 ,*, 陶泽兴, 葛全胜中国科学院地理科学与资源研究所中国科学院陆地表层格局与模拟重点实验室, 北京 100101
,*, 陶泽兴, 葛全胜中国科学院地理科学与资源研究所中国科学院陆地表层格局与模拟重点实验室, 北京 100101Effects of climate variation on the first leaf dates of 39 woody species and their thermal requirements in Xi’an, China
Huan-Jiong WANG,*, Ze-Xing TAO, Quan-Sheng GEKey Laboratory of Land Surface Pattern and Simulation, Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing, 100101, China通讯作者: 通讯作者Author for correspondence (E-mail:jshe@pku.edu.cn)
编委: 李敏
收稿日期:2019-07-8接受日期:2019-09-15网络出版日期:2019-10-20
| 基金资助: |
Received:2019-07-8Accepted:2019-09-15Online:2019-10-20
| Fund supported: |

摘要
过去几十年来暖春等异常气候事件发生的频次和强度显著增加, 使植物春季物候期发生了明显变化。但异常气候事件对植物春季物候积温需求的影响仍不清楚, 限制了对未来物候变化预测精度的提升。该研究利用西安植物园1963-2018年39种木本植物的展叶始期和相应气象数据, 首先根据3-4月平均气温划分了偏冷年、正常年和偏暖年, 对比了冷暖年相对于正常年的展叶始期变化。其次, 利用3种积温算法计算了各植物逐年的展叶始期积温需求, 比较了积温需求在冷暖年和正常年的差异。最后, 评估了传统积温模型在模拟偏冷或偏暖年展叶始期时的误差。结果表明, 所有植物的展叶始期在偏暖年比正常年平均早8.6天, 而在偏冷年平均晚8.2天。在偏暖年, 大多数物种展叶始期的积温需求(以5 ℃为阈值, 平均257.5度日)显著高于正常年(平均195.1度日); 在偏冷年的积温需求(平均168.0度日)低于正常年, 但在统计上差异不显著。就不同类群而言, 古老类群相对于年轻类群在偏冷年的推迟天数更多, 积温需求变化较小, 但在偏暖年无显著差异。不同生活型间物候与积温需求变化也无显著差异。造成偏暖年积温需求增加的可能原因是偏暖年冬季气温较高, 导致植物受到的冷激程度减轻, 从而抑制了后续的展叶。在正常年, 积温模型模拟木本植物展叶始期的平均误差仅为0.4-1.9天。在偏暖年和偏冷年, 模拟值分别比观测值平均早4.1天和晚3.0天。因此在预测未来物候变化时, 需要考虑气候波动条件下的积温需求变化。
关键词:
Abstract
Aims The frequency and intensity of exceptional climatic events such as warm spring have increased significantly over the past few decades and exerted a significant impact on the spring phenophases of plants. However, the influence of exceptional climatic events on the thermal requirements of spring phenophases is still unclear, which limits the predictive accuracy of the future phenological changes. Here we aim to demonstrate how the first leaf dates of woody plants and their associated thermal requirements change under exceptional climatic conditions and how exceptional climatic conditions affect the ability of the growing degree model to predict leaf unfolding date. Methods Using data on the first leaf date of 39 woody species at Xi’an Botanical Garden from 1963 to 2018 and the corresponding meteorological data, this study firstly classified each year into the cold year, normal year and warm year. Subsequently, we analyzed the phenological change in the years with abnormal climate compared to the years with normal climate. Second, three kinds of algorithms were used to calculate the thermal requirements of the first leaf date for each plant, and the difference in the thermal requirements between years with abnormal climate and normal climate was compared. Finally, the error of the traditional growing degree day model in the simulation of the first leaf date in exceptional climatic conditions was assessed. Important findings For all plant species, the first leaf date was earlier in warm years than that in normal years with a mean advance of 8.6 days, and it was later in cold years with a mean delay of 8.2 days. In warm years, the thermal requirement of the first leaf date (257.5 degree days on average) was significantly higher than that in normal years (195.1 degree days on average, p < 0.05) for most species. However, in cold years, the thermal requirement (168.0 degree days on average) was lower than in normal years (not statistically significant) for most species. In cold years, the ancient group delayed by more in first leaf date and showed smaller changes in thermal requirement than the young group, but there was no significant difference in warm years.There were no significant differences in changes of first leaf date and thermal requirement among different life forms. The high temperature in the previous winter caused plants to receive less chilling, and thus reduced the thermal requirement in the following year. The first leaf date of woody plants simulated by the growing degree day model was 4.1 days earlier than the observed date in warm years, and was 3.0 days later than the observed date in cold years. Therefore, when predicting the future phenological changes, it is necessary to consider changes in the thermal requirement under exceptional climatic conditions; otherwise, it will overestimate the promotion effects of climate warming on the leaf unfolding date.
Keywords:
PDF (1285KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
王焕炯, 陶泽兴, 葛全胜. 气候波动对西安39种木本植物展叶始期及其积温需求的影响. 植物生态学报, 2019, 43(10): 877-888. DOI: 10.17521/cjpe.2019.0178
WANG Huan-Jiong, TAO Ze-Xing, GE Quan-Sheng.
1951到2012年, 全球地表平均温度以每10年0.12 ℃的速度升高了0.73 ℃, 这与人类活动造成的温室气体浓度增加密切相关(IPCC, 2013)。气候变暖改变了异常天气和气候事件发生的频次和强度(Stott, 2016)。例如, 中国地区热浪发生的频次和强度在过去50年里均有增加(Wang et al., 2012)。南美洲的夏季日数在1950-2010年时段内显著增加, 而寒冷日数则降低(de los Milagros Skansi et al., 2013)。这类高温事件出现的频次在未来百年可能继续增加(Chen & Sun, 2015)。异常的高温、干旱或霜冻等气候事件对陆地生态系统产生了显著影响, 例如生产力下降(Frank et al., 2015)、群落组成改变和生物多样性降低(Smith, 2011)等。
植物物候是反映气候变化对生态系统影响的重要指标。过去几十年, 北半球温带大部分地区的植物春季物候期都出现了提前的趋势(Gonsamo et al., 2013; Ge et al., 2015; Templ et al., 2017)。尤其是展叶始期的提前, 改变了生态系统的碳循环(Xia et al., 2015)和水分利用效率(Jin et al., 2017)。现有研究多利用长时间物候观测序列, 从年际变化的角度揭示物候与气候因子之间的关系(Bolmgren et al., 2013; 黄文婕等, 2017; Wang et al., 2019)。近年来, 部分研究开始注意到异常气候事件对植物物候的影响。例如, 与2009年正常年份相比, 2016年春季暖旱时北京地区荚蒾属(Viburnum)植物的花期整体提前(许聪聪等, 2017)。美国马萨诸塞州和威斯康星州2012年的暖春使55种植物出现了百年历史上最早的花期(Ellwood et al., 2013)。美国伊利诺伊州莫顿植物园478种植物在2012年的展叶始期比正常年平均提前了31天(Fahey, 2016)。由此可见, 当前异常气候事件对物候的影响研究只集中在少数几个地区, 亟需开展多个物种物候对异常气候事件的响应差异研究。
在北半球温带地区, 温度是驱动木本植物展叶始期变化的最主要因子(Dai et al., 2014; Jochner et al., 2016; 周彤等, 2018)。木本植物在秋季短日照和低温的影响下停止生长, 形成冬芽, 进入自然休眠期(Anderson et al., 2010)。芽需要一段时间的冷激处理才能打破自然休眠, 进入生态休眠期(Lang et al., 1987)。在生态休眠期, 只要温度回升到适合生长的温度, 植物就恢复生长。在中高纬度地区, 由于冬季气温足够低, 大多数情况下都能够满足打破自然休眠的冷激需求(Chmielewski & G?tz, 2017), 植物的展叶始期主要取决于春季的热量需求(Chuine et al., 2013)。春季的热量需求主要通过积温来衡量。积温模型又称生长度日(GDD)模型, 被广泛地用于预测木本植物的春季物候期(Hunter & Lechowicz, 1992;仲舒颖等, 2017)。该模型假设当春季积温达到某一固定阈值时, 植物开始展叶或开花, 且这一阈值在年与年之间是固定的(Clark et al., 2014)。但是, 有研究开始注意到, 部分植物开花的积温需求在冷年或暖年会发生改变(Carter et al., 2017; 许聪聪等, 2017)。如果积温需求是不稳定的, 在利用GDD模型预测未来气候变化背景下的物候变化时会出现较大误差。因此有必要深入研究在气候波动下展叶始期的积温需求变化。
本研究利用中国物候观测网西安站的气象数据划分了偏冷年和偏暖年, 并对比了39种木本植物的展叶始期及其积温需求在气候异常年份与正常年份的差异。本文拟解决的科学问题是: (1)气候波动对木本植物展叶始期及其积温需求会产生怎样的影响; (2)异常气候条件下积温模型对展叶始期的预测能力是否发生变化。
1 材料和方法
1.1 物候与气象数据
西安位于渭河流域中部关中盆地, 属于典型的暖温带半湿润大陆性季风气候, 年平均气温14.3 ℃, 年降水量557 mm (1981-2010年平均值)。冬季寒冷而少雨雪, 其中1月气温最低, 降水最少, 多年平均气温只有0.3 ℃, 降水量只有6.5 mm; 夏季炎热多雨, 其中7月气温最高, 降水最多, 多年平均气温27.1 ℃, 降水量101.7 mm。过去50年来, 西安出现了显著的增温趋势(金丽娜等, 2014)。由于西安已经成为关中平原城市群的中心城市, 城市人口和建成区面积一直增加, 热岛效应使城区的增温趋势强于郊区(刘宇峰等, 2015)。因此, 受热岛效应影响, 西安的增温幅度比自然生态系统未来可能出现的变化更早, 是研究异常气候对植物物候影响的理想地区。本文所用的物候数据来源于中国物候观测网西安站点, 共采用了1963-2018年39种木本植物的展叶始期数据(表1)。依据《中国物候观测方法》(宛敏谓和刘秀珍, 1979), 展叶始期(又称开始展叶期)定义为观测植株上的芽从芽苞中发出卷曲着的或按叶脉褶叠着的小叶, 出现第一批有一、二片的叶片平展时。选定的观测植株均位于陕西省西安植物园内。由于经费短缺等原因, 物候数据存在缺测年份, 尤其在1969-1972和1997-2002年2个时段没有观测数据。为了保证冷暖年和正常年物候对比的准确性, 我们只保留了展叶始期观测年数在30年及以上的物种, 共涉及25科38属的39种植物, 这些物种多年平均的展叶始期在3月15日到4月20日之间。根据《中国植物志》(Wu & Raven, 2013), 确定各植物的生活型, 将这些物种按照生活型分为3组, 包括25种乔木, 13种灌木或小乔木, 1种藤本(表1)。为研究植物系统发育对物候响应的影响, 本研究利用Cam Webb等编写的软件Phylomatic (http://phylodiversity.net/phylomatic/), 基于39种植物的名录生成进化树。由于Phylomatic集成了已有进化树骨架(Zanne et al., 2014), 直接生成了有枝长的进化树。根据枝长获取了各物种的分化时间(表1)。最后, 利用k-means聚类方法对各物种按照分化时间分为古老类群、中间类群和年轻类群。
本研究所用的气象数据来源于中国气象数据网(https://data.cma.cn/)中的中国地面气候资料日值数据集(V3.0)。其中, 使用了西安和泾河两站1963- 2018年日平均、最高和最低气温数据。2005年位于西安市未央路上的西安基准气候站(城区)迁至20 km以外的泾河(郊区)。两站共同观测年份只有2007和2008年, 之后只有泾河站有观测数据。由于西安站距物候观测点(西安植物园)的距离更近(约10 km), 因此本文利用两站的月气温差异, 将泾河各月的日气温均一化到西安(郑景云和卞娟娟, 2012)。由于积温算法需用到逐小时气温数据, 本研究利用已有算法将一天中气温的上升和下降过程用正弦曲线模拟得到西安逐小时气温数据(Chow & Levermore, 2007)。
1.2 研究方法
1.2.1 气候异常年识别和物候变化对比根据以往结果, 西安木本植物的展叶始期主要与3或4月的平均气温显著相关(白洁等, 2010)。因此, 本研究利用3-4月平均气温划分冷暖年。首先, 去掉所有植物都没有物候观测记录的年份, 将1963-2018年间3-4月最冷的6年作为偏冷年, 最暖的6年作为偏暖年。由于展叶始期数据存在缺测年份, 只有划分6个冷年或暖年才能保证计算异常气候年份平均展叶始期时至少有2个样本。从3-4月平均气温的变化趋势来看, 研究区呈每10年0.86 ℃的显著增温趋势(图1A), 因此划分出的偏冷年(1963、1965、1976、1979、1988和1996年)主要分布在早期, 划分出的偏暖年(2004、2006、2013、2014、2016和2018年)主要分布在近10年, 其余年份为正常年。对比3-4月平均气温发现, 偏暖年比正常年气温高3.15 ℃, 偏冷年比正常年气温低2.24 ℃ (图1B)。假设3-4月的平均气温符合正态分布, 本文划分出的偏暖年和偏冷年出现概率分别为9.3%和14.4%。
在划分出气候异常年后, 对每种植物分别计算偏暖年、偏冷年和正常年的展叶始期平均值, 并利用双样本t检验检测冷暖年份的展叶始期是否与正常年有显著差异。由于异常气候年的样本量远小于正常年, 因此在检验时假设2组样本的方差不相等。另外, 利用方差分析确定冷暖年的物候变化是否在生活型和类群间有显著差异。
Table 1
表1
表1西安39种木本植物及展叶始期
Table 1
| 编号 No. | 物种 Species | 生活型 Life form | 观测年数 N | 分化时间(百万年) Differentiation time (Ma) | 展叶始期(月-日) First leaf date (month-day) |
|---|---|---|---|---|---|
| 1 | 垂柳 Salix babylonica | 乔木 Tree | 42 | 32.4a | 03-15 |
| 2 | 牡丹 Paeonia suffruticosa | 灌木 Shrub | 42 | 115.3b | 03-18 |
| 3 | 木瓜 Chaenomeles sinensis | 灌木或小乔木 Shrub or small tree | 34 | 3.0a | 03-19 |
| 4 | 紫丁香 Syringa oblata | 灌木或小乔木 Shrub or small tree | 42 | 11.2a | 03-19 |
| 5 | 山桃 Amygdalus davidiana | 乔木 Tree | 41 | 82.2b | 03-22 |
| 6 | 杜梨 Pyrus betulifolia | 乔木 Tree | 32 | 3.0a | 03-24 |
| 7 | 连翘 Forsythia suspensa | 灌木 Shrub | 31 | 15.2a | 03-25 |
| 8 | 毛樱桃 Cerasus tomentosa | 灌木 Shrub | 32 | 41.1a | 03-25 |
| 9 | 迎春花 Jasminum nudiflorum | 灌木 Shrub | 38 | 15.2a | 03-25 |
| 10 | 枫杨 Pterocarya stenoptera | 乔木 Tree | 32 | 12.1a | 03-26 |
| 11 | 灯台树 Cornus controversa | 乔木 Tree | 32 | 105.6b | 03-29 |
| 12 | 榛 Corylus heterophylla | 灌木或小乔木 Shrub or small tree | 32 | 49.3a | 03-29 |
| 13 | 蜡梅 Chimonanthus praecox | 灌木 Shrub | 32 | 120.6b | 03-30 |
| 14 | 水杉 Metasequoia glyptostroboides | 乔木 Tree | 32 | 290.0c | 04-01 |
| 15 | 胡桃 Juglans regia | 乔木 Tree | 38 | 12.1a | 04-01 |
| 16 | 栾树 Koelreuteria paniculata | 乔木 Tree | 40 | 46.1a | 04-02 |
| 17 | 紫荆 Cercis chinensis | 灌木 Shrub | 42 | 69.2b | 04-02 |
| 18 | 日本樱花 Cerasus yedoensis | 乔木 Tree | 38 | 41.1a | 04-02 |
| 19 | 玉兰 Yulania denudate | 乔木 Tree | 41 | 120.6b | 04-03 |
| 20 | 银杏 Ginkgo biloba | 乔木 Tree | 32 | 290.0c | 04-04 |
| 21 | 色木槭 Acer pictum subsp. mono | 乔木 Tree | 42 | 46.1a | 04-04 |
| 22 | 枸橘 Poncirus trifoliata | 小乔木 Small tree | 34 | 49.9a | 04-05 |
| 23 | 悬铃木 Platanus orientalis | 乔木 Tree | 37 | 136.9b | 04-05 |
| 24 | 柿 Diospyros kaki | 乔木 Tree | 39 | 105.6b | 04-06 |
| 25 | 毛白杨 Populus tomentosa | 乔木 Tree | 39 | 32.4a | 04-07 |
| 26 | 女贞 Ligustrum lucidum | 乔木 Tree | 31 | 11.2a | 04-07 |
| 27 | 紫藤 Wisteria sinensis | 藤本 Liana | 40 | 36.1a | 04-07 |
| 28 | 刺槐 Robinia pseudoacacia | 乔木 Tree | 41 | 36.1a | 04-07 |
| 29 | 文冠果 Xanthoceras sorbifolium | 灌木或小乔木 Shrub or small tree | 30 | 46.7a | 04-08 |
| 30 | 桑 Morus alba | 乔木 Tree | 42 | 54.8a | 04-08 |
| 31 | 白蜡树 Fraxinus chinensis | 乔木 Tree | 34 | 21.7a | 04-09 |
| 32 | 构树 Broussonetia papyrifera | 乔木 Tree | 30 | 54.8a | 04-09 |
| 33 | 臭椿 Ailanthus altissima | 乔木 Tree | 42 | 49.9a | 04-09 |
| 34 | 槐 Sophora japonica | 乔木 Tree | 37 | 53.9a | 04-09 |
| 35 | 木槿 Hibiscus syriacus | 灌木 Shrub | 36 | 69.2b | 04-12 |
| 36 | 黄连木 Pistacia chinensis | 乔木 Tree | 31 | 70.9b | 04-14 |
| 37 | 紫薇 Lagerstroemia indica | 灌木或小乔木 Shrub or small tree | 40 | 111.7b | 04-14 |
| 38 | 乌桕 Sapium sebiferum | 乔木 Tree | 32 | 100.6b | 04-18 |
| 39 | 梧桐 Firmiana simplex | 乔木 Tree | 30 | 69.2b | 04-20 |
新窗口打开|下载CSV
1.2.2 积温需求对比
本研究共采用了3种积温计算方法。第一种算法是目前最常用的算法, 即将积温计算为高于一定阈值的逐日温度累加和(Hunter & Lechowicz, 1992)。具体公式为:
图1
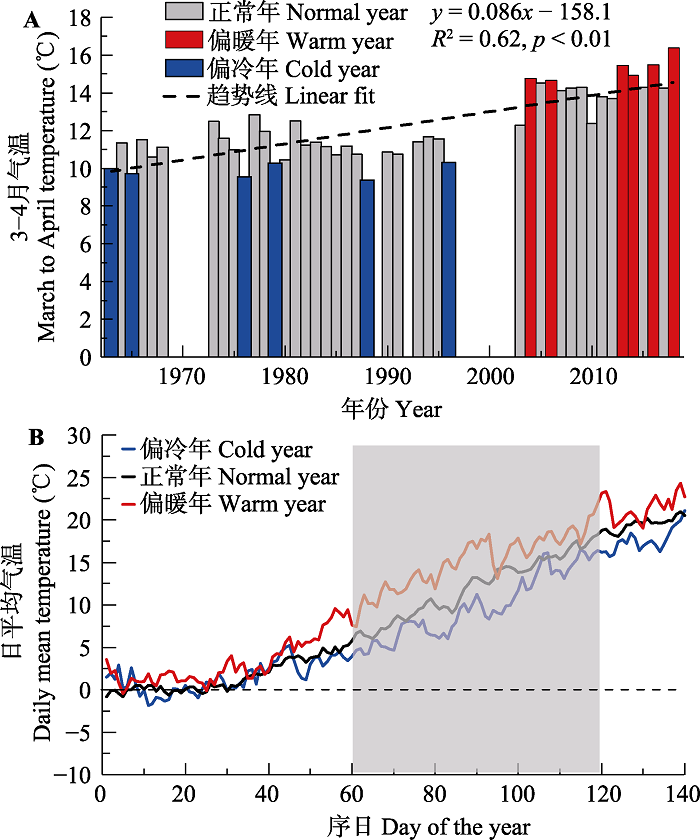 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1西安地区偏冷年、偏暖年和气候正常年的划分。A, 1963-2018年3-4月平均气温逐年变化。B, 异常气候年和正常年的逐日平均气温对比。
Fig. 1Division of the cold years, warm years, and normal years. A, The March to April average temperature from 1963 to 2018. B, The daily mean temperature in years with abnormal and normal climate.
式中GDD为第一种算法计算得到的积温。t1代表展叶始期, t0为积温开始累积的日期, 本文设为1月1日。x(t)为第t天的日平均气温。Tb为阈值或基础温度, 根据以往研究设为5 ℃ (Cannell & Smith, 1983), 以便于不同物种积温需求间的对比。
上述计算积温的方法假设发育速率与温度呈线性关系, 有实验证据表明发育速率与温度的关系是非线性的(Sarvas, 1972)。因此, 许多研究采取第二种方法计算积温(H?nninen, 1990):
式中GDDS为第二种算法计算得到的积温。其他参数与公式(1)一致。只有x(t)大于0 ℃的时候才累积积温。
第三种计算积温的方法是生长度时。与其他两种方法不同, 该方法根据逐小时的气温计算积温。另外, 该方法假设存在发育速率的下限温度、上限温度和最适温度, 高于或低于最适温度时, 发育速率较低。具体公式(Anderson et al., 1986; Luedeling et al., 2009)为:
式中GDH为生长度时; TL为发育的下限温度; Tu为发育的最适温度; Tc为发育的上限温度。根据Anderson等(1986), TL、Tu和Tc分别设置为4、25和36 ℃。F指其他因子(水分等)对发育速度的限制作用。由于西安植物园在植物缺水时会进行灌溉处理, 因此可认为本研究中的植物物候不受水分条件的限制, 因此将F设为1。Th代表一天内第h小时的气温。
利用公式(3), 首先计算GDH的日累积量。将日累积量除以24 h可将度时的单位转换到度日。最后再计算GDH的逐日累积和。具体公式为:
式中GDDH为第三种算法计算得到的积温。t0与前两种算法一样均设置为1月1日; t(1)指第t天第1小时的平均气温, t(2)指第t天第2小时的平均气温, 其他时刻的气温与此类似。
对于每个物种每年的展叶始期, 分别利用上述3种方法计算展叶始期的积温需求。其次, 对每种植物, 分别计算偏暖年、偏冷年和正常年的展叶始期积温需求均值, 并利用t检验检测异常气候年的展叶始期积温需求是否与正常年有显著差异。另外, 利用方差分析确定冷暖年的积温需求变化是否在生活型和类群间有显著差异。
1.2.3 积温模型的建立及模拟效果检验
本研究利用正常年的积温需求建立物候模型, 检验该模型是否能准确地模拟气候异常年的展叶始期。由于3种算法计算得到的积温需求显著相关, 因此本研究只采用以第一种积温算法为基础的GDD模型建模。在建模过程中, 与计算积温需求时不同, 不再将3个参数(t0、Tb和积温需求阈值GDDb)视为固定值, 而是通过拟合的方式确定3个参数。拟合时输入数据为正常年的展叶始期和日平均气温, 具体方法为: (1)设定t0取值范围在1月1日和1月31日之间(以1天为步长), Tb在0 ℃和10 ℃之间(以1 ℃为步长); (2)以每组t0和Tb为输入参数, 计算所有正常年展叶始期的积温需求; (3)将所有年积温需求的平均值作为GDDb; (3)以每组t0、Tb和GDDb为输入参数, 模拟每一年的展叶始期, 并计算模拟值和观测值的均方根误差(RMSE)。RMSE最小的那组参数作为选定的最优参数。
在确定各物种的模型参数后, 分别模拟出各物种对应年份的展叶始期。利用正常年的观测值与模拟值, 计算RMSE和R2, 用以检验模型模拟效果。为计算以正常年数据拟合的物候模型能否准确模拟气候异常年的展叶始期, 分别计算偏暖年和偏冷年展叶始期模拟值与观测值的误差, 并用单样本t检验计算误差的平均值是否显著不等于0。
2 结果
2.1 气候异常年展叶始期的变化
对西安的39种植物, 偏暖年的展叶始期均早于正常年(图2A), 所有物种平均提前8.6天。不同物种在偏暖年展叶始期的提前天数存在差异, 范围在1.9天(槐(Sophora japonica))到13.1天(乌桕(Sapium sebiferum))之间(图2B)。有33个物种在偏暖年的展叶始期均值与正常年存在显著差异(p < 0.05), 占研究物种总数的84.6%。在偏冷年, 各植物的展叶始期均晚于正常年, 所有物种平均推迟8.2天(图2A)。在偏冷年展叶始期推迟最少的物种是牡丹(Paeonia suffruti cosa)仅0.9天, 推迟最多的物种是银杏(Ginkgo biloba), 达15.4天。有16个物种(41.0%)在偏冷年的展叶始期与正常年存在显著差异。因此, 异常气候尤其是 春季偏暖事件对大多数物种的展叶始期产生了显著影响。图2
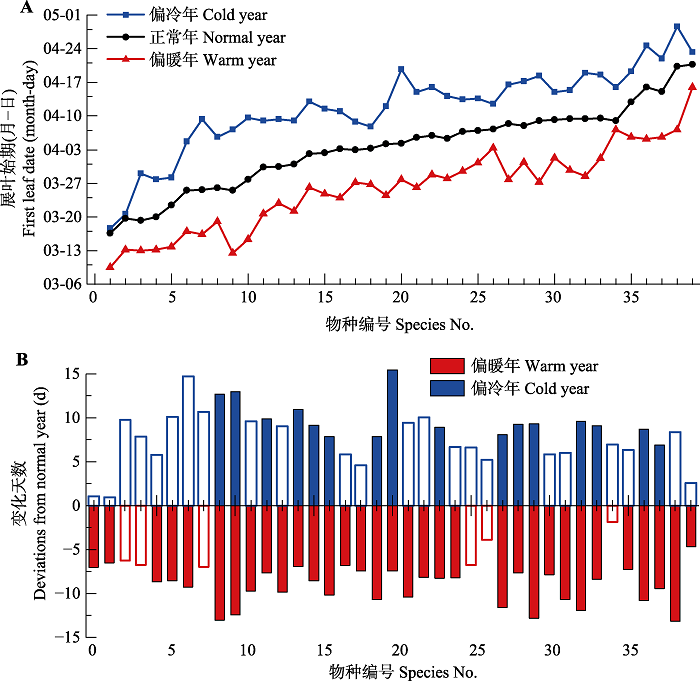 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2西安地区异常气候年和正常年39种木本植物的展叶始期对比。A, 各物种在偏冷年、正常年和偏暖年的平均展叶始期。B, 异常气候年与正常年展叶始期的差异。实心柱代表差异显著(p < 0.05)。物种编号同
Fig. 2Difference in first leaf date (FLD) of 39 woody species between the years with abnormal and normal climate in Xi?an. A, The average FLD in cold years, normal years, and warm years for each species. B, The difference in FLD between the years with abnormal and normal climate. The solid bars represent that the difference is significant (p < 0.05). See
方差分析显示, 不同类群在偏冷年的展叶始期变化存在显著差异, 具体体现在古老类群偏冷年推迟的天数更多, 但在偏暖年不同类群间的物候变化无明显差异(表2)。无论是在偏冷年或偏暖年, 不同生活型间的展叶始期变化无显著差异。
2.2 气候异常年的积温需求变化
图3显示了3种算法计算的积温需求在冷暖年与正常年的差异。总体来看, 无论哪种算法, 所有物种在偏暖年比正常年都需要更多的积温展叶。例如, 对于算法1, 所有物种在偏暖年展叶始期的平均积温需求是257.5度日, 而在正常年的均值只有195.1度日, 其他算法的结果也与此类似。值得注意的是, t检验结果表明, 大多数物种在偏暖年的平均积温需求与正常年存在显著差异。对算法1, 显著差异的物种占82.1%, 而算法2和算法3显著差异的物种占比稍低, 分别为64.1%和56.4%。3种算法都表明, 偏冷年展叶始期的积温需求低于正常年(图3)。例如, 对于算法1, 所有物种在偏冷年展叶始期的平均积温需求是168.0度日, 小于正常年的均值(195.1度日), 但是在偏冷年的降幅(27.1度日)明显低于偏暖年的增幅(62.4度日)。虽然对大多数物种, 展叶始期的积温需求在偏冷年比正常年更小, 但t检验结果表明, 只有在算法1下3个物种表现出显著差异(图3A)。
对3种算法的对比(图4)表明, 算法间计算得到的积温需求只有数量上的差异, 而两两间呈显著线性正相关关系(R2 > 0.99)。因此在后续的建模与分析中, 以最为常用的算法1结果为准。通过方差分析对比不同类群在冷暖年的积温需求变化, 发现古老类群在偏冷年的积温需求变化弱于中间和年轻类群(其中与中间类群的差异显著), 但在偏暖年三者之间无显著差异。无论是在偏冷年或偏暖年, 不同生活型间的积温需求变化无显著差异。
2.3 模型建立以及在气候异常年的模拟效果
利用正常年的数据, 对每一种植物建立了展叶始期积温模型(图5A)。结果表明, 该模型能够较准确模拟展叶始期的年际变化。方差解释量(R2)的均值达到了0.34, 且显著的占比达79.5% (p < 0.05)。RMSE的范围在3.6天(玉兰(Yulania denudate))到9.5天(槐(Sophora japonica))之间, 平均值只有5.6天。为对比该模型在异常气候年的模拟偏差, 本文计算了气候异常年与正常年模拟误差(模拟值减观测值)的均值(图4B)。结果表明, 在正常年, 模型的平均误差在0.4天到1.9天之间, 且没有任何物种的平均误差与0有显著差异。但在偏暖年, 模型模拟的展叶始期在大多数情况下(37个物种, 占94.9%)都早于观测值, 误差在-11.6天到1.3天之间, 平均误差为-4.1天。另外, 有18个物种的平均误差与0差异显著(p < 0.05), 表明展叶始期在偏暖年的预测值比观测值显著偏早, 模型高估了气候变暖对展叶始期的提前效应。展叶始期在偏冷年的模拟值在32个物种(82.1%)中都晚于观测值, 误差在-4.7天到11.0天之间, 平均误差为3.0天。但只有2个物种的平均误差与0差异显著(p < 0.05)。因此, 展叶始期在偏冷年的 预测值在多数情况下比观测值偏晚, 但差异并不显著。
Table 2
表2
表2西安39种木本植物不同类群和生活型的展叶始期及其积温需求变化对比(平均值±标准偏差)
Table 2
| 变量 Variable | 类群 Group | 生活型 Life form | ||||
|---|---|---|---|---|---|---|
| 年轻 Young | 中间 Intermediate | 古老 Ancient | 乔木 Tree | 灌木或小乔木 Shrub or small tree | 藤本 Liana | |
| 物种数量 N | 24 | 13 | 2 | 25 | 13 | 1 |
| 偏冷年物候变化(天) PC in the cold year (day) | 8.6 ± 2.9a | 6.7 ± 2.6b | 13.2 ± 3.2a,b | 7.9 ± 3.0 | 8.8 ± 3.4 | 8.1 |
| 偏暖年物候变化(天) PC in the warm year (day) | -8.6 ± 2.7 | -8.8 ± 2.2 | -7.2 ± 0.4 | -8.5 ± 2.6 | -8.5 ± 2.3 | -11.6 |
| 偏冷年积温需求变化(度日) CTR in the cold year (degree day) | -26.2 ± 18.7 | -32.9 ± 16.4a | 0.7 ± 35.8a | -29.1 ± 21.5 | -23.2 ± 16.2 | -25.6 |
| 偏暖年积温需求变化(度日) CTR in the warm year (degree day) | 62.2 ± 31.8 | 61.5 ± 26.7 | 70.7 ± 6.1 | 67.7 ± 32.6 | 54.3 ± 18.8 | 36.4 |
新窗口打开|下载CSV
图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3西安地区气候异常年和正常年39种木本植物展叶始期的积温需求对比。A, 算法1结果。B, 算法2结果。C, 算法3结果。实心点代表异常气候年与正常年差异显著(p < 0.05)。
Fig. 3Comparison of thermal requirements for the first leaf date (FLD) of 39 woody species between the years with abnormal and normal climate in Xi?an. A, The results of the first method. B, The results of the second method. C, The results of the third method. The solid circles represent that the difference is significant (p < 0.05).
图4
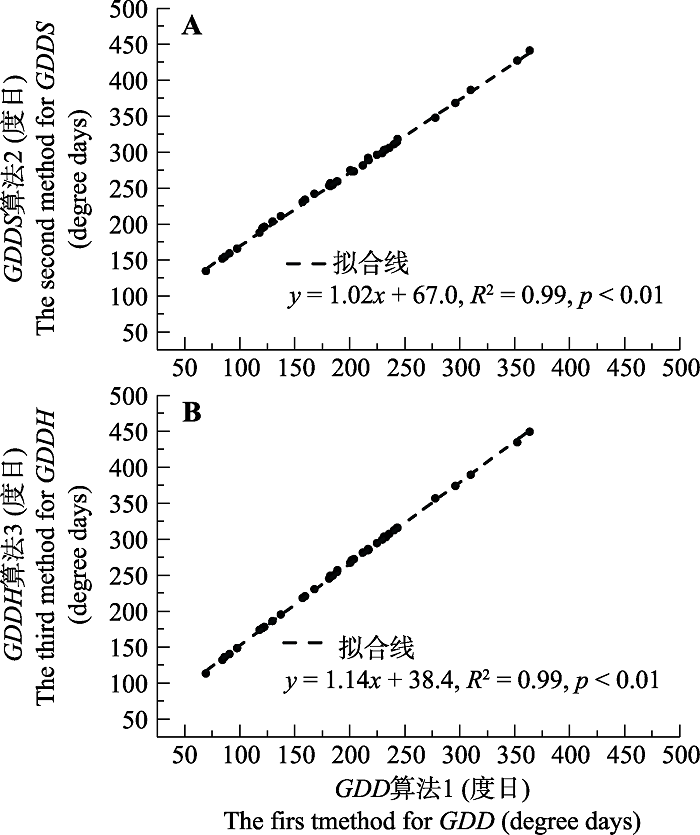 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4不同算法计算得到的西安39种木本植物展叶始期积温需求对比。A, 算法1与算法2。B, 算法1与算法3。黑点表示各物种在正常年的积温需求。算法同
Fig. 4Comparisons among different methods for calculating thermal requirements for the first leaf date (FLD) of 39 woody species in Xi?an. A, method 1 vs. method 2. B, method 1 vs. method 3. Black circles represent the mean thermal requirement in normal years for each species. See
图5
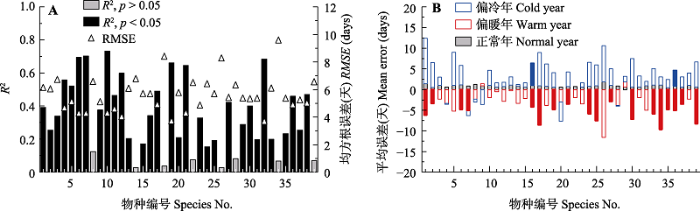 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5西安39种木本植物展叶始期的积温模型模拟效果。A, 模型的拟合优度(R2)和均方根误差(RMSE)。B, 各物种在异常气候年和正常年的模拟误差。实心柱代表异常气候年的误差显著不为0 (p < 0.05)。物种编号同
Fig. 5Effect of growing degree day model for simulating the first leaf date (FLD) of 39 woody species in Xi?an. A, Goodness of fit (R2) and root mean square error (RMSE) of the models. B, Error of the model in simulating the FLD in the years with abnormal and normal climate. The solid bars represent that the mean error was significant from 0 (p < 0.05). See
3 讨论
本研究发现气候波动使植物的春季物候期出现了较大变化, 西安39种植物的展叶始期在偏暖年比偏冷年平均早16.8天之多。该研究结果与其他地区的结果类似。例如, 北京地区部分荚蒾属(Viburnum)植物的始花期在2016年的偏暖年比2010年的偏冷年提前了1个月以上(许聪聪等, 2017)。2010和2012年的暖春使美国东北部落叶阔叶林的展叶始期比正常年提前了2星期(Friedl et al., 2014)。在气候变暖下展叶始期提前越明显的物种, 生长季和光合作用时间延长越多, 能够在群落竞争中占据优势(Cleland et al., 2012)。因此, 展叶始期对气候波动的响应在一定程度上反映了植物对气候变化的适应能力。但是, 气候变暖下提前的展叶始期并不一定对植物生长永远有利, 在展叶始期之后发生的晚霜冻事件会严重影响植物生长, 例如2011年5月发生在德国的晚霜冻事件使得落叶阔叶林的新叶全部凋落(Menzel et al., 2015)。本研究发现, 在偏冷年, 古老类群(银杏和水杉(Metasequoia glyptostroboides))展叶始期推迟天数明显大于年轻类群。可能是因为古老类群进化出了避免被晚霜冻伤害的适应机制, 以避免在偏冷年的霜冻风险。
本文结果表明偏暖年木本植物展叶的积温需求高于正常年, 在偏冷年的积温需求略低于正常年, 也与其他结果相类似。例如美国白蜡树(Fraxinus americana)在偏暖的2012年, 展叶始期的积温需求显著高于其他年份(Menzel et al., 2015)。偏暖条件积温需求的增加有两个可能的影响因素。第一个原因是冬季冷激的影响。多数情况下, 展叶或开花的积温需求与前一年秋冬季植物受到的冷激程度呈负指数关系。例如, 在美国, 早期的研究发现北美云杉(Picea sitchensis)芽开放所需积温随秋冬季冷激日数(低于5 ℃的日数)的增加而呈指数下降(Cannell & Smith, 1983)。在美国东南部对桃(Prunus persica)的实验表明, 冷激增强会导致花芽开放的积温需求减少(Okie & Blackburn, 2011)。本研究中, 偏暖年比正常年的秋冬季气温(前一年11月到当年2月)高1.3 ℃, 而偏冷年比正常年秋冬季平均气温仅低0.3 ℃。这是大多数物种在偏暖年的积温需求显著大于正常年, 而偏冷年小于正常年的原因。另一种可能的原因是光周期对展叶始期存在影响(Way & Montgomery, 2015)。对光周期响应敏感的植物, 虽然计算出的偏暖年积温需求增加, 但增加的积温被当时较低的昼长所抵消, 而在偏冷年, 植物展叶始期推后, 临近展叶时的昼长较长, 促进了积温对生长发育的影响, 从而使计算出的积温需求降低。在两种原因中, 冷激的影响可能更大, 因为Laube等(2014)的控制实验结果表明, 冷激增强使积温需求减少,促进了36种木本植物发芽, 但长的日照时间只使约1/3的植物芽开放期出现轻微提前。
目前, 很多研究均采用常规的GDD模型预测木本植物的春季物候期(张爱英等, 2014; 徐韵佳等, 2017)。常规的GDD模型在检验时是所有输入数据一起检验, 没有考虑在异常气候条件下的模型预测效果。在本研究中, 以正常年物候数据拟合的积温模型对偏暖年展叶始期的预测结果显著早于观测到的日期。这主要是因为在偏暖年的积温需求显著高于正常年, 因此常规的物候模型高估了高温对展叶始期的提前作用。特别是在当前的陆地生物圈模型中, 仍以传统的GDD模型为主预测植被生长季开始期(Richardson et al., 2012), 在未来气候进一步变暖的情景下, 模型对物候乃至植被生长力的预测结果可能会产生较大误差。当前已有部分物候模型考虑了冬季冷激对积温需求的影响(仲舒颖等, 2017; Zhang et al., 2018), 建议在下一代的陆地生物圈模型中采用考虑积温需求变化的物候模型。
4 结论
为研究气候波动对植物展叶始期及其积温需求的影响, 本研究利用1963-2018年西安39种木本植物的展叶始期数据, 采用多种方法量化了展叶始期的积温需求; 并根据3-4月平均气温划分了偏冷年、偏暖年与正常年, 对比了气候异常年与正常年的展叶始期及其积温需求, 主要得到以下结论:(1)所有木本植物的展叶始期在偏暖年比正常年提前, 平均达8.6天; 在偏冷年比正常年推迟, 平均推迟8.2天。提前或推迟的幅度在物种间不同, 但对大多数物种, 偏暖年展叶始期与正常年的差异显著。古老类群在偏冷年展叶始期的推迟天数更多。
(2)在偏暖年, 大多数物种展叶始期的积温需求高于正常年, 且差异显著; 大多数物种在偏冷年的积温需求低于正常年, 但差异不显著。可能原因是在偏暖年冬季气温较高使植物在前一年冬季受到的冷激程度减轻, 限制了植物在春季的芽开放, 增加了积温需求。
(3) GDD模型模拟的木本植物展叶始期在偏暖年比观测值平均早4.1天。这主要是因为植物在偏暖年的积温需求更高, 而常规的积温模型假设积温需求在年际间保持不变。IPCC报告预测全球气温在21世纪仍将持续升高, 高温事件的发生频次会持续增加(IPCC, 2013), 因此在预测未来物候变化时, 需要考虑在春季偏暖情形下的积温需求变化, 否则会高估气候变暖对展叶始期的影响。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 2]
[本文引用: 1]
DOI:10.3773/j.issn.1005-264x.2010.11.004URL [本文引用: 1]

根据1963–2007年中国物候观测网西安观测站的物候和气温、降水资料,分析了西安站34种木本植物春季展叶始期、展叶盛期、始花期和盛花期等4个关键物候期的变化趋势、对气候变化的阶段响应特点及其与气温、降水变化的关系。结果表明,1963年以来,西安地区气温呈显著上升趋势,特别是1994年前后,气温发生明显突变,上升趋势更加明显;西安春季物候变化主要呈现提前趋势。在45年中,观测到的34种植物的展叶始期平均提前1天,展叶盛期平均提前1.4天,始花期平均提前9天,盛花期平均提前12天;以突变点为界,34个物种1995–2007年的4种物候期比1963–1994年平均提前了4.34±0.77天;春季物候期的早晚主要受春季气温的影响,特别是春季物候期发生当月和上一月的平均气温对物候期的影响最为显著。叶物候和物候发生期前一月的降水量有较为明显的相关关系,花物候期和降水的关系不明显。
DOI:10.3773/j.issn.1005-264x.2010.11.004URL [本文引用: 1]
根据1963–2007年中国物候观测网西安观测站的物候和气温、降水资料,分析了西安站34种木本植物春季展叶始期、展叶盛期、始花期和盛花期等4个关键物候期的变化趋势、对气候变化的阶段响应特点及其与气温、降水变化的关系。结果表明,1963年以来,西安地区气温呈显著上升趋势,特别是1994年前后,气温发生明显突变,上升趋势更加明显;西安春季物候变化主要呈现提前趋势。在45年中,观测到的34种植物的展叶始期平均提前1天,展叶盛期平均提前1.4天,始花期平均提前9天,盛花期平均提前12天;以突变点为界,34个物种1995–2007年的4种物候期比1963–1994年平均提前了4.34±0.77天;春季物候期的早晚主要受春季气温的影响,特别是春季物候期发生当月和上一月的平均气温对物候期的影响最为显著。叶物候和物候发生期前一月的降水量有较为明显的相关关系,花物候期和降水的关系不明显。
DOI:10.1007/s00484-012-0560-8URL [本文引用: 1]
Phenological shifts linked to global warming reflect the ability of organisms to track changing climatic conditions. However, different organisms track global warming differently and there is an increasing interest in the link between phenological traits and plant abundance and distribution. Long-term data sets are often used to estimate phenological traits to climate change, but so far little has been done to evaluate the quality of these estimates. Here, we use a 73-year long data series of first flowering dates for 25 species from north-temperate Sweden to evaluate (i) correlations between first flowering dates and year for different time periods and (ii) linear regression models between first flowering date and mean monthly temperatures in preceding months. Furthermore, we evaluate the potential of this kind of data to estimate the phenological temperature sensitivities (i.e. number of days phenological change per degree temperature change, beta(60)) in such models. The sign of the correlations between first flowering dates and year were highly inconsistent among different time periods, highlighting that estimates of phenological change are sensitive to the specific time period used. The first flowering dates of all species were correlated with temperature, but with large differences in both the strength of the response and the period(s) of the year that were most strongly associated with phenological variation. Finally, our analyses indicated that legacy data sets need to be relatively long-term to be useful for estimating phenological temperature sensitivities (beta(60)) for inter-specific comparisons. In 10-year long observation series only one out of 24 species reached a parts per thousand yen80 % probability of estimating temperature sensitivity (beta(60)) within a +/- 1 range, and 17 out of 24 species reached a parts per thousand yen80 % probability when observation series were 20 years or shorter. The standard error for beta(60) ranged from 0.6 to 2.0 for 10-year long observation series, and 19 out of 24 species reached SE < 1 after 15 years. In general, late flowering species will require longer time series than early flowering species.
DOI:10.2307/2403139URL [本文引用: 2]
DOI:10.1007/s00442-017-3838-zURLPMID:28224350 [本文引用: 1]
The frequency of extreme warm years is increasing across the majority of the planet. Shifts in plant phenology in response to extreme years can influence plant survival, productivity, and synchrony with pollinators/herbivores. Despite extensive work on plant phenological responses to climate change, little is known about responses to extreme warm years, particularly at the intraspecific level. Here we investigate 43 populations of white ash trees (Fraxinus americana) from throughout the species range that were all grown in a common garden. We compared the timing of leaf emergence during the warmest year in U.S. history (2012) with relatively non-extreme years. We show that (a) leaf emergence among white ash populations was accelerated by 21 days on average during the extreme warm year of 2012 relative to non-extreme years; (b) rank order for the timing of leaf emergence was maintained among populations across extreme and non-extreme years, with southern populations emerging earlier than northern populations; (c) greater amounts of warming units accumulated prior to leaf emergence during the extreme warm year relative to non-extreme years, and this constrained the potential for even earlier leaf emergence by an average of 9 days among populations; and (d) the extreme warm year reduced the reliability of a relevant phenological model for white ash by producing a consistent bias toward earlier predicted leaf emergence relative to observations. These results demonstrate a critical need to better understand how extreme warm years will impact tree phenology, particularly at the intraspecific level.
DOI:10.1002/joc.2015.35.issue-10URL [本文引用: 1]
[本文引用: 1]
DOI:10.1177/0143624407078642URL [本文引用: 1]
[本文引用: 1]
DOI:10.1111/1365-2435.12309URL [本文引用: 1]
;Increases in primary production may occur if plants respond to climate warming with prolonged growing seasons, but not if local adaptation, cued by photoperiod, limits phenological advance. It has been hypothesized that trees with diffuse-porous xylem anatomy and early successional species may respond most to warming. Within species, northern populations may respond most due to the fact that growing seasons are relatively short. Species most sensitive to spring temperature may show little overall response to warming if reduced chilling in fall/winter offsets accelerated winter/spring development.
Because current thermal models consider only highly aggregated variables, for example degree-days or chilling units (temperature sums for a season or year), they may not accurately represent warming effects. We show that assumptions contained in current thermal (degree-day) models are unrealistic for climate change analysis. Critical threshold parameters are not identifiable, and they do not actually have much to do with thresholds for development. Traditional models further overlook the discrete nature of observations, observation error and the continuous response of phenological development to temperature variation. An alternative continuous development model (CDM) that addresses these problems is applied to a large experimental warming study near northern and southern boundaries of 15 species in the eastern deciduous forest of the USA, in North Carolina and Massachusetts.
Results provide a detailed time course of phenological development, including vernalization during winter and warming in spring, and challenge the basic assumptions of thermal models. Where traditional models find little evidence of a chilling effect (most are insignificant or have the wrong sign), the continuous development model finds evidence of chilling effects in most species.
Contrary to the hypothesis that northern populations respond most, we find southern populations are most responsive. Because northern populations already have a compressed period for spring development, they may lack flexibility to further advance development. A stronger response in the southern range could allow residents to resist northward migration of immigrants as climate warms. If potential invaders fail to exploit a prolonged growing season to the same degree as residents, then there is a resident advantage.
Hypothesized effects of warming for xylem anatomy and successional status are not supported by the 15 species in this study.
URL [本文引用: 1]
Earlier spring phenology observed in many plant species in recent decades provides compelling evidence that species are already responding to the rising global temperatures associated with anthropogenic climate change. There is great variability among species, however, in their phenological sensitivity to temperature. Species that do not phenologically "track" climate change may be at a disadvantage if their growth becomes limited by missed interactions with mutualists, or a shorter growing season relative to earlier-active competitors. Here, we set out to test the hypothesis that phenological sensitivity could be used to predict species performance in a warming climate, by synthesizing results across terrestrial warming experiments. We assembled data for 57 species across 24 studies where flowering or vegetative phenology was matched with a measure of species performance. Performance metrics included biomass, percent cover, number of flowers, or individual growth. We found that species that advanced their phenology with warming also increased their performance, whereas those that did not advance tended to decline in performance with warming. This indicates that species that cannot phenologically "track" climate may be at increased risk with future climate change, and it suggests that phenological monitoring may provide an important tool for setting future conservation priorities.
DOI:10.1007/s00484-013-0679-2URL [本文引用: 1]
Leaf phenology has been shown to be one of the most important indicators of the effects of climate change on biological systems. Few such studies have, however, been published detailing the relationship between phenology and climate change in Asian contexts. With the aim of quantifying species' phenological responsiveness to temperature and deepening understandings of spatial patterns of phenological and climate change in China, this study analyzes the first leaf date (FLD) and the leaf coloring date (LCD) from datasets of four woody plant species, Robinia pseudoacacia, Ulmus pumila, Salix babylonica, and Melia azedarach, collected from 1963 to 2009 at 47 Chinese Phenological Observation Network (CPON) stations spread across China (from 21A degrees to 50A degrees N). The results of this study show that changes in temperatures in the range of 39-43 days preceding the date of FLD of these plants affected annual variations in FLD, while annual variations in temperature in the range of 71-85 days preceding LCD of these plants affected the date of LCD. Average temperature sensitivity of FLD and LCD for these plants was -3.93 to 3.30 days A degrees C-1 and 2.11 to 4.43 days A degrees C-1, respectively. Temperature sensitivity of FLD was found to be stronger at lower latitudes or altitude as well as in more continental climates, while the response of LCD showed no consistent pattern. Within the context of significant warming across China during the study period, FLD was found to have advanced by 5.44 days from 1960 to 2009; over the same period, LCD was found to have been delayed by 4.56 days. These findings indicate that the length of the growing season of the four plant species studied was extended by a total of 10.00 days from 1960 to 2009. They also indicate that phenological response to climate is highly heterogeneous spatially.
DOI:10.1016/j.gloplacha.2012.11.004URL [本文引用: 1]
Here we show and discuss the results of an assessment of changes in both area-averaged and station-based climate extreme indices over South America (SA) for the 1950-2010 and 1969-2009 periods using high-quality daily maximum and minimum temperature and precipitation series. A weeklong regional workshop in Guayaquil (Ecuador) provided the opportunity to extend the current picture of changes in climate extreme indices over SA.
Our results provide evidence of warming and wetting across the whole SA since the mid-20th century onwards. Nighttime (minimum) temperature indices show the largest rates of warming (e.g. for tropical nights, cold and warm nights), while daytime (maximum) temperature indices also point to warming (e.g. for cold days, summer days, the annual lowest daytime temperature), but at lower rates than for minimums. Both tails of night-time temperatures have warmed by a similar magnitude, with cold days (the annual lowest nighttime and daytime temperatures) seeing reductions (increases). Trends are strong and moderate (moderate to weak) for regional-averaged (local) indices, most of them pointing to a less cold SA during the day and warmer night-time temperatures.
Regionally-averaged precipitation indices show clear wetting and a signature of intensified heavy rain events over the eastern part of the continent. The annual amounts of rainfall are rising strongly over south-east SA (26.41 mm/decade) and Amazonia (16.09 mm/decade), but north-east Brazil and the western part of SA have experienced non-significant decreases. Very wet and extremely days, the annual maximum 5-day and 1-day precipitation show the largest upward trends, indicating an intensified rainfall signal for SA, particularly over Amazonia and south-east SA Local trends for precipitation extreme indices are in general less coherent spatially, but with more general spatially coherent upward trends in extremely wet days over all SA. (C) 2012 Elsevier B.V.
DOI:10.1371/journal.pone.0053788URLPMID:23342001 [本文引用: 1]
Flowering times are well-documented indicators of the ecological effects of climate change and are linked to numerous ecosystem processes and trophic interactions. Dozens of studies have shown that flowering times for many spring-flowering plants have become earlier as a result of recent climate change, but it is uncertain if flowering times will continue to advance as temperatures rise. Here, we used long-term flowering records initiated by Henry David Thoreau in 1852 and Aldo Leopold in 1935 to investigate this question. Our analyses demonstrate that record-breaking spring temperatures in 2010 and 2012 in Massachusetts, USA, and 2012 in Wisconsin, USA, resulted in the earliest flowering times in recorded history for dozens of spring-flowering plants of the eastern United States. These dramatic advances in spring flowering were successfully predicted by historical relationships between flowering and spring temperature spanning up to 161 years of ecological change. These results demonstrate that numerous temperate plant species have yet to show obvious signs of physiological constraints on phenological advancement in the face of climate change.
URLPMID:693155 [本文引用: 1]
We conducted a retrospective study by telephone interview (10 to 22 months later) of 26 families who had experienced a perinatal death. Six of 26 mothers had a prolonged grief reaction (12 to 20 months). Those mothers with a surviving twin or subsequent pregnancy less than five months following the death were at higher risk for a prolonged grieving period than were those without subsequent pregnancy or one more than six months later. Half of the families obtained information about the cause of death and risk of recurrence only during hospitalization; subsequent contact, weeks to months later, provided additional information for the other half. Twenty-two of 26 mothers met predetermined criteria for having an adequate understanding of cause of death and risk of recurrence; four of 26 knew neither. Sixty percent of the mothers who had adequate understanding and who had no prolonged grief response felt totally dissatisfied or only partially satisfied with the information they received and the way they received it. Follow-up contact by phone or in person increased understanding significantly; mothers who had had in-person follow-up were more likely to be satisfied with the information they received.
DOI:10.1111/gcb.12916URLPMID:25752680 [本文引用: 1]
Extreme droughts, heat waves, frosts, precipitation, wind storms and other climate extremes may impact the structure, composition and functioning of terrestrial ecosystems, and thus carbon cycling and its feedbacks to the climate system. Yet, the interconnected avenues through which climate extremes drive ecological and physiological processes and alter the carbon balance are poorly understood. Here, we review the literature on carbon cycle relevant responses of ecosystems to extreme climatic events. Given that impacts of climate extremes are considered disturbances, we assume the respective general disturbance-induced mechanisms and processes to also operate in an extreme context. The paucity of well-defined studies currently renders a quantitative meta-analysis impossible, but permits us to develop a deductive framework for identifying the main mechanisms (and coupling thereof) through which climate extremes may act on the carbon cycle. We find that ecosystem responses can exceed the duration of the climate impacts via lagged effects on the carbon cycle. The expected regional impacts of future climate extremes will depend on changes in the probability and severity of their occurrence, on the compound effects and timing of different climate extremes, and on the vulnerability of each land-cover type modulated by management. Although processes and sensitivities differ among biomes, based on expert opinion, we expect forests to exhibit the largest net effect of extremes due to their large carbon pools and fluxes, potentially large indirect and lagged impacts, and long recovery time to regain previous stocks. At the global scale, we presume that droughts have the strongest and most widespread effects on terrestrial carbon cycling. Comparing impacts of climate extremes identified via remote sensing vs. ground-based observational case studies reveals that many regions in the (sub-)tropics are understudied. Hence, regional investigations are needed to allow a global upscaling of the impacts of climate extremes on global carbon-climate feedbacks.
DOI:10.1088/1748-9326/9/5/054006URL [本文引用: 1]
DOI:10.1111/gcb.12648URLPMID:24895088 [本文引用: 1]
The change in the phenology of plants or animals reflects the response of living systems to climate change. Numerous studies have reported a consistent earlier spring phenophases in many parts of middle and high latitudes reflecting increasing temperatures with the exception of China. A systematic analysis of Chinese phenological response could complement the assessment of climate change impact for the whole Northern Hemisphere. Here, we analyze 1263 phenological time series (1960-2011, with 20+ years data) of 112 species extracted from 48 studies across 145 sites in China. Taxonomic groups include trees, shrubs, herbs, birds, amphibians and insects. Results demonstrate that 90.8% of the spring/summer phenophases time series show earlier trends and 69.0% of the autumn phenophases records show later trends. For spring/summer phenophases, the mean advance across all the taxonomic groups was 2.75 days decade(-1) ranging between 2.11 and 6.11 days decade(-1) for insects and amphibians, respectively. Herbs and amphibians show significantly stronger advancement than trees, shrubs and insect. The response of phenophases of different taxonomic groups in autumn is more complex: trees, shrubs, herbs and insects show a delay between 1.93 and 4.84 days decade(-1), while other groups reveal an advancement ranging from 1.10 to 2.11 days decade(-1) . For woody plants (including trees and shrubs), the stronger shifts toward earlier spring/summer were detected from the data series starting from more recent decades (1980s-2000s). The geographic factors (latitude, longitude and altitude) could only explain 9% and 3% of the overall variance in spring/summer and autumn phenological trends, respectively. The rate of change in spring/summer phenophase of woody plants (1960s-2000s) generally matches measured local warming across 49 sites in China (R=-0.33, P&lt;0.05).
DOI:10.1038/srep02239URLPMID:23867863 [本文引用: 1]
The timing of crucial events in plant life cycles is shifting in response to climate change. We use phenology records from PlantWatch Canada 'Citizen Science' networks to study recent rapid shifts of flowering phenology and its relationship with climate. The average first flower bloom day of 19 Canadian plant species has advanced by about 9 days during 2001-2012. 73% of the rapid and unprecedented first bloom day advances are explained by changes in mean annual national temperature, allowing the reconstruction of historic flower phenology records starting from 1948. The overall trends show that plant flowering in Canada is advancing by about 9 days per °C. This analysis reveals the strongest biological signal yet of climate warming in Canada. This finding has broad implications for niche differentiation among coexisting species, competitive interactions between species, and the asynchrony between plants and the organisms they interact with.
[本文引用: 1]
DOI:10.18306/dlkxjz.2017.08.010URL [本文引用: 1]
Temperature sensitivity of phenophases can reflect how and to what degree plants could tract climate change, and is related to the ability of plants to adapt to climate change. Investigating the temperature sensitivity of phenophases of different plant species could help us to identify species that are sensitive to climate change. To date, the studies about temperature sensitivity of first flowering date (FFD) mainly focused on the temperate area, and fewer studies focused on the subtropical area. We selected Guiyang City, located in subtropical China, as the study area, and analyzed the temporal changes in FFD of plants and their temperature sensitivities based on phenological observation data of 60 typical woody plants from 1980 to 2014. In addition, we evaluated the impact of the length of time series on the stability of the estimates of temperature sensitivity. The results show that: (1) Guiyang City experienced notable climate change with significantly increased annual mean temperature during the study period. The warming of spring and autumn was stronger than summer and winter. (2) FFD of 53 species (88.3%) advanced during the study period with 13 species (21.7%) significantly advancing (P<0.05). Most trends of FFD were between -4 and -2 d/decade. The overall advancing trend for FFD of 60 species was 2.89 d/decade. (3) FFD was significantly and negatively correlated with mean temperature during the optimum period for most species (88.3%). Most temperature sensitivities of FFD ranged from -8 to -4 d/°C. The overall temperature sensitivity for FFD of all species was -5.75 d/°C. (4) Sample size clearly affected stability of the estimates of temperature sensitivity. The time series of 15 years could make the difference of estimates less than 2 d/°C with a probability of 99%. Thus, time series should be as long as possible to be used in estimating the temperature sensitivity of flowering phenology with a stability that is sufficient for interspecific comparisons.
DOI:10.18306/dlkxjz.2017.08.010URL [本文引用: 1]
Temperature sensitivity of phenophases can reflect how and to what degree plants could tract climate change, and is related to the ability of plants to adapt to climate change. Investigating the temperature sensitivity of phenophases of different plant species could help us to identify species that are sensitive to climate change. To date, the studies about temperature sensitivity of first flowering date (FFD) mainly focused on the temperate area, and fewer studies focused on the subtropical area. We selected Guiyang City, located in subtropical China, as the study area, and analyzed the temporal changes in FFD of plants and their temperature sensitivities based on phenological observation data of 60 typical woody plants from 1980 to 2014. In addition, we evaluated the impact of the length of time series on the stability of the estimates of temperature sensitivity. The results show that: (1) Guiyang City experienced notable climate change with significantly increased annual mean temperature during the study period. The warming of spring and autumn was stronger than summer and winter. (2) FFD of 53 species (88.3%) advanced during the study period with 13 species (21.7%) significantly advancing (P<0.05). Most trends of FFD were between -4 and -2 d/decade. The overall advancing trend for FFD of 60 species was 2.89 d/decade. (3) FFD was significantly and negatively correlated with mean temperature during the optimum period for most species (88.3%). Most temperature sensitivities of FFD ranged from -8 to -4 d/°C. The overall temperature sensitivity for FFD of all species was -5.75 d/°C. (4) Sample size clearly affected stability of the estimates of temperature sensitivity. The time series of 15 years could make the difference of estimates less than 2 d/°C with a probability of 99%. Thus, time series should be as long as possible to be used in estimating the temperature sensitivity of flowering phenology with a stability that is sufficient for interspecific comparisons.
DOI:10.1006/jtbi.2000.2178URLPMID:11082304 [本文引用: 2]
Accurate plant phenology (seasonal plant activity driven by environmental factors) models are vital tools for ecosystem simulation models and for predicting the response of ecosystems to climate change. Since the early 1970s, efforts have concentrated on predicting phenology of the temperate and boreal forests because they represent one-third of the carbon captured in plant ecosystems and they are the principal ecosystems with seasonal patterns of growth on Earth (one-fifth of the plant ecosystems area). Numerous phenological models have been developed to predict the growth timing of temperate or boreal trees. They are in general empirical, nonlinear and non-nested. For these reasons they are particularly difficult to fit, to test and to compare with each other. The methodological difficulties as well as the diversity of models used have greatly slowed down their improvement. The aim of this study was to show that the most widely used models simulating vegetative or reproductive phenology of trees are particular cases of a more general model. This unified model has three main advantages. First, it allows for a direct estimation of (i) the response of bud growth to either chilling or forcing temperatures and (ii) the periods when these temperatures affect the bud growth. Second, it can be simplified according to standard statistical tests for any particular species. Third, it provides a standardized framework for phenological models, which is essential for comparative studies as well as for robust model identification.
[本文引用: 2]
DOI:10.3390/rs9070664URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s00484-016-1146-7URLPMID:26942933 [本文引用: 1]
Over a large temperature range, the statistical association between spring phenology and temperature is often regarded and treated as a linear function. There are suggestions that a sigmoidal relationship with definite upper and lower limits to leaf unfolding and flowering onset dates might be more realistic. We utilised European plant phenological records provided by the European phenology database PEP725 and gridded monthly mean temperature data for 1951-2012 calculated from the ENSEMBLES data set E-OBS (version 7.0). We analysed 568,456 observations of ten spring flowering or leafing phenophases derived from 3657 stations in 22 European countries in order to detect possible nonlinear responses to temperature. Linear response rates averaged for all stations ranged between -7.7 (flowering of hazel) and -2.7?days?°C-1 (leaf unfolding of beech and oak). A lower sensitivity at the cooler end of the temperature range was detected for most phenophases. However, a similar lower sensitivity at the warmer end was not that evident. For only ~14?% of the station time series (where a comparison between linear and nonlinear model was possible), nonlinear models described the relationship significantly better than linear models. Although in most cases simple linear models might be still sufficient to predict future changes, this linear relationship between phenology and temperature might not be appropriate when incorporating phenological data of very cold (and possibly very warm) environments. For these cases, extrapolations on the basis of linear models would introduce uncertainty in expected ecosystem changes.
[本文引用: 1]
DOI:10.1111/gcb.12360URL [本文引用: 1]
It is well known that increased spring temperatures cause earlier onset dates of leaf unfolding and flowering. However, a temperature increase in winter may be associated with delayed development when species' chilling requirements are not fulfilled. Furthermore, photosensitivity is supposed to interfere with temperature triggers. To date, neither the relative importance nor possible interactions of these three factors have been elucidated. In this study, we present a multispecies climate chamber experiment to test the effects of chilling and photoperiod on the spring phenology of 36 woody species. Several hypotheses regarding their variation with species traits (successional strategy, floristic status, climate of their native range) were tested. Long photoperiods advanced budburst for one-third of the studied species, but magnitudes of these effects were generally minor. In contrast to prior hypotheses, photosensitive responses were not restricted to climax or oceanic species. Increased chilling length advanced budburst for almost all species; its effect greatly exceeding that of photoperiod. Moreover, we suggest that photosensitivity and chilling effects have to be rigorously disentangled, as the response to photoperiod was restricted to individuals that had not been fully chilled. The results indicate that temperature requirements and successional strategy are linked, with climax species having higher chilling and forcing requirements than pioneer species. Temperature requirements of invasive species closely matched those of native species, suggesting that high phenological concordance is a prerequisite for successful establishment. Lack of chilling not only led to a considerable delay in budburst but also caused substantial changes in the chronological order of species' budburst. The results reveal that increased winter temperatures might impact forest ecosystems more than formerly assumed. Species with lower chilling requirements, such as pioneer or invasive species, might profit from warming winters, if late spring frost events would in parallel occur earlier.
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.agrformet.2009.06.013URL [本文引用: 1]
DOI:10.3389/fpls.2015.00110URLPMID:25759707 [本文引用: 2]
Damage by late spring frost is a risk deciduous trees have to cope with in order to optimize the length of their growing season. The timing of spring phenological development plays a crucial role, not only at the species level, but also at the population and individual level, since fresh new leaves are especially vulnerable. For the pronounced late spring frost in May 2011 in Germany, we studied the individual leaf development of 35 deciduous trees (mainly European beech Fagus sylvatica L.) at a mountainous forest site in the Bayerischer Wald National Park using repeated digital photographs. Analyses of the time series of greenness by a novel Bayesian multiple change point approach mostly revealed five change points which almost perfectly matched the expected break points in leaf development: (i) start of the first greening between day of the year (DOY) 108-119 (mean 113), (ii) end of greening, and (iii) visible frost damage after the frost on the night of May 3rd/4th (DOY 123/124), (iv) re-sprouting 19-38 days after the frost, and (v) full maturity around DOY 178 (166-184) when all beech crowns had fully recovered. Since frost damage was nearly 100%, individual susceptibility did not depend on the timing of first spring leaf unfolding. However, we could identify significant patterns in fitness linked to an earlier start of leaf unfolding. Those individuals that had an earlier start of greening during the first flushing period had a shorter period of recovery and started the second greening earlier. Thus, phenological timing triggered the speed of recovery from such an extreme event. The maximum greenness achieved, however, did not vary with leaf unfolding dates. Two mountain ashes (Sorbus aucuparia L.) were not affected by the low temperatures of -5°C. Time series analysis of webcam pictures can thus improve process-based knowledge and provide valuable insights into the link between phenological variation, late spring frost damage, and recovery within one stand.
URL [本文引用: 1]
Response to chilling temperatures is a critical factor in the suitability of peach [Prunus persica (L.) Batsch] cultivars to moderate climates such as in the southeastern United States. Time of bloom depends on the innate chilling requirement of the cultivar as well as the timing and duration of cold and warm temperatures experienced by the buds. Most current chilling models have considered dormancy break a sequential process: after appropriate chilling is received, a fixed amount of heat accumulation will produce bloom. This research shows that as chilling exposure increases, the time and heat accumulation required for peach floral budbreak decreases in an exponential relationship. When chilling is ample, a sequential, two-part dormancy model is adequate to describe budbreak, and increased chilling has little effect on heat requirement. In lower-chilling situations relative to a particular cultivar, the curvilinear relationship between chilling and heat requirement results in more heat being required for budbreak. This relationship, in conjunction with the range of chilling requirement found among the buds on a tree, results in an extended, asymmetrical budbreak.
DOI:10.1111/j.1365-2486.2011.02562.xURL [本文引用: 1]
[本文引用: 1]
DOI:10.1111/j.1365-2745.2011.01798.xURL [本文引用: 1]
P>1. Growing recognition of the importance of climate extremes as drivers of contemporary and future ecological dynamics has led to increasing interest in studying these locally and globally important phenomena.
2. Many ecological studies examining the impacts of what are deemed climate extremes, such as heat waves and severe drought, do not provide a definition of extremity, either from a statistical context based on the long-term climatic record or from the perspective of the response of the system - are the effects extreme (unusual or profound) in comparison to normal variability?;3. A synthetic definition of an extreme climatic event (ECE) is proposed that includes 'extremeness' in both the driver and the response: an ECE is as an episode or occurrence in which a statistically rare or unusual climatic period alters ecosystem structure and/or function well outside the bounds of what is considered typical or normal variability. This definition is accompanied by a mechanistic framework based on the concept that extreme response thresholds associated with significant community change and altered ecosystem function must be crossed in order for an ECE to occur.
4. Synthesis. A definition and mechanistic framework for ECEs is used to identify priorities for future research that will enable ecologists to more fully assess the ecological consequences of climate extremes for ecosystem structure and function today and in a future world where their frequency and intensity are expected to increase.
DOI:10.1126/science.aaf7271URLPMID:27339968 [本文引用: 1]
DOI:10.1007/s00484-017-1312-6URLPMID:28220255 [本文引用: 1]
Long-term changes of plant phenological phases determined by complex interactions of environmental factors are in the focus of recent climate impact research. There is a lack of studies on the comparison of biogeographical regions in Europe in terms of plant responses to climate. We examined the flowering phenology of plant species to identify the spatio-temporal patterns in their responses to environmental variables over the period 1970-2010. Data were collected from 12 countries along a 3000-km-long, North-South transect from northern to eastern Central Europe.Biogeographical regions of Europe were covered from Finland to Macedonia. Robust statistical methods were used to determine the most influential factors driving the changes of the beginning of flowering dates. Significant species-specific advancements in plant flowering onsets within the Continental (3 to 8.3 days), Alpine (2 to 3.8 days) and by highest magnitude in the Boreal biogeographical regions (2.2 to 9.6 days per decades) were found, while less pronounced responses were detected in the Pannonian and Mediterranean regions. While most of the other studies only use mean temperature in the models, we show that also the distribution of minimum and maximum temperatures are reasonable to consider as explanatory variable. Not just local (e.g. temperature) but large scale (e.g. North Atlantic Oscillation) climate factors, as well as altitude and latitude play significant role in the timing of flowering across biogeographical regions of Europe. Our analysis gave evidences that species show a delay in the timing of flowering with an increase in latitude (between the geographical coordinates of 40.9 and 67.9), and an advance with changing climate. The woody species (black locust and small-leaved lime) showed stronger advancements in their timing of flowering than the herbaceous species (dandelion, lily of the valley). In later decades (1991-2010), more pronounced phenological change was detected than during the earlier years (1970-1990), which indicates the increased influence of human induced higher spring temperatures in the late twentieth century.
[本文引用: 1]
[本文引用: 1]
DOI:10.1127/0941-2948/2012/0330URL [本文引用: 1]
DOI:10.1007/s00484-017-1377-2URLPMID:28547481 [本文引用: 1]
Existing evidence demonstrates that the first flowering date (FFD) of most plant species became earlier in response to temperature increase over the past several decades. However, the studies on changes in flowering duration (FD) were limited. By using the non-parametric Theil-Sen estimator, this study investigated the temporal trends in 127 time series of FFD, end of flowering date (EFD), and FD of 97 woody plants from 1963 to 2014 at three sites (Harbin, Beijing, and Xi'an) in North China. The relationship between flowering phenophases and temperature was analyzed using two phenological models. The results showed that most of FFD and EFD time series exhibited an apparent advancing trend. Among them, trends of 52.0% (40.9%) of FFD (EFD) time series were significant (P?<?0.05). FFD and EFD time series (95.3 and 89.8%, respectively) responded negatively and significantly to preseason temperature (P?<?0.05). The direction of FD changes varied among sites and species. On average, a shortening trend of FD was observed at Harbin (-0.51?days?decade-1), with 7.5% of species significantly. However, FD on average extended by 0.42 and 0.93?days?decade-1 at Beijing (24.5% significantly) and Xi'an (28.9% significantly), respectively. The regression models could simulate the interannual changes in FFD and EFD with the mean goodness of fit (R2) ranging from 0.37 to 0.67, but fail to simulate the changes in FD accurately (R2 ranging from 0.09 to 0.18). The growing degree day model could improve the R2 for simulating FFD and EFD except for FD. Therefore, more phenological models need to be tested, and more drivers of FD need to be further investigated.
DOI:10.1111/pce.12431URLPMID:25142260 [本文引用: 1]
Increasing temperatures should facilitate the poleward movement of species distributions through a variety of processes, including increasing the growing season length. However, in temperate and boreal latitudes, temperature is not the only cue used by trees to determine seasonality, as changes in photoperiod provide a more consistent, reliable annual signal of seasonality than temperature. Here, we discuss how day length may limit the ability of tree species to respond to climate warming in situ, focusing on the implications of photoperiodic sensing for extending the growing season and affecting plant phenology and growth, as well as the potential role of photoperiod in controlling carbon uptake and water fluxes in forests. We also review whether there are patterns across plant functional types (based on successional strategy, xylem anatomy and leaf morphology) in their sensitivity to photoperiod that we can use to predict which species or groups might be more successful in migrating as the climate warms, or may be more successfully used for forestry and agriculture through assisted migration schemes.
[本文引用: 1]
DOI:10.1073/pnas.1413090112URLPMID:25730847 [本文引用: 1]
Terrestrial gross primary productivity (GPP) varies greatly over time and space. A better understanding of this variability is necessary for more accurate predictions of the future climate-carbon cycle feedback. Recent studies have suggested that variability in GPP is driven by a broad range of biotic and abiotic factors operating mainly through changes in vegetation phenology and physiological processes. However, it is still unclear how plant phenology and physiology can be integrated to explain the spatiotemporal variability of terrestrial GPP. Based on analyses of eddy-covariance and satellite-derived data, we decomposed annual terrestrial GPP into the length of the CO2 uptake period (CUP) and the seasonal maximal capacity of CO2 uptake (GPPmax). The product of CUP and GPPmax explained &gt;90% of the temporal GPP variability in most areas of North America during 2000-2010 and the spatial GPP variation among globally distributed eddy flux tower sites. It also explained GPP response to the European heatwave in 2003 (r(2) = 0.90) and GPP recovery after a fire disturbance in South Dakota (r(2) = 0.88). Additional analysis of the eddy-covariance flux data shows that the interbiome variation in annual GPP is better explained by that in GPPmax than CUP. These findings indicate that terrestrial GPP is jointly controlled by ecosystem-level plant phenology and photosynthetic capacity, and greater understanding of GPPmax and CUP responses to environmental and biological variations will, thus, improve predictions of GPP over time and space.
[本文引用: 3]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
DOI:10.1038/nature12872URL [本文引用: 1]
Early flowering plants are thought to have been woody species restricted to warm habitats(1-3). This lineage has since radiated into almost every climate, with manifold growth forms(4). As angiosperms spread and climate changed, they evolved mechanisms to cope with episodic freezing. To explore the evolution of traits underpinning the ability to persist in freezing conditions, we assembled a large species-level database of growth habit (woody or herbaceous; 49,064 species), as well as leaf phenology (evergreen or deciduous), diameter of hydraulic conduits (that is, xylem vessels and tracheids) and climate occupancies (exposure to freezing). To model the evolution of species' traits and climate occupancies, we combined these data with an unparalleled dated molecular phylogeny (32,223 species) for land plants. Here we show that woody clades successfully moved into freezing-prone environments by either possessing transport networks of small safe conduits(5) and/or shutting down hydraulic function by dropping leaves during freezing. Herbaceous species largely avoided freezing periods by senescing cheaply constructed aboveground tissue. Growth habit has long been considered labile(6), but we find that growth habit was less labile than climate occupancy. Additionally, freezing environments were largely filled by lineages that had already become herbs or, when remaining woody, already had small conduits (that is, the trait evolved before the climate occupancy). By contrast, most deciduous woody lineages had an evolutionary shift to seasonally shedding their leaves only after exposure to freezing (that is, the climate occupancy evolved before the trait). For angiosperms to inhabit novel cold environments they had to gain new structural and functional trait solutions; our results suggest that many of these solutions were probably acquired before their foray into the cold.
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/gcb.14002URLPMID:29197142 [本文引用: 1]
Constraints of temperature on spring plant phenology are closely related to plant growth, vegetation dynamics, and ecosystem carbon cycle. However, the effects of temperature on leaf onset, especially for winter chilling, are still not well understood. Using long-term, widespread in?situ phenology observations collected over China for multiple plant species, this study analyzes the quantitative response of leaf onset to temperature, and compares empirical findings with existing theories and modeling approaches, as implemented in 18 phenology algorithms. Results show that the growing degree days (GDD) required for leaf onset vary distinctly among plant species and geographical locations as well as at organizational levels (species and community), pointing to diverse adaptation strategies. Chilling durations (CHD) needed for releasing bud dormancy decline monotonously from cold to warm areas with very limited interspecies variations. Results also reveal that winter chilling is a crucial component of phenology models, and its effect is better captured with an index that accounts for the inhomogeneous effectiveness of low temperature to chilling rate than with the conventional CHD index. The impact of spring warming on leaf onset is nonlinear, better represented by a logistical function of temperature than by the linear function currently implemented in biosphere models. The optimized base temperatures for thermal accumulation and the optimal chilling temperatures are species-dependent and average at 6.9 and 0.2°C, respectively. Overall, plants' chilling requirement is not a constant, and more chilling generally results in less requirement of thermal accumulation for leaf onset. Our results clearly demonstrate multiple deficiencies of the parameters (e.g., base temperature) and algorithms (e.g., method for calculating GDD) in conventional phenology models to represent leaf onset. Therefore, this study not only advances our mechanistic and quantitative understanding of temperature controls on leaf onset but also provides critical information for improving existing phenology models.
[本文引用: 1]
[本文引用: 1]
DOI:10.18402/resci.2017.11.10URL [本文引用: 2]
Developing flowering phenological models is conducive to the accurate simulation of flowering periods of ornamental plants and could provide basis for seasonal flowering tourism events. We investigated four widespread ornamental plants with high aesthetic value,namely,Amygdalus persica,Armeniaca vulgaris,Cercis chinensis and Syringa oblate. Using the first flowering date (FFD)and end of flowering date (EFD)data of these species and corresponding meteorological data at 42 sites,we developed and validated the spatiotemporal model of FFD and EFD and reconstructed data series of FFD,EFD and flowering duration (FD)of the four species over their distribution area from 1962 to 2013. At last,we analyzed spatiotemporal patterns of mean phenophases and phenological changes. The results showed that the spatiotemporal model was able to simulate the flowering phenology accurately on large spatial and temporal scales with root-mean-square-error of about 4–6 days. The simulated mean FFD,EFD and FD followed certain geographical gradients. Latitude was the major factor influencing flowering phenology. The FFD and EFD were delayed by 1.23–4.46 days and the FD was extended by 0.07–1.47 days per degree increase of latitude. Over the past 50 years,the mean FFD and EFD of all species became earlier with a rate of 0.95–1.61 days decade-1. The advance of S. oblate FFD and EFD exhibited no obvious spatial pattern,while the changes of FFD and EFD for the other three species showed a noticeable spatial variation with clearer advance in the north than in the south. The FD of S. oblate extended by 0.20 days decade-1,while the other three species showed very weak trends of -0.01–0.07 days decade-1. The changes of FD showed strong spatial heterogeneity. These results provided a scientific basis for simulating flowering phenology of typical ornamental plants and assessing their phenological responses to climate change.
DOI:10.18402/resci.2017.11.10URL [本文引用: 2]
Developing flowering phenological models is conducive to the accurate simulation of flowering periods of ornamental plants and could provide basis for seasonal flowering tourism events. We investigated four widespread ornamental plants with high aesthetic value,namely,Amygdalus persica,Armeniaca vulgaris,Cercis chinensis and Syringa oblate. Using the first flowering date (FFD)and end of flowering date (EFD)data of these species and corresponding meteorological data at 42 sites,we developed and validated the spatiotemporal model of FFD and EFD and reconstructed data series of FFD,EFD and flowering duration (FD)of the four species over their distribution area from 1962 to 2013. At last,we analyzed spatiotemporal patterns of mean phenophases and phenological changes. The results showed that the spatiotemporal model was able to simulate the flowering phenology accurately on large spatial and temporal scales with root-mean-square-error of about 4–6 days. The simulated mean FFD,EFD and FD followed certain geographical gradients. Latitude was the major factor influencing flowering phenology. The FFD and EFD were delayed by 1.23–4.46 days and the FD was extended by 0.07–1.47 days per degree increase of latitude. Over the past 50 years,the mean FFD and EFD of all species became earlier with a rate of 0.95–1.61 days decade-1. The advance of S. oblate FFD and EFD exhibited no obvious spatial pattern,while the changes of FFD and EFD for the other three species showed a noticeable spatial variation with clearer advance in the north than in the south. The FD of S. oblate extended by 0.20 days decade-1,while the other three species showed very weak trends of -0.01–0.07 days decade-1. The changes of FD showed strong spatial heterogeneity. These results provided a scientific basis for simulating flowering phenology of typical ornamental plants and assessing their phenological responses to climate change.
DOI:10.17521/cjpe.2017.0305URL [本文引用: 1]
长期以来, 气候与植物物候关系的研究大多基于线性模型, 但植被物候对气候变化的响应可能是非线性的。该文利用非线性模型——生存分析模型来分析时间序列中过去事件(气候因子)对目的变量(物候)的作用: 用生存分析模型分析了春季气温和降水量对内蒙古草地、青藏高原草甸和欧洲地区木本植物返青期的影响。其中, 内蒙古与青藏高原的物候信息来自遥感数据, 欧洲地区物候信息为实测数据。蒙特卡洛方法用于拟合模型参数。结果表明: 生存分析模型适合对上述不同研究对象的物候影响因素进行分析, 并能模拟非线性效应; 在内蒙古草地, 春季气温和降水对春季返青有很大的影响, 而青藏高原草甸和欧洲地区木本植物返青期对春季降水响应较小, 对春季气温变化的响应随Holdridge干燥度指数上升而下降; 在预测返青期时发现: 春季平均气温提高1 ℃会导致上述地区返青期提前1-6天; 而春季气温与降水的增加会导致返青期发生明显的非线性变化, 这种非线性效应无法基于线性模型模拟出来。结果说明生存分析模型既能用于分析不同尺度下植物物候与气候的关系, 也能用于模型预测, 尤其适合探讨大幅度气候变化对物候的非线性影响。
DOI:10.17521/cjpe.2017.0305URL [本文引用: 1]
长期以来, 气候与植物物候关系的研究大多基于线性模型, 但植被物候对气候变化的响应可能是非线性的。该文利用非线性模型——生存分析模型来分析时间序列中过去事件(气候因子)对目的变量(物候)的作用: 用生存分析模型分析了春季气温和降水量对内蒙古草地、青藏高原草甸和欧洲地区木本植物返青期的影响。其中, 内蒙古与青藏高原的物候信息来自遥感数据, 欧洲地区物候信息为实测数据。蒙特卡洛方法用于拟合模型参数。结果表明: 生存分析模型适合对上述不同研究对象的物候影响因素进行分析, 并能模拟非线性效应; 在内蒙古草地, 春季气温和降水对春季返青有很大的影响, 而青藏高原草甸和欧洲地区木本植物返青期对春季降水响应较小, 对春季气温变化的响应随Holdridge干燥度指数上升而下降; 在预测返青期时发现: 春季平均气温提高1 ℃会导致上述地区返青期提前1-6天; 而春季气温与降水的增加会导致返青期发生明显的非线性变化, 这种非线性效应无法基于线性模型模拟出来。结果说明生存分析模型既能用于分析不同尺度下植物物候与气候的关系, 也能用于模型预测, 尤其适合探讨大幅度气候变化对物候的非线性影响。
Validation of chill unit and flower bud phenology models for ‘Montmorency’ sour cherry
2
1986
... 第三种计算积温的方法是生长度时.与其他两种方法不同, 该方法根据逐小时的气温计算积温.另外, 该方法假设存在发育速率的下限温度、上限温度和最适温度, 高于或低于最适温度时, 发育速率较低.具体公式(
... 式中GDH为生长度时; TL为发育的下限温度; Tu为发育的最适温度; Tc为发育的上限温度.根据
1
2010
... 在北半球温带地区, 温度是驱动木本植物展叶始期变化的最主要因子(
西安木本植物物候与气候要素的关系
1
2010
... 根据以往结果, 西安木本植物的展叶始期主要与3或4月的平均气温显著相关(
西安木本植物物候与气候要素的关系
1
2010
... 根据以往结果, 西安木本植物的展叶始期主要与3或4月的平均气温显著相关(
One man, 73 years, and 25 species. Evaluating phenological responses using a lifelong study of first flowering dates
1
2013
... 植物物候是反映气候变化对生态系统影响的重要指标.过去几十年, 北半球温带大部分地区的植物春季物候期都出现了提前的趋势(
Thermal time, chill days and prediction of budburst in Picea sitchensis.
2
1983
... 式中GDD为第一种算法计算得到的积温.t1代表展叶始期, t0为积温开始累积的日期, 本文设为1月1日.x(t)为第t天的日平均气温.Tb为阈值或基础温度, 根据以往研究设为5 ℃ (
... 本文结果表明偏暖年木本植物展叶的积温需求高于正常年, 在偏冷年的积温需求略低于正常年, 也与其他结果相类似.例如美国白蜡树(Fraxinus americana)在偏暖的2012年, 展叶始期的积温需求显著高于其他年份(
Warmest extreme year in US history alters thermal requirements for tree phenology
1
2017
... 在北半球温带地区, 温度是驱动木本植物展叶始期变化的最主要因子(
Changes in climate extreme events in China associated with warming
1
2015
... 1951到2012年, 全球地表平均温度以每10年0.12 ℃的速度升高了0.73 ℃, 这与人类活动造成的温室气体浓度增加密切相关(
Identification and timing of dormant and ontogenetic phase for sweet cherries in Northeast Germany for modelling purposes
1
2017
... 在北半球温带地区, 温度是驱动木本植物展叶始期变化的最主要因子(
New algorithm for generating hourly temperature values using daily maximum, minimum and average values from climate models
1
2007
... 本研究所用的气象数据来源于中国气象数据网(https://data.cma.cn/)中的中国地面气候资料日值数据集(V3.0).其中, 使用了西安和泾河两站1963- 2018年日平均、最高和最低气温数据.2005年位于西安市未央路上的西安基准气候站(城区)迁至20 km以外的泾河(郊区).两站共同观测年份只有2007和2008年, 之后只有泾河站有观测数据.由于西安站距物候观测点(西安植物园)的距离更近(约10 km), 因此本文利用两站的月气温差异, 将泾河各月的日气温均一化到西安(
1
... 在北半球温带地区, 温度是驱动木本植物展叶始期变化的最主要因子(
Tree phenology responses to winter chilling, spring warming, at north and south range limits
1
2014
... 在北半球温带地区, 温度是驱动木本植物展叶始期变化的最主要因子(
Phenological tracking enables positive species responses to climate change
1
2012
... 本研究发现气候波动使植物的春季物候期出现了较大变化, 西安39种植物的展叶始期在偏暖年比偏冷年平均早16.8天之多.该研究结果与其他地区的结果类似.例如, 北京地区部分荚蒾属(Viburnum)植物的始花期在2016年的偏暖年比2010年的偏冷年提前了1个月以上(
The spatial pattern of leaf phenology and its response to climate change in China
1
2014
... 在北半球温带地区, 温度是驱动木本植物展叶始期变化的最主要因子(
Warming and wetting signals emerging from analysis of changes in climate extreme indices over South America
1
2013
... 1951到2012年, 全球地表平均温度以每10年0.12 ℃的速度升高了0.73 ℃, 这与人类活动造成的温室气体浓度增加密切相关(
Record-breaking early flowering in the eastern United States
1
2013
... 植物物候是反映气候变化对生态系统影响的重要指标.过去几十年, 北半球温带大部分地区的植物春季物候期都出现了提前的趋势(
Variation in responsiveness of woody plant leaf out phenology to anomalous spring onset
1
2016
... 植物物候是反映气候变化对生态系统影响的重要指标.过去几十年, 北半球温带大部分地区的植物春季物候期都出现了提前的趋势(
Effects of climate extremes on the terrestrial carbon cycle: Concepts, processes and potential future impacts
1
2015
... 1951到2012年, 全球地表平均温度以每10年0.12 ℃的速度升高了0.73 ℃, 这与人类活动造成的温室气体浓度增加密切相关(
A tale of two springs: Using recent climate anomalies to characterize the sensitivity of temperate forest phenology to climate change
1
2014
... 本研究发现气候波动使植物的春季物候期出现了较大变化, 西安39种植物的展叶始期在偏暖年比偏冷年平均早16.8天之多.该研究结果与其他地区的结果类似.例如, 北京地区部分荚蒾属(Viburnum)植物的始花期在2016年的偏暖年比2010年的偏冷年提前了1个月以上(
Phenological response to climate change in China: A meta-analysis
1
2015
... 植物物候是反映气候变化对生态系统影响的重要指标.过去几十年, 北半球温带大部分地区的植物春季物候期都出现了提前的趋势(
Citizen science: Linking the recent rapid advances of plant flowering in Canada with climate variability
1
2013
... 植物物候是反映气候变化对生态系统影响的重要指标.过去几十年, 北半球温带大部分地区的植物春季物候期都出现了提前的趋势(
Modelling bud dormancy release in trees from cool and temperate regions
1
1990
... 上述计算积温的方法假设发育速率与温度呈线性关系, 有实验证据表明发育速率与温度的关系是非线性的(
贵阳木本植物始花期对温度变化的敏感度
1
2017
... 植物物候是反映气候变化对生态系统影响的重要指标.过去几十年, 北半球温带大部分地区的植物春季物候期都出现了提前的趋势(
贵阳木本植物始花期对温度变化的敏感度
1
2017
... 植物物候是反映气候变化对生态系统影响的重要指标.过去几十年, 北半球温带大部分地区的植物春季物候期都出现了提前的趋势(
Predicting the timing of budburst in temperate trees
2
1992
... 在北半球温带地区, 温度是驱动木本植物展叶始期变化的最主要因子(
... 本研究共采用了3种积温计算方法.第一种算法是目前最常用的算法, 即将积温计算为高于一定阈值的逐日温度累加和(
2
... 1951到2012年, 全球地表平均温度以每10年0.12 ℃的速度升高了0.73 ℃, 这与人类活动造成的温室气体浓度增加密切相关(
... (3) GDD模型模拟的木本植物展叶始期在偏暖年比观测值平均早4.1天.这主要是因为植物在偏暖年的积温需求更高, 而常规的积温模型假设积温需求在年际间保持不变.IPCC报告预测全球气温在21世纪仍将持续升高, 高温事件的发生频次会持续增加(
Phenology plays an important role in the regulation of terrestrial ecosystem water-use efficiency in the northern hemisphere
1
2017
... 植物物候是反映气候变化对生态系统影响的重要指标.过去几十年, 北半球温带大部分地区的植物春季物候期都出现了提前的趋势(
西安近63年气候变化特征综合分析
1
2014
... 西安位于渭河流域中部关中盆地, 属于典型的暖温带半湿润大陆性季风气候, 年平均气温14.3 ℃, 年降水量557 mm (1981-2010年平均值).冬季寒冷而少雨雪, 其中1月气温最低, 降水最少, 多年平均气温只有0.3 ℃, 降水量只有6.5 mm; 夏季炎热多雨, 其中7月气温最高, 降水最多, 多年平均气温27.1 ℃, 降水量101.7 mm.过去50年来, 西安出现了显著的增温趋势(
西安近63年气候变化特征综合分析
1
2014
... 西安位于渭河流域中部关中盆地, 属于典型的暖温带半湿润大陆性季风气候, 年平均气温14.3 ℃, 年降水量557 mm (1981-2010年平均值).冬季寒冷而少雨雪, 其中1月气温最低, 降水最少, 多年平均气温只有0.3 ℃, 降水量只有6.5 mm; 夏季炎热多雨, 其中7月气温最高, 降水最多, 多年平均气温27.1 ℃, 降水量101.7 mm.过去50年来, 西安出现了显著的增温趋势(
Can we detect a nonlinear response to temperature in European plant phenology?
1
2016
... 在北半球温带地区, 温度是驱动木本植物展叶始期变化的最主要因子(
Endo-, para-, and ecodormancy: Physiological terminology and classification for dormancy research
1
1987
... 在北半球温带地区, 温度是驱动木本植物展叶始期变化的最主要因子(
Chilling outweighs photoperiod in preventing precocious spring development
1
2014
... 本文结果表明偏暖年木本植物展叶的积温需求高于正常年, 在偏冷年的积温需求略低于正常年, 也与其他结果相类似.例如美国白蜡树(Fraxinus americana)在偏暖的2012年, 展叶始期的积温需求显著高于其他年份(
1993-2012年西安城区城市热岛效应强度变化趋势及影响因素分析
1
2015
... 西安位于渭河流域中部关中盆地, 属于典型的暖温带半湿润大陆性季风气候, 年平均气温14.3 ℃, 年降水量557 mm (1981-2010年平均值).冬季寒冷而少雨雪, 其中1月气温最低, 降水最少, 多年平均气温只有0.3 ℃, 降水量只有6.5 mm; 夏季炎热多雨, 其中7月气温最高, 降水最多, 多年平均气温27.1 ℃, 降水量101.7 mm.过去50年来, 西安出现了显著的增温趋势(
1993-2012年西安城区城市热岛效应强度变化趋势及影响因素分析
1
2015
... 西安位于渭河流域中部关中盆地, 属于典型的暖温带半湿润大陆性季风气候, 年平均气温14.3 ℃, 年降水量557 mm (1981-2010年平均值).冬季寒冷而少雨雪, 其中1月气温最低, 降水最少, 多年平均气温只有0.3 ℃, 降水量只有6.5 mm; 夏季炎热多雨, 其中7月气温最高, 降水最多, 多年平均气温27.1 ℃, 降水量101.7 mm.过去50年来, 西安出现了显著的增温趋势(
Validation of winter chill models using historic records of walnut phenology
1
2009
... 第三种计算积温的方法是生长度时.与其他两种方法不同, 该方法根据逐小时的气温计算积温.另外, 该方法假设存在发育速率的下限温度、上限温度和最适温度, 高于或低于最适温度时, 发育速率较低.具体公式(
Patterns of late spring frost leaf damage and recovery in a European beech (
2
2015
... 但是, 气候变暖下提前的展叶始期并不一定对植物生长永远有利, 在展叶始期之后发生的晚霜冻事件会严重影响植物生长, 例如2011年5月发生在德国的晚霜冻事件使得落叶阔叶林的新叶全部凋落(
... 本文结果表明偏暖年木本植物展叶的积温需求高于正常年, 在偏冷年的积温需求略低于正常年, 也与其他结果相类似.例如美国白蜡树(Fraxinus americana)在偏暖的2012年, 展叶始期的积温需求显著高于其他年份(
Increasing chilling reduces heat requirement for floral budbreak in Peach
1
2011
... 本文结果表明偏暖年木本植物展叶的积温需求高于正常年, 在偏冷年的积温需求略低于正常年, 也与其他结果相类似.例如美国白蜡树(Fraxinus americana)在偏暖的2012年, 展叶始期的积温需求显著高于其他年份(
Terrestrial biosphere models need better representation of vegetation phenology: Results from the North American Carbon Program Site Synthesis
1
2012
... 目前, 很多研究均采用常规的GDD模型预测木本植物的春季物候期(
Investigations on the annual cycle of development on forest trees: Active period
1
1972
... 上述计算积温的方法假设发育速率与温度呈线性关系, 有实验证据表明发育速率与温度的关系是非线性的(
An ecological perspective on extreme climatic events: A synthetic definition and framework to guide future research
1
2011
... 1951到2012年, 全球地表平均温度以每10年0.12 ℃的速度升高了0.73 ℃, 这与人类活动造成的温室气体浓度增加密切相关(
How climate change affects extreme weather events
1
2016
... 1951到2012年, 全球地表平均温度以每10年0.12 ℃的速度升高了0.73 ℃, 这与人类活动造成的温室气体浓度增加密切相关(
Phenological patterns of flowering across biogeographical regions of Europe
1
2017
... 植物物候是反映气候变化对生态系统影响的重要指标.过去几十年, 北半球温带大部分地区的植物春季物候期都出现了提前的趋势(
1
... 本文所用的物候数据来源于中国物候观测网西安站点, 共采用了1963-2018年39种木本植物的展叶始期数据(
1
... 本文所用的物候数据来源于中国物候观测网西安站点, 共采用了1963-2018年39种木本植物的展叶始期数据(
Extreme climate in China: Facts, simulation and projection
1
2012
... 1951到2012年, 全球地表平均温度以每10年0.12 ℃的速度升高了0.73 ℃, 这与人类活动造成的温室气体浓度增加密切相关(
Changes in flowering phenology of woody plants from 1963 to 2014 in North China
1
2019
... 植物物候是反映气候变化对生态系统影响的重要指标.过去几十年, 北半球温带大部分地区的植物春季物候期都出现了提前的趋势(
Photoperiod constraints on tree phenology, performance and migration in a warming world
1
2015
... 本文结果表明偏暖年木本植物展叶的积温需求高于正常年, 在偏冷年的积温需求略低于正常年, 也与其他结果相类似.例如美国白蜡树(Fraxinus americana)在偏暖的2012年, 展叶始期的积温需求显著高于其他年份(
1
... 本文所用的物候数据来源于中国物候观测网西安站点, 共采用了1963-2018年39种木本植物的展叶始期数据(
Joint control of terrestrial gross primary productivity by plant phenology and physiology
1
2015
... 植物物候是反映气候变化对生态系统影响的重要指标.过去几十年, 北半球温带大部分地区的植物春季物候期都出现了提前的趋势(
荚蒾属植物花期物候对春季异常气象事件的响应
3
2017
... 植物物候是反映气候变化对生态系统影响的重要指标.过去几十年, 北半球温带大部分地区的植物春季物候期都出现了提前的趋势(
... 在北半球温带地区, 温度是驱动木本植物展叶始期变化的最主要因子(
... 本研究发现气候波动使植物的春季物候期出现了较大变化, 西安39种植物的展叶始期在偏暖年比偏冷年平均早16.8天之多.该研究结果与其他地区的结果类似.例如, 北京地区部分荚蒾属(Viburnum)植物的始花期在2016年的偏暖年比2010年的偏冷年提前了1个月以上(
荚蒾属植物花期物候对春季异常气象事件的响应
3
2017
... 植物物候是反映气候变化对生态系统影响的重要指标.过去几十年, 北半球温带大部分地区的植物春季物候期都出现了提前的趋势(
... 在北半球温带地区, 温度是驱动木本植物展叶始期变化的最主要因子(
... 本研究发现气候波动使植物的春季物候期出现了较大变化, 西安39种植物的展叶始期在偏暖年比偏冷年平均早16.8天之多.该研究结果与其他地区的结果类似.例如, 北京地区部分荚蒾属(Viburnum)植物的始花期在2016年的偏暖年比2010年的偏冷年提前了1个月以上(
1978-2014年牡丹江地区植物花期变化及模型模拟
1
2017
... 目前, 很多研究均采用常规的GDD模型预测木本植物的春季物候期(
1978-2014年牡丹江地区植物花期变化及模型模拟
1
2017
... 目前, 很多研究均采用常规的GDD模型预测木本植物的春季物候期(
Three keys to the radiation of angiosperms into freezing environments
1
2014
... 本文所用的物候数据来源于中国物候观测网西安站点, 共采用了1963-2018年39种木本植物的展叶始期数据(
物候模型在北京观赏植物开花期预测中的适用性
1
2014
... 目前, 很多研究均采用常规的GDD模型预测木本植物的春季物候期(
物候模型在北京观赏植物开花期预测中的适用性
1
2014
... 目前, 很多研究均采用常规的GDD模型预测木本植物的春季物候期(
New insights on plant phenological response to temperature revealed from long-term widespread observations in China
1
2018
... 目前, 很多研究均采用常规的GDD模型预测木本植物的春季物候期(
类型变更的相邻气象观测站的日气温资料整合
1
2012
... 本研究所用的气象数据来源于中国气象数据网(https://data.cma.cn/)中的中国地面气候资料日值数据集(V3.0).其中, 使用了西安和泾河两站1963- 2018年日平均、最高和最低气温数据.2005年位于西安市未央路上的西安基准气候站(城区)迁至20 km以外的泾河(郊区).两站共同观测年份只有2007和2008年, 之后只有泾河站有观测数据.由于西安站距物候观测点(西安植物园)的距离更近(约10 km), 因此本文利用两站的月气温差异, 将泾河各月的日气温均一化到西安(
类型变更的相邻气象观测站的日气温资料整合
1
2012
... 本研究所用的气象数据来源于中国气象数据网(https://data.cma.cn/)中的中国地面气候资料日值数据集(V3.0).其中, 使用了西安和泾河两站1963- 2018年日平均、最高和最低气温数据.2005年位于西安市未央路上的西安基准气候站(城区)迁至20 km以外的泾河(郊区).两站共同观测年份只有2007和2008年, 之后只有泾河站有观测数据.由于西安站距物候观测点(西安植物园)的距离更近(约10 km), 因此本文利用两站的月气温差异, 将泾河各月的日气温均一化到西安(
中国典型观赏植物花期模型建立及过去花期变化模拟
2
2017
... 在北半球温带地区, 温度是驱动木本植物展叶始期变化的最主要因子(
... 目前, 很多研究均采用常规的GDD模型预测木本植物的春季物候期(
中国典型观赏植物花期模型建立及过去花期变化模拟
2
2017
... 在北半球温带地区, 温度是驱动木本植物展叶始期变化的最主要因子(
... 目前, 很多研究均采用常规的GDD模型预测木本植物的春季物候期(
中国草地和欧洲木本植物返青期对气温和降水变化的响应: 基于生存分析的研究
1
2018
... 在北半球温带地区, 温度是驱动木本植物展叶始期变化的最主要因子(
中国草地和欧洲木本植物返青期对气温和降水变化的响应: 基于生存分析的研究
1
2018
... 在北半球温带地区, 温度是驱动木本植物展叶始期变化的最主要因子(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}