高的水分含量可能对植物产生更严重的低温损伤.研究表明, 木质部通过寒冷驯化 后其直径少量收缩(Améglio et al., 2001; Charra- Vaskou et al., 2016; Lintunen et al., 2016).例如, 欧洲赤松(Pinus sylvestris)和欧洲山杨(Populus tremula)木质部直径经寒冷驯化后分别收缩了0.5%和1.0%, 小于渗透胁迫和干旱胁迫引起的收缩量(Lintunen et al., 2016).此外, 植物通过寒冷驯化, 可以降低质外体的含水量, 并将水分转移到冰储存位点, 同时不同冰区之间形成隔离带(Willick et al., 2019), 从而尽可能减少低温损伤. ...
Temperature effects on xylem sap osmolarity in walnut trees: Evidence for a vitalistic model of winter embolism repair 1 2004
... 修复是植物应对冻融栓塞的又一重要策略(Preston & Sandve, 2013; Wang et al., 2018).有研究表明, 采用正压(主要包括根压和茎压)恢复水力功能的植物可能具有更强的抗冷冻疲劳能力(即木质部冰冻后对水力功能丢失更敏感的现象)(Niu et al., 2017; Yin et al., 2018), 这是植物对冻融栓塞的一种主动修复.Hao等(2013)发现, 早春纸桦(Betula papyrifera)树干水分含量明显高于生长季, 表明根压在修复冻融栓塞导管的过程中具有重要作用.Améglio等(2002)报道, 冬季胡桃(Juglans regia) PLC可高达到100%, 但是由于冬季和春季茎压修复, 木质部冻融栓塞程度有一定的降低.另外, 近期研究表明, 能产生木质部正压的植物, 其抗冻融栓塞能力可能不受导管直径的影响(Niu et al., 2017).根压和茎压主要来源于糖类水解成的蔗糖被运输到栓塞导管中而产生渗透浓度梯度(Nardini et al., 2011; Cao et al., 2012), 从而使水分流向栓塞导管, 即渗透调节假说(金鹰等, 2016).两者的不同之处在于: 根压主要在无蒸腾流情况下产生作用(Swaef et al., 2013), 且修复栓塞作用较小(Salleo et al., 2004), 特别是对高大植物上层木质部栓塞的修复作用很小(Westhoff et al., 2008).而茎压产生的渗透梯度可能来自韧皮部的糖类(韧皮部卸载假说, Salleo et al., 2004; Hölttä et al., 2018), 因此在多数情况下均有茎压存在, 这也被称为“新的再冲水” (novel refilling)假说(Brodersen & McElrone, 2013; 金鹰等, 2016).Améglio等(2004)发现, 胡桃在深冬时茎中淀粉含量最低, 在发芽前茎中再次合成淀粉, 这不仅表明胡桃在冬季低温下进行淀粉和蔗糖的转换, 而且蔗糖累积是其冻融栓塞修复中的关键.张海燕等(2013)也发现, 12个温带树种老枝的淀粉含量变化具有相同趋势. ...
Freezing avoidance by supercooling in Olea europaea cultivars: The role of apoplastic water, solute content and cell wall rigidity 4 2015
... 植物还可以通过木质部过冷却的方式避免低温及冰冻损伤(Arias et al., 2017).过冷却指植物使细胞中液体的冰点温度远低于液态水冰点的能力(Arias et al., 2015).Rasmussen和MacKenzie (1972)指出, 没有任何结晶核的纯水结冰温度低至-38.1 ℃.因此, 植物细胞内异质结晶核含量影响其过冷却点的高低(Losso et al., 2018).过冷却能力对植物应对突发性的冰冻事件(如热带植物遭遇突发性寒流)十分重要(Reyes-Díaz et al., 2006).Arias等(2015)发现, 在寒冷环境中驯化过的油橄榄(Olea europaea)栽培种, 其叶片冰点温度低至-13 ℃ (远低于环境最低温度), 而且有更高的叶密度和细胞壁刚性, 从而使其溶质渗透性和组织可塑性更高, 质外体水分含量更低; 进一步研究发现, 从夏季到冬季该树种茎最低冰点逐渐降低, 而且其木质部抗空穴化能力与叶片和茎的过冷却能力之间存在权衡(Arias et al., 2017). ...
Cold temperature extremes during spring do not limit the range shift of Mediterranean pines into regions with intermittent frost 2 2016
... 常绿植物保证叶片安全越冬对其之后的生长具有关键作用(Ball et al., 2004), 因此我们还可以通过分析叶片抗低温胁迫能力来评价植物的低温耐性.目前评价叶片抗低温胁迫能力的指标主要有: 叶片电解质渗出率(Climent et al., 2009; Hoermiller et al., 2018)、可溶性糖含量(Fernández-Pérez et al., 2018)、黎明前叶水势(Taneda & Tateno, 2005)、叶细胞弹性模量(细胞壁刚性越高, 细胞弹性模量越大)(Zhang et al., 2016)等.Fernández-Pérez等(2018)报道, 伊比利亚半岛6个松属树种的分布与其叶片低温忍耐能力有关, 生长在寒冷环境中的针叶的电解质渗出率和可溶性糖含量较低, 且黎明前叶水势对针叶低温损伤有影响.Martínez-Vilalta等(2016)整合分析全球尺度121个研究中177个物种非结构性碳(淀粉和可溶性糖)数据发现, 北方和温带生物区系物种叶片非结构性碳含量最高, 而且在入冬过程中逐步增加.Zhang等(2016)发现, 叶片细胞壁强度的增加, 降低了质外体冰冻对细胞膜的物理损伤, 使植物在较低的温度下得以存活.上述几个评价指标中, 电解质渗出率是最常用的评价植物组织损伤的指标(Charrier et al., 2013; Arias et al., 2015; Mayr & Améglio, 2016).通常采用电解质渗出率达50%时所对应的温度(LT50)来评价植物的低温抗性: LT50越低, 则低温抗性越强(Kreyling et al., 2015; Bachofen et al., 2016; Arias et al., 2017).例如, Arias等(2017)报道, 油橄榄的LT50从夏季到冬季呈现逐步降低的趋势, 这表明随着温度的降低, 通过不断驯化, 油橄榄的低温抗性逐步增强. ...
... 木质部低温抗性、干旱抗性和水力效率三者间是否存在多元权衡现象?目前对此问题的综合研究较少, 但已有研究指出, 干旱显著增加植物的低温抗性(Kreyling et al., 2012; Walter et al., 2013; Sierra-?Almeida et al., 2016; Sperling et al., 2017).这是因为植物通过驯化改变了细胞对脱水的忍耐能力(Buchner & Neuner, 2011), 其中包括基因表达的改变导致的细胞膜结构变化或者细胞化学成分变化(Verhoeven et al., 2018).例如, Medeiros和Pockman (2011)报道, 在-15 ℃环境中干旱处理使Larrea tridentate植株细胞死亡减少、叶留存量增加、水力导度和气体交换速率升高, 这表明干旱处理显著增加了其低温抗性.Hoffman等(2012)也发现, 干旱处理能够显著增加黑麦草(Lolium perenne)的低温抗性.然而, Kong和Henry (2016)研究发现, 早春低温处理的草地早熟禾(Poa pratensis)的抗旱性显著增加(生物量和存活率增加), 但夏季干旱处理并未增加其低温抗性, 这表明干旱处理对植物低温抗性无影响的现象也存在.同样, Bachofen等(2016)也发现, 干旱处理没有增加地中海松属3个树种的低温抗性, 而且推测该现象可能仅在短期极端条件下发生.Yin等(2018)同时对低温抗性、干旱抗性和水力效率进行研究发现, 抗干旱栓塞与水力效率存在权衡, 抗冷冻疲劳与水力效率间也存在权衡现象, 支持多元权衡现象.然而, 当植物同时面对干旱和低温胁迫时(如高山树线植物)(Mayr et al., 2007; Charrier et al., 2017), 多次冻融交替结合干旱胁迫可能引起PLC增加(Mayr et al., 2003; Charrier et al., 2014).Mayr等(2006)报道, 生长在林线处的针叶树种, 每个冬季经历至少100次冻融交替, 再加上干旱胁迫, 二者共同引起欧洲云杉(Picea abies) PLC高达100%.因此, 生长在低温和干旱胁迫共存环境中的植物, 其PLC可能更高, 间接表明三者间存在一定程度的权衡.综上所述, 我们构建了低温抗性、干旱抗性和水力效率之间多元权衡概念图(图2). ...
Structural changes in acclimated and unacclimated leaves during freezing and thawing 2 2004
... 常绿树种茎抗低温胁迫能力可能还与叶片低温抗性有关(Ball et al., 2004; Fernández-Pérez et al., 2018; Verhoeven et al., 2018).“脆弱性分割假说”指出, 植物面临胁迫时, 其叶片(或小枝)可能比其茎表现得更为脆弱, 通过牺牲 “造价低廉”的末端器官保护“造价昂贵”的茎干, 以避免栓塞发生(Zimmermann, 1983; Tyree & Ewers, 1991).已存在大量有关干旱胁迫下的“脆弱性分割假说”验证研究(Hao et al., 2008; Johnson et al., 2012; Pivovaroff et al., 2014; Zhu et al., 2019), 但还不清楚低温胁迫下是否也会发生该现象.Fernández-Pérez等(2018)发现, 6个松属(Pinus)树种的针叶低温抗性对其分布范围的预测能力大于茎干木质部抗冻融栓塞能力.在北半球温带森林中, 冬季留存叶片因日间光照而使其日间温度高于0 ℃, 从而可能延长光合作用的时间(Feild & Brodribb, 2001; Sevanto et al., 2006; Lindfors et al., 2015; Bowling et al., 2018), 生产的有机物可用于早春修复冻融栓塞和细胞损伤.不过, 此时树干仍处于冰冻状态, 导致叶片可能比茎经历更多次的冻融循环, 从而加剧叶片的冻融栓塞(Feild & Brodribb, 2001; Charra-Vaskou et al., 2016), 降低茎干的冻融栓塞程度, 在一定程度上对主茎起到保护作用(Charrier et al., 2017). ...
... 常绿植物保证叶片安全越冬对其之后的生长具有关键作用(Ball et al., 2004), 因此我们还可以通过分析叶片抗低温胁迫能力来评价植物的低温耐性.目前评价叶片抗低温胁迫能力的指标主要有: 叶片电解质渗出率(Climent et al., 2009; Hoermiller et al., 2018)、可溶性糖含量(Fernández-Pérez et al., 2018)、黎明前叶水势(Taneda & Tateno, 2005)、叶细胞弹性模量(细胞壁刚性越高, 细胞弹性模量越大)(Zhang et al., 2016)等.Fernández-Pérez等(2018)报道, 伊比利亚半岛6个松属树种的分布与其叶片低温忍耐能力有关, 生长在寒冷环境中的针叶的电解质渗出率和可溶性糖含量较低, 且黎明前叶水势对针叶低温损伤有影响.Martínez-Vilalta等(2016)整合分析全球尺度121个研究中177个物种非结构性碳(淀粉和可溶性糖)数据发现, 北方和温带生物区系物种叶片非结构性碳含量最高, 而且在入冬过程中逐步增加.Zhang等(2016)发现, 叶片细胞壁强度的增加, 降低了质外体冰冻对细胞膜的物理损伤, 使植物在较低的温度下得以存活.上述几个评价指标中, 电解质渗出率是最常用的评价植物组织损伤的指标(Charrier et al., 2013; Arias et al., 2015; Mayr & Améglio, 2016).通常采用电解质渗出率达50%时所对应的温度(LT50)来评价植物的低温抗性: LT50越低, 则低温抗性越强(Kreyling et al., 2015; Bachofen et al., 2016; Arias et al., 2017).例如, Arias等(2017)报道, 油橄榄的LT50从夏季到冬季呈现逐步降低的趋势, 这表明随着温度的降低, 通过不断驯化, 油橄榄的低温抗性逐步增强. ...
Leaf hydraulic vulnerability is related to conduit dimensions and drought resistance across a diverse range of woody angiosperms 1 2010
... 木质部的水力效率-安全权衡是植物适应环境和进化的结果(Blackman et al., 2010; Jin et al., 2016), 影响植物的竞争力和生产力(Nardini & Luglio, 2014).水力效率一般用植物水力导度表示, 而水力安全常用抗干旱栓塞能力(P50)表示(Santiago et al., 2018).Nardini等(2012)报道, 来自温带的6种植物(槭属(Acer)和栎属(Quercus)各3种)的水力导度、P50和比叶质量之间存在显著相关性, 支持水力效率-安全权衡, 且增加叶碳投入可以提高水力丢失的忍耐能力.Nardini和Luglio (2014)整合130个木本被子植物叶片水力数据后也发现存在叶水力效率-安全权衡关系, 而且生长在低降水量地区的植物的P50 (负值)和叶水力导度均较低.Gleason等(2016)综合分析335种被子植物和89种裸子植物的茎水力数据, 发现虽然水力导度和抗干旱栓塞能力之间存在弱相关性(支持水力效率-安全权衡), 但没有发现高效率和高安全性的植物.因此, 植物普遍存在不同程度的水力效率-安全权衡关系, 但其关系的紧密程度受导管特征(Lens et al., 2011; Hajek et al., 2014)、木质材性(金鹰和王传宽, 2016)、树高(Prendin et al., 2018)等因素的影响. ...
Limitations to winter and spring photosynthesis of a Rocky Mountain subalpine forest 3 2018
... 北方森林和温带森林几乎占全球森林总面积的1/3 (Reich et al., 2016), 其植物面临的最主要环境胁迫之一是冬季低温.低温使植物地上器官木质部中的水分逐步冻结(Niu et al., 2017), 所以植物如何最大程度减少低温胁迫引起的损伤对其安全越冬(Hacke et al., 2015; Kreyling et al., 2015; Charrier et al., 2017)、生长、生存以及分布均具有重要作用(Schreiber et al., 2013; Charrier et al., 2015; Niu et al., 2017).低温时期冻融诱导形成的栓塞是低温胁迫对植物产生损伤的重要因素(Zanne et al., 2014; Zhang et al., 2018).许多研究发现, 植物抗冻融栓塞能力不仅影响其在低温时期的损伤程度和碳消耗(Bowling et al., 2018; Yin et al., 2018), 还显著影响随后生长季中的水分供应能力(冷冻疲劳, Feng et al., 2015; Zhang et al., 2018).但与干旱栓塞相比, 目前对木质部冻融栓塞的形成以及植物响应和调节机制的理解不足(Choat et al., 2011; Yin et al., 2018).为了推进该领域的研究, 本文先综述木质部冻融栓塞的形成机制和植物的逃避、忍耐、修复等3种冻融栓塞的应对策略, 然后总结植物低温抗性的生理表现、评价和影响因子, 在此基础上讨论低温抗性、干旱抗性和水力效率之间的多元权衡关系, 最后提出今后该领域中的优先研究问题. ...
... 常绿树种茎抗低温胁迫能力可能还与叶片低温抗性有关(Ball et al., 2004; Fernández-Pérez et al., 2018; Verhoeven et al., 2018).“脆弱性分割假说”指出, 植物面临胁迫时, 其叶片(或小枝)可能比其茎表现得更为脆弱, 通过牺牲 “造价低廉”的末端器官保护“造价昂贵”的茎干, 以避免栓塞发生(Zimmermann, 1983; Tyree & Ewers, 1991).已存在大量有关干旱胁迫下的“脆弱性分割假说”验证研究(Hao et al., 2008; Johnson et al., 2012; Pivovaroff et al., 2014; Zhu et al., 2019), 但还不清楚低温胁迫下是否也会发生该现象.Fernández-Pérez等(2018)发现, 6个松属(Pinus)树种的针叶低温抗性对其分布范围的预测能力大于茎干木质部抗冻融栓塞能力.在北半球温带森林中, 冬季留存叶片因日间光照而使其日间温度高于0 ℃, 从而可能延长光合作用的时间(Feild & Brodribb, 2001; Sevanto et al., 2006; Lindfors et al., 2015; Bowling et al., 2018), 生产的有机物可用于早春修复冻融栓塞和细胞损伤.不过, 此时树干仍处于冰冻状态, 导致叶片可能比茎经历更多次的冻融循环, 从而加剧叶片的冻融栓塞(Feild & Brodribb, 2001; Charra-Vaskou et al., 2016), 降低茎干的冻融栓塞程度, 在一定程度上对主茎起到保护作用(Charrier et al., 2017). ...
... 冻的树干(Charrier et al., 2017)无法及时补充水分丢失(Bowling et al., 2018), 从而使植物叶片和小枝的木质部含水量降低.另外, 含水量变化还可能会引起质外体组织冰冻时的体积发生变化(Lintunen et al., 2016).低温胁迫使木质组织首先脱水收缩, 再经历液态水冰冻时的体积膨大(+9%)(Améglio et al., 2001).在冰冻前, 植物主动降低质外体含水量, 增加溶质浓度, 此时脱水引起的体积减小能够降低过冷却点(Lintunen et al., 2013; Willick et al., 2019).因此, 冰冻前含水量越低(体积越小), 植物对低温胁迫的忍耐力越强(Lintunen et al., 2018).当冰冻发生时, 由于水分由液态转为固态的体积增加, 对周围细胞壁产生机械压力, 植物需构建更强健的细胞壁(Zhang et al., 2016; Ployet et al., 2018)和改变细胞壁上蛋白组分(Willick et al., 2018)来抵御低温胁迫, 因此, 质外体较高的水分含量可能对植物产生更严重的低温损伤.研究表明, 木质部通过寒冷驯化 后其直径少量收缩(Améglio et al., 2001; Charra- Vaskou et al., 2016; Lintunen et al., 2016).例如, 欧洲赤松(Pinus sylvestris)和欧洲山杨(Populus tremula)木质部直径经寒冷驯化后分别收缩了0.5%和1.0%, 小于渗透胁迫和干旱胁迫引起的收缩量(Lintunen et al., 2016).此外, 植物通过寒冷驯化, 可以降低质外体的含水量, 并将水分转移到冰储存位点, 同时不同冰区之间形成隔离带(Willick et al., 2019), 从而尽可能减少低温损伤. ...
Maintenance of xylem network transport capacity: A review of embolism repair in vascular plants 2 2013
... 常绿植物保证叶片安全越冬对其之后的生长具有关键作用(Ball et al., 2004), 因此我们还可以通过分析叶片抗低温胁迫能力来评价植物的低温耐性.目前评价叶片抗低温胁迫能力的指标主要有: 叶片电解质渗出率(Climent et al., 2009; Hoermiller et al., 2018)、可溶性糖含量(Fernández-Pérez et al., 2018)、黎明前叶水势(Taneda & Tateno, 2005)、叶细胞弹性模量(细胞壁刚性越高, 细胞弹性模量越大)(Zhang et al., 2016)等.Fernández-Pérez等(2018)报道, 伊比利亚半岛6个松属树种的分布与其叶片低温忍耐能力有关, 生长在寒冷环境中的针叶的电解质渗出率和可溶性糖含量较低, 且黎明前叶水势对针叶低温损伤有影响.Martínez-Vilalta等(2016)整合分析全球尺度121个研究中177个物种非结构性碳(淀粉和可溶性糖)数据发现, 北方和温带生物区系物种叶片非结构性碳含量最高, 而且在入冬过程中逐步增加.Zhang等(2016)发现, 叶片细胞壁强度的增加, 降低了质外体冰冻对细胞膜的物理损伤, 使植物在较低的温度下得以存活.上述几个评价指标中, 电解质渗出率是最常用的评价植物组织损伤的指标(Charrier et al., 2013; Arias et al., 2015; Mayr & Améglio, 2016).通常采用电解质渗出率达50%时所对应的温度(LT50)来评价植物的低温抗性: LT50越低, 则低温抗性越强(Kreyling et al., 2015; Bachofen et al., 2016; Arias et al., 2017).例如, Arias等(2017)报道, 油橄榄的LT50从夏季到冬季呈现逐步降低的趋势, 这表明随着温度的降低, 通过不断驯化, 油橄榄的低温抗性逐步增强. ...
Effects of environmental factors and management practices on microclimate, winter physiology, and frost resistance in trees 1 2015
... 北方森林和温带森林几乎占全球森林总面积的1/3 (Reich et al., 2016), 其植物面临的最主要环境胁迫之一是冬季低温.低温使植物地上器官木质部中的水分逐步冻结(Niu et al., 2017), 所以植物如何最大程度减少低温胁迫引起的损伤对其安全越冬(Hacke et al., 2015; Kreyling et al., 2015; Charrier et al., 2017)、生长、生存以及分布均具有重要作用(Schreiber et al., 2013; Charrier et al., 2015; Niu et al., 2017).低温时期冻融诱导形成的栓塞是低温胁迫对植物产生损伤的重要因素(Zanne et al., 2014; Zhang et al., 2018).许多研究发现, 植物抗冻融栓塞能力不仅影响其在低温时期的损伤程度和碳消耗(Bowling et al., 2018; Yin et al., 2018), 还显著影响随后生长季中的水分供应能力(冷冻疲劳, Feng et al., 2015; Zhang et al., 2018).但与干旱栓塞相比, 目前对木质部冻融栓塞的形成以及植物响应和调节机制的理解不足(Choat et al., 2011; Yin et al., 2018).为了推进该领域的研究, 本文先综述木质部冻融栓塞的形成机制和植物的逃避、忍耐、修复等3种冻融栓塞的应对策略, 然后总结植物低温抗性的生理表现、评价和影响因子, 在此基础上讨论低温抗性、干旱抗性和水力效率之间的多元权衡关系, 最后提出今后该领域中的优先研究问题. ...
Monitoring of freezing dynamics in trees: A simple phase shift causes complexity 5 2017
... 北方森林和温带森林几乎占全球森林总面积的1/3 (Reich et al., 2016), 其植物面临的最主要环境胁迫之一是冬季低温.低温使植物地上器官木质部中的水分逐步冻结(Niu et al., 2017), 所以植物如何最大程度减少低温胁迫引起的损伤对其安全越冬(Hacke et al., 2015; Kreyling et al., 2015; Charrier et al., 2017)、生长、生存以及分布均具有重要作用(Schreiber et al., 2013; Charrier et al., 2015; Niu et al., 2017).低温时期冻融诱导形成的栓塞是低温胁迫对植物产生损伤的重要因素(Zanne et al., 2014; Zhang et al., 2018).许多研究发现, 植物抗冻融栓塞能力不仅影响其在低温时期的损伤程度和碳消耗(Bowling et al., 2018; Yin et al., 2018), 还显著影响随后生长季中的水分供应能力(冷冻疲劳, Feng et al., 2015; Zhang et al., 2018).但与干旱栓塞相比, 目前对木质部冻融栓塞的形成以及植物响应和调节机制的理解不足(Choat et al., 2011; Yin et al., 2018).为了推进该领域的研究, 本文先综述木质部冻融栓塞的形成机制和植物的逃避、忍耐、修复等3种冻融栓塞的应对策略, 然后总结植物低温抗性的生理表现、评价和影响因子, 在此基础上讨论低温抗性、干旱抗性和水力效率之间的多元权衡关系, 最后提出今后该领域中的优先研究问题. ...
... 常绿树种茎抗低温胁迫能力可能还与叶片低温抗性有关(Ball et al., 2004; Fernández-Pérez et al., 2018; Verhoeven et al., 2018).“脆弱性分割假说”指出, 植物面临胁迫时, 其叶片(或小枝)可能比其茎表现得更为脆弱, 通过牺牲 “造价低廉”的末端器官保护“造价昂贵”的茎干, 以避免栓塞发生(Zimmermann, 1983; Tyree & Ewers, 1991).已存在大量有关干旱胁迫下的“脆弱性分割假说”验证研究(Hao et al., 2008; Johnson et al., 2012; Pivovaroff et al., 2014; Zhu et al., 2019), 但还不清楚低温胁迫下是否也会发生该现象.Fernández-Pérez等(2018)发现, 6个松属(Pinus)树种的针叶低温抗性对其分布范围的预测能力大于茎干木质部抗冻融栓塞能力.在北半球温带森林中, 冬季留存叶片因日间光照而使其日间温度高于0 ℃, 从而可能延长光合作用的时间(Feild & Brodribb, 2001; Sevanto et al., 2006; Lindfors et al., 2015; Bowling et al., 2018), 生产的有机物可用于早春修复冻融栓塞和细胞损伤.不过, 此时树干仍处于冰冻状态, 导致叶片可能比茎经历更多次的冻融循环, 从而加剧叶片的冻融栓塞(Feild & Brodribb, 2001; Charra-Vaskou et al., 2016), 降低茎干的冻融栓塞程度, 在一定程度上对主茎起到保护作用(Charrier et al., 2017). ...
... 冻的树干(Charrier et al., 2017)无法及时补充水分丢失(Bowling et al., 2018), 从而使植物叶片和小枝的木质部含水量降低.另外, 含水量变化还可能会引起质外体组织冰冻时的体积发生变化(Lintunen et al., 2016).低温胁迫使木质组织首先脱水收缩, 再经历液态水冰冻时的体积膨大(+9%)(Améglio et al., 2001).在冰冻前, 植物主动降低质外体含水量, 增加溶质浓度, 此时脱水引起的体积减小能够降低过冷却点(Lintunen et al., 2013; Willick et al., 2019).因此, 冰冻前含水量越低(体积越小), 植物对低温胁迫的忍耐力越强(Lintunen et al., 2018).当冰冻发生时, 由于水分由液态转为固态的体积增加, 对周围细胞壁产生机械压力, 植物需构建更强健的细胞壁(Zhang et al., 2016; Ployet et al., 2018)和改变细胞壁上蛋白组分(Willick et al., 2018)来抵御低温胁迫, 因此, 质外体较高的水分含量可能对植物产生更严重的低温损伤.研究表明, 木质部通过寒冷驯化 后其直径少量收缩(Améglio et al., 2001; Charra- Vaskou et al., 2016; Lintunen et al., 2016).例如, 欧洲赤松(Pinus sylvestris)和欧洲山杨(Populus tremula)木质部直径经寒冷驯化后分别收缩了0.5%和1.0%, 小于渗透胁迫和干旱胁迫引起的收缩量(Lintunen et al., 2016).此外, 植物通过寒冷驯化, 可以降低质外体的含水量, 并将水分转移到冰储存位点, 同时不同冰区之间形成隔离带(Willick et al., 2019), 从而尽可能减少低温损伤. ...
... 木质部低温抗性、干旱抗性和水力效率三者间是否存在多元权衡现象?目前对此问题的综合研究较少, 但已有研究指出, 干旱显著增加植物的低温抗性(Kreyling et al., 2012; Walter et al., 2013; Sierra-?Almeida et al., 2016; Sperling et al., 2017).这是因为植物通过驯化改变了细胞对脱水的忍耐能力(Buchner & Neuner, 2011), 其中包括基因表达的改变导致的细胞膜结构变化或者细胞化学成分变化(Verhoeven et al., 2018).例如, Medeiros和Pockman (2011)报道, 在-15 ℃环境中干旱处理使Larrea tridentate植株细胞死亡减少、叶留存量增加、水力导度和气体交换速率升高, 这表明干旱处理显著增加了其低温抗性.Hoffman等(2012)也发现, 干旱处理能够显著增加黑麦草(Lolium perenne)的低温抗性.然而, Kong和Henry (2016)研究发现, 早春低温处理的草地早熟禾(Poa pratensis)的抗旱性显著增加(生物量和存活率增加), 但夏季干旱处理并未增加其低温抗性, 这表明干旱处理对植物低温抗性无影响的现象也存在.同样, Bachofen等(2016)也发现, 干旱处理没有增加地中海松属3个树种的低温抗性, 而且推测该现象可能仅在短期极端条件下发生.Yin等(2018)同时对低温抗性、干旱抗性和水力效率进行研究发现, 抗干旱栓塞与水力效率存在权衡, 抗冷冻疲劳与水力效率间也存在权衡现象, 支持多元权衡现象.然而, 当植物同时面对干旱和低温胁迫时(如高山树线植物)(Mayr et al., 2007; Charrier et al., 2017), 多次冻融交替结合干旱胁迫可能引起PLC增加(Mayr et al., 2003; Charrier et al., 2014).Mayr等(2006)报道, 生长在林线处的针叶树种, 每个冬季经历至少100次冻融交替, 再加上干旱胁迫, 二者共同引起欧洲云杉(Picea abies) PLC高达100%.因此, 生长在低温和干旱胁迫共存环境中的植物, 其PLC可能更高, 间接表明三者间存在一定程度的权衡.综上所述, 我们构建了低温抗性、干旱抗性和水力效率之间多元权衡概念图(图2). ...
Direct measurements of intervessel pit membrane hydraulic resistance in two angiosperm tree species 1 2006
... 形成的气泡更小, 而且小气泡更容易在融化期重新溶解进入水中(Hacke et al., 2015).Pittermann和Sperry等(2006)报道, 管胞越窄的裸子树种气泡形成的空穴化越小, 并且得出冻融栓塞引起的木质部负压随着管胞直径的增加而增加.之后, Choat等(2011)对11种被子植物的研究也发现, 导管直径越大、长度越长的植物, 冻融引起的水力导度丢失量(PLC)越多、造成的栓塞越大.这可能是因为直径较大的导管具有更少的单位长度导管末端, 造成其纹孔数量减少、栓塞抗性降低(Choat et al., 2006).因此, 构建直径更小的导管(或管胞), 是植物应对冻融栓塞最直接的方式(Pittermann & Sperry, 2006; Jiménez-Castillo & Lusk, 2013; Hacke et al., 2017; Gleason et al., 2018). ...
Triggers of tree mortality under drought 2 2018
... 栓塞是降低木质部内水分运输效率的关键因子(Choat et al., 2012; 金鹰等, 2016), 主要包括干旱栓塞和冻融栓塞(Améglio et al., 2002; Willson & Jackson, 2006; Venturas et al., 2017).两种栓塞形成的主要机理均为空气进入导管或管胞, 从而阻断木质部水柱, 形成栓塞(Sperry & Sullivan, 1992), 而不同之处在于引发空气进入的过程(Hacke & Sperry, 2001).根据内聚力-张力理论, 木质部导管内水分持续运输动力主要来自叶片蒸腾失水引起的拉力, 因此导管中的水分始终处于亚稳态(Sperry & Love, 2015).当蒸腾加剧和/或土壤水分供应不足而使木质部导管内负压超过一定阈值时, 就会造成其水柱断裂, 致使周围组织中的气体进入导管, 从而形成干旱栓塞(Choat et al., 2018).而冻融栓塞主要发生在木质部经历多次冻融循环的过程中(Sperry & Sullivan, 1992; Feild & Brodribb, 2001; Charra-?Vaskou et al., 2016).当木质部水分冰冻时, 水中原有溶解气体因在冰中的溶解度很低而逸散到导管中(Lintunen et al., 2014).而当冰冻融化时, 导管中的留存气体或者重新溶进水中(气泡压力大于大气压时, Hacke & Sperry, 2001), 或者由于冰冻压力的释放(冰融化后体积减小)等使气体在导管中扩大(Lemoine et al., 1999; Mayr et al., 2002; Pittermann & Sperry, 2006), 多次冻融交替之后, 导管中气体不断增加和扩张, 最终引发冻融栓塞, 该现象也被称为“融化-扩张假说” (Mayr & Sperry, 2010; Mayr & Améglio, 2016).干旱栓塞和冻融栓塞均会阻碍植物水分从土壤到叶片的运输, 从而降低叶片的光合速率(Brodribb et al., 2007; Reich, 2014; Jin et al., 2016), 导致植物生产力降低甚至死亡(McDowell et al., 2008; Hoffmann et al., 2011; McDowell, 2011; Choat et al., 2018). ...
... ; Choat et al., 2018). ...
Global convergence in the vulnerability of forests to drought 1 2012
... 栓塞是降低木质部内水分运输效率的关键因子(Choat et al., 2012; 金鹰等, 2016), 主要包括干旱栓塞和冻融栓塞(Améglio et al., 2002; Willson & Jackson, 2006; Venturas et al., 2017).两种栓塞形成的主要机理均为空气进入导管或管胞, 从而阻断木质部水柱, 形成栓塞(Sperry & Sullivan, 1992), 而不同之处在于引发空气进入的过程(Hacke & Sperry, 2001).根据内聚力-张力理论, 木质部导管内水分持续运输动力主要来自叶片蒸腾失水引起的拉力, 因此导管中的水分始终处于亚稳态(Sperry & Love, 2015).当蒸腾加剧和/或土壤水分供应不足而使木质部导管内负压超过一定阈值时, 就会造成其水柱断裂, 致使周围组织中的气体进入导管, 从而形成干旱栓塞(Choat et al., 2018).而冻融栓塞主要发生在木质部经历多次冻融循环的过程中(Sperry & Sullivan, 1992; Feild & Brodribb, 2001; Charra-?Vaskou et al., 2016).当木质部水分冰冻时, 水中原有溶解气体因在冰中的溶解度很低而逸散到导管中(Lintunen et al., 2014).而当冰冻融化时, 导管中的留存气体或者重新溶进水中(气泡压力大于大气压时, Hacke & Sperry, 2001), 或者由于冰冻压力的释放(冰融化后体积减小)等使气体在导管中扩大(Lemoine et al., 1999; Mayr et al., 2002; Pittermann & Sperry, 2006), 多次冻融交替之后, 导管中气体不断增加和扩张, 最终引发冻融栓塞, 该现象也被称为“融化-扩张假说” (Mayr & Sperry, 2010; Mayr & Améglio, 2016).干旱栓塞和冻融栓塞均会阻碍植物水分从土壤到叶片的运输, 从而降低叶片的光合速率(Brodribb et al., 2007; Reich, 2014; Jin et al., 2016), 导致植物生产力降低甚至死亡(McDowell et al., 2008; Hoffmann et al., 2011; McDowell, 2011; Choat et al., 2018). ...
Xylem traits mediate a trade-off between resistance to freeze-thaw-induced embolism and photosynthetic capacity in overwintering evergreens 5 2011
... 北方森林和温带森林几乎占全球森林总面积的1/3 (Reich et al., 2016), 其植物面临的最主要环境胁迫之一是冬季低温.低温使植物地上器官木质部中的水分逐步冻结(Niu et al., 2017), 所以植物如何最大程度减少低温胁迫引起的损伤对其安全越冬(Hacke et al., 2015; Kreyling et al., 2015; Charrier et al., 2017)、生长、生存以及分布均具有重要作用(Schreiber et al., 2013; Charrier et al., 2015; Niu et al., 2017).低温时期冻融诱导形成的栓塞是低温胁迫对植物产生损伤的重要因素(Zanne et al., 2014; Zhang et al., 2018).许多研究发现, 植物抗冻融栓塞能力不仅影响其在低温时期的损伤程度和碳消耗(Bowling et al., 2018; Yin et al., 2018), 还显著影响随后生长季中的水分供应能力(冷冻疲劳, Feng et al., 2015; Zhang et al., 2018).但与干旱栓塞相比, 目前对木质部冻融栓塞的形成以及植物响应和调节机制的理解不足(Choat et al., 2011; Yin et al., 2018).为了推进该领域的研究, 本文先综述木质部冻融栓塞的形成机制和植物的逃避、忍耐、修复等3种冻融栓塞的应对策略, 然后总结植物低温抗性的生理表现、评价和影响因子, 在此基础上讨论低温抗性、干旱抗性和水力效率之间的多元权衡关系, 最后提出今后该领域中的优先研究问题. ...
... 与动物不同, 植物的固定性使得其更多地采用“被动”的忍耐策略, 例如不同的木质结构、调整组织水分含量和溶质含量等, 以减少低温损伤(Mayr et al., 2014; Arias et al., 2015; Lintunen et al., 2018).被子植物和裸子植物解剖结构的差异, 使二者对低温胁迫的忍耐能力不同(Pittermann & Sperry, 2003; Lintunen et al., 2016).被子植物的导管比裸子植物的管胞结构具有更高的水力效率, 但是其木质部抗冻融栓塞能力较低(Feild & Brodribb, 2001; Lintunen et al., 2018).Davis等(1999)报道, 当导管直径大于40 μm时, 一次冻融交替使木质部栓塞程度高达95%; Feild和Brodribb (2001)也发现, 常绿针叶植物和导管较小的被子植物经历一次冻融交替后, 其茎水力导度降低最少(几乎为0), 而其他被子植物则平均降低18%-70%.可见, 被子植物木质部忍耐低温能力远低于裸子植物.植物经过多次冻融交替后, 导管或周围组织中产生的气泡可能在整个冬季留存, 直到春季才能重新进入融化水中或者气泡扩张形成栓塞(Hacke & Sperry, 2001; Venturas et al., 2017; Medeiros & Danielson, 2018), 所以导管内水分含量直接影响植物冻融栓塞程度(Choat et al., 2011).而水分含量与导管直径紧密相关(Lens et al., 2013).因此构建直径更小的导管是最直接增加抗冻融栓塞能力的方法.裸子植物直径较小的管胞结构能够在很大程度上减少冻融栓塞的形成, 不仅因为在管胞中 ...
... 形成的气泡更小, 而且小气泡更容易在融化期重新溶解进入水中(Hacke et al., 2015).Pittermann和Sperry等(2006)报道, 管胞越窄的裸子树种气泡形成的空穴化越小, 并且得出冻融栓塞引起的木质部负压随着管胞直径的增加而增加.之后, Choat等(2011)对11种被子植物的研究也发现, 导管直径越大、长度越长的植物, 冻融引起的水力导度丢失量(PLC)越多、造成的栓塞越大.这可能是因为直径较大的导管具有更少的单位长度导管末端, 造成其纹孔数量减少、栓塞抗性降低(Choat et al., 2006).因此, 构建直径更小的导管(或管胞), 是植物应对冻融栓塞最直接的方式(Pittermann & Sperry, 2006; Jiménez-Castillo & Lusk, 2013; Hacke et al., 2017; Gleason et al., 2018). ...
... 只有当Px小于P*时, 气泡才有可能扩大并形成空穴.由公式(1)可知, 木质部导管越大, 越容易形成直径较大的气泡, 气泡压力越小, 在较低张力下气泡越可能扩大, 因而对冻融栓塞的抵抗更加脆弱(Feild & Brodribb, 2001; Choat et al., 2011; Hacke et al., 2017).裸子植物管胞直径一般比被子植物导管直径小(Hacke et al., 2017), 因此裸子植物的抗冻融栓塞能力较强.但在管道直径相同时, 裸子植物与被子植物具有相似的抗冻融栓塞能力(Pittermann & Sperry, 2003, 2006; Hacke et al., 2015). ...
Investigations concerning cavitation and frost fatigue in clonal 84K poplar using high-resolution cavitron measurements 3 2015
... 北方森林和温带森林几乎占全球森林总面积的1/3 (Reich et al., 2016), 其植物面临的最主要环境胁迫之一是冬季低温.低温使植物地上器官木质部中的水分逐步冻结(Niu et al., 2017), 所以植物如何最大程度减少低温胁迫引起的损伤对其安全越冬(Hacke et al., 2015; Kreyling et al., 2015; Charrier et al., 2017)、生长、生存以及分布均具有重要作用(Schreiber et al., 2013; Charrier et al., 2015; Niu et al., 2017).低温时期冻融诱导形成的栓塞是低温胁迫对植物产生损伤的重要因素(Zanne et al., 2014; Zhang et al., 2018).许多研究发现, 植物抗冻融栓塞能力不仅影响其在低温时期的损伤程度和碳消耗(Bowling et al., 2018; Yin et al., 2018), 还显著影响随后生长季中的水分供应能力(冷冻疲劳, Feng et al., 2015; Zhang et al., 2018).但与干旱栓塞相比, 目前对木质部冻融栓塞的形成以及植物响应和调节机制的理解不足(Choat et al., 2011; Yin et al., 2018).为了推进该领域的研究, 本文先综述木质部冻融栓塞的形成机制和植物的逃避、忍耐、修复等3种冻融栓塞的应对策略, 然后总结植物低温抗性的生理表现、评价和影响因子, 在此基础上讨论低温抗性、干旱抗性和水力效率之间的多元权衡关系, 最后提出今后该领域中的优先研究问题. ...
... 木质部结构是影响植物忍耐低温胁迫的主要因素(Hacke et al., 2015, 2017; Lintunen et al., 2018).被子植物采用多个细胞死亡后相互连接构成的导管进行水分运输, 长度在1 mm到几米不等; 裸子植物则依靠单个细胞伸长形成的管胞进行水分运输, 长度在2-6 mm之间(Tyree & Ewers, 1991), 而且管胞直径一般小于导管(Sperry et al., 2006).这种木质部结构差异导致裸子植物和被子植物响应冻融栓塞时产生显著PLC差异(Feild & Brodribb, 2001; Feng et al., 2015; Fernández-Pérez et al., 2018).以往研究表明, 植物在入冬前PLC逐步增加(Zhang et al., 2018), 而导管或管胞的直径差异是植物PLC不同的主要原因(Pittermann & Sperry, 2003; Hacke et al., 2015, 2017).在所有季节中, 裸子植物的PLC均小于被子植物(Granda et al., 2014).例如, 被子植物葡萄牙栎(Quercus faginea)抗空穴化能力较差, 夏季PLC达到80%, 冬季进一步达到90%, 而裸子植物香桧(Juniperus thurifera)和欧洲黑松(Pinus nigra)夏季PLC几乎为0, 冬季仅增加到3%-9% (Granda et al., 2014). ...
Distribution of pines in the Iberian Peninsula agrees with species differences in foliage frost tolerance, not with vulnerability to freezing-induced xylem embolism 6 2018
... 常绿树种茎抗低温胁迫能力可能还与叶片低温抗性有关(Ball et al., 2004; Fernández-Pérez et al., 2018; Verhoeven et al., 2018).“脆弱性分割假说”指出, 植物面临胁迫时, 其叶片(或小枝)可能比其茎表现得更为脆弱, 通过牺牲 “造价低廉”的末端器官保护“造价昂贵”的茎干, 以避免栓塞发生(Zimmermann, 1983; Tyree & Ewers, 1991).已存在大量有关干旱胁迫下的“脆弱性分割假说”验证研究(Hao et al., 2008; Johnson et al., 2012; Pivovaroff et al., 2014; Zhu et al., 2019), 但还不清楚低温胁迫下是否也会发生该现象.Fernández-Pérez等(2018)发现, 6个松属(Pinus)树种的针叶低温抗性对其分布范围的预测能力大于茎干木质部抗冻融栓塞能力.在北半球温带森林中, 冬季留存叶片因日间光照而使其日间温度高于0 ℃, 从而可能延长光合作用的时间(Feild & Brodribb, 2001; Sevanto et al., 2006; Lindfors et al., 2015; Bowling et al., 2018), 生产的有机物可用于早春修复冻融栓塞和细胞损伤.不过, 此时树干仍处于冰冻状态, 导致叶片可能比茎经历更多次的冻融循环, 从而加剧叶片的冻融栓塞(Feild & Brodribb, 2001; Charra-Vaskou et al., 2016), 降低茎干的冻融栓塞程度, 在一定程度上对主茎起到保护作用(Charrier et al., 2017). ...
... ), 但还不清楚低温胁迫下是否也会发生该现象.Fernández-Pérez等(2018)发现, 6个松属(Pinus)树种的针叶低温抗性对其分布范围的预测能力大于茎干木质部抗冻融栓塞能力.在北半球温带森林中, 冬季留存叶片因日间光照而使其日间温度高于0 ℃, 从而可能延长光合作用的时间(Feild & Brodribb, 2001; Sevanto et al., 2006; Lindfors et al., 2015; Bowling et al., 2018), 生产的有机物可用于早春修复冻融栓塞和细胞损伤.不过, 此时树干仍处于冰冻状态, 导致叶片可能比茎经历更多次的冻融循环, 从而加剧叶片的冻融栓塞(Feild & Brodribb, 2001; Charra-Vaskou et al., 2016), 降低茎干的冻融栓塞程度, 在一定程度上对主茎起到保护作用(Charrier et al., 2017). ...
... 木质部结构是影响植物忍耐低温胁迫的主要因素(Hacke et al., 2015, 2017; Lintunen et al., 2018).被子植物采用多个细胞死亡后相互连接构成的导管进行水分运输, 长度在1 mm到几米不等; 裸子植物则依靠单个细胞伸长形成的管胞进行水分运输, 长度在2-6 mm之间(Tyree & Ewers, 1991), 而且管胞直径一般小于导管(Sperry et al., 2006).这种木质部结构差异导致裸子植物和被子植物响应冻融栓塞时产生显著PLC差异(Feild & Brodribb, 2001; Feng et al., 2015; Fernández-Pérez et al., 2018).以往研究表明, 植物在入冬前PLC逐步增加(Zhang et al., 2018), 而导管或管胞的直径差异是植物PLC不同的主要原因(Pittermann & Sperry, 2003; Hacke et al., 2015, 2017).在所有季节中, 裸子植物的PLC均小于被子植物(Granda et al., 2014).例如, 被子植物葡萄牙栎(Quercus faginea)抗空穴化能力较差, 夏季PLC达到80%, 冬季进一步达到90%, 而裸子植物香桧(Juniperus thurifera)和欧洲黑松(Pinus nigra)夏季PLC几乎为0, 冬季仅增加到3%-9% (Granda et al., 2014). ...
... 评价植物抗冻融栓塞能力最直接的指标是冻融栓塞引起水力导度丢失的百分比, 即木质部最大水力导度与低温下水力导度之差和最大水力导度之比(Charrier et al., 2013; Niu et al., 2017), 与干旱PLC类似.冻融栓塞常常会引起植物水力导度丢失现象.Granda等(2014)整合75个研究中的210个记录发现, 植物PLC从夏季到冬季逐步增加.Charrier等(2013)也发现, 冬季植物PLC最高, 且PLC可能受海拔限制.然而, Fernández-Pérez等(2018)报道, 低温胁迫引起的6个松属树种的PLC较低, 而且没有发现PLC与低温损伤(叶黎明前水势)之间的关系. ...
... 常绿植物保证叶片安全越冬对其之后的生长具有关键作用(Ball et al., 2004), 因此我们还可以通过分析叶片抗低温胁迫能力来评价植物的低温耐性.目前评价叶片抗低温胁迫能力的指标主要有: 叶片电解质渗出率(Climent et al., 2009; Hoermiller et al., 2018)、可溶性糖含量(Fernández-Pérez et al., 2018)、黎明前叶水势(Taneda & Tateno, 2005)、叶细胞弹性模量(细胞壁刚性越高, 细胞弹性模量越大)(Zhang et al., 2016)等.Fernández-Pérez等(2018)报道, 伊比利亚半岛6个松属树种的分布与其叶片低温忍耐能力有关, 生长在寒冷环境中的针叶的电解质渗出率和可溶性糖含量较低, 且黎明前叶水势对针叶低温损伤有影响.Martínez-Vilalta等(2016)整合分析全球尺度121个研究中177个物种非结构性碳(淀粉和可溶性糖)数据发现, 北方和温带生物区系物种叶片非结构性碳含量最高, 而且在入冬过程中逐步增加.Zhang等(2016)发现, 叶片细胞壁强度的增加, 降低了质外体冰冻对细胞膜的物理损伤, 使植物在较低的温度下得以存活.上述几个评价指标中, 电解质渗出率是最常用的评价植物组织损伤的指标(Charrier et al., 2013; Arias et al., 2015; Mayr & Améglio, 2016).通常采用电解质渗出率达50%时所对应的温度(LT50)来评价植物的低温抗性: LT50越低, 则低温抗性越强(Kreyling et al., 2015; Bachofen et al., 2016; Arias et al., 2017).例如, Arias等(2017)报道, 油橄榄的LT50从夏季到冬季呈现逐步降低的趋势, 这表明随着温度的降低, 通过不断驯化, 油橄榄的低温抗性逐步增强. ...
... )等.Fernández-Pérez等(2018)报道, 伊比利亚半岛6个松属树种的分布与其叶片低温忍耐能力有关, 生长在寒冷环境中的针叶的电解质渗出率和可溶性糖含量较低, 且黎明前叶水势对针叶低温损伤有影响.Martínez-Vilalta等(2016)整合分析全球尺度121个研究中177个物种非结构性碳(淀粉和可溶性糖)数据发现, 北方和温带生物区系物种叶片非结构性碳含量最高, 而且在入冬过程中逐步增加.Zhang等(2016)发现, 叶片细胞壁强度的增加, 降低了质外体冰冻对细胞膜的物理损伤, 使植物在较低的温度下得以存活.上述几个评价指标中, 电解质渗出率是最常用的评价植物组织损伤的指标(Charrier et al., 2013; Arias et al., 2015; Mayr & Améglio, 2016).通常采用电解质渗出率达50%时所对应的温度(LT50)来评价植物的低温抗性: LT50越低, 则低温抗性越强(Kreyling et al., 2015; Bachofen et al., 2016; Arias et al., 2017).例如, Arias等(2017)报道, 油橄榄的LT50从夏季到冬季呈现逐步降低的趋势, 这表明随着温度的降低, 通过不断驯化, 油橄榄的低温抗性逐步增强. ...
Stem water transport and freeze-thaw xylem embolism in conifers and angiosperms in a Tasmanian treeline heath 8 2001
... 栓塞是降低木质部内水分运输效率的关键因子(Choat et al., 2012; 金鹰等, 2016), 主要包括干旱栓塞和冻融栓塞(Améglio et al., 2002; Willson & Jackson, 2006; Venturas et al., 2017).两种栓塞形成的主要机理均为空气进入导管或管胞, 从而阻断木质部水柱, 形成栓塞(Sperry & Sullivan, 1992), 而不同之处在于引发空气进入的过程(Hacke & Sperry, 2001).根据内聚力-张力理论, 木质部导管内水分持续运输动力主要来自叶片蒸腾失水引起的拉力, 因此导管中的水分始终处于亚稳态(Sperry & Love, 2015).当蒸腾加剧和/或土壤水分供应不足而使木质部导管内负压超过一定阈值时, 就会造成其水柱断裂, 致使周围组织中的气体进入导管, 从而形成干旱栓塞(Choat et al., 2018).而冻融栓塞主要发生在木质部经历多次冻融循环的过程中(Sperry & Sullivan, 1992; Feild & Brodribb, 2001; Charra-?Vaskou et al., 2016).当木质部水分冰冻时, 水中原有溶解气体因在冰中的溶解度很低而逸散到导管中(Lintunen et al., 2014).而当冰冻融化时, 导管中的留存气体或者重新溶进水中(气泡压力大于大气压时, Hacke & Sperry, 2001), 或者由于冰冻压力的释放(冰融化后体积减小)等使气体在导管中扩大(Lemoine et al., 1999; Mayr et al., 2002; Pittermann & Sperry, 2006), 多次冻融交替之后, 导管中气体不断增加和扩张, 最终引发冻融栓塞, 该现象也被称为“融化-扩张假说” (Mayr & Sperry, 2010; Mayr & Améglio, 2016).干旱栓塞和冻融栓塞均会阻碍植物水分从土壤到叶片的运输, 从而降低叶片的光合速率(Brodribb et al., 2007; Reich, 2014; Jin et al., 2016), 导致植物生产力降低甚至死亡(McDowell et al., 2008; Hoffmann et al., 2011; McDowell, 2011; Choat et al., 2018). ...
... 与动物不同, 植物的固定性使得其更多地采用“被动”的忍耐策略, 例如不同的木质结构、调整组织水分含量和溶质含量等, 以减少低温损伤(Mayr et al., 2014; Arias et al., 2015; Lintunen et al., 2018).被子植物和裸子植物解剖结构的差异, 使二者对低温胁迫的忍耐能力不同(Pittermann & Sperry, 2003; Lintunen et al., 2016).被子植物的导管比裸子植物的管胞结构具有更高的水力效率, 但是其木质部抗冻融栓塞能力较低(Feild & Brodribb, 2001; Lintunen et al., 2018).Davis等(1999)报道, 当导管直径大于40 μm时, 一次冻融交替使木质部栓塞程度高达95%; Feild和Brodribb (2001)也发现, 常绿针叶植物和导管较小的被子植物经历一次冻融交替后, 其茎水力导度降低最少(几乎为0), 而其他被子植物则平均降低18%-70%.可见, 被子植物木质部忍耐低温能力远低于裸子植物.植物经过多次冻融交替后, 导管或周围组织中产生的气泡可能在整个冬季留存, 直到春季才能重新进入融化水中或者气泡扩张形成栓塞(Hacke & Sperry, 2001; Venturas et al., 2017; Medeiros & Danielson, 2018), 所以导管内水分含量直接影响植物冻融栓塞程度(Choat et al., 2011).而水分含量与导管直径紧密相关(Lens et al., 2013).因此构建直径更小的导管是最直接增加抗冻融栓塞能力的方法.裸子植物直径较小的管胞结构能够在很大程度上减少冻融栓塞的形成, 不仅因为在管胞中 ...
Vessel scaling in evergreen angiosperm leaves conforms with Murray’s law and area-filling assumptions: Implications for plant size, leaf size and cold tolerance 1 2018
... 形成的气泡更小, 而且小气泡更容易在融化期重新溶解进入水中(Hacke et al., 2015).Pittermann和Sperry等(2006)报道, 管胞越窄的裸子树种气泡形成的空穴化越小, 并且得出冻融栓塞引起的木质部负压随着管胞直径的增加而增加.之后, Choat等(2011)对11种被子植物的研究也发现, 导管直径越大、长度越长的植物, 冻融引起的水力导度丢失量(PLC)越多、造成的栓塞越大.这可能是因为直径较大的导管具有更少的单位长度导管末端, 造成其纹孔数量减少、栓塞抗性降低(Choat et al., 2006).因此, 构建直径更小的导管(或管胞), 是植物应对冻融栓塞最直接的方式(Pittermann & Sperry, 2006; Jiménez-Castillo & Lusk, 2013; Hacke et al., 2017; Gleason et al., 2018). ...
Weak tradeoff between xylem safety and xylem-specific hydraulic efficiency across the world’s woody plant species 1 2016
... 木质部的水力效率-安全权衡是植物适应环境和进化的结果(Blackman et al., 2010; Jin et al., 2016), 影响植物的竞争力和生产力(Nardini & Luglio, 2014).水力效率一般用植物水力导度表示, 而水力安全常用抗干旱栓塞能力(P50)表示(Santiago et al., 2018).Nardini等(2012)报道, 来自温带的6种植物(槭属(Acer)和栎属(Quercus)各3种)的水力导度、P50和比叶质量之间存在显著相关性, 支持水力效率-安全权衡, 且增加叶碳投入可以提高水力丢失的忍耐能力.Nardini和Luglio (2014)整合130个木本被子植物叶片水力数据后也发现存在叶水力效率-安全权衡关系, 而且生长在低降水量地区的植物的P50 (负值)和叶水力导度均较低.Gleason等(2016)综合分析335种被子植物和89种裸子植物的茎水力数据, 发现虽然水力导度和抗干旱栓塞能力之间存在弱相关性(支持水力效率-安全权衡), 但没有发现高效率和高安全性的植物.因此, 植物普遍存在不同程度的水力效率-安全权衡关系, 但其关系的紧密程度受导管特征(Lens et al., 2011; Hajek et al., 2014)、木质材性(金鹰和王传宽, 2016)、树高(Prendin et al., 2018)等因素的影响. ...
Leaf and stem physiological responses to summer and winter extremes of woody species across temperate ecosystems 3 2014
... 木质部结构是影响植物忍耐低温胁迫的主要因素(Hacke et al., 2015, 2017; Lintunen et al., 2018).被子植物采用多个细胞死亡后相互连接构成的导管进行水分运输, 长度在1 mm到几米不等; 裸子植物则依靠单个细胞伸长形成的管胞进行水分运输, 长度在2-6 mm之间(Tyree & Ewers, 1991), 而且管胞直径一般小于导管(Sperry et al., 2006).这种木质部结构差异导致裸子植物和被子植物响应冻融栓塞时产生显著PLC差异(Feild & Brodribb, 2001; Feng et al., 2015; Fernández-Pérez et al., 2018).以往研究表明, 植物在入冬前PLC逐步增加(Zhang et al., 2018), 而导管或管胞的直径差异是植物PLC不同的主要原因(Pittermann & Sperry, 2003; Hacke et al., 2015, 2017).在所有季节中, 裸子植物的PLC均小于被子植物(Granda et al., 2014).例如, 被子植物葡萄牙栎(Quercus faginea)抗空穴化能力较差, 夏季PLC达到80%, 冬季进一步达到90%, 而裸子植物香桧(Juniperus thurifera)和欧洲黑松(Pinus nigra)夏季PLC几乎为0, 冬季仅增加到3%-9% (Granda et al., 2014). ...
... 几乎为0, 冬季仅增加到3%-9% (Granda et al., 2014). ...
... 评价植物抗冻融栓塞能力最直接的指标是冻融栓塞引起水力导度丢失的百分比, 即木质部最大水力导度与低温下水力导度之差和最大水力导度之比(Charrier et al., 2013; Niu et al., 2017), 与干旱PLC类似.冻融栓塞常常会引起植物水力导度丢失现象.Granda等(2014)整合75个研究中的210个记录发现, 植物PLC从夏季到冬季逐步增加.Charrier等(2013)也发现, 冬季植物PLC最高, 且PLC可能受海拔限制.然而, Fernández-Pérez等(2018)报道, 低温胁迫引起的6个松属树种的PLC较低, 而且没有发现PLC与低温损伤(叶黎明前水势)之间的关系. ...
The hydraulic architecture of conifers 5 2015
... 北方森林和温带森林几乎占全球森林总面积的1/3 (Reich et al., 2016), 其植物面临的最主要环境胁迫之一是冬季低温.低温使植物地上器官木质部中的水分逐步冻结(Niu et al., 2017), 所以植物如何最大程度减少低温胁迫引起的损伤对其安全越冬(Hacke et al., 2015; Kreyling et al., 2015; Charrier et al., 2017)、生长、生存以及分布均具有重要作用(Schreiber et al., 2013; Charrier et al., 2015; Niu et al., 2017).低温时期冻融诱导形成的栓塞是低温胁迫对植物产生损伤的重要因素(Zanne et al., 2014; Zhang et al., 2018).许多研究发现, 植物抗冻融栓塞能力不仅影响其在低温时期的损伤程度和碳消耗(Bowling et al., 2018; Yin et al., 2018), 还显著影响随后生长季中的水分供应能力(冷冻疲劳, Feng et al., 2015; Zhang et al., 2018).但与干旱栓塞相比, 目前对木质部冻融栓塞的形成以及植物响应和调节机制的理解不足(Choat et al., 2011; Yin et al., 2018).为了推进该领域的研究, 本文先综述木质部冻融栓塞的形成机制和植物的逃避、忍耐、修复等3种冻融栓塞的应对策略, 然后总结植物低温抗性的生理表现、评价和影响因子, 在此基础上讨论低温抗性、干旱抗性和水力效率之间的多元权衡关系, 最后提出今后该领域中的优先研究问题. ...
... 形成的气泡更小, 而且小气泡更容易在融化期重新溶解进入水中(Hacke et al., 2015).Pittermann和Sperry等(2006)报道, 管胞越窄的裸子树种气泡形成的空穴化越小, 并且得出冻融栓塞引起的木质部负压随着管胞直径的增加而增加.之后, Choat等(2011)对11种被子植物的研究也发现, 导管直径越大、长度越长的植物, 冻融引起的水力导度丢失量(PLC)越多、造成的栓塞越大.这可能是因为直径较大的导管具有更少的单位长度导管末端, 造成其纹孔数量减少、栓塞抗性降低(Choat et al., 2006).因此, 构建直径更小的导管(或管胞), 是植物应对冻融栓塞最直接的方式(Pittermann & Sperry, 2006; Jiménez-Castillo & Lusk, 2013; Hacke et al., 2017; Gleason et al., 2018). ...
... 木质部结构是影响植物忍耐低温胁迫的主要因素(Hacke et al., 2015, 2017; Lintunen et al., 2018).被子植物采用多个细胞死亡后相互连接构成的导管进行水分运输, 长度在1 mm到几米不等; 裸子植物则依靠单个细胞伸长形成的管胞进行水分运输, 长度在2-6 mm之间(Tyree & Ewers, 1991), 而且管胞直径一般小于导管(Sperry et al., 2006).这种木质部结构差异导致裸子植物和被子植物响应冻融栓塞时产生显著PLC差异(Feild & Brodribb, 2001; Feng et al., 2015; Fernández-Pérez et al., 2018).以往研究表明, 植物在入冬前PLC逐步增加(Zhang et al., 2018), 而导管或管胞的直径差异是植物PLC不同的主要原因(Pittermann & Sperry, 2003; Hacke et al., 2015, 2017).在所有季节中, 裸子植物的PLC均小于被子植物(Granda et al., 2014).例如, 被子植物葡萄牙栎(Quercus faginea)抗空穴化能力较差, 夏季PLC达到80%, 冬季进一步达到90%, 而裸子植物香桧(Juniperus thurifera)和欧洲黑松(Pinus nigra)夏季PLC几乎为0, 冬季仅增加到3%-9% (Granda et al., 2014). ...
... ; Hacke et al., 2015, 2017).在所有季节中, 裸子植物的PLC均小于被子植物(Granda et al., 2014).例如, 被子植物葡萄牙栎(Quercus faginea)抗空穴化能力较差, 夏季PLC达到80%, 冬季进一步达到90%, 而裸子植物香桧(Juniperus thurifera)和欧洲黑松(Pinus nigra)夏季PLC几乎为0, 冬季仅增加到3%-9% (Granda et al., 2014). ...
... 只有当Px小于P*时, 气泡才有可能扩大并形成空穴.由公式(1)可知, 木质部导管越大, 越容易形成直径较大的气泡, 气泡压力越小, 在较低张力下气泡越可能扩大, 因而对冻融栓塞的抵抗更加脆弱(Feild & Brodribb, 2001; Choat et al., 2011; Hacke et al., 2017).裸子植物管胞直径一般比被子植物导管直径小(Hacke et al., 2017), 因此裸子植物的抗冻融栓塞能力较强.但在管道直径相同时, 裸子植物与被子植物具有相似的抗冻融栓塞能力(Pittermann & Sperry, 2003, 2006; Hacke et al., 2015). ...
Functional and ecological xylem anatomy 3 2001
... 栓塞是降低木质部内水分运输效率的关键因子(Choat et al., 2012; 金鹰等, 2016), 主要包括干旱栓塞和冻融栓塞(Améglio et al., 2002; Willson & Jackson, 2006; Venturas et al., 2017).两种栓塞形成的主要机理均为空气进入导管或管胞, 从而阻断木质部水柱, 形成栓塞(Sperry & Sullivan, 1992), 而不同之处在于引发空气进入的过程(Hacke & Sperry, 2001).根据内聚力-张力理论, 木质部导管内水分持续运输动力主要来自叶片蒸腾失水引起的拉力, 因此导管中的水分始终处于亚稳态(Sperry & Love, 2015).当蒸腾加剧和/或土壤水分供应不足而使木质部导管内负压超过一定阈值时, 就会造成其水柱断裂, 致使周围组织中的气体进入导管, 从而形成干旱栓塞(Choat et al., 2018).而冻融栓塞主要发生在木质部经历多次冻融循环的过程中(Sperry & Sullivan, 1992; Feild & Brodribb, 2001; Charra-?Vaskou et al., 2016).当木质部水分冰冻时, 水中原有溶解气体因在冰中的溶解度很低而逸散到导管中(Lintunen et al., 2014).而当冰冻融化时, 导管中的留存气体或者重新溶进水中(气泡压力大于大气压时, Hacke & Sperry, 2001), 或者由于冰冻压力的释放(冰融化后体积减小)等使气体在导管中扩大(Lemoine et al., 1999; Mayr et al., 2002; Pittermann & Sperry, 2006), 多次冻融交替之后, 导管中气体不断增加和扩张, 最终引发冻融栓塞, 该现象也被称为“融化-扩张假说” (Mayr & Sperry, 2010; Mayr & Améglio, 2016).干旱栓塞和冻融栓塞均会阻碍植物水分从土壤到叶片的运输, 从而降低叶片的光合速率(Brodribb et al., 2007; Reich, 2014; Jin et al., 2016), 导致植物生产力降低甚至死亡(McDowell et al., 2008; Hoffmann et al., 2011; McDowell, 2011; Choat et al., 2018). ...
... ).而当冰冻融化时, 导管中的留存气体或者重新溶进水中(气泡压力大于大气压时, Hacke & Sperry, 2001), 或者由于冰冻压力的释放(冰融化后体积减小)等使气体在导管中扩大(Lemoine et al., 1999; Mayr et al., 2002; Pittermann & Sperry, 2006), 多次冻融交替之后, 导管中气体不断增加和扩张, 最终引发冻融栓塞, 该现象也被称为“融化-扩张假说” (Mayr & Sperry, 2010; Mayr & Améglio, 2016).干旱栓塞和冻融栓塞均会阻碍植物水分从土壤到叶片的运输, 从而降低叶片的光合速率(Brodribb et al., 2007; Reich, 2014; Jin et al., 2016), 导致植物生产力降低甚至死亡(McDowell et al., 2008; Hoffmann et al., 2011; McDowell, 2011; Choat et al., 2018). ...
... 与动物不同, 植物的固定性使得其更多地采用“被动”的忍耐策略, 例如不同的木质结构、调整组织水分含量和溶质含量等, 以减少低温损伤(Mayr et al., 2014; Arias et al., 2015; Lintunen et al., 2018).被子植物和裸子植物解剖结构的差异, 使二者对低温胁迫的忍耐能力不同(Pittermann & Sperry, 2003; Lintunen et al., 2016).被子植物的导管比裸子植物的管胞结构具有更高的水力效率, 但是其木质部抗冻融栓塞能力较低(Feild & Brodribb, 2001; Lintunen et al., 2018).Davis等(1999)报道, 当导管直径大于40 μm时, 一次冻融交替使木质部栓塞程度高达95%; Feild和Brodribb (2001)也发现, 常绿针叶植物和导管较小的被子植物经历一次冻融交替后, 其茎水力导度降低最少(几乎为0), 而其他被子植物则平均降低18%-70%.可见, 被子植物木质部忍耐低温能力远低于裸子植物.植物经过多次冻融交替后, 导管或周围组织中产生的气泡可能在整个冬季留存, 直到春季才能重新进入融化水中或者气泡扩张形成栓塞(Hacke & Sperry, 2001; Venturas et al., 2017; Medeiros & Danielson, 2018), 所以导管内水分含量直接影响植物冻融栓塞程度(Choat et al., 2011).而水分含量与导管直径紧密相关(Lens et al., 2013).因此构建直径更小的导管是最直接增加抗冻融栓塞能力的方法.裸子植物直径较小的管胞结构能够在很大程度上减少冻融栓塞的形成, 不仅因为在管胞中 ...
An ecophysiological and developmental perspective on variation in vessel diameter 5 2017
... 形成的气泡更小, 而且小气泡更容易在融化期重新溶解进入水中(Hacke et al., 2015).Pittermann和Sperry等(2006)报道, 管胞越窄的裸子树种气泡形成的空穴化越小, 并且得出冻融栓塞引起的木质部负压随着管胞直径的增加而增加.之后, Choat等(2011)对11种被子植物的研究也发现, 导管直径越大、长度越长的植物, 冻融引起的水力导度丢失量(PLC)越多、造成的栓塞越大.这可能是因为直径较大的导管具有更少的单位长度导管末端, 造成其纹孔数量减少、栓塞抗性降低(Choat et al., 2006).因此, 构建直径更小的导管(或管胞), 是植物应对冻融栓塞最直接的方式(Pittermann & Sperry, 2006; Jiménez-Castillo & Lusk, 2013; Hacke et al., 2017; Gleason et al., 2018). ...
... 木质部结构是影响植物忍耐低温胁迫的主要因素(Hacke et al., 2015, 2017; Lintunen et al., 2018).被子植物采用多个细胞死亡后相互连接构成的导管进行水分运输, 长度在1 mm到几米不等; 裸子植物则依靠单个细胞伸长形成的管胞进行水分运输, 长度在2-6 mm之间(Tyree & Ewers, 1991), 而且管胞直径一般小于导管(Sperry et al., 2006).这种木质部结构差异导致裸子植物和被子植物响应冻融栓塞时产生显著PLC差异(Feild & Brodribb, 2001; Feng et al., 2015; Fernández-Pérez et al., 2018).以往研究表明, 植物在入冬前PLC逐步增加(Zhang et al., 2018), 而导管或管胞的直径差异是植物PLC不同的主要原因(Pittermann & Sperry, 2003; Hacke et al., 2015, 2017).在所有季节中, 裸子植物的PLC均小于被子植物(Granda et al., 2014).例如, 被子植物葡萄牙栎(Quercus faginea)抗空穴化能力较差, 夏季PLC达到80%, 冬季进一步达到90%, 而裸子植物香桧(Juniperus thurifera)和欧洲黑松(Pinus nigra)夏季PLC几乎为0, 冬季仅增加到3%-9% (Granda et al., 2014). ...
... , 2017).在所有季节中, 裸子植物的PLC均小于被子植物(Granda et al., 2014).例如, 被子植物葡萄牙栎(Quercus faginea)抗空穴化能力较差, 夏季PLC达到80%, 冬季进一步达到90%, 而裸子植物香桧(Juniperus thurifera)和欧洲黑松(Pinus nigra)夏季PLC几乎为0, 冬季仅增加到3%-9% (Granda et al., 2014). ...
... 只有当Px小于P*时, 气泡才有可能扩大并形成空穴.由公式(1)可知, 木质部导管越大, 越容易形成直径较大的气泡, 气泡压力越小, 在较低张力下气泡越可能扩大, 因而对冻融栓塞的抵抗更加脆弱(Feild & Brodribb, 2001; Choat et al., 2011; Hacke et al., 2017).裸子植物管胞直径一般比被子植物导管直径小(Hacke et al., 2017), 因此裸子植物的抗冻融栓塞能力较强.但在管道直径相同时, 裸子植物与被子植物具有相似的抗冻融栓塞能力(Pittermann & Sperry, 2003, 2006; Hacke et al., 2015). ...
... ).裸子植物管胞直径一般比被子植物导管直径小(Hacke et al., 2017), 因此裸子植物的抗冻融栓塞能力较强.但在管道直径相同时, 裸子植物与被子植物具有相似的抗冻融栓塞能力(Pittermann & Sperry, 2003, 2006; Hacke et al., 2015). ...
Trade-offs between xylem hydraulic properties, wood anatomy and yield in Populus 1 2014
... 木质部的水力效率-安全权衡是植物适应环境和进化的结果(Blackman et al., 2010; Jin et al., 2016), 影响植物的竞争力和生产力(Nardini & Luglio, 2014).水力效率一般用植物水力导度表示, 而水力安全常用抗干旱栓塞能力(P50)表示(Santiago et al., 2018).Nardini等(2012)报道, 来自温带的6种植物(槭属(Acer)和栎属(Quercus)各3种)的水力导度、P50和比叶质量之间存在显著相关性, 支持水力效率-安全权衡, 且增加叶碳投入可以提高水力丢失的忍耐能力.Nardini和Luglio (2014)整合130个木本被子植物叶片水力数据后也发现存在叶水力效率-安全权衡关系, 而且生长在低降水量地区的植物的P50 (负值)和叶水力导度均较低.Gleason等(2016)综合分析335种被子植物和89种裸子植物的茎水力数据, 发现虽然水力导度和抗干旱栓塞能力之间存在弱相关性(支持水力效率-安全权衡), 但没有发现高效率和高安全性的植物.因此, 植物普遍存在不同程度的水力效率-安全权衡关系, 但其关系的紧密程度受导管特征(Lens et al., 2011; Hajek et al., 2014)、木质材性(金鹰和王传宽, 2016)、树高(Prendin et al., 2018)等因素的影响. ...
Stem and leaf hydraulics of congeneric tree species from adjacent tropical savanna and forest ecosystems 1 2008
... 常绿树种茎抗低温胁迫能力可能还与叶片低温抗性有关(Ball et al., 2004; Fernández-Pérez et al., 2018; Verhoeven et al., 2018).“脆弱性分割假说”指出, 植物面临胁迫时, 其叶片(或小枝)可能比其茎表现得更为脆弱, 通过牺牲 “造价低廉”的末端器官保护“造价昂贵”的茎干, 以避免栓塞发生(Zimmermann, 1983; Tyree & Ewers, 1991).已存在大量有关干旱胁迫下的“脆弱性分割假说”验证研究(Hao et al., 2008; Johnson et al., 2012; Pivovaroff et al., 2014; Zhu et al., 2019), 但还不清楚低温胁迫下是否也会发生该现象.Fernández-Pérez等(2018)发现, 6个松属(Pinus)树种的针叶低温抗性对其分布范围的预测能力大于茎干木质部抗冻融栓塞能力.在北半球温带森林中, 冬季留存叶片因日间光照而使其日间温度高于0 ℃, 从而可能延长光合作用的时间(Feild & Brodribb, 2001; Sevanto et al., 2006; Lindfors et al., 2015; Bowling et al., 2018), 生产的有机物可用于早春修复冻融栓塞和细胞损伤.不过, 此时树干仍处于冰冻状态, 导致叶片可能比茎经历更多次的冻融循环, 从而加剧叶片的冻融栓塞(Feild & Brodribb, 2001; Charra-Vaskou et al., 2016), 降低茎干的冻融栓塞程度, 在一定程度上对主茎起到保护作用(Charrier et al., 2017). ...
Investigating xylem embolism formation, refilling and water storage in tree trunks using frequency domain reflectometry 1 2013
... 修复是植物应对冻融栓塞的又一重要策略(Preston & Sandve, 2013; Wang et al., 2018).有研究表明, 采用正压(主要包括根压和茎压)恢复水力功能的植物可能具有更强的抗冷冻疲劳能力(即木质部冰冻后对水力功能丢失更敏感的现象)(Niu et al., 2017; Yin et al., 2018), 这是植物对冻融栓塞的一种主动修复.Hao等(2013)发现, 早春纸桦(Betula papyrifera)树干水分含量明显高于生长季, 表明根压在修复冻融栓塞导管的过程中具有重要作用.Améglio等(2002)报道, 冬季胡桃(Juglans regia) PLC可高达到100%, 但是由于冬季和春季茎压修复, 木质部冻融栓塞程度有一定的降低.另外, 近期研究表明, 能产生木质部正压的植物, 其抗冻融栓塞能力可能不受导管直径的影响(Niu et al., 2017).根压和茎压主要来源于糖类水解成的蔗糖被运输到栓塞导管中而产生渗透浓度梯度(Nardini et al., 2011; Cao et al., 2012), 从而使水分流向栓塞导管, 即渗透调节假说(金鹰等, 2016).两者的不同之处在于: 根压主要在无蒸腾流情况下产生作用(Swaef et al., 2013), 且修复栓塞作用较小(Salleo et al., 2004), 特别是对高大植物上层木质部栓塞的修复作用很小(Westhoff et al., 2008).而茎压产生的渗透梯度可能来自韧皮部的糖类(韧皮部卸载假说, Salleo et al., 2004; Hölttä et al., 2018), 因此在多数情况下均有茎压存在, 这也被称为“新的再冲水” (novel refilling)假说(Brodersen & McElrone, 2013; 金鹰等, 2016).Améglio等(2004)发现, 胡桃在深冬时茎中淀粉含量最低, 在发芽前茎中再次合成淀粉, 这不仅表明胡桃在冬季低温下进行淀粉和蔗糖的转换, 而且蔗糖累积是其冻融栓塞修复中的关键.张海燕等(2013)也发现, 12个温带树种老枝的淀粉含量变化具有相同趋势. ...
Drought-induced xylem pit membrane damage in aspen and balsam poplar 1 2016
... 木质部的水力效率-安全权衡是植物适应环境和进化的结果(Blackman et al., 2010; Jin et al., 2016), 影响植物的竞争力和生产力(Nardini & Luglio, 2014).水力效率一般用植物水力导度表示, 而水力安全常用抗干旱栓塞能力(P50)表示(Santiago et al., 2018).Nardini等(2012)报道, 来自温带的6种植物(槭属(Acer)和栎属(Quercus)各3种)的水力导度、P50和比叶质量之间存在显著相关性, 支持水力效率-安全权衡, 且增加叶碳投入可以提高水力丢失的忍耐能力.Nardini和Luglio (2014)整合130个木本被子植物叶片水力数据后也发现存在叶水力效率-安全权衡关系, 而且生长在低降水量地区的植物的P50 (负值)和叶水力导度均较低.Gleason等(2016)综合分析335种被子植物和89种裸子植物的茎水力数据, 发现虽然水力导度和抗干旱栓塞能力之间存在弱相关性(支持水力效率-安全权衡), 但没有发现高效率和高安全性的植物.因此, 植物普遍存在不同程度的水力效率-安全权衡关系, 但其关系的紧密程度受导管特征(Lens et al., 2011; Hajek et al., 2014)、木质材性(金鹰和王传宽, 2016)、树高(Prendin et al., 2018)等因素的影响. ...
木本植物木质部栓塞修复机制: 研究进展与问题 1 2016
... 木质部的水力效率-安全权衡是植物适应环境和进化的结果(Blackman et al., 2010; Jin et al., 2016), 影响植物的竞争力和生产力(Nardini & Luglio, 2014).水力效率一般用植物水力导度表示, 而水力安全常用抗干旱栓塞能力(P50)表示(Santiago et al., 2018).Nardini等(2012)报道, 来自温带的6种植物(槭属(Acer)和栎属(Quercus)各3种)的水力导度、P50和比叶质量之间存在显著相关性, 支持水力效率-安全权衡, 且增加叶碳投入可以提高水力丢失的忍耐能力.Nardini和Luglio (2014)整合130个木本被子植物叶片水力数据后也发现存在叶水力效率-安全权衡关系, 而且生长在低降水量地区的植物的P50 (负值)和叶水力导度均较低.Gleason等(2016)综合分析335种被子植物和89种裸子植物的茎水力数据, 发现虽然水力导度和抗干旱栓塞能力之间存在弱相关性(支持水力效率-安全权衡), 但没有发现高效率和高安全性的植物.因此, 植物普遍存在不同程度的水力效率-安全权衡关系, 但其关系的紧密程度受导管特征(Lens et al., 2011; Hajek et al., 2014)、木质材性(金鹰和王传宽, 2016)、树高(Prendin et al., 2018)等因素的影响. ...
Hydraulic safety margins and embolism reversal in stems and leaves: Why are conifers and angiosperms so different? 1 2012
... 常绿树种茎抗低温胁迫能力可能还与叶片低温抗性有关(Ball et al., 2004; Fernández-Pérez et al., 2018; Verhoeven et al., 2018).“脆弱性分割假说”指出, 植物面临胁迫时, 其叶片(或小枝)可能比其茎表现得更为脆弱, 通过牺牲 “造价低廉”的末端器官保护“造价昂贵”的茎干, 以避免栓塞发生(Zimmermann, 1983; Tyree & Ewers, 1991).已存在大量有关干旱胁迫下的“脆弱性分割假说”验证研究(Hao et al., 2008; Johnson et al., 2012; Pivovaroff et al., 2014; Zhu et al., 2019), 但还不清楚低温胁迫下是否也会发生该现象.Fernández-Pérez等(2018)发现, 6个松属(Pinus)树种的针叶低温抗性对其分布范围的预测能力大于茎干木质部抗冻融栓塞能力.在北半球温带森林中, 冬季留存叶片因日间光照而使其日间温度高于0 ℃, 从而可能延长光合作用的时间(Feild & Brodribb, 2001; Sevanto et al., 2006; Lindfors et al., 2015; Bowling et al., 2018), 生产的有机物可用于早春修复冻融栓塞和细胞损伤.不过, 此时树干仍处于冰冻状态, 导致叶片可能比茎经历更多次的冻融循环, 从而加剧叶片的冻融栓塞(Feild & Brodribb, 2001; Charra-Vaskou et al., 2016), 降低茎干的冻融栓塞程度, 在一定程度上对主茎起到保护作用(Charrier et al., 2017). ...
Prior exposure to freezing stress enhances the survival and recovery of Poa pratensis exposed to severe drought 1 2016
... 木质部低温抗性、干旱抗性和水力效率三者间是否存在多元权衡现象?目前对此问题的综合研究较少, 但已有研究指出, 干旱显著增加植物的低温抗性(Kreyling et al., 2012; Walter et al., 2013; Sierra-?Almeida et al., 2016; Sperling et al., 2017).这是因为植物通过驯化改变了细胞对脱水的忍耐能力(Buchner & Neuner, 2011), 其中包括基因表达的改变导致的细胞膜结构变化或者细胞化学成分变化(Verhoeven et al., 2018).例如, Medeiros和Pockman (2011)报道, 在-15 ℃环境中干旱处理使Larrea tridentate植株细胞死亡减少、叶留存量增加、水力导度和气体交换速率升高, 这表明干旱处理显著增加了其低温抗性.Hoffman等(2012)也发现, 干旱处理能够显著增加黑麦草(Lolium perenne)的低温抗性.然而, Kong和Henry (2016)研究发现, 早春低温处理的草地早熟禾(Poa pratensis)的抗旱性显著增加(生物量和存活率增加), 但夏季干旱处理并未增加其低温抗性, 这表明干旱处理对植物低温抗性无影响的现象也存在.同样, Bachofen等(2016)也发现, 干旱处理没有增加地中海松属3个树种的低温抗性, 而且推测该现象可能仅在短期极端条件下发生.Yin等(2018)同时对低温抗性、干旱抗性和水力效率进行研究发现, 抗干旱栓塞与水力效率存在权衡, 抗冷冻疲劳与水力效率间也存在权衡现象, 支持多元权衡现象.然而, 当植物同时面对干旱和低温胁迫时(如高山树线植物)(Mayr et al., 2007; Charrier et al., 2017), 多次冻融交替结合干旱胁迫可能引起PLC增加(Mayr et al., 2003; Charrier et al., 2014).Mayr等(2006)报道, 生长在林线处的针叶树种, 每个冬季经历至少100次冻融交替, 再加上干旱胁迫, 二者共同引起欧洲云杉(Picea abies) PLC高达100%.因此, 生长在低温和干旱胁迫共存环境中的植物, 其PLC可能更高, 间接表明三者间存在一定程度的权衡.综上所述, 我们构建了低温抗性、干旱抗性和水力效率之间多元权衡概念图(图2). ...
Cold tolerance of tree species is related to the climate of their native ranges 2 2015
... 北方森林和温带森林几乎占全球森林总面积的1/3 (Reich et al., 2016), 其植物面临的最主要环境胁迫之一是冬季低温.低温使植物地上器官木质部中的水分逐步冻结(Niu et al., 2017), 所以植物如何最大程度减少低温胁迫引起的损伤对其安全越冬(Hacke et al., 2015; Kreyling et al., 2015; Charrier et al., 2017)、生长、生存以及分布均具有重要作用(Schreiber et al., 2013; Charrier et al., 2015; Niu et al., 2017).低温时期冻融诱导形成的栓塞是低温胁迫对植物产生损伤的重要因素(Zanne et al., 2014; Zhang et al., 2018).许多研究发现, 植物抗冻融栓塞能力不仅影响其在低温时期的损伤程度和碳消耗(Bowling et al., 2018; Yin et al., 2018), 还显著影响随后生长季中的水分供应能力(冷冻疲劳, Feng et al., 2015; Zhang et al., 2018).但与干旱栓塞相比, 目前对木质部冻融栓塞的形成以及植物响应和调节机制的理解不足(Choat et al., 2011; Yin et al., 2018).为了推进该领域的研究, 本文先综述木质部冻融栓塞的形成机制和植物的逃避、忍耐、修复等3种冻融栓塞的应对策略, 然后总结植物低温抗性的生理表现、评价和影响因子, 在此基础上讨论低温抗性、干旱抗性和水力效率之间的多元权衡关系, 最后提出今后该领域中的优先研究问题. ...
... 常绿植物保证叶片安全越冬对其之后的生长具有关键作用(Ball et al., 2004), 因此我们还可以通过分析叶片抗低温胁迫能力来评价植物的低温耐性.目前评价叶片抗低温胁迫能力的指标主要有: 叶片电解质渗出率(Climent et al., 2009; Hoermiller et al., 2018)、可溶性糖含量(Fernández-Pérez et al., 2018)、黎明前叶水势(Taneda & Tateno, 2005)、叶细胞弹性模量(细胞壁刚性越高, 细胞弹性模量越大)(Zhang et al., 2016)等.Fernández-Pérez等(2018)报道, 伊比利亚半岛6个松属树种的分布与其叶片低温忍耐能力有关, 生长在寒冷环境中的针叶的电解质渗出率和可溶性糖含量较低, 且黎明前叶水势对针叶低温损伤有影响.Martínez-Vilalta等(2016)整合分析全球尺度121个研究中177个物种非结构性碳(淀粉和可溶性糖)数据发现, 北方和温带生物区系物种叶片非结构性碳含量最高, 而且在入冬过程中逐步增加.Zhang等(2016)发现, 叶片细胞壁强度的增加, 降低了质外体冰冻对细胞膜的物理损伤, 使植物在较低的温度下得以存活.上述几个评价指标中, 电解质渗出率是最常用的评价植物组织损伤的指标(Charrier et al., 2013; Arias et al., 2015; Mayr & Améglio, 2016).通常采用电解质渗出率达50%时所对应的温度(LT50)来评价植物的低温抗性: LT50越低, 则低温抗性越强(Kreyling et al., 2015; Bachofen et al., 2016; Arias et al., 2017).例如, Arias等(2017)报道, 油橄榄的LT50从夏季到冬季呈现逐步降低的趋势, 这表明随着温度的降低, 通过不断驯化, 油橄榄的低温抗性逐步增强. ...
Cold hardiness of Pinus nigra Arnold as influenced by geographic origin, warming, and extreme summer drought 1 2012
... 木质部低温抗性、干旱抗性和水力效率三者间是否存在多元权衡现象?目前对此问题的综合研究较少, 但已有研究指出, 干旱显著增加植物的低温抗性(Kreyling et al., 2012; Walter et al., 2013; Sierra-?Almeida et al., 2016; Sperling et al., 2017).这是因为植物通过驯化改变了细胞对脱水的忍耐能力(Buchner & Neuner, 2011), 其中包括基因表达的改变导致的细胞膜结构变化或者细胞化学成分变化(Verhoeven et al., 2018).例如, Medeiros和Pockman (2011)报道, 在-15 ℃环境中干旱处理使Larrea tridentate植株细胞死亡减少、叶留存量增加、水力导度和气体交换速率升高, 这表明干旱处理显著增加了其低温抗性.Hoffman等(2012)也发现, 干旱处理能够显著增加黑麦草(Lolium perenne)的低温抗性.然而, Kong和Henry (2016)研究发现, 早春低温处理的草地早熟禾(Poa pratensis)的抗旱性显著增加(生物量和存活率增加), 但夏季干旱处理并未增加其低温抗性, 这表明干旱处理对植物低温抗性无影响的现象也存在.同样, Bachofen等(2016)也发现, 干旱处理没有增加地中海松属3个树种的低温抗性, 而且推测该现象可能仅在短期极端条件下发生.Yin等(2018)同时对低温抗性、干旱抗性和水力效率进行研究发现, 抗干旱栓塞与水力效率存在权衡, 抗冷冻疲劳与水力效率间也存在权衡现象, 支持多元权衡现象.然而, 当植物同时面对干旱和低温胁迫时(如高山树线植物)(Mayr et al., 2007; Charrier et al., 2017), 多次冻融交替结合干旱胁迫可能引起PLC增加(Mayr et al., 2003; Charrier et al., 2014).Mayr等(2006)报道, 生长在林线处的针叶树种, 每个冬季经历至少100次冻融交替, 再加上干旱胁迫, 二者共同引起欧洲云杉(Picea abies) PLC高达100%.因此, 生长在低温和干旱胁迫共存环境中的植物, 其PLC可能更高, 间接表明三者间存在一定程度的权衡.综上所述, 我们构建了低温抗性、干旱抗性和水力效率之间多元权衡概念图(图2). ...
Mechanism of freeze-induced embolism in Fagus sylvatica L 1 1999
... 栓塞是降低木质部内水分运输效率的关键因子(Choat et al., 2012; 金鹰等, 2016), 主要包括干旱栓塞和冻融栓塞(Améglio et al., 2002; Willson & Jackson, 2006; Venturas et al., 2017).两种栓塞形成的主要机理均为空气进入导管或管胞, 从而阻断木质部水柱, 形成栓塞(Sperry & Sullivan, 1992), 而不同之处在于引发空气进入的过程(Hacke & Sperry, 2001).根据内聚力-张力理论, 木质部导管内水分持续运输动力主要来自叶片蒸腾失水引起的拉力, 因此导管中的水分始终处于亚稳态(Sperry & Love, 2015).当蒸腾加剧和/或土壤水分供应不足而使木质部导管内负压超过一定阈值时, 就会造成其水柱断裂, 致使周围组织中的气体进入导管, 从而形成干旱栓塞(Choat et al., 2018).而冻融栓塞主要发生在木质部经历多次冻融循环的过程中(Sperry & Sullivan, 1992; Feild & Brodribb, 2001; Charra-?Vaskou et al., 2016).当木质部水分冰冻时, 水中原有溶解气体因在冰中的溶解度很低而逸散到导管中(Lintunen et al., 2014).而当冰冻融化时, 导管中的留存气体或者重新溶进水中(气泡压力大于大气压时, Hacke & Sperry, 2001), 或者由于冰冻压力的释放(冰融化后体积减小)等使气体在导管中扩大(Lemoine et al., 1999; Mayr et al., 2002; Pittermann & Sperry, 2006), 多次冻融交替之后, 导管中气体不断增加和扩张, 最终引发冻融栓塞, 该现象也被称为“融化-扩张假说” (Mayr & Sperry, 2010; Mayr & Améglio, 2016).干旱栓塞和冻融栓塞均会阻碍植物水分从土壤到叶片的运输, 从而降低叶片的光合速率(Brodribb et al., 2007; Reich, 2014; Jin et al., 2016), 导致植物生产力降低甚至死亡(McDowell et al., 2008; Hoffmann et al., 2011; McDowell, 2011; Choat et al., 2018). ...
Testing hypotheses that link wood anatomy to cavitation resistance and hydraulic conductivity in the genus Acer 1 2011
... 木质部的水力效率-安全权衡是植物适应环境和进化的结果(Blackman et al., 2010; Jin et al., 2016), 影响植物的竞争力和生产力(Nardini & Luglio, 2014).水力效率一般用植物水力导度表示, 而水力安全常用抗干旱栓塞能力(P50)表示(Santiago et al., 2018).Nardini等(2012)报道, 来自温带的6种植物(槭属(Acer)和栎属(Quercus)各3种)的水力导度、P50和比叶质量之间存在显著相关性, 支持水力效率-安全权衡, 且增加叶碳投入可以提高水力丢失的忍耐能力.Nardini和Luglio (2014)整合130个木本被子植物叶片水力数据后也发现存在叶水力效率-安全权衡关系, 而且生长在低降水量地区的植物的P50 (负值)和叶水力导度均较低.Gleason等(2016)综合分析335种被子植物和89种裸子植物的茎水力数据, 发现虽然水力导度和抗干旱栓塞能力之间存在弱相关性(支持水力效率-安全权衡), 但没有发现高效率和高安全性的植物.因此, 植物普遍存在不同程度的水力效率-安全权衡关系, 但其关系的紧密程度受导管特征(Lens et al., 2011; Hajek et al., 2014)、木质材性(金鹰和王传宽, 2016)、树高(Prendin et al., 2018)等因素的影响. ...
Embolism resistance as a key mechanism to understand adaptive plant strategies 2 2013
... 与动物不同, 植物的固定性使得其更多地采用“被动”的忍耐策略, 例如不同的木质结构、调整组织水分含量和溶质含量等, 以减少低温损伤(Mayr et al., 2014; Arias et al., 2015; Lintunen et al., 2018).被子植物和裸子植物解剖结构的差异, 使二者对低温胁迫的忍耐能力不同(Pittermann & Sperry, 2003; Lintunen et al., 2016).被子植物的导管比裸子植物的管胞结构具有更高的水力效率, 但是其木质部抗冻融栓塞能力较低(Feild & Brodribb, 2001; Lintunen et al., 2018).Davis等(1999)报道, 当导管直径大于40 μm时, 一次冻融交替使木质部栓塞程度高达95%; Feild和Brodribb (2001)也发现, 常绿针叶植物和导管较小的被子植物经历一次冻融交替后, 其茎水力导度降低最少(几乎为0), 而其他被子植物则平均降低18%-70%.可见, 被子植物木质部忍耐低温能力远低于裸子植物.植物经过多次冻融交替后, 导管或周围组织中产生的气泡可能在整个冬季留存, 直到春季才能重新进入融化水中或者气泡扩张形成栓塞(Hacke & Sperry, 2001; Venturas et al., 2017; Medeiros & Danielson, 2018), 所以导管内水分含量直接影响植物冻融栓塞程度(Choat et al., 2011).而水分含量与导管直径紧密相关(Lens et al., 2013).因此构建直径更小的导管是最直接增加抗冻融栓塞能力的方法.裸子植物直径较小的管胞结构能够在很大程度上减少冻融栓塞的形成, 不仅因为在管胞中 ...
Dynamics of leaf gas exchange, chlorophyll fluorescence and stem diameter changes during freezing and thawing of Scots pine seedlings 1 2015
... 常绿树种茎抗低温胁迫能力可能还与叶片低温抗性有关(Ball et al., 2004; Fernández-Pérez et al., 2018; Verhoeven et al., 2018).“脆弱性分割假说”指出, 植物面临胁迫时, 其叶片(或小枝)可能比其茎表现得更为脆弱, 通过牺牲 “造价低廉”的末端器官保护“造价昂贵”的茎干, 以避免栓塞发生(Zimmermann, 1983; Tyree & Ewers, 1991).已存在大量有关干旱胁迫下的“脆弱性分割假说”验证研究(Hao et al., 2008; Johnson et al., 2012; Pivovaroff et al., 2014; Zhu et al., 2019), 但还不清楚低温胁迫下是否也会发生该现象.Fernández-Pérez等(2018)发现, 6个松属(Pinus)树种的针叶低温抗性对其分布范围的预测能力大于茎干木质部抗冻融栓塞能力.在北半球温带森林中, 冬季留存叶片因日间光照而使其日间温度高于0 ℃, 从而可能延长光合作用的时间(Feild & Brodribb, 2001; Sevanto et al., 2006; Lindfors et al., 2015; Bowling et al., 2018), 生产的有机物可用于早春修复冻融栓塞和细胞损伤.不过, 此时树干仍处于冰冻状态, 导致叶片可能比茎经历更多次的冻融循环, 从而加剧叶片的冻融栓塞(Feild & Brodribb, 2001; Charra-Vaskou et al., 2016), 降低茎干的冻融栓塞程度, 在一定程度上对主茎起到保护作用(Charrier et al., 2017). ...
Anatomical regulation of ice nucleation and cavitation helps trees to survive freezing and drought stress 1 2013
... 冻的树干(Charrier et al., 2017)无法及时补充水分丢失(Bowling et al., 2018), 从而使植物叶片和小枝的木质部含水量降低.另外, 含水量变化还可能会引起质外体组织冰冻时的体积发生变化(Lintunen et al., 2016).低温胁迫使木质组织首先脱水收缩, 再经历液态水冰冻时的体积膨大(+9%)(Améglio et al., 2001).在冰冻前, 植物主动降低质外体含水量, 增加溶质浓度, 此时脱水引起的体积减小能够降低过冷却点(Lintunen et al., 2013; Willick et al., 2019).因此, 冰冻前含水量越低(体积越小), 植物对低温胁迫的忍耐力越强(Lintunen et al., 2018).当冰冻发生时, 由于水分由液态转为固态的体积增加, 对周围细胞壁产生机械压力, 植物需构建更强健的细胞壁(Zhang et al., 2016; Ployet et al., 2018)和改变细胞壁上蛋白组分(Willick et al., 2018)来抵御低温胁迫, 因此, 质外体较高的水分含量可能对植物产生更严重的低温损伤.研究表明, 木质部通过寒冷驯化 后其直径少量收缩(Améglio et al., 2001; Charra- Vaskou et al., 2016; Lintunen et al., 2016).例如, 欧洲赤松(Pinus sylvestris)和欧洲山杨(Populus tremula)木质部直径经寒冷驯化后分别收缩了0.5%和1.0%, 小于渗透胁迫和干旱胁迫引起的收缩量(Lintunen et al., 2016).此外, 植物通过寒冷驯化, 可以降低质外体的含水量, 并将水分转移到冰储存位点, 同时不同冰区之间形成隔离带(Willick et al., 2019), 从而尽可能减少低温损伤. ...
Bursts of CO2 released during freezing offer a new perspective on avoidance of winter embolism in trees 1 2014
... 栓塞是降低木质部内水分运输效率的关键因子(Choat et al., 2012; 金鹰等, 2016), 主要包括干旱栓塞和冻融栓塞(Améglio et al., 2002; Willson & Jackson, 2006; Venturas et al., 2017).两种栓塞形成的主要机理均为空气进入导管或管胞, 从而阻断木质部水柱, 形成栓塞(Sperry & Sullivan, 1992), 而不同之处在于引发空气进入的过程(Hacke & Sperry, 2001).根据内聚力-张力理论, 木质部导管内水分持续运输动力主要来自叶片蒸腾失水引起的拉力, 因此导管中的水分始终处于亚稳态(Sperry & Love, 2015).当蒸腾加剧和/或土壤水分供应不足而使木质部导管内负压超过一定阈值时, 就会造成其水柱断裂, 致使周围组织中的气体进入导管, 从而形成干旱栓塞(Choat et al., 2018).而冻融栓塞主要发生在木质部经历多次冻融循环的过程中(Sperry & Sullivan, 1992; Feild & Brodribb, 2001; Charra-?Vaskou et al., 2016).当木质部水分冰冻时, 水中原有溶解气体因在冰中的溶解度很低而逸散到导管中(Lintunen et al., 2014).而当冰冻融化时, 导管中的留存气体或者重新溶进水中(气泡压力大于大气压时, Hacke & Sperry, 2001), 或者由于冰冻压力的释放(冰融化后体积减小)等使气体在导管中扩大(Lemoine et al., 1999; Mayr et al., 2002; Pittermann & Sperry, 2006), 多次冻融交替之后, 导管中气体不断增加和扩张, 最终引发冻融栓塞, 该现象也被称为“融化-扩张假说” (Mayr & Sperry, 2010; Mayr & Améglio, 2016).干旱栓塞和冻融栓塞均会阻碍植物水分从土壤到叶片的运输, 从而降低叶片的光合速率(Brodribb et al., 2007; Reich, 2014; Jin et al., 2016), 导致植物生产力降低甚至死亡(McDowell et al., 2008; Hoffmann et al., 2011; McDowell, 2011; Choat et al., 2018). ...
Xylem diameter changes during osmotic stress, desiccation and freezing in Pinus sylvestris and Populus tremula 5 2016
... 与动物不同, 植物的固定性使得其更多地采用“被动”的忍耐策略, 例如不同的木质结构、调整组织水分含量和溶质含量等, 以减少低温损伤(Mayr et al., 2014; Arias et al., 2015; Lintunen et al., 2018).被子植物和裸子植物解剖结构的差异, 使二者对低温胁迫的忍耐能力不同(Pittermann & Sperry, 2003; Lintunen et al., 2016).被子植物的导管比裸子植物的管胞结构具有更高的水力效率, 但是其木质部抗冻融栓塞能力较低(Feild & Brodribb, 2001; Lintunen et al., 2018).Davis等(1999)报道, 当导管直径大于40 μm时, 一次冻融交替使木质部栓塞程度高达95%; Feild和Brodribb (2001)也发现, 常绿针叶植物和导管较小的被子植物经历一次冻融交替后, 其茎水力导度降低最少(几乎为0), 而其他被子植物则平均降低18%-70%.可见, 被子植物木质部忍耐低温能力远低于裸子植物.植物经过多次冻融交替后, 导管或周围组织中产生的气泡可能在整个冬季留存, 直到春季才能重新进入融化水中或者气泡扩张形成栓塞(Hacke & Sperry, 2001; Venturas et al., 2017; Medeiros & Danielson, 2018), 所以导管内水分含量直接影响植物冻融栓塞程度(Choat et al., 2011).而水分含量与导管直径紧密相关(Lens et al., 2013).因此构建直径更小的导管是最直接增加抗冻融栓塞能力的方法.裸子植物直径较小的管胞结构能够在很大程度上减少冻融栓塞的形成, 不仅因为在管胞中 ...
... 植物还有一种忍耐低温胁迫的策略是, 通过改变细胞膜内外的溶质浓度, 使细胞内溶质浓度增加(形成低水势), 从而降低细胞外溶质浓度(Mayr et al., 2007; Lintunen et al., 2016).当冰冻发生时, 细胞外水分首先冻结(主要发生在细胞壁表面、纤维、导管内或者细胞外的空间)(Willick et al., 2018), 冰形成的低水势使细胞内水分进一步流出, 细胞内溶质浓度增加, 从而避免细胞内冰冻导致的细胞死亡(McCully et al., 2004), 能够暂时保护细胞不受冰冻损伤(Mayr & Améglio, 2016). ...
... 冻的树干(Charrier et al., 2017)无法及时补充水分丢失(Bowling et al., 2018), 从而使植物叶片和小枝的木质部含水量降低.另外, 含水量变化还可能会引起质外体组织冰冻时的体积发生变化(Lintunen et al., 2016).低温胁迫使木质组织首先脱水收缩, 再经历液态水冰冻时的体积膨大(+9%)(Améglio et al., 2001).在冰冻前, 植物主动降低质外体含水量, 增加溶质浓度, 此时脱水引起的体积减小能够降低过冷却点(Lintunen et al., 2013; Willick et al., 2019).因此, 冰冻前含水量越低(体积越小), 植物对低温胁迫的忍耐力越强(Lintunen et al., 2018).当冰冻发生时, 由于水分由液态转为固态的体积增加, 对周围细胞壁产生机械压力, 植物需构建更强健的细胞壁(Zhang et al., 2016; Ployet et al., 2018)和改变细胞壁上蛋白组分(Willick et al., 2018)来抵御低温胁迫, 因此, 质外体较高的水分含量可能对植物产生更严重的低温损伤.研究表明, 木质部通过寒冷驯化 后其直径少量收缩(Améglio et al., 2001; Charra- Vaskou et al., 2016; Lintunen et al., 2016).例如, 欧洲赤松(Pinus sylvestris)和欧洲山杨(Populus tremula)木质部直径经寒冷驯化后分别收缩了0.5%和1.0%, 小于渗透胁迫和干旱胁迫引起的收缩量(Lintunen et al., 2016).此外, 植物通过寒冷驯化, 可以降低质外体的含水量, 并将水分转移到冰储存位点, 同时不同冰区之间形成隔离带(Willick et al., 2019), 从而尽可能减少低温损伤. ...
... ; Lintunen et al., 2016).例如, 欧洲赤松(Pinus sylvestris)和欧洲山杨(Populus tremula)木质部直径经寒冷驯化后分别收缩了0.5%和1.0%, 小于渗透胁迫和干旱胁迫引起的收缩量(Lintunen et al., 2016).此外, 植物通过寒冷驯化, 可以降低质外体的含水量, 并将水分转移到冰储存位点, 同时不同冰区之间形成隔离带(Willick et al., 2019), 从而尽可能减少低温损伤. ...
... )木质部直径经寒冷驯化后分别收缩了0.5%和1.0%, 小于渗透胁迫和干旱胁迫引起的收缩量(Lintunen et al., 2016).此外, 植物通过寒冷驯化, 可以降低质外体的含水量, 并将水分转移到冰储存位点, 同时不同冰区之间形成隔离带(Willick et al., 2019), 从而尽可能减少低温损伤. ...
Drivers of apoplastic freezing in gymnosperm and angiosperm branches 5 2018
... 在长期的进化过程中, 植物(尤其是中高纬度、高山和亚高山木本植物)面对冻融栓塞, 逐渐形成了不同的应对策略(Lintunen et al., 2018; Wang et al., 2018; Willick et al., 2018; Yin et al., 2018), 主要包括逃避、忍耐和修复等3种应对策略(图1). ...
... 与动物不同, 植物的固定性使得其更多地采用“被动”的忍耐策略, 例如不同的木质结构、调整组织水分含量和溶质含量等, 以减少低温损伤(Mayr et al., 2014; Arias et al., 2015; Lintunen et al., 2018).被子植物和裸子植物解剖结构的差异, 使二者对低温胁迫的忍耐能力不同(Pittermann & Sperry, 2003; Lintunen et al., 2016).被子植物的导管比裸子植物的管胞结构具有更高的水力效率, 但是其木质部抗冻融栓塞能力较低(Feild & Brodribb, 2001; Lintunen et al., 2018).Davis等(1999)报道, 当导管直径大于40 μm时, 一次冻融交替使木质部栓塞程度高达95%; Feild和Brodribb (2001)也发现, 常绿针叶植物和导管较小的被子植物经历一次冻融交替后, 其茎水力导度降低最少(几乎为0), 而其他被子植物则平均降低18%-70%.可见, 被子植物木质部忍耐低温能力远低于裸子植物.植物经过多次冻融交替后, 导管或周围组织中产生的气泡可能在整个冬季留存, 直到春季才能重新进入融化水中或者气泡扩张形成栓塞(Hacke & Sperry, 2001; Venturas et al., 2017; Medeiros & Danielson, 2018), 所以导管内水分含量直接影响植物冻融栓塞程度(Choat et al., 2011).而水分含量与导管直径紧密相关(Lens et al., 2013).因此构建直径更小的导管是最直接增加抗冻融栓塞能力的方法.裸子植物直径较小的管胞结构能够在很大程度上减少冻融栓塞的形成, 不仅因为在管胞中 ...
... ; Lintunen et al., 2018).Davis等(1999)报道, 当导管直径大于40 μm时, 一次冻融交替使木质部栓塞程度高达95%; Feild和Brodribb (2001)也发现, 常绿针叶植物和导管较小的被子植物经历一次冻融交替后, 其茎水力导度降低最少(几乎为0), 而其他被子植物则平均降低18%-70%.可见, 被子植物木质部忍耐低温能力远低于裸子植物.植物经过多次冻融交替后, 导管或周围组织中产生的气泡可能在整个冬季留存, 直到春季才能重新进入融化水中或者气泡扩张形成栓塞(Hacke & Sperry, 2001; Venturas et al., 2017; Medeiros & Danielson, 2018), 所以导管内水分含量直接影响植物冻融栓塞程度(Choat et al., 2011).而水分含量与导管直径紧密相关(Lens et al., 2013).因此构建直径更小的导管是最直接增加抗冻融栓塞能力的方法.裸子植物直径较小的管胞结构能够在很大程度上减少冻融栓塞的形成, 不仅因为在管胞中 ...
... 木质部结构是影响植物忍耐低温胁迫的主要因素(Hacke et al., 2015, 2017; Lintunen et al., 2018).被子植物采用多个细胞死亡后相互连接构成的导管进行水分运输, 长度在1 mm到几米不等; 裸子植物则依靠单个细胞伸长形成的管胞进行水分运输, 长度在2-6 mm之间(Tyree & Ewers, 1991), 而且管胞直径一般小于导管(Sperry et al., 2006).这种木质部结构差异导致裸子植物和被子植物响应冻融栓塞时产生显著PLC差异(Feild & Brodribb, 2001; Feng et al., 2015; Fernández-Pérez et al., 2018).以往研究表明, 植物在入冬前PLC逐步增加(Zhang et al., 2018), 而导管或管胞的直径差异是植物PLC不同的主要原因(Pittermann & Sperry, 2003; Hacke et al., 2015, 2017).在所有季节中, 裸子植物的PLC均小于被子植物(Granda et al., 2014).例如, 被子植物葡萄牙栎(Quercus faginea)抗空穴化能力较差, 夏季PLC达到80%, 冬季进一步达到90%, 而裸子植物香桧(Juniperus thurifera)和欧洲黑松(Pinus nigra)夏季PLC几乎为0, 冬季仅增加到3%-9% (Granda et al., 2014). ...
... 冻的树干(Charrier et al., 2017)无法及时补充水分丢失(Bowling et al., 2018), 从而使植物叶片和小枝的木质部含水量降低.另外, 含水量变化还可能会引起质外体组织冰冻时的体积发生变化(Lintunen et al., 2016).低温胁迫使木质组织首先脱水收缩, 再经历液态水冰冻时的体积膨大(+9%)(Améglio et al., 2001).在冰冻前, 植物主动降低质外体含水量, 增加溶质浓度, 此时脱水引起的体积减小能够降低过冷却点(Lintunen et al., 2013; Willick et al., 2019).因此, 冰冻前含水量越低(体积越小), 植物对低温胁迫的忍耐力越强(Lintunen et al., 2018).当冰冻发生时, 由于水分由液态转为固态的体积增加, 对周围细胞壁产生机械压力, 植物需构建更强健的细胞壁(Zhang et al., 2016; Ployet et al., 2018)和改变细胞壁上蛋白组分(Willick et al., 2018)来抵御低温胁迫, 因此, 质外体较高的水分含量可能对植物产生更严重的低温损伤.研究表明, 木质部通过寒冷驯化 后其直径少量收缩(Améglio et al., 2001; Charra- Vaskou et al., 2016; Lintunen et al., 2016).例如, 欧洲赤松(Pinus sylvestris)和欧洲山杨(Populus tremula)木质部直径经寒冷驯化后分别收缩了0.5%和1.0%, 小于渗透胁迫和干旱胁迫引起的收缩量(Lintunen et al., 2016).此外, 植物通过寒冷驯化, 可以降低质外体的含水量, 并将水分转移到冰储存位点, 同时不同冰区之间形成隔离带(Willick et al., 2019), 从而尽可能减少低温损伤. ...
Xylem sap chemistry: Seasonal changes in timberline conifers Pinus cembra, Picea abies, and Larix decidua 1 2018
... 植物还可以通过木质部过冷却的方式避免低温及冰冻损伤(Arias et al., 2017).过冷却指植物使细胞中液体的冰点温度远低于液态水冰点的能力(Arias et al., 2015).Rasmussen和MacKenzie (1972)指出, 没有任何结晶核的纯水结冰温度低至-38.1 ℃.因此, 植物细胞内异质结晶核含量影响其过冷却点的高低(Losso et al., 2018).过冷却能力对植物应对突发性的冰冻事件(如热带植物遭遇突发性寒流)十分重要(Reyes-Díaz et al., 2006).Arias等(2015)发现, 在寒冷环境中驯化过的油橄榄(Olea europaea)栽培种, 其叶片冰点温度低至-13 ℃ (远低于环境最低温度), 而且有更高的叶密度和细胞壁刚性, 从而使其溶质渗透性和组织可塑性更高, 质外体水分含量更低; 进一步研究发现, 从夏季到冬季该树种茎最低冰点逐渐降低, 而且其木质部抗空穴化能力与叶片和茎的过冷却能力之间存在权衡(Arias et al., 2017). ...
Dynamics of non-structural carbohydrates in terrestrial plants: A global synthesis 1 2016
... 常绿植物保证叶片安全越冬对其之后的生长具有关键作用(Ball et al., 2004), 因此我们还可以通过分析叶片抗低温胁迫能力来评价植物的低温耐性.目前评价叶片抗低温胁迫能力的指标主要有: 叶片电解质渗出率(Climent et al., 2009; Hoermiller et al., 2018)、可溶性糖含量(Fernández-Pérez et al., 2018)、黎明前叶水势(Taneda & Tateno, 2005)、叶细胞弹性模量(细胞壁刚性越高, 细胞弹性模量越大)(Zhang et al., 2016)等.Fernández-Pérez等(2018)报道, 伊比利亚半岛6个松属树种的分布与其叶片低温忍耐能力有关, 生长在寒冷环境中的针叶的电解质渗出率和可溶性糖含量较低, 且黎明前叶水势对针叶低温损伤有影响.Martínez-Vilalta等(2016)整合分析全球尺度121个研究中177个物种非结构性碳(淀粉和可溶性糖)数据发现, 北方和温带生物区系物种叶片非结构性碳含量最高, 而且在入冬过程中逐步增加.Zhang等(2016)发现, 叶片细胞壁强度的增加, 降低了质外体冰冻对细胞膜的物理损伤, 使植物在较低的温度下得以存活.上述几个评价指标中, 电解质渗出率是最常用的评价植物组织损伤的指标(Charrier et al., 2013; Arias et al., 2015; Mayr & Améglio, 2016).通常采用电解质渗出率达50%时所对应的温度(LT50)来评价植物的低温抗性: LT50越低, 则低温抗性越强(Kreyling et al., 2015; Bachofen et al., 2016; Arias et al., 2017).例如, Arias等(2017)报道, 油橄榄的LT50从夏季到冬季呈现逐步降低的趋势, 这表明随着温度的降低, 通过不断驯化, 油橄榄的低温抗性逐步增强. ...
Freezing stress in tree xylem 3 2016
... 栓塞是降低木质部内水分运输效率的关键因子(Choat et al., 2012; 金鹰等, 2016), 主要包括干旱栓塞和冻融栓塞(Améglio et al., 2002; Willson & Jackson, 2006; Venturas et al., 2017).两种栓塞形成的主要机理均为空气进入导管或管胞, 从而阻断木质部水柱, 形成栓塞(Sperry & Sullivan, 1992), 而不同之处在于引发空气进入的过程(Hacke & Sperry, 2001).根据内聚力-张力理论, 木质部导管内水分持续运输动力主要来自叶片蒸腾失水引起的拉力, 因此导管中的水分始终处于亚稳态(Sperry & Love, 2015).当蒸腾加剧和/或土壤水分供应不足而使木质部导管内负压超过一定阈值时, 就会造成其水柱断裂, 致使周围组织中的气体进入导管, 从而形成干旱栓塞(Choat et al., 2018).而冻融栓塞主要发生在木质部经历多次冻融循环的过程中(Sperry & Sullivan, 1992; Feild & Brodribb, 2001; Charra-?Vaskou et al., 2016).当木质部水分冰冻时, 水中原有溶解气体因在冰中的溶解度很低而逸散到导管中(Lintunen et al., 2014).而当冰冻融化时, 导管中的留存气体或者重新溶进水中(气泡压力大于大气压时, Hacke & Sperry, 2001), 或者由于冰冻压力的释放(冰融化后体积减小)等使气体在导管中扩大(Lemoine et al., 1999; Mayr et al., 2002; Pittermann & Sperry, 2006), 多次冻融交替之后, 导管中气体不断增加和扩张, 最终引发冻融栓塞, 该现象也被称为“融化-扩张假说” (Mayr & Sperry, 2010; Mayr & Améglio, 2016).干旱栓塞和冻融栓塞均会阻碍植物水分从土壤到叶片的运输, 从而降低叶片的光合速率(Brodribb et al., 2007; Reich, 2014; Jin et al., 2016), 导致植物生产力降低甚至死亡(McDowell et al., 2008; Hoffmann et al., 2011; McDowell, 2011; Choat et al., 2018). ...
... 植物还有一种忍耐低温胁迫的策略是, 通过改变细胞膜内外的溶质浓度, 使细胞内溶质浓度增加(形成低水势), 从而降低细胞外溶质浓度(Mayr et al., 2007; Lintunen et al., 2016).当冰冻发生时, 细胞外水分首先冻结(主要发生在细胞壁表面、纤维、导管内或者细胞外的空间)(Willick et al., 2018), 冰形成的低水势使细胞内水分进一步流出, 细胞内溶质浓度增加, 从而避免细胞内冰冻导致的细胞死亡(McCully et al., 2004), 能够暂时保护细胞不受冰冻损伤(Mayr & Améglio, 2016). ...
... 常绿植物保证叶片安全越冬对其之后的生长具有关键作用(Ball et al., 2004), 因此我们还可以通过分析叶片抗低温胁迫能力来评价植物的低温耐性.目前评价叶片抗低温胁迫能力的指标主要有: 叶片电解质渗出率(Climent et al., 2009; Hoermiller et al., 2018)、可溶性糖含量(Fernández-Pérez et al., 2018)、黎明前叶水势(Taneda & Tateno, 2005)、叶细胞弹性模量(细胞壁刚性越高, 细胞弹性模量越大)(Zhang et al., 2016)等.Fernández-Pérez等(2018)报道, 伊比利亚半岛6个松属树种的分布与其叶片低温忍耐能力有关, 生长在寒冷环境中的针叶的电解质渗出率和可溶性糖含量较低, 且黎明前叶水势对针叶低温损伤有影响.Martínez-Vilalta等(2016)整合分析全球尺度121个研究中177个物种非结构性碳(淀粉和可溶性糖)数据发现, 北方和温带生物区系物种叶片非结构性碳含量最高, 而且在入冬过程中逐步增加.Zhang等(2016)发现, 叶片细胞壁强度的增加, 降低了质外体冰冻对细胞膜的物理损伤, 使植物在较低的温度下得以存活.上述几个评价指标中, 电解质渗出率是最常用的评价植物组织损伤的指标(Charrier et al., 2013; Arias et al., 2015; Mayr & Améglio, 2016).通常采用电解质渗出率达50%时所对应的温度(LT50)来评价植物的低温抗性: LT50越低, 则低温抗性越强(Kreyling et al., 2015; Bachofen et al., 2016; Arias et al., 2017).例如, Arias等(2017)报道, 油橄榄的LT50从夏季到冬季呈现逐步降低的趋势, 这表明随着温度的降低, 通过不断驯化, 油橄榄的低温抗性逐步增强. ...
Embolism formation during freezing in the wood of Picea abies 2 2007
... 植物还有一种忍耐低温胁迫的策略是, 通过改变细胞膜内外的溶质浓度, 使细胞内溶质浓度增加(形成低水势), 从而降低细胞外溶质浓度(Mayr et al., 2007; Lintunen et al., 2016).当冰冻发生时, 细胞外水分首先冻结(主要发生在细胞壁表面、纤维、导管内或者细胞外的空间)(Willick et al., 2018), 冰形成的低水势使细胞内水分进一步流出, 细胞内溶质浓度增加, 从而避免细胞内冰冻导致的细胞死亡(McCully et al., 2004), 能够暂时保护细胞不受冰冻损伤(Mayr & Améglio, 2016). ...
... 木质部低温抗性、干旱抗性和水力效率三者间是否存在多元权衡现象?目前对此问题的综合研究较少, 但已有研究指出, 干旱显著增加植物的低温抗性(Kreyling et al., 2012; Walter et al., 2013; Sierra-?Almeida et al., 2016; Sperling et al., 2017).这是因为植物通过驯化改变了细胞对脱水的忍耐能力(Buchner & Neuner, 2011), 其中包括基因表达的改变导致的细胞膜结构变化或者细胞化学成分变化(Verhoeven et al., 2018).例如, Medeiros和Pockman (2011)报道, 在-15 ℃环境中干旱处理使Larrea tridentate植株细胞死亡减少、叶留存量增加、水力导度和气体交换速率升高, 这表明干旱处理显著增加了其低温抗性.Hoffman等(2012)也发现, 干旱处理能够显著增加黑麦草(Lolium perenne)的低温抗性.然而, Kong和Henry (2016)研究发现, 早春低温处理的草地早熟禾(Poa pratensis)的抗旱性显著增加(生物量和存活率增加), 但夏季干旱处理并未增加其低温抗性, 这表明干旱处理对植物低温抗性无影响的现象也存在.同样, Bachofen等(2016)也发现, 干旱处理没有增加地中海松属3个树种的低温抗性, 而且推测该现象可能仅在短期极端条件下发生.Yin等(2018)同时对低温抗性、干旱抗性和水力效率进行研究发现, 抗干旱栓塞与水力效率存在权衡, 抗冷冻疲劳与水力效率间也存在权衡现象, 支持多元权衡现象.然而, 当植物同时面对干旱和低温胁迫时(如高山树线植物)(Mayr et al., 2007; Charrier et al., 2017), 多次冻融交替结合干旱胁迫可能引起PLC增加(Mayr et al., 2003; Charrier et al., 2014).Mayr等(2006)报道, 生长在林线处的针叶树种, 每个冬季经历至少100次冻融交替, 再加上干旱胁迫, 二者共同引起欧洲云杉(Picea abies) PLC高达100%.因此, 生长在低温和干旱胁迫共存环境中的植物, 其PLC可能更高, 间接表明三者间存在一定程度的权衡.综上所述, 我们构建了低温抗性、干旱抗性和水力效率之间多元权衡概念图(图2). ...
Frost drought in conifers at the alpine timberline: Xylem dysfunction and adaptations 1 2006
... 木质部低温抗性、干旱抗性和水力效率三者间是否存在多元权衡现象?目前对此问题的综合研究较少, 但已有研究指出, 干旱显著增加植物的低温抗性(Kreyling et al., 2012; Walter et al., 2013; Sierra-?Almeida et al., 2016; Sperling et al., 2017).这是因为植物通过驯化改变了细胞对脱水的忍耐能力(Buchner & Neuner, 2011), 其中包括基因表达的改变导致的细胞膜结构变化或者细胞化学成分变化(Verhoeven et al., 2018).例如, Medeiros和Pockman (2011)报道, 在-15 ℃环境中干旱处理使Larrea tridentate植株细胞死亡减少、叶留存量增加、水力导度和气体交换速率升高, 这表明干旱处理显著增加了其低温抗性.Hoffman等(2012)也发现, 干旱处理能够显著增加黑麦草(Lolium perenne)的低温抗性.然而, Kong和Henry (2016)研究发现, 早春低温处理的草地早熟禾(Poa pratensis)的抗旱性显著增加(生物量和存活率增加), 但夏季干旱处理并未增加其低温抗性, 这表明干旱处理对植物低温抗性无影响的现象也存在.同样, Bachofen等(2016)也发现, 干旱处理没有增加地中海松属3个树种的低温抗性, 而且推测该现象可能仅在短期极端条件下发生.Yin等(2018)同时对低温抗性、干旱抗性和水力效率进行研究发现, 抗干旱栓塞与水力效率存在权衡, 抗冷冻疲劳与水力效率间也存在权衡现象, 支持多元权衡现象.然而, 当植物同时面对干旱和低温胁迫时(如高山树线植物)(Mayr et al., 2007; Charrier et al., 2017), 多次冻融交替结合干旱胁迫可能引起PLC增加(Mayr et al., 2003; Charrier et al., 2014).Mayr等(2006)报道, 生长在林线处的针叶树种, 每个冬季经历至少100次冻融交替, 再加上干旱胁迫, 二者共同引起欧洲云杉(Picea abies) PLC高达100%.因此, 生长在低温和干旱胁迫共存环境中的植物, 其PLC可能更高, 间接表明三者间存在一定程度的权衡.综上所述, 我们构建了低温抗性、干旱抗性和水力效率之间多元权衡概念图(图2). ...
Uptake of water via branches helps timberline conifers refill embolized xylem in late winter 1 2014
... 与动物不同, 植物的固定性使得其更多地采用“被动”的忍耐策略, 例如不同的木质结构、调整组织水分含量和溶质含量等, 以减少低温损伤(Mayr et al., 2014; Arias et al., 2015; Lintunen et al., 2018).被子植物和裸子植物解剖结构的差异, 使二者对低温胁迫的忍耐能力不同(Pittermann & Sperry, 2003; Lintunen et al., 2016).被子植物的导管比裸子植物的管胞结构具有更高的水力效率, 但是其木质部抗冻融栓塞能力较低(Feild & Brodribb, 2001; Lintunen et al., 2018).Davis等(1999)报道, 当导管直径大于40 μm时, 一次冻融交替使木质部栓塞程度高达95%; Feild和Brodribb (2001)也发现, 常绿针叶植物和导管较小的被子植物经历一次冻融交替后, 其茎水力导度降低最少(几乎为0), 而其他被子植物则平均降低18%-70%.可见, 被子植物木质部忍耐低温能力远低于裸子植物.植物经过多次冻融交替后, 导管或周围组织中产生的气泡可能在整个冬季留存, 直到春季才能重新进入融化水中或者气泡扩张形成栓塞(Hacke & Sperry, 2001; Venturas et al., 2017; Medeiros & Danielson, 2018), 所以导管内水分含量直接影响植物冻融栓塞程度(Choat et al., 2011).而水分含量与导管直径紧密相关(Lens et al., 2013).因此构建直径更小的导管是最直接增加抗冻融栓塞能力的方法.裸子植物直径较小的管胞结构能够在很大程度上减少冻融栓塞的形成, 不仅因为在管胞中 ...
Winter at the alpine timberline. Why does embolism occur in Norway spruce but not in stone pine? 1 2003
... 木质部低温抗性、干旱抗性和水力效率三者间是否存在多元权衡现象?目前对此问题的综合研究较少, 但已有研究指出, 干旱显著增加植物的低温抗性(Kreyling et al., 2012; Walter et al., 2013; Sierra-?Almeida et al., 2016; Sperling et al., 2017).这是因为植物通过驯化改变了细胞对脱水的忍耐能力(Buchner & Neuner, 2011), 其中包括基因表达的改变导致的细胞膜结构变化或者细胞化学成分变化(Verhoeven et al., 2018).例如, Medeiros和Pockman (2011)报道, 在-15 ℃环境中干旱处理使Larrea tridentate植株细胞死亡减少、叶留存量增加、水力导度和气体交换速率升高, 这表明干旱处理显著增加了其低温抗性.Hoffman等(2012)也发现, 干旱处理能够显著增加黑麦草(Lolium perenne)的低温抗性.然而, Kong和Henry (2016)研究发现, 早春低温处理的草地早熟禾(Poa pratensis)的抗旱性显著增加(生物量和存活率增加), 但夏季干旱处理并未增加其低温抗性, 这表明干旱处理对植物低温抗性无影响的现象也存在.同样, Bachofen等(2016)也发现, 干旱处理没有增加地中海松属3个树种的低温抗性, 而且推测该现象可能仅在短期极端条件下发生.Yin等(2018)同时对低温抗性、干旱抗性和水力效率进行研究发现, 抗干旱栓塞与水力效率存在权衡, 抗冷冻疲劳与水力效率间也存在权衡现象, 支持多元权衡现象.然而, 当植物同时面对干旱和低温胁迫时(如高山树线植物)(Mayr et al., 2007; Charrier et al., 2017), 多次冻融交替结合干旱胁迫可能引起PLC增加(Mayr et al., 2003; Charrier et al., 2014).Mayr等(2006)报道, 生长在林线处的针叶树种, 每个冬季经历至少100次冻融交替, 再加上干旱胁迫, 二者共同引起欧洲云杉(Picea abies) PLC高达100%.因此, 生长在低温和干旱胁迫共存环境中的植物, 其PLC可能更高, 间接表明三者间存在一定程度的权衡.综上所述, 我们构建了低温抗性、干旱抗性和水力效率之间多元权衡概念图(图2). ...
Freeze-thaw-induced embolism in Pinus contorta: Centrifuge experiments validate the ‘thaw-?expansion hypothesis’ but conflict with ultrasonic emission data 1 2010
... 栓塞是降低木质部内水分运输效率的关键因子(Choat et al., 2012; 金鹰等, 2016), 主要包括干旱栓塞和冻融栓塞(Améglio et al., 2002; Willson & Jackson, 2006; Venturas et al., 2017).两种栓塞形成的主要机理均为空气进入导管或管胞, 从而阻断木质部水柱, 形成栓塞(Sperry & Sullivan, 1992), 而不同之处在于引发空气进入的过程(Hacke & Sperry, 2001).根据内聚力-张力理论, 木质部导管内水分持续运输动力主要来自叶片蒸腾失水引起的拉力, 因此导管中的水分始终处于亚稳态(Sperry & Love, 2015).当蒸腾加剧和/或土壤水分供应不足而使木质部导管内负压超过一定阈值时, 就会造成其水柱断裂, 致使周围组织中的气体进入导管, 从而形成干旱栓塞(Choat et al., 2018).而冻融栓塞主要发生在木质部经历多次冻融循环的过程中(Sperry & Sullivan, 1992; Feild & Brodribb, 2001; Charra-?Vaskou et al., 2016).当木质部水分冰冻时, 水中原有溶解气体因在冰中的溶解度很低而逸散到导管中(Lintunen et al., 2014).而当冰冻融化时, 导管中的留存气体或者重新溶进水中(气泡压力大于大气压时, Hacke & Sperry, 2001), 或者由于冰冻压力的释放(冰融化后体积减小)等使气体在导管中扩大(Lemoine et al., 1999; Mayr et al., 2002; Pittermann & Sperry, 2006), 多次冻融交替之后, 导管中气体不断增加和扩张, 最终引发冻融栓塞, 该现象也被称为“融化-扩张假说” (Mayr & Sperry, 2010; Mayr & Améglio, 2016).干旱栓塞和冻融栓塞均会阻碍植物水分从土壤到叶片的运输, 从而降低叶片的光合速率(Brodribb et al., 2007; Reich, 2014; Jin et al., 2016), 导致植物生产力降低甚至死亡(McDowell et al., 2008; Hoffmann et al., 2011; McDowell, 2011; Choat et al., 2018). ...
Winter-drought induced embolism in Norway spruce (Picea abies) at the Alpine timberline 1 2002
... 栓塞是降低木质部内水分运输效率的关键因子(Choat et al., 2012; 金鹰等, 2016), 主要包括干旱栓塞和冻融栓塞(Améglio et al., 2002; Willson & Jackson, 2006; Venturas et al., 2017).两种栓塞形成的主要机理均为空气进入导管或管胞, 从而阻断木质部水柱, 形成栓塞(Sperry & Sullivan, 1992), 而不同之处在于引发空气进入的过程(Hacke & Sperry, 2001).根据内聚力-张力理论, 木质部导管内水分持续运输动力主要来自叶片蒸腾失水引起的拉力, 因此导管中的水分始终处于亚稳态(Sperry & Love, 2015).当蒸腾加剧和/或土壤水分供应不足而使木质部导管内负压超过一定阈值时, 就会造成其水柱断裂, 致使周围组织中的气体进入导管, 从而形成干旱栓塞(Choat et al., 2018).而冻融栓塞主要发生在木质部经历多次冻融循环的过程中(Sperry & Sullivan, 1992; Feild & Brodribb, 2001; Charra-?Vaskou et al., 2016).当木质部水分冰冻时, 水中原有溶解气体因在冰中的溶解度很低而逸散到导管中(Lintunen et al., 2014).而当冰冻融化时, 导管中的留存气体或者重新溶进水中(气泡压力大于大气压时, Hacke & Sperry, 2001), 或者由于冰冻压力的释放(冰融化后体积减小)等使气体在导管中扩大(Lemoine et al., 1999; Mayr et al., 2002; Pittermann & Sperry, 2006), 多次冻融交替之后, 导管中气体不断增加和扩张, 最终引发冻融栓塞, 该现象也被称为“融化-扩张假说” (Mayr & Sperry, 2010; Mayr & Améglio, 2016).干旱栓塞和冻融栓塞均会阻碍植物水分从土壤到叶片的运输, 从而降低叶片的光合速率(Brodribb et al., 2007; Reich, 2014; Jin et al., 2016), 导致植物生产力降低甚至死亡(McDowell et al., 2008; Hoffmann et al., 2011; McDowell, 2011; Choat et al., 2018). ...
The management of extracellular ice by petioles of frost-resistant herbaceous plants 1 2004
... 植物还有一种忍耐低温胁迫的策略是, 通过改变细胞膜内外的溶质浓度, 使细胞内溶质浓度增加(形成低水势), 从而降低细胞外溶质浓度(Mayr et al., 2007; Lintunen et al., 2016).当冰冻发生时, 细胞外水分首先冻结(主要发生在细胞壁表面、纤维、导管内或者细胞外的空间)(Willick et al., 2018), 冰形成的低水势使细胞内水分进一步流出, 细胞内溶质浓度增加, 从而避免细胞内冰冻导致的细胞死亡(McCully et al., 2004), 能够暂时保护细胞不受冰冻损伤(Mayr & Améglio, 2016). ...
... 栓塞是降低木质部内水分运输效率的关键因子(Choat et al., 2012; 金鹰等, 2016), 主要包括干旱栓塞和冻融栓塞(Améglio et al., 2002; Willson & Jackson, 2006; Venturas et al., 2017).两种栓塞形成的主要机理均为空气进入导管或管胞, 从而阻断木质部水柱, 形成栓塞(Sperry & Sullivan, 1992), 而不同之处在于引发空气进入的过程(Hacke & Sperry, 2001).根据内聚力-张力理论, 木质部导管内水分持续运输动力主要来自叶片蒸腾失水引起的拉力, 因此导管中的水分始终处于亚稳态(Sperry & Love, 2015).当蒸腾加剧和/或土壤水分供应不足而使木质部导管内负压超过一定阈值时, 就会造成其水柱断裂, 致使周围组织中的气体进入导管, 从而形成干旱栓塞(Choat et al., 2018).而冻融栓塞主要发生在木质部经历多次冻融循环的过程中(Sperry & Sullivan, 1992; Feild & Brodribb, 2001; Charra-?Vaskou et al., 2016).当木质部水分冰冻时, 水中原有溶解气体因在冰中的溶解度很低而逸散到导管中(Lintunen et al., 2014).而当冰冻融化时, 导管中的留存气体或者重新溶进水中(气泡压力大于大气压时, Hacke & Sperry, 2001), 或者由于冰冻压力的释放(冰融化后体积减小)等使气体在导管中扩大(Lemoine et al., 1999; Mayr et al., 2002; Pittermann & Sperry, 2006), 多次冻融交替之后, 导管中气体不断增加和扩张, 最终引发冻融栓塞, 该现象也被称为“融化-扩张假说” (Mayr & Sperry, 2010; Mayr & Améglio, 2016).干旱栓塞和冻融栓塞均会阻碍植物水分从土壤到叶片的运输, 从而降低叶片的光合速率(Brodribb et al., 2007; Reich, 2014; Jin et al., 2016), 导致植物生产力降低甚至死亡(McDowell et al., 2008; Hoffmann et al., 2011; McDowell, 2011; Choat et al., 2018). ...
Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? 1 2008
... 栓塞是降低木质部内水分运输效率的关键因子(Choat et al., 2012; 金鹰等, 2016), 主要包括干旱栓塞和冻融栓塞(Améglio et al., 2002; Willson & Jackson, 2006; Venturas et al., 2017).两种栓塞形成的主要机理均为空气进入导管或管胞, 从而阻断木质部水柱, 形成栓塞(Sperry & Sullivan, 1992), 而不同之处在于引发空气进入的过程(Hacke & Sperry, 2001).根据内聚力-张力理论, 木质部导管内水分持续运输动力主要来自叶片蒸腾失水引起的拉力, 因此导管中的水分始终处于亚稳态(Sperry & Love, 2015).当蒸腾加剧和/或土壤水分供应不足而使木质部导管内负压超过一定阈值时, 就会造成其水柱断裂, 致使周围组织中的气体进入导管, 从而形成干旱栓塞(Choat et al., 2018).而冻融栓塞主要发生在木质部经历多次冻融循环的过程中(Sperry & Sullivan, 1992; Feild & Brodribb, 2001; Charra-?Vaskou et al., 2016).当木质部水分冰冻时, 水中原有溶解气体因在冰中的溶解度很低而逸散到导管中(Lintunen et al., 2014).而当冰冻融化时, 导管中的留存气体或者重新溶进水中(气泡压力大于大气压时, Hacke & Sperry, 2001), 或者由于冰冻压力的释放(冰融化后体积减小)等使气体在导管中扩大(Lemoine et al., 1999; Mayr et al., 2002; Pittermann & Sperry, 2006), 多次冻融交替之后, 导管中气体不断增加和扩张, 最终引发冻融栓塞, 该现象也被称为“融化-扩张假说” (Mayr & Sperry, 2010; Mayr & Améglio, 2016).干旱栓塞和冻融栓塞均会阻碍植物水分从土壤到叶片的运输, 从而降低叶片的光合速率(Brodribb et al., 2007; Reich, 2014; Jin et al., 2016), 导致植物生产力降低甚至死亡(McDowell et al., 2008; Hoffmann et al., 2011; McDowell, 2011; Choat et al., 2018). ...
Renewed interest in whole-plant physiology sheds light on the complexity of plant stress response architecture 1 2018
... 与动物不同, 植物的固定性使得其更多地采用“被动”的忍耐策略, 例如不同的木质结构、调整组织水分含量和溶质含量等, 以减少低温损伤(Mayr et al., 2014; Arias et al., 2015; Lintunen et al., 2018).被子植物和裸子植物解剖结构的差异, 使二者对低温胁迫的忍耐能力不同(Pittermann & Sperry, 2003; Lintunen et al., 2016).被子植物的导管比裸子植物的管胞结构具有更高的水力效率, 但是其木质部抗冻融栓塞能力较低(Feild & Brodribb, 2001; Lintunen et al., 2018).Davis等(1999)报道, 当导管直径大于40 μm时, 一次冻融交替使木质部栓塞程度高达95%; Feild和Brodribb (2001)也发现, 常绿针叶植物和导管较小的被子植物经历一次冻融交替后, 其茎水力导度降低最少(几乎为0), 而其他被子植物则平均降低18%-70%.可见, 被子植物木质部忍耐低温能力远低于裸子植物.植物经过多次冻融交替后, 导管或周围组织中产生的气泡可能在整个冬季留存, 直到春季才能重新进入融化水中或者气泡扩张形成栓塞(Hacke & Sperry, 2001; Venturas et al., 2017; Medeiros & Danielson, 2018), 所以导管内水分含量直接影响植物冻融栓塞程度(Choat et al., 2011).而水分含量与导管直径紧密相关(Lens et al., 2013).因此构建直径更小的导管是最直接增加抗冻融栓塞能力的方法.裸子植物直径较小的管胞结构能够在很大程度上减少冻融栓塞的形成, 不仅因为在管胞中 ...
Drought increases freezing tolerance of both leaves and xylem of 1 2011
... 木质部低温抗性、干旱抗性和水力效率三者间是否存在多元权衡现象?目前对此问题的综合研究较少, 但已有研究指出, 干旱显著增加植物的低温抗性(Kreyling et al., 2012; Walter et al., 2013; Sierra-?Almeida et al., 2016; Sperling et al., 2017).这是因为植物通过驯化改变了细胞对脱水的忍耐能力(Buchner & Neuner, 2011), 其中包括基因表达的改变导致的细胞膜结构变化或者细胞化学成分变化(Verhoeven et al., 2018).例如, Medeiros和Pockman (2011)报道, 在-15 ℃环境中干旱处理使Larrea tridentate植株细胞死亡减少、叶留存量增加、水力导度和气体交换速率升高, 这表明干旱处理显著增加了其低温抗性.Hoffman等(2012)也发现, 干旱处理能够显著增加黑麦草(Lolium perenne)的低温抗性.然而, Kong和Henry (2016)研究发现, 早春低温处理的草地早熟禾(Poa pratensis)的抗旱性显著增加(生物量和存活率增加), 但夏季干旱处理并未增加其低温抗性, 这表明干旱处理对植物低温抗性无影响的现象也存在.同样, Bachofen等(2016)也发现, 干旱处理没有增加地中海松属3个树种的低温抗性, 而且推测该现象可能仅在短期极端条件下发生.Yin等(2018)同时对低温抗性、干旱抗性和水力效率进行研究发现, 抗干旱栓塞与水力效率存在权衡, 抗冷冻疲劳与水力效率间也存在权衡现象, 支持多元权衡现象.然而, 当植物同时面对干旱和低温胁迫时(如高山树线植物)(Mayr et al., 2007; Charrier et al., 2017), 多次冻融交替结合干旱胁迫可能引起PLC增加(Mayr et al., 2003; Charrier et al., 2014).Mayr等(2006)报道, 生长在林线处的针叶树种, 每个冬季经历至少100次冻融交替, 再加上干旱胁迫, 二者共同引起欧洲云杉(Picea abies) PLC高达100%.因此, 生长在低温和干旱胁迫共存环境中的植物, 其PLC可能更高, 间接表明三者间存在一定程度的权衡.综上所述, 我们构建了低温抗性、干旱抗性和水力效率之间多元权衡概念图(图2). ...
Refilling embolized xylem conduits: Is it a matter of phloem unloading? 1 2011
... 修复是植物应对冻融栓塞的又一重要策略(Preston & Sandve, 2013; Wang et al., 2018).有研究表明, 采用正压(主要包括根压和茎压)恢复水力功能的植物可能具有更强的抗冷冻疲劳能力(即木质部冰冻后对水力功能丢失更敏感的现象)(Niu et al., 2017; Yin et al., 2018), 这是植物对冻融栓塞的一种主动修复.Hao等(2013)发现, 早春纸桦(Betula papyrifera)树干水分含量明显高于生长季, 表明根压在修复冻融栓塞导管的过程中具有重要作用.Améglio等(2002)报道, 冬季胡桃(Juglans regia) PLC可高达到100%, 但是由于冬季和春季茎压修复, 木质部冻融栓塞程度有一定的降低.另外, 近期研究表明, 能产生木质部正压的植物, 其抗冻融栓塞能力可能不受导管直径的影响(Niu et al., 2017).根压和茎压主要来源于糖类水解成的蔗糖被运输到栓塞导管中而产生渗透浓度梯度(Nardini et al., 2011; Cao et al., 2012), 从而使水分流向栓塞导管, 即渗透调节假说(金鹰等, 2016).两者的不同之处在于: 根压主要在无蒸腾流情况下产生作用(Swaef et al., 2013), 且修复栓塞作用较小(Salleo et al., 2004), 特别是对高大植物上层木质部栓塞的修复作用很小(Westhoff et al., 2008).而茎压产生的渗透梯度可能来自韧皮部的糖类(韧皮部卸载假说, Salleo et al., 2004; Hölttä et al., 2018), 因此在多数情况下均有茎压存在, 这也被称为“新的再冲水” (novel refilling)假说(Brodersen & McElrone, 2013; 金鹰等, 2016).Améglio等(2004)发现, 胡桃在深冬时茎中淀粉含量最低, 在发芽前茎中再次合成淀粉, 这不仅表明胡桃在冬季低温下进行淀粉和蔗糖的转换, 而且蔗糖累积是其冻融栓塞修复中的关键.张海燕等(2013)也发现, 12个温带树种老枝的淀粉含量变化具有相同趋势. ...
Leaf hydraulic capacity and drought vulnerability: Possible trade-offs and correlations with climate across three major biomes 2 2014
... 木质部的水力效率-安全权衡是植物适应环境和进化的结果(Blackman et al., 2010; Jin et al., 2016), 影响植物的竞争力和生产力(Nardini & Luglio, 2014).水力效率一般用植物水力导度表示, 而水力安全常用抗干旱栓塞能力(P50)表示(Santiago et al., 2018).Nardini等(2012)报道, 来自温带的6种植物(槭属(Acer)和栎属(Quercus)各3种)的水力导度、P50和比叶质量之间存在显著相关性, 支持水力效率-安全权衡, 且增加叶碳投入可以提高水力丢失的忍耐能力.Nardini和Luglio (2014)整合130个木本被子植物叶片水力数据后也发现存在叶水力效率-安全权衡关系, 而且生长在低降水量地区的植物的P50 (负值)和叶水力导度均较低.Gleason等(2016)综合分析335种被子植物和89种裸子植物的茎水力数据, 发现虽然水力导度和抗干旱栓塞能力之间存在弱相关性(支持水力效率-安全权衡), 但没有发现高效率和高安全性的植物.因此, 植物普遍存在不同程度的水力效率-安全权衡关系, 但其关系的紧密程度受导管特征(Lens et al., 2011; Hajek et al., 2014)、木质材性(金鹰和王传宽, 2016)、树高(Prendin et al., 2018)等因素的影响. ...
... 和比叶质量之间存在显著相关性, 支持水力效率-安全权衡, 且增加叶碳投入可以提高水力丢失的忍耐能力.Nardini和Luglio (2014)整合130个木本被子植物叶片水力数据后也发现存在叶水力效率-安全权衡关系, 而且生长在低降水量地区的植物的P50 (负值)和叶水力导度均较低.Gleason等(2016)综合分析335种被子植物和89种裸子植物的茎水力数据, 发现虽然水力导度和抗干旱栓塞能力之间存在弱相关性(支持水力效率-安全权衡), 但没有发现高效率和高安全性的植物.因此, 植物普遍存在不同程度的水力效率-安全权衡关系, 但其关系的紧密程度受导管特征(Lens et al., 2011; Hajek et al., 2014)、木质材性(金鹰和王传宽, 2016)、树高(Prendin et al., 2018)等因素的影响. ...
Trade-offs between leaf hydraulic capacity and drought vulnerability: Morpho-?anatomical bases, carbon costs and ecological consequences 1 2012
... 木质部的水力效率-安全权衡是植物适应环境和进化的结果(Blackman et al., 2010; Jin et al., 2016), 影响植物的竞争力和生产力(Nardini & Luglio, 2014).水力效率一般用植物水力导度表示, 而水力安全常用抗干旱栓塞能力(P50)表示(Santiago et al., 2018).Nardini等(2012)报道, 来自温带的6种植物(槭属(Acer)和栎属(Quercus)各3种)的水力导度、P50和比叶质量之间存在显著相关性, 支持水力效率-安全权衡, 且增加叶碳投入可以提高水力丢失的忍耐能力.Nardini和Luglio (2014)整合130个木本被子植物叶片水力数据后也发现存在叶水力效率-安全权衡关系, 而且生长在低降水量地区的植物的P50 (负值)和叶水力导度均较低.Gleason等(2016)综合分析335种被子植物和89种裸子植物的茎水力数据, 发现虽然水力导度和抗干旱栓塞能力之间存在弱相关性(支持水力效率-安全权衡), 但没有发现高效率和高安全性的植物.因此, 植物普遍存在不同程度的水力效率-安全权衡关系, 但其关系的紧密程度受导管特征(Lens et al., 2011; Hajek et al., 2014)、木质材性(金鹰和王传宽, 2016)、树高(Prendin et al., 2018)等因素的影响. ...
Divergence in strategies for coping with winter embolism among co-occurring temperate tree species: The role of positive xylem pressure, wood type and tree stature 6 2017
... 北方森林和温带森林几乎占全球森林总面积的1/3 (Reich et al., 2016), 其植物面临的最主要环境胁迫之一是冬季低温.低温使植物地上器官木质部中的水分逐步冻结(Niu et al., 2017), 所以植物如何最大程度减少低温胁迫引起的损伤对其安全越冬(Hacke et al., 2015; Kreyling et al., 2015; Charrier et al., 2017)、生长、生存以及分布均具有重要作用(Schreiber et al., 2013; Charrier et al., 2015; Niu et al., 2017).低温时期冻融诱导形成的栓塞是低温胁迫对植物产生损伤的重要因素(Zanne et al., 2014; Zhang et al., 2018).许多研究发现, 植物抗冻融栓塞能力不仅影响其在低温时期的损伤程度和碳消耗(Bowling et al., 2018; Yin et al., 2018), 还显著影响随后生长季中的水分供应能力(冷冻疲劳, Feng et al., 2015; Zhang et al., 2018).但与干旱栓塞相比, 目前对木质部冻融栓塞的形成以及植物响应和调节机制的理解不足(Choat et al., 2011; Yin et al., 2018).为了推进该领域的研究, 本文先综述木质部冻融栓塞的形成机制和植物的逃避、忍耐、修复等3种冻融栓塞的应对策略, 然后总结植物低温抗性的生理表现、评价和影响因子, 在此基础上讨论低温抗性、干旱抗性和水力效率之间的多元权衡关系, 最后提出今后该领域中的优先研究问题. ...
... ; Niu et al., 2017).低温时期冻融诱导形成的栓塞是低温胁迫对植物产生损伤的重要因素(Zanne et al., 2014; Zhang et al., 2018).许多研究发现, 植物抗冻融栓塞能力不仅影响其在低温时期的损伤程度和碳消耗(Bowling et al., 2018; Yin et al., 2018), 还显著影响随后生长季中的水分供应能力(冷冻疲劳, Feng et al., 2015; Zhang et al., 2018).但与干旱栓塞相比, 目前对木质部冻融栓塞的形成以及植物响应和调节机制的理解不足(Choat et al., 2011; Yin et al., 2018).为了推进该领域的研究, 本文先综述木质部冻融栓塞的形成机制和植物的逃避、忍耐、修复等3种冻融栓塞的应对策略, 然后总结植物低温抗性的生理表现、评价和影响因子, 在此基础上讨论低温抗性、干旱抗性和水力效率之间的多元权衡关系, 最后提出今后该领域中的优先研究问题. ...
... 修复是植物应对冻融栓塞的又一重要策略(Preston & Sandve, 2013; Wang et al., 2018).有研究表明, 采用正压(主要包括根压和茎压)恢复水力功能的植物可能具有更强的抗冷冻疲劳能力(即木质部冰冻后对水力功能丢失更敏感的现象)(Niu et al., 2017; Yin et al., 2018), 这是植物对冻融栓塞的一种主动修复.Hao等(2013)发现, 早春纸桦(Betula papyrifera)树干水分含量明显高于生长季, 表明根压在修复冻融栓塞导管的过程中具有重要作用.Améglio等(2002)报道, 冬季胡桃(Juglans regia) PLC可高达到100%, 但是由于冬季和春季茎压修复, 木质部冻融栓塞程度有一定的降低.另外, 近期研究表明, 能产生木质部正压的植物, 其抗冻融栓塞能力可能不受导管直径的影响(Niu et al., 2017).根压和茎压主要来源于糖类水解成的蔗糖被运输到栓塞导管中而产生渗透浓度梯度(Nardini et al., 2011; Cao et al., 2012), 从而使水分流向栓塞导管, 即渗透调节假说(金鹰等, 2016).两者的不同之处在于: 根压主要在无蒸腾流情况下产生作用(Swaef et al., 2013), 且修复栓塞作用较小(Salleo et al., 2004), 特别是对高大植物上层木质部栓塞的修复作用很小(Westhoff et al., 2008).而茎压产生的渗透梯度可能来自韧皮部的糖类(韧皮部卸载假说, Salleo et al., 2004; Hölttä et al., 2018), 因此在多数情况下均有茎压存在, 这也被称为“新的再冲水” (novel refilling)假说(Brodersen & McElrone, 2013; 金鹰等, 2016).Améglio等(2004)发现, 胡桃在深冬时茎中淀粉含量最低, 在发芽前茎中再次合成淀粉, 这不仅表明胡桃在冬季低温下进行淀粉和蔗糖的转换, 而且蔗糖累积是其冻融栓塞修复中的关键.张海燕等(2013)也发现, 12个温带树种老枝的淀粉含量变化具有相同趋势. ...
... 可高达到100%, 但是由于冬季和春季茎压修复, 木质部冻融栓塞程度有一定的降低.另外, 近期研究表明, 能产生木质部正压的植物, 其抗冻融栓塞能力可能不受导管直径的影响(Niu et al., 2017).根压和茎压主要来源于糖类水解成的蔗糖被运输到栓塞导管中而产生渗透浓度梯度(Nardini et al., 2011; Cao et al., 2012), 从而使水分流向栓塞导管, 即渗透调节假说(金鹰等, 2016).两者的不同之处在于: 根压主要在无蒸腾流情况下产生作用(Swaef et al., 2013), 且修复栓塞作用较小(Salleo et al., 2004), 特别是对高大植物上层木质部栓塞的修复作用很小(Westhoff et al., 2008).而茎压产生的渗透梯度可能来自韧皮部的糖类(韧皮部卸载假说, Salleo et al., 2004; Hölttä et al., 2018), 因此在多数情况下均有茎压存在, 这也被称为“新的再冲水” (novel refilling)假说(Brodersen & McElrone, 2013; 金鹰等, 2016).Améglio等(2004)发现, 胡桃在深冬时茎中淀粉含量最低, 在发芽前茎中再次合成淀粉, 这不仅表明胡桃在冬季低温下进行淀粉和蔗糖的转换, 而且蔗糖累积是其冻融栓塞修复中的关键.张海燕等(2013)也发现, 12个温带树种老枝的淀粉含量变化具有相同趋势. ...
... 评价植物抗冻融栓塞能力最直接的指标是冻融栓塞引起水力导度丢失的百分比, 即木质部最大水力导度与低温下水力导度之差和最大水力导度之比(Charrier et al., 2013; Niu et al., 2017), 与干旱PLC类似.冻融栓塞常常会引起植物水力导度丢失现象.Granda等(2014)整合75个研究中的210个记录发现, 植物PLC从夏季到冬季逐步增加.Charrier等(2013)也发现, 冬季植物PLC最高, 且PLC可能受海拔限制.然而, Fernández-Pérez等(2018)报道, 低温胁迫引起的6个松属树种的PLC较低, 而且没有发现PLC与低温损伤(叶黎明前水势)之间的关系. ...
Boreal and temperate trees show strong acclimation of respiration to warming 1 2016
... 北方森林和温带森林几乎占全球森林总面积的1/3 (Reich et al., 2016), 其植物面临的最主要环境胁迫之一是冬季低温.低温使植物地上器官木质部中的水分逐步冻结(Niu et al., 2017), 所以植物如何最大程度减少低温胁迫引起的损伤对其安全越冬(Hacke et al., 2015; Kreyling et al., 2015; Charrier et al., 2017)、生长、生存以及分布均具有重要作用(Schreiber et al., 2013; Charrier et al., 2015; Niu et al., 2017).低温时期冻融诱导形成的栓塞是低温胁迫对植物产生损伤的重要因素(Zanne et al., 2014; Zhang et al., 2018).许多研究发现, 植物抗冻融栓塞能力不仅影响其在低温时期的损伤程度和碳消耗(Bowling et al., 2018; Yin et al., 2018), 还显著影响随后生长季中的水分供应能力(冷冻疲劳, Feng et al., 2015; Zhang et al., 2018).但与干旱栓塞相比, 目前对木质部冻融栓塞的形成以及植物响应和调节机制的理解不足(Choat et al., 2011; Yin et al., 2018).为了推进该领域的研究, 本文先综述木质部冻融栓塞的形成机制和植物的逃避、忍耐、修复等3种冻融栓塞的应对策略, 然后总结植物低温抗性的生理表现、评价和影响因子, 在此基础上讨论低温抗性、干旱抗性和水力效率之间的多元权衡关系, 最后提出今后该领域中的优先研究问题. ...
Arabidopsis thaliana avoids freezing by supercooling 1 2006
... 植物还可以通过木质部过冷却的方式避免低温及冰冻损伤(Arias et al., 2017).过冷却指植物使细胞中液体的冰点温度远低于液态水冰点的能力(Arias et al., 2015).Rasmussen和MacKenzie (1972)指出, 没有任何结晶核的纯水结冰温度低至-38.1 ℃.因此, 植物细胞内异质结晶核含量影响其过冷却点的高低(Losso et al., 2018).过冷却能力对植物应对突发性的冰冻事件(如热带植物遭遇突发性寒流)十分重要(Reyes-Díaz et al., 2006).Arias等(2015)发现, 在寒冷环境中驯化过的油橄榄(Olea europaea)栽培种, 其叶片冰点温度低至-13 ℃ (远低于环境最低温度), 而且有更高的叶密度和细胞壁刚性, 从而使其溶质渗透性和组织可塑性更高, 质外体水分含量更低; 进一步研究发现, 从夏季到冬季该树种茎最低冰点逐渐降低, 而且其木质部抗空穴化能力与叶片和茎的过冷却能力之间存在权衡(Arias et al., 2017). ...
Tolerance and resilience of forest species to frost in restoration planting in southern Brazil 1 2018
... 另一种评价植物低温耐性的生理特征是低温对植物的损伤程度, 即将经过低温处理后的植物置于适宜环境中生长, 之后观察枯落叶(或者死根)的百分比, 以此来判断植物低温忍耐能力(Carles et al., 2011; Rorato et al., 2018).Toca等(2018)报道, 经历冬季(低温处理)后的松树幼苗, 其小枝低温忍耐能力明显增加, 且氮添加显著影响植物低温忍耐能力, 但正负效应均存在.土壤养分是否影响植物低温耐性还有待于深入研究. ...
New evidence for a role of vessel-associated cells and phloem in the rapid xylem refilling of cavitated stems of Laurus nobilis L 2 2004
... 修复是植物应对冻融栓塞的又一重要策略(Preston & Sandve, 2013; Wang et al., 2018).有研究表明, 采用正压(主要包括根压和茎压)恢复水力功能的植物可能具有更强的抗冷冻疲劳能力(即木质部冰冻后对水力功能丢失更敏感的现象)(Niu et al., 2017; Yin et al., 2018), 这是植物对冻融栓塞的一种主动修复.Hao等(2013)发现, 早春纸桦(Betula papyrifera)树干水分含量明显高于生长季, 表明根压在修复冻融栓塞导管的过程中具有重要作用.Améglio等(2002)报道, 冬季胡桃(Juglans regia) PLC可高达到100%, 但是由于冬季和春季茎压修复, 木质部冻融栓塞程度有一定的降低.另外, 近期研究表明, 能产生木质部正压的植物, 其抗冻融栓塞能力可能不受导管直径的影响(Niu et al., 2017).根压和茎压主要来源于糖类水解成的蔗糖被运输到栓塞导管中而产生渗透浓度梯度(Nardini et al., 2011; Cao et al., 2012), 从而使水分流向栓塞导管, 即渗透调节假说(金鹰等, 2016).两者的不同之处在于: 根压主要在无蒸腾流情况下产生作用(Swaef et al., 2013), 且修复栓塞作用较小(Salleo et al., 2004), 特别是对高大植物上层木质部栓塞的修复作用很小(Westhoff et al., 2008).而茎压产生的渗透梯度可能来自韧皮部的糖类(韧皮部卸载假说, Salleo et al., 2004; Hölttä et al., 2018), 因此在多数情况下均有茎压存在, 这也被称为“新的再冲水” (novel refilling)假说(Brodersen & McElrone, 2013; 金鹰等, 2016).Améglio等(2004)发现, 胡桃在深冬时茎中淀粉含量最低, 在发芽前茎中再次合成淀粉, 这不仅表明胡桃在冬季低温下进行淀粉和蔗糖的转换, 而且蔗糖累积是其冻融栓塞修复中的关键.张海燕等(2013)也发现, 12个温带树种老枝的淀粉含量变化具有相同趋势. ...
Coordination and trade-offs among hydraulic safety, efficiency and drought avoidance traits in Amazonian rainforest canopy tree species 1 2018
... 木质部的水力效率-安全权衡是植物适应环境和进化的结果(Blackman et al., 2010; Jin et al., 2016), 影响植物的竞争力和生产力(Nardini & Luglio, 2014).水力效率一般用植物水力导度表示, 而水力安全常用抗干旱栓塞能力(P50)表示(Santiago et al., 2018).Nardini等(2012)报道, 来自温带的6种植物(槭属(Acer)和栎属(Quercus)各3种)的水力导度、P50和比叶质量之间存在显著相关性, 支持水力效率-安全权衡, 且增加叶碳投入可以提高水力丢失的忍耐能力.Nardini和Luglio (2014)整合130个木本被子植物叶片水力数据后也发现存在叶水力效率-安全权衡关系, 而且生长在低降水量地区的植物的P50 (负值)和叶水力导度均较低.Gleason等(2016)综合分析335种被子植物和89种裸子植物的茎水力数据, 发现虽然水力导度和抗干旱栓塞能力之间存在弱相关性(支持水力效率-安全权衡), 但没有发现高效率和高安全性的植物.因此, 植物普遍存在不同程度的水力效率-安全权衡关系, 但其关系的紧密程度受导管特征(Lens et al., 2011; Hajek et al., 2014)、木质材性(金鹰和王传宽, 2016)、树高(Prendin et al., 2018)等因素的影响. ...
Sixteen years of winter stress: An assessment of cold hardiness, growth performance and survival of hybrid poplar clones at a boreal planting site 1 2013
... 北方森林和温带森林几乎占全球森林总面积的1/3 (Reich et al., 2016), 其植物面临的最主要环境胁迫之一是冬季低温.低温使植物地上器官木质部中的水分逐步冻结(Niu et al., 2017), 所以植物如何最大程度减少低温胁迫引起的损伤对其安全越冬(Hacke et al., 2015; Kreyling et al., 2015; Charrier et al., 2017)、生长、生存以及分布均具有重要作用(Schreiber et al., 2013; Charrier et al., 2015; Niu et al., 2017).低温时期冻融诱导形成的栓塞是低温胁迫对植物产生损伤的重要因素(Zanne et al., 2014; Zhang et al., 2018).许多研究发现, 植物抗冻融栓塞能力不仅影响其在低温时期的损伤程度和碳消耗(Bowling et al., 2018; Yin et al., 2018), 还显著影响随后生长季中的水分供应能力(冷冻疲劳, Feng et al., 2015; Zhang et al., 2018).但与干旱栓塞相比, 目前对木质部冻融栓塞的形成以及植物响应和调节机制的理解不足(Choat et al., 2011; Yin et al., 2018).为了推进该领域的研究, 本文先综述木质部冻融栓塞的形成机制和植物的逃避、忍耐、修复等3种冻融栓塞的应对策略, 然后总结植物低温抗性的生理表现、评价和影响因子, 在此基础上讨论低温抗性、干旱抗性和水力效率之间的多元权衡关系, 最后提出今后该领域中的优先研究问题. ...
Wintertime photosynthesis and water uptake in a boreal forest 1 2006
... 常绿树种茎抗低温胁迫能力可能还与叶片低温抗性有关(Ball et al., 2004; Fernández-Pérez et al., 2018; Verhoeven et al., 2018).“脆弱性分割假说”指出, 植物面临胁迫时, 其叶片(或小枝)可能比其茎表现得更为脆弱, 通过牺牲 “造价低廉”的末端器官保护“造价昂贵”的茎干, 以避免栓塞发生(Zimmermann, 1983; Tyree & Ewers, 1991).已存在大量有关干旱胁迫下的“脆弱性分割假说”验证研究(Hao et al., 2008; Johnson et al., 2012; Pivovaroff et al., 2014; Zhu et al., 2019), 但还不清楚低温胁迫下是否也会发生该现象.Fernández-Pérez等(2018)发现, 6个松属(Pinus)树种的针叶低温抗性对其分布范围的预测能力大于茎干木质部抗冻融栓塞能力.在北半球温带森林中, 冬季留存叶片因日间光照而使其日间温度高于0 ℃, 从而可能延长光合作用的时间(Feild & Brodribb, 2001; Sevanto et al., 2006; Lindfors et al., 2015; Bowling et al., 2018), 生产的有机物可用于早春修复冻融栓塞和细胞损伤.不过, 此时树干仍处于冰冻状态, 导致叶片可能比茎经历更多次的冻融循环, 从而加剧叶片的冻融栓塞(Feild & Brodribb, 2001; Charra-Vaskou et al., 2016), 降低茎干的冻融栓塞程度, 在一定程度上对主茎起到保护作用(Charrier et al., 2017). ...
Drought increases the freezing resistance of high-elevation plants of the Central Chilean Andes 1 2016
... 木质部低温抗性、干旱抗性和水力效率三者间是否存在多元权衡现象?目前对此问题的综合研究较少, 但已有研究指出, 干旱显著增加植物的低温抗性(Kreyling et al., 2012; Walter et al., 2013; Sierra-?Almeida et al., 2016; Sperling et al., 2017).这是因为植物通过驯化改变了细胞对脱水的忍耐能力(Buchner & Neuner, 2011), 其中包括基因表达的改变导致的细胞膜结构变化或者细胞化学成分变化(Verhoeven et al., 2018).例如, Medeiros和Pockman (2011)报道, 在-15 ℃环境中干旱处理使Larrea tridentate植株细胞死亡减少、叶留存量增加、水力导度和气体交换速率升高, 这表明干旱处理显著增加了其低温抗性.Hoffman等(2012)也发现, 干旱处理能够显著增加黑麦草(Lolium perenne)的低温抗性.然而, Kong和Henry (2016)研究发现, 早春低温处理的草地早熟禾(Poa pratensis)的抗旱性显著增加(生物量和存活率增加), 但夏季干旱处理并未增加其低温抗性, 这表明干旱处理对植物低温抗性无影响的现象也存在.同样, Bachofen等(2016)也发现, 干旱处理没有增加地中海松属3个树种的低温抗性, 而且推测该现象可能仅在短期极端条件下发生.Yin等(2018)同时对低温抗性、干旱抗性和水力效率进行研究发现, 抗干旱栓塞与水力效率存在权衡, 抗冷冻疲劳与水力效率间也存在权衡现象, 支持多元权衡现象.然而, 当植物同时面对干旱和低温胁迫时(如高山树线植物)(Mayr et al., 2007; Charrier et al., 2017), 多次冻融交替结合干旱胁迫可能引起PLC增加(Mayr et al., 2003; Charrier et al., 2014).Mayr等(2006)报道, 生长在林线处的针叶树种, 每个冬季经历至少100次冻融交替, 再加上干旱胁迫, 二者共同引起欧洲云杉(Picea abies) PLC高达100%.因此, 生长在低温和干旱胁迫共存环境中的植物, 其PLC可能更高, 间接表明三者间存在一定程度的权衡.综上所述, 我们构建了低温抗性、干旱抗性和水力效率之间多元权衡概念图(图2). ...
Acclimation of Pistacia integerrima trees to frost in semi-arid environments depends on autumn’s drought 1 2017
... 木质部低温抗性、干旱抗性和水力效率三者间是否存在多元权衡现象?目前对此问题的综合研究较少, 但已有研究指出, 干旱显著增加植物的低温抗性(Kreyling et al., 2012; Walter et al., 2013; Sierra-?Almeida et al., 2016; Sperling et al., 2017).这是因为植物通过驯化改变了细胞对脱水的忍耐能力(Buchner & Neuner, 2011), 其中包括基因表达的改变导致的细胞膜结构变化或者细胞化学成分变化(Verhoeven et al., 2018).例如, Medeiros和Pockman (2011)报道, 在-15 ℃环境中干旱处理使Larrea tridentate植株细胞死亡减少、叶留存量增加、水力导度和气体交换速率升高, 这表明干旱处理显著增加了其低温抗性.Hoffman等(2012)也发现, 干旱处理能够显著增加黑麦草(Lolium perenne)的低温抗性.然而, Kong和Henry (2016)研究发现, 早春低温处理的草地早熟禾(Poa pratensis)的抗旱性显著增加(生物量和存活率增加), 但夏季干旱处理并未增加其低温抗性, 这表明干旱处理对植物低温抗性无影响的现象也存在.同样, Bachofen等(2016)也发现, 干旱处理没有增加地中海松属3个树种的低温抗性, 而且推测该现象可能仅在短期极端条件下发生.Yin等(2018)同时对低温抗性、干旱抗性和水力效率进行研究发现, 抗干旱栓塞与水力效率存在权衡, 抗冷冻疲劳与水力效率间也存在权衡现象, 支持多元权衡现象.然而, 当植物同时面对干旱和低温胁迫时(如高山树线植物)(Mayr et al., 2007; Charrier et al., 2017), 多次冻融交替结合干旱胁迫可能引起PLC增加(Mayr et al., 2003; Charrier et al., 2014).Mayr等(2006)报道, 生长在林线处的针叶树种, 每个冬季经历至少100次冻融交替, 再加上干旱胁迫, 二者共同引起欧洲云杉(Picea abies) PLC高达100%.因此, 生长在低温和干旱胁迫共存环境中的植物, 其PLC可能更高, 间接表明三者间存在一定程度的权衡.综上所述, 我们构建了低温抗性、干旱抗性和水力效率之间多元权衡概念图(图2). ...
Size and function in conifer tracheids and angiosperm vessels 1 2006
... 木质部结构是影响植物忍耐低温胁迫的主要因素(Hacke et al., 2015, 2017; Lintunen et al., 2018).被子植物采用多个细胞死亡后相互连接构成的导管进行水分运输, 长度在1 mm到几米不等; 裸子植物则依靠单个细胞伸长形成的管胞进行水分运输, 长度在2-6 mm之间(Tyree & Ewers, 1991), 而且管胞直径一般小于导管(Sperry et al., 2006).这种木质部结构差异导致裸子植物和被子植物响应冻融栓塞时产生显著PLC差异(Feild & Brodribb, 2001; Feng et al., 2015; Fernández-Pérez et al., 2018).以往研究表明, 植物在入冬前PLC逐步增加(Zhang et al., 2018), 而导管或管胞的直径差异是植物PLC不同的主要原因(Pittermann & Sperry, 2003; Hacke et al., 2015, 2017).在所有季节中, 裸子植物的PLC均小于被子植物(Granda et al., 2014).例如, 被子植物葡萄牙栎(Quercus faginea)抗空穴化能力较差, 夏季PLC达到80%, 冬季进一步达到90%, 而裸子植物香桧(Juniperus thurifera)和欧洲黑松(Pinus nigra)夏季PLC几乎为0, 冬季仅增加到3%-9% (Granda et al., 2014). ...
What plant hydraulics can tell us about responses to climate-change droughts 1 2015
... 栓塞是降低木质部内水分运输效率的关键因子(Choat et al., 2012; 金鹰等, 2016), 主要包括干旱栓塞和冻融栓塞(Améglio et al., 2002; Willson & Jackson, 2006; Venturas et al., 2017).两种栓塞形成的主要机理均为空气进入导管或管胞, 从而阻断木质部水柱, 形成栓塞(Sperry & Sullivan, 1992), 而不同之处在于引发空气进入的过程(Hacke & Sperry, 2001).根据内聚力-张力理论, 木质部导管内水分持续运输动力主要来自叶片蒸腾失水引起的拉力, 因此导管中的水分始终处于亚稳态(Sperry & Love, 2015).当蒸腾加剧和/或土壤水分供应不足而使木质部导管内负压超过一定阈值时, 就会造成其水柱断裂, 致使周围组织中的气体进入导管, 从而形成干旱栓塞(Choat et al., 2018).而冻融栓塞主要发生在木质部经历多次冻融循环的过程中(Sperry & Sullivan, 1992; Feild & Brodribb, 2001; Charra-?Vaskou et al., 2016).当木质部水分冰冻时, 水中原有溶解气体因在冰中的溶解度很低而逸散到导管中(Lintunen et al., 2014).而当冰冻融化时, 导管中的留存气体或者重新溶进水中(气泡压力大于大气压时, Hacke & Sperry, 2001), 或者由于冰冻压力的释放(冰融化后体积减小)等使气体在导管中扩大(Lemoine et al., 1999; Mayr et al., 2002; Pittermann & Sperry, 2006), 多次冻融交替之后, 导管中气体不断增加和扩张, 最终引发冻融栓塞, 该现象也被称为“融化-扩张假说” (Mayr & Sperry, 2010; Mayr & Améglio, 2016).干旱栓塞和冻融栓塞均会阻碍植物水分从土壤到叶片的运输, 从而降低叶片的光合速率(Brodribb et al., 2007; Reich, 2014; Jin et al., 2016), 导致植物生产力降低甚至死亡(McDowell et al., 2008; Hoffmann et al., 2011; McDowell, 2011; Choat et al., 2018). ...
Xylem embolism in response to freeze-thaw cycles and water stress in ring-porous, diffuse-porous, and conifer species 2 1992
... 栓塞是降低木质部内水分运输效率的关键因子(Choat et al., 2012; 金鹰等, 2016), 主要包括干旱栓塞和冻融栓塞(Améglio et al., 2002; Willson & Jackson, 2006; Venturas et al., 2017).两种栓塞形成的主要机理均为空气进入导管或管胞, 从而阻断木质部水柱, 形成栓塞(Sperry & Sullivan, 1992), 而不同之处在于引发空气进入的过程(Hacke & Sperry, 2001).根据内聚力-张力理论, 木质部导管内水分持续运输动力主要来自叶片蒸腾失水引起的拉力, 因此导管中的水分始终处于亚稳态(Sperry & Love, 2015).当蒸腾加剧和/或土壤水分供应不足而使木质部导管内负压超过一定阈值时, 就会造成其水柱断裂, 致使周围组织中的气体进入导管, 从而形成干旱栓塞(Choat et al., 2018).而冻融栓塞主要发生在木质部经历多次冻融循环的过程中(Sperry & Sullivan, 1992; Feild & Brodribb, 2001; Charra-?Vaskou et al., 2016).当木质部水分冰冻时, 水中原有溶解气体因在冰中的溶解度很低而逸散到导管中(Lintunen et al., 2014).而当冰冻融化时, 导管中的留存气体或者重新溶进水中(气泡压力大于大气压时, Hacke & Sperry, 2001), 或者由于冰冻压力的释放(冰融化后体积减小)等使气体在导管中扩大(Lemoine et al., 1999; Mayr et al., 2002; Pittermann & Sperry, 2006), 多次冻融交替之后, 导管中气体不断增加和扩张, 最终引发冻融栓塞, 该现象也被称为“融化-扩张假说” (Mayr & Sperry, 2010; Mayr & Améglio, 2016).干旱栓塞和冻融栓塞均会阻碍植物水分从土壤到叶片的运输, 从而降低叶片的光合速率(Brodribb et al., 2007; Reich, 2014; Jin et al., 2016), 导致植物生产力降低甚至死亡(McDowell et al., 2008; Hoffmann et al., 2011; McDowell, 2011; Choat et al., 2018). ...
... ).而冻融栓塞主要发生在木质部经历多次冻融循环的过程中(Sperry & Sullivan, 1992; Feild & Brodribb, 2001; Charra-?Vaskou et al., 2016).当木质部水分冰冻时, 水中原有溶解气体因在冰中的溶解度很低而逸散到导管中(Lintunen et al., 2014).而当冰冻融化时, 导管中的留存气体或者重新溶进水中(气泡压力大于大气压时, Hacke & Sperry, 2001), 或者由于冰冻压力的释放(冰融化后体积减小)等使气体在导管中扩大(Lemoine et al., 1999; Mayr et al., 2002; Pittermann & Sperry, 2006), 多次冻融交替之后, 导管中气体不断增加和扩张, 最终引发冻融栓塞, 该现象也被称为“融化-扩张假说” (Mayr & Sperry, 2010; Mayr & Améglio, 2016).干旱栓塞和冻融栓塞均会阻碍植物水分从土壤到叶片的运输, 从而降低叶片的光合速率(Brodribb et al., 2007; Reich, 2014; Jin et al., 2016), 导致植物生产力降低甚至死亡(McDowell et al., 2008; Hoffmann et al., 2011; McDowell, 2011; Choat et al., 2018). ...
Cavitation fatigue and its reversal in sunflower ( Helianthus annuus L.) 1 2002
Ecological stress memory and cross stress tolerance in plants in the face of climate extremes 1 2013
... 木质部低温抗性、干旱抗性和水力效率三者间是否存在多元权衡现象?目前对此问题的综合研究较少, 但已有研究指出, 干旱显著增加植物的低温抗性(Kreyling et al., 2012; Walter et al., 2013; Sierra-?Almeida et al., 2016; Sperling et al., 2017).这是因为植物通过驯化改变了细胞对脱水的忍耐能力(Buchner & Neuner, 2011), 其中包括基因表达的改变导致的细胞膜结构变化或者细胞化学成分变化(Verhoeven et al., 2018).例如, Medeiros和Pockman (2011)报道, 在-15 ℃环境中干旱处理使Larrea tridentate植株细胞死亡减少、叶留存量增加、水力导度和气体交换速率升高, 这表明干旱处理显著增加了其低温抗性.Hoffman等(2012)也发现, 干旱处理能够显著增加黑麦草(Lolium perenne)的低温抗性.然而, Kong和Henry (2016)研究发现, 早春低温处理的草地早熟禾(Poa pratensis)的抗旱性显著增加(生物量和存活率增加), 但夏季干旱处理并未增加其低温抗性, 这表明干旱处理对植物低温抗性无影响的现象也存在.同样, Bachofen等(2016)也发现, 干旱处理没有增加地中海松属3个树种的低温抗性, 而且推测该现象可能仅在短期极端条件下发生.Yin等(2018)同时对低温抗性、干旱抗性和水力效率进行研究发现, 抗干旱栓塞与水力效率存在权衡, 抗冷冻疲劳与水力效率间也存在权衡现象, 支持多元权衡现象.然而, 当植物同时面对干旱和低温胁迫时(如高山树线植物)(Mayr et al., 2007; Charrier et al., 2017), 多次冻融交替结合干旱胁迫可能引起PLC增加(Mayr et al., 2003; Charrier et al., 2014).Mayr等(2006)报道, 生长在林线处的针叶树种, 每个冬季经历至少100次冻融交替, 再加上干旱胁迫, 二者共同引起欧洲云杉(Picea abies) PLC高达100%.因此, 生长在低温和干旱胁迫共存环境中的植物, 其PLC可能更高, 间接表明三者间存在一定程度的权衡.综上所述, 我们构建了低温抗性、干旱抗性和水力效率之间多元权衡概念图(图2). ...
The interaction between nonstructural carbohydrate reserves and xylem hydraulics in Korean pine trees across an altitudinal gradient 2 2018
... 在长期的进化过程中, 植物(尤其是中高纬度、高山和亚高山木本植物)面对冻融栓塞, 逐渐形成了不同的应对策略(Lintunen et al., 2018; Wang et al., 2018; Willick et al., 2018; Yin et al., 2018), 主要包括逃避、忍耐和修复等3种应对策略(图1). ...
... 修复是植物应对冻融栓塞的又一重要策略(Preston & Sandve, 2013; Wang et al., 2018).有研究表明, 采用正压(主要包括根压和茎压)恢复水力功能的植物可能具有更强的抗冷冻疲劳能力(即木质部冰冻后对水力功能丢失更敏感的现象)(Niu et al., 2017; Yin et al., 2018), 这是植物对冻融栓塞的一种主动修复.Hao等(2013)发现, 早春纸桦(Betula papyrifera)树干水分含量明显高于生长季, 表明根压在修复冻融栓塞导管的过程中具有重要作用.Améglio等(2002)报道, 冬季胡桃(Juglans regia) PLC可高达到100%, 但是由于冬季和春季茎压修复, 木质部冻融栓塞程度有一定的降低.另外, 近期研究表明, 能产生木质部正压的植物, 其抗冻融栓塞能力可能不受导管直径的影响(Niu et al., 2017).根压和茎压主要来源于糖类水解成的蔗糖被运输到栓塞导管中而产生渗透浓度梯度(Nardini et al., 2011; Cao et al., 2012), 从而使水分流向栓塞导管, 即渗透调节假说(金鹰等, 2016).两者的不同之处在于: 根压主要在无蒸腾流情况下产生作用(Swaef et al., 2013), 且修复栓塞作用较小(Salleo et al., 2004), 特别是对高大植物上层木质部栓塞的修复作用很小(Westhoff et al., 2008).而茎压产生的渗透梯度可能来自韧皮部的糖类(韧皮部卸载假说, Salleo et al., 2004; Hölttä et al., 2018), 因此在多数情况下均有茎压存在, 这也被称为“新的再冲水” (novel refilling)假说(Brodersen & McElrone, 2013; 金鹰等, 2016).Améglio等(2004)发现, 胡桃在深冬时茎中淀粉含量最低, 在发芽前茎中再次合成淀粉, 这不仅表明胡桃在冬季低温下进行淀粉和蔗糖的转换, 而且蔗糖累积是其冻融栓塞修复中的关键.张海燕等(2013)也发现, 12个温带树种老枝的淀粉含量变化具有相同趋势. ...
The mechanisms of refilling of xylem conduits and bleeding of tall birch during spring 1 2008
... 修复是植物应对冻融栓塞的又一重要策略(Preston & Sandve, 2013; Wang et al., 2018).有研究表明, 采用正压(主要包括根压和茎压)恢复水力功能的植物可能具有更强的抗冷冻疲劳能力(即木质部冰冻后对水力功能丢失更敏感的现象)(Niu et al., 2017; Yin et al., 2018), 这是植物对冻融栓塞的一种主动修复.Hao等(2013)发现, 早春纸桦(Betula papyrifera)树干水分含量明显高于生长季, 表明根压在修复冻融栓塞导管的过程中具有重要作用.Améglio等(2002)报道, 冬季胡桃(Juglans regia) PLC可高达到100%, 但是由于冬季和春季茎压修复, 木质部冻融栓塞程度有一定的降低.另外, 近期研究表明, 能产生木质部正压的植物, 其抗冻融栓塞能力可能不受导管直径的影响(Niu et al., 2017).根压和茎压主要来源于糖类水解成的蔗糖被运输到栓塞导管中而产生渗透浓度梯度(Nardini et al., 2011; Cao et al., 2012), 从而使水分流向栓塞导管, 即渗透调节假说(金鹰等, 2016).两者的不同之处在于: 根压主要在无蒸腾流情况下产生作用(Swaef et al., 2013), 且修复栓塞作用较小(Salleo et al., 2004), 特别是对高大植物上层木质部栓塞的修复作用很小(Westhoff et al., 2008).而茎压产生的渗透梯度可能来自韧皮部的糖类(韧皮部卸载假说, Salleo et al., 2004; Hölttä et al., 2018), 因此在多数情况下均有茎压存在, 这也被称为“新的再冲水” (novel refilling)假说(Brodersen & McElrone, 2013; 金鹰等, 2016).Améglio等(2004)发现, 胡桃在深冬时茎中淀粉含量最低, 在发芽前茎中再次合成淀粉, 这不仅表明胡桃在冬季低温下进行淀粉和蔗糖的转换, 而且蔗糖累积是其冻融栓塞修复中的关键.张海燕等(2013)也发现, 12个温带树种老枝的淀粉含量变化具有相同趋势. ...
Ice segregation in the crown of winter cereals: Evidence for extraorgan and extratissue freezing 2 2019
... 冻的树干(Charrier et al., 2017)无法及时补充水分丢失(Bowling et al., 2018), 从而使植物叶片和小枝的木质部含水量降低.另外, 含水量变化还可能会引起质外体组织冰冻时的体积发生变化(Lintunen et al., 2016).低温胁迫使木质组织首先脱水收缩, 再经历液态水冰冻时的体积膨大(+9%)(Améglio et al., 2001).在冰冻前, 植物主动降低质外体含水量, 增加溶质浓度, 此时脱水引起的体积减小能够降低过冷却点(Lintunen et al., 2013; Willick et al., 2019).因此, 冰冻前含水量越低(体积越小), 植物对低温胁迫的忍耐力越强(Lintunen et al., 2018).当冰冻发生时, 由于水分由液态转为固态的体积增加, 对周围细胞壁产生机械压力, 植物需构建更强健的细胞壁(Zhang et al., 2016; Ployet et al., 2018)和改变细胞壁上蛋白组分(Willick et al., 2018)来抵御低温胁迫, 因此, 质外体较高的水分含量可能对植物产生更严重的低温损伤.研究表明, 木质部通过寒冷驯化 后其直径少量收缩(Améglio et al., 2001; Charra- Vaskou et al., 2016; Lintunen et al., 2016).例如, 欧洲赤松(Pinus sylvestris)和欧洲山杨(Populus tremula)木质部直径经寒冷驯化后分别收缩了0.5%和1.0%, 小于渗透胁迫和干旱胁迫引起的收缩量(Lintunen et al., 2016).此外, 植物通过寒冷驯化, 可以降低质外体的含水量, 并将水分转移到冰储存位点, 同时不同冰区之间形成隔离带(Willick et al., 2019), 从而尽可能减少低温损伤. ...
Tissue-specific changes in apoplastic proteins and cell wall structure during cold acclimation of winter wheat crowns 3 2018
... 在长期的进化过程中, 植物(尤其是中高纬度、高山和亚高山木本植物)面对冻融栓塞, 逐渐形成了不同的应对策略(Lintunen et al., 2018; Wang et al., 2018; Willick et al., 2018; Yin et al., 2018), 主要包括逃避、忍耐和修复等3种应对策略(图1). ...
... 植物还有一种忍耐低温胁迫的策略是, 通过改变细胞膜内外的溶质浓度, 使细胞内溶质浓度增加(形成低水势), 从而降低细胞外溶质浓度(Mayr et al., 2007; Lintunen et al., 2016).当冰冻发生时, 细胞外水分首先冻结(主要发生在细胞壁表面、纤维、导管内或者细胞外的空间)(Willick et al., 2018), 冰形成的低水势使细胞内水分进一步流出, 细胞内溶质浓度增加, 从而避免细胞内冰冻导致的细胞死亡(McCully et al., 2004), 能够暂时保护细胞不受冰冻损伤(Mayr & Améglio, 2016). ...
... 冻的树干(Charrier et al., 2017)无法及时补充水分丢失(Bowling et al., 2018), 从而使植物叶片和小枝的木质部含水量降低.另外, 含水量变化还可能会引起质外体组织冰冻时的体积发生变化(Lintunen et al., 2016).低温胁迫使木质组织首先脱水收缩, 再经历液态水冰冻时的体积膨大(+9%)(Améglio et al., 2001).在冰冻前, 植物主动降低质外体含水量, 增加溶质浓度, 此时脱水引起的体积减小能够降低过冷却点(Lintunen et al., 2013; Willick et al., 2019).因此, 冰冻前含水量越低(体积越小), 植物对低温胁迫的忍耐力越强(Lintunen et al., 2018).当冰冻发生时, 由于水分由液态转为固态的体积增加, 对周围细胞壁产生机械压力, 植物需构建更强健的细胞壁(Zhang et al., 2016; Ployet et al., 2018)和改变细胞壁上蛋白组分(Willick et al., 2018)来抵御低温胁迫, 因此, 质外体较高的水分含量可能对植物产生更严重的低温损伤.研究表明, 木质部通过寒冷驯化 后其直径少量收缩(Améglio et al., 2001; Charra- Vaskou et al., 2016; Lintunen et al., 2016).例如, 欧洲赤松(Pinus sylvestris)和欧洲山杨(Populus tremula)木质部直径经寒冷驯化后分别收缩了0.5%和1.0%, 小于渗透胁迫和干旱胁迫引起的收缩量(Lintunen et al., 2016).此外, 植物通过寒冷驯化, 可以降低质外体的含水量, 并将水分转移到冰储存位点, 同时不同冰区之间形成隔离带(Willick et al., 2019), 从而尽可能减少低温损伤. ...
Xylem cavitation caused by drought and freezing stress in four co-occurring Juniperus species 1 2006
... 栓塞是降低木质部内水分运输效率的关键因子(Choat et al., 2012; 金鹰等, 2016), 主要包括干旱栓塞和冻融栓塞(Améglio et al., 2002; Willson & Jackson, 2006; Venturas et al., 2017).两种栓塞形成的主要机理均为空气进入导管或管胞, 从而阻断木质部水柱, 形成栓塞(Sperry & Sullivan, 1992), 而不同之处在于引发空气进入的过程(Hacke & Sperry, 2001).根据内聚力-张力理论, 木质部导管内水分持续运输动力主要来自叶片蒸腾失水引起的拉力, 因此导管中的水分始终处于亚稳态(Sperry & Love, 2015).当蒸腾加剧和/或土壤水分供应不足而使木质部导管内负压超过一定阈值时, 就会造成其水柱断裂, 致使周围组织中的气体进入导管, 从而形成干旱栓塞(Choat et al., 2018).而冻融栓塞主要发生在木质部经历多次冻融循环的过程中(Sperry & Sullivan, 1992; Feild & Brodribb, 2001; Charra-?Vaskou et al., 2016).当木质部水分冰冻时, 水中原有溶解气体因在冰中的溶解度很低而逸散到导管中(Lintunen et al., 2014).而当冰冻融化时, 导管中的留存气体或者重新溶进水中(气泡压力大于大气压时, Hacke & Sperry, 2001), 或者由于冰冻压力的释放(冰融化后体积减小)等使气体在导管中扩大(Lemoine et al., 1999; Mayr et al., 2002; Pittermann & Sperry, 2006), 多次冻融交替之后, 导管中气体不断增加和扩张, 最终引发冻融栓塞, 该现象也被称为“融化-扩张假说” (Mayr & Sperry, 2010; Mayr & Améglio, 2016).干旱栓塞和冻融栓塞均会阻碍植物水分从土壤到叶片的运输, 从而降低叶片的光合速率(Brodribb et al., 2007; Reich, 2014; Jin et al., 2016), 导致植物生产力降低甚至死亡(McDowell et al., 2008; Hoffmann et al., 2011; McDowell, 2011; Choat et al., 2018). ...
长白山阔叶树种木质部环孔和散孔结构特征的分化导致其水力学性状的显著差异 7 2018
... 北方森林和温带森林几乎占全球森林总面积的1/3 (Reich et al., 2016), 其植物面临的最主要环境胁迫之一是冬季低温.低温使植物地上器官木质部中的水分逐步冻结(Niu et al., 2017), 所以植物如何最大程度减少低温胁迫引起的损伤对其安全越冬(Hacke et al., 2015; Kreyling et al., 2015; Charrier et al., 2017)、生长、生存以及分布均具有重要作用(Schreiber et al., 2013; Charrier et al., 2015; Niu et al., 2017).低温时期冻融诱导形成的栓塞是低温胁迫对植物产生损伤的重要因素(Zanne et al., 2014; Zhang et al., 2018).许多研究发现, 植物抗冻融栓塞能力不仅影响其在低温时期的损伤程度和碳消耗(Bowling et al., 2018; Yin et al., 2018), 还显著影响随后生长季中的水分供应能力(冷冻疲劳, Feng et al., 2015; Zhang et al., 2018).但与干旱栓塞相比, 目前对木质部冻融栓塞的形成以及植物响应和调节机制的理解不足(Choat et al., 2011; Yin et al., 2018).为了推进该领域的研究, 本文先综述木质部冻融栓塞的形成机制和植物的逃避、忍耐、修复等3种冻融栓塞的应对策略, 然后总结植物低温抗性的生理表现、评价和影响因子, 在此基础上讨论低温抗性、干旱抗性和水力效率之间的多元权衡关系, 最后提出今后该领域中的优先研究问题. ...
... 在长期的进化过程中, 植物(尤其是中高纬度、高山和亚高山木本植物)面对冻融栓塞, 逐渐形成了不同的应对策略(Lintunen et al., 2018; Wang et al., 2018; Willick et al., 2018; Yin et al., 2018), 主要包括逃避、忍耐和修复等3种应对策略(图1). ...
... 北方森林和温带森林几乎占全球森林总面积的1/3 (Reich et al., 2016), 其植物面临的最主要环境胁迫之一是冬季低温.低温使植物地上器官木质部中的水分逐步冻结(Niu et al., 2017), 所以植物如何最大程度减少低温胁迫引起的损伤对其安全越冬(Hacke et al., 2015; Kreyling et al., 2015; Charrier et al., 2017)、生长、生存以及分布均具有重要作用(Schreiber et al., 2013; Charrier et al., 2015; Niu et al., 2017).低温时期冻融诱导形成的栓塞是低温胁迫对植物产生损伤的重要因素(Zanne et al., 2014; Zhang et al., 2018).许多研究发现, 植物抗冻融栓塞能力不仅影响其在低温时期的损伤程度和碳消耗(Bowling et al., 2018; Yin et al., 2018), 还显著影响随后生长季中的水分供应能力(冷冻疲劳, Feng et al., 2015; Zhang et al., 2018).但与干旱栓塞相比, 目前对木质部冻融栓塞的形成以及植物响应和调节机制的理解不足(Choat et al., 2011; Yin et al., 2018).为了推进该领域的研究, 本文先综述木质部冻融栓塞的形成机制和植物的逃避、忍耐、修复等3种冻融栓塞的应对策略, 然后总结植物低温抗性的生理表现、评价和影响因子, 在此基础上讨论低温抗性、干旱抗性和水力效率之间的多元权衡关系, 最后提出今后该领域中的优先研究问题. ...
... 在长期的进化过程中, 植物(尤其是中高纬度、高山和亚高山木本植物)面对冻融栓塞, 逐渐形成了不同的应对策略(Lintunen et al., 2018; Wang et al., 2018; Willick et al., 2018; Yin et al., 2018), 主要包括逃避、忍耐和修复等3种应对策略(图1). ...
Divergent hydraulic strategies to cope with freezing in co-occurring temperate tree species with special reference to root and stem pressure generation 1 2018
... 木质部低温抗性、干旱抗性和水力效率三者间是否存在多元权衡现象?目前对此问题的综合研究较少, 但已有研究指出, 干旱显著增加植物的低温抗性(Kreyling et al., 2012; Walter et al., 2013; Sierra-?Almeida et al., 2016; Sperling et al., 2017).这是因为植物通过驯化改变了细胞对脱水的忍耐能力(Buchner & Neuner, 2011), 其中包括基因表达的改变导致的细胞膜结构变化或者细胞化学成分变化(Verhoeven et al., 2018).例如, Medeiros和Pockman (2011)报道, 在-15 ℃环境中干旱处理使Larrea tridentate植株细胞死亡减少、叶留存量增加、水力导度和气体交换速率升高, 这表明干旱处理显著增加了其低温抗性.Hoffman等(2012)也发现, 干旱处理能够显著增加黑麦草(Lolium perenne)的低温抗性.然而, Kong和Henry (2016)研究发现, 早春低温处理的草地早熟禾(Poa pratensis)的抗旱性显著增加(生物量和存活率增加), 但夏季干旱处理并未增加其低温抗性, 这表明干旱处理对植物低温抗性无影响的现象也存在.同样, Bachofen等(2016)也发现, 干旱处理没有增加地中海松属3个树种的低温抗性, 而且推测该现象可能仅在短期极端条件下发生.Yin等(2018)同时对低温抗性、干旱抗性和水力效率进行研究发现, 抗干旱栓塞与水力效率存在权衡, 抗冷冻疲劳与水力效率间也存在权衡现象, 支持多元权衡现象.然而, 当植物同时面对干旱和低温胁迫时(如高山树线植物)(Mayr et al., 2007; Charrier et al., 2017), 多次冻融交替结合干旱胁迫可能引起PLC增加(Mayr et al., 2003; Charrier et al., 2014).Mayr等(2006)报道, 生长在林线处的针叶树种, 每个冬季经历至少100次冻融交替, 再加上干旱胁迫, 二者共同引起欧洲云杉(Picea abies) PLC高达100%.因此, 生长在低温和干旱胁迫共存环境中的植物, 其PLC可能更高, 间接表明三者间存在一定程度的权衡.综上所述, 我们构建了低温抗性、干旱抗性和水力效率之间多元权衡概念图(图2). ...
Three keys to the radiation of angiosperms into freezing environments 2 2014
... 北方森林和温带森林几乎占全球森林总面积的1/3 (Reich et al., 2016), 其植物面临的最主要环境胁迫之一是冬季低温.低温使植物地上器官木质部中的水分逐步冻结(Niu et al., 2017), 所以植物如何最大程度减少低温胁迫引起的损伤对其安全越冬(Hacke et al., 2015; Kreyling et al., 2015; Charrier et al., 2017)、生长、生存以及分布均具有重要作用(Schreiber et al., 2013; Charrier et al., 2015; Niu et al., 2017).低温时期冻融诱导形成的栓塞是低温胁迫对植物产生损伤的重要因素(Zanne et al., 2014; Zhang et al., 2018).许多研究发现, 植物抗冻融栓塞能力不仅影响其在低温时期的损伤程度和碳消耗(Bowling et al., 2018; Yin et al., 2018), 还显著影响随后生长季中的水分供应能力(冷冻疲劳, Feng et al., 2015; Zhang et al., 2018).但与干旱栓塞相比, 目前对木质部冻融栓塞的形成以及植物响应和调节机制的理解不足(Choat et al., 2011; Yin et al., 2018).为了推进该领域的研究, 本文先综述木质部冻融栓塞的形成机制和植物的逃避、忍耐、修复等3种冻融栓塞的应对策略, 然后总结植物低温抗性的生理表现、评价和影响因子, 在此基础上讨论低温抗性、干旱抗性和水力效率之间的多元权衡关系, 最后提出今后该领域中的优先研究问题. ...
... 修复是植物应对冻融栓塞的又一重要策略(Preston & Sandve, 2013; Wang et al., 2018).有研究表明, 采用正压(主要包括根压和茎压)恢复水力功能的植物可能具有更强的抗冷冻疲劳能力(即木质部冰冻后对水力功能丢失更敏感的现象)(Niu et al., 2017; Yin et al., 2018), 这是植物对冻融栓塞的一种主动修复.Hao等(2013)发现, 早春纸桦(Betula papyrifera)树干水分含量明显高于生长季, 表明根压在修复冻融栓塞导管的过程中具有重要作用.Améglio等(2002)报道, 冬季胡桃(Juglans regia) PLC可高达到100%, 但是由于冬季和春季茎压修复, 木质部冻融栓塞程度有一定的降低.另外, 近期研究表明, 能产生木质部正压的植物, 其抗冻融栓塞能力可能不受导管直径的影响(Niu et al., 2017).根压和茎压主要来源于糖类水解成的蔗糖被运输到栓塞导管中而产生渗透浓度梯度(Nardini et al., 2011; Cao et al., 2012), 从而使水分流向栓塞导管, 即渗透调节假说(金鹰等, 2016).两者的不同之处在于: 根压主要在无蒸腾流情况下产生作用(Swaef et al., 2013), 且修复栓塞作用较小(Salleo et al., 2004), 特别是对高大植物上层木质部栓塞的修复作用很小(Westhoff et al., 2008).而茎压产生的渗透梯度可能来自韧皮部的糖类(韧皮部卸载假说, Salleo et al., 2004; Hölttä et al., 2018), 因此在多数情况下均有茎压存在, 这也被称为“新的再冲水” (novel refilling)假说(Brodersen & McElrone, 2013; 金鹰等, 2016).Améglio等(2004)发现, 胡桃在深冬时茎中淀粉含量最低, 在发芽前茎中再次合成淀粉, 这不仅表明胡桃在冬季低温下进行淀粉和蔗糖的转换, 而且蔗糖累积是其冻融栓塞修复中的关键.张海燕等(2013)也发现, 12个温带树种老枝的淀粉含量变化具有相同趋势. ...
温带12个树种新老树枝非结构性碳水化合物浓度比较 1 2013
... 修复是植物应对冻融栓塞的又一重要策略(Preston & Sandve, 2013; Wang et al., 2018).有研究表明, 采用正压(主要包括根压和茎压)恢复水力功能的植物可能具有更强的抗冷冻疲劳能力(即木质部冰冻后对水力功能丢失更敏感的现象)(Niu et al., 2017; Yin et al., 2018), 这是植物对冻融栓塞的一种主动修复.Hao等(2013)发现, 早春纸桦(Betula papyrifera)树干水分含量明显高于生长季, 表明根压在修复冻融栓塞导管的过程中具有重要作用.Améglio等(2002)报道, 冬季胡桃(Juglans regia) PLC可高达到100%, 但是由于冬季和春季茎压修复, 木质部冻融栓塞程度有一定的降低.另外, 近期研究表明, 能产生木质部正压的植物, 其抗冻融栓塞能力可能不受导管直径的影响(Niu et al., 2017).根压和茎压主要来源于糖类水解成的蔗糖被运输到栓塞导管中而产生渗透浓度梯度(Nardini et al., 2011; Cao et al., 2012), 从而使水分流向栓塞导管, 即渗透调节假说(金鹰等, 2016).两者的不同之处在于: 根压主要在无蒸腾流情况下产生作用(Swaef et al., 2013), 且修复栓塞作用较小(Salleo et al., 2004), 特别是对高大植物上层木质部栓塞的修复作用很小(Westhoff et al., 2008).而茎压产生的渗透梯度可能来自韧皮部的糖类(韧皮部卸载假说, Salleo et al., 2004; Hölttä et al., 2018), 因此在多数情况下均有茎压存在, 这也被称为“新的再冲水” (novel refilling)假说(Brodersen & McElrone, 2013; 金鹰等, 2016).Améglio等(2004)发现, 胡桃在深冬时茎中淀粉含量最低, 在发芽前茎中再次合成淀粉, 这不仅表明胡桃在冬季低温下进行淀粉和蔗糖的转换, 而且蔗糖累积是其冻融栓塞修复中的关键.张海燕等(2013)也发现, 12个温带树种老枝的淀粉含量变化具有相同趋势. ...
Seasonality of cavitation and frost fatigue in Acer mono Maxim 5 2018
... 北方森林和温带森林几乎占全球森林总面积的1/3 (Reich et al., 2016), 其植物面临的最主要环境胁迫之一是冬季低温.低温使植物地上器官木质部中的水分逐步冻结(Niu et al., 2017), 所以植物如何最大程度减少低温胁迫引起的损伤对其安全越冬(Hacke et al., 2015; Kreyling et al., 2015; Charrier et al., 2017)、生长、生存以及分布均具有重要作用(Schreiber et al., 2013; Charrier et al., 2015; Niu et al., 2017).低温时期冻融诱导形成的栓塞是低温胁迫对植物产生损伤的重要因素(Zanne et al., 2014; Zhang et al., 2018).许多研究发现, 植物抗冻融栓塞能力不仅影响其在低温时期的损伤程度和碳消耗(Bowling et al., 2018; Yin et al., 2018), 还显著影响随后生长季中的水分供应能力(冷冻疲劳, Feng et al., 2015; Zhang et al., 2018).但与干旱栓塞相比, 目前对木质部冻融栓塞的形成以及植物响应和调节机制的理解不足(Choat et al., 2011; Yin et al., 2018).为了推进该领域的研究, 本文先综述木质部冻融栓塞的形成机制和植物的逃避、忍耐、修复等3种冻融栓塞的应对策略, 然后总结植物低温抗性的生理表现、评价和影响因子, 在此基础上讨论低温抗性、干旱抗性和水力效率之间的多元权衡关系, 最后提出今后该领域中的优先研究问题. ...
... ; Zhang et al., 2018).但与干旱栓塞相比, 目前对木质部冻融栓塞的形成以及植物响应和调节机制的理解不足(Choat et al., 2011; Yin et al., 2018).为了推进该领域的研究, 本文先综述木质部冻融栓塞的形成机制和植物的逃避、忍耐、修复等3种冻融栓塞的应对策略, 然后总结植物低温抗性的生理表现、评价和影响因子, 在此基础上讨论低温抗性、干旱抗性和水力效率之间的多元权衡关系, 最后提出今后该领域中的优先研究问题. ...
... 木质部结构是影响植物忍耐低温胁迫的主要因素(Hacke et al., 2015, 2017; Lintunen et al., 2018).被子植物采用多个细胞死亡后相互连接构成的导管进行水分运输, 长度在1 mm到几米不等; 裸子植物则依靠单个细胞伸长形成的管胞进行水分运输, 长度在2-6 mm之间(Tyree & Ewers, 1991), 而且管胞直径一般小于导管(Sperry et al., 2006).这种木质部结构差异导致裸子植物和被子植物响应冻融栓塞时产生显著PLC差异(Feild & Brodribb, 2001; Feng et al., 2015; Fernández-Pérez et al., 2018).以往研究表明, 植物在入冬前PLC逐步增加(Zhang et al., 2018), 而导管或管胞的直径差异是植物PLC不同的主要原因(Pittermann & Sperry, 2003; Hacke et al., 2015, 2017).在所有季节中, 裸子植物的PLC均小于被子植物(Granda et al., 2014).例如, 被子植物葡萄牙栎(Quercus faginea)抗空穴化能力较差, 夏季PLC达到80%, 冬季进一步达到90%, 而裸子植物香桧(Juniperus thurifera)和欧洲黑松(Pinus nigra)夏季PLC几乎为0, 冬季仅增加到3%-9% (Granda et al., 2014). ...
 ,, 王传宽
,, 王传宽 ,*森林生态系统可持续经营教育部重点实验室, 哈尔滨 150040
,*森林生态系统可持续经营教育部重点实验室, 哈尔滨 150040
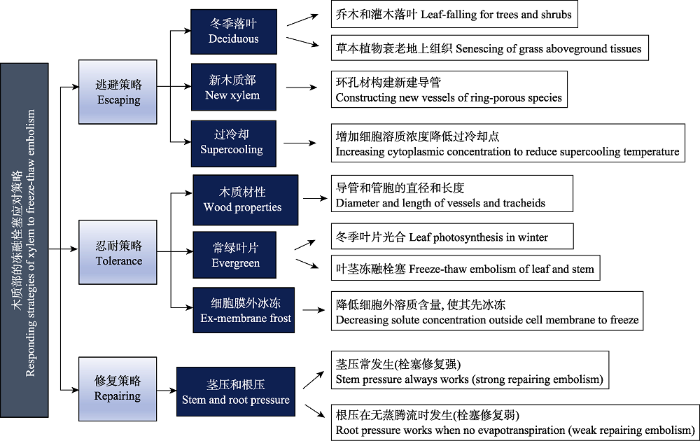 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT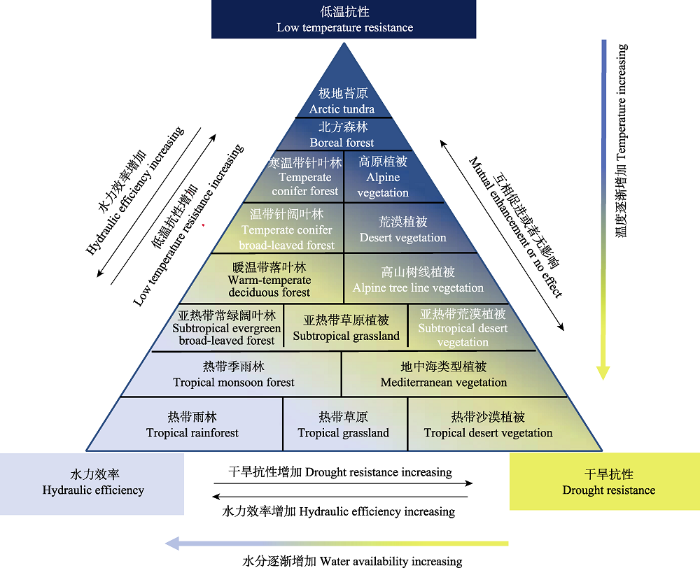 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}