 ,, 李亚楠, 卜海燕, 张世挺, 齐威
,, 李亚楠, 卜海燕, 张世挺, 齐威 ,*兰州大学生命科学学院草地农业生态系统国家重点实验室, 兰州 730000
,*兰州大学生命科学学院草地农业生态系统国家重点实验室, 兰州 730000Response of leaf traits of common broad-leaved woody plants to environmental factors on the eastern Qinghai-Xizang Plateau
Ji-Hong YANG,, Ya-Nan LI, Hai-Yan BU, Shi-Ting ZHANG, Wei QI,*State Key Laboratory of Grassland and Agro-Ecosystems, School of Life Sciences, Lanzhou University, Lanzhou 730000, China通讯作者: 通信作者 Corresponding author (qiw@lzu.edu.cn)
编委: 李敏
收稿日期:2019-07-8接受日期:2019-10-2网络出版日期:2019-10-20
| 基金资助: |
Received:2019-07-8Accepted:2019-10-2Online:2019-10-20
| Fund supported: |

摘要
叶片性状-环境关系对于预测气候变化对植物的影响至关重要。该研究以青藏高原东缘常见阔叶木本植物为研究对象, 从47个样点采集了332个物种共666个种群的叶片, 测量了15个叶片性状, 调查了该区域木本植物叶片性状的变异程度, 并从种内和种间水平探讨了叶片性状对环境的响应及适应策略。结果表明, 反眏叶片大小的性状均具有较高的变异, 其中, 叶片面积是变异程度最大的性状。除气孔密度外, 大多数叶片性状与海拔显著相关。气候是叶片性状变异的重要驱动因素, 3.3%-29.5%的叶片性状变异由气候因子组合解释。其中, 气温对叶片性状变异解释度最高, 日照时间能解释大部分叶片性状的变异, 而降水量对叶片性状变异的解释度相对较小。与环境(海拔和气候因子)显著相关的叶片性状在种内明显少于种间水平, 可能是植物性状之间的协同变化与权衡使种内性状变异比较小, 从而减弱了种内叶片性状与环境因子的相关性。研究结果总体表明,叶片性状与木本植物对环境的适应策略密切相关, 植物通过选择小而厚的叶片和较短的叶柄以适应高海拔的 环境。
关键词:
Abstract
Aims Leaf trait-environment relationships are critical for predicting the effects of climate change on plants. Our objective was to reveal the response of leaf traits of common broad-leaved woody plants to environmental factors on the eastern Qinghai-Xizang Plateau. Methods We measured 15 leaf traits of 332 species from 666 populations collected at 47 sites on the eastern Qinghai-Xizang Plateau. We investigated the extent of leaf trait variation in this area, and explored the response and adaptation strategies of leaf traits to environment at intra- and inter-species levels. Important findings Traits related to leaf size exhibited relatively high variation, and the leaf area was the most variant trait. Most leaf traits were significantly associated with elevation, except stomatal density. Climatic factors were important drivers of leaf trait variation because they explained 3.3%-29.5% of leaf trait variation. Meantime, temperature had the highest interpretation degree of leaf trait variation, and sunshine hours could explain the variation of most leaf traits. However, the interpretation degree of precipitation was relatively weak. In addition, the significant relationships between leaf traits and environmental (altitude and climatic) factors at intra-species level were far less than at inter-species levels. The reason for the result may be the coordinated variation and trade-off between plant traits, which make the variation of intra-species traits relatively small, and thus weaken the correlation between intra-plant leaf traits and environmental factors. Overall, leaf traits were closely related to woody plant adaptation strategies to the environment, and small, thick leaves and short petioles were selected for high-altitude plants to adapt to harsh environments such as strong winds and low temperature.
Keywords:
PDF (2904KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
杨继鸿, 李亚楠, 卜海燕, 张世挺, 齐威. 青藏高原东缘常见阔叶木本植物叶片性状对环境因子的响应. 植物生态学报, 2019, 43(10): 863-876. DOI: 10.17521/cjpe.2019.0174
YANG Ji-Hong, LI Ya-Nan, BU Hai-Yan, ZHANG Shi-Ting, QI Wei.
植物与其生存环境的关系是植物生态学研究的核心问题。植物通过调节外部形态和内部生理特性以响应和适应环境的变化, 同时改变植物对生态系统功能的贡献。基于性状的研究方法已被广泛用于植物适应策略的研究(Westoby et al., 2002; Rozendaal et al., 2006)。叶片性状与植物对资源的获取和利用密切相关, 可以反映植物适应环境变化所形成的生存策略, 并且具有易观察、易测定等优势(Royer et al., 2010; Scoffoni et al., 2011)。因此, 研究植物叶片沿环境梯度的变化可以更好地认识植物对环境的响应和适应机制(宋璐璐等, 2011; 王常顺和汪诗平, 2015)。
本研究以青藏高原东缘常见阔叶木本植物为研究对象, 调查该区域阔叶木本植物叶片性状的变异程度, 并从种内和种间水平探究叶片性状与环境(海拔和气候)的关系, 以期为深入解释植物对环境的响应方式和适应策略提供科学依据。我们提出了以下问题: (1)青藏高原东缘常见阔叶木本植物叶片性状的变异程度如何?(2)叶片性状随环境(海拔和气候)的变化模式, 以及它们在种内和种间水平上是否相似?(3)叶片性状随海拔梯度的变化模式会受到气候因子的影响吗?(4)在多个气候因子中, 影响叶片性状变异的主导因子是什么?
1 材料和方法
1.1 研究区概况
研究区位于青藏高原东北缘与西秦岭山脉的过渡地区(101.38°-104.33° E, 33.45°-35.08° N)的甘南藏族自治州和陇南市。该区域处于我国温带季风、亚热带季风和高原山地气候带的汇合区域以及半湿润和半干旱地区的分界线上。境内高差明显, 形成完整的亚热带-暖温带-中温带-亚高山-高山的垂直气候带, 相关气象或环境因子, 如年平均气温、无霜期、降水量、日照时间和土壤特征随海拔呈明显的梯度变化, 地带性木本植被也随海拔呈明显的地带性变化(表1), 是研究山地植物分布与特征变异的理想场所。1.2 采样方法
于2018年6-9月在研究区进行野外采样, 在每个海拔高度(间隔100 m)根据不同的植被类型设置1-3个样点, 共设置了47个采样点。使用GPS定位, 记录每个采样点的海拔和经纬度(图1)。在研究区选择了666个种群, 共332个物种, 隶属于47科102属(依据APG IV系统)。选择晴朗的天气, 于9:00-10:00进行采样。采样时, 在每个样点对每个物种随机选择生长健康、发育程度相近的3个不同个体, 每个个体选取位于冠层外围的3个当年生枝, 采集每个当年生枝上的完全成熟的, 无明显破损的所有叶片(10-30个叶片), 将采自3个不同个体的叶片混匀后放入便携式小冰箱带回室内。Table 1
表1
表1青藏高原东缘研究区各海拔段气候和植被特征
Table 1
| 海拔段 Altitude belt (m) | 气候带 Climatic zone | 年平均气温 Mean annual air temperature (℃) | 无霜期 Frost-free period (d) | 生长季 Growing season (d) | 地带性木本植被类型 Zonal woody vegetation type |
|---|---|---|---|---|---|
| 1 600-1 900 | 亚热带-暖温带 Subtropical-warm temperate | 11-15 | 200-240 | 230-270 | 温带-北亚热带阔叶林 Temperate-North subtropical broad-leaved forest |
| 1 900-2 200 | 暖温带 Warm temperate | 8-12 | 160-210 | 210-250 | 温带落叶阔叶林 Temperate deciduous broad-leaved forest |
| 2 200-2 500 | 暖温带-中温带 Warm temperate-medium temperature | 5-9 | 120-170 | 190-230 | 温带落叶阔叶林及针阔混交林 Temperate deciduous broad-leaved forest and coniferous and broad-leaved mixed forest |
| 2 500-2 800 | 中温带 Medium temperature | 3-7 | 80-130 | 170-210 | 温带针阔混交林 Temperate coniferous and broad-leaved mixed forest |
| 2 800-3 100 | 中温带-亚高山带 Medium temperate-subalpine | 1-5 | 40-90 | 150-190 | 温带及亚高山针阔混交林 Temperate and subalpine coniferous and broad-leaved mixed forest |
| 3 100-3 400 | 亚高山带 subalpine zone | -1-3 | 0-50 | 140-170 | 亚高山针阔混交林 Subalpine coniferous and broad-leaved mixed forest |
| 3 400-3 700 | 亚高山带-高寒带 Subalpine-alpine | -3-1 | 0-20 | 120-160 | 高寒及亚高山灌丛 Alpine and subalpine shrub |
| 3 700-4 000 | 高寒带 Alpine | -5- -1 | 0 | 100-140 | 高寒灌丛 Alpine shrub |
新窗口打开|下载CSV
1.3 气象数据的搜集
气象信息来源于中国气象数据网(http://data.?cma.cn/)。从该网站下载了距离各采样点最近的区县气象台2017-2018年的气象数据, 然后利用各样点实测经纬度信息, 通过ArcGIS 10.2 (https://www.arcgis.com/), 使用Kriging插值法获得每个样点的气象数据预测值。初步提取了10个气候变量, 包括4个温度变量: 年平均气温(MAT)、最热月最高气温(MTWA)、最冷月最低气温(MTCO)和生长季平均气温(MGST); 4个降水量变量: 年降水量(MAP)、最湿月降水量(PWE)、最干月降水量(PDR)、生长季平均降水量(MGSP); 2个日照时间变量: 年日照时间(MAS)和生长季日照时间(MGSS)。对不满足正态分布的MAT、MGST、MGSP和MGSS进行了对数转换, 然后分析了气候因子间的相关关系。对于高度相关(r > 0.80)的成对气候因子, 排除其中可以最大化被其他气候因子解释的因子以降低共线性。最终选取了7个因子(MTCO、MGST、MAP、PWE、PDR、MGSP、MGSS)作为分析用的气候因子。然后对选取的气候因子进行主成分分析(PCA)以减少气候数据的维数。我们提取了两个主成分(PC1和PC2), 共同解释了气候数据总方差的78.00%。PC1占方差的52.54%, 其中降水因子和日照时间因子(MAP、PWE、PDR和MGSS)具有较高的成分载荷; PC2解释了25.46%的方差, 温度因子(MTCO和MGST)具有最高的成分载荷。图1
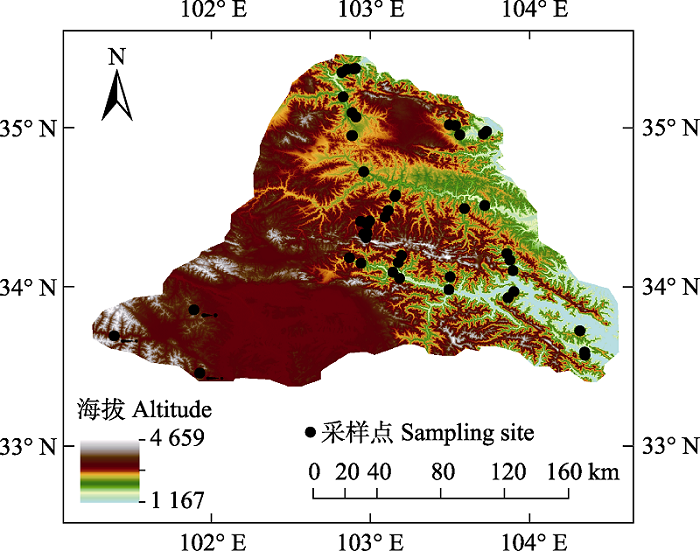 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1青藏高原东缘常见阔叶木本植物采样点分布图。
Fig. 1Distribution of sampling sites of common broad-leaved woody plants on the eastern Qinghai-Xizang Plateau.
1.4 叶片性状的测定
1.4.1 叶片形态性状和叶片含水量的测定取出10片较为完整、完全展开的叶片用于性状指标的测定。清除叶片表面粉尘后, 用电子天平(精度0.000 1 g)称其鲜质量。用螺旋测微仪(精度0.001 mm)测量叶片厚度, 测量时避开叶片主脉, 分别测量叶片前、中、末端, 最后取三者的平均值作为所测叶片的叶厚度值。然后将叶片平铺于扫描仪上进行扫描, 结合叶片扫描图片和Image J 1.52a软件(https://imagej.net/)测量叶片长度、叶片宽度、叶片面积和叶柄长度。将扫描后的叶片放入70 ℃烘箱, 48 h后取出, 用电子天平(精度0.000 1 g)称其干质量, 然后结合叶片干质量和叶片面积计算比叶面积, 结合叶片干质量和鲜质量计算叶片含水量。
比叶面积=叶片面积/叶片干质量
叶片含水量=(叶片鲜质量-叶片干质量)/叶片鲜质量
叶片长宽比=叶片长度/叶片宽度
1.4.2 气孔性状的测定
采用印迹法和透明胶带结合的改良方法测定气孔性状(段云峰等, 2008)。随机选择3个成熟叶片, 将透明指甲油涂抹于叶片下表皮中部(避开主脉和大的次级脉), 涂抹面积大约1 cm2, 静置, 待指甲油干后用透明胶带粘取下指甲油薄片, 置于载玻片上制成3张临时装片。在实验室内使用显微镜及其配置的摄像装置进行观察、测量并拍照。在40倍物镜、10倍目镜的条件下, 每个装片随机选择5个视野记录气孔数量, 根据视野内的气孔数量和视野面积计算气孔密度。同时, 在每个视野中选择3个气孔, 利用配套的分析软件测量气孔长度和气孔宽度。
气孔密度=视野内的气孔数量/视野面积
气孔长宽比=气孔长度/气孔宽度
气孔面积=气孔长度×气孔宽度× π × 1/4
气孔面积指数=气孔密度×气孔面积
1.5 数据处理
本研究分为种内分析和种间分析。在种内分析中, 选择区域广泛分布的物种(74个), 每个物种至少采集3个不同海拔段的种群, 以分析同一物种在不同种群间叶片性状的变异情况。在种间分析中, 选择区域内的常见物种(332个), 分析不同物种间的叶片性状变异。对多次采样的物种, 其叶性状及环境特征(海拔和气候因子)按多次采样的平均值用于种间分析。所有数据分析在R 3.5.2 (https://www.r-project.? org/)中进行。首先, 获得所有物种各性状平均值的描述性统计量, 以了解叶片性状的变异程度(对应科学问题1)。除叶片含水量(LWC)外, 对所有叶片性状进行了对数转换以保证数据符合正态分布。在种内分析中, 使用相关分析探究每个物种叶片性状与环境(海拔和两个气候主成分)的关系, 并且对每个性状分别记录正相关和负相关的物种数, 最后进行卡方检验以探究所有物种的总体趋势。在种间分析中, 首先使用回归分析探索叶片性状与环境(海拔和2个气候主成分)的关系。回归分析时, 选用线性方程和二次方程拟合模型, 并且根据模型解释度(R2)的大小选择最优模型(对应科学问题2)。由于PC1和PC2都与海拔显著相关(PC1: r = -0.548, p < 0.001; PC2: r = -0.575; p < 0.001), 所以使用偏相关分析(控制气候主成分)确定海拔和叶片性状之间的关系(对应科学问题3)。最后, 以7个气候因子作为自变量, 使用多元线性回归分析探究叶片性状与气候因子的关系。在初始模型拟合之后, 使用向后逐步法依据最小赤池信息准则(AIC)确定最优模型(对应科学问题4)。
2 结果和分析
2.1 叶片性状的变异特征
变异系数最大的5个性状分别是: 叶片面积、叶柄长度、单位叶柄长度的叶片面积、叶片宽度和叶片长度, 其中, 叶片面积是变异最大的叶片性状 (表2)。2.2 种内叶片性状与环境的关系
大部分物种的叶片长度(LL)、叶片宽度(LW)和叶片面积(LA)随海拔的升高而降低, 而气孔长度(SL)和气孔宽度(SW)随海拔升高而增加(表3), 说明在高海拔环境中大多数木本植物通常比其低海拔种群有更小的叶片和更大的气孔。对于两个气候主成分, 种内叶片性状与PC1没有一致的相关趋势, 而多数物种单位叶柄长度的叶片面积(LA/PL)随PC2的增加而增加(表3)。PC2中具有最高载荷值的气候因子是温度因子(MTCO和MGST), 并且载荷值都为正, 说明随着最低气温和生长季平均气温的升高, LA/PL增加, 即叶片面积支持效率增加。Table 2
表2
表2青藏高原东缘常见阔叶木本植物叶片性状的描述性统计结果
Table 2
| 性状 Traits | 物种数量 N | 平均值 Mean | 最小值 Min | 最大值 Max | 标准误差 SE | 变异系数 CV | 偏度 Skewness |
|---|---|---|---|---|---|---|---|
| LL (cm) | 329 | 7.54 | 0.80 | 40.68 | 0.26 | 0.63 | 2.24 |
| LW (cm) | 329 | 4.79 | 0.36 | 27.18 | 0.21 | 0.81 | 1.85 |
| LL/LW | 329 | 2.02 | 0.26 | 12.97 | 0.07 | 0.62 | 3.24 |
| LA (cm2) | 330 | 32.77 | 0.36 | 1 312.49 | 4.68 | 2.59 | 11.43 |
| LT (mm) | 314 | 0.18 | 0.05 | 0.44 | 0.003 | 0.28 | 1.25 |
| LWC | 280 | 0.60 | 0.33 | 0.85 | 0.01 | 0.14 | -0.28 |
| PL (cm) | 310 | 2.13 | 0.19 | 18.89 | 0.13 | 1.08 | 2.64 |
| SLA (cm2·g-1) | 320 | 173.49 | 46.47 | 392.15 | 3.29 | 0.34 | 0.93 |
| LA/PL (cm2·cm-1) | 310 | 15.05 | 1.61 | 109.17 | 0.75 | 0.87 | 3.17 |
| SD (No·mm-2) | 318 | 279.93 | 32.55 | 896.50 | 7.43 | 0.47 | 1.35 |
| SL (μm) | 320 | 26.05 | 10.77 | 64.99 | 0.41 | 0.28 | 0.77 |
| SW (μm) | 320 | 18.69 | 3.79 | 43.77 | 0.35 | 0.33 | 0.16 |
| SL/SW | 320 | 1.46 | 0.55 | 3.04 | 0.02 | 0.20 | 1.08 |
| SA (μm2) | 320 | 412.56 | 34.19 | 2 234.30 | 13.61 | 0.59 | 2.10 |
| SPI | 318 | 0.10 | 0.01 | 0.34 | 0.003 | 0.54 | 1.13 |
新窗口打开|下载CSV
Table 3
表3
表3青藏高原东缘常见阔叶木本植物种内叶片性状与环境因子相关性的卡方检验结果
Table 3
| 性状 Traits | 海拔 Altitude | PC1 | PC2 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N(+) | N(-) | χ2 | p | N(+) | N(-) | χ2 | p | N(+) | N(-) | χ2 | p | |
| LL | 20 | 50 | 12.86 | <0.01 | 40 | 30 | 1.43 | 0.23 | 36 | 34 | 0.06 | 0.81 |
| LW | 22 | 47 | 9.06 | <0.01 | 39 | 29 | 1.47 | 0.23 | 38 | 31 | 0.71 | 0.40 |
| LL/LW | 28 | 41 | 2.45 | 0.12 | 38 | 31 | 0.71 | 0.40 | 31 | 38 | 0.71 | 0.40 |
| LA | 19 | 51 | 14.63 | <0.01 | 39 | 31 | 0.91 | 0.34 | 41 | 29 | 2.06 | 0.15 |
| LT | 36 | 25 | 1.98 | 0.16 | 33 | 26 | 0.83 | 0.36 | 25 | 36 | 1.98 | 0.16 |
| PL | 22 | 37 | 3.81 | 0.05 | 34 | 27 | 0.80 | 0.37 | 27 | 32 | 0.42 | 0.52 |
| LWC | 21 | 25 | 0.35 | 0.56 | 26 | 20 | 0.78 | 0.38 | 20 | 26 | 0.78 | 0.38 |
| SLA | 36 | 30 | 0.55 | 0.46 | 35 | 31 | 0.24 | 0.62 | 30 | 36 | 0.55 | 0.46 |
| LA/PL | 23 | 38 | 3.69 | 0.06 | 30 | 31 | 0.02 | 0.90 | 39 | 22 | 4.74 | 0.03 |
| SD | 27 | 35 | 1.03 | 0.31 | 27 | 35 | 1.03 | 0.31 | 34 | 28 | 0.58 | 0.45 |
| SL | 40 | 24 | 4.00 | 0.04 | 33 | 31 | 0.06 | 0.80 | 30 | 34 | 0.25 | 0.61 |
| SW | 40 | 24 | 4.00 | 0.04 | 33 | 31 | 0.06 | 0.80 | 35 | 29 | 0.56 | 0.45 |
| SL/SW | 33 | 31 | 0.06 | 0.80 | 34 | 30 | 0.25 | 0.62 | 36 | 28 | 1.00 | 0.32 |
| SA | 36 | 28 | 1.00 | 0.32 | 32 | 32 | 0.00 | 1.00 | 31 | 33 | 0.06 | 0.80 |
| SPI | 31 | 33 | 0.06 | 0.80 | 30 | 34 | 0.25 | 0.62 | 38 | 26 | 2.25 | 0.13 |
新窗口打开|下载CSV
2.3 种间叶片性状与环境的关系
2.3.1 叶片性状与海拔的关系在种间水平, 海拔和10个叶片性状(LL、LW、LL/LW、LA、PL、LA/PL、SL、SW、SA和SPI存在显著线性关系, 其中, 只有LL/LW随海拔升高而增加, 其余9个叶片性状都随海拔升高而降低(图2)。海拔与叶片厚度(LT)、比叶面积(SLA)、气孔长宽比(SL/SW)存在明显的曲线关系(图2), 其中, SLA和SL/SW随海拔升高先增加后降低, 而LT随海拔升高先降低后增加。这些结果表明, 高海拔地区的木本物种往往比低海拔物种具有更短窄小的叶片, 更短的叶柄、更小的叶面积支持效率(LA/PL)和更短、更窄的气孔。
图2
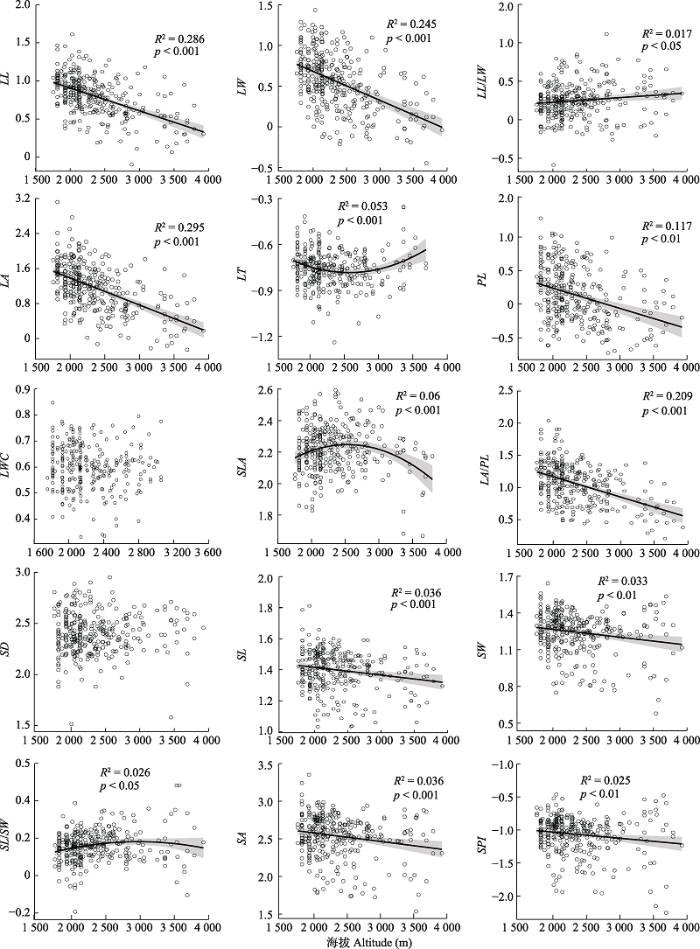 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2青藏高原东缘常见阔叶木本植物叶片性状的海拔变异模式。性状同
Fig. 2Leaf trait patterns of common broad-leaved woody plants along altitude on the eastern Qinghai-Xizang Plateau. See
然而, 当消除气候因子的影响后, 部分叶片性状和海拔之间的显著关系消失了(图2; 表4)。当控制PC2的影响时, LL/LW、SL、SW、SA和SPI与海拔的显著关系消失了, 但在控制PC1的影响时这些显著关系仍然存在(表4)。因此, 叶片长宽比和气孔性状(气孔长度、气孔宽度、气孔面积和气孔面积指 数)的海拔模式受到PC2代表的气候变量(气温)的影响。
2.3.2 叶片性状与气候的关系
回归分析显示, 12种叶片性状与PC1、PC2显著相关, 并且这些性状与PC1、PC2的最优拟合模型均为直线模型。其中, PC1与8种叶片性状(LL、LW、LA、PL、LA/PL、SL、SW和SA)呈正相关关系(图3)。PC2与11种叶片性状(LL、LW、LA、LT、PL、LWC、LA/PL、SL、SW、SA和SPI)呈正相关关系, 但与LL/LW呈负相关关系(图4)。
多元回归分析结果(表5)表明, 气温、降水量和光照时间对叶片性状都有明显的影响。例如, 叶片长度随着生长季平均气温的增加而降低, 随最冷月最低温度、最干月降水量和生长季平均日照时间的增加而增加(R2 = 0.29, p < 0.001)。在单个气候因子中, 有4个变量能解释较多的叶片性状变异(表5): 最冷月最低气温、最干月降水量、生长季平均日照时间能解释10个叶片性状和生长季平均气温能解释8个叶片性状。
3 讨论
3.1 叶片性状变异
植物的性状值常因其生境的不同而不同(Violle et al., 2007; Albert et al., 2011)。因此, 植物性状变异程度反映了植物对环境条件的适应范围(Reich et al., 1997)。在本研究中, 各叶片性状变异程度不同, 这与以前的研究结果一致(Cornelissen et al., 2011; Barros et al., 2012)。我们发现变异最大的5个叶片性状是叶片面积、叶柄长度、单位叶柄长度的叶片面积、叶片宽度和叶片长度, 而这些性状都是表征叶片大小的性状, 即叶片大小变异程度较高。这与前人的研究结果一致, 比如Díaz等(2016)发现叶片大小在全球范围内可达到6个数量级的差异。此外, 已有研究表明, 具有高变异性的性状通常与植物资源获取能力有关(Rozendaal et al., 2006; Violle et al., 2009)。叶片大小直接影响着植物对光的截取和对碳的获取能力, 并且叶片大小可以通过影响叶片边界层厚度进而影响叶片温度调节和蒸腾速率, 因此影响叶片热量和水平衡(Wright et al., 2017)。Table 4
表4
表4控制PC1和PC2影响后的海拔与叶片性状的偏相关系数
Table 4
| 性状 Traits | 海拔(控制PC1) Altitude (control PC1) | 海拔(控制PC2) Altitude (control PC2) | 海拔(控制PC1和PC2) Altitude (control PC1and PC2) |
|---|---|---|---|
| LL | -0.472** | -0.382** | -3.00** |
| LW | -0.467** | -0.323** | -0.289** |
| LL/LW | 0.168** | 0.043 | 0.086 |
| LA | -0.491** | -0.366** | -0.297** |
| LT | 0.014 | 0.069 | 0.118* |
| PL | -0.305** | -0.205** | -0.161** |
| LWC | -0.074 | 0.068 | 0.082 |
| SLA | -0.071 | -0.115 | -0.127* |
| LA/PL | -0.404** | -0.334** | -0.269** |
| SD | 0.015 | 0.021 | -0.010 |
| SL | -0.151** | -0.095 | -0.049 |
| SW | -0.146** | -0.073 | -0.029 |
| SL/SW | 0.055 | 0.000 | -0.012 |
| SA | -0.153** | -0.086 | -0.042 |
| SPI | -0.140* | -0.064 | -0.041 |
新窗口打开|下载CSV
图3
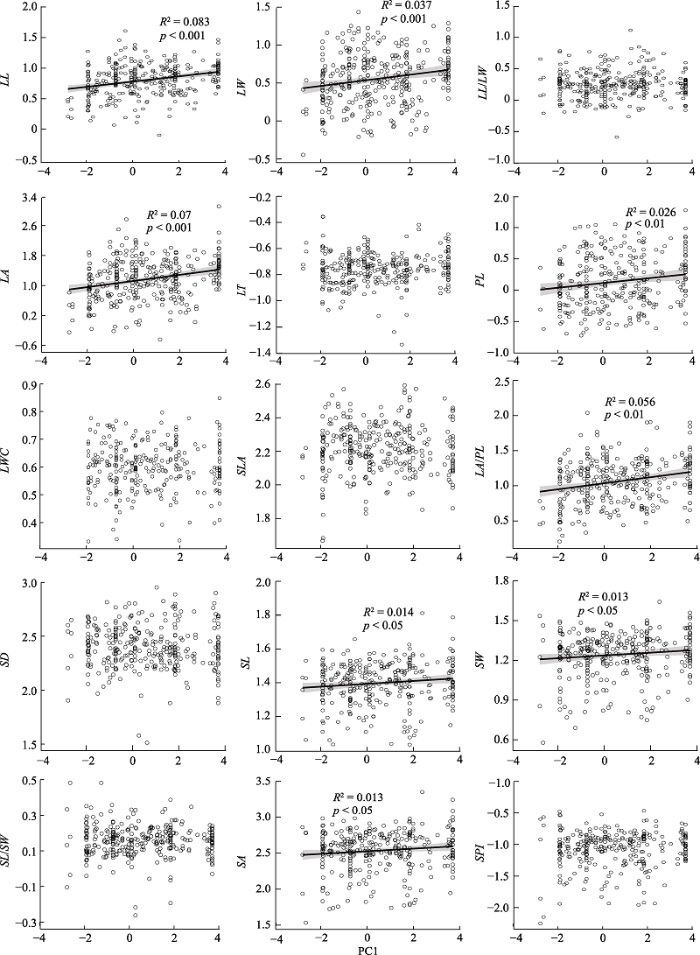 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3气候因子(第一主成分PC1)对青藏高原东缘常见阔叶木本植物叶片性状的影响。性状同
Fig. 3Effect of climatic factors (first principal component PC1) on leaf traits of common broad-leaved woody plants on the eastern Qinghai-Xizang Plateau. See
图4
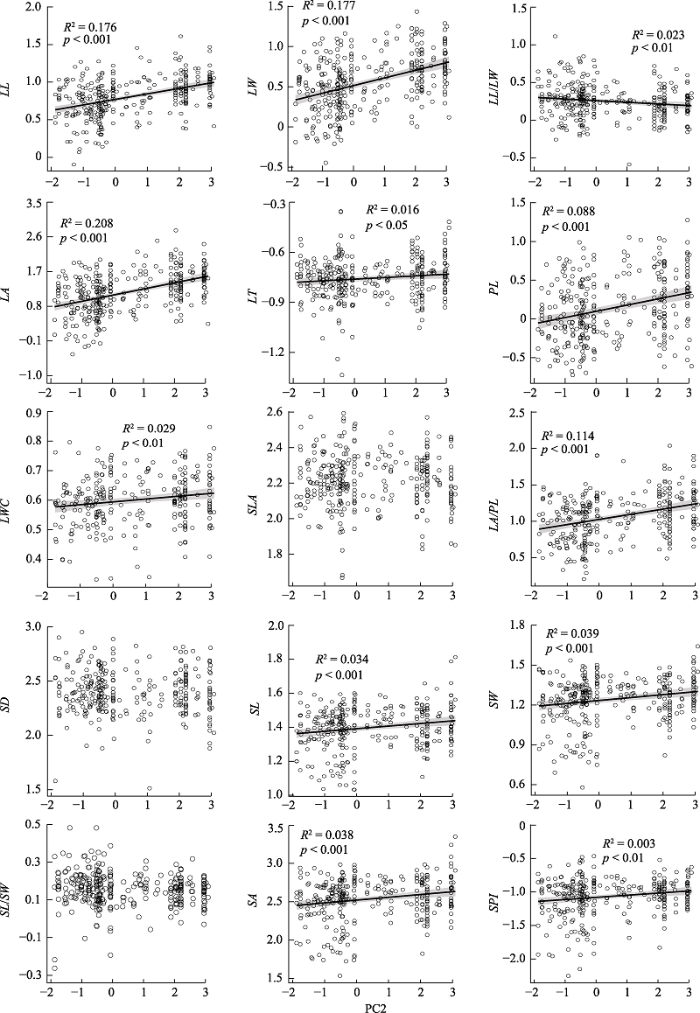 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4气候因子(第二主成分PC2)对青藏高原东缘常见阔叶木本植物叶片性状的影响。性状同
Fig. 4Effect of climatic factors (second principal component PC2) on leaf traits of common broad-leaved woody plants on the eastern Qinghai-Xizang Plateau. See
Table 5
表5
表5青藏高原东缘常见阔叶木本植物叶片性状与气候因子的多元线性回归模型
Table 5
| 线性模型 Linear regression model | AIC | R2 | p |
|---|---|---|---|
| LL = -0.16 + 0.13MTCO + 0.03MGSS - 0.14MGST + 0.20PDR | -983.45 | 0.29 | <0.001 |
| LW = -0.65 + 0.13MTCO + 0.30PDR + 0.03MGSS - 0.12MGST | -788.19 | 0.23 | <0.001 |
| LL/LW = 0.96 + 0.006PWE - 0.22PDR - 0.0006MAP + 0.03MGSS | -1 008.31 | 0.04 | <0.01 |
| LA = -0.91 + 0.26MTCO + 0.52MGSS + 0.47PDR - 0.26MGST | -507.75 | 0.30 | <0.001 |
| LT = 0.49 - 0.02PWE - 0.22MGST + 0.11MTCO + 0.03MGSS + 0.39PDR - 0.01 MGSP + 0.002MAP | -1 333.81 | 0.08 | <0.001 |
| PL = -0.94 + 0.05MTCO + 0.01MGSS + 0.16PDR | -587.23 | 0.11 | <0.001 |
| LWC = 0.37 + 0.01MGST + 0.0002MAP | -1 382.27 | 0.04 | <0.01 |
| SLA = 0.05 + 0.02MGSP - 0.17MTCO + 0.34MGST - 0.05MGSS + 0.02PWE - 0.56PDR -0.002MAP | -1 237.94 | 0.08 | <0.001 |
| LA/PL = -0.16 + 0.133MTCO + 0.03MGSS - 0.14MGST + 0.22PDR | -766.38 | 0.19 | <0.001 |
| SL = 1.10 + 0.01MTCO + 0.002MGSS + 0.05PDR | -1 355.05 | 0.06 | <0.001 |
| SW = 1.42 + 0.01MTCO | -1 154.49 | 0.05 | <0.001 |
| SL/SW = 0.005 + 0.002MGSS + 0.01PDR - 0.002PWE - 0.005MGST | -1 553.84 | 0.03 | <0.05 |
| SA = 2.83 + 0.02MTCO | -819.19 | 0.05 | <0.001 |
| SPI = 3.19 + 0.02MTCO | -813.97 | 0.04 | <0.001 |
新窗口打开|下载CSV
3.2 海拔与叶片性状
在本研究中, 叶片长度、叶片宽度和叶面积随海拔的升高而降低, 这在种内和种间分析中是一致的, 说明减小叶片大小既是木本植物对高海拔环境的可塑性反应, 也是其适应性结果, 与H?lscher等(2002)和McDonald等(2003)的结果相同。这可能是因为在高海拔的低温环境下, 小叶片的呼吸和蒸腾成本更低, 可以降低植物的维持消耗。其次, 高海拔环境中风速较大, 小叶片与空气的接触面积更小, 使其不容易受到风力的撕扯, 从而减少机械损伤(Givnish et al., 1984)。此外, 随着叶片面积增加, 叶片的边界层阻力增加, 导致叶片与土壤、空气和周围植被的热交换减少。这意味着在其他条件相同的情况下, 大叶片与环境的温差比小叶片更大, 从而使得大叶片在寒冷地区更容易遭受霜冻的破坏(Wright et al., 2017)。种间分析结果表明, 叶柄随海拔升高而变短, 可能是因为在低海拔环境中森林群落内植物间光竞争激烈, 长叶柄有利于增加叶片对光的拦截面积和效率, 而高海拔环境中光资源充足, 植物间光竞争较弱, 则不需发育较长的叶柄(Niiemets et al., 2004); 再加上短叶柄会使叶片受风力影响时的力矩较短, 从而叶片受到风力的拉扯作用就较小。叶片厚度和比叶面积的种间海拔变异式样是非直线的。两者在低海拔地区变化不明显, 在高海拔地区呈明显的海拔变异趋势, 这可能是因为低海拔地区的环境条件比较温和, 植物受到的环境胁迫较小, 其叶片特征的环境变异也相对和缓。叶片厚度在海拔2 700 m后随海拔升高而增加, 较厚的叶片可以通过增加单位叶片面积的氮含量和光合组织的体积增加强辐照下的光合能力, 而且较厚的叶片具有保温和耐辐射的作用, 以适应高海拔的寒冷气候和近日照的环境条件(Niinemets, 2001)。比叶面积能够反映植物在不同环境下获取资源的能力。通常认为, 生长在养分丰富的环境中的植物具有较大的比叶面积; 而生长在养分贫瘠的环境中的植物具有较小的比叶面积(Westoby et al., 2002)。本研究中, 比叶面积在海拔2 600 m之后随海拔升高而降低。随海拔升高, 土壤养分缺乏, 这样的环境更适合比叶面积较低的植物生存, 它们可以将更多的干物质投入用于抵御不良环境。此外, 随海拔升高, 土壤温度降低, 限制了植物根系对土壤水分的吸收, 从而植物受到的水分胁迫(即生理干旱)逐渐增大(Hultine & Marshall, 2000)。而较小的比叶面积意味着叶片会形成厚度较大而面积较小的叶片, 使得叶片内部水分向叶片表面扩散的距离或阻力增大, 降低植物内部水分散失, 从而缓解植物受到的水分胁迫(胡耀升等, 2015)。
对于气孔长度、气孔宽度与海拔的关系, 种内和种间分析出现了相反的结果。在种间分析中, 气孔长度、气孔宽度与海拔呈负相关关系, 这可能是因为在高海拔地区生境恶劣, 如低温、大风和高紫外线辐射等, 小的气孔有利于植物快速打开和关闭以快速应对多变的环境条件(Liu et al., 2019), 并且在叶片总气孔面积不变的情况下, 较小的气孔使得叶片蒸腾时的阻力增加, 从而可以减少水分的散失, 减缓叶片表面温度的下降。然而, 在种内分析中, 大多数物种的气孔长度、气孔宽度与海拔呈正相关关系。这可能是因为用于种内气孔性状测量的物种共64个, 其中有50个是灌木, 乔木较少。在低海拔环境中, 灌木多处于林下层, 其叶片所处的微环境中光照较弱, 随着海拔升高, 灌木逐渐成为群落中的优势种, 其叶片所处的环境中光资源充足, 因此, 光照可能会影响气孔长度和气孔宽度。这与徐浩杰等(2012)的研究结果一致, 他们发现分布在半阳坡的烈香杜鹃(Rhododendron anthopogonoides)的叶片气孔长度相比半阴坡的烈香杜鹃有增大趋势。
3.3 气候与叶片性状
种内分析中, 叶片性状与气候的关系没有统一趋势(表3)。在种间分析中, 没有任何单个气候因子可以解释所有叶片性状变异(表5), 气候和叶片性状变异之间的关联比较复杂。我们的研究结果表明, PC1与8个叶片性状显著相关(图3), PC2与11个叶片性状显著相关(图4), 并且3.3%-29.5%的叶片性状变异是由气候因子组合解释的(表5), 因此, 气候是叶片性状变异的重要驱动因素, 这与先前的从区域到全球尺度的研究结果(Niinemets, 2001; Ordo?ez et al., 2009; Xiao et al., 2015)一致。种间结果表明, 除了气孔密度, 所有叶片性状变异都可以由气候因子解释(图3, 图4; 表5)。其中, 对于同一性状, PC2对其的解释度总是大于PC1 (图3, 图4), 说明PC2代表的气候变量(气温)对叶片性状变异的影响大于PC1所代表的气候因子(光照时间和降水量)。此外, 我们发现在多元回归模型中气温和光照时间对叶片性状变异具有较高的解释度(表5), 所以在本研究中, 气温是叶片性状变异的主导因子, 光照时间可以解释大部分的叶片性状变异, 而降水量对叶片性状变异的解释度相对较小。由于本研究区集中在高海拔环境, 降水相对充足。因此, 降水量可能不是叶片性状变异的限制因子。此外, 相对于其他气候因子, 气温对气孔性状的解释度最高(表5), 同时, 偏相关分析结果也表明气孔性状(气孔长度、气孔宽度、气孔面积和气孔面积指数)的海拔变异模式受到气温的影响, 所以对于气孔性状的变异, 气温可能是其主导因子。
种内与环境(海拔和气候因子)显著相关的叶片性状明显少于种间水平(表3; 图2, 图3, 图4)。Siefert等(2015)对全球数据进行meta分析后发现, 种内变异的相对程度取决于性状分类的规模(例如植株水平与器官水平性状, 生理生化与形态学性状), 而不是生活型和环境。另外, 国内外****在植物功能性状沿环境梯度的变化规律以及不同性状之间的相互关系方面开展了大量的研究工作。许多研究表明, 在环境梯度下, 植物性状之间具有明显的协同变化规律和权衡关系(Funk & Cornwell, 2013; 李西良等, 2015), 性状之间的协同变化与权衡可能使种内变异比较小, 从而造成种内叶片性状变化与环境因子间的相关性较差。
4 结论
本文探讨了青藏高原东缘常见阔叶木本植物对海拔和气候因子的响应及适应策略, 为植物与环境关系的研究提供了参考依据。在研究区域, 叶片性状变异程度不同, 叶片大小具有较高的变异。大多数叶片性状受到海拔梯度和气候变化的影响。随着海拔的升高, 为适应随之而变的气候环境, 植物叶片变得小而厚, 叶柄变短, 以降低植物的维持消耗以及外力作用所导致的损伤。气候是叶片性状变异的重要驱动因素, 气温是叶片性状变异的主导因子, 光照时间也能解释大部分的叶片性状变异, 而降水量对叶片性状变异的解释度相对较小。另外, 对于气孔性状变异, 气温是其主导因子。这项工作提供了区域尺度上叶片性状的变异情况, 并显示了叶片性状-气候关系。同时, 已有证据表明土壤条件和植物的系统发育背景都对叶片性状有影响, 因此需要进一步研究以区分遗传-土壤-气候三个维度的关系, 探究由环境变化引起的叶片性状变异的来源。致谢 感谢兰州大学高寒草甸与湿地生态系统定位研究站提供野外工作条件, 以及草地农业生态系统国家重点实验室提供实验设备条件。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1111/j.1461-0248.2006.01006.xURLPMID:17257101

Plant functional traits vary both along environmental gradients and among species occupying similar conditions, creating a challenge for the synthesis of functional and community ecology. We present a trait-based approach that provides an additive decomposition of species' trait values into alpha and beta components: beta values refer to a species' position along a gradient defined by community-level mean trait values; alpha values are the difference between a species' trait values and the mean of co-occurring taxa. In woody plant communities of coastal California, beta trait values for specific leaf area, leaf size, wood density and maximum height all covary strongly, reflecting species distributions across a gradient of soil moisture availability. Alpha values, on the other hand, are generally not significantly correlated, suggesting several independent axes of differentiation within communities. This trait-based framework provides a novel approach to integrate functional ecology and gradient analysis with community ecology and coexistence theory.
DOI:10.1111/j.1654-1103.2005.tb02378.xURL
DOI:10.1111/j.1600-0706.2011.19672.xURL [本文引用: 1]
Functional diversity (FD) is a key facet of biodiversity used to address central questions in ecology. Despite recent methodological advances, FD remains a complex concept and no consensus has been reached either on how to quantify it, or on how it influences ecological processes. Here we define FD as the distribution of trait values within a community. When and how to account for intraspecific trait variability (ITV) when measuring FD remains one of the main current debates. It remains however unclear to what extent accounting for population-level ITV would modify FD quantification and associated conclusions. In this paper, we address two critical questions: (1) How sensitive are different components of FD to the inclusion of population-level ITV? (2) Does the omission of ITV obscure the understanding of ecological patterns? Using a mixture of empirical data and simulation experiments, we conducted a sensitivity analysis of four commonly used FD indices (community weighted mean traits, functional richness, Rao's quadratic entropy, Petchey and Gaston's FD index) and their relationships with environmental gradients and species richness, by varying both the extent (plasticity or not) and the structure (contingency to environmental gradient due to local adaptation) of population-level ITV. Our results suggest that ITV may strongly alter the quantification of FD and the detection of ecological patterns. Our analysis highlights that 1) species trait values distributions within communities are crucial to the sensitivity to ITV, 2) ITV structure plays a major role in this sensitivity and 3) different indices are not evenly sensitive to ITV, the single-trait FD from Petchey and Gaston being the most sensitive among the four metrics tested. We conclude that the effects of intraspecific variability in trait values should be more systematically tested before drawing central conclusions on FD, and suggest the use of simulation studies for such sensitivity analyses.
DOI:10.1111/j.1438-8677.2011.00474.xURLPMID:21972934 [本文引用: 1]
The Brazilian Atlantic Forest is a typically multi-layer tropical forest, while cerrado (savanna) is a patchy habitat with different physiognomy. Despite these differences, both habitats have high light heterogeneity. Functional traits of Dalbergia nigra and D. miscolobium from the Atlantic Forest and cerrado, respectively, were evaluated under shade (25% of full sunlight) and full sunlight in a nursery experiment. We hypothesised that both species should benefit from high phenotypic plasticity in relation to light. Plasticity was estimated using the relative distance phenotypic index (RDPI). D. miscolobium had lower shoot growth under both light conditions, suggesting it has low competitive capacity in the forest environment, which could explain its limited ability to expand over areas of Atlantic Forest. The studied species exhibited photoprotection strategies under high light and improved light capture under low light. Stomatal conductance, ETR(max) (maximum electron transport rate), PPFD(sat) (saturating photosynthetically active photon flux density), chlorophyll and carotenoid content had higher RDPI than stem morphological traits. Although both species showed considerable phenotypic plasticity, D. miscolobium had higher RDPI for eight of 11 evaluated traits. This high plasticity could be one of the factors that explain the occurrence of this species in a wide range of environmental conditions, from open grassland to dense woodlands, and it could also reflect its adaptation to high light. D. nigra also had considerable plasticity and good growth performance in both shade and full sunlight, but its absence in areas of cerrado suggests that factors other than light limit its occurrence in these habitats.
DOI:10.1111/j.1365-2435.2010.01765.xURL [本文引用: 1]
P>1. Interspecific variation in plant functional traits is fast becoming popular as a tool for understanding and predicting ecosystem biogeochemistry as dependent on vegetation composition. Leaf pH has recently been shown to be a promising new candidate trait for this purpose. But how robust is leaf pH as a species trait in the face of environmental variation? We hypothesized that inherent interspecific variation in leaf pH should be greater than phenotypic variation of given species in response to soil environments.
2. We tested this hypothesis in a temperate herbaceous flora by growing 23 species experimentally in three soils of contrasting pH (ranging by almost three pH units) and related chemistry.
3. As predicted, there was large and consistent variation in leaf pH among these species, which was robust to the differences between soil types. Indeed both the species rankings and the absolute species values for leaf pH were remarkably constant in comparisons between soil types.
4. The fact that a given species can maintain a leaf pH very different from that of their soil environment, combined with the great interspecific variation in leaf pH, indicates that leaf pH really is largely a species-specific trait. Linked with recent field evidence we suggest that interspecific variation in leaf pH, while easy and cheap to assess, has important predictive power of biogeochemical properties and processes in ecosystems.
DOI:10.1038/nature16489URLPMID:26700811 [本文引用: 1]
Earth is home to a remarkable diversity of plant forms and life histories, yet comparatively few essential trait combinations have proved evolutionarily viable in today's terrestrial biosphere. By analysing worldwide variation in six major traits critical to growth, survival and reproduction within the largest sample of vascular plant species ever compiled, we found that occupancy of six-dimensional trait space is strongly concentrated, indicating coordination and trade-offs. Three-quarters of trait variation is captured in a two-dimensional global spectrum of plant form and function. One major dimension within this plane reflects the size of whole plants and their parts; the other represents the leaf economics spectrum, which balances leaf construction costs against growth potential. The global plant trait spectrum provides a backdrop for elucidating constraints on evolution, for functionally qualifying species and ecosystems, and for improving models that predict future vegetation based on continuous variation in plant form and function.
DOI:10.7668/hbnxb.2008.S1.019URL [本文引用: 1]
In order to facilitate the process of plant stomatal development and physiology study, we report a simplified method for observing stomata by using transparent tape to shave off mesophyll cells and obtain epidermis from leaf.Compared with other methods: by tearing or adhering, this method is simple, efficient and wide applied. It has a good application on soybeans, wheat, corn and other crops. It is suitable for studying different developmental stages, distribution,dynamics and morphological studies of stomata.
DOI:10.7668/hbnxb.2008.S1.019URL [本文引用: 1]
In order to facilitate the process of plant stomatal development and physiology study, we report a simplified method for observing stomata by using transparent tape to shave off mesophyll cells and obtain epidermis from leaf.Compared with other methods: by tearing or adhering, this method is simple, efficient and wide applied. It has a good application on soybeans, wheat, corn and other crops. It is suitable for studying different developmental stages, distribution,dynamics and morphological studies of stomata.
DOI:10.1890/12-1602.1URL [本文引用: 1]
The leaf economics spectrum (LES) has revolutionized the way many ecologists think about quantifying plant ecological trade-offs. In particular, the LES has connected a clear functional trade-off (long-lived leaves with slow carbon capture vs. short-lived leaves with fast carbon capture) to a handful of easily measured leaf traits. Building on this work, community ecologists are now able to quickly assess species carbon-capture strategies, which may have implications for community-level patterns such as competition or succession. However, there are a number of steps in this logic that require careful examination, and a potential danger arises when interpreting leaf-trait variation among species within communities where trait relationships are weak. Using data from 22 diverse communities, we show that relationships among three common functional traits (photosynthetic rate, leaf nitrogen concentration per mass, leaf mass per area) are weak in communities with low variation in leaf life span (LLS), especially communities dominated by herbaceous or deciduous woody species. However, globally there are few LLS data sets for communities dominated by herbaceous or deciduous species, and more data are needed to confirm this pattern. The context-dependent nature of trait relationships at the community level suggests that leaf-trait variation within communities, especially those dominated by herbaceous and deciduous woody species, should be interpreted with caution.
DOI:10.1093/aob/mct147URLPMID:23864002 [本文引用: 1]
Epiphytism imposes physiological constraints resulting from the lack of access to the nutrient sources available to ground-rooted plants. A conspicuous adaptation in response to that lack is the phytotelm (plant-held waters) of tank-bromeliad species that are often nutrient-rich. Associations with terrestrial invertebrates also result in higher plant nutrient acquisition. Assuming that tank-bromeliads rely on reservoir-assisted nutrition, it was hypothesized that the dual association with mutualistic ants and the phytotelm food web provides greater nutritional benefits to the plant compared with those bromeliads involved in only one of these two associations.
DOI:10.1111/j.1469-8137.2006.01704.xURLPMID:16684242
Leaf mass per area (LMA), nitrogen concentration (on mass and area bases, N(mass) and N(area), respectively), photosynthetic capacity (A(mass) and A(area)) and photosynthetic nitrogen use efficiency (PNUE) are key foliar traits, but few data are available from cold, high-altitude environments. Here, we systematically measured these leaf traits in 74 species at 49 research sites on the Tibetan Plateau to examine how these traits, measured near the extremes of plant tolerance, compare with global patterns. Overall, Tibetan species had higher leaf nitrogen concentrations and photosynthetic capacities compared with a global dataset, but they had a slightly lower A(mass) at a given N(mass). These leaf trait relationships were consistent with those reported from the global dataset, with slopes of the standardized major axes A(mass)-LMA, N(mass)-LMA and A(mass)-N(mass) identical to those from the global dataset. Climate only weakly modulated leaf traits. Our data indicate that covarying sets of leaf traits are consistent across environments and biogeographic regions. Our results demonstrate functional convergence of leaf trait relationships in an extreme environment.
DOI:10.1046/j.1439-0337.2002.02031.xURL [本文引用: 1]
Mit dem Ziel die Baumartenwahl für die Begründung von Mischwäldern zu unterstützen, haben wir das Wachstums und die Blatteigenschaften vier verschiedener Baumarten (Fagus sylvatica L., Acer pseudoplatanus L., Tilia platyphyllos Scop., Fraxinus excelsior L.) entlang eines Hanggradienten untersucht. Die Untersuchung wurde in Nordhessen auf einem Basaltstandort durchgeführt. Am Unterhang wies der nährstoffreiche Boden einen Skelettanteil von 10 Vol.% und am Oberhang von 25% auf (0–10 cm Bodentiefe). Bergahorne, Buchen und Sommerlinden wurden nach dem Zusammenbruch eines Fichtenbestandes gepflanzt. Eschen fanden sich als Naturverjüngung im Bestand ein. Die untersuchten Bäume waren etwa 6 Jahre alt.In beiden Untersuchungsjahren nahm der Höhenzuwachs der untersuchten Baumarten entlang des Gradienten ab. Die Reduktion war am höchsten beim Bergahorn, dessen Zuwachs am Oberhang nur 16% des Zuwachses am Unterhang betrug (Buche 33%, Sommerlinde 48%, Esche 47%). Die Blattgrößen und die Blattfläche pro Baum nahmen vom Unterhang zum Oberhang ab, während die Stomatadichten anstiegen, was eine Reaktion auf abnehmende Wasserverfügbarkeit anzeigt. Innerhalb einer Art wurden keine signifikanten Unterschiede der
 13C-Werte von Sonnenblättern an den verschiedenen Hangpositionen gefunden. Dies deutet darauf hin, dass es auf Blattebene keine Änderung der Wassernutzungseffizienz der Photosynthese gab. Die N- und K-Gehalte der Blätter waren am Unterhang am höchsten und am Mittel- und Oberhang geringer, was nicht die chemischen Eigenschaften des Bodens widerspiegelt. Wir nehmen an, dass die Pflanzenernährung durch eine abnehmende Bodenwasserverfügbarkeit eingeschränkt war. Diesen Ergebnissen zufolge sind Esche und Sommerlinde besser für den Anbau auf nährstoffreichen Böden mit einer begrenzten Wasserversorgung geeignet als Bergahorn; Buche nahm eine Mittelstellung ein.
13C-Werte von Sonnenblättern an den verschiedenen Hangpositionen gefunden. Dies deutet darauf hin, dass es auf Blattebene keine Änderung der Wassernutzungseffizienz der Photosynthese gab. Die N- und K-Gehalte der Blätter waren am Unterhang am höchsten und am Mittel- und Oberhang geringer, was nicht die chemischen Eigenschaften des Bodens widerspiegelt. Wir nehmen an, dass die Pflanzenernährung durch eine abnehmende Bodenwasserverfügbarkeit eingeschränkt war. Diesen Ergebnissen zufolge sind Esche und Sommerlinde besser für den Anbau auf nährstoffreichen Böden mit einer begrenzten Wasserversorgung geeignet als Bergahorn; Buche nahm eine Mittelstellung ein.[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s004420050986URL [本文引用: 1]
The natural ratio of stable carbon isotopes (δ13C) was compared to leaf structural and chemical characteristics in evergreen conifers in the north-central Rockies, United States. We sought a general model that would explain variation in δ13C across altitudinal gradients. Because variation in δ13C is attributed to the shifts between supply and demand for carbon dioxide within the leaf, we measured structural and chemical variables related to supply and demand. We measured stomatal density, which is related to CO2 supply to the chloroplasts, and leaf nitrogen content, which is related to CO2 demand. Leaf mass per area was measured as an intermediate between supply and demand. Models were tested on four evergreen conifers: Pseudotsuga menziesii, Abies lasiocarpa, Picea engelmannii, and Pinus contorta, which were sampled across 1800 m of altitude. We found significant variation among species in the rate of δ13C increase with altitude, ranging from 0.91‰ km–1 for A. lasiocarpa to 2.68‰ km–1 for Pinus contorta. Leaf structure and chemistry also varied with altitude: stomatal density decreased, leaf mass per area increased, but leaf nitrogen content (per unit area) was constant. The regressions on altitude were particularly robust in Pinus contorta. Variables were derived to describe the balance between supply and demand; these variables were stomata per gram of nitrogen and stomata per gram of leaf mass. Both derived variables should be positively related to internal CO2 supply and thus negatively related to δ13C. As expected, both derived variables were negatively correlated with δ13C. In fact, the regression on stomatal density per gram was the best fit in the study (r 2=0.72, P<0.0001); however, the relationships were species specific. The only general relationship observed was between δ13C and LMA: δ13C (‰)=–32.972+ 0.0173×LMA (r 2=0.45, P<0.0001). We conclude that species specificity of the isotopic shift indicates that evergreen conifers demonstrate varying degrees of functional plasticity across environmental gradients, while the observed convergence of δ13C with LMA suggests that internal resistance may be the key to understanding inter-specific isotopic variation across altitude.
DOI:10.3724/SP.J.1259.2015.00159URL [本文引用: 1]
DOI:10.3724/SP.J.1259.2015.00159URL [本文引用: 1]
DOI:10.3724/SP.J.1258.2012.00088URL
Leaf morphology is closely related to the specific environment and provides the most useful characteristics to understand plant response and adaptation strategy to environmental change. Leaf morphology plasticity is obviously related to the temporal and spatial variation of environmental variables, which are useful to plants to enhance their ability to survive. Consequently, for many years, studies on plant physiology, plant ecology and physiological ecology focused on leaf morphology. We establish a simple category of leaf morphology classification. Simultaneously, based on the principle of material and energy balances, we systematically review the relationships among environment, leaf morphology and energy balances (or material changes), and emphasize that leaf morphology responded or adapted to lower water availability and higher radiation (or temperature) in arid ecosystems. In conclusion, we submit and discuss existing problems in leaf morphology based on the weaknesses of previous studies.
DOI:10.3724/SP.J.1258.2012.00088URL
Leaf morphology is closely related to the specific environment and provides the most useful characteristics to understand plant response and adaptation strategy to environmental change. Leaf morphology plasticity is obviously related to the temporal and spatial variation of environmental variables, which are useful to plants to enhance their ability to survive. Consequently, for many years, studies on plant physiology, plant ecology and physiological ecology focused on leaf morphology. We establish a simple category of leaf morphology classification. Simultaneously, based on the principle of material and energy balances, we systematically review the relationships among environment, leaf morphology and energy balances (or material changes), and emphasize that leaf morphology responded or adapted to lower water availability and higher radiation (or temperature) in arid ecosystems. In conclusion, we submit and discuss existing problems in leaf morphology based on the weaknesses of previous studies.
DOI:10.1038/s41598-019-42335-2URLPMID:30967600 [本文引用: 1]
Leaf functional traits have attracted the attention of ecologists for several decades, but few studies have systematically assessed leaf morphological traits (termed &quot;economic traits&quot;), stomatal (termed &quot;hydraulic&quot;), and anatomical traits of entire forest communities, thus it is unclear whether their relationships are consistent among trees, shrubs, and herbs, and which anatomical traits should be assigned to economical or hydraulic traits. In this study, we collected leaf samples of 106 plant species in temperate forests and 164 plant species in subtropical forests and determined nine key functional traits. We found that functional traits differed between temperate and subtropical forests. Leaf traits also differed between different plant functional groups, irrespective of forest type; dry matter content, stomatal density, and cell tense ratio followed the order trees?&gt;?shrubs?&gt;?herbs, whereas specific leaf area and sponginess ratio showed the opposite pattern. The correlations of leaf traits were not consistent among trees, shrubs, and herbs, which may reflect different adaptive strategies. Principal component analysis indicated that leaf economics and hydraulic traits were uncoupled in temperate and subtropical forests, and correlations of anatomical traits and economic and hydraulic traits were weak, indicating anatomical traits should be emphasized in future studies.
DOI:10.1046/j.1365-2435.2003.00698.xURL [本文引用: 1]
DOI:10.1890/0012-9658(2001)082[0453:GSCCOL]2.0.CO;2URL [本文引用: 2]
DOI:10.1093/treephys/24.2.141URLPMID:14676031
Leaf architecture, stand leaf area index and canopy light interception were studied in 13 poplar clones growing in a second rotation of a coppice plantation, to determine the role of leaf architectural attributes on canopy light-harvesting efficiency and to assess biomass investment in leaf support tissue. Stand leaf area index (L) varied from 2.89 to 6.99, but L was only weakly associated with canopy transmittance (TC). The weak relationship between TC and L was a result of a higher degree of foliage aggregation at larger values of L, leading to lower light-interception efficiency in stands with greater total leaf area. We observed a strong increase in leaf aggregation and a decrease in light-harvesting efficiency with decreasing mean leaf petiole length (PL) but not with leaf size, possibly because, in cordate or deltoid poplar leaves, most of the leaf area is located close to the petiole attachment to the lamina. Although PL was the key leaf characteristic of light-harvesting efficiency, clones with longer petioles had larger biomass investments in petioles, and there was a negative relationship between PL and L, demonstrating that enhanced light harvesting may lead to an overall decline in photosynthesizing leaf surface. Upper-canopy leaves were generally larger and had greater dry mass (MA) and nitrogen per unit area (NA) than lower-canopy leaves. Canopy plasticity in MA and NA was higher in clones with higher foliar biomass investment in midrib, and lower in clones with relatively longer petioles. These relationships suggest that there is a trade-off between photosynthetic plasticity and biomass investment in support, and also that high light-harvesting efficiency may be associated with lower photosynthetic plasticity. Our results demonstrate important clonal differences in leaf aggregation that are linked to leaf structure and biomass allocation patterns within the leaf.
DOI:10.1111/geb.2009.18.issue-2URL [本文引用: 1]
DOI:10.1111/fec.2015.29.issue-1URL
DOI:10.1073/pnas.94.25.13730URLPMID:9391094 [本文引用: 1]
Despite striking differences in climate, soils, and evolutionary history among diverse biomes ranging from tropical and temperate forests to alpine tundra and desert, we found similar interspecific relationships among leaf structure and function and plant growth in all biomes. Our results thus demonstrate convergent evolution and global generality in plant functioning, despite the enormous diversity of plant species and biomes. For 280 plant species from two global data sets, we found that potential carbon gain (photosynthesis) and carbon loss (respiration) increase in similar proportion with decreasing leaf life-span, increasing leaf nitrogen concentration, and increasing leaf surface area-to-mass ratio. Productivity of individual plants and of leaves in vegetation canopies also changes in constant proportion to leaf life-span and surface area-to-mass ratio. These global plant functional relationships have significant implications for global scale modeling of vegetation-atmosphere CO2 exchange.
DOI:10.3732/ajb.0900290URLPMID:21622407 [本文引用: 1]
Many key aspects of early angiosperms are poorly known, including their ecophysiology and associated habitats. Evidence for fast-growing, weedy angiosperms comes from the Early Cretaceous Potomac Group, where angiosperm fossils, some of them putative herbs, are found in riparian depositional settings. However, inferences of growth rate from sedimentology and growth habit are somewhat indirect; also, the geographic extent of a weedy habit in early angiosperms is poorly constrained. Using a power law between petiole width and leaf mass, we estimated the leaf mass per area (LMA) of species from three Albian (110-105 Ma) fossil floras from North America (Winthrop Formation, Patapsco Formation of the Potomac Group, and the Aspen Shale). All LMAs for angiosperm species are low (&lt;125 g/m(2); mean = 76 g/m(2)) but are high for gymnosperm species (&gt;240 g/m(2); mean = 291 g/m(2)). On the basis of extant relationships between LMA and other leaf economic traits such as photosynthetic rate and leaf lifespan, we conclude that these Early Cretaceous landscapes were populated with weedy angiosperms with short-lived leaves (&lt;12 mo). The unrivalled capacity for fast growth observed today in many angiosperms was in place by no later than the Albian and likely played an important role in their subsequent ecological success.
DOI:10.1111/fec.2006.20.issue-2URL [本文引用: 2]
DOI:10.1104/pp.111.173856URL [本文引用: 1]
Across plant species, leaves vary enormously in their size and their venation architecture, of which one major function is to replace water lost to transpiration. The leaf hydraulic conductance (K-leaf) represents the capacity of the transport system to deliver water, allowing stomata to remain open for photosynthesis. Previous studies showed that K-leaf relates to vein density (vein length per area). Additionally, venation architecture determines the sensitivity of K-leaf to damage; severing the midrib caused K-leaf and gas exchange to decline, with lesser impacts in leaves with higher major vein density that provided more numerous water flow pathways around the damaged vein. Because xylem embolism during dehydration also reduces K-leaf, we hypothesized that higher major vein density would also reduce hydraulic vulnerability. Smaller leaves, which generally have higher major vein density, would thus have lower hydraulic vulnerability. Tests using simulations with a spatially explicit model confirmed that smaller leaves with higher major vein density were more tolerant of major vein embolism. Additionally, for 10 species ranging strongly in drought tolerance, hydraulic vulnerability, determined as the leaf water potential at 50% and 80% loss of K-leaf, was lower with greater major vein density and smaller leaf size (vertical bar r vertical bar = 0.85-0.90; P < 0.01). These relationships were independent of other aspects of physiological and morphological drought tolerance. These findings point to a new functional role of venation architecture and small leaf size in drought tolerance, potentially contributing to well-known biogeographic trends in leaf size.
DOI:10.1111/ele.12508URLPMID:26415616 [本文引用: 1]
Recent studies have shown that accounting for intraspecific trait variation (ITV) may better address major questions in community ecology. However, a general picture of the relative extent of ITV compared to interspecific trait variation in plant communities is still missing. Here, we conducted a meta-analysis of the relative extent of ITV within and among plant communities worldwide, using a data set encompassing 629 communities (plots) and 36 functional traits. Overall, ITV accounted for 25% of the total trait variation within communities and 32% of the total trait variation among communities on average. The relative extent of ITV tended to be greater for whole-plant (e.g. plant height) vs. organ-level traits and for leaf chemical (e.g. leaf N and P concentration) vs. leaf morphological (e.g. leaf area and thickness) traits. The relative amount of ITV decreased with increasing species richness and spatial extent, but did not vary with plant growth form or climate. These results highlight global patterns in the relative importance of ITV in plant communities, providing practical guidelines for when researchers should include ITV in trait-based community and ecosystem studies.
DOI:10.1007/s11258-011-0009-xURL
Community structure and leaf traits are important elements of terrestrial ecosystems. Changes of community structure and leaf traits are of particular use in the study of the influence of climate change on terrestrial ecosystems. Patterns of community structure (including species richness, above- and below-ground biomass) and leaf traits (including leaf mass per area (LMA), nitrogen content both on mass and area bases (Nmass and Narea), and foliar δ13C) from 19 grassland plots along an altitudinal transect at Hongchiba in Chongqing, China, were analyzed. Species richness along the altitudinal transect had a hump-shaped pattern. Above-ground biomass had a quadratic decrease along the altitudinal gradient whereas below-ground biomass had the opposite pattern. Change of above-ground biomass of various taxonomic groups with altitude was also studied. Poaceae showed strong negative relationships and Asteraceae showed a hump-shaped relationship with increase of altitude. Five common species of the grassland, Trifolium pratense, Geranium wilfordii,Aster tataricus, Leontopodium leontopodioides, and Spiraea prunifolia, were particularly studied for variation of leaf traits along the altitudinal gradient. Averaged for all species, LMA, Narea and foliar δ13C had positive correlations with altitude. Nmass did not change significantly as altitude increased. LMA and Narea showed significant positive relationships with foliar δ13C. The adaptive features of leaf traits among different species were not consistent. The study highlights specific adaptation patterns in relation to altitude for different plant species, provides further insights into adaptive trends of community structure and leaf traits in a specific ecological region filling a gap in the definition of global patterns, and adds to the understanding of how adaptive patterns of plants may respond to global climate change.
DOI:10.11820/dlkxjz.2011.11.014URL [本文引用: 1]
A variety of environment factors show continuous gradient changes under an altitude gradient, so it is very significant to reveal the responses of the plants traits under the climatic change under altitude gradient and the results can also provide an foundation to understand the relationship between the terrestrial ecosystem and the global change. This article puts emphases on the changes of leaf span, specific leaf area, leaf nitrogen content, chlorophyll, stomata conductance, carboxylation efficiency, water use efficiency and leaf δ13C along an altitude gradient. This article also explores some uncertainties on the interpretation of mechanisms of leaf traits along the altitude gradient. This review provides a good knowledge for directing the relevant studies in China.
DOI:10.11820/dlkxjz.2011.11.014URL [本文引用: 1]
A variety of environment factors show continuous gradient changes under an altitude gradient, so it is very significant to reveal the responses of the plants traits under the climatic change under altitude gradient and the results can also provide an foundation to understand the relationship between the terrestrial ecosystem and the global change. This article puts emphases on the changes of leaf span, specific leaf area, leaf nitrogen content, chlorophyll, stomata conductance, carboxylation efficiency, water use efficiency and leaf δ13C along an altitude gradient. This article also explores some uncertainties on the interpretation of mechanisms of leaf traits along the altitude gradient. This review provides a good knowledge for directing the relevant studies in China.
DOI:10.1007/s00442-009-1333-xURL [本文引用: 1]
Although of primary importance to explain plant community structure, general relationships between plant traits, resource depletion and competitive outcomes remain to be quantified across species. Here, we used a comparative approach to test whether instantaneous measurements of plant traits can capture both the amount of resources depleted under plant cover over time (competitive effect) and the way competitors perceived this resource depletion (competitive response). We performed a large competition experiment in which phytometers from a single grass species were transplanted within 18 different monocultures grown in a common-garden experiment, with a time-integrative quantification of light and water depletion over the phytometers’ growing season. Resource-capturing traits were measured on both phytometers (competitive response traits) and monocultures (competitive effect traits). The total amounts of depleted light and water availabilities over the season strongly differed among monocultures; they were best estimated by instantaneous measurements of height and rooting depth, respectively, performed when either light or water became limiting. Specific leaf area and leaf water potential, two competitive response traits measured at the leaf level, were good predictors of changes in phytometer performance under competition, and reflected the amount of light and water, respectively, perceived by plants throughout their lifespan. Our results demonstrated the relevance of instantaneous measures of plant traits as indicators of resource depletion over time, validating the trait-based approach for competition ecology.
DOI:10.1111/oik.2007.116.issue-5URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s11676-016-0290-6URL
DOI:10.11821/dlxb201511004URL
To explore the large-scale changes in leaf functional traits from the species level to community level and their influencing factors in Eastern China, we investigated leaf area (LA), leaf thickness (LT), specific leaf area (SLA) and leaf dry matter content (LDMC) across 847 species from nine typical forest ecosystems along the North-South Transect of Eastern China (NSTEC) between July and August in 2013, and we also calculated the community weighted means (CWM) of leaf traits on the basis of the relative dominance of each species within the community. Our results showed that, for all 847 species, the means (± SE) of LA, LT, SLA, and LDMC were 2860.01 ± 135.37 mm2, 0.17 ± 0.003 mm, 20.15 ± 0.43 m2 kg-1 and 316.73 ± 3.81 mg g-1, respectively. In addition, SLA increased and LDMC decreased with the increasing latitude, whereas no clear latitudinal trends of LA and LT were found (R2 = 0.02-0.06), as latitudinal changes were mainly affected by plant functional types. At the community level, more significant spatial patterns of leaf traits were observed (R2 = 0.46-0.71). Generally, as latitude increased, LACWM and SLACWM varied nonlinearly, while LTCWM increased and LDMCCWM decreased linearly, which were influenced by climate and soil N. This study improved the biogeographical database of leaf functional traits from tropical to cold-temperate forests in eastern China and emphasized the importance of scaling-up in the study of plant traits.
DOI:10.11821/dlxb201511004URL
To explore the large-scale changes in leaf functional traits from the species level to community level and their influencing factors in Eastern China, we investigated leaf area (LA), leaf thickness (LT), specific leaf area (SLA) and leaf dry matter content (LDMC) across 847 species from nine typical forest ecosystems along the North-South Transect of Eastern China (NSTEC) between July and August in 2013, and we also calculated the community weighted means (CWM) of leaf traits on the basis of the relative dominance of each species within the community. Our results showed that, for all 847 species, the means (± SE) of LA, LT, SLA, and LDMC were 2860.01 ± 135.37 mm2, 0.17 ± 0.003 mm, 20.15 ± 0.43 m2 kg-1 and 316.73 ± 3.81 mg g-1, respectively. In addition, SLA increased and LDMC decreased with the increasing latitude, whereas no clear latitudinal trends of LA and LT were found (R2 = 0.02-0.06), as latitudinal changes were mainly affected by plant functional types. At the community level, more significant spatial patterns of leaf traits were observed (R2 = 0.46-0.71). Generally, as latitude increased, LACWM and SLACWM varied nonlinearly, while LTCWM increased and LDMCCWM decreased linearly, which were influenced by climate and soil N. This study improved the biogeographical database of leaf functional traits from tropical to cold-temperate forests in eastern China and emphasized the importance of scaling-up in the study of plant traits.
DOI:10.1146/annurev.ecolsys.33.010802.150452URL [本文引用: 2]
DOI:10.1111/1365-2745.12598URL
DOI:10.1126/science.aal4760URLPMID:28860384 [本文引用: 3]
Leaf size varies by over a 100,000-fold among species worldwide. Although 19th-century plant geographers noted that the wet tropics harbor plants with exceptionally large leaves, the latitudinal gradient of leaf size has not been well quantified nor the key climatic drivers convincingly identified. Here, we characterize worldwide patterns in leaf size. Large-leaved species predominate in wet, hot, sunny environments; small-leaved species typify hot, sunny environments only in arid conditions; small leaves are also found in high latitudes and elevations. By modeling the balance of leaf energy inputs and outputs, we show that daytime and nighttime leaf-to-air temperature differences are key to geographic gradients in leaf size. This knowledge can enrich &quot;next-generation&quot; vegetation models in which leaf temperature and water use during photosynthesis play key roles.
URL [本文引用: 1]
利用光学显微镜,研究陇西黄土高原兴隆山自然保护区2 600~3 100 m海拔带内高山杜鹃(Rhododendron lapponicum)和烈香杜鹃(Rhododendron anthopogonoides)(高山杜鹃分布在北坡,烈香杜鹃分布在南坡)叶片气孔长度和密度随海拔高度的变化情况,并通过建立大气温度、大气CO2浓度等环境因子与海拔高度之间的变化关系,探讨了杜鹃叶片气孔长度和密度随海拔高度升高而发生变化的原因。结果表明:海拔高度由低到高,高山杜鹃叶片气孔长度有减小趋势,气孔密度有显著增加趋势;烈香杜鹃叶片气孔长度随海拔高度升高先增大后减小,气孔密度先减小后增大。海拔高度的变化能够引起大气温度、大气CO2浓度和地势等的变化。高山杜鹃通过增加叶片气孔长度并减小叶片气孔密度来适应环境温度和CO2浓度的变化,烈香杜鹃则通过增加叶片气孔长度,并减小叶片气孔密度达到对环境温度、CO2浓度和土壤湿度的最大适合度。
URL [本文引用: 1]
利用光学显微镜,研究陇西黄土高原兴隆山自然保护区2 600~3 100 m海拔带内高山杜鹃(Rhododendron lapponicum)和烈香杜鹃(Rhododendron anthopogonoides)(高山杜鹃分布在北坡,烈香杜鹃分布在南坡)叶片气孔长度和密度随海拔高度的变化情况,并通过建立大气温度、大气CO2浓度等环境因子与海拔高度之间的变化关系,探讨了杜鹃叶片气孔长度和密度随海拔高度升高而发生变化的原因。结果表明:海拔高度由低到高,高山杜鹃叶片气孔长度有减小趋势,气孔密度有显著增加趋势;烈香杜鹃叶片气孔长度随海拔高度升高先增大后减小,气孔密度先减小后增大。海拔高度的变化能够引起大气温度、大气CO2浓度和地势等的变化。高山杜鹃通过增加叶片气孔长度并减小叶片气孔密度来适应环境温度和CO2浓度的变化,烈香杜鹃则通过增加叶片气孔长度,并减小叶片气孔密度达到对环境温度、CO2浓度和土壤湿度的最大适合度。
DOI:10.1080/17429145.2014.990063URL
Plant functional traits built the relationships between plant diversity, species composition, and physiology along with the environmental changes, thus influencing soil microbial community. As the sensitivity indicators, soil microbial biomass and plant functional traits responses soil micro-organism and plant characteristics in direct way. Ten plant functional traits of 149 species and soil microbial biomass (carbon, nitrogen, and phosphorus) were analyzed across the different vegetation types (forest, forest-steppe, and steppe) that are divided by environmental gradient (temperature and precipitation), aimed to find the correlations among them. Our results confirmed the greatest values of plant functional traits (except the leaf density and the fine root density) that were distributed in the steppe zone, mainly due to the different mean annual temperature and mean annual precipitation conditions. For different plant growth forms, the plant functional traits were significant differences among the vegetation zones. The advantages of higher rate nutrient cycling, plentiful biomass supplements, and favorite habit conditions lead to the forest-steppe zone with the highest C-mic and N-mic concentrations. The canonical correlation analysis indicated that leaf nitrogen, root nitrogen, and fine root densities were correlated with root exudate and tissue which affected the concentrations of soil organic carbon (SOC) and total nitrogen (N), consequently impacting soil microbial biomass carbon (C-mic) and soil microbial biomass nitrogen (N-mic). Soil is the medium that connects micro-organism and plant root system that influenced leaf nitrogen, root nitrogen, and fine root density of plant functional traits, the concentrations of SOC and total N that plant feedback are consequently influencing C-mic and N-mic.
A trait-based approach to community assembly: Partitioning of species trait values into within- and among-community components
2007
Variation in leaf traits through seasons and N-?availability levels and its consequences for ranking grassland species
2005
On the importance of intraspecific variability for the quantification of functional diversity
1
2012
... 植物的性状值常因其生境的不同而不同(
Phenotypic plasticity to light of two congeneric trees from contrasting habitats: Brazilian Atlantic Forest versus cerrado (savanna)
1
2011
... 植物的性状值常因其生境的不同而不同(
Leaf pH as a plant trait: Species- driven rather than soil-driven variation
1
2011
... 植物的性状值常因其生境的不同而不同(
The global spectrum of plant form and function
1
2016
... 植物的性状值常因其生境的不同而不同(
一种获得叶片表皮观察气孔的简易方法及其应用
1
2008
... 采用印迹法和透明胶带结合的改良方法测定气孔性状(
一种获得叶片表皮观察气孔的简易方法及其应用
1
2008
... 采用印迹法和透明胶带结合的改良方法测定气孔性状(
Leaf traits within communities: Context may affect the mapping of traits to function
1
2013
... 种内与环境(海拔和气候因子)显著相关的叶片性状明显少于种间水平(
Carnivory in the bromeliad
1
1984
... 在本研究中, 叶片长度、叶片宽度和叶面积随海拔的升高而降低, 这在种内和种间分析中是一致的, 说明减小叶片大小既是木本植物对高海拔环境的可塑性反应, 也是其适应性结果, 与
Leaf traits variation in
2014
长白山阔叶木本植物叶片形态性状沿海拔梯度的响应特征
2014
长白山阔叶木本植物叶片形态性状沿海拔梯度的响应特征
2014
A test of the generality of leaf trait relationships on the Tibetan Plateau
2006
Growth and leaf traits of four broad-leaved tree species along a hillside gradient
1
2002
... 在本研究中, 叶片长度、叶片宽度和叶面积随海拔的升高而降低, 这在种内和种间分析中是一致的, 说明减小叶片大小既是木本植物对高海拔环境的可塑性反应, 也是其适应性结果, 与
西藏紫花针茅叶功能性状沿降水梯度的变化
2012
西藏紫花针茅叶功能性状沿降水梯度的变化
2012
长白山森林不同演替阶段比叶面积及其影响因子
1
2015
... 叶片厚度和比叶面积的种间海拔变异式样是非直线的.两者在低海拔地区变化不明显, 在高海拔地区呈明显的海拔变异趋势, 这可能是因为低海拔地区的环境条件比较温和, 植物受到的环境胁迫较小, 其叶片特征的环境变异也相对和缓.叶片厚度在海拔2 700 m后随海拔升高而增加, 较厚的叶片可以通过增加单位叶片面积的氮含量和光合组织的体积增加强辐照下的光合能力, 而且较厚的叶片具有保温和耐辐射的作用, 以适应高海拔的寒冷气候和近日照的环境条件(
长白山森林不同演替阶段比叶面积及其影响因子
1
2015
... 叶片厚度和比叶面积的种间海拔变异式样是非直线的.两者在低海拔地区变化不明显, 在高海拔地区呈明显的海拔变异趋势, 这可能是因为低海拔地区的环境条件比较温和, 植物受到的环境胁迫较小, 其叶片特征的环境变异也相对和缓.叶片厚度在海拔2 700 m后随海拔升高而增加, 较厚的叶片可以通过增加单位叶片面积的氮含量和光合组织的体积增加强辐照下的光合能力, 而且较厚的叶片具有保温和耐辐射的作用, 以适应高海拔的寒冷气候和近日照的环境条件(
Altitude trends in conifer leaf morphology and stable carbon isotope composition
1
2000
... 叶片厚度和比叶面积的种间海拔变异式样是非直线的.两者在低海拔地区变化不明显, 在高海拔地区呈明显的海拔变异趋势, 这可能是因为低海拔地区的环境条件比较温和, 植物受到的环境胁迫较小, 其叶片特征的环境变异也相对和缓.叶片厚度在海拔2 700 m后随海拔升高而增加, 较厚的叶片可以通过增加单位叶片面积的氮含量和光合组织的体积增加强辐照下的光合能力, 而且较厚的叶片具有保温和耐辐射的作用, 以适应高海拔的寒冷气候和近日照的环境条件(
中国东部南北样带暖温带区栎属树种叶片形态性状对气候条件的响应
2013
中国东部南北样带暖温带区栎属树种叶片形态性状对气候条件的响应
2013
放牧对草原植物功能性状及其权衡关系的调控
1
2015
... 种内与环境(海拔和气候因子)显著相关的叶片性状明显少于种间水平(
放牧对草原植物功能性状及其权衡关系的调控
1
2015
... 种内与环境(海拔和气候因子)显著相关的叶片性状明显少于种间水平(
干旱区叶片形态特征与植物响应和适应的关系
2012
干旱区叶片形态特征与植物响应和适应的关系
2012
甘肃省摩天岭北坡木本植物叶性状变异及关联
2018
甘肃省摩天岭北坡木本植物叶性状变异及关联
2018
Variation in leaf morphological, stomatal, and anatomical traits and their relationships in temperate and subtropical forests
1
2019
... 对于气孔长度、气孔宽度与海拔的关系, 种内和种间分析出现了相反的结果.在种间分析中, 气孔长度、气孔宽度与海拔呈负相关关系, 这可能是因为在高海拔地区生境恶劣, 如低温、大风和高紫外线辐射等, 小的气孔有利于植物快速打开和关闭以快速应对多变的环境条件(
Leaf-size divergence along rainfall and soil-nutrient gradients: Is the method of size reduction common among clades?
1
2003
... 在本研究中, 叶片长度、叶片宽度和叶面积随海拔的升高而降低, 这在种内和种间分析中是一致的, 说明减小叶片大小既是木本植物对高海拔环境的可塑性反应, 也是其适应性结果, 与
Global-scale climatic controls of leaf dry mass per area, density, and thickness in trees and shrubs
2
2001
... 叶片厚度和比叶面积的种间海拔变异式样是非直线的.两者在低海拔地区变化不明显, 在高海拔地区呈明显的海拔变异趋势, 这可能是因为低海拔地区的环境条件比较温和, 植物受到的环境胁迫较小, 其叶片特征的环境变异也相对和缓.叶片厚度在海拔2 700 m后随海拔升高而增加, 较厚的叶片可以通过增加单位叶片面积的氮含量和光合组织的体积增加强辐照下的光合能力, 而且较厚的叶片具有保温和耐辐射的作用, 以适应高海拔的寒冷气候和近日照的环境条件(
... 种内分析中, 叶片性状与气候的关系没有统一趋势(
Petiole length and biomass investment in support modify light interception efficiency in dense poplar plantations
2004
A global study of relationships between leaf traits, climate and soil measures of nutrient fertility
1
2009
... 种内分析中, 叶片性状与气候的关系没有统一趋势(
Radial changes in wood specific gravity of tropical trees: Inter- and intraspecific variation during secondary succession
2015
From tropics to tundra: Global convergence in plant functioning
1
1997
... 植物的性状值常因其生境的不同而不同(
Leaf economic traits from fossils support a weedy habit for early angiosperms
1
2010
... 植物与其生存环境的关系是植物生态学研究的核心问题.植物通过调节外部形态和内部生理特性以响应和适应环境的变化, 同时改变植物对生态系统功能的贡献.基于性状的研究方法已被广泛用于植物适应策略的研究(
Plasticity in leaf traits of 38 tropical tree species in response to light; relationships with light demand and adult stature
2
2006
... 植物与其生存环境的关系是植物生态学研究的核心问题.植物通过调节外部形态和内部生理特性以响应和适应环境的变化, 同时改变植物对生态系统功能的贡献.基于性状的研究方法已被广泛用于植物适应策略的研究(
... 植物的性状值常因其生境的不同而不同(
Decline of leaf hydraulic conductance with dehydration: Relationship to leaf size and venation architecture
1
2011
... 植物与其生存环境的关系是植物生态学研究的核心问题.植物通过调节外部形态和内部生理特性以响应和适应环境的变化, 同时改变植物对生态系统功能的贡献.基于性状的研究方法已被广泛用于植物适应策略的研究(
A global meta-analysis of the relative extent of intraspecific trait variation in plant communities
1
2015
... 种内与环境(海拔和气候因子)显著相关的叶片性状明显少于种间水平(
Adaptive characteristics of grassland community structure and leaf traits along an altitudinal gradient on a subtropical mountain in Chongqing, China
2012
植物叶片性状沿海拔梯度变化研究进展
1
2011
... 植物与其生存环境的关系是植物生态学研究的核心问题.植物通过调节外部形态和内部生理特性以响应和适应环境的变化, 同时改变植物对生态系统功能的贡献.基于性状的研究方法已被广泛用于植物适应策略的研究(
植物叶片性状沿海拔梯度变化研究进展
1
2011
... 植物与其生存环境的关系是植物生态学研究的核心问题.植物通过调节外部形态和内部生理特性以响应和适应环境的变化, 同时改变植物对生态系统功能的贡献.基于性状的研究方法已被广泛用于植物适应策略的研究(
Competition, traits and resource depletion in plant communities
1
2009
... 植物的性状值常因其生境的不同而不同(
Let the concept of trait be functional!
1
2007
... 植物的性状值常因其生境的不同而不同(
植物叶片性状对气候变化的响应研究进展
1
2015
... 植物与其生存环境的关系是植物生态学研究的核心问题.植物通过调节外部形态和内部生理特性以响应和适应环境的变化, 同时改变植物对生态系统功能的贡献.基于性状的研究方法已被广泛用于植物适应策略的研究(
植物叶片性状对气候变化的响应研究进展
1
2015
... 植物与其生存环境的关系是植物生态学研究的核心问题.植物通过调节外部形态和内部生理特性以响应和适应环境的变化, 同时改变植物对生态系统功能的贡献.基于性状的研究方法已被广泛用于植物适应策略的研究(
Variations in leaf functional traits among plant species grouped by growth and leaf types in Zhenjiang, China
2017
中国森林叶片功能属性的纬度格局及其影响因素
2015
中国森林叶片功能属性的纬度格局及其影响因素
2015
Plant ecological strategies: Some leading dimensions of variation between species
2
2002
... 植物与其生存环境的关系是植物生态学研究的核心问题.植物通过调节外部形态和内部生理特性以响应和适应环境的变化, 同时改变植物对生态系统功能的贡献.基于性状的研究方法已被广泛用于植物适应策略的研究(
... 叶片厚度和比叶面积的种间海拔变异式样是非直线的.两者在低海拔地区变化不明显, 在高海拔地区呈明显的海拔变异趋势, 这可能是因为低海拔地区的环境条件比较温和, 植物受到的环境胁迫较小, 其叶片特征的环境变异也相对和缓.叶片厚度在海拔2 700 m后随海拔升高而增加, 较厚的叶片可以通过增加单位叶片面积的氮含量和光合组织的体积增加强辐照下的光合能力, 而且较厚的叶片具有保温和耐辐射的作用, 以适应高海拔的寒冷气候和近日照的环境条件(
Leaf traits of African woody savanna species across climate and soil fertility gradients: Evidence for conservative versus acquisitive resource-use strategies
2016
Global climatic drivers of leaf size
3
2017
... 植物的性状值常因其生境的不同而不同(
... 在本研究中, 叶片长度、叶片宽度和叶面积随海拔的升高而降低, 这在种内和种间分析中是一致的, 说明减小叶片大小既是木本植物对高海拔环境的可塑性反应, 也是其适应性结果, 与
... 种内分析中, 叶片性状与气候的关系没有统一趋势(
Insights into the differences in leaf functional traits of heterophyllous
2015
杜鹃叶片气孔长度和密度对海拔变化的响应
1
2012
... 对于气孔长度、气孔宽度与海拔的关系, 种内和种间分析出现了相反的结果.在种间分析中, 气孔长度、气孔宽度与海拔呈负相关关系, 这可能是因为在高海拔地区生境恶劣, 如低温、大风和高紫外线辐射等, 小的气孔有利于植物快速打开和关闭以快速应对多变的环境条件(
杜鹃叶片气孔长度和密度对海拔变化的响应
1
2012
... 对于气孔长度、气孔宽度与海拔的关系, 种内和种间分析出现了相反的结果.在种间分析中, 气孔长度、气孔宽度与海拔呈负相关关系, 这可能是因为在高海拔地区生境恶劣, 如低温、大风和高紫外线辐射等, 小的气孔有利于植物快速打开和关闭以快速应对多变的环境条件(
Plant functional traits and soil microbial biomass in different vegetation zones on the Loess Plateau
2014
Altitudinal variation in leaf construction cost and energy content of
2012

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}